
İnsular Bölgenin Anatomisi ve Cerrahi Yaklaşım Yolları The ...

This article was downloaded by: [Universita Studi la Sapienza]On: 14 May 2014, At: 07:19Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Italian Journal of ZoologyPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/tizo20
Morphometric and genetic divergence in island andmainland populations of Anolis nebulosus (Squamata:Polychrotidae) from Jalisco (Mexico): an instance ofinsular gigantismG. Senczuka, A. Garcíab, P. Colangeloa, F. Annesia & R. Castigliaa
a Dipartimento di Biologia e Biotecnologie “Charles Darwin”, Università di Roma “LaSapienza”, sede di Anatomia Comparata, Rome, Italyb Estación de Biología Chamela Instituto de Biología, Universidad Nacional Autónoma deMéxico, Jalisco, MexicoPublished online: 13 May 2014.
To cite this article: G. Senczuk, A. García, P. Colangelo, F. Annesi & R. Castiglia (2014): Morphometric and geneticdivergence in island and mainland populations of Anolis nebulosus (Squamata: Polychrotidae) from Jalisco (Mexico): aninstance of insular gigantism, Italian Journal of Zoology, DOI: 10.1080/11250003.2014.911973
To link to this article: http://dx.doi.org/10.1080/11250003.2014.911973
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of theContent. Any opinions and views expressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information. Taylor and Francis shall not be liable forany losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use ofthe Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Morphometric and genetic divergence in island and mainlandpopulations of Anolis nebulosus (Squamata: Polychrotidae) from Jalisco(Mexico): an instance of insular gigantism
G. SENCZUK1, A. GARCÍA2, P. COLANGELO1, F. ANNESI1, & R. CASTIGLIA1*
1Dipartimento di Biologia e Biotecnologie “Charles Darwin”, Università di Roma “La Sapienza”, sede di AnatomiaComparata, Rome, Italy, and 2Estación de Biología Chamela Instituto de Biología, Universidad Nacional Autónoma deMéxico, Jalisco, Mexico
(Received 10 October 2013; accepted 23 March 2014)
AbstractThe clouded anole Anolis nebulosus (Squamata: Polychrotidae) is widespread on the Pacific coast of Mexico. The species alsoinhabits Don Panchito, a small islet located near the coast of the Chamela-Cuixmala Biosphere Reserve in the state ofJalisco. We studied the extent of intraspecific differences in morphology (absolute size and body proportions) and inmtDNA sequences (16S and NDH2) between the population living on the islet (N = 18 for morphometry; N = 12 formtDNA) and the one on the facing mainland (N = 38 for morphometry; N = 16 for mtDNA). The individuals on the isletare larger than those on the mainland with little overlap in size for either males (islet: 52.79 ± 1.82 mm; mainland:40.96 ± 2.99 mm) or females (islet: 46.18 ± 3.24 mm; mainland 37.14 ± 2.13 mm). The presence of insular gigantism,as here found in A. nebulosus, seems uncommon in the genus and could be explained as a combination of low predationpressure and higher intraspecific competition on the island. Moreover, we found that sexual dimorphism (SD) is higher inthe island population than in the mainland one. The molecular analysis shows the absence of shared haplotypes between theisland and mainland populations. Ten mtDNA haplotypes belonged to the mainland population and three to the islandpopulation. The shape of the minimum spanning network and of the mismatch distribution indicates a single colonizationevent. These molecular data indicate a certain degree of isolation of the island population notwithstanding its proximity tothe coast. The morphological characteristics of the anoles on Don Panchito match with the expectation of the so-called“reversed island syndrome” theory, which predicts an increased body size and sexual dimorphism in lizards living on verysmall islands characterized by unpredictable environmental conditions.
Keywords: Anoles, body size, reversed island syndrome, mtDNA, Norops
Introduction
Island populations of lizards represent an idealmodel to study morphological changes and how theparticular selective pressures on islands can shapephenotypic traits (Herrel et al. 2008; Losos &Ricklefs 2009). The body size of animals living onislands often differs from that of their mainland rela-tives, with larger species becoming smaller and smal-ler species becoming larger, the so-called “islandrule” (Van Valen 1973). Recent data and analyses,however, have challenged this view (Meiri et al.2008). In fact, insular species often evolve othersubstantial differences compared to the mainlandones, a phenomenon called “island syndrome” (IS).
The IS predicts that insular taxa, living in densepopulations, will display characteristics includinglow aggressivity, small sexual dimorphism, lowfecundity and large size (Novosolov et al. 2013).Recently, Raia et al. (2010) shed new light on the
effect of insularism in reptiles and formulated a newtheory predicting that insular populations, in parti-cular unpredictable environmental conditions,would display an array of trait shifts opposite to theIS, which they called the “reversed island syndrome”(RIS). Such highly uncertain conditions, which maybe common on very small islands, will keep popula-tion density low or fluctuating and lizards couldevolve either large or small body size and allocate
*Correspondence: R. Castiglia, Dipartimento di Biologia e Biotecnologie “Charles Darwin”, Università di Roma “La Sapienza”, sede di Anatomia Comparata,via Borelli 50, 00161 Rome, Italy. Tel: +39 06 49918092. Fax: +39 06 4457516. Email: [email protected]
Italian Journal of Zoology, 2014, 1–11http://dx.doi.org/10.1080/11250003.2014.911973
© 2014 Unione Zoologica Italiana
Dow
nloa
ded
by [
Uni
vers
ita S
tudi
la S
apie
nza]
at 0
7:19
14
May
201
4

more energy to reproduction, by increasing sexualdimorphism and aggressiveness.
Anole lizards (Squamata: Iguania: Polychrotidae)are distributed throughout the Caribbean islands andmainland Northern, Central and South America(Losos 2009). Recently, the genus Anolis was thesubject of a systematic revision resulting in its splitinto eight different genera (Nicholson et al. 2012).However, this systematic arrangement has been hotlycriticized (e.g. Poe 2013), and in this paper we adoptthe classic view that anole lizards belong to a singlegenus encompassing nearly 380 species (Losos2009). Anoles exhibit a wide array of ecological andmorphological specializations and are a classic exam-ple of vertebrate adaptive radiation, with the pre-sence of “ecomorphs” (habitat-specialized species)that converge in morphology and behavior(Williams 1972; Losos et al. 1998). In addition,differences in relative limb length can be shaped bynatural selection as an adaptation to different habi-tats in populations of the same species (Calsbeeket al. 2007), and it has been experimentally demon-strated that this also occurs in offspring growing indifferent environments, due to morphological plasti-city (Losos et al. 2000; Kolbe & Losos 2005). Thissuggests that anoles are prone to rapid intraspecificmorphological changes and islands may represent anideal natural laboratory to study such events.
Examples of intraspecific size differences betweenisland and mainland populations are scarce. Islandpopulations of Anolis marmoratus (Duméril &Bibron, 1837) and Anolis aeneus (Gray, 1840) tendto be smaller than their mainland counterparts(Schwartz & Henderson 1991; Roughgarden 1995),while Anolis equestris (Merrem, 1820) is larger onsmall islands than on larger ones (Rodriguez-Schettino 1999). Interestingly, the opposite patterncan be observed in different populations of the samespecies. Thus, Anolis luteogularis (Noble & Hassler,1935) is smaller on some islands but not on otherswith respect to mainland populations (Rodriguez-Schettino 1999). In other instances, differences insize were not detected, for example in the compar-ison of island and mainland populations of Anolisbremeri (Barbour, 1914) and Anolis maynardi(Garman, 1888) (Herrel et al. 2011).
The present study focuses on Anolis nebulosus(Wiegmann, 1834), a mainland species widespreadon the Pacific coast of Mexico from Sonora toOaxaca. A. nebulosus also inhabits Don Panchito, anislet located near the coast of the Chamela-CuixmalaBiosphere Reserve in the state of Jalisco (Mexico).This population is known to consist of larger speci-mens than those living on the mainland (García,pers. obs.). However, a detailed analysis of the
morphological characteristics of this population islacking.This background provides a good opportunity to
study the extent of intraspecific morphological diver-gence between island and mainland populations inthe framework of the IS and RIS theories. Morespecifically, we think that the presently studied con-text could favor morphological characteristics com-patible with the RIS rather than with the IS; in fact,the RIS should more likely manifest itself at intra-specific levels, and on very small islands because oftheir unpredictable environment and resources(Novosolov et al. 2013).We compared the morphometric characteristics of
Anolis individuals living on the islet and those on thefacing mainland. The morphological comparisonincluded the absolute size and body proportions.We used these data (i) to examine differencesbetween the two populations and (ii) to assess theextent of sexual dimorphism in body size and bodyproportions. The morphological analysis wascoupled with a molecular one. We used thesequences of two mtDNA genes (i) to confirm theconspecificity of the two samples, (ii) to estimate thedegree of isolation of the islet and (iii) to understandthe modality (multiple vs. single) of colonization ofthe islet.
Materials and methods
Specimens and study area
The sampling took place in the Chamela-CuixmalaBiosphere Reserve (19° 22' 03” – 19° 35' 11” N and104° 56' 13” – 105° 03' 25” W) during July 2010.Thirty-eight adult specimens (21 males and 17females) were collected from different localities scat-tered on the mainland and 18 adult specimens (ninemales and nine females) on Don Panchito (Figure 1).All animals were captured by hand, weighed with aprecision scale (Pesola®) and carried back to the fieldstation for morphometric measurements and forremoval of a small tissue sample (the tip of the tail).All the specimens were released at the capture loca-tion the following day.Don Panchito is one of eight islets included in the
Chamela Bay islands nature sanctuary. This islet(3.30 ha) is situated 0.46 km from the coast and isthe only one where A. nebulosus has been found(García, pers. obs.). It is characterized by shrubbyvegetation with Leguminosae and Euphorbiaceatrees. Other lizards present on the island areAspidoscelis lineatissima (Cope, 1878), Ctenosaura pec-tinata (Wiegmann, 1834), Iguana iguana (Linnaeus,1758), Urosaurus bicarinatus (Duméril, 1856) and
2 G. Senczuk et al.
Dow
nloa
ded
by [
Uni
vers
ita S
tudi
la S
apie
nza]
at 0
7:19
14
May
201
4

Phyllodactylus lanei (Smith, 1935) (SEMARNAT2002). The climate of the area is characterized by asingle wet season and a severe dry season that lastsfrom October to June (Bullock 1986).Geomorphological studies indicate that the islandsof Chamela Bay were formed 18,000–7000 years ago(Curray et al. 1969; Ramı́rez-Herrera et al. 2004).Therefore, this frame is the most ancient limit for theage of this isolated population.
Genetic analysis
Genomic DNA was extracted from tail tissues bymeans of the universal extraction protocol of Aljanabiand Martinez (1997) with incubation at 56°C withproteinase K and precipitation of DNA with isopropa-nol. Fragments of two mtDNA genes, the NADHdehydrogenase 2 gene and flanking regions (NDH2)and the 16S rDNA gene (16S), were amplified by astandard polymerase chain reaction (PCR) methodwith the primers L4437: 5’-AAG-CTT-TCG-GGC-CCA-TAC-C-3’, H5934: 5’-AGR-GTG-CCA-ATG-TCT-TTG-TGR-TT-3’ for NDH2 and 16SA-L: 5’-CGC-CTG-TTT-ATC-AAA-AAC-AT-3’, 16SB-H5’-CCG-GTC-TGA-ACT-CAG-ATC-ACG-3’ for16S. PCR products (approximately 600 bp forNDH2 and 500 bp for 16S) were purified with aSureClean (Bioline) purification kit. Sequencing reac-tions were run under Big-Dye™ Terminator cyclingconditions. Sequences were aligned using ClustalX(Thompson et al. 1997) and a few adjustments weremade by eye for 16S.
Nucleotide (p) and haplotype (h) diversity, anddistance matrices based on pairwise differences,were estimated using Arlequin ver. 3.5 (Excoffier &Lischer 2010) and DnaSP ver. 5.10.1 (Librado &Rozas 2009).Because genetic variation within species is often
low, networks are a more appropriate way of asses-sing genetic variation than using the more commonphylogenetic tree-building algorithms. Thus we usedArlequin ver. 3.5 to construct a minimum spanningnetwork to connect haplotypes.In an island population, the genetic signature of an
expansion may be indicative of the last colonizationevent. Therefore, we assessed the population expan-sion on Don Panchito and on the mainland by mis-match distribution (MMD) analysis performed withDnaSP ver. 5.10.1. The significance of the differencebetween the expected and observed mismatch distri-bution was tested by goodness-of-fit with the para-metric bootstrap approach (Schneider & Excoffier1999). We calculated the parameter Tau (τ) of themismatch distribution, corresponding to the mode ofthe curve, with DnaSP ver. 5.10.1. Tau estimates thetime (in unit of mutational time) since the beginningof the expansion (Rogers & Harpending 1992).
Morphometric analysis
The following measures were taken for each indivi-dual: snout-vent length (SVL), from the tip of thesnout to the anterior end of the cloaca; head length(HeaL), from the anterior margin of the rostral scale
Figure 1. Aerial view of Chamela Bay islands nature sanctuary with Don Panchito islet indicated by the arrow.
Insular gigantism in an anole lizard 3
Dow
nloa
ded
by [
Uni
vers
ita S
tudi
la S
apie
nza]
at 0
7:19
14
May
201
4

to the anterior margin of the ear opening; headheight (HeaH), measured at the level of the eyes;mouth length (MoL), from the anterior margin ofthe rostral scale to the union of the supralabial andinfralabial scales; snout length (SL), from the ante-rior margin of the rostral scales to the anterior mar-gin of the eye; interorbital distance (IntD); trunklength (TrL), measured between the insertion ofthe extremities; forelimb length (ForLiL), from thepoint of insertion into the abdomen to the humero-radio-ulnar joint; forearm length (ForL); hindlimblength (HinL), from the point of insertion into theabdomen to the femoro-tibial joint; tibia length(TibL). All measurements were taken on the rightside by the same person (G. Senczuk) using a digitalcaliper (Storm® ± 0.01 mm). Each variable wasmeasured twice; an additional measurement (occa-sionally two additional measurements) was takenwhen the two original measurements were not within5% of each other. The mean of the two/three mea-sures was then used.
To estimate the body size of the lizard populationswe used the mean SVL, the upper decile SVL(UDL), and the size of the largest individuals (seeMeiri 2007 for a discussion of the various methodsfor estimating body size in lizards).
All variables were log-transformed and tested formultivariate normality (Doornick-Hansen test,P = 0.0002); a morphological matrix was then cre-ated including sex and geographic location (main-land – island populations) as explanatory variables.Next, we performed a principal component analysis(PCA) to reduce the multivariate matrix to a bivari-ate framework and to explore the distribution of theindividuals in the morphospace.
Differences in size variation among groups werefirst evaluated by two-way analysis of variance(ANOVA) using sex and geographic location asgrouping factors and SVL as response variable.Then multivariate analysis of covariance(MANCOVA) was performed, using the raw values,to evaluate the occurrence of significant morphologi-cal differences between sexes, geographic locationsand their interactions; SVL was included as covariateto assess the effect of size on the overall morphological
differences observed between sexes and geographiclocation. Successively, in order to investigate the dif-ferences in body proportions between mainland andisland populations, we controlled for the effect of thesize running a two-way analysis of covariance(ANCOVA) for all the characters, using the SVL ascovariate. Finally, we did a multivariate regression tocreate a matrix of size-corrected variables. This newmultivariate matrix was then used to perform a PCAon size correct variables.To quantify sexual dimorphism (SD) in the exam-
ined populations, we calculated squaredMahalanobis distances (D2) of both absolute andsize-corrected matrices; this statistic allows us tocalculate the distance between groups, relative tothe amount of within-group pooled variation in thedirection of the differences between groups.The morphometric analysis was performed with R
(Ihaka & Gentleman 1996) and PAST (Hammeret al. 2001) statistical software packages.
Results
Mitochondrial DNA
The sequenced fragments were 573 bp long forNDH2 and 438 bp long for 16S. The 16S geneshowed very low variability (three haplotypes andthree substitutions out of 15 specimens) (Genbankaccession numbers KJ606218–KJ606220).Therefore, it was removed from the subsequent ana-lysis since it was considered uninformative for thisscale of study. For NDH2, the 28 individuals pre-sented 15 polymorphic sites and 13 unique haplo-types (hap1-13, Genbank accession numbersKJ588374–KJ588386) (Table I). Ten haplotypesbelonged to the mainland population and three tothe island population. There were no shared haplo-types between the island and mainland populations.Accordingly, the mainland population had high hap-lotype (h = 0.925 ± 0.002) and nucleotide diversity(π = 0.0036 ± 0.0005), while the island populationshowed lower genetic indexes (h = 0.439 ± 0.158;π = 0.0008 ± 0.0003).The MSN connecting haplotypes (Figure 2)
showed a star-like relationship in the haplotypes
Table I. Number of individuals per locality, number of haplotypes for each locality, number of polymorphic sites, nucleotide and haplotypediversity.
No. ofindividuals
No. ofhaplotypes
No. ofpolymorphic sites
Nucleotide diversityπ ± SD
Haplotype diversityh ± SD
Mean no. of pairwisedifferences (± SD)
Islet 12 3 3 0.0008 (± 0.0003) 0.439 (± 0.158) 0.470 (± 0.436)Continent 16 10 12 0.0058 (± 0.0019) 0.934 (± 0.043) 3.316 (± 1.783)Total 28 13 15 0.0034 (± 0.0003) 0.878 (± 0.048) 2.837 (± 1.535)
4 G. Senczuk et al.
Dow
nloa
ded
by [
Uni
vers
ita S
tudi
la S
apie
nza]
at 0
7:19
14
May
201
4

from the mainland, with hap10 in a central positionand the others differing by one to three substitutions.In the island population, hap-11 was the most com-mon haplotype, present in nine of 12 specimens.The island population differed from the mainlandpopulation by a single G-A transition at posi-tion 474.
Figure 3 shows the MMD values for the island andmainland populations. The MMD values do notdiffer significantly from the Rogers (1995) suddenexpansion model (all p values not significant), mean-ing that the differences between the data and thetheoretical predictions are not significant (accordingto Harpending’s raggedness index). The mainlandpopulation has a bell-shaped unimodal distribution,as expected for populations with an expansion, withτ = 2.10. The island population has an L-shapeddistribution with τ = 0.47 suggesting a very recentdemographic expansion.
Morphometrics
Table II reports the mean and standard deviation forall the absolute measures. The island specimenswere distinctly larger than those on the mainlandfor all the measures (Figure 4). The mean SVL was52.8 mm in males (max SVL = 55.8 mm;UDL = 55.48) and 46.2 mm in females (maxSVL = 48.40 mm, UDL = 47.31 mm), while in themainland population it was 41.0 mm in males (maxSVL = 45.1 mm, UDL = 44.3 mm) and 37.1 mm infemales (max SVL = 41.5 mm, UDL = 40.47 mm).The mean weight was 3.0 g in males and 2.4 g infemales for the island population and 1.5 g in malesand 1.1 g in females for the mainland one.
The PCA showed a clear separation of the indivi-duals into four groups which displace according togeographic location along the first component(83.3% of total variance), and according to the sexalong the second component (4.4% of total variance)(Figure 5).Two-way ANOVA for SVL showed significant
differences between geographic locations(F = 167.66, P < 0.0001) and between sexes(F = 39.60, P < 0.0001) but no significant interac-tion (Table II).The MANCOVA for all variables, including sex
and geographic location as factors and SVL as cov-ariate, indicated significant differences in size (Wilks’Lambda = 0.0209, P < 0.0001) between sexes(Wilks’ Lambda = 0.338; P = 0.0001) and betweenthe geographic location (Wilks’ Lambda = 0.596;P = 0.016). We didn’t find any significant interactionamong factors.Mahalanobis distances on the log-transformed
multivariate matrix indicated greater sexual
Figure 2. Minimum spanning tree connecting haplotypes. Whitecircles: mainland haplotypes; gray circles: insular haplotypes.
Figure 3. Frequency distributions of pairwise differences, betweenobserved and expected distributions under the model of popula-tion growth-decline for (a) mainland and (b) island populations.
Insular gigantism in an anole lizard 5
Dow
nloa
ded
by [
Uni
vers
ita S
tudi
la S
apie
nza]
at 0
7:19
14
May
201
4
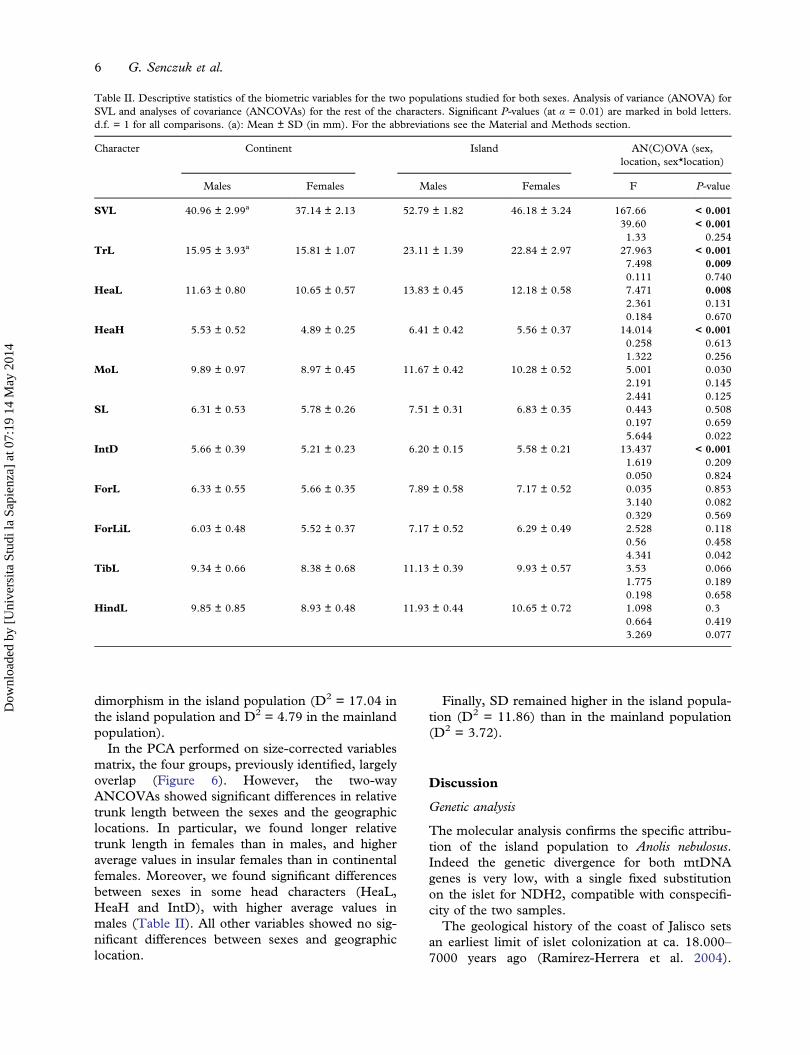
dimorphism in the island population (D2 = 17.04 inthe island population and D2 = 4.79 in the mainlandpopulation).
In the PCA performed on size-corrected variablesmatrix, the four groups, previously identified, largelyoverlap (Figure 6). However, the two-wayANCOVAs showed significant differences in relativetrunk length between the sexes and the geographiclocations. In particular, we found longer relativetrunk length in females than in males, and higheraverage values in insular females than in continentalfemales. Moreover, we found significant differencesbetween sexes in some head characters (HeaL,HeaH and IntD), with higher average values inmales (Table II). All other variables showed no sig-nificant differences between sexes and geographiclocation.
Finally, SD remained higher in the island popula-tion (D2 = 11.86) than in the mainland population(D2 = 3.72).
Discussion
Genetic analysis
The molecular analysis confirms the specific attribu-tion of the island population to Anolis nebulosus.Indeed the genetic divergence for both mtDNAgenes is very low, with a single fixed substitutionon the islet for NDH2, compatible with conspecifi-city of the two samples.The geological history of the coast of Jalisco sets
an earliest limit of islet colonization at ca. 18.000–7000 years ago (Ramı́rez-Herrera et al. 2004).
Table II. Descriptive statistics of the biometric variables for the two populations studied for both sexes. Analysis of variance (ANOVA) forSVL and analyses of covariance (ANCOVAs) for the rest of the characters. Significant P-values (at α = 0.01) are marked in bold letters.d.f. = 1 for all comparisons. (a): Mean ± SD (in mm). For the abbreviations see the Material and Methods section.
Character Continent Island AN(C)OVA (sex,location, sex*location)
Males Females Males Females F P-value
SVL 40.96 ± 2.99a 37.14 ± 2.13 52.79 ± 1.82 46.18 ± 3.24 167.66 < 0.00139.60 < 0.0011.33 0.254
TrL 15.95 ± 3.93a 15.81 ± 1.07 23.11 ± 1.39 22.84 ± 2.97 27.963 < 0.0017.498 0.0090.111 0.740
HeaL 11.63 ± 0.80 10.65 ± 0.57 13.83 ± 0.45 12.18 ± 0.58 7.471 0.0082.361 0.1310.184 0.670
HeaH 5.53 ± 0.52 4.89 ± 0.25 6.41 ± 0.42 5.56 ± 0.37 14.014 < 0.0010.258 0.6131.322 0.256
MoL 9.89 ± 0.97 8.97 ± 0.45 11.67 ± 0.42 10.28 ± 0.52 5.001 0.0302.191 0.1452.441 0.125
SL 6.31 ± 0.53 5.78 ± 0.26 7.51 ± 0.31 6.83 ± 0.35 0.443 0.5080.197 0.6595.644 0.022
IntD 5.66 ± 0.39 5.21 ± 0.23 6.20 ± 0.15 5.58 ± 0.21 13.437 < 0.0011.619 0.2090.050 0.824
ForL 6.33 ± 0.55 5.66 ± 0.35 7.89 ± 0.58 7.17 ± 0.52 0.035 0.8533.140 0.0820.329 0.569
ForLiL 6.03 ± 0.48 5.52 ± 0.37 7.17 ± 0.52 6.29 ± 0.49 2.528 0.1180.56 0.4584.341 0.042
TibL 9.34 ± 0.66 8.38 ± 0.68 11.13 ± 0.39 9.93 ± 0.57 3.53 0.0661.775 0.1890.198 0.658
HindL 9.85 ± 0.85 8.93 ± 0.48 11.93 ± 0.44 10.65 ± 0.72 1.098 0.30.664 0.4193.269 0.077
6 G. Senczuk et al.
Dow
nloa
ded
by [
Uni
vers
ita S
tudi
la S
apie
nza]
at 0
7:19
14
May
201
4

Therefore, the population of A. nebulosus on DonPanchito could not have been separated before thisperiod.
Evaluation of the time of isolation by means of theavailable molecular data is difficult. The evolutionarytime is too short to apply a molecular clock to
estimate the age of the population since the singlefixed substitution characterizing the island popula-tion could generate a biased estimation. In fact, thissubstitution may have occurred and become fixed onthe island since islet formation up to the present dayor, alternatively, may have derived from an ancientcolonizing haplotype not yet sampled on themainland.The MMD analysis shows an L-shaped distribu-
tion on the island indicative of a very recent bottle-neck compared to a more ancient expansiondetected on the mainland.Some shared haplotypes could be expected
given the strong swimming abilities of manylizards and the short distance between the isletand the mainland. Yet none was detected.Instead, the shape of the MSN indicates a singlecolonization event.Together, these genetic data indicate a relatively
recent colonization of Don Panchito from the main-land, and an absence of current female gene flow.Genetic studies of small islet populations in otherlizard species showed a similar pattern with lowgenetic divergence between islet and mainland (e.g.Funk & Fa 2006; Terrasa et al. 2009).
Figure 4. Two specimens representing approximately the meansize for males in the two populations. Above: a male individualfrom mainland. Below: a male from Don Panchito island.
Figure 5. Principal component analysis (PCA) on log-transformed characters. Black circles: continental males; white circles: continentalfemales; black squares: insular males; white squares: insular females. At the top right, direction and length of the variable vectors areshown.
Insular gigantism in an anole lizard 7
Dow
nloa
ded
by [
Uni
vers
ita S
tudi
la S
apie
nza]
at 0
7:19
14
May
201
4

Size differences and ecological considerations
The present study confirms the previous extempora-neous observation that Anolis nebulosus individuals onDon Panchito island are larger than those of thefacing mainland (Figure 4) and we quantified thisdifference for the first time. Indeed, the size differ-ences between the islet and mainland populations ofA. nebulosus are remarkable, with little overlap in sizefor either males or females. Hence, the maximumsize and mass are higher in the island specimens thanin the mainland population. There is little informa-tion about the size of A. nebulosus from other main-land populations. In a population from Nayarit(Mexico), adult males and females averaged 42 mmand 36 mm respectively in SVL. The maximum sizewas 50 mm for males and 46 mm for females(Jenssen 1970). In females from Guayavas (Jalisco),SVL ranged from 38.8 to 42.1 mm (McDiarmid1963). Therefore, it is plausible that specimens ofA. nebulosus inhabiting Don Panchito are at theupper limit of size for this species.
The presence of insular gigantism, as found in A.nebulosus, seems uncommon in the genus Anolis.Comparable size differences have been reported onlyin A. marmoratus, albeit with smaller-sized popula-tions being found on islands (Roughgarden 1995).
Insular gigantism in vertebrates has been attribu-ted to different causes, including a light predationregime, high food availability and strong density-
dependent intraspecific competition (Meiri et al.2004; Dayan & Simberloff 2005; Meiri 2008). Incertain cases, a positive correlation has been demon-strated between the abundant presence of seabirdsand body sizes in island lizards (Sanchez-Piñero &Polis 2000; Pafilis et al. 2009).In light of this, the large size of the anoles on Don
Panchito could be explained as a combination ofseveral factors.First of all, predators of Anolis (snakes and small
carnivores) are virtually absent on the islet (Garcíaand Senczuk, pers. obs.). Moreover, the presence onthe island of breeding seabirds such as Sula leucoga-ster (Boddaert, 1783) and Pelecanus occidentalis(Linnaeus, 1766) (Senczuk, pers. obs.) may provideadditional protection from predators. On the main-land, A. nebulosus individuals have a very short effec-tive lifespan (Lister & García Aguayo 1992) and itcannot be ruled out that the low predation pressureon the island allows a longer life and thus larger size.The effect of predation on body size was suggested,for example, in Podarcis siculus klemmeri (Lanza &Capolongo, 1972) (Raia et al. 2010), where theabsence of specific predators could explain thelonger longevity of insular lizards, thus the measuredlarge size of individuals.Secondly, the presence of breeding seabirds may
provide large amounts of nutrients (fecal material,food scraps and carcasses), enhancing arthropod
Figure 6. Principal component analysis (PCA) on size-corrected characters. Black circles: continental males; white circles: continentalfemales; black squares: insular males; white squares: insular females. At the top right, direction and length of the variable vectors are shown.
8 G. Senczuk et al.
Dow
nloa
ded
by [
Uni
vers
ita S
tudi
la S
apie
nza]
at 0
7:19
14
May
201
4

densities. Therefore under this condition of foodavailability, the resident lizards community couldpositively increase body size (Raia & Meiri 2006).
Differences in sexual dimorphism
Anoles vary considerably in the extent of sexualdimorphism in body size (Schoener 1969; Stamps1983; Butler et al. 2000), ranging from species inwhich the sexes are the same size to others in whichadult males are more than three times the mass ofadult females. The sexes also differ in eco-morpho-logically relevant proportions of various parts of thebody, under the hypothesis that the habitat placesconstraints on morphology (Schoener 1967; Butler& Losos 2002; Butler et al. 2007). Moreover, withinspecies, males and females differ in the same mor-phological traits that distinguish the ecomorphs andfor which the ecological significance of morphologi-cal variation is well understood as a combination offunctional differences and sexual selection (Losos1990; Butler & Losos 2002).
Our morphometric analysis showed clear sexualdimorphism in almost all the absolute size traits,with males being larger than females. We alsofound that the extent of SD is greater on the islandthan on the mainland. In Anolis, sexual dimorphismin size diversifies faster in small-island lineages thanin mainland and large-island populations, mostprobably due to the high intraspecific resource com-petition and male-male competition (Thomas et al.2009). The male anoles on Don Panchito investapproximately 95% of their time in courting females(compared to 49% for mainland males), on averagedewlapping and headbobbing much more thanmainland males (Lister & García Aguayo 1992).
When we controlled for the body size effect, wefound longer relative trunk length in females than inmales, and higher average values in insular femalesthan in continental females. The longer trunk infemales is in accordance with studies carried out onsome Lacertidae species (Olsson et al. 2002;Schwarzkopf 2005; Kaliontzopoulou et al. 2007).This trait is related to a reproductive advantage infacilitating more space for eggs allocation and pro-moting clutch size (Cox et al. 2003). Thus, higheraverage values of the trunk length in insular femalescould explain the main differences detected in theextent of SD between island and mainland and itcould be correlated to an increased investment inreproduction.
Finally, we found larger head-size characters(HeaL, HeaH and IntD) in male lizards that couldbe advantageous in male-male competition, duringterritorial fighting, and enable males to successfully
grasp and hold a female during copulation (Gvozdik& Van Damme 2003). Moreover, a large head allowsmales to eat bigger and harder prey items comparedto females, thus reducing intersexual competition forfood (Schoener 1967; Stamps 1977; Preest 1994).
The Anole of Don Panchito, an example of “reversedisland syndrome”?
The RIS theory predicts a series of morphological,physiological and behavioural evolutionary changesthat were studied in detail in the Italian wall lizardPodarcis sicula klemmeri, living on a small islet inSouthern Italy (Raia et al. 2010; Monti et al.2013). The characteristics showed by this populationrespect to their mainland relatives include higheraggressivity, pronounced sexual dimorphism andphysiological correlates of increased investment inreproduction. These traits could be explained as anadaptation to a very small island with environmentalunpredictable conditions, and at a fluctuating popu-lation size. However, a recent meta-analysis on 641species of lizards found only weak support for awidespread occurrence of the RIS. The authors sus-pected that, if such a pattern is generalized, it is morelikely to manifest itself at intra-specific levels and onvery small islands (Novosolov et al. 2013).Some of the characteristics held by the island
anoles studied in this paper match the predictionsof the RIS. These are an increase in body size and insexual dimorphism in insular lizards. In addition,previous data indicate that the male anoles on DonPanchito invest more time in courting females com-pared to the mainland ones (Lister & García Aguayo1992). This is in accordance with the RIS scenariosince, in the presence of low predation pressure andin non-predictable environmental conditions, anamount of energy is moved from growth to repro-duction. As a consequence, sexual dimorphismincreases, with males becoming larger and femalessmaller (because of the very large amount of energymoved from growth to the clutch mass). Thus, underall the RIS conditions, the insular populationbecomes larger than the related one on the mainlandbecause of the increased sexual dimorphism (mainlyinvolving males) and the greater longevity in absenceof predators (involving both sexes).The climatic regime of the Chamela forest is char-
acterized by pronounced seasonal fluctuations andthis area has also large fluctuations in arthropodnumbers and biomass (Lister & García Aguayo1992). The mainland population of A. nebulosus inthe Chamela forest experiences high predation ratesand low annual survivorship. Seasonality and preda-tion turned out to be the major forces shaping
Insular gigantism in an anole lizard 9
Dow
nloa
ded
by [
Uni
vers
ita S
tudi
la S
apie
nza]
at 0
7:19
14
May
201
4

foraging strategies, activity levels and social beha-viour (Lister & García Aguayo 1992).
Even if there are still no comparable data on thepopulations of the anoles living in Don Panchito, it ispossible that the island population is seasonally sub-jected to high environmental stress. This condition,together with an hypothesised low predation pressureon the island, could determined the evolutionarychanges observed in Don Panchito, under the RIS.
Acknowledgements
We wish to thank Cuauhtemoc Hernando MorenoSalazar, Jesus Eduardo Olivan Pliego, Laura KarenRodríguez Lavín and Ramón López Castillo for theirvaluable help in the field, and Emma Mason,Emanuela Solano and Oscar Flores-Villela for criti-cally reading the manuscript. Thanks are extended totwo anonymous reviewers who greatly improved theprevious version of the manuscript with their com-ments on the evolution of island lizards. This workhas been partly supported by funds from “Progetti diRicerca di Università” to R.C.
References
Aljanabi SM, Martinez I. 1997. Universal and rapid salt-extractionof high quality genomic DNA for PCR-based techniques.Nucleic Acids Research 25:4692–4693. doi:10.1093/nar/25.22.4692.
Bullock SH. 1986. Climate of Chamela, Jalisco, and trends in thesouth coastal region of Mexico. Archives for Meteorology,Geophysics, and Bioclimatology Series B 36:297–316.doi:10.1007/BF02263135.
Butler MA, Losos JB. 2002. Multivariate sexual dimorphism,sexual selection, and adaptation in Greater Antillean Anolislizards. Ecological Monographs 72:541–559. doi:10.1890/0012-9615(2002)072[0541:MSDSSA]2.0.CO;2.
Butler MA, Sawyer SA, Losos JB. 2007. Sexual dimorphism andadaptive radiation in Anolis lizards. Nature 447:202–205.doi:10.1038/nature05774.
Butler MA, Schoener TW, Losos JB. 2000. The relationshipbetween sexual size dimorphism and habitat type in GreaterAntillean Anolis. Evolution 54:259–272.
Calsbeek R, Smith TB, Bardeleben C. 2007. Intraspecific varia-tion in Anolis sagrei mirrors the adaptive radiation of GreaterAntillean anoles. Biological Journal of the Linnean Society90:189–199. doi:10.1111/j.1095-8312.2007.00700.x.
Cox RM, Skelly SL, John-Alder HB. 2003. A comparative test ofadaptive hypotheses for sexual size dimorphism in lizards.Evolution 57:1653–1669.
Curray JR, Emmel FJ, Crampton PJS. 1969. Holocene history of astrand plain lagoonal coast, Nayarit, Mexico. In: LagunasCosteras, Un Simposio. Memorias del SimposioInternacional, Lagunas Costeras. UNAM-UNESCO, Nov.28–30, 1967, Mexico D.F. pp. 63–100.
Dayan T, Simberloff D. 2005. Ecological and community-widecharacter displacement: the next generation. Ecology Letters8:875–894. doi:10.1111/j.1461-0248.2005.00791.x.
Excoffier L, Lischer HEL. 2010. Arlequin suite ver 3.5: A newseries of programs to perform population genetics analysesunder Linux and Windows. Molecular Ecology Resources10:564–567. doi:10.1111/j.1755-0998.2010.02847.x.
Excoffier L, Schneider S. 1999. Why hunter–gatherer populationsdo not show signs of Pleistocene demographic expansions.Proceedings of the National Academy of Sciences 96:10597–10602. doi:10.1073/pnas.96.19.10597.
Funk SM, Fa JE. 2006. Phylogeography of the endemic St. Luciawhiptail lizard Cnemidophorus vanzoi: Conservation genetics atthe species boundary. Conservation Genetics 7:651–663.doi:10.1007/s10592-005-9068-7.
Gvozdík L, Van Damme R. 2003. Evolutionary maintenance ofsexual dimorphism in head size in the lizard Zootoca vivipara: atest of two hypotheses. Journal of Zoology 259:7–13.doi:10.1017/S0952836902003308.
Hammer Ø, Harper DAT, Ryan PD. 2001. PAST-Palaeontological Statistics Software Package for Educationand Data Analysis. Palaeontologia Electronica 4:9 pp.
Herrel A, Cottam MD, Godbeer K, Sanger T, Losos JB. 2011. Anecomorphological analysis of native and introduced popula-tions of the endemic lizard Anolis maynardi of the CaymanIslands. Breviora 522:1–10. doi:10.3099/0006-9698-522.1.1.
Herrel A, Huyghe K, Vanhooydonck B, Backeljau T, BreugelmansK, Grbac I, Van Damme R, Irschick DJ. 2008. Rapid large-scale evolutionary divergence in morphology and performanceassociated with exploitation of a different dietary resource.Proceedings of the National Academy of Sciences USA105:4792–4795.
Ihaka R, Gentleman R. 1996. R: A language for data analysis andgraphics. Journal of Computational and Graphical Statistics5:299–314.
Jenssen TA. 1970. The Ethoecology of Anolis nebulosus (Sauria,Iguanidae). Journal of Herpetology 4:1–38. doi:10.2307/1562700.
Kaliontzopoulou A, Carretero MA, Llorente GA. 2007.Multivariate and geometric morphometrics in the analysis ofsexual dimorphism variation in Podarcis lizards. Journal ofMorphology 268:152–165. doi:10.1002/jmor.10494.
Kolbe JJ, Losos JB. 2005. Hind-limb length plasticity in Anolis caroli-nensis. Journal ofHerpetology 39:674–678. doi:10.1670/87-05N.1.
Librado P, Rozas J. 2009. DnaSP v5: A software for comprehen-sive analysis of DNA polymorphism data. Bioinformatics25:1451–1452. doi:10.1093/bioinformatics/btp187.
Lister BC, García Aguayo A. 1992. Seasonality, predation, andthe behaviour of a tropical mainland anole. The Journal ofAnimal Ecology 61:717–733. doi:10.2307/5626.
Losos JB. 1990. A phylogenetic analysis of character displacementin Caribbean Anolis lizards. Evolution 44:558–569.doi:10.2307/2409435.
Losos JB. 2009. Lizard in an evolutionary tree: Ecology andadaptive radiation of Anoles. Oakland, CA: University ofCalifornia press.
Losos JB, Creer DA, Glossip D, Goellner R, Hampton A, RobertsG, Haskell N, Taylor P, Etling J. 2000. Evolutionary implica-tions of phenotypic plasticity in the hindlimb of the lizardAnolis sagrei. Evolution 54:301–305.
Losos JB, Jackman TR, Larson A, de Queiroz K, Rodríguez-Schettino L. 1998. Contingency and determinism in replicatedadaptive radiations of island lizards. Science 279:2115–2118.doi:10.1126/science.279.5359.2115.
Losos JB, Ricklefs RE. 2009. Adaptation and diversification onislands. Nature 457:830–836. doi:10.1038/nature07893.
McDiarmid RW. 1963. A collection of reptiles and amphibiansfrom the highland faunal assemblage of western Mexico.Contributions in Science 68:1–15.
10 G. Senczuk et al.
Dow
nloa
ded
by [
Uni
vers
ita S
tudi
la S
apie
nza]
at 0
7:19
14
May
201
4

Meiri S. 2007. Size evolution in island lizards. Global Ecology andBiogeography 16:702–708. doi:10.1111/j.1466-8238.2007.00327.x.
Meiri S. 2008. Evolution and ecology of lizard body sizes. GlobalEcology and Biogeography 17:724–734. doi:10.1111/j.1466-8238.2008.00414.x.
Meiri S, Cooper N, Purvis A. 2008. The island rule: Made to bebroken? Proceedings of the Royal Society B: BiologicalSciences 275:141–148.
Meiri S, Dayan T, Simberloff D. 2004. Body size of insularcarnivores: little support for the island rule. The AmericanNaturalist 163:469–479. doi:10.1086/382229.
Monti DM, Raia P, Vroonen J, Maselli V, Van Damme R,Fulgione D. 2013. Physiological change in an insular lizardpopulation confirms the reversed island syndrome. BiologicalJournal of the Linnean Society 108:144–150. doi:10.1111/j.1095-8312.2012.02019.x.
Nicholson KE, Crother BI, Guyer C, Savage MJ. 2012. It is timefor a new classification of anoles (Squamata: Dactyloidae).Zootaxa 3477:1–108.
Novosolov M, Raia P, Meiri S. 2013. The island syndrome inlizards. Global Ecology and Biogeography 22:184–191.doi:10.1111/j.1466-8238.2012.00791.x.
Olsson M, Shine R, Wapstra E, Ujvari B, Madsen T. 2002. Sexualdimorphism in lizard body shape: The roles of sexual selectionand fecundity selection. Evolution 56:1538–1542.
Pafilis P, Meiri S, Foufopoulos J, Valakos E. 2009. Intraspecificcompetition and high food availability are associated with insu-lar gigantism in a lizard. Naturwissenschaften 96:1107–1113.doi:10.1007/s00114-009-0564-3.
Poe S. 2013. 1986 Redux: New genera of anoles (Squamata:Dactyloidae) are unwarranted. Zootaxa 3626:295–299.doi:10.11646/zootaxa.3626.2.7.
Preest MR. 1994. Sexual size dimorphism and feeding energetics inAnolis carolinensis: Why do females take smaller prey than males?.Journal of Herpetology 28:292–298. doi:10.2307/1564527.
Raia P, Guarino FM, TuranoM, Polese G, Rippa D, Carotenuto F,MontiDM,CardiM,FulgioneD. 2010.The blue lizard spandreland the island syndrome. BMC Evolutionary Biology. 10:289.
Raia P, Meiri S. 2006. The island rule in large mammals:Paleontology meets ecology. Evolution 60:1731–1742.
Ramı́rez-Herrera MT, Kostoglodovb V, Urrutia-Fucugauchi J.2004. Holocene-emerged notches and tectonic uplift alongthe Jalisco coast, Southwest Mexico. Geomorphology58:291–304. doi:10.1016/j.geomorph.2003.07.004.
Rodrıguez-Schettino L. 1999. The iguanid lizards of Cuba.Gainesville, FL: The University of Florida Press.
Rogers AR. 1995. Genetic evidence for a Pleistocene populationexplosion. Evolution 49:608–615. doi:10.2307/2410314.
Rogers AR, Harpending H. 1992. Population growth makes wavesin the distribution of pairwise genetic differences. MolecularBiology and Evolution 9:552–569.
Roughgarden J. 1995. Anolis lizards of the Caribbean: Ecology,evolution, and plate tectonics. Oxford: Oxford UniversityPress.
Sanchez-Piñero F, Polis GA. 2000. Bottom-up dynamics ofallochthonous input: Direct and indirect effects of seabirdson islands. Ecology 81:3117–3132.
Schoener TW. 1967. The ecological significance of sexualdimorphism in size in the lizard Anolis conspersus. Science155:474–477. doi:10.1126/science.155.3761.474.
Schoener TW. 1969. Size patterns in West Indian Anolis lizards: I.Size and species diversity. Systematic Zoology 18:386–401.doi:10.2307/2412183.
Schwartz A, Henderson RW. 1991. Amphibians and reptiles of theWest Indies: Descriptions, distributions, and natural history.Gainesville, FL: University of Florida Press.
Schwarzkopf L. 2005. Sexual dimorphism in body shape withoutsexual dimorphism in body size in water skinks (Eulamprusquoyii). Herpetologica 61:116–123. doi:10.1655/04-66.
SEMARNAT. 2002. DECRETO por el que se declara área naturalprotegida con la categoría de santuario a las islas La Pajarera,Cocinas, Mamut, Colorada, San Pedro, San Agustín, SanAndrés y Negrita, y los islotes Los Anegados, Novillas, Mosca ySubmarino, situadas en la Bahía de Chamela, frente a las costasdelMunicipio deLaHuerta, Estado de Jalisco, con una superficietotal de 1,981-43-93.200 hectáreas. Diario Oficial de laFederacion. 13 de junio del 2002.
Stamps JA. 1977. The Relationship between ResourceCompetition, Risk, and Aggression in a Tropical TerritorialLizard. Ecology 58:349–358. doi:10.2307/1935609.
Stamps JA. 1983. The relationship between ontogenetic habitatshifts, competition and predator avoidance in a juvenile lizard(Anolis aeneus). Behavioral Ecology and Sociobiology 12:19–33. doi:10.1007/BF00296929.
Terrasa B, Pérez-Mellado V, Brown RP, Picornell A, Castro JA,Ramon MM. 2009. Foundations for conservation of intraspe-cific genetic diversity revealed by analysis of phylogeographicalstructure in the endangered endemic lizard Podarcis lilfordi.Diversity and Distributions 15:207–221. doi:10.1111/j.1472-4642.2008.00520.x.
Thomas GH, Meiri S, Phillimore AB. 2009. Body size diversifica-tion in Anolis: novel environment and island effects. Evolution63:2017–2030. doi:10.1111/j.1558-5646.2009.00694.x.
Thompson JD, Gibson TJ, Plewniak JM, Jeanmougin F, HigginsDG. 1997. The clustal_X windows interface: flexible strategiesfor multiple sequence alignment aided by quality analysis tools.Nucleic Acids Research 25:4876–4882. doi:10.1093/nar/25.24.4876.
Van Valen LM. 1973. A new evolutionary law. EvolutionaryTheory 1:1–30.
Williams EE. 1972. The origin of faunas. Evolution of lizardcongeners in a complex island fauna: A trial analysis.Evolutionary Biology 6:47–89.
Insular gigantism in an anole lizard 11
Dow
nloa
ded
by [
Uni
vers
ita S
tudi
la S
apie
nza]
at 0
7:19
14
May
201
4
Copyright © 2022 FDOKUMEN




















