
Spisulosine (ES-285) induces prostate tumor PC-3 and LNCaP cell death by de novo synthesis of...
9
Spisulosine (ES-285) induces prostate tumor PC-3 and LNCaP cell death by de novo synthesis of ceramide and PKCζ activation Ana M. Sánchez a,1 , Sophie Malagarie-Cazenave a,1 , Nuria Olea a , Diana Vara a , Carmen Cuevas b , Inés Díaz-Laviada a, ⁎ a Department of Biochemistry and Molecular Biology, School of Medicine, University of Alcalá, 28871 Alcalá de Henares, Madrid, Spain b PharmaMar, 28770 Colmenar Viejo, Madrid, Spain Received 14 May 2007; received in revised form 21 January 2008; accepted 6 February 2008 Available online 14 February 2008 Abstract During the past decades, intense attention has been focused on the anti-tumor properties of marine compounds which some of them have been revealed as potent apoptotic inducers. In the present work, we studied the mechanism of action of a new compound, Spisulosine (ES-285), isolated from the sea mollusc Spisula polynyma, in the prostate tumor PC-3 and LNCaP cell lines. Spisulosine inhibited cell proliferation with an IC 50 of 1 μM in both cell lines, although it was more effective in the androgen-independent PC-3 cells. The anti-proliferative effect induced by Spisulosine in prostate cells was independent of peroxisome proliferator activated receptor gamma (PPARγ) and phosphatidylinositol 3-kinase/ (PI3K/Akt), Jun N-terminal kinase (JNK), p38 or classical protein kinase C (PKCs) pathways, as it was inferred from the results obtained with specific inhibitors of these routes. However, Spisulosine treatment of prostate cells induced an increase in the intracellular ceramide levels, that was totally blocked by the ceramide synthase inhibitor Fumonisin B1, indicating that the ceramide accumulation came from the de novo biosynthesis. Spisulosine also induced in both PC-3 and LNCaP cells, an activation of the atypical PKC isoform, PKCζ, which is one of the target proteins of ceramide. These results indicate that the marine compound Spisulosine inhibits the growth of the prostate PC-3 and LNCaP cells through intracellular ceramide accumulation and PKCζ activation. © 2008 Elsevier B.V. All rights reserved. Keywords: Spisulosine; Ceramide; PKCζ; Cancer; Prostate 1. Introduction Numerous studies have demonstrated that suppression of apoptosis or defects in apoptotic pathway contribute to expansion of aberrant clones leading to cancer development. It is generally assumed that tumor growth occurs when cells lose the normal balance between cell proliferation and apoptosis. Therefore, cancer chemotherapy has been focused on anti- proliferative strategies. During the past decades, intense attention has been focused on the anti-tumor properties of marine natural products such as Pachymatismin, derived from marine sponge, Didemnin B, ET-743 and Aplidin, derived from tunicates, or Bryostatins, derived from bryozoan (Jimeno, 2002). Spisulosine (PharmaMar, ES-285) is a novel anti-cancer agent isolated from the sea mollusc Spisula polynyma which has been suggested to display anti-tumor activities in several types of human tumors. It has been previously described that Spisulosine inhibits the growth of monkey kidney cells in culture and promotes the disassembly of stress fibbers (Cuadros et al., 2000). However the mechanism whereby Spisulosine induces cell death remains elusive. Recent research reveals that sphingolipids can cause growth arrest or death in a variety of cultured tumor cell lines (Cuvillier and Levade, 2003). Ceramide, in particular, has emerged as a new lipid signalling molecule capable of regulating a variety of cellular processes including cell growth arrest and apoptosis. There are two main routes to generate ceramide: the de novo pathway and the degradation of membrane sphingomyelin by Available online at www.sciencedirect.com European Journal of Pharmacology 584 (2008) 237 – 245 www.elsevier.com/locate/ejphar ⁎ Corresponding author. Tel.: +34 91 885 5141. E-mail address: [email protected] (I. Díaz-Laviada). 1 Both authors contributed equally to this study. 0014-2999/$ - see front matter © 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.ejphar.2008.02.011
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Spisulosine (ES-285) induces prostate tumor PC-3 and LNCaP cell death by de novo synthesis of...

Available online at www.sciencedirect.com
gy 584 (2008) 237–245www.elsevier.com/locate/ejphar
European Journal of Pharmacolo
Spisulosine (ES-285) induces prostate tumor PC-3 and LNCaP cell death byde novo synthesis of ceramide and PKCζ activation
Ana M. Sánchez a,1, Sophie Malagarie-Cazenave a,1, Nuria Olea a, Diana Vara a,Carmen Cuevas b, Inés Díaz-Laviada a,⁎
a Department of Biochemistry and Molecular Biology, School of Medicine, University of Alcalá, 28871 Alcalá de Henares, Madrid, Spainb PharmaMar, 28770 Colmenar Viejo, Madrid, Spain
Received 14 May 2007; received in revised form 21 January 2008; accepted 6 February 2008Available online 14 February 2008
Abstract
During the past decades, intense attention has been focused on the anti-tumor properties of marine compounds which some of them have beenrevealed as potent apoptotic inducers. In the present work, we studied the mechanism of action of a new compound, Spisulosine (ES-285), isolatedfrom the sea mollusc Spisula polynyma, in the prostate tumor PC-3 and LNCaP cell lines. Spisulosine inhibited cell proliferation with an IC50 of1 μM in both cell lines, although it was more effective in the androgen-independent PC-3 cells. The anti-proliferative effect induced bySpisulosine in prostate cells was independent of peroxisome proliferator activated receptor gamma (PPARγ) and phosphatidylinositol 3-kinase/(PI3K/Akt), Jun N-terminal kinase (JNK), p38 or classical protein kinase C (PKCs) pathways, as it was inferred from the results obtained withspecific inhibitors of these routes. However, Spisulosine treatment of prostate cells induced an increase in the intracellular ceramide levels, thatwas totally blocked by the ceramide synthase inhibitor Fumonisin B1, indicating that the ceramide accumulation came from the de novobiosynthesis. Spisulosine also induced in both PC-3 and LNCaP cells, an activation of the atypical PKC isoform, PKCζ, which is one of the targetproteins of ceramide. These results indicate that the marine compound Spisulosine inhibits the growth of the prostate PC-3 and LNCaP cellsthrough intracellular ceramide accumulation and PKCζ activation.© 2008 Elsevier B.V. All rights reserved.
Keywords: Spisulosine; Ceramide; PKCζ; Cancer; Prostate
1. Introduction
Numerous studies have demonstrated that suppression ofapoptosis or defects in apoptotic pathway contribute toexpansion of aberrant clones leading to cancer development.It is generally assumed that tumor growth occurs when cells losethe normal balance between cell proliferation and apoptosis.Therefore, cancer chemotherapy has been focused on anti-proliferative strategies. During the past decades, intenseattention has been focused on the anti-tumor properties ofmarine natural products such as Pachymatismin, derived frommarine sponge, Didemnin B, ET-743 and Aplidin, derived from
⁎ Corresponding author. Tel.: +34 91 885 5141.E-mail address: [email protected] (I. Díaz-Laviada).
1 Both authors contributed equally to this study.
0014-2999/$ - see front matter © 2008 Elsevier B.V. All rights reserved.doi:10.1016/j.ejphar.2008.02.011
tunicates, or Bryostatins, derived from bryozoan (Jimeno,2002). Spisulosine (PharmaMar, ES-285) is a novel anti-canceragent isolated from the sea mollusc Spisula polynyma whichhas been suggested to display anti-tumor activities in severaltypes of human tumors. It has been previously described thatSpisulosine inhibits the growth of monkey kidney cells inculture and promotes the disassembly of stress fibbers (Cuadroset al., 2000). However the mechanism whereby Spisulosineinduces cell death remains elusive.
Recent research reveals that sphingolipids can cause growtharrest or death in a variety of cultured tumor cell lines (Cuvillierand Levade, 2003). Ceramide, in particular, has emerged as anew lipid signalling molecule capable of regulating a variety ofcellular processes including cell growth arrest and apoptosis.There are two main routes to generate ceramide: the de novopathway and the degradation of membrane sphingomyelin by

Fig. 2. Cell death induction by Spisulosine in prostate cells. Prostate cancer PC-3and LNCaP cells as well as human prostate epithelial RWPE-1 cells wereincubated in the absence or presence of Spisulosine (ES-285) for 48 h. A. Effectof Spisulosine on cell viability. MTT cell viability assay was performed ac-cording to Materials and methods. Absorbance ratio at 630 nm/560 nm wascalculated. Data are expressed as % of control values and are the mean±S.D. ofthree different experiments performed in triplicate. B. Effect of Spisulosine oncell cycle progression. Cells were treated with either vehicle control or 1 μMSpisulosine (ES-285). After 48 h, cells were collected, and then cellular DNAwas stained with propidium iodide as detailed in Materials and methods. Flowcytometric analysis was then performed for cell cycle distribution. The figureshows propidium iodide fluorescence pattern for cell cycle distribution. Num-bers in the upper right corner indicate the % of subG1 cells. The experiment wasperformed twice with similar results.
238 A.M. Sánchez et al. / European Journal of Pharmacology 584 (2008) 237–245
sphingomyelinases. De novo biosynthesis includes condensa-tion of serine with a palmitoyl CoA by the enzyme serinepalmitoyltransferase, a reduction to sphinganine and a furtherformation of dihydroceramide by dihydroceramide synthase(also named ceramide synthase). Inhibitors of de novo ceramidesynthesis have been widely used to investigate the role of thispathway in cell physiology. Fumonisin B1, a mycotoxin pro-duced by Fusarium verticillioides, is a reportedly specific in-hibitor of ceramide synthase which has shown to inhibitceramide-induced apoptosis in many cells (Son et al., 2007; Leeet al., 2007) including prostate tumor cells (Jiang et al., 2004;Eto et al., 2003). Intracellular ceramide accumulation has beendescribed to occur as a consequence of several stressing stimuliand it drives death-signalling cascades (Levade et al., 2002).One of the best-characterized intracellular targets of ceramideaction is the atypical protein kinase C (PKC) isoform, PKCζ,which has been shown to be involved in apoptotic responses.Binding of PKCζ to ceramide, triggers a conformational changeand activation of PKCζ by phosphorylation (Wang et al., 2005),which stimulates downstream targets as Jun N-terminal kinase(JNK) activation (Castrillo et al., 2003) or nuclear factor-kappaB (NFκB) inhibition (Wang et al., 2005).
In this study, Spisulosine was tested for its anti-tumor po-tential on human androgen-independent cancer PC-3 cells aswell as in human androgen-dependent LNCaP cells. We dem-onstrate here that Spisulosine inhibits prostate cell growth withan IC50 of 1–10 μM through intracellular accumulation of cer-amide and activation of PKCζ.
2. Materials and methods
2.1. Materials
Spisulosine (ES-285, molecular formula C18H40ClNO, MW321 g/mol), was obtained from PharmaMar (Tres Cantos,Madrid). GW 9662 was from Alexis Corporation (San Diego,CA, USA). Bisindolylmaleimide I, JNK inhibitor I, SB 203580,Myriocin and PKCζ inhibitor, were purchased from Calbio-chem (Darmstadt, Germany). Fumonisin B1 and other agentswere from Sigma (St. Louis, MO, USA).
2.2. Cell cultures
Human prostatic carcinoma (PC-3), lymph node carcinomaof the prostate (LNCaP) and human prostate epithelial RWPE-1cells, were purchased from American Type Culture Collection(ATCC CRL-1435, CRL-1740 and CRL-11609, respectively)(Rockville, MD, USA). PC-3 and LNCaP cells were routinelygrown in RPMI 1640 medium supplemented with 100 IU/ml
Fig. 1. Structure of the compound Spisulosine (ES-285).
penicillin G sodium, 100 μg/ml sptreptomycin sulfate and0.25 μg/ml amphotericin B (Invitrogen, Paisley, UK) and either10% foetal calf serum for PC-3 cells or 5% foetal calf serum forLNCaP cells. RWPE-1 cells were maintained in the Keratino-cyte-serum free medium (K-SFM) containing 5 ng/ml humanrecombinant epithelial growth factor (hrEGF), 0.05 mg/mlbovine pituitary extract (Invitrogen), 100 U/ml Penicillin G,and 100 μg/ml Streptomycin. Cells were seeded at a density of

Fig. 3. Spisulosine induces apoptosis in prostate cancer cells. FITC-Annexin Vand propidium iodide staining of PC-3 and LNCaP cells treated with 1 μM Spisulosine(ES-285). Numbers in the upper right corner of each graph indicate the number of apoptotic cells (Annexin+/PI+). Western blot of procaspase-3 is shown on the right.Tubulin is used as loading control. The experiment was performed twice with similar results.
239A.M. Sánchez et al. / European Journal of Pharmacology 584 (2008) 237–245
10,000 cells/cm2. One day prior to the experiments, the serum-containing medium was removed and a chemically definedmedium consisting of RPMI 1640 supplemented with 5 μg/mlinsulin, 5 μg/ml transferrin and 5 ng/ml sodium selenite wasadded.
2.3. MTT cell viability assay
After treatments according to figure legends, cell viabilitywas measured with a 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT) colorimetric assay (BoehringerMannheim, Mannheim, Germany) based on the cleavage of theyellow tetrazolium salt MTT to purple formazan crystals by
Table 1Effect of different inhibitors of signalling proteins on the action of ES-285 onprostate cells
Treatment Cell viability (% of control)
PC-3 LNCaP
Control 100 1001 μM ES-285 54.29±4,8 69.23±6.31 μM ES-285+2 μM BIM 48.55±9.8 62.75±5.41 μM ES-285+5 μM GW 9662 55.76±8.1 72.45±8.31 μM ES-285+100 μM Fumonisin B1 80.56±3.1a 95.36±9.4a
1 μM ES-285+0.1 μM JNK Inhibitor I 53.25±9.8 60.73±7.71 μM ES-285+10 μM LY 294002 52.82±11.0 58.20±12.31 μM ES-285+10 μM SB 203580 58.48±11.7 63.34±9.9
Prostate PC-3 and LNCaP cells were incubated with 1 μMES-285 for 48 h in thepresence of the broad PKCs inhibitor bisindolylmaleimide I (BIM), the PPARγinhibitor GW 9662, the ceramide synthase inhibitor Fumonisin B1, the JNKinhibitor I, the PI-3K inhibitor LY 294002, and the p38 MAPK inhibitor SB203580, at the doses indicated. Cell viability was analyzed by the MTT color-imetric assay (aPb0.05).
metabolic active cells. The formazan crystals formed were sol-ubilized in 10% SDS, 0.01 MHCl and monitored by absorbanceat 570 nm with a reference wavelength of 630 nm.
2.4. Flow cytometry and propidium iodide staining
Control or Spisulosine-treated cells were harvested in 0.1%Nonidet P-40 and 0.5 mg/ml RNase for 30 min and stained with0.05 mg/ml propidium iodide (PI) to indicate relative DNAcontent. Cell samples were analyzed by flow cytometry in aFACScan (Beckton Dickinson, CA, Costa Rica).
Fig. 4. Examination of the cytotoxic effect of Fumonisin B1 in prostate PC-3cells. Cells were incubated with 1 μM Spisulosine (ES-285) in the presence ofincreasing concentrations of Fumonisin B1 for 48 h. Then, a MTT cell viabilityassay was performed according to Materials and methods. Absorbance ratio at630 nm/560 nm was calculated. Data are expressed as % of control values andare the mean±S.D. of three different experiments performed in triplicate.

240 A.M. Sánchez et al. / European Journal of Pharmacology 584 (2008) 237–245
2.5. Apoptosis assessment by Annexin V staining
Prostate cells were cultured in 35 mm well plates as describeabove, in the presence or absence of capsaicin according to theexperiment. The cells were then washed twice with PBS andincubated in 0.5 ml of Binding Buffer (10 mM HEPES pH 7.4,150 mM NaCl, 2.5 mM CaCl2, 1 mM MgCl2, and 4% BSA),with 4 μg/ml Annexin V-FITC for 15 min. Cells were thenwashed in PBS and resuspended in Binding Buffer with 0.6 μg/ml propidium iodide, (Calbiochem, USA). 20,000 cells of eachsample were analyzed by flow cytometry in a FACScan (BecktonDickinson, CA, Costa Rica).
2.6. Lipid extraction and ceramide quantification
Ceramide quantification in cell lipid extracts was performedaccording to the method described by Bielawska et al. (2001), forceramide quantification in cultured cells. Briefly, cell pellets weresuspended in 0.6 ml distilled water, and cells were disruptedat 4 °C by sonication. Lipids were extracted with chloroform/methanol using the procedure of Bligh and Dyer (1959) andceramide content was determined by phosphorylation withE. coli
Fig. 5. Time-course of ceramide accumulation induced by Spisulosine inprostate tumor cells. PC-3 and LNCaP cells were incubated with 1 μMSpisulosine for the indicated times. Then intracellular ceramide levels weremeasured in the cell lipid extract by the diacylglycerol kinase method asdescribed in Materials and methods. Data are expressed as % of control valuesand are the mean±S.D. of three different experiments performed in triplicate.
Fig. 6. Spisulosine induces intracellular ceramide accumulation in prostatetumor cells. PC-3 and LNCaP cells were incubated with 1 μM or 10 μMSpisulosine (ES-285) for 48 h in the presence or absence of 100 μM FumonisinB1 (Fumo). Then, intracellular ceramide levels were measured in the cell lipidextract by the diacylglycerol kinase method as described in Materials andmethods. Data are expressed as % of control values and are the mean±S.D. ofthree different experiments performed in triplicate.
diacylglycerol kinase (Calbiochem) and [32P]γ-ATP (6000 Ci/mmol; Perkin-Elmer, Barcelona). Products of this reaction werepurified by thin-layer chromatography and quantified as pre-viously reported (Malagarie-Cazenave et al., 2004).
2.7. Western blot analysis
Cultured PC-3 and LNCaP cells were pooled and lysedin a lysis buffer (50 mM Tris–HCl, pH 7.4, 5 mM ethylene-diaminetetraacetic acid (EDTA), 1 mM EGTA, 10 mM 2-mercaptoethanol) containing protease inhibitors (5 μg/mlleupeptin, 5 μg/ml aprotinin, 10 μg/ml soybean trypsin inhibitorand 1 mM phenylmethylsulfonyl fluoride) and disrupted by twofreeze–thaw cycles and sonication. Particulate fraction wasisolated by centrifugation at 100,000 g for 1 h. Western blotanalysis was performed as previously described (Diaz-Laviadaet al., 1991) with minor modifications. 20 μg of particulatefraction were resolved on an 8% acrylamide gel and the proteinswere then transferred to a nitrocellulose membrane. The mem-brane was blocked with 3% fat-free milk and incubatedwith anti-caspase 3 antibody or anti-PKCζ antibody (Sta Cruz

Fig. 7. Fumonisin B1 andMyriocin inhibit the anti-proliferative effect of ES-285in PC-3 and LNCaP prostate cells. A. Cells were incubated for 48 h with 1 μMor10 μM Spisulosine (ES-285) in the presence or absence of 100 μM FumonisinB1. B. Cells were incubated for 48 h with 1 μM or 10 μM Spisulosine (ES-285)in the presence or absence of 500 nMMyriocin. Then, a MTTcell viability assaywas performed according to Materials and methods. Absorbance ratio at630 nm/560 nm was calculated. Data are expressed as % of control values andare the mean±S.D. of three different experiments performed in triplicate.
241A.M. Sánchez et al. / European Journal of Pharmacology 584 (2008) 237–245
Biotechnology, Sta Cruz, CA, USA) (1:1000 dilution) overnightat 4 °C. After washing, the membranes were incubated for 1 hwith a secondary horseradish peroxidase-conjugated antibodyand developed with Enhanced Chemiluminiscence substrate(Millipore, Madrid, Spain).
2.8. Statistical analysis
Data are presented as the mean±S.D. of the number ofexperiments indicated. Statistical comparisons among groupswere made with a t test and the difference was considered to bestatistically significant when the P value was b0.05.
3. Results
3.1. Spisulosine inhibits prostate PC-3 and LNCaP cell growth
Spisulosine (Fig. 1) is a synthetic molecule originally foundin the mollusc Mactromeris polynyma. To study the effect ofSpisulosine in prostate tumor cells, we compared the androgen-resistant PC-3 cells and the androgen-sensitive LNCaP cellswith human prostate epithelial RWPE-1 cells. PC-3 cells wereisolated from a human androgen-insensitive prostatic adeno-carcinoma metastatic to bone and they represent the androgen-refractory phase of advanced prostate cancer, whereas LNCaPcells were isolated from a lymph node prostate cancer whichresponds to androgen stimulation for growth and hence rep-resents an early and treatable cancer. The RWPE-1 cell line wasimmortalized with human papillomavirus 18 from histologicalnormal prostate epithelial cells and they retain normal epithelialcell morphology (Bello et al., 1997). Cell lines were treated withdifferent doses of Spisulosine and then, cell viability wasassessed by MTT colorimetric assay. As shown in Fig. 2A,Spisulosine induced cell growth arrest with an IC50 of 1 μMat 48 h of treatment. Studies of cell cycle by flow cytometryrevealed that Spisulosine increased the number of cells insubG1, suggesting a cytotoxic effect (Fig. 2B). As it can beinferred from Fig. 2, Spisulosine was more efficacious inprostate cancer cells than in normal prostate epithelial cells.Whereas in PC-3 cells the maximal cell growth inhibitionobtained was of 95% and subG1 cells increased up to nearly30% in the androgen-sensitive LNCaP cells, the growth wasinhibited to 45% and the subG1 cell increased to 18.5%.Spisulosine was less efficacious in control RWPE-1 cells inwhich cell growth inhibition reached 50% and subG1 cellsincreased to 10% (Fig. 2). So, we decided to continue the studywith the prostate cancer PC-3 and LNCaP cells.
In order to ascertain whether Spisulosine induced apoptoticcell death, we determined the translocation of phosphatidylser-ine to the external layer of the membrane by Annexin Vstaining. As shown in Fig. 3, 1 μM Spisulosine increased thenumber of Annexin V+/PI+ cells, which were interpreted as inthe late stage of apoptosis. We further investigated the effect ofSpisulosine on caspase-3 activation which is considered as ahallmark of apoptosis. Spisulosine increased the cleavage ofprocaspase-3 both in PC-3 as in LNCaP cells, although thedecrease in procaspase-3 was more pronounced in PC-3 cells
(Fig. 3). This data indicate that Spisulosine induce apoptosis inprostate cancer cells.
In order to analyze the mechanism of action of ES-285,prostate cancer cells were incubated with the compound in thepresence of inhibitors of different signalling key molecules.We use the broad PKC inhibitor bisindolylmaleimide I, theperoxisome proliferator activated receptor gamma (PPARγ)antagonist GW 9662, the ceramide synthase inhibitor Fumoni-sin B1, the phosphatidylinositol 3-kinase (PI-3K) inhibitor LY294002, and inhibitors for the JNK and p38 stress-relatedmitogen-activated protein (MAP) kinases (JNK inhibitor Iand SB 203580, respectively). None of the different inhibitorstested, with the exception of the ceramide synthase inhibitor

242 A.M. Sánchez et al. / European Journal of Pharmacology 584 (2008) 237–245
Fumonisin B1, could block the growth inhibitory effect inducedby ES-285 (Table 1). To confirm these results and to test thedose of Fumonisin B1 to employ in further experiments, weincubate PC-3 cells with 1 μM ES-285 in the presence ofdifferent concentrations of Fumonisin B1. As it can be observedin Fig. 4, Fumonisin B1 reversed the effect of ES-285 from aconcentration of 10 μM to 100 μM. None of these doses ofFumonisin B1 were toxic for cells, as were corroborated by acell viability titration assay (data not shown). Fumonisin B1 is awell known inhibitor the ceramide synthase enzyme (Sorianoet al., 2005) and therefore our results pointed to a role forceramide synthesis in the mechanism whereby Spisulosine in-duced inhibition of prostate cell growth.
3.2. ES-285 induces ceramide production in PC-3 and LNCaPprostate cells
To corroborate this hypothesis and as ceramide has beenidentified as an intracellular mediator of stress signalling thatleads to cell death (Andrieu-Abadie and Levade, 2002), weanalyzed the intracellular ceramide levels after ES-285 treat-ment. In both cell lines PC-3 and LNCaP, an increase in
Fig. 8. Spisulosine-induced apoptosis is prevented by Fumonisin B1. FITC-Annex100 μM Fumonisin B1 prior the treatment with 1 μM Spisulosine (ES-285). Numbe(Annexin+/PI+). The experiment was performed twice with similar results.
ceramide levels after 24 h Spisulosine treatment could bedetected (Fig. 5). Accumulation of ceramide in prostate cellswas a late phenomenon since the time-course experimentsrevealed that ceramide levels started to elevate from 24 h ofSpisulosine treatment and was maximal at 48 h (Fig. 5). Longertimes were very difficult to measure since the quantity of cells inthe plate was insufficient to isolate enough ceramide.
The increase in intracellular ceramide was prevented with thepre-treatment with Fumonisin B1 (Fig. 6). However, intracel-lular ceramide levels after Spisulosine treatment were higher inPC-3 cells than in LNCaP cells. This finding could explain thehigher anti-proliferative effect induced by the marine compoundin PC-3 cells. The fact that increased ceramide levels weretotally abolished with the ceramide synthase inhibitor confirmsthat ceramide production after Spisulosine treatment comesfrom de novo synthesis and not from sphingomyelin hydrolysis.
3.3. Ceramide is involved in the Spisulosine-induced prostatecell death
To investigate whether ceramide synthesis was involvedin the anti-proliferative effect of Spisulosine, we tested the
in V and propidium iodide staining of PC-3 and LNCaP cells pre-treated withrs in the upper right corner of each graph indicate the number of apoptotic cells
243A.M. Sánchez et al. / European Journal of Pharmacology 584 (2008) 237–245
effect of the marine compound in the presence of the ceramidesynthase inhibitor Fumonisin B1. As shown in Fig. 7A,Fumonisin B1 prevented the Spisulosine-induced cell deathin PC-3 and LNCaP cells at both doses of 1 μM and of 10 μMSpisulosine studied. To further study the involvement ofceramide synthesis in the action mechanism of ES-285, wetested the effect of Myriocin, a serine palmitoyltransferase in-hibitor. Results in Fig. 7B show that the anti-proliferative effectof Spisulosine in PC-3 and LNCaP cells was prevented by500 nM Myriocin.
The Spisulosine-induced apoptosis was also preventedby the inhibitor Fumonisin B1 as can be inferred from Figs. 8and 9. Fumonisin B1 reduced the Spisulosine-induced apoptoticcells to control values both in PC-3 as in LNCaP cells (Fig. 8).Furthermore, Fumonisin B1 reduced the caspase-3 cleavagepromoted by Spisulosine treatment (Fig. 9). These data indicatethat de novo synthesis of ceramide is involved in the anti-proliferative effect of Spisulosine.
3.4. Spisulosine induces PKCζ activation through a mechanisminvolving ceramide production
Ceramide generated in the cells is capable of modifying theactivity of a number of proteins including kinases such as JNK,
Fig. 9. Spisulosine-induced caspase-3 cleavage is prevented by Fumonisin B1.PC-3 and LNCaP cells were pre-treated with 100 μM Fumonisin B1 prior thetreatment with 1 μM Spisulosine (ES-285). Then Western blot for procaspase-3was performed according to Materials and methods. The corresponding den-sitometric analysis has been normalized to tubulin levels in each case, and it isexpressed as the mean±S.D. of two independent experiments.
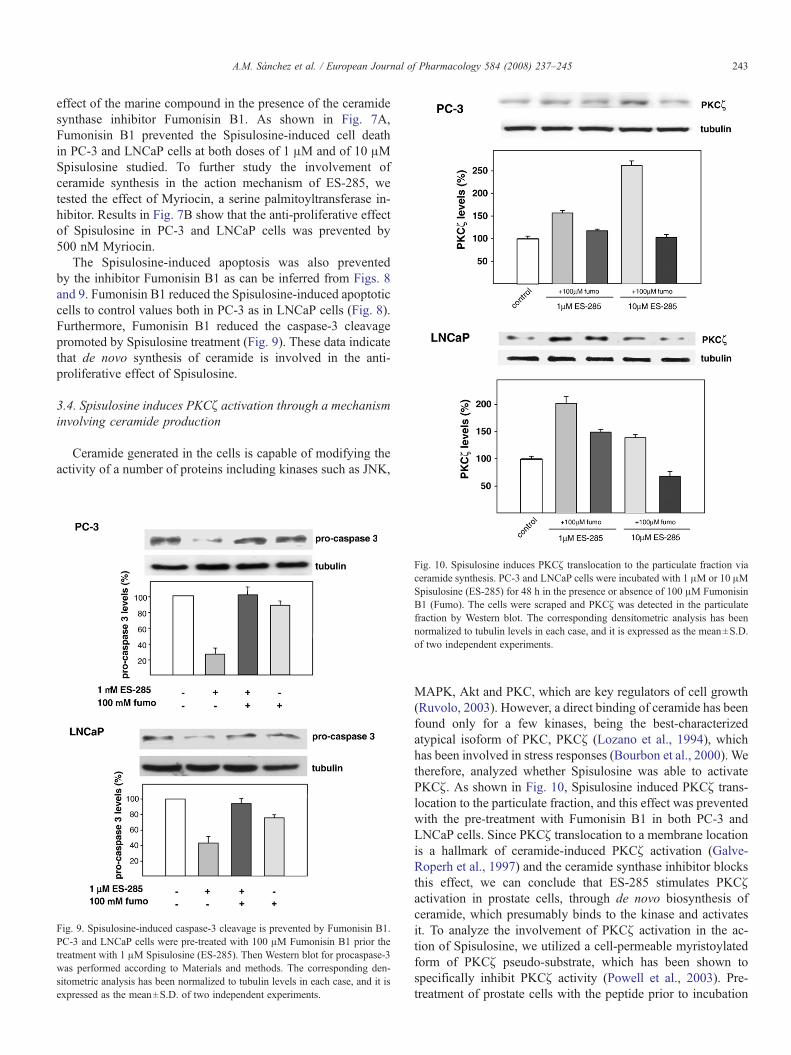
Fig. 10. Spisulosine induces PKCζ translocation to the particulate fraction viaceramide synthesis. PC-3 and LNCaP cells were incubated with 1 μM or 10 μMSpisulosine (ES-285) for 48 h in the presence or absence of 100 μM FumonisinB1 (Fumo). The cells were scraped and PKCζ was detected in the particulatefraction by Western blot. The corresponding densitometric analysis has beennormalized to tubulin levels in each case, and it is expressed as the mean±S.D.of two independent experiments.
MAPK, Akt and PKC, which are key regulators of cell growth(Ruvolo, 2003). However, a direct binding of ceramide has beenfound only for a few kinases, being the best-characterizedatypical isoform of PKC, PKCζ (Lozano et al., 1994), whichhas been involved in stress responses (Bourbon et al., 2000). Wetherefore, analyzed whether Spisulosine was able to activatePKCζ. As shown in Fig. 10, Spisulosine induced PKCζ trans-location to the particulate fraction, and this effect was preventedwith the pre-treatment with Fumonisin B1 in both PC-3 andLNCaP cells. Since PKCζ translocation to a membrane locationis a hallmark of ceramide-induced PKCζ activation (Galve-Roperh et al., 1997) and the ceramide synthase inhibitor blocksthis effect, we can conclude that ES-285 stimulates PKCζactivation in prostate cells, through de novo biosynthesis ofceramide, which presumably binds to the kinase and activatesit. To analyze the involvement of PKCζ activation in the ac-tion of Spisulosine, we utilized a cell-permeable myristoylatedform of PKCζ pseudo-substrate, which has been shown tospecifically inhibit PKCζ activity (Powell et al., 2003). Pre-treatment of prostate cells with the peptide prior to incubation

Fig. 11. The PKCζ inhibitor prevents the anti-proliferative effect of Spisulosinein prostate cells. PC-3 and LNCaP cells were incubated with 1 μMor 10 μMES-285 for 48 h in the presence or absence of 1 μM PKCζ pseudo-substrate peptideinhibitor (PKCζ INH). Then, a MTT cell viability assay was performedaccording to Materials and methods. Absorbance ratio at 630 nm/560 nm wascalculated. Data are expressed as % of control values and are the mean±S.D. oftwo different experiments performed in triplicate.
244 A.M. Sánchez et al. / European Journal of Pharmacology 584 (2008) 237–245
with Spisulosine, prevented the effect of the marine compoundon prostate cell viability (Fig. 11).
4. Discussion
Sphingoid bases have been described to exert an anti-proliferative effect in several cell lines (Pettus et al., 2002). Themarine compound Spisulosine has a molecular structure whichresembles sphingosine and it has been shown to prevent thegrowth of kidney transformed cells (Cuadros et al., 2000). Theincreased apoptotic response to Spisulosine is of interest sinceSpisulosine is currently under consideration in clinical trialsfor cancer therapy. Therefore, we investigated the effect ofSpisulosine in tumor prostate cell growth. We found thatSpisulosine inhibits the growth of the prostate tumor cell linesPC-3 and LNCaP. The compound had a higher effect on theandrogen-resistant PC-3 cells than on the androgen-responsiveLNCaP cells. Since PC-3 cells represent an advanced andtreatment-resistant prostate cancer, the compound Spisulosinecould have a role as prostate anti-tumoral agent. The mechanismof action of Spisulosine in prostate cells seems to be inde-pendent of the stress-related MAP kinases as well as of PPARγreceptor, PI3K/Akt and classical PKCs, since specific inhibitorsof these routes were unable to block the effect of Spisulosine.However, the ceramide synthase inhibitor Fumonisin B1 as wellas the serine palmitoyltransferase inhibitor Myriocin, were ableto prevent the effect induced by Spisulosine, pointing to a denovo synthesis of ceramide in the mechanism of action ofthis compound in prostate cells. In fact, intracellular ceramidelevels were increased after 48 h of the treatment with 1 μMSpisulosine. Ceramide generation is a near universal feature of
programmed cell death (Pettus et al., 2002; Ruvolo, 2003). It ispossible that during at least some apoptotic events, ceramidemay be required to activate stress-signal cascades that lead tocell death, while concurrently, suppressing growth and survivalpathways in the dying cell. Spisulosine induced an increaseof ceramide synthesis that was related with the inhibitionof prostate cell growth. Moreover, ceramide levels increasedhigher in PC-3 cells than in LNCaP cells, corresponding withthe higher efficacy of Spisulosine in PC-3 cells. The mechanismwhereby Spisulosine induces a ceramide increase is not wellunderstood. One possibility is that Spisulosine could be a sub-strate for ceramide synthase since its molecular structure is verysimilar to D-erythro-sphingosine. Studies performed with [3H]-Spisulosine showed that when this compound was up-taken bycultured cells it was transformed into a more hydrophobic mol-ecule (Fernández-Sousa, personal communication). However,even if Spisulosine was transformed into a pseudo-ceramide,it will probably be detected in a separate fraction with thediacylglycerol kinase analytical method. As this was not thecase, we can suppose that Spisulosine induces de novo synthesisof ceramide. De novo synthesis of ceramide has been previouslyreported to mediate cell death in other cell types. In glioma cells,cannabinoids induce apoptosis via de novo synthesis of ceramide(Velasco et al., 2005). In addition, other studies support the viewthat G-protein-coupled receptors may generate a functional-ly active pool of ceramide via enhanced synthesis de novo(Lehtonen et al., 1999). This indicates that de novo synthesis ofceramide could be a common feature in the mechanism of actionof several anti-tumor drugs.
PKCζ has been implicated in the control of many cell func-tions including apoptosis induction in many cell types (Sorianoet al., 2005), so we investigate whether PKCζ was involved inthe growth inhibition induced by Spisulosine in prostate cells.Our data show that de novo synthesis of ceramide plays arelevant role in the activation of PKCζ by ES-285. It has beenreported previously that PKCζ directly interacts with ceramideand modulates the cellular response to TNF (Andrieu-Abadieand Levade, 2002). Furthermore, ceramide has been demon-strated to increase PKCζ activity towards hnRNPA1, a knownregulator of alternative splicing, which phosphorylates andmodifies the alternative splicing of specific genes, such as Bcl-x,resulting in promotion of apoptosis (Pettus et al., 2002). More-over, ceramide also increases the affinity of PKCζ towards itsinhibitor protein PAR-4, which is a mediator apoptosis withspecial relevance in prostate cells (Wang et al., 2005). Our studyshows that the marine compound Spisulosine inhibits prostatetumor cells growth by a mechanism that involves de novo syn-thesis of ceramide and PKCζ activation. These findings throwlight to the mechanism of action of Spisulosine and reinforce thepotential therapeutic role of the marine compound as anti-tumoralagent for prostate cancer treatment.
Acknowledgements
This work was supported by the Spanish MEC (SAF2005-00602), by UAH-CAM (CCG06-UAH/SAL-0562) and byComunidad de Madrid (S-SAL-0261-2006). SM-C is a fellow

245A.M. Sánchez et al. / European Journal of Pharmacology 584 (2008) 237–245
of Juan de la Cierva programme from Spanish MEC. AMS isan FPU fellow from the Spanish MEC. NO is a fellow of theUniversity of Alcalá.
References
Andrieu-Abadie, N., Levade, T., 2002. Sphingomyelin hydrolysis during apo-ptosis. Biochim. Biophys. Acta 1585, 126–134.
Bello, D., Webber, M.M., Kleinman, H.K., Wartinger, D.D.R.J.S., 1997. An-drogen responsive adult human prostatic epithelial cell lines immortalizedby human papillomavirus 18. Carcinogenesis 18, 1215–1223.
Bielawska, A., Perry, D.K., Hannun, Y.A., 2001. Determination of ceramidesand diglycerides by the diglyceride kinase assay. Anal. Biochem. 298,141–150.
Bligh, E.G., Dyer, W.J., 1959. A rapid method of total lipid extraction andpurification. Can. J. Biochem. Physiol. 37, 911–917.
Bourbon, N.A., Yun, J., Kester, M., 2000. Ceramide directly activates proteinkinase C zeta to regulate a stress-activated protein kinase signaling complex.J. Biol. Chem. 275, 35617–35623.
Castrillo, A., Traves, P.G., Martin-Sanz, P., Parkinson, S., Parker, P.J., Bosca, L.,2003. Potentiation of protein kinase C zeta activity by 15-deoxy-delta(12,14)-prostaglandin J(2) induces an imbalance between mitogen-activatedprotein kinases and NF-kappa B that promotes apoptosis in macrophages.Mol. Cell. Biol. 23, 1196–1208.
Cuadros, R., Montejo de Garcini, E., Wandosell, F., Faircloth, G., Fernandez-Sousa, J.M., Avila, J., 2000. The marine compound spisulosine, an inhibitorof cell proliferation, promotes the disassembly of actin stress fibers. CancerLett. 152, 23–29.
Cuvillier, O., Levade, T., 2003. Enzymes of sphingosine metabolism as potentialpharmacological targets for therapeutic intervention in cancer. Pharmacol.Res. 47, 439–445.
Diaz-Laviada, I., Larrodera, P., Nieto, J.L., Cornet, M.E., Diaz-Meco, M.T.,Sanchez, M.J., Guddal, P.H., Johansen, T., Haro, A., Moscat, J., 1991.Mechanism of inhibition of adenylate cyclase by phospholipase C-catalyzedhydrolysis of phosphatidylcholine. Involvement of a pertussis toxin-sensitive G protein and protein kinase C. J. Biol. Chem. 266, 1170–1176.
Eto,M., Bennouna, J., Hunter, O.C., Hershberger, P.A., Kanto, T., Johnson, C.S.,Lotze, M.T., Amoscato, A.A., 2003. C16 ceramide accumulates followingandrogen ablation in LNCaP prostate cancer cells. Prostate 57, 66–79.
Galve-Roperh, I., Haro, A., Diaz-Laviada, I., 1997. Ceramide-induced trans-location of protein kinase C zeta in primary cultures of astrocytes. FEBSLett. 415, 271–274.
Jiang, Q., Wong, J., Fyrst, H., Saba, J.D., Ames, B.N., 2004. g-Tocopherol orcombinations of vitamin E forms induce cell death in human prostate cancer
cells by interrupting sphingolipid synthesis. Proc. Natl. Acad. Sci. USA 101,17825–17830.
Jimeno, J.M., 2002. A clinical armamentarium of marine-derived anti-cancercompounds. Anticancer Drugs 13 (Suppl 1), S15–S19.
Lee, W.K., Torchalski, B., Thévenod, F., 2007. Cadmium-induced ceramideformation triggers calpain-dependent apoptosis in cultured kidney proximaltubule cells. Am. J. Physiol. Cell Physiol. 293, C839–C847.
Lehtonen, J.Y., Horiuchi, M., Daviet, L., Akishita, M., Dzau, V.J., 1999. Ac-tivation of the de novo biosynthesis of sphingolipids mediates angiotensin IItype 2 receptor-induced apoptosis. J. Biol. Chem. 274, 16901–16906.
Levade, T., Malagarie-Cazenave, S., Gouaze, V., Segui, B., Tardy, C., Betito, S.,Andrieu-Abadie, N., Cuvillier, O., 2002. Ceramide in apoptosis: a revisitedrole. Neurochem. Res. 27, 601–607.
Lozano, J., Berra, E., Municio, M.M., Diaz-Meco, M.T., Dominguez, I., Sanz,L., Moscat, J., 1994. Protein kinase C zeta isoform is critical for kappa B-dependent promoter activation by sphingomyelinase. J. Biol. Chem. 269,19200–19202.
Malagarie-Cazenave, S., Segui, B., Leveque, S., Garcia, V., Carpentier, S., Altie,M.F., Brouchet, A., Gouaze, V., Andrieu-Abadie, N., Barreira, Y., Benoist,H., Levade, T., 2004. Role of FAN in tumor necrosis factor-alpha andlipopolysaccharide-induced interleukin-6 secretion and lethality in D-galactosamine-sensitized mice. J. Biol. Chem. 279, 18648–18655.
Pettus, B.J., Chalfant, C.E., Hannun, Y.A., 2002. Ceramide in apoptosis: anoverview and current perspectives. Biochim. Biophys. Acta 1585, 114–124.
Powell, D.J., Hajduch, E., Kular, G., Hundal, H.S., 2003. Ceramide disables 3-phosphoinositide binding to the pleckstrin homology domain of proteinkinase B (PKB)/Akt by a PKCzeta-dependent mechanism. Mol. Cell. Biol.23, 7794–7808.
Ruvolo, P.P., 2003. Intracellular signal transduction pathways activated byceramide and its metabolites. Pharmacol. Res. 47, 383–392.
Son, J.H., Yoo, H.H., Kim, D.H., 2007. Activation of the novo syntheticpathway of ceramides is responsible for the initiation of hydrogen peroxide-induced apoptosis in HL-60 cells. J. Toxicol. Environ. Health 70,1310–1318.
Soriano, J.M., Gonzalez, L., Catala, A.I., 2005. Mechanism of action ofsphingolipids and their metabolites in the toxicity of fumonisin B1. Prog.Lipid Res. 44, 345–356.
Velasco, G., Galve-Roperh, I., Sanchez, C., Blazquez, C., Haro, A., Guzman,M., 2005. Cannabinoids and ceramide: two lipids acting hand-by-hand. LifeSci. 77, 1723–1731.
Wang, G., Silva, J., Krishnamurthy, K., Tran, E., Condie, B.G., Bieberich, E.,2005. Direct binding to ceramide activates protein kinase Czeta before theformation of a pro-apoptotic complex with PAR-4 in differentiating stemcells. J. Biol. Chem. 280, 26415–26424.




















