
Submerged Racialization in American 21st-Century Children's ...

陸水学雑誌 (Japanese Journal of Limnology)67: 69- 79(2006)
69
原 著〔Original article〕
2002 年夏の琵琶湖南湖における沈水植物の
現存量と種組成の場所による違い
芳賀 裕樹 1), 大塚 泰介 1), 松田 征也 1), 芦谷美奈子 1)
Spatial distributions of biomass and species composition in submerged macrophytes
in the southern basin of Lake Biwa in summer of 2002
Hiroki HAGA1), Taisuke OHTSUKA1), Masanari MATSUDA1), and Minako ASHIYA1)
Abstract Spatial distributions of the biomass and species composition in submerged macrophytes in the southern basin of Lake Biwa were surveyed in September, 2002. Macrophyte samples were collected from 52 sites by SCUBA. The total biomass (dry weight) of and the vegetated area of macrophytes in the southern basin during the study period were estimated as 10735 ± 3030 t, and 43 km2, respectively. Potamogeton maackianus was dominant, and Hydrilla verticillata, Ceratophyllum demersum, Egeria densa and Myriophyllum spicatum were also abundant in the basin. Biomass of these five species accounted for 99% of the total macrophyte biomass. P. maackianus and C. demersum showed high niche overlap (Pianka's α = 0.71). Biomass of P. maackianus was positively correlated with the average transparency/water depth ratio (Spearman's ρ = 0.50), suggesting that P. maackianus tended to grow at the bottom under strong light. Biomass of C. demersum showed a positive correlation with transparency (ρ = 0.44). Biomass of E. densa was negatively correlated with the sediment diameter (φmean)(ρ = -0.40). The quantitative composition of macrophyte species in 2002 was quite different from that of 1936 when macrophytes were flourishing in the southern basin. Key Words: Lake Biwa, southern basin, submerged macrophyte, biomass, species composition
摘 要
琵琶湖南湖の 52地点で ,2002年 9月に潜水による沈水植物の定量採取を行った。沈水植物は 43地点
に出現した。平均現存量と出現地点数から ,調査時の南湖全体の沈水植物の現存量は 10735 ± 3030 t,分
布面積は 43 km2と推定された。センニンモの現存量が最大で ,これにクロモ ,マツモ ,オオカナダモ ,ホ
ザキノフサモを加えた 5種で沈水植物全体の現存量の 99%を占めた。センニンモとマツモの分布は高い
重なり合いを示した(Pianka's α = 0.71)。沈水植物の現存量と環境要因について Spearmanの順位相関係
数を求めた。センニンモの現存量は平均透明度/水深比との相関が最も強く(ρ = 0.50),湖底の相対的
な光の強さが現存量の大きさを規定する可能性が示された。マツモの現存量は透明度と正の相関を(ρ
= 0.44),オオカナダモの現存量は底質の平均粒径 (φmean)と負の相関を示した(ρ = -0.40)。南湖の沈水
植物の現存量の歴史的変遷を整理した。2002年の沈水植物の量的な種組成は ,かつて沈水植物が豊富だっ
た 1936年と大きく異なっていた。
キーワード:琵琶湖 南湖 沈水植物 現存量 種組成
(2004年 10月 7日受付;2006年 4月 4日受理)
1)滋賀県立琵琶湖博物館 〒 525-0001滋賀県草津市下物町 1091番地 Lake Biwa Museum,Oroshimo-cho 1091,Kusatsu,Shiga 525-0001,Japan

70
芳賀 裕樹ら 2002年夏の琵琶湖南湖における沈水植物の現存量と種組成の場所による違い
71
はじめに
沈水植物は湖沼生態系の主要な基礎生産者であり ,同
時に ,その物理的構造によって湖沼の物質循環や化学
的・物理的環境 ,生物相 ,食物連鎖に大きな影響を与え
る(Carpenter and Lodge, 1986; Scheffer and Jeppesen, 1997;
Takamura et al., 2003)。日本の多くの湖沼では ,富栄養化
や湖岸の改変など人間活動の影響で沈水植物が減少また
は消滅している(Hamabata and Kobayashi, 2002; Takamura
et al., 2003)。ところが ,日本最大の湖である琵琶湖で
は ,1994年以降に沈水植物が増加している(大村ら ,
2001; Hamabata and Kobayashi, 2002)。
琵琶湖の副湖盆である南湖は ,沈水植物が生育可能な
水深 0-7 mの水域の面積の 48%を占め ,琵琶湖全体の沈
水植物現存量の 40 %が存在したことがある(生嶋ら ,
1962)。南湖は琵琶湖で最大規模の沿岸帯といってよく ,
沈水植物の分布上きわめて重要な水域である。南湖の沈
水植物は , 1930- 40年代にはほぼ全域に分布していた
(山口 , 1943)。夏には水深 4 m以深で沈水植物が見ら
れなくなるが(山口 , 1943),その場合でも分布面積は
27 km2程度あったと推定されている(大塚ら , 2004)。
1953年の分布面積は 23 km2(滋賀県水産試験場 , 1954;
生嶋ら , 1962)で 1930- 40年代と同程度だった。しか
し 1964年には最低値 0.6 km2を記録し(生嶋 , 1966),
以後 1994年まで 6 km2以下の状態が続いた(大村ら ,
2001; Hamabata and Kobayashi, 2002)。沈水植物の分布面
積は 1995年から増加に転じ ,1994年に 6 km2だったも
のが 1995年には 9 km2,1997年には 16 km2, 2000年には
29 km2, 2001年には 32 km2と急速に拡大した(滋賀県水
産試験場 , 1998; 大村ら , 2001; Hamabata and Kobayashi,
2002; 大塚ら , 2004)。2001年の沈水植物の現存量(乾燥
重量)は 7100 tと推定され ,記録上では過去最大となっ
た(大塚ら , 2004)。Hamabata and Kobayashi (2002)は ,
1994年以降の沈水植物の増加を回復(recovery)と表現
し , 湖への栄養塩負荷の減少と 1994年と 2000年の夏に
起こった水位低下が沈水植物の回復をもたらしたと述べ
ている。
1994年以降の南湖の沈水植物の種組成については ,
1995年に潜水による深度別の定量採取調査(滋賀県水
産試験場 , 1998), 1997年に潜水による目視ベルトト
ランセクト調査(今本ら , 1998), 2000年に航空写真
と魚群探知機を用いた調査(Hamabata and Kobayashi,
2002)が行われている。1995年の調査では(滋賀県水
産試験場 , 1998),沈水植物全体の現存量は 2500 tと推
定され ,コカナダモ (Elodea nuttallii)が全体の 42%を
占め優占種であること ,ついでセンニンモ (Potamogeton
maackianus)が全体の 38%を占めることが報告された。
1997年の調査では (今本ら , 1998),コドラート内で優
占種となった頻度はセンニンモが 25%で最も高く ,以
下 ,クロモ(Hydrilla verticillata),マツモ (Ceratophyllum
demersum),ホザキノフサモ (Myriophyllum spicatum),コ
カナダモの順だった。
2000年の調査では(Hamabata and Kobayashi, 2002),
クロモ ,オオカナダモ(Egeria densa),ホザキノフサモ
が南湖の主要な沈水植物であった。これらの結果から ,
1995年から 2000年の間に主要な沈水植物が交代したこ
とが示唆されるが ,調査方法が異なるために厳密な比較
はできない。また ,南湖全体の沈水植物の現存量が 1995
年から 2001年の間に 2.8倍に増加していることから ,順
位だけでなく個別の種の現存量も変化していることが予
想される。従って南湖の沈水植物の現状を把握し ,過去
の状況と比較するには 1995年の調査と比較可能な定量
採取調査が必要である。
沈水植物の種別の水平分布と環境要因の対応について
は ,1986年に浜端(1991 a, b)が ,1997年に今本ら(1998)
が解析を行っている。これらの解析では出現頻度や被度
が用いられており ,現存量はまだ検討されていない。種
別の現存量の分布は出現頻度や被度の分布とともに沈水
植物群落の形成過程を考える重要な指標である。
本研究では ,南湖の沈水植物の種別の現存量を把握し ,
種別の現存量の水平分布ならびに現存量の分布と環境要
因の対応を明らかにすることを目的として 2002年 9月
に潜水による定量採取を行った。また ,得られた結果を
過去の記録と比較し ,沈水植物の量的な変遷に関する検
討も行った。
方 法
沈水植物の採取は 2002年 9月 9日から 11日にかけ
て行った。9月は南湖の沈水植物の現存量が年間で最大
に達する時期である(生嶋 , 1966; 大村ら , 2001)。南湖
全域を 1 kmメッシュに区切り ,その格子点 52地点を採
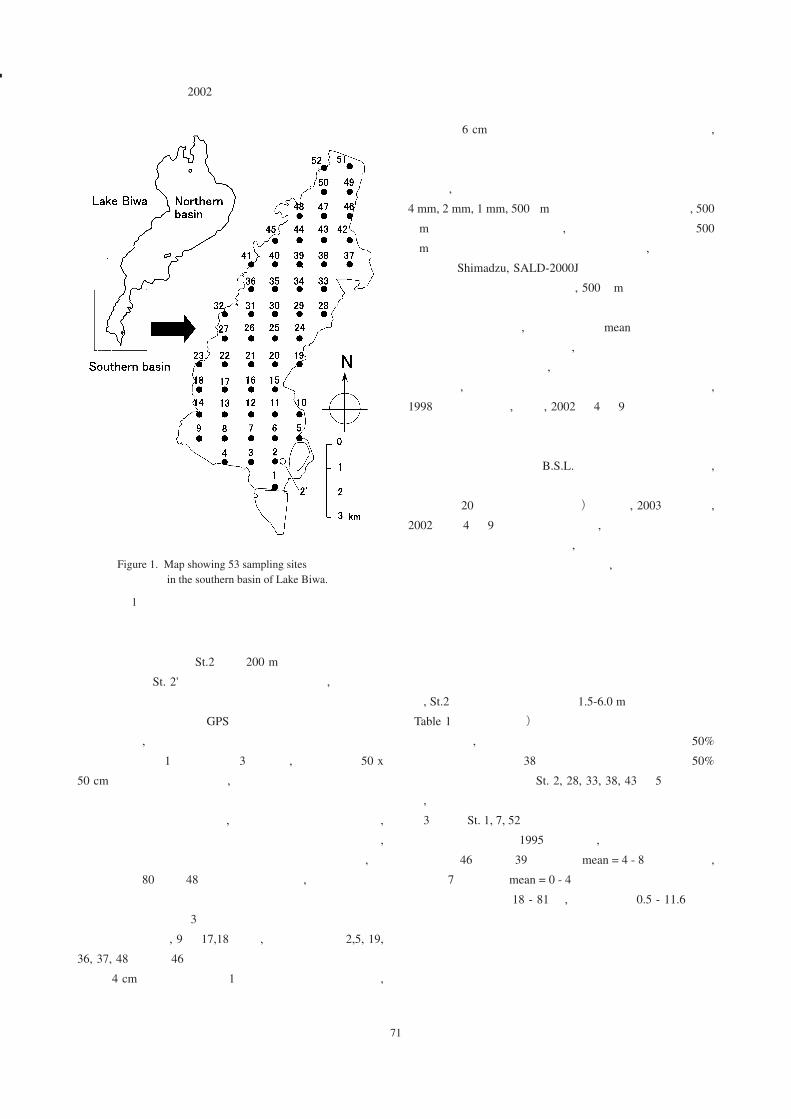
取地点とした(Fig. 1)。St. 2は東西 500 m×南北 500 m,
水深 12 mの浚渫くぼ地と重なった。この地点は水深が
著しく深いため ,潜水による沈水植物の採取は行わず ,
魚群探知機の映像で沈水植物がないことを確認した。
St.2のデータは沈水植物の現存量と分布面積の推定に用
いた。しかし ,人為的に作られた特異な地形であるため ,
70
芳賀 裕樹ら 2002年夏の琵琶湖南湖における沈水植物の現存量と種組成の場所による違い
71
沈水植物の分布と環境要因の関係の検討には用いなかっ
た。参考記録として St.2の東 200 mの位置で沈水植物
を採取した(St. 2')。記録は解析には用いず ,結果のみ
表に示した。
ディファレンシャル GPSを用いて調査船を調査地点
に到達させ ,アンカーで係留して潜水によるコドラート
採取を行った。1地点につき 3回ずつ ,ランダムに 50 x
50 cmのコドラートを置き ,その中に生えている沈水植
物を根も含めて採取した。採取した沈水植物は水中でナ
イロン製の土のう袋につめ ,船上で水を切って持ち帰り ,
分別まで冷蔵庫で保管した。試料を実験室で水洗いの後 ,
種類別に分別した。分別後の試料の湿重量を測定後 ,紙
袋に入れて 80 ℃で 48時間以上乾燥させ ,恒量になって
から乾燥重量を測定した。全ての試料の分別と計量が終
了するまでの期間は 3ヶ月だった。
底質の採取は , 9月 17,18日に ,調査地点番号 2,5, 19,
36, 37, 48を除く 46地点で行った。簡易コアサンプラー
(直径 4 cm)による採取を 1地点につき1本ずつ行い ,
表層から 6 cmの深さまでの泥をスパーテルで採取し ,
プラスチック製のサンプル容器に密封して分析まで冷蔵
庫で保存した。試料の一部を用いて含水比と強熱減量を
測定し ,残りの試料を用いて粒度組成の分析を行った。
4 mm, 2 mm, 1 mm, 500 μmの金属篩で底質を分別し , 500
μm以上の画分については ,乾燥重量を計測した。500
μmのメッシュを通過した底質については ,粒度分布測
定装置(Shimadzu, SALD-2000J)を用いてサイズ画分ご
との粒子数の寄与率を算出し , 500 μm未満の底質の乾
燥重量を乗じて各サイズ画分の乾燥重量を推定した。以
上のデータを統合し ,平均粒径(φ mean)を求めた。
前述の底質の分析のほかに , 沈水植物の現存量分布と
対応する環境要因として ,各採取地点から最も近い岸ま
での距離 ,夏に南湖で卓越する北風および西風(今本ら ,
1998)の吹送距離 ,水深 , 2002年 4- 9月の平均透明度
を求めた。岸までの距離と吹送距離は地図上で算出した。
水深は調査時に測深錘で計測した値を当日の平均水位を
用いて琵琶湖基準水位〔B.S.L.〕に換算した。透明度は ,
滋賀県および国土交通省近畿地方建設局が毎月行って
いる南湖 20地点の観測データ (滋賀県 , 2003)を用い ,
2002年の 4- 9月データを平均し ,スプライン外挿法で
調査地点の値を求めた。また ,平均透明度を水深で除し
た値(平均透明度/水深比)を求め ,湖底の相対的な光
の強度とした。
結 果
調査地点の環境
琵琶湖基準水位を水面とした場合の調査地点の水深
は , St.2の浚渫くぼ地を除けば 1.5-6.0 mの範囲にあった
(Table 1)。底質は泥 (シルトおよび粘土 )と砂が主な
成分であり ,一部にれきが混ざった。底質の重量の 50%
を泥が占めた地点は 38地点だった。底質の重量の 50%
以上を砂が占めたのは St. 2, 28, 33, 38, 43の 5地点であ
り ,いずれも南湖の中央よりも東岸側に位置した。残り
の 3地点(St. 1, 7, 52)は泥・砂・れきが混合した底質だっ
た。西條・三田村(1995)に従い ,平均粒径で底質を区
分すると 46地点中 39地点がφ mean = 4 - 8のシルト質 ,
残りの 7地点がφ mean = 0 - 4の砂質に区分された。
底質の含水比は 18 - 81% ,強熱減量は 0.5 - 11.6 %の
範囲にあった。
Figure 1. Map showing 53 sampling sites in the southern basin of Lake Biwa.
図 1. 琵琶湖南湖における調査地点

72
芳賀 裕樹ら 2002年夏の琵琶湖南湖における沈水植物の現存量と種組成の場所による違い
73
Table 1. Water depths (B.S.L.) and sediment characteristics at the 52 sampling sites
in the southern basin of Lake Biwa on September 2002.
表 1. 2002年 9月の南湖 52調査地点における水深(琵琶湖基準水位時に換算),ならびに底泥の性状
Station number
Water depth
Sediment characteristics
Sediment composition (weight %) φ meanWater
contentIgnision
loss
(m) Clay(φ >=8)
Silt(φ =<8-4)
Sand(φ =<4-0)
Grain (φ<0) (%) (%)
1 2.5 3 18 40 39 0.8 13 0.52 11.8 - - - - - - -3 3.8 28 62 10 1 6.7 70 7.74 3.2 10 60 30 0 5.2 73 10.85 2.0 - - - - - - -6 4.1 17 67 16 0 6 76 8.67 4.2 4 19 46 31 6 77 8.48 4.5 15 68 17 0 5.9 77 9.59 3.8 10 52 21 17 3.7 75 11.6
10 2.5 12 72 17 0 5.7 70 7.611 4.0 20 71 10 0 6.4 71 8.612 4.6 17 73 10 0 6.2 76 8.913 4.6 18 75 7 0 6.3 79 914 4.0 13 65 22 0 5.7 69 9.315 2.5 10 53 36 1 5 59 5.416 4.4 17 74 9 0 6.3 71 8.617 4.5 15 74 11 0 6.1 76 8.418 3.4 11 69 20 0 5.6 79 10.019 1.5 - - - - - - -20 3.8 21 69 10 0 6.4 66 7.321 4.5 24 70 6 0 6.7 72 7.622 4.6 18 74 8 0 6.4 73 8.323 3.2 17 66 16 0 6 68 9.724 2.8 15 58 27 0 5.6 76 8.025 4.5 27 62 10 0 6.6 69 6.826 4.7 19 75 6 0 6.4 81 7.627 4.0 12 74 14 0 5.8 81 9.328 2.3 0 42 58 0 1.9 28 0.929 4.5 11 58 31 0 5.3 55 5.130 4.0 33 65 1 0 7.3 68 5.831 5.0 18 76 5 0 6.4 64 6.832 3.2 6 53 41 0 4.7 72 8.533 3.5 0 16 72 12 1.8 33 2.634 4.4 31 61 8 0 6.9 68.3 6.335 4.3 16 65 17 2 5.9 71.3 8.136 3.7 - - - - - - -37 2.0 - - - - - - -38 2.9 2 12 56 29 1.1 30.5 2.839 3.8 15 62 23 0 5.7 70.6 6.940 4.9 17 73 10 0 6.3 72.0 7.841 2.5 12 68 19 0 5.7 69.4 6.842 2.4 10 56 34 0 5.1 52.5 3.843 3.5 4 23 63 10 2.2 35.5 1.844 4.5 15 69 16 0 6 72.0 7.945 3.0 13 67 19 0 5.8 67.1 6.646 2.4 11 61 28 0 5.3 43.6 3.747 4.2 12 64 24 0 5.6 63.1 6.848 5.2 - - - - - - -49 3.5 9 40 44 7 4.1 58.3 5.950 6.0 19 71 11 0 6.3 62.7 6.651 5.2 12 69 19 0 5.7 68.0 7.452 3.3 5 32 25 37 3.4 55.6 5.9

72
芳賀 裕樹ら 2002年夏の琵琶湖南湖における沈水植物の現存量と種組成の場所による違い
73
沈水植物の現存量と分布
沈水植物は 52地点中 43地点に出現した (Fig. 2, Table
2,ただし参考地点 St. 2'を除く )。沈水植物が分布しな
かった地点は東岸の 2地点と St.2の浚渫くぼ地を除き ,
南湖中央部に集中した (Fig. 2)。沈水植物の現存量は南
部と北部で大きく ,現存量が特に大きな地点は北部に多
く見られた。
全調査地点の沈水植物の現存量の平均値と 95%信頼
限界は 208 ± 59 g m-2だった。現存量の平均値に南湖の
水面面積 51.6 km2(芳賀 , 2006)を乗じて南湖全体の沈
水植物の現存量を推定すると 10735 ± 3030 tとなった。
なお ,この値は沈水植物と共に採取された糸状藻類を除
いた値である。糸状藻類も含めた場合の現存量は 11239
± 3142 t だった。
全調査地点に対する沈水植物出現地点の比率から ,調
査時の沈水植物の分布面積は 43 km2と推定された。
出現した沈水植物の種類と現存量
調査時にコドラート内に出現した沈水植物はセンニン
モ ,クロモ ,マツモ ,オオカナダモ ,ホザキノフサモ ,コ
カナダモ ,ネジレモ (Vallisneria asiatica var. biwaensis),
コウガイモ (Vallisneria denseserrulata),オオササエビモ
(Potamogeton anguillanus)の 9種だった(Table 2)。ササ
バモ(Potamogeton malaianus)はしばしば観察されたがコ
ドラート内には出現しなかった。
沈水植物が出現した地点では , 2種から 7種が同時に
出現し ,平均出現種数は 4.6だった。コドラートあたり
では 1種から 7種出現し ,平均出現種数は 3.6だった。
種別の出現地点数はセンニンモが 41地点で最も多く ,
以下クロモ(40地点),マツモ(38地点),オオカナダモ(33
地点),ホザキノフサモ(25地点)の順となった(Table 2)。
現存量で優占した地点数もセンニンモが 21地点で最も
多く ,ついでクロモ(15地点),オオカナダモ(4地点),
マツモ(2地点),ホザキノフサモ(1地点)の順となった。
出現地点における平均現存量はセンニンモが 110 g m-2
で最大となり ,続いてクロモ(57 g m-2), オオカナダモ(39
g m-2),マツモ (35 g m-2), ホザキノフサモ (31 g m-2)の順
となった。
各沈水植物について ,沈水植物が分布しなかった地点
も含めて平均現存量を算出し ,南湖の水面面積を乗じて
南湖全体での現存量を推定した。センニンモが 4827 t,
クロモが 2417 t,マツモが 1424 t, オオカナダモが 1088 t,
ホザキノフサモが 820 tとなり ,これら 5種類で沈水植
物全体の現存量の 99%を占めた。コウガイモ ,オオササ
エビモ , コカナダモ ,ネジレモの現存量はそれぞれ 67 t,
39 t, 41 t, 12 tであり ,前述の 5種類に較べて 1桁から 2
桁小さい値をとった。沈水植物ではないが ,糸状藻類(複
数の種類を含む)は 23地点に出現し ,南湖全体におけ
る現存量は 504 tで ,出現地点数 ,現存量とも主要な 5種
の沈水植物に次ぐ位置を占めた。糸状藻類は沈水植物の
ない地点には出現しなかった。
主要な沈水植物種の現存量の分布
量的に主要な 5種類の沈水植物の分布は種によって異
なった(Fig. 3)。センニンモは南部より北部 ,沖よりも
沿岸に現存量の大きい地点が多く出現した。これに対し
クロモは北部の 3地点を除き ,沿岸よりも沖に現存量の
大きな地点が分布した。マツモは西岸沿いの 3地点で現
存量が大きかったが ,他の地点では現存量は小さかった。
ホザキノフサモは北部に現存量の大きな地点が集中し ,
南部では出現しても現存量は小さかった。オオカナダモ
は南部に現存量の多い地点が集中し ,北部東岸や西岸の
一部にやや現存量の大きな地点があった。
Figure 2. Distribution of total macrophyte biomass. Length of the bar represents biomass of macrophyte.
図 2.沈水植物全体の現存量分布。 棒の長さは現存量を表している。
74
芳賀 裕樹ら 2002年夏の琵琶湖南湖における沈水植物の現存量と種組成の場所による違い
75
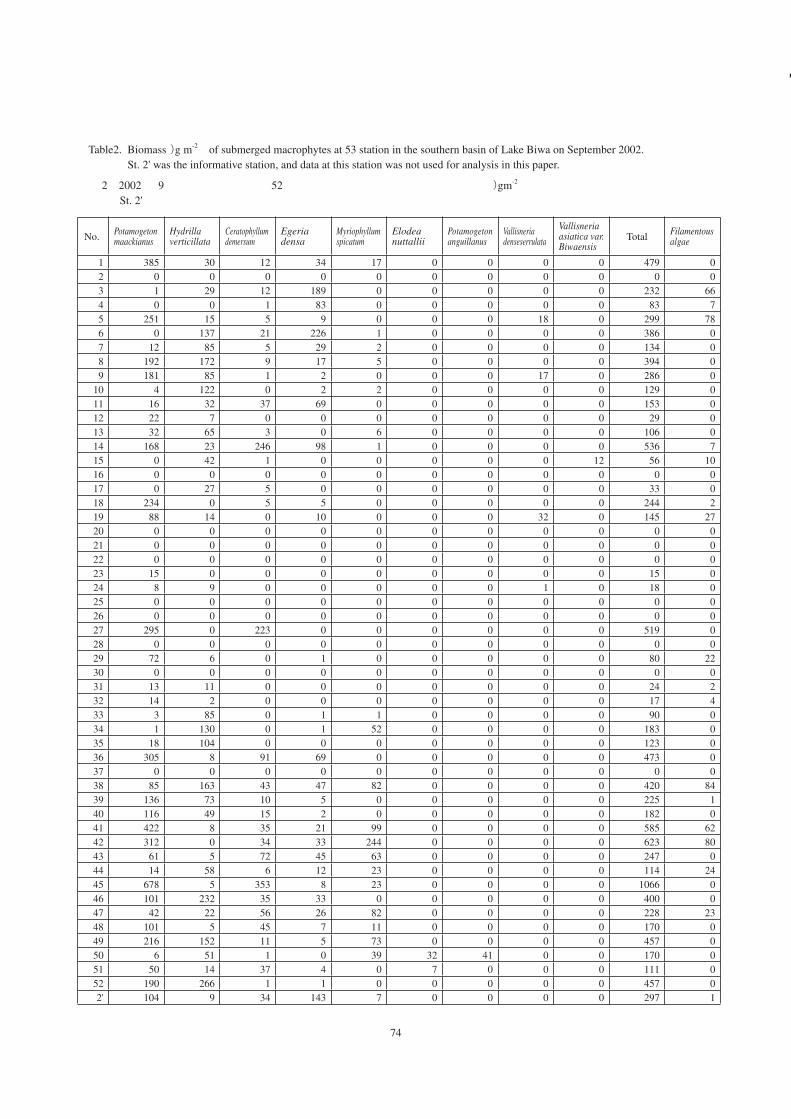
Table2. Biomass (g m-2 ) of submerged macrophytes at 53 station in the southern basin of Lake Biwa on September 2002. St. 2' was the informative station, and data at this station was not used for analysis in this paper.
表 2.2002年 9月に観測された南湖 52地点における種類別の沈水植物の現存量 (gm-2 )。 St. 2'は参考地点で、データは本文中の解析には用いていない。
No. Potamogeton maackianus
Hydrilla verticillata
Ceratophyllum demersum
Egeria densa
Myriophyllum spicatum
Elodea nuttallii
Potamogeton anguillanus
Vallisneria denseserrulata
Vallisneria asiatica var. Biwaensis
Total Filamentous algae
1 385 30 12 34 17 0 0 0 0 479 0 2 0 0 0 0 0 0 0 0 0 0 0 3 1 29 12 189 0 0 0 0 0 232 66 4 0 0 1 83 0 0 0 0 0 83 7 5 251 15 5 9 0 0 0 18 0 299 78 6 0 137 21 226 1 0 0 0 0 386 0 7 12 85 5 29 2 0 0 0 0 134 0 8 192 172 9 17 5 0 0 0 0 394 0 9 181 85 1 2 0 0 0 17 0 286 0
10 4 122 0 2 2 0 0 0 0 129 0 11 16 32 37 69 0 0 0 0 0 153 0 12 22 7 0 0 0 0 0 0 0 29 0 13 32 65 3 0 6 0 0 0 0 106 0 14 168 23 246 98 1 0 0 0 0 536 7 15 0 42 1 0 0 0 0 0 12 56 10 16 0 0 0 0 0 0 0 0 0 0 017 0 27 5 0 0 0 0 0 0 33 018 234 0 5 5 0 0 0 0 0 244 219 88 14 0 10 0 0 0 32 0 145 2720 0 0 0 0 0 0 0 0 0 0 021 0 0 0 0 0 0 0 0 0 0 022 0 0 0 0 0 0 0 0 0 0 023 15 0 0 0 0 0 0 0 0 15 024 8 9 0 0 0 0 0 1 0 18 025 0 0 0 0 0 0 0 0 0 0 026 0 0 0 0 0 0 0 0 0 0 027 295 0 223 0 0 0 0 0 0 519 028 0 0 0 0 0 0 0 0 0 0 029 72 6 0 1 0 0 0 0 0 80 2230 0 0 0 0 0 0 0 0 0 0 031 13 11 0 0 0 0 0 0 0 24 232 14 2 0 0 0 0 0 0 0 17 433 3 85 0 1 1 0 0 0 0 90 034 1 130 0 1 52 0 0 0 0 183 035 18 104 0 0 0 0 0 0 0 123 036 305 8 91 69 0 0 0 0 0 473 037 0 0 0 0 0 0 0 0 0 0 038 85 163 43 47 82 0 0 0 0 420 8439 136 73 10 5 0 0 0 0 0 225 140 116 49 15 2 0 0 0 0 0 182 041 422 8 35 21 99 0 0 0 0 585 6242 312 0 34 33 244 0 0 0 0 623 8043 61 5 72 45 63 0 0 0 0 247 044 14 58 6 12 23 0 0 0 0 114 2445 678 5 353 8 23 0 0 0 0 1066 046 101 232 35 33 0 0 0 0 0 400 047 42 22 56 26 82 0 0 0 0 228 2348 101 5 45 7 11 0 0 0 0 170 049 216 152 11 5 73 0 0 0 0 457 050 6 51 1 0 39 32 41 0 0 170 051 50 14 37 4 0 7 0 0 0 111 052 190 266 1 1 0 0 0 0 0 457 02' 104 9 34 143 7 0 0 0 0 297 1

74
芳賀 裕樹ら 2002年夏の琵琶湖南湖における沈水植物の現存量と種組成の場所による違い
75
考 察
2002 年の南湖の沈水植物の現存量と分布範囲
大塚ら(2004)は 2001年の南湖における沈水植物の
分布面積と現存量を ,それぞれ 32km2と 7100 t と推定し
た。これらの値は南湖の水面面積を実際よりも大きく見
積もったために過大評価になっており ,実際の沈水植物
の分布面積と現存量はそれぞれ 29 km2, 6500 tに修正さ
れる(芳賀 , 2006)。いずれにせよ ,本調査で明らかになっ
た 2002年の分布面積 43 km2と現存量 10735 ± 3030 tは
2001年の推定値を大きく上回った。
大塚ら(2004)は透明度と水深 ,補償深度/水深比を
用いて沈水植物が分布可能な水域を推定するモデルを作
成し ,2001年 7月には分布可能な水域のほとんどに沈水
植物が進出したと述べた。大塚ら(2004)の沈水植物の
分布範囲と本調査の結果を比較すると , 8地点で新たに
沈水植物が出現していた(地点番号 12, 13, 17, 22, 29, 31,
33, 34)。調査時に沈水植物が根付いていたことを確認し
ているので ,これらはすべて新たな分布地点と考えられ
る。この分布域の拡大が大塚ら(2004)のモデルで説明
可能かどうかを , 2002年の透明度と水位のデータ(滋賀
県 , 2003)を用いて検討した。新たに沈水植物が出現し
た 8地点のうち ,大塚ら(2004)のモデルで分布可能と
推定されたのは 3地点に過ぎなかった。この矛盾は ,大
塚ら(2004)のモデルで用いられた光の条件が必要以上
に厳しかったことを示唆する。まず補償深度/水深比だ
が ,一般に沈水植物が生長可能な補償深度は透明度の 2
倍とされ(生嶋ら , 1962; 生嶋 , 1972; Chambers and Kalff,
1985),水温が上昇する夏にはこの比は低下する(生嶋 ,
1972)。従って , 補償深度/水深比を大塚ら(2004)の
用いた 2.0より大きくすることは妥当ではない。大塚ら
(2004)は ,沈水植物の初期成長には湖底に光が十分に
届く期間が 2- 3ヶ月程度必要と仮定し , 4- 8月の有
光深度の中央値を分布の可否の推定に用いた。この仮定
条件を緩和し ,上位から 2番目の有光深度を沈水植物の
分布の可否の推定に用いると , 2002年の新たな分布地点
8地点のうち 6地点までが分布可能と推定された。この
結果は ,沈水植物の初期成長に必要な ,湖底に光が十分
に届く期間が大塚ら(2004)の仮定より短いことを示し
ている。2002年の沈水植物の分布範囲が限界に達した
とする根拠はないので ,分布はさらに拡大する可能性も
ある。その場合 ,沈水植物の分布に必要な ,湖底に光が
十分に届く期間の長さは ,さらに短く ,数週間~ 1ヶ月
程度となる。
2002年の南湖全体の沈水植物の現存量の推定値は
2001年のそれより約 4200 t大きくなった。調査方法は
異なるものの ,この増加幅は 95%信頼限界の幅より大き
く ,現存量が増加したことは間違いないと思われる。新
たな沈水植物の分布地点の現存量をゼロとした場合の南
湖全体の現存量を実際の現存量の推定値から差し引き ,
沈水植物の分布の拡大によって増加した現存量を計算す
ると 635 tとなった。この結果は , 2002年と 2001年の
現存量の差が ,分布の拡大だけでは説明できないことを
示している。2002年の沈水植物の分布地点のうち , 2001
年にも沈水植物が分布していたと考えられる 35地点に
ついて平均現存量を計算すると 252± 64 g m-2となった。
一方 , 2001年の沈水植物の分布範囲における平均現存量
は 224 g m-2と推定された。これらの結果から ,統計的に
有意な差はないものの ,既存の分布範囲でも沈水植物の
現存量が増加した可能性が示された。大塚ら(2004)は ,
南湖の沈水植物の現存量は 9月頃に最大に達することか
Figure 3. Biomass distribution of the five major macrophyte species. Length of the bar represents biomass of macrophyte.
図 3. 主要な沈水植物 5種の現存量分布。 棒の長さは現存量を表している。

76
芳賀 裕樹ら 2002年夏の琵琶湖南湖における沈水植物の現存量と種組成の場所による違い
77
ら 7月末に観測された 2001年の値は年間の最大現存量
より小かった可能性があると述べた。2002年に見られ
た既存分布地点の現存量の増加の原因には ,観測した月
の違いも含まれる可能性がある。
沈水植物の種組成
本調査で出現した沈水植物は 9種であり ,1995年の 12
種(滋賀県水産試験場 , 1998),1997年の 18種(今本ら ,
1998)よりは少なかった。調査手法や調査地点数が類似
している水産試験場の 1995年の調査と比較すると ,本
調査で出現した 9種はふたつの調査に共通し ,ササバモ ,
イバラモ(Najas marina),シャジクモ類 (Characeae)が
本調査で出現しなかった。ただし ,ササバモは採取され
なかったものの ,存在は目視で確認されている。
沈水植物が分布した地点では ,1地点あたりの出現種
数は 4.6,コドラートあたりの平均出現種数は 3.6であっ
た。1種のみ出現した地点はなく ,また 1種しか出現し
なかったコドラートも 11%にとどまった。これらの数
値から ,南湖の沈水植物群落は基本的に混合群落である
と考えられた。過去の記録においても 1995年の調査(滋
賀県水産試験場 , 1998)で 2種以上出現した地点は全体
の 86%を占めている。また ,生嶋ら(1962)は 1953年
の琵琶湖の沈水植物について 50× 50 cm あるいは 1×
1 m のコドラート内には同時に 3種出現する場合が最も
多いことから ,琵琶湖の沈水植物の植生は混合群落が基
本だと述べている。
量的に主要な 5種の沈水植物について Pianka (1973)
の式によりニッチの重なり合い指数を計算した (Table
3)。センニンモとマツモの重なり合い指数は大きく(α
= 0.71),両者が共存する傾向が示された。マツモとク
ロモ(α = 0.13)の重なり合い指数は小さかった。クロ
モとマツモが共に出現した地点は少なくなかったが ,二
つの沈水植物の現存量がともに大きい地点はなかった
(Fig. 3)。クロモとマツモは成育に適した環境が異なる
か ,または競合関係があるかのいずれかであることが示
唆された。
沈水植物の現存量分布と環境要因
主要な 5種の沈水植物の現存量分布と環境要因の関係
を検討した (Table 4)。種によって現存量がゼロとなる
地点が多く現れ ,データが二変量正規分布に従うことが
期待できないため ,相関の指標には Spearmanの順位相
関係数を用いた。
センニンモの現存量は ,平均透明度/水深比と最も強
い相関を示し(ρ = 0.50),湖底の相対的な光の強さが
大きいほど ,センニンモの現存量が大きくなる傾向が示
された。センニンモの現存量は平均透明度とも正の相
関を示した (ρ = 0.45)。一方 ,センニンモの現存量は
西風および北風の吹送距離とは負の相関を示した(ρ =
-0.35, -0.43)。西風および北風の吹送距離と平均透明度
の間には負の相関があり(ρ =-0.65, -0.63),風の影響が
弱い南湖北部や西岸沿いで平均透明度が高い傾向が見ら
Table 4. Spearman's rank correlation coefficients between biomass of five major macrophytes and environmental factors.
表 4.量的に主要な 5種の沈水植物の現存量と環境要因の間の Spearmanの順位相関係数
Potamogeton maackianus
Hydrilla verticillata
Ceratophyllum demersum Egeria densa Myriophyllum
spicatumPhysical parameters (n = 51)
Transparency (m) 0.45 0.28 0.44 0.24 0.32
Water depth (m) -0.22 0.06 -0.05 -0.28 0.06
Transparency / Water depth 0.50 0.13 0.27 0.39 0.16
Distance to the shore (km) -0.29 0.10 -0.14 -0.19 0.17
Fetch for west wind (km) -0.35 0.07 -0.31 -0.06 -0.08
Fetch for north wind (km) -0.43 0.06 -0.25 0.10 -0.19
Sediment characteristics (n = 46)
Water content (%) -0.13 -0.14 -0.09 -0.23 -0.26
Ignition loss (%) -0.03 -0.13 -0.01 -0.11 -0.26
Mean diameter(φmean) -0.45 -0.24 -0.31 -0.40 -0.25
Table 3. Pianka's niche overlap index between five major macrophyte species.
表 3. 量的に主要な 5種の沈水植物に関する Pianka (1973)のニッチ重なり合い指数
Potamogeton maackianus
Hydrilla verticillata
Egeria densa
Hydrilla verticillata 0.33
Ceratophyllum demersum 0.71 0.13
Egeria densa 0.24 0.34 0.30
Myriophyllum spicatum 0.46 0.23 0.22 0.19

76
芳賀 裕樹ら 2002年夏の琵琶湖南湖における沈水植物の現存量と種組成の場所による違い
77
れた。これらの結果から ,南湖北部や西岸の沿岸部でセ
ンニンモの現存量が大きい傾向は(Fig. 3),直接的には
平均透明度が高くて湖底が明るいこと ,間接的には風の
影響を受けにくいことが原因であると考えられた。セン
ニンモの現存量と底質の平均粒径 (φ mean)は負の相
関(ρ =- 0.45)を示し ,底質の平均粒径が大きいほどセ
ンニンモの現存量が大きい傾向が見られた。底質の平均
粒径と ,透明度/水深比あるいは透明度との関係は明ら
かではなかった。
マツモの現存量は透明度と正の相関を示した(ρ =
0.44)。オオカナダモの現存量は底質の平均粒径と負の
相関を示した(ρ = -0.40)。クロモとホザキノフサモの
現存量は ,本研究で取り上げた環境要因と明らかな相関
を示さなかった。
南湖における沈水植物組成の歴史変遷
本調査で得られた 2002年 9月の南湖における沈水植
物の現存量と種組成を過去の記録と比較し ,その位置づ
けを試みた(Table 5)。表中の 1936年のデータは ,南湖
南部(本研究における St. 1 - 14に相当)で得られた水
深ごとの平均現存量密度(山口 , 1938)を南湖全体に適
用可能と仮定して ,1953年の深度別湖底面積(滋賀県水
産試験場 , 1954)を乗じて算出した。滋賀県水産試験場
(1972, 1998)のデータについては湿重量で表示されてい
るので ,本研究で得られた乾湿重量比の平均値 0.086を
用いて乾重量に換算した。報告ごとに南湖の面積が異な
るため ,表中にそれらの値を示した。ただし ,面積が異
なっても以下の議論に大きな影響はないので ,特に補正
は行わないことを断っておく。
山口(1938)の報告から推定された 1936年の南湖の
沈水植物の現存量は約 4000 t である(Table 5)。1953年
から 1970年代半ばまでの沈水植物の現存量は 500 - 800
tと低い状態が続いた。1953年の現存量は沈水植物の衰
退期にあたる 11月に観測され ,過小評価の可能性があ
る(大塚ら , 2004)。また ,1964年の著しい現存量の低
下は湖岸の埋め立てで発生した濁水が原因とされてい
る(生嶋 , 1966)。このため ,沈水植物の現存量が減少し
た正確な時期や ,直接的な原因は不明である。ただし ,
Table 5. Temporal change in macrophyte biomass (t) in the southern basin of Lake Biwa.
表 5.琵琶湖南湖の沈水植物の現存量 (t)の歴史的変遷
Investigation periods 1936 1958 1964 1969 1974 1995 2001 2002
P. maackianus 521 <1 11 962 4827
Potamogeton perfoliatus 58
P. malaianus 93 17 7
Potamogeton anguillanus 28 41
H. verticillata 442 313 11 156 2417
C. demersum 838 110 75 1424
M. spicatum 307 2 3 820
V. denseserrulata 505 5 91 67
V. asiatica var. biwaensis 788 200 36 12
Najas marina 352 30 5
Najas japonica 13
E. densa 621 83 1088
E. nuttallii 118 23 1053 39
Characeae 23 6 1
filamentous algae 508
Total 3940 553 11 802 666 2501 **6500 ***10735
Area of southern basin 57 57 - 55 - 52 **51.6 51.6
Reference * 1 2 3 4 5 6 7 8
* 1, estimated from Yamaguti (1938); 2, Shiga Prefectural Fisheries and Experimental Station (1954), Ikusima et al.(1962); 3, Ikusima(1966), 4, Shiga Prefectural Fisheries and Experimental Station (1972), 5, Tanimizu and Miura (1976), 6, Shiga Prefectural Fisheries and Experimental Station (1998), 7, Ohtsuka et al. (2004); 8, This study.
** Recalculated using area of 51.6 km2.***Biomass of filamentous algae was not included.
* 1, 山口(1938)より推定 ; 2, 滋賀県水産試験場(1954), 生嶋ら (1962); 3, 生嶋(1966); 4, 滋賀県水産試験場(1972); 5, 谷水・三浦 (1976); 6, 滋賀県水産試験場 (1998); 7, 大塚ら (2004); 8, 本調査。
** 水面面積を 51.6 km2として再計算。*** 糸状藻類の現存量を含まない。

78
芳賀 裕樹ら 2002年夏の琵琶湖南湖における沈水植物の現存量と種組成の場所による違い
79
少なくとも 1960年代半ばには沈水植物の現存量が少な
い期間が始まっていたと思われる。1960年代後半には
コカナダモ ,1970年代半ばにはオオカナダモといった
外来性の沈水植物が爆発的に増加したが(谷水・三浦 ,
1976),沈水植物全体の現存量は 1936年よりも低いまま
だった。1994年の渇水による水位低下を契機として沈
水植物の現存量は増加に転じ(Hamabata and Kobayashi,
2002),1995年には 1936年の現存量の 6割に相当する
2500 tが記録された。1995年以降も沈水植物の現存量は
増加し続け ,本調査が行われた 2002年には 1936年の 2.7
倍の現存量が観測された。
次に量的な種組成の変遷について述べる。まず量的
に主要な沈水植物の種数に注目し , Table 5から全体の
90%以上を占めるのに必要な種数を計算すると 1936年
は 7種 ,1969年は 4種 ,1974年は 1種 ,1995年は 2種 ,2002
年は 5種となった。これらの結果から ,南湖の沈水植物
相は 1971年に最も単純化が進み ,その後最近になって
再び多様な状態に戻ったことがわかる。量的に主要な種
を 1936年と 2002年で比較すると ,共通なのはセンニン
モ ,クロモ ,マツモ ,ホザキノフサモである。これらの
種の 2002年の現存量は 1936年のそれよりむしろ大きく
なっている。一方 ,ネジレモとコウガイモ ,イバラモ ,サ
サバモの現存量は 2002年には著しく低下している。こ
のように ,1936年に量的に主要だった 7種の消長は ,2002
年に現存量が増加したグループと減少したグループに二
分される。ロゼット型のネジレモやコウガイモ ,あるい
は有茎型でも背丈の低いイバラモは ,有茎型で茎を長く
伸ばすセンニンモやクロモ ,ホザキノフサモに比べて透
明度の低下に弱いと考えられている(浜端 ,1996)。2002
年の透明度が 1936年に及ばないと仮定すれば ,センニ
ンモ ,クロモ ,マツモ ,ホザキノフサモの現存量が増え ,
ネジレモ ,コウガイモ ,イバラモの現存量が回復しない
原因の説明になる。しかし ,1936年には夏期には水深
4 mより深所に沈水植物がまれだったのに対し (山口 ,
1938),2002年には水深 4 mより深所にセンニンモやク
ロモの進出が見られた(Tables 1, 2)。このため ,1936年
に比べて 2002年の透明度が低いという説は根拠が弱い。
ほかの環境要因についても情報が不足しており ,現時点
では在来種が現存量の増減でふたつのグループに分かれ
た原因は明らかではない。
外来種であるオオカナダモやコカナダモは比較的短期
間に現存量が著しく変動している (Table 5)。これらの
種の現存量が変動する原因はわからないが ,その変動は
今後の南湖の沈水植物の現存量や種組成に大きな影響を
与えるかもしれない。2002年の沈水植物全体の現存量
に占めるオオカナダモの割合は 11%に過ぎなかったが ,
その現存量は 1088 t であり ,オオカナダモが沈水植物全
体の 93%を占めた 1974年の現存量より 2倍近く大きい。
また 1088 tという現存量は 1936年のいずれの在来種の
現存量よりも大きい。
南湖の沈水植物は 1950年代から 1990年代前半に
一度衰退し ,1994 年以降に回復した(Hamabata and
Kobayashi, 2002)。ただし ,2002年と 1936年を比較する
と ,沈水植物全体の現存量が 2.7倍になっている点 ,在
来種の中で現存量が増えたグループと減ったグループが
分かれた点 ,および ,外来種であるオオカナダモが量的
に主要な種として加わっている点が異なっている。南湖
の沈水植物の量的な組成が 1936年の状態に近づくのか ,
それとも異なる方向に進むのかを見守るため ,今後も継
続的な調査が必要であろう。
謝 辞
調査にあたって ,中山隆志 ,町田典三 ,保井浩二の各
氏に採集の協力をいただいた。底質の分析では ,滋賀県
立大学の分析機器をお借りするとともに三田村緒佐武教
授 ,吉田晴子さんに分析時の助力をいただいた。滋賀県
河港課からは ,水位のデータをいただいた。早狩進氏に
は ,粒度分析データ解析用のエクセルアドインの提供を
いただいた。論文の執筆にあたって琵琶湖博物館の楠岡
泰主任学芸員 , M. J. Grygier専門学芸員に貴重な助言を
いただいた。本研究の一部は ,河川環境管理財団平成 14
年度研究助成金により行われた。
参考文献
Carpenter, S. R. and D. M. Lodge (1986): Effects of submerged
macrophytes on ecosystem processes. Aquatic Botany, 16:
341 - 370.
Chambers, P.A. and J. Kalff (1985): Depth distribution and
biomass of submersed aquatic macrophyte communities in
relation to Secchi depth. Canadian Journal of Fisheries and
Aquatic Sciences, 42:701-709.
芳賀裕樹 (2006):琵琶湖南湖の面積について .陸水学雑
誌 , 67: 123-126.
浜端悦治(1991 a):琵琶湖の沈水植物群落に関する研
究(1)潜水調査による種組成と分布 . 日本生態学会誌 ,
41: 125-139.

78
芳賀 裕樹ら 2002年夏の琵琶湖南湖における沈水植物の現存量と種組成の場所による違い
79
浜端悦治(1991 b):琵琶湖の沈水植物群落に関する研
究(2)魚群探知機と船上からの採集による分布調査 ,
滋賀県自然誌 , 滋賀県自然誌編集委員会(編著):
1295-1310. 滋賀県自然保護財団 .
浜端悦治 (1996): 沈水植物の特性 , 河川環境と水辺植物
-植生の保全と管理 -, 奥田重俊佐々木寧 (編): 71-92.
ソフトサイエンス社,東京.
Hamabata, E. and Y. Kobayashi (2002): Present status of
submerged macrophyte growth in Lake Biwa: Recent
recovery following a summer decline in the water level.
Lakes & Reservoirs: Research and Management, 7:
331-338.
生嶋功・古川優・池田准蔵 (1962):琵琶湖の水生高等
植物の現存量.千葉大学文理学部紀要 , 3:483-494.
生嶋功 (1966):水草班中間報告 琵琶湖の水生高等植
物.びわ湖生物資源調査団中間報告 (一般調査の
部 ),313-341.
生嶋功(1972):水界植物群落の物質生産 I-水生植物- ,
生態学講座 7.共立出版 , 東京 .
今本博臣・加藤正典・堀家建司・原稔明 (1998): 琵琶
湖の湖岸環境に関する研究 I. 沈水植物の種組成と分
布 . 応用生態工学 , 1: 7-20.
大塚泰介・桑原泰典・芳賀裕樹 (2004): 琵琶湖南湖にお
ける沈水植物群落の分布および現存量-魚群探知機を
用いた推定- . 陸水学雑誌 , 65: 13-20.
大村朋広・福士富之信・林日出喜・堀家健司 (2001):
琵琶湖の湖岸環境に関する研究Ⅱ.沈水植物の群落面
積と季節変化.生態学リサーチマネージメントをアシ
ストする勉強会 第1回 発表論文集 : 27-37.応用生態
工学研究会 ,東京 .
Pianka, E. R. (1973): The structure of lizard communities.
Annual review of ecology and systematics. 4: 53 - 74.
西條八束・三田村緒佐武 (1995):新編湖沼調査法 .講談
社 ,東京 .
Scheffer, M. and E. Jeppesen (1997): Alternative Stable
States. In Structuring role of macrophytes in lake
ecosystems, E. Jeppesen, M. Søndergaard, M. Søndergaard
and K. Christoffersen (Eds.) :397-406. Springer-Verlag,
New York.
滋賀県(2003):環境白書-資料編-.平成 15年(2003
年)版 , 滋賀県 .
滋賀県水産試験場 (1954):琵琶湖水位低下対策(水産
生物)調査報告書.昭和 28年度総合開発調査 , 滋賀県.
滋賀県水産試験場 (1972):昭和 44年度 琵琶湖沿岸帯
調査報告書 , 滋賀県.
滋賀県水産試験場 (1998):平成 7年度 琵琶湖沿岸帯
調査報告書 , 滋賀県.
谷水久利雄 ; 三浦泰蔵 (1976): びわ湖における沈水植物
群集に関する研究 1. 南湖における侵入種オオカナダ
モの分布と生産能 . 生理生態 , 17: 283-290.
Takamura, N., Y. Kadono, M. Fukushima, M. Nakagawa, and
B.O. Kim (2003): Effects of aquatic macrophytes on water
quality and phytoplankton communities in shallow lakes.
Ecological Research, 18: 381-395.
山口久直(1938):琵琶湖南部に於ける高等水生植物の
生態分布.生態学研究 , 4: 17- 26.
山口久直(1943):琵琶湖の水草(琵琶湖沿岸帯生物群
集の研究 1).陸水学雑誌 , 13: 92-104.
Copyright © 2022 FDOKUMEN




















