
Use of Customized 3D-Printed Titanium Augment ... - Frontiers
Upload
independentCategory
view
1download
0
doi:10.1016/j.jmb.2006.01.060 J. Mol. Biol. (2006) 358, 172–192
SRY and Human Sex Determination: The Basic Tailof the HMG Box Functions as a Kinetic Clampto Augment DNA Bending
Nelson B. Phillips1†, Agnes Jancso-Radek1†, Varda Ittah2
Rupinder Singh1, Ging Chan1, Elisha Haas2 and Michael A. Weiss1*
1Department of BiochemistryCase School of Medicine, CaseWestern Reserve UniversityCleveland, OH 44106-4935USA
2Faculty of Life Sciences, BarIlan University, Ramat Gan52900, Israel
0022-2836/$ - see front matter q 2006 E
†N.B.P. & A.J.-R. contributed equPresent address: A. Jancso-RadekAbbreviations used: 6-FAM, 6-iso
high mobility group; GMSA, gel mosubstance (also designated Anti-Mueffect spectroscopy; PGE, permutatichromatography; SOX, SRY-relatedchromosome; SRY-p and SRY-pD, dorhodamine; TBP, TATA-binding protare designated by standard single-leindicated. Consensus position 1 coritalics, and proteins in capital letterE-mail address of the correspond
Human testis-determining factor SRY contains a high-mobility-group(HMG) box, an a-helical DNA-binding domain that binds within anexpanded minor groove to induce DNA bending. This motif is flanked onthe C-terminal end by a basic tail, which functions both as a nuclearlocalization signal and accessory DNA-binding element. Whereas theHMG box is broadly conserved among otherwise unrelated transcriptionfactors, tails differ in sequence and mode of DNA binding. Contrastingexamples are provided by SRY and lymphoid enhancer factor 1 (LEF-1):whereas the SRY tail remains in the minor groove distal to the HMG box,the LEF-1 tail binds back across the center of the bent DNA site. The LEF-1tail relieves electrostatic repulsion that would otherwise be incurred withinthe compressed major groove to enable sharp DNA bending with highaffinity. Here, we demonstrate that the analogous SRY tail functions as a“kinetic clamp” to regulate the lifetime of the bent DNA complex. As inLEF-1, partial truncation of the distal SRY tail reduces specific DNA affinityand DNA bending, but these perturbations are modest: binding is reducedby only 1.8-fold, and bending by only 7–108. “Tailed” and truncated SRYcomplexes exhibit similar structures (as probed by NMR) and distributionsof long-range conformational substates (as probed by time-resolvedfluorescence resonance energy transfer). Surprisingly, however, the SRYtail retards dissociation of the protein–DNA complex by 20-fold. Themarked and compensating changes in rates of association and dissociationobserved on tail truncation, disproportionate to perturbations in affinity orstructure, suggest that this accessory element functions as a kinetic clampto regulate the lifetime of the SRY-DNA complex. We speculate that thekinetic stability of a bent DNA complex is critical to the assembly andmaintenance of a sex-specific transcriptional pre-initiation complex.
q 2006 Elsevier Ltd. All rights reserved.
Keywords: gene regulation; protein structure; intersex abnormalities;gonadal dysgenesis; human development
*Corresponding authorlsevier Ltd. All rights reserved.
ally to the work., Epicentre Technologies, Madison, WI 53713, USA.mer of aectamido fluorescein; FRET, fluorescence resonance energy transfer; HMG,bility-shift assay; LEF-1, lymphoid enhancer factor 1; MIS, Mullerian inhibitingllerian Hormone); NLS, nuclear localization signal; NOESY, nuclear Overhauseron gel electrophoresis; rp-HPLC, reverse-phase high-performance liquidHMG box; SRY, protein encoded by the sex-determining region of the Ymains containing residues 2–86 and 2–78, respectively; TAMRA, tetramethyl-ein; TCF-1a, T-cell factor 1a and homologue of LEF-1; tr, time resolved. Amino acidstter code. Residue numbers in SRY refer to an HMG box consensus unless otherwiseresponds to residue 56 of intact human SRY. Genes and DNA sites are indicated ins.ing author: [email protected]

A Kinetic Clamp in SRY Augments DNA Bending 173
Introduction
Sexual dimorphism provides a model of a geneticswitch between alternative programs of develop-ment.1 The male phenotype in eutherian mammalsis determined by Sry (sex-determining region of theY chromosome),2 a gene on the short arm of the Ychromosome. SRY contains a sequence-specificHMG box,3 a conserved motif of DNA bindingand DNA bending.4 Assignment of SRY as thetestis-determining factor (TDF) is supported bystudies of transgenic murine models5 and humanintersex abnormalities.6 SRY belongs to an extendedfamily of SRY-related HMG-box proteins, desig-nated SOX.7 Members of this family play critical butdiverse roles in mammalian organogenesis, includ-ing development of the heart8 and central nervoussystem.9 Autosomal gene Sox9 functions down-stream of Sry in the testis-determining pathway.10,11
Clinical mutations in either Sry6,12 or Sox913,14 areassociated with pure gonadal dysgenesis leadingto XY sex reversal. The HMG boxes of SRY, SOX9,and other members of the SOX family exhibitsimilar specific DNA-binding and DNA-bendingproperties.15–18
How SRY regulates testicular differentiation isnot well understood. Comparison of Sry alleles hasdemonstrated marked divergence outside of theHMG box.19 Selective conservation of a specificDNA-bending motif led to the proposal that SRYfunctions as an architectural transcription factor.3,20
This proposal is supported by clinical observationsthat almost all point mutations causing human sexreversal occur de novo in the HMG box and impairspecific DNA binding20,21 or DNA bending.22 Thatthe primary function of SRY is site-specific DNAbending is suggested by studies of chimeric murineSry (mSry) transgenes in XX mice.23,24 Participationof SRY-interacting proteins in multi-protein-DNAcomplexes25–28 and splicing-related RNAcomplexes29 has also been proposed.30 Althoughtarget genes for SRY-mediated transcriptional regu-lation have not been defined, the dramatic effects ofHMG boxes on DNA structure (Figure 1(a) and (b))are proposed to contribute to assembly of sex-specific transcriptional pre-initiation complexes(Figure 1(c)).30,31 Studies of chimeric SRY trans-genes have shown that SRY-directed sex reversal inXX mice readily tolerates modest changes inthe molecular properties of the HMG box,23,24
including subtle changes in sequence specificityand extent of DNA bending.32,33
The HMG box exhibits an L-shaped structurein which three a-helices and an N-terminal b-strandpack to form major and minor “wings”(Figure 1(a)).34,35 Themotif was originally describedin the non-sequence-specific DNA-bindingdomains of abundant non-histone chromosomalproteins.31 Two classes of HMG boxes are distin-guished by their DNA-binding properties. Whereasnon-histone chromosomal proteins (such as HMG1and HMG2) typically contain two or more boxesthat recognize distorted DNA structures with weak
or absent sequence specificity, sequence-specificarchitectural transcription factors (such aslymphoid enhancer factor 1; LEF-1)36,37 containone HMG box that recognizes specific DNAsequences. Specific and non-specific domains eachdock within a widened minor groove: the angularwings provide a template for DNA bending.37–41
The extent of bending varies but in each case theprotein binds on the outside of the DNA bend tocompress the major groove. Structural models ofsequence-specific HMG box-DNA complexes areprovided by SRY (Figure 1(a))38,42 and LEF-1(Figure 1(b)).37 DNA bend angles in thesestructures are 548 and 1178, respectively. Thereported precision of the DNA bend angle in theSRY complex is G28. The physical significance ofsuch precision is not well characterized.The structure of an SRY–DNA complex has
provided a basis for analysis of mutationsassociated with sex reversal (46, XY pure gonadaldysgenesis).38 The present study focuses on the roleof a conserved basic region C-terminal to the HMGbox (red segment in Figure 1(a) and Figure 2(a)).Comprising residues 125–140 in human SRY(residues 70–85 in an HMG box consensussequence; Figure 2(b)), this basic C-terminal tailalso contains a nuclear localization signal43,44 andsites of clinical mutations.30 Although SRYand LEF-1 contain analogous tails, the two proteins exhibitdifferent patterns of basic residues, proline andglycine residues (upper panel and last line ofFigure 2(b), respectively). These divergent featurescorrespond to distinct structural roles. Whereas theSRY tail remains in the DNA minor groove distal tothe HMG box and is well ordered (Figure 1(a)),42 theLEF-1 tail crosses back across the major groove inthe center of the DNA site and is not well ordered(Figure 1(b)).37 The basic side-chains of the LEF-1tail (KKRK; green in Figure 2(b)) are proposed toalleviate electrostatic repulsion that would other-wise occur between neighboring DNA phospho-diester charges in the compressed major groove.37
Truncation of the LEF-1 tail markedly impairs bothDNA bending (from 1308 to 528) and specific DNAaffinity (by at least 100-fold).45–48
To define the contribution of the SRY tail to DNAbinding and DNA bending, we have investigatedSRY domains containing either intact or truncatedtails. These domains (designated “tailed” or trun-cated, respectively) correspond to the polypeptidesemployed in two successive NMR studies of theSRY–DNA complex.38,42 We have characterizedtailed and truncated domains in otherwise identicalcontexts and by multiple independent biochemicaland biophysical assays. As expected,49 the structureand stability of the free domain are not perturbedby truncation of its disordered tail; respective DNA-bound structures are also similar. As in LEF-1,45–48
the SRY tail augments specific DNA binding andbending, but in SRY these effects are modest: DNAbinding is reduced by less than twofold and DNAbending by only 7–108. Further, binding of the taildoes not damp long-range fluctuations in DNA

Figure 1. Sequence-specific HMG box–DNA complexes (stereo pairs) and enhanceosome model. (a) SRY–DNAcomplex (Protein Data Bank accession code 1J46).42 Ribbon model of protein is shown in green (HMG box) and red (tail;residues 70–85). DNA is shown as gray sticks. The L-shaped structure of the HMG box contains a major wing (consistingof a-helix 1, a-helix 2, the first turn of a-helix 3, and connecting loops) and minor wing (consisting of the N-terminalb-strand, the remainder of a-helix 3, and C-terminal tail). (b) LEF1–DNA complex (PDB accession code 2LEF).37 A ribbonmodel of the protein is shown in blue (HMG box) and red (tail; residues 70–85). DNA is shown as gray sticks. Theasterisk (right panel) indicates binding of the LEF-1 tail within the compressed major groove. Cantilever side-chains I13(SRY) and M13 (LEF-1; M11 in PDB entry) are shown as orange sticks at the center of complexes. (c) Schematic model ofSRY-directed assembly of a male-specific transcriptional pre-initiation complex. The sharp DNA bend is clamped by theSRY tail (red; asterisk). Right: activated transcription occurs in the presence of bound SRY (green) as the specific DNAbend permits assembly of the enhanceosome.93 Left: activated transcription is off in the absence of SRY due todisassembly of the DNA–multiprotein complex, including activator–coactivator complexes. Putative factor X (purple atright) is proposed to bind cooperatively with SRY to an adjoining DNA site.94
174 A Kinetic Clamp in SRY Augments DNA Bending

Figure 2. Tail sequences and mode of DNA binding. (a) Ribbon model of SRY HMG box with selected side-chains aslabeled. A disordered tail is shown as broken red line. (b) Tail sequences among mammalian SRY alleles (upper group)and mouse LEF-1 (last line). SRYalleles are as follows: h, human; m, mouse; b, bovine; g, goat; e, equine; p, porcine; ando, ovine. Purple and red bars indicate the positions of tail; purple (residues 70–78) delimits the truncated tail. The asteriskindicates R133, the site of sex-reversal mutation R133W.44,60 The arrowhead indicates the C terminus of the truncateddomain used in a previous NMR study38 and here. Boxes indicate residues discussed in the text. Broken boxes indicatedivergent residues in the LEF-1/TCF-1 family. Basic side-chains are highlighted in green. (c) Position of the SRY tail inthe protein–DNA complex (stereo pairs, views rotated by 908 about the vertical axis). Protein is shown as sticks, andDNA in space-filling representation. Residues 70–78 (purple) are present in the truncated tail whereas residues 79–85(red) are included in the “tailed” domain. The DNA backbone is shown in blue, and base-pairs in dark gray. In the upperpanel the green sphere indicates a para-oxygen atom of Y74; the orange sphere is adjoining the H1 0 atom of deoxyribose(distance 3.0 A). DNA contacts are made by K73, Y74, P76, R78, K79, and K81.42
A Kinetic Clamp in SRY Augments DNA Bending 175

Table 1. Thermodynamic stabilities and FRET analysis
A. Thermodynamic parametersa
Protein Cmid (M) m (kcal molK1 MK1) DGu (kcal/mol) Tm (8C)
Intact tail 2.2G0.1 1.8G0.1 4.0G0.1 39.0G0.5Truncated 2.2G0.1 1.8G0.1 4.0G0.1 40.0G0.5
B. FRET distance-distribution analysisb
Sample Temp (8C) Peak (A) Mean (A) FWHM (A)c c2,d
Free DNA 4 62.6 (62.5–62.7) 62.7 (62.5–62.7) 16.7 (14.5–17.0) 1.4215 61.7 (61.6–61.8) 61.7 (61.6–61.8) 14.6 (14.5–14.7) 1.45
Tailed complex 4 51.9 (51.6–52.0) 52.0 (51.7–52.1) 20.2 (19.8–20.6) 1.7315 51.5 (51.2–51.6) 51.5 (51.3–51.7) 19.0 (18.8–19.4) 1.75
Truncated complex 4 54.0 (53.8–54.2) 54.1 (53.9–54.3) 22.0 (21.5–24.4) 1.6315 53.8 (53.6–53.9) 53.8 (53.6–54.0) 20.3 (18.1–20.8 1.61
a Tm was obtained by temperature denaturation experiments monitored by CD at 222 nm. Steady-state tryptophan fluorescence andautomated guanidine titrations were otherwise used to determine thermodynamic parameters (see Materials and Methods). Cmid isdefined as that concentration of guanidine at which 50% of the protein is unfolded. The m value, defined as the derivative of DGu withrespect to denaturant concentration, is related to the difference in the solvent accessible surface area between unfolded and native states.The statistical confidence of fitting is in each case characterized by an R value greater than 0.9998. Thermodynamic parameters obtainedby CD-detected guanidine titration of the intact domain are essentially identical to those obtained by fluorescence.
b Measurements were performed at 4 8C and 15 8C. Each analysis included two independent repeats of each experiment analyzedsimultaneously, a procedure which reduced the statistical uncertainties of the parameters. The peak of the end-to-end distributionfunction and its mean are provided. The values in parentheses represent possible errors in parameters calculated as described.91
Average Ro value was 58.3 A for the free DNA at 4 8C; the specific Ro value suitable for each experiment was introduced using theradiative lifetime in order to account for the variation of the values of Ro with the lifetime components of the donor.
c FWHM indicates the full width at half maximum of the distribution.d Global c2 derives from joint analysis of four donor fluorescence decay curves; its value is raised by the large number of experiments
forced into joint analysis.
176 A Kinetic Clamp in SRY Augments DNA Bending
bending as probed by time-resolved fluorescenceresonance energy transfer (tr-FRET).33 Despite thesesimilarities, marked changes are observed in ratesof SRY–DNA association and dissociation: the tailretards dissociation of the protein–DNA complexby 20-fold. Because this perturbation is dispropor-tionate to changes in affinity or structure, wepropose that the SRY tail functions as a “kineticclamp” to regulate the lifetime of the bent DNAcomplex and in turn the assembly and maintenanceof a sex-specific transcriptional pre-initiationcomplex.
Results
This study employs human SRY domainsconsisting of the HMG box (residues 56–124 in theintact protein) and C-terminal basic regions ofdifferent length (residues 125–133 and 125–141).These correspond to residues 2–78 and 2–86 in aconsensus HMG-box numbering scheme.38,42 TheDNA-bending properties of the tailed fragment aresimilar to those of full-length SRY.20,33 As expected,the domains exhibit similar secondary structures asinferred from far-ultraviolet CD spectra. Trpfluorescence spectra are likewise identical,suggesting that the mean environments of thethree Trp side-chains in the major wing (W15,W43, and W52; residue numbers refer to HMGconsensus) are unperturbed by truncation of thetail. Thermal and denaturant-induced unfoldingtransitions, as monitored by CD and intrinsic Trpfluorescence, respectively, likewise overlap and
yield similar estimates of transition midpointtemperatures (Tm) and free energies of unfolding(DGu; Table 1A). 1H-NMR spectra of the domainsare essentially identical with the exception of sharpresonances at near random-coil chemical shiftsconsistent with the presence (or absence) of adisordered tail.
The SRY tail modestly enhances DNA bindingand DNA bending
Specific protein–DNA binding (site 5 0-TGATTGTTCA-3 0 and complement; core consensussequence underlined) was initially investigatedusing a gel mobility-shift assay (GMSA). A 36 bpDNA probe was employed to provide sufficientnon-specific flanking DNA sequences to permitevaluation of higher-order complexes.33,50,51 Inaccord with prior studies33,50,51 the fragments bindto the specific DNA site at nanomolar proteinconcentrations (complex c1; Figure 3(a)) and formadditional complexes (bands c2 and c3) at higherprotein concentrations. The qualitative pattern ofshifted bands is similar in the presence of an intactor truncated tail; 50% occupancy of the c1 site underthese conditions is observed at respective proteinconcentrations of ca 8 nM (intact tail) and 20 nM(truncated tail). The specific affinity of the truncatedfragment is thus reduced by ca 2.5-fold. Essentiallyidentical results are obtained at 4 8C and 15 8C.Although dissociation constants inferred fromGMSA studies are approximate and can beinfluenced both by off-rate effects and by theconcentration of unlabeled competitor DNA,

Figure 3. GMSA DNA-bindingand DNA-bending assays. (a)Apparent specific DNA affinity ofthe tailed domain (right-handlanes) is ca 2.5-fold higher thanthat of truncated fragment (left-hand lanes). Complex (designatedc1 below) indicates a specific 1:1complex; c2 and c3 indicate higher-order complexes. Protein concen-trations are (a) 0 nM, (b) 2.5 nM, (c)5 nM, (d) 10 nM, (e) 20 nM, (f)40 nM and (g) 80 nM. Quantifi-cation (in order of increa-singprotein concentration): with intacttail, percent shifts are (a) 0%, (b)31% c1, (c) 37% c1, (d) 75% c1 and!1% c2, (e) 89% c1 and 2% c2, (f)82% c1 and 16% c2; with truncatedtail, (a) 0%, (b) 4% c1, (c) 5% c1, (d)24% c1, (e) 56% c1, and (f) 78% c1and 8% c2. (b) and (c) DNAbending by full-length SRY andtailed domain: (b) PGE and (c)plot of relative electrophoreticmobility as function of flexuredisplacement in DNA probes. Cor-responding permutation probesa–-f (with the consensus sequence5 0-ATTGTT at varying distancesfrom the ends of the 146 bp probeas described in Materials andMethods) are shown with full-length SRY (left-hand set) andwith tailed domain (right-handset). Studies employed DNA site50-CCCATTGTTCTC-3 0 and comp-
lement (core consensus sequence underlined; site 2 in (d)). Inferred DNA bend angles with full-length SRYor the taileddomain are 69.7(G1)8 and 69.2(G1)8, respectively, in this gel system (6.5% polyacrylamide with 29:1 ratio ofacrylamide:bis-acrylamide at 4 8C). (d) DNA sequences of duplex sites employed in DNA bending assays (see Table 2).Sites 1–3 share the core consensus element 5 0-ATTGTT-3 0 and complement (boxed) but differ at flanking positions. Site 4contains transversions at indicated positions (asterisk and arrow). Derived from a SOX9 element in MIS promoter,57,58 a14 bp duplex containing site 4 (5 0-TTTGTG) was employed in structures shown in Figures 1(a) and 2(c).42
A Kinetic Clamp in SRY Augments DNA Bending 177
the ratio of specific DNA binding affinities isconsistently between 2 and 3 (tailed:truncateddomains). Because the truncated complex is lesslong lived, possible GMSA artifacts due to pre-mature dissociation of specific complexes duringelectrophoresis would exaggerate the apparentdifference between fragments, yielding an upperbound of 3 for the ratio of dissociation constants.The net contribution of the missing residues at thetail-DNA interface (Figure 2(c)) to the free energy ofspecific DNA binding is thus low (DDGb!0.7 kcal/mol). Absent interactions include multiple saltbridges and potential hydrogen bonds to thephosphodiester backbone (by R78, K79 and K81)and vanderWaals interactionswithDNAbases (P76and R78; Figure 2(c)).42 FRET-basedmeasurement ofthe ratio of dissociation constants at 15 8C yields avalue of 1.8G0.1 (final section of Results, below).
SRY-induced DNA bending was initially eval-uated using permutation gel electrophoresis(PGE).52 In this approach53,54 the mobilities of
DNA probes are observed to depend on theposition (“flexure displacement”) of the specifictarget site, enabling induced DNA bend angles tobe inferred.22,33,51 Three consensus DNA siteswere investigated in which the core recognitionelement (5 0-ATTGTT-3 0 and complement) isimbedded in different sequence contexts (sites1–3 in Figure 3(d)). Site 1, as previouslycharacterized,12,51,55,56 is a high-affinity variant ofa SOX9 target element in the promoter of theMullerian Inhibiting Substance gene (MIS).57,58 Site2 is a high-affinity variant of a LEF-1 targetelement (designated TCR-11) previouslyemployed in studies of SRY.32 Site 3 is a high-affinity SRY site employed by Goodfellow andco-workers.22 Site 4 is the SOX9 target site in theMIS promoter employed in structural studies ofthe SRY–DNA complex;38,42 5 0- and 3 0 non-consensus base-pairs are indicated inFigure 3(d) by an arrow and asterisk, respectively.Relative to a consensus 5 0-ATTGTT-3 0 DNA site,

Table 2. Electrophoretic DNA-bending studiesa
A. Comparison of full-length SRY and tailed domainb
Site qCtail (deg.) qfull (deg.) Dq
1 tgATTGTTca 69 70 1c
B. Comparison of tailed and truncated complexesc
Site qCtail (deg.) qtrunc (deg.) Dq (deg.)
1 tgATTGTTca 79 69 102 ccATTGTTct 87 80 73 gcATTGTTat 83 NDc ND4 tgTTTGTGca 68 49 19
C. Comparison of native and mutant domainsd
Site Fragment Mutation qwild-type qmutant Dq
1 SRY-p K81A 79 75 41 SRY-p R78W 79 73 61 SRY-pD R78W 69 58 11
a Tailed and truncated SRY domains are designated SRY-p and SRY-pD, respectively. DNA sites are depicted in Figure 3(d). InferredDNA bend angles qfull, qCtail and qtrunc, respectively, refer to full-length SRY and SRY domains containing an intact C-terminal tail(ending at residue 85) or truncated tail (ending at residue 78). Angles are reproducible toG18 on multiple gels of the same composition;precision of Dq is G28.
b Full-length SRY (qfull; column 3) and SRY-p (qCtail; column 2). Change in apparent bend angle of SRY-p between lines 3 and 6 is dueto difference in gel composition (in line 3 part A 6.5% acrylamide with 29:1 ratio of acrylamide to bis-acrylamide whereas followingstudies employed 10% acrylamide with 58:1 ratio). Differences (Dq) (but not necessarily absolute values) are robust to changes in gelcomposition.
c ND, values not determined; estimate of qCtail in line 8 was based on four probes (a, c, d, and f; see Materials and Methods).d Positions 78 and 81 in the HMG box correspond to residues 133 and 136 in full-length human SRY. P300-mediated acetylation of
K136 in full-length SRY regulates nuclear localization.78 Samples were run side-by-side in the same gel in duplicate.
178 A Kinetic Clamp in SRY Augments DNA Bending
the affinity of variant site 4 for either fragment isdecreased by threefold.12
A summary of PGE results is provided in Table 2.Although calibration can vary with gel composition,differences between bend angles of variantcomplexes, as investigated side-by-side in the samegel, are robust to gel composition. DNA bending bythe tailed HMG domain is similar to that induced byfull-length SRY (Dq!38; Figure 3(b) and Table 2A) inaccord with previous studies.20,33 Binding of SRYdomains containing intact or truncated tails, how-ever, yields distinct patterns of PGE mobilities(Figure 4(a) and (b)). Inferred bend angles (qCtail
and qtrunc; columns 2 and 3 in Table 2B) anddifferences between intact and truncated complexes(Dq; column 4) demonstrate that for consensus DNAsites (sites 1 and 2), truncation of the tail leads to 7–108decrements in bend angle. For the non-consensus siteemployed in previous structural studies (5 0-TTTGTG-30 and complement; site 4 in Figure 3(d)),the extent of impairment is significantly larger (Dq198). Comparison of an additional variant site (50-ATTGTG-3 0 and complement; variant in bold)suggests that the 30 T/G substitution is responsiblefor this additional decrement.
Mutations in tail modestly impair DNA bending
The contribution of a conserved lysine residue inthe truncated segment (K81; boxed in Figure 2(b))was evaluated through study of a K81A analogue.This substitution lies at the tail–DNA interface(Figure 2(c)) and impairs DNA bending to a smallbut measurable extent (Dq 48; Table 2C), yielding
a DNA bend angle (758) almost midway betweentailed and truncated complexes (798 and 698,respectively; Table 2B). Effects on specific DNAaffinity are less than 1.5-fold. Sex-reversal mutationR78W (R133W in full-length SRY) likewise occurs ata conserved DNA contact site in the tail(Figure 2(b)) and (c)). Previous studies haveestablished that this clinical variant, investigatedin the context of a tailed domain, exhibits wild-typeaffinity for site 1 and only a small decrement inDNA bend angle (Dq 68; Figure 4(c)).44 R133 is theC-terminal residue of the truncated domain (arrow-head in Figure 2(b)). Because this fragment has beenused to characterize many sex-reversal mutationsand species variants,22,38,59 we reinvestigated theR133W mutation in that context. Whereas themutation does not affect the affinity of the taileddomain, it impairs specific binding of the truncatedfragment by twofold relative to the wild-typetruncated fragment. Further, truncation of the tailalmost doubles the apparent bending defect due tothe mutation (588 versus 698; Figure 4(d) andTable 2C). An intact tail thus partially restores anative DNA bend angle otherwise perturbed bymutation of an adjoining site in the protein.Gonadal dysgenesis in this patient is attributed toimpaired nuclear import of SRY.44,60
FRET studies of DNA bendingand conformational fluctuations
Bending of site 1 in a 15 bp DNA duplex(5 0-TCGGTGATTGTTCAG-3 0 upper strandand complement) was characterized by tr-FRET.33

Figure 4. DNA bending by SRYdomains and sex-reversal variants.(a) PGE and (b) plot of relativeelectrophoretic mobility as functionof flexure displacement in DNAprobes. Binding of the taileddomain is shown in the left-handportion of (a) (filled triangles in(b)); binding of the truncateddomain is shown in the right-hand portion (squares in (b)).Studies employed consensus DNAsite 2 in Figure 3(d).69 InferredDNA bend angles with intact SRYand tailed domain are 71(G1)8 and61(G1)8, respectively, in this gelsystem (8.5% polyacrylamide with29:1 ratio of acrylamide:bis-acryl-amide at 4 8C). Although estimatesof apparent bend angles candepend on gel composition, differ-ences (Dq) are robust to percent
polyacrylamide and the ratio of bis-acrylamide. (c) and (d) Tail truncation alters apparent effects of the sex-reversalR78Wmutation (residue 133 in intact human SRY) on DNA bending. PGE of wild-type and variant with (c) intact tail and(d) truncated tail. Corresponding permutation probes a–f are shown in each panel for wild-type (left-hand set) andvariant (right-hand set). Studies employed the consensus DNA site 1 (Figure 3(d)). Inferred DNA bend angles within thisgel system (10% polyacrylamide with 58:1 ratio of acrylamide:bis-acrylamide at 4 8C) are (c) 798 (wild-type) and 738(mutant) for intact tail, and (d) 698 (wild-type) and 588 (mutant) for truncated tail. Truncation of tail thus accentuates theapparent DNA bending decrement (DDq) from 68 to 118. Protein concentrations were 100 nM in (c) and 300 nM in (d).Whereas mutation does not perturb specific affinity of the tailed domain, it impairs binding of the truncated fragment bythreefold.
A Kinetic Clamp in SRY Augments DNA Bending 179
End-to-end DNA distances are sensitive to meanDNA bending and fluctuations in bend angle(Figure 5(a) and (b)). The DNA probe was labeledwith a fluorescent donor (fluorescein; 6-FAM) at one5 0 end and/or a fluorescent acceptor (tetramethyl-rhodamine; TAMRA) at the other 5 0 end(Figure 5(b)).33,51 Time-resolved fluorescence aniso-tropy measurements (Supplementary Data) demon-strate that the shortest correlation-time componentand its pre-exponents are similar among thesamples. Although differences between time-resolved anisotropies are small, the short com-ponents in the truncated complex are closer to thoseof the free DNA than are the correspondingcorrelation times in the tailed complex.
Steady-state FRET efficiency is enhanced onbinding of SRY (Figure 5(c)). As predicted by PGEstudies, qualitative comparison of tailed (greenspectrum in Figure 5(c)) and truncated (bluespectrum) complexes demonstrates that thisenhancement is slightly larger in the presence ofthe tail: steady-state FRET efficiencies are 51(G1)%(free DNA), 66(G1)% (truncated complex), and69(G1)% (tailed complex). Time-resolved measure-ments of the decay of donor fluorescence(Figure 5(d)) demonstrate a significant change indonor lifetime in the double-labeled DNA site onbinding of either the tailed or truncated SRYdomain. Because the reduction is more marked inthe tailed complex (green decay curve inFigure 5(d)), qualitative inspection of these datareveals that its binding shortens the mean
end-to-end distance in the DNA measurably morethan does binding of the truncated domain (bluedecay curve in Figure 5(d)). Global analyses ofrespective donor decay curves in singly and doublylabeled samples yields estimates of end-to-enddistance distributions in the free and bound states(Figure 5(e)). Parameters are given in Table 1B. Onbinding of the tailed domain, the mean end-to-enddistance of the DNA at 15 8C was reduced from61.7 A (red Gaussian distribution in Figure 5(e)) to51.5 A (green distribution). By contrast, on bindingof the truncated domain, the mean distance wasreduced only to 53.8 A (blue distribution).Although such reductions of end-to-end distancereflect both DNA bending and correlated changesin DNA unwinding, the predominant contributionis due to DNA bending.33,61,62 The tr-FRET thuscorroborates PGE studies that the tail accentuatesDNA bending. Skewed Gaussian modeling alsoindicates a significant broadening of the end-to-enddistributions in each complex relative to the freeDNA.Whereas the breadth of the distribution in thefree DNA (full width of the Gaussian at half-maximal height ca 14.6 A) is consistent with theallowed range offlexible linker configurations (A. R.Srinivasan & W. Olson, personal communication),the widths increase to ca 20 A in both complexes(Figure 5(e) and Table 1B). Such broadeningsuggests that the bound DNA structures exhibit adistribution of conformational substates and inturn a range of populated DNA bend angles;interconversion among substates must be in fast
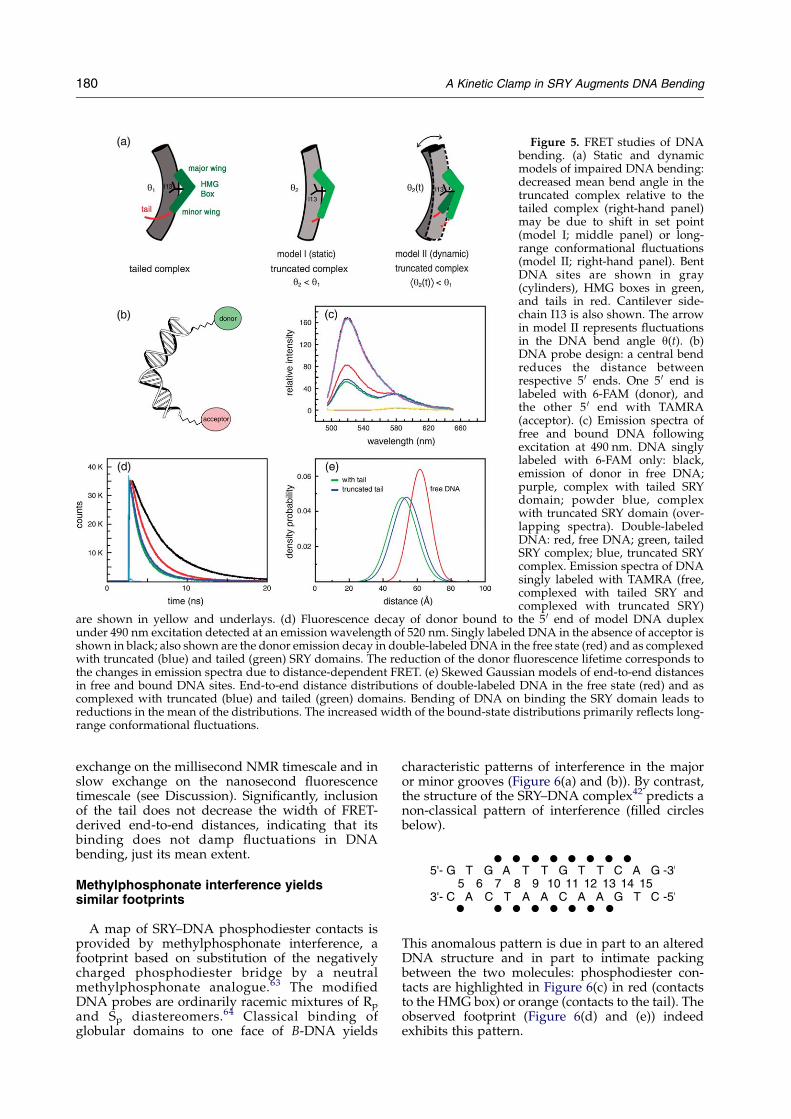
Figure 5. FRET studies of DNAbending. (a) Static and dynamicmodels of impaired DNA bending:decreased mean bend angle in thetruncated complex relative to thetailed complex (right-hand panel)may be due to shift in set point(model I; middle panel) or long-range conformational fluctuations(model II; right-hand panel). BentDNA sites are shown in gray(cylinders), HMG boxes in green,and tails in red. Cantilever side-chain I13 is also shown. The arrowin model II represents fluctuationsin the DNA bend angle q(t). (b)DNA probe design: a central bendreduces the distance betweenrespective 5 0 ends. One 5 0 end islabeled with 6-FAM (donor), andthe other 5 0 end with TAMRA(acceptor). (c) Emission spectra offree and bound DNA followingexcitation at 490 nm. DNA singlylabeled with 6-FAM only: black,emission of donor in free DNA;purple, complex with tailed SRYdomain; powder blue, complexwith truncated SRY domain (over-lapping spectra). Double-labeledDNA: red, free DNA; green, tailedSRY complex; blue, truncated SRYcomplex. Emission spectra of DNAsingly labeled with TAMRA (free,complexed with tailed SRY andcomplexed with truncated SRY)
are shown in yellow and underlays. (d) Fluorescence decay of donor bound to the 5 0 end of model DNA duplexunder 490 nm excitation detected at an emission wavelength of 520 nm. Singly labeled DNA in the absence of acceptor isshown in black; also shown are the donor emission decay in double-labeled DNA in the free state (red) and as complexedwith truncated (blue) and tailed (green) SRY domains. The reduction of the donor fluorescence lifetime corresponds tothe changes in emission spectra due to distance-dependent FRET. (e) Skewed Gaussian models of end-to-end distancesin free and bound DNA sites. End-to-end distance distributions of double-labeled DNA in the free state (red) and ascomplexed with truncated (blue) and tailed (green) domains. Bending of DNA on binding the SRY domain leads toreductions in the mean of the distributions. The increased width of the bound-state distributions primarily reflects long-range conformational fluctuations.
180 A Kinetic Clamp in SRY Augments DNA Bending
exchange on the millisecond NMR timescale and inslow exchange on the nanosecond fluorescencetimescale (see Discussion). Significantly, inclusionof the tail does not decrease the width of FRET-derived end-to-end distances, indicating that itsbinding does not damp fluctuations in DNAbending, just its mean extent.
Methylphosphonate interference yieldssimilar footprints
A map of SRY–DNA phosphodiester contacts isprovided by methylphosphonate interference, afootprint based on substitution of the negativelycharged phosphodiester bridge by a neutralmethylphosphonate analogue.63 The modifiedDNA probes are ordinarily racemic mixtures of Rp
and Sp diastereomers.64 Classical binding ofglobular domains to one face of B-DNA yields
characteristic patterns of interference in the majoror minor grooves (Figure 6(a) and (b)). By contrast,the structure of the SRY–DNA complex42 predicts anon-classical pattern of interference (filled circlesbelow).
This anomalous pattern is due in part to an alteredDNA structure and in part to intimate packingbetween the two molecules: phosphodiester con-tacts are highlighted in Figure 6(c) in red (contactsto the HMG box) or orange (contacts to the tail). Theobserved footprint (Figure 6(d) and (e)) indeedexhibits this pattern.

Figure 6.Methylphosphonate interference footprint of SRY–DNA complexes. (a) and (b) Classical backbone footprintof major andminor-groove DNA-binding proteins: binding of a globular domain to one face of B-DNA is associated witha staggered pattern of phosphate contacts (red arrowheads) whose orientation depends on which groove isoccupied.95,96 (c) Structure of the SRY–DNA complex42 indicates a non-classical pattern of contiguous phosphatecontacts to both strands due in part to DNA bending and unwinding. Phosphodiester groups contacted by the HMG boxand tail are shown in red and orange, respectively. Non-contacted phosphodiester groups are shown in black. Sites ofcontact not evident in the footprint are indicated by blackened non-bridging oxygen atoms. The HMG box is shown as agreen ribbon, and tail in brown. U and L designate the upper and lower strand, respectively. The triangle indicates thesite of cantilever insertion. The side-chain of residues I13 (cantilever) and Y74 are depicted in light blue. (d) and (e)Methylphosphonate interference footprint and sequence of DNA site. In accord with structure, sites of interference occuralong the upper strand (d) and lower stand (e) at the same base-pairs. Phosphodiester positions across the 15 bp DNAsite in complementary strands are designated by a common number based on the position of the nucleoside in the upperstrand. Base-pairs are numbered 1–15 from the left; i.e. 5 0-G1, G2, G3, etc. in the upper strand, and 3 0-C1, C2, C3, etc. inthe lower strand. The triangle between 8 and 9 indicates the site of cantilever insertion. (f) and (g) GMSA studies of 15 bp(f) and 12 bp (g) DNA probes containing single methylphosphonate modifications. (f) Comparison of upper strand sites7–12. (g) Comparison of selected sites in upper and lower strands. Unmodified DNA site is in each case designated wt.Arrows in (f) and (g) indicate differential effect of tail truncation at the 3 0 end of the core DNA site; triangles indicate thesite of I13 insertion. The asterisk in (g) indicates sites where modification enhances binding of the truncated domain.(f) Lanes 7 indicate modification at nucleoside A7, lanes 8 at T8, lanes 9 at T9, etc.; lanes wt contain an unmodified DNAsite. (g) Lanes 13 0 indicate modification at nucleoside A12 of bottom strand, lanes 14 0 at G13 of bottom strand, and lanes15 0 at T14. Quantification of percent shifts with intact tail, (lane 7) 53, (8) 10, (9) none detected, (10) 5, (11) 10, (12) 27, (wt)60, (12) 13, (13) 7, (14) 8, (13 0) 31, (14 0) 40, (15 0) 49, and (wt) 49%. Percent shifts with truncated tail: (lane 7) 13, (8) 1, (9)none detected, (10) 1, (11) 3, (12) 21, (wt) 20, (12) 60, (13) 15, (14) 17, (13 0) 74, (14 0) 72, (15 0) 79, and (wt) 50%. The truncateddomain appears to bind DNA probes with modifications at positions 13 0, 140, and 15 0 with slightly higher affinity (!1.5-fold increase) than it binds to unmodified DNA. A fivefold differential binding of tailed and truncated domains occurs atposition 12 (arrows).
A Kinetic Clamp in SRY Augments DNA Bending 181

182 A Kinetic Clamp in SRY Augments DNA Bending
Structural contacts not observed as sites of methyl-phosphonate interference (open circles above) occurat two flanking sites on the lower strand, contactedby R31 (A5) and K81 (G13). Almost completeinterference (O50-fold decrement in DNA binding)is observed at sites adjoining the partial inter-calation of I13 (part of the hydrophobic wedge ofthe HMG box;38 arrowhead above and in Figure 6(c)and (f)). These essential features are unaffected bytruncation of the tail (illustrated for the upper standin Figure 6(f)), implying similar modes of DNAbinding. A striking difference is nonethelessobserved at position 12 in the upper strand (arrowabove and in Figure 6(f) and (g)). Whereasmethylphosphonate substitution at this site impairsbinding of the tailed domain by fivefold, nosignificant interference is observed to binding ofthe truncated domain. This DNA site adjoins Y74 inthe proximal tail (blue side-chain in Figure 6(c); seealso Figure 2). We speculate that this loss ofinterference reflects loosening of the adjoiningSRY–DNA interface due to the absence of distaltail–DNA contacts.
A subtle difference between tailed and truncatedfootprints is observed at positions 13 0–15 0 inthe lower strand (i.e. the phosphate groups ofbases 3 0-AGT; see inset between Figure 6(d) and (e)).At these sites the neutral modification has negli-gible effect on binding of the tailed domain butenhances binding of the truncated domain (aster-isks below and in Figure 6(g)).
Although the extent of enhancement is small(!50%), this observation suggests that thenegative charges of the native DNA backboneslightly destabilize the truncated complex,presumably due to electrostatic repulsion. Theintact tail alleviates this effect.
Tailed and truncated SRY–DNA complexesexhibit similar NMR features
Similarity of DNA bend angles and methylpho-sphonate footprints suggest that tailed and trun-cated complexes exhibit similar structures. Probesof DNA structure are provided by imino resonances(Figure 7(a)). In the free DNA guanine iminoresonances (base-pairs 1, 3, 7, 10, and 12; numberingscheme inset in Figure 7(a)) are clustered between12 and 13 ppm, whereas thymidine resonances (bp2, 5, 6, 8, 9, and 11) are clustered between 13.4 ppmand 14.0 ppm (spectrum i in Figure 7(a)). Terminal
imino resonances are broad due to fraying of thedouble helix. Addition of either SRY domain toform a specific 1:1 complex results in markedchanges in chemical shifts (spectra ii and iii inFigure 7). Despite their differences in electrophor-etic behavior (Figure 4(a) and (b)), tailed andtruncated complexes exhibit similar patterns ofchanges in chemical shift (“complexation shifts”;Dd). In each case G7 is shifted upfield, for example,and T8 downfield. Further and in accord withprevious studies,55 insertion of the “cantilever”side-chain of I13 between T5 and T6 is associatedwith a dramatic upfield shift of T5 into theguanidine region (arrows in Figure 7(a)). Subtledifferences in complexation shifts are nonethelessobserved between the two complexes. In general,complexation shifts in the tailed complex areslightly larger than in the truncated complex(broken vertical lines in Figure 7(a); SupplementaryData). In addition, the broad terminal resonances ofG1 and G12 (asterisks in spectrum ii) are furtherbroadened and essentially unobservable in thetruncated complex.
NMR spectra of tailed and truncated complexesare also notable for complexation shifts of side-chains at the protein–DNA interface. (i) Partialintercalation of the I13 cantilever side-chain isassociated with an upfield shift of its methylresonances (spectra v and vi in Figure 7(b)).55,56
The resonance position of I13 d-CH3 (which insertsmost deeply in the double helix)42 is nearK1.2 ppmin both complexes; the precise shift is slightly moreupfield in the tailed complex (Dd 0.04 ppm; verticallines between spectra v and vi). The slightattenuation of the I13 complexation shift is similarin magnitude to that observed among imino DNAresonances. (ii) Resolved probes of the major wing(formed by a-helices 1, 2 and proximal portion ofhelix 3) are provided by the exchangeable indoleNH resonances of Trp side-chains (W15, W43 andW52; Figure 7(c), scheme vii). The indole resonancesof the free domain are shown in Figure 7(c)(spectrum viii) and in the two complexes inFigure 7(a) (right-hand portion of spectra ii andiii). Of these side-chains, only W43 contacts theDNA backbone: the ring lies in a crevice with itsedge near the site of I13 insertion.42 In both tailedand truncated complexes the indole resonanceof W43 is shifted downfield by 1 ppm on DNAbinding (spectra ii and iii). Together, resolved NMRprobes provided by DNA imino resonances andselected interfacial side-chains demonstrate thatpartial truncation of the tail does not alter the modeof DNA recognition. 2D-NMR studies of tailed andtruncated complexes further indicate that therespective bound structures of the HMG box aresimilar.
The SRY tail acts as a kinetic clamp
Modest enhancement of the specific DNA affinityof the tailed SRY domain (relative to the truncateddomain) may in principle reflect faster association,

Figure 7. NMR studies of SRY–DNA complexes. (a) 600 MHz 1H-NMR spectra of site 1 DNA iminoresonances (12–14 ppm) andTrp indole NH resonances(10–11 ppm) at 25 8C. (i) Spectrumof free DNA (12 bp; sequenceinset); assignments are as indi-cated. Broadening of G1 and G12terminal resonances is due to DNAfraying. (ii) and (iii) Correspondingspectral regions of specific 1:1complex with intact (ii) or trun-cated (iii) tails exhibit large com-plexation shifts. A similar patternof imino resonances in the twocomplexes indicates similarity ofstructures. A trend toward smallercomplexation shifts is observed inthe truncated complex (verticallines; see Supplementary Data).The arrow indicates the anomalousupfield position of T5 at a site of I13insertion. Asterisks in spectrum (ii)near 12.8–13.0 ppm highlightbroad resonances of G1 and G12and a sharp resonance of G3 in thepresence of intact tail; in thetruncated complex these reson-ances are further broadened. Trpindole NH resonances in the twocomplexes (10–11 ppm) exhibitessentially identical chemical shiftsand relative linewidths. Theasterisk in spectrum (ii) near10 ppm indicates a novel exchange-able resonance assigned to para-OH
of Y74 (see Figure 2(c)), not observed in the truncated complex. (b) Upfield region of 1H-NMR spectra containingaliphatic resonances, primarily in protein. (iv) The spectrum of free protein contains few up-field shifted methylresonances. DSSwas used as a chemical-shift standard; the broken line indicates region upfield of acquisition. (v) and (vi)Corresponding spectra of protein–DNA complexes with intact (v) or truncated (vi) tails exhibit similar patterns ofup-field shifted aliphatic spin systems, including I13 (inserted between DNA base-pairs), L46 (near W15 and W43 in thecore of the major wing) and V5 (near H65, Y69 and Y72 in the mini-core of the minor wing). The complexation shift of I13d-CH3 is slightly attenuated in the truncated complex (dotted vertical lines). (c) Ribbon model of the free domainshowing Trp side-chains (vii) and corresponding indole NH resonances (viii); (ix) NOESY spectrum of the free taileddomain contains prominent NOEs from L46 methyl groups to W43 indole NH; no other Trp-methyl NOEs are observed.The spectrum was recorded at 25 8C at 600 MHz with mixing time of 200 ms.
A Kinetic Clamp in SRY Augments DNA Bending 183
slower dissociation, or effects on both rates.Previous NMR titration studies of SRY–DNAbinding established that dissociation is slow onthe NMR timescale.55 Exchange cross-peaksbetween bound and free DNA resonances arereadily observable at 40 8C in NOESY spectraat intermediate stoichiometries.33,51,55 Whereasbinding of the truncated domain remains in theslow-exchange region, exchange cross-peaks areaccentuated, qualitatively indicating more rapiddissociation of the truncated complex. Dissociationrates of the protein–DNA complexes were thereforecharacterized using gel and FRET-based kineticassays. GMSAwas adapted to monitor competitionbetween the labeled DNA site and a 100-fold excessof unlabeled DNA competitor containing the same
target sequence. At equilibrium the presence of thecompetitor essentially eliminates radiographicdetection of the shifted band. Respective complexeswere formed at 4 8C in the absence of competitor;competitor DNA was then added. Aliquots wereloaded on to a running gel (Figure 8(a) and (b)). Ineach gel lane a contains the free DNA probe andlane b the respective SRY–DNA complex prior toaddition of competitor DNA. Whereas for thetruncated complex competition is essentially com-plete within 24 s (lane k in Figure 8(b)), the tailedcomplex remains detectable for 29 min (lanes c–f inFigure 8(a)). Dissociation rates were calculatedbased on the time course of competition(Figure 8(c)). The off-rate of the tailed complex is0.002 sK1 under these conditions. For the truncated

Figure 8. Kinetic studies of SRY. (a) and (b) Kinetic GMSA of tailed SRY–DNA complex (a) and truncated complex (b)at 4 8C following addition of 100-fold excess unlabeled competitor DNA. Gel lanes show selected time intervals asindicated: (a) lane a, free probe; b, preformed tailed complex at time 0; c, 21 s; d, 2.5 min; e, 5 min; f, 12.5 min; g, 30 min; h,40 min; i, 60 min. (b) Lane a, free probe; b, preformed truncated complex at time 0; k, 24 s; l, 2 min;m, 3 min; n, 6.3 min; o,18 min; p, 30 min; q, 50 min. (c) Plot of natural logarithm of the ratio of bound DNAt to bound DNAt(0) as a function oftime; the off-rate koff is given by the slope.45 Blue squares and the blue fitted line represent the tailed complex; red circlesand the red fitted line represent the upper limit to off-rate of the truncated complex, based on the assumption thatapproximately 25% of the shifted complex remained at the initial 24 s time point. (d) and (e) Stopped-flow FRET assay.(d) Stopped-flow apparatus coupled to the fluorimeter and schematic representation of the experimental design.Dissociation is achieved by rapidly mixing pre-formed complex (in one syringe) with a 20-fold excess of unmodified12 bp DNA (final mixed stoichiometries) containing the same target site (from other syringe). (e) Representative data at15 8C showing the time-dependent increase in donor fluorescence due to dissociation of FRET-labeled SRY–DNAcomplexes: tailed complex (blue) and truncated complex (red). Dissociation reactions were monitored for 40 (truncated)and 120 s until equilibrium was reached. Off-rates were estimated based on initial slopes. Kinetic GMSA and stopped-flow FRET studies employed site 1 (Figure 3(d)).
184 A Kinetic Clamp in SRY Augments DNA Bending
domain an upper limit on the off-rate is obtained,O0.042 sK1. The ratios of these rates demonstratethat binding of the tail retards dissociation of theSRY–DNA by O20-fold. Since the DNA-bindingaffinity (Kd) of the tailed domain is !3-fold greaterthan that of the truncated domain under the sameconditions, the tail must also retard the associationrate but to a lesser extent.
To further characterize dissociation rates, astopped-flow kinetic FRET assay was developedas described in Materials and Methods(Figure 8(d)). Tailed or truncated domains werecomplexed with the same double-labeled DNAduplex as employed in tr-FRET studies (above).Use of a 15-bp DNA site restricts binding to thehigh-affinity 1:1 complex as verified by GMSA (seecontrol lane in Figure 6(f)). As illustrated inFigure 8(e), the increase in donor fluorescence
with time reflects the dissociation rate.65 Rateconstants (kobs) were estimated by the initial slopesin five to seven replicates. Under these conditionsthe truncated complex dissociates markedly morerapidly than does the wild-type domain; koffZ0.045(G0.007) sK1 for the tailed complex and0.90(G0.12) sK1 for the truncated complex.Although specific affinities are similar at 4 8C and15 8C as assessed by GMSA, off-rates at 15 8C aresignificantly faster than those observed by GMSA at4 8C. The ratio of FRET-defined off-rates is 20G5 inaccord with the limiting electrophoretic estimate.Dissociation constants pertaining to the bindingof the tailed or truncated domains to the same(donor, acceptor)-labeled 15 bp DNA site at 15 8Cwere measured in the same buffer by steady-stateFRET titration (Supplementary Data). RespectiveFRET-derived dissociation constants are 21.0(G

A Kinetic Clamp in SRY Augments DNA Bending 185
0.1) nM (tailed complex) and 37.0(G0.1) nM (trun-cated complex). These values (higher than theapparent SRY–DNA dissociation constants observedby GMSA in Figure 3 due to the shorter length ofthe FRET-labeled DNA) imply respective on-ratesof 2.1(G0.3)!106 MK1 sK1 (tailed complex) and2.4(G0.3)!107 MK1 sK1 (truncated complex).Truncation of the tail thus accelerates both associa-tion and dissociation, and to similar extents.
Discussion
Sequence-specific architectural transcriptionfactors contain clusters of basic residues C-terminalto the HMG box (Figure 2). Disordered in the freeprotein, this tail has dual functions in nuclearimport43,44,60,66,67 and DNA binding.37,42,47
The contribution of a basic tail to DNA bendingwas first visualized in the LEF-1/DNA complex(Figure 1(b)): although not well ordered within thecompressed major groove, the basic side-chains areinterposed between negative phosphodiestercharges.37 The LEF-1 tail was thus proposed toalleviate electrostatic repulsion that would other-wise be incurred at the crux of the bent DNA site.This mechanism is supported by the markedlylower affinity and reduced bending observed ontruncation of the LEF-1/TCF tail.47 Here, we soughtto test the contribution of the analogous SRY tail tospecific DNA binding and DNA bending. Whereaspartial truncation of the SRY tail is compatible withhigh-affinity DNA binding and bending, our resultshighlight the contribution of the SRY tail to thekinetic stability of the complex.
SRY tail functions as a kinetic clamp
The SRY-related and LEF-1 families differ in DNAsequence specificity, extent of DNA bending, andpattern of basic residues in respective tails. Whereasthe LEF-1 tail folds back across the major groovenear the center of the DNA bend, the SRY tailremains in the minor groove (Figures 1(a) and2(c)).42 Such structural differences, which precludeanalogous electrostatic roles, may account for themore shallow bend angle of the SRY complex as theSRY-directed bend angle is similar to that of atruncated LEF-1 complex.37,47,48,68–70 Because theextent of major-groove compression is less markedin the SRY complex, its tail is apparently not neededat the crux to alleviate electrostatic repulsion.Accordingly, the C-terminal portion of the SRY tailonly modestly enhances specific DNA binding andDNA bending. FRET analysis corroborates differ-ences in mean bend angles; although the bentcomplexes exhibit a range of conformationalsubstates, giving rise to a distribution of DNAbend angles. From the dynamic perspective, thecomplexes exhibit significant long-range fluctu-ations that are fast on the millisecond NMRtimescale and slow on the nanosecond fluorescencetimescale. Binding of the tail does not constrain the
amplitude of such fluctuations, but an effect on theirtimescale cannot be excluded.Surprisingly, kinetic studies of protein–DNA
dissociation demonstrate that binding of the tailretards dissociation by 20-fold, which is dispropor-tionate to the twofold change in affinity. The SRYtail thus retards both association and dissociation,the latter to a greater extent. The estimated on-rateat 15 8C is at least 100-fold slower than the diffusionlimit, presumably due to kinetic steps required forDNA reorganization. On-rates slower than thediffusion limit are a general feature of DNA-bending proteins: the estimated on-rates of thetailed LEF-1 domain (1.1!105 MK1 sK1)45 and theTATA-binding protein TBP (1.66!105 MK1 sK1)71 at20 8C are tenfold slower than that of the tailed SRYdomain (2.1!106 MK1 sK1) at 15 8C. These proteinsnevertheless exhibit different kinetic lifetimes: bothLEF-1–DNA complex (off-rate 1.1!10K4 sK1)45 andTBP (off-rate 7!10K4 sK1) dissociate more slowly,commensurate with their higher specific DNA-binding affinities (Kd 1 and 4 nM, respectively).Although among these proteins relative kineticstabilities mirror relative DNA bend angles, it isnot known in general whether more highly bentDNA–protein complexes would tend to exhibitlonger lifetimes.The kinetic importance of DNA reorganization is
suggested by profound differences betweenbinding of HMG1 domains to unmodified (andstraight) DNA or cis-platin-distorted (and bent)DNA adducts. Such adducts in part mimic thebound state of HMG box complexes; furtherbending of the cis-platin-DNA adduct occurs onprotein binding. Surface plasmon resonance studiesof HMG1 and HMG2 (each containing two HMGbox domains) indicate that bidentate binding to a30 bp oligonucleotide duplex exhibited kineticrate constants of 2!104 MK1 sK1 (kon) and w6–7!10K2 sK1 (koff).
72 Stopped-flow kinetic studies of thebinding of the HMG-1A domain to cis-platin-modified DNA showed that the association rate isnear the diffusion limit. Similarly, dissociation ratesof cis-platin-modified DNA complexes are signifi-cantly faster than those of the unmodified DNAcomplexes.65 The high enthalpies of activationobserved in studies of TBP (ca 30 kcal/mol)73 andsuggested here indicate that dissociation requirespartial unfolding of the protein or DNA or both inthe transition state.
Biological importance of the SRY tail
The marked acceleration of protein–DNAdissociation on truncation of the tail suggests thatthis structure acts as a kinetic clamp to prolong thelifetime of the complex. We propose that thislifetime, independent of the equilibrium constant,is a critical parameter in SRY-mediated transcrip-tional regulation. Transcriptional initiation underkinetic rather than thermodynamic control has beenproposed for the TBP–TATA complex formation.73
Kinetic control of biological processes has been

186 A Kinetic Clamp in SRY Augments DNA Bending
observed in diverse pathways, from the mitogenicsignaling pathway of insulin74 to DNA replica-tion.75 Analogous kinetic principles occur inpharmacology: the anti-cancer potency of DNAintercalating agents, mediated by inhibition oftranscriptional elongation, depends on the lifetimeof the intercalative DNA complex.76,77 By analogy,we imagine that the kinetic stability of a strategicSRY-induced DNA bend in turn enables theassembly of a multi-protein–DNA complex(Figure 1(c)). Recent studies of chimeric Srytransgenes in XX mice23,24 suggest by contrast thatthe modest effects of the tail on specific DNAaffinity and DNA bending are unlikely to be ofregulatory significance. The extent of DNA bendingin the truncated complex is similar to that of thenative murine SRY–DNA complex. Since the HMGboxes of human and murine SRY are equallycompetent to induce sex reversal in transgenicmice,24 the lower bend angle in the truncatedcomplex is likely to fall within a functional rangeof values.33 Similarly, the modest decrement inspecific DNA binding observed on truncation of thetail is similar to the decrements observed in bindingof murine SRY to some SRY target sites.32,33
Maintenance of an SRY complex through mul-tiple rounds of transcriptional initiation would beexpected to enhance potency of transcriptionalactivation. This hypothesis is supported by pre-vious studies of an SRY variant containing thechimpanzee-specific cantilever substitution I13F.51
The aromatic cantilever prolongs the lifetime of thetailed SRY complex to an extent similar to itsenhanced transcriptional activation.12 The canti-lever substitution does not perturb equilibriumproperties (specific DNA affinity, DNA-bindingspecificity, DNA bending, and thermodynamicstability); cellular expression and nuclear locali-zation remain unchanged.51 We suggest a model ofkinetic regulation wherein the net accumulation ofmRNA is controlled by the rate of SRY dissociationfrom an activated pre-initiation complex. Thisperspective suggests that the length and sequenceof the SRY tail has been conserved amongstmammalian SRY and SOX proteins by the dualconstraints of nuclear localization and kineticstabilization. A further constraint may be imposedby P300-mediated acetylation of K81 (residue 136 inintact SRY) proposed to regulate nuclear import.78
The biological importance of the SRY tail isdemonstrated by human mutations associatedwith partial or pure gonadal dysgenensis.79–82
Role of tail in DNA bending
The present studies of SRY-directed DNAbending demonstrate that the tail augments DNAbending. The similar bend angles induced by thetailed domain and full-length human SRY indicatethat inclusion of additional N or C-terminalsequences does not further enhance DNA bending.The present PGE results rationalize the lower DNAbend angles observed in the two NMR structures
relative to previous PGE studies20,83 and FRETanalysis:51 the variant DNA site employed in thestructural studies (5 0-TTTGTG-3 0 and complement)is markedly less bent than a consensus target site(5 0-ATTGTT-3 0 and complement). Using identicalgel systems (10% polyacrylamide with a 58:1 ratioof acrylamide to bis-acrylamide) and core targetsites imbedded in equivalent sequence contexts(site 1 versus site 4 in Figure 3(d)), the decrement inbend angle is 118 (798 versus 688; lines 6 and 9 inTable 2). Although estimating the DNA bend angleis difficult in the original SRY–DNA structure due tothe short length of the duplex, the apparentlyshallower bend angle38 is consistent with themarked effect of partial tail truncation on bendingof this variant DNA site: 498 (line 9 in Table 2),leading to a decrement of 308 relative to a consensustailed complex. It is possible that the NMR-definedbend angle was further attenuated by end effectsdue to the short DNA site employed (8 bp).38
NMR features of tailed and truncated SRY–DNAcomplexes with a 12 bp consensus site (site 1 inFigure 3(d)) are broadly similar. Despite the 108difference in mean DNA bend angle as inferredfrom PGE (Table 2B), imino 1H-NMR spectra of thebound DNA exhibit an almost identical pattern ofcomplexation shifts. Since these shifts reflectchanges in ring currents of neighboring bases (dueto changes in roll, twist, and propeller twist withinthe double helix) and possible changes in thestrength of Watson–Crick hydrogen bonds (due tochanges in base-pair geometry), this shared patternsuggests that local distortions induced in DNAstructure by the tailed or truncated domains arealmost identical. The truncated complex none-theless exhibits a subtle trend toward attenuatedcomplexation shifts. The distributed nature of thesechanges suggests that augmented bending resultsfrom multiple small effects rather than introductionor reorganization of a single DNA kink. Wespeculate that the slight attenuation of complexa-tion shifts reflects in part enhanced flexibility.
Whereas the present NMR parameters (chemicalshifts, linewidths and NOEs) represent local para-meters, distance distribution analysis provides twoglobal parameters: the mean and width of thedistribution.61 In the free DNA this distance is ca62 A at 15 8C (Table 1B) consistent with thecanonical structure of B-DNA (including effects ofthe linkers). Binding of the tailed domain leads to adramatic reduction in end-to-end distance (51.5 A)whereas the effect of the truncated domain is lessmarked (53.8 A). The length reduction (Dl) in thetruncated complex (Dl 7.9(G0.3) A) is thus only ca78% of that seen in the tailed complex (Dl 10.2(G0.3) A). Because end-to-end distances are influ-enced by both DNA bending and DNA unwinding,however, it is not possible to rigorously interpretthese distances in relation to DNA bend angles. Itwould be of future interest to extend the presentFRET studies by permuting the position of the coretarget site within the 15 bp DNA duplex. Byanalogy to PGE,53,84 the relationship between

A Kinetic Clamp in SRY Augments DNA Bending 187
FRET efficiency and “flexure displacement” mayenable the effects of DNA bending and DNAunwinding to be distinguished.
The FRET-derived distribution of end-to-enddistances (FWHM values; Table 1B) is broadenedin both complexes relative to the free DNA. Thewidth of the free distribution is consistentwith mobility of the two linkers withoutadditional contribution from possible DNA flexi-bility61,85 (K. Srinivasan & W. K. Olson, personalcommunication). Although bending can in itselfcause an increase in the apparent width due togeometric factors, the observed FWHM values aresufficiently large as to suggest that a range of DNAbend angles are populated in solution. Since NMRspectra contain a single set of protein and DNAresonances, transitions between distinct bentconformers must be fast on the timescale of NMRchemical shifts (!1 ms). Since no intramoleculardiffusion could be detected in the FRET exper-iments,86 global fluctuations must also be slowerthan the lifetime of the excited state of the donor, i.e.slower than 5 ns. Such fast exchange rationalizeswhy electrophoretic methods give rise to a singlewell-defined band and not multiple shifted 1:1complexes or diffuse tails. These observationssuggest that DNA bend angles observed in crystalstructures or inferred from simulated-annealingmodels may under-estimate the extent of flexibilityin solution.
Concluding remarks
The HMG boxes of SRY and LEF-1 are eachextended by C-terminal basic tails that provide bothnuclear localization signals43,44,60,78 and accessoryDNA-binding elements.37,42,46,48 Although the SRYand LEF-1 tails differ in sequence andmode of DNAbinding, in each case they significantly modifycritical aspects of the bent protein–DNA complexes.Dual use of a basic peptide segment in twounrelated processes represents an elegant economyof protein design. Mutations in the SRY tail areassociated with human sex reversal due to impair-ment of one or both of these functions. It would beof future interest to investigate whether suchmutations may impair the ability of the tail tofunction as a kinetic clamp without affectingnuclear localization or equilibrium DNA-bindingproperties. Correlation between phenotype and thelifetime of an SRY–DNA complex could provideevidence for the kinetic regulation of transcriptionalinitiation by a strategic DNA bend.
Materials and Methods
Plasmids
The full-length human SRY coding region was clonedby PCR and ligated into pET-15b (Novagen, Inc.). Thecloning site contains codons for six histidine residues andthrombin site following initiator methionine as described
by the vendor. Substitutions were introduced into thecoding region in phage M13mp19RF by oligonucleotide-directed mutagenesis, and mutants were cloned into anexpression plasmid.51,87 Constructions were verified byDNA sequencing.
Protein purification
The tailed SRY HMG box (residues 2–85 in HMG-boxconsensus) and truncated fragment (residues 2–78) wereexpressed in Escherichia coli strain BL21(DE3)pLysS asthrombin-cleavable fusion proteins (with staphylococcalnuclease; SN).51,87 Purity was O98% as assessed by SDS-polyacrylamide gel electrophoresis. Results of massspectrometry were in accord with expected values.Fragments contain two additional N-terminal residues(GK1 and S0) derived from thrombin site in fusionprotein.55 Full-length SRY was purified by passage overNi-NTA resin (Qiagen, Inc.).88 The protein was elutedfrom the column in a buffer containing 6 M guanidine–HCl and 20 mM Tris–HCl (pH 8.0) and dialyzed into150 mM NaCl, 20 mM sodium phosphate (pH 7.0), 1 mMEDTA, and 3 mM DTT. DNA bending assays wereperformed without cleavage of histidine tag.
DNA-binding assay
The 36 bp DNA sites (5 0-CATACTGCGGGGGTGATTGTTCAGGATCATACTGCG-3 0 and complement)containing consensus sequence ATTGTTwere purchasedfrom Oligos, Etc. (Wilsonville, OR). One strand waslabeled with [g-33P]ATP (ICN Biomedicals, Inc.). Theduplex probe was annealed, and analyzed by GMSA asdescribed.55,57 Each reaction contained 1–32 nM proteinand less than 1 nM labeled DNA in 10 mM potassiumphosphate (pH 7.0), 30 mg/ml bovine serum albumin(BSA), 50 mM KCl, 2.5 mM MgCl2, 5% (v/v) glycerol and4 mM dithiothreitol; the reaction was incubated in 12.5 mlreactions for 1 h on ice. Only specific binding is observedunder these conditions at protein concentrations tested.0.45X TBE/polyacrylamide gels (8%) were pre-run for45–60 min at 10 V/cm at 4 8C. TB buffer is the samewithout EDTA. After sample loading, the gel was run at15 V/cm for 1 h at 4 8C. 1x TBE electrophoresis bufferconsists of 100 mM Tris base, 100 mM boric acid, and2 mM EDTA (pH 8.0 as tetrasodium salt).
DNA-bending assay
DNA sites containing a consensus SRY binding site 5 0-ATTGTT-3 0 (sites1–4 inFigure3)wereclonedbetweenXbaIand SalI sites of pBend2.52 Probes of equal length, with thebinding site at varying distance from the ends weregenerated by PCR, and 5 0-labeled with [g-33P]ATP and T4polynucleotide kinase. 10 ml binding reactions contained50 mMKCl, 20 mMTris–HCl (pH 7.4), 5 mMMgCl2, 50 ngpoly(dI-dC), 10% glycerol, approximately 1 nM 33P-labeled DNA probe and purified SRY–HMG box protein(60 nM).ToassessreproducibilityofPGEestimatesofDNAbend angle in different gels of same composition, thewild-type tailed domain was assayed six times in the 10%/29:1system. Inferred angles were 77.88, 78.58, 78.68, 79.28, 79.28,and 80.58, givingmean value of 798 and standarddeviationof G18. Absolute calibration of PGE-based estimates ofDNAbendangles isnotwellestablishedinrelationtothree-dimensional structures. A DNA bend angle of 54.0(G1.7)8was observed in the NMR structure (Figure 1(a)) ofa specific complex with a variant DNA site (5 0-

188 A Kinetic Clamp in SRY Augments DNA Bending
GTGTTTGTGCA-3 0 andcomplement; core siteunderlinedwith non-consensus bases in bold).42 The present PGEanalysis of this site yields an estimate of 678 in an 8% gelsystem containing a 29:1 ratio of acrylamide to bis-acrylamide.
Electrophoretic assay of dissociation kinetics
Stabilities of the tailed and truncated SRY–DNAcomplexes were determined by GMSA in the presence of100-fold excess unlabeled competitor DNA. Respectiveproteins were incubated with 0.7 nM 33P-labeled 36-mercontaining the SRY binding site in 140 mM KCl, 10 mMpotassium phosphate and 10 mM Tris–HCl (pH 8.4) with8% glycerol, 33 mg/ml BSA, and 1 mM DTT. Afterincubation on ice for 1 h, a 100-fold excess of cold DNAwas added, and 15 ml aliquots were loaded onto a running8% TB/gel at 36 different time intervals for each protein.Initial time intervals were 10 s apart; intervals wereprogressively lengthened until samples were loaded at10 min intervals. The gel was loaded over 70 min.Dissociation rateswere determined by plotting the naturallogarithm of ratio of bound DNAt to bound DNAt0 as afunction of time with koff equaling the slope of the line.45
Circular dichroism
Spectra were obtained using a spectropolarimeter (AvivBiomedical, Lakewood, New Jersey) equipped withthermister control for automated analysis of thermalunfolding. The samples were made 30 mM in 140 mMKCland 10 mM potassium phosphate (pH 7.4) in a 1 mmpath-length quartz cuvette.
Methylphosphonate interference assays
GMSA of 12 bp DNA duplex probes containing singlemethylphosphonate modifications in either top or bottomstrands were performed with either tailed or truncatedSRY domains. Oligonucleotides each containing a singlemethylphosphonate modification were synthesized byOligos Etc., Inc. (Wilsonville, OR).
Fluorescence spectroscopy
Spectra were measured using an Aviv model ATF 105Fluorometer (Aviv Biomedical, Lakewood, New Jersey).For FRET studies excitation and emission wavelengthswere 490 nm and 520 nm, respectively, for the donor, and560 nm and 585 nm for the acceptor. The quantum yieldof the donor was determined for each sample using areference solution of fluorescein in 0.1 M NaOH (0.85).89
The calculated quantum yield of the DNA-coupled donorwas 0.83 at 4 8C for the single-labeled free DNA andvaried between 0.8 and 0.9 for all other samples. Forsterdistance, Ro, was calculated accordingly (Table 1B; 58.3 Aat 4 8C for the free DNA). For protein denaturationstudies, guanidine titrations were monitored at 4 8C byintrinsic Trp fluorescence as measured at emissionwavelength 390 nm following excitation at 270 nm. Forsuch studies tailed and truncated domains were made1 mM in 140 mM KCl and 10 mM potassium phosphate(pH 7.4) in the titrating cuvette. The same concentrationof SRY was used in the titrant reservoir containing 7.2 Mguanidine–HCl in the above buffer. Experimental curveswere fitted to a two-state model as described.51,90
Fluorescence control experiments to verify the
predominance of an intramolecular FRET mechanismwere as described33 (Supplementary Data).
FRET sample preparation
Studies of DNA bending employed a 15 bp DNAduplex (5 0-TCGGTGATTGTTCAG-3 0 and complement;target site underlined). Cantilever insertion is between bp8 and 9. To ensure mobility of probes, fluorophores weretethered to DNA by flexible linkers. Three sampleswere prepared as described33: (i) 6-FAM tethered to the5 0-phosphate of the upper strand by a hexanyl linker;(ii) TAMRA tethered to the 5 0-phosphate of the lowerstrand by an analogous linker; and (iii) a double-labeledderivative. DNA probes were made 5 mM in 10 mMpotassium phosphate and 10 mM Tris–HCl (pH 8.4)containing 140 mM KCl, 1 mM EDTA and 1 mM DTT.
Steady-state FRET efficiencies
Steady-state measurements were conducted at 25 8Cusing an AVIV spectrofluorimeter (model ATF-105). Theexcitation frequency was 490 nm. FRET efficiencies werecalculated by the fluorescence intensity at 520 nm of thedoubly labeled DNA relative to the singly labeled 6-FAM-DNA adduct (i.e. containing only the donor). Calculationswere done after correction for differences in concen-trations of labeled DNAs. The small enhancement insteady-state FRETefficiency of the tailed complex relativeto the truncated complex was corroborated by time-resolved analysis of fluorescence lifetimes and supportstheir interpretation in relation to excitation transfer.
Time-resolved fluorescence measurements
A home-built system at Bar Ilan University employstime-correlated single-photon counting as described.33
The emission wavelength was selected by a double0.125 m subtractive monochromator. The donor emissionwas collected at 520 nm. The acceptor emission wascollected at 585 nm. The response of the system yielded apulse width of 35 ps. FRET studies were conducted bothat 15 8C and 4 8C.
Global analysis of tr-FRET
Distance distributions were obtained from globalanalysis of experimental decay curves using Marquardtnon-linear least-squares.91 Four (or two) theoretical decaycurves were calculated for each set of FRET experimentsand fitted to corresponding experimental curves.91 Thesignificance of inferred parameters was assessed based onfour indicators: global c2 values; distributions ofresiduals; autocorrelation of the residuals; and errorintervals of the calculated parameters.91
Fluorescence titration for Kd determination
Steady-state FRET-based titration experiments wereused to determine the dissociation constants (Kd) of tailedand truncated complexes at 15 8C. Varying SRY concen-trations were titrated in the FRET buffer described abovewith 25 nM of the 15 bp DNA doubly labeled with 6-FAMand TAMRA. Emission spectra were recorded from500 nm to 650 nm after excitation at 490 nm. Kd valueswere determined by plotting change in fluorescenceintensity at 520 nm against total protein concentration.

A Kinetic Clamp in SRY Augments DNA Bending 189
Data were fit with a single-site ligand-binding equation(Erithacus Software).
Stopped-flow kinetic FRET studies
Competition stopped-flow FRET studies were per-formed with an Aviv double-mixing stopped-flowapparatus with a thermo-electric temperature controller.Data were obtained with an Aviv ATF 105 spectro-fluorometer. Kinetic experiments were carried out byplacing in one syringe a 1:1 complex of 1 mM of SRYand a15 bp duplex (in FRET buffer) containing DNA site 1(Figure 3) labeled at respective 5 0 ends with donor andacceptor. Displacement was achieved by rapidly mixingcomplex with 20-fold excess (final stoichiometries) ofunlabeled 12 bp DNA with same consensus site from asecond syringe. In labeled SRY complex donor emission isreduced by FRET; on mixing with excess unlabeled DNA,donor emission increases as protein leaves the preformedcomplex and binds to competitor DNA. Fluorescenceemission was monitored at 520 nm following excitationat 490 nm. The initial slope of each trace was determinedto obtain observed rate constants. The reported koff isthe average of five to seven traces; kon values werecalculated from observed koff and Kd values accordingto: KdZkoff/kon.
NMR spectroscopy
Spectra of free domains were observed at 25 8C in10 mM deuterated acetic acid (pH 4.5) and 140 mM KCl;such spectra are similar in the pH range 4–8. Spectra ofcomplexes were obtained at 40 8C in 10 mM potassiumphosphate (pH 6.0) and 50 mM KCl. Assignments wereobtained as described.38,51,55,92
Acknowledgements
We thank H. Chen, C.- Y. King; C. Kung, andE. Rivera for participation in early stages of work;N. Narayana for statistical analysis of SRY struc-tures; P. K. Donahoe, Q. X. Hua, D. N. Jones, C.- Y.King, and L. A. Labeots for discussion; and E.Collins and S. Price for preparation of manuscript.This work was supported in part by an equipmentgrant from the Israel Science Foundation to E.H.(number 553-99), a grant from the United States/Israel Binational Foundation (to E.H. and M.A.W.;number 98–362) and in its early stages by theNational Institutes of Health (to M.A.W.;GM051558). These studies are a contribution fromthe Cleveland Center for Structural Biology.
Supplementary Data
Supplementary data associated with this article,comprising seven Figures giving CD, fluorescence,and NMR data and providing additional moleculargraphics; description of fluorescence controlstudies; and three Tables summarizing DNAbend angles, fluorescence parameters, and NMR
chemical shifts, can be found, in the online version,at doi:10.1016/j.jmb.2006.01.060
References
1. Goodfellow, P. N. & Lovell-Badge, R. (1993). SRYand sex determination in mammals. Annu. Rev. Genet.27, 71–92.
2. Sinclair, A. H., Berta, P., Palmer, M. S., Hawkins, J. R.,Griffiths, B. L., Smith, M. J. et al. (1990). A gene fromthe human sex-determining region encodes a proteinwith homology to a conserved DNA-binding motif.Nature, 346, 240–244.
3. Gubbay, J., Collignon, J., Koopman, P., Capel, B.,Economou, A., Munsterberg, A. et al. (1990). A genemapping to the sex-determining region of the mouseY chromosome is a member of a novel family ofembryonically expressed genes. Nature, 346, 245–250.
4. Ner, S. S. (1992). HMGs everywhere. Curr. Biol. 2,208–210.
5. Koopman, P., Gubbay, J., Vivian, N., Goodfellow, P. &Lovell-Badge, R. (1991). Male development of chro-mosomally female mice transgenic for Sry. Nature,351, 117–121.
6. Berta, P., Hawkins, J. R., Sinclair, A. H., Taylor, A.,Griffiths, B. L., Goodfellow, P. N. & Fellous, M. (1990).Genetic evidence equating SRY and the testis-determining factor. Nature, 348, 448–450.
7. Wegner, M. (1999). From head to toes: the multiplefacets of Sox protein. Nucl. Acids Res. 6, 1409–1420.
8. Schilham,M.W., Oosterwegel, M. A., Moerer, P., Ya, J.,de Boer, P. A., van deWetering, M. et al. (1996). Defectsin cardiac outflow tract formation and pro-B-lympho-cyte expansion in mice lacking Sox-4. Nature, 380,711–714.
9. Pevny, L. H. & Lovell-Badge, R. (1997). Sox genes findtheir feet. Curr. Opin. Genet. Dev. 7, 338–344.
10. Koopman, P. (1999). Sry and Sox9: mammalian testis-determining genes. Cell. Mol. Life Sci. 55, 839–856.
11. Sekido, R., Bar, I., Narvaez, V., Penny, G. & Lovell-Badge, R. (2004). SOX9 is up-regulated by thetransient expression of SRY specifically in Sertoli cellprecursors. Dev. Biol. 274, 271–279.
12. Haqq, C. M., King, C. Y., Ukiyama, E., Falsafi, S.,Haqq, T. N., Donahoe, P. K. & Weiss, M. A. (1994).Molecular basis of mammalian sexual determination:activation of Mullerian inhibiting substance geneexpression by SRY. Science, 266, 1494–1500.
13. Foster, J. W. & Graves, J. A. (1994). An SRY-relatedsequence on the marsupial X chromosome: impli-cations for the evolution of the mammalian testis-determining gene. Proc. Natl Acad. Sci. USA, 91,1927–1931.
14. Wagner, T., Wirth, J., Meyer, J., Zabel, B., Held, M.,Zimmer, J. et al. (1994). Autosomal sex reversal andcampomelic dysplasia are caused by mutations inand around the SRY-related gene SOX9. Cell, 79,1111–1120.
15. Harley, V. R., Lovell-Badge, R. & Goodfellow, P. N.(1994). Definition of a consensus DNA binding site forSRY. Nucl. Acids Res. 22, 500–501.
16. Mertin, S., McDowall, S. G. & Harley, V. R. (1999). TheDNA-binding specificity of SOX9 and other SOXproteins. Nucl. Acids Res. 27, 1359–1364.
17. Kamachi, Y., Cheah, K. S. & Kondoh, H. (1999). Themechanism of regulatory target selection by the SOX

190 A Kinetic Clamp in SRY Augments DNA Bending
HMG domain proteins as revealed by comparison ofSOX1/2 and SOX9. Mol. Cell. Biol. 19, 107–120.
18. Preiss, S., Argentaro, A., Clayton, A., John, A., Jans,D. A., Ogata, T. et al. (2001). Compound effects ofpoint mutations causing campomelic dysplasia/autosomal sex reversal upon SOX9 structure, nucleartransport, DNA binding, and transcriptional acti-vation. J. Biol. Chem. 276, 27864–27872.
19. Whitfield, L. S., Lovell-Badge, R. & Goodfellow, P. N.(1993). Rapid sequence evolution of the mammaliansex-determining gene SRY. Nature, 364, 713–715.
20. Ferrari, S., Harley, V. R., Pontiggia, A., Goodfellow,P. N., Lovell-Badge, R. & Bianchi, M. E. (1992). SRY,like HMG1, recognizes sharp angles in DNA. EMBO J.11, 4497–4506.
21. Mitchell, C. L. & Harley, V. R. (2002). Biochemicaldefects in eight SRY missense mutations causingXY gonadal dysgenesis. Mol. Genet. Metab. 77,217–225.
22. Pontiggia, A., Rimini, R., Harley, V. R., Goodfellow,P. N., Lovell-Badge, R. & Bianchi, M. E. (1994). Sex-reversing mutations affect the architecture of SRY–DNA complexes. EMBO J. 13, 6115–6124.
23. Bergstrom, D. E., Young, M., Albrecht, K. H. & Eicher,E. M. (2000). Related function of mouse SOX3, SOX9,and SRY HMG domains assayed by male sexdetermination. Genesis, 28, 111–124.
24. Lovell-Badge, R., Canning, C. & Sekido, R. (2002). Sex-determining genes in mice: building pathways. In TheGenetics and Biology of Sex Determination (Chadwick, D.& Goode, J., eds), Wiley, West Sussex, UK.
25. Harley, V. R., Lovell-Badge, R., Goodfellow, P. N. &Hextall, P. J. (1996). The HMG box of SRY is acalmodulin binding domain. FEBS Letters, 391, 24–28.
26. Poulat, F., Barbara, P. S., Desclozeaux, M., Soullier, S.,Moniot, B., Bonneaud, N. et al. (1997). The humantestis determining factor SRY binds a nuclear factorcontaining PDZ protein interaction domains. J. Biol.Chem. 272, 7167–7172.
27. Matsuzawa-Watanabe, Y., Inoue, J. & Semba, K.(2003). Transcriptional activity of testis-determiningfactor SRY is modulated by the Wilms’ tumor 1 geneproduct, WT1. Oncogene, 22, 7900–7904.
28. Oh, H. J., Li, Y. & Lau, Y. (2005). Sry associates with theheterochromatin protein 1 complex by Interactingwith a KRAB domain protein. Biol. Reprod. 72,407–415.
29. Ohe, K., Lalli, E. & Sassone-Corsi, P. (2002). A directrole of SRY and SOX proteins in pre-mRNA splicing.Proc. Natl Acad. Sci. USA, 99, 1146–1151.
30. Weiss, M. A. (2005). Molecular mechanisms of malesex determination: the enigma of SRY. In DNAConformation and Transcription (Ohyama, T., ed.), pp.1–15, Landes Bioscience, Georgetown, TX.
31. Bewley, C. A., Gronenborn, A. M. & Clore, G. M.(1998). Minor groove-binding architectural proteins:structure, function, and DNA recognition. Annu. Rev.Biophys. Biomol. Struct. 27, 105–131.
32. Giese, K., Pagel, J. & Grosschedl, R. (1994). DistinctDNA-binding properties of the high mobility groupdomain of murine and human SRY sex-determiningfactors. Proc. Natl Acad. Sci. USA, 91, 3368–3372.
33. Phillips, N. B., Nikolskaya, T., Jancso-Radek, A.,Ittah, V., Jiang, F., Singh, R. et al. (2004). Sry-directed sex reversal in transgenic mice is robust toenhanced DNA bending: comparison of human andmurine in HMG boxes. Biochemistry, 43, 7066–7081.
34. Read, C. M., Cary, P. D., Crane-Robinson, C., Driscoll,P. C. & Norman, D. G. (1993). Solution structure of a
DNA-binding domain from HMG1. Nucl. Acids Res.21, 3427–3436.
35. Weir, H.M., Kraulis, P. J., Hill, C. S., Raine, A. R., Laue,E. D. & Thomas, J. O. (1993). Structure of the HMGbox motif in the B-domain of HMG1. EMBO J. 12,311–319.
36. Verbeek, S., Izon, D., Hofhuis, F., Robanus-Maan-dag, E., te Riele, H., van de Wetering, M. et al.(1995). An HMG-box-containing T-cell factorrequired for thymocyte differentiation. Nature, 374,70–74.
37. Love, J. J., Li, X., Case, D. A., Giese, K., Grosschedl, R.& Wright, P. E. (1995). Structural basis for DNAbending by the architectural transcription factor LEF-1. Nature, 376, 791–795.
38. Werner, H.M., Huth, J. R., Gronenborn, A.M. & Clore,G. M. (1995). Molecular basis of human 46X,Y sexreversal revealed from the three-dimensional solutionstructure of the human SRY–DNA complex. Cell, 81,705–714.
39. Ohndorf, U. M., Rould, M. A., He, Q., Pabo, C. O. &Lippard, S. J. (1999). Basis for recognition of cisplatin-modified DNA by high-mobility-group proteins.Nature, 399, 708–712.
40. Murphy, F. V., Sweet, R. M. & Churchill, M. E. (1999).The structure of a chromosomal high mobility groupprotein–DNA complex reveals sequence-neutralmechanisms important for non-sequence-specificDNA recognition. EMBO J. 18, 6610–6618.
41. Allain, F. H., Yen, Y. M., Masse, J. E., Schultze, P.,Dieckmann, T., Johnson, R. C. & Feigon, J. (1999).Solution structure of the HMGprotein NHP6A and itsinteraction with DNA reveals the structural determi-nants for non-sequence-specific binding. EMBO J. 18,2563–2579.
42. Murphy, E. C., Zhurkin, V. B., Louis, J. M., Cornilescu,G. & Clore, G. M. (2001). Structural basis for SRY-dependent 46-X,Y sex reversal: modulation of DNAbending by a naturally occurring point mutation.J. Mol. Biol. 312, 481–499.
43. Sudbeck, P. & Scherer, G. (1997). Two independentnuclear localization signals are present in the DNA-binding high-mobility group domains of SRY andSOX9. J. Biol. Chem. 272, 27848–27852.
44. Li, B., Zhang, W., Chan, G., Jancso-Radek, A., Liu, S. &Weiss, M. A. (2001). Human sex reversal due toimpaired nuclear localization of SRY. A clinicalcorrelation. J. Biol. Chem. 276, 46480–46484.
45. Giese, K., Amsterdam, A. & Grosschedl, R. (1991).DNA-binding properties of the HMG domain of thelymphoid-specific transcriptional regulator LEF-1.Genes Dev. 5, 2567–2578.
46. Carlsson, P., Waterman, M. L. & Jones, K. A. (1993).The hLEF/TCF-1a HMG protein contains a context-dependent transcriptional activation domain thatinduces the TCRa enhancer in T cells. Genes Dev. 7,2418–2430.
47. Read, C. M., Cary, P. D., Preston, N. S., Lnenicek-Allen, M. & Crane-Robinson, C. (1994). The DNAsequence specificity of HMG boxes lies in the minorwing of the structure. EMBO J. 13, 5639–5646.
48. Lnenicek-Allen,M., Read, C.M. &Crane-Robinson, C.(1996). The DNAbend angle and binding affinity of anHMG box increased by the presence of short terminalarms. Nucl. Acids Res. 24, 1047–1051.
49. Weiss, M. A. (2001). Floppy SOX: mutual induced fitin HMB (High-Mobility Group) Box-DNA recog-nition. Mol. Endocrinol. 15, 353–362.

A Kinetic Clamp in SRY Augments DNA Bending 191
50. Teo, S. H., Grasser, K. D. & Thomas, J. O. (1995).Differences in the DNA-binding properties of theHMG-box domains of HMG1 and the sex-determin-ing factor SRY. Eur. J. Biochem. 3, 943–950.
51. Ukiyama, E., Jancso-Radek, A., Li, B., Milos, L.,Zhang, W., Phillips, N. B. et al. (2001). SRY andarchitectural gene regulation: the kinetic stability of abent protein–DNA complex can regulate its transcrip-tional potency. Mol. Endocrinol. 15, 363–377.
52. Kim, J., Zwieb, C., Wu, C. & Adhya, S. (1989). Bendingof DNA by gene-regulatory proteins: constructionand use of a DNA bending vector. Gene, 85, 15–23.
53. Wu, H. M. & Crothers, D. M. (1984). The locus ofsequence-directed and protein-induced DNA bend-ing. Nature, 308, 509–513.
54. Snyder, U. K., Thompson, J. F. & Landy, A. (1989).Phasing of protein-induced DNA bends in a recombi-nation complex. Nature, 341, 255–257.
55. King, C. Y. & Weiss, M. A. (1993). The SRY high-mobility-group box recognizes DNA by partialintercalation in the minor groove: a topologicalmechanism of sequence specificity. Proc. Natl Acad.Sci. USA, 90, 11990–11994.
56. Peters, R., King, C. Y., Ukiyama, E., Falsafi, S.,Donahoe, P. K. & Weiss, M. A. (1995). An SRYmutation causing human sex reversal resolves ageneral mechanism of structure-specific DNA recog-nition: application to the four-way DNA junction.Biochemistry, 14, 4569–4576.
57. Haqq, C. M., King, C. Y., Donahoe, P. K. & Weiss,M. A. (1993). SRY recognizes conserved DNA sites insex-specific promoters. Proc. Natl Acad. Sci. USA, 90,1097–1101.
58. De Santa Barbara, P., Bonneaud, N., Boizet, B.,Desclozeaux, M., Moniot, B., Sudbeck, P. et al. (1998).Direct interaction of SRY-related protein SOX9 andsteroidogenic factor 1 regulates transcription of thehuman anti-Mullerian hormone gene. Mol. Cell. Biol.18, 6653–6665.
59. Pontiggia, A., Whitfield, S., Goodfellow, P. N., Lovell-Badge, R. & Bianchi, M. E. (1995). Evolutionaryconservation in the DNA-binding and -bendingproperties of HMG-boxes from SRY proteins ofprimates. Gene, 154, 277–280.
60. Harley, V. R., Layfield, S., Mitchell, C. L., Forwood,J. K., John, A. P., Briggs, L. J. et al. (2003). Defectiveimportin beta recognition and nuclear import of thesex-determining factor SRY are associated with XYsex-reversing mutations. Proc. Natl Acad. Sci. USA,100, 7045–7050.
61. Parkhurst, K. M., Brenowitz, M. & Parkhurst, L. J.(1996). Simultaneous binding and bending ofpromoter DNA by the TATA binding protein: realtime kinetic measurements. Biochemistry, 35,7459–7465.
62. Dragan, A. I., Read, C. M., Makeyeva, E. N.,Milgotina, E. L., Churchill, M. E., Crane-Robinson,C. & Privalov, P. L. (2004). DNA binding and bendingby HMG boxes: energetic determinants of specificity.J. Mol. Biol. 343, 371–393.
63. Botfield, M. C. & Weiss, M. A. (1994). Bipartite DNArecognition by the human Oct-2 POU domain: POUs-specific phosphate contacts are analogous to those ofbacteriophage lambda repressor. Biochemistry, 33,2349–2355.
64. Labeots, L. A. &Weiss, M. A. (1997). Electrostatics andhydration at the homeodomain–DNA interface:chemical probes of an interfacial water cavity. J. Mol.Biol. 269, 113–128.
65. Jamieson, E. R. & Lippard, S. J. (2000). Stopped-flowfluorescence studies of HMG-domain protein bindingto cisplatin-modified DNA. Biochemistry, 39,8426–8438.
66. Forwood, J. K., Harley, V. & Jans, D. A.(2001). The C-terminal nuclear localizationsignal of the sex-determining region Y (SRY)high mobility group domain mediates nuclearimport through importin beta 1. J. Biol. Chem.276, 46575–46582.
67. Gasca, S., Canizares, J., De Santa Barbara, P., Mejean,C., Poulat, F., Berta, P. & Boizet-Bonhoure, B. (2002).A nuclear export signal within the highmobility group domain regulates the nucleocytoplas-mic translocation of SOX9 during sexualdetermination. Proc. Natl Acad. Sci. USA, 99,11199–111204.
68. Giese, K., Cox, J. & Grosschedl, R. (1992). The HMGdomain of lymphoid enhancer factor 1 bends DNAand facilitates assembly of functional nucleoproteinstructures. Cell, 69, 185–195.
69. Giese, K., Kingsley, C., Kirshner, J. R. & Grosschedl, R.(1995). Assembly and function of a TCR alphaenhancer complex is dependent on LEF-1-inducedDNA bending and multiple protein–protein inter-actions. Genes Dev. 9, 995–1008.
70. Giese, K., Pagel, J. & Grosschedl, R. (1997). Functionalanalysis of DNA bending and unwinding by the highmobility group domain of LEF-1. Proc. Natl Acad. Sci.USA, 94, 12845–12850.
71. Perez-Howard, G. M., Weil, P. A. & Beechem, J. M.(1995). Yeast TATA binding protein interaction withDNA: fluorescence determination of oligomeric state,equilibrium binding, on-rate, and dissociation kin-etics. Biochemistry, 34, 8005–8017.
72. Yamamoto, A., Ando, Y., Yoshioka, K., Saito, K.,Tanabe, T., Shirakawa, H. & Yoshida, M. (1997).Difference in affinity for DNA between HMG proteins1 and 2 determined by surface plasmon resonancemeasurements. J. Biochem. (Tokyo), 122, 586–594.
73. Parkhurst, K. M., Richards, R. M., Brenowitz, M. &Parkhurst, L. J. (1999). Intermediate species posses-sing bent DNA are present along the pathway toformation of a final TBP-TATA complex. J. Mol. Biol.289, 1327–1341.
74. Shymko, R. M., De Meyts, P. & Thomas, R. (1997).Logical analysis of timing-dependent receptor signal-ling specificity: application to the insulin receptormetabolic and mitogenic signalling pathways. Bio-chem. J. 2, 463–469.
75. Bowman, G. D., Goedken, E. R., Kazmirski, S. L.,O’Donnell, M. & Kuriyan, J. (2005). DNA polymeraseclamp loaders and DNA recognition. FEBS Letters,579, 863–867.
76. Mueller, W. & Corothers, D. (1968). Studies of thebinding of actinomycin and related compounds toDNA. J. Mol. Biol. 35, 251–290.
77. White, R. J. & Phillips, D. R. (1989). Drug–DNAdissociation kinetics. In vitro transcription andsodium dodecyl sulphate sequestration. Biochem.Pharmacol. 2, 331–334.
78. Thevenet, L., Mejean, C., Moniot, B., Bonneaud, N.,Galeotti, N., Aldrian-Herrada, G. et al. (2004).Regulation of human SRY subcellular distributionby its acetylation/deacetylation. EMBO J. 23,3336–3345.
79. Poulat, F., Soullier, S., Goze, C., Heitz, F., Calas, B. &Berta, P. (1994). Description and functional impli-

192 A Kinetic Clamp in SRY Augments DNA Bending
cations of a novel mutation in the sex-determininggene SRY. Hum. Mutat. 3, 200–204.
80. Schmitt-Ney, M., Thiele, H., Kaltwasser, P.,Bardoni, B., Cisternino, M. & Scherer, G. (1995).Two novel SRY missense mutations reducingDNA binding identified in XY females and theirmosaic fathers. Am. J. Hum. Genet. 56, 862–869.
81. Veitia, R., Ion, A., Barbaux, S., Jobling, M. A.,Souleyreau, N., Ennis, K. et al. (1997). Mutations andsequence variants in the testis-determining region ofthe Y chromosome in individuals with a 46,XY femalephenotype. Hum. Genet. 99, 648–652.
82. Lundberg, Y., Ritzen, M., Harlin, J. & Wedell, A.(1998). Novel missense mutation (P131R) in the HMGbox of SRY in XY sex reversal. Hum. Mutat. Suppl. 1,S328.
83. Chow, C. S., Whitehead, J. P. & Lippard, S. J. (1994).HMG domain proteins induce sharp bends incisplatin-modified DNA. Biochemistry, 33,15124–15130.
84. Thompson, J. F. & Landy, A. (1988). Empiricalestimation of protein-induced DNA bending angles:applications to lambda site-specific recombinationcomplexes. Nucl. Acids Res. 16, 9687–9705.
85. Srinivasan, A. R., Torres, R., Clark, W. & Olson, W. K.(1987). Base sequence effects in double helical DNA. I.Potential energy estimates of local base morphology.J. Biomol. Struct. Dynam. 5, 459–496.
86. Haas, E. (2005). The study of protein folding anddynamics by determination of intramolecular dis-tance distributions and their fluctuations usingensemble and single-molecule FRET measurements.Chemphyschem. 6, 858–870.
87. Hinck, A. P., Walkenhorst, W. F., Westler, W. M.,Choe, S. & Markley, J. L. (1993). Overexpressionand purification of avian ovomucoid third domainsin Escherichia coli. Protein Eng. 6, 221–227.
88. Cohen, D. R., Sinclair, A. H. & McGovern, J. D. (1994).SRY protein enhances transcription of Fos-related
antigen 1 promoter. Proc. Natl Acad. Sci. USA, 91,4372–4376.
89. Dawson, W. R. & Windsor, M. W. (1968). Flourescenceyields of aromatic compounds. J. Phys. Chem. 72,3251–3260.
90. Sosnick, T. R., Fang, X. & Shelton, V. M. (2000).Application of circular dichroism to study RNAfolding transitions. Methods Enzymol. 317,393–409.
91. Beechem, J. M. & Haas, E. (1989). Simultaneousdetermination of intramolecular distance distri-butions and conformational dynamics by globalanalysis of energy transfer measurements. Biophys. J.55, 1225–1236.
92. Werner, M. H., Bianchi, M. E., Gronenborn, A. M. &Clore, G.M. (1995). NMR spectroscopic analysis of theDNA conformation induced by the human testisdetermining factor SRY. Biochemistry, 34, 11998–12004.
93. Maniatis, T., Falvo, J. V., Kim, T. H., Kim, T. K., Lin,C. H., Parekh, B. S. & Wathelet, M. G. (1998).Structure and function of the interferon-betaenhanceosome. Cold Spring Harbor Symp. Quant.Biol. 63, 609–620.
94. Williams, D. C. J., Cai, M. & Clore, G. M. (2004).Molecular basis for synergistic transcriptional acti-vation by Oct1 and Sox2 revealed from the solutionstructure of the 42-kDa Oct1.Sox2.Hoxb1-DNA tern-ary transcription factor complex. J. Biol. Chem. 279,1449–1457.
95. Bushman, F. D., Anderson, J. E., Harrison, S. C. &Ptashne, M. (1985). Ethylation interference andX-ray crystallography identify similar interactionsbetween 434 repressor and operator. Nature, 316,651–653.
96. Zhu, L., Wilken, J., Phillips, N. B., Narendra, U.,Chan, G., Stratton, S. M. et al. (2000). Sexualdimorphism in diverse metazoans is regulated bya novel class of intertwined zinc fingers. Genes Dev.14, 1750–1764.
Edited by J. O. Thomas
(Received 27 May 2005; received in revised form 8 December 2005; accepted 7 January 2006)Available online 6 February 2006
Copyright © 2022 FDOKUMEN




















