
Social malleability in cowbirds: New measures reveal new evidence of plasticity in the eastern...
12
Journal of Comparative Psychology Copyright 1996 by the American Psychological Association, Inc. 1996, Vol. 110, No. 1, 15-26 0735-7036/96/$3.00 Social Malleability in Cowbirds: New Measures Reveal New Evidence of Plasticity in the Eastern Subspecies (Molothrus ater ater) Meredith J. West, Andrew P. King, and Todd M. Freeberg Indiana University Bloomington The social experiences of young Molothrus ater ater cowbirds were manipulated in a 2-year study. In the 1 st year, males were housed with pairs of canaries. The males were tested in 3 social contexts. Also, vocal repertoires were recorded and played back to females. In contrast to a previous study of the M. a. artemisiae subspecies, the males did not vocalize to the canaries in courtship tests (T. M. Freeberg, A. P. King, & M. J. West, 1995) but showed incompetent courtship of female cowbirds. In their 2nd year, half of the males were housed with older males and female cowbirds, and half were housed with only females. Those exposed to older males courted much more successfully than did those deprived of such experience. All males developed new repertoires, and song potencies did not correlate across years. The data reveal intraspecific variation in the ontogeny of mate recognition but intraspecific dependence on social learning to acquire courtship skills. In the experiments reported here, we manipulated juve- nile Molothrus ater ater cowbirds' opportunities for social and vocal interaction during their 1st and 2nd years. We bad three goals. First, we looked for intraspecific variation in the ontogeny of mate recognition in cowbirds. Second, we examined interrelationships between vocal capacity and vo- cal performance. Third, we investigated the role social experience with older conspecific males may play in con- solidating the courtship skills of younger males. Our interest in intraspecific variation in mate recognition derived from field and laboratory studies suggesting con- siderable ontogenetic variation among cowbird subspecies, especially in processes related to vocal communication (Dufty & McChrystal, 1992; Eastzer, 1988; Eastzer, King, & West, 1985; King & West, 1987, 1990; O'Loghlen & Rothstein, 1993; Rothstein, Yokel, & Fleischer, 1986, 1988). Most recently, we had found evidence to suggest that other social capacities, including species and mate recogni- tion, might also show variation by population. In a study of cowbirds captured in South Dakota (SD), we documented the development of atypical vocal and courtship patterns in juvenile males housed with canaries (Serinus canaria) or female cowbirds during their 1st year (Freeberg, King, & West, 1995). The males housed with canaries had failed to show recognition of appropriate mates, allocating a majority Meredith J. West, Departments of Biology and Psychology, Indiana University, Bloomington; Andrew P. King, Department of Psychology, Indiana University, Bloomington; Todd M. Freeberg, Department of Biology, Indiana University, Bloomington. This work was supported by grants from the National Science Foundation. We thank Eva Sanders for her help in collecting data. We thank the personnel of Custer State Park (South Dakota) and the Fall River Feedlots (South Dakota) for assistance in capturing birds under Federal Permit PRT 767881. Correspondence concerning this article should be addressed to Meredith J. West, Department of Psychology, Indiana University, Bloomington, Indiana 47405. Electronic mail may be sent via Internet to [email protected]. 15 of their vocal behavior during the breeding season to male and female canaries, even though female conspecifics were always present. The results were all the more surprising in that the males had been captured as juveniles with other cowbirds and thus were not naive. Three of the five males had also incorporated canary calls and trills into their vocal repertoires. The second group of SD males, which had been housed with female cowbirds, vocalized more toward con- specific females when tested within the confines of a flight cage but did not show sustained singing, chasing, or atten- tion to females when housed in a large aviary. Their court- ship success rate was no higher than that of the males that had been housed with canaries. In contrast, wild-caught, adult SD males showed selective and persistent courtship of females from their own population. These data suggested that mate recognition systems in cowbirds are open at the species level. Several lines of evidence suggested that males of the eastern subspecies, M. a. ater, would be less disrupted by the social manipulations used in Freeberg et al. (1995). First, we had raised eastern M. a. ater with canaries in several studies (e.g., West & King, 1986). Although they, like the SD males, had copied some canary sounds, the M. a. ater males had deleted these sounds before their songs crystallized. Second, field studies suggest that l st-year M. a. ater males are as successful in obtaining consortships as adult males (Dufty, 1982; Rothstein et al., 1986), whereas differential success between yearlings and adults had been reported for a western population of M. a. artemisiae (Yo- kel, 1989). Birds of the latter population also do not com- plete development of their vocal repertoires until their 2nd year (O'Loghlen & Rothstein, 1993). In previous studies of captive colonies of M. a. ater, we also had found no differ- ences in copulatory success or vocal repertoire size between yearlings and adults (West, King, & Eastzer, 1981a, 1981b; West, King, & Harrocks, 1983). Finally, attempts to hybrid- ize the two subspecies, using birds collected from the same capture site as in Freeberg et al. (1995) and in the present
Transcript of Social malleability in cowbirds: New measures reveal new evidence of plasticity in the eastern...

Journal of Comparative Psychology Copyright 1996 by the American Psychological Association, Inc. 1996, Vol. 110, No. 1, 15-26 0735-7036/96/$3.00
Social Malleability in Cowbirds: New Measures Reveal New Evidence of Plasticity in the Eastern Subspecies (Molothrus ater ater)
Mered i th J. West , A n d r e w P. King, and T o d d M. F reebe rg Indiana University Bloomington
The social experiences of young Molothrus ater ater cowbirds were manipulated in a 2-year study. In the 1 st year, males were housed with pairs of canaries. The males were tested in 3 social contexts. Also, vocal repertoires were recorded and played back to females. In contrast to a previous study of the M. a. artemisiae subspecies, the males did not vocalize to the canaries in courtship tests (T. M. Freeberg, A. P. King, & M. J. West, 1995) but showed incompetent courtship of female cowbirds. In their 2nd year, half of the males were housed with older males and female cowbirds, and half were housed with only females. Those exposed to older males courted much more successfully than did those deprived of such experience. All males developed new repertoires, and song potencies did not correlate across years. The data reveal intraspecific variation in the ontogeny of mate recognition but intraspecific dependence on social learning to acquire courtship skills.
In the experiments reported here, we manipulated juve- nile Molothrus ater ater cowbirds' opportunities for social and vocal interaction during their 1st and 2nd years. We bad three goals. First, we looked for intraspecific variation in the ontogeny of mate recognition in cowbirds. Second, we examined interrelationships between vocal capacity and vo- cal performance. Third, we investigated the role social experience with older conspecific males may play in con- solidating the courtship skills of younger males.
Our interest in intraspecific variation in mate recognition derived from field and laboratory studies suggesting con- siderable ontogenetic variation among cowbird subspecies, especially in processes related to vocal communication (Dufty & McChrystal, 1992; Eastzer, 1988; Eastzer, King, & West, 1985; King & West, 1987, 1990; O'Loghlen & Rothstein, 1993; Rothstein, Yokel, & Fleischer, 1986, 1988). Most recently, we had found evidence to suggest that other social capacities, including species and mate recogni- tion, might also show variation by population. In a study of cowbirds captured in South Dakota (SD), we documented the development of atypical vocal and courtship patterns in juvenile males housed with canaries (Serinus canaria) or female cowbirds during their 1 st year (Freeberg, King, & West, 1995). The males housed with canaries had failed to show recognition of appropriate mates, allocating a majority
Meredith J. West, Departments of Biology and Psychology, Indiana University, Bloomington; Andrew P. King, Department of Psychology, Indiana University, Bloomington; Todd M. Freeberg, Department of Biology, Indiana University, Bloomington.
This work was supported by grants from the National Science Foundation. We thank Eva Sanders for her help in collecting data. We thank the personnel of Custer State Park (South Dakota) and the Fall River Feedlots (South Dakota) for assistance in capturing birds under Federal Permit PRT 767881.
Correspondence concerning this article should be addressed to Meredith J. West, Department of Psychology, Indiana University, Bloomington, Indiana 47405. Electronic mail may be sent via Internet to [email protected].
15
of their vocal behavior during the breeding season to male and female canaries, even though female conspecifics were always present. The results were all the more surprising in that the males had been captured as juveniles with other cowbirds and thus were not naive. Three of the five males had also incorporated canary calls and trills into their vocal repertoires. The second group of SD males, which had been housed with female cowbirds, vocalized more toward con- specific females when tested within the confines of a flight cage but did not show sustained singing, chasing, or atten- tion to females when housed in a large aviary. Their court- ship success rate was no higher than that of the males that had been housed with canaries. In contrast, wild-caught, adult SD males showed selective and persistent courtship of females from their own population. These data suggested that mate recognition systems in cowbirds are open at the species level.
Several lines of evidence suggested that males of the eastern subspecies, M. a. ater, would be less disrupted by the social manipulations used in Freeberg et al. (1995). First, we had raised eastern M. a. ater with canaries in several studies (e.g., West & King, 1986). Although they, like the SD males, had copied some canary sounds, the M. a. ater males had deleted these sounds before their songs crystallized. Second, field studies suggest that l st-year M. a. ater males are as successful in obtaining consortships as adult males (Dufty, 1982; Rothstein et al., 1986), whereas differential success between yearlings and adults had been reported for a western population of M. a. artemisiae (Yo- kel, 1989). Birds of the latter population also do not com- plete development of their vocal repertoires until their 2nd year (O'Loghlen & Rothstein, 1993). In previous studies of captive colonies of M. a. ater, we also had found no differ- ences in copulatory success or vocal repertoire size between yearlings and adults (West, King, & Eastzer, 1981a, 1981b; West, King, & Harrocks, 1983). Finally, attempts to hybrid- ize the two subspecies, using birds collected from the same capture site as in Freeberg et al. (1995) and in the present

16 WEST, KING, AND FREEBERG
study, suggested reproductive incompatibili t ies between the two groups. M. a. ater females housed with M. a. artemisiae males produced 68% fewer viable eggs than did M. a. ater females housed with M. a. ater males (King & Freeberg, 1995). Although these latter data did not suggest which population would be more malleable, they pointed to phy- logenetic differences of sufficient magnitude to affect on- togenetic processes underlying mate recognition systems.
In the present study, we addressed these potential in- traspecific differences. We asked whether M. a. ater cow- birds, deprived of exposure to conspecifics during vocal development, would develop competent reproductive skills. Would they be successful in courting potential or actual mates? We focused on the role of heterospecific influence (as opposed to the influence of female cowbirds) because it had produced the more extreme outcome.
To test these questions regarding social capacities in M. a. ater birds, we conducted two experiments. In Experiment 1, we used a protocol similar to the one we had used with the SD population to determine whether juvenile Indiana (IN) cowbirds from the M. a. ater subspecies were socially malleable. We also played back the males ' vocalizations to females to examine relationships between song potency and song use. In the males ' 2nd year (Experiment 2), we ma- nipulated exposure to adult males to isolate further the role of social learning in the ontogeny of courtship skills.
E x p e r i m e n t 1
We studied juvenile male cowbirds collected in IN, near the center of the range for M. a. ater (Friedmann, Kiff, & Rothstein, 1977). We housed males individually with pairs of adult male and female canaries from late summer until the following breeding season. All of the juvenile males were captured while adult IN cowbirds were still in resi- dence, and thus the juveniles were not socially or acousti- cally naive. We tested the males in three phases designed to allow them to display courtship behaviors in increasingly more complex social settings. In Phase 1, we observed the males ' reactions to new canaries or female cowbirds within the confines of a flight cage. In Phase 2, we recorded patterns of courtship in a large indoor-outdoor aviary. In Phase 3, we also recorded courtship patterns in the aviary setting, but normally reared adult IN males were present as well.
M e t h o d
Juvenile males. Nine juvenile male cowbirds, determined by plumage and date of capture to be between 50 and 100 days of age, served as the canary-housed (CH) subjects. They were collected in July 1992 in Monroe County, IN, at the laboratory. They were housed individually with pairs of adult canaries (S. canaria). All resided in sound-attenuating chambers (West, King, Eastzer, & Staddon, 1979) until the following May. One male (banded GN; see later discussion) developed difficulty flying during Phase 1 and at the onset of Phase 2; we removed this bird from the experiment. None of the bird's data are reported for the 1st year. He did recover and was used in Experiment 2.
Other participants. One yearling and 11 IN adult males, along with 19 females, participated in Phase 1, 2, or 3. The females served as potential recipients of courtship in Phase 1, 2, or 3, and none were used in more than one phase. The males (hereafter IN normal males) served as potential competitors during Phase 3. We also recorded vocalizations from some of these males while they were in their home aviary, and we used their vocalizations in playback tests.
Twelve female cowbirds, originally captured in Orange County, North Carolina (NC), also served as potential recipients in Phase 2. All of the NC birds were housed in an indoor-outdoor home aviary (9.1 × 18.3 X 3.4 m) from March 15, 1990, to July 1993. We had not used them in the Phase 1 test because of time constraints. The birds were adults and had been in the laboratory since 1989. Twelve females, originally captured in Fall River and Custer counties (SD), also served as potential recipients in Phases 1 and 2. When not being used in this experiment, they resided in an indoor-outdoor aviary of the same dimensions described earlier.
Two canaries, 1 male and l female, were used in Phase 1. Each test aviary also contained 5 canaries at all times during Phases 2 and 3 to serve as potential recipients of social behavior. Alto- gether, 10 canaries were rotated through the aviaries. Five starlings (Sturnus vulgaris), 2 adults and 3 juveniles, also resided in the testing aviary throughout Phases 2 and 3. The canaries were rotated to avoid stress resulting from variable outdoor weather conditions. The starlings were not rotated because they were breeding in the aviary.
Playback subjects. Nine adult IN females, captured from the same Monroe County site, served as subjects for the playback of the yearling and adult IN males' vocalizations in May and June of 1993. The females had been housed together before testing in an aviary with IN males. The males were removed a month before playback testing. The females were transferred to sound-attenuat- ing chambers 3 weeks before playback testing. During the play- back tests, 8 females were housed in pairs and 1 was housed alone.
All birds used in the study were banded with colored leg rings to permit individual identification. Abbreviations of these leg bands are used to identify individuals in Tables 3-6 and elsewhere in the text (e.g., LN for a male banded with light blue-dark green bands). All of the subjects were maintained on a modified version of the Bronx Zoo diet for omnivorous birds and were given millet, canary seed, and vitamin-treated water daily.
Testing environments. The 9 juvenile males resided in the sound-attenuating chambers until May 11, 1993. On that date, the males (hereafter CH yearlings) were moved into two indoor flight cages (1.8 x 2.4 x 1.8 m), with 5 males in one cage and 4 in another. They remained so housed until May 25, 1993, when they were moved into either a testing or holding aviary. While in the cages, both groups could see and hear each other and canaries in adjoining cages.
The Phase 2 and Phase 3 testing aviaries were unfamiliar to all of the males. The testing aviaries' outdoor section measured 9.1 x 18.3 X 3.4 m, and the indoor section measured 9.1 x 3.0 × 2.7 m; the indoor-outdoor holding aviaries measured 3.1 X 10.9 x 3.4 m. When not being observed in a testing aviary during Phase 2 or 3, the yearling males were housed in a separate holding aviary. The IN normal males and the female conspecifics and canaries used in Phases 2 and 3 were returned to home aviaries when not being observed in a test aviary.
Phase 1: Responses to unfamiliar individuals. To observe the CH yearlings' behavior toward unfamiliar canaries or female cow- birds, we housed 2 canaries and 2 IN females in an indoor flight cage (1.8 x 2.4 x 1.8 m) for 10 days beginning on May 15, 1993.

SOCIAL MALLEABILITY IN COWBIRDS 17
None had been housed with the CH males. Twice daily, we introduced 1 of the CH males for 20 min, with each male partic- ipating in two sessions (on different days) in a random order. We videotaped the sessions; two coders also observed each session, noting each vocalization and whether it was directed to an indi- vidual. To direct a vocalization (song or whistle), a male had to move to within 0.3 m of a recipient and turn his head and body toward the recipient while vocalizing. All other vocalizations were scored as undirected. After all of the males had been exposed twice to IN females and canaries, they were exposed in an identical manner to 2 SD females and 2 new canaries (one trial for each male). Thus, each male participated in three sessions in Phase 1.
Phase 2: Behavior in the presence of other CH males. To assess the males' responsiveness to females and canaries while also in the presence of similarly experienced CH yearling males, we rotated the birds through a testing aviary beginning on May 26, 1993, and ending on June 15, 1993. At all times, we observed 6 of the yearling males in the testing aviary. At the onset of Phase 2, the aviary also contained 6 SD females, 6 IN females, 5 canaries, and 5 starlings. On June 7, 1993, 6 NC females were added. We removed a male and replaced him with another male after repeated singing and chasing of the same bird for 3 days (see later discus- sion) or after approximately 7 days. We removed females and replaced them with other females from the same respective pop- ulations if they had formed consortships (see later discussion). Each male was removed one or two times (3 were removed twice and 5 were removed once) in Phase 2; males averaged 12.7 days in the aviaries (range = 10 to 16 days).
Two observers recorded vocal behavior and copulations from 0645 to 0945 hr, 7 days a week, using 15-min units of focal sampling per male. To be scored as directing vocalizations on a given day to a class of birds (e.g., to IN or SD females), a male had to produce a minimum of two or more directed songs or whistles while facing and moving within 0.3 m of the individual and had to follow the vocalization with a chase if the recipient departed. We defined chasing as flying toward the female immediately after the female's departure. Males potentially could interact with more than one class per day. We calculated the number of days on which a male displayed directed vocal behavior to each of the classes of potential recipients (Table 1). We chose to report data by day to capture the male's persistence and to make comparisons with previously collected data (Freeberg et al., 1995). Thus, a male that vocalized 6 times to a female on one day and 8 times on the next day was scored as having directed social behavior on 2 days, whereas a male that vocalized 14 times on only one day directed social behavior on one day. In addition, we measured the males' latency in days to vocalize toward each class and their rate of vocalizing to males and females.
We also calculated the number of consortships for each male. A consort day was scored if a male sang at least 10 songs to females in a morning, of which at least one third were to a particular female. For a consortship to be established, 3 consecutive consort days with the same female had to occur. If a copulation occurred on the 2nd day of a developing consortship, we judged that the pair of birds formed a consortship. We scored a copulation if the female adopted a copulatory posture and the male mounted the female. Once a consortship was established, we removed either the male or the female from the aviary and replaced it with a bird of the same class (i.e., a male with another male or an IN female with another IN female). We also calculated the males' consort persistence level (CPL), which was the number of consort days divided by the number of days in the aviary (Eastzer et al., 1985).
Phase 3: Behavior in the presence of adult male cowbirds. To assess responsiveness when experienced males were also present, we used methods similar to those of Phase 2 except that the IN normal males were added: We included the yearling males, IN normal males, IN females, canaries, and starlings. No SD or NC females were present. Phase 3 began on June 16, 1993, and ended on July 2, 1993. At all times, the aviary contained 4 yearling males and 4 normal IN males, 8 IN females, 4 starlings, and 4 canaries. The CH males averaged 8.5 days in residence (range = 7 to 10 days). They were removed two or three times (4 were removed twice and 4 were removed three times). The normal IN males averaged 5.2 days (range = 3 to 7 days) in the testing aviary and were removed one or two times (9 were removed once, 2 were removed twice, and 1 remained for the duration). We used the same criterion for removing and replacing males as in Phase 2. We removed and replaced 6 of the 8 IN females on June 25, 1993, to stimulate responsiveness in the males being tested.
Recording and playback procedures. The males were recorded in their home chambers in early May when their vocalizations had crystallized. We used a Sony TCD-D10 PRO digital audiotape recorder for recording. We then dubbed the recordings chosen for playback to an Otari MX III half-track recorder at 15 in./s. We played back songs using the Otari recorder, a Urei 537 t/a-octave equalizer, and a Crown D75 power amplifier through JBL 2105 speakers located in each female's chamber. The sound pressure levels of the vocalizations were 85 -+ 2 dB (a weighted impulse reading at 0.8 m from the speaker as recorded by a B & K 2209 sound pressure meter), comparable to typical sound levels for vocalizing in this acoustic environment.
We chose vocalizations for the CH males on the basis of record- ing quality from 1,206 songs and whistles (average per male = 134, range = 102 to 239) for the CH males and 614 recordings of the normal IN males (average per male = 62, range = 7 to 189). For both groups of males, both songs and flight whistles were
Table 1 Mean Percentage of Days Vocalizing to Each Class of Recipients in Phases 2 and 3
Phase and subject
Recipient
SD and NC Canaries IN females females Males
M %Range M %Range M %Range M %Range
Phase 2: CH yearlings 6 0-20 21 0-50 14 0-54 40 14-94 Phase 3:
CH yearlings 0 0 13 0-43 0 44 0-89 IN normal males 0 0 78 0-100 53 0-100
Note. SD and NC females were not present in Phase 3. IN = Indiana; SD = South Dakota; NC = North Carolina; CH = canary housed.

| 8 WEST, KING, AND FREEBERG
included (Rothstein et al., 1988). The normal IN males were recorded in their home aviary in May 1993.
We played six vocalizations per day to the females, with one vocalization per trial, separated in time by 90 min. We played the vocalizations in varying orders each day, but each vocalization was presented an equal number of times at different times of day. We played back each vocalization eight times over the course of the experiment. We scored a positive response if a female adopted a copulatory posture within 1 s from onset of the sound (i.e., if the female arched her neck and back and separated the feathers around the cloacal area).
Resul~
Phase 1: Responses to unfamiliar individuals. Five of the 8 CH yearlings directed vocal behavior toward canaries, allocating a mean of 6% (range = 3% to 50%) of their vocalizations to them. All of the males vocalized to the IN and SD females, yielding means of 24% and 23%, respec- tively (combined range = 4% to 55%). The largest propor- tion of each of 8 males' vocalizations was scored as undi- rected, with a mean of 69% (range = 30% to 94%). A Friedman analysis of variance ranking mean proportions of vocalizations to canaries, vocalization to females (average response to IN and SD combined), or undirected vocaliza- tions was significant, X~r = 17.1, p < .001, indicating con- cordance among the males in their allocation of vocal behavior.
Phase 2: Responses to females in the presence of other males. Only 3 of the 8 CH males directed vocal behavior to canaries in the testing aviary (Table 1). The 8 males vocalized on most days to one another. Seven of the 8 vocalized to IN females, 6 vocalized to SD females, and 1 vocalized to NC females (Table 1). A Friedman analysis of variance showed significant agreement among the males in their allocation of vocal behavior toward recipients, X~ = 15.34, p < .01. The only significant difference between classes of recipients (after control for multiple comparisons; see Siegel & Castellan, 1988) involved vocalizing to other males versus vocalizing to canaries (T = 0, p < .008). None of the males vocalized to starlings.
Latency to vocalize to the various classes showed con- siderable variability, with CH males tending to vocalize first to other males and then to canaries or IN females (Table 2).
A Friedman analysis of variance failed to reveal a signifi- cant preference for any class, ~ = 5.2, ns).
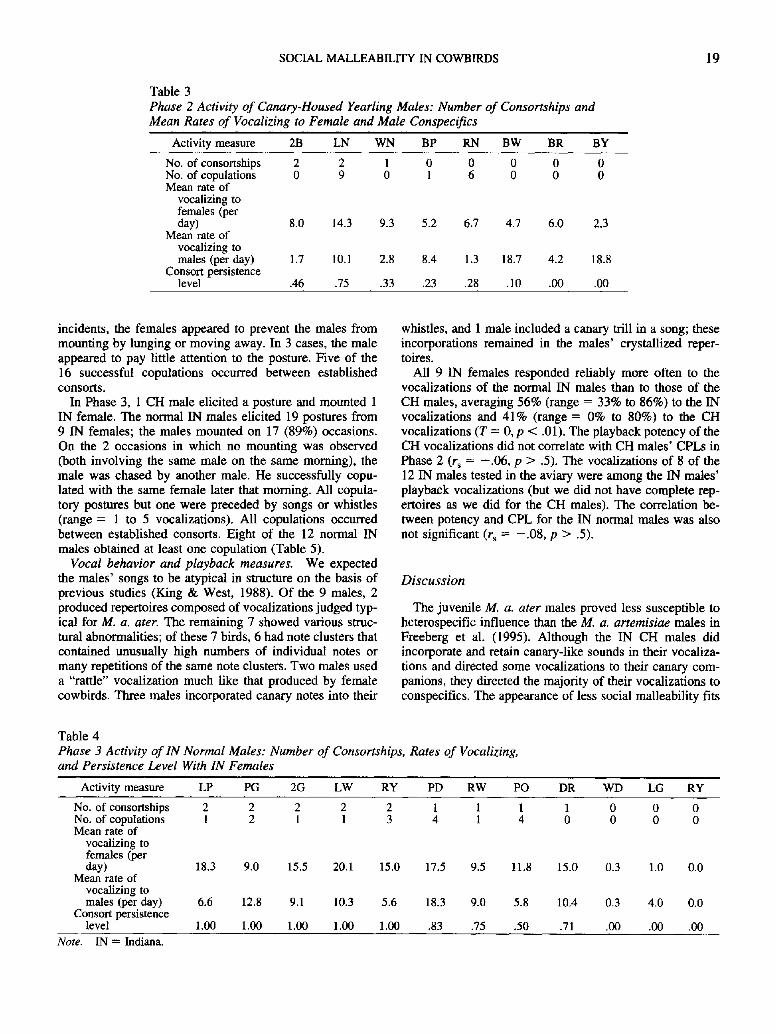
Three of the 8 CH males succeeded in forming consort- ships (Table 3). One of the males (2B) consorted with the same IN female in two different introductions as a result of an error in reintroducing a familiar female. Male LN con- sorted with 1 NC and 1 SD female. Male WN consorted with 1 SD female. Directing of vocalizations to females in Phase 1 was not correlated with the same measure in Phase 2 (r~ = - .34 , p > .50).
Phase 3: Responses in the presence of IN normal males. The wild-caught normal IN males commenced courtship activities sooner (Table 2) and were more selective and more successful (Tables 2 and 4). The IN yearling's (LP) courtship was comparable to that of the adults (Table 4). The IN normal males showed significant agreement in their allocation of vocal behavior, directing the most activity toward the IN females, X~ = 11.4, p < .001. The IN normal males' CPLs were high, averaging 86%, in comparison with a CPL of 35% for the CH males that consorted at all in Phase 2 and 22% for those that consorted in Phase 3. The difference in Phase 3 CPLs was significant (Mann-Whitney test; U -- 8, p < .001). A comparison of the CH males' Phase 2 CPL and the IN normal males' Phase 3 CPL also produced a significant difference (U = 14, p < .01). None of the CH males obtained consortships in Phase 3 (Table 5).
Copulatory performance. We did not use number of copulations as a primary measure of competence in Phase 2 because the technique of removing consort pairs after 3 days disrupts typical copulatory patterns (see Table 3 for the frequency of successful copulations in Phase 2). Moreover, because the SD and NC females did not have access to males from their respective populations, their copulatory behavior was difficult to interpret. We did look at copula- tory events, however, to explore any differences between CH and IN males in their reactions to copulatory postures. In Phase 2, 3 CH males successfully mounted females; a 4th CH male never did, although he elicited two postures; and the remaining 4 neither copulated nor elicited copulatory postures. Twenty-seven copulatory postures were observed in Phase 2. All were preceded by vocalizations (range = 1 to 4 songs or whistles). The males mounted females, how- ever, after only 16 (60%) of the postures. For 8 of these
Table 2 Mean Latency in Days for Males to Vocalize to Each Class of Recipient After First Introduction to Testing Aviary
Recipient
NC and SD Canaries IN females females Males
Latency Latency Latency Latency Phase and subject M range M range M range M range
Phase 2: CH yearlings 1.8 0-5 5.0 3-8 6.5 1-15 2.4 1 ~ Phase 3: IN normal 1.5 1-3 1.4 1-3
males Note. NC and SD females were not present in Phase 3. IN = Indiana; NC = North Carolina; SD = South Dakota; CH = canary housed.
SOCIAL MALLEABILITY IN COWBIRDS
Table 3 Phase 2 Activity of Canary-Housed Yearling Males: Number of Consortships and Mean Rates of Vocalizing to Female and Male Conspecifics
Activity measure 2B LN WN BP RN BW BR BY
No. of consortships 2 2 1 0 0 0 0 0 No. of copulations 0 9 0 1 6 0 0 0 Mean rate of
vocalizing to females (per day) 8.0 14.3 9.3 5.2 6.7 4.7 6.0 2.3
Mean rate of vocalizing to males (per day) 1.7 10.1 2.8 8.4 1.3 18.7 4.2 18.8
Consort persistence level .46 .75 .33 .23 .28 .10 .00 .00
19
incidents, the females appeared to prevent the males from mounting by lunging or moving away. In 3 cases, the male appeared to pay little attention to the posture. Five of the 16 successful copulations occurred between established consorts.
In Phase 3, 1 CH male elicited a posture and mounted 1 IN female. The normal IN males elicited 19 postures from 9 IN females; the males mounted on 17 (89%) occasions. On the 2 occasions in which no mounting was observed (both involving the same male on the same morning), the male was chased by another male. He successfully copu- lated with the same female later that morning. All copula- tory postures but one were preceded by songs or whistles (range = 1 to 5 vocalizations). All copulations occurred between established consorts. Eight of the 12 normal IN males obtained at least one copulation (Table 5).
Vocal behavior and playback measures. We expected the males' songs to be atypical in structure on the basis of previous studies (King & West, 1988). Of the 9 males, 2 produced repertoires composed of vocalizations judged typ- ical for M. a. ater. The remaining 7 showed various struc- tural abnormalities; of these 7 birds, 6 had note clusters that contained unusually high numbers of individual notes or many repetitions of the same note clusters. Two males used a "rattle" vocalization much like that produced by female cowbirds. Three males incorporated canary notes into their
whistles, and 1 male included a canary trill in a song; these incorporations remained in the males' crystallized reper- toires.
All 9 IN females responded reliably more often to the vocalizations of the normal IN males than to those of the CH males, averaging 56% (range = 33% to 86%) to the IN vocalizations and 41% (range = 0% to 80%) to the CH vocalizations (T = 0, p < .01). The playback potency of the CH vocalizations did not correlate with CH males' CPLs in Phase 2 (r s = - .06, p > .5). The vocalizations of 8 of the 12 IN males tested in the aviary were among the IN males' playback vocalizations (but we did not have complete rep- ertoires as we did for the CH males). The correlation be- tween potency and CPL for the IN normal males was also not significant (r s = - .08, p > .5).
Discussion
The juvenile M. a. ater males proved less susceptible to heterospecific influence than the M. a. artemisiae males in Freeberg et al. (1995). Although the IN CH males did incorporate and retain canary-like sounds in their vocaliza- tions and directed some vocalizations to their canary com- panions, they directed the majority of their vocalizations to conspecifics. The appearance of less social malleability fits
Table 4 Phase 3 Activity of IN Normal Males: Number of Consortships, Rates of Vocalizing, and Persistence Level With IN Females
Activity measure LP PG 2G LW RY PD RW PO DR WD LG RY
No. of consortships 2 2 2 2 2 1 1 1 1 0 0 0 No. of copulations 1 2 1 1 3 4 1 4 0 0 0 0 Mean rate of
vocalizing to females (per day) 18.3 9.0 15.5 20.1 15.0 17.5 9.5 11.8 15.0 0.3 1.0 0.0
Mean rate of vocalizing to males (per day) 6.6 12.8 9.1 10.3 5.6 18.3 9.0 5.8 10.4 0.3 4.0 0.0
Consort persistence level 1.00 1.00 1.00 1.00 1.00 .83 .75 .50 .71 .00 .00 .00
Note. IN = Indiana.

20 WEST, KING, AND FREEBERG
Table 5 Phase 3 Activity of Canary-Housed Yearling Males: Number of Consortships, Rates of Vocalizing, and Consort Persistence Level With Indiana Females
Activity measure 2B LN WN BP RN BW BR BY
No. of consortships 0 0 0 0 0 0 0 0 Mean rate of
vocalizing to females (per day) 8.1 10.1 1.2 9.6 0.1 4.4 1.4 1.0
Mean rate of vocalizing to males (per day) 1.7 6.5 0.2 17.1 0.3 17.9 5.6 14.4
Consort persistence level .14 .28 .00 .22 .00 .22 .00 .00
with other findings suggesting differences in ontogenetic patterns between populations of M. a. ater and M. a. arte- misiae (O'Loghlen & Rothstein, 1993).
The CH males directed approximately 25% of their vo- calizations to females when in the testing aviary. They thus courted slightly more than had the SD males housed with canaries; those CH males had directed an average of 19% (range = 0% to 100%) of their vocalizations to female cowbirds (Freeberg et al., 1995). Part of the difference was the greater allocation of behavior by the SD CH males to canaries; more than 50% of their days in the aviaries were spent directing vocalizations to canaries, in comparison with 6% for the IN CH males. Despite the greater opportu- nities, the consort persistence levels of the IN CH males were below 50% for all but 1 male, suggesting that these males used their time and vocalizations inefficiently with respect to attracting potential mates. Furthermore, relative to the IN normal males' behavior in the aviary, the high incidence of failures to mount females in copulatory pos- tures suggested that the IN CH males' attentiveness to female signals was deficient.
Although we measured courtship from the perspective of the male, females' behavior contributes to a male's success. Females must remain still for males to direct vocalizations to them and must permit the male to vocalize from very short distances for his signal to have maximal impact (King, West, Eastzer, & Staddon, 1981). Thus, part of the males' difficulties here may have been that the females, given less than optimal choices of males, displayed less stable respon- siveness. Of the five consortships achieved by the CH males, only two were with IN females, and no copulations occurred. As noted earlier, the females often lunged at the males even after adopting copulatory postures, a response we did not see with more experienced males, although it does rarely occur.
The vocalizations of the CH males were less effective than the songs of normal males at eliciting copulatory postures from the playback females. This finding fits with the aviary observations but may appear to conflict with previously published data from this laboratory suggesting that acoustically naive male cowbirds produce potent sig- nals (Freeberg et al., 1995; King & West, 1977; West et al., 1979). Some of those earlier males had, however, been
housed with female cowbirds, a social condition we now know has direct effects on song potency (Freeberg et al., 1995; King & West, 1983; West & King, 1985, 1988a). Moreover, in the original studies, we compared the songs of the naive males with only a small set of songs from normal males. We now know that a normal male's repertoire may contain songs varying widely in potency, making the choice of playback songs especially important (Freeberg et al., in press; West, King, & Freeberg, 1994). The data reported in the present study derive from males raised without female conspecifics and included in playback tests of more normal males, thus accounting for some of the differences.
Earlier, we had speculated that the parasitic reproductive behavior of cowbirds might have led to selection for female responsiveness to songs developed under a wide range of rearing conditions for males and females (King & West, 1977). Playback tests of vocal precursors to stereotyped song also suggested that cowbird songs were different than those in nonparasitic species. Even plastic song elicited copulatory responding from captive male-deprived females, suggesting that males need only minimal acoustic experi- ence to shape their songs into a form that is above a female cowbird's perceptual threshold (West & King, 1988b). All of these findings suggested a communicative safety net for male cowbirds' exploitation of females' reflexive respon- siveness to vocal or social stimulation. The present data, and those of Freeberg et al. (1995), expose a serious weakness in such reasoning: Males do not appear to use their song reflexively to interact with females. As such, the idea of a safety net for mate attraction becomes untenable. The ab- sence of species-typical experience produces behavioral handicaps that would greatly reduce a male cowbird's op- portunity for reproductive success. The next experiment was designed to attempt to rehabilitate some of the skills necessary for successful courtship in the males.
Exper iment 2
We looked further at the effects of social experience in the second experiment, again generally modeling the design after that of Freeberg et al. (1995). Thus, at the end of the CH males' first breeding season, we randomly assigned the

SOCIAL MALLEABILITY IN COWBIRDS 21
males to two different social groups. Half of the males resided with IN female conspecifics and each other but were deprived of contact with experienced, adult male conspe- cifics (the deprived group). The other half also resided with females and each other but were provided with experienced, adult males as well (the experienced group). We had used these same conditions in the study of SD males with one exception: In that study, the social manipulation was con- founded by density, the 5 experienced males were housed with 13 other birds (5 adult males as well as 8 females), and the 5 deprived males were housed with only 4 additional females (Freeberg et al., 1995). Here we attempted to solve the problem by rotating in and out a number of different adult IN males in smaller numbers. Thus, at any given time, the experienced group was housed with only a few adult males, making densities across the two aviaries roughly comparable.
Experiment 2 also offered an opportunity to deal with a concern about the effects of changes in housing on social behavior. Most studies of songbirds whose acoustic expe- riences have been manipulated restrict measures of outcome to acoustic analyses of songs and playbacks to male or female conspecifics. The present data, those of Freeberg et al. (1995), and those of Will iams, Kilander, and Sotanski (1993) suggest that such measures may not be sufficient to capture developmental outcomes. Part of the problem in using more extended assessments is that the species ' breed- ing biology often cannot be simulated, or the birds may be too stressed by the changes from the secluded conditions of deprivation housing to the more complex conditions of living in a captive flock to show courtship at all (but see Wil l iams et al., 1993). Although the CH males studied in Experiment 1 displayed clear inadequacies in their courtship behavior, we are inclined to think that many of the defi- ciencies reflected inadequacies in their social learning his- tories rather than reactions to the changed housing condi- tions. First, all of the males resumed singing within 1 day of being placed in the flight cages. Second, all sang during the cage tests. Third, they showed typical patterns of social flocking when foraging, both in the cages and then in the aviary. Fourth, the frequency with which they vocalized in the aviaries was not substantially different from that of normal males. What differed was the recipient of their vocalizations and how often they directed these vocaliza- tions. In the second experiment, however, all males lived in indoor -ou tdoor aviaries quite similar to those in which they were tested. Because their housing was comparable, any group differences could not be attributed to stress. We followed the birds through their second breeding season, recording the same measures of courtship. We predicted that experienced males, by virtue of their exposure to older, adult males, would perform better than the deprived males in courtship tests.
M e ~ o d
Yearling males. The 9 males used in Experiment 1, now com- ing into their 2nd year, served as subjects. These birds were randomly divided into two experimental groups on September 6,
1993. Both groups were housed with sets of IN female cowbirds, but the experienced group of 4 yearlings was provided with social contact with older, adult male cowbirds, whereas the deprived group consisted only of 5 of the yearlings and female cowbirds.
Adult males. Nine different adult IN males were housed, for varying lengths of time, with the experienced group from Septem- ber 6, 1993, through May 15, 1994. All of these adult males had been captured at the same IN site in the previous year. While in the laboratory before their use in the present study, they were housed in large indoor-outdoor aviaries with other male and female cowbirds. The adult males were cycled through the winter housing aviary (see later discussion) containing the experienced group such that, on average, there were 3.2 adult males (range = 2 to 5) per day. Each adult male spent an average of 90.2 days (range = 19 to 155) in the winter housing aviary with the experienced group.
Females. Eleven different IN females were housed with the experienced group, and 10 different IN females were housed with the deprived yearlings, for varying lengths of time, from Septem- ber 6, 1993, through May 15, 1994. All of the females had been captured at the same IN site on March 10, 1993, or from March to April 1994. Females in the laboratory before their use in the present study were housed in large indoor-outdoor aviaries with other male and female cowbirds. Whereas adult males were rotated in and out of the experienced aviary, new females were added only when resident females died or were moved to be used in other studies. Altogether, females housed with the experienced group spent an average of 136.3 days (range = 42 to 251) in the winter housing aviary; an average of 6.0 (range = 5 to 9) females were present per day. Females housed with deprived males spent an average of 99.8 days (range = 52 to 251) in the winter housing aviary, with an average of 4.0 (range = 2 to 6) females present per day. These manipulations produced a sex ratio of roughly 5 males to 4 females for the deprived group and approximately 7 males to 6 females for the experienced group (corresponding to a 1.2:1 male:female ratio in each group).
Playback subjects. Eight IN females served as playback sub- jects; all had served as playback subjects in the previous year. Between breeding seasons, they were housed in aviaries and moved to the sound-attenuating chambers 3 weeks before testing.
Aviary participants. During the breeding season aviary test, 20 different IN females served as recipients of the experienced and deprived males' vocal and courtship behavior. One of these fe- males had been housed over the winter with the experienced group; the other 19 females were unfamiliar with the males. The 19 females had been caught at the same IN sites in March or July 1993 or in April 1994 and had been housed since the time of their capture in aviaries with other cowbirds. Twelve different juncos (Junco hyemalis) were rotated through the testing aviary as part of another experiment. The testing aviary also contained 2 different male starlings (S. vulgaris).
Winter social housing. The experienced and deprived males, and the birds of their respective conditions, were housed in iden- tical indoor-outdoor aviaries (3.1 × 10.9 × 3.4 m). Birds in the two winter aviaries were visually isolated from one another. All other details of care were the same as in Experiment 1.
Breeding season testing and housing. The courtship effective- ness of experienced and deprived males was tested in a large indoor-outdoor aviary (9.1 × 18.3 × 3.4 m). When birds were not being tested in this aviary, they were kept in a small outdoor holding cage. All other details of care were the same as in Exper- iment 1.
Aviary test of experienced and deprived males' abilities to pair with conspecificfemales. From May 16-30 and June 5-15, 1994, we observed the courtship behaviors of experienced and deprived

22 WEST, KING, AND FREEBERG
males. At all times in the testing aviary, there were 6 males (typically 3 from each group but sometimes 2 and 4) and 6 IN females. From 0630 to 1030 every morning, we observed each male twice in two 15-min focal sampling sessions using the same measures described for Experiment 1.
Once a consortship was established, either the male or the female was removed and replaced with a different male or female. The 4 experienced males were introduced to the testing aviary an average of 3.5 times (range = 3 to 4), and each bird averaged 16.5 days (range = 13 to 21) in the aviary. The 5 deprived males were introduced an average of 3.4 times (range = 3 to 4), and each male averaged 16.2 days (range = 14 to 21) in the aviary.
Recording and playback procedures. We used the same pro- cedures and equipment as in Experiment 1. We obtained all vo- calizations of the experienced and deprived males in May 1994, immediately before the males' second breeding season, when the males were in their respective winter housing aviaries. When sufficient recordings were obtained from a male, the bird was temporarily removed in an attempt to obtain more recordings from the other males in the aviary. We tested two or three vocalizations from each of the males (a total of 20 songs and whistles). We obtained copulatory responsiveness measures using the same play- back procedures as in Experiment 1.
Results
The males from the experienced group were uniformly more successful than the deprived males at consorting and copulating with IN females (Table 6). The experienced males obtained a mean of 4.5 (range = 4 to 6) consortships, and the deprived males obtained a mean of 1 (range = 0 to 2). Altogether, 18 different females were involved in con- sortships; all were unfamiliar to the males before testing, and no male consorted with the same female twice. Each experienced male vocalized more to females than any of the deprived males. Singing to males did not differ by group. The CPLs for the two groups were nonoverlapping, with the experienced males averaging 73% and the deprived males averaging 18% (Table 6). We recorded 34 copulations in- volving experienced males and 2 involving deprived males. All copulations occurred between consorting males and females. Copulatory postures were preceded by one to five vocalizations, usually songs and whistles. In addition, fe-
males adopted copulatory postures on 13 occasions in which males did not mount; these cases involved 4 experienced and 3 deprived males. In 11 cases, other males interfered; in 2 cases, no obvious cause was identified.
The vocalizations of the deprived males were signifi- cantly more effective at eliciting copulatory postures from the playback females than were those of the experienced males (T = 1, p < .05). Seven of the 8 playback females responded more to deprived males' vocalizations. The de- prived males' average potency was 76% (range = 44% to 100%), and the experienced males' average potency was 68% (range = 42% to 95%).
Comparison with the 1st year. Both groups of males changed their vocal repertoires, and there were some dif- ferences between the groups in song type retention and song sharing. Song sharing was established by making zero crossings: plots of each song that were visually compared and sorted on a light table by one observer. Two additional observers then reviewed the comparisons. Agreement as to "shared" or "nonshared" status was 100%. To be shared, songs had to have the same number and pattern of note clusters and the same contour of whistle structures (see West & King, 1986, for more details on determining song types and sharing). The 4 experienced males produced 10 songs in their 1st year; 1 male retained both of his types into his 2nd year while adding a third. In the 2nd year, the experienced males possessed a total of 14 songs (range = 3 to 4). One of the males (RN) shared two song types with 2 different IN adults. Two song types were shared among 2 or more experienced yearlings. In their 1st year, the deprived males had produced 11 songs; in their 2nd year, they pro- duced 13 (range = 1 to 4), retaining 4 from the 1st year. Five song types were shared by 2 or more males, with one song type shared by 4 of the 5. Thus, the experienced males demonstrated less retention of song types than the deprived males (two song types as opposed to four song types), and song sharing was slightly higher among the deprived males than the experienced males (five song types as opposed to two song types shared by at least 2 males).
The 2nd-year males' success or lack thereof could not be predicted by measures of an individual's lst-year perfor-
Table 6 Aviary Activity of Experienced and Deprived Males: Number of Consortships, Rates of Vocalizing, and Consort Persistence Level With Indiana Females
Experienced group Deprived group
Activity measure RN WN BW LN BY 2B BR BP GN
No. of consortships 6 4 4 4 2 2 1 0 0 No. of copulations 13 5 13 3 1 0 1 0 0 Mean rate of
vocalizing to females (per day) 9.5 8.0 8.9 6.2 4.1 3.9 3.0 3.1 2.8
Mean rate of vocalizing to males (per day) 4.7 3.5 12.9 8.3 4.8 7.9 0.7 13.1 13.8
Consort persistence level .69 .69 .89 .65 .22 .31 .13 .14 .08

SOCIAL MALLEABILITY IN COWBIRDS 23
mance (Tables 3, 5, and 6). CPLs between years were not significantly correlated (r s = .32, p > .5). Rates of vocal- izing to females were also not correlated (r~ = . 14, p > .5). Vocal potencies, as tested by playback, were not correlated across years (r s = - .38, p > .5).
Discussion
The data indicate that social experience after an M. a. ater male's 1st year can have a significant effect in structuring social and vocal behavior quite similar to that found in the previous study of M. a. artemisiae males (Freeberg et al., 1995). The data from both studies suggest that increased competence is not simply a product of age in either subspe- cies. Both experienced and deprived males had access to females and to one another, but the males provided with opportunities to observe and interact in the presence of older males were clearly more successful in the breeding season. In that the older males were not present during testing, their impact presumably derived from accumulated social expe- riences over the fall, winter, and spring. In the present study, sex ratio and density were better controlled, increasing the possibility that the difference originated from specific ex- periences with the older males. We now have to study this time period in detail to determine the nature of the social interactions involved. Aggression between yearling and older males has been suggested as a possible mechanism influencing song sharing in laboratory and field settings (Casey & Baker, 1993; Payne, 1981; Payne & Payne, 1993). It may also be that adult-yearling experiences facilitate transmission of other song-related skills such as attentive- ness to females. Observations of the males in the present study during daily care and periodic recording revealed no obvious aggression between males, but chases between males and perch displacements are common in our captive aviaries, and males do challenge one another if a new male approaches another male's consort (West & King, 1980).
The social experiences of both groups during the 2nd year also influenced vocal development: All males turned over their repertoires and acquired new vocalizations. No signs of the males' lst-year experiences (i.e., housing with canar- ies) were evident in their 2nd year, a finding also in accord with the data from Freeberg et al. (1995). The males in the experienced group retained fewer songs from their 1st year and shared fewer songs with one another and with adults. Thus, vocal modification and improvisation may also be important mechanisms of social change.
A feature of male-male vocal interactions requiring fur- ther study is song copying. The most successful experienced male in the present study was the only male to copy the adult males, making two of his songs unique in the testing aviary (we found an effect similar to that found by Freeberg et al., 1995). The other 3 experienced males shared some songs with one another. Song sharing was higher among the deprived males, as was playback potency. Previous aviary observations and playback studies suggest that potent songs may release aggression and countersinging among males (West & King, 1980). Thus, the more potent songs (as
revealed by playback) may be advantageous only if males can compete physically with other males. Male cowbirds may have to defend song types. The high incidence of male-male singing may be a context in which males "sam- pie" each other's repertoires. Those males intimidated by such challenges may cease to compete or refrain from producing certain song types. The 3 IN adults in Experiment 1 that did not sing in Phase 3 had the highest song potencies (as tested by playback). Previous studies suggest that social responses by female cowbirds may help shape males' rep- ertoires (West & King, 1988a). Males' responses to one another may also guide repertoire development and the skills associated with using vocal signals.
General Discussion
The findings of both experiments prompt us to view vocal ontogeny (i.e., the process responsible for the transmission of species-typical acoustic patterns) in a new light. The data from the present study and Freeberg et al. (1995) suggest that the absence of adequate social skills can overwhelm advantages or disadvantages associated with song potency. Whereas much experimental effort has gone into determin- ing the role of social and vocal stimulation in vocal acqui- sition, far less effort has been expended to study the onto- genetic processes responsible for vocal performance. We believe that the findings reported here for cowbirds apply to many other songbirds as well. Song is one of the most conspicuous traits of male songbirds during the breeding season and thus represents a logical candidate in the study of mate attraction. However, male birds must perform in many other ways to be reproductively successful; singing may only create the opportunity to engage in further assess- ment by a potential mate. The challenge now is to learn how the varied facets of male competence become integrated.
The data from this study and that of Freeberg et al. (1995) suggest that female songbirds may perceive songs differently as a function of context. When song is the only cue, as in playback studies, females show considerable agreement with one another as to which songs are poten- tially most stimulating. But when vocal behaviors are only one of a hierarchy of cues, as in the aviary setting, other features such as attentiveness and persistence may become preferred indicators of male quality. Payne and Payne (1993) concluded that female assessment of male indigo buntings (Passerina cyanea) rests on multiple cues, includ- ing age, plumage, and song. Moreover, the way in which these characteristics are weighted may change from year to year and across ecologies. The findings of Williams et al. (1993) offer converging evidence of the dissociability of vocal and reproductive processes. In their study, juvenile zebra finches (Taeniopygia guttata) were housed with or without adult male conspecifics. Although members of the latter group displayed the expected abnormalities in song structure, they attracted mates (although not as frequently as more normally reared males). Their courtship was compe- tent, and two of the three males reared young. The untutored males had been housed with male siblings and thus had had

24 WEST, KING, AND FREEBERG
some social experience with conspecifics. In all songbirds studied to date, exposure to conspecific adults appears es- sential for males to develop normal song repertoires (Kroodsma & Baylis, 1982). The present data suggest that such exposure may also play a role in learning to use such repertoires and to acquire other requisite skills (see also Kroodsma, 1988; Spector, McKim, & Kroodsma, 1989). In Williams et al. (1993), the greater success of the more normally reared males was attributed to "slight advantages in all aspects of pair formation and breeding" (p. 701).
Thus, we suggest that a general conclusion from these studies of cowbirds, buntings, and zebra finches is that the ontogeny of song must be viewed as part of a broader process of social development. Identification of social skills requires different measures of outcome than those tradition- ally used to assess vocal development. What is needed are tests of actual courtship and mating, as opposed to acoustic measures or playback tests alone. Acoustic analyses can reveal much about the degree of accurate vocal transmission between generations. Playback techniques can identify pos- sible features by which the birds perceive and discriminate among vocal signals. Neither method, however, can reveal how the signals are embedded into the broader suite of behaviors necessary to connect attraction of females to copulation with females.
Although it would be ideal to be able to dissect these mechanisms in field settings, confinement is often neces- sary if the outcomes are to be witnessed (but see Payne & Payne, 1993, for a thorough field assessment). The results from the present study, however, indicate that such confined settings also need to be evaluated in terms of how they might influence outcomes. For example, the Phase l tests in flight cages suggested that all of the CH males were capable of directing songs to females. In Phase 2, however, many of these same males infrequently exercised this ability. More- over, there was no correlation between frequency of direct- ing vocalizations to IN females between Phases 1 and 2. We found a comparable difference in Freeberg et al. (1995). The flight cage limits the female's ability to meter here distance from males and may thus bias the results to suggest more competence in the males than actually exists.
The testing aviaries do afford the females more opportu- nities to avoid males, an option many seemed to have chosen, as evidenced by the low levels of consortship seen in Phase 2. The aviary setting also revealed more about the social dynamics of vocal production. The aviaries in which the birds were tested were large enough to permit vocaliza- tions to degrade over distances known to affect copulatory responding by captive M. a. a t e r females (King et al., 1981). For example, an increase in distance between the signal and perceiver from 0.3 to 3.0 m reduced potency by greater than 50%. The length of the testing aviary was 18.3 m. More- over, because the aviaries were outdoors, the males had to compete with other sounds in the environment, including other birds inside and outside the aviaries, to be heard by the females. As a result, for a male to "expose" the potency of his song, he had to vocalize very close to the female. Singing in the aviary in and of itself was not sufficient; the males had to approach a female and vocalize before she
departed. Female cowbirds rarely approach males (in an- other study, one of us [Todd M. Freeberg] witnessed three instances of SD females approaching SD males to within 0.5 m over 7 weeks of courtship; 33 females and 32 males were involved). Thus, the burden falls to the male to maintain proximity. When male cowbirds sing to one another (as opposed to exchanging flight whistles; see Dufty & Pugh, 1994), they do so in close proximity, typically within 0.5 m (Lowther, 1993). Thus, by housing males with canaries in their I st year or with females and other naive males in the 2nd year (the deprived group), we may have denied the juveniles and yearlings opportunities to observe and expe- rience the typical distances males use when directing vocalizations.
In that male cowbirds contribute no parental care, no defense of territories, and no assistance in finding nests to be parasitized, the question arises as to what qualities fe- male cowbirds use to evaluate potential mates. That females do, in fact, assess potential mates seems clear from (a) field and captive studies showing that females typically do not copulate unless courted for several days and (b) the absence of evidence of copulations in the field between unpaired birds in many populations (West et al., 1981b; Yokel, 1986). These life history attributes suggest that a male's social skills may be the only "resources" females can eval- uate. These skills would include vocal content, but they would also include how well the vocalizations were used, as determined by persistent vocalizing and attentiveness to females in the face of male-male competition (which, in turn, would include vocal interactions between males).
Female cowbirds have one of the highest ratios of sperm storage tubule length in any passerine species studied to date (Briskie & Montogomerie, 1993). This capacity means that males must guard females to ensure their paternity. Such attention to mates must be expended throughout the entire breeding season because females cycle and recycle through the stages of ovulation, fertilization, and egg laying (Jackson & Roby, 1992; Scott & Ankney, 1983). Because males outnumber females in many populations, competition is likely. Vocal and social plasticity during and beyond the 1st year may then be advantageous to allow males to tune their repertoires and their skills to suit local conditions, perhaps analogous to the advantages wild rats receive by using social learning to adjust diet preferences (Galef, in press).
Because cowbirds are brood parasites, it has always been tempting to suppose that their development differs from that of other passerines. Typically, the assumption has been made that learning must play less of a role to avoid costly errors (Alcock, 1984; King & West, 1977; Lehrman, 1974; Mayr, 1974; Todd & Miller, 1993). We originally proposed that male cowbirds possess innately effective songs and females an innate preference for such songs, providing "an independent system designed to insure identification during the most important context, the breeding season" (King & West, 1977, p. 1004). We were wrong. The linkage between male vocal production and female perception is subservient to learned behaviors that create functional couplings be- tween the two domains. As we have argued elsewhere,

SOCIAL MALLEABILITY IN COWBIRDS 25
understanding mate recognition systems in many songbirds now requires an increased emphasis on how behaviors, regardless of their origins, become connected. Such pro- cesses appear to vary geographically in several songbird taxa, reflecting the impact of ecological and phylogenetic variables on the dynamic assembly of mate recognition systems (Nelson, Marler, & Palleroni, in press; West & King, in press).
R e f e r e n c e s
Alcock, J. (1984). Animal behavior: An evolutionary approach (3rd ed.). Sunderland, MA: Sinauer.
Briskie, J.V., & Montogomerie, R. (1993). Patterns of sperm storage in relation to sperm competition in passerine birds. Condor, 95, 442-454.
Casey, R. M., & Baker, M. C. (1993). Aggression and song devel- opment in white-crowned sparrows. Condor, 95, 723-728.
Dufty, A.M., Jr., & McChrystal, R. (1992). Vocalizations and copulatory attempts in free-living brown-headed cowbirds. Jour- nal of Field Ornithology, 63, 16-25.
Dufty, A. M. J. (1982). Movements and activities of radio-tracked brown-headed cowbirds. Auk, 99, 319-327.
Dufty, A. M. J., & Pugh, J. K. (1994). Response of male brown- headed cowbirds to broadcast of complete or partial flight whis- tles. Auk, 111, 734-739.
Eastzer, D.H. (1988). Geographic variation in cowbird song. Chapel Hill: University of North Carolina.
Eastzer, D.H., King, A.P., & West, M.J. (1985). Patterns of courtship between cowbird subspecies: Evidence for positive assortment. Animal Behaviour, 33, 30-39.
Freeberg, T. M., King, A. P., & West, M.J. (1995). Social mal- leability in cowbirds (Molothrus ater artemisiae): Species and mate recognition in the first 2 years of life. Journal of Compar- ative Psychology, 109, 357-367.
Friedmann, H. K., Kiff, L. F., & Rothstein, S. I. (1977). A further contribution to knowledge of the host relations of the parasitic cowbirds. Smithsonian Contributions to Zoology, 235, 1-75.
Galef, B.G., Jr. (in press). Why behaviors that animals learn socially are locally adaptive, Animal Behaviour.
Jackson, N. H., & Roby, D. H. (1992). Fecundity and egg-laying patterns of captive yearling brown-headed cowbirds. Condor, 94, 585-589.
King, A.P., & Freeberg, T.M. (1995). [Egg laying in hybrid populations of cowbirds]. Unpublished raw data.
King, A.P., & West, M.J. (1977). Species identification in the North American cowbird: Appropriate responses to abnormal song. Science, 195, 1002-1004.
King, A. P., & West, M. J. (1983). Epigenesis of cowbird song: A joint endeavor of males and females. Nature, 305, 704-706.
King, A. P., & West, M. J. (1987). Different outcomes of synergy between song production and song perception in the same sub- species (Molothrus ater ater). Developmental Psychobiology, 20, 177-187.
King, A.P., & West, M. J. (1988). Searching for the functional origins of cowbird song in eastern brown-headed cowbirds (Mo- lothrus ater ater). Animal Behaviour, 36, 1575-1588.
King, A.P., & West, M.J. (1990). Variation in species-typical behavior: A contemporary theme for comparative psychology. In D. A. Dewsbury (Ed.), Contemporary issues in comparative psychology (pp. 331-339). Sunderland, MA: Sinauer.
King, A.P., & West, M.J., Eastzer, D. H., & Staddon, J. E. R. (1981). An experimental investigation of the bioacoustics of
cowbird song. Behavioral Ecology and Sociobiology, 9, 211- 217.
Kroodsma, D. E. (1988). Song types and their use: Developmental flexibility of the male blue-winged warbler. Ethnology, 79, 235-247.
Kroodsma, D.E., & Baylis, J.R. (1982). Appendix: A world survey of evidence for vocal learning in birds. In D.E. Kroodsma & E. H. Miller (Eds.), Acoustic communication in birds (pp, 311-337). New York: Academic Press.
Lehrman, D. S. (1974). Can psychiatrists use ethology? In N. F. White (Ed.), Ethology and psychiatry (pp. 187-196). Toronto: University of Toronto Press.
Lowther, P. E. (1993). Brown-headed cowbird (Molothrus ater). In A. Poole & F. Gill (Eds.), The birds of North America, No. 47 (pp. 1-24). Washington, DC: Academy of Natural Sciences.
Mayr, E. (1974). Behavioral programs and evolutionary strategies. American Scientist, 62, 650-659.
Nelson, D. A., Marler, P., & Palleroni, A. (in press). A compara- tive approach to vocal learning: Intraspecific variation in the learning process. Animal Behaviour.
O'Loghlen, A. L., & Rothstein, S. I. (1993). An extreme example of delayed vocal development: Song learning in a population of wild brown-headed cowbirds. Animal Behaviour, 46, 293-304.
Payne, R. B. (1981). Song learning and social interaction in indigo buntings. Animal Behaviour, 29, 688-697.
Payne, R. B., & Payne, L. L. (1993). Song copying and cultural transmission in indigo buntings. Animal Behaviour, 46, 1045- 1065.
Rothstein, S. I., Yokel, D. A., & Fleischer, R. C. (1986). Social dominance, mating, and spacing systems, female fecundity and vocal dialects in captive and free-ranging brown-headed cow- birds. Current Ornithology, 3, 127-185.
Rothstein, S.I., Yokel, D.A., & Fleischer, R.C. (1988). The agonistic and sexual functions of vocalizations of male brown- headed cowbirds (Molothrus ater). Animal Behaviour, 36, 73- 86.
Scott, D. M., & Ankney, C. D. (1983). The laying cycle of brown- headed cowbirds: Passerine chickens? Auk, 100, 583-592.
Siegel, S., & Castellan, N. J. J. (1988). Nonparametric statistics for the behavioral sciences (2nd ed.). New York: McGraw-Hill.
Spector, D. A., McKim, L. K., & Kroodsma, D. E. (1989). Yellow warblers are able to learn songs and situations in which to use them. Animal Behaviour, 38, 723-725.
Todd, P. M., & Miller, G. A. (1993). Parental guidance suggested: How parental imprinting evolves through sexual selection as an adaptive learning mechanism. Adaptive Behavior, 2, 5-47.
West, M.J., & King, A. P. (1980). Enriching cowbird song by social deprivation. Journal of Comparative and Physiological Psychology, 94, 263-270.
West, M.J., & King, A.P. (1985). Social guidance of vocal learning by female cowbirds: Validating its functional signifi- cance. Ethology, 70, 225-235.
West, M. J., & King, A. P. (1986). Song repertoire development in male cowbirds (Molothrus ater): Its relation to female assess- ment of song. Journal of Comparative Psychology, 100, 296- 303.
West, M. J., & King, A. P. (1988a). Female visual displays affect the development of male song in the cowbird. Nature, 334, 244 -246.
West, M.J., & King, A.P. (1988b). Vocalizations of juvenile cowbirds (Molothrus ater ater) evoke copulatory responses from females. Developmental Psychobiology, 21, 543-552.
West, M. J., & King, A. P. (in press). Eco-gen-actics: A systems approach to the ontogeny of avian communication. In D.E.

26 WEST, KING, AND FREEBERG
Kroodsma & E. H. Miller (Eds.), The evolution and ecology of avian acoustic communication. Ithaca, NY: Cornell University Press.
West, M. J., & King, A. P., & Eastzer, D. H. (1981a). The cow- bird: Reflections on development from an unlikely source. American Scientist, 69, 57-66.
West, M. J., & King, A. P., & Eastzer, D. H. (1981b). Validating the female bioassay of cowbird song: Relating differences in song potency to mating success. Animal Behaviour, 29, 490- 501.
West, M. J., & King, A.P., Eastzer, D. H., & Staddon, J. E. R. (1979). A bioassay of isolate cowbird song. Journal of Compar- ative and Physiological Psychology, 93, 124-133.
West, M. J., & King, A. P., & Freeberg, T. M. (1994). The nature and nurture of neophenotypes. In L. A. Real (Ed.), Behavioral mechanisms in evolutionary ecology (pp. 238-257). Chicago: University of Chicago Press.
West, M.J., & King, A. P., & Harrocks, T. H. (1983). Cultural transmission of cowbird song: Measuring its development and outcome. Journal of Comparative Psychology, 97, 327-337.
Williams, H., Kilander, K., & Sotanski, M. L. (1993). Untutored song, reproductive success and song learning. Animal Behav- iour, 45, 695-705.
Yokel, D.A. (1986). Monogamy and brood parasitism: An un- likely pair. Animal Behaviour, 34, 1348-1358.
Yokel, D. A. (1989). Intrasexual aggression and the mating behav- ior of brown-headed cowbirds: Their relation to population densities and sex ratios. Condor, 91, 43-51.
Received April 15, 1995 Revision received July 24, 1995
Accepted August 7, 1995
New Editors Appointed, 1997-2002
The Publications and Communications Board of the American Psychological Association announces the appointment of four new editors for 6-year terms beginning in 1997.
As of January 1, 1996, manuscripts should be directed as follows:
For the Journal of Consulting and Clinical Psychology, submit manuscripts to Philip C. Kendall, PhD, Department of Psychology, Weiss Hall, Temple University, Philadelphia, PA 19122.
For the Journal of Educational Psychology, submit manuscripts to Michael Pressley, PhD, Department of Educational Psychology and Statistics, State University of New York, Albany, NY 12222.
For the Interpersonal Relations and Group Processes section of the Journal of Personality and Social Psychology, submit manuscripts to Chester A. Insko, PhD, Incoming Editor JPSP--IRGP, Department of Psychology, CB #3270, Davie Hall, University of North Carolina, Chapel Hill, NC 27599-3270.
As of March 1, 1996, manuscripts should be directed as follows:
• For Psychological Bulletin, submit manuscripts to Nancy Eisenberg, PhD, Depart- ment of Psychology, Arizona State University, Tempe, AZ 85287.
Manuscript submission patterns make the precise date of completion of 1996 volumes uncertain. Current editors Larry E. Beutler, PhD; Joel R. Levin, PhD; and Norman Miller, PhD, respectively, will receive and consider manuscripts until December 31, 1995. Current editor Robert J. Sternberg, PhD, will receive and consider manuscripts until February 28, 1996. Should 1996 volumes be com- pleted before the dates noted, manuscripts will be redirected to the new editors for consideration in 1997 volumes.




















