
Spatial distribution patterns of the hyporheic invertebrate communities in a polluted river in...
20
PRIMARY RESEARCH PAPER Spatial distribution patterns of the hyporheic invertebrate communities in a polluted river in Romania Oana Teodora Moldovan • Erika Levei • Constantin Marin • Manuela Banciu • Horia Leonard Banciu • Claudia Pavelescu • Traian Brad • Mirela Cı ˆmpean • Ioana Meleg • Sanda Iepure • Ioan Povara ˘ Received: 1 June 2010 / Revised: 12 February 2011 / Accepted: 26 February 2011 / Published online: 13 March 2011 Ó Springer Science+Business Media B.V. 2011 Abstract The purpose of this study was to examine the sensitivity, in a field situation, of the hyporheic fauna to pollution by heavy metals and also to test the use of oxidative stress enzymes produced by this fauna as a sensitive indicator of oxidative stress generated by chemical contamination. This was done by surveying the patterns of distribution, structure, and composition of hyporheic invertebrate communities in one of the most polluted rivers in Romania. Twelve permanent sampling stations with differing water qualities were established along a 180 km transect of the Aries ¸ River. Data on hyporheic invertebrate abundance and richness, chemistry of the surface and hyporheic water and interstitial suspended particles were analyzed via mul- tifactorial analyses. In the downstream, more polluted stations, epigean species were less abundant and hyporheic communities, especially macrocrustaceans and oligochetes, became dominant. The higher levels of hyporheic invertebrate biodiversity in the moderately polluted stations compared to highly polluted, and the increase of the number of some hyporheos (especially macrocrustaceans) in the moderately polluted stations, suggested that the hyporheic fauna was more tolerant of heavy metal pollution than the surface water fauna of the area. However, the different richness and abundance of hyporheic fauna in sites of similar water chemistry suggested that additional factors, such as sediment structure are shaping the spatial distribution of hypor- heic fauna. Strong correlations between superoxide dismutase (SOD) activity in pooled tissues extracts and some chemical parameters suggest that oxidative stress enzymes may prove to be sensitive indicators of chemical pollution in hyporheic zones. Keywords Interstitial fauna Crustacea Heavy metals Oxidative stress enzymes Aries ¸ River Introduction The surface water/groundwater interface of river alluvia, the interstitial or the hyporheic zone (Creuze ´ Handling editor: Stuart Anthony Halse O. T. Moldovan (&) T. Brad I. Meleg S. Iepure ‘‘Emil Racovitza’’ Institute of Speleology, Romanian Academy, Clinicilor 5, 400006 Cluj-Napoca, Romania e-mail: [email protected] M. Banciu H. L. Banciu M. Cı ˆmpean Faculty of Biology and Geology, ‘‘Babes ¸-Bolyai’’ University, Cluj-Napoca, Romania C. Pavelescu Romanian Waters National Administration, Somes ¸ Tisa Division, Cluj-Napoca, Romania E. Levei National Institute of Research and Development for Optoelectronics, Research Institute for Analytical Instrumentation, Bucharest, Romania C. Marin I. Povara ˘ ‘‘Emil Racovitza’’ Institute of Speleology, Romanian Academy, Bucures ¸ti, Romania 123 Hydrobiologia (2011) 669:63–82 DOI 10.1007/s10750-011-0651-2
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Spatial distribution patterns of the hyporheic invertebrate communities in a polluted river in...

PRIMARY RESEARCH PAPER
Spatial distribution patterns of the hyporheic invertebratecommunities in a polluted river in Romania
Oana Teodora Moldovan • Erika Levei • Constantin Marin • Manuela Banciu •
Horia Leonard Banciu • Claudia Pavelescu • Traian Brad • Mirela Cımpean •
Ioana Meleg • Sanda Iepure • Ioan Povara
Received: 1 June 2010 / Revised: 12 February 2011 / Accepted: 26 February 2011 / Published online: 13 March 2011
� Springer Science+Business Media B.V. 2011
Abstract The purpose of this study was to examine
the sensitivity, in a field situation, of the hyporheic fauna
to pollution by heavy metals and also to test the use of
oxidative stress enzymes produced by this fauna as a
sensitive indicator of oxidative stress generated by
chemical contamination. This was done by surveying
the patterns of distribution, structure, and composition
of hyporheic invertebrate communities in one of the
most polluted rivers in Romania. Twelve permanent
sampling stations with differing water qualities were
established along a 180 km transect of the Aries River.
Data on hyporheic invertebrate abundance and richness,
chemistry of the surface and hyporheic water and
interstitial suspended particles were analyzed via mul-
tifactorial analyses. In the downstream, more polluted
stations, epigean species were less abundant and
hyporheic communities, especially macrocrustaceans
and oligochetes, became dominant. The higher levels of
hyporheic invertebrate biodiversity in the moderately
polluted stations compared to highly polluted, and the
increase of the number of some hyporheos (especially
macrocrustaceans) in the moderately polluted stations,
suggested that the hyporheic fauna was more tolerant of
heavy metal pollution than the surface water fauna of the
area. However, the different richness and abundance of
hyporheic fauna in sites of similar water chemistry
suggested that additional factors, such as sediment
structure are shaping the spatial distribution of hypor-
heic fauna. Strong correlations between superoxide
dismutase (SOD) activity in pooled tissues extracts and
some chemical parameters suggest that oxidative stress
enzymes may prove to be sensitive indicators of
chemical pollution in hyporheic zones.
Keywords Interstitial fauna � Crustacea � Heavy
metals � Oxidative stress enzymes � Aries River
Introduction
The surface water/groundwater interface of river
alluvia, the interstitial or the hyporheic zone (Creuze
Handling editor: Stuart Anthony Halse
O. T. Moldovan (&) � T. Brad � I. Meleg � S. Iepure
‘‘Emil Racovitza’’ Institute of Speleology,
Romanian Academy, Clinicilor 5, 400006 Cluj-Napoca,
Romania
e-mail: [email protected]
M. Banciu � H. L. Banciu � M. Cımpean
Faculty of Biology and Geology, ‘‘Babes-Bolyai’’
University, Cluj-Napoca, Romania
C. Pavelescu
Romanian Waters National Administration, Somes Tisa
Division, Cluj-Napoca, Romania
E. Levei
National Institute of Research and Development for
Optoelectronics, Research Institute for Analytical
Instrumentation, Bucharest, Romania
C. Marin � I. Povara
‘‘Emil Racovitza’’ Institute of Speleology, Romanian
Academy, Bucuresti, Romania
123
Hydrobiologia (2011) 669:63–82
DOI 10.1007/s10750-011-0651-2

des Chatelliers et al., 1994), is a spatially fluctuating
ecotone zone of high diversity and variability.
Horizontal and vertical gradients of oxygen, temper-
ature, light, and other environmental parameters
define this ecotone, which is complex to study and
the physical boundaries of which are difficult to
define (Boulton et al., 1998). The hyporheic zone acts
both as source and sink for nutrients, potentially
regulating biotic productivity (Moser et al., 2003),
and has an impact on the fate and transport of mining-
derived pollutants, influencing both surface water and
groundwater (Bourg & Bertin, 1993). The hyporheic
zone can be viewed as an ‘‘archive’’ of past episodes
of pollutants passing through its sediments, and it can
also function as a barrier preventing or retarding the
movement of pollutants between surface and ground-
water (Fuller & Harvey, 2000). Accordingly, the
study of the hyporheic zone benefits from a multi-
disciplinary approach that takes into account hydro-
logical, hydrogeological, chemical, microbiological,
and faunal variables.
It has been suggested that groundwater fauna are
sensitive indicators of aquifer contamination, including
contamination with heavy metals (e.g., Malard et al.,
1994, 1996; Notenboom et al., 1994; Boulton, 2000a).
Although the impact of contaminants on groundwater
invertebrates has been extensively investigated (review
in Notenboom et al., 1994; Mosslacher & Notenboom,
1999), the results are somewhat contradictory. Hypor-
heic animals seem to be less sensitive than their epigean
relatives, at least for groups such as typical hyporheic
isopods and amphipods (Plenet & Gibert, 1994; Plenet,
1995; Plenet et al., 1996; Plenet, 1999; Williams &
Fulthorpe, 2003). However, more recently, El Adnani
et al. (2007) found that hyporheic fauna may show both
qualitative and quantitative changes downstream from
metal sulfide tailings containing Pb, Zn and Cu (see also
Pyle & Mirza, 2007).
Perhaps a more useful indicators of groundwater
contamination are the oxidative stress enzymes
generated by aquatic invertebrates in response to
chemical contamination and/or abnormal physical
parameters of their surroundings (Cantu-Medellın
et al., 2009). High intracellular concentration of
reactive oxygen species (ROS), such as glutathione
S-transferase, glutathione peroxidase, glutathione
reductase, catalase, and superoxide dismutase
(SOD), may provide an estimate of the impact of
heavy metal pollution on the metabolic status of
aquatic macroinvertebrates such as bivalves (Geret
et al., 2002; Manduzio et al., 2004; Cravo et al.,
2009) or microinvertebrates such as cladocerans and
copepods (Fan et al., 2009; Wong et al., 2010).
In this study, we examined the sensitivity of
hyporheic Crustacea to pollution and compared the
utility of using hyporheic invertebrate composition
and oxidative stress enzymes as indicators of pollu-
tion. Our study site was one of the most polluted rives
in Romania, as a result of mining-related water
contamination. We assessed the impact of pollution
on the spatial distribution patterns of hyporheic
invertebrates. Pollution impact was investigated
using invertebrate abundance and species richness,
with emphasis on hyporheic Crustacea, by empha-
sizing the changes in biological hyporheic commu-
nities in each station in correlation to chemical
composition of both interstitial water and sediments.
The most polluted sites, presumably located in the
middle part of the studied transect, were expected to
have less species and fewer individuals than the less
polluted locations, although the extent of decline
should be dependent on the sensitivity of hyporheic
invertebrates to pollution, with some studies predict-
ing minimal impact (see above). Two hypotheses
were tested along the studied longitudinal transect.
The ecological and evolutionary implications of
our findings in the context of the exchanges between
surface and hyporheic communities are discussed,
emphasizing the importance of habitat heterogeneity
and biological interactions.
Materials and methods
Study area
The study area was in Transylvania, Romania, in the
so-called ‘Golden Quadrilateral’, where important
gold resources of Europe are concentrated. The
mining activities in the region have led to extensive
and high levels of pollution of the Aries River, one of
the main rivers in north-western Romania (Fig. 1).
While most of the pollution of the Aries River is from
leakages and unchecked discharges of the non-treated
acid waste-water, other pollution sources include
waste dumps and settlings from mines, sewage waters
from towns along the river and pesticides and
fertilisers from agricultural activities (Forray, 2001).
64 Hydrobiologia (2011) 669:63–82
123

Survey planning
Twelve permanent stations (Table 1) were estab-
lished along a 180 km longitudinal transect of the
eastward flowing river (Fig. 1). Three of the stations
(1–3) were selected upstream of Baia de Aries, the
locality known as the most polluted with heavy
metals along the Aries River. The locations of the
stations were chosen according to accessibility to
the river, state of conservation of alluvia (many of the
sandy beaches are exploited by industry) and by the
presence of fauna, as established in preliminary
samplings. The sampling stations were located in
the unconsolidated detritic deposits of the floodplain,
and interstitial waters of the flood plain are prone to
pollution from surface waters during flooding peri-
ods. The lithology and granulometry of alluvia
deposits is non-homogenous, being a mixture of fine
sands, oozy sands, gravels and boulders, without clear
vertical distribution and highly dynamic through
time.
Standpipes of galvanized iron 20 mm in diameter,
with perforations at 1.3–1.4 m depth, were perma-
nently fixed in the alluvia at distances of 1–5 m from
the river. A manual diaphragm pump was installed
and water was pumped according to the technique
described by Bou (1974). Through our method of
pumping, 0.3 m3 of water was extracted from sedi-
ments. In each sample, very fine sand, rich organic
matter and some sawdust were collected. From each
sampling station two replicate samples for fauna and
interstitial and surface water were taken monthly
from February 2008 to March 2009 (a total of 168
samples). Three samples (March, July and October
2008) were taken from each sampling station for the
chemical analysis of suspended particles.
Analysis of fauna and of physico-chemical
parameters
From each sample the first 20 l of water was filtered
through a planktonic net of 160 lm and fixed in 96�ethanol or 4% formaldehyde. Collected invertebrates
were sorted into major taxonomical groups with the
adults subsequently sorted to species level for most
taxa. The identification of individuals was performed
according to the specific methods for each group.
Invertebrates were also assigned to one of the
Fig. 1 Aries River basin with its localization in north-western Romania. Sampling stations and input of pollutants along the studied
sector are marked on the map
Hydrobiologia (2011) 669:63–82 65
123

following ecological classifications: (1) typical hy-
porheos restricted to groundwater and rarely occur-
ring near the surface; (2) epigeans that are usually
restricted to benthos; and (3) transitionals that can be
found in the above mentioned habitats, but rarely
occurring in deep groundwater.
Temperature (�C), pH, and electrical conductivity
(EC) (lS/cm) of surface and interstitial waters were
measured on site using Multiparameter Combo
Hanna Instruments HI 98129. Oxygen concentration
in water was not measured because of the errors
introduced by the pumping method.
For chemical analysis, surface and interstitial
waters were collected in 1-l acid-washed polyethyl-
ene bottles. During the transportation to the labora-
tory, the samples were kept at 4�C.
The concentrations of Al, Cr, Ni, Cu, Zn, As, Sr,
Cd, Pb, Na, Mg, K, Ca, Mn, Fe, Ti, Cl-, NO2-,
NO3-, SO4
2- were analyzed and the chemical
oxygen demand (COD) was measured. For total
heavy metal concentration, 100 ml unfiltered water
samples were digested on a sand bath with 5 ml
Ultrapur� 60% Nitric acid. After cooling, the solution
was filtered through 0.45-lm pore-diameter filters
and made up to 100 ml with ultrapure water.
Depending on their concentration, metals were
quantified using Inductively Coupled Plasma Optical
Emission Spectrometry (ICP-OES OPTIMA
5300DV, Perkin-Elmer, USA) or Inductively Cou-
pled Plasma Mass Spectrometry (ICP-MS ELAN
DRC II, Perkin-Elmer, USA).
Anions measurement was taken from samples
filtered on 0.45 lm pore-diameter cellulose nitrate
membrane filters by ion chromatography (761Com-
pact IC System, Metrohm, Switzerland).
A COD test was used as an indirect measure of the
amount of organic compounds in sampled water. The
method is based on the oxidation in strong acid
Table 1 General
information about the
sampling stations along the
Aries River
No. Station Distance from
headwaters
(km)
River
side
Distance
from
water (m)
UTM
coordinates
Relief
1 Scarisoara 1 21.5 Left 7.80 34 T 0643874 Base of the inferior
terrace5146460
2 Scarisoara 2 26.45 Left 2.50 34 T 0646710 High river meadow
5145965
3 Vadu Motilor 44.27 Left 10.50 34 T 0652692 Low river meadow
5139174
4 Baia de Aries 81.70 Left 3.50 34 T 0675486 High river meadow
5139031
5 Sartas 84.56 Left 1.20 34 T 0676858 Base of the inferior
terrace5140089
6 Brazesti 1 87.04 Left 2.50 34 T 0677960 Minor river bed
5140621
7 Brazesti 2 89.56 Right 4.50 34 T 0679404 Minor river bed
5141915
8 Salciua de Jos 101.42 Left 6.15 34 T 0687603 Minor river bed
5142005
9 Lunca Aries 109.54 Left 12.30 34 T 0688565 High river meadow
5145950
10 Vidolm 117.29 Right 13.15 34 T 0692634 Low river meadow
5150484
11 Buru 1 129.00 Left 5.00 34 T 0700074 Low river meadow
5153340
12 Buru 2 129.92 Left 5.55 34 T 0700886 Terrace
5153629
66 Hydrobiologia (2011) 669:63–82
123

Ta
ble
2C
hem
istr
yo
fin
ters
titi
alw
ater
s(I
),su
rfac
ew
ater
s(S
),an
dS
PM
(P)
inth
e1
2m
on
ito
red
stat
ion
s
Sta
tio
ns
I Al
(mg
/l)
Cr
(mg
/l)
Ni
(mg
/l)
Cu
(mg
/l)
Zn
(mg
/l)
Sr
(mg
/l)
Pb
(mg
/l)
Na
(mg
/l)
Mg
(mg
/l)
K (mg
/l)
Ca
(mg
/l)
Mn
(mg
/l)
Fe
(mg
/l)
Ti
(mg
/l)
Cl-
(mg
/l)
NO
2-
(mg
/l)
NO
3-
(mg
/l)
SO
42-
(mg
/l)
CO
D
(mg
/l)
10
.1\
0.0
1\
0.0
10
.10
.40
.3\
0.0
19
3.7
11
.81
2.2
92
.72
.00
.4\
0.0
12
10
.30
.12
.71
3.9
9.8
21
.04
.53
.23
2.4
39
.16
6.4
69
.44
5.2
6.0
4.7
31
.21
.82
.30
.21
04
.11
.54
.07
.75
.0
31
.33
.11
.81
4.6
17
.92
6.6
29
.65
6.1
7.1
5.4
42
.61
.92
.60
.21
32
.01
.44
.49
.49
.3
41
.02
.11
.51
7.5
21
.13
7.7
38
.25
7.2
6.6
4.6
50
.21
.72
.00
.11
47
.60
.83
.71
2.9
12
.5
51
.41
.91
.41
6.0
19
.43
4.7
34
.95
2.7
7.3
4.6
62
.61
.82
.80
.21
33
.80
.73
.28
0.9
23
.2
62
.80
.80
.67
.39
.41
6.0
15
.93
4.1
9.2
4.9
88
.52
.96
.00
.27
6.4
0.3
6.5
23
7.3
51
.4
72
.40
.90
.77
.99
.81
7.5
17
.53
0.7
8.0
4.1
68
.73
.25
.00
.27
2.8
0.3
5.1
15
2.7
37
.6
83
.00
.60
.55
.87
.31
2.8
12
.82
4.9
9.4
4.2
76
.03
.76
.20
.25
6.5
0.3
6.4
18
1.1
61
.0
93
.10
.40
.33
.74
.78
.38
.22
0.4
10
.13
.86
7.3
4.0
6.5
0.2
42
.10
.27
.71
32
.05
5.7
10
3.0
0.3
0.3
3.2
3.9
7.2
7.0
16
.11
0.6
3.2
66
.33
.76
.30
.23
3.5
0.3
6.8
20
3.0
56
.7
11
2.9
0.3
0.2
2.3
2.8
5.2
5.1
14
.01
0.7
3.0
69
.82
.45
.90
.22
7.2
0.3
7.0
21
6.1
57
.3
12
2.2
0.2
0.2
1.8
2.1
3.9
3.8
10
.98
.72
.86
3.0
1.8
4.4
0.1
23
.40
.48
.82
32
.23
7.1
Sta
tio
ns
S Al
(mg
/l)
Cr
(mg
/l)
Ni
(mg
/l)
Cu
(mg
/l)
Zn
(mg
/l)
Sr
(mg
/l)
Pb
(mg
/l)
Na
(mg
/l)
Mg
(mg
/l)
K (mg
/l)
Ca (mg/l)
Mn
(mg
/l)
Fe
(mg
/l)
Ti
(mg
/l)
Cl
(mg
/l)
NO
2
(mg
/l)
NO
3
(mg
/l)
SO
4
(mg
/l)
CO
D
10
.1\
0.0
1\
0.0
1\
0.0
10
.10
.1\
0.0
18
.23
.31
.83
2.2
0.3
0.3
0.1
6.5
0.2
2.6
6.0
8.6
20
.1\
0.0
1\
0.0
1\
0.0
10
.10
.0\
0.0
13
.62
.70
.72
7.0
\0
.01
0.2
\0
.01
5.6
0.4
2.3
5.7
4.0
30
.1\
0.0
1\
0.0
1\
0.0
10
.10
.0\
0.0
14
.94
.70
.92
8.8
\0
.01
0.3
\0
.01
5.1
0.2
1.9
6.1
4.3
41
.5\
0.0
1\
0.0
10
.20
.20
.1\
0.0
15
.06
.11
.93
8.8
0.4
0.9
0.1
5.0
0.2
2.0
47
.12
.9
51
.4\
0.0
1\
0.0
10
.10
.20
.1\
0.0
14
.96
.92
.14
3.3
1.0
0.7
0.1
5.9
0.2
2.2
45
.94
.2
61
.8\
0.0
1\
0.0
10
.20
.20
.1\
0.0
17
.96
.22
.24
4.1
0.8
1.2
8.0
5.3
0.6
2.1
43
.73
.2
70
.9\
0.0
1\
0.0
10
.50
.50
.1\
0.0
15
.56
.72
.06
2.0
0.3
0.9
0.1
4.9
0.2
2.4
41
.74
.1
81
.3\
0.0
1\
0.0
10
.10
.20
.1\
0.0
14
.75
.41
.93
8.5
0.4
0.9
0.1
6.1
0.4
2.5
46
.32
.3
90
.9\
0.0
1\
0.0
10
.10
.20
.1\
0.0
14
.55
.71
.84
2.1
0.4
0.7
0.1
6.0
0.4
2.4
63
.25
.1
10
0.9
\0
.01
\0
.01
0.1
0.2
0.1
\0
.01
4.8
6.0
2.0
45
.10
.41
.00
.15
.50
.32
.14
3.1
3.4
11
0.5
\0
.01
\0
.01
0.1
0.1
0.1
\0
.01
5.1
6.2
2.1
41
.40
.20
.70
.17
.60
.74
.34
3.5
6.1
12
0.4
\0
.01
\0
.01
0.1
0.2
0.1
\0
.01
4.7
6.0
1.9
42
.00
.30
.50
.15
.20
.42
.54
4.6
3.8
Hydrobiologia (2011) 669:63–82 67
123

solution with a known excess of potassium perman-
ganate (KMnO4), titration of the remaining unre-
duced KMnO4 with sodium oxalate solution and
calculation of the consumed KMnO4.
A direct method of measuring the concentrations
of Ba, Mn, Cr, Ni, Cu, Zn, Cd, Pb, Sb, and As
adsorbed on the surface of particles in suspension in
interstitial water was used. Separation of suspended
particulate matter (SPM) was realized in situ. When
collected, samples were filtered by means of a
Chromatography Research Supplies filtering system,
provided with a manual Nalgene vacuum pump. Pre-
weighed (Kern 770-60) MF-Millipore Membrane of
mixed cellulose esters of 0.45 lm porosity and
47 mm diameter were used for filtering. The mem-
branes were dried at room temperature and then
weighed to the nearest 0.01 mg to determine the mass
of SPM in the known volume of filtered water. The
filters were then digested at room temperature for
48 h with 5.0 ml of 1:1 solution Ultrapur� 60%
Nitric acid and Ultrapur� 30% Hydrochloric acid. A
2 ml aliquot of the digest solution (not including any
visible solid) was diluted to 5 ml with ultra-pure
water and analyzed by atomic absorption spectros-
copy (AAS). This process was repeated for duplicate
unused filters and the average elemental content in
the filter blanks was subtracted from the sample
results. The Mn, Cu, and Zn concentrations for the
digested SPM were determined by means of standard
flame-atomic absorption spectrometry methods and
Ba, Cr, Ni, Cd, Pb, Sb and As concentrations by
standard electrothermal-AAS methods. The determi-
nations were carried out with a Perkin-Elmer atomic
absorption spectrometer model AAnalyst 700. The
calibration lines were traced using solutions prepared
from standard solutions CertiPUR�. The methods
accuracy, precision and sensitivity have been tested
by using the Perkin-Elmer groundwater and waste-
water pollution control certified reference materials.
Determination of superoxide dismutase (SOD)
activity
Biological samples for the assay of SOD activity
were collected at four different sampling stations (2,
3, 8, and 12) in May and November 2008 (Table 3).
Biological samples were prepared for SOD analysis
according to the method described by de Oliveira
et al. (2005). Organisms (all individuals found at thatTa
ble
2co
nti
nu
ed
Sta
tio
ns
P Ba
(mm
ol/
kg
)
Mn
(mm
ol/
kg
)
Cr
(mm
ol/
kg
)
Ni
(mm
ol/
kg
)
Cu
(mm
ol/
kg
)
Zn
(mm
ol/
kg
)
Pb
(mm
ol/
kg
)
As
(mm
ol/
kg
)
Sb
(mm
ol/
kg
)
13
.83
.40
.20
.21
.70
.60
.20
.30
.1
22
.11
2.1
0.7
0.3
1.0
1.9
0.2
0.1
0.1
39
.22
2.7
1.7
0.5
0.0
78
.31
.90
.00
.0
41
.61
.51
.00
.50
.71
.51
.50
.30
.2
50
.91
.50
.70
.80
.80
.50
.60
.20
.1
61
.11
3.5
1.0
0.6
2.9
2.8
0.2
0.8
0.1
71
.21
.60
.80
.70
.70
.40
.20
.10
.1
81
.41
.71
.20
.81
.60
.50
.30
.20
.7
90
.71
2.2
0.4
0.5
0.6
4.9
0.2
1.5
0.0
10
1.7
1.4
0.4
0.4
0.8
0.4
0.2
0.1
0.1
11
1.1
2.5
0.5
0.4
0.9
0.5
0.8
0.2
0.1
12
0.8
20
.40
.60
.52
.31
2.9
0.3
0.2
0.0
68 Hydrobiologia (2011) 669:63–82
123

Ta
ble
3N
um
ber
of
ind
ivid
ual
so
fea
chfa
un
ag
rou
p(a
)an
do
fth
eh
yp
orh
eic
Cru
stac
eaan
dO
lig
och
aeta
(b)
inth
e1
2st
atio
ns
on
the
Ari
esR
iver
(su
b-a
du
lts
no
tin
clu
ded
inb
)
Sta
tion
Gro
up
Rot-
ifer
a
Tar
di-
gra
da
Nem
-
atoda
Oli
go-
chae
ta
Aca
riG
as-
tropoda
Cycl
op-
oid
a
Har
pac
-
tico
ida
Ost
ra-
coda
Am
ph-
ipoda
Isopoda
Bat
hy-
nel
lace
a
Inse
ct
larv
ae
R Indiv
idual
s
R Gro
ups
a 124
019
66
10
062
65
76
01
030
353
9
20
08
24
19
141
943
26
414
012
597
10
30
123
26
34
28
29
31
25
230
76
242
12
40
03
17
00
20
10
01
226
6
50
083
96
00
67
00
043
10
245
6
60
15
90
0294
24
00
00
315
6
70
04
76
22
0561
450
52
610
36
1388
10
80
02
81
10
31
082
03
15
3218
8
90
05
21
30
259
20
14
15
46
16
5404
10
10
00
117
20
174
216
18
33
82
273
10
11
00
06
60
92
22
45
55
127
9
12
00
328
80
95
247
25
52
12
7279
10
RIn
div
idual
s24
2156
467
105
29
1646
144
360
142
1194
110
88
4467
Sta
tion
Spec
ies
Nip
harg
us
transy
l-va
nic
us
Nip
harg
us
rom
anic
us
Bogid
-ie
lla
sp.
Bath
y-nel
lasp
.
Mic
ro-
charo
nsp
.
Pro
ase
llus
sp.
Aca
nth
o-
cycl
ops
sp.
Aca
nth
o-
cycl
ops
b.
bis
-aet
osu
s
Dia
cy-
clops
clandes
-ti
nus
Ela
phoid
-el
lael
aphoid
es
Para
ste-
noca
ris
sp.
1
Para
ste-
noca
ris
sp.
2
Tri
cho-
dri
lus
sp.
R Indi-
vid
ual
s
R Spec
ies
b 10
00
01
00
00
21
00
527
3
228
00
0412
10
01
11
15
450
8
30
50
19
00
20
00
00
17
4
40
00
10
00
02
00
08
11
3
50
00
43
00
00
10
05
18
67
4
60
00
00
00
00
00
01
11
716
37
03
601
90
00
00
022
688
6
80
00
13
30
00
00
00
50
66
3
92
13
016
42
40
10
10
07
12
107
9
10
018
08
33
021
42
20
02
14
140
8
11
03
05
50
00
00
00
922
4
12
619
112
50
00
00
00
01
89
6
RIn
div
idual
s52
95
1102
1156
14
21
54
722
115
145
1685
Hydrobiologia (2011) 669:63–82 69
123

time in the station) from an entire sample were
lysed in 0.5 M sucrose, buffered at pH 7.4 with
20 mM Tris–HCl (7 ml/g of tissue), at 4�C. Phen-
ylmethanesulfonyl fluoride (PMSF), as protease
inhibitor, was prepared as 100 mM stock solution in
isopropanol. PMSF was added to the lysis buffer
(10 ll/ml of buffer). Samples were homogenized in
lysis buffer using a Potter-Teflon homogenizer. The
homogenates were incubated for 30 min on ice and
centrifuged at 10,000g, 4�C, for 10 min. The
supernatants were collected and the total protein
content of the lysates was determined colorimetri-
cally using Coomassie Brilliant Blue G-250 (Sigma)
as a dye (Bradford, 1976) and bovine serum albumin
as a standard.
The SOD activity in lysates was determined
indirectly by using a SOD Assay Kit (Biochemika,
Sigma-Aldrich) to measure inhibition of the reduction
of highly water-soluble tetrazolium salt, WST-1
(2-(4-Iodophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophe-
nyl)-2H-tetrazolium, monosodium salt) by superoxide
anion (O2-) produced by xanthine oxidase activity.
Upon reduction of WST-1 with the superoxide anion,
a water-soluble formazan dye was developed that
could be measured colorimetrically at 450 nm. The
rate of the reduction with O2- is linearly related to
the inhibition of xanthine oxidase activity by SOD
activity. As the absorbance at 450 nm is proportional
to the amount of superoxide anion, the SOD activity
as an inhibition activity can be quantified by
measuring the decrease in the color development at
450 nm with a reference wavelength of 630 nm
(Ukeda et al., 2002). The assay was performed in
96-well plates by using a Stat Fax 2100 Microplate
Reader (Awareness Technology, Palm City, FL). For
each sample, SOD activity was determined for the
same amount of protein (10 lg). Blanks and samples
were prepared according to the manufacturer’s
instructions. The activity of SOD was expressed as
a rate of inhibition of xanthine oxidase activity. SOD
activity for each experiment was determined in
triplicate.
Statistical analysis
The log-transformated data used for analyses corre-
spond to four categories: physico-chemical charac-
teristics of interstitial waters, metal concentrations,
and COD of interstitial waters, metal concentrations
in sediments and abundance of hyporheic Crustacea
species. Statistical analyses, such as multiple factor
analysis (MFA), principal component analysis (PCA)
and Mantel tests were performed using XLSTAT
Version 2010. The MFA is a synthesis of principal
component analysis and multiple correspondence
analysis and allows the study of the relationships
between the observations, the variables, and tables
(Pages, 2004). Mantel tests were used to compare the
chemistry of surface and interstitial waters in the 12
stations, by using Spearman correlation and 10,000
permutations.
Superoxide dismutase activity for each experimen-
tal group was determined in triplicate. The final
results represent mean ± SD of three measurements.
For statistical analysis, a value of P \ 0.05 was
considered significant. The differences between the
SOD activities in different samples at the same
season were analyzed by one-way ANOVA with
Bonferroni correction for multiple comparisons using
GraphPad Prism v.4.02 for Windows, GraphPad
Software (San Diego, CA).
Results
Water and SPM chemical features
Temperature and pH of interstitial and surface water
were similar at each sampling station. Water pH was
generally lower (0.5 pH units) at stations situated
downstream of the Baia de Aries mining area
(Table 2).
Heavy metal concentrations of interstitial waters,
especially Cu, Pb, Zn, and Fe, varied among different
stations (Table 2), with high levels recorded at
Stations 2, 4, 5, and 6. Heavy metal concentrations
in interstitial waters were much higher in these
stations than in the corresponding surface water
samples. At the other stations, heavy metal concen-
trations were only slightly higher in interstitial waters
and not significantly different from levels in surface
waters.
A PCA of the composition of heavy metals (Cr,
Ni, Cu, Zn, Pb, and Mn) in surface and interstitial
waters and SPM (Fig. 2) showed a clear separation of
SPM samples along the first axis (72% of the
variation). Interstitial and surface samples are sepa-
rated along the second axis (10%) with very low
70 Hydrobiologia (2011) 669:63–82
123
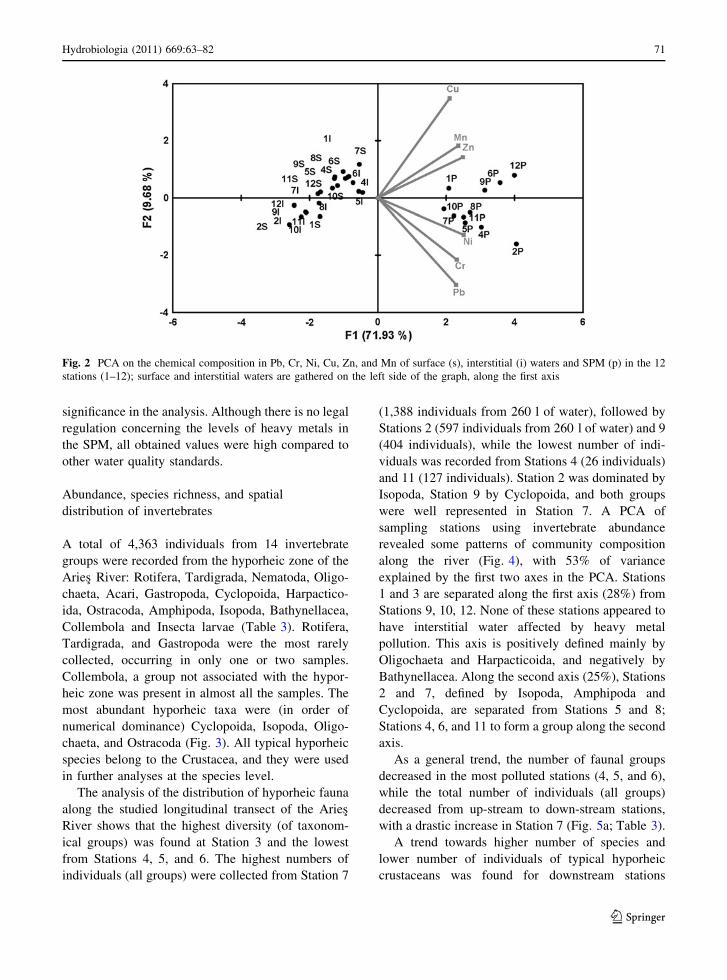
significance in the analysis. Although there is no legal
regulation concerning the levels of heavy metals in
the SPM, all obtained values were high compared to
other water quality standards.
Abundance, species richness, and spatial
distribution of invertebrates
A total of 4,363 individuals from 14 invertebrate
groups were recorded from the hyporheic zone of the
Aries River: Rotifera, Tardigrada, Nematoda, Oligo-
chaeta, Acari, Gastropoda, Cyclopoida, Harpactico-
ida, Ostracoda, Amphipoda, Isopoda, Bathynellacea,
Collembola and Insecta larvae (Table 3). Rotifera,
Tardigrada, and Gastropoda were the most rarely
collected, occurring in only one or two samples.
Collembola, a group not associated with the hypor-
heic zone was present in almost all the samples. The
most abundant hyporheic taxa were (in order of
numerical dominance) Cyclopoida, Isopoda, Oligo-
chaeta, and Ostracoda (Fig. 3). All typical hyporheic
species belong to the Crustacea, and they were used
in further analyses at the species level.
The analysis of the distribution of hyporheic fauna
along the studied longitudinal transect of the Aries
River shows that the highest diversity (of taxonom-
ical groups) was found at Station 3 and the lowest
from Stations 4, 5, and 6. The highest numbers of
individuals (all groups) were collected from Station 7
(1,388 individuals from 260 l of water), followed by
Stations 2 (597 individuals from 260 l of water) and 9
(404 individuals), while the lowest number of indi-
viduals was recorded from Stations 4 (26 individuals)
and 11 (127 individuals). Station 2 was dominated by
Isopoda, Station 9 by Cyclopoida, and both groups
were well represented in Station 7. A PCA of
sampling stations using invertebrate abundance
revealed some patterns of community composition
along the river (Fig. 4), with 53% of variance
explained by the first two axes in the PCA. Stations
1 and 3 are separated along the first axis (28%) from
Stations 9, 10, 12. None of these stations appeared to
have interstitial water affected by heavy metal
pollution. This axis is positively defined mainly by
Oligochaeta and Harpacticoida, and negatively by
Bathynellacea. Along the second axis (25%), Stations
2 and 7, defined by Isopoda, Amphipoda and
Cyclopoida, are separated from Stations 5 and 8;
Stations 4, 6, and 11 to form a group along the second
axis.
As a general trend, the number of faunal groups
decreased in the most polluted stations (4, 5, and 6),
while the total number of individuals (all groups)
decreased from up-stream to down-stream stations,
with a drastic increase in Station 7 (Fig. 5a; Table 3).
A trend towards higher number of species and
lower number of individuals of typical hyporheic
crustaceans was found for downstream stations
Fig. 2 PCA on the chemical composition in Pb, Cr, Ni, Cu, Zn, and Mn of surface (s), interstitial (i) waters and SPM (p) in the 12
stations (1–12); surface and interstitial waters are gathered on the left side of the graph, along the first axis
Hydrobiologia (2011) 669:63–82 71
123

(Fig. 5b; Table 4). The local distribution of Crustacea
species was variable. For example, highly polluted
Stations 4 and 5 supported a very low diversity of
Crustacea. Station 5 was dominated by a cosmopol-
itan epigean species, Paracyclops fimbriatus. Down-
stream stations were dominated by the transitional
Diacyclops languidoides, with more than 200 indi-
viduals in August in Station 8. Of the typical
hyporheic cyclopoids, two species of Acanthocyclops
were well represented in Station 10. Harpacticoida
was present only in Stations 1, 2, 9, and 10, where
hyporheic species were best represented (Paraste-
nocaris sp., Bryocamptus tauricus, Elaphoidella
elaphoides). All the macrocrustaceans collected are
typical hyporheic species. The dominant isopod was a
species of Microcharon, an interstitial genus, which
was abundant in Stations 2 and 7 (with more than 250
individuals counted in a single sample at Station 7,
December 2008) (Fig. 6), Less abundant were species
of Niphargus (Amphipoda), Bogidiella (Amphipoda),
Fig. 3 Ranking of the
taxonomical groups based
on their abundance
cumulated in all 12
sampling stations
Fig. 4 PCA of the
relationships between
stations (1–12) according to
common fauna groups
72 Hydrobiologia (2011) 669:63–82
123

Bathynella (Bathynellacea) and Proasellus (Isopoda).
Except for N. transylvanicus (relatively abundant
especially in Station 2), all other macrocrustaceans
were recorded in higher numbers downstream of
Station 4, particularly from Station 5 (Bathynella sp.)
and Station 7 (N. romanicus).
The other well-represented group was Oligochae-
ta, with the dominant species belonging to the
typically interstitial genus Trichodrilus. It was most
abundant downstream of Station 4, reaching maxi-
mum numbers in Stations 7 and 8 (Table 3b). The
other identified oligochete species were recorded in
low numbers.
Correlation analyses between invertebrate
communities and water/sediment quality
The general MFA combines results given by the
PCAs on physico-chemical features, on chemistry of
the water, on chemistry of the SPM and on typical
hyporheic species of Crustacea. In terms of interstitial
water, Station 1 was well-separated from the other
stations in the PCA (Fig. 7a) by high values of
conductivity. Stations 2, 3, 10, and 11 were positively
correlated with pH along the first axis (50%), while
Stations 4, 5, and 6 were negatively correlated with
pH. Station 7 was isolated along the second axis
Fig. 5 a Spatial
distributions of fauna
groups (squares) and
number of individuals
(circles) (a) and of
hyporheic Crustacea species
(squares) and number of
individuals (circles) (b);
polynomial trendlines and
equations are also
represented
Hydrobiologia (2011) 669:63–82 73
123

Table 4 List of fauna (major groups and species) found in the
hyporheic of the Aries River
Taxa E T H
Tardigrada
Oligochaeta
Amphichaeta sp.
Aulodrilus limnobius
Aulodrilus sp.
Cernosvitoviella sp. *
Cognettia glandulosa
Eiseniella tetraedra *
Fridericia bisetosa *
Haplotaxis gordioides *
Lumbriculus variegatus *
Nais communis *
Pristina aequiseta foreli *
Pristinella jenkinae *
Rhyacodrilus coccineus *
Stylodrilus heringianus *
Trichodrilus sp. *
Acari
Hydrachnidia
Athienemanniidae
Phreatohydracarusmosticus
*
Aturidae
Lethaxona sp. *
Hygrobatidae
Atractides cf latipalpis *
Atractides sp.
Lebertiidae
Lebertia insignis *
Momoniidae
Momonisia phreatica *
Stygomononia latipes *
Sperchontidae
Sperchon glandulosus *
Torrenticolidae
Torrenticola amplexa *
Halacarida
Gastropoda
Amphipoda
Bogidiella sp. *
Niphargus romanicus *
N. transsylvanicus *
Ostracoda
Bathynellacea
Table 4 continued
Taxa E T H
Bathynella sp. *
Cyclopoida
Acanthocyclops sp. *
A. balcanicus bisaetosus *
A. venustus *
A. vernalis *
Diacyclops sp.
D. bisetosus *
D. languidus *
D. clandestinus ssp. *
D. languidoides hypnicola *
Eucyclops s. serrulatus *
Megacyclops viridis *
Paracyclops fimbriatus *
Harpacticoida
Atheyella crassa *
Bryocamptus cuspidatus *
B. dacicus *
B. tauricus *
B. weberi cf. tauricus *
Elaphoidella elaphoides *
Moraria aff. alpina
Paracamptus schmeili
Parastenocaris sp. 1 *
Parastenocaris sp. 2 *
Isopoda
Microcharon sp. *
Proasellus sp. *
Collembolla *
Insecta (larvae) *
Coleoptera
Scirtidae
Diptera
Athericidae
Ceratopogonidae
Chironomidae
Limoniidae
Tabanidae
Ephemeroptera
Baetidae
Heptageniidae
Plecoptera
Leuctridae
Trichoptera
74 Hydrobiologia (2011) 669:63–82
123

(35%) and characterized by temperature. The other
stations were defined by two or all three of the
physico-chemical features. In terms of interstitial
water chemistry (Fig. 7b), Stations 4, 5, 6, and 8 were
separated from all other stations except 1 along the
first axis (34.5%). Station 1 was separated along the
second axis (21.5%) by high levels of Cl, Na, and K.
In terms of hyporheic crustacean composition
(Fig. 7c), Stations 2 and 7 defined by amphipods,
isopods, and harpacticoids were separated along the
first axis (27%) from Stations 4 and 5. Stations 9 and
10 form another group defined on the second axis
(23%) by cyclopoids, while all other stations were
grouped by the presence of Bogidiella sp. and
Elaphoidella elaphoides. In the PCA of SPM chem-
istry (Fig. 7d), only Stations 3, 4, and 8 had a distinct
position. Stations 3 and 4 are grouped along the first
Fig. 6 a Spatial
distribution of hyporheic
crustaceans identified along
the longitudinal transect of
the Aries River; b without
the dominant isopod
Microcharon sp.
Table 4 continued
Taxa E T H
Brachycentridae
Glossosomatidae
E epigean (surface), T transitional, H hyporheic
Hydrobiologia (2011) 669:63–82 75
123

axis (42%) because of high content of Cr, Pb, Zn, and
Ba; Station 8 belongs to the second axis (22%) by the
content of Sb. In the overall MFA, along the first axis
(23%), Stations 4, 5, 6, and 8 were separated from
Stations 2, 3, and 12. For all these stations, the
chemical features of water and SPM had strong
impact on their position on the plot. Station 1 was
separated from Stations 7, 9, 10, and 11 along the
second axis (Fig. 7e), with lower significance in the
analysis (17%). Station 1 was defined by fauna and
physico-chemical features, while Stations 7, 9, and 10
were defined by fauna composition.
Indirect measurement of SOD activity
in biological samples
Statistical analysis of SOD activity in biological
samples from different sampling sites collected
during May 2008, showed that only the sample from
Station 12 differed statistically. SOD activity was two
to three times higher in samples from Stations 2, 3,
and 8 than Station 12 (P \ 0.03) (Table 5). SOD
activity in samples collected during May was most
strongly correlated with COD (Spearman correlation
coefficient [ 0.9) (Fig. 8a). The normal value of
COD is lower than 3 mg O2/dm3, according to SR
EN ISO 8467/2001.
From the biological samples collected during
November 2008, the activity of SOD in the organisms
isolated from Station 2, was significantly higher than
the SOD activity in the other samples (P \ 0.001).
SOD activity was also significantly different
(P \ 0.001) from one season to another. During late
spring (May), the SOD activity was much higher than
those recorded in organisms sampled during the fall
season (November). For the latter, the strongest
positive correlation was with lead concentration
(Spearman correlation coefficient [ 0.9) (Fig. 8b).
Lead concentration only exceeded the acceptable
level (10 lg/dm3, according to SR ISO 17294-2/
2003) at Station 2.
Discussion
Water quality
Significant differences between the heavy metal
content of surface and interstitial waters were evident
at only a few stations (2, 4, 5, and 6) along the studied
longitudinal transect of the Aries River. At the other
stations, interstitial water was only slightly higher in
heavy metals than the surface water, but richer in
nitrites and nitrates due to the biogeochemical
processes that occur there (Brunke & Gonser, 1997;
Hinkle et al., 2001). Pollution of surface waters
probably occurs in pulses, and past episodes of
pollution can be detected by analyzing the SPM
chemistry. Much higher concentrations of heavy
metals are being carried adsorbed on the SPM active
surface, as compared to concentrations of the corre-
sponding dissolved ions which are carried in the
aqueous solution (Morel, 1983; Munk et al., 2002).
The SPM values differed from one sampling site to
another, suggesting distinct recharge mechanisms for
each specific sampling site.
Spatial patterns of fauna distribution and impact
of heavy metals
This study of the interstitial biodiversity and impact
of heavy metal pollution along the longitudinal
transect revealed spatial patterns of fauna distribution
illustrating the complexity and patchiness of the
hyporheic zone, even at depths over one meter. The
Aries River hyporheic fauna was diverse, with 14
invertebrate groups identified, compared to other
rivers of Transylvania, such as the Somes and the
Table 5 SOD activities in biological samples collected during May and November 2008
Month 2008 SOD activity (% inhibition of xanthin oxidase activity)
Station 2 Station 3 Station 8 Station 12
May 43.792 ± 5.673 33.026 ± 2.151 45.995 ± 7.711 13.431 ± 2.390
November 24.911 ± 2.802 5.272 ± 1.054 3.322 ± 2.916 6.372 ± 4.963
The results represent the mean of three independent measurements ± standard deviation (SD)
76 Hydrobiologia (2011) 669:63–82
123

Cris (10 and 14 groups, respectively) (Moldovan
et al., 2005). Most of the sampled individuals were
Crustacea, especially the co-dominant Cyclopo-
ida and Isopoda, belonging to both the micro- and
macro-interstitial faunas. Harpacticoida was abun-
dant only in clean waters, where hyporheic and
interstitial species were best represented. The mac-
rocrustaceans are all groundwater species, some of
Fig. 7 PCAs on observations (stations 1–12) and the physical
and chemical features (a), on interstitial water chemistry (b),
on hyporheic crustaceans (c), and on SPM chemistry (d); the
variable space of the total MFA (e) and the total MFA (f);
T temperature, EC electrical conductivity, P physico-chemical
features, C interstitial water chemistry, F hyporheic Crustacea,
S SPM chemistry
Hydrobiologia (2011) 669:63–82 77
123

which are better adapted to narrow interstices due to
their finer shape, while the larger species can exploit
larger spaces. Another abundant group was Oligo-
chaeta with the main representative being a common
hyporheic genus, Trichodrilus. These dominant
groups were recorded from different periods or/and
stations, a pattern previously reported for other rivers
(Danielopol et al., 1997).
Using multiple sampling stations with differing
water qualities enabled the structure of biological
communities in sites with natural and polluted waters
to be compared. Station 1 was not affected by heavy
metal pollution due to its geographical position
upstream of the main pollutant inflow. However,
Station 1 (with high concentrations of Na, K, and Cl)
revealed the effects of another human impact: salt
used during winter on a nearby road. There were
downstream stations (Stations 9, 10 and 12) whose
communities were not affected by heavy metal
pollution due to water dilution. Species richness and
abundance of individuals were also variable among
the unpolluted stations.
In the most polluted stations (Stations 4 and 11),
fewer individuals were collected during the study
compared with more than 1,300 individuals from
moderately polluted Station 7, downstream of the
polluted area. In some samples, polluted stations
almost completely lacked fauna. Macrocrustaceans,
however, were dominant in the moderately polluted
stations, such as Stations 2 and 7. Such lack of
sensitivity to sediment toxicity has been reported for
other benthic macroinvertebrates, which are more
sensitive to water pH (Van Damme et al., 2008).
The highest numbers of hyporheic Crustacea (both
individuals and species) were found in the moderately
polluted stations (Stations 2 and 7) and less polluted,
downstream stations (Stations 9, 10, and 12). The
most polluted stations had both fewer faunal groups
and typical hyporheic Crustacea (Stations 4, 5, and
6). The highest number of individuals was found in
moderately polluted stations (Stations 2 and 7)
followed by the unpolluted stations (Stations 3, 9,
10, and 12).
The ‘‘intermediate disturbance hypothesis’’
(Connell, 1978) states that intermediate levels of
disturbance will lead to the greatest species diversity.
In an undisturbed or rarely disturbed environment,
the most competitively dominant species will take
over, whereas in a highly disturbed environment, only
species that can cope with the disturbance will
survive. Between these two extremes, both types of
species populate the same place, thus enhancing the
species diversity. This theory may be applied to our
study of the Aries River, where the moderately
disturbed stations (Stations 2, 7, and 9), supported the
highest diversity and number of individuals.
The general spatial pattern of occurrence of
invertebrates, and also the pattern of hyporheic
crustaceans, can be explained by water chemistry.
However, neither interstitial water nor SPM chemis-
try could explain the differences between stations
with similar level of pollution and there appear to be
additional factors affecting species occurrence. One
of these is the heterogeneous grain size distribution in
the Aries alluvia, which is beneficial for hyporheic
fauna (Bretschko, 1991; Schmid-Araya, 1998; Chafiq
Fig. 8 Evaluation of the correlation between SOD activity and the two chemical parameters for different sampling stations:
correlation is positive between COD and SOD activity in samples collected during May 2008 (a) and during November 2008 (b)
78 Hydrobiologia (2011) 669:63–82
123

et al., 1999). Different authors have emphasized
sediments as a significant factor in the interactions
between biotic and abiotic elements of surface and
interstitial waters (Hendricks & White, 1995; Brunke
& Gonser, 1997; Dole-Olivier, 1998). Others high-
light the importance of organic matter (Griebler &
Mosslacher, 2003; Datry et al., 2005; Hahn &
Matzke, 2005), or bacteria (Brown et al., 2003;
Griebler & Mosslacher, 2003; Febria et al., 2009) in
shaping the biological communities of groundwater
and hyporheic zone.
Typical interstitial communities are composed of
fauna with different degrees of adaptations to micro-
habitats and to various (micro-) environmental needs.
The presence of extremely tiny (Parastenocaris sp.)
and large crustaceans (isopods and amphipods) in the
same sample emphasizes the heterogeneity of the
hyporheic habitat. Local impacts, organic matter
input, sediment structure, and many other features
of the physical–chemical-biological microcosm will
favor some species and some assemblages of species
(see review in Bork et al., 2009).
Ecological and evolutionary implications
The general axiom in groundwater ecology is that
typical hyporheic animals have low ecological toler-
ance (Dole-Olivier et al., 2009), and can survive only in
relatively stable habitats. Our results from the Aries
River show that colonization of different levels of the
hyporheic zone is not necessarily a result of stressful
conditions or climate, such as heavy metal pollution,
but exists more because of lower levels of stress. The
stressing condition can be, in this case, competition
from surface species or from other hyporheic repre-
sentatives. In the Aries River interstitial, heavy metal
pollution is the limiting factor for surface benthic and
for hyporheic species sensitive to pollution, but will
indirectly ‘promote’ the development of populations of
pollution-resistant species. Macrocrustaceans and
some microcrustaceans (harpacticoids) in intermediate
stations may have developed larger populations due to
a lack of competition for resources from benthic
surface species or from other hyporheos.
Originating from epigean organisms with wide
ecological tolerance and retaining plasticity for
vertical and horizontal migrations in a highly patchy
and heterogeneous three-dimensional biotope,
groundwater species are opportunistic in their attempt
to colonize lower levels of the interstitial habitat
(Rouch & Danielopol, 1987). In stressful conditions,
species with higher adaptability, resistance or mobil-
ity will flourish. Reduction of competition and
predation will give opportunistic or ‘neutral’ species
the possibility to develop larger populations.
The interstitial habitat is an ecotone with a contin-
uous interplay between surface and subsurface
communities colonizing each vertical layer of a three-
dimensional environment (Boulton, 2000b; Hakenkamp
& Palmer, 2000). The balance may favor epigean or
hyporheic species, depending on water quality and
microbiota, because the interstitial habitat is extremely
dynamic in space and time (Datry et al., 2005; Hahn,
2006), and interspecific competition is intense.
For the Aries River surface waters it has been found
that the number of individuals and species of benthic
macroinvertebrate groups decrease drastically in
response to pollution (Momeu et al., 2007). However,
hyporheic fauna flourish in the moderately polluted
locations. The low metabolic rates of hyporheic
species versus intense metabolism of surface species
(e.g. Culver, 1982; Danielopol et al., 1994; Mosslacher
& Creuze des Chatelliers, 1996; Malard & Hervant,
1999) is one of the factors favouring hyporheic species
and enabling them to resist episodes of water pollution
for longer periods. In such situations, the hyporheic
fauna will also invade those superficial layers depleted
of surface fauna.
In general terms, the hyporheic fauna is more
tolerant of heavy metal pollution than animals of the
same taxonomical groups in surface water. The
hyporheic Crustacea of the Aries River appeared to
be insensitive to heavy metals, probably until
concentrations reach lethal levels. Plenet (1995)
described a similar situation, with no relationship
between hyporheic amphipod abundance and metal
levels suggesting that these taxa could not be used as
biomonitors for metal contamination. Macrocrusta-
ceans are resistant to heavy metal pollution and can
survive and develop larger populations by lack of
competition and predation from surface representa-
tives which are normally present in the upper layers
of the interstitial habitat.
SOD activity significance
The indirect measurements of SOD activity in
organisms collected from various sampling sites
Hydrobiologia (2011) 669:63–82 79
123

along Aries River have provided an interesting
insight on the response of a whole interstitial
invertebrate community to an environmental chal-
lenge from one particular, or a group, of environ-
mental factors. The assay of SOD activity in
biological samples has the advantages of low cost,
speed and the need of a very small amount of
biomass. The strong correlations found between SOD
activity in pooled tissues extract and some chemical
parameters, such as Pb concentration and COD,
suggesting that the oxidative stress enzymes may be
sensitive indicators of chemical pollution in intersti-
tial zones and could be used in assessing and
monitoring pollution. The values of the SOD activity
are congruent to those obtained for the distribution of
hyporheic communities along the studied transect.
Stations with both fewer fauna groups and species
have higher SOD activity. Until present, experiments
were undertaken only on whole communities and the
method is refined by using Oligochaeta (Rusu et al.,
2010). Further investigations will focus on macro-
crustacean species or other taxa sensitive to heavy
metal pollution, though difficulties arise from the
need to collect a sufficient number of individuals
from different stations in the same period of time.
Final remarks
Heavy metals associated with mining can pollute
surface streams, groundwater and alluvial aquifers,
which in turn will impact on both the hydrological
exchange and biological activity of the hyporheic
zone (Hancock, 2002). The reduction of both the
number of invertebrate groups and crustacean species
in the polluted sectors of the Aries River supports
this. However, water chemistry was not the only
determinant of hyporheic biodiversity, as was also
emphasized by Dole-Olivier et al. (2009). There are
factors related to hydraulic conductivity, availability
of food resources, physical habitat fragmentation, and
biological interactions that can be also important.
The presence of high biodiversity levels in the
moderately polluted stations compared to strongly
polluted stations, or even cleaner waters, and the
increase of the number of some hyporheos (especially
macrocrustaceans and oligochetes) in the intermedi-
ate stations suggests that factors other than just water
chemistry influence their spatial distribution. On the
Aries River, such changes over a relatively short
longitudinal transect, may be explained by (1) higher
tolerance of some hyporheic species compared to
surface fauna in response to heavy metal pollution;
(2) biotic interactions between surface and subterra-
nean species; and (3) the physical habitat fragmen-
tation given by extreme diverse granulometry of the
Aries alluvia.
Further research into the effects of pollution and
other environmental impacts on interstitial habitats
needs to focus both on the identification of hyporheic
species (such as micro-crustaceans) and of oxidative
stress enzymes that can be used as indicators of
chemical pollution in interstitial zones.
Acknowledgments We are grateful to Frank Fiers for useful
discussions and suggestions, to Geza Rajka, Akos Nagy and
Nimrod Nemeth for help with samplings, to Vlad Paul for insect
larvae identification, and to Mihai Terente for the map. Claire
Stephens was very helpful in improving the content and the
English of the manuscript. Stuart Halse, Koen Martens and two
anonymous reviewers made valuable remarks improving
considerably the manuscript quality. This study was funded
through the grants 31_032/2007 (CNMP, Ministry of Education,
Research and Innovation, Romania) and SYNTHESYS BE-TAF
4681 (European Union-funded Integrated Activities grant).
References
Bork, J., S. E. Berkhoff, S. Bork & H. J. Hahn, 2009. Using
subsurface metazoan fauna to indicate groundwater–sur-
face water interactions in the Nakdong River floodplain.
South Korea Hydrogeology Journal 17: 61–75.
Bou, C., 1974. Recherches sur les eaux souterraines. 25—
Methodes de recolte dans les eaux souterraines interstiti-
elles. Annales de Speleologie 29: 611–619.
Boulton, A. J., 2000a. River ecosystem health down under:
assessing ecological condition in riverine groundwater
zones in Australia. Ecosystem Health 6: 118–198.
Boulton, A. J., 2000b. The subsurface macrofauna. In Jones, J.
B. & P. J. Mulholland (eds), Streams and Ground Waters.
Academic Press, San Diego: 337–361.
Boulton, A. J., S. Findlay, P. Marmonier, E. H. Stanley & H.
M. Valett, 1998. The functional significance of the hyp-
orheic zone in streams and rivers. Annual Review of
Ecology, Evolution, and Systematics 29: 59–81.
Bourg, A. C. M. & C. Bertin, 1993. Biogeochemical processes
during the infiltration of river water into an alluvial
aquifer. Environmental Science & Technology 27:
661–666.
Bradford, M. M., 1976. A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing
the principle of protein-dye binding. Analytical Bio-
chemistry 72: 248–254.
Bretschko, G., 1991. Bedsediments, groundwater and
stream limnology. Verhandlungen der Internationalen
80 Hydrobiologia (2011) 669:63–82
123

Vereinigung fur Theoretische und Angewandte Limnologie
24: 1957–1960.
Brown, R. J., S. D. Rundle, T. H. Hutchinson, T. D. Williams
& M. B. Jones, 2003. Small-scale detritus-invertebrate
interactions: Influence of detrital biofilm composition on
development and reproduction in a meiofaunal copepod.
Archiv fur Hydrobiologie 157: 117–129.
Brunke, M. & T. Gonser, 1997. The ecological significance of
exchange processes between rivers and groundwater.
Freshwater Biology 37: 1–33.
Cantu-Medellın, N., N. O. Olguın-Monroy, L. C. Mendez-
Rodrıguez & T. Zenteno-Savın, 2009. Antioxidant
enzymes and heavy metal levels in tissues of the black
chocolate clam Megapitaria squalida in Bahıa de La Paz,
Mexico. Archives of Environmental Contamination and
Toxicology 56: 60–66.
Chafiq, M., J. Gibert & C. Claret, 1999. Interactions among
sediments, organic matter, and microbial activity in the
hyporheic zone of an intermittent stream. Canadian
Journal of Fisheries and Aquatic Sciences 56(3): 487–495.
Connell, J. H., 1978. Diversity in tropical rain forests and coral
reefs. Science 199: 1302–1310.
Cravo, A., B. Lopes, A. Serafim, R. Company, L. Barreira, T.
Gomes & M. J. Bebianno, 2009. A multibiomarker
approach in Mytilus galloprovincialis to assess environ-
mental quality. Journal of Environmental Monitoring
11(9): 1673–1686.
Creuze des Chatelliers, M., D. Poinsart & J.-P. Bravard, 1994.
Geomorphology of alluvial groundwater ecosystems. In
Gibert, J., D. L. Danielopol & J. A. Stanford (eds),
Groundwater Ecology. Academic Press, San Diego:
158–185.
Culver, D. C., 1982. Cave Life: Evolution and Ecology. Har-
vard University Press, Cambridge.
Danielopol, D. L., M. Creuze des Chatteliers, F. Mosslacher, P.
Pospisil & R. Popa, 1994. Adaption of Crustacea to inter-
stitial habitats: a practical agenda for ecological studies. In
Gibert, J., D. L. Danielopol & J. A. Stanford (eds),
Groundwater Ecology. Academic, New York: 218–243.
Danielopol, D. L., R. Rouch, P. Pospisil, P. Torreiter & F.
Moßlacher, 1997. Ecotonal animal assemblages; their
interest for groundwater studies. In Gibert, J., J. Mathieu
& F. Fournier (eds), Groundwater/Surface Water Eco-
tones: Biological and Hydrological Interactions and
Management Options. Cambridge University Press,
Cambridge: 11–20.
Datry, T., F. Malard & J. Gibert, 2005. Response of inverte-
brate assemblages to increased groundwater recharge rates
in a phreatic aquifer. Journal of the North American
Benthological Society 24: 461–477.
de Oliveira, U. O., A. S. da Rosa Araujo, A. Bello0-Klein, R.
S. M. da Silvaa & L. C. Kucharski, 2005. Effects of
environmental anoxia and different periods of reoxygen-
ation on oxidative balance in gills of the estuarine crab
Chasmagnathus granulate. Comparative Biochemistry
and Physiology Part B 140: 51–57.
Dole-Olivier, M.-J., 1998. Surface water-groundwater
exchanges in three dimensions on a backwater of the
Rhone River. Freshwater Biology 40: 93–109.
Dole-Olivier, M.-J., F. Malard, D. Martin, T. Leferbure &
J. Gibert, 2009. Relationships between environmental
variables and groundwater biodiversity at the regional
scale. Freshwater Biology 54: 797–813.
El Adnani, M., A. Ait Boughrous, M. Y. Khebiza, A. El
Gharmali, M. L. Sbai, A. S. Errouane, L. Loukili Idrissi &
A. Nejmeddine, 2007. Impact of mining wastes on the
physicochemical and biological characteristics of
groundwater in a mining area in Marrakech (Morocco).
Environmental Technology 28: 71–82.
Fan, W. H., G. Tang, C. M. Zhao, Y. Duan & R. Zhang, 2009.
Metal accumulation and biomarker responses in Daphniamagna following cadmium and zinc exposure. Environ-
mental Toxicology & Chemistry 28(2): 305–310.
Febria, C. M., R. R. Fulthorpe & D. D. Williams, 2009.
Characterizing seasonal changes in physicochemistry and
bacterial community composition in hyporheic sediments.
Hydrobiologia. doi:10.1007/s10750-009-9882-x.
Forray, F. L., 2001. Aplicarea analizei factoriale ın studiul
poluarii raului Aries (Muntii Apuseni). Studia Universi-
tatis Babes-Bolyai (Geologia) 46: 47–58.
Fuller, C. C. & J. W. Harvey, 2000. Reactive uptake of trace
metals in the hyporheic zone of a mining-contaminated
stream, Pinal Creek, Arizona. Environmental Science &
Technology 34: 1150–1155.
Geret, F., A. Serafim, L. Barreira & M. J. Bebianno, 2002.
Effect of cadmium on antioxidant enzyme activities and
lipid peroxidation in the gills of the clam Ruditapes de-cussates. Biomarkers 7(3): 242–256.
Griebler, C. & F. Mosslacher, 2003. Grundwasser-eine oko-
systemische Betrachtung. In Griebler, C. & F. Mosslacher
(eds), Grundwasserokologie. UTB-Facultas Verlag, Wien:
253–310.
Hahn, H. J., 2006. The GW-fauna-index: a first approach to a
quantitative ecological assessment of groundwater habi-
tats. Limnologica 36: 119–137.
Hahn, H. J. & D. Matzke, 2005. A comparison of stygofauna
communities inside and outside groundwater bores. Lim-
nologica 35: 31–44.
Hakenkamp, C. C. & M. A. Palmer, 2000. The ecology of
hyporheic meiofauna. In Jones, J. B. & P. J. Mulholland
(eds), Streams and Ground Waters. Academic, San Diego:
307–335.
Hancock, P. J., 2002. Human impacts on the stream-ground-
water exchange zone. Environmental Management 29(6):
763–781.
Hendricks, S. P. & D. S. White, 1995. Seasonal biogeochem-
ical patterns in surface water, subsurface hyporheic, and
riparian ground water in a temperate stream ecosystem.
Archiv fur Hydrobiologie 134: 459–490.
Hinkle, S., J. Duff, F. Triska, A. Laenen, E. Gates, K. Bencala,
D. Wentz & S. Silva, 2001. Linking hyporheic flow and
nitrogen cycling near the Willamette River—a large river
in Oregon, USA. Journal of Hydrology 244: 157–180.
Malard, F. & F. Hervant, 1999. Oxygen supply and the adaptations
of animals in groundwater. Freshwater Biology 41: 1–30.
Malard, F., J.-L. Reygrobellet, J. Mathieu & M. Lafont, 1994.
The use of invertebrate communities to describe ground-
water flow and contaminant transport in a fractured rock
aquifer. Archiv fur Hydrobiologie 131(1): 93–110.
Malard, F., S. Plenet & J. Gibert, 1996. The use of inverte-
brates in ground water monitoring: a rising research field.
Ground Water Monitoring and Remediation 16: 103–113.
Hydrobiologia (2011) 669:63–82 81
123

Manduzio, H., T. Monsinjon, C. Galap, F. Leboulenger &
B. Rocher, 2004. Seasonal variations in antioxidant
defences in blue mussels Mytilus edulis collected from a
polluted area: major contributions in gills of an inducible
isoform of Cu/Zn-superoxide dismutase and of glutathi-
one S-transferase. Aquatic Toxicology 70(1): 83–93.
Moldovan, O., Iepure, S. & A. Persoiu, 2005. Biodiversity and
protection of Romanian karst areas: the example of
interstitial fauna. In Stevanovic, Z. & P. Milanovic (eds),
Water Resources and Environmental Problems in Karst.
Proceedings International Conference & Field Seminar,
Beograd & Kotor, 13–19 September 2005: 831–836.
Momeu, L., K. W. Battes, F. Pricope, A. Avram, K. P. Battes,
M. Cımpean, D. Ureche & I. Stoica, 2007. Preliminary
data on algal, macroinvertebrate and fish communities
from the Aries catchment area, Transylvania, Romania.
Studia Universitatis Babes-Bolyai (Biologıa) 52: 25–36.
Morel, F. M. M., 1983. Principles of Aquatic Chemistry.
Wiley, New York.
Moser, D. P., J. K. Fredrickson, D. R. Geist, E. V. Arntzen, A.
D. Peacock & S.-M. W. Li, 2003. Biogeochemical pro-
cesses and microbial characteristics across groundwater-
surface water boundaries of the Hanford Reach of the
Colombia River. Environmental Science & Technology
37: 5127–5134.
Mosslacher, F. & J. Notenboom, 1999. Groundwater biomon-
itoring. In Gerhardt, A. (ed.), Biomonitoring of Polluted
Water. Environmental Science Forum 96: 119–140.
Mosslacher, F. & M. Creuze des Chatelliers, 1996. Physio-
logical and behavioural adaptations of an epigean and a
hyporheic dwelling population of Asellus aquaticus (L.)
(Crustacea, Isopoda). Archiv fur Hydrobiologie 138:
187–198.
Munk, L., G. Faure, D. Pride & J. Bigham, 2002. Sorption of
trace metals to an aluminum precipitate in a stream
receiving acid rock-drainage; Snake River, Summit
County, Colorado. Applied Geochemistry 17: 421–430.
Notenboom, J., S. Plenet & M.-J. Turquin, 1994. Groundwater
contamination and its impact on groundwater animals and
ecosystems. In Gibert, J., D. L. Danielopol & J. A. Stan-
ford (eds), Groundwater Ecology. Academic Press, San
Diego: 477–504.
Pages, J., 2004. Multiple factor analysis: main features and
application to sensory data. Revista Colombiana de Est-
adistica 27: 1–26.
Plenet, S., 1995. Freshwater amphipods as biomonitors of
metal pollution in surface and interstitial aquatic systems.
Freshwater Biology 33: 127–137.
Plenet, S., 1999. Metal accumulation by an epigean and a
hypogean freshwater amphipod: considerations for water
quality assessment. Water Environment Research 71:
1298–1309.
Plenet, S. & J. Gibert, 1994. Invertebrate community responses
to physical and chemical factors at the river/aquifer
interaction zone. I. Upstream from the city of Lyon. Ar-
chiv fur Hydrobiologie 132: 165–189.
Plenet, S., H. Hugueny & J. Gibert, 1996. Invertebrate com-
munity responses to physical and chemical factors at the
river/aquifer interaction zone. II. Downstream from the
city of Lyon. Archiv fur Hydrobiologie 136: 65–88.
Pyle, G. G. & R. S. Mirza, 2007. Copper-impaired chemo-
sensory function and behavior in aquatic animals. Human
Ecological Risk Assessment 13: 492–505.
Rouch, R. & D. Danielopol, 1987. L’origine de la faune
aquatique souterraine, entre le paradigme du refuge et le
modele de la colonization active. Stygologia 3: 345–372.
Rusu, M., H. L. Banciu, M. Banciu, T. Brad & O. T. Moldovan,
2010. Oxidative stress enzymes as biomarkers of heavy
metal pollution in interstitial invertebrates. Studia Uni-
versitatis Babes-Bolyai 2: 61–66.
Schmid-Araya, J. M., 1998. Small-sized invertebrates in a
gravel stream: community structure and variability of
benthic rotifers. Freshwater Biology 39: 25–39.
Ukeda, H., T. Shimamura, M. Tsubouchi, Y. Harada, Y. Nakai
& M. Sawamura, 2002. Spectrophotometric assay ofsuperoxide anion formed in Maillard reaction based on
highly water-soluble tetrazolium salt. Analytical Sciences
18: 1151–1154.
Van Damme, P. A., C. Hamel, A. Ayala & L. Bervoets, 2008.
Macroinvertebrate community response to acid mine
drainage in rivers of the High Andes (Bolivia). Environ-
mental Pollution 156: 1061–1068.
Williams, D. D. & R. R. Fulthorpe, 2003. Using invertebrate
and microbial communities to assess the condition of the
hyporheic zone of a river subject to 80 years of contam-
ination by chlorobenzenes. Canadian Journal of Zoology
8: 789–802.
Wong, S. W., P. T. Leung, A. B. Djurisic & K. M. Leung,
2010. Toxicities of nano zinc oxide to five marine
organisms: influences of aggregate size and ion solubility.
Analytical and Bioanalytical Chemistry 396(2): 609–618.
82 Hydrobiologia (2011) 669:63–82
123




















