
Structural diversity of the patatin gene family in potato cv. Desiree
12
Mol Gen Genet (1988) 212:325-336 MGG © Springer-Verlag 1988 Structural diversity of the patatin gene family in potato cv. Desiree David Twell* and Gert Ooms AFRC Institute of Arable Crops Research, Biochemistry Department, Rothamsted Experimental Station, Harpenden, Herts AL5 2JQ, UK Summary. We have used a combined genetical and molecu- lar approach to study the structural diversity of the patatin gene family in tetraploid Solanum tuberosum L. cv. Desiree (2n=4x = 48). Nine dihaploid derivatives (2n= 2x = 24) of cv. Desiree were isolated by gynogenesis through prickle pollination with S. phureja Juz et Buk. Patatin DNA se- quences in Desiree and in the dihaploids were examined by probing Southern blots of restriction endonucleases Hin- dIII and XbaI digested DNA with patatin cDNA region- specific gene probes and by more detailed examination (re- striction endonuclease mapping and partial DNA sequenc- ing) of 10 patatin genomic clones in bacteriophage 2 re- placement vector EMBL4. This provided positive identifica- tion for most individual patatin gene family members and some estimate of their organisation and diversity. Most of the 64-72 patatin DNA copies showed little allelic variation based on HindlII and XbaI restriction fragment length polymorphism (RFLP) mapping and did not appear to be very tightly clustered. Four of the 6-8 class I patatin genes (without a characteristic 22 bp insert in their untranslated leader DNA), showed apparent allelic homogeneity, whilst the remaining class I genes comigrated with a single class II patatin gene RFLP subclass, Of the isolated clones, 4 con- tained apparent pseudogenes lacking 5' control sequences and exon-I DNA while another clone contained a patatin gene truncated at the 5' region due to the cloning event. The remaining 5 all contained class I! genes (with the 22 bp insert) and these showed varying degrees of sequence ho- mology for 400 bp of conserved 5' coding and non-coding DNA (from 77%-95%). In one case the extent of homology differed, with complete sequence divergence upstream of position - 80 from the start of transcription. The structural diversity of the patatin gene family is discussed in relation to expression of individual patatin genes and the use of cv. Desiree as a host for potato transformation experiments. Key words: Patatin - Solanum tuberosum - Gynogenesis - Expression signals Introduction The major soluble protein of potato (Solanum tuberosum) tubers is patatin, a family of glycoproteins with a molecular * Present address: Plant Gene Expression Centre, 800 Buchanon Street, Albany, CA 94710, USA Offprint requests to: G. Ooms weight of about 40 kDa (Racusen and Foote 1980; Paiva et al. 1983). Patatin has been detected in all potato cultivars examined so far, can account for up to 40% of the soluble tuber protein and possesses a lipid acyl hydrolase (LAH) activity (Racusen and Foote 1980; Racusen 1984; Rosahl et al. 1987). Generally, it exhibits considerable charge heter- ogeneity (Park et al. 1983) and, although most abundant in potato tubers, it is also present at low levels in roots and stems of normally (field) grown plants (Pikaard et al. 1987). Patatin can also be artificially induced in petioles of axillary node cuttings with excised meristems under spe- cific conditions i.e. short daylength (Paiva et al. 1983). Be- cause of its prominence in tubers and inducibility under tuberization stimulating conditions, patatin has been pro- posed as a useful biochemical marker to study molecular events prior to visible tuber formation (Park et al. 1985). Patatin gene expression is at least partly controlled at the transcriptional level. This is shown by: (1) run-off RNA analysis (Rosahl et al. 1986), (2) quantitative comparison of steady state RNA levels in different potato organs (Mig- nery et al. 1984; Rosahl et al. 1986) and (3) by transforma- tion of potato with a transcriptional fusion gene, containing patatin 5' control sequences, the reporter gene cat and nos 3' control sequences, which gave tuber specific chloram- phenicol acetyltransferase expression (Twell and toms 1987). A complication in the molecular analysis of patatin expression is the large size of the gene family of which estimates vary from 20-40 copies per tetraploid potato ge- nome (Bevan et al. 1986). Although several different patatin genes characterised so far show considerable DNA homolo- gy (Rosahl etal. 1986; Bevan etal. 1986; Pikaard etal. 1986), individual gene members may show considerable variation in levels of expression and regulation of tissue specificity. This is partly suggested by more advanced stu- dies in plants, such as in the Rubisco gene family in petunia (with only eight members) which indicated that a wide vari- ation in expression of individual members of a gene family is common (Dean et al. 1985b; 1987; Turner et al. 1986), but also by initial analysis on potato of patatin mRNA and protein. For example, in cv. Superior, 50% of the pata- tin mRNA in tubers has been designated class I RNA, based on the presence of a 22 bp insert within the untrans- lated leader sequence, while the other 50% is class II RNA, without insert (Pikaard et al. 1987). In contrast, class II RNA constitutes 100% of the patatin mRNA population in roots, which is only present at about 1% of the level in tubers. Furthermore, tuber proteins from a number of
Transcript of Structural diversity of the patatin gene family in potato cv. Desiree

Mol Gen Genet (1988) 212:325-336 MGG © Springer-Verlag 1988
Structural diversity of the patatin gene family in potato cv. Desiree David Twel l* and Gert Ooms AFRC Institute of Arable Crops Research, Biochemistry Department, Rothamsted Experimental Station, Harpenden, Herts AL5 2JQ, UK
Summary. We have used a combined genetical and molecu- lar approach to study the structural diversity of the patatin gene family in tetraploid Solanum tuberosum L. cv. Desiree (2n=4x = 48). Nine dihaploid derivatives (2n= 2x = 24) of cv. Desiree were isolated by gynogenesis through prickle pollination with S. phureja Juz et Buk. Patatin DNA se- quences in Desiree and in the dihaploids were examined by probing Southern blots of restriction endonucleases Hin- dIII and XbaI digested DNA with patatin cDNA region- specific gene probes and by more detailed examination (re- striction endonuclease mapping and partial DNA sequenc- ing) of 10 patatin genomic clones in bacteriophage 2 re- placement vector EMBL4. This provided positive identifica- tion for most individual patatin gene family members and some estimate of their organisation and diversity. Most of the 64-72 patatin DNA copies showed little allelic variation based on HindlII and XbaI restriction fragment length polymorphism (RFLP) mapping and did not appear to be very tightly clustered. Four of the 6-8 class I patatin genes (without a characteristic 22 bp insert in their untranslated leader DNA), showed apparent allelic homogeneity, whilst the remaining class I genes comigrated with a single class II patatin gene RFLP subclass, Of the isolated clones, 4 con- tained apparent pseudogenes lacking 5' control sequences and exon-I DNA while another clone contained a patatin gene truncated at the 5' region due to the cloning event. The remaining 5 all contained class I! genes (with the 22 bp insert) and these showed varying degrees of sequence ho- mology for 400 bp of conserved 5' coding and non-coding DNA (from 77%-95%). In one case the extent of homology differed, with complete sequence divergence upstream of position - 80 from the start of transcription. The structural diversity of the patatin gene family is discussed in relation to expression of individual patatin genes and the use of cv. Desiree as a host for potato transformation experiments.
Key words: Patatin - Solanum tuberosum - Gynogenesis - Expression signals
Introduction
The major soluble protein of potato (Solanum tuberosum) tubers is patatin, a family of glycoproteins with a molecular
* Present address: Plant Gene Expression Centre, 800 Buchanon Street, Albany, CA 94710, USA
Offprint requests to: G. Ooms
weight of about 40 kDa (Racusen and Foote 1980; Paiva et al. 1983). Patatin has been detected in all potato cultivars examined so far, can account for up to 40% of the soluble tuber protein and possesses a lipid acyl hydrolase (LAH) activity (Racusen and Foote 1980; Racusen 1984; Rosahl et al. 1987). Generally, it exhibits considerable charge heter- ogeneity (Park et al. 1983) and, although most abundant in potato tubers, it is also present at low levels in roots and stems of normally (field) grown plants (Pikaard et al. 1987). Patatin can also be artificially induced in petioles of axillary node cuttings with excised meristems under spe- cific conditions i.e. short daylength (Paiva et al. 1983). Be- cause of its prominence in tubers and inducibility under tuberization stimulating conditions, patatin has been pro- posed as a useful biochemical marker to study molecular events prior to visible tuber formation (Park et al. 1985).
Patatin gene expression is at least partly controlled at the transcriptional level. This is shown by: (1) run-off RNA analysis (Rosahl et al. 1986), (2) quantitative comparison of steady state RNA levels in different potato organs (Mig- nery et al. 1984; Rosahl et al. 1986) and (3) by transforma- tion of potato with a transcriptional fusion gene, containing patatin 5' control sequences, the reporter gene cat and nos 3' control sequences, which gave tuber specific chloram- phenicol acetyltransferase expression (Twell and t o m s 1987). A complication in the molecular analysis of patatin expression is the large size of the gene family of which estimates vary from 20-40 copies per tetraploid potato ge- nome (Bevan et al. 1986). Although several different patatin genes characterised so far show considerable DNA homolo- gy (Rosahl etal. 1986; Bevan etal. 1986; Pikaard etal. 1986), individual gene members may show considerable variation in levels of expression and regulation of tissue specificity. This is partly suggested by more advanced stu- dies in plants, such as in the Rubisco gene family in petunia (with only eight members) which indicated that a wide vari- ation in expression of individual members of a gene family is common (Dean et al. 1985b; 1987; Turner et al. 1986), but also by initial analysis on potato of patatin m R N A and protein. For example, in cv. Superior, 50% of the pata- tin mRNA in tubers has been designated class I RNA, based on the presence of a 22 bp insert within the untrans- lated leader sequence, while the other 50% is class II RNA, without insert (Pikaard et al. 1987). In contrast, class II RNA constitutes 100% of the patatin mRNA population in roots, which is only present at about 1% of the level in tubers. Furthermore, tuber proteins from a number of

326
potato cultivars show a wide range of LAH isoforms, both within and between cultivars, indicating significant differ- ences in the structure and expression of individual patatin genes (Galliard and Dennis 1974; Racusen 1984).
For several reasons, we have focused our analysis on potato cultivar Desiree as a model cultivar in which the molecular complexity of patatin gene structure and expres- sion can be examined. Desiree is a widely grown cultivar with the entire genetic complexity of other tetraploid culti- vars; it is among the more amenable cultivars for classical genetic studies and because of its high regeneration capacity (Webb et al. 1983) it is particularly suitable for tissue cul- ture procedures, including Agrobacterium transformation.
Here, we present an initial description of the structural complexity of the patatin gene family in Desiree and a series of dihaploid plants derived from Desiree by 'prickle pollin- ation' with S. phureja (Uijtewaal et al. •987). Total DNA was examined by Southern blot hybridisations using patatin cDNA sequences and class I and class II sub-family specific oligonucleotides as gene probes. Ten random patatin genes were examined in more detail by cloning in 2 EMBL4 (Frischauff et al. 1983), restriction endonuclease mapping and partial DNA sequence analysis.
Materials and methods
Plants and growth conditions. Potato cultivar Desiree and dihaploid derivatives were micropropagated on Murashige and Skoog (MS) medium (Murashige and Skoog 1962) con- taining 2% sucrose at 25 ° C 8 h dark; 16 h light. Dihaploid derivatives of Desiree were produced by induction of parth- enocarpic seed development by pollination of emasculated Desiree flowers with pollen from S. phureja IVP48 (prickle pollination; Hermsen and Verdenius 1973; Uijtewaal et al. 1987). Nine seeds lacking the S. phureja embryo spot marker (Hermsen and Verdenius 1973) germinated on ster- ile MS medium containing 2% sucrose. Chromosome numbers were determined as described by Karp et al. (1984) and 8 were shown to contain 24 chromosomes per cell (nor- mal karyotype 2n =4x =48). The dihaploid line PDH 528 also derived from Desiree was obtained from Dr. G.R. Mackay (Scottish Plant Breeding Station, Pentlandfield, Roslin, Midlothian, Scotland).
Enzymes and chemicals. Restriction enzymes, Klenow poly- merase, T4 DNA ligase and AMV reverse transcriptase were obtained from Amersham International, BRL or Pharmacia and were used according to the manufacturers' instructions. DNase-free RNase, proteinase K and phen- anthroline hydrate were from Sigma.
Rapid, small scale isolation of potato DNA. Genomic DNA of Desiree and dihaploid derivatives was isolated by a rapid mini-isolation procedure which gave high molecular weight DNA of good quality as determined by gel electrophoresis and restriction endonuclease cleavage. Approximately 300 mg of leaf tissue was ground using a pestle and mortar under liquid nitrogen. To the frozen tissue 0.5 ml TES (100mM Tris-HC1, pH 8.8; 150raM EDTA; 0.1% SDS; 10 mM phenanthroline hydrate) and 150 gl of a proteinase K (1 mg/ml) solution was added. The homogenate was transferred to a 1.5 ml microcentrifuge tube and incubated at 37°C for 2 h. Following centrifugation for 2 min, the supernatant was phenol extracted by gently mixing with
250 gl phenol for 5 min, then 250 t~l chloroform and 25 lal 5 M NaC1 was added and mixing continued for a further 5 min. After centrifugation, the aqueous phase was extract- ed twice with an equal volume of chloroform and the DNA precipitated by adding an equal volume of ethanol at - 2 0 ° C. The DNA was pelleted, briefly vacuum dried, re- dissolved in 300 gl 10 mM Tris-HC1, pH 8.0; 1 mM EDTA, then 3 gg of DNase free ribonuclease A was added and incubated for 30 rain at 37 ° C. The mixture was phenol/ chloroform extracted once, chloroform extracted twice as described above and the DNA precipitated with ethanol. Yields of 40-60 gg DNA per sample were routinely ob- tained.
Isolation of patatin genomic clones. Recombinant bacterio- phage lambda EMBL4 clones containing patatin genes were isolated as previously described (Twell and Ooms 1987). In short, a genomic library of potato cv. Desiree DNA (isolated from leaves) in 2 EMBL4 (Frischauff et al. 1983) was screened with the nick translated insert of patatin cDNA clone pGM203 (Mignery et al. 1984) and hybridising plaques purified by several rounds of plaque hybridisation according to Benton and Davis (1977).
DNA sequencing. The dideoxy chain termination method was used for DNA sequencing according to standard meth- ods (Sanger et al. 1977). Templates for sequencing were prepared by direct cloning of restriction fragments into bac- teriophage M13 and pUC plasmid vectors (Yanisch-Perron et al. 1985) and transformation into Escherichia coli strains JMI01 and JM83, respectively, using standard methods (Maniatis et al. 1982). Some regions of DNA were se- quenced in double stranded plasmid vectors (Chen and See- burg 1985) using synthetic oligonucleotides complementary to sequences present within the 5' flanking and N-terminal coding DNA of the patatin gene. These oligonucleotides (5'-CCAACTATAAATTCACTTTCC-3', 5 '-GGTTGA- TATTTAAGTGAAAGG-3 ' and 5'-GGAACAGTCAC- CATTTCTCCC-3') were synthesised on an Applied Biosys- terns DNA synthesiser model 380A. Computer analysis and comparison of DNA sequences was performed on a VAXll/750 computer using the Staden (1980) and Univer- sity of Wisconsin (Devereaux et al. 1984) DNA sequence analysis software.
Isolation ofpatatin cDNA clones. Poly(A +) RNA from po- tato (cv. Desiree) tubers was isolated as described by Burrell et al. (1985). A cDNA library was constructed from tuber poly(A +) DNA in bacteriophage 2gtl0 essentially as de- scribed by Huynh et al. (1985). The cDNA library was screened by plaque hybridisation to the nick translated (Rigby et al. 1977) insert of patatin cDNA clone pGM203. Hybridising plaques were purified by two further rounds of hybridisation and DNA isolated from small scale liquid lysates according to Maniatis et al. (1982).
3 2p labelling of DNA probes. Isolated fragments of patatin cDNA clone pGM01 (Mignery et al. 1984) were labelled to high specific activity (0.5-I x 109 dpm/gg) by random primer directed synthesis according to Feinberg and Vogel- stein (1983) for use in Southern blot hybridisations (South- ern 1975). Oligonucleotides I29 and $27 were labelled to high specific activity with ?:-[32p]ATP and T4 polynucleo- tide kinase for use in Southern blot hybridisations accord-

327
ing to Maniatis et al. (1982). In short, 50 ng of oligonucleo- tide was added to 50-100 gCi 7-[3/p]ATP (Amersham Inter- national, 3,000 Ci/mmol) in a volume of 10 gl, 10 U of T4 polynucleotide kinase was added and the reaction incubated for 60 min at 37 ° C. Unincorporated y-[32p]ATP was re- moved by passage through a I ml column of Sephadex G15. Specific activities of oligonucleotide probes obtained were 4-8 X 10 9 dpm/gg.
Southern blot hybridisations. Five to ten microgram samples of potato (cv. Desiree) or dihaploid derivative DNA, were digested with restriction endonucleases, electrophoresed in 0.75% agarose gels using standard procedures (Maniatis et al. 1982) and blotted onto Biodyne A nylon membranes according to the manufacturer's instructions. Some blots were hybridised to ~-[32P]dATP labelled patatin cDNA pGM01 DNA fragments. Hybridisation conditions were as described by Burrell et al. (1985), except that formamide was omitted from the hybridisation solution and the tem- perature was raised to 65 ° C. Blots were washed three times in 2 x S S C (1 x SSC is 0.15 M NaC1, 0.015 M Sodium- citrate), 0.1% sodium dodecylsulphate at room temperature for 10 rain each, followed by three washes in 0.1-0.3 x SSC, 0.1% SDS at 65°C each for 30 min, before exposure to Fuji X-ray film for 4-5 days at - 7 0 ° C. Different blots were hybridised with y-[32p]ATP labelled oligonucleotides I29 or $27 under the same hybridisation solution, at 53 ° C overnight. These blots were washed twice for 30 rain at 53 ° C with 6 x SSC 0.1% SDS and subsequently in the same solution at 58°C for 10 min. Blots were exposed to Fuji X-ray film for 10-14 days.
Results
Dihaploids from cv. Desiree
From potato cv. Desiree (2n-- 4x = 48), we isolated 8 dihap- loid derivatives (2n = 2x = 24) by pollination of emasculated Desiree flowers with pollen from S. phureja IVP48 (prickle pollination). Seeds lacking the embryo-spot marker were grown into mature plants and, by cytological examination of root tips, confirmed to have the reduced chromosome number of 24 chromosomes per cell. The plants were clearly distinguishable in a range of morphological characteristics (not shown) and were distinct from the parental cv. Desiree. They were numbered DDH1 to DDH8.
Complexity and distribution of patatin DNA in cv. Desiree
The strategy we followed to provide an initial description of the patatin multigene family of cv. Desiree was essentially based on the principles of restriction fragment length poly- morphism (RFLP) mapping (Helentjaris et al. 1985). On the one hand, use of the above described Desiree dihaploids was expected to provide a genetic simplification of the com- plexity of the Desiree patatin gene family, and to give some insight into the chromosomal segregation of patatin related sequences. On the other hand, selection of particular restric- tion endonucleases was required to obtain useful variation in the gel separation of restriction endonuclease fragments detectable in Southern blot experiments with patatin gene probes. For this purpose we selected HindIII and XbaI which both gave, upon hybridisation analysis with full- length patatin cDNA probe pGM01 (Mignery et al. 1984),
a wide range of distinct and often well separated fragments with a molecular weight range of 1.4 to 12 kb (Fig. 1 A, B). Substantial variation was present in the intensity of the bands, indicating comigration of multiple patatin re- lated restriction endonuclease fragments in a number of instances. Perhaps surprisingly, the hybridisation pat tern of cDNA clone pGM01 was nearly identical for all the dihaploids and the parent cv. Desiree with both HindIII and XbaI digested DNA. This suggested that the patatin gene complements of each of the four haploid genomes of cv. Desiree are very similar in both structure and copy number.
Hybridisations with short patatin gene probes
Sequence information obtained in previous studies for four genomic patatin clones (Rosahl et al. 1986b; Bevan et al. 1986; Pikaard et al. 1986) has indicated that patatin genes invariably contain six introns which contribute to the length of patatin genomic DNA (of at least 4.2 kb) and to the RFLP observed between patatin genes. This partly explains the complexity of the hybridisation patterns obtained using full length patatin cDNA probe pGM01. Therefore, to ob- tain further simplification of the hybridisation patterns in Fig. 1, we also used shorter fragments derived from pGM01 as probes: a 253 bp HindIII-BamHI fragment and an 80 bp DraI-HindIII fragment from the central and 5' regions of pGM01, respectively. As expected from the hybridisations with pGM01, the patterns obtained with the shorter probes were nearly identical for Desiree DNA and for DNA from all of the dihaploid derivatives. Furthermore, the hybridisa- tions gave much simpler patterns. From these simplified patterns we estimated the total patatin gene copy number in cv. Desiree, and divided the gene family into subclasses (see Table 1). In these estimates we made the fundamental assumption that the clearly visible bands in XbaI and Hin- dIII patterns represented a copy number per tetraploid ge- home of four patatin genes or multiples thereof. This as- sumption was based on: (1) comparison of the intensity of the bands with reconstruction lanes, (2) on the above described genetic evidence suggesting little alMic variation, and (3) on the high specificity of the probes used, which we expected to detect all patatin related sequences in the Desiree genome with equal or similar efficiency under our hybridisation conditions. This latter expectation was based on the high degree of DNA homology between all patatin sequences published so far (cDNAs and genomics), and was experimentally confirmed for patatin sequences in cv. Desiree by varying the stringency of the hybridisation con- ditions (not shown). Thus, in the HindIII digest of cv. De- siree DNA, probed with the 253 bp central probe, were 6 bands representing 24 putative patatin gene copies plus 1 very strong band representing 6--10 multiples of 4 putative gene copies. Similarly, in the cv. Desiree XbaI digest were 5 bands representing 4 putative patatin gene copies plus 2 bands, one with 5-6 and the other 6-7 sets of 4 putative gene copies. Hybridisations with the even smaller 146 bp DraI-HinfI fragment, in a HindIII digest gave 9 four-copy bands, plus 1 band with two, and I with three or four sets, respectively. In the XbaI digest were approximately 9 four- copy bands, 1 band with two and I with three or four sets of patatin related sequences.
One implication of the assumption that the four haploid genomes contain essentially identical alleles for all the ma-
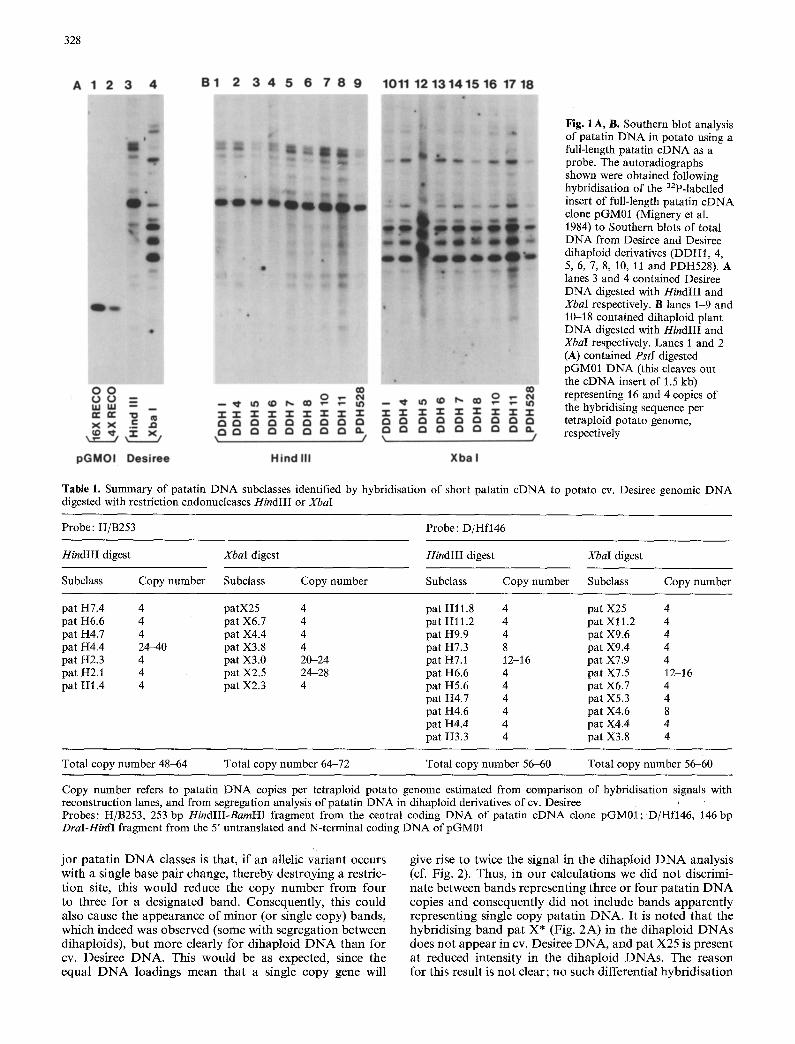
328
Fig. 1 A, B. Southern blot analysis of patatin DNA in potato using a full-length patatin cDNA as a probe. The autoradiographs shown were obtained following hybridisation of the azP-labelled insert of full-length patatin cDNA clone pGM01 (Mignery et al. 1984) to Southern blots of total DNA from Desiree and Desiree dihaploid derivatives (DDH1, 4, 5, 6, 7, 8, 10, 11 and PDH528). A lanes 3 and 4 contained Desiree DNA digested with HindIII and XbaI respectively. B lanes 1-9 and 10-18 contained dihaploid plant DNA digested with HindIII and XbaI respectively. Lanes 1 and 2 (A) contained PstI digested pGM01 DNA (this cleaves out the cDNA insert of 1.5 kb) representing 16 and 4 copies of the hybridising sequence per tetraploid potato genome, respectively
Table 1. Summary of patatin DNA subclasses identified by hybridisation of short patatin cDNA to potato cv. Desiree genomic DNA digested with restriction endonucleases HindIII or Xbal
Probe: H/B253 Probe: D/Hf146
HindlII digest XbaI digest HindlII digest XbaI digest
Subclass Copy number Subclass Copy number Subclass Copy number Subclass Copy number
pat H7.4 4 patX25 4 pat H6.6 4 pat X6.7 4 pat H4.7 4 pat X4.4 4 pat H4.4 24-40 pat X3.8 4 pat H2.3 4 pat X3.0 20-24 pat H2.1 4 pat X2.5 24-28 pat H1.4 4 pat X2.3 4
pat Hl l .8 4 pat X25 4 pat Hl l .2 4 pat Xl l .2 4 pat H9.9 4 pat X9.6 4 pat H7.3 8 pat X9.4 4 pat H7.1 12-16 pat X7.9 4 pat H6.6 4 pat X7.5 12-16 pat H5.6 4 pat X6.7 4 pat H4.7 4 pat X5.3 4 pat H4.6 4 pat X4.6 8 pat H4.4 4 pat X4.4 4 pat It3.3 4 pat X3.8 4
Total copy number 48-64 Total copy number 64-72 Total copy number 56-60 Total copy number 56-60
Copy number refers to patatin DNA copies per tetraploid potato genome estimated from comparison of hybridisation signals with reconstruction lanes, and from segregation analysis of patatin DNA in dihaploid derivatives of cv. Desiree Probes: H/B253, 253 bp HindIII-BamHI fragment from the central coding DNA of patatin cDNA clone pGM01; D/Hf146, 146 bp DraI-Hinfl fragment from the 5' untranslated and N-terminal coding DNA of pGM01
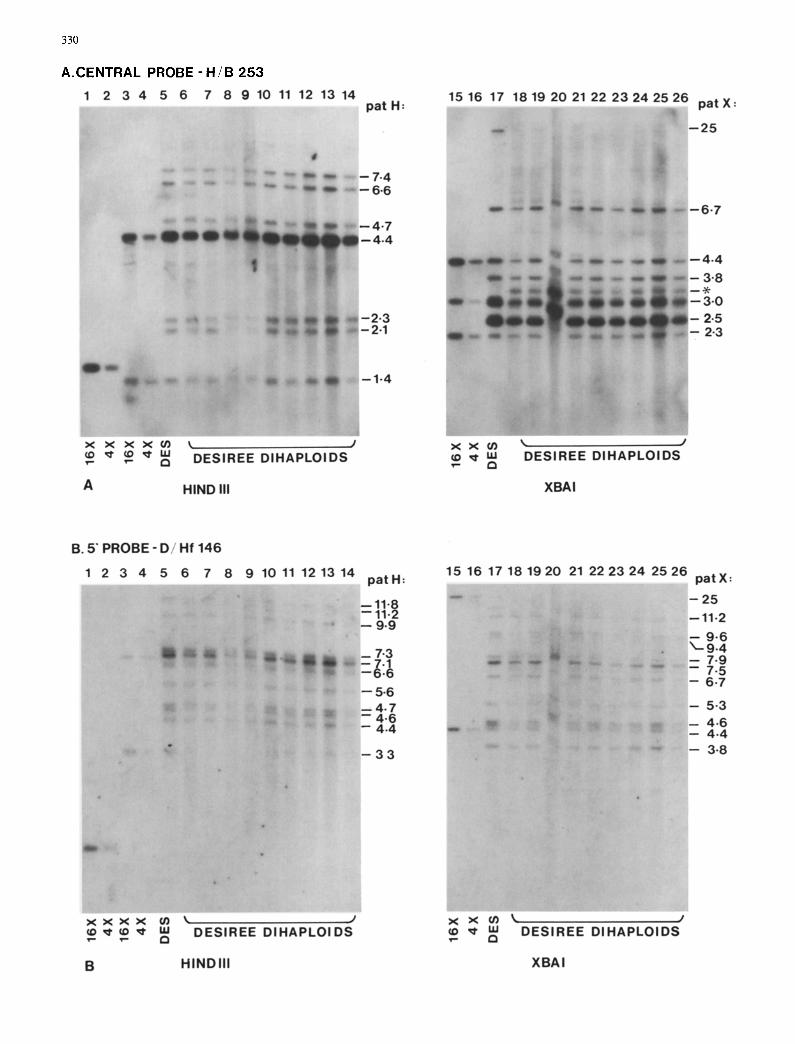
jo r pa ta t in D N A classes is that, i f an allelic var iant occurs with a single base pair change, thereby destroying a restric- t ion site, this would reduce the copy number from four to three for a designated band. Consequently, this could also cause the appearance of minor (or single copy) bands, which indeed was observed (some with segregation between dihaploids), but more clearly for d ihaplo id D N A than for cv. Desiree D N A . This would be as expected, since the equal D N A loadings mean that a single copy gene will
give rise to twice the signal in the d ihaploid D N A analysis (cf. Fig. 2). Thus, in our calculations we did not discrimi- nate between bands representing three or four pata t in D N A copies and consequently did not include bands apparent ly representing single copy pa ta t in D N A . I t is noted that the hybridising band pa t X* (Fig. 2A) in the d ihaploid D N A s does not appear in cv. Desiree D N A , and pa t X25 is present at reduced intensity in the d ihaploid DNAs . The reason for this result is not clear; no such differential hybr idisat ion

329
was observed for HindIII digested DNA with the 5' and central probes or with 5' cDNA probes.
Isolation of patatin genomic clones
To enable a more detailed analysis of the heterogeneity at the molecular level between individual members of the patatin gene family, a library of Desiree genomic DNA was established in bacteriophage lambda EMBL4. Screen- ing of approximately 2 x 105 recombinant phages of the unamplified library with patatin cDNA clone pGM203 (Mignery et al. 1984) gave 42 hybridising plaques. Ten of these clones chosen at random were purified and character- ised by restriction endonuclease mapping as shown in Fig. 3. The orientation and extent of the sequences with patatin homology was determined by Southern blot hybrid- isatiou experiments in which patatin 5' and 3' specific DNA fragments of eDNA clone pGM01 were used as probes. Based on similarities of restriction sites, both within and outside regions of patatin homology, these genomic clones were classified into 5 groups of related clones: (1) LPOT1 and LPOT4; (2) LPOT3, LPOT27, LPOT37 and LPOT43; (3) LPOT5 and LPOT23 ; (4) LPOT2 and (5) LPOT6. None of the clones contained more than one DNA region with patatin related sequences. Despite attempts to differentiate between these groups on the basis of variation in stringency of hybridisation conditions with patatin cDNA sequences from pGM01, no differential signals were obtained (data not shown). This suggested very high sequence homology between these clones within their coding DNA, and con- firmed the results of hybridisations of pGM01 with total Desiree DNA described above. Subsequently, DNA frag- ments containing patatin homology from all of the genomic clones were subcloned into pUC and M13mp vectors for further analysis with the exception of LPOT4, which, due to the particular cloning event of genomic DNA into 2EMBL4, did not contain 5' patatin DNA sequences.
5' truncated patatin pseudogenes
One group of clones with patatin DNA related sequences (LPOT3, 27, 37 and 43) showed remarkably similar restric- tion endonuclease maps, despite the fact that the clones clearly differed in insert size and orientation, which proved that they originated from independent cloning events. Only LPOT27 and LPOT43 showed minor but clear differences in their restriction endonuclease maps, positively revealing their distinct origin from within the Desiree genome. Com- parison of migration positions of restriction endonuclease HindIII and XbaI fragments that contained patatin DNA homology from the purified clones with similar fragments from total Desiree DNA, demonstrated that the cloned DNA corresponded with the single major HindIII band pat H4.4 and the major XbaI band pat X3.0 (Fig. 2). It is therefore likely that the clones represented members of a major group of at least 24 very closely related amplified sequences highly homologous to patatin DNA. A second characteristic of all four clones was that they showed no homology with 5' specific patatin eDNA probes, such as a 146 bp DraI-HinfI DNA fragment from cDNA clone pGM01 which corresponded to exon 1 DNA, and various DNA sequencing primers including ones specific for 5' un- transcribed flanking DNA.
The location of the discontinuity of the homology with
patatin DNA was found by comparison of the nucleotide sequence determined for two contiguous restriction endonu- clease fragments from LPOT3 (a 2.1 kb KpnI-HindIII and a 0.9 kb KpnI fragment; see Fig. 3) with published sequence data for patatin genomic clone pgT5 (Rosahl et al. 1986b). The 2.1 kb KpnI-HindIII fragment from LPOT3 contained exon 2, intron 2 and exon 3 sequences showing complete conservation of size and organisation relative to pgT5, ex- cept that intron 2 of LPOT3 was approximately 700 bp compared with 337 bp for pgT5. Within approximately 1100 bp of the 5' border of exon 2 of LPOT3, extensive homology was detected with the 389 bp intron 1 sequence of pgT5. This homology terminated 38 bp upstream of the 2.1 kb KpnI-HindIII fragment of LPOT3 within the contig- uous 0.9 kb KpnI fragment. The point of complete sequence divergence between LPOT3 and pgT5 corresponded to a position 12 bp downstream of the 3' border of exon 1 from pgT5. Furthermore, sequence comparison did not reveal additional minor reactions or weak homology between pa- tatin exon 1 sequences and DNA in the 0.9 kb KpnI frag- ment of LPOT3. This confirmed the absence of exon 1 ho- mology detectable via hybridisation. Thus, it seems unlikely that in the remaining 3-4 kb of DNA in LPOT3, upstream of the point of divergence, additional patatin related se- quences are present. We did, however, find homology in this region of LPOT3 DNA (as in LPOT 27, 37 and 43) with a 750 bp DraI fragment derived from 5' flanking DNA of genomic patatin clone LPOTI (Twell and t o m s 1987) extending 740 bp upstream of its transcriptional start site. Although this hybridising DNA was not further examined, the finding was consistent with the generation of a deletion in the 5' region of the patatin clones, thereby classifying them almost certainly as pseudogenes.
5' flanking regions of patatin DNA in LPOT1, 2, 5, 6 and 23
The DNA sequence of possible 5' regulatory signals and N-terminal coding DNA for the putative patatin genes in LPOTI, 2, 5, 6 and 23 was determined from subcloned DNA fragments, mainly using patatin DNA specific se- quencing primers. These DNA sequence data are summa- rized in Fig. 4A. Included is the previously established DNA sequence at the 5' end of the patatin gene from lambda clone LPOT1, which has been shown to have a functional 5' end (Twell and t o m s 1987). The DNA se- quence data for the other clones show perfect conservation of TATA and CAAT box homologies at identical positions - 2 5 and - 6 3 relative to the start of transcription as found in LPOT1. LPOT2, 5 and 23 also had the previously identi- fied 8 bp GC-rich repeats at position - 124 and - 161 and the 26 bp AT-rich repeats at position -2 2 0 and -260, although LPOT5 did not contain the most 5' copy of the AT-rich repeat due to an apparent 58 bp deletion in this region. In fact, LPOT1, 2, 5 and 23 are very homologous for all the Y-flanking DNA sequenced (up to position - 344 from the site of initiation of transcription). LPOT6, in con- trast, shows complete divergence upstream of position - 79, Thus, based on 5' sequence data alone, LPOT6 appears to represent a divergent subclass of patatin related DNA, confirming the separate classification based on differences detected using restriction endonuclease mapping (Fig. 3). Furthermore, LPOT6 has at the position of the normal ATG translation initiation eodon a GTG triplet which, if
330
A.CENTRAL PROBE-H/B 253

331
I 2 k b I ATG TAA
I ' ' , , , , , - . . . . . . . . . . .
XX X EHH H B E H E
LPOT 3 ,~N-,NWJI L ~ I~( I I, X
. E . E, . l . LPOT 27 ~ . . , . ~ I~ " KX
E H E H E LPOT 37 .vw~,,,~ I~ k~( x
E H E H B/HE E/B ,,~ I II LPOT 43 '.~.,,N.~ K KX X X
E/B B H i H E H e/a LPOT5 ~ ? XK X
E/B H B H E EIB LPOT 23 ~ , , ,
K I( " XK L P O T 2 ~ H H BH IE , f~ H B ~ , , , / v v ~ v v
K H E
, ' J . . . . // . . . . LPOT6 ~ I~ ~ t~ E x ;~
5' ) 3' Patat in
Fig. 3. Restriction endonuclease maps and patatin DNA homology within cv. Desiree patatin genomic clones. The solid bars show the maximum extent of homology with patatin cDNA sequences as determined by Southern blot hybridisation analysis. For LPOTI, the position of the ATG translational initiation codon and the TAA translational termination codon are shown, as determined by DNA sequence analysis. The orientation of patatin sequences is shown below the restriction maps (5' --* 3'). Restriction endonuclease sites are indicated as single letter abbreviations: B, BamHI ; E, EeoRI ; H, HindIII ; K, KpnI ; X, XbaI
the LPOT6 gene is functional, would shift expected initia- tion of translation 36 bp downstream to the next, in-frame A T G codon.
Computer comparison of sequence homology within 414 bp of 5' flanking D N A (314 bp untranscribed 5' D N A plus 100 bp of untranslated leader DNA) in LPOT1, 2, 5 and 23, showed similar relatedness of D N A in LPOTI with LPOT5 (95%), LPOT2 (91%) and LPOT23 (90%) as between LPOT23 and LPOT5 (94%) or LPOT2 (87%). However, the extent of homology between these regions in LPOT5 and LPOT2 was only 77%.
The sequence data is therefore consistent with the results from hybridisation. LPOT6 is obviously a distinct represen- tative of a subgroup with probably four members, identified in HindI I I and XbaI digested Desiree D N A by a patatin central probe as pat H1.4 and pat X4.4, respectively (Fig. 2A). Similarly, the divergence between LPOT2 and LPOT5 5' sequences confirmed that they were members of different patatin subclasses (with probably 4 and up to 24 members respectively) hybridising to pat H6.6 or pat H7.4 and pat X25 or to pat H4.7 and pat X.25, respectively, using the same probe.
Class I and class H patat in genes
A complementary way of classifying individual members of the patatin multigene family, rather than by structural differences, is by biological differences (i,e. expression). Based on the presence or absence of a 22 bp insert just upstream of the translational initiation codon, Pikaard et al. (1987) discriminated between expressed patatin genes without such an insert (class I genes) and those with the insert (class II genes). From lambda cloning and D N A se- quencing, examples of both class I and class II genes have been isolated (Bevan et al. 1986; Rosahl et al. 1986b). How- ever, amongst the patatin c D N A clones isolated so far there are only clones without the insert (class I; Mignery et al. 1984; Nakamura et al. 1986; Rosahl et al. 1986b). The oc- currence of such an insert in patatin related D N A of cv. Desiree was examined by hybridisations using two synthetic oligonucleotide probes. One probe, I29, was a 29 bp oligo- nucleotide specific for insert D N A and its sequence was determined from patatin c D N A clone pPOT301 (Fig. 4B). The second probe, $27, was a 27 bp oligonucleotide specific for D N A sequence spanning or surrounding the insert
Fig. 2A, B. Southern blot analysis of patatin DNA in potato using short patatin cDNA sequences as probes. A Autoradiographs resulting from hybridisation of Southern blots with the 32p-labelled 253 bp HindIII-BamHI fragment (H/B 253) isolated from the central coding DNA patatin cDNA clone pGM01 (Mignery et al. 1984). B Autoradiographs resulting from hybridisation of the same Southern blots shown in A (following washing to remove the probe H/B253) with the 146 bp DraI-Hinfl fragment (D/Hf 146) from the 5' untranslated and N-terminal coding region of pGM01. Lanes 5 14 and 17-26 contained HindIII and Xbal digested plant DNA, respectively. Lanes 5 and 17, Desiree DNA, lanes 6-14 and 18-26 dihaploid DNA in the following order: DDH1, 4, 5, 6, 7, 8, 10, 11 and PDH528. Lanes 1 and 2, 16 and 4 copy reconstructions. Lanes 3 and 4, 16 and 4 copy reconstructions containing HindIII digested DNA from patatin genomic clones LPOT2, LPOT5 and LPOT6. Internal hybridising restriction endonuclease fragments are shown for A LPOT5, 4.4 kb; LPOT6, 1.4 kb and B LPOT2, 3.3 kb; LPOT5, 7.1 kb, Similarly, lanes 15 and 16 contained 16 and 4 copy reconstructions of XbaI digested LPOT6, LPOT23 and LPOT43 DNA with internal hybridising fragments of A LPOT6, 4.4 kb; LPOT23, 2.3 kb; LPOT43, 3.0 kb and B LPOT6, 4.4 kb. The hybridising DNA fragment sizes are shown to the right of each autoradiograph, each band being designated pat H:x or pat X:x (where x is the fragment size in kbp) depending on the restriction endonuclease used; H, HindIII; X, XbaI

332
LPOT 1 LPOT 2 LPOT 5 LPOT 6 L P O T 2 3 consen
#
TTTTGAGAAA GATTTGAGGG ATaGTTATAT CAGgAATCAA TCTAAGCATT CCTAATACAT . cGAAATTC ATGGTATTAG TAATGCATGC ATTATTaTTT-245 TTTTGAGAAA GATTTGAGGG ATaGTTATta t tcatgTaAt TCTAAaCATT CoTAATACAT TCtGgAATTC ATGGTATTAG TAATGCATGC ATTtTaTTTT TTTTGAGAAA GATTTGAGGG ATEGTTATAT CAGTAATCAA TCTAAGCATT CCTAATACAT . . . . . . . . . . . . . . . . . . . . = . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . i . . . . . . . . . . . . . . . . . . . . TTTTGAGAAA GATTT~AGGG ATtGTTATAT CAGTAATCAA TCTAAGCATT CCTAAT..ca TCoaAAcTTC ATGGTATTAG "AATGCATGC ATTATTTTTT TTTTGAGAAA GATTTGAGGG AT-GTTATAT CAGTAATCAA TCTAAGCATT CCTAATACAT TC-GAAATTC ATGGTATTAG TAATGCATGC ATTATTTTTT
LPOT 1 LPOT 2 LPOT 5 LPOT 6 L P O T 2 3 consen
CTAATA~TAT TACACAAAGT ITAATGCATAT ATTATTTTTT CTgAT~CACT CTATCAAAEG ACTCCTTatT G T G T T A G c C ~ C C AACTATAAAT -145 tTAATA~TAT TACACAAAGT ITAATGCATAT ATTATTTTTT CTAATA~ACT CTATCAAACG ACTCCTTGET GTGTTAGcOG GGCCCA~ACC AACTATAAAT . . . . . . ~ . . . . . . . . . . . . ~ IaAATtCATAT ATTATTTTTT CTAATA~ACT CTATCAAACG ACTCCTTGcT GTGTTAG~C~ ~CCCA~CC AACTATAAAT . . . . . . ~ . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . I . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . !. .GaaaAg~aa AAooAaAAca CTAATA~TAT TACACAAAGT ITAATGCATAT ATTATTTTTT CTAATA~ACT CTActAAACG ACTCCTTGcT GTGT..atC~ GcCCaAa~CC AACTATAAtT CTAATA~TAT TACACAAAGT ITAATGCATAT ATTATTTTTT CTAATA~ACT CTATCAAACG ACTCCTTG-T GTGTTAG-E~3 GGCCCA~ACC AACTATAAAT
LPOT 1 LPOT 2 LPOT 5 LPOT 6 L P O T 2 3 CORSet;
LPOT 1 LPOT 2 LPOT 5 LPOT 6 LPOT23 consen
TCACTTTCCT TCAAc~ . . . . GGCCCAC~A TCCACGTGTC AAAaTTTGAA cGGATTcTCA TACAAC. TTT TCACTTTCCr TCAATT~ . . . . GGCCCAC~A TCCACGTGTC AAAaTTTaAA cGGATTTTCA TACAACTTTT TCACTTTtCT TCAAT~G . . . . GGCCCAC~A TCCACGTGTC 9AAtTTTGAA tGGATTTTCA TACAACTTTT TCACaaTCtg aCAgrT~cgo tGcaaagt~A gtgAgGTaat AAAcaTcact aatAgacatc actAACTTTT cactTTcCCT TCAAT~G . . . . GaCCCAC~A TCCACGTGTC gAAtTTTGAA tGGATTTTCA TACAACTTTT TCACTTTCCT TCAATT~ . . . . G GCCCAC~A TCCACGTGTC AAA-TTTGAA -GGATTTTCA TACAACTTTT
+1 CTCAA. CTtG TTTACGTGA~ TATATATACC ATGC.TTGTT ATA~TTCAA AGCACCCCcA AAAAAAATcA CTCAA.CTAG TTTACGTGc~ TATATATA~C ATGCETTGTT ATAT~cTCAA AGCAC . . . . . . . . . . . . . cA CTCAAcCTAG TTTACGTGA~ TATATATA~C ATGC. TTGTT ATAT~TTCAA AGCA. CCC~A AAAAAAATTA C. CtAaCTtG TTTACGTGcE TATATATA~C AcGC. TTGTT ATAT~cTCAA AGCA...tcA AcAAAAtTTA CTCAA.CTAG TTTACGTG~ TATATATA~C ATGC. TTGTT ATA~GTTCAA AGCA.CCC~A AAAAAAATTA CTCAA-CTAG TTTACGTGA~ TATAr°TAlC ATGC-TTGTT ATAT~TTCAA AGCACCCC-A AAAAAAATTA
ATTGGTTATG ~AAACTCAA AAT~AAATTT - 4 5 ATTGGTTATG T~AAACTCAA AA~AAATTT ATTGGTTATG T~AAACTCAA AA~AAAhTTT ATTGGTTATG ~CAAACTCAA AA~AAAATTT ATTGGTTATG TFAAACTCAA AA'II~AAATTT ATTGGTTATG T~AAACTCAA AA~AAATTT
AAAACACTTT GAGATATTAG TTTTTATTtA +56 AcAAaAtTT~ aAcAaATTAG TTTTTATTAA AAtACACTTT GAGATATTAG TTTTTATTAA AAAACACTTT GAGATATTAG TTTTTATTAA AAtACACTTT GAGATATTAG TTTTTATTAA AAAACACTTT GAGATATTAG TTTTTATTAA
LPOT I TTATAATCTG CAAA~CA ACTAgTAAAT CTTTTtTAAT TTIAtTTTTT ATGATATTAG CAACTACTAG TTC +130 LPOT 2 TTATAATCTG CAAA~TG~CA ACTACTAAAT CTTTTcTAAT TTTAATTTTT ATGATGTTAG ~AACTACTAc TTC LPOT 5 TTATAATCTG CAA~ATG~CA ACTACTAAAT aTTTTacAAT TTTAATTTTT AT~AT~TTAG CAACTACTAG TTC LOOT 6 TTATAATCTG CAA~gTG~CA ACTACTAAAT CTTTTtTAAT TTTAATTTTT ATGATATTAS CAACTACTAG TTC LPOT23 TTATPATCTG GAA A~AT~GCA ACTACTAAAT aTTTTacAAT TTTAATTTTT ATGATGTTAG CAACTACTAG TTC oonsen TTATAATCTG CAA AA~GCA ACTACTAAAT CTTTT-TAAT TTTAATTTTT ATGATGTTAG CAACTACTAG TTC 3' A
5 t 3 I
2 1 b p I N S E R T • . . . . . .
° oT 300 . . . . . . . . TorT o r ArTOo T+OrCT CAO COACr cr AO ATr ocoo OA r O O OT AAC C AOT ' I I I I ~ ' I I I ~ ' , ' , I ~ ~ l ~ I I ~ I : I ~ I I l l l I I I I I I l l I I I I I l I I I I I I l l l I I l | I I I l l I I I I I I I I I I I l I I I I l l l l
pPOT 301 CACTTTG~ATATTA~TTTTTATTAATTA~AATCTGCAAI~?TG,~CAACTA CTAAATATTTTACAATTTTAATTTTTATGATGTTAGCAACTACTAGTTC
pPOT 302 AAAAACACTTTGA . . . . . . . . . . . . . . . . . . . . . °CATTTGCAAA~C AACTACTAAATCTGTTTTAGTTTTAATTTTTATGATATTAGCAACTACTAGTTC
B Fig. 4A, B. DNA sequences of the 5' flanking and N-terminal coding regions of cv. Desiree patatin genomic clones and cDNAs. A 5' flanking and N-terminal coding sequences of genomic clones aligned, and gaps (shown as dots) introduced to maximise homology. Lower case letters indicate deviations from the derived consensus (consen) for the five clones. Features indicated: transcriptional initiation site (+~, see Rosahl et al. 1986b; Bevan et al. 1986); boxed sequences: ATG translational initiation codon, TATA and CAAT box homology and 86 bp GC-rich and 26 bp AT-rich direct repeats B 5' untranslated leader and N-terminal coding sequence of patatin eDNA clones pPOT300, 301, and 302. The translational initiation codon and the 19-21 bp insert sequence (21 bp insert) found in class II patatin eDNA clones (pPOT300 and 301) are boxed. A gap is introduced into the 5' untranslated sequence of pPOT302 to maximise homology and show the relative position of the 21 bp insert
D N A and its sequence was determined from patatin e D N A clone pPOT302 (Fig. 4B).
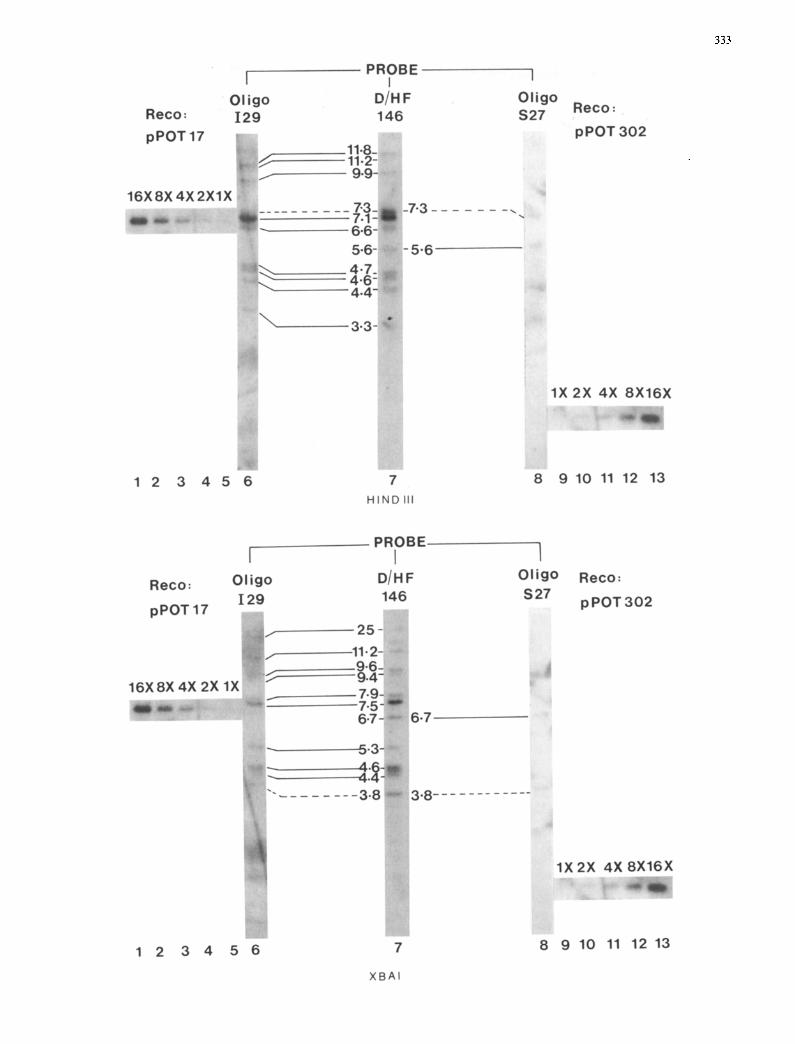
The hybridisation patterns with the class II probe I29 (Fig. 5) were essentially identical to that obtained with the 5' specific 146 bp DraI-HinJI probe described above; but no hybridisation was observed to pat H5.6 and pat X6.7 and the relative intensities of pat H7.3 and pat X3.8 were
significantly reduced. Comparison with reconstruction lanes also gave comparable estimates for copy numbers. This suggested that the vast majority (approximately 48-54 copies) o f full length Desiree patatin D N A contains the insert and it is present in all of the subclasses with multiple members o f full length patatin D N A , except sub- classes pat H5.6 and pat X6.7, each representing approxi-
Fig. 5. Southern blot analysis of class I and class II patatin DNA in cv. Desiree using class-specific oligonucleotide probes. The autoradio- graphs shown were obtained following hybridisation of the 32p-labelled class I ($27) or class II (I29) specific patatin gene oligonucleotide probes, or the 146 bp DraI-HinfI fragment (D/HF146) from the 5' untranslated and N-terminal coding region of patatin eDNA clone pGM0I to Southern blots of Desiree DNA. The nucleotide sequences of $27 and I29 were complementary to the coding strands in the 5' untranslated leader DNA of patatin eDNA clones pPOT302 and pPOT300 respectively. Their DNA sequences were as follows: $27, 5'-CATTTTGCAAATGTTCAAAGTGTTTTT-3'; I29, 5'-GCAGATTATAATTAATAAAAACTAATATC-3'- Top: Lanes 6, 7 and 8; Desiree DNA digested with HindIII and hybridised with I29, D/HF146 and $27, respectively. Bottom: Lanes 6, 7 and 8; Desiree DNA digested with XbaI and hybridised with I29, D/HF146 and $27, respectively. Lanes 1 5, copy number reconstructions containing class II patatin genomic subclone pPOT17 (7.6 kb BamHI-HindlII fragment from LPOT23 containing 5' flanking and N-terminal coding patatin sequences in pUC18) DNA digested with HindIII respresenting 16, 8, 4, 2 and 1 copy per tetraploid potato genome. Similarly, lanes 9-13 represent copy number reconstructions containing EeoRI digested class I patatin eDNA clone pPOT302. The sizes of hybridising fragments in kbp are shown adjacent to lane 7; corresponding bands of similar relative intensity as in lane 7 are linked by solid lines and those with reduced relative intensity are linked by dashed lines
333

334
mately 4 copies. Hybridisation of class I probe $27 to a Southern blot with HindIII and XbaI digested DNA showed it to be present in Desiree DNA, but at a low copy number of only 6-8 copies per tetraploid genome. Two hybridising fragments were reproducibly detected in two independent hybridisation experiments in the XbaI digested DNA corresponding to pat X6.7 and pat X3.8 (see Fig. 5, bottom), the latter with reduced intensity, which were not detected (pat X6.7) or detected at reduced intensity (pat X3.8) with I29. The HindIII pattern with $27 gave two clear bands corresponding to pat H7.3 and pat H5.6 (see Fig. 5, top), the former with reduced relative intensity, which were not detected (pat H5.6) or detected at reduced relative intensity (pat H7.3) with I29. These results strongly suggest that pat H5.6 or pat X6.7 detected using $27 and the patatin 5' 146 bp Dral-Hinfl probe, contain exclusively class I patatin DNA, each with about 4 copies per tetra- ploid genome; and also that pat H7.3 and pat X3.8 detected using the same probes contain both class I and class II pata- tin DNA.
An initial demonstration that both classes of genes are expressed in Desiree, came from cloning and sequence anal- yses of three near full-length patatin cDNA clones gener- ated from tuber RNA. Two clones (pPOT300 and pPOT301; class II) contained the insert and one did not (pPOT302, class I). The 5' DNA sequences of the three cDNA clones are shown in Fig. 4B.
Discussion
Segregation of patatin DNA
Southern blot analysis of patatin related sequences in geno- mic DNA of potato cv. Desiree and in nine dihaploid lines derived from cv. Desiree by prickle pollination with S. phur- eja pollen, gave near identical hybridisation patterns (Fig. 1). Assuming random segregation of patatin loci over egg cells induced to develop into the dihaploid plants, this suggests little allelic variation between these patatin loci on each of the four haploid genomes. Such apparent allelic homogeneity of patatin D N A is perhaps surprising, given that potato is known to maintain a high degree of heterozy- gosity and that patatin hybridisation patterns for different varieties show significant variation (Bevan et al. 1986; D. Twell unpublished observations). On the other hand, there is no reason to doubt the assumed random segregation. If not random or nearly random it would have to be partic- ularly extreme or total in order to provide an alternative explanation for the observed hybridisation results. This is difficult to reconcile with the substantial phenotypic vari- ability among the dihaploid plants and the detected segrega- tion of minor bands (Fig. 1) representing low, perhaps sing- le copy, patatin D N A loci. It will be of interest, however, to examine the segregation of patatin loci more closely by other genetical or molecular approaches, such as analysis of dihaploid or monohaploid derivatives isolated via anther culture using patatin and other gene probes, analysis of progeny from crosses between Desiree and other tetraploid potato cultivars, or in situ hybridisation of patatin probes to potato chromosomes, of which the feasibility has been demonstrated for very low copy number loci in tobacco (Mouras et al. 1987). Our data do not answer the question of whether the detected patatin sequences are dispersed over a number of chromosomes or clustered at one or more loci, the latter organisation has been shown for the Rubisco SSU and Cab binding protein gene families of pea (Polans
et al. 1985). None of the isolated genomic clones contained multiple patatin genes, indicating no extremely tight cluster- ings within the 15-20 kb fragments cloned. Such tight clus- tering has been noted for some members of plant multigene families, including the maize zein genes (Spena et al. 1983; Heidecker and Messing 1986) soybean leghemoglobin genes (Lee et al. 1983), petunia Rubisco SSU genes (Dean et al. 1985a), Arabidopsis Cab genes (Leutwiler et al. 1986), to- mato wound inducible protease inhibitors I and II (Lee et al. 1986), and Phaseolus glutamine synthetase genes (B. Forde, personal communication).
Patatin DNA copy number
From our combined molecular and genetical analysis we estimated that there are between 64-72 patatin DNA copies in the tetraploid genome of cv. Desiree, giving a gene family size of 16-18 members per haploid genome. This estimate was based in part on the assumption that all of the patatin DNA copies should be detectable using the central 253 bp HindIII-BamHI hybridisation probe (Fig. 2A and Table 1). Due to the presence of multiple comigrating patatin DNA subclasses detected using this probe (i.e. pat H4.4, pat X3.0 and pat X2.5; Fig. 2A), a more accurate quantitation of total copy number was not possible. The less pronounced comigration of patatin DNA subclasses detectable using the 5' 146 bp DraI-HinfI probe (Fig. 2B) facilitated more accurate copy number estimates, although none of the 5' truncated patatin pseudogenes (i.e. LPOT3, 27, 37 and 43) hybridise to this probe. Thus, the discrepancy between the estimates of total patatin DNA copy number summarised in Table 1, using both the central (64-72 copies) and 5' (56-60 copies) probes, suggests that the 5' truncated pseu- dogenes account for between 4-16 patatin DNA copies per tetraploid genome of cv. Desiree. The patatin gene family size appears to differ between different potato cultivars, as estimates of 5-10 patatin family members have been made for cv. Maris Piper (Bevan et al. 1986) and we have also observed copy number variation in a number of other cultivars (unpublished data). Comparison with other plant storage protein gene families places patatin among the mod- erately sized families such as the barley B hordein genes with 11-13 members per haploid genome (Kreis et al. 1983) in contrast with the larger zein gene family of maize with about 120 members (Viotti et al. 1979).
Patatin DNA diversity and classification
Although patatin DNA family members showed little allelic variation based on segregation patterns in cv. Desiree di- haploids, the different allelic groups were often clearly dis- tinguishable on the basis of their restriction fragment length of polymorphism (RFLP). The use of restriction endonucle- ases HindIII and XbaI and different patatin cDNA regions as probes (Fig. 2) showed various types of structural differ- ences between cloned gene members. These included differ- ences in percentage DNA sequence homology within con- served analogous 5' flanking regions (from 96% between LPOTI and LPOT5 to 77% between LPOT2 and LPOT5; Fig. 4A), differences in restriction endonuclease maps (for all clones except LPOT3 and LPOT37) and the occurrence of complete DNA sequence divergence at analogous posi- tions (such as in 5' flanking DNA of LPOT6 compared with LPOT1, 2, 5 and 23 and the apparent deletion of exon I plus 5' regulatory DNA in clones LPOT3, 27, 37 and 43).

335
Thus, the group of clones LPOT3, 27, 37 and 43 appear to represent four pseudogenes of a subclass of approximate- ly 20-24 patatin gene family members which were identified as pat X3.0 and pat H.4.4, using the central HindIII-BamHI fragment of patatin cDNA clone pGM01 as a gene probe. Whether these apparent pseudogenes are indeed non-ex- pressed or under control of other expression signals is not known. This can be examined by transformation experi- ments using translational fusions; but it has to be kept in mind that steady-state analysis of RNA from tubers and other plant organs showed transcripts with patatin DNA homology only at the size of patatin in mRNA (Mignery et al. 1984; Rosahl et at. 1986a). Wether any of the other cloned genes are pseudogenes, expressed or non-expressed functional genes is also difficult to predict from their se- quence alone. For LPOT1, we demonstrated in transforma- tion experiments that a 3.8 kb Y-flanking DNA fragment directed tuber-specific expression of the reporter gene cat, which strongly suggested that LPOT1 contained an active patatin gene. The high degree of DNA homology between Y-flanking sequences in LPOT1 and LPOT2, 5 and 23 sug- gests that these latter clones may also contain functional genes. These genes, however, are all representatives of dif- ferent allelic groups, identified by RFLP mapping. The most clearly distinct clone with identified 5' flanking DNA, LPOT6, also differed in known expression signals, such as the first correctly positioned ATG translation initiation co- don. LPOT6 therefore may contain a patatin pseudogene similar to two genomic clones described by Pikaard et al. (1986). Both these clones showed a high degree of homology to patatin cDNA and at least some LPOTI Y-flanking DNA; but also contained single nucleotide changes result- ing in frame shift mutations. Such non-functional genes are common within large multigene families, several being identified in plants including examples within the soybean leghaemoglobin (Brisson and Verma 1982) and the pea le- gumin (Bown et al. 1985) gene families.
Class I and I I patatin genes
A further distinction can be made concerning the classifica- tion of individual members of the patatin gene family based on the presence or absence of a 22 bp insert sequence, an apparent expression marker. The length of the insert is be- tween 19-22 bp depending on how ambiguities near the borders are interpreted and which genes are compared (cf. Fig. 3). The copy numbers of class I genes (without insert) and class II genes (with insert) in cv. Desiree are 48-54 and 6-8, respectively. The majority of class II genes and one class I gene subclass appear to show allelic homogenei- ty; although one patatin gene subclass contains representa- tives of both classes.
In other cultivars the relative number of insert-plus to insert-minus genes may be different. Pikaard et at. (1987) reported the isolation of more than 20 patatin genomic clones from cv. Superior and approximately one-half of the clones were of each type (G.A. Mignery, in prepara- tion). Rosahl et al. (1986b) examined one, and Bevan et al. (1986) two (identical) randomly chosen patatin clones from potato haploid line AM 80/5793 and Maris Piper, respec- tively. The first one was with and the second two both without an insert.
The importance of the 22 bp insert as an expression marker, or as a regulatory element, was suggested by steady-state RNA analysis of tuber and root RNA from
cv. Superior by primer-extension and 5' S1 nuclease map- ping (Pikaard et al. 1987). Fifty percent of tuber patatin RNA was with and 50% without the insert, but in roots, with only 1% of the tuber patatin RNA, all were with an insert. It remains to be seen whether similar expression is common for different stages of plant growth, how it com- pares for different varieties and what the contribution and regulation of expression is for individual gene members. Insight into the natural diversity of expression of endoge- nous patatin genes will be of use in transformation experi- ments aimed at analysis of patatin gene expression signals and their use in directing tuber specific expression of foreign genes.
More advanced studies in animal cells have shown that both cis- and trans-acting factors are involved in gene regu- lation, and that competition for trans-acting factors by en- dogenous genes may play an important role (Sch61er and Gross 1984; Dynan and Tjian 1985; Ptashne 1986; Mania- tis et al. 1987). This suggests that better characterisation of the host plant in transformation studies is likely to be of increasing importance, probably even more so in a genet- ically heterogenous plant such as potato. Because of the suitability of cv. Desiree in transformation studies with tet- raploid potato, we considered it useful to obtain at least some insight into the structural diversity and complexity of the patatin gene family, at present of interest in any potato variety, but our approach is particular for cv. De- siree.
Acknowledgements. We thank Ange[a Karp for kindly performing chromosome counts of dihaploid potato lines, Mike Burrell for valuable practical advice on cDNA cloning and members of the Biochemistry department for helpful discussions. DT was sup- ported by an AFRC Studentship.
References
Benton WD, Davis RW (1977) Screening Agt recombinant clones by hybridisation to single plaques in situ. Science 196:180-182
Bevan M, Barker R, Goldsbrough A, Jarvis M, Kavanagh T, Itur- riaga G (1986) The structure and transcription start site of a major potato tuber protein gene. Nucleic Acids Res 14:4625-4638
Bown D, Levasseur M, Croy RD, Boulter D, Gatehouse JA (1985) Sequence of a pseudogene in the legumin gene family of pea (Pisum sativum L.) Nucleic Acids Res 13: 4527-4528
Brisson N, Verma DPS (1982) Soybean leghaemaglobin gene fami- ly: normal, pseudo, and trucated genes. Proc Natl Acad Sci USA 79:4055-4059
Burrell MM, Twell D, Karp A, Ooms G (1985) Expression of shoot-inducing Ti-TL-DNA in differentiated tissues of potato (Solanum tuberosum cv. Maris Bard). Plant Mol Biol 5:213-222
Chen EY, Seeburg PH (1985) Supercoil sequencing: a fast and simple method for sequencing plasmid DNA. DNA 4:165-170
Dean C, Van Den Elzen P, Tamaki S, Dunsmuir P, Bedbrook J (1985a) Linkage and homology analysis divides the eight genes for the small subunit of petunia ribulose 1,5-bisphosphate carboxylase into three gene families. Proc Natl Acad Sci USA 82: 4964-4968
Dean C, Van Den Elzen P, Tamaki S, Dunsmuir P, Bedbrook J (1985b) Differential expression of the eight genes of the pe- tunia ribulose bisphosphate carboxylase small subunit multi- gene family. EMBO J 4:3055-3061
Dean C, Favreau M, Dunsmuir P, Bedbrook J (1987) Confirmation of the relative expression levels of the Petunia (Mitchell) rbcS genes. Nucleic Acids Res 15: 4655-4668
Devereaux J, Haeberli P, Smithies O (1984) A comprehensive set of sequence analysis programs for the VAX. Nucleic Acids Res 12:387-395

336
Dynan WS, Tjian R (1985) Control of eukaryotic messenger RNA synthesis by sequence-specific DNA-binding proteins. Nature 316:774-778
Feinberg AP, Vogelstein B (1983) A technique for radiolabelling DNA restriction endonuclease fragments to high specific activi- ty. Anal Biochem 132: 6-13
Frischauff AM, Lehrach H, Poustka A, Murray N (1983) Lambda replacement vectors carrying polylinker sequences. J Mol Biol 170: 827-842
Galliard T, Dennis S (1974) Isoenzymes of lipolytic acyl hydrolase and esterase in potato tuber. Phytochemistry 13: 2463-2468
Heidecker G, Messing J (1986) Structural analysis of plant genes. Annu Rev Plant Physiol 37:439-466
Helentjaris T, King G, Slocum M, Siedenstrang C, Wegman S (1985) Restriction fragment polymorphisms as probes for plant diversity and their development as tools for applied plant breed- ing. Plant Mol Biol 5:109-118
Hermsen JGTh, Verdenius J (1973) Selection from Solarium tubero- sum group Phureja of genotypes combining high frequency hap- loid induction with homozygosity for embryo-spot. Euphytica 22: 244-259
Huynh TV, Young RA, Davis RW (1985) Constructing and screen- ing cDNA libraries in 2gtl0 and 2gtll . In: Glover GM (ed) DNA Cloning, vol. I A practical approach. IRL Press, Oxford, pp 49-78
Karp A, Risiott R, Jones MGK, Bright SWJ (1984) Chromosome doubling in monohaploid and dihaploid potatoes by regenera- tion from cultured leaf explants. Plant Cell Tissue Organ Cul- ture 3:363-373
Kreis M, Shewry PR, Forde BG, Rahman S, Miflin BJ (1983) Molecular analysis of a mutation conferring the high lysine phenotype on the grain of barley (Hordeum vulgare). Cell 34:161-167
Lee JS, Brown GG, Verma DPS (1983) Chromosomal arrangement of leghaemoglobin genes in soybean. Nucleic Acids Res 11 : 5541-5553
Lee JS, Brown WE, Graham JS, Pearce G, Fox EA, Dreher TW, Ahern KG, Pearson GD, Ryan CA (1986) Molecular character- ization and phylogenetic studies of a wound-inducible protein- ase inhibitor I gene in Lycopersieon species. Proc Natl Acad Sci USA 83 : 7277-7281
Leutwiler LS, Meyerowitz EM, Tobin EM (1986) Structure and expression of three light-harvesting chlorophyll a/b-binding protein genes in Arabidopsis thaliana. Nucleic Acids Res 14:4051-4064
Maniatis T, Fritsch EF, Sambrook J (1982) Molecular cloning: a laboratory manual. Cold Spring Harbor Laboratory Press, New York
Maniatis T, Goodbourn S, Fischer JA (1987) Regulation of induc- ible and tissue-specific gene expression. Science 236:1237-1245
Miguery GA, Pikaard CS, Hannapel D J, Park WD (1984) Isolation and sequence analysis of cDNAs for the major potato tuber protein, patatin. Nucleic Acids Res 12:7987-8000
Mouras A, Saul MW, Essad S, Potrykus I (1987) Localization by in situ hybridization of a low copy chimaeric resistance gene introduced into plants by direct gene transfer. Mol Gen Genet 207: 204-209
Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassay with tobacco tissue cultures. Physiol Plant 15: 473-497
Nakamura K, Hattori T, Asahi T (1986) Direct immunological identification of full length eDNA clones for plant protein with- out gene fusion to E. eoli protein. FEBS Lett 198:16-20
Paiva E, Lister RM, Park WD (1983) Induction and accumulation of major tuber proteins of potato in stems and petioles. Plant Physiol 71 : 161-168
Park WD, Blackwood C, Mignery GA, Bermodson MA, Lister RM (1983) Analysis of the heterogeneity of the 40000 molecu- lar weight tuber glycoprotein of potatoes by immunological methods and by NH2-terminal sequence analysis. Plant Physiol 71 : 156-160
Park WD, Hannapel D J, Mignery GA, Pikaard CS (1985) Molecu- lar approaches to the study of the major tuber proteins. In: Li PH (ed) Potato physiology. Academic Press, London, pp 261-278
Pikaard CS, Mignery GA, Ma DP, Stark VJ, Park WD (1986) Sequence of two apparent pseudogenes of the major potato tuber protein patatin. Nucleic Acids Res 14:5564-5566
Pikaard CS, Brusca JS, Hannapel DJ, Park WD (1987) The two classes of genes for the major potato tuber protein, patatin, are differentially expressed in tubers and roots. Nucleic Acids Res 15:1979-1994
Polans NO, Weeden NF, Thompson WF (1985) Inheritance, orga- nization, and mapping of rbcS and cab multigene families in pea. Proc Natl Acad Sci USA 82:5083-5087
Ptashne M (1986) Gene regulation by proteins acting nearby and at a distance. Nature 322:697-701
Racusen D (1984) Lipid acyl hydrolase of patatin. Can J. Bot 62 : 1640-1644
Racusen D, Foote M (1980) A major soluble glycoprotein of po- tato. J Food Biochem 4:43-52
Rigby PW, Dieckmann M, Rhodes C, Berg P (1977) Labelling deoxyribonucleic acid to high specific activity in vitro by nick translation with DNA polymerase I. J Mol Biol 113:237-251
Rosahl S, Eckes P, Schell J, Willmitzer L (1986a) Organ-specific gene expression in potato: isolation and characterisafion of tuber-specific cDNA sequences. Mol Gen Genet 202 : 368-373
Rosahl S, Schmidt R, Schell J, Willmitzer L (1986b) Isolation and characterization of a gene from Solanum tuberosum encod- ing patatin, the major storage protein of potato tubers. Mol Gen Genet 203 : 214-220
Rosahl S, Schell J, Willmitzer L (1987) Expression of a tuber- specific storage protein in transgenic tobacco plants: demon- stration of an esterase activity. EMBO J 6:1155-1159
Sanger F, Nicklen S, Coulson A (1977) DNA sequencing with chain terminating inhibitors. Proc Natl Acad Sci USA 74: 5463-5467
Sch61er HR, Gross H (1984) Specific interaction between enhancer- containing molecules and cellular components. Cell 36:403- 411
Southern EM (1975) Detection of specific sequences among DNA fragments separated by gel electrophoresis. J Mol Biol 98 : 503-517
Spena V, Viotti A, Pirrotta V (1983) Two adjacent genomic zein sequences : structure, organization and tissue-specific restriction pattern. J Mol Biol 169: 799-811
Staden R (1980) Sequence data handling by computer. Nucleic Acids Res 8 : 3673-3694
Turner NE, Clark WG, Tabor G J, Hironaka CM, Fraley RT, Shah DM (1986) The genes encoding the small subunit of ribu- lose-l,5-bisphosphate carboxylase are expressed differentially in petunia leaves. Nucleic Acids Res 14:3325-3342
Twell D, Ooms G (1987) The 5'-flanking DNA of a patatin gene directs tuber specific expression of a chimaeric gene in potato. Plant Mol Biol 9:365-375
Uijtewaal BA, Huigen DJ, Hermsen JGTh (1987) Production of potato monohaploids (2n = x = 12) through prickle pollination. Theor Appl Genet 73 : 751-758
Viotti A, Sala E, Marotta R, Alberi P, Balducci C, Soave C (1979) Genes and mRNAs coding for zein polypeptides in Zea mays. Eur J Biochem 102:211-222
Webb KJ, Osifo EO, Henshaw GG (1983) Shoot regeneration from leaflet discs of six cultivars of potato (Solanum tuberosum subsp. tuberosum). Plant Sci Lett 30:1-8
Yanisch-Perron C, Vieira J, Messing J (1985) Improved M13 phage cloning vectors and host strains: nucleotide sequences of M13 mpl8 and pUC19 vectors. Gene 33:103-119
Communica ted by J. Schell
Received October 8, 1987




















