
STAT5 regulates the self-renewal capacity and differentiation of human memory B cells and controls...
11
STAT5 regulates the self-renewal capacity and differentiation of human memory B cells and controls Bcl-6 expression Ferenc A Scheeren 1–3 , Marianne Naspetti 2,3 , Sean Diehl 1 , Remko Schotte 1,2 , Maho Nagasawa 1,2 , Erwin Wijnands 1 , Ramon Gimeno 1 , Florry A Vyth-Dreese 2 , Bianca Blom 1,2 & Hergen Spits 1,2 It is unknown how B cells that mature during a germinal center reaction ‘decide’ between plasma or memory cell fate. Here we describe a previously unknown subpopulation of B cells in the human germinal center that is characterized by tyrosine phosphorylated transcriptional activator STAT5. These cells had an activated centrocyte phenotype and had abundant expression of BCL6 but low expression of PRDM1, both encoding transcriptional repression proteins. Using RNA interference and ectopic expression of constitutively activated forms of STAT5, we demonstrate here a function for STAT5 in the self-renewal of B cells in vitro. STAT5b isoform seemed to directly upregulate Bcl-6, and ectopic expression of Bcl-6 in B cells resulted in self-renewal and inhibition of plasma cell differentiation. These data indicate that activation of STAT5 is involved in regulation of memory B cell differentiation. During the T cell–dependent B cell response that takes place in germinal centers, B cells undergo extensive proliferation followed by selection and maturation. Long-term memory B cells and antibody- forming plasma cells form during the germinal center reaction 1,2 . The mechanisms by which these two populations of B cells are generated in germinal centers are incompletely understood. The affinity of the B cell receptor (BCR) seems to be one of the factors that determine plasma or memory B cell differentiation 3 . Evidence also exists showing other extracellular signals can regulate memory versus plasma cell differentiation. The tumor necrosis factor family members CD40 and OX40L are involved in memory cell differentiation and plasma cell development, respectively 4 . Differentiation of plasma cells is driven by PRDM1, the gene encoding the transcriptional repressor protein Blimp-1 (refs. 5–8), and Xbp1, the gene encoding the X-box transcription factor XBP-1 (ref. 9). In contrast, memory B cells express Bcl-6, a transcriptional repressor that inhibits differentiation into plasma cells, most likely by downregulating Blimp-1. These findings indicate that the choice between memory B cell and plasma cell fate in germinal centers involves regulation of BCL6 versus PRDM1 and XBP1. Moreover, whereas plasma cells are terminally differentiated cells that are unable to enter the cell cycle, memory B cells should maintain their self- renewal potential. Indeed, in contrast to plasma cells, memory B cell populations expand extensively in vitro when stimulated through CD40 and cytokines 10 . However, such in vitro–expanded B cell populations have a limited replication potential. Apparently the in vitro conditions do not prevent the expanded memory B cell populations from undergoing terminal differentiation accompanied by cell cycle arrest 11 . In accordance with this, ectopic expression of BCL6 in human B cells results in maintenance of their in vitro expansion potential 12 , suggesting that Bcl-6 protein is required not only for the generation of memory cells but also for maintenance of the potency to self-renew, as proposed before 13 . Assuming that continued high expression of Bcl-6 is an ‘instructed’ event, the question arises regarding which signals are required to maintain Bcl- 6 expression. In addition to CD40 triggering, cytokines are also needed for differentiation and expansion of memory cell populations 10 . Interleukin 2 (IL-2), IL-4 and IL-10 are the most efficient cytokines in promoting continued cell cycle progression of CD40- or surface immunoglobulin–stimulated naive and memory B cells in vitro. Although these cytokines are not essential for memory B cell differ- entiation, they might trigger signaling pathways that, together with CD40-linked signaling, are involved in memory cell differentiation in vivo. IL-2, IL-4 and IL-10 induce growth of B cells through activation of Janus kinase (JAK)–signal transducer of activation and transcription (STAT) pathways. IL-4 activates STAT6 and STAT5 (refs. 14,15), whereas IL-2 and IL-10 activate STAT3 and STAT5 (ref. 16). STAT6 promotes the choice of the immunoglobulin E (IgE) isotype during class-switch recombination 17,18 , whereas STAT3 is linked to plasma Published online 13 February 2005; doi:10.1038/ni1172 1 Department of Cell Biology and Histology of the Academic Medical Center, University of Amsterdam, 1105 AZ Amsterdam, Netherlands. 2 Division of Immunology, Netherlands Cancer Institute, 1066 CX Amsterdam, Netherlands. 3 These authors contributed equally to this work. Correspondence should be addressed to H.S. ([email protected]) NATURE IMMUNOLOGY VOLUME 6 NUMBER 3 MARCH 2005 303 ARTICLES © 2005 Nature Publishing Group http://www.nature.com/natureimmunology
Transcript of STAT5 regulates the self-renewal capacity and differentiation of human memory B cells and controls...

STAT5 regulates the self-renewal capacity anddifferentiation of human memory B cells and controlsBcl-6 expression
Ferenc A Scheeren1–3, Marianne Naspetti2,3, Sean Diehl1, Remko Schotte1,2, Maho Nagasawa1,2,Erwin Wijnands1, Ramon Gimeno1, Florry A Vyth-Dreese2, Bianca Blom1,2 & Hergen Spits1,2
It is unknown how B cells that mature during a germinal center reaction ‘decide’ between plasma or memory cell fate. Here
we describe a previously unknown subpopulation of B cells in the human germinal center that is characterized by tyrosine
phosphorylated transcriptional activator STAT5. These cells had an activated centrocyte phenotype and had abundant expression
of BCL6 but low expression of PRDM1, both encoding transcriptional repression proteins. Using RNA interference and ectopic
expression of constitutively activated forms of STAT5, we demonstrate here a function for STAT5 in the self-renewal of B cells
in vitro. STAT5b isoform seemed to directly upregulate Bcl-6, and ectopic expression of Bcl-6 in B cells resulted in self-renewal
and inhibition of plasma cell differentiation. These data indicate that activation of STAT5 is involved in regulation of memory
B cell differentiation.
During the T cell–dependent B cell response that takes place ingerminal centers, B cells undergo extensive proliferation followed byselection and maturation. Long-term memory B cells and antibody-forming plasma cells form during the germinal center reaction1,2. Themechanisms by which these two populations of B cells are generated ingerminal centers are incompletely understood. The affinity of the Bcell receptor (BCR) seems to be one of the factors that determineplasma or memory B cell differentiation3. Evidence also exists showingother extracellular signals can regulate memory versus plasma celldifferentiation. The tumor necrosis factor family members CD40 andOX40L are involved in memory cell differentiation and plasma celldevelopment, respectively4.
Differentiation of plasma cells is driven by PRDM1, the geneencoding the transcriptional repressor protein Blimp-1 (refs. 5–8),and Xbp1, the gene encoding the X-box transcription factor XBP-1(ref. 9). In contrast, memory B cells express Bcl-6, a transcriptionalrepressor that inhibits differentiation into plasma cells, most likely bydownregulating Blimp-1. These findings indicate that the choicebetween memory B cell and plasma cell fate in germinal centersinvolves regulation of BCL6 versus PRDM1 and XBP1. Moreover,whereas plasma cells are terminally differentiated cells that are unableto enter the cell cycle, memory B cells should maintain their self-renewal potential. Indeed, in contrast to plasma cells, memory B cellpopulations expand extensively in vitro when stimulated throughCD40 and cytokines10. However, such in vitro–expanded B cell
populations have a limited replication potential. Apparently thein vitro conditions do not prevent the expanded memory B cellpopulations from undergoing terminal differentiation accompaniedby cell cycle arrest11. In accordance with this, ectopic expression ofBCL6 in human B cells results in maintenance of their in vitroexpansion potential12, suggesting that Bcl-6 protein is required notonly for the generation of memory cells but also for maintenance ofthe potency to self-renew, as proposed before13. Assuming thatcontinued high expression of Bcl-6 is an ‘instructed’ event, thequestion arises regarding which signals are required to maintain Bcl-6 expression. In addition to CD40 triggering, cytokines are also neededfor differentiation and expansion of memory cell populations10.Interleukin 2 (IL-2), IL-4 and IL-10 are the most efficient cytokinesin promoting continued cell cycle progression of CD40- or surfaceimmunoglobulin–stimulated naive and memory B cells in vitro.Although these cytokines are not essential for memory B cell differ-entiation, they might trigger signaling pathways that, together withCD40-linked signaling, are involved in memory cell differentiationin vivo.
IL-2, IL-4 and IL-10 induce growth of B cells through activation ofJanus kinase (JAK)–signal transducer of activation and transcription(STAT) pathways. IL-4 activates STAT6 and STAT5 (refs. 14,15),whereas IL-2 and IL-10 activate STAT3 and STAT5 (ref. 16). STAT6promotes the choice of the immunoglobulin E (IgE) isotype duringclass-switch recombination17,18, whereas STAT3 is linked to plasma
Published online 13 February 2005; doi:10.1038/ni1172
1Department of Cell Biology and Histology of the Academic Medical Center, University of Amsterdam, 1105 AZ Amsterdam, Netherlands. 2Division of Immunology,Netherlands Cancer Institute, 1066 CX Amsterdam, Netherlands. 3These authors contributed equally to this work. Correspondence should be addressed to H.S.([email protected])
NATURE IMMUNOLOGY VOLUME 6 NUMBER 3 MARCH 2005 30 3
A R T I C L E S©
2005
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eim
mun
olog
y
cell differentiation6. The function of STAT5 in B cell development anddifferentiation is less well defined. Two forms of STAT5 (a and b) existthat are encoded by two different, tandemly linked genes. They haveboth unique and redundant functions in the response of cells to a widevariety of growth factors19. The bone marrow of STAT5-deficient miceis hypocellular and these mice are lymphopenic. In competitiverepopulation assays, the hematopoietic stem cells of STAT5-deficientmice have a strongly reduced ability to repopulate the B and T cellcompartments20. Although mature B cells are present in STAT5-deficient mice, their numbers are substantially reduced in peripheralblood, although not in the spleen or bone marrow21. The reducednumbers of pro– and pre–B cells in the STAT5-deficient mice andtheir reduced responsiveness to IL-7, which is essential for early B celldevelopment in mice, provides one explanation for the decreasednumbers of mature B cells21. Transgenic expression of activatedSTAT5b results in a substantial increase in pro–B cells in thesemice22. However, STAT5 is also involved in promoting survival ofmature lymphocytes. In T cells, IL-2-mediated STAT5 activationcorrelates with the induction of genes encoding the antiapoptoticfactors Bcl-XL and Bcl-2 (ref. 23). Thus, it is likely that STAT5isoforms participate in regulation of antiapoptotic genes in primaryB cells as well.
Here we describe a previously unknown function of STAT5 inmemory B cell differentiation. STAT5 was activated in situ in a smallproportion of B cells present in the human tonsil. PhosphorylatedSTAT5 (pSTAT5) was expressed in a small population of germinalcenter B cells that expressed CD95, CD25 and CD69 and low amountsof CD77 and lacked IgD expression, suggesting that they wereactivated centrocytes. The CD25+ B cells had high expression ofBcl-6 mRNA and low expression of Blimp-1 mRNA compared withthat of bulk centrocytes. This observation prompted us to examine theinvolvement of STAT5a and STAT5b in the differentiation and pro-liferation of human B cells in vitro. Using RNA interference, we foundthat decreased STAT5 expression impeded the expansion potential ofhuman B cells. In addition, the introduction of constitutively active
mutants of STAT5a or STAT5b into human B cells enhanced theproliferative activity and greatly extended their replicative lifespan invitro, indicating that terminal differentiation was inhibited by STAT5activation. Using a STAT5b construct that could be regulated, wefound that in primary B cells, BCL6 and BCL2L1, the gene encodingthe antiapoptotic factor Bcl-xL, were directly regulated by STAT5b.Control of activity of a BCL6 reporter construct by STAT5b wasdependent on the presence of STAT5 binding sites in the BCL6promoter. In addition, transcription of the Blimp-1-encoding genePRDM1 was blocked by STAT5b. These data strongly suggest thatmemory cell differentiation in germinal centers involves activation ofSTAT5, which inhibits apoptosis and induces Bcl-6.
RESULTS
Activated STAT5 in human germinal center B cells
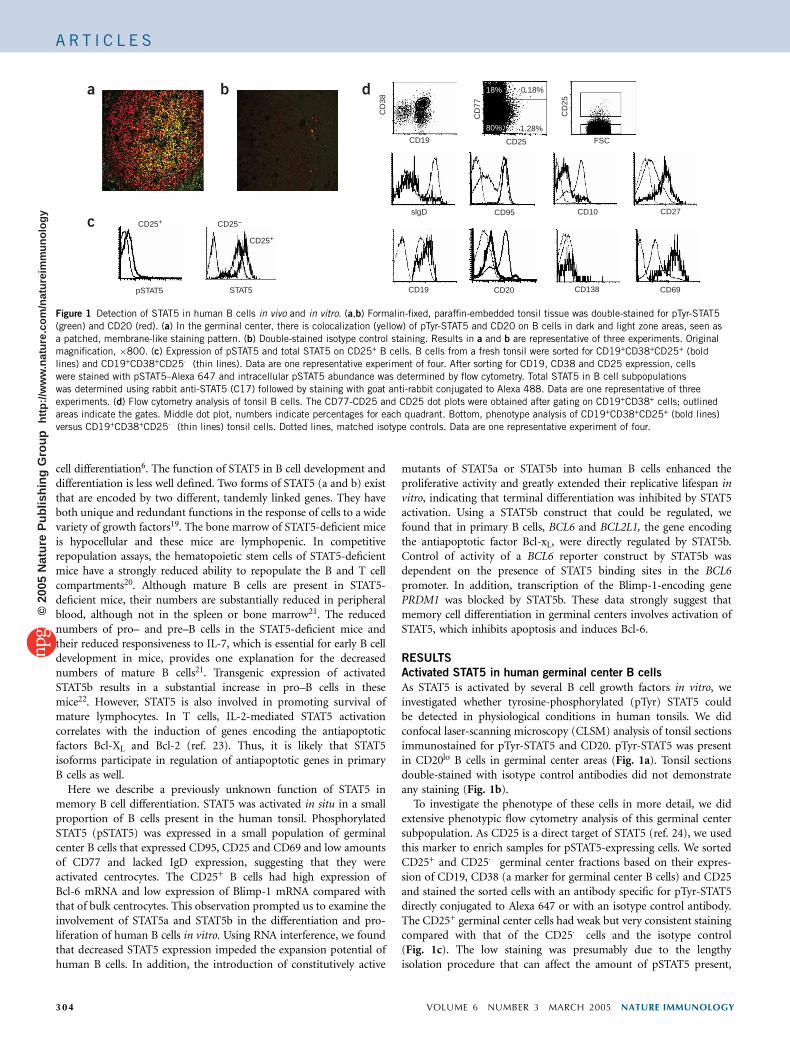
As STAT5 is activated by several B cell growth factors in vitro, weinvestigated whether tyrosine-phosphorylated (pTyr) STAT5 couldbe detected in physiological conditions in human tonsils. We didconfocal laser-scanning microscopy (CLSM) analysis of tonsil sectionsimmunostained for pTyr-STAT5 and CD20. pTyr-STAT5 was presentin CD20lo B cells in germinal center areas (Fig. 1a). Tonsil sectionsdouble-stained with isotype control antibodies did not demonstrateany staining (Fig. 1b).
To investigate the phenotype of these cells in more detail, we didextensive phenotypic flow cytometry analysis of this germinal centersubpopulation. As CD25 is a direct target of STAT5 (ref. 24), we usedthis marker to enrich samples for pSTAT5-expressing cells. We sortedCD25+ and CD25� germinal center fractions based on their expres-sion of CD19, CD38 (a marker for germinal center B cells) and CD25and stained the sorted cells with an antibody specific for pTyr-STAT5directly conjugated to Alexa 647 or with an isotype control antibody.The CD25+ germinal center cells had weak but very consistent stainingcompared with that of the CD25� cells and the isotype control(Fig. 1c). The low staining was presumably due to the lengthyisolation procedure that can affect the amount of pSTAT5 present,
CD69
CD27CD10CD95
CD138CD20CD19
slgD
CD19 CD25 FSC
CD
25
CD
77
CD
38
18%
80%
0.18%
1.28%
STAT5pSTAT5
CD25+ CD25–
CD25+
c
da b
Figure 1 Detection of STAT5 in human B cells in vivo and in vitro. (a,b) Formalin-fixed, paraffin-embedded tonsil tissue was double-stained for pTyr-STAT5
(green) and CD20 (red). (a) In the germinal center, there is colocalization (yellow) of pTyr-STAT5 and CD20 on B cells in dark and light zone areas, seen as
a patched, membrane-like staining pattern. (b) Double-stained isotype control staining. Results in a and b are representative of three experiments. Original
magnification, �800. (c) Expression of pSTAT5 and total STAT5 on CD25+ B cells. B cells from a fresh tonsil were sorted for CD19+CD38+CD25+ (bold
lines) and CD19+CD38+CD25� (thin lines). Data are one representative experiment of four. After sorting for CD19, CD38 and CD25 expression, cells
were stained with pSTAT5–Alexa 647 and intracellular pSTAT5 abundance was determined by flow cytometry. Total STAT5 in B cell subpopulations
was determined using rabbit anti-STAT5 (C17) followed by staining with goat anti-rabbit conjugated to Alexa 488. Data are one representative of three
experiments. (d) Flow cytometry analysis of tonsil B cells. The CD77-CD25 and CD25 dot plots were obtained after gating on CD19+CD38+ cells; outlined
areas indicate the gates. Middle dot plot, numbers indicate percentages for each quadrant. Bottom, phenotype analysis of CD19+CD38+CD25+ (bold lines)
versus CD19+CD38+CD25� (thin lines) tonsil cells. Dotted lines, matched isotype controls. Data are one representative experiment of four.
3 04 VOLUME 6 NUMBER 3 MARCH 2005 NATURE IMMUNOLOGY
A R T I C L E S©
2005
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eim
mun
olog
y

most likely through phosphatase activities. We also analyzed the totalamount of STAT5 present in the CD19+CD25+ and CD19+CD25�
germinal center (CD38+) populations by cytoplasmic staining. TheCD25+ population expressed twofold more STAT5 than did theCD25� population (Fig. 1c). From these observations we concludethat the CD25+ B cells are contained in the germinal center cells thatexpress pSTAT5.
To further analyze this population, we compared the phenotypes ofCD19+CD38+CD25+ cells versus CD19+CD38+CD25� cells (Fig. 1d).Germinal center CD25+ cells were almost completely negative for cellsurface IgD (sIgD). As sIgD is expressed on recent germinal centerimmigrants, this result indicated that germinal center CD25+ B cellswere not recent germinal center immigrants. Furthermore, we foundhigh expression of CD95 on CD25+ germinal center B cells, inaccordance with the finding that CD95 is expressed only on IgD�
germinal center B cells25. Germinal center B cells can be separated intotwo populations by CD77 expression, with centroblasts being CD77hi
and centrocytes being CD77lo. Freshly isolated germinal center CD25+
B cells had low expression of CD77, indicating that they representedcentrocytes (Fig. 1d). CD25+ B cells had only low expression of CD10,which is commonly regarded as a marker for germinal center B cells.CD10 is absent on memory B cells26. Thus, the CD25+ B cellsexpressed the germinal center marker CD38 and were only weaklyCD10+, indicative of a memory cell type. CD20 expression wassubstantially lower in the germinal center CD25+ B cells (Fig. 1d),consistent with the results of the CLSM analysis. The germinal centerCD25+ cells abundantly expressed the major histocompatibility com-plex class II molecule HLA-DR and the activation antigen CD69(Fig. 1d). In addition, approximately 50% of these cells expressedCD70. These results indicate that the CD25+ B cells were activatedin situ. Less than 2% of the germinal center CD25+ B cells expressedCD138, a plasma cell marker, and had abundant expression of CD27.
To confirm that the germinal center CD25+ B cell was a memoryB cell and not a plasma cell, we examined the mRNA expression ofBCL6, which encodes a known inhibitor of plasma cell differentiation(Fig. 2). Because the germinal center CD25+ B cells had a phenotypesimilar to that of centrocytes, we compared their expression of BCL6with that of bulk centrocytes (CD19+CD38+CD77lo). The germinalcenter CD25+ B cells had eightfold higher BCL6 transcript abundancethan did bulk centrocytes, as determined by real-time PCR. Inconcordance with the increased BCL6 expression in CD25+ germinalcenter cells compared with that of bulk centrocytes, the expression ofPRDM1 was one fourth that of CD25+ germinal center cells (Fig. 2).The lower PRDM1 expression in CD25+ B cells was expected, as Bcl-6protein directly downregulates transcription of the gene encodingBlimp-1 (refs. 5,27). As PRDM1, encoding Blimp-1, and BCL6 are‘master genes’ for plasma cell and memory cell differentiation7,8,respectively, our data indicate that germinal center CD25+ cellsrepresent activated B cells that are committed to the memory cell fate.
STAT5 ‘knockdown’ reduces human B cell proliferation
In contrast to plasma cells, memory B cells show self-renewingcapacity and can proliferate extensively in vitro after engagement ofCD40 and cytokine receptors. Cytokines such as IL-2, IL-10 and IL-4induce proliferation of B cells and activate STAT5. As pSTAT5 waspresent in a small population of germinal center B cells withcharacteristics of memory cells, we examined the function of STAT5in proliferation of B cells. We chose to ‘knock down’ STAT5 expressionby specifically targeting STAT5 for degradation using small interferingRNA (siRNA). We designed a self-inactivating (SIN) retroviral con-struct expressing green fluorescent protein under control of the pgk
promoter and STAT5 siRNA oligonucleotides (1365 and 1668) undercontrol of the pol3 promoter tested their knockdown efficiency in cellsof the human Burkitt lymphoma cell line Raji, which endogenouslyexpresses STAT5. We transduced Raji B cells with retroviruses expres-sing STAT5(1365) or STAT5(1668) siRNA or a control pSIN-GFPvirus and sorted the cells based on GFP expression. STAT5(1668)siRNA very efficiently reduced STAT5 protein expression comparedwith control-transduced Raji cells (Fig. 3a). STAT5(1365) siRNA wasless efficient but also substantially reduced STAT5 expression (Fig. 3a).To analyze the effect of STAT5 on B cell proliferation, we transducedthe STAT5(1668) siRNA probe in parallel with the control pSIN-GFPvirus into CD19+ sorted peripheral blood B cells and cultured these onCD40L-expressing mouse L-cell fibroblasts (CD40L-L cells) along withIL-2 and IL-4. We sorted GFP+ B cells and cultured them for 3 d, andthen determined their proliferation by [3H]thymidine incorporation.Reducing STAT5 protein expression reduced the proliferative capacityof the B cells by 50% compared with that of B cells expressing controlpSIN-GFP (Fig. 3b).
Human naive and memory B cells can be cultured in vitro for only alimited time after engagement of CD40 in the presence of cytokines,including IL-2, IL-4 and IL-10 (refs. 10,28). The results of the siRNAexperiment (Fig. 3b) indicated that activation of STAT5 was impor-tant for in vitro expansion of CD40L-activated B cell populations. Thisresult raises the issue of why B cells cultured with STAT5 activatingcytokines eventually stop proliferating. We therefore determinedwhether the amount of STAT5 protein present in B cells could be alimiting factor, which may cause nonresponsiveness to growth factors.We sorted human CD19+ B cells and cultured them on CD40L-L cellstogether with IL-2 and IL-4. We collected samples after 2 and 17 d ofculture and determined the amount of STAT5 protein. STAT5 proteinwas reduced in B cells after 17 d of culture compared with cultures atday 2 (Fig. 3c). Thus, the decrease in proliferation of CD40-activatedB cells correlated with a decrease in STAT5 protein abundance. Thesedata support the conclusion from our siRNA experiments that STAT5activation is important for in vitro B cell population expansion and atthe same time provide an explanation for the observation thatcultured B cells stop proliferating despite the presence of STAT5-activating cytokines.
Effect of STAT5 on primary human B cell population expansion
The identification of a small population of activated pSTAT5+
memory B cells and the observation that ‘knocking down’ STAT5expression limited the proliferative potential of primary B cells
CD25+CD77loCD25+CD77lo0
0.2
0.4
0.6
0.8
1
1.212
10
8
6
4
2
0
Rel
ativ
e B
CL6
exp
ress
ion
Rel
ativ
e P
RD
M1
expr
essi
on
Figure 2 Quantitative real-time PCR of BCL6 and PRDM1. Data represent
BCL6 and PRDM1 expression of tonsillar CD19+CD38+CD25+ B cells
relative to CD19+CD38+CD77lo B cells. All values were normalized to ACTB.
The error bars represent the standard deviation of triplicate samples of one
representative experiment of three.
NATURE IMMUNOLOGY VOLUME 6 NUMBER 3 MARCH 2005 30 5
A R T I C L E S©
2005
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eim
mun
olog
y

prompted us to examine the effects of ectopic expression of STAT5 inthe regulation of the survival, expansion and differentiation of humanB cell populations. We compared the effects of wild-type (WT) andconstitutively active (CA) mutants of STAT5a and STAT5b on theproliferation of human B cells. We cocultured purified tonsil CD19+
B cells with CD40L-L cells plus IL-2 and IL-4. At day 7 we transducedthe cells with retrovirus expressing CA-STAT5a-internal ribosomalentry site (IRES)–GFP, CA-STAT5b-IRES-GFP or WT-STAT5b-IRES-GFP or retrovirus containing ‘empty’ expression vector (control-IRES-GFP). Transduction efficiencies were 5–20% of the total B cell cultures.The percentages of GFP+ cells in the cultures transduced with CA-STAT5b-GFP (Fig. 4a) or CA-STAT5a-GFP (data not shown)increased over time. At weeks 5–6, the B cells transduced withcontrol-IRES-GFP and the untransduced B cells started to die, whereasat week 6 (5 weeks after transduction), more than 95% of the culturestransduced with CA-STAT5b-GFP were GFP+ and continued toproliferate. These GFP+ B cells could be kept in culture for morethan 300 d (data not shown). B cells transduced with WT-STAT5b-GFP selectively survived but their populations did not expand sub-stantially (Fig. 4a,b), indicating that activation of STAT5 was requiredfor expansion of the B cell populations. The expanded CA-STAT5b-GFP+ B cell population expressed CD19 but not CD20 (Fig. 4c). Toensure that the replicative lifespan extension of cells transduced withCA-STAT5b-GFP was not due to Epstein-Barr virus transformation,
we analyzed latent membrane protein 1 and Epstein Barr virus nuclearantigen 1 and 2 mRNA by RT-PCR and confirmed that these viralgenes were not expressed (data not shown). The B cells transducedwith CA-STAT5b-GFP expressed several rearranged immunoglobulinheavy-chain variable gene segments joined to Cm (Fig. 4d) or Cgconstant regions (data not shown), as determined by RT-PCR. Thisresult indicated that the transduced B cell lines were not monoclonal.These data collectively show that constitutive expression of activeSTAT5 can rescue in vitro–cultured human B cells from cell death andcan extend their replicative life span. Furthermore, these data suggestSTAT5 activation in B cells may inhibit terminal differentiation–induced cell cycle arrest.
Activation of STAT5 by tamoxifen in human B cells
To further confirm that constitutive activation of STAT5b, rather thana secondary transforming event, leads to an extended proliferativeresponse of human B cells, we examined the effect on B cellproliferation of a STAT5b construct that could be regulated. Forthis, we prepared a fusion of estrogen receptor (ER) with CA-STAT5b or WT-STAT5b. We generated recombinant retrovirus con-structs containing CA-STAT5b-ER or WT-STAT5b-ER upstream ofIRES-DNGFR, a truncated, signaling-incompetent mutant of thenerve growth factor receptor (NGFR). After introduction into cells,the estrogen receptor fusion protein is expressed in the cytoplasm as
Actin
STAT5
172Time (d)
86420
[3H]thymidine incorporation (103 c.p.m.)
si-CONTROL–GFP
si-STAT5(1668)–GFP
1 2 3
Actin
STAT5
a b c
Figure 3 Reduction of STAT5 expression limits the proliferative potential of primary human B cells. (a) Knockdown efficiency of siRNA probes (1365 and
1668) specifically targeting STAT5 mRNA. The Raji B cell line was transduced with pSIN-control-GFP (lane 1), pSIN-siSTAT5(1365)-GFP (lane 2) or pSIN-
siSTAT5(1668)-GFP (lane 3). Total cell lysates of GFP+ cells were analyzed for STAT5 expression. The blot was stripped and reprobed for actin as a loading
control. This experiment was repeated three times with the same results. (b) Proliferative response of human B cells transduced with either pSIN-control-GFP
or pSIN-siSTAT5(1668)-GFP. GFP+ B cells were sorted and were cultured for 3 d in duplicate in a 96-well plate with CD40L-L cells, IL-2 and IL-4, with the
last 18 h in the presence of [3H]thymidine (1m Ci/well). Incorporation of [3H]thymidine is expressed as counts per minute (c.p.m.) The error bars represent
the standard deviation among three experiments. (c) Human primary B cells were cultured (time, above lanes) on CD40L-L cells with IL-2 and IL-4, and
total cell lysates were analyzed by immunoblot for the abundance of STAT5. The blot was stripped and reprobed with a monoclonal antibody to total actin
(broad isoform range) to confirm equal loading of the lanes. Data are one representative of three experiments.
VH-Cµ
VH5VH4VH3VH2VH1/7
CD19 CD20
3532272218158
Time after transduction (d)
3532272218158
Time after transduction (d)
0
100
200
300
400
Abs
olut
e ce
ll nu
mbe
r (×
104 )
30
25
50
75
100
GF
P+ c
ells
(%
)
a
d
b c
Figure 4 Expression of CA-STAT5b leads to survival and expansion of B cell populations, whereas expression of WT-STAT5b results in survival only.
(a) CD19+ B cells were sorted from tonsil tissue, cultured with CD40L, IL-2 and IL-4, and transduced with control-IRES-GFP (open squares), WT-STAT5b-
IRES-GFP (filled circles) or CA-STAT5b-IRES-GFP (open circles), then were cultured further with CD40L, IL-2 and IL-4. Data are the percentages of GFP+
cells as determined by flow cytometry (time, horizontal axis) and are one representative experiment of three. (b) Absolute cell number counts determinedbased on the percentages of GFP+ cells; symbols as in a. (c) Flow cytometry showing the expression of CD19 and CD20 (bold lines) compared with isotype-
matched control antibody staining (thin lines) on B cells transduced with CA-STAT5b-IRES-GFP and cultured for 3 months in CD40L, IL-2 and IL-4.
(d) RT-PCR analysis of VH–Cm gene recombination in B cells transduced with CA-STAT5b-IRES-GFP cultured for 3 months as described in c. Results are
representative of three independent experiments.
3 06 VOLUME 6 NUMBER 3 MARCH 2005 NATURE IMMUNOLOGY
A R T I C L E S©
2005
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eim
mun
olog
y

an inactive complex sequestered by heat-shock proteins. After incuba-tion with 4-hydroxytamoxifen (4HT), the estrogen receptor fusionprotein dissociates from heat shock proteins and translocates to thenucleus29. Treatment of CA-STAT5b-ER-IRES-DNGFR+ B cells with4HT followed by staining with an estrogen receptor–specific antibodydemonstrated, by CLSM analysis, that CA-STAT5b-ER localized to thenucleus as expected in a 4HT-dependent way (Fig. 5a). Immunoblotanalysis confirmed the presence of the fusion protein of the expectedmolecular weight in the transduced cells (Fig. 5b).
We then transduced human tonsillar B cells with the CA-STAT5b-ER-IRES-DNGFR construct and examined the effect of 4HT on theproliferative capacity of these B cells. After culture of tonsillar B cellstransduced with CA-STAT5b-ER-IRES-DNGFR in the presence ofCD40L, IL-2 and IL-4, growth selection of CA-STAT5b-ER expressingcells (DNGFR+) occurred only in the presence of 4HT (Fig. 5c).Removal of 4HT from CA-STAT5b-ER-IRES-DNGFR+ cells resulted intermination of the growth of the B cells and eventual death (Fig. 5d).The CA-STAT5b-ER-IRES-DNGFR+ B cells cultured with CD40L, IL-2and IL-4 in the absence of 4HT initially proliferated, but they ceasedto grow and then died after 7–20 d. When 4HT was added again after7 d, the cells survived and then continued to grow (Fig. 5d). Theseresults indicated that CA-STAT5b-induced expansion of the tonsilB cell populations was not a consequence of irreversible geneticchanges induced by transformation of the B cells by CA-STAT5b butwas dependent on continued functional expression of CA-STAT5b inthe nucleus. Some of the tonsil B cells transduced with CA-STAT5b-ER-IRES-DNGFR and cultured for 3 months in CD40L, IL-2, IL-4 and4HT expressed cell surface IgM (Supplementary Fig. 1 online). Thecells were negative for cell surface IgD and a small percentage (1–2%)expressed IgG but lacked IgM. Furthermore, the CA-STAT5b-ER-IRES-DNGFR+ B cells expressed CD19, CD38, CD40, CD70, CD80and HLA-DR but were negative for CD20 and CD27 (SupplementaryFig. 1 online). CD25 was highly expressed on these cells (Supplemen-tary Fig. 1 online), consistent with our observation that CD25+ B cellsin the germinal center expressed pSTAT5 and the fact that STAT5bdirectly controls transcription of the gene encoding CD25 (IL2RA)24.
Notably, expression of the WT-STAT5b-ER fusion protein in B cellsresulted in a 4HT-dependent growth effect and phenotype similar tothat resulting from expression of the CA-STAT5b-ER protein in B cells(Supplementary Fig. 2 online). Presumably this result was due todimerization of estrogen receptor domains, as has been found forSTAT6-ER29, creating STAT5b-ER dimers that can be translocated tothe nucleus. Although IgG+ cells were present in the cultures of theB cells transduced with CA-STAT5b-ER (Supplementary Fig. 2online), we wanted to confirm that STAT5b-ER transduction led tocontinued expansion of purified IgG+ memory cell populations.Transduction of WT-STAT5b-ER resulted in a 4HT-dependent expan-sion of IgG+ B cell populations, whereas IgG+ B cells transduced withcontrol-GFP did not survive beyond 40 d (Supplementary Fig. 2online). We also transduced total B cells with WT-STAT5-ER, sortedIgM+ and IgG+ cells to 96% purity after 19 d of culture, and culturedthese cells in the presence of 4HT. Fifteen days later we noted 30- and200-fold expansion of the IgM+ and IgG+ cell populations, respec-tively. Flow cytometry of the IgG+ and IgM+ B cells expressingWT-STAT5b-ER at day 36 in culture (the control-GFP transducedcells already died by then) showed that these cells maintained sIgexpression (Supplementary Fig. 2 online). Collectively, these dataindicate that both IgM+ and IgG+ B cells transduced with activeSTAT5 maintain their in vitro proliferative capacity for prolongedperiods of time.
Bcl-6 extends the lifespan of normal human B cells
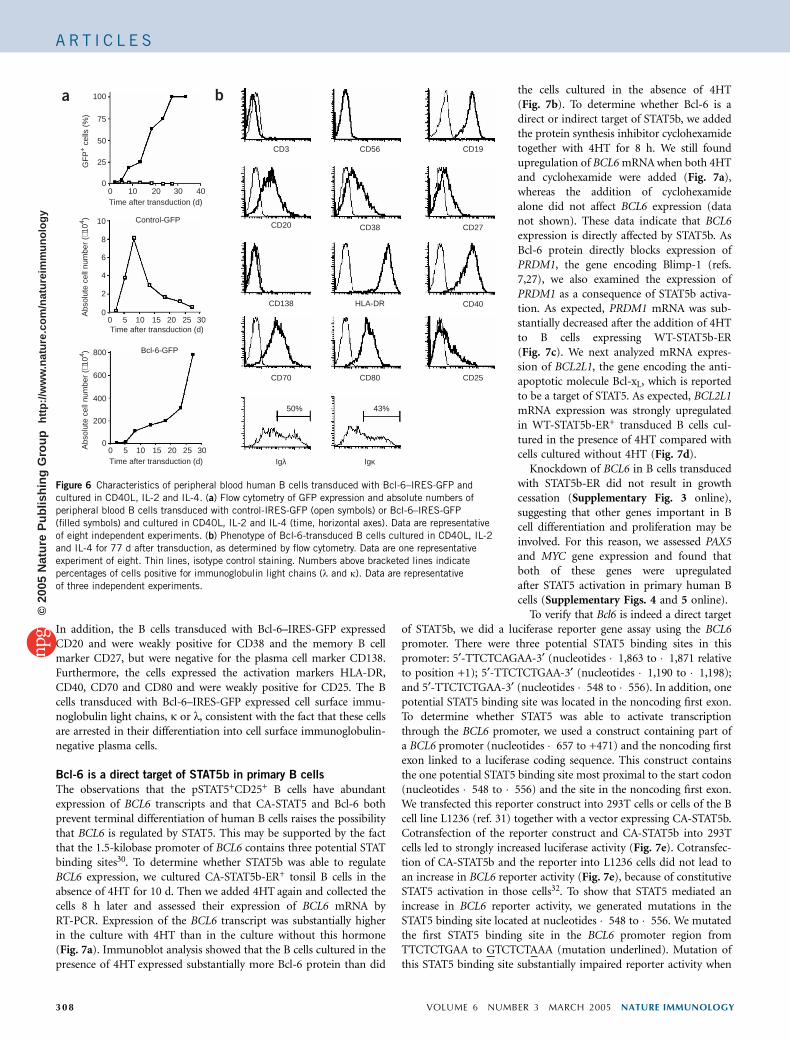
Ectopic expression of Bcl-6 in human tonsil B cells from youngchildren results in prevention of eventual cell cycle arrest of B cellswhen they are cultured in CD40L, IL-2 and IL-4 (ref. 12). To extendthose findings, we investigated whether Bcl-6 also maintains the self-renewal capacity of adult peripheral blood B cells. Therefore weintroduced Bcl-6–IRES-GFP into human B cells cultured withCD40L-L cells, IL-2 and IL-4. Expression of Bcl-6 resulted in a growthadvantage of the peripheral B cells expressing the marker GFP starting10–13 d after transduction of Bcl-6 (Fig. 6a). Phenotypically,B cells transduced with Bcl-6–IRES-GFP expressed CD19 (Fig. 6b).
Figure 5 CA-STAT5b-ER localizes to the nucleus
and induces B cell population expansion in a
4HT-dependent way. (a) B cells transduced with
CA-STAT5b-ER-IRES-DNGFR were stained with
anti-NGFR–fluorescein isothiocyanate (green)
and anti–estrogen receptor–Texas Red (red)
in the absence (left) or presence (right) of
4HT and were analyzed by CLSM. Imagesare representative of two experiments.
(b) Immunoblot analysis of cellular extracts of
CA-STAT5b-ER-IRES-DNGFR+ B cells with an
anti-STAT5 (right) or anti-estrogen receptor (left).
This experiment was done twice with similar
results. (c) Tonsil B cells were cultured in
CD40L, IL-2 and IL-4 and were transduced with
CA-STAT5b-ER-IRES-DNGFR (circles) or with
control-IRES-DNGFR (squares). The cultures were
split (downward arrow); one part was cultured
without 4HT (open symbols) and the other
was cultured with 1 mM 4HT (filled symbols).
After transduction (time, horizontal axes), the
percentages of transduced cells were determined
by flow cytometry after staining with an
allophycocyanin-conjugated antibody to NGFR. Absolute cell numbers were determined based on the percentages of DNGFR+ cells. Data are representative
of five independent experiments. (d) B cells were transduced with CA-STAT5b-ER-IRES-DNGFR and 30 d later were cultured for 7 d in the absence of 4HT.
Equal numbers of cells were then cultured in the presence (filled squares) or absence (open squares) of 4HT and absolute cell numbers were determined.
Data are one representative experiment of three.
252015105Time after 4HT
Time after tranduction (d)Time after tranduction (d) withdrawal or addition (d)
100
75
50
25
0
100
75
50
25
0
Abs
olut
e ce
ll nu
mbe
r (×
104 )
Abs
olut
e ce
ll nu
mbe
r (×
104 )
48423935292217
400
300
200
100
0494136292822173
∆NG
FR
+ c
ells
(%
)
STAT5
STAT5–ER
STAT5
ERkDa
115
93
–4HT +4HTa
c d
b
NATURE IMMUNOLOGY VOLUME 6 NUMBER 3 MARCH 2005 30 7
A R T I C L E S©
2005
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eim
mun
olog
y
In addition, the B cells transduced with Bcl-6–IRES-GFP expressedCD20 and were weakly positive for CD38 and the memory B cellmarker CD27, but were negative for the plasma cell marker CD138.Furthermore, the cells expressed the activation markers HLA-DR,CD40, CD70 and CD80 and were weakly positive for CD25. The Bcells transduced with Bcl-6–IRES-GFP expressed cell surface immu-noglobulin light chains, k or l, consistent with the fact that these cellsare arrested in their differentiation into cell surface immunoglobulin-negative plasma cells.
Bcl-6 is a direct target of STAT5b in primary B cells
The observations that the pSTAT5+CD25+ B cells have abundantexpression of BCL6 transcripts and that CA-STAT5 and Bcl-6 bothprevent terminal differentiation of human B cells raises the possibilitythat BCL6 is regulated by STAT5. This may be supported by the factthat the 1.5-kilobase promoter of BCL6 contains three potential STATbinding sites30. To determine whether STAT5b was able to regulateBCL6 expression, we cultured CA-STAT5b-ER+ tonsil B cells in theabsence of 4HT for 10 d. Then we added 4HT again and collected thecells 8 h later and assessed their expression of BCL6 mRNA byRT-PCR. Expression of the BCL6 transcript was substantially higherin the culture with 4HT than in the culture without this hormone(Fig. 7a). Immunoblot analysis showed that the B cells cultured in thepresence of 4HT expressed substantially more Bcl-6 protein than did
the cells cultured in the absence of 4HT(Fig. 7b). To determine whether Bcl-6 is adirect or indirect target of STAT5b, we addedthe protein synthesis inhibitor cyclohexamidetogether with 4HT for 8 h. We still foundupregulation of BCL6 mRNA when both 4HTand cyclohexamide were added (Fig. 7a),whereas the addition of cyclohexamidealone did not affect BCL6 expression (datanot shown). These data indicate that BCL6expression is directly affected by STAT5b. AsBcl-6 protein directly blocks expression ofPRDM1, the gene encoding Blimp-1 (refs.7,27), we also examined the expression ofPRDM1 as a consequence of STAT5b activa-tion. As expected, PRDM1 mRNA was sub-stantially decreased after the addition of 4HTto B cells expressing WT-STAT5b-ER(Fig. 7c). We next analyzed mRNA expres-sion of BCL2L1, the gene encoding the anti-apoptotic molecule Bcl-xL, which is reportedto be a target of STAT5. As expected, BCL2L1mRNA expression was strongly upregulatedin WT-STAT5b-ER+ transduced B cells cul-tured in the presence of 4HT compared withcells cultured without 4HT (Fig. 7d).
Knockdown of BCL6 in B cells transducedwith STAT5b-ER did not result in growthcessation (Supplementary Fig. 3 online),suggesting that other genes important in Bcell differentiation and proliferation may beinvolved. For this reason, we assessed PAX5and MYC gene expression and found thatboth of these genes were upregulatedafter STAT5 activation in primary human Bcells (Supplementary Figs. 4 and 5 online).
To verify that Bcl6 is indeed a direct targetof STAT5b, we did a luciferase reporter gene assay using the BCL6promoter. There were three potential STAT5 binding sites in thispromoter: 5¢-TTCTCAGAA-3¢ (nucleotides �1,863 to �1,871 relativeto position +1); 5¢-TTCTCTGAA-3¢ (nucleotides �1,190 to �1,198);and 5¢-TTCTCTGAA-3¢ (nucleotides �548 to �556). In addition, onepotential STAT5 binding site was located in the noncoding first exon.To determine whether STAT5 was able to activate transcriptionthrough the BCL6 promoter, we used a construct containing part ofa BCL6 promoter (nucleotides �657 to +471) and the noncoding firstexon linked to a luciferase coding sequence. This construct containsthe one potential STAT5 binding site most proximal to the start codon(nucleotides �548 to �556) and the site in the noncoding first exon.We transfected this reporter construct into 293T cells or cells of the Bcell line L1236 (ref. 31) together with a vector expressing CA-STAT5b.Cotransfection of the reporter construct and CA-STAT5b into 293Tcells led to strongly increased luciferase activity (Fig. 7e). Cotransfec-tion of CA-STAT5b and the reporter into L1236 cells did not lead toan increase in BCL6 reporter activity (Fig. 7e), because of constitutiveSTAT5 activation in those cells32. To show that STAT5 mediated anincrease in BCL6 reporter activity, we generated mutations in theSTAT5 binding site located at nucleotides �548 to �556. We mutatedthe first STAT5 binding site in the BCL6 promoter region fromTTCTCTGAA to GTCTCTAAA (mutation underlined). Mutation ofthis STAT5 binding site substantially impaired reporter activity when
IgκIgλ
43%50%
CD40
CD70 CD80 CD25
HLA-DRCD138
CD27CD38
CD3 CD56 CD19
CD20
302520151050Time after transduction (d)
302520151050Time after transduction (d)
Time after transduction (d)403020100
100
75
50
25
0
GF
P+ c
ells
(%
)
10
8
6
4
2
0Abs
olut
e ce
ll nu
mbe
r (×
104 )
Abs
olut
e ce
ll nu
mbe
r (×
104 ) 800
600
400
200
0
Control-GFP
Bcl-6-GFP
a b
Figure 6 Characteristics of peripheral blood human B cells transduced with Bcl-6–IRES-GFP and
cultured in CD40L, IL-2 and IL-4. (a) Flow cytometry of GFP expression and absolute numbers of
peripheral blood B cells transduced with control-IRES-GFP (open symbols) or Bcl-6–IRES-GFP
(filled symbols) and cultured in CD40L, IL-2 and IL-4 (time, horizontal axes). Data are representative
of eight independent experiments. (b) Phenotype of Bcl-6-transduced B cells cultured in CD40L, IL-2
and IL-4 for 77 d after transduction, as determined by flow cytometry. Data are one representative
experiment of eight. Thin lines, isotype control staining. Numbers above bracketed lines indicate
percentages of cells positive for immunoglobulin light chains (l and k). Data are representative
of three independent experiments.
3 08 VOLUME 6 NUMBER 3 MARCH 2005 NATURE IMMUNOLOGY
A R T I C L E S©
2005
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eim
mun
olog
y

CA-STAT5b was cotransfected in L1236 cells (Fig. 7e, right). Weobtained similar results when we transfected CA-STAT5b and thereporter constructs into 293T cells (Fig. 7e, left). These data demon-strate that the STAT5 binding site at nucleotides �548 to �556 in theBCL6 promoter is functional. Collectively, our results show thatSTAT5 directly regulates Bcl-6 mRNA and protein expression.
DISCUSSION
Here we have documented a function for STAT5 in the regulation ofproliferation and differentiation of memory B cells. A small popula-tion of germinal center CD25+ B cells expressed pSTAT5 and increasedamounts of total STAT5 and had an activated phenotype. TheseCD25+pSTAT5+ cells are probably centrocytes that are committed tothe memory B cell lineage, based on their surface phenotype, theirabundance of BCL6 mRNA and their low PRDM1 mRNA expression.As Blimp-1 is required for and Bcl-6 inhibits plasma cell differentia-tion8, these data indicate that the CD25+ cells are directed to thememory cell fate. Consistent with this, these B cells have highexpression of CD27. Ligation of CD27 on B cells enhances memorycell differentiation33 and inhibition of terminal differentiation ofactivated B cells into plasma cells34.
We propose an important function for STAT5 in B cell proliferationbased on the following observations. Knockdown STAT5 expression byRNA interference in B cells reduced their expansion potential. STAT5protein abundance was downregulated in B cells after prolongedin vitro culture with CD40L and cytokines. Introduction of constitu-tively active mutants of STAT5a or STAT5b or of STAT5b-ER fusionproteins substantially extended human B cell lifespan in vitro. How-ever, cells expressing STAT5b-ER cultured with 4HT required CD40Land either IL-2 or IL-4 for optimal growth. Thus, activation of STAT5in this system was by itself not sufficient to fully replace the signalsprovided by growth factors. Obviously, other signal transductionmolecules triggered by cytokines and CD40 ligation, such as PI-3K,NF-kB and p38 kinase35,36, are likely to induce cell cycle progression.
Although STAT5b-ER+ B cell populations cultured with 4HT, butwithout CD40L or cytokines, did not expand, they showed a survivaladvantage relative to that of cells cultured without 4HT (data notshown). This was likely to be the result of STAT5b-induced Bcl-xL
protein expression, because both B cells transduced with WT-STAT5-ER and those transduced with CA-STAT5-ER expressed BCL2L1mRNA but not BCL2 mRNA or mRNA of genes encoding otherantiapoptotic molecules after 4HT addition (data not shown). AsBCL2L1 expression in B cells transduced with WT-STAT5-ER or CA-STAT5-ER is dependent on 4HT, BCL2L1 is a direct target of STAT5,consistent with the finding that STAT5 consensus binding sites arepresent in the promoter of BCL2L1, encoding Bcl-xL (ref. 37). Theobservations that active STAT5a and STAT5b prolonged the in vitroproliferation of IgM+ and memory IgG+ B cells and that cessation of Bcell proliferation in vitro correlated with decreased STAT5 proteinabundance strongly suggest that activated STAT5 prevent terminaldifferentiation–induced cell cycle arrest.
We have confirmed published finding that Bcl-6 overexpressionextends human tonsillar B cell lifespan in vitro12 and have demon-strated that Bcl-6 also extended the lifespan of adult peripheral bloodB cells. As the effects of CA-STAT5b and Bcl-6 on B cell proliferationwere comparable (data not shown) and screening of the human BCL6promoter30 showed the presence of three potential STAT5 bindingsites37, we hypothesized that STAT5 may regulate Bcl-6 expression.Using an inducible STAT5b-ER construct, we demonstrated that Bcl-6was regulated by STAT5b. Bcl-6 was upregulated by the preformedCA-STAT5b-ER protein even in the presence of the protein synthesisinhibitor cyclohexamide, indicating that BCL6 is a direct target ofSTAT5b. Moreover, we were able to demonstrate that at least one ofthe STAT5 binding sites in the BCL6 promoter was functional, asmutation of this site strongly diminished the capacity of STAT5bto induce expression of a luciferase reporter downstream of theBCL6 promoter. The demonstration that STAT5b directly regulatesBCL6 provides insight into the mechanisms of the effect of STAT5b
Figure 7 Induction of BCL6 expression by
STAT5. (a) RT-PCR analysis of BCL6 and
hypoxanthine guanine phosphoribosyl transferase
(HPRT1) in CA-STAT5b-ER+ B cells cultured in
the presence of CD40L, IL-2, IL-4 and constant
4HT (+4HT (constant)). Cells were deprived of
4HT for 10 d (�4HT), then 4HT was added back
for 8 h in the presence (+4HT +CH) or absence(+4HT) of cyclohexamide. (b) Immunoblot
analysis of the expression of Bcl-6 in CA-STAT5b-
ER–transduced B cells cultured for 12 d in the
constant presence (+4HT) or absence of 4HT
(�4HT). Actin, loading control. (c) CD19+
WT-STAT5B-ER+ B cells were cultured in the
presence of CD40L, IL-2, IL-4, and 4HT, then
were maintained for 11 d in the continuous
presence (+4HT constant) or absence of 4HT
(�4HT). PRDM1, BCL6 and HPRT1 were
analyzed by RT-PCR. (d) WT-STAT5B-ER+
B cells were cultured in the presence of C40L,
IL-2, IL-4 and 4HT, were deprived of 4HT for
11 d, and then were cultured for 48 h in the
presence (+4HT 48h) or absence (�4HT) of 4HT.
BCL2L1 and HPRT1 were analyzed by RT-PCR.
(e) Dual luciferase reporter assay. One potential
STAT5 binding site in the BCL6 promoter region
was mutated. Two different clones with the same mutation (BCL6* and BCL6**) were subcloned into the pGL3–firefly luciferase backbone and withLZRS-CA-STAT5b into 293T cells (left) or the B cell line L1236 (right). Error bars represent standard deviation among measurements. All data are
representative of two or three independent experiments.
+
+++
+
+
+
+++
+
+
+
–––––
– – – – –
––––
–
pGL3–Bcl-6**
pGL3–Bcl-6*
pGL3–Bcl-6
LZRS–CA–STAT5b
1.2
1
0.8
0.6
0.4
0.2
0
L1236293T8
7
6
5
4
3
2
1
0
Rel
ativ
e lu
cife
rase
uni
ts
HPRT1HPRT1
HPRT1
BCL2L1
BCL6BCL6
PRDM1
–4HT
–4HT
–4HT
+4HT
+4HT (c
onsta
nt)
+4HT 4
8h
+4HT +
CH
+4HT
+4HT (c
onsta
nt)
–4HT
Actin
Bcl-6
–4HT (10d)a
e
b c
d
NATURE IMMUNOLOGY VOLUME 6 NUMBER 3 MARCH 2005 30 9
A R T I C L E S©
2005
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eim
mun
olog
y

activation on B cells. Bcl-6 controls B cell differentiation in partby repressing Blimp-1 (ref. 7), which drives plasma cell differentia-tion5–8. Bcl-6 protein binds to the PRDM1 promoter in vivo, indicat-ing that the gene encoding Blimp-1 is a direct target of Bcl-6-mediatedtranscriptional repression27. In correlation with that, BCL6-deficientmice lack germinal centers and show increased antibody responses andincreased numbers of Blimp-1+ antibody-forming cells27. Blimp-1inhibits transcription of MYC, which is essential for cell cycleprogression of B cells cultured with CD40L and IL-4 (ref. 38). It istherefore likely that ectopic expression of active STAT5b or Bcl-6 in Bcells turns off Blimp-1 protein expression and therefore enablescontinued expression of MYC. Our observations that activation ofCA-STAT5b-ER by 4HT upregulates MYC mRNA expression in B cellsis in agreement with that.
Our findings are consistent with a published model6,13 showing thatBcl-6 prevents terminal differentiation of germinal center B cells andmemory B cells, thereby preventing cell cycle arrest. Our findingstherefore suggest that activated STAT5b inhibits terminal differentia-tion of the mature B cells at least in part by regulating Bcl-6. However,the induction of BCL6 by STAT5 is unlikely to be the sole reason forprevention of cell cycle arrest and inhibition of terminal differentiationof STAT5b-transduced B cells, as we found that ‘knocking down’ BCL6expression by siRNA in B cells transduced with CA-STAT5b-ER didnot affect expansion of the cell populations cultured with CD40L, IL-2and IL-4. Moreover, Pax5, which stimulates the proliferation ofmature B cells and inhibits differentiation into plasma cells bysuppressing XBP-1 (refs. 39,40), was also upregulated by STAT5.This suggests that Pax5 could be responsible for the inhibition ofterminal differentiation by active STAT5 when Bcl-6 is downregulated.The idea that upregulation of BCL6 by active STAT5b is not the onlyexplanation for the effects of continued activation of STAT5b oncultured human B cells is consistent with some other observations. Anotable difference between STAT5b-transduced and Bcl-6-transducedB cells is that the latter cells consistently express sIg, whereas STAT5btransduced B cells gradually lose sIg expression after long-term in vitroculture (data not shown); the mechanism for this is not yet known.Moreover, although the cell surface expression of many B cell activa-tion antigens, such as CD40, CD70, CD80 and CD86, is very similar,STAT5b-transduced B cells do not express CD20 and CD27 and havemuch higher expression of CD25 than do Bcl-6-transduced B cells.The absence of the memory marker CD27 on STAT5b-transduced Bcells cultured in vitro in contrast to its expression on freshly isolatedCD25+ pSTAT5+ B cells ex vivo is puzzling. This is not due to in vitrogrowth selection of CD27� cells, as purified CD27+ and CD27� cellpopulations transduced with activated STAT5b expand in a similarway. However, CD27 was downregulated in CD27+ cells transducedwith STAT5-ER and cultured with 4HT by an as-yet-unknownmechanism. It is likely that the differences in expression of certaincell surface antigens are due to different transcriptional programs thatare turned on or off by continued (over)expression of Bcl-6 andactivated STAT5b.
The identity of the triggering agents that activate STAT5 in vivo areunknown. IL-4 is involved in isotype switching and thus in memorycell formation, but this activity is mediated through STAT6. Analysisof mice deficient in IL-2 or IL-10, cytokines that can activate STAT5,have not demonstrated essential functions for these cytokines in thegeneration of memory B cells, but the possibility that they haveredundant functions with respect to STAT5 activation-inducedmemory B cell generation cannot be excluded. Triggering the BCRcan activate STAT5 (ref. 41), raising the possibility that the BCR isinvolved in activation of STAT5 leading to memory cell differentiation.
The affinity of the BCR is one of the factors that determine plasma ormemory B cell differentiation3.
Our data showed that STAT5b regulates BCL6, but as germinalcenter B cells that do not express activated STAT5 can be Bcl-6+, thisindicates that activated STAT5 isoforms are not strictly required forBcl-6 expression. Our data, however, indicate that STAT5 isoforms domodulate Bcl-6 transcription, thereby facilitating commitment tomemory B cells. Until now, it was not appreciated that activatedSTAT5 regulates BCL6. Although BCL6-deficient mice lack germinalcenters42–44, the B cells in these mice are able to produce IgM andIgG1, indicating that there is capacity for isotype switching45; however,the immunoglobulins produced lack somatic hypermutations, a hall-mark of memory B cell responses21,45. It is unknown whether germinalcenters are formed in STAT5-deficient mice and whether somatichypermutations occur in the Ig genes of these mice. Analysis of the Bcell responses in mice conditionally deficient in both STAT5a andSTAT5b in B cells is needed for further analysis of the function ofSTAT5 in memory B cell responses.
METHODSB cell isolation. B cells were obtained from tonsils or peripheral blood of adults.
Tonsillectomies were done in the Department of Surgery for Children of the
Free University of Amsterdam and from the Academic Medical Center
(Amsterdam, Netherlands). The use of this tissue was approved by the medical
ethical committees of our institutions and was contingent on informed consent.
Samples were depleted of T cells using microbeads coated with antibody to CD4
(anti-CD4) and anti-CD8 (Miltenyi Biotec). Next, the cells were incubated with
fluorescein isothiocyanate–conjugated anti-CD19 (DAKO) and phycoerythrin-
conjugated anti-CD3 (Becton Dickinson), followed by sorting of the
CD19+CD3� population with a MoFlo (Cytomation/DAKO), a FacStarPlus
(Becton Dickinson) or a FacsARIA (Becton Dickinson). The resulting B cells
were more than 99% pure after reanalysis. B cells isolated from tonsil or
peripheral blood with CD19 microbeads (Miltenyi) yielded identical results.
Retroviral constructs and production of recombinant retrovirus. Constitu-
tively active mutants of STAT5a and STAT5b have been described46. DNA
encoding these mutants and WT-STAT5b was obtained from T. Kitamura
(IMSUT, Tokyo, Japan). BCL6 was identified in a senescence rescue screen in
mouse fibroblasts as an inhibitor of anti-proliferative p19ARF-p53 signaling12.
cDNA encoding these proteins was ligated into Lazarus (LZRS)-linker-IRES-
GFP vector that has been published47. For knockdown experiments, the
pSUPER construct48 was adapted. For the identification of transduced cells
by flow cytometry, a GFP-expressing cassette was added to the pSUPER
construct such that the polymerase III H1-RNA promoter (pol3) for the
transcription of the siRNA probe and the phosphoglycerin kinase (pgk)1
promoter driving GFP expression were in opposite directions. The siRNA
sequences specifically targeting STAT5 mRNA (1365, 5¢-GCAGCAGACCAT
CATCCTG-3¢; 1668, 3¢-GACCCAGACCAAGTTTGCA-5¢) and BCL6 mRNA
(789, 5¢-TGTGTGCCACAGCAATATC-3¢; 970, 3¢-GATGAGATTGCCCTG
CATT-5¢) were designed using the Ambion website (http://www.ambion.com).
Sequences were inserted into the BglII-HindIII sites of pSUPER-GFP.
Subsequently, the pol3-siRNA-pgk-GFP cassette was subcloned into the self-
inactivating version of the LZRS retroviral construct.
The retroviral plasmids were transfected into the helper virus–free ampho-
tropic producer cell line Phoenix-A, a derivative of the human embryonic
kidney cell line 293 (a gift from G. Nolan, Stanford University, Palo Alto,
California), with Fugene-6 (Roche Diagnostics Netherlands), according to
manufacturer’s protocols. Then, 2 d later, selection of transfected cells was
begun by the addition of 2 mg/ml of puromycin (Becton Dickinson Clontech
Laboratories). At 10–14 d after transfection, 6 � 106 cells were plated per 10-cm
Petri dish (Becton Dickinson Discovery Labware) overnight in 10 ml of
complete medium (Iscove’s medium (Life Technologies BV) plus 8% FCS,
penicillin and streptomycin without puromycin). The medium was refreshed
the next morning and 24 h later retroviral supernatant was collected, centri-
fuged and frozen in cell-free aliquots at �70 1C. This approach affords a
3 10 VOLUME 6 NUMBER 3 MARCH 2005 NATURE IMMUNOLOGY
A R T I C L E S©
2005
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eim
mun
olog
y

reproducible rapid, large-scale and high-titer retroviral production of more
than 3 � 106 infectious virus particles/ml.
The constitutively active and wild-type STAT5b-ER fusion constructs were
made as follows: PCR was used to introduce a BglII site in place of the stop
codon in the N604H STAT5b mutant46 or WT-STAT5b cDNA. An XhoI-BglII
digestion product was generated, which was ligated with the product of a
BamHI-EcoRI digestion of the c-terminal part of the estrogen receptor
(provided by H. Kurata, DNAX Institute, Palo Alto, California)29. This
construct was cloned into LZRS-CA-STAT5b or WT-STAT5b to create LZRS-
CA-STAT5b-ER and LZRS-WT-STAT5b-ER. In all LZRS constructs, the STAT5
genes were upstream of IRES-DNGFR (a signaling-incompetent mutant of
NGFR; provided by C. Bonini, St. Raphael Hospital, Milan, Italy) or IRES-GFP.
A monoclonal antibody to NGFR (ME20.4, Chromaprobe) was used to
visualize DNGFR-expressing cells.
Retroviral transduction. The ‘recombinant human fibronectin fragments CH-
296 transduction’ procedure (RetroNectin; Takara) was done as described47.
Non–tissue culture-treated 24-well plates (Costar) were coated with 0.3 ml of
30 mg/ml of recombinant human fibronectin fragment CH-296 at 25 1C for 2 h
or overnight at 4 1C. The CH-296 solution was removed, followed by
incubation for 30 min at 25 1C with 2% human serum albumin in PBS,
followed by one wash with PBS. B cells (5 � 105) were plated in 0.25 ml
complete medium mixed with 0.25 ml of thawed retroviral supernatant and
polybrene (final concentration, 4 mg/ml) and were incubated for 6 h at 37 1C.
Next, 0.25 ml of supernatant was removed and 0.25 ml of fresh retroviral
supernatant plus polybrene was added, followed by incubation at 37 1C
overnight. The next morning (approximately 16 h later), cells were washed
and transferred to 24-well tissue culture–treated plates (Costar) with irradiated
(80 Gy) CD40L-L cells expressing CD40L (CD154), plus IL-2 (20 U/ml) and
IL-4 (50 ng/ml).
Cell culture. B cells were cultured in complete medium at 37 1C in humidified
air containing 5% CO2. Irradiated CD40L-L cells were seeded at a density of
5 � 104 cells per well in 24-well tissue culture–treated plates (Costar). Sorted
B cells (5 � 105) were added together with IL-2 (20 U/ml) and IL-4 (10 ng/ml;
R&D Systems). After 1 week, the cells were used for retroviral transduction.
After transduction, B cells were cultured again with irradiated CD40L-L cells,
IL-2 and IL-4.
Immunohistological staining for pTyr-STAT5 and CD20. Human tonsil
tissues were obtained from children undergoing tonsillectomy, and immuno-
histological analysis of pTyr-STAT5 and CD20 was done with immunofluor-
escence and CLSM analysis as described49, with minor modifications49.
Formalin-fixed, paraffin-embedded human tonsil sections, after citrate retrie-
val, were preincubated in 5% (volume/volume) normal goat serum (Central
Laboratory of the Netherlands Red Cross Blood Transfusion Service, Sanguin).
Subsequently, sections were incubated at 4 1C overnight with rabbit anti-
pTyr-STAT5b (Cell Signaling Technology) and mouse anti-CD20 (L26,
DAKO), followed by incubations in biotinylated goat anti-rabbit IgG (DAKO),
streptavidin–biotin-conjugated–horseradish peroxidase complex (ABC proto-
col; DAKO) and tyramide–AlexaFluor 568 (Molecular Probes Europe). CD20
was visualized with AlexaFluor 633–conjugated goat anti-mouse IgG (Mole-
cular Probes). Between incubations, sections were rinsed extensively in PBS
containing 1% BSA (Sigma Aldrich). Matched isotype control antibodies
(normal rabbit IgG and mouse IgG1 for pSTAT5 and CD20 staining, respec-
tively) were included as negative controls. Confocal fluorescence images were
obtained on a Leica TCS SP (Leica Microsystems) confocal system, equipped
with a Kr/HeNe laser combination. Images were obtained with a 40�,
1.25–numerical aperture objective. Color photomicrographs were obtained
from electronic overlays.
For localization of STAT5 expression, B cells transduced with CA-STAT5b-
ER-IRES-DNGFR were attached onto polysine-coated coverslips. After fixation
with 3.7% formaldehyde at 25 1C for 15 min, cells were permeablized for 45 s
with 0.1% Triton X-100. Primary antibodies were mouse anti-NGFR and rabbit
anti–estrogen receptor (MC-20, Santa Cruz Biotechnology). Secondary anti-
bodies were fluorescein isothiocyanate–anti-mouse (BD) and Texas Red–anti-
rabbit (Molecular Probes). Confocal fluorescence images were obtained on a
Leica TCS SP (Leica).
Phenotyping of B cells. Monoclonal antibodies (all IgG1 unless otherwise
indicated) to the human molecules IgD (IA6-2, IgG2a), IgG (G18-145), CD3
(SK7), CD10 (HI10a), CD19 (4G7, SJ25C1, ID3 (IgG2a), or HIB19), CD20
(2H7, IgG2b), CD25 (2A3), CD27 (L128), CD38 (HB7), CD40 (5C3), CD45
(2D1), CD56 (NCAM16.2, IgG2b or B159), CD69 (L78), CD70 (Ki24, IgG3),
CD80 (L307.4), CD86 (It2.2, IgG2b), CD95 (DX2) and HLA-DR (L243; IgG2a;
BD-Pharmingen) were directly labeled with fluorescein isothiocyanate,
phycoerythrin, peridinine chlorophyll protein, allophycocyanin, phycoery-
thrin-indotricarbocyanine or allophycocyanin-indotricarbocyanine. Antibodies
to IgM (rabbit polyclonal), k light chain (clone A8B5), l light chain (rabbit
polyclonal) and CD138 (clone MI15), directly labeled with phycoerythrin
(DAKO), were used for flow cytometry analysis. CD77 was measured with anti-
CD77 (clone 38.13; rat IgM; Coulter-Immunotech) followed by phycoerythrin–
anti-rat IgM (BD). Stained cells were analyzed with a FACSCalibur or LSR II
(Becton Dickinson Immunocytometry Systems) and flow cytometry data
were processed with CellQuest computer software (Becton Dickinson Immuno-
cytometry Systems).
Intracellular staining for pSTAT5 or total STAT5 was done, after methanol
fixation, using an Alexa 647–conjugated monoclonal antibody specific for
pSTAT5 (Y694) (pSTAT5, clone 47, IgG1, BD) or rabbit anti-STAT5 (clone
C17 from Santa Cruz Biotechnology), respectively. For visualization of total
STAT5 staining, a rabbit-specific Alexa 488–conjugated goat secondary
antibody (Molecular Probes) was used. For the pSTAT5 detection, fresh
tonsils were sorted for CD19-phycoerythrin-indotricarbocyanine, CD38-
fluorescein isothiocyanate, CD25-phycoerythrin and CD19-phycoerythrin-
indotricarbocyanine, CD38–fluorescein isothiocyanate and CD25-
phycoerythrin. After the cell sort cells were fixed with 90% methanol, washed,
and stained with anti-pSTAT5 for flow cytometry. For the STAT5 detection, the
following tonsil B cell fractions, CD19+CD38� (non–germinal center B cells),
CD19+CD38+CD77+ (centroblasts), CD19+CD38+CD77lo (centrocytes),
CD19+CD38+CD25+ and CD19+CD38+CD25�, were sorted with the following
antibodies: CD19-phycoerythrin-indotricarbocyanine, CD38-allophycocyanin,
CD25-phycoerythrin, and CD77 followed by phycoerythrin–anti–rat IgM (BD).
RT-PCR. Total RNA was isolated from thawed pellets using the RNeasy mini kit
(Qiagen). RNA was reverse-transcribed in a volume of 20 ml containing
5� first-strand buffer, 500 mM dNTPs, 25 mg/l oligo(dT) and 200 U superscript
II RT (Life Technologies). A portion of the cDNA solution (1 ml) was amplified
by PCR in a 50-ml solution containing 20 mM Tris-HCl, 50 mM KCl, 1.5 mM
MgCl2, 5 mM dNTPs, 2.5 U Taq DNA polymerase (Life Technologies) and
30 pmol of each primer. PCR conditions were as follows: a 7-minute
denaturing step at 94 1C followed by 30 cycles of 30 s at 94 1C, 30 s at
62 1C (HPRT1), 52 1C (LMP-1), 58 1C (EBNA1/2) or 58 1C (Bcl-6), and 30 s at
72 1C, and a final 7-minute extension at 72 1C. The oligonucleotides used for
RT-PCR were as follows: HPRT1 forward (5¢-TATGGACAGGACTGAACGTC
TTGC-3¢) and HPRT1 reverse (5¢-GACACAAACATGATTCAAATCCCTGA-3¢);
LMP-1 forward: 5¢-GCGACTCTGCTGGAAATGAT-3¢) and LMP-1 reverse (5¢-GACATGGTAATGCCTAGAAG-3¢); EBNA1/2 forward (5¢-AGCAAGAAGAG
GAGGTGGTAAG-3¢) and EBNA1/2 reverse (5¢-GGCTCAAAGTGGTCTC
TAATGC-3¢); BCL6 forward (5¢-AAGGGTCTGGTTAGTCCACAG-3¢) and
BCL6 reverse (5¢-GGTCACACTTGTAGGGTTTGTC-3¢); PRDM1 forward (5¢-CAACAACTTTGGCCTCTTCC-3¢) and PRDM1 reverse (5¢-GCATTCATGTG
GCTTTTCTC-3¢); and BCL2L1 forward (5¢-AATGGCAACCCATCCTG
GCA-3¢) and BCL2L1 reverse (5¢-TTCTCCTGGATCCAAGGCTC-3¢). The
PCR conditions to detect VH genes were similar to those described50, including
a 7-minute denaturing step at 94 1C followed by 35 cycles of 1 min at 94 1C,
1 min at 60 1C and 1 min at 72 1C, and a final 10-minute extension at 72 1C. The
oligonucleotides used for RT-PCR were as follows: VH1-7 forward (5¢-TCTG
GGGCTGAGGTGAAGAA-3¢); VH2 forward (5¢-ACCTTGAAGGAGTCTGGT
CCT-3¢); VH3 forward (5¢-GGGGGTCCCTGAGACTCTC-3¢); VH4 forward (5¢-GCCCAGGACTGGTGAAGC-3¢); VH5 forward (5¢-CTGGTGCAGTCTGGAG
CAG-3¢); Cm reverse (5¢-GAGGATCCGGGTGCTGCTGATGTCAGA-3¢); and
Cg reverse (5¢-GGGTCTAGACAGGCAGCCCAGGGCCGCTGTGC-3¢). Quan-
titative real-time PCR analysis was done with a iCycler PCR (BioRad). The iQ
SYBR Green Supermix of BioRad was used for amplification. After incubation
at 95 1C for 6 min, 40 cycles of amplification were done. Each cycle consisted of
30 s at 95 1C, 30 s at 60 1C and 30 s at 72 1C. For each sample, serial dilutions
NATURE IMMUNOLOGY VOLUME 6 NUMBER 3 MARCH 2005 31 1
A R T I C L E S©
2005
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eim
mun
olog
y

were made to determine the PCR efficiency. The oligonucleotides used for real
time RT-PCR were as follows: BCL6 forward (5¢-CTGGCTTTTGTGACG
GAAAT-3¢) and BCL6 reverse (5¢-AACCTGAAAACCCACACTCG-3¢); PRDM1
forward (5¢-GTGTCAGAACGGGATGAACA-3¢) and PRDM1 reverse (5¢-GCTCGGTTGCTTTAGACTGC-3¢); and ACTB (b-actin) forward (5¢-GGATG
CAGAAGGAGATCACTG-3¢) and ACTB (b-actin) reverse (5¢-CGATCCA
CACGGAGTACTTG-3¢).
Immunoblotting. Cell extracts were prepared in radioimmunoprecipitation
assay lysis buffer (150 mM NaCl, 1% Nonidet-P40, 0.1% SDS, 0.5% deoxy-
cholic acid, 50 mM Tris-HCl pH 8.0) supplemented with a protease inhibitor
‘cocktail’ (Boehringer Ingelheim BV). Proteins were transferred to Protran
nitrocellulose transfer membranes (Schleicher and Schuell BioScience). Primary
antibodies, conjugated to horseradish persoxidase, used for immunoblotting
were to Bcl-6 (C-19), STAT5 (C-17), ER (MC-20) and actin (I-19), all from
Santa Cruz Biotechnology. For the determination of STAT5 expression, equal
amounts of protein were loaded using a Bradford test (BCA protein assay
reagent kit (Pierce Biotechnology). For the other immunoblots, blots actin used
as loading control. Detection of proteins was done with enhanced chemilumi-
nescence (Pierce).
Luciferase reporter transactivation assay. Cotransfection of the reporter
construct, the constitutively active Renilla reniformis luciferase–producing
vector prL-CMV (Promega) and the expression vector LZRS in 293T cells
was done with the Fugene transfection reagent in accordance with the
manufacturer’s instructions (Roche). Detection of the firefly (Photinus pyralis)
and R. reniformis luciferases was done with the Dual Luciferase assay kit
according to manufacturer’s instructions (Promega). The BCL6 promoter
region (nucleotides �657 to +471) in the pGL3 basic vector was provided by
S. Hirosawa (Tokyo Medical and Dental University, Tokyo, Japan). Mutations
of the STAT5 binding site in the BCL6 promoter fragments were generated by a
two-step PCR approach. The first-step PCR was done with the Bcl-6(F) or Bcl-
6(R) primer and the corresponding GLprimer2 and RVprimer4 (Promega):
Bcl-6(F), 5¢-GAACATGTCTCTAAAGTGCAGGA-3¢, and Bcl-6(R), 5¢-TCCTGCACTTTAGAGACATGTTC-3¢. The corresponding PCR products were
then purified, mixed and reamplified with the GLprimer2 and RVprimer4
primers. The amplified PCR fragments were cloned directly into the
pCR2�1TOPO vector (Invitrogen), according to the manufacturer’s instruc-
tions. Sequencing with primers M13(F) and M13(R) was done with an ABI
sequencer (Perkin Elmer) with the dye-terminator cycle-sequencing kit (Perkin
Elmer). The mutated BCL6 promoter region was subcloned in pGL3 via SacI
and BglII digestion.
Note: Supplementary information is available on the Nature Immunology website.
ACKNOWLEDGMENTSWe thank A. Bakker (Netherlands Cancer Institute, Amsterdam, Netherlands)for help in cloning; B. Hooibrink (Academic Medical Center, Amsterdam,Netherlands), A. Pfauth and F. van Diepen (Netherlands Cancer Institute,Amsterdam, Netherlands) for help with cell sorting; T. Dellemijn, L. Oomenand L. Brooks (Netherlands Cancer Institute, Amsterdam, Netherlands) for helpwith CLSM analyses of the tonsil sections and transduced B cell samples; N. vander Stoep (Leiden University Medical Center, Leiden, Netherlands) for help withthe luciferase assays; and the Department of Otolaryngology, Academic MedicalCenter, Amsterdam, Netherlands (W. Fokkens) for providing tonsil tissue.
COMPETING INTERESTS STATEMENTThe authors declare competing financial interests (see the Nature Immunologywebsite for details).
Received 5 November 2004; accepted 20 January 2005
Published online at http://www.nature.com/natureimmunology/
1. Klein, U. et al. Transcriptional analysis of the B cell germinal center reaction. Proc.Natl. Acad. Sci. USA 100, 2639–2644 (2003).
2. Calame, K.L., Lin, K.I. & Tunyaplin, C. Regulatory mechanisms that determine thedevelopment and function of plasma cells. Annu. Rev. Immunol. 21, 205–230 (2003).
3. Tarlinton, D.M. & Smith, K.G. Dissecting affinity maturation: a model explainingselection of antibody-forming cells and memory B cells in the germinal centre.Immunol. Today 21, 436–441 (2000).
4. Stuber, E. & Strober, W. The T cell-B cell interaction via OX40–OX40L is necessaryfor the T cell-dependent humoral immune response. J. Exp. Med. 183, 979–989(1996).
5. Turner, C.A., Jr., Mack, D.H. & Davis, M.M. Blimp-1, a novel zinc finger-containingprotein that can drive the maturation of B lymphocytes into immunoglobulin-secretingcells. Cell 77, 297–306 (1994).
6. Reljic, R., Wagner, S.D., Peakman, L.J. & Fearon, D.T. Suppression of signal transducerand activator of transcription 3- dependent B lymphocyte terminal differentiation byBCL-6. J. Exp. Med. 192, 1841–1848 (2000).
7. Shaffer, A.L. et al. BCL-6 represses genes that function in lymphocyte differentiation,inflammation, and cell cycle control. Immunity 13, 199–212 (2000).
8. Shaffer, A.L. et al. Blimp-1 orchestrates plasma cell differentiation by extinguishing themature B cell gene expression program. Immunity 17, 51–62 (2002).
9. Reimold, A.M. et al. Plasma cell differentiation requires the transcription factor XBP-1.Nature 412, 300–307 (2001).
10. Arpin, C. et al. Generation of memory B cells and plasma cells in vitro. Science 268,720–722 (1995).
11. Arpin, C., Banchereau, J. & Liu, Y.J. Memory B cells are biased towards terminaldifferentiation: a strategy that may prevent repertoire freezing. J. Exp. Med. 186,931–940 (1997).
12. Shvarts, A. et al. A senescence rescue screen identifies BCL6 as an inhibitor of anti-proliferative p19(ARF)-p53 signaling. Genes Dev. 16, 681–686 (2002).
13. Fearon, D.T., Manders, P. & Wagner, S.D. Arrested differentiation, the self-renewingmemory lymphocyte, and vaccination. Science 293, 248–250 (2001).
14. Lischke, A. et al. The interleukin-4 receptor activates STAT5 by a mechanism that reliesupon common gamma-chain. J. Biol. Chem. 273, 31222–31229 (1998).
15. Rolling, C., Treton, D., Pellegrini, S., Galanaud, P. & Richard, Y. IL4 and IL13 receptorsshare the g c chain and activate STAT6, STAT3 and STAT5 proteins in normal humanB cells. FEBS Lett. 393, 53–56 (1996).
16. Leonard, W.J. & O’Shea, J.J. Jaks and STATs: biological implications. Annu. Rev.Immunol. 16, 293–322 (1998).
17. Shimoda, K. et al. Lack of IL-4-induced TH2 response and IgE class switching in micewith disrupted Stat6 gene. Nature 380, 630–633 (1996).
18. Kaplan, M.H., Schindler, U., Smiley, S.T. & Grusby, M.J. STAT6 is required formediating responses to IL-4 and for development of TH2 cells. Immunity 4, 313–319 (1996).
19. Teglund, S. et al. STAT5a and STAT5b proteins have essential and nonessential, orredundant, roles in cytokine responses. Cell 93, 841–850 (1998).
20. Bunting, K.D. et al. Reduced lymphomyeloid repopulating activity from adult bonemarrow and fetal liver of mice lacking expression of STAT5. Blood 99, 479–487(2002).
21. Sexl, V. et al. STAT5a/b contribute to interleukin 7-induced B-cell precursor expansion,but abl- and bcr/abl-induced transformation are independent of stat5. Blood 96,2277–2283 (2000).
22. Burchill, M.A. et al. Distinct effects of STAT5 activation on CD4+ and CD8+ T cellhomeostasis: development of CD4+CD25+ regulatory T cells versus CD8+ memoryT cells. J. Immunol. 171, 5853–5864 (2003).
23. Lord, J.D., McIntosh, B.C., Greenberg, P.D. & Nelson, B.H. The IL-2 receptor promoteslymphocyte proliferation and induction of the c-myc, bcl-2, and bcl-x genes through thetrans-activation domain of STAT5. J. Immunol. 164, 2533–2541 (2000).
24. John, S., Robbins, C.M. & Leonard, W.J. An IL-2 response element in the human IL-2receptor alpha chain promoter is a composite element that binds STAT5, Elf-1, HMG-I(Y) and a GATA family protein. EMBO J. 15, 5627–5635 (1996).
25. Martinez-Valdez, H. et al. Human germinal center B cells express the apoptosis-inducing genes Fas, c-myc, P53, and Bax but not the survival gene bcl-2. J. Exp. Med.183, 971–977 (1996).
26. Angelin-Duclos, C., Cattoretti, G., Lin, K.I. & Calame, K. Commitment of B lympho-cytes to a plasma cell fate is associated with Blimp-1 expression in vivo. J. Immunol.165, 5462–5471 (2000).
27. Tunyaplin, C. et al. Direct repression of prdm1 by Bcl-6 inhibits plasmacytic differ-entiation. J. Immunol. 173, 1158–1165 (2004).
28. Banchereau, J., de Paoli, P., Valle, A., Garcia, E. & Rousset, F. Long-term human B celllines dependent on interleukin-4 and antibody to CD40. Science 251, 70–72 (1991).
29. Kurata, H., Lee, H.J., O’Garra, A. & Arai, N. Ectopic expression of activated Stat6induces the expression of TH2- specific cytokines and transcription factors in devel-oping TH1 cells. Immunity 11, 677–688 (1999).
30. Ohashi, K., Miki, T., Hirosawa, S. & Aoki, N. Characterization of the promoter region ofhuman BCL-6 gene. Biochem. Biophys. Res. Commun. 214, 461–467 (1995).
31. Wolf, J. et al. Peripheral blood mononuclear cells of a patient with advanced Hodgkin’slymphoma give rise to permanently growing Hodgkin-Reed Sternberg cells. Blood 87,3418–3428 (1996).
32. Hinz, M. et al. Nuclear factor kappaB-dependent gene expression profiling of Hodgkin’sdisease tumor cells, pathogenetic significance, and link to constitutive signal transdu-cer and activator of transcription 5a activity. J. Exp. Med. 196, 605–617 (2002).
33. Raman, V.S., Akondy, R.S., Rath, S., Bal, V. & George, A. Ligation of CD27 on B cellsin vivo during primary immunization enhances commitment to memory B cellresponses. J. Immunol. 171, 5876–5881 (2003).
34. Raman, V.S., Bal, V., Rath, S. & George, A. Ligation of CD27 on murine B cellsresponding to T-dependent and T-independent stimuli inhibits the generation of plasmacells. J. Immunol. 165, 6809–6815 (2000).
35. Andjelic, S. et al. Phosphatidylinositol 3-kinase and NF-kB/Rel are at the divergenceof CD40-mediated proliferation and survival pathways. J. Immunol. 165, 3860–3867(2000).
3 12 VOLUME 6 NUMBER 3 MARCH 2005 NATURE IMMUNOLOGY
A R T I C L E S©
2005
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eim
mun
olog
y

36. Dadgostar, H. et al. Cooperation of multiple signaling pathways in CD40-regulatedgene expression in B lymphocytes. Proc. Natl. Acad. Sci. USA 99, 1497–1502(2002).
37. Ehret, G.B. et al. DNA binding specificity of different STAT proteins. Comparison ofin vitro specificity with natural target sites. J. Biol. Chem. 276, 6675–6688 (2001).
38. Lin, Y., Wong, K. & Calame, K. Repression of c-myc transcription by Blimp-1, aninducer of terminal B cell differentiation. Science 276, 596–599 (1997).
39. Usui, T. et al. Overexpression of B cell-specific activator protein (BSAP/Pax-5) in a lateB cell is sufficient to suppress differentiation to an Ig high producer cell with plasmacell phenotype. J. Immunol. 158, 3197–3204 (1997).
40. Reimold, A.M. et al. Plasma cell differentiation requires the transcription factorXBP-1. Nature 412, 300–307 (2001).
41. Karras, J.G., Wang, Z., Coniglio, S.J., Frank, D.A. & Rothstein, T.L. Antigen-receptorengagement in B cells induces nuclear expression of STAT5 and STAT6 proteins thatbind and transactivate an IFN-g activation site. J. Immunol. 157, 39–47 (1996).
42. Dent, A.L., Shaffer, A.L., Yu, X., Allman, D. & Staudt, L.M. Control of inflammation,cytokine expression, and germinal center formation by BCL-6. Science 276, 589–592(1997).
43. Ye, B.H. et al. The bcl-6 proto-oncogene controls germinal-centre formation andTH2- type inflammation. Nat. Genet. 16, 161–170 (1997).
44. Fukuda, T. et al. Disruption of the bcl-6 gene results in an impaired germinal centerformation. J. Exp. Med. 186, 439–448 (1997).
45. Toyama, H. et al. Memory B cells without somatic hypermutation are generated frombcl-6-deficient B cells. Immunity 17, 329–339 (2002).
46. Ariyoshi, K. et al. Constitutive activation of STAT5 by a point mutation in the SH2domain. J. Biol. Chem. 275, 24407–24413 (2000).
47. Heemskerk, M.H. et al. Inhibition of T cell and promotion of natural killer celldevelopment by the dominant negative helix loop helix factor Id3. J. Exp. Med. 186,1597–1602 (1997).
48. Brummelkamp, T.R., Bernards, R. & Agami, R. A system for stable expression of shortinterfering RNAs in mammalian cells. Science 296, 550–553 (2002).
49. Vyth-Dreese, F.A., Dellemijn, T.A., Majoor, D. & de Jong, D. Localization in situ of theco-stimulatory molecules B7.1, B7.2, CD40 and their ligands in normal humanlymphoid tissue. Eur. J. Immunol. 25, 3023–3029 (1995).
50. Guikema, J.E. et al. Multiple myeloma related cells in patients undergoing autologousperipheral blood stem cell transplantation. Br. J. Haematol. 104, 748–754 (1999).
NATURE IMMUNOLOGY VOLUME 6 NUMBER 3 MARCH 2005 31 3
A R T I C L E S©
2005
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eim
mun
olog
y




















