
Stable Activation of Phosphatidylinositol 3-Kinase in the T Cell Immunological Synapse Stimulates...
12
of June 13, 2013. This information is current as Nuclear Exclusion and Cell Growth Control Synapse Stimulates Akt Signaling to FoxO1 3-Kinase in the T Cell Immunological Stable Activation of Phosphatidylinositol Terry G. Unterman, Alain Trautmann and Georges Bismuth Stéphanie Fabre, Valérie Lang, Julie Harriague, Aude Jobart, http://www.jimmunol.org/content/174/7/4161 2005; 174:4161-4171; ; J Immunol Material Supplementary ml http://www.jimmunol.org/content/suppl/2005/03/16/174.7.4161.DC1.ht References http://www.jimmunol.org/content/174/7/4161.full#ref-list-1 , 21 of which you can access for free at: cites 47 articles This article Subscriptions http://jimmunol.org/subscriptions is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/ji/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/cgi/alerts/etoc Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 2005 by The American Association of 9650 Rockville Pike, Bethesda, MD 20814-3994. The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on June 13, 2013 http://www.jimmunol.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Stable Activation of Phosphatidylinositol 3-Kinase in the T Cell Immunological Synapse Stimulates...

of June 13, 2013.This information is current as
Nuclear Exclusion and Cell Growth ControlSynapse Stimulates Akt Signaling to FoxO13-Kinase in the T Cell Immunological Stable Activation of Phosphatidylinositol
Terry G. Unterman, Alain Trautmann and Georges BismuthStéphanie Fabre, Valérie Lang, Julie Harriague, Aude Jobart,
http://www.jimmunol.org/content/174/7/41612005; 174:4161-4171; ;J Immunol
MaterialSupplementary
mlhttp://www.jimmunol.org/content/suppl/2005/03/16/174.7.4161.DC1.ht
Referenceshttp://www.jimmunol.org/content/174/7/4161.full#ref-list-1
, 21 of which you can access for free at: cites 47 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2005 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

Stable Activation of Phosphatidylinositol 3-Kinase in the T CellImmunological Synapse Stimulates Akt Signaling to FoxO1Nuclear Exclusion and Cell Growth Control1
Stephanie Fabre,* Valerie Lang,* Julie Harriague,† Aude Jobart,* Terry G. Unterman,‡
Alain Trautmann,* and Georges Bismuth2*
We have previously reported at the single cell level that PI3K is activated after conjugate formation between T lymphocytes andAPCs. However, in contrast to cells exposed to an asymmetrical signal that usually increase 3�-phosphoinositides (3�-PI) transientlyin the region of the activated receptors, T cells contacting APC accumulate 3�-PI across their whole plasma membrane far beyondthe region of the immunological synapse (IS). Importantly, this effect is maintained over time, for hours, and although PI3K-dependent pathways translate in various cell types extracellular stimuli into a wide range of biological events, in primary T cellsthis stability is mostly required for cell division induced by Ag. Using imaging methodologies, the present article elucidates themolecular mechanisms responsible for this particular functioning of the PI3K pathway in primary human T lymphocytes inter-acting with APCs, especially with dendritic cells. The results reveal that the IS unremittingly recruits PI3K to maintain high 3�-PIlevels in T cells through phosphotyrosine-dependent mechanisms, suggesting a major participation of class Ia PI3K. This persistentactivation of PI3K results in the Akt-dependent sequestration of the FoxO transcription factor, FoxO1, outside the nucleus of Tcells interacting with APCs. Using an active form of FoxO1, we demonstrate that this compartmentalization process can affect Tcell growth after Ag recognition. We conclude that the need for sustained PI3K signaling within the consolidated IS is probablyan undemanding tactic used by primary T cells critical for initiating cell cycle progression through the prolonged inactivation ofFoxO1, one important factor that can control cell quiescence. The Journal of Immunology, 2005, 174: 4161–4171.
A ntigen-specific activation of T lymphocytes by APC isassociated with T cell polarization toward the APC andthe formation of a highly specialized junction, the im-
munological synapse (IS),3 characterized by a distinct segregationof Ag receptors and adhesion and signaling molecules (1–4).These phenomena require extensive remodeling of the T cell cy-toskeleton and are associated with a tyrosine phosphorylation cas-cade that can activate numerous cellular effectors. Among these,PI3Ks are key enzymes, the function of which in the immunesystem has been highlighted over the past few years, primarily in
studies using a genetic approach (5–9). However, direct demon-stration that PI3K is activated in resting T cells during IS forma-tion has only been recently reported (10, 11).
The PI3K family consists of three main classes of enzymes, I, II,and III, that phosphorylate membrane phosphatidylinositol (PtdIns),PtdIns 4-phosphate, or PtdIns 4,5-bisphosphate (PtdIns(4,5)P2) on theD3 position of the inositol ring. Class I PI3Ks, the best studied, areactivated by extracellular stimuli, and class IA PI3Ks are recruiteddownstream of receptors associated with tyrosine kinases such asthe BCR and the TCR for Ag, cytokine receptors, or costimulatorymolecules, whereas class IB PI3Ks act downstream of G protein-coupled receptors such as chemokine receptors (7, 9, 12). Class IAPI3Ks are heterodimeric enzymes constituted by a regulatory sub-unit, generally p85�, and a p110 catalytic subunit in tight interac-tion. The Src homology 2 domains of the regulatory subunit arerecruited on the consensus motif YxxM phosphorylated upon ty-rosine kinase-coupled receptor engagement (13), thereby convey-ing the p110 subunit close to the plasma membrane to catalyze theformation of PtdIns(3,4)P2 and PtdIns 3,4,5-triphosphate(PtdIns(3,4,5)P3), two phosphoinositides that are almost undetect-able in resting cells. This process allows the recruitment and ac-tivation of effector proteins containing a pleckstrin homology (PH)domain specific for these 3�-phosphoinositides (3�-PI) (14), includ-ing the serine-threonine kinase Akt, which can operate fused withthe GFP as a probe to dynamically follow 3�-PI in living cells.
This strategy was successfully used to reveal the existence of anintense and sustained production of 3�-PI after conjugate formationbetween a T cell and an APC in both human and murine primaryT cell systems (10, 11). In the presence of Ag, GFP-Akt-PH ac-cumulated to the T cell plasma membrane rapidly upon contactinitiation. Moreover, 3�-PI was increased for hours (11), a resultconfirmed in a more recent study (15), showing that its activation
*Departement de Biologie Cellulaire, Institut Cochin, Institut National de la Sante etde la Recherche Medicale, Unite 567, Centre National de la Recherche Scientifique,Unite Mixte de Recherche 8104, Universite Rene Descartes, and Laboratoire Label-lise par la Ligue Nationale contre le Cancer, Paris, France; †Unite d’ImmunologieVirale, Institut Pasteur, Paris, France; and ‡Departments of Medicine and Physiologyand Biophysics, University of Illinois College of Medicine and Veterans Affairs Chi-cago Health Care System, Chicago, IL 60612
Received for publication July 30, 2004. Accepted for publication January 24, 2005.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by the Ministere de l’Education Nationale et de La Re-cherche (to S.F.), the Association pour la Recherche sur le Cancer (to V.L.), Sidaction(to J.H.), and grants from the Ligue Nationale contre le Cancer, Institut National dela Sante et de la Recherche Medicale, Centre National de la Recherche Scientifique,the National Institutes of Health (DK41430; to T.G.U.), and the Department of Vet-erans Affairs Merit Review Program (to T.G.U.).2 Address correspondence and reprint requests to Dr. Georges Bismuth, Departementde Biologie Cellulaire, Institut Cochin, 22 rue Mechain, 75014, Paris, France. E-mailaddress: [email protected] Abbreviations used in this paper: IS, immunological synapse; CFP, cyan fluorescentprotein; DC, dendritic cell; FLIP, fluorescence loss in photobleaching; FRAP, fluo-rescence recovery after photobleaching; PH, pleckstrin homology; 3�-PI, 3�-phospho-inositide; PtdIns, phosphatidylinositol; PtdIns(4,5)P2, PtdIns 4,5-bisphosphate;PtdIns(3,4,5)P3, PtdIns 3,4,5-triphosphate; pTyr, phosphotyrosine; SAg, superanti-gen; YFP, yellow fluorescent protein.
The Journal of Immunology
Copyright © 2005 by The American Association of Immunologists, Inc. 0022-1767/05/$02.00
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

by Ag was a long-lasting process representing a striking feature ofPI3K metabolism in T cells. It was also found that PtdIns(3,4,5)P3
accumulated not only at the contact zone, but also in the plasmamembrane outside the IS when T cell-APC contact continued, sug-gesting that PI3K activation may be not restricted to the IS. Thisis in contrast with the current view of 3�-PI accumulation estab-lished in different cellular models, such as fibroblasts or neutro-phils, where the regulation of PI3K has been extensively studied,and this process is thought to be limited to the activated pole of thecell (16–19). Interestingly, it also appeared that this prolongedPI3K activation was essential for T cell proliferation, but not for ISformation and stability or for early signals triggered by the T cell/APC contact, including increased phosphotyrosine (pTyr) accumu-lation and calcium responses (10, 11).
Based on these observations, we determined the mechanismsunderlying the persistent accumulation of PtdIns(3,4,5)P3 ob-served in T cells contacting APCs, and how the asymmetrical ac-tivation of the T cell through the IS may account for the increasein PtdIns(3,4,5)P3 in the whole plasma membrane. We also askedwhether any downstream signaling pathway of critical importancefor cell division is activated by Ag in T cells that requires thissustained PI3K metabolism. The results of this study reveal thatthe IS unremittingly recruits PI3K to maintain high PtdIns(3,4,5)P3
levels in T cells, and that this sustained activation of PI3K metab-olism results in the sequestration of FoxO transcription factorsoutside the nucleus, a compartmentalization process important forT cells to undergo cell growth after Ag recognition.
Materials and MethodsCells
Human T lymphocytes were isolated from blood donors by Ficoll densitygradient centrifugation, followed by negative depletion on magnetic beads(T cell negative isolation kit; Dynal Biotech). Peripheral blood monocytes,purified with the negative monocyte isolation kit (Dynal Biotech), werederived into dendritic cells (DCs) in RPMI 1640 (Biochrom), 10% FCS inthe presence of 20 ng/ml IL-4 (PeproTech), and 800 U/ml GM-CSF (Leu-comax; Novartis/Shering-Plough) during 6 days before maturation with 5�g/ml LPS (Sigma-Aldrich). The Raji B cell line was cultured in RPMI1640, 10% FCS, 50 U/ml penicillin, and 50 �g/ml streptomycin. T8.1 is amurine T cell hybridoma expressing a human-mouse chimeric TCR spe-cific for the tetanus toxin peptide tt830–843 restricted to HLA-DRB1*1102.They were cultured in DMEM (Invitrogen Life Technologies), 10% FCS,50 U/ml penicillin, 50 �g/ml streptomycin, 50 �M 2-ME (complete me-dium) supplemented with 400 nM methotrexate, and 1 mg/ml G-418. AT8.1 clone stably expressing the fusion protein GFP-Akt-PH was culturedin complete medium supplemented with 1 �g/ml puromycin. Murine fi-broblasts L625.7 expressing human MHC class II molecules were used asAPC and maintained in complete medium containing 250 �g/ml G-418.
Plasmid constructs
GFP-Akt-PH was a gift from Dr. T. Meyer (Stanford University School ofMedicine, Stanford, CA). Cyan fluorescent protein (CFP)-Akt-PH was ob-tained by subcloning Akt-PH into the pECFP-C1 fusion vector (BD Clon-tech) as an EcoRI restriction enzyme fragment. p85� DNA was obtainedby PCR from a construct provided by Dr. E. Howard-Cofield (Universityof Kent, Canterbury, U.K.) and cloned into pCR2.1-TOPO (Invitrogen LifeTechnologies). p85� was then subcloned into the pEGFP-C1 and thepYFP-C1 fusion vectors (BD Clontech), as an EcoRI restriction enzyme frag-ment, resulting in expression of fusion proteins with GFP or yellow fluores-cence protein (YFP) being N-terminal. FoxO1-GFP and the triple mutantT24A/S256A/S319A FoxO1-GFP constructs were previously described (20).
Cell transfection and flow cytometric analysis
Freshly prepared human primary T cells were transfected with Nucleofec-tor technology (Amaxa) according to the manufacturer’s instructions with5 �g of the indicated construct. Expression levels were controlled by FACSanalysis (FACScan cytometer; BD Biosciences). For measurements of cellapoptosis, transfected T cells were grown in complete medium in 96-well,round-bottom plates (2 � 105 cells/well) and analyzed after 12 or 36 h ofculture at 37°C. Cells were washed once in PBS-2% FCS, then in PBS
alone and resuspended in 150 �l of annexin V binding buffer (10 mMHEPES (pH 7.4), 140 mM NaCl, and 2.5 mM CaCl2). Cells were labeledfor 15 min at room temperature with 5 �l of annexin V-PE (BD Pharm-ingen) and subjected to FACS analysis.
For activation assays, 24 h after transfection with the indicated con-structs, T cells were cultured as described above in the presence of 0.5 �105 irradiated Raji B cells (10,000 rad) with or without superantigen (SAg)at 20 ng/ml. After 6 or 24 h of culture, CD69 expression was measured byFACS with a PE-conjugated CD69 Ab (BD Pharmingen). Cell growth andnucleic acid contents were measured after 48 or 72 h of activation under theculture conditions described above by FACS analysis of unlabeled cellsuspensions or after overnight incorporation of propidium iodide (10 �g/ml) in cells fixed with PBS-1% paraformaldehyde (Sigma-Aldrich),respectively.
Fluorescence imaging
DCs or Raji B cells were plated on polylysine-coated glass coverslipsmounted on 30-mm petri dishes and incubated for 30 min at 37°C in RPMI1640 in the presence of recombinant staphylococcal enterotoxin E, staph-ylococcal enterotoxin A, staphylococcal enterotoxin B, and staphylococcalenterotoxin C3 (Toxin Technology) SAg at 200 ng/ml each. Transfected Tcells were then added to APCs and incubated at 37°C in RPMI 1640–0.5%FCS for various times. Conjugates were fixed for 10 min at room temper-ature in PBS-4% paraformaldehyde, washed twice in PBS, and stored at4°C in PBS. Studies with conjugates were performed on an EclipseTE3200 inverted microscope (Nikon) with a �60 oil objective. Imageswere collected with a cooled CCD camera (CoolSNAPfx; Roper Scientific)and the Metavue Imaging system (Universal Imaging), then analyzed withMetaMorph software (Universal Imaging).
For immunofluorescent staining, paraformaldehyde-fixed cells were in-cubated for 20 min in PBS-0.1 M glycine and permeabilized in PBS-0.1%Triton X-100 for 10 min at room temperature. After three washes in PBS,they were incubated in PBS containing 0.2% BSA (Sigma-Aldrich) for 45min. Cells were then stained with anti-phosphotyrosine 4G10 Ab (UpstateBiotechnology), followed by incubation with Rhodamine Red-X-conju-gated secondary Ab (Jackson ImmunoResearch Laboratories) at 4°C for 30min. After three washes in PBS-0.2% BSA and one in PBS, coverslipswere finally mounted in Mowiol (Sigma-Aldrich). Quantitative analysis offluorescence intensity was performed on 12-bit images after backgroundsubtraction using the MetaMorph software.
Video imaging
All time-lapse experiments were conducted using Raji B cells as surrogateAPCs because of the higher frequency of contacts they formed with restinghuman T cells compared with DCs in live video-recording experiments.Thirty minutes before the experiment, Raji B cells were plated on glasscoverslips mounted on 30-mm petri dishes and incubated at 37°C in pres-ence of a mixture of SAg (recombinant staphylococcal enterotoxin E,staphylococcal enterotoxin A, staphylococcal enterotoxin B, and staphy-lococcal enterotoxin C3; 200 ng/ml each). Transfected T cells were thenadded to the APCs in a final volume of 200 �l. Transmitted light and GFPfluorescence images were acquired sequentially every 10 s on an invertedEclipse TE300 microscope (Nikon) equipped with a �20 or a �40 oilobjective and the Metafluor imaging system (Universal Imaging). Videoswere made with Premiere software (Adobe Systems).
Photobleaching measurements
T8.1 GFP-Akt-PH/L625.7 conjugates formed in presence of Ag (1 �g/mlpeptide tt830–843) were analyzed after 1 h of interaction. Fluorescence re-covery after photobleaching (FRAP) analysis was conducted on a LeicaTCS SP2 AOBS confocal laser-scanning microscope equipped with a ther-mostated chamber at 37°C, using the 488 nm line of a 20-mW argon laserand a �63 oil objective. Prebleaching and recovery images were collectedat 208-ms time intervals scanning a single Z-section of the whole cell at10% laser transmission. Photobleaching was conducted at 100% lasertransmission for three iterations on a small membrane region at the rear ofthe cell. Quantitative analysis was performed on the resulting 12-bit imagesusing Metamorph Imaging software. The average fluorescence intensity inthe bleached region was measured, normalized as a percentage of the initialfluorescence, and plotted against time using Kaleida Graph software (Syn-ergy Software). The diffusion coefficient at the membrane was approxi-mated using the equation D � �2�/4�D, where � is the radius of thebleached area, �D is the time constant of diffusion, and � is a correctionfactor for the amount of bleaching (estimated at 1.4 in the case of a 70%bleach).
For fluorescence loss in photobleaching (FLIP) experiments, T8.1 GFP-Akt-PH cells contacting L625.7 APCs were repeatedly photobleached
4162 PI3K ACTIVATION AND FoxO1 NUCLEAR EXCLUSION IN T CELLS
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

within a small membrane region outside the IS, at the rear of the cell, orwithin the IS. Between the bleaching periods, the cell was imaged with lowintensity light. Practically, we recorded one 8-bit image every 2.012 s at7% laser transmission, in turn with 1.006-s photobleaching pulses at 100%laser transmission. The Metamorph software was then used to measure theloss in fluorescence intensity as a function of time in different membraneregions of the T cell adjacent to the bleached one or in the whole T cell.
ResultsPI3K recruitment to the IS induced by Ag recognition
The spatio-temporal localization of PI3K during T cell-Ag recog-nition and its relationship to the distribution of PtdIns(3,4,5)P3
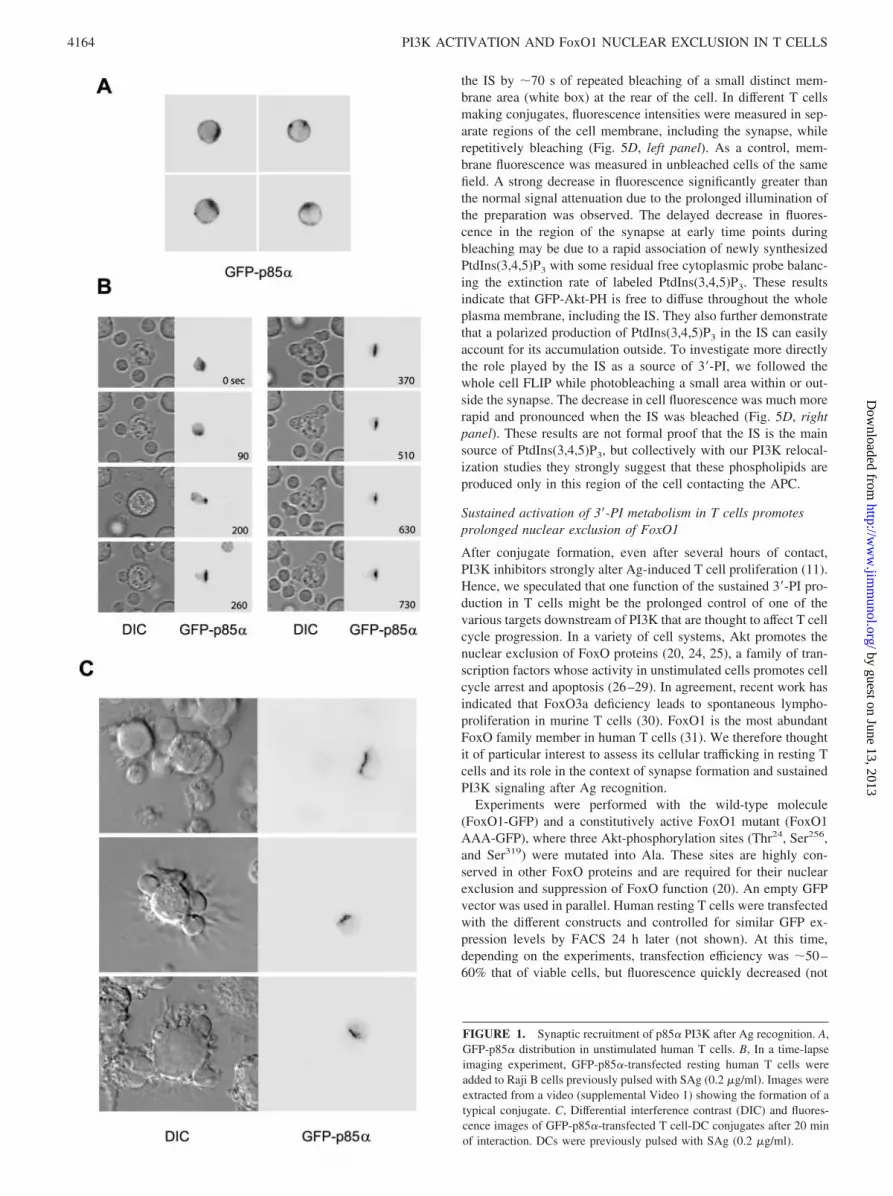
outside the IS observed in living T cells interacting with APCshave not been previously examined. To assess PI3K localizationduring this process, we performed dynamic analyses using a GFPfusion protein containing p85�, the main regulatory subunit of theclass IA PI3K expressed in T cells. In unstimulated human periph-eral blood T cells, this probe has a diffuse cytoplasmic distribution,with some cells showing additionally a few hot spots of fluores-cence at the plasma membrane (Fig. 1A). Major changes occurredin T cells contacting Ag-pulsed APCs. Fig. 1B shows a series ofimages extracted from a video (supplemental Video 1)4 in whichan IS is formed between a T cell and an APC (in this case a RajiB cell). Rapidly, within a few seconds after the initial contact, theprobe began to accumulate at the IS. In �1 min, GFP-p85� waslargely removed from the cytoplasm. Identical results were ob-served with Ag-pulsed DCs as APCs. Fig. 1C shows several fixedT-DC conjugates analyzed after 20 min of interaction, illustratingthe marked and exclusive redistribution of GFP-p85� to the IS, asituation found with almost all conjugates.
Sustained 3�-PI metabolism in T cells contacting DCs correlateswith prolonged PI3K recruitment and phosphotyrosineaccumulation at the IS
PI3K metabolism is activated for hours in murine T cells contact-ing APCs, as measured by GFP-Akt-PH accumulation at theplasma membrane (10, 11). Whether this biological phenomenonis also true for human T cells has not been examined. Mainly, itsrelationship with a prolonged PI3K recruitment to the IS is un-known. To explore this, resting human T cells were transfectedwith GFP-p85� and incubated several hours with Ag-pulsed DCs.The cellular localization of the probe was then examined on fixedconjugates. The same cells were analyzed for their pTyr contents.As shown in Fig. 2A, GFP-p85� was persistently recruited to theIS, and the probe never accumulated outside the contact zone. Tcells with GFP-p85� relocalized to the IS were also strongly la-beled by the pTyr Ab at the different time points studied. Thispattern of costaining was observed in �100% of cells showing asignificant GFP-p85� relocalization at the synaptic region. More-over, a high correlation between GFP-p85� and pTyr fluorescencelevels was found (Fig. 2B), making clear the relationship betweenthe two processes. This superimposed and restricted labeling at thesynapse highly contrasts with the very homogeneous distributionof GFP-Akt-PH at the T cell plasma membrane within and outsidethe contact zone in these long-lasting conjugates (Fig. 2C).
Quantitative analyses also were conducted to follow the evolu-tion of pTyr in T-DC synapses after various periods of incubation.The number of conjugates with increased pTyr was found to berather stable (Fig. 3A). In these conjugates, pTyr decreasedthroughout the experiment, but never vanished (Fig. 3B). Similarresults were obtained with the T8.1 hybridoma T cell interactingwith MHC class II-expressing fibroblast APCs presenting a tetanus
toxin peptide (not shown) and previously used to demonstrate sus-tained 3�-PI metabolism upon Ag recognition in T cells (10).
IS is the major site of 3�-PI production in T cells
As stated above (see introduction), T cells contain various PI3Ksthat can be activated by phosphotyrosine-dependent and -indepen-dent mechanisms, and a deficiency in p110�, the catalytic subunitof class IB PI3K activated by G protein-coupled receptors, hasbeen shown to affect the proliferative response of murine T cellsinduced by anti-CD3 Ab (21). Thus, showing a parallel accumu-lation of pTyr-dependent class IA PI3Ks members such as p85�and pTyr within the IS does not exclude the recruitment of PI3Ksfrom other classes outside this particular location to activate 3�-PIsynthesis. It has been reported that when overexpressed in cells,p85� inhibits pTyr-associated PI3K activity (22). One proposedmechanism is that the occupation of phosphorylation sites by themonomeric p85� form competitively limits interaction with theactive heterodimer. We thus reasoned that if 3�-PI synthesis in-duced by Ag recognition originated from the synapse only, espe-cially through the pTyr accumulated within this site, overexpres-sion of p85� should inhibit the increase in 3�-PI in the whole cell.To evaluate this, we expressed a YFP-p85� molecule in resting Tcells and analyzed the plasma membrane accumulation of a co-transfected CFP-Akt-PH probe after Ag recognition. Fig. 4 showsvarious 2-h conjugates from the same preparation with T cellsexpressing significant amounts of CFP-Akt-PH, but different levelsof YFP-p85�. The results showed that translocation of Akt-PH tothe plasma membrane was strongly impaired in the presence of a highlevel of YFP-p85�. This clearly suggests that the IS is the main, if notthe exclusive, source of 3�-PI continuously synthesized through apTyr-dependent process in T cells contacting APCs.
To support this finding, a different strategy was used based on pho-tobleaching methodologies. We assumed that if PtdIns(3,4,5)P3 wasprimarily produced at the IS, its rapid diffusion from this place tothe rest of the plasma membrane should be detectable. To explorethis possibility, we first examined fluorescence recovery of mem-brane GFP-Akt-PH after photobleaching. Experiments were per-formed with T8.1 hybridoma T cells stably expressing GFP-Akt-PH (10), whose size makes laser beam focusing easier toperform than in resting T cells and also exhibits a sustained accu-mulation of GFP-Akt-PH in the entire plasma membrane (see Fig.5A). After 1 h of interaction, several T8.1-APC conjugates wereselected on which a small membrane region located at the rear ofthe T cell (outlined box in Fig. 5A) was photobleached for 624 ms,resulting in 70% probe fluorescence extinction. The average fluo-rescence intensity in the bleached area was then quantified onwhole T cell images collected at different time intervals at lowlaser power. We observed that membrane fluorescence of GFP-Akt-PH was very rapidly restored after photobleaching with a timeconstant (�D) of 3.1 � 0.80 s (n � 11; Fig. 5B). These data allowus to estimate a value for the PtdIns(3,4,5)P3 diffusion coefficientof �0.92 � 0.37 �m2/s, which is of the same order as the valuerecently determined using a different strategy in a fibroblast T cellmodel (0.37 � 0.25 �m2/s) (23).
FLIP is the decrease/disappearance of fluorescence in a definedregion adjacent to a repetitively bleached region in living cells; asa result, this technique can indicate whether a molecule is rapidlyexchanged between separate cell compartments or various regionsof a given compartment, like the plasma membrane. We thereforeused this method to test whether labeled phospholipids diffusingthrough the bleached area in the FRAP experiments come from thepool accumulated at the IS. A typical experiment is shown in Fig.5C, in which is shown a rapid darkening of the cell in and outside4 The online version of this article contains supplemental material.
4163The Journal of Immunology
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from
the IS by �70 s of repeated bleaching of a small distinct mem-brane area (white box) at the rear of the cell. In different T cellsmaking conjugates, fluorescence intensities were measured in sep-arate regions of the cell membrane, including the synapse, whilerepetitively bleaching (Fig. 5D, left panel). As a control, mem-brane fluorescence was measured in unbleached cells of the samefield. A strong decrease in fluorescence significantly greater thanthe normal signal attenuation due to the prolonged illumination ofthe preparation was observed. The delayed decrease in fluores-cence in the region of the synapse at early time points duringbleaching may be due to a rapid association of newly synthesizedPtdIns(3,4,5)P3 with some residual free cytoplasmic probe balanc-ing the extinction rate of labeled PtdIns(3,4,5)P3. These resultsindicate that GFP-Akt-PH is free to diffuse throughout the wholeplasma membrane, including the IS. They also further demonstratethat a polarized production of PtdIns(3,4,5)P3 in the IS can easilyaccount for its accumulation outside. To investigate more directlythe role played by the IS as a source of 3�-PI, we followed thewhole cell FLIP while photobleaching a small area within or out-side the synapse. The decrease in cell fluorescence was much morerapid and pronounced when the IS was bleached (Fig. 5D, rightpanel). These results are not formal proof that the IS is the mainsource of PtdIns(3,4,5)P3, but collectively with our PI3K relocal-ization studies they strongly suggest that these phospholipids areproduced only in this region of the cell contacting the APC.
Sustained activation of 3�-PI metabolism in T cells promotesprolonged nuclear exclusion of FoxO1
After conjugate formation, even after several hours of contact,PI3K inhibitors strongly alter Ag-induced T cell proliferation (11).Hence, we speculated that one function of the sustained 3�-PI pro-duction in T cells might be the prolonged control of one of thevarious targets downstream of PI3K that are thought to affect T cellcycle progression. In a variety of cell systems, Akt promotes thenuclear exclusion of FoxO proteins (20, 24, 25), a family of tran-scription factors whose activity in unstimulated cells promotes cellcycle arrest and apoptosis (26–29). In agreement, recent work hasindicated that FoxO3a deficiency leads to spontaneous lympho-proliferation in murine T cells (30). FoxO1 is the most abundantFoxO family member in human T cells (31). We therefore thoughtit of particular interest to assess its cellular trafficking in resting Tcells and its role in the context of synapse formation and sustainedPI3K signaling after Ag recognition.
Experiments were performed with the wild-type molecule(FoxO1-GFP) and a constitutively active FoxO1 mutant (FoxO1AAA-GFP), where three Akt-phosphorylation sites (Thr24, Ser256,and Ser319) were mutated into Ala. These sites are highly con-served in other FoxO proteins and are required for their nuclearexclusion and suppression of FoxO function (20). An empty GFPvector was used in parallel. Human resting T cells were transfectedwith the different constructs and controlled for similar GFP ex-pression levels by FACS 24 h later (not shown). At this time,depending on the experiments, transfection efficiency was �50–60% that of viable cells, but fluorescence quickly decreased (not
FIGURE 1. Synaptic recruitment of p85� PI3K after Ag recognition. A,GFP-p85� distribution in unstimulated human T cells. B, In a time-lapseimaging experiment, GFP-p85�-transfected resting human T cells wereadded to Raji B cells previously pulsed with SAg (0.2 �g/ml). Images wereextracted from a video (supplemental Video 1) showing the formation of atypical conjugate. C, Differential interference contrast (DIC) and fluores-cence images of GFP-p85�-transfected T cell-DC conjugates after 20 minof interaction. DCs were previously pulsed with SAg (0.2 �g/ml).
4164 PI3K ACTIVATION AND FoxO1 NUCLEAR EXCLUSION IN T CELLS
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

shown). We first controlled, with annexin V, T cell apoptosis in-duced by FoxO1 overexpression. As shown in Fig. 6A, a small butsignificantly increased staining compared with the GFP empty vec-tor was observed with the mutant and to a lesser extent with thewild-type FoxO1 molecule 36 h posttransfection. We next fol-lowed the impact of FoxO1 overexpression on CD69 expression, ausual parameter for monitoring activation of primary T cells earlyafter Ag recognition (Fig. 6B). Only the mutant slightly impairedCD69 expression after 6 and 24 h of coculture with APCs, but notenough to suggest that the level of FoxO1 activity is critical in theinitial steps of T cell activation. We then assessed the impact ofFoxO1 overexpression on T cell growth induced by Ag recogni-tion. The increase in T cell size was measured on GFP-positive and-negative viable T cells within the forward and side scatter lym-phocyte gate delineated on the whole cell population after a 2-dayculture with APCs (Fig. 6C). The results showed that cells trans-fected with FoxO1 AAA-GFP were much smaller than FoxO1-
GFP- or GFP-expressing cells. No such difference was seen withinthe GFP-negative cell populations. The nucleic acid content mea-sured with an intercalating dye was also strongly reduced in cellsexpressing the constitutively active mutant (Fig. 6D), suggestingthat the decrease in cell size is because FoxO1 AAA-GFP-express-ing cells are in a different T cell cycle phase than those expressingempty GFP or FoxO1-GFP. Intriguingly, we repeatedly found apartial, but significant, alteration of these biological parameters inthe fluorescent-negative cell population gated from FoxO1 AAA-GFP-transfected samples. This might be indicative of an indirectsuppressive effect of FoxO1 AAA-expressing cells on the whole Tcell population through an as yet unknown mechanism. However,we could not exclude that some cells initially transfected no longerexpressed the transgene at this stage of the assay.
We next monitored the localization of FoxO1 in unstimulated Tcells and in T cells contacting Ag-pulsed DCs. The predominantnuclear localization of FoxO1-GFP was obvious in unstimulated T
FIGURE 2. Persistent recruitment of p85� PI3-K to the IS correlates with pTyr accumulation. A, GFP-p85�-transfected T cells were fixed after 30 min,2 h, or 4 h of interaction with SAg (0.2 �g/ml)-pulsed DCs and probed with anti-pTyr Ab. Differential interference contrast (DIC) and fluorescence imagesof the same conjugates are shown. B, Conjugates obtained as described in A were fixed after 2 h of interaction and labeled with the anti-pTyr Ab. TheZ-series images were then analyzed using Metamorph software; for each conjugate, the T-DC contact zone was delineated, and average fluorescenceintensity was quantified. Only the Z-section with the maximal value was selected for each labeling. Pairs of values were then copied out on the dot plotdepicted in this study (n � 85). The significance of the correlation between the two variables was assessed using Pearson’s r test (r � 0.825; p � 0.01).C, GFP-Akt-PH-transfected T cells were fixed after 4 h of interaction with SAg (0.2 �g/ml)-pulsed DCs and labeled with the anti-pTyr Ab. DIC andfluorescence images of the same conjugates are shown.
4165The Journal of Immunology
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

cells (Fig. 7A). After Ag recognition, FoxO1-GFP was totally ex-cluded from the nucleus, a feature found in �80% of transfectedT cells with extensive contacts with DCs. This is illustrated in Fig.7B, left panel, which shows several T-DC conjugates analyzedafter 20 min of interaction. It should be noted that many cellsexhibited a higher accumulation in discrete regions of the cyto-plasm, but not systematically and not always in the synaptic re-gion. This nuclear exclusion was never observed with the FoxO1AAA-GFP mutant that inhibits cell proliferation (Fig. 7B, rightpanel). Shown in Fig. 7C are sequential images extracted from atime-lapse experiment (supplemental Video 2), indicating addi-tionally that translocation of FoxO1 is not immediate after thebeginning of the contact, in contrast to the translocation of Akt,which usually starts concurrently with contact establishment (10).Moreover, it can be seen that the redistribution of FoxO1 is notpolarized at any time point. Prolonged incubations were next per-formed to analyze the localization of FoxO1 after several hours ofinteraction between T cells and DCs. A similar translocation ofFoxO1-GFP was observed, as illustrated in the conjugates shownin Fig. 7D, left panel, and also in the quantitative analysis in Fig.7D, right panel, demonstrating that the very large proportion of Tcells with nuclear exclusion of FoxO1-GFP was fully maintainedover time. The PI3K inhibitor wortmannin added on preformedT-DC conjugates antagonized this sustained cytoplasmic retentionof FoxO1 (Fig. 7E). Together, these findings demonstrate that in Tcells contacting APCs, FoxO1 is strongly and enduringly unbal-anced toward the cytoplasm downstream of PI3K and Akt.
DiscussionUntil recently, limited information has been available regardingthe role of PI3K in T lymphocytes after IS formation. Recent stud-ies using imaging approaches have revealed that this specific cell-to-cell interaction is a powerful trigger for PI3K activation (10,11). These studies also revealed that, unlike other systems, con-tinuous accumulation of 3�-PI is required for Ag-induced prolif-eration of T cells. Thus, understanding how this prolonged accu-mulation of PtdIns(3,4,5)P3 is implemented by the IS and how itcan result in T cell proliferation is of critical importance.
3�-PI content increases immediately at the IS after Ag recogni-tion in T cells contacting APCs (10, 11). Subsequently, 3�-PIsrapidly accumulate far beyond the contact zone. Because PI3Krecruitment at the plasma membrane is essential to catalyze thephosphorylation of membrane phosphoinositides (8, 13), we ex-amined the trafficking of one of its main regulatory subunit, p85�,during conjugate formation. Our results show that PI3K recruit-ment during T cell Ag recognition is a highly polarized processand is localized primarily to the region of the IS. This was foundat the earliest time points of conjugate formation, in agreementwith our previous observations that a GFP-Akt-PH probe begins totranslocate to the T cell plasma membrane at the onset of the re-sponse in the small contact area of the T cell with the APC (10).In this study we found that p85� remained strictly concentrated inthis membrane region when the contact continues. A role for the ISin maintaining the 3�-PI signal for hours in murine T cells has beenreported (11). Continuous activation is probably involved, becausedestabilizing preformed T cell-APC conjugates with an MHC classII-peptide specific Ab reverses the membrane localization of GFP-Akt PH (15). We found that persistent activation also takes placeat the T-APC synapse in human T cells, illustrated by the sustainedincrease in pTyr that occurs at the contact zone. Although thespecific mechanisms accounting for this accumulation of pTyr re-main to be clarified, this is likely to explain the prolonged local-ization of GFP-p85� at this site. It is worth noting that incubating
FIGURE 3. Sustained pTyr levels at the synapse between human T cellsand DCs. A, Quantitative analysis showing the percentage of T cell-DCconjugates with increased pTyr at the synapse relative to the total numberof conjugates after different periods of interaction. Data represent themean � SD of three independent experiments with �100 cells analyzed ineach. B, Distribution of pTyr levels in T cells interacting with DCs fordifferent periods of incubation (n � 80). The pTyr level is shown as theaverage fluorescence intensity within the whole T cell, quantified usingMetamorph software. Polynomial curves fitting these distributions also areshown.
FIGURE 4. p85� PI3-K overexpression inhibits 3�-PI synthesis. Humanresting T cells were cotransfected with CFP-Akt-PH and YFP-p85�, andfluorescence was analyzed on fixed conjugates after 2 h of interaction withRaji B cells in the presence of SAg (0.2 �g/ml). Three typical conjugatesfrom the same cell preparation with T cells expressing different levels ofYFP-p85� are shown.
4166 PI3K ACTIVATION AND FoxO1 NUCLEAR EXCLUSION IN T CELLS
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

preformed conjugates with the Src kinase inhibitor PP2 efficientlyreversed both pTyr labeling and GFP-Akt-PH translocation (notshown). We do not believe that the SAg presentation to human Tcells used in our study explains this continued signaling, becausesustained pTyr has already been shown in a human CD4� T cellclone activated by a nominal Ag (32). Moreover, increased pTyrand 3�-PI lipid synthesis are seen with the T8.1 hybridoma murineT cell model specific for a given peptide (not shown). This sug-gests that human and murine T cells may use similar signalingstrategies to maintain sustained changes in 3�-PI metabolism. In
this case, one should expect to find persistent changes pTyr accu-mulation in the two separate murine TCR transgenic models inwhich sustained PI3K activation has been observed (11, 15). Thiswill require additional experiments, but it will be interesting totest this hypothesis with various agonist peptide systems, be-cause pTyr increased only transiently in another TCR transgenic Tcell model (33).
The precise class(s) of PI3K that is activated by the TCR is amatter of controversy. For example, the examination of PI3K p85�knockout mice has failed to show a significant effect on T cell
FIGURE 5. Rapid diffusion of 3�-PI from the syn-apse in the whole plasma membrane. A, Representativeimages showing a typical FRAP experiment on GFP-Akt-PH in T8.1 murine T cells after 1 h of contact withL625.7 fibroblasts loaded with specific Ag. The T cellwas imaged at regular intervals before and after a624-ms bleach pulse at maximum laser power focusedon a small region at the rear of the cell (outlined box).t � 0 s corresponds to the beginning of the recoveryperiod. B, Fluorescence intensity, normalized as a per-centage of the initial fluorescence, from quantitativeFRAP analysis was plotted against time. Mean valuesfor 11 representative cells from two independent exper-iments are shown. C, FLIP experiments were performedon GFP-Akt-PH T8.1 cells after 1 h of contact withAg-loaded L625.7 fibroblasts. Cells were repeatedlybleached in the indicated zone (white box) and imagedbetween consecutive bleach pulses. A detail of thebleached area with higher magnification is shown in theright corner of each panel. D, Left panel, Time courseof mean fluorescence intensities measured in eight sep-arate cells in bleached areas and in regions of equal arealocated within the synapse and on the side of the Tcells, as illustrated by the blue and red boxes in C.Fluorescence in neighboring unbleached cells was plot-ted as a control. Right panel, In this representative ex-periment, independent cells with similar fluorescencelevels and contacting L625.7 APCs were repeatedlyphotobleached within a small membrane region outsidethe IS, at the rear of the cell, or within the IS. The lossof fluorescence intensity as a function of time in thewhole T cell was then plotted. In the two panels, fluo-rescence intensities were normalized as a percentage ofthe initial fluorescence.
4167The Journal of Immunology
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

activation (34, 35), and a compensatory role for p85� has beendiscussed (36). Although our results do not resolve this issue, thefinding that competing for PI3K pTyr-binding sites within the ISinhibits GFP-Akt-PH membrane recruitment (Fig. 4) supports theconcept that the binding of class IA PI3K to pTyr contained withinthe IS plays an important role in the activation of 3�-PI metabolism inT cells. Together with the photobleaching data, our experiments in-dicate that the broad distribution of GFP-Akt-PH in the whole plasma
membrane is due to continuous spreading of PtdIns(3,4,5)P3 out of theIS, the major site of PtdIns(3,4,5)P3 production in T cells contactingAPCs. This conclusion is supported by the disappearance of mem-brane GFP-Akt-PH in the whole cell after a few photobleaching iter-ations given within the synapse (Fig. 5D).
Maintaining PI3K in an active state is required to induce T cellproliferation in response to Ag stimulation (11). Several 3�-PI-dependent pathways that control cell division have been described
FIGURE 6. Constitutively active FoxO1 inhibits Ag-induced T cell growth, but not early T cell activation. A, Human resting T cells were nucleoporatedwith vectors expressing GFP alone or either FoxO1-GFP or the constitutively active FoxO1 AAA-GFP protein. Cell apoptosis was measured by FACS withannexin V after 12- and 36-h cell culture without any stimulus on the GFP-gated T cell populations. Data represent the mean � SD of three independentexperiments. �, p � 0.05; ��, p � 0.01 (by Student’s t test, comparing either FoxO1 or FoxO1 AAA to empty vector). B, T cells were nucleoporated asdescribed in A, and 24 h posttransfection they were incubated for the indicated periods of time with irradiated SAg-pulsed (20 ng/ml) or unpulsed Raji Bcells as APCs. CD69 expression was then analyzed on the GFP-gated T cell populations by FACS. Data represent the mean � SD of three independentexperiments. �, p � 0.1; ��, p � 0.01 (by Student’s t test, comparing either FoxO1 or FoxO1 AAA to empty vector). C, Twenty-four hours posttransfectionT cells were incubated with irradiated SAg-pulsed (20 ng/ml) Raji B cells. T cell size increase was then measured after a 2-day culture on GFP-positiveand -negative viable T cells within the forward and side scatter lymphocyte gate R1 delineated on the whole cell population and positioned according tothe forward and side scatter profile of untransfected T cells incubated with APCs (upper panel). A total of 2 � 103 cells accumulated in each dot plot. Thedata shown are representative of four different experiments. D, In a separate experiment, after 3 days of stimulation in the presence of APCs, cells werelabeled with propidium iodide to stain nucleic acids. Viable T cells gated on R1 (left panels) were analyzed for red (PI) and green (GFP) fluorescence. Thedata shown are representative of two different experiments.
4168 PI3K ACTIVATION AND FoxO1 NUCLEAR EXCLUSION IN T CELLS
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

FIGURE 7. Ag-induced, PI3K-dependent, nuclear exclusion of FoxO1 in T cells. A, FoxO1-GFP distribution in resting human T cells. B, Distributionof FoxO1-GFP or FoxO1 AAA-GFP mutant in human T cells after 30 min of interaction with SAg (0.2 �g/ml)-pulsed DCs. C, Left, In a time-lapse imagingexperiment, FoxO1-GFP-transfected cells were added to SAg-pulsed (0.2 �g/ml) Raji B cells. Images were extracted from a video (supplemental Video2) showing the formation of a typical conjugate. Right, Fluorescence intensities across the black line (shown in the corresponding images) are plotted asline intensity histograms after two-dimensional deconvolution on 12-bit images. D, Left, DIC and FoxO1-GFP fluorescence images of T-DC conjugates after4 h of interaction. Right, Quantitative analysis showing the percentage of T cells in conjugates with a cytoplasmic or a nuclear localization of FoxO1-GFPafter different periods of incubation. Results are for �100 cells analyzed from randomly acquired fields. E, Left, DIC and FoxO1-GFP fluorescence imagesof Ag-induced T-DC conjugates after 4 h of interaction. Wortmannin (100 nM) was added during the last 2 h of contact. Right, Quantitative analysisshowing the percentage of T cells with a cytoplasmic or a nuclear localization of FoxO1-GFP with or without wortmannin. Results are shown for �100cells in each group analyzed from randomly acquired fields.
4169The Journal of Immunology
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

(37). Recent studies in several cell systems have shown that Aktsuppresses transactivation and promotes nuclear exclusion ofFoxO transcription factors (20, 24, 25). Unopposed, FoxO proteinspromote cell cycle arrest, at least in part by promoting the synthe-sis of cell cycle inhibitory factors, including the cyclin-dependentkinase inhibitor, p27Kip1, and p21 (26–29, 38) and suppressingthe expression of cyclin D1 (39). FoxO proteins also can stimulatethe expression of proapoptotic molecules, including Bim and Fasligand (26). Recently, some information regarding the function ofFoxO proteins in T cells has been made available. FoxO proteinshave been shown to induce cell cycle arrest in Jurkat leukemic Tcells (29, 31) and are phosphorylated downstream of the IL-2R toregulate IL-2-mediated cell survival in the CTLL-2 murine T cellline (26). In addition, mice defective in FoxO3a have been gener-ated that exhibit spontaneous inflammatory syndrome and lympho-proliferation mainly composed of T cells (30). In the present studywe thus asked whether the ability of sustained 3�-PI signaling throughthe IS to trigger a T cell response after Ag encounter involves theregulation of another FoxO transcription factor, FoxO1.
We found that FoxO1 is translocated to the cytoplasmic com-partment when human T cells interact with APCs, dependent onthe residues that are phosphorylated by Akt. Kinetic analysis re-veals that translocation of FoxO1 begins after the initiation of cellconjugation (see Fig. 7B and supplemental Video 2) and the firstaccumulation of Akt-PH at the plasma membrane (10), consistentwith the concept that the binding and subsequent activation of Aktat the plasma membrane are required for the redistribution ofFoxO1. Video-imaging records also suggest that FoxO1 does notpreferentially translocate into the cytoplasm region facing theAPC. This lack of directionality contrasts with the recent findingthat full-length Akt preferentially localized to the IS in a murine Tcell clone (40). Our results also indicate that maintaining FoxO1outside the nucleus for an extended time is dependent on sustainedPI3K activity. This process appears to be critical for initiating cellcycle progression in T cells after Ag recognition, because the ex-pression of constitutively active FoxO1 strongly antagonized Tcell growth and cell cycle progression, as recently shown in Bcells, a cell system in which some cell apoptosis is induced byactive FoxO molecules (41). Interestingly, it has been observedwith specific Abs that the PI3K-Akt pathway links CD28 costimu-lation of human resting T cells to cell cycle progression with aparallel decrease in p27Kip1, a well-known target of FoxO tran-scription factors, whereas CD3 has a less critical role (42). Con-tinuous engagement of CD28 within the IS might therefore alsopromote strong support for the T cell to proliferate through thispathway.
Quiescence in lymphocytes is an actively maintained state thatreflects the effects of multiple factors known to function as tumorsuppressor genes, rather than just a default pathway (43–45), andin T cells, PI3K probably promotes survival and proliferation by itscoordinate and concurrent actions on several molecular targets in-volved in these processes (36, 45). We have shown in this reportthat through its effects on the function of factors such as FoxO1,the PI3K-Akt pathway certainly helps T cells escape from thiscontrol after Ag recognition. Obviously, many other effectors ofPI3K are probably involved to cooperatively trigger these biolog-ical processes with FoxO1; additional studies will be necessary todiscover its specific contribution. The functions of the so-calledmature or stabilized synapse formed by T cells with APCs are stillincompletely understood. Studies have called attention to the rolethat this structure plays in modulating and slowing down the T cellresponse to Ag (46). In this study we have shown that the IS alsocan act as a very effective platform for promoting the sustainedrecruitment and activation of PI3K, thereby suppressing the func-
tion of FoxO proteins, resulting in T cell proliferation. Interest-ingly, it has recently been reported that sustained Akt signalinginduced by late-acting and inducible costimulatory molecules alsoregulates long term T cell survival for several days after Ag en-counter (47). Thus, the need for sustained TCR and/or costimula-tory signaling(s) is probably a straightforward means used by Tcells and APCs to maintain continuous activation of PI3K-depen-dent metabolic cascades to shape the different steps of the immuneresponse.
AcknowledgmentsWe thank J. Delon, E. Donnadieu, and C. Randriamampita for their com-ments on the manuscript.
DisclosuresThe authors have no financial conflict of interest.
References1. Shaw, A. S., and M. L. Dustin. 1997. Making the T cell receptor go the distance:
a topological view of T cell activation. Immunity 6:361.2. Grakoui, A., S. K. Bromley, C. Sumen, M. M. Davis, A. S. Shaw, P. M. Allen,
and M. L. Dustin. 1999. The immunological synapse: a molecular machine con-trolling T cell activation. Science 285:221.
3. Monks, C. R. F., B. A. Freiberg, H. Kupfer, N. Sciaky, and A. Kupfer. 1999.Three-dimensional segregation of supramolacular activation clusters in T cells.Nature 395:82.
4. van der Merwe, P. A. 2002. Formation and function of the immunological syn-apse. Curr. Opin. Immunol. 14:293.
5. Ward, S. G., and D. A. Cantrell. 2001. Phosphoinositide 3-kinases in T lympho-cyte activation. Curr. Opin. Immunol. 13:332.
6. Okkenhaug, K., and B. Vanhaesebroeck. 2003. PI3K in lymphocyte develop-ment, differentiation and activation. Nat. Rev. Immunol. 3:317.
7. Koyasu, S. 2003. The role of PI3K in immune cells. Nat. Immunol. 4:313.8. Seminario, M. C., and R. L. Wange. 2002. Signaling pathways of D3-phospho-
inositide-binding kinases in T cells and their regulation by PTEN. Semin. Immu-nol. 14:27.
9. Fruman, D. A., and L. C. Cantley. 2002. Phosphoinositide 3-kinase in immuno-logical systems. Semin. Immunol. 14:7.
10. Harriague, J., and G. Bismuth. 2002. Imaging antigen-induced PI3K activation inT cells. Nat. Immunol. 3:1090.
11. Costello, P. S., M. Gallagher, and D. A. Cantrell. 2002. Sustained and dynamicinositol lipid metabolism inside and outside the immunological synapse. Nat.Immunol. 3:1082.
12. Katso, R., K. Okkenhaug, K. Ahmadi, S. White, J. Timms, and M. D. Waterfield.2001. Cellular function of phosphoinositide 3-kinases: implications for develop-ment, homeostasis, and cancer. Annu. Rev. Cell. Dev. Biol. 17:615.
13. Carpenter, C. L., K. R. Auger, M. Chanudhuri, M. Yoakim, B. Schaffhausen,S. Shoelson, and L. C. Cantley. 1993. Phosphoinositide 3-kinase is activated byphosphopeptides that bind to the SH2 domains of the 85-kDa subunit. J. Biol.Chem. 268:9478.
14. Rebecchi, M. J., and S. Scarlata. 1998. Pleckstrin homology domains: a commonfold with diverse functions. Annu. Rev. Biophys. Biomol. Struct. 27:503.
15. Huppa, J. B., M. Gleimer, C. Sumen, and M. M. Davis. 2003. Continuous T cellreceptor signaling required for synapse maintenance and full effector potential.Nat. Immunol. 4:749.
16. Haugh, J. M., F. Codazzi, M. Teruel, and T. Meyer. 2000. Spatial sensing infibroblasts mediated by 3� phosphoinositides. J. Cell Biol. 151:1269.
17. Meili, R., C. Ellsworth, S. Lee, T. B. Reddy, H. Ma, and R. A. Firtel. 1999.Chemoattractant-mediated transient activation and membrane localization of Akt/PKB is required for efficient chemotaxis to cAMP in Dictyostelium. EMBO J.18:2092.
18. Servant, G., O. D. Weiner, P. Herzmark, T. Balla, J. W. Sedat, and H. R. Bourne.2000. Polarization of chemoattractant receptor signaling during neutrophil che-motaxis. Science 287:1037.
19. Wang, F., P. Herzmark, O. D. Weiner, S. Srinivasan, G. Servant, andH. R. Bourne. 2002. Lipid products of PI(3)Ks maintain persistent cell polarityand directed motility in neutrophils. Nat. Cell. Biol. 4:513.
20. Zhang, X., L. Gan, H. Pan, S. Guo, X. He, S. T. Olson, A. Mesecar, S. Adam, andT. G. Unterman. 2002. Phosphorylation of serine 256 suppresses transactivationby FKHR (FOXO1) by multiple mechanisms: direct and indirect effects on nu-clear/cytoplasmic shuttling and DNA binding. J. Biol. Chem. 277:45276.
21. Sasaki, T., J. Irie-Sasaki, R. G. Jones, A. J. Oliveira-dos-Santos, W. L. Stanford,B. Bolon, A. Wakeham, A. Itie, D. Bouchard, I. Kozieradzki, et al. 2000. Func-tion of PI3K� in thymocyte development, T cell activation, and neutrophil mi-gration. Science 287:1040.
22. Ueki, K., P. Algenstaedt, F. Mauvais-Jarvis, and C. R. Kahn. 2000. Positive andnegative regulation of phosphoinositide 3-kinase-dependent signaling pathwaysby three different gene products of the p85� regulatory subunit. Mol. Cell. Biol.20:8035.
23. Schneider, I. C., and J. M. Haugh. 2004. Spatial analysis of 3� phosphoinositidesignaling in living fibroblasts. II. Parameter estimates for individual cells fromexperiments. Biophys J. 86:599.
4170 PI3K ACTIVATION AND FoxO1 NUCLEAR EXCLUSION IN T CELLS
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

24. Rena, G., S. Guo, S. C. Cichy, T. G. Unterman, and P. Cohen. 1999. Phosphor-ylation of the transcription factor forkhead family member FKHR by proteinkinase B. J. Biol. Chem. 274:17179.
25. Biggs, W. H., III, J. Meisenhelder, T. Hunter, W. K. Cavenee, and K. C. Arden.1999. Protein kinase B/Akt-mediated phosphorylation promotes nuclear exclu-sion of the winged helix transcription factor FKHR1. Proc. Natl. Acad. Sci. USA96:7421.
26. Stahl, M., P. F. Dijkers, G. J. Kops, S. M. Lens, P. J. Coffer, B. M. Burgering, andR. H. Medema. 2002. The forkhead transcription factor FoxO regulates transcrip-tion of p27Kip1 and Bim in response to IL-2. J. Immunol. 168:5024.
27. Dijkers, P. F., R. H. Medema, C. Pals, L. Banerji, N. S. Thomas, E. W. Lam,B. M. Burgering, J. A. Raaijmakers, J. W. Lammers, L. Koenderman, et al. 2000.Forkhead transcription factor FKHR-L1 modulates cytokine-dependent transcrip-tional regulation of p27KIP1. Mol. Cell. Biol. 20:9138.
28. Nakamura, N., S. Ramaswamy, F. Vazquez, S. Signoretti, M. Loda, andW. R. Sellers. 2000. Forkhead transcription factors are critical effectors of celldeath and cell cycle arrest downstream of PTEN. Mol. Cell. Biol. 20:8969.
29. Medema, R. H., G. J. Kops, J. L. Bos, and B. M. Burgering. 2000. AFX-likeForkhead transcription factors mediate cell-cycle regulation by Ras and PKBthrough p27kip1. Nature 404:782.
30. Lin, L., J. D. Hron, and S. L. Peng. 2004. Regulation of NF-�B, Th activation,and autoinflammation by the forkhead transcription factor Foxo3a. Immunity21:203.
31. Leenders, H., S. Whiffield, C. Benoist, and D. Mathis. 2000. Role of the forkheadtranscription family member, FKHR, in thymocyte differentiation. Eur. J. Immu-nol. 30:2980.
32. Muller, S., S. Demotz, C. Bulliard, and S. Valitutti. 1999. Kinetics and extent ofprotein tyrosine kinase activation in individual T cells upon antigenic stimulation.Immunology 97:287.
33. Lee, K. H., A. D. Holdorf, M. L. Dustin, A. C. Chan, P. M. Allen, and A. S. Shaw.2002. T cell receptor signaling precedes immunological synapse formation. Sci-ence 295:1539.
34. Fruman, D. A., S. B. Snapper, C. M. Yballe, L. Davidson, J. Y. Yu, F. W. Alt,and L. C. Cantley. 1999. Impaired B cell development and proliferation in ab-sence of phosphoinositide 3-kinase p85�. Science 283:393.
35. Suzuki, H., Y. Terauchi, M. Fujiwara, S. Aizawa, Y. Yazaki, T. Kadowaki, andS. Koyasu. 1999. Xid-like immunodeficiency in mice with disruption of the p85�subunit of phosphoinositide 3-kinase. Science 283:390.
36. Kane, L. P., and A. Weiss. 2003. The PI-3 kinase/Akt pathway and T cell acti-vation: pleiotropic pathways downstream of PIP3. Immunol. Rev. 192:7.
37. Vanhaesebroeck, B., S. J. Leevers, K. Ahmadi, J. Timms, R. Katso,P. C. Driscoll, R. Woscholski, P. J. Parker, and M. D. Waterfield. 2001. Synthesisand function of 3-phosphorylated inositol lipids. Annu. Rev. Biochem. 70:535.
38. Seoane, J., H. V. Le, L. Shen, S. A. Anderson, and J. Massague. 2004. Integrationof Smad and forkhead pathways in the control of neuroepithelial and glioblas-toma cell proliferation. Cell 117:211.
39. Schmidt, M., S. Fernandez de Mattos, A. van der Horst, R. Klompmaker,G. J. Kops, E. W. Lam, B. M. Burgering, and R. H. Medema. 2002. Cell cycleinhibition by FoxO forkhead transcription factors involves downregulation ofcyclin D. Mol. Cell. Biol. 22:7842.
40. Kane, L. P., M. N. Mollenauer, and A. Weiss. 2004. A proline-rich motif in theC terminus of Akt contributes to its localization in the immunological synapse.J. Immunol. 172:5441.
41. Yusuf, I., X. Zhu, M. G. Kharas, J. Chen, and D. A. Fruman. 2004. Optimal B cellproliferation requires phosphoinositide 3-kinase-dependent inactivation of FOXOtranscription factors. Blood 104:784.
42. Appleman, L. J., A. A. van Puijenbroek, K. M. Shu, L. M. Nadler, andV. A. Boussiotis. 2002. CD28 costimulation mediates down-regulation ofp27kip1 and cell cycle progression by activation of the PI3K/PKB signalingpathway in primary human T cells. J. Immunol. 168:2729.
43. Burgering, B. M., and G. J. Kops. 2002. Cell cycle and death control: long liveForkheads. Trends Biochem. Sci. 27:352.
44. Birkenkamp, K. U., and P. J. Coffer. 2003. FOXO transcription factors as regu-lators of immune homeostasis: molecules to die for? J. Immunol. 171:1623.
45. Yusuf, I., and D. A. Fruman. 2003. Regulation of quiescence in lymphocytes.Trends Immunol. 24:380.
46. Lee, K. H., A. R. Dinner, C. Tu, G. Campi, S. Raychaudhuri, R. Varma,T. N. Sims, W. R. Burack, H. Wu, J. Wang, et al. 2003. The immunologicalsynapse balances T cell receptor signaling and degradation. Science 302:1218.
47. Song, J., S. Salek-Ardakani, P. R. Rogers, M. Cheng, L. Van Parijs, and M. Croft.2004. The costimulation-regulated duration of PKB activation controls T celllongevity. Nat. Immunol. 5:150.
4171The Journal of Immunology
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from




















