
Effects of dredging on benthic diatom assemblages in a lowland stream

Structural heterogeneity and tree spatial patterns in an old-growth deciduous
lowland forest in Cantabria, northern Spain
Vicente Rozas*Departamento de Ecologıa, Centro de Investigaciones Forestales y Ambientales de Lourizan, Apartado 127,36080, Pontevedra, Spain; *Author for correspondence (e-mail: [email protected]; phone:+34-986-805005; fax: +34-986-856420)
Received 11 March 2005; accepted in revised form 28 November 2005
Key words: Fagus sylvatica, Ilex aquifolium, Quercus robur, Ripley’s K-function, Spatial autocorrelation,Temperate deciduous forest
Abstract
Old-growth deciduous forests in western Europe, for the most part, consist of small tracts that often may beatypical due to human disturbance, poor soil productivity or inaccessibility. In addition, very little infor-mation on tree age distributions, structural heterogeneity and tree spatial patterns appears to be availablefor west-European forests. Characterization of the structural features of tree populations in these old-growth stands can provide the basis to design conservation plans and also inform on how present forestsmight look in the absence of human interference. Four old-growth stands in a deciduous forest in theCantabrian lowlands, northern Spain, were surveyed to determine forest structure and spatial patterns.Live and dead trees were identified, measured and mapped, and live trees were cored for age estimation.Structural heterogeneity was analyzed by means of the spatial autocorrelation of tree diameter, height andage, and the uni- and bivariate spatial patterns of trees were analyzed. The dominant species, Fagussylvatica and Quercus robur, showed reverse-J shaped size distributions but discontinuous age distribu-tions, with maximum ages of 255 –270 yr. Tree ages suggested that the forest was largely modified by pastchanges in forest-use, especially by temporal variation in grazing intensity. Spatial autocorrelation revealedthat former parkland stands were heterogeneous with respect to tree height only, while high forest standswere composed of patches of even-aged and even-sized trees. Young trees were clumped at varying dis-tances and establishment occurred preferentially in canopy gaps, except for Ilex aquifolium that mainlyoccurred beneath mature Quercus trees. Surviving trees became less intensely clumped in the dominantspecies, and more strongly clumped in understorey ones, which may have been due to the effects ofintraspecific competition and of canopy trees on tree survival, respectively. The spatial associations betweenspecies varied within the forest, probably as a consequence of specific establishment preferences andcompetitive interactions.
Introduction
The structure of a forest stand refers to the relativeabundance and spatial dispersion of trees in termsof size and age (Veblen 1992). A general definition
of old-growth for temperate forests includes arelatively high degree of patchiness and heteroge-neity, dead trees and logs, relatively old age, re-verse-J shaped size distribution and multicohortage distribution, but not necessarily without any
Plant Ecology (2006) 185:57 –72 � Springer 2005
DOI 10.1007/s11258-005-9084-1

evidence of human activity (Foster et al. 1996;Oliver and Larson 1996; Peterken 1996). Consid-erable effort has been directed toward character-izing the structure and spatial patterns oftemperate old-growth forests (e.g., Williamson1975; Turner and Franz 1985; Manabe et al. 2000;Antos and Parish 2002; Miyadokoro et al. 2003).However, there is a notable lack of comparativeinformation for west-European deciduous forests,where very little information on age structures andtree spatial distribution appears to be available.
In northern Spain, as in other European coun-tries, widespread deforestation for land reclama-tion, for pasture, and for exotic tree plantation,make well-preserved, old-growth forests very rare.These forests, for the most part, consist of smalltracts that often are atypical due to anthropogenicdisturbance, poor soil productivity or inaccessi-bility. Nevertheless, characterization of the com-position and structural characteristics of treepopulations in these old-growth stands can pro-vide the basis to design conservation plans, andalso provide information on how present forestsmight look in the absence of significant humaninterference (Foster et al. 1996; Chokkalingamand White 2001).
Studies of some old-growth forests show that alarge variety of processes can give rise to foreststructure. It is commonly accepted that old-growthtemperate forests are largely structured by distur-bances (Duncan and Stewart 1991; Oliver andLarson 1996). Major disturbances can initiate newforest stands, but only small-scale disturbances areevident in a near-steady state (Lusk and Smith 1998;Antos and Parish 2002). The spatial patchinesscreated by local disturbances, at the scale of indi-vidual or small groups of trees, causes small-scaleheterogeneity in seedling recruitment, stand struc-ture and spatial associations within and betweentree species (Frelich et al. 1993; Park et al. 2005).
In dense temperate forests, the ecological pro-cesses of seed-dispersal, seedling establishment andsurvival are influenced by the change in conditions– light environment, soil parameters, behavior ofhoarding animals and herbivores – from beneathclosed canopy to the center of canopy gaps. Patchydisturbances and gap-phase regeneration create avariety of favorable microsites that might result inclumped spatial patterns of juveniles and negativeassociations with adults (Leemans 1991; Ku-uluvainen and Rouvinen 2000; Arevalo and
Fernandez-Palacios 2003). The presence of differ-ent forest floor microsites, topographic positions,ground cover and overstorey types, also tend tocreate clumped distributions (Duncan 1993;Manabe and Yamamoto 1997).
Analyses of the reconstructed spatial patterningof live and dead trees (Kenkel 1988; Duncan 1991;He and Duncan 2000) and long-term studies ontree spatial dynamics (Ward et al. 1996; Kenkelet al. 1997; Aldrich et al. 2003) have shown theimportance of density-dependent processes andsuccessional status in determining spatial shiftsthrough time. Inter-tree competition in the stemexclusion stage, and the accumulated effects ofwinds, pathogens, droughts, defoliating outbreaksand other diseases in the old-growth stage, causethe death of trees (Oliver and Larson 1996). Lossof trees usually leads to a weakly aggregated,random or uniform distribution (Stewart and Rose1990; Ward et al. 1996). Moreover, ingrowth ofnew trees of shade-tolerant, late-successional spe-cies can cause either less-clumped spatial distri-butions in the absence of significant exogenousdisturbances (e.g., McDonald et al. 2003) or morestrongly clumped distributions in semi-natural,formerly managed forests (e.g., Wolf 2005).
I studied the structure and tree spatial patterns inan old-growth deciduous forest in the Cantabrianlowlands, northern Spain. I conducted this researchto report onheterogeneity in structural traits, and toevaluate the idea that tree populations start offclumped andbecomeprogressivelymore uniformasthe forest ages. The following hypotheses were tes-ted: (1) tree species display reverse-J shaped sizedistributions and all-aged populations, (2) foreststands are structurally heterogeneous – i.e. trees ofsimilar size or age will tend to occur together withineven-sized or even-aged patches –, (3) tree speciespartitioned establishment sites with respect tooverstorey conditions, (4) trees are clumped andbecome more regularly distributed with time, and(5) the different tree species and age-classes arenegatively associated.
Material and methods
Study site
The study site is called the Caviedes forest, a110 ha broadleaved remnant that belongs to the
58

Oyambre Natural Park in the western lowlands ofCantabria, northern Spain (43�20¢ N, 04�18¢ W).It is part of the 2000 ha Corona Forest, now lar-gely composed of plantations of exotic species,principally Eucalyptus globulus and Pinus radiata.The forest is located on a gentle north-east ori-ented slope, with altitudes ranging from 40 to240 m. The dominant soil type is deep sandybrown earth, with bedrock of Cretaceous sand-stone and clay (Guitian et al. 1985). The climate isAtlantic, with temperate, wet winters and periodsof summer drought in occasional years (Allue1990). Annual precipitation averages 1200 mm,and mean annual temperature is 14 �C.
Fagus sylvatica and Quercus robur (Fagaceae)are the tallest canopy species. Forest understoreyis composed of Corylus avellana (Corylaceae),Crataegus monogyna, Pyrus cordata (Rosaceae),Frangula alnus (Rhamnaceae), Ilex aquifolium(Aquifoliaceae), and Salix atrocinerea (Salicaceae).Botanical nomenclature follows Tutin et al. (1964 –1980). In the text only genus names of the tree speciesare used. Characteristics of old-growth, such asstanding and fallen dead trees and logs, woody deb-ris, uprootedand snapped trees, soilmound-and-pithtopography, large hollow trees and canopy gaps,were encountered throughout the forest. The studiedforest represents the largest and best-preservedremnant with old-growth attributes in the Canta-brian lowlands. As in other European forest reserves(e.g., Koop and Hilgen 1987; Peterken 1996; Mottaet al. 2002), high forest stands, along with formeropen parklands used as pasture by cattle, in whichtrees have abundantly recruited, coexist in Caviedes.There is no record of tree cutting since 1951, but thepresence of cut stumps and charred wood indicateintense prior human use. A detailed analysis of dis-turbance and forest-use histories in Caviedes hasbeen carried out (Rozas 2003a).
Field procedures
I selected four stands as being representative of thevariability of forest composition and structure.Stands P1 (50 m�50 m) and P2 (50 m�50 m)have a structure corresponding to former park-land, whereas stands H1 (50 m�60 m) and H2(50 m�110 m) have a high forest structure. Totalstand densities range between 940 and 1169 livetrees per ha, and total basal areas range between28.9 and 39.7 m2 per ha (Rozas 2003a). Fagus and
Ilex show the highest densities of live trees in allstands, while Fagus and Quercus display the largestbasal areas. All live and dead trees (DBH ‡ 5 cm,measured at 1.3 m above ground) and saplings(2 cm £ DBH<5 cm) were identified and labeled.DBH and the height of live saplings and trees wereregistered. The x and y coordinates to the centre ofeach labeled stem were measured to the nearest0.1 m within 10 m�10 m quadrats.
To estimate tree ages, I cored all live Quercusand Fagus trees, and a random sample of morethan 50% of the live Ilex, Frangula, Pyrus andSalix trees. No age data were obtained for Corylus,as they always have several stems, nor for Cra-taegus, as only a few individuals were found in asingle stand. Cores were taken with incrementborers at 20 cm above ground in trees withDBH<30 cm, and 100 cm when DBH ‡ 30 cm.Because of the effort involved in detailed survey-ing, mapping, coring, and core processing, thestudy was confined to two former parkland standsand two high forest stands.
Laboratory procedures
Cores were air dried, glued onto wooden mountsand surfaced with successively finer grades ofsandpaper, until the growth rings were clearlyvisible. I dated and measured tree-ring series ofFagus and Quercus, and the dates were verifiedagainst master site chronologies (Rozas 2003a). Inother species, the number of rings on the cores wascalculated by ring counting. Methods used toestimate the number of rings missing toward thepith, and to correct for the time to grow to sam-pling height, have been previously described (Ro-zas 2003a, b). To estimate the ages of uncored andhollow trees, I used age-diameter regressionequations calibrated from cored trees (Stewart andRose 1990; Taylor et al. 1996). In Fagus andQuercus, age –diameter equations of two differentage-classes were used because these producedbetter results than equations of the population as awhole (Rozas 2003b).
Structure analysis
DBH distributions of 5-cm classes were quanti-fied by using the power, negative exponential and
59

Weibull functions, which reliably characterize well-balanced, reverse-J shaped distributions (Veblen1992; Oliver and Larson 1996). The Weibull func-tion can assume a wide variety of shapes, which areregulated by the shape parameter c. At values of c<1,the function is a descending monotonic curve, whilefor values of c>1 the curve is unimodal (Lorimer andKrug 1983). Age and height structures of tree popu-lations were examined on height –age scatter plots.
I studied the spatial structure of tree sizes and ageswith the Moran’s I coefficient. This coefficient is ameasure of the autocorrelation of the values of aquantitative variable for all pairs of points separatedby a given spatial lag (Legendre and Fortin 1989).When applied to trees, the spatial autocorrelationcan reveal scales of heterogeneity of the foreststructure related to the size and distribution of even-sized or even-aged tree patches (Duncan and Stewart1991; Frelich et al. 1993; Kuuluvainen et al. 1998).The hypothesis of spatial independence of tree sizesandages in a stand,was testedon correlogramsof thestandard normal deviates (SND) of I coefficientscalculated by 5-m distance classes, up to inter-treedistances of 50 m, against the critical values for astandard normal distribution. SND values above1.96 indicate significant positive autocorrelation ata=0.05, values below )1.96 indicate negative auto-correlation. SinceSNDvalueswere calculated for tendistance classes in each correlogram, a correlogramwas considered statistically significant only if at leastone value was outside the )2.81 to 2.81 range,corresponding to the Bonferroni-corrected signifi-cance level a0=0.005 (Legendre and Fortin 1989). Adetailed description of the ways of computing andtesting Moran’s I coefficients can be found inDuncan and Stewart (1991). Correlograms werecomputed forDBH, tree height and age.All trees in astand were taken together in the analyses.
Spatial arrangement analysis
To investigate the degree of association of juvenileswith different overstorey conditions, I calculated themean number of juveniles (saplings+young trees)foundwithin 5 mof oldQuercus orFagus trees, deadQuercus or Fagus canopy trees, and former opensites. Young trees are those younger than 80 yr, oldtrees are those older than 150 yr. Dead trees withDBH>20 cm were considered to be former canopytrees. Former open sites were defined as points
greater than 10 m from any old live or dead tree.Focal trees or points closer than 5 m to the neareststand boundary were not included in the analyses.
To analyze spatial patterns, I used the univariateRipley’s K function (Ripley 1977), which reflects thetype and intensity of pattern at different inter-treedistances d.K (d)was calculated at 1-m intervals, up toa maximum distance of 25 m. The 95% confidenceenvelopes for testing spatial randomness were gener-ated from 100 Monte Carlo iterations of randomprocesses. Details of the calculation of K(d), and thecorrection of the edge effect, are available in Moeur(1993) and Dixon (2002). I tested the hypothesis thatjuveniles (saplings+young trees) and old trees tend tobe more regularly spaced, by analyzing the univariatespatial patterns for both pre-mortality (live+deadstems) and post-mortality (live stems only) data sets(Kenkel 1988; He and Duncan 2000). The transfor-mation L(d)=(K(d)/p)0.5 linearizes the function andstabilizes its variance, and L(d) –d has an expectedvalue of zero under the assumption of spatial ran-domness. The results of pre- and post-mortality spa-tial patterns were plotted as L(d) –d against d.Significant positive and negative L(d) –d values indi-cated clumping and regularity, respectively.
The bivariate extension of Ripley’s K function(Lotwick and Silverman 1982) was used to analyzespatial associations between young trees of differentspecies in the pre-mortality populations, and alsobetween young and old trees. For details on the cal-culation and testing of K12(d) see Moeur (1993) andDixon (2002). As with the univariate function, thetransformation L12(d) = (K12(d)/p)
0.5 linearizes thefunction and stabilizes its variance, andL12(d) –d hasan expected value of zero under the assumption ofspatial independence between two sets of trees. The95% confidence envelopes for testing independencewere generated from 100 random toroidal shifts ofone set of trees with respect to the other (Dixon2002). Significant positive and negative L12(d) –dvalues indicated positive and negative association,i.e. spatial attraction and repulsion, respectively.
Results
Populations structure
The maximum diameter measured was ca. 80 cmfor Fagus and 130 cm for Quercus, whereas
60

non-dominant species were restricted to the smal-ler diameter classes (Figure 1). Diameter distribu-tions of Fagus and Quercus populations roughlyshowed a descending reverse-J shaped pattern,which displayed the best fits to the exponential,power or Weibull functions depending on thestand (r2 ranged from 0.52 to 0.90, and the cparameter in Weibull functions ranged from 0.17to 0.79; Table 1). The exception was Quercusin stand P2, which showed a flat diameterdistribution. Fagus in stands H1 and H2 showeda compound diameter distribution, which wasdescending in the smaller diameter classes, and hada broad peak in the larger ones (Figure 1). TheWeibull function did not show a convergent solu-tion when it was fit to the compound distribution(Table 1). Maximum diameters for the other
species were 17.6 cm in Ilex, 7.2 cm in Corylus,9.7 cm in Crataegus, 9.4 cm in Frangula, 43.5 cmin Pyrus, and 27.4 cm in Salix.
Fagus and Quercus reached heights of ca. 30 m(Figure 2). Ilex reached at most 12 m, and formeda sub-canopy that usually did not exceed 8 m.Maximum heights of the other species were 8.1 min Corylus, 7.5 m in Crataegus, 10.7 m in Frangula,15.9 m in Pyrus, and 16.4 m in Salix. Age –diameterregressions for the different species and age classeswere all significant (Table 2). The obtainedregression equations, which were used to estimateage of uncored trees, were of lineal, lineal-through-the-origin and power types. The Fagus and Quercuspopulations consisted of two age groups (Figure 2).The younger group ranged from 25 to 104 yr forFagus, and from 20 to 64 yr for Quercus; the older
H2
H1
P2
P1
Fagus
0
20
40
60
Quercus Ilex Other
0
20
40
60
0
20
40
60
Fre
quen
cy (
%)
0 20 40 60 80
0
20
40
60
0 20 40 60 80 100 120
Diameter (cm)
0 20 40 0 20 40
Figure 1. Diameter frequency distribution in stands P1, P2, H1 and H2. Other species include Corylus, Crataegus, Frangula, Pyrus and
Salix.
61

group ranged from 150 to 255 yr for Fagus, andfrom 156 to 269 yr for Quercus. Ages in Ilex rangedbetween 19 and 112 yr, and in the other species
from 12 to 36 yr in Frangula, 17 –65 yr in Pyrus,and 11–59 yr in Salix.
Spatial autocorrelation
In stands P1 and P2, correlograms of tree heightwere significant (p<0.001). These correlogramsshowed positive autocorrelation at inter-tree dis-tances less than 10 –15 m, which correspondedto the distances between trees of similar heightwithin a patch, and negative between 20 and40 m, which revealed the distances between pat-ches of dissimilar height (Figure 3). However,correlograms for DBH and tree age were notsignificant in stands P1 and P2. In stands H1 andH2, all the calculated correlograms were statistically
Table 1. R-square coefficients for the power, exponential, and
Weibull models applied to dbh frequency distributions of Fagus
and Quercus. The shape parameter value c in the Weibull
equations is shown.
Species Stand Power Exponential Weibull c
Fagus P1 0.72** 0.60** 0.90*** 0.26
P2 0.75*** 0.83*** 0.65*** 0.43
H1 0.85*** 0.77***
H2 0.85*** 0.66***
Quercus P1 0.36* 0.53** 0.11 0.79
H1 0.52* 0.35 0.49* 0.55
H2 0.62*** 0.36* 0.86*** 0.17
*p<0.05, **p<0.01, ***p<0.001.
Fagus
0
10
20
30Quercus Ilex Other
P1
0
10
20
30
P2
0
10
20
30
Hei
ght (
m)
H1
0 100 200 300
0
10
20
30
0 100 200 300
Age (yr)
0 100 200 0 100 200
H2
Figure 2. Height age scatter-diagrams in stands P1, P2, H1 and H2. Other species include Frangula, Pyrus and Salix.
62

significant (p<0.001). Positive autocorrelation atdistances less than 5–10 m indicated association ofsimilarly sized and aged trees within a patch (Figure3). Negative autocorrelation at intermediate (20 –35 m) or large (40–50 m) spatial scale correspondedto distances between patches of dissimilarly sizedand aged trees.
Space partitioning
Fagus juveniles were abundant in the proximity ofdead Quercus trees, and also in open sites, whilethey were scarce near old Fagus trees (H=18.92,
Table 2. Regression equations for estimating tree age (y, yr) from
stem diameter (x, cm) for different tree species and age ranges (yr).
n and %: number of cored trees and percentage of the whole sample
of live trees, respectively, used to calculate each equation.
Species Age
range
n % Equation r2 p
Fagus 25 –104 379 99.2 y=31.971 x0.212 0.325 <0.001
150 –255 81 92.0 y=117.014 x0.163 0.161 <0.001
Frangula 12 –32 17 54.8 y=3.787 x 0.867 <0.001
Ilex 19 –70 51 51.0 y=6.568 x 0.848 <0.001
Pyrus 17 –59 16 69.5 y=1.795 x 0.847 <0.001
Quercus 20 –64 87 100.0 y=17.072 x0.344 0.622 <0.001
156 –236 29 85.3 y=148.908
+0.934 x0.186 0.019
Salix 11 –52 29 52.7 y=2.380 x 0.924 <0.001
H1
P1
DBH AgeHeight
-10
-5
0
5
10
P2
-10
-5
0
5
10
-10
-5
0
5
10SN
D
0 10 20 30 40 50-10
-5
0
5
10
0 10 20 30 40 50
Distance (m)
0 10 20 30 40 50
H2
Figure 3. Correlograms for DBH, height and age of all trees in stands P1, P2, H1 and H2. SND values outside the shaded area indicate
significant autocorrelation (p<0.05), positive for values above 1.96, negative for values below )1.96.
63

p<0.001, Kruskal –Wallis test; Table 3). Quercuspreferentially occupied former open sites, whichshowed a significantly higher number of stems(H=87.19, p<0.001). The mean number of stemsindicated that Ilex showed a preference for oldQuercus trees (H=23.60, p<0.001), while the otherunderstorey species were found in greater numbers
in open sites, followed by canopy gaps caused bydead Quercus trees (H=89.96, p<0.001).
Spatial patterns
Fagus juveniles were significantly clumped, andthe scale of clumping did not vary greatly as a
Table 3. Mean number (1S.E.) of juveniles found within 5 m from old trees (>150 yr), large dead trees (>20 cm DBH), and open
sites. Values in a row followed by the same letter are not significantly different at p=0.001 according to Kruskal –Wallis test. n=total
sample number of target points.
Old Fagus Old Quercus Dead Fagus Dead Quercus Open sites
n=69 n=29 n=35 n=19 n=55
Fagus 2.00 (0.36) a 3.59 (0.61) ab 3.29 (0.77) ab 6.16 (1.49) b 4.24 (0.62) b
Quercus 0.03 (0.02) a 0.00 (0.00) a 0.51 (0.12) a 1.32 (0.83) ab 2.31 (0.37) b
Ilex 0.99 (0.19) a 3.97 (1.09) b 0.43 (0.14) a 0.79 (0.32) a 0.76 (0.16) a
Other 0.14 (0.05) a 0.93 (0.32) ab 0.40 (0.11) a 1.58 (0.64) b 2.91 (0.33) c
-4-202468
10
P2
-2
-1
0
1
2
P1
Other
-2
0
2
4
6
8
H2
-4
-2
0
2
4
H1
Fagus IlexQuercus
-1
0
1
2
3
4
-2-101234
-6-4-202468
-1
0
1
2
-2-1012345
-1012345L(
d)-d
-4
-2
0
2
4
-2
0
2
4
6
8
0 5 10 15 20 25-1
0
1
2
3
0 5 10 15 20 25
Distance (m)
-202468
101214
0 5 10 15 20 25-2
0
2
4
6
0 5 10 15 20 25
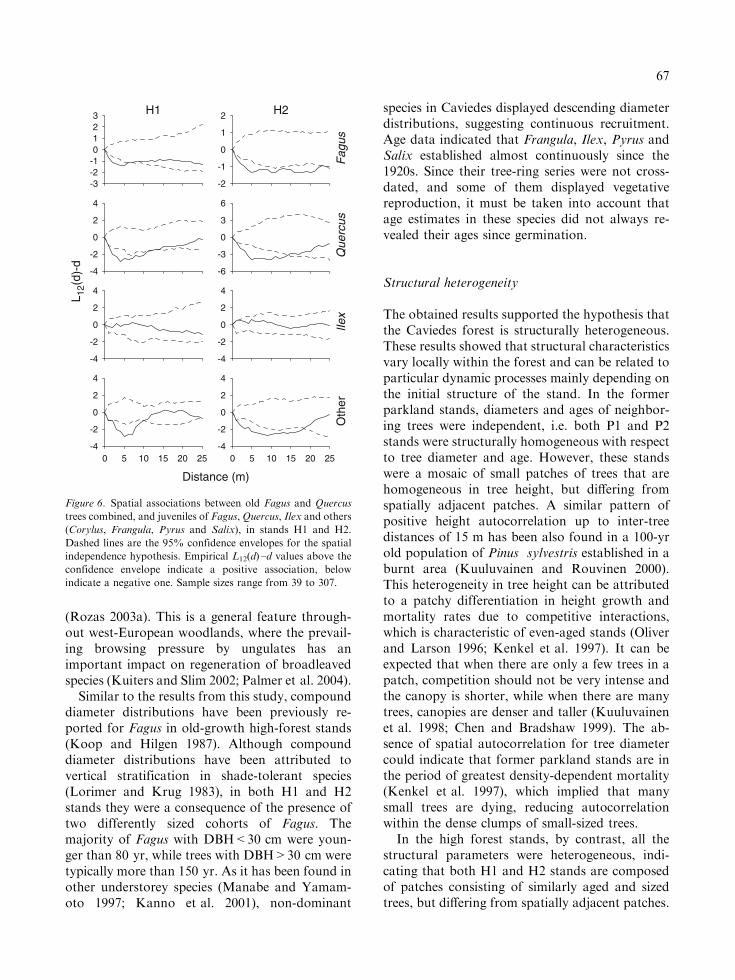
Figure 4. Spatial point patterns for juveniles in stands P1, P2, H1 and H2. Dark and thin continuous lines are the empirical L(d) –d
values for live+dead and live stems, respectively. Long and short dashed lines are respectively the pre- and post-mortality 95%
confidence envelopes for the spatial randomness hypothesis. Empirical values above the confidence envelope indicate clustering, below
indicate regularity. Sample sizes range from 19 to 307. Other species include Corylus, Crataegus, Frangula, Pyrus and Salix.
64

consequence of mortality (Figure 4). SurvivingFagus and Quercus juveniles were less intenselyclumped than live+dead juveniles. In stands P1and H1, the scale of clumping in Quercus juvenilesdiffered when dead trees were included, while instand H2 they were clumped from 1 to 25 m.Quercus in stand P2 were too few to analyze forspatial pattern. Only live Ilex were analyzed be-cause few dead individuals were found. Ilex stemsin stand P1 were clumped from 1 to 18 m andshowed a regular pattern from 21 to 25 m, while inthe other stands Ilex stems were significantlyclumped at almost all distances: 1 –23 m in P2, 3 –25 m in H1 and 2 –25 m in H2 (Figure 4). Ingeneral, mortality of the other understorey speciesincreased the intensity of clumping, and preservedor extended the scale of clumping (from 1 –4 m to1 –13 m in P1, from 4 –16 m to 3 –25 m in P2, and2 –25 m for both pre- and post-mortality stems inH2). The exception was stand H1, in which pre-mortality stems were clumped from 1 to 9 m, whilesurvivors showed a random pattern from 1 to25 m (Figure 4). This exception could be a con-sequence of the smaller sample size of understoreyspecies in this stand.
In old Fagus and Quercus trees combined, theintensity and scale of clumping significantly de-creased as a consequence of mortality (Figure 5).In stand H1, pre-mortality trees were significantlyclumped from 4 to 13 m, while the survivorsshowed a significantly regular pattern at 3 m only.In stand H2, pre-mortality stems showed clumpingfrom 1 to 18 m, and the post-mortality data setwas clumped only at 1 m.
Spatial associations
Spatial associations between juveniles variedamong the stands (Table 4). In stand P1, Fagusand Quercus showed positive association at 2 –6 m. In stands H1 and H2, Fagus and Quercuswere always spatially independent. Fagus and Ilexwere spatially independent in stands P1 and P2. Bycontrast, Fagus and Ilex showed positive associa-tion from 1 to 21 m in stand H1, and positiveassociation at 1 m and negative at 17 –25 m, instand H2 (Table 4). Fagus and other species(Corylus, Crataegus, Frangula, Pyrus and Salix)showed negative association at 3 m and 14 –15 min stand P1, and showed also negative association
in stand H1 at 11 –25 m. However, Fagus andother species were always independent in stands P2and H2. Quercus and Ilex were spatially indepen-dent in stands P1 and H2, but in stand H1 showednegative association at 4 –9 m (Table 4). In standP1, Quercus and other species were negativelyassociated at 11 –14 m. By contrast, Quercus andothers were positively associated in stand H1 at 2 –7 m, and in stand H2 at 3 –25 m. Finally, Ilex andother species were spatially independent in standsP1 and H2, but negatively associated at varyingdistances in stands P2 and H1 (Table 4).
Juveniles of Fagus, Quercus and other species(Corylus, Frangula, Pyrus and Salix) were nega-tively associated with old trees in stands H1 andH2, but Ilex and old trees were always spatiallyindependent (Figure 6). The distances of negativeassociation were smaller in stand H1 than in H2.In stand H1, Fagus juveniles showed negativeassociation with old trees at 1 –5 m, Quercus at 2 –11 m, and other species at 3 –10 m. In stand H2,
-2
-1
0
1
2
L(d)
-d
0 5 10 15 20 25
Distance (m)
-2
-1
0
1
2
L(d)
-d
H1
H2
Figure 5. Spatial point patterns for old Fagus and Quercus trees
combined in stands H1 and H2. Sample sizes range from 43 to
92. Format and interpretation as in Figure 4.
65

the distances of negative association were 1 –22 mfor Fagus, 3 –17 m for Quercus, and 2 –18 m forother species.
Discussion
Populations structure
Regarding the first hypothesis, Fagus and Quercusshowed reverse-J shaped diameter distribution,which is in accordance with idealized, well-balanced populations structure for old-growthforest (Veblen 1992; Oliver and Larson 1996).However, populations of Fagus and Quercus werenot all-aged, showing two clear age groups sepa-rated by periods of 85 –185 yr without treerecruitment. Deficiency of trees dating from theseperiods may be due to lack of tree establishment,
but also to high mortality of trees regenerating atthis time (Johnson et al. 1994). Nonetheless, thisresult was comparable to those previously ob-tained from other old-growth forests (Chokkalin-gam and White 2001; Motta et al. 2002),suggesting that balanced diameter distributions donot necessarily mean that continuous regenerationhas occurred. The particular structure of tree agesin the Caviedes forest can be attributed to themanagement history, especially to temporal vari-ation in grazing pressure. Evidence supporting theidea that intense browsing, by domestic and wildungulates, can cause large gaps in the age struc-tures of forest trees has been previously reported(Hytteborn et al. 1987; Abrams and Orwig 1996;Motta et al. 2002). In the Caviedes forest, Fagus,Quercus, Ilex and the other species mainly estab-lished in periods of reduced grazing intensity, i.e.from the 1740s to the 1830s and since the 1920s
Table 4. Interaction between juveniles in stands P1, P2, H1 and H2. Spatial associations (species 1/species 2) for pre-mortality data
sets, significant at p=0.05 according to bivariate Ripley’s K-function, are shown. Other species include Corylus, Crataegus, Frangula,
Pyrus and Salix.
Interaction Distance (m)
1 5 10 15 20 25
Stand P1
Fagus/Quercus Æ + + + + + Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ ÆFagus/Ilex Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ ÆFagus/Other Æ Æ ) Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ ) ) Æ Æ Æ Æ Æ Æ Æ Æ Æ ÆQuercus/Ilex Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ ÆQuercus/Other Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ ) ) ) ) Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ ÆIlex/Other Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ
Stand P2
Fagus/Ilex Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ ÆFagus/Other Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ ÆIlex/Other Æ Æ Æ Æ Æ ) Æ Æ ) ) Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ
Stand H1
Fagus/Quercus Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ ÆFagus/Ilex + + + + + + + + + + + + + + + + + + + + + Æ Æ Æ ÆFagus/Other Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ ) ) ) ) ) ) ) ) ) ) ) ) ) ) )Quercus/Ilex Æ Æ Æ ) ) ) ) ) ) Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ ÆQuercus/Other Æ + + + + + + Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ ÆIlex/Other Æ Æ ) ) Æ Æ ) ) Æ Æ Æ Æ Æ Æ Æ ) Æ Æ ) ) ) ) ) ) )
Stand H2
Fagus/Quercus Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ ÆFagus/Ilex + Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ ) ) ) ) ) ) ) ) )Fagus/Other Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ ÆQuercus/Ilex Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ ÆQuercus/Other Æ Æ + + + + + + + + + + + + + + + + + + + + + + +
Ilex/Other Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ Æ
+=positive association, Æ=spatial independence, )=negative association. Sample sizes range from 19 to 307.
66
(Rozas 2003a). This is a general feature through-out west-European woodlands, where the prevail-ing browsing pressure by ungulates has animportant impact on regeneration of broadleavedspecies (Kuiters and Slim 2002; Palmer et al. 2004).
Similar to the results from this study, compounddiameter distributions have been previously re-ported for Fagus in old-growth high-forest stands(Koop and Hilgen 1987). Although compounddiameter distributions have been attributed tovertical stratification in shade-tolerant species(Lorimer and Krug 1983), in both H1 and H2stands they were a consequence of the presence oftwo differently sized cohorts of Fagus. Themajority of Fagus with DBH<30 cm were youn-ger than 80 yr, while trees with DBH>30 cm weretypically more than 150 yr. As it has been found inother understorey species (Manabe and Yamam-oto 1997; Kanno et al. 2001), non-dominant
species in Caviedes displayed descending diameterdistributions, suggesting continuous recruitment.Age data indicated that Frangula, Ilex, Pyrus andSalix established almost continuously since the1920s. Since their tree-ring series were not cross-dated, and some of them displayed vegetativereproduction, it must be taken into account thatage estimates in these species did not always re-vealed their ages since germination.
Structural heterogeneity
The obtained results supported the hypothesis thatthe Caviedes forest is structurally heterogeneous.These results showed that structural characteristicsvary locally within the forest and can be related toparticular dynamic processes mainly depending onthe initial structure of the stand. In the formerparkland stands, diameters and ages of neighbor-ing trees were independent, i.e. both P1 and P2stands were structurally homogeneous with respectto tree diameter and age. However, these standswere a mosaic of small patches of trees that arehomogeneous in tree height, but differing fromspatially adjacent patches. A similar pattern ofpositive height autocorrelation up to inter-treedistances of 15 m has been also found in a 100-yrold population of Pinus sylvestris established in aburnt area (Kuuluvainen and Rouvinen 2000).This heterogeneity in tree height can be attributedto a patchy differentiation in height growth andmortality rates due to competitive interactions,which is characteristic of even-aged stands (Oliverand Larson 1996; Kenkel et al. 1997). It can beexpected that when there are only a few trees in apatch, competition should not be very intense andthe canopy is shorter, while when there are manytrees, canopies are denser and taller (Kuuluvainenet al. 1998; Chen and Bradshaw 1999). The ab-sence of spatial autocorrelation for tree diametercould indicate that former parkland stands are inthe period of greatest density-dependent mortality(Kenkel et al. 1997), which implied that manysmall trees are dying, reducing autocorrelationwithin the dense clumps of small-sized trees.
In the high forest stands, by contrast, all thestructural parameters were heterogeneous, indi-cating that both H1 and H2 stands are composedof patches consisting of similarly aged and sizedtrees, but differing from spatially adjacent patches.
H1 H2
-3-2-10123
-2
-1
0
1
2
Fag
us
-6
-3
0
3
6
Que
rcus
-4
-2
0
2
4
-4
-2
0
2
4
L 12(
d)-d
-4
-2
0
2
4
lIex
0 5 10 15 20 25-4
-2
0
2
4
0 5 10 15 20 25
Distance (m)
-4
-2
0
2
4
Oth
er
Figure 6. Spatial associations between old Fagus and Quercus
trees combined, and juveniles of Fagus, Quercus, Ilex and others
(Corylus, Frangula, Pyrus and Salix), in stands H1 and H2.
Dashed lines are the 95% confidence envelopes for the spatial
independence hypothesis. Empirical L12(d) –d values above the
confidence envelope indicate a positive association, below
indicate a negative one. Sample sizes range from 39 to 307.
67

These structural units might be considered as ele-mentary functional patches of the forest mosaic,according to the patch-dynamics paradigm, thatconsider the disturbance-caused canopy openingsas the starting points of the silvigenetic cycle(Oliver and Larson 1996; Busing 1998). Fine-scaletree falls sometimes control the regenerationdynamics of mesic temperate forests, creating amosaic of gap, building and mature phases (Koopand Hilgen 1987; Veblen 1992). Evidence of even-aged patches of trees has been previously obtainedin other forests for particular tree species (Duncanand Stewart 1991; Lusk and Smith 1998; Mast andVeblen 1999; Rogers 1999), and also in the Cav-iedes forest for Fagus and Quercus (Rozas 2003a).The size of patches of even-aged trees revealed bycorrelograms, 5 m in stand H1 and 10 m in standH2, would indicate the prevalent size of formercanopy openings in these stands. The lack of even-aged patches in the former parkland stands indi-cates that development of this heterogeneity is aslow process requiring at least 150 –200 yr.
Overstorey conditions and tree spatial distribution
Dominant tree species showed a partially over-lapping spatial distribution among the differentforest habitats. Fagus was more abundant in for-mer open sites and in canopy gaps formed byQuercus, probably due to the existence of ad-vanced regeneration, while Quercus was moreabundant in former open sites. The relative lack ofFagus juveniles near old conspecifics could be aconsequence of the low survival of seedlings be-neath old Fagus trees during the years followinggermination (Peltier et al. 1997; Szwagrzyk et al.2001). But the abundance of Fagus juveniles wasrelatively high in all forest environments, whichindicate that this species is a generalist in itsregeneration behavior. By contrast, Quercus wasalmost restricted to old and recent canopy gaps,which is consistent with gap-obligate regenerationbehavior (Rozas 2003a).
Among understorey trees, Ilex was significantlymore abundant beneath old Quercus trees thanunder other overstorey types. Ilex regeneration ishighly influenced by habitat heterogeneity, withthe intensity of seed rain, seed germination, andseedling survival being greater beneath intermedi-ate shaded tree canopies (Obeso and Fernandez-
Calvo 2002; Arrieta and Suarez 2005). Spatialpatterns of Ilex reproduce the spatial dispositionof the canopies of old Quercus trees. In formerparkland stands, Ilex was clumped at small andintermediate scales, and randomly or regularlydistributed at larger scales, due to the sparse dis-tribution of old trees. By contrast, in high-foreststands, safe sites for Ilex were less intenselyclumped on a small spatial scale, but more in-tensely clumped at intermediate and larger scales,due to the existence of extensive patches of oldtrees (Rozas 2003a).
Other understorey species were found mainly informer open sites, and near dead Quercus trees.Spatial dispersion of shade-intolerant understoreyspecies is typically dependent on the distribution ofcanopy gaps (Manabe and Yamamoto 1997),probably due to the combined effect of establish-ment preferences and a differential probability ofsurvival. The analysis of spatial distribution in allunderstorey species, considered as a whole, prob-ably masked their differences in regenerationbehavior. Fleshy-fruited species (i.e. Crataegus,Frangula, Pyrus) use to be particularly abundantunder and near forest edges, where perching bybirds are common, while species dispersed bycaching animals (i.e. Corylus, Quercus) tend to bemore abundant in open areas (Kollmann andSchill 1996). Also Salix, the only wind-dispersedspecies, was occasionally found where other treeswere absent, in poorly-drained soil and on up-rooted mounds, probably because of the lowprobability of seedling establishment on the litterlayer.
Tree spatial patterns and associations
As in previous investigations of spatial patterns inold-growth forests (e.g., Moeur 1993; Busing 1998;Manabe et al. 2000; Chokkalingam and White2001), this present work provides sound evidencefor clumped distribution of juveniles. This is aconsequence of the contrasting suitable site con-ditions for establishment and survival (Frelichet al. 1993). The intensity of clumping was foundto change with the scale of the analysis, and itvaried among species and stands. The importanceof density-dependent mortality in leading towardsa less aggregated distribution has been repeatedlyemphasized elsewhere (Ward et al. 1996; He et al.
68

1997; Chen and Bradshaw 1999). The tendency formortality to lead to a less intensely aggregateddistribution of juveniles of Fagus and Quercus isconsistent with an outcome expected from self-thinning, in which density-dependent processescould influence tree survival (Kenkel 1988; Dun-can 1991).
In contrast, a more aggregated post-mortalitydistribution in shade-intolerant understorey spe-cies was evident. The intensity of clumping canincrease after fires due to the distribution of sur-vivors in discrete, isolated clumps protected fromfire damage (Rebertus et al. 1989; Park 2003). Butmore probably, the shift toward a more aggregateddistribution in understorey species could be aconsequence of interspecific neighborhood effecton tree survival (He and Duncan 2000). Strongeraggregation than expected could result fromhigher tree survival in favorable canopy openings,away from dominant canopy trees.
The trend from clumped to random distributionin old trees agreed only partially with results fromstudies on spatial dynamics, which suggests auniform pattern for dominant trees (Turner andFranz 1985; Stewart and Rose 1990; Ward et al.1996). Several works suggested that the final pat-tern for mature trees use to be random, whileregular patterns are rare, with the exceptions ofvery stable environments without significantexogenous disturbances (Sterner et al. 1986; Heet al. 1997; Antos and Parish 2002) or stands witha long history of human intervention and man-agement (Wolf 2005). The observed random pat-tern of survivors was consistent with the outcomeexpected from mortality of maturing trees inducedby winds, pathogens, droughts, defoliating out-breaks and other diseases.
Studies of mature and old-growth forests haveshown that establishment occurs preferentially incanopy gaps, leading to characteristic negativeassociation of juveniles and old trees at theapproximate scale of a gap (Leemans 1991; Moeur1993; Busing 1998). Strong segregation of patchesof juveniles and old conspecifics has been previ-ously revealed for Fagus and Quercus in the Cav-iedes forest (Rozas 2003a). This present workrevealed that shade-intolerant understory specieswere also negatively associated with old trees.Segregation of juveniles and old trees occurred atwider distances in stand H2 (17 –22 m) than in H1(5 –11 m), which is consistent with a larger size of
patches of even-aged trees in stand H2, as revealedby autocorrelation analyses (Figure 3).
The obtained spatial associations suggested thatinteractions between tree species vary within theforest and could be related to particular dynamicprocesses, such as tree establishment and compe-tition. Negative associations among species estab-lished simultaneously can be indicative of thepartitioning of establishment sites and/or inter-specific competition (Duncan 1993; Rejmanek andLeps 1996). Competition could explain the nega-tive associations found between Fagus and otherspecies in stands P1 and H1, and between Quercusand other species in stand P1. However, estab-lishment site partitioning can be the reason fornegative associations between Ilex and other spe-cies in stands P2 and H1, and between Quercus andIlex in stand H1. Positive associations at smalldistances could be expected when the species havesimilar requirements for establishment sites(Manabe et al. 2000), as occurred with Fagus andQuercus in stand P1, Fagus and Ilex in stand H1,and Quercus and other species in stands H1 andH2. The results reported here agree with studiesshowing that independence is a common spatialrelationship among different tree species (Duncan1993; Szwagrzyk and Czerwczak 1993; Rogers1999; Park 2003). Spatial independence, through acertain differentiation in their establishment sitepreferences, can contribute to avoid interspecificcompetition and promote long-term coexistence oftree species (Duncan 1991). Further analyses, fo-cused on the study of the relationships betweenneighborhood density, tree growth and survival(He and Duncan 2000), are needed to ascertain therole of competition in causing the observed spatialpatterns and associations.
General implications for forest restoration
As almost all forest in Europe has been managed,forest restoration has always to deal with standswith a composition, structure and function devel-oped through a history of repeated, intenseanthropogenic disturbance (Wolf 2005). Anywood,which is to be treated as a natural reserve and re-stored, will inherit a structure created by pastmanagement that should be preserved as a startingpoint for the future forest structure. Age distribu-tions in the studied stands revealed that the
69

prevailing browsing pressure by ungulates is adecisive factor for successful regeneration ofbroadleaved species. The first step for restoringshould be the control of wild and domestic ungu-lates at population levels that match browsingpressures in natural temperate forests (Kuiters andSlim 2002). However, blanket recommendationsapplicable to any site based upon a thresholdungulate density, which will allow regeneration,cannot be made (Palmer et al. 2004). Naturalregeneration is a desirable option, but if it does notoccur tree planting should be carried out. In agree-ment with results obtained from other formerlymanaged forests (Motta et al. 2002; Aldrich et al.2003; Wolf 2005), this study has shown thatthe clumped distribution of recruits seemed to bethe more natural state after cessation of humanintervention. Contrary to Spanish legislation onforest restoration, planting should not be asevenly spaced as in productive plantations, butassembled in clumps of contrasting density. Toguarantee an uneven-aged structure of tree popu-lations, clumps should not cover the completesite. The recommendation is to reserve areasfor natural establishment in the future. Thisadvice would also help to preserve the spatialheterogeneity in stand structure, as it has beenfound in the studied stands.
Acknowledgements
The author thanks J.R. Arevalo, J.J. Camareroand J.M. Olano for helpful comments and sug-gestions on an earlier manuscript, and K.McKenney for English language assistance. Thecritical review of three anonymous referees greatlyenhanced the quality of the paper. Spatial auto-correlation and spatial pattern analyses were per-formed with the spatial analysis programs writtenby R.P. Duncan. The Junta Vecinal de Caviedesand the Servicio de Montes y Conservacion de laNaturaleza de Cantabria gave permission fortaking cores.
References
Abrams M.D. and Orwig D.A. 1996. A 300-year history of
disturbance and canopy recruitment for co-occurring white
pine and hemlock on the Allegheny Plateau, USA. J. Ecol.
84: 353 –363.
Aldrich P.R., Parker G.R., Ward J.S. and Michler C.H. 2003.
Spatial dispersion of trees in an old-growth temperate hard-
wood forest over 60 years of succession. For. Ecol. Manage.
180: 475 –491.
Allue J.L. 1990. Atlas fitoclimatico de Espana. Ministerio de
Agricultura, Pesca y Alimentacion, Madrid.
Antos J.A. and Parish R. 2002. Structure and dynamics of a
nearly steady-state subalpine forest in south-central British
Columbia, Canada. Oecologia 130: 126 –135.
Arevalo J.R. and Fernandez-Palacios J.M. 2003. Spatial pat-
terns of trees and juveniles in a laurel forest of Tenerife,
Canary Islands. Plant Ecol. 165: 1 –10.
Arrieta S. and Suarez F. 2005. Spatial patterns of seedling
emergence and survival as a critical phase in holly (Ilex
aquifolium L.) woodland recruitment in Central Spain. For.
Ecol. Manage. 205: 267 –282.
Busing R.T. 1998. Composition, structure and diversity of cove
forest stands in the Great Smoky Mountains: a patch
dynamics perspective. J. Veg. Sci. 9: 881 –890.
Chen J. and Bradshaw G.A. 1999. Forest structure in space: a
case study of an old growth spruce-fir forest in Changbaishan
Natural Reserve, PR China. For. Ecol. Manage. 120: 219 –
233.
Chokkalingam U. and White A. 2001. Structure and spatial
patterns of trees in old-growth northern hardwood and mixed
forests of northern Maine. Plant Ecol. 156: 139 –160.
Dixon P.M. 2002. Ripley’s K function. In: El-Shaaraui A.H.
and Piergorsch W.W. (eds.), The Encyclopedia of Environ-
metrics, Wiley, New York, pp. 1796 –1803.
Duncan R.P. 1991. Competition and the coexistence of species
in a mixed podocarp stand. J. Ecol. 79: 1073 –1084.
Duncan R.P. 1993. Flood disturbance and the coexistence of
species in a lowland podocarp forest, south Westland, New
Zealand. J. Ecol. 81: 403 –416.
Duncan R.P. and Stewart G.H. 1991. The temporal and spatial
analysis of tree age distributions. Can. J. For. Res. 21: 1703 –
1710.
Foster D.R., Orwig D.A. and McLachlan J.S. 1996. Ecological
and conservation insights from reconstructive studies of
temperate old-growth forests. Trend. Ecol. Evol. 11: 419 –
424.
Frelich L.E., Calcote R.R., Davis M.B. and Pastor J. 1993.
Patch formation and maintenance in an old-growth hemlock-
hardwood forest. Ecology 74: 513 –527.
Guitian F., Carballas T., Dıaz-Fierros F. and Plata M. 1985.
Suelos naturales de Cantabria. CSIC, Instituto de Investi-
gaciones Agrobiologicas de Galicia, Santiago de Compostela.
He F. and Duncan R.P. 2000. Density-dependent effects on tree
survival in an old-growth Douglas fir forest. J. Ecol. 88: 676 –
688.
He F., Legendre P. and LaFrankie J.V. 1997. Distribution
patterns of tree species in a Malaysian tropical rain forest. J.
Veg. Sci. 8: 105 –114.
Hytteborn H., Packham J.R and Verwijst T. 1987. Tree pop-
ulation dynamics, stand structure and species composition in
the montane virgin forest of Vallibacken, northern Sweden.
Vegetatio 72: 3 –19.
Johnson E.A., Miyanishi K. and Kleb H. 1994. The hazards of
interpretation of static age structures as shown by stand
70

reconstructions in a Pinus contorta-Picea engelmannii forest.
J. Ecol. 82: 923 –931.
Kanno H., Hara M., Hirabuki Y., Takehara A. and Seiwa K.
2001. Population dynamics of four understorey shrub species
during a 7-yr period in a primary beech forest. J. Veg. Sci. 12:
391 –400.
Kenkel N.C. 1988. Pattern of self-thinning in jack pine: testing
the random mortality hypothesis. Ecology 69: 1017 –1024.
Kenkel N.C., Hendrie M.L. and Bella I.E. 1997. A long-term
study of Pinus banksiana population dynamics. J. Veg. Sci. 8:
241 –254.
Kollmann J. and Schill H.-P. 1996. Spatial patterns of dispersal,
seed predation and germination during colonization of
abandoned grassland by Quercus petraea and Corylus avell-
ana. Vegetatio 125: 193 –205.
Koop H. and Hilgen P. 1987. Forest dynamics and regeneration
mosaic shifts in unexploited beech (Fagus sylvatica) stands at
Fontainebleau (France). For. Ecol. Manage. 20: 135 –150.
Kuiters A.T. and Slim P.A. 2002. Regeneration of mixed
deciduous forest in a Dutch forest-heathland, following a
reduction of ungulate densities. Biol. Conserv. 105: 65 –74.
Kuuluvainen T. and Rouvinen S. 2000. Post-fire understorey
regeneration in boreal Pinus sylvestris forest sites with dif-
ferent fire histories. J. Veg. Sci. 11: 801 –812.
Kuuluvainen T., Jarvinen E., Hokkanen T.J., Rouvinen S. and
Heikkinen K. 1998. Structural heterogeneity and spatial
autocorrelation in a natural mature Pinus sylvestris domi-
nated forest. Ecography 21: 159 –174.
Leemans R. 1991. Canopy gaps and establishment patterns of
spruce (Picea abies (L.) Karst.) in two old-growth coniferous
forests in central Sweden. Vegetatio 93: 157 –165.
Legendre P. and Fortin M.-J. 1989. Spatial pattern and eco-
logical analysis. Vegetatio 80: 107 –138.
Lorimer C.G. and Krug A.G. 1983. Diameter distributions in
even-aged stands of shade-tolerant and midtolerant tree
species. Am. Midl. Nat. 109: 331 –345.
Lotwick H.W. and Silverman B.W. 1982. Methods for ana-
lyzing spatial processes of several types of points. J. R. Stat.
Soc. B 44: 405 –413.
Lusk C.H. and Smith B. 1998. Life history differences and tree
species coexistence in an old-growth New Zealand rain forest.
Ecology 79: 795 –806.
Manabe T. and Yamamoto S. 1997. Spatial distribution of
Eurya japonica in an old-growth evergreen broad-leaved
forest, SW Japan. J. Veg. Sci. 8: 761 –772.
Manabe T., Nishimura N., Miura M. and Yamamoto S. 2000.
Population structure and spatial patterns for trees in a tem-
perate old-growth evergreen broad-leaved forest in Japan.
Plant Ecol. 151: 181 –197.
Mast J.N. and Veblen T.T. 1999. Tree spatial patterns and
stand development along the pine-grassland ecotone in the
Colorado Front Range. Can. J. For. Res. 29: 575 –584.
McDonald R.I., Peet R.K. and Urban D.L. 2003. Spatial pat-
terns of Quercus regeneration limitation and Acer rubrum
invasion in a Piedmont forest. J. Veg. Sci. 14: 441 –450.
Miyadokoro T., Nishimura N. and Yamamoto S. 2003. Pop-
ulation structure and spatial patterns of major trees in a
subalpine old-growth coniferous forest, central Japan. For.
Ecol. Manage. 182: 259 –272.
Moeur M. 1993. Characterizing spatial patterns of trees using
stem-mapped data. Forest Science 39: 756 –775.
Motta R., Nola P. and Piussi P. 2002. Long-term investigations
in a strict forest reserve in the eastern Italian Alps: spatio-
temporal origin and development in two multi-layered sub-
alpine stands. J. Ecol. 90: 495 –507.
Obeso J.R. and Fernandez-Calvo I.C. 2002. Fruit removal, py-
rene dispersal, post-dispersal predation and seedling estab-
lishment of a bird-dispersed tree. Plant Ecol. 165: 223 –233.
Oliver C.D. and Larson B.C. 1996. Forest Stand Dynamics.
Wiley & Sons, New York, Update edition.
Palmer S.C.F., Mitchell R.J., Truscott A.-M. and Welch D.
2004. Regeneration failure in Atlantic oakwoods: the roles of
ungulate grazing and invertebrates. For. Ecol. Manage. 192:
251 –265.
Park A. 2003. Spatial segregation of pines and oaks under
different fire regimes in the Sierra Madre Occidental. Plant
Ecol. 169: 1 –20.
Park A., Kneeshaw D., Bergeron Y. and Leduc A. 2005. Spatial
relationships and tree species associations across a 236-year
boreal mixedwood chronosequence. Can. J. For. Res. 35:
750 –761.
Peltier A., Touzet M.-C., Armengaud C. and Ponge J.-F. 1997.
Establishment of Fagus sylvatica and Fraxinus excelsior in an
old-growth beech forest. J. Veg. Sci. 8: 13 –20.
Peterken G.F. 1996. Natural Woodland. Ecology and Conser-
vation in Northern Temperate Regions. Cambridge Univer-
sity Press, Cambridge.
Rebertus A.J., Williamson B. and Moser E.B. 1989. Fire-in-
duced changes in Quercus laevis spatial pattern in Florida
sandhills. J. Ecol. 77: 638 –650.
Rejmanek M. and Leps J. 1996. Negative associations can re-
veal interspecific competition and reversal of competitive
hierarchies during succession. Oikos 76: 161 –168.
Ripley B.D. 1977. Modelling spatial patterns. J. R. Stat. Soc. B
39: 172 –212.
Rogers H.M. 1999. Stand dynamics of Dacrydium cupressinum
dominated forest on glacial terraces, south Westland, New
Zealand. For. Ecol. Manage. 117: 111 –128.
Rozas V. 2003a. Regeneration patterns, dendroecology, and
forest-use history in an old-growth beech-oak lowland forest
in Northern Spain. For. Ecol. Manage. 182: 175 –194.
Rozas V. 2003b. Tree age estimates in Fagus sylvatica and
Quercus robur: testing previous and improved methods. Plant
Ecol. 167: 193 –212.
Sterner R.W., Ribic C.A. and Schatz G.E. 1986. Testing for life
historical changes in spatial patterns of four tropical tree
species. J. Ecol. 74: 621 –633.
Stewart G.H. and Rose A.B. 1990. The significance of life
history strategies in the developmental history of mixed
beech (Nothofagus) forests, New Zealand. Vegetatio 87:
101 –114.
Szwagrzyk J. andCzerwczakM. 1993. Spatial patterns of trees in
natural forests of East-Central Europe. J. Veg. Sci. 4: 469 –476.
Szwagrzyk J., Szewczyk J. and Bodziarczyk J. 2001. Dynamics
of seedling banks in beech forest: results of a 10-year study on
germination, growth and survival. For. Ecol. Manage. 141:
237 –250.
Taylor A.H., Zisheng Q. and Jie L. 1996. Structure and
dynamics of subalpine forests in the Wang Lang Natural
Reserve, Sichuan, China. Vegetatio 124: 25 –38.
Turner D.P. and Franz E.H. 1985. Size class structure and
tree dispersion patterns in old-growth cedar-hemlock
71

forests of the northern Rocky Mountains (USA). Oecologia
68: 52 –56.
Tutin T.G., Heywood V.H., Burges N.A., Valentine D.H.,
Walters S.M. and Webb D.A. 1964 –1980. Flora Europaea,
Vols. 1 –5. Cambridge University Press, Cambridge.
Veblen T.T. 1992. Regeneration dynamics. In: Glenn-Lewin
D.C., Peet R.K. and Veblen T.T. (eds.), Plant Succession:
Theory and Prediction, Chapman and Hall, London, pp.
152 –187.
Ward J.S., Parker G.R. and Ferrandino F.J. 1996. Long-term
spatial dynamics in an old-growth deciduous forest. For.
Ecol. Manage. 83: 189 –202.
Williamson G.B. 1975. Pattern and seral composition in an old-
growth beech-maple forest. Ecology 56: 727 –731.
Wolf A. 2005. Fifty year record of change in tree spatial pat-
terns within a mixed deciduous forest. For. Ecol. Manage.
215: 212 –223.
72
Copyright © 2022 FDOKUMEN




















