
Sorafenib inhibits growth, migration, and angiogenic potential of ectopic endometrial mesenchymal...
12
Sorafenib inhibits growth, migration, and angiogenic potential of ectopic endometrial mesenchymal stem cells derived from patients with endometriosis Aldo Moggio, M.Sc., a Giulia Pittatore, M.D., b Paola Cassoni, M.D., Ph.D., c Gian Luigi Marchino, M.D., b Alberto Revelli, M.D., Ph.D., c and Benedetta Bussolati, M.D., Ph.D. a a Department of Internal Medicine, Molecular Biotechnology Center; b Physiopathology of Reproduction and IVF Unit, Department of Gynecological and Obstetrical Sciences; and c Department of Biomedical Sciences and Human Oncology, University of Torino, Torino, Italy Objective: To characterize the proliferation, migration, and angiogenic properties of mesenchymal stem cells (MSC) from ectopic and eutopic endometrial tissue and to investigate the effect of the tyrosine kinase inhibitor sorafenib. Design: In vitro studies. Setting: University hospital and research center. Patient(s): Patients receiving surgical treatment of endometriosis (n ¼ 4) and control patients without endometriosis (n ¼ 2) under- going surgery for benign gynecologic diseases. Intervention(s): Mesenchymal stem cell lines were isolated from ectopic and eutopic endometrial tissue, and sorafenib was adminis- tered to them. Main Outcome Measure(s): Proliferation, migration, invasion of endometrial MSC, and expression of ezrin, vascular endothelial growth factor, and hypoxia-inducible factor-1a (HIF-1a) were measured. Result(s): Ectopic endometrial MSC from patients with endometriosis showed a higher proliferation, migration, and angiogenic ability than eutopic MSC from the same patient or control MSC from patients without endometriosis. Sorafenib reduced the proliferation, mo- tility, ezrin phosphorylation, vascular endothelial growth factor release, and HIF-1a expression of ectopic MSC. Conclusion(s): The increased proliferative, migratory, and angiogenic phenotype of ectopic MSC may be reverted by treatment with sorafenib. Targeting of the MSC population involved in sustaining the ectopic lesions might be useful in eradicating endometriotic implants. (Fertil Steril Ò 2012;98:1521–30. Ó2012 by American Society for Reproductive Medicine.) Key Words: Endometriosis, mesenchymal stem cells, sorafenib, VEGF, ezrin, HIF-1a Discuss: You can discuss this article with its authors and with other ASRM members at http:// fertstertforum.com/moggioa-sorafenib-mesenchymal-stem-cells/ Use your smartphone to scan this QR code and connect to the discussion forum for this article now.* * Download a free QR code scanner by searching for “QR scanner” in your smartphone’s app store or app marketplace. E ndometriosis is a common gyne- cologic disorder characterized by the growth of endometrial tissue outside the uterine cavity (1). It affects 6%–10% of all women and 35%–50% of women with pelvic pain and infertility (2). Many hypotheses have been made about the histologic origin of endometri- osis. First, Sampson's theory postulated that endometriotic implants may arise from retrograde menstruation of endo- metrial tissue through the fallopian tubes into the peritoneal cavity (3, 4). In addition, genetic and immunologic factors seem to play important roles in the pathogenesis of endometriosis, because they may be involved in the survival of cells inside the ectopic lesions (5–7). In the last decade, cell Received March 20, 2012; revised July 11, 2012; accepted August 3, 2012; published online September 12, 2012. A.M. has nothing to disclose. G.P. has nothing to disclose. P.C. has nothing to disclose. G.L.M. has noth- ing to disclose. A.R. has nothing to disclose. B.B. has nothing to disclose. This work was supported by Italian Ministry of University and Research grants PRIN08 and ex60% to B.B. Reprint requests: Benedetta Bussolati, M.D., Ph.D., University of Torino, Molecular Biotechnology Centre, Department of Internal Medicine, C.so Dogliotti 14, 10126 Torino, Italy (E-mail: [email protected]). Fertility and Sterility® Vol. 98, No. 6, December 2012 0015-0282/$36.00 Copyright ©2012 American Society for Reproductive Medicine, Published by Elsevier Inc. http://dx.doi.org/10.1016/j.fertnstert.2012.08.003 VOL. 98 NO. 6 / DECEMBER 2012 1521
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Sorafenib inhibits growth, migration, and angiogenic potential of ectopic endometrial mesenchymal...

Sorafenib inhibits growth, migration,and angiogenic potential of ectopicendometrial mesenchymal stem cellsderived from patientswith endometriosis
Aldo Moggio, M.Sc.,a Giulia Pittatore, M.D.,b Paola Cassoni, M.D., Ph.D.,c Gian Luigi Marchino, M.D.,bAlberto Revelli, M.D., Ph.D.,c and Benedetta Bussolati, M.D., Ph.D.a
a Department of Internal Medicine, Molecular Biotechnology Center; b Physiopathology of Reproduction and IVF Unit,Department of Gynecological and Obstetrical Sciences; and c Department of Biomedical Sciences and Human Oncology,University of Torino, Torino, Italy
Objective: To characterize the proliferation, migration, and angiogenic properties of mesenchymal stem cells (MSC) from ectopic andeutopic endometrial tissue and to investigate the effect of the tyrosine kinase inhibitor sorafenib.Design: In vitro studies.Setting: University hospital and research center.Patient(s): Patients receiving surgical treatment of endometriosis (n ¼ 4) and control patients without endometriosis (n ¼ 2) under-going surgery for benign gynecologic diseases.Intervention(s): Mesenchymal stem cell lines were isolated from ectopic and eutopic endometrial tissue, and sorafenib was adminis-tered to them.Main Outcome Measure(s): Proliferation, migration, invasion of endometrial MSC, and expression of ezrin, vascular endothelialgrowth factor, and hypoxia-inducible factor-1a (HIF-1a) were measured.Result(s): Ectopic endometrial MSC from patients with endometriosis showed a higher proliferation, migration, and angiogenic abilitythan eutopic MSC from the same patient or control MSC from patients without endometriosis. Sorafenib reduced the proliferation, mo-tility, ezrin phosphorylation, vascular endothelial growth factor release, and HIF-1a expression of ectopic MSC.
Use your smartphone
Conclusion(s): The increased proliferative, migratory, and angiogenic phenotype of ectopicMSC may be reverted by treatment with sorafenib. Targeting of the MSC population involvedin sustaining the ectopic lesions might be useful in eradicating endometriotic implants. (FertilSteril� 2012;98:1521–30. �2012 by American Society for Reproductive Medicine.)Key Words: Endometriosis, mesenchymal stem cells, sorafenib, VEGF, ezrin, HIF-1a
Discuss: You can discuss this article with its authors and with other ASRM members at http://fertstertforum.com/moggioa-sorafenib-mesenchymal-stem-cells/
to scan this QR codeand connect to thediscussion forum forthis article now.*
* Download a free QR code scanner by searching for “QRscanner” in your smartphone’s app store or app marketplace.
E ndometriosis is a common gyne-cologic disorder characterized bythe growth of endometrial tissue
Received March 20, 2012; revised July 11, 2012; accep12, 2012.
A.M. has nothing to disclose. G.P. has nothing to discling to disclose. A.R. has nothing to disclose. B.B
This work was supported by Italian Ministry of UniveB.B.
Reprint requests: Benedetta Bussolati, M.D., Ph.D.,Centre, Department of Internal Medicine, [email protected]).
Fertility and Sterility® Vol. 98, No. 6, December 2012Copyright ©2012 American Society for Reproductivehttp://dx.doi.org/10.1016/j.fertnstert.2012.08.003
VOL. 98 NO. 6 / DECEMBER 2012
outside the uterine cavity (1). It affects6%–10% of all women and 35%–50%ofwomenwithpelvic pain and infertility
ted August 3, 2012; published online September
ose. P.C. has nothing to disclose. G.L.M. has noth-. has nothing to disclose.rsity and Research grants PRIN08 and ex60% to
University of Torino, Molecular Biotechnology.so Dogliotti 14, 10126 Torino, Italy (E-mail:
0015-0282/$36.00Medicine, Published by Elsevier Inc.
(2). Many hypotheses have been madeabout the histologic origin of endometri-osis. First, Sampson's theory postulatedthat endometriotic implants may arisefrom retrograde menstruation of endo-metrial tissue through the fallopiantubes into the peritoneal cavity (3, 4).In addition, genetic and immunologicfactors seem to play important roles inthe pathogenesis of endometriosis,because they may be involved in thesurvival of cells inside the ectopiclesions (5–7). In the last decade, cell
1521

ORIGINAL ARTICLE: ENDOMETRIOSIS
populations displaying stem cell markers/properties wereidentified in the basalis layer of endometrium (8–17), addingnew perspectives to the pathogenesis of endometriosis (18).Endometrial stem cells were concordantly isolated andcharacterized as cells with mesenchymal origin and multi-differentiative properties (13–17) and were implicated incyclic endometrial regeneration (9, 10, 15–17). Stem cellswith a mesenchymal phenotype were also isolated fromectopic endometrial implants (19). It was proposed thatendometrial stem cells, shed through the fallopian tubeduring menstruation, could be responsible for theestablishment of endometrial implants (18). However, thebiological mechanisms triggering the characteristics ofmesenchymal stem cells (MSC) in the eutopic and ectopicendometrium still deserve further investigation.
Endometriosis is not a cancer; however, it is characterizedby some of the adaptive properties of tumor cells, such as mi-gration, invasion, and angiogenesis. Indeed, survival and pro-liferation of endometrial lesions strictly depend on theformation of new blood vessels, which provide oxygen andnutrient supply (19–21). Accordingly, antiangiogenictreatment and vascular-disrupting agents could representa possible therapeutic strategy against this pathology (22, 23).
In the present study, we isolated MSC from ectopicendometrial tissue to evaluate their characteristics in termsof proliferation, migration, ezrin phosphorylation, hypoxia-inducible factor-1a (HIF-1a), and vascular endothelialgrowth factor (VEGF) expression with respect to MSC ob-tained from eutopic tissue of the same patient. Subsequently,we tested the effect of sorafenib, a multi-tyrosine kinase in-hibitor (24), as a possible tool to modulate their growth, mi-gration, and angiogenic properties.
MATERIALS AND METHODSMSC Isolation and Culture
The cell lines were obtained from four patients receiving sur-gery for treatment of ovarian and peritoneal endometriosisand from two control patients undergoing polypectomy andmyomectomy (Supplemental Table 1, available online) inthe Department of Obstetrics and Gynecology, University ofTorino. Two samples were collected from the same patientwith endometriosis—one of eutopic endometrium and one ofectopic implant. This study was approved by the Ethics Re-view Board of Ospedale Infantile Regina Margherita SantAnna (Torino). Preoperative informed consent was obtainedfrom each patient. Tissues were processed enzimaticallywith 0.1% type I collagenase (Sigma-Aldrich) for 30 minutesin a 37�C incubator. Later, cell aggregates were filteredthrough 60-mm and 120-mm meshes. Cells were seeded ata density of 1.0–1.5� 105 viable cells (80% viable cells deter-mined by trypan blue) per cm2 in endothelial cell basal me-dium with an EGM-MV kit (Lonza; containing epidermalgrowth factor, hydrocortisone, bovine brain extract, and 5%fetal calf serum [FCS]) previously described for MSC isolation(25). Dead cells were poured off 72 hours later and, after 5–7days, cell clones were typically observed. Cells were passagedat confluence and after 2–3 days in the subsequent passages.The endometrial MSC obtained (eutopic MSC, n ¼ 4; ectopic
1522
MSC, n ¼ 4; control MSC, n ¼ 2) were cultured for at least20 passages to test the self-renewal ability typical of MSC.Bone marrow–derived MSC were cultured in mesenchymalstem cells basal medium (Lonza).
Flow Cytometric Analysis
Cytometric analysis was performed using FACScan (BectonDickinson) as previously described (25). Details are providedin Supplemental Materials and Methods (available online).
Real-Time Polymerase Chain Reaction Analysis
Gene expression was performed by quantitative real timereverse transcription-polymerase chain reaction (RT-PCR)with Applied Biosystems StepOne, as previously described(26). Details are provided in Supplemental Materials andMethods.
Immunofluorescence and Immunohistochemistry
Immunofluorescence was performed on chamber slides(Sigma) on which cells were fixed in 4% paraformaldehydecontaining 2% sucrose for 15 minutes at 4�C, permeabilizedwith 0.1% Triton X-100 (Sigma) for 8 minutes at 4�C, andthen incubated overnight at 4�C with the appropriate anti-bodies. Anti–pan-cytokeratin (Biomeda), anti-vimentin(Sigma), mouse anti–E-cadherin (DakoCytomation), and rab-bit anti-estrogen receptor (ER) (clone SP1; Ventana-Diapath)antibodies were used. Primary antibodies were detected usinganti-mouse or rabbit secondary antibodies conjugated withAlexa Fluor 488 or Texas Red (Molecular Probes). Hoescht33258 dye (Sigma) was added for nuclear staining, and imag-ingwas performedusing an LSM5Pascal confocalmicroscope(Carl Zeiss International). Substitution with an unrelated rab-bit serum or mouse IgG served as negative control. For immu-nohistochemistry, cell lines were harvested by cell scraperbefore reaching confluence, washed, and fixed in 4%neutral-buffered formalin (Histo-Line Laboratories) at roomtemperature. Afterward, cell pellets were processed to paraffinembedding with an automatic processor (ASP 300; Leica Mi-crosystems). One section from each block was stainedwith he-matoxylin and eosin. Immunohistochemistry was performedon additional sections using an automated slide-processingplatform (BenchMark AutoStainer; Ventana) with predilutedanti-ER monoclonal antibody (clone SP1; Ventana-Diapath).
In Vitro Differentiation
The MSC lines were seeded in differentiating conditions, aspreviously described (25). For in vitro osteogenic differenti-ation, cells were cultured (50,000 cells per well) in osteo-genic medium (Lonza) for 2 weeks, and differentiation wasanalyzed by staining with alizarin red (Lonza) for 20 min-utes. For epithelial differentiation, cells were seeded at50,000 cells per well with Roswell Park Memorial Institute1640 medium (Lonza) with addition of 10% FCS, 20 ng/mL hepatocyte growth factor, 10 ng/mL fibroblast growthfactor-4, and 10�9 M dexamethasone (all from Sigma). After2 weeks of differentiation, cells were subjected to immuno-fluorescence. Expression of epithelial markers was analyzed
VOL. 98 NO. 6 / DECEMBER 2012

Fertility and Sterility®
semi-quantitatively by measuring fluorescence intensity bydigital image analysis after subtraction of the backgroundfluorescence, using the LSM image analysis program (CarlZeiss International). Ten microscopic fields (magnification,�40) were analyzed for each marker.
Drugs and Reagents
Sorafenib (BAY-9006), provided by Bayer Pharmaceuticals,was dissolved in polyethylene glycol 400 (Sigma) at a finalconcentration of 10 mM and stored at �20�C. The drug wasdiluted in the culture medium to the desired concentration,range 1–10 mM, as previously described to be effectivein vitro (27). 17b-Estradiol (Sigma) was dissolved in absoluteethanol and subsequently diluted in phenol-free Roswell ParkMemorial Institute 1640 medium (Lonza). Solvent vehicle wasadded to cultures as control.
Cell Proliferation Assay
The cells were seeded at 2,500 cells per well into 96-wellplates. Deoxyribonucleic acid synthesis was detected as incor-poration of 5-bromo-2-deoxyuridine (BrdU) into the cellularDNA after 48 hours of culture in expansion medium. Cellswere fixed with ethanol/HCl and incubated with nuclease.The BrdU incorporated into the DNA was detected using ananti-BrdU peroxidase-conjugated antibody and visualizedwith a soluble chromogenic substrate (Roche Applied Sci-ence). Optical density was measured with an ELISA reader (Bi-oRad) at 415 nm. In selected experiments, cells were treatedwith sorafenib (2.5–10 mM), 10 nM E2, or vehicle alone.
Migration and Invasion Assay
Cells were seeded in transwell chambers with 8-mm pore, ata concentration of 50,000 cells per well in 200-mL endothelialcell basal medium. For invasion assay, cells were seeded ona filter coated with Matrigel (1:5 dilution). In selected exper-iments, cells were treated with sorafenib (5 mM) or with vehi-cle alone. After 24 hours the cells on the underside of thefilters were fixed with methanol for 10 minutes and stainedwith crystal violet solution for 20 minutes. The numbers ofcells migrated and invaded to the underside of the membranewere quantitatively determined by ImageJ software (freelyavailable at http://rsbweb.nih.gov/ij/).
Cytokine Assay
Vascular endothelial growth factor was analyzed by multi-plex cytokine array, based on fluorescently dyed micro-spheres (Bio-Plex; BioRad). Mesenchymal stem cells werecultured to a confluence of 90%. Cells were then exposed toa-minimal essential medium with 0.5% albumin (Sigma) for12 hours. The supernatant was collected, centrifuged (1,200rpm, 4�C, 5 minutes), and frozen at �20�C.
Western Blot Analysis
Western blot analysis was performed as previously described(26). For protein analysis, cells were lysed at 4�C for 30minutes in RIPA buffer (20 nM Tris-Hcl, 150 nM NaCl, 1%
VOL. 98 NO. 6 / DECEMBER 2012
deoxycholate, 0.1% sodium dodecyl sulfate, and 1% TritonX-100, pH 7.8) supplemented with protease and phosphataseinhibitors cocktail and phenylmethylsulfonyl fluoride(Sigma). Aliquots of the cell lysates containing 40 mg protein,as quantified by the Bradford BioRad Protein Assay (BioRad),were run on 10% sodium dodecyl sulfate–polyacrylamide gelelectrophoresis and transferred onto polyvinylidene difluor-ide (PVDF) membrane by the iBlot Dry Blotting System (Invi-trogen). Primary antibodies were anti-actin (Santa CruzBiotechnology), anti-ezrin, anti-fosfo-ezrin-radixin-moesin(ERM) proteins, and anti–hypoxia-inducible factor-1a(HIF-1a) from Cell Signaling Technology. After incubation,membranes were stained with secondary antibodies (Pierce)conjugated with horseradish peroxidase (Pierce). Resultswere analysed with Quantity One software (BioRad). Quanti-fication of the bands was done by Quantity One software (Bi-oRad), and the relative quantification of the band wascalculated through the ratio band-specific antibody/band ac-tin. Every experiment was repeated three times.
Statistical Analysis
Results were expressed as means � SD. Analysis of variancewith Dunnett's multicomparison test or Student's t test wereperformed. Significance was set at P< .05.
RESULTSIsolation and Characterization of Ectopic, Eutopic,and Control MSC
We isolated and generated MSC lines from biopsy specimensof eutopic (Euto-MSC) and ectopic (Ecto-MSC) endometriumof women with endometriosis (n ¼ 4) (SupplementalTable 1), as described in Materials and Methods. ControlMSC lines were obtained from biopsy specimens of womenwithout endometriosis (Ctrl-MSC; n ¼ 2). The Euto-MSC,Ecto-MSC, and Ctrl-MSC lines were phenotypically and func-tionally characterized and were kept in culture until the 20th–28th passage, showing the self-renewal ability typical ofmesenchymal stem cells (14). Experiments were performedon cells between the 3rd and 10th culture passages.
By FACS analysis, all lines were highly positive for theMSC markers CD105, CD73, and CD44 (Fig. 1A;Supplemental Table 1). The expression of CD29 and of the em-bryonic stem cell marker SSEA4 was variable among lines(Supplemental Table 1). The absence of contaminating cellswas shown by the lack of CD34, CD45, and CD14 expressionby endometrial MSC lines (Supplemental Table 1). The pheno-type of these lines was also confirmed by expression of themesenchymal marker vimentin but not of the epithelialmarker cytokeratin by all cell lines, as evaluated by immuno-fluorescence staining (Fig. 2). Moreover, all MSC lines wereable to differentiate in osteoblasts, as shown by stainingwith alizarin red (Fig. 2). No differentiation was observedinto adipocytes in all endometrial MSC lines (data not shown).We also evaluated the expression of embryonic stem cellstranscription factors by quantitative real time RT-PCR analy-sis. Klf and c-Myc and c-Myc genes were overexpressed in theEcto-MSC with respect to Euto-MSC derived from the samepatient, as well as with respect to Ctrl-MSC (Fig. 1B). No
1523
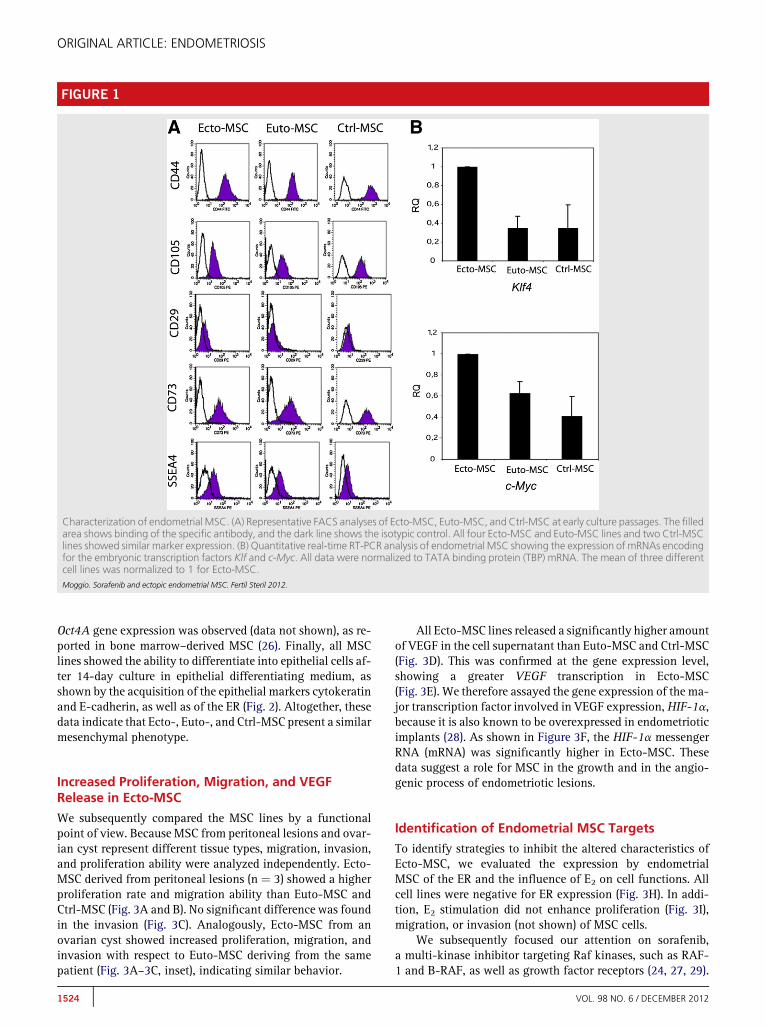
FIGURE 1
Characterization of endometrial MSC. (A) Representative FACS analyses of Ecto-MSC, Euto-MSC, and Ctrl-MSC at early culture passages. The filledarea shows binding of the specific antibody, and the dark line shows the isotypic control. All four Ecto-MSC and Euto-MSC lines and two Ctrl-MSClines showed similar marker expression. (B) Quantitative real-time RT-PCR analysis of endometrial MSC showing the expression of mRNAs encodingfor the embryonic transcription factors Klf and c-Myc. All data were normalized to TATA binding protein (TBP) mRNA. The mean of three differentcell lines was normalized to 1 for Ecto-MSC.Moggio. Sorafenib and ectopic endometrial MSC. Fertil Steril 2012.
ORIGINAL ARTICLE: ENDOMETRIOSIS
Oct4A gene expression was observed (data not shown), as re-ported in bone marrow–derived MSC (26). Finally, all MSClines showed the ability to differentiate into epithelial cells af-ter 14-day culture in epithelial differentiating medium, asshown by the acquisition of the epithelial markers cytokeratinand E-cadherin, as well as of the ER (Fig. 2). Altogether, thesedata indicate that Ecto-, Euto-, and Ctrl-MSC present a similarmesenchymal phenotype.
Increased Proliferation, Migration, and VEGFRelease in Ecto-MSC
We subsequently compared the MSC lines by a functionalpoint of view. Because MSC from peritoneal lesions and ovar-ian cyst represent different tissue types, migration, invasion,and proliferation ability were analyzed independently. Ecto-MSC derived from peritoneal lesions (n ¼ 3) showed a higherproliferation rate and migration ability than Euto-MSC andCtrl-MSC (Fig. 3A and B). No significant difference was foundin the invasion (Fig. 3C). Analogously, Ecto-MSC from anovarian cyst showed increased proliferation, migration, andinvasion with respect to Euto-MSC deriving from the samepatient (Fig. 3A–3C, inset), indicating similar behavior.
1524
All Ecto-MSC lines released a significantly higher amountof VEGF in the cell supernatant than Euto-MSC and Ctrl-MSC(Fig. 3D). This was confirmed at the gene expression level,showing a greater VEGF transcription in Ecto-MSC(Fig. 3E). We therefore assayed the gene expression of the ma-jor transcription factor involved in VEGF expression,HIF-1a,because it is also known to be overexpressed in endometrioticimplants (28). As shown in Figure 3F, the HIF-1a messengerRNA (mRNA) was significantly higher in Ecto-MSC. Thesedata suggest a role for MSC in the growth and in the angio-genic process of endometriotic lesions.
Identification of Endometrial MSC Targets
To identify strategies to inhibit the altered characteristics ofEcto-MSC, we evaluated the expression by endometrialMSC of the ER and the influence of E2 on cell functions. Allcell lines were negative for ER expression (Fig. 3H). In addi-tion, E2 stimulation did not enhance proliferation (Fig. 3I),migration, or invasion (not shown) of MSC cells.
We subsequently focused our attention on sorafenib,a multi-kinase inhibitor targeting Raf kinases, such as RAF-1 and B-RAF, as well as growth factor receptors (24, 27, 29).
VOL. 98 NO. 6 / DECEMBER 2012

FIGURE 2
Characterization and differentiation of endometrial MSC. (A) Representative confocal immunofluorescencemicrographs showing the expression ofvimentin and the absence of cytokeratin by Ecto-MSC, Euto-MSC, and Ctrl-MSC in basal culture conditions. When cultured in osteogenic medium(14 days), MSC lines presented osteogenic differentiation, as shown by alizarin red positivity.When cultured in epithelial differentiatingmedium (14days), MSC lines acquired positivity for cytokeratin, E-cadherin, and ER. Nuclei were stainedwith Hoechst dye 33342. Original magnification:�630.All four Ecto-MSC and Euto-MSC lines and two Ctrl-MSC lines showed similar results. (B) Semiquantitative analysis of the expression of epithelialmarkers as detected by immunofluorescence staining after 14 days of differentiation of endometrial MSC. Data are mean � SD of all cell lines.Student's t test was performed. *P<.05 vs. day 0.Moggio. Sorafenib and ectopic endometrial MSC. Fertil Steril 2012.
Fertility and Sterility®
We evaluated the expression of the sorafenib targets CD117(c-Kit), VEGF-R2 and -3, and CD140 (platelet growthfactor-b receptor), as well as of the Raf-1 downstream proteinezrin. This cytoskeletal protein was previously shown to beinvolved in migration of endometrial stromal cells of patientswith endometriosis (30). All endometrial MSC lines expressedCD140, and not c-Kit or VEGF-R2 and VEGF-R3 (Fig. 3G).Moreover, the expression of the activated phosphorylated
VOL. 98 NO. 6 / DECEMBER 2012
form of ezrin was higher in Ecto-MSC with respect to Euto-MSC and Ctrl-MSC (Fig. 4A).
Effect of Sorafenib on Endometrial MSC Lines
Endometrial MSC lines were treated with sorafenib at 2.5–10mM for 24 hours. Cytotoxicity tests using Annexin V/propi-dium iodide staining showed that sorafenib was not cytotoxic
1525

FIGURE 3
Functional properties and targeting of endometrial MSC. Evaluation of proliferation (A), migration (B), and invasion (C) of Ecto-MSC, Euto-MSC,and Ctrl-MSC from peritoneal lesions (A–C) (n ¼ 3) or from an ovarian cyst (A–C, inset). Values are mean � SD of three different experimentsperformed in triplicate. (D) Evaluation of VEGF release in the culture supernatant of MSC lines. Values are mean � SD of two differentexperiments performed with two different cell lines in duplicate. (E, F) Quantitative RT-PCR analysis of endometrial MSC showing theexpression of mRNAs encoding for VEGF and HIF-1a. Data were normalized to TBP mRNA. The mean of three different cell lines wasnormalized to 1 for Ecto-MSC. Ecto-MSC showed significant higher proliferation, migration, VEGF, and HIF-1a expression than Euto-MSC orCtrl-MSC. Analysis of variance with Dunnett's comparison test was performed. *P<.05, Ecto-MSC vs. Euto-MSC or Ctrl-MSC. (G)Representative FACS analyses of sorafenib-target receptors in MSC lines, showing binding of the specific antibody (filled area) and of theisotypic control (dark line). Only CD140 was expressed on MSC lines. All Ecto-MSC (n ¼ 4), Euto-MSC (n ¼ 4), and Ctrl-MSC (n ¼ 2) lines weretested and showed similar marker expression. (H) Representative micrographs showing the absence of the estrogen receptor by MSC lines byimmunohistochemistry. Magnification: x 250. All MSC lines were analyzed with similar results. (I) Evaluation of proliferation of MSC lines afterstimulation with 10 nM E2 with respect to vehicle alone. Basal culture condition (10% FCS) was used as positive control. Values are mean � SDof three different experiments performed with three different cell lines in triplicate.Moggio. Sorafenib and ectopic endometrial MSC. Fertil Steril 2012.
ORIGINAL ARTICLE: ENDOMETRIOSIS
at the used doses (7.5 mM sorafenib: Ecto-MSC 0.37%� 0.4%;Euto-MSC 0.42% � 0.5%; Ctrl-MSC 0.17% � 0.1% AnnexinV/propidium iodide–positive cells).
1526
Sorafenib treatment induced a reduction of the increasedphosphorylation of ezrin observed in the Ecto-MSC, evalu-ated as phospho-ezrin/ezrin ratio (Fig. 4B). Moreover,
VOL. 98 NO. 6 / DECEMBER 2012

FIGURE 4
Effect of sorafenib on endometrial MSC. (A, B) Western blot micrograph and densitometric analysis of phospho-ezrin (P-Ezrin) and ezrin expressionin Ecto-MSC, Euto-MSC, and Ctrl-MSC. (A) Ecto-MSC showed higher P-ezrin levels. Analysis of variance with Dunnett's comparison test wasperformed. *P<.05, Ecto-MSC vs. Euto-MSC or Ctrl-MSC. (B) Treatment with 2.5–7.5 mM sorafenib reduced both P-ezrin/ezrin ratio andprotein expression levels. Data, shown as arbitrary units, are representative of three different experiments and were normalized to actinexpression. (C–E) Sorafenib treatment also inhibited the increased migration (C) and proliferation (D) of Ecto-MSC, as well as VEGF mRNAexpression (E), evaluated by quantitative real-time RT-PCR analysis and normalized to TBP mRNA. (C, D) Data are mean � SD of three differentexperiments performed with three different cell lines in triplicate. (E) Data are mean of three different cell lines normalized to 1 for vehicle inEcto-MSC. (F) Western blot micrograph and densitometric analysis showing the reduction of HIF-1a expression in Ecto-MSC treated with 5 mMsorafenib. Data are shown as arbitrary units and are representative of two different experiments performed on different cell lines and werenormalized to actin expression. (B–F) Student's t test was performed: *P<.05, sorafenib vs. vehicle.Moggio. Sorafenib and ectopic endometrial MSC. Fertil Steril 2012.
Fertility and Sterility®
Sorafenib decreased both the basic and activated forms of ez-rin in all endometrial MSC lines. In particular, Euto-MSC andCtrl-MSC showed ezrin reductionwhen treatedwith a concen-tration of 5 mM sorafenib, whereas Euto-MSCwere responsive
VOL. 98 NO. 6 / DECEMBER 2012
only at 7.5 mM sorafenib (Fig. 4B). Because ezrin modulatescell migration, we evaluated whether sorafenib treatment ofthe Ecto-MSC could affect the observed increase in their mi-gratory properties. Sorafenib, at the concentration of 5 mM,
1527

ORIGINAL ARTICLE: ENDOMETRIOSIS
reduced cell migration. (Fig. 4C) but not cell invasion (datanot shown). In addition, sorafenib induced a dose-dependent decrease in the proliferation of Ecto-, Euto-, andCtrl-MSC, with a prominent effect on the observed increasedproliferation of Ecto-MSC (Fig. 4D). No significant effect ofsorafenib (5 mM) was observed on the proliferation of bonemarrow–derived MSC (vehicle: 0.63 � 0.11; sorafenib 0.67� 0.03 o.d.; n ¼ 3). Finally, treatment with sorafenib (5mM) induced a significant reduction of HIF-1a and VEGF ex-pression in Ecto-MSC (Fig. 4E and F). These data indicate thatsorafenib is effective on MSC lines and may act in particularby reducing the increasedmigration, proliferation, and angio-genic properties of MSC from ectopic lesions.
DISCUSSIONIn the present study, we isolated stem cell lines from ectopicand eutopic endometrium of women complaining of endome-triosis, as well as from the normal endometrium of controlsubjects without endometriosis. These cells showed similarmesenchymal phenotype but different functional properties.In particular, Ecto-MSC from both peritoneal lesions andovarian cyst displayed increased proliferation and invasioncapacity in comparison with Ctrl-MSC and Euto-MSC derivedfrom the same patient. Moreover, Ecto-MSC showed an in-creased VEGF release and HIF-1a expression. The antiangio-genic drug sorafenib was able to reduce the increasedproliferation and motility of Ecto-MSC, possibly actingthrough the inhibition of the Raf-dependent cytoskeletal pro-tein ezrin, shown to be overexpressed in Ecto-MSC, and re-duced the observed increased VEGF and HIF-1a expression.
A stem cell population with mesenchymal characteristicswas previously reported to reside in the human endometriumas well as in ectopic endometrial implants (8–17, 19). Herein,we isolated MSC from both eutopic and ectopic endometriumof the same patient, and we used the normal endometrium ofhealthy women as control. The phenotypic characterizationshowed a comparable mesenchymal phenotype of MSCfrom control, eutopic, and ectopic endometrial tissue; infact, they expressed the mesenchymal surface markersCD105, CD44, CD73, the stromal cytoplasmic markervimentin, and showed osteogenic and epithelialdifferentiative ability, as described for bone marrow–derived MSC (31). Moreover, they expressed the stemembryonic marker SSEA-4 (31) and the stem transcriptionfactors (32) c-Myc and Klf-4 but not Oct4-A. This result isin contrast with the identified expression of Oct4 in endome-trial MSC cells (33). However, this discrepancy can be ex-plained by the possible detection in previous studies ofdifferent Oct4 isoforms that can be generated by alternativesplicing. Because only the Oct-4A isoform is considered tobe stem cell specific, we restricted our analysis to this isoform(34, 35). Indeed, MSC from bone marrow are known to lackOct-4A (27, 36). Interestingly, MSC from ectopicendometriosis lesions displayed altered functionalproperties, such as increased proliferation and migration,with respect not only to MSC obtained from theendometrium of normal subjects, but also to MSC from theeutopic endometrium derived from the same patient. These
1528
data are in accordance with the reported increase inproliferation and matrix invasion of stromal endometrialcells in endometriosis (19, 37). In addition, we found anincreased proangiogenic phenotype of MSC derived fromectopic endometriosis implants: they expressed in basalculture conditions HIF-1a, known to transcriptionallyactivate proangiogenic cell signals, including VEGF. Indeed,VEGF was present at high levels in the supernatant of MSCfrom ectopic lesions, whereas it was detectable in that ofMSC from normal and eutopic endometrium. Thisobservation parallels the increased angiogenesis found inendometrial ectopic implants (21, 38) and supports thenotion that MSC may alter their phenotype within theectopic environment.
Different hypotheses could explain the altered phenotypedetected in ectopic MSC with respect to eutopic or controlMSC. Mesenchymal stem cells, once located in the microenvi-ronment of the ectopic lesion, may undergo a selection pro-cess that could lead to the survival of the MSC cells/cloneswith enhanced migratory, proliferative, and angiogenic prop-erties. Alternatively, it can be postulated that the extrauterinemicroenvironment found in ectopic sites might epigeneticallymodulate the endometrial MSC, thus changing their charac-teristics. Indeed, the importance of epigenetic changesinduced by the microenvironment is gaining great consider-ation in the pathogenesis of cancer and several other humandiseases (39). Accordingly, various epigenetic aberrationshave been described in endometriosis (40).
The endometrial MSC population is supposed to be in-volved in the growth and maintenance of the endometrial le-sions (18). As a consequence, as postulated for tumors (41),the eradication of the disease cannot be achieved without tar-geting ectopic MSC with specific treatments. Because wefound that endometrial MSC do not express ER, nor are sen-sitive to estrogen treatment, the current use of estrogen deple-tion therapy in endometriosis (42) is likely to spare MSC andtarget only ER-positive differentiated endometrial cells, rep-resenting the majority of the lesion. This could explain whytreatments inducing a prolonged hypoestrogenic status (e.g.,GnRH analogues) are able to reduce symptoms of endometri-osis but are rarely effective in eradicating the disease, theirsuspension being often followed by endometriosis recurrenceafter months.
As a possible pharmacologic agent for endometrial MSC,we investigated the effect of sorafenib, an antiangiogenicdrug currently used in tumor therapy that targets the Raf/MEK/ERK pathway and receptor tyrosine kinases involvedin tumor progression and angiogenesis (29). EndometrialMSC indeed expressed CD140, a known target of sorafenib(29). In addition, MSC overexpressed the activated form of ez-rin, another possible target of sorafenib, owing to its inhibi-tion of Raf-1/ROCKII/ezrin signaling pathway (27). Ezrinwas reported to play a major role in the regulation of cell mor-phology, migration, and attachment, and its basal and acti-vated forms were shown to be overexpressed in tissues aswell as in isolated MSC from ectopic endometrial implants(30, 43).
We observed that sorafenib treatment inhibited the in-creased phosphorylation of ezrin in ectopic MSC, and
VOL. 98 NO. 6 / DECEMBER 2012

Fertility and Sterility®
consequently it was also effective in limiting the increasedmigration of ectopic MSC. Moreover, it reduced the increasedproliferation and VEGF release found in ectopic MSC. The in-hibitory effect of sorafenib on ectopic MSC proliferation andthe VEGF release may be explained by the effect on a tyrosinekinases target of sorafenib, such as CD140. This mechanismwas reported for tumor cell lines, where the inhibition of re-ceptor signaling pathways reduced the cytokine-induced pro-liferation and HIF-1a transcriptional activity (44). Activationof HIF-1amay also be induced in normoxic conditions by cy-tokines (45). The possible response of MSC to sorafenib couldbe influenced by the individual phenotype of endometrialMSC and by fluctuation of receptor expression. This pointcould deserve further investigation.
Our data indicate that sorafenib could exert a multileveleffect on mesenchymal cells, limiting their attachment,growth, invasion, and maintenance in endometriosis. Sorafe-nib treatment seemed to be able to normalize ectopic MSC,conferring to them a phenotype similar to the one observedin normal and eutopic endometrium.
In conclusion, our results demonstrate that the increasedproliferative, migratory, and angiogenic phenotype present inectopic MSC cells may be reverted by the treatment with themulti-tyrosine kinase inhibitor sorafenib. Targeting thestem cell population that sustains the endometriosis ectopiclesions may be relevant in achieving the complete eradicationof endometriotic implants.
REFERENCES1. Bulun SE. Endometriosis. N Engl J Med 2009;360:268–79.2. Giudice LC. Clinical practice. Endometriosis. N Engl J Med 2010;362:
2389–98.3. Sampson JA. Peritoneal endometriosis due to menstrual dissemination of
endometrial tissue into the peritoneal cavity. Am J Obstet Gynecol 1927;14:422–69.
4. D'Hooghe TM, Debrock S. Endometriosis, retrograde menstruation andperitoneal inflammation in women and in baboons. Hum Reprod Update2002;8:84–8.
5. Stefansson H, Geirsson RT, Steinthorsdottir V, Jonsson H, Manolescu A,Kong A, et al. Genetic factors contribute to the risk of developing endome-triosis. Hum Reprod 2002;17:555–9.
6. Christodoulakos G, Augoulea A, Lambrinoudaki I, Sioulas V, Creatsas G.Pathogenesis of endometriosis: the role of defective ‘‘immunosorveillance’’.Eur J Contracept Reprod Health Care 2007;12:194–202.
7. Vigan�o P, Somigliana E, Vignali M, Busacca M, Blasio AM. Genetics ofendometriosis: current status and prospects. Front Biosci 2007;12:3247–55.
8. Chan RW, Schwab KE, Gargett CE. Clonogenicity of human endometrial ep-ithelial and stromal cells. Biol Reprod 2004;70:1738–50.
9. Chan RW, Gargett CE. Identification of label-retaining cells in mouse endo-metrium. Stem Cells 2006;24:1529–38.
10. Schwab KE, Chan RW, Gargett CE. Putative stem cell activity of human en-dometrial epithelial and stromal cells during the menstrual cycle. Fertil Steril2005;84:1124–30.
11. Cervell�o I, Martínez-Conejero JA, Horcajadas JA, Pellicer A, Sim�on C. Identi-fication, characterization and co-localization of label-retaining cell popula-tion in mouse endometrium with typical undifferentiated markers. HumReprod 2007;22:45–51.
12. Kato K, YoshimotoM, Kato K, Adachi S, Yamayoshi A, Arima T, Asanoma K,et al. Characterization of side-population cells in human normal endome-trium. Hum Reprod 2007;22:1214–23.
VOL. 98 NO. 6 / DECEMBER 2012
13. Dimitrov R, Timeva T, Kyurkchiev D, Stamenova M, Shterev A, Kostova P,Zlatkov V, et al. Characterization of clonogenic stromal cells isolated fromhuman endometrium. Reproduction 2008;135:551–8.
14. Gargett CE, Schwab KE, Zillwood RM, Nguyen HP, Wu D. Isolation and cul-ture of epithelial progenitors and mesenchymal stem cells from human en-dometrium. Biol Reprod 2009;80:1136–45.
15. Masuda H, Matsuzaki Y, Hiratsu E, OnoM, Nagashima T, Kajitani T, Arase T,et al. Stem cell-like properties of the endometrial side population: implica-tion in endometrial regeneration. PLoS One 2010;5:e10387.
16. Cervell�o I, Gil-Sanchis C, Mas A, Delgado-Rosas F, Martínez-Conejero JA,Gal�an A, et al. Human endometrial side population cells exhibit genotypic,phenotypic and functional features of somatic stem cells. PLoS One 2010;5:e10964.
17. Cervell�o I, Mas A, Gil-Sanchis C, Peris L, Faus A, Saunders PT, Martìnez-Romero A, et al. Reconstruction of endometrium from human endometrialside population cell lines. PLoS One 2011;6:e21221.
18. Oliveira FR, Dela Cruz C, Del Puerto HL, Vilamil QT, Reis FM, Camargos AF.Stem cells: are they the answer to the puzzling etiology of endometriosis?Histol Histopathol 2012;27:23–9.
19. Kao AP, Wang KH, Chang CC, Lee JN, Long CY, Chen HS, Tsai CF, et al.Comparative study of human eutopic and ectopic endometrial mesenchy-mal stem cells and the development of an in vivo endometriotic invasionmodel. Fertil Steril 2011;95:1308–15.
20. Laschke MW, Menger MD. In vitro and in vivo approaches to study angio-genesis in the pathophysiology and therapy of endometriosis. Hum ReprodUpdate 2007;13:331–42.
21. Taylor RN, Yu J, Torres PB, Schickedanz AC, Park JK, Mueller MD, Sidell N.Mechanistic and therapeutic implications of angiogenesis in endometriosis.Reprod Sci 2009;16:140–6.
22. Hull ML, Charnock-Jones DS, Chan CL, Bruner-Tran KL, Osteen KG, Tom BD,et al. Antiangiogenic agents are effective inhibitors of endometriosis. J ClinEndocrinol Metab 2003;88:2889–99.
23. Van Langendonckt A, Donnez J, Defr�ere S, Dunselman GA, Groothuis PG.Antiangiogenic and vascular-disrupting agents in endometriosis: pitfallsand promises. Mol Hum Reprod 2008;14:259–68.
24. Iyer R, Fetterly G, Lugade A, Thanavala Y. Sorafenib: a clinical and pharma-cologic review. Expert Opin Pharmacother 2010;11:1943–55.
25. Bruno S, Bussolati B, Grange C, Collino F, di Cantogno LV, Herrera MB,Biancone L, et al. Isolation and characterization of resident mesenchymalstem cells in human glomeruli. Stem Cells Dev 2009;18:867–80.
26. Bussolati B, Moggio A, Collino F, Aghemo G, D'Armento G, Grange C, et al.Hypoxia modulates the undifferentiated phenotype of human renal innermedullary CD133þ progenitors through Oct4/miR-145 balance. Am J Phys-iol Renal Physiol 2012;302:116–28.
27. Pignochino Y, Grignani G, Cavalloni G, Motta M, Tapparo M, Bruno S, et al.Sorafenib blocks tumour growth, angiogenesis and metastatic potential inpreclinical models of osteosarcoma through a mechanism potentially involv-ing the inhibition of ERK1/2, MCL-1 and ezrin pathways. Mol Cancer 2009;8:118.
28. Goteri G, Lucarini G, Montik N, Zizzi A, Stramazzotti D, Fabris G, et al. Ex-pression of vascular endothelial growth factor (VEGF), hypoxia induciblefactor-1alpha (HIF-1alpha), and microvessel density in endometrial tissuein women with adenomyosis. Int J Gynecol Pathol 2009;28:157–63.
29. Wilhelm SM, Carter C, Tang L, Wilkie D, McNabola A, Rong H, et al. BAY43-9006 exhibits broad spectrum oral antitumor activity and targets theRAF/MEK/ERK pathway and receptor tyrosine kinases involved in tumorprogression and angiogenesis. Cancer Res 2004;64:7099–109.
30. Yotova I, Quan P, Gaba A, Leditznig N, Pateisky P, Kurz C, et al. Raf-1 levelsdetermine themigration rate of primary endometrial stromal cells of patientswith endometriosis. J Cell Mol Med 2012;16:2127–39.
31. Chamberlain G, Fox J, Ashton B, Middleton J. Concise review: mesenchymalstem cells: their phenotype, differentiation capacity, immunological fea-tures, and potential for homing. Stem Cells 2007;25:2739–49.
32. Chambers I, Tomlinson SR. The transcriptional foundation of pluripotency.Development 2009;136:2311–22.
33. Matthai C, Horvat R, Noe M, Nagele F, Radjabi A, van Trotsenburg M, et al.Oct-4 expression in human endometrium. Mol Hum Reprod 2006;12:7–10.
1529

ORIGINAL ARTICLE: ENDOMETRIOSIS
34. Wang X, Dai J. Concise review: isoforms of OCT4 contribute to the confus-ing diversity in stem cell biology. Stem Cells 2010;28:885–93.
35. Liedtke S, Stephan M, K€ogler G. Oct4 expression revisited: potential pitfallsfor datamisinterpretation in stemcell research. BiolChem2008;389:845–50.
36. Pierantozzi E, Gava B, Manini I, Roviello F, Marotta G. Pluripotency regula-tors in human mesenchymal stem cells: expression of NANOG but not ofOCT-4 and SOX-2. Stem Cells Dev 2011;20:915–23.
37. Klemmt PA, Carver JG, Koninckx P, McVeigh EJ, Mardon HJ. Endometrialcells from women with endometriosis have increased adhesion and prolifer-ative capacity in response to extracellular matrix components: towardsa mechanistic model for endometriosis progression. Hum Reprod 2007;22:3139–47.
38. McLaren J. Vascular endothelial growth factor and endometriotic angiogen-esis. Hum Reprod Update 2000;6:45–55.
39. Feinberg AP. Phenotypic plasticity and the epigenetics of human disease.Nature 2007;447:433–40.
1530
40. Guo SW. Epigenetics of endometriosis. Mol Hum Reprod 2009;15:587–607.41. Velasco-Velasquez MA, Popov VM, Lisanti MP, Pestell RG. The role of breast
cancer stem cells in metastasis and therapeutic implications. Am J Pathol2011;179:2–11.
42. Olive DL. Medical therapy of endometriosis. Semin Reprod Med 2003;21:209.
43. Ornek T, Fadiel A, Tan O, Naftolin F, Arici A. Regulation and activation of ez-rin protein in endometriosis. Hum Reprod 2008;23:2104–12.
44. Nilsson MB, Zage PE, Zeng L, Xu L, Cascone T, Wu HK, Saigal B,et al. Multiple receptor tyrosine kinases regulate HIF-1alpha andHIF-2alpha in normoxia and hypoxia in neuroblastoma: implicationsfor antiangiogenic mechanism of multikinase inhibitors. Oncogene2010;29:2938–49.
45. Kuschel A, Simon P, Tug S. Functional regulation of HIF-1a under nor-moxia—is there more than post-translational regulation? J Cell Physiol2012;227:514–24.
VOL. 98 NO. 6 / DECEMBER 2012

Fertility and Sterility®
0 0
SUPPLEMENTAL MATERIALS AND METHODSReal-Time Polymerase Chain Reaction AnalysisTotal RNA was extracted with Trizol reagent (Invitrogen) ac-cording to the manufacturer's instructions, and its qualitywas evaluated with Nanodrop 2000 (Thermo Scientific) and2100 Bioanalyzer (Agilent Technologies). Briefly, first-strand complementary DNA (cDNA) was produced from 200ng of total RNA, using the High Capacity cDNA Reverse Tran-scription Kit (Applied Biosystems). Real-time PCR experi-ments were performed in 20-mL reaction mixturecontaining 5 ng of cDNA template, the sequence-specific ol-igonucleotide primers purchased fromMWG-Biotech, and thePower SYBR Green PCR Master Mix (Applied Biosystems).Relative quantization of the products was performed usingthe 48-well StepOne Real-Time PCR System (Applied Biosys-tems). Thermal cycling conditions were as follows: activationof AmpliTaq Gold DNA Polymerase LD at 95�C for 10 min-utes, followed by 45 cycles of amplification at 95�C for 15seconds and 60�C for 1 minute, and a final incubation at95�C for 15 seconds. TATA binding protein (TBP) messengerRNAwas used to normalize RNA inputs. Fold change was cal-culated respect to control for all samples through the formula:
RQ ¼ 2�½ðDCtsampleÞ�ðDCtreferenceÞ�; where : DCt
¼ Ctspecific�primer � CtTBP:
Sequence-Specific Oligonucleotide Primers
Human Oct4A: forward, 50-AGC AGG AGT CGG GGT GG-30,and reverse, 50-CTG GGA CTC CTC CGGGTT-30; human KLF4:
VOL. 98 NO. 6 / DECEMBER 2012
forward, 5 - CCA TTA CCA AGA GCT CAT GCC-3 , and re-verse, 50-GGG CCA CGA TCG TCT TCC-30; human c-Myc: for-ward, 50-CAG CGA CTC TGA GGA GGA ACA-30, and reverse,50-TGA GGA GGT TTG CTG TGG C-30; human VEGF-A: for-ward, 50-TCA TCA CGA AGT GGT GAA GTT CA-30, and re-verse, 50-TCA GGG TAC TCC TGG AAG ATG TC-30; humanhypoxia-inducible factor (HIF): forward, 50-GTCGACACAGCCTGGATATGAA-30, and reverse, 50-CATATCATGATGAGTTTTGGTCAGATG-30; and human TBP: forward,50-TGT GCA CAG GAG CCA AGA GT-30 (nt 938-957 iso1),and reverse, 50-ATT TTC TTG CTG CCA GTC TGG-30 (nt988-968 iso1).
Flow Cytometric Analysis
The cells suspensions were incubated with antibodies for 30minutes a 4�C in 100 mL of phosphate-buffered saline withaddition of 0.1% bovine serum albumin (Sigma). The follow-ing monoclonal antibodies, all fluorescein isothiocyanate orphycoerythrin conjugated, were used: anti-CD117, -CD140,-CD45, -CD73, -CD29, -CD34, -CD14 (Becton Dickinson),-SSEA-4, vascular endothelial growth factor receptor(VEGF-R) 2 or 3 (R&D Systems), -CD44 (BioLegend), and-CD105 (Miltenyi Biotec). Fluorescein isothiocyanate or phy-coerythrin mouse nonimmune isotypic IgG (R&D Systems) areused as control. To evaluate sorafenib cytotoxicity, 10 � 106
cells were double-stained with fluorescein isothiocyanate–conjugated Annexin V and propidium iodide for 15 minutesat 20�C (Invitrogen) according to the manufacturer's instruc-tion. At each experimental point, 10,000 cells were analyzedon a FACScan (Becton Dickinson).
1530.e1

SUPPLEMENTAL TABLE 1
Characterization of the endometrial cell lines.
Patient no. Cell line Sample CD105 CD73 CD44 CD29 SSEA-4 CD34 CD45 CD14
1 Ecto-MSC1 Endometriotic ovarian cyst 82.1 � 5.7 92.4 � 7.9 99.8 � 6.8 23.3 � 2.6 64.8 � 6.3 1.14 � 0.5 1.33 � 0.3 0.95 � 0.1Euto-MSC1 Endometrium 97.3 � .3.9 99.4 � 2.4 99.7 � 3.3 45.5 � 3.1 20.4 � 2.6 1.32 � 0.2 0.95 � 0.2 2.08 � 0.6
2 Ecto-MSC2 Endometriotic peritoneal lesion 79.8 � 8.5 93.1 � 8.9 99.9 � 7.4 56.8 � 5.0 56.8 � 5.9 0.44 � 0.3 0.66 � 0.1 1.05 � 0.7Euto-MSC2 Endometrium 67.3 � 4.2 99.1 � 3.3 99.7 � 5.0 17.8 � 1.5 36.3 � 2.0 1.02 � 0.1 1.75 � 0.1 1.49 � 0.2
3 Ecto-MSC3 Endometriotic peritoneal lesion 99.3 � 8.5 99.7 � 2.3 99.8 � 2.7 29.4 � 3.4 30.7 � 1.6 1.02 � 0.3 0.66 � 0.1 0.25 � 0.1Euto-MSC3 Endometrium 98.1 � 3.4 86.8 � 5.9 99.8 � 3.1 3.4 � 0.2 11.5 � 1.3 0.50 � 0.2 1.47 � 0.1 1.76 � 0.5
4 Ecto-MSC4 Endometriotic peritoneal lesion 95.0 � 2.7 99.5 � 3.7 99.4 � 1.9 8.5 � 1.3 79.8 � 5.6 2.01 � 0.1 0.97 � 0.2 0.34 � 0.1Euto-MSC4 Endometrium 99.0 � 5.7 99.2 � 4.5 99.6 � 6.8 11.8 � 0.8 69.0 � 2.6 0.65 � 0.2 1.26 � 0.1 2.17 � 0.3
5 Ctrl-MSC1 Normal endometrium 99.9 � 1.3 99.9 � 0.7 100 � 0.9 30.4 � 0.5 49.9 � 2.3 1.34 � 0.1 0.48 � 0.5 1.47 � 0.56 Ctrl-MSC2 Normal endometrium 99.9 � 2.7 99.9 � 2.3 99.9 � 3.0 4.7 � 0.4 31.3 � 0.8 1.97 � 0.3 0.52 � 0.2 1.32 � 0.2Note: Endometrial MSC lines were generated from four patients with endometriosis (ectopic and eutopic lesions) and from two control patients. In the table, the tissue origin and the percentage of cells positive for mesenchymal markers are shown (data are mean� SD,n ¼ 3).
Moggio. Sorafenib and ectopic endometrial MSC. Fertil Steril 2012.
1530.e2
VOL.98
NO.6
/DEC
EMBER
2012
ORIG
INALARTIC
LE:ENDOMETRIO
SIS




















