
Stability Analysis in Plant Breeding
24
Plant Breeding 101, 1—23 (1988) © 1988 Paul Parey Scientific Publishers, Berlin and Hamburg ISSN 0179-9541 Review Department of Crop Genetics and Breeding, The Swedish University of Agricultural Sciences, Svalov (Sweden); Institute of Crop Science and Plant Breeding, Christian-Albrechts-Umversitdt, Kiel (Germany, T.R.) Stability Analysis in Plant Breeding H. C. BECKER and J. LEON With S figures and 6 tables Received April 12, 1988 / Accepted April 21, 1988 ]. Introduction Successful new varieties must show high per- formance for yield and other essenti,al ag- ronomic traits. Moreover, their superiority ,should be reliible 'Over a wide range of envi- ronmental conditions. Plant breeders generally agree on the importance of a high yield stabih- ty, but there is less accord on the most appro- priate definition of 'stability' and on methods to measure and to improve yield stabihty. The basic cause of differences between geno- types in their yield stability is the wide O'Ccur- reoce of genotype-environment interactions (GE-interactions), i.e. the ranking of geno- types depends on the particular environmental conditiotis where they are grown. These in- teractions of genotypes with environments can be partly understood as a result of a differential reaction to environmental stress factors like drought or diseases, and consequently resist- ance breeding is of significance in improving yield stability. But generally, only a minor pan of the GE- interactions can be attributed to known envi- ronmental determinants and the major part is just an inexplicable quantity in the statistical analysis of yield trials. When discussing these unexpected variations in yield the term "phenotypic stability" is often used to refer to fluctuations in the phenotypic expression of yield while the geno'typic composition of the varieties or populations remains stable. Phenotypic stability has been extensively studied by biometncians wliO' have developed numerous methods to analyse this phenome- non and by breedmg researchers who per- formed and published hundreds of experi- ments. Nevertheless, up to now a statistical measure of yield stability is hardly used in practical breeding pogrammes. It is the aim of the present paper to review the methods of stability analysis and to discuss the prospects and limitations of improving }-ield stability with the aid of biometrics. We will confine ourselves to methods which clas- sify genotypes; very similar approaches can be used to classif)' environments according to their suitability for yield testing, but this prob- lem is beyond the scope of this contribution. 2. Basic concepts of stability The terms phenotypic stability, jdeld stability, and adaptation are often used m quite different senses. Therefore DORST (1957) remarked: "The word adaptation has a great adaptabili- ty." And recently LIN et al. (1986) amplified: "It (the concept of stability) is defined in many ways depending on how the scientist wishes to look at the problem." In fact, depending on the goal and on the character under considera- tion, two different concepts of stability exist, which are termed as the static concept of stabil- ity and as the dynamic concept of stability (LEON 1985). Both concepts of stability are U.S. Copyright Ckaraacc Center Code Sutemen.: 0 1 7 9 - 9 5 4 1 / 8 8 / 0 1 0 1 - 0 0 0 1 $ 0 2 . 5 0 / 0
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Stability Analysis in Plant Breeding

Plant Breeding 101, 1—23 (1988)© 1988 Paul Parey Scientific Publishers, Berlin and HamburgISSN 0179-9541
Review
Department of Crop Genetics and Breeding,The Swedish University of Agricultural Sciences, Svalov (Sweden);Institute of Crop Science and Plant Breeding,Christian-Albrechts-Umversitdt, Kiel (Germany, T.R.)
Stability Analysis in Plant Breeding
H. C. BECKER and J. LEON
With S figures and 6 tables
Received April 12, 1988 / Accepted April 21, 1988
]. IntroductionSuccessful new varieties must show high per-formance for yield and other essenti,al ag-ronomic traits. Moreover, their superiority,should be reliible 'Over a wide range of envi-ronmental conditions. Plant breeders generallyagree on the importance of a high yield stabih-ty, but there is less accord on the most appro-priate definition of 'stability' and on methodsto measure and to improve yield stabihty.
The basic cause of differences between geno-types in their yield stability is the wide O'Ccur-reoce of genotype-environment interactions(GE-interactions), i.e. the ranking of geno-types depends on the particular environmentalconditiotis where they are grown. These in-teractions of genotypes with environments canbe partly understood as a result of a differentialreaction to environmental stress factors likedrought or diseases, and consequently resist-ance breeding is of significance in improvingyield stability.
But generally, only a minor pan of the GE-interactions can be attributed to known envi-ronmental determinants and the major part isjust an inexplicable quantity in the statisticalanalysis of yield trials. When discussing theseunexpected variations in yield the term"phenotypic stability" is often used to refer tofluctuations in the phenotypic expression ofyield while the geno'typic composition of thevarieties or populations remains stable.
Phenotypic stability has been extensivelystudied by biometncians wliO' have developednumerous methods to analyse this phenome-non and by breedmg researchers who per-formed and published hundreds of experi-ments. Nevertheless, up to now a statisticalmeasure of yield stability is hardly used inpractical breeding pogrammes.
It is the aim of the present paper to reviewthe methods of stability analysis and to discussthe prospects and limitations of improving}-ield stability with the aid of biometrics. Wewill confine ourselves to methods which clas-sify genotypes; very similar approaches can beused to classif)' environments according totheir suitability for yield testing, but this prob-lem is beyond the scope of this contribution.
2. Basic concepts of stabilityThe terms phenotypic stability, jdeld stability,and adaptation are often used m quite differentsenses. Therefore DORST (1957) remarked:"The word adaptation has a great adaptabili-ty." And recently LIN et al. (1986) amplified:"It (the concept of stability) is defined in manyways depending on how the scientist wishes tolook at the problem." In fact, depending onthe goal and on the character under considera-tion, two different concepts of stability exist,which are termed as the static concept of stabil-ity and as the dynamic concept of stability(LEON 1985). Both concepts of stability are
U.S. Copyright Ckaraacc Center Code Sutemen.: 01 7 9 - 9 5 4 1 / 8 8 / 0 1 01 -0001 $ 0 2 . 5 0 / 0

BECKER and LEON
valuable but their application depends on thetrait considered.
With regard to the static concept a stablegenotyp'e possesses an unchanged performanceregardless of any variation of the etiviron-mental conditions. This stable genotype showsno deviation from the expected character level,that means its variance among environments iszero.
Unlike this static concept, where a stablegenotype has a constant performance level, thedynamic concept permits a predictable re-sponse to environments and a stable genotypeaccording to the dynamic concept has no devi-ation from this response to environments. Foreach envirO'nment the performance of a stablegenotype corresponds completely to the esti-mated level or the prediction. In the dynamicconcept of stability, it is not required that thegenotypic response to environmental condi-tions should be equal for all genotypes. Whatis important, ho^n^ever, is the agreement of theestimated or predicted level with the level ofperformance actually measured when definingstability'. BECKER (1981a) termed this type ofstability the agronomic concept and distin-guished it from the biological concept of stabil-ity, which is equivalent to the static concept.
Numerous methods have been proposed toestimate phenotypic stability or to analyse GE-interactions, which are strongly related to sta-bility according to the dynamic concept. Utii-variate parametric stability statistics measureuncertaint)' in the respective biometrical analy-sis. In additioti, univanate nonparametric sta-bility statistics have been proposed, which arebased on rank orders of genotypes and whichdo not need any as,sumptions about distribu-tion of obser\'ed values or of variancehomogeneity. Multivariate techniques have al-so been introduced for stability analysis. Thesemultivariate methods provide further informa-tion on the real multivariate response of geno-types to environments.
3. Biometrical methods to estimate sta-bility
The following chapters describe stability statis-tics of the static concept and afterwards statis-tics of the dynamic concept, for which para-metric and nonparametric measures are pro-posed. Because only the most important statis-
tics are considered, the reader is referred to LINet al. (1986), WESTCOTT (1986), HUH.\' (1987a)and N.«s.'\R and HUHN (1987) for further litera-ture and mformation.
In order to present the methods of stabilityanalysis, a two-way linear model is assumedfor convenience as follows:
X;i = fi - e, + gi + (ge)., + Ejjwhere Xij is the observed phenotypic meanvalue of genotype i (i = 1, . . ., G) in environ-ment j (j = 1, . . ., E), and u, e,, g,, (ge)., and E;,represent the overall population mean, the ef-fect of the j-th environment, the effect of thei-th genotype, the effect of the interactionbetween the i-th genotype and the ]-th envi-ronment and the mean random error of the l-thgenotype in the )-tli environment, respectively,with X,, Xj and X. denoting the marginalmeans of genotype i, environment j and theoverall mean, respectively.
J.I Static
Phenotypic stability according to the staticconcept was measured as carK' as 1917 byRoE.MER and later on bv numerous authorsusing the variance of a genotype across envi-
. I (X., - %fronments:
environmental variance:E-1
This 'environmental variance of genotypes' de-tects all deviations from the genotypic mean.Well known significance tests for comparingvariances may be applied for an assessment ofgenotypes.
Using the environmental variance, a desir-able genotype will not react at all in changingenvironmental conditions. This concept of sta-bility is useful for traits, the levels of whichhave to be maintained at all costs, e.g, iorquality traits, for resistance against diseases, orfor stress characters like winter hardiness. Butfor yield, a breeder aims at finding genotypeswhich are stable and high-yielding simultane-ously. Stability evaluated by means of the sta-tic concept, however, is usually associatedwith a relatively poor yield level. Therefore thedynamic concept is recommended for studiesof yield stability.
3.2 Dynamic concept
Frequently with yield or other quantitativetraits most genotypes react similarly to favour-

Stability Analysis in Plant Breeding
Fig. 1. Graphical re'presentation of GE-interactions: the stability statistic ecova-lence (W,) is the sum of squares of thedeviations from the upper straight line
120-
100-
80
60
40
20
yield
able or unfa^i'ourable environmental condi-tions. This average response to environmentsresults in var)'ing mean trait levels among envi-ronments. According to the dynamic conceptonl}' the deviations 'of a genotype from thisgeneral reaction are considered as a contribu-tion to instability, because the general responseof all genotypes may be interpreted as an envi-ronmental effect.
If the usual biometrical two-way model isapplied for anaK'smg a series of trials withgenot)'pes tested m several environments, esti-mates of genotypic and environmental effectsare obtained. These effects can only explainpart of the variation if GE-interactions exist.These unexplained deviations represent the un-certainty of an estimation or ot a prediction ofthe phenotypic value of genotype i in environ-ment j . Therefore, these GE-interaction effectsdescribe the uncertainty or instability m themodel, or vice versa they measure stabilitywith genotypes possessing small contributionsto GE-mteractions as being more stable thangenotypes with larger contributions. All stabil-ity procedures based on quantifying GE-m-teraction effects belong to the dynamic stabili-ty concept. This also holds true for non-parametric stability statistics, which are basedon rank orders, for environmental variation ofthe ranks of a genotype is predominantlycaused by GE-interactions.
20 40 60 60 100 120
E n v i r o n m e n t a l m e a n ( X , j )
3.2.1 Partitioning of GE-interactions
S.-\L.vioN (1951) and PL.MSTED and PETERSON(1959) computed an analysis of variance forevery pair of genotypes to estimate the interac-tion \'ariance for ever}' combination of twogenotypes. The mean of the interaction var-iances obtained for each genotype was used asan indication of the contribution of that geno-type to the total GE-interactions (WESTCOTT1986).
WRiCKt (1962) proposed using the GE-in-teraction effects for e,ach genotype, squaredand summed across all environments, as a sta-bility measure. This statistic, termed ecova-lence (W;), is far more simple to compute and'more directly related to the GE-interactionsthan the statistics proposed by SAEMON (1951)and PLAISTED and PETERSON (1959) and may be
estimated as follows:ecovalence: W, = X (X,j - X,, - X,, -f X.,)-
Because ecovalence measures the contributionof a genotype to the GE-interactions, a geno-type -nqth Wi = 0 is regarded as stable. Accord-in" to the meaning of the word ecovalence tliisstable genot}'pe possesses a high ecovalence(low values of W; = high ecovalence).
Figure 1 presents a numerical example of aplot of yields of genotype l in various environ-ments against the respective mean of environ-

BECKER and LEON
120-
lOO-
80-
60-
40-
20-
y i e l d
y
be
Fig. 2. Graphical representation of theregression approach
20 40 60 80 100 120
Environmental mean'[>t.J
ments. The lower straight line estimates thea-s'erage yield of all genot}'pes simpl}' usinginformation about the general mean (u) and theenvironmental effects (ej), while the upperstraight line additionally takes into account thegenotypic effect (gj and therefore estimates the}-ield of genotype i. Deviations of yields fromthe upper straight line are the GE-interactioneffects of genotype l and these deviations,squared and summed across environmentsconstitute the ecovalence.
SHUKLA (1972 a) proposed an unbiased esti-mate of the variance of (ge),, + E|J for genorv'pei (LIN et al. 1986). This stability statistic istermed 'stability variance' (of) and is estimatedas follows:
1G(G-l)
I (X,, - X,, - X, + x,y- 2Z (Xi, - X, - X, + X,.) ]
The stability variance is a linear combinationof the ecovalence and therefore both W; and a]are equivalent for ranking purposes (WRICKEand WEBER 1980):
f Wj(E-1) (G-2)
Mean squares GE-interaction
G-2
As the stability variance is the difference be-tween two sums of s,qu,ares it can be negative,but negative estimates of variances arc notuncommon m variance components problems.Negative estimates of at may be taken as equalto zero as usual (SHUKLA 1972 a).
Although the distribution of W, and a] is notknown, homogeneity of the estimates can betested using SHUKLA'S (1972 b) approximate test(EIN et al. 1986).
3.2.2 Regression approach
Apph'ing the usual biometrical model, it isassumed that the effects are independent ofeach other. This assumption is fulfilled whenregarding all the genotypes together and nocovariance exists between the effects of envi-ronments and of GE-interactions. Consideringeach genotype separately, however, thiscovariance may be different from zero (COM-STOCK and MOLL 1963). The regressi'On coeffi-cient is a standardized description of thiscovariance.
STRINGHELD and SALTER (1934) were prob-ably the first to calculate a linear regressioncoefficient to characterize the specific responseof genotypes to varying climatic factors. Thisregression technique has been described andelaborated by Y,ATES and COCHRAN (1938), FIN-
LAY and WILKINSON (1963), EBERHART and RUS-
SELL (1966) and PERKINS and JINKS (1968).

Stability Analysis in Plant Breeding
The same example as presented in Figure 1has been analysed by regression (Fig. 2). Thedeviations between actual and predicted valuesnow decrease by the amount of covariancebetween environmental and GE-interaction ef-fects. The straight line Y = fi + hfi, + g, fits thedata better than does the line Y = ^ + ej -t- g^.
The effects of GE-mteraction may be e'X-pressed as follows:
(ge),, = |3,e, + d,
where (3j is the linear regression coefficient forgenotype i and dj, a deviation. Two slightlydifferent regression techniques are proposed toexplain part of GE-interactions. Either GE-interaction effects may be regressed on envi-ronmental effects (PI, PERKINS and JINKS 1968),or Xjj-values may be regressed on means ofenvironments (b;, FIM.AY and WIEKINSON
1963); both statistics are equivalent (|3i = b; —1). In addition to the coefficient of regression,the deviation mean squares (s^J describe thecontribution of genotype i to GE-interactions(EBERHART and RUSSEEE 1966):
coefficient of regression:
2 (X,, - X , -X,.+ X..] (X,, - X )bi = 1 +-J -^^
Z (X, - XJdeviation mean squares:
- (b, - ' (X,, - X..)-]
SHUKLA (1972 a) proposed a linear combina-tion of deviation mean squares as a stabilitystatistic. This statistic, denoted as sf, is anunbiased estimate of the variance of (d;, + £;,)and for ranking purposes is equivalent to thedeviation mean squares.
Both statistics of the regression method, b;and sji, are used in different ways to assess thereaction of genotypes to varying environ-mental conditions. While sj, is strongly relatedto the remaining unpredictable part of variabil-ity of any genotype and therefore is consideredas a stability parameter, the coefficient of re-gression bj characterizes the specific responseof genotypes to environmental effects and maybe regarded as response parameter (BREESE1969). Genotypes which do not react to vary-ing environmental factors show zero bj-valuesand would be stable according to the static
concept. On the other hand genotypes pos-sessing an average response to changing envi-ronmental conditions show bj-values of one.For ranking purposes the choice of desired b,-value depends on the specific goal, while inde-pendently of the objective, deviation meansquares of stable genotypes are zero.
PiNTHUs (1973) proposed to use the coeffi-cient of determination (rf) instead of deviationmean squares to estimate stability of geno-types, because rf is strongly related to sjj(BECKER 1981b):
coefficient of determination: rf = 1 — S'di
The application of rf and b; has the advantagethat both statistics are independent of the unitsof measurement.
In spite of the advant,age of a meaningfulinterpretation of the regression coefficient,serious criticisms can be made:
1. The environmental index is not independ-ent of the data analysed, because it is extractedfrom the whole set of data. FREEMAN and PER-KINS (1971) suggested that the regression ap-proach should be based on the use of an inde-pendent measure of the environment. Inde-pendent measures may be obtained by omit-ting the regressed genotype or by an additionaltest of a control group of genot)'pes. However,on testing a large number of genotypes theresults differed little when applying independ-ent measures if compared to the usual environ-mental mean (FRIPP and C.WEN 1971, SNO.- D
and ARTHUR 1976). Furthermore the bias will,as shown by WRICHT (1976), only be present inthe case of rand,O'm genotyp'ic effects. Fo'rmodels with random genor}'pic effects,WRIGHT (1976) proposed a modified estimateof regression parameters which are free frombias.
2. Regression coefficients are biased becausethe assumption of the regression analysis, thatthe independent variable, in this case the envi-ronmental mean, is measured "without error,could not be met (SPRENT 1969). This biasdepends on the number of genotypes testedand on the ratio of environmental variance toerror variance (SPRENT 1969). In well-designedexperiments (e.g. a large number of replica-tions and/or contrasting environmental condi-tions) this ratio will be large and bias will bestnall (HARDWICK and WOOD 1972, WRICHT

BECKER and LEON
1976). Furthermore, because with replicatedtrials error variance is estimated from the ex-periment, a correction is possible using T.M'S(1971) a; and Ji;, which can be regarded as aspecial form of bi and sji, when the environ-mental index is assumed to be random (LIN ex.al. 1986).
3. A frequent experience in the analysis ofreplicated experiments is that error variance arenot homogeneous between sites (SKR0PPA1984). However, applying weighted regressionanalysis with reciprocals of the error variancesas vi^eights, the absolute values of the regres-sion coefficients did not change nor did theirerrors (HUHN 1980, SKR0PPA 1984, LEON 1985).
Reviewing these statistical considerations,SK]10PP.>\ (1984) stated that, with Iarg6.numbersof genotypes, environments and replications,with no outliers of either genotypic or envi-ronmental effects and with homogeneous errorvariances of genot}'pes, as usually assumed inanalysis of variance, the regression approachshould be statistically valid.
Testing significance in the regression ap-proach depends upon even more assumptionsthan is usual in the analysis of variance (FREE-MAN 1973). FREEMAN (1973) recommends thatthe significance test of regression coefficientsshould be based on individual error termsrather than on pooled deviations because ofpossible heterogeneity of deviation meansquares. In an analysis of variance the GE-interactions 'Can be subdivided into a part duet'Q heterogeneity of b; and a remainder due topoO'led d,evi,ations from regressiO'n (FINLAY and
WILKINSON 1963, PERKINS and JINKS 1968). H
heterogeneity of bj does not reach significance,the usefulness of the regression approach forinterpretation of the data is doubtful. Forfurther significance tests see e.g. EBERHART andRUSSEEE (1966), FREEMAN and PERKINS (1971)
and SMUKEA (1972 a, b).
HANSON (1970) proposed a stability statisticwhich is founded on the regression approach.This measure was termed as 'genot}'pic stabili-ty' (Df), because it includes that part of thevariance of environmental effects which couldbe reduced by breeding and selection (UTZ1972). Assuming that the differences betweenregression coefficients are completely amen-able to breeding procedures, the minimum ob-served b (bmin) is used to define a stable geno-type which does not deviate from the straightline (Y = u + h„,,„e, + g;).
Therefore, the 'genotypic stability' statisticis expressed as follows:genotypic stability:
Df = 2 (X,, - X, - b j X,,, + h^,, XJ'
Environmental variance, ecovalence, and de-viation mean squares can be considered as spe-cial 'Cases of 'genotypic stability'. If b jn = 0,Df/(E-r) measures environmental v,ariance, ifbn,,n = 1, Df m,easures ecovalence and if b jp =bi, than Df/(E-2) measures deviation meansqu,ares (UTZ 1972). Although 'genotypic sta-bilitv-' was recommended by several authors, l:has not been widely used and has not beensubjected to further examination of its statisti-cal characteristics.
4-
3-
2-
1-
TYPE 1
4-
3-
2-
TYPE2 TYPE 3
E, EJ E, EJ E, EJ
Fig. 3. Graphical representation of some of HALDANE'S GE-interactions types for the genotypes Aand B and the environments E, and E2 (adapted from ALLARD and BRADSHAW 1964)
Stability Analysis in Plant Breeding
3.2.3 Nonparametric methods
Using parametric stability statistics and theirsignificance tests, assumptions have to be madeabout distribution and variance homogeneity.Nonparametric stability measures, however,are distribution-free and are not affected byoutliers as are parametric estimates (NASSARand HUHN 1987). Nonparametric stabilitystatistics may be based upon ranks rather thanon measured X;,-values.
HALDANE (1946) used rank orders to classifyGE-mteraction types. Figure 3 shows threeout of SIX possible interaction types for twoenvironments (Ej and E2) and two genotj'pes(A and B). Since HALDANE (1946) ranks themeasured Xjj-values according to their amountregardless of whether they belong to differentenvironments or to various genotypes, thenumber of possible interaction types will bevery large when many genotypes and manyenvironments have to be considered. Thisproblem is reduced when, for each environ-ment, the genotypic rank orders are estab-lished separately and so differences or variabil-ity of ranks of genotype i may be applied toquantify phenotypic stability of the respectivegenotype. However as shown in Figure 3 onlyHaldane's interaction type 3 results in a chang-ing of genotypic rank orders. Type 1 does notpossess GE-mteraction effects as defined bythe biometrical model and consequently rankorders are not affected. Despite existing GE-interaction effects, no difference in rank orders
occurs for type 2. This fact has to be taken intoaccount if using rank orders to quantify stabili-ty. Therefore, as much as possible of the GE-interaction effects have to be considered whenapplying nonparametric stability statisticsbased on ranks.
Usually rank orders of each environment aredetermined by Xjj-values, which may be ex-pressed as follo'ws:
X,, = u + e, + g;+ (ge)i,; + E,,
Because fi and e, are constant for environment), these ranks (r'jj) are based O'n:
gi + (ge)ii + h,
Phenotypic stability, however, should bemeasured independently of genotypic effects,so that ranks hzve to be based on corrected Xjj-values: (X,j - X,.) (NASSAR and HUHN 1987).
Therefore these ranks (rjj) depend on GE-in-teraction and on error effects only:
(ge)., + Eii
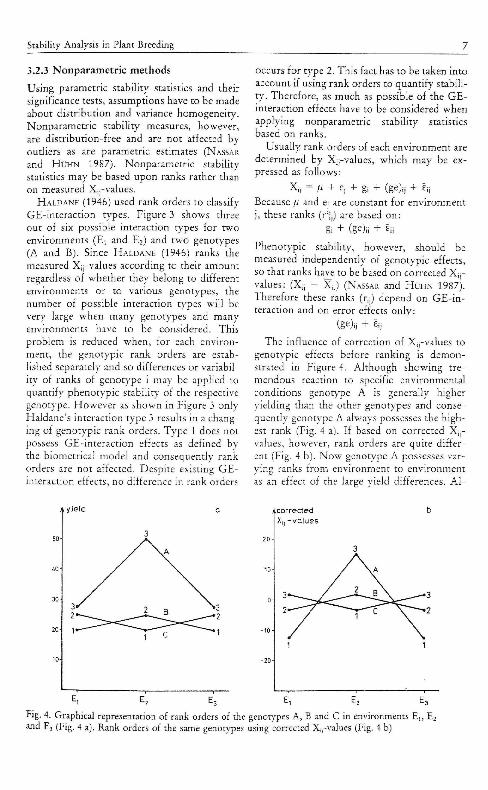
The influence of correction of X,,-values togenotypic effects before ranking is demon-strated in Figure 4. Although showing tre-mendous reaction to specific environmentalconditions genotype A is generally higheryielding than the other genotypes and conse-quently genotype A always possesses the high-est rank (Fig. 4 a). If based on corrected X;,-values, however, rank orders are quite differ-ent (Fig. 4 b). Now genotype A possesses var-ying ranks from environment to environmentas an effect of the large yield differences. AI-
yjeld
20-
10-
0-
10-
20-
corrected
/1
3A.
/ \ *
2 B \
b
._..-.—3
\ " «'2
\1
Fig. 4. Graphical representation of rank orders of the genotypes A, B and C in environments E,,and EJ (Fig. 4 a). Rank orders of the same genot)'pes using corrected X;;-values (Fig. 4 b)

8
Table 1. Sutnmary of univariate s
Statistics
tability statisi
Symbol
BECKER and LEON
Stabilityconcept
Parametric:
Environmental varianceEcovalenceRegression coefficientDeviation mean squaresCoefficient of determination
Nonparametric:
Mean rank differenceVariance of ratiks
W,
S;4
Static
dynamicstatic/dynamicdynamicdvnamic
dynamicdvnamic
though genotypes B and C are equal for theiryield differences among environments, geno-type B IS regarded as stable concerning rankdifferences, while genotype C is classified assimilar to genotype A. This is a result of thedependence of ranking on the range of geno-types. Because the yield of genotype C de-creases in environment £7 while the yields ofgenotypes A and B increase, rank differencesof genotype C increase compared to genoti'peB. This pattern is in accordance with the higherGE-interaction of genotype C.
Nonparametric stabihtj' statistics based onrank orders (of corrected Xjj-values) have beenproposed by HUHN (1979) and N.WS.' R andHUHN (1987). As described above, rank ordersof genotypes are established for each environ-ment separately. Then the var^'ing ranks ofgenotype i are used to estimate the phenotypicstabilit)' of genotypes. HUHN (1979) and N.«-s. R and HUHN (1987) proposed two stability
mean rank difference: S,l = —, ! r., - rf \
E (E-l)/
variance of ranks:
here
S,4 =E-1
;, is the rank of genotype i in environ-ment j , based on corrected X.j-values. Sjl issimilar to the Gmi-measure of variability butbased on ranks (for further information on theGini-measure of variability see e.g. KENDALLand STL'ART 1958). Significance tests for bothstatistics are given by NASSAR and HUHN(1987).
HUHN (1979) and THOMSON and CUNNING-
H.\M (1979) calculated variance of ranks basedon uncorrected X,j-values. However these pa-rameters may show significant differences instabilirv' between genotypes when in fact thereis no GE-interaction (NASS.AR and HUHN 1987).Eurrhermore, nonlinear relationships betweenX;-values and Sjl* and S'|4'"'" (each based onuncorrected X;J-values) occur because the mag-nitude of rank differences of genotype 1 de-pends on the distribution of genotypes or, mother words, on the frequency of genotypes ofsimilar Xi-value (EEON 1985). Using correctedX,|-values for ranking purposes (NASSAR andHUHN 1987) causes these nonlinear relation-ships to disappear (LEON 1985).
With ranks based on corrected X,;-values asa prerequisite, the nonparametric stabilitystatistics S,l and Si4 permit a meaningful in-terpretation of the results. These stability esti-mates are distribution-free and no assumptionon the distribution of the X,,-values is neces-sary. Eurther advantages are that the non-parametric stabihty statistics are expected to beless sensitive to errors of measurements thanparametric estimates and that the addition ordeletion of one or of a few observations is notlikely to cause great variation in the estimate aswould be the case for parametric stabihtystatistics (NASSAR and HUHN 1987). Loss ofinformation, however, is associated with theapplication of rank orders to estimate pheno-typic stability because some minor GE-inter-action effects do not result in varied rank or-ders, even if ranks are based on corrected X,j-values.

Stability Analysis in Plant Breeding
3.3 Relationship between different methods
Table 1 gives an O'verview of the most com-monly-used univariate stability parameters andtheir underlying stability concepts. Only rela-tionships among these particular measures willbe discussed; for information concerning otherparameters see LIN et al. {1986).
The relationships hetween si,, Wi, b,, and sj;have been pointed out hy SCHNELL (1967) andWRICKE and WEBER (1980):
environmental variance:
+ (b. - I)- I. e] + X dy/(E-l)
ecovalence: W, = (b, - 1)^ Z e] + Z df,
where 2 e = Z (X,, - X_.)': sum of squares ofenvironmentaleffects
and Z df, : sum of squareddeviations fromregression, whichIS just another ex-pression of s;ii.
The coefficient of determination (rf), proposedby PiNTHUS (1973) instead of Sji, is related tothe statistics of the regression approach as fol-lows :
coefficient ofdetermination: rf =
But rank orders of rf and Z df, are equal orsimilar, respectively, only if all regressioncoefficients are equal (bi = . . . = b, = . . . =be) or the variance of bj-values is small. Forfurther information on the biometncal rela-tionships between various parametric stabilitystatistics see also UTZ (1972).
The nonparametnc statistics Sil and Si4 dif-fer in weighting differences of ranks, becauseS;4 may be expressed as follows:variance of ranks:
S,4 =E-1 2E (E-l)/2
which therefore equals S;!, except that differ-ences are squared by S;4 instead of using abso-lute differences as with Sil.
Besides theoretical relationships between thestability statistics empirical correlations are al-
so useful for quantifying the influence of eachterm of the equation. Table 2 gives rank corre-lation coefficients among parametric stabilitystatistics (for further information about thematerial see BECKER 1981b).
For ranking purposes, high r^-values andlow values of s ,, Wj, b, and sji are regarded asbeing desired. High correlation coefficients ex-ist between b, and s ; and among the threeparameters sj^, W, and rf. The other correla-tions are small to moderate and not consistentfor the various crops. The magnitudes of thecorrelations presented in Table 1 correspondto results reported in the literature (for reviewsee e.g. WEBER and WRICKE 1987).
Because ecovalence may be partitioned intothe two components (b; - 1) 2 ef and Z d\and, because the first component representsthe covariance between GE-interaction effectsand environmental effects, the high correlationcoefficients among W, and Sji show that thiscovariance usually explains only a small part ofW,, as pointed out by WRICKE and WEBER
(1980).The strong correlation of W; and sg, to rf is
due to the relatively large variability of sj;compared to the variability of bf Z ef. How-ever, when the range of environments is ver}'limited and therefore variation of b; is high,correlations of W, and sj; tO' T\ are only mediumand correlation coefficients between rf and bjmay be moderate to high (V. siLj and MILAS1984). The high correlation between s i and b;IS similarly explained. Because Z ej is constantfor all genoti'pes, differences in s ; dependchiefly on differences in the term 2 (b; — 1)Z ef, a linear function of bj (BECKER 1981 b).
The nonparametric stability statistics Sil andS;4 are strongly correlated (r > 0.95) (LEON1985). Exact biometrical relationships of thesetwo nonparametnc stabilit}' measure to para-metric stability statistics like s i, W;, b,, and S3ihave not so far been elaborated. Tbereforeempirical correlations give important informa-tion about these relationships. Table 3 showsrank correlation coefficients between these twogrO'ups of stability statistics for 22 successiveyears of winter wheat trials (for further infor-mation on material see LEON 1985).
Obviously W; and sJ are strongly correlatedwith both nonparametric statistics and theserelationships are much stronger than the re-spective relationships with s ;. Correlation

10
Table 2. Rank correlation coefficients among stability
s;,
statistics for
W,
grain yii:ld (BECKER
4.
BECKER and
1981b)
rf
L£ON
w,
maizebarleyoats
maizebarleyoats
maizebarleyoats
maize
barley
oats
0.99='*0.96 =-=-0.95='=^
- 0 . 0 5- 0 . 0 4
0.36= =
0.050.210.58= *
-oao0.030.4Q=^*
O.OC0.270 . 6 1 * *
0.94**0.94**0.96**
- 0 . 4 8 * *- 0 . 3 6 *
0.04
- 0 . 3 8 *- 0 . 0 8
0.33='-
0.83**0 . 9 1 * *0.90**
0.90**0.87**0.93**
*, *,, ** Significant at P = 0.10, 0.05, 0.01, respectively
coefficients between s ; and nonparametncstatistics are almost exactly as high as the cor-relation coefficient of sj; to W; or to Sj;. Thismay be explained by the fact that s ; detectseffects which are also measured by Wj and si,.
Therefore the nonparametric stability statis-tics measure stabilitv' accordmg tO' the dynamicconcept. The size of correlation coefficientsbetween W; and nonparametric stabilit)' statis-tics indicates that most but not all GE-mterac-tion effects result in different ranks of thecorrected X,,-values. Other comparisons be-tween nonparametnc and parametric stabilitystatistics, reported in the literature, are notcomparable because they do not take into ac-count the necessary correction of Xjrvalues.
3.4 Multivariate methods
In addition to univariate stability statistics,multivariate methods of analysing GE-mterac-tion have been proposed. The wide range ofmultivariate methods include multivariate
Table 3. Rank correlation coefficients between uni-Yariate parametric and nonparametric stability statis-tics for -winter wheat trials conducted from 1961 to1983
S,l
S,4
(L£ON 1985)
si
0.23=-0.24='
W.
0.84*
0.84*
Sd,
0.80='0.82*
Significantly different from zero at P = 0.05
analysis of variance (MANOVA) (CALINSKI etal. 1987a, b), cluster analysis, principal com-ponent analysis, geometrical metbods,stochastic dominance and methods using exter-nal information on environments or genotypes(for review see EIN et al. 1986, WESTCOTT 1986,HuHN 1987a). Because of these recently pub-lished reviews we will only make some generalremarks on multivariate methods of analysingGE-interaction.
Considering each environment as a variableand genotypes as objects or replications, themultivariate techniques might be used to de-scribe the performance of genotypes. TheMANOVA-technique uses the methods de-scribed in chapter 3.2 in a multivariate manner(CALINSKI et al. 1987 a, b). The aim of variousclassification methods is to assign genotypesintO' qualitatively homogeneous stability sub-sets. Within subsets no significant GE-interac-tions occur, while differences among subsetsare due to GE-interactions. To achieve thesesubsets, many different dissimilarity measuresand clustering strategies have been proposed.At least 10 different dissimilarity measures andeight different clustering strategies exist andchoosing between them can result in differentcluster groups so that the acceptance or rejec-tion of any particular choice may be difficult tojustify (WESTCOTT 1986). Furthermore, in-terpretation of the results is not directly mterms of stability. However, if a well-knowncultivar is included in the test it can be used as a

Stability Analysis in Plant Breeding 11
paradigm for the other genotypes in the samesubset (LIN et al. 1986).
Applying principal component analysis, thevariables (here environments) are combined sothat the first new variable, the first principalcomponent, explains the maximum of var-iance. The second princtpal component ex-plains the maximum of remaining variance andso on. The principal components are inde-pendent of eacii other. Genotypes are charac-terized by their principal component values.However, the chief difficulty with this ap-proach is the interpretation of resulting princi-pal components, which may not bear any obvi-ous relationship to environmental conditions(WESTCOTT 1986).
In so-called geometrical methods, the basicaim is to represent each genotype by a point msome Euclidean space so that genotypes whichare similar to one other are represented bypoints which are close together. The config-uration of points is then investigated in anattemp't to detect any underlying structure(WESTCOTT 1986). These geometrical methods,like principal coordinates analysis, non-metricmultidimensional scaling, biplot method andcorrespondence analysis are reiatively unex-plored (for hterature on examples see WEST.COTT 1986). The same is true for the method ofusing stochastic dominance to analyse GE-interactions.
Taking account of recorded informationabout the environments, such as data on forexample, rainfall, temperature or global radia-tion at different stages throughout the growthseason, might partly explain GE-interactioneffects. However, environmental measure-ments are very seldom available and often sev-eral of them are lntercorrelated. As an exampleof using recorded information on environ-ments, AAST\'EIT and MARTENS (1986) explaineda considerable part of GE-interaction by ap-plying the relatively new partial least squares(PLS) regression method, which may beunderstood as separate principal componentanalysis of several y-variables (Y) and of sever-al x-vanables (X) at the same time, with com-ponent estimates providing prediction of Yfrom X. Eor an assessment of the PLS regres-sion method and other relatively unexploredmultivariate methods for analysing GE-in-teractions, more research is needed.
4. Estimating stability in breeding mate-rial
After presenting methods of stability analysis,mainly from a biometrical point of view, wewdl now discuss the problems which arisewhen these methods are applied to the estima-tion of stability m a plant breeding pro-gramme.
4.1 Appropriate biometricai method
By far the most frequently used method is thatinvolving regressions. Opinions differ aboutwhich value of b; should be achieved. FINEAYand WiLKiNSON (1963) defined a genotype withb, = 0 as stable; this definition is according tothe static concept but it "would be preferable touse the environmental variance instead, whichis highly correlated with b: but avoids thestatistical problems connected with the regres-sion approach. EBERHART and RUSSELL (1966),on the contrary, defined a genotype with b, =1 to be stable; this definition is in accordancewith the dynamic concept, but it would bepreferable to use the ecovalence instead whichcombines bi and si, into one parameter.
In conclusion, the regression approach is oflittle use if b; is included in the definition of"stability". Consequently, most authors con-sider b; not as a measure of stability but asadditional information on the average responseof a genotype to advantageousness of environ-mental conditions. This approach is schemati-cally presented in Figure 5.
Usually only a small part of the GE-mterac-tions can be explained by a heterogeneity ofregression lines. Eor this reason the usefulnessof the regression approach is sometimes ques-tioned. Tlie small numerical contribution of
High Yield Stability
Adapted 10 Adapted CoLaw yi&l^iing — — High YralzlmgEnioronmenfs I En^ironmnnts
Low Yield Stab l i i t y
I
Fig. 5. Interpretation of the parameters b, and Sj; ofthe regression approach (adapted from HAUFE andGEIDEE 1978)

12 BECKER and LEON
differences in b; to the total interactions isbecause in a normal series of trials most envi-ronments have about average yield levels andmost genotypes have values of b; close to 1.But, nevertheless, if there is interest in the fewexceptional genotypes which are speciallyadapted to extremely low- or high-yieldmgenvironments, the regression approach may bevery useful. Even small differences in the valueof bi become important if environments areused m which yield levels are far below orabove the average. Such environments are rareof course and often there will be no commer-cial interest in developing special varieties forsuch environments.
In summar}', there is little reason to rejectthe popular regression approach and we don'tsee any method to estimate yield stabilitywhich would generally be preferable. The en-vironmental variance does not provide infor-mation of interest for most plant breedingpurposes. The information given by the ecova-lence IS rather similar to S2, but the regressionapproach gives additional information on bj,which at least might sometimes be of interest.However, as already emphasized, the regres-sion analysis should only be used if there isinterest in estimating and interpreting the valueof b,, otherwise the ecovalence (or, as an equi-valent, the stability variance) is preferable as amore direct measure of GE-interactions. Thevariance of ranks is of interest in situationswhere the assumptions of parametric statisticsare not fulfilled and certainly more experiencewith nonparametric methods would be desir-able.
In recent years there has been an increasingamount of discussion of multivariate analyses.Nevertheless, examples for the application ofthese methods to normal breeding material arevery rare and most published experiments arenothing but numerical examples with fewselected genotypes. Thus, at present, none ofthe various multivariate methods can be re-commended as a routine procedure for breed-ing programmes.
A problem which affects any method ofanalysing GE-interactions is the use of trans-formations. FINEAY and WILKINSON (1963) andmany others used a logarithmic transformationof the original data. The argument of FINEAYand WILKINSON (1963) was that this achievedlinearity and homogeneity of errors. This is
statistical reasoning and it must still depend onthe specific set of data as to whether or not atransformation is necessary.
But there are fundamental grounds for theuse of logarithmic transformations irrespectiveof these statistical arguments. Imagine twogenotypes A and B, and m all environments Ahas exactly twice the yield of B. When compar-ing these two genotypes, A will show twice thevalue of bi and four times the value of varianceparameters hke sji, sj; and W;, though therelative reaction of both genotypes to environ-ments is identical. Using logarithmic transfor-mations will result in the same values of allparameters for the two genotypes. Usually inpopulation genetics it is not the absolute varia-bility but tbe relative or "intrinsic" variabilitywhich IS of interest, and thus it is common rouse logarithmic transformations or coefficientsof variation which have the same effect(LEWONTIN 1966). A logarithmic transforma-tion of data is of little consequence for practicalbreeding where only genoti'pes with similaryield level are compared. But at least some-times in breeding research logarithmic trans-formations should be considered; e.g. resultsfrom comp'arisons of inbred lines and hybridsare largely influenced by the scale of measure-ment, also the frequently-o^bserved positivecorrelation between b, and average yield level ispartly due to a scaling effect (BREESE and HILL1973).
Finally we need to refer to the main limita-tion of any stability analysis, that is the influ-ence of the material on the results. AccordingTO the dynamic concept, stability is defined lorelation to the varieties under test and thisselection of genotypes may be arbitrary. Theuse of check varieties to assess the environ-mental index IS no real solution, for then theresults depend on the choice of check varieties.
In normal breeding programmes the influ-ence of the material used on the results ob-tained is not a serious problem because newvarieties are compared with each other or withchecks, and the choice of material is obvious.But, nevertheless, one should be careful ininterpreting large deviations from regression as"instability", for it simply means that a geno-type reacts quite differently to environmentswhen compared with the majority of geno-types. Thus for example a very early maizevariety will probably be "unstable" if tested

Stability Analysis in Plant Breeding 13
together with many late varieties, and so will asix-row barley if indoded m a trial with two-row types.
It should be noticed that the influence ofmaterial on stabihty estimates is not character-istie of the regression approach but arisesequally with any other method for the analysisof GE-interactions, including multivariatemethods. The basic cause for this problem liesin the nature of an interaction. If we considerone only genotype, no interaction with envi-ronments exists; an interaction can only arisefrom comparing at least two genotypes, andthus GE-interactions m principle refer tospecific material.
LIN et al. (1986) doubt the value of theregressiO'n approach because the environmentalinde.x is not measurable prior to the experimentand they favour the use of external climaticfactors to develop a prediction model. From amore practical point of view, however, it isusually well known m advance if a location willgive an especially low or high yield level.
The use of external variables to describe anenvironment has man,}' advantages. Some ex-amples exist for the successful use of climaticvariables; e.g. differences between maize geno-types m East Africa could be described bylinear regressions on the altitude of the loca-tion (EBERH.^RT et al. 1973), or varieties ofdurum wheat m Cyprus were characterized bya regression on annual rainfall (H.'iDjiCHRiS-TODOUl.ou and K.-\Ri 1984). But in most situa-tions, the yield level of an environment is notdetermined by just a few limiting environ-mental factors. Also, reactions to growth fac-tors are usually more likely to be optimalfunctions than linear relations (KNIGHT 1970),and again the optimum depends upon on thematerial used. Thus, the choice of material wilialways influence an analysis of GE-interac-tions and has to be considered when discussingthe results.
4.2 Number and choice of environments
A major problem in selecting for yield stabilityIS the poor repeatability of the statistical para-meters estimated. Generally, genotypes rankdifferently in different years and results calcu-lated from different sets of locations or fromdifferent years are only slightly correlated (forliterature see WEBER and WRICKE 1987, LEON
and BECKER 1988). If stability is estimated fromseveral locations within one year only, geno-type-year interactions are not taken into ac-count and hence a significant difference be-tween two genotypes based on one-year resultswill not necessarily occur again in the follow-ing years. In trials with large genotype-yearinteractions LEON and BECKER (1988) observeda very low repeatability of single-year esti-mates of stability, even if the number of loca-tions was 15 to 20.
In experiments performed in more than oneyear it would be desirable to distinguish be-tween stability considering years and stabilityconsidering locations m order to get a morecomplete characterization of a genotype (B.»iR-RAF-i et al 1981). But most experiments coveronly two or three years and thus it is usually-assumed that years and locations can replaceone another. In a number of experiments thereactions to years and to locations have beenobserved to be similar (PEDERSEN and SEIF 1975,BARR.'IH et al. 1981, HOPPE 1982, A. STA'EIT and
AA.STVEIT 1984), but this correspondence doesnot always exist (WERMKE 1966).
In multi-year trials it is either possible tocalculate a regression on the locations averagedovev the various years (e.g. Y.ATES and GOGH-RAN 1938), or to consider each year-locationcombination as a "macroenvironment" (e.g.EBERH.ART and RUSSEEE 1966). Regressions onlocations and regressions on macroenviron-ments usually lead to similar rankings of geno-types; an interpretation of b, is often easier ifcalculated from a regression on locations but aregression on macroenvironments is morepowerful in detecting differences m sj: and thuswill be preferable in most cases (BECKER 1984).
For selection purposes not only is the statis-tical accuracy ol the estimated parameters ofimportance but also the genetic variabilityavailable. This is taken into considerationwhen applying the concept of heritability. Theheritability of stabihty measures is rather low;in performance trials Vi'ith wheat and barley at12 locations in Germany, F.R., hentability ofb; and S3j was between 0.11 and 0.27 if based onone-year results and between 0.27 and 0.52 ifbased on three-year results (BECKER 1987).
In order to increase the number of environ-ments available artifical "environments" areoften created by the variation of cultural prac-tices like sowing date, fertilization, irrigation

14 BECKER and LEON
cHiOlo
Land r aces ofself -poUinaiors
Line mixtures
Pure lines
Populations
Double crosses
Three-way crosses
Single crossesClones
Fig. 6. Schematical represen-tation of the genetic structureof various t)'pes of varieties(adapted from BECKER 1983)
homozygou sHeterozygosity
heterozygous
etc. Surprisingly enough, hardly any experi-mental evidence exists to justify this proce-dure. The only direct comparison between in-teractions with macroenvironments and in-teractions with cultural practices was pub-lished for carrots (DOWKER et al. 1978). Here,both t}'pes of environmental influence resultedin similar rankings of genotypes; this agreeswith more indirect indications from wheat(D.« and ]A]N 1971, BAINS 1976), but partlydisagrees with results in tobacco (PERKINS andJINKS 1971).
In conclusion, it will usually be impossibleto calculate useful stabihty measures from afew environments only. Locations, years andcultural practices will sometimes result m simi-lar reactions of a genotype and thus can replaceeach other, btit this depends upon the materialand the geographic region and should not gen-erally be taken for granted.
5. Selection for improved stability
Yield stability, like any other trait, can beinfluenced at various phases during the breed-ing process: the choice of the type of variety isof importance, the choice of parents has aninfluence and, finally, during development of
the variety direct and indirect selection can beapplied.
5.1 Type of variety
Types ot varieties differ m,ainly in two re-spects: the degree of heterozygosity of theplants and the amount of genetic heterogeneitywithin the variety. All possible types of varie-ties can be characterized by these two dimen-sions as shown in a diagram propO'Sed bySCHNELE (SCHNEEE 1982, ScHNEEE and Bl CKER
1986), see Figure 6.Sometimes at least different types of varieties
are possible and yield stability is one of thefeatures to be considered if discussing, forexample, the choice between lines and hybridsin self-pollinating species or between popula-tions and hybrids in cross-pollinating crops.
It is to be expected that the genetic structureof a population will influence the size of GE-interactions. It is a classical concept of popula-tion genetics that heterozygous genotypes areless susceptible to environmental influencesthan homozygous ones, and that heterogene-ous populations are better buffered thanhomogeneous ones (LERNER 1954, LEWONTIN1957); the application of this concept to plant

Stability Analysis in Plant Breeding 15
Table 4. Relative sizes of GE-jnteraction mean squares and respective coefficients of variation (CV) forhomozygous lines (Hom) and their heterozygous single crosses (Het)
Crcip
Maize
RyeSorghum
Wheat
Mean squareHom
100100100IQQ100100100
Het
66556677699427
Hom
100100100100100100100
cvHet
41247274668446
Reference
RowE and ANDREW 1964WAHLE 1977
MAJISU and DOGGETT 1972P.1TANOTHAI and ATKFNS 1974RAO and RAO 1978WrDNER and LEBSOCK 1973JATASRA and PARODA 1981
breeding was discussed b)- AU.ARD and BRAD-SH.w (1964).
Table 4 summarizes experiments where thesize of GE-interactions was estimated inhomozygous lines and in their heterozygoussingle crosses. The table gives the mean squaresof GE-interactions and also the respectivecoefficients of variation whieh take into ac-count the very different yield level of lines andhybrids.
In all experiinents, the homoz)'gous geno-types show larger GE-interactions than theheterozygous ones. This lower stability ofhomozygous lines is especially pronounced inmaize and rye and reflects the high phenotypicinstability caused by inbreeding in naturallyoutbreeding crops. But the same trend can alsobe observed in sorghum and wheat and couldbe an additional argument in favour of hybridvarieties in self-pollinated crops.
To investigate the effect of geneticheterogeneity, two main types of experimentshave been performed. One was to compare the
mixture of homozygous lines with the compo-nents grown in pure stands. Such mixtureshave smaller s3,-values and consequently ahigher yield stability than their components aswas observed m several crops, e.g. barley(CLAY and ALLARD 1969), oats (QUALSET and
GR.^NGER 1970, PFAHLER and LINSKENS 1979),
sorghum (KOFOID et al. 1978), cotton (EEASTERand TuRCOTTE 1973), soy beans (SCHUTZ andBRIM 1971, WALKER and EEHR 1978), chickpeas
(SINGH and BAINS 1984), and rapeseed (LEON
and DiEPENBROCK 1987). Theoretically, theyield stability of a mixture can be superior toany of the components grown in pure stands(MARSHALI and BROWN 1973).
A second type of experiment to evaluate theinfluence of genetic heterogeneity was thecomparison of different hybrid types. Whenanalysing yield trials in maize, SPR.^GUE andEEDERER (1951) had already obser^'ed lowerGE-interactions in double crosses than in sin-gle crosses. Experiments with a balanced struc-ture of the material are listed m Table 5. All
Table 5. Relative sjzes of GE-interaction mean squares m single crosses (2W), three-way crosses (3W) anddouble crosses (4\V)
Crop
Maize
SorghumRye
2W
100100100
roo100100
—
GE-interactions3W
44
70
9780
100
4«'
6S4557
76
References
EBERHART et al. 1964
EBERH.'IRT and RUSSELL 1969
WE.ATHERSPOON 1970
SCHNELL and BECKER 1986*
GEIGER et al. 1987
P.\TANOTHAi and ATKINS 1974BECKER et al. 1982
" Average ecovalence instead of mean squares

16 BECKER and LEON
500
400
300
200-
100-
Genotype-environmentinteractions ( CV,rel.)
Fig. 7. Influence of genetic structure onthe relative amounts of GE-interactionsin sorghum (REiCH and ATKINS 1970),rapeseed (LEON 1987), and maize(SCHNELL and BECKER 1986)
Pure stand Blended Pure stand Blended
Homozygous lines Single crosses
results indieate an advantage of the heterogene-ous three-"way and double crosses when com-pared to the homogeneous single crosses.
Three experiments are published where allfour possible co'inbinations of homo- andheterozygosity with homo- and heterogeneityare compared. The results are summarizedgraphically in Figure 7. In all studies theheterogeneous mixture of hybrids showed thesmallest GE-interactions. In sorghum andrapeseed the mixtures of homozygous hneswere more stable than the single crosses in purestands, demonstrating a prevailing importanceof heterogeneity. In maize, on the other hand,keterozygosit}' is a prerequisite for sufficientyield stability, since inbred lines are extremelysusceptible to environmental influences, evenif grown in mixtures.
In Tables 4 and 5 and in Figure 7 only theGE-interactions have been considered. Insome of the experiments mentioned, these in-teractions were partitioned according to theregression approach. Theoretically, the regres-sion coefficient of a heterogeneous mixtureshould be approximately similar to the averageof the regression coefficients of the compo-nents (FlO'HN 1987 b), and with heterozygosityone might expect lower bj-values with hybridsthan with inbred lines, for hybrids usually
suffer less from extreme!)- unfavourable stressenvironments. Experimental data on the influ-ence of heterogeneity and heteroz)'gosity on b,give no clear picture, however, and will not bediscussed in detail here.
The results presented so far favour a type ofvariety which is as heterozygous and asheterogeneous as possible, but there are twoaspects which modify these conclusions. First,the yield level has alsO' to be considered, andthe chances of finding a high-yieldmghomogeneous genotype are greater than offinding a high-yielding heterogeneous mixture,unless very strong positive competition effectsare assumed.
Moreover, all the results presented so farrefer to the average stability of a type of varietybut genetic variability also exists within eachgroup. As for any other character, the geno-typic variance for the trait "yield stabiLty" isexpected to be larger among homogeneous va-rieties than among heterogeneous ones, andlarger between inbred lines than between hy-brids. Thus, O'ver thirty years ago. JINKS andMATHER (1955) emphasized the possibility thathomozygous genotypes may exist which are asstable as a heterozygous and heterogeneouspopulation. But, as discussed m chapter 4.2,stability is a trait with a very low heritability.

Stability Analysis in Plant Breeding 17
40-
30-
20-
10-
0-
Porentswith b-<1
r
—
—
U- 40-
30-
20-
10-
o-
r -
Porentswith b| >1
n .0 1 Pn,,L 2 b,
Fig, 8. Distribution of brvalues m F4 progenies oftv.'o crosses in wheat (adapted from B. lNS 1976)
so even it pure lines with good stability ma\'exist it will be extremely difficult to identifythem. If feasible, the choice of a heterogeneousand heterozygous type of variety is a simpleand reliable way to sxoid the risk of developingcultivars with poor yield stability.
5.2 Choice of parents
If the type of variety is determined, the nextstep m the breeding procedure is to choose theparental material. A lot of data from yield trialsof the parents will often be available, and thusinformation on stability could be consideredfor the choice of parents.
In several experiments the inheritance of b;and sg; was investigated. B.41NS (1976) crossedwheat lines which were known to differ wideh'in their value.s of bj and Sj; and developedunseiected F4-lines. Yield and stability of par-ents and of F4-Iines were estimated in inde-pendent trials. The b; values of the progeniesclearly reflected the parental values, as shownfor two crosses as examples in Figure 8. For S3,some associations between parents and prog-enies were observed too, but the relationshipswere less clear.
Similar conclusions were drawn by FINLAY(1964), who analysed the values of bj in an F2-diallel m barley and observed large and signifi-cant estimates for general combining abiiit}'(gca) and nonsignificant values for specificcombining ability (sea). This is in agreementwith several studies with maize (EBERHART andRUSSELL 1969, DHILLON and SINGH 1977,
GEIGER et al. 1987), where the gca variance ofbi was always much more important than thesea variance, whereas for sji the gca neverreached significance. Consequently, it waspossible to predict the b, of three-way crossesvery reliably from the gca of the respectivesingle crosses (r = 0.84*''"), whereas a corre-sponding prediction of sj, was not possible(GEIGER et al. 1987). Also it was observed inbromegrass, that the contribution of gca-ef-fects was more important for b, than for sj,(TAN and TAN 1980).
In conclusion, if \'arieties with exceptionallyhigh or low values of b| are desired, the choiceot parents could be an effective way of reach-ing this objective. The results with sji are lessconvincing and more experiments are neces-sary. In many of the studies mentioned onlyfew environments were included and the poorcorrelation between parents and progenies forsg, may be due to unrehable estimates of sg;. Inthis connection also, multivariate methodsshould be investigated further, for it could be apromising strateg}' to combine parents withdiverse patterns of GF-mteractions to obtainprogenies with superior yield stability.
5.3 Direct and indirect selection
The response to selection for improved yieldstability will be limited by the rather lowheritability of this trait (cf. 4.2). Thus duringthe development of new varieties selection forstability will hardl)- be efficient and estima-tions of biometrical measures for stability willbe of little use. For the same reasons the use-fulness of a "cross prediction" of stability fromearly generations (POWELL et al. 1986) is some-what doubtful. In the final comparative trialsfor licensing, however, a stability measure canprovide valuable additional information on thevarieties under test.
If varieties with either a low or a high b, areobtained, it might sometimes be possible tobegin selection in early generations. In such
Ham Breeding, Vol. 101 (1)

18 BECKER and LEON
situations it would be advisable to choose siteswith very different yield levels, for with agiven number of environments the statisticalerror of bj is inversely proportional to thevariance between the environments.
Due to the difficulties with direct selectionfor yield stability,, the possibilities of indirectselection gain importance. Such indirect selec-tion is usually practised when resistanceagainst diseases, lodging etc. is considered, butfew information is available on the effects ofplant morphology or yield components onyield stability.
A ver}' striking example for a relationshipbetween plant architecture and yield stabilitywas observ-ed in maize, where two-ear typeswere found to have lower values for b, and forsa, than one-ear types (RUSSELL and EBERHART1968, CROSS 1977). Under unfavourable condi-tions, plants of the one-ear type sometimesproduce no ear. Under similar stress condi-tions two-ear types are able to produce at leastone ear and hence tend to be superior to one-ear types in low-yield environments and havehigher yield stability.
Only a few similar examples can be found inthe literature. Maize genotypes with late matu-rity tend to be better adapted to high-yieldmgenvironments than early ones, i.e. they show ahigher bj (LOFFLER et al. 1986). In field beans,small-seeded types were observed to be morestable than large-seeded genotypes (DANTUMAet al. 1983). These results cannot be gener-alized for other crops of course, but theyshould encourage more work m this directionwhich would be very helpful for practicalbreediog.
In the examples mentioned so far the stabili-ty of yield was related to the mean perform-ance of other traits. Sometimes, not only meanvalues are considered but also stability analysesof secondary traits are performed. Instead ofreviewing the literature we just want to makesome critical comments on this type of investi-gation.
It is of course possible to calculate statisticalmeasures of GE-interactions for any quantita-tive trait. But one should remember that thesemethods were originally proposed for estimat-ing yield stability and a stereotyped transfer toother traits can be misleading. In a regressionanalysis for traits like yield components orplant height, the environmental index is ex-
pressed in the units of the secondary trait andcannot be interpreted as a measure "whether anenvironment is 'favourable' or 'unfavourable'.Consequently, for analysing the stability ofsuch traits, b, is of little value and either theecovalence should be chosen or a regressionshould be calculated on the environmental val-ues of yield instead of the secondary trait(HEINRICH et al. 1983).
Moreover, the importance of these types ofstudy is generally doubtful, for stability ofyield components is not necessarily a breedingaim. As pointed out by WESTERMAN and LAW-RENCE (1970) stability of a complex trait mayoften be the result O'f a flexibility of primarycharacters, and BAINS and GUPTA (1972) andSINGH and BAINS (1984) found no relationshipbetween stability of yield and stability of yieldcomponents.
Final!)', it should be remembered that fortraits like disease resistance or quality, theapplication of the dynamic concept of stabilityis not justified. For such traits the static eon-cept is preferable, tor example a "stable" bak-ing quahty or winter hardiness implies a con-stant performance of these traits not affectedby the environment instead of a reaction toenvironments exactly like the average of allgenotypes.
6. Phenotypic stability and performancelevel
Phenotypic stability is usually measured inde-pendently of the respective performance level.For an assessment of genotypes, however,both traits have to be considered and thereforemethods of simultaneous estimation of per-formance and phenotypic stability are requiredto select among genotypes.
Stability statistics which consider deviationsfrom a hypothetical, desired genotype (SEP.'IHI1974) or from the site highest yield [JENSEN1976) are almost perfectly associated with per-formance level and show httle or no correla-tions with stability statistics (LEON 1986) andtherefore cannot be recommended for a simul-taneous selection of yield level and phenotypicstability.
BARAH et al. (1981) adapted the decisiontheory under risk, which was developed byeconomists, to the problem of simultaneousselection for yield and yield stabihty. For theexample given by BARAH et al. (1981) nomarked difference exists between yield-based

Stability Analysis in Plant Breeding 19
Table 6.
S.3
Rank correlation coefficients
Yield
0.62''
0.88-
of S,3 and S
0.14
-0.04
i6 with yield and
W,
0.59''
0.43''
stability statistics
0.0.
b,
,21''
(LEON 1986)
si
0.58''0.42''
rankings and rankings derived from the com-bined measure.s.
But information of performance and stabili-ty may also be combined by applying a selec-tion index or using nonparametric statistics asproposed by HUHN (1979). As with the meanrank difference and variance of ranks thesenonparametric measures are based on ranks.When phenotypic stability has to be combinedwith performance level, the Xij-values mustnot be corrected for the genotypic effects be-fore ranking (these uncorrected ranks aretermed r';',), because information about traitlevel would be lost. HL'HN (1979) proposedtwo nonparametric statistics for the simultane-ous estimation of performance and stability,which are expressed as follows:
r';
I r^-, - I
These statistics measure stabihty in units ofthe mean rank of the i-th genotype. Using S|3the differences between rank and mean rankare weighted with themselves avoiding the pos-sibility that a lot ot smaller rank differencesmay lead to the same Si3-value as a few largerdifferences. Tests of significance have not yetbeen proposed.
Empirical correlations ot grain yield andstability statistics with Sj3 and Sj6 are given inTable 6 (for further information see LEON1986). Sj3 and S,6 show considerable correla-tion with grain )'ield and with the stabilitystatistics Wj and sj,. Sj6 is more strongly associ-ated with grain yield and less with the stabilitystatistics than S,3. Therefore LEON (I9S6)stated that the choice between the nonparamet-nc statistics Sj3 and Si6 can be used to givemore weight to either performance o.r stability,like a weight in an index.
7. General conclusions and future pros-pects
When considering the biometrical methodsavailable to estimate GE-interactions, the con-
clusions of HILL (1975) are still valid today:" . . . the linear regression technique will con-tinue to play an important part m furtheringour understanding of GE-mteractions because,despite Its imperfections, it does have the twinmerits of simplicity and biological relevance.. . . But, this technique is not the plant breed-er's panacea, nor is it intended as a substitutefor physiology . . . Like every other model itwill sometimes fail."
The various multivariate methods whichhave been proposed will often allow a moredetailed analysis of GE-interactions, but theywill not replace the regression approach. Thehigh complexity of these methods, which isusually regarded to be their mam advantage,paradoxically is the main obstacle to their usein plant breeding. Multivariate methods aretoo sophisticated to provide any simple meas-ure of )'ield stability which allows a ranking ofgenot}-pes.
The most severe limitation of the regressionapproach is the poor repeatability of b, and sj,and the large number of environments neededfor a reliable estimate, but no other method hasbeen proven to be superior m this respect. Acertain improvement may be expected in futureby the use of nonparametric methods, whichare less susceptible to outliers. But one shouldbe aware of the statistical fact that any estimateof a parameter for variability is less reliable byfar than the estimate of a mean value. Thus,selection for yield stability will alu^ays remaina difficult task.
Nevertheless, even with the presentmethodology, many important problems canbe tackled:, e.g. more information is desirableon the stability of hybrids vs. inbreds m self-pollination crops; for the optimal choice ofparents the possibility of considering stabilitymeasures, including multivariate anahrses ofGE-interaction, are worthy of notice; and,finally, the prospects of indirect selection foryield stability should be investigated morethoroughly. Perhaps tO' understand why onegenotype is more stable than another crop

20 BECKER and LEON
physiology could provide a most significantcontribution.
We thank Prof. F. \V. ScHNELL and Prof. H, H.GEIGER. Stuttgart-Hohenheim, and Prof. M.HUHN, Kiel, lor many stimulating discussions onstability analysis and Mrs. B. KETELSEN for assist-ance with preparing the drawings.
8. ReferencesAASTVEIT, A . H . , and K. AAST\.'EIT, 1984: Genetic
variation of developmetital stability in barley.Hereditas 101, 155—170.
, and H. MARTENS, 1986: ANOVA interac-tions interpreted by partial least squares regres-sions. Biometrics 42, 829—844.
ALLARD, R. W. , and A. D. BRADSHAW, 1964: Impli-
cations of genotype-environmental interactions inapplied plant breeding. Crop Sci. 4, 503—508.
BAINS, K, S., 1976: Parent dependent genotype Xenvironment interaction in crosses of spring ^ 'heat.Heredity 36, 163—171.
, and V. P. GUPTA, 1972: Stability of yield andyield cO'iriponems in bread wheat. Ind. j . Gen.Plant Breed. 32, 3 0 ^ 3 1 2 .
BAR,\H, B. C , H . P, Bns'SWANGER, B. S. RANA, and
N. G. P. !L\O, 1981: The use of risk aversion inplant breeding: concept and application. Euphytica30, 451—+58.
BECKER, H . C , 19Sla: Correlations among somestatistical measures of phenotypic stability. Euphy-tica 30, 835—S4C.
, 1981 b: Biometrical and empirical relationsbetween different concepts of phenotypic stability.In: G.\LL, JS, A. (ed.), Quantitative Genetics andBreeding Methods, 307—314. Versailles: I.N.R.A.
, 1983: Ziichtensche Moglichkeiten zur Ver-besserung der Ertragssicherhert. Vortr. Pflanzen-ziichtg. 3, 203—225.
, 1984: Analysis of genotype x environmentinteractions with partitioning of the environmentaleffects into effects of locations and years. Vortr.Pflanzenziichtg. 7, 209—218.
, 1987: Zur Heritabilitat statistischer Mai5-zahlen fiir die Ertragssicherheit. Vortr. Pflanzen-ziichtg. 12, 134—144.
, H. H. GEIGER, and K. MORGENSTERN, 1982:
Performance and phenotypic stability of differenthybrid types in winter r\'e. Crop Sci. 22, 340—344.
BREESE, E, L., 1969: The measurement and signifi-cance of genotype-environment interactions ingrasses. Heredity 24, 27—44.
, and J. HILL, 1973: Regression analysis ofinteractions between competing species. Heredity31, 181—200.
GALINSKI, T., S. CZAJKA, and Z. KACZMAREK,
1987a: A model for the analysi.s of a series ofexperiments repeated at several places over a periodof vears. I. Theory. Biuietyn Oceny Odmian XII,7—33.
, , and , 1987 b: A model for theanalysis ol a series of e.xpei'iments repeated at sever-al places over a period of years. IL Example. Biule-tvn Oceny Odmian XII, 35—71.
GLAY, R. E,, and R. W. ALL.ARD, 1969: A compari-
son of the performance of homogeneous andheterogeneous barley populations. Grop Sci. 9,407—412.
GOMSTOCK, R. E., and R. H. MOLL, 1963: Geno-ti'pe-environment interaction. In: HANSON, W.D,, and H. E. ROBINSON (eds.). Statistical Geneticsand Plant Breeding, 164—196, Washington: Nat.Acad, SCI. Nat. Res, Council Pubi, 9S2.
GROSS. H . Z,. 1977: interrelationship among yieldstability and I'ield components in early maize.Crop Sci. 17, 741—745.
DANTUMA, G., E. VON KITTLITZ, M. FRAUEN, and
D. A. BOND, 1983: Yield, yield stability and meas-urements of morphological and phenotypicalcharacters of faba bean {Viaa faba L.) varietiesgrown m a wide range of environments m WesternEurope. Z, Pflanzenziichtg. 90, 85—105.
D.\S, P. K., and H, K. JAIN, 1971: Studies on
adaptation in wheat, II. Uni-local testing forphenotypic stability and varietal adaptability. Ind,j . Gen. Plant Breed. 31, 77—85.
DHIILON, B. S., and J. SINGH, 1977: Estimation and
inheritance of stability parameters of grain Yield mmaize. J. .Agric. Sci. Camb. 88, 257—265,
DOR.ST, j . C , 1957: .adaptation. Euphytica fc,247—254.
DowKER, B, D,, J. G. JACXSON, and K. PHELPS,
1978: Variation studies in carrots as an aid tobreeding. VI. Genotype-environment interactionsin contrasting field environments. J, Hort, Sci. 53,131—137.
EBERHART, S. A., L. H. PENNY, and M, N, HARRI-
SON, 1973: Genotype by environment interactionsin maize in Eastern Africa. East Afric. Agric. Eor,J. 39, 61—71.
, and W. A, RusSELL, 1966: Stability parame-ters for comparing varieties. Grop Sci. 6, 36—40.
, and , 1969: Yield and stability for a 10-line diallel of single-cross and double-cross maizehybrids. Grop Sci. 9, 357—361,
— , , and L, H, PENNY, 1964: Double crosshybrid prediction in maize when epistasis is pre-sent. Crop Sci. 4, 363—366.
FEASTER, G. V,, and E, L, TuRCO-lTE. 1973: Yieldstability m doubled haploids of American primacotton. Crop Sci, 13, 232—233.
ElNL,\Y, K. W,, 1964: Adaptation — its measure-ment and significance in barley breeding. In: Barley

Stability Analysis in Plant Breeding 21
Genetics I, 351—359. ^X'agenmgenr Proc. 1st Int.Barley Genetics Symposium.
, and G. N. WILKINSON, 1963: The analysis ofadaptation in a piant breeding programme. Austr.]. Agric. Res. 14, 742—754.
FREEM.^N, G . H . , 1973: Statistical methods for theanalysis of genotype-environment interactions.Heredity J l , 339—354.
, and J. M. PERKINS, 1971: Environmental andgenotype-environmental components of variabili-ty. VIII. Relations between genotype.s grown indifferent environments and measures of these envi-ronments. Heredity 27, 15—23.
FRIPP, Y. J., and C. E. C.WEN, 1971: Genotype-
ent'ironmenta! interactions in Schizophyllum com-mune. I. Analysis and character. Heredity 27,393—407.
GEIGER, H . H . , A . E . MELCHINGER, und G. SEITZ,
1987: Vorhersage der phanotypischen Stabilitat 'onDreiweghybriden bei Mais. Vortr. Pflanzenzuchtg.12, 145—155.
HADJIGHRISTODOULOU, A . , and A. K.-VK!, 1984:
Karpasia, a high yielding new du,rum, wheat varietywith improved gram qualit)' characteristics. Techn.Bull. Agric. Res. Inst. 57, 1—7, Ministry of Agric.and Nat. Resources Nicosia, Cyprus.
HALDANE, | . B . S., 1946: The interaction of natureand nurture. Ann. Eugenics 13, 197—205.
HANSON:, W . D . , 1970: Genotypic .stability. Theor.Appl. Gen. 40, 2 2 ^ 2 3 1 .
HARDWICK, R . C , and J. T. W O O D , 1972: Regres-
sion methods for studymg genotype-environmentinteraction. Heredity 28, 209—222.
HAUFE, W . . und H. GEIDEL, 1978: Zur Beurteilung
der Ertragssicherheit von Sorten und Zuchtstam-men. 1. Definitionen, Stabilitatsparameter und de-ren Interpretationsmoghchkeiten. Z. Pflanzen-zuchtg. 80, 24—37.
HEINRICH, G . M . , C . A . FRANCJS, and J. I). EAS-
TON, 1983: Stability of grain sorghum yield compo-nents across diverse environments. Crop Sci. 23,209—212.
HILL, ]., 1975: Genotype-environment mteraction— a challenge for plant breeding, j . Agric. Sci.(Camb.) 85, 477—493.
HoppE, H.-D., 1982: Untersuchungen der Wechsel-wirkungen Sorten — Orte — Jahre am Beispiel vonHaferpriifungen in den Jahren 1973—1978. Tag.Ber., Akad. Eandwirtsch! Wiss. DDR, Berlin 203,,57—67.
H U H N , M . , 1979: Beitrage zur Erfassung derphanotypischen Stabilitat. i. Vorschlag einiger aufRanginformationen beruhender Stabditiitsparame-ter. EDV in Medizin und Biologie 10, 112—117.
, 1980: Beitrage zur Erfassung der phanotypi-schen Stabilitat. II. Modifikationen des Regres-sionsansatzes durch Verwendung gewogener Re-gressionsmodelle sowie Anderungen der Bezugs-basis. EDV in Medizin und Biologie 11, 113—120.
, 1987a: Quantitative Erfassung, der Ertrags-,sicherheit — aktueller Stand, offene Fragen undmogliche Entwicklungen. Vortr. Pflanzenziichtg.12,37—61.
, 1987b: Phenotypic stability of mixtures —Relations between the stability parameters of amixture and its components. Biom. J. 29,703—719.
JATASRA, D . S., and R. S. PARODA, 1981: Genotype-
environment interaction in segregating generationsof wheat, ind. J. Gen. Plant Breed. 41, 12—17.
JENSEN, N . F. , 1976: Floating checks for plantbreeding nurseries. Cereal Res. Comm. 4,285—295.
JINKS, J. L., and K. MATHER, 1955: Stability in
development of heterozygotes and homozygotes.Proc. Rov. Soc. Ser. B 143, 561—578.
KENDAIL, M . G. , and A. STUART, 195S: The Ad-
vanced Theory of Statistics. Vol. 1: DistributionTkeor}-. London: Charles Griffin and CompanyLimited.
KNIGHT, R. , 1970: The measurement and interpreta-tion of genotype-environment interactions. Euphy-tica 19,"225—235.
KOFOID, K. D., W. M. Ross, and R. F. MUMM,1978: Yield stability of sorghum random-matingpopulations. Crop Sci. 18,, 677—679.
LEON, J., 1985: Beitrage zur Erfassung der phan'oty-pischen Stabilitat unter besonderer Beriicksichti-g!jng unterschiedlicher Heterogenitats- undHeterozygotiegrade sowie einer zusammenfassen-den Beurteilung von Ertragshohe und Ertrags-sicherheit. Dissertation, Christian-Albrechts-Uni-versit,at Kiel.
, 1986: Methods of simultaneous estimation ofyield and yieid stability. In: Biometrics m PlantBreeding. Proc. 6th Meeting EUCARPIA SectionBiometrics in Plant Breeding, Birmingham, UK,299—308.
, 1987: Ertrag und Ertragssicherheit ver-schiedener ,Sortenstrukturen bei Winterraps. Vortr.Pdanzenzuchtg. 12, 62—75.
, and H. C. BECKER, 19SS: Repeatability ofsome statistical measures of phenotypic stability —Correlations between single year results and multi-vcar results. Plant Breeding 100, 137—142.
, and W. DiEPENEROCK, 1987: Yielding abilityof pure stands and equal proportion blends ofrapeseed {Brassiea napus L.) with double low quali-ty. J. Agron. Crop Sci. 159, 82—89.
LERNER, 1. M., 1954: Genetic Homeostasis. Edin-bourg,, London: Oliver & Boyd.
EEWONTIN, R. C , 1957: The adaptation of popula-tions to various environments. Cold Spring Harb.Symp. Quant. Eiol. 22, 395—408.
_ 1966: On the measurement of relative varia-bilitv. Svst. Zool. 15, 141—142.

22 BECKER and LEON
LIN, C . S,, M . R. BINNS, and- L. P. LEFKOVITCH,
1986: Stability analysis: Where do we stand? CropSci. 26,, 894—900. '
LOFFLER, C . M , M. T. S.AL.'\BERRY, and J. C.
MAGGIO, 1986: Stability and genetic improvementof maize yield in Argentina. Euphytica J5,4 4 9 ^ 5 8 .
MAJISU, B . N . , and H. DOGGETT, 1972: The yieldstabilit)' of sorghum varieties and hybrids in EastAfrican environments. East Afr. Agric. For. J. 38,179—192.
M.\RSHALL, D. R., and A. H. D. BROWN, 1973:Stability of perfortnance of mixtures and multilines.Euphytica 22, 405—412.
NASS.\R, R,., and M. HUHN, 1987: Studies on esti-mation of phenotypic stability: Test of significancefor nonparametric measures of phenotypic stabili-ty. Biometrics 43, 45—53.
PATANOTHAI, A. , and R. E. ATKINS, 1974: Yield
stabihty of single cross and three-way hybrids ofgram sorghum. Crop Sci. 14, 287—290.
PEDERSON, D . G., and E. SEJF, 1975: Compensating
for site and year effects in estimating the true yieldof wheat genotypes. Austr. J. Agric. Res. 26,7,83—790.
PERKINS, J. M., and J. L. JINKS,. 196S: Environ-
mental and genotype-environmental componentsof variability. III. Multiple lines and crosses.Heredity 23, 339—356.
, and , 1971: Specifity of the interaction ofgenotypes with contrasting environments. Heredi-ty 26, 463—474.
PFAHLER, P . L., and H. E. LiNSKENS, 1979: Yieldstabilit}' and population diversity in oats (Avenasp.). Theor. AppL Genet. 54, 1—5.
PiNTHUS, M. J., 1973: Estimate of genotypic value:A proposed method. Euphytica 22, 121—123.
PLAETED, R. I . , and L. C. PETERSON, 1959: Atechnique for evaluating the ability of selections toyield consistently in different locations or seasons,Amer. Potato]. 36, 381—385.
POWELL, W., P. D. S. CALIGARI, M. S. PHILLIPS,
and J. L. JLN'KS, 1986: The measurement and in-terpretation of genotype by environment interac-tions in spring barley (Hordeum vulgare). Heredity56,, 255—262.
QUALSET, C . O . , and R. M. GRANGER, 1970: Fre-
quency-dependent stability of performance in oats.Crop Sci. 10, 386—389.
RAO, S. S., and K. V. RAO, 1978: Genotypic stabili-ty of sorg,hum varieties and hybrids. Ind. J. Agric.Sci. 48, 691—695.
REICH, \V. H . , and R. E. ATKINS, 1970: Yield
stability of four population tj'pes of grain sorghum.Sorghum bicolor (L.) Moench, in different environ-ments. Crop Sci. 10, 511—517.
ROEMER, T., 1917: Sind die ertragsreichen Sortenertragssicherer? Mitt. DLG 32, 87—89.
ROWE, P. R., and R. H. ANDREW, 1964: Phenotypicstabihty for a systematic series of corn genotypes.Crop Sci. 4, 563—564.
RUSSELL, W . A., and S. A. EBERH.ART, 196S: Test-
crosses of one- and two-ear types of corn beltinbreds. II. Stability of performance in differentenvironments. Crop Sci. 8, 248—251.
SALJMON, S. C , 1951: Analysis of variance and long-time variety tests of wheat. Agron. J. 43, 562—570.
SCHNELL, F. W., 1967: Die Methoden zur Erfassungder phanotypischen Stabilitat von Zuchtsorten.Rundschreiben 1,/1967, AG Biomeine, DLG-Pflan-zenzuchtabt., 18—21.
, 1982: A synoptic study of the methods andcategories of plant breeding. Z. Pflanzenziichtg. 89,1—18.
, and H. G. BECKER, 1986: Yield and yieldstability in a balanced system of widely differingpopulation structures in Zea mays L. Plant Breed-ing 97, 30—38.
SCHUTZ, W. M., and C. A. BRIM, 1971: Inter-genotypic competition m soybeans. III. An evalua-tion of stability in multiline mixtures. Crop Sci. 11,684—689.
SEPAHI, A. , 1974: Estimating stability and selectionindex. Iran, J. Agric. Res. 2, 121—127.
SHUKL.*!, C . K., 1972a: Some statistical aspects ofpartitioning genotype-environmental componentsof variability. Heredity 29, 237—245.
, 1972 b: An invariant test for the homogeneityof variances in a two-wa\' classification. Biometrics28, 1063-1072.
SINGH, H . , and K. S. BAINS, 1984: Stability analysisof varietal mixtures in chickpea [Cieer arietinumL.). Z. Pflanzenzuchtg. 92, 159—167.
SKR0PPA, T., 1984: A critical evaluation of methodsavailable to estimate the genotype x environmentinteraction. Studia Forestalia Suecica 166, 3—14.
SNOAD, B., and A. E. ARTHCR, 1976: The use of
regression techniques for predicting the response ofpeas to environments. Theor. Appl. Genetics 47,9—19.
SPRAGUE, G . F., and W. T. FEDERER, 1951: A
comparison of variance components m corn }'ieldtrials. 11. Error, year X variety, location X varietyand variety components. Agron. J. 42, 535—541.
SPRENT, P., 1969: Models in regression and relatedtopics. London: Methuen.
STRINGFIELD, G . H . , and R. M. SALTER, 1934:
Differential response of corn varieties to fertilitylevels and to seasons. J. Agnc. Res. 49, 991—lOod.
T.'\J, G. G. C , 1971: Genotypic stability analysisand its application to potato regional trails. CropSci. 11, 184—190.
TAN, W . - K . , and G. Y. TAN, 1980: Combiningability analyses of stability parameters and forageyield in smooth bromegrass. Theor. Appl. Genet.58, 71-74.

Stability Analysis in Plant Breedmg 2,3
THOMSON, N . J., and R. B. CUNNINGHAM, 1979:
Genotype x environment mteractions and evalua-tion of cotton cultivars. Austr. j . Agric. Res. 30,105—112.
UTZ, H . P. , 1972: Die Zerlegung der Genotyp XUmwelt Interaktion. EDV in Medizin und Biologie3, 52—59.
VASIL], D . , and S. MILAS, 1984: Relationship be-
tween yield stability parameters estimated with dif-ferent methods for some maize and wheat geno-types. Vortr. Pflanzenziichtg. 7, 266—279.
WAHLE, G . , 1977: Analyse der Heterosis des Ertragsund der Ertragskomponenten bei Wmterroggen[Secale eereale L.). Dissertation, UniversitatHohenheim.
WALKER, A. K., and W. R. FEHR, 1978: Yield
stability of soybean mixtures and ,multiple purestands. Grop Sci. 18, 719—723.
WEATHERSPOON, J . H . , 1970: Comparative yields ofsingle, three-way, and double crosses of maize.Crop Sci. 10, 157—159.
WEBER, W . E. , und G. WRICKE, 1987: Wie zuverlas-
sig sind Schatzungen von Parametern der phanoty-pischen Stabilitat? Vortr. Pflanzenzuchtg. 12,120—133,
WERMRL, M. , 1966: Untersuchungen uber dieokologische Streubreite einiger Sorten des W'el-schen ^'eidelgrases [Lolium mukiflorum Lam.italieum). Z. Acker- u. Pflanzenbau 123, 309—331.
WESTCOTT, B., 1986: Some methods of analysinggenotype-environment interactions. Heredity 56,243—253.
WESTER.MAN, J. M., and M. J. LAWRENCE, 1970:Genotype-environment interaction and develop-mental regulation in Arabtdopsis tbahana. I. Inbredlines; description. Heredity 25, 609—627.
WlDNER, ]. N., and K. L. LEBSOCK, 1973: Combin-ing ability in durum 'ft'heat. I. Agronomic charac-teristics. Grop Sci. 13, 164—167.
WRICKE, G . , 1962: tjber eine Methode zur Erfas-sung der okologischen Streubreite in Feldver-suchen. Z. Pflanzenzuchtg. 47, 92—96.
, und •*'. E. WEBER, 1980: Erweiterte Analysevon Wechselwirkungen m Versuchsserien. In:KoPCKE, W., and K. UBERLA (eds.), Biometrie —heute und morgen, 87—95. Berlin—Heidelberg—New York: Spnnger-Verlag.
WRIGHT, A. J. 1976: Bias in the estimation of regres-sion coefficients in the analysis of genotype-envi-ronmental interaction. Heredity 37, 299—303.
Y.WES, F., and W. G. CoCHRAN, 1938: The analysisof groups of experiments. J. Agr. Sci. 28, 556—580.
Authors' addresses: Dr. H. C. BECKER, The S'n'ed-ish University of Agricultural Sciences, Departmentof Crop Genetics and Breeding, S-26800 Svalov(Sw'eden); Dr. J. LEON,, Institute of Crop Scienceand Plant Breedmg, Christian-AIbrechts-UniversitatKiel, Olshausenstr. 40, D-2300 Kiel 1 (Germany,F.R.).





















