
StGA2ox1 is induced prior to stolon swelling and controls GA levels during potato tuber development
12
StGA2ox1 is induced prior to stolon swelling and controls GA levels during potato tuber development Bjorn Kloosterman 1,* , Christina Navarro 2 , Gerard Bijsterbosch 1 , Theo Lange 3 , Salome ´ Prat 2 , Richard G. F. Visser 1 and Christian W. B. Bachem 1 1 Graduate School Experimental Plant Sciences, Laboratory of Plant Breeding, Department of Plant Sciences, Wageningen University and Research Centre, PO Box 386, 6700 AJ Wageningen, The Netherlands, 2 Centro Nacional de Biotecnologı´a, Consejo Superior de Investigaciones Cientı´ficas, Campus Universidad Auto ´ noma de Madrid, c/Darwin 3, 28049 Madrid, Spain, and 3 Institute of Plant Biology, Technical University of Braunschweig, D-38106 Braunschweig, Germany Received 21 March 2007; revised 9 June 2007; accepted 25 June 2007. * For correspondence (fax +31 317 483457; e-mail [email protected]). Summary The formation and growth of a potato (Solanum tuberosum) tuber is a complex process regulated by different environmental signals and plant hormones. In particular, the action of gibberellins (GAs) has been implicated in different aspects of potato tuber formation. Here we report on the isolation and functional analysis of a potato GA 2-oxidase gene (StGA2ox1) and its role in tuber formation. StGA2ox1 is upregulated during the early stages of potato tuber development prior to visible swelling and is predominantly expressed in the subapical region of the stolon and growing tuber. 35S-over-expression transformants exhibit a dwarf phenotype, reduced stolon growth and earlier in vitro tuberization. Transgenic plants with reduced expression levels of StGA2ox1 showed normal plant growth, an altered stolon swelling phenotype and delayed in vitro tuberization. Tubers of the StGA2ox1 suppression clones contain increased levels of GA 20 , indicating altered GA metabolism. We propose a role for StGA2ox1 in early tuber initiation by modifying GA levels in the subapical stolon region at the onset of tuberization, thereby facilitating normal tuber development and growth. Keywords: GA 2-oxidase, potato tuber development, gibberellin metabolism. Introduction Potato (Solanum tuberosum) plants have the capability of producing underground storage organs called tubers. In general, tuberization is promoted by short days with the degree of response largely dependent on the genotype and physiological age of the plant (Ewing and Struik, 1992). Plants induced to tuberize produce a graft transmissible signal in the leaves that is transported basipetally to the growing stolon tip where it promotes tuberization (Gregory, 1956; Jackson et al., 1998). The exact nature of this trans- missible signal is still unknown but it is most likely based on a mixture of both inducing and inhibiting types of signals (Ewing and Struik, 1992; Jackson, 1999; Martinez-Garcia et al., 2001). Interestingly, there are several lines of evidence suggesting a similar regulatory pathway for both flowering and tuberization photoperiodic responses in plants (reviewed in Rodriguez-Falcon et al., 2006). Gibberellins (GAs) have long been implicated to have a regulatory role in potato stolon growth and tuber initiation (Booth, 1963; Ewing, 1987; Kumar and Wareing, 1974). Applications of biologically active GAs or inhibitors of GA biosynthesis have been shown to either delay or promote tuber formation under tuber-inducing conditions (Jackson and Prat, 1996; Vreugdenhil and Struik, 1989). Furthermore, several studies have reported the existence of cross-talk between the perception of day length and GA metabolism (Amador et al., 2001; Chen et al., 2003; Jackson et al., 2000). In the last decade, significant progress has been made in identifying the genes involved in GA metabolism (for review see Hedden and Phillips, 2000; Hedden and Proebsting, 1999; Lange, 1998). In potato, Northern blot analysis of three isolated GA20-oxidase genes showed expression of all three genes in both stolon and tuber tissue (Carrera et al., 1999), 362 ª 2007 The Authors Journal compilation ª 2007 Blackwell Publishing Ltd The Plant Journal (2007) 52, 362–373 doi: 10.1111/j.1365-313X.2007.03245.x
Transcript of StGA2ox1 is induced prior to stolon swelling and controls GA levels during potato tuber development

StGA2ox1 is induced prior to stolon swelling and controlsGA levels during potato tuber development
Bjorn Kloosterman1,*, Christina Navarro2, Gerard Bijsterbosch1, Theo Lange3, Salome Prat2, Richard G. F. Visser1 and
Christian W. B. Bachem1
1Graduate School Experimental Plant Sciences, Laboratory of Plant Breeding, Department of Plant Sciences,
Wageningen University and Research Centre, PO Box 386, 6700 AJ Wageningen, The Netherlands,2Centro Nacional de Biotecnologıa, Consejo Superior de Investigaciones Cientıficas, Campus Universidad Autonoma de
Madrid, c/Darwin 3, 28049 Madrid, Spain, and3Institute of Plant Biology, Technical University of Braunschweig, D-38106 Braunschweig, Germany
Received 21 March 2007; revised 9 June 2007; accepted 25 June 2007.*For correspondence (fax +31 317 483457; e-mail [email protected]).
Summary
The formation and growth of a potato (Solanum tuberosum) tuber is a complex process regulated by different
environmental signals and plant hormones. In particular, the action of gibberellins (GAs) has been implicated
in different aspects of potato tuber formation. Here we report on the isolation and functional analysis of a
potato GA 2-oxidase gene (StGA2ox1) and its role in tuber formation. StGA2ox1 is upregulated during the early
stages of potato tuber development prior to visible swelling and is predominantly expressed in the subapical
region of the stolon and growing tuber. 35S-over-expression transformants exhibit a dwarf phenotype,
reduced stolon growth and earlier in vitro tuberization. Transgenic plants with reduced expression levels of
StGA2ox1 showed normal plant growth, an altered stolon swelling phenotype and delayed in vitro
tuberization. Tubers of the StGA2ox1 suppression clones contain increased levels of GA20, indicating altered
GA metabolism. We propose a role for StGA2ox1 in early tuber initiation by modifying GA levels in the
subapical stolon region at the onset of tuberization, thereby facilitating normal tuber development and
growth.
Keywords: GA 2-oxidase, potato tuber development, gibberellin metabolism.
Introduction
Potato (Solanum tuberosum) plants have the capability of
producing underground storage organs called tubers. In
general, tuberization is promoted by short days with the
degree of response largely dependent on the genotype and
physiological age of the plant (Ewing and Struik, 1992).
Plants induced to tuberize produce a graft transmissible
signal in the leaves that is transported basipetally to the
growing stolon tip where it promotes tuberization (Gregory,
1956; Jackson et al., 1998). The exact nature of this trans-
missible signal is still unknown but it is most likely based on
a mixture of both inducing and inhibiting types of signals
(Ewing and Struik, 1992; Jackson, 1999; Martinez-Garcia
et al., 2001). Interestingly, there are several lines of evidence
suggesting a similar regulatory pathway for both flowering
and tuberization photoperiodic responses in plants
(reviewed in Rodriguez-Falcon et al., 2006).
Gibberellins (GAs) have long been implicated to have a
regulatory role in potato stolon growth and tuber initiation
(Booth, 1963; Ewing, 1987; Kumar and Wareing, 1974).
Applications of biologically active GAs or inhibitors of GA
biosynthesis have been shown to either delay or promote
tuber formation under tuber-inducing conditions (Jackson
and Prat, 1996; Vreugdenhil and Struik, 1989). Furthermore,
several studies have reported the existence of cross-talk
between the perception of day length and GA metabolism
(Amador et al., 2001; Chen et al., 2003; Jackson et al., 2000).
In the last decade, significant progress has been made in
identifying the genes involved in GA metabolism (for review
see Hedden and Phillips, 2000; Hedden and Proebsting,
1999; Lange, 1998). In potato, Northern blot analysis of three
isolated GA20-oxidase genes showed expression of all three
genes in both stolon and tuber tissue (Carrera et al., 1999),
362 ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd
The Plant Journal (2007) 52, 362–373 doi: 10.1111/j.1365-313X.2007.03245.x

yet little is known about the corresponding enzyme activities
in these tissues. Over-expression of StGA20ox1 results in
plants that show delayed tuber formation, whilst suppres-
sion clones are semi-dwarfed and tuberize earlier under
short-day conditions (Carrera et al., 2000). Whether biolog-
ically active GAs or the precursors themselves are part of the
transmissible signal or play a role in the production,
sensitivity or transport of the tuberization-promoting or
-inhibiting signals within the plant remains to be resolved.
The identification of novel genes regulating GA levels
through transcriptional control (StBEL5, POTH1) or altering
GA sensitivity (PHOR1) in potato plants, which were shown
to affect tuberization onset (Amador et al., 2001; Chen et al.,
2003; Rosin et al., 2003), indicates the existence of a complex
mechanism for controlling GA levels during the formation of
potato tubers.
Another important line of evidence linking GAs to the
regulation of tuber development is the observation of a
decrease in the levels of GA1 within the subapical region of
in vitro grown microtubers prior to visible swelling (Xu et al.,
1998a). When potato plants are induced to tuberize, stolon
growth ceases and the reduction in the levels of GAs is
thought to result in the longitudinal reorientation of the cell
microtubules, allowing lateral cell expansion and division
(Fujino et al., 1995; Shibaoka, 1993; Xu et al., 1998b). The
regulatory mechanism by which levels of bioactive GAs are
reduced prior to stolon swelling is, however, still unknown.
In many plant species, bioactive GAs are inactivated
through 2b-hydroxylation to produce inactive forms of
GAs. This step is catalysed by a member of the 2-oxoglut-
arate-dependent dioxygenase family, GA 2-oxidase (Thomas
et al., 1999). Gibberellin 2-oxidase has also been shown to be
able to react with precursors of active GAs, i.e. GA20 (Martin
et al., 1999). Similar to the GA 20-oxidase and GA 3-oxidase
gene families, the GA 2-oxidase gene family consists of
multiple members with often overlapping functions. In
recent years, a large number of GA 2-oxidases have been
identified in various plant species and are considered to be
important regulators of GA metabolism (Hedden, 2001).
In a previous experiments we found strong upregulation
of a potato GA 2-oxidase family member after the switch
from long- to short-day conditions within the stolon tip
(Kloosterman et al., 2005). Here we present data on the
analysis of the expression pattern of the isolated potato GA
2-oxidase (StGA2ox1) and its role in the process of tuber
formation.
Results
StGA2ox1 is upregulated prior to tuber development
and is located within the subapical stolon region
Large-scale gene expression analysis of potato tuber
development has shown differential expression of many
genes during the early stages of tuber formation (Kloos-
terman et al., 2005). One striking expression profile that was
observed in this study was the upregulation of a potato
expressed sequence tag (EST) with homology to a GA 2-
oxidase. Sequence analysis of the corresponding cDNA
clone revealed a complete open reading frame of 340 amino
acids, coding for a protein with a predicted molecular weight
of 38 kDa. The deduced amino acid sequence showed high
levels of homology to other GA 2-oxidase genes isolated
[Nicotiana tabacum, NtGA2ox5 (gbjABO70986) and Nerium
oleander, NoGA2ox3 (gbjAAT72916)], sharing respectively
80.5% and 73.5% of sequence similarity at the amino acid
level. The isolated potato gene described here was named
StGA2ox1. Quantitative real-time polymerase chain reaction
(RT-PCR) analysis of StGA2ox1 expression during tuber
formation (Figure 1a,b) confirms a large increase in tran-
script levels at tuber organogenesis (Stage 3, 72 � 10-fold)
and further tuber growth (Stage 7, 128 � 4-fold) in com-
parison with the non-induced stolon at day 0 (Stage 1).
Expression levels in the tuber gradually decrease when it
reaches its final size (Figure 1a,b; Stage 8). Analysis of
StGA2ox1 tissue specificity shows that the corresponding
gene is predominantly expressed in the tuber while minor
expression was found in root, stem and stolon (Figure 1c).
To determine the precise location of StGA2ox1 transcrip-
tion during tuber development, in situ hybridization was
performed on longitudinal sections of swelling stolons and
very young tubers (<0.8 cm). StGA2ox1 transcripts were
detected within the subapical region of the stolon but not in
the apical meristem (Figure 2). In the swelling stolon, signal
is observed surrounding the vascular bundle in the region
where cell division and expansion are thought to initiate.
During stolon swelling, lower levels of transcript were also
observed in the central pith and possibly the cortex
(Figure 2a). In the longitudinal cross section of a very young
tuber, the region of StGA2ox1 expression is extended to
regions of high mitotic activity associated with tuber growth
and initiation of the perimedullary region (Figure 2b). This is
in good agreement with the upregulated expression pattern
that we observed at tuber initiation and the absence of any
significant expression in dormant tubers, where no further
tuber growth occurs. Thus, StGA2ox1 primary expression in
the subapical stolon region at the early stages of tuber
formation suggests a primary role for this gene in regulating
GA levels during tuber development.
Over-expression and suppression of StGA2ox1 expression
results in altered growth characteristics and tuber
morphology
To further elucidate the function of StGA2ox1 we produced
35S-CaMV over-expression and suppression (post-tran-
scriptional gene silencing, PTGS) clones of potato var.
Karnico. After initial screening, the transgenic clones
Role for a GA 2-oxidase in potato tuber development 363
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 52, 362–373

showing the severest phenotypes were selected. These were
confirmed for the presence of at least one construct insertion
and grown in 10 replicates for further analysis. Plants were
kept under long-day conditions (16-h light) for a period of
6 weeks after which they were switched to short days (8-h
light) to promote tuber formation. Tuber induction in the
Karnico variety is not under strict photoperiod control and
some tuber formation was observed in both transgenic and
control plants under long-day conditions. Individual repli-
cates of each clone did not tuberize synchronously and
therefore we were not able to statistically determine a dif-
ference in the time point of tuberization between the trans-
genic or control plants. However, several clear phenotypic
differences were observed.
Transgenic plants over-expressing StGA2ox1 are semi-
dwarfed and have smaller leaves (Figure 3a,b). These
plants exhibited reduced stolon growth, with on average
fewer and shorter stolons, and formed small tuber initia-
tions directly on the underground stem or developing from
lateral stolon buds (Figure 3e). Formation of these small
(a)
(b)
(c)
Figure 1. Quantitative real-time polymerase chain reaction measuring
StGA2ox1 gene expression during potato tuber development and tissue
specificity.
(a) Representation of the tuber development range consisting of eight
developmental stages harvested over a period of 25 days after a switch from
a 16-h to a 8-h light period (from Kloosterman et al., 2005; ªBlackwell
Publishing, reprinted with permission).
(b) Relative gene expression of StGA2ox1 during the potato tuber develop-
mental stages. Expression levels (DDCt) are represented relative to potato ubi3
gene and the first harvest (Stage 1).
(c) Comparison of the relative expression levels of StGA2ox1 in the different
tissues. StGA2ox1 expression levels (fold change) were calculated relative to
the tissues in which expression levels was highest (Tuber). Tissues studied
include 3-week-old tuber, root, stem, stolon, non-tuberizing stolon tip from LD
plants (NTS), shoot apex, petiole, leaf and dormant tuber (stored for 4 weeks
at 4�C). Error bars indicate the SD between three replicated measurements.
(a) (b)
(c) (d)
Figure 2. In situ localization of StGA2ox1 expression in longitudinal sections
of swelling stolons (a, c) and young tubers (b, d).
Sections were hybridized with StGA2ox1 antisense (a, b) and sense probes (c,
d). The presence of StGA2ox1 mRNA is indicated by the dark stain under
bright-field microscopy. Bars correspond to 1 mm.
364 Bjorn Kloosterman et al.
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 52, 362–373

tuber incipients coincides with stolon initiation, 4 weeks
after planting, but these did not continue to increase in
size. As in control plants, tuber development and further
tuber growth was observed after 6 weeks of growth under
long-day conditions. On average, a significant (P < 0.05)
reduction in total tuber yield was observed in the over-
expression clones (O5, 96 � 8 g; O8, 117 � 16 g; O6,
146 � 3 g) in comparison with the control plants
(226 � 12 g), these clones having either fewer or smaller
tubers (Figure 3j,k).
The suppression clones did not show marked changes in
the aerial part of the plant (Figure 3c). However, they
produced ‘elongated’ tuber swellings or had a higher degree
of stolon branching in the underground parts (Figure 3f–i).
The elongated swellings were most clearly visible during the
early stages of tuber development (Figure 3g). This pheno-
type becomes less distinct during secondary growth (Fig-
ure 3g–i), as tubers produced by these clones show a more
normal morphology after full maturation (Figure 3j–l). Total
tuber yield was on average reduced in the two severest
StGA2ox1 suppression clones (S9, 151 � 24.6 g; S10,
148 � 21.3 g), although this effect was less severe than that
observed for the over-expression clones. A significant
reduction in tuber yield could not be observed for the third
suppression clone (S15), and thus no conclusions about a
negative influence of StGA2ox1 suppression on tuber yield
can be made. The number of tubers did not change
significantly in these clones.
Tubers from both sets of transgenic plants do not differ in
the duration of their dormancy period in comparison to the
control tubers when stored at 18�C. However, sprout growth
(i.e. internode length) was reduced in the StGA2ox1 over-
expression clones, which is in agreement with dwarf
phenotype of these clones (Figure 3m,n).
For confirmation of altered StGA2ox1 expression in the
transgenic clones, transcript levels of StGA2ox1 were
determined in active sink tubers using quantitative (q)RT-
PCR and they are plotted relative to the expression levels
found in control tubers (Figure 4). The transcript levels of
StGA2ox1 in the over-expression clones O5 and O8 were
(a) (c)(b)
(d) (e) (f)
(g) (h) (i)
(j) (k) (l) (m) (n)
Figure 3. Observed phenotypes for StGA2ox1
over-expression clones O5, O6, O8 and suppres-
sion clones S9, S10, S15 and untransformed
control.
(a) Photograph of 4-week-old plants of control
and three independent over-expression clones
(O5, O6 and O8) grown under controlled envi-
ronmental conditions. (b) Reduced leaf size
(4 weeks old) of StGA2ox1 over-expression
clones (O5 and O8) relative to the untransformed
control. (c) Comparison between plant growth of
StGA2ox1 suppression clone S15 and untrans-
formed control plant. (d)–(f) Stolon growth and
tuber set in the untransformed control (d), over-
expression clone O5 (e) and StGA2ox1 suppres-
sion clone S9 (f). The presence of small tuber
incipients in the over-expression clone is indi-
cated with arrows (e). (g)–(i) Different growth
stages of the elongated tuber swelling pheno-
type observed in StGA2ox1 suppression clones.
(j)–(l) Tuber yield of control and transgenic plants
harvested after the plants had completely died
off. Sprouting of tubers stored in the dark from
untransformed control plants (m) and StGA2ox1
over-expression clone O8 (n).
Figure 4. Fold-change in the expression level of StGA2ox1 in tubers of the
over-expression (O5, O8) and suppression clones (S10, S15) relative to the
untransformed control.
Error bars indicate the SD between three replicated measurements.
Role for a GA 2-oxidase in potato tuber development 365
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 52, 362–373

increased respectively, by 13.2 � 0.7- and 9.9 � 1.3-fold,
whilst in the suppression clones S10 and S15 transcript
levels were reduced by 3.9 � 0.3- and 3.6 � 0.3-fold. Hence,
although the expression levels in the suppression clones are
reduced, low levels of StGA2ox1 transcripts were still
detected in these plants and therefore low levels of
StGA2ox1 activity would remain in these clones throughout
tuber growth.
Gibberellin levels in tubers of StGA2ox1 over-expression
and PTGS clones
Gibberellin levels were measured in active sink tubers of
both over-expression and suppression clones and untrans-
formed control plants in order to correlate the observed
phenotypes with altered levels of bioactive GAs. An increase
of GA20 together with a smaller increase in GA1 and GA8 was
observed in the tubers of both suppression clones, S10 and
S15 (Table 1). Measurements of GA levels in tubers at a later
stage of tuber growth also showed elevated levels of GA20 in
both suppression clones S10 (22 ng g)1 FW) and S15
(87 ng g)1 FW) as compared with control plants (0.01 ng g)1
FW), thus further confirming these results.
No clear differences were found for GA metabolites of the
non-13-hydroxylation pathway (Table 1; GA12, GA9, GA51,
GA4 and GA34) and therefore it is assumed that StGA2ox1 is
primarily active in the early 13-hydroxylation pathway
(Table 1; GA53, GA20, GA29, GA1 and GA8).
Interestingly, no obvious difference in GA content could
be observed in the over-expression clones (O5 and O8) in
comparison with control plants (Table 1). A small and
consistent reduction in the levels of GA29 was observed
in both over-expression clones, though its significance
remains unclear. The levels of GA5, GA3 and GA29 catabolite
were not determined in the transgenic plants and therefore
we were unable to determine whether this part of the GA
pathway had changed (Figure 5).
Expression levels of GA metabolism genes are altered
during development and in StGA2ox1 transgenic plants
In order to further investigate GA metabolism during tuber
development we looked at the expression levels of known
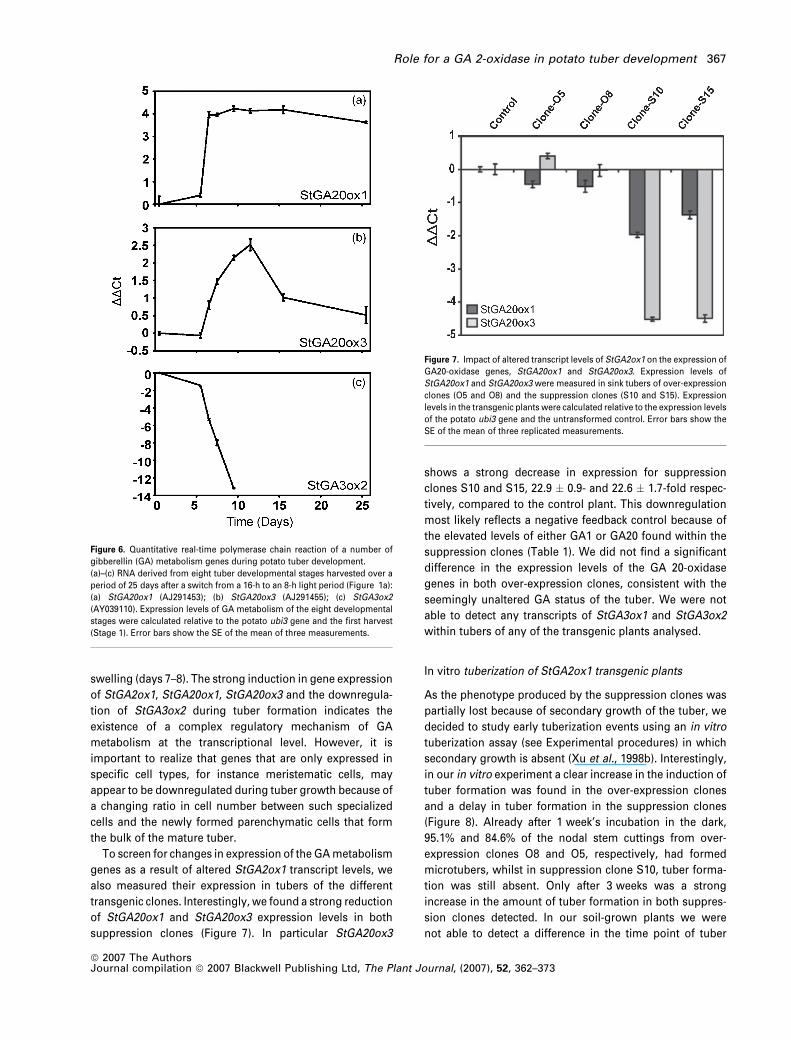
potato GA 20-oxidase and GA 3-oxidase genes (Figure 6).
We found a similar expression profile for StGA20ox1 as was
observed for StGA2ox1 during the eight developmental
stages, with the strongest increase of expression levels
occurring in the swelling stage (15.6 � 2.0-fold change rel-
ative to day 0), after which expression levels remain rela-
tively constant (Figure 6a). StGA20ox3 transcript levels are
also increased at tuber organogenesis but the expression
profile is characterized by a more gradual increase, reaching
peak levels at stage 6 (Figure 6b; 5.7 � 0.9-fold increase). In
potato, two GA 3b-hydroxylase genes have been isolated
from sprouts, StGA3ox1 (gbjAAK91507) and StGA3ox2
(gbjAAK91506), but little is known about their expression
levels or activity within the plant. Using qRT-PCR we were
able to detect expression of one of the GA 3b-hydroxylase
genes (StGA3ox2) in the non-tuberizing stolon tips and early
stages of tuber development (Figure 6c). After the switch
from long-day to short-day conditions (Figure 1a; Stages 1
and 2), transcript levels of StGA3ox2 decrease until no
transcripts could be detected from tuber Stage 5 onwards.
Like StGA2ox1 and StGA20ox1, expression levels of
StGA3ox2 have already changed after the switch from long-
to short-day conditions prior to visible swelling and with the
strongest change in expression levels occurring at stolon
Table 1. Endogenous gibberellin (GA) levels in potato tubers fromuntransformed control plants and StGA2ox1 over-expression andsuppression clones
GA levels (ng g)1 FW)
Control Clone O5 Clone O8 Clone S10 Clone S15
Non-13-hydroxylation pathwayGA12 0.03 n.d. 0.05 0.02 n.d.GA9 0.01 n.d. 0.02 0.01 n.d.GA51 0.04 0.01 0.03 0.03 n.d.GA4 0.06 0.16 0.05 0.04 0.05GA34 0.01 0.01 0.01 0.01 0.02
13-Hydroxylation pathwayGA53 0.04 0.01 0.01 0.01 0.01GA20 0.01 n.d. n.d. 0.49a 0.33a
GA29 0.27 0.13 0.15 0.23 0.24GA1 0.01 0.02 0.04 0.09a 0.10a
GA8 0.03 0.01 0.02 0.23a 0.21a
FW, fresh weight; n.d., no dilution of the internal standard.aStatistically significant differences between control plants andStGA2ox1 suppression lines, Student’s t-test (P < 0.05).
Figure 5. Schematic overview of the major genes involved in the 13-hydrox-
ylation pathway in plants.
Steps catalysed by the different gibberellins (GA)-oxidase genes are
indicated; GA20-oxidase (20ox), GA3-oxidase (3ox), GA 2-oxidase (2ox).
Biologically active GAs are boxed.
366 Bjorn Kloosterman et al.
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 52, 362–373
swelling (days 7–8). The strong induction in gene expression
of StGA2ox1, StGA20ox1, StGA20ox3 and the downregula-
tion of StGA3ox2 during tuber formation indicates the
existence of a complex regulatory mechanism of GA
metabolism at the transcriptional level. However, it is
important to realize that genes that are only expressed in
specific cell types, for instance meristematic cells, may
appear to be downregulated during tuber growth because of
a changing ratio in cell number between such specialized
cells and the newly formed parenchymatic cells that form
the bulk of the mature tuber.
To screen for changes in expression of the GA metabolism
genes as a result of altered StGA2ox1 transcript levels, we
also measured their expression in tubers of the different
transgenic clones. Interestingly, we found a strong reduction
of StGA20ox1 and StGA20ox3 expression levels in both
suppression clones (Figure 7). In particular StGA20ox3
shows a strong decrease in expression for suppression
clones S10 and S15, 22.9 � 0.9- and 22.6 � 1.7-fold respec-
tively, compared to the control plant. This downregulation
most likely reflects a negative feedback control because of
the elevated levels of either GA1 or GA20 found within the
suppression clones (Table 1). We did not find a significant
difference in the expression levels of the GA 20-oxidase
genes in both over-expression clones, consistent with the
seemingly unaltered GA status of the tuber. We were not
able to detect any transcripts of StGA3ox1 and StGA3ox2
within tubers of any of the transgenic plants analysed.
In vitro tuberization of StGA2ox1 transgenic plants
As the phenotype produced by the suppression clones was
partially lost because of secondary growth of the tuber, we
decided to study early tuberization events using an in vitro
tuberization assay (see Experimental procedures) in which
secondary growth is absent (Xu et al., 1998b). Interestingly,
in our in vitro experiment a clear increase in the induction of
tuber formation was found in the over-expression clones
and a delay in tuber formation in the suppression clones
(Figure 8). Already after 1 week’s incubation in the dark,
95.1% and 84.6% of the nodal stem cuttings from over-
expression clones O8 and O5, respectively, had formed
microtubers, whilst in suppression clone S10, tuber forma-
tion was still absent. Only after 3 weeks was a strong
increase in the amount of tuber formation in both suppres-
sion clones detected. In our soil-grown plants we were
not able to detect a difference in the time point of tuber
Figure 6. Quantitative real-time polymerase chain reaction of a number of
gibberellin (GA) metabolism genes during potato tuber development.
(a)–(c) RNA derived from eight tuber developmental stages harvested over a
period of 25 days after a switch from a 16-h to an 8-h light period (Figure 1a):
(a) StGA20ox1 (AJ291453); (b) StGA20ox3 (AJ291455); (c) StGA3ox2
(AY039110). Expression levels of GA metabolism of the eight developmental
stages were calculated relative to the potato ubi3 gene and the first harvest
(Stage 1). Error bars show the SE of the mean of three measurements.
Figure 7. Impact of altered transcript levels of StGA2ox1 on the expression of
GA20-oxidase genes, StGA20ox1 and StGA20ox3. Expression levels of
StGA20ox1 and StGA20ox3 were measured in sink tubers of over-expression
clones (O5 and O8) and the suppression clones (S10 and S15). Expression
levels in the transgenic plants were calculated relative to the expression levels
of the potato ubi3 gene and the untransformed control. Error bars show the
SE of the mean of three replicated measurements.
Role for a GA 2-oxidase in potato tuber development 367
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 52, 362–373

initiation, although early small tuber incipients were obs-
erved in the over-expression clones. Moreover, morpho-
logically significant differences between the individual
transgenic clones and control plants could be observed
(Figure 9). In the over-expression clones, a large percentage
of the formed tubers (78.9%) are sessile where stolon growth
is largely inhibited. The suppression clones however, have a
much larger percentage of lateral tuber formation (43.1%)
and produce a small number of sessile tubers (7.6%) in
comparison to the control plants. Furthermore, while tubers
of the control plantlets generally have a round shape in
which the first and second nodes are incorporated in the
radial swelling, the suppression clones frequently have an
elongated phenotype between the first and second nodes of
the in vitro tuber similar to the phenotype observed in the
in vivo plants (Figure 3g). In addition, lateral tubers were
sometimes formed directly on the tuber eyes.
In an additional experiment, we applied different concen-
trations of GA3 and the GA biosynthesis inhibitor, ancym-
idol, to the tuber-inducing medium (Figure 9). The control
plants supplemented with 0.1 lM GA3 produced fewer
sessile tubers (17.0%) in comparison with the normal
tuber-inducing medium (47.0%) and had an increased
number of lateral tubers, indicating that lateral tuber forma-
tion is at least in part regulated by GA levels. Reduction in
the number of sessile tubers is further enhanced in medium
supplemented with 1.0 lM GA3 where the total number of
tubers formed decreases. Supplementing the over-expres-
sion clone O8 with 0.1 lM GA3 does not seem to have a large
effect on the number of sessile tubers formed, 78.9% and
72.1% respectively. However, when adding 1.0 lM GA3 the
percentage of sessile tubers formed decreased significantly
(42.5%) together with an increase in longitudinal stolon
growth. These findings are consistent with the inhibiting
effect of GA3 on tuber formation in potato. Because of the
expected increase of StGA2ox1 activity in the over-expres-
sion clones, higher levels of GA3 supplementation may be
required to cause a significant effect on tuber formation. In
suppression clone S15, a reduction in the percentage of
apical tubers was observed relative to the amount of GA3
supplied, but this did not lead to an increase in lateral tuber
formation as was found for the control. In conjunction with
these observations, when supplementing the tuber-inducing
medium with 0.5 lM ancymidol, a GA synthesis inhibitor,
stolon growth is largely absent in the over-expression clones
and control, while in the suppression clones the effect is
much smaller and longitudinal stolon growth persists. If GA
synthesis in the stolon is absent because of the presence of
an inhibitor, GA levels present within in the nodal stem
cuttings at the time point of harvest appear to be longer lived
in the suppression clones compared with the over-expres-
sion and control plantlets, possibly because of a reduction in
StGA2ox1 activity.
Taken together these findings suggest that GA levels are
reduced in the over-expression clones, resulting in inhibition
of stolon growth and formation of sessile tubers, whilst in
Figure 8. In vitro tuberization of single node
cuttings from untransformed control plants
(C, ), StGA2ox1 over-expression clones (O8,
and O5, ) and suppression clones (S10, and
S15, ). A large number of single node cuttings
(n) were collected from 4-week-old in vitro grown
plantlets and incubated on tuber-inducing med-
ium in the dark at 18�C. The relative percentage
of tuber formation was scored on a weekly basis.
Figure 9. Distribution of the type of tubers
scored in an in vitro tuberization experiment of
untransformed control plants (C), StGA2ox1
over-expression clone (O8) and StGA2ox1 sup-
pression clone (S15). Single node cuttings were
harvested from 4-week-old in vitro grown plant-
lets and incubated in the dark (18�C) on tuber-
inducing medium (TI) or tuber-inducing medium
supplemented with either 0.1 lM GA3, 1.0 lM
GA3 or 0.5 lM ancymidol and scored after
6 weeks. Images on the right panel illustrate the
type of tubers that were scored.
368 Bjorn Kloosterman et al.
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 52, 362–373

the suppression clones GA levels remain high within the
stolon tip delaying tuber formation and promoting lateral
stolon growth and subsequent lateral tuber formation.
Discussion
StGA2ox1 is expressed during tuber formation in the
subapical region affecting tuber growth
StGA2ox1 is strongly upregulated at stolon swelling and
tuber formation, indicating a role in regulating GA levels at
tuber organogenesis. It has been well documented that GA
levels promote longitudinal cell division and elongation
within plants by affecting the orientation of microtubules
(Fujino et al., 1995; Sanz et al., 1996; Shibaoka, 1993). Xu
et al. (1998b) found that prior to in vitro tuber formation a
strong reduction in GA1 levels was associated with a reori-
entation of the plane of cell division and expansion, result-
ing in radial swelling of the subapical part of the stolon. In
our study, transcription of StGA2ox1 is strongly upregulated
at tuber formation and shown to be restricted to the sub-
apical region of the stolon tip in regions where active cell
divisions take place (Figure 2).
When reducing the number of StGA2ox1 transcripts in the
suppression clones we observe an elongated tuber pheno-
type. In these plants, the required reduction of GA levels at
tuber organogenesis may be insufficient, resulting in only a
partial reorientation of the plane of cell division and expan-
sion and thereby causing the elongated swelling morpho-
logy (Figure 3g–i). During secondary tuber growth, cell
divisions are randomly orientated in the perimedullary
region, often masking the initial elongated swelling pheno-
type. Together, these findings clearly indicate an important
role for StGA2ox1 in regulating the plane of cell division by
controlling GA levels within the subapical stolon region in
the early stages of tuber organogenesis. StGA2ox1 is
considered to be a primary response gene (i.e. a tuber
identity gene) as its expression specifically increases in the
subapical region only in cells undergoing transition to tuber
fate after tuber formation is induced. This is in agreement
with the absence of a significant difference in the time point
of tuberization between the control plants and the StGA2ox1
over-expression and suppression clones in the in vivo
experiment.
In sharp contrast, a clear difference in the time point of
tuberization was observed in the in vitro tuberization exper-
iment (Figure 8) in which either a clear induction or delay in
the rate of in vitro tuber formation was observed for the
over-expression and suppression clones, respectively, con-
sistent with altered GA content (Jackson and Prat, 1996;
Vreugdenhil and Struik, 1989; Xu et al., 1998a). Under in
vitro conditions, high sucrose levels are thought to be the
major signalling component in tuber formation, as sucrose
concentrations correlate negatively with GA levels in the
stolon tip (Xu et al., 1998a). Hence, under in vitro conditions
the time point of tuber formation appears to rely primarily
on sucrose/GA interactions within the stolon tip (Xu et al.,
1998a) whilst under in vivo conditions additional regulatory
factors are required to induce tuber formation. Interestingly,
the ratio of lateral in vitro tuber formation was significantly
increased in the suppression clones (Figure 9), indicating
the presence of a gradient of biologically active GAs within
the stolon. It has been suggested that concentrations of GA1
vary throughout the stolon, with the highest concentration
located in the stolon tip (Jackson, 1999).
Similarly, we did not observe a difference in the duration
of the dormancy period for both StGA2ox1 over-expression
and suppression clones in comparison with tubers of the
control plants. However, we did find that early sprout growth
(i.e. internode length) is reduced in the StGA2ox1 over-
expression clones consistent with the dwarfed plant pheno-
type (Figure 3a,m,n). We therefore conclude that StGA2ox1
does not play a role in regulating the length of the dormancy
period within the tuber.
Reduced transcript levels of StGA2ox1 result in increased
levels of GA20, GA1 and GA8
Within the suppression clones an apparently large increase
of GA20 and a less obvious increase of GA1 and GA8 were
observed (Table 1). The increased substrate availability of
GA20 most likely results in an increased flux towards the
synthesis of both GA1 and GA8. As the levels of the biolo-
gically inactive GA8 are elevated in the StGA2ox1 suppres-
sion clones and not in the over-expression clones, it seems
unlikely that StGA2ox1 acts directly on GA1. Surprisingly, we
did not find a significant increase or reduction of bioactive
GAs in the studied tubers of the over-expression clones.
However, the phenotypes of these plants, semi-dwarfed,
reduced stolon growth and earlier in vitro tuberization, are
consistent with a reduction in overall bioactive GA content,
indicating that very small changes in GA homeostasis may
already be sufficient to alter plant morphology.
The oxidation of GA29 to produce GA29 catabolite has
been shown to be catalysed by a GA 2-oxidase in pea shoots
(Lester et al., 1999; Martin et al., 1999). We did find a small
reduction in the levels of GA29 in the over-expression clones
suggesting a role for StGA2ox1 in this conversion, but no
supporting evidence was found with data obtained for the
suppression clones in which no difference were observed.
In maize shoots it was shown that GA3 can be synthesized
from GA20 via GA5 (Fujioka et al., 1990; Spray et al., 1996).
Abdala et al. (2002) measured significant levels of GA3 in
potato foliage, root, stolon and tuber. However, there are no
detailed data available on changing GA3 levels during the
transition of a stolon into a tuber as was reported for GA1
levels (Xu et al., 1998a) and as a result the role of GA3 in
tuber formation remains unclear. The GA 2-oxidase gene
Role for a GA 2-oxidase in potato tuber development 369
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 52, 362–373

family in general consists of multiple members which are
often functionally redundant (Hedden and Proebsting, 1999;
Thomas et al., 1999) and therefore we cannot exclude
complementation of StGA2ox1 activity by a different family
member in the transgenic plants.
Transcriptional control of GA metabolism genes during
potato tuber development
A long-standing issue has been the understanding of how
endogenous GA levels are controlled during the process of
tuber formation. Based on the observed phenotype in the
StGA2ox1 suppression clones and its subapical localization,
inactivation of bioactive GAs or GA intermediates through
the activity of StGA2ox1 is an important regulatory step in
controlling endogenous GA levels in the subapical stolon
region. However, no StGA2ox1 transcript signal was
detected in the stolon apex, indicating that another control
mechanism is required for reducing GA content in the stolon
apex as reported by Xu et al. (1998a). We found that the
transcript levels of an enzyme catalysing the final step in the
biosynthesis of bioactive GAs, StGA3ox2, was strongly
downregulated at tuber onset (Figure 6c). Both the upreg-
ulation of a GA inactivation gene (StGA2ox1) and the
downregulation of a GA biosynthesis gene (StGA3ox2)
would allow a rapid reduction in GA content within the
swelling stolon required for normal tuber formation. Inter-
estingly, two genes involved in the synthesis of GA-precur-
sor GA20 (StGA20ox1 and StGA20ox3), show an upregulated
expression profile during the early stages of tuber devel-
opment, although less strong in comparison to StGA2ox1
(Figures 1b and 6a,b).
As both StGA20ox1 and StGA20ox3 transcript levels
have been shown to be under negative feedback control of
GA levels (Carrera et al., 1999), the increased expression of
both genes at tuber organogenesis may be the result of
feedback regulation because of a reduction in the concen-
tration of specific GAs. Similarly, the observed decrease in
the transcription levels of the same genes in the transgenic
tubers of StGA2ox1 suppression clones, having increased
levels of GA20, GA1 and GA8 (Table 1), can be explained by
the same feedback mechanism (Figure 7). Conversely,
several studies have shown that changes in StGA20ox1
transcript levels in the stolon can be directly altered
through over-expression of homeobox genes POTH1 and
StBEL5 that are able to bind to the StGA20ox promoter,
reducing its activity (Chen et al., 2003, 2004). As both
StBEL5 and StGA20ox1 are upregulated at tuber formation,
these findings bring up an interesting paradox as to the
primary transcriptional control mechanism and relative
importance of StGA20ox1 during tuber development within
the stolon and developing tuber. StGA20ox1 is highly
expressed in the leaves and a primary role for this gene in
controlling tuber induction in the leaves is further sup-
ported by the finding that over-expression of StGA20ox1
under control of a leaf specific promoter (STLS1) delays
tuber formation in a similar fashion as constitutive (CaMV
35S) over-expression lines (Carrera et al., 2000).
The relatively strong increase and decrease in transcript
levels of StGA2ox1 and StGA3ox2 at tuber onset raises the
question whether there is a common mechanism controlling
their expression. When decapitating pea shoots, the reduc-
tion of auxin levels throughout the plant leads to a decrease
of PsGA3ox1 expression and an increase of PsGA2ox1
expression levels (Ross et al., 2000). The effects can be
opposed by the application of indole-3-acetic acid (IAA)
indicating a regulatory role in GA metabolism. These
findings may support a similar role for IAA in regulating
StGA3ox2 and StGA2ox1 expression levels during potato
tuber development.
The changes in gene expression already observed after
the switch to short-day conditions within the stolon tip prior
to visible swelling implies that transcription of StGA2ox1,
StGA20ox1 and StGA3ox2 is closely associated with the
tuber induction signal. However, the mechanism by which a
tuber induction signal produced in the leaves translates to
GA metabolism or sensitivity within the stolon tip is still
under investigation, although much progress has been
made in recent years.
As mentioned, StBEL5 and its protein partner POTH1 can
alter tuber formation by mediating hormone levels in the
stolon tip. It was recently shown that transcripts of StBEL5
are upregulated in response to short-day conditions and are
present in phloem cells, capable of moving across a graft
union towards the stolon tip revealing a long-distance
signalling pathway (Banerjee et al., 2006). Different KNOX
proteins have been implicated in regulation of GA catabolic
enzymes to confine GA activity to the differentiating leaf
primordia (Jasinski et al., 2005). In another study, a MADS-
box transcription factor, AGAMOUS-like 15, has been shown
to bind to the regulatory region of AtGA2ox6, controlling its
expression level (Wang et al., 2004). Whether the expression
level of StGA2ox1, and thereby its activity, is also controlled
through similar types of regulatory protein needs to be
investigated. In addition, post-transcriptional control of
gene expression, protein modification and degradation,
provides the plant with additional tools for fine-tuning
of the GA metabolism pathway during potato tuber
development.
The data presented in this paper on the expression of
StGA2ox1 and other GA metabolism genes during potato
tuber development provide novel insights into GA-regulated
potato tuber formation at the organ level. From our expres-
sion studies in transgenic plants, we conclude that
StGA2ox1 fulfils a central role in the transition from longi-
tudinal stolon growth to tuber initiation by regulating GA
levels in the subapical stolon region facilitating radial
growth.
370 Bjorn Kloosterman et al.
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 52, 362–373

Experimental procedures
Cloning of StGA2ox1 and sequence analysis
The sequence of the potato GA 2-oxidase was obtained bysequencing cDNA clone BI176613 deriving from an in vitro grownmicrotuber EST library (cv. Bintje). Sequence analysis revealed thatthe cDNA was full length and coded for a protein of 340 amino acids insize and was named StGA2ox1 (gb|EU003995). The complete codingsequence of StGA2ox1 was amplified by PCR (forward primer5¢-CACCTATGGTTGTTTTGTCTCA-3¢ and reverse primer 5¢-TGTCA-TGATTGAGCATTCT-3¢). The forward primer contains a 5¢-CACCpartial overhang to facilitate cloning of the PCR product inpENTR/SD/D-TOPO vector (Invitrogen, http://www.invitrogen.com/).StGA2ox1 was further subcloned by a LR recombination reactioninto Gateway plant transformation destination vector pk7WG2 andpk7GWIWG2 (Plant Systems Biology, University of Ghent, Belgium)(Karimi et al., 2002). The two resulting expression constructs werenamed pk7GA2ox1_O and pk7GAox1_S. pk7GA2ox_O expressionresults in heterologous expression of StGA2ox1 gene under controlof the 35S-CaMV promoter. The Pk7GA2ox_S expression constructconsists of a full-length StGA2ox1 inverted repeat harbouring anintron behind the 35S-CaMV promoter that when expressedproduces a hairpin structure resulting in PTGS.
Plant transformation and regeneration
pk7GA2ox1_O and pk7GAox1_S were transformed into Agrobacte-rium tumefaciens strain Agl0 using electroporation. In vitro shootsof the S. tuberosum cv. Karnico were used for A. tumefaciens-mediated transformation (Visser et al., 1991). After regeneration ofin vitro shoots on selective kanamycin Murashige and Skoog (MS)medium (100 mg L)1) (Murashige and Skoog, 1962), 45 indepen-dent suppression and 9 over-expression clones were transferred tosoil and grown in the greenhouse to check for altered expression ofStGA2ox1. Integration of at least one copy of either construct in theselected transgenic plants was confirmed by Southern blottingusing part of the 35S-CaMV promoter as probe. From these trans-genic plants, three strong over-expression and suppression cloneswere selected for further analysis. Plants regenerated fromuntransformed in vitro shoots were used as wild-type controls.
Plant material growth conditions and tuber harvest
Ten replicates of the in vitro control plants and of the three selectedover-expression and suppression clones were transferred to soiland grown in a climate chamber (16-h light 20�C and 8-h dark at18�C) for a period of 6 weeks after which plants were transferred toshort-day conditions (8-h light and 16-h dark period). Tubers fromtwo repeats of each plant were harvested 6 and 12 weeks after theswitch to short days and immediately frozen in liquid nitrogen.Frozen tubers were ground into a fine powder and used forexpression studies and GA measurements. Remaining plants wereleft to senescence and die after which the number of tubers and totaltuber weight was determined from at least three repeats from eachtransgenic clone and control plants. Significance for difference intotal tuber weight between the different transgenic clones andcontrol plant was determined using Student’s t-test (P < 0.05).
RNA isolation and quantitative RT-PCR
For expression studies within the different transgenic clones andcontrol plants, total RNA was isolated from tubers as described by
Bachem et al. (1996). For expression studies of the GA-oxidasegenes during the potato tuber developmental time series and in thedifferent potato tissues, total RNA was used from a previous study(Kloosterman et al., 2005). One additional tissue sample wasincluded, namely the shoot apex, and was harvested from 8-week-old greenhouse-grown plants (S. tuberosum cv. Karnico).
Relative expression levels of genes were determined by real-timeqRT-PCR on a Perkin Elmer Abi Prism 7700 sequence detector(Perkin Elmer, http://www.perkinelmer.com/) following the protocoldescribed in Kloosterman et al. (2005). Potato ubiquitin primers(ubi3) were used as a control. Relative quantification of the targetRNA expression level and SD was calculated from three replicatedmeasurements of the same sample using the comparative Ctmethod according to User Bulletin no. 2 (ABI Prism 7700 SequenceDetection System, December 1997, Applied Biosystems, http://www.appliedbiosystems.com/). The primer sequences for thegenes studied are as follows: StGA20ox1 (gb|CAC13036) forwardprimer 5¢-CGGCCCAACAAGCATCTAAG-3¢, reverse primer 5¢-AAGC-CATGACTCCGACACG-3¢; StGA20ox3 (gb|CAC13038) forwardprimer 5¢-GCAATGCCATGAGCACCC-3¢, reverse primer 5¢-GGCTC-AATCCCAAAAGTTCCA-3¢. StGA3ox2 (gb|AAK91506) forwardprimer 5¢-AGCTCATGTGGTCCGAAGGA-3¢; reverse primer 5¢-CGG-ACAAGCCGGGTAAGAAT-3¢; StGA2ox1 (gb|EU003995) forwardprimer 5¢-AGGCACAGAGTGATCGCAGAT-3¢, reverse primer5¢-TGGTGGCCCTCCAAAGTAAA-3¢; ubi3 (gb|L22576) forward pri-mer, 5¢-TCCGACACCATCGACAATGT-3¢, reverse primer 5¢-CGACCA-TCCTCAAGCTGCTT-3¢.
In situ hybridizations of StGA2ox1
Tissue preparation and in situ hybridization experiments were car-ried out according to Jackson et al. (1991), with small modificationsas described in Bradley et al. (1993) and Coen et al. (1990). Probesused to detect the StGA2ox1 transcript were prepared from a pGEM-T-Easy vector (Promega, http://www.promega.com/) that containeda 786-bp fragment of the StGA2ox1 cDNA inserted in sense and inantisense orientations. This insert was obtained by PCR amplifica-tion using the primers 5¢-TTTTCCAACAAACAACTATGGTTG-3¢ and5¢-CTGCAATGAGTCACCAACATTG-3¢. After T7 transcription, theprobe was hydrolysed for 40 min at 60�C in carbonate buffer, toobtain fragments of approximately 180 bp.
The GA measurements
For quantitative determination of endogenous GAs in tubers of thetransgenic clones (O5, O8, S10 and S15) and untransformed con-trols (C), tubers from three independent replicates were harvestedfrom 12-week-old plants, mixed and immediately frozen in liquidnitrogen before being ground into a fine powder. Five grams of FWtubers of each sample was spiked with 16, 17-d2-GA standards (5 ngeach; from Prof. L. Mander, Canberra, Australia). Samples wareextracted, purified, derivatized and analysed by gas chromatogra-phy mass spectrometry using selected ion monitoring as describedelsewhere (Lange et al., 2005). Significance for difference in GAcontent between the transgenic clones and control plant wasdetermined using Student’s t-test (P < 0.05).
In vitro tuberization assay
A modified version of in the in vitro tuberization method describedby Hendriks et al. (1991) was used for microtuber production.Instead of using in vivo-grown plants grown for single node harvest,in vitro plantlets were used. The upper three nodal stem sections
Role for a GA 2-oxidase in potato tuber development 371
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 52, 362–373

containing a single axillary bud from 4-week-old in vitro-grownplantlets were harvested from two independent over-expressionand suppression clones and control plants. For each clone, morethan 100 stem sections (n) were put directly on the tuber-inducingmedium and incubated in the dark (18�C). Tuber-inducing mediumconsists of modified MS medium (Murashige and Skoog, 1962),containing 1/10 part of the standard amount of KNO3 and NH4NO3,8% w/v sucrose and 0.8% w/v agar with a final pH of 5.8. Tuberformation was scored weekly over a period of 6 weeks. In addition,around 50 stem sections of the control and different transgenicclones were incubated on tuber-inducing medium supplementedwith either 0.1 lM GA3, 1.0 lM GA3 or 1.0 lM ancymidol and scoredfor tuberization. As a non-tuberizing control, plantlets were incu-bated on tuber-inducing medium containing 1% w/v of sucrose. Theproportion of tuber formation and types of tuber (p) for each treat-ment was calculated relative to the number (n) of incubated stemsections with variance Var = p(1 – p)/n.
Acknowledgements
The authors acknowledge support from EU-SOL, EPS (GraduateSchool of Experimental Plant Sciences), Wageningen Universityand Plant Research International through its twinning projectprogramme. We thank Anja Liebrandt for performing the GAmeasurements and Niek Appeldoorn for critical reading of themanuscript.
References
Abdala, G., Castro, G., Miersch, O. and Pearce, D. (2002) Changes injasmonates and gibberellins during development of potato plants(Solanum tuberosum L.). Plant Growth Regul. 36, 121–126.
Amador, V., Monte, E., Garcia-Martinez, J.L. and Prat, S. (2001)Gibberellins signal nuclear import of PHOR1, a photoperiod-responsive protein with homology to Drosophila armadillo. Cell,106, 343–354.
Bachem, C.W., van der Hoeven, R.S., de Bruijn, S.M., Vreugdenhil,
D., Zabeau, M. and Visser, R.G. (1996) Visualization of differentialgene expression using a novel method of RNA fingerprintingbased on AFLP: analysis of gene expression during potato tuberdevelopment. Plant J. 9, 745–753.
Banerjee, A.K., Chatterjee, M., Yu, Y., Suh, S.G., Miller, W.A. and
Hannapel, D.J. (2006) Dynamics of a mobile RNA of potato in-volved in a long-distance signaling pathway. Plant Cell, 18, 3443–3457.
Booth, A. (1963) The role of growth substances in the developmentof stolons. In The Growth of the Potato (Ivins, J.D. and Milthorpe,F.L., eds). London: Butterworths, pp. 99–112.
Bradley, D., Carpenter, R., Sommer, H., Hartley, N. and Coen, E.
(1993) Complementary floral homeotic phenotypes result fromopposite orientations of a transposon at the plena locus ofAntirrhinum. Cell, 72, 85–95.
Carrera, E., Jackson, S.D. and Prat, S. (1999) Feedback control anddiurnal regulation of gibberellin 20-oxidase transcript levels inpotato. Plant Physiol. 119, 765–774.
Carrera, E., Bou, J., Garcia-Martinez, J.L. and Prat, S. (2000) Chan-ges in GA 20-oxidase gene expression strongly affect stemlength, tuber induction and tuber yield of potato plants. Plant J.22, 247–256.
Chen, H., Rosin, F.M., Prat, S. and Hannapel, D.J. (2003) Interactingtranscription factors from the three-amino acid loop extensionsuperclass regulate tuber formation. Plant Physiol. 132, 1391–1404.
Chen, H., Banerjee, A.K. and Hannapel, D.J. (2004) The tandemcomplex of BEL and KNOX partners is required for transcriptionalrepression of ga20ox1. Plant J. 38, 276–284.
Coen, E.S., Romero, J.M., Doyle, S., Elliott, R., Murphy, G. and
Carpenter, R. (1990) Floricaula: a homeotic gene required forflower development in antirrhinum majus. Cell, 63, 1311–1322.
Ewing, E.E. (1987) The role of hormones in potato (Solanumtuberosum L.) tuberization. In Plant Hormones and Their Role inPlant Growth and Development (Davies, P.J., ed). Boston: Marti-nus Nijhoff Publishers, pp. 515–538.
Ewing, E.E. and Struik, P.C. (1992) Tuber formation in potato:induction, initiation, and growth. Hortic. Rev. 14, 89–198.
Fujino, K., Koda, Y. and Kikuta, Y. (1995) Reorientation of corticalmicrotubules in the sub-apical region during tuberization insingle note stem segments of potato in culture. Plant Cell Physiol.36, 348–353.
Fujioka, S., Yamane, H., Spray, C.R., Phinney, B.O., Gaskin, P.,
MacMillian, J. and Takahashi, N. (1990) Gibberellin A3 is bio-synthesized from gibberellin A20 via gibberellin A5 in shoots ofZea Mays L. Plant Physiol. 94, 127–131.
Gregory, L.E. (1956) Some factors for tuberization in the potato. AnnBot. 41, 281–288.
Hedden, P. (2001) Gibberellin metabolism and its regulation. J. PlantGrowth Regul. 20, 317–318.
Hedden, P. and Phillips, A.L. (2000) Gibberellin metabolism: newinsights revealed by the genes. Trends Plant Sci. 5, 523–530.
Hedden, P. and Proebsting, W.M. (1999) Genetic analysis of gib-berellin biosynthesis. Plant Physiol. 119, 365–370.
Hendriks, T., Vreugdenhil, D. and Stiekema, W.J. (1991) Patatinand four serine proteinase inhibitor genes are differentiallyexpressed during potato tuber development. Plant Mol. Biol.17, 385–394.
Jackson, S.D. (1999) Multiple signaling pathways control tuberinduction in potato. Plant Physiol. 119, 1–8.
Jackson, S.D. and Prat, S. (1996) Control of tuberization in potato bygibberellins and phytochrome B. Physiol. Plant, 98, 407–412.
Jackson, D., Culianez-Macia, F., Prescott, A.G., Roberts, K. and
Martin, C. (1991) Expression patterns of myb genes from Antir-rhinum flowers. Plant Cell, 3, 115–125.
Jackson, S.D., James, P., Prat, S. and Thomas, B. (1998) Phyto-chrome B affects the levels of a graft-transmissible signalinvolved in tuberization. Plant Physiol. 117, 29–32.
Jackson, S.D., James, P.E., Carrera, E., Prat, S. and Thomas, B.
(2000) Regulation of transcript levels of a potato gibberellin 20-oxidase gene by light and phytochrome B. Plant Physiol. 124,423–430.
Jasinski, S., Piazza, P., Craft, J., Hay, A., Woolley, L., Rieu, I., Phillips,
A., Hedden, P. and Tsiantis, M. (2005) KNOX action in Arabidopsisis mediated by coordinate regulation of cytokinin and gibberellinactivities. Curr. Biol. 15, 1560–1565.
Karimi, M., Inze, D. and Depicker, A. (2002) GATEWAY vectors forAgrobacterium-mediated plant transformation. Trends Plant Sci.7, 193–195.
Kloosterman, B., Vorst, O., Hall, R.D., Visser, R.G.F. and Bachem,
C.W. (2005) Tuber on a chip: differential gene expression duringpotato tuber development. Plant Biotechnol. J. 3, 505–519.
Kumar, D. and Wareing, P.F. (1974) Studies on tuberization ofSolanum andigena. II. Growth hormones and tuberization. NewPhytol. 73, 833–840.
Lange, T. (1998) Molecular biology of gibberellin synthesis. Planta,204, 409–419.
Lange, T., Kappler, J., Fischer, A., Frisse, A., Padeffke, T., Schmidtke,
S. and Lange, M.J. (2005) Gibberellin biosynthesis in developingpumpkin seedlings. Plant Physiol. 139, 213–223.
372 Bjorn Kloosterman et al.
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 52, 362–373

Lester, D.R., Ross, J.J., Smith, J.J., Elliott, R.C. and Reid, J.B. (1999)Gibberellin 2-oxidation and the SLN gene of Pisum sativum. PlantJ. 19, 65–73.
Martin, D.N., Proebsting, W.M. and Hedden, P. (1999) The SLENDERgene of pea encodes a gibberellin 2-oxidase. Plant Physiol. 121,775–781.
Martinez-Garcia, J.F., Garcia-Martinez, J.L., Bou, J. and Prat, S.
(2001) The interaction of gibberellins and photoperiod in thecontrol of potato tuberization. J. Plant Growth Regul. 20, 377–386.
Murashige, T. and Skoog, F. (1962) A revised medium for rapidgrowth and bio-assays with tobacco tissue cultures. Physiol.Plant, 15, 473–497.
Rodriguez-Falcon, M., Bou, J. and Prat, S. (2006) Seasonal control oftuberization in potato: conserved elements with the floweringresponse. Annu. Rev. Plant Biol. 57, 151–180.
Rosin, F.M., Hart, J.K., Horner, H.T., Davies, P.J. and Hannapel, D.J.
(2003) Overexpression of a knotted-like homeobox gene of potatoalters vegetative development by decreasing gibberellin accu-mulation. Plant Physiol. 132, 106–117.
Ross, J.J., O’Neill, D.P., Smith, J.J., Kerckhoffs, L.H. and Elliott, R.C.
(2000) Evidence that auxin promotes gibberellin A1 biosynthesisin pea. Plant J. 21, 547–552.
Sanz, M.J., Mingo-Castel, A., Lammeren, A.A.M.V. and
Vreugdenhil, D. (1996) Changes in the microtubular cytoskele-ton precede in vitro tuber formation in potato. Protoplasma,191, 46–54.
Shibaoka, H. (1993) Regulation by gibberellins of the orientation ofcortical microtubules in plant cells. Aust. J. Plant Physiol. 20, 461–470.
Spray, C.R., Kobayashi, M., Suzuki, Y., Phinney, B.O., Gaskin, P. and
MacMillan, J. (1996) The dwarf-1 (dt) Mutant of Zea mays blocksthree steps in the gibberellin-biosynthetic pathway. Proc. NatlAcad. Sci. USA, 93, 10515–10518.
Thomas, S.G., Phillips, A.L. and Hedden, P. (1999) Molecular cloningand functional expression of gibberellin 2-oxidases, multifunc-tional enzymes involved in gibberellin deactivation. Proc. NatlAcad. Sci. USA, 96, 4698–4703.
Visser, R.G., Stolte, A. and Jacobsen, E. (1991) Expression of achimaeric granule-bound starch synthase-GUS gene in trans-genic potato plants. Plant Mol. Biol. 17, 691–699.
Vreugdenhil, D. and Struik, P.C. (1989) An integrated view ofthe hormonal regulation of tuber formation in potato (Solanumtubersosum). Physiol. Plant, 75, 525–531.
Wang, H., Caruso, L.V., Downie, A.B. and Perry, S.E. (2004) Theembryo MADS domain protein AGAMOUS-Like 15 directly reg-ulates expression of a gene encoding an enzyme involved ingibberellin metabolism. Plant Cell, 16, 1206–1219.
Xu, X., van Lammeren, A.A., Vermeer, E. and Vreugdenhil, D. (1998a)The role of gibberellin, abscisic acid, and sucrose in the regulationof potato tuber formation in vitro. Plant Physiol. 117, 575–584.
Xu, X., Vreugdenhil, D. and van Lammeren, A.A.M. (1998b) Celldivision and cell enlargement during potato tuber formation.J. Exp. Bot. 49, 573–582.
Role for a GA 2-oxidase in potato tuber development 373
ª 2007 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 52, 362–373




















