
Environmentally cued parturition in a desert rattlesnake, Crotalus atrox

Snake Venomics of the Central American Rattlesnake Crotalus
simus and the South American Crotalus durissus Complex Points to
Neurotoxicity as an Adaptive Paedomorphic Trend along Crotalus
Dispersal in South America
Juan J. Calvete,*,† Libia Sanz,† Pedro Cid,† Pilar de la Torre,† Marietta Flores-Dıaz,‡
M. Cristina Dos Santos,§ Adolfo Borges,| Adolfo Bremo,⊥ Yamileth Angulo,‡ Bruno Lomonte,‡
Alberto Alape-Giron,‡,# and Jose Marıa Gutierrez‡
Instituto de Biomedicina de Valencia, CSIC, Jaume Roig 11, 46010 Valencia, Spain, Instituto Clodomiro Picado,Facultad de Microbiologıa, Universidad de Costa Rica, San Jose, Costa Rica, Instituto de Ciencias Biologicas,
Universidade Federal do Amazonas, Manaus, Brazil, Instituto de Medicina Experimental, Universidad Centralde Venezuela, Caracas, Venezuela, Departamento de Sanidad Animal, Programa de Ciencias, Universidad
Nacional Experimental Francisco de Miranda, Coro, Venezuela., and Centro de Investigaciones en EstructurasMicroscopicas, Universidad de Costa Rica
Received September 28, 2009
We report a comparative venomic and antivenomic characterization of the venoms of newborn andadult specimens of the Central American rattlesnake, Crotalus simus, and of the subspecies cumanensis,durissus, ruruima, and terrificus of South American Crotalus durissus. Neonate and adult C. simusshare about 50% of their venom proteome. The venom proteome of 6-week-old C. simus ispredominantly made of the neurotoxic heterodimeric phospholipase A2 (PLA2 crotoxin) (55.9%) andserine proteinases (36%), whereas snake venom Zn2+-metalloproteinases (SVMPs), exclusively of classPIII, represent only 2% of the total venom proteins. In marked contrast, venom from adult C. simuscomprises toxins from 7 protein families. A large proportion (71.7%) of these toxins are SVMPs, two-thirds of which belong to the PIII class. These toxin profiles correlate well with the overall biochemicaland pharmacological features of venoms from adult (hemorrhagic) and newborn (neurotoxic) C. simusspecimens. The venoms of the South American Crotalus subspecies belong to one of two distinctphenotypes. C. d. cumanensis exhibits high levels of SVMPs and low lethal potency (LD50), whereas C.d. subspecies terrificus, ruruima, and durissus have low SVMP activity and high neurotoxicity to mice.Their overall toxin compositions explain the outcome of envenomation by these species. Further, inall C. simus and C. durissus venoms, the concentration of neurotoxins (crotoxin and crotamine) is directlyrelated with lethal activity, whereas lethality and metalloproteinase activity show an inverse relationship.The similar venom toxin profiles of newborn C. simus and adult C. durissus terrificus, ruruima, anddurissus subspecies strongly suggests that the South American taxa have retained juvenile venomcharacteristics in the adult form (paedomorphism) along their North-South stepping-stone dispersal.The driving force behind paedomorphism is often competition or predation pressure. The increasedconcentration of the neurotoxins crotoxin and crotamine in South American rattlesnake venoms stronglyargues that the gain of neurotoxicity and lethal venom activities to mammals may have representedthe key axis along which overall venom toxicity has evolved during Crotalus durissus invasion of SouthAmerica. The paedomorphic trend is supported by a decreasing LNC (lethal neurotoxicity coefficient,defined as the ratio between the average LD50 of the venom and the crotoxin + crotamine concentration)along the North-South axis, coincident with the evolutionary dispersal pattern of the Neotropicalrattlesnakes. The indistinguisable immunoreactivity patterns of Costa Rican and Venezuelan polyvalentantivenoms toward C. simus and C. durissus venoms strongly suggest the possibility of using theseantivenoms indistinctly for the management of snakebites by adult C. simus and by certain C. d.cumanensis populations exhibiting a hemorrhagic venom phenotype. The antivenomic results alsoexplain why the antivenoms effectively neutralize the hemorrhagic activity of adult C. simus venomsbut does not protect against adult C. durissus sp. and newborn C. simus envenomations. Theidentification of evolutionary trends among tropical Crotalus, as reported here, may have an impact indefining the mixture of venoms for immunization to produce an effective pan-American anti-Crotalusantivenom.
528 Journal of Proteome Research 2010, 9, 528–544 10.1021/pr9008749 2010 American Chemical SocietyPublished on Web 10/28/2009

Keywords: snake venomics • Crotalus simus • Crotalus durissus • snake venom protein families •ontogenetic shift • crotoxin • paedomorphism • antivenomics • antibothropic and anticrotalic ABCantivenom • polyvalent (Crotalinae) ICP antivenom
IntroductionThe pitvipers comprise the subfamily Crotalinae of the family
Viperidae, found in Asia and the Americas. Pitvipers aredistinguished by the presence of a heat-sensing pit organlocated in an indentation of the upper jaw, between the eyeand the nostril on either side of the head. This infrared receptoris used to detect the distance and direction of warm-bloodedprey.1,2 Pitvipers represent the only viperids found in theAmericas. They appear to have dispersed into the New Worldas a single lineage from Asia during the late Oligocene or theearly Miocene, between 22-24 Mya,3 at a time when easternNorth America and Eurasia were widely separated across theAtlantic, whereas northeastern Asia and Alaska remainedconnected via the Bering land bridge. The colonization of theNew World was followed by rapid adaptative radiation in themiddle-late Miocene and early Pliocene (16-3.6 Mya) givingrise to most of the approximately 126 currently recognizedspecies included in the 12 genera of New World pitvipers.3,4
Genera Crotalus and Sistrurus diverged from their commonancestor with Agkistrodon5 and segregated into the monophyl-etic clade of the rattlesnakes6,7 which likely had its origin ∼ 20Mya in the Sierra Madre Occidental in the north-central portionof the Mexican Plateau.8-10 The rattlesnakes as a groupdispersed northward into North America and southward intoSouth America,11,12 and the genus Crotalus groups venomouspitvipers widely distributed discontinuously from southernCanada to northern Argentina.13-15 It comprises approximately70 species and subspecies (http://www.reptile-database.org) ofrattlesnakes. The scientific name Crotalus derives from theGreek, κFoτRλ�ν, meaning “castanet”. Indeed, except for a fewisland populations that have lost it, rattlesnakes are easilyrecognized by the presence of a “rattle” at the tip of the tail,used as a warning device when threatened.
The assemblage of forms currently known as the subspeciesof the Neotropical rattlesnake, Crotalus durissus, constitute aset of closely related parapatric forms, whose mutual relationsand mechanisms of speciation still remain largely elusive.Wuster and co-workers16 have traced the dispersal of C.durissus in South America, revealing a classical pattern ofstepwise colonization progressing from a northern center oforigin in Mexico to northern South America and across theAmazon Basin. The biogeographical data also suggested anancient basal cladogenesis in the Central American C. simusclade dated back to the late Miocene/early Pliocene (6.4-6.7Mya), and a relatively recent (1.7-1.1 Mya) basal SouthAmerican dispersal across a central trans-Amazonian corridorduring the middle Pleistocene (1.1-1.0 Mya) followed by latervicariance due to expanding rain forests, and dispersal intonortheastern Brazil.16,17
Identifying the molecular basis of adaptations in naturalpopulations is an important yet largely unrealized goal in
evolutionary biology.18-20 Such information is of broad sig-nificance because it addresses fundamental questions aboutthe connection between genotype and phenotype for fitness-related traits. Venoms represent critical trophic adaptations thatallowed advanced snakes to transition from a mechanical(constriction) to a chemical (venom) means of subduing anddigesting prey which are often larger than themselves, and assuch, venom proteins have multiple functions including im-mobilizing, paralyzing, killing, and digesting prey. Given thecentral role that diet has played in the adaptive radiation ofsnakes,21 venom represents a key factor in the diversificationof these animals. Here we used a venomics approach22-24 tocompare the relative abundance of toxin families in the venomsof the closely related congeneric species Crotalus simus, theCentral American rattlesnake, and the subspecies terrificus,cumanensis, ruruima and durissus of the South AmericanNeotropical rattlesnake, C. durissus.
Besides ecological and taxonomical implications, knowledgeof the natural history and toxin composition of venoms is offundamental importance for the treatment of bite victims andin the selection of specimens for the preparation of venompools for antivenom production.24 Although considered non-aggressive, ophidian accidents caused by Central and SouthAmerican Crotalus species constitute a serious medical problemin many parts of their range.25 Envenoming by adult C. simusin Central America produce clinical effects reminiscent of mostrattlesnake bites in North America.25 Venoms from adultspecimens of C. simus from Costa Rica present high proteolytic,hemorrhagic, and edematogenic activities and are devoid ofneurotoxic activity.26,27 Local symptoms may be severe, withpain, massive swelling, blistering, and necrosis that sometimeslead to physicians to perform fasciotomies and in some casesamputations. Systemic effects are relatively mild or moderate28,29
involving hemostatic disturbances (hypofibrinogenemia) butspontaneous systemic bleeding is most unusual, as are renalfailure and neurotoxicity. Consequently fatalities due to C.simus bites are rare. Only the venom from neonates containscrotoxin, a heterodimeric PLA2 molecule exhibiting presynaptic�-neurotoxicity typically found in C. durissus venoms,30 thatproduces systemic neurotoxicity and myalgic symptoms, to-gether with coagulation disturbances and frequent renal failureaccompanied by acute tubular necrosis.25-27,31-34 In contrast,the South American Neotropical rattlesnakes generally lackproteolytic, hemorrhagic and edematogenic activities35-38 andenvenomations by these species are very different from thoseof Nearctic species. C. d. cumanensis (Venezuela) is an exeption,and bites by this snake are characterized by local manifesta-tions, that is, edema, bleeding, and necrosis, in addition tosystemic neurotoxicity, myotoxicity, and coagulopathy.39 SouthAmerican rattlesnake venoms are notorious for their ability tocause systemic myotoxicity and disable neuromuscular trans-mission38,40 causing progressive paralysis. This symptomatologyis reminiscent of elapid and sea snake envenoming.25 Neuro-toxicity upon Crotalus bites is mainly attributable to thepresence of high concentrations of crotoxin. Venoms from C.d. cumanensis, C. d. durissus, and from certain populations ofC. d. terrificus and C. d. ruruima also contain variable amountsof crotamine, a myonecrotic toxin acting on voltage-sensitiveNa+ channels, paralyzing the prey, thus securing the meal and
* To whom correspondence should be addressed. Juan J. Calvete, Institutode Biomedicina de Valencia, C.S.I.C., Jaume Roig 11, 46010 Valencia, Spain.Phone: +34 96 339 1778. Fax: +34 96 369 0800. E-mail: [email protected].
† Instituto de Biomedicina de Valencia.‡ Facultad de Microbiologıa, Universidad de Costa Rica.§ Universidade Federal do Amazonas.| Universidad Central de Venezuela.⊥ Universidad Nacional Experimental Francisco de Miranda.# Centro de Investigaciones en Estructuras Microscopicas, Universidad
de Costa Rica.
Venomics of the Tropical Rattlesnake Complex research articles
Journal of Proteome Research • Vol. 9, No. 1, 2010 529

avoiding injury to the snake.41,42 Hemorrhagins may be presentin the venom, but any corresponding effects are completelyovershadowed by the startling and serious neurotoxic symp-toms.25
Adequate treatment of snakebite envenoming is criticallydependent on the parenteral administration of an appropriateantivenom to neutralize the venom-induced pathology.24 Ananticrotalic antivenom produced at Instituto Butantan (SaoPaulo, Brazil) against C. d. terrificus venom showed a very higheffectiveness in the neutralization of the lethal, myotoxin, andneurotoxic effects of venoms of C. durissus subspecies andnewborn C. simus.27 However, this antivenom failed to neutral-ize the hemorrhagic activity of C. simus, C. d. cumanensis, andC. d. ruruima (yellow) venoms.27,43 In contrast, a polyvalentantivenom produced by Instituto Clodomiro Picado (San Jose,Costa Rica) effectively neutralized the hemorrhagic activity ofCrotalus venoms. The Costa Rican antivenom was effective inthe neutralization of the lethality of venoms of adult C. simusbut not against C. durissus venoms, and showed a very lowneutralizing activity against newborn C. simus envenoming.27
Here we sought to investigate the molecular basis of theimmunoreactivity of the Costa Rican antivenom toward thevenoms of adult and 6-week-old specimens of C. simus andadult C. durissus using an antivenomics approach.23,24,44-47 Ourresults show that crotoxin and crotamine are poorly recognizedby the antivenom. We hypothesize that neurotoxicity associatedwith an increased expression of crotoxin and crotaminerepresented a paedomorphic trend and an adaptive trait alongCrotalus dispersal and diversification in South America. Thiswork illustrates how the knowledge of evolutionary trends mayhave an impact in formulating the mixture of venoms forimmunization aimed to produce an improved antivenomeffective against envenoming by rattlesnakes from Central andSouth America.
Experimental Section
Venoms and Antivenom. Crude venoms of adult and 6-weekold Crotalus simus (previously classified as C. durissus duris-sus48) were pooled from specimens collected in Costa Rica andfrom their captive-born offspring, all kept at the serpentariumof the Instituto Clodomiro Picado, University of Costa Rica inSan Jose. The venom from adult animals was collected by snakebiting on a parafilm-wrapped jar. The venom from neonateswas obtained by aspiration from the fangs using an Eppendorfpipet. Crude venoms were centrifuged at low speed to removecells and debris, lyophilized, weighed on a microbalance, andstored at -20 °C until used. Venom from C. d. cumanensis waspooled from adult specimens collected at the Adıcora savanna(Bocaına sector), Paraguana peninsula, Falcon State, Venezuela.Specimens were kept in captivity in the serpentarium of theDepartment of Animal Health, Veterinary Sciences Programme,School of Health Sciences, Universidad Nacional ExperimentalFrancisco de Miranda, Coro, Venezuela. The pooled “whitevenom” was collected from six adult specimens of C. d. ruruimacaptured at Monte Cristo Farm, in the municipality of Boa Vista,state of Roraima. The venom of C. d. ruruima may belong toone of two varieties, named white or yellow, according to theircolor appearance. The pool of white venom investigated in thiswork has been shown to display lethal, coagulant, myotoxic,edematogenic and hemolytic activities very similar to C. d.terrificus venoms.43 The yellow variety possess neurotoxicactivity and, in addition, induces hemorrhage, necrosis, caseinol-ysis, and crotamine-induced paralysis.43,49 Venoms of C. d.
durissus (formerly C. d. dryinas) from French Guyana and C.durissus terrificus (Brazil) were purchased from Latoxan (Va-lence, France). The antibothropic and anticrotalic (ABC) horseantivenom used for the antivenomic study (Batch L138, expirydate: September 2009) was raised against a mixture of thevenoms of Bothrops colombiensis and C. d. cumanensis in theCentro de Biotecnologıa, Facultad de Farmacia, UniversidadCentral de Venezuela (UCV). Inmunoglobulins were purifiedby ammonium sulfate precipitation and F(ab′)2 fragments wereproduced by pepsin digestion. The polyvalent (Crotalinae)antivenom (batch 4201007POLQ, expiry date: October 2010)manufactured at the Instituto Clodomiro Picado (ICP), Uni-versidad de Costa Rica, was produced by immunizing horseswith a mixture of equal amounts of the venoms of Bothropsasper, Crotalus simus, and Lachesis stenophrys obtained fromadult specimens kept in captivity at the ICP serpentarium.50
Figure 1. Characterization of the venom proteome of Crotalussimus from Costa Rica.Reverse-phase HPLC separations of theproteins from pooled venom of (A) adult and (B) 6-week oldspecimens of C. simus, respectively. Two milligrams of totalvenom proteins were applied to a Lichrosphere RP100 C18
column, which was then developed with the following chromato-graphic conditions: isocratically (5% B) for 10 min, followed by5-15% B for 20 min, 15-45% B for 120 min, and 45-70% B for20 min. Fractions were collected manually and submitted toN-terminal sequencing and molecular determination by SDS-PAGE, ESI and MALDI mass spectrometry. The results are shownin Table 1. Inserted in panels A and B are SDS-polyacrylamideelectrophoretic gels showing the protein composition of thereverse-phase HPLC separated venom protein fractions displayedin the respective panel and run under nonreduced (upper panels)and reduced (lower panels) conditions. Molecular mass markers(in kDa) are indicated at the side of each gel. Protein bands wereexcised and characterized by mass fingerprinting and CID-MS/MS of selected doubly or triply charged peptide ions (Table 1).
research articles Calvete et al.
530 Journal of Proteome Research • Vol. 9, No. 1, 2010

Table 1. Assignment of the Reverse-Phase Fractions of Crotalus simus Venom Isolated from Adult (Figure 1A) and 6-Week Old(Figure 1B) Specimens, to Protein Families by N-Terminal Edman Sequencing, Mass Spectrometry, and Collision-InducedFragmentation by nESI-MS/MS of Selected Peptide Ions from In-Gel Digested Protein Bands (Separated by SDS-PAGE, SeeInserted Panels in Figure 1A and B)a
peptide ion
HPLC fraction Cs- N-terminal sequence molecular mass m/z Z MS/MS-derived sequence protein family
1 TPPAGPDGGPR 1021.5 Bradykinin-inhibitory peptide [Q2PE51]2,3 n.p.4 EECDCGSPANPCCDAAT 7325 Disintegrin Durissin 4-72 [P68521]5 GEECDCGSPANPCCDAA 7104 Disintegrin Durissin 3-70 [P68521]
AGEECDCGSPANPCCDA 7290 Disintegrin Durissin 3-70 [P68521]6 EAGEECDCGSPANPCCD 7639 458.7 2 GTVCRPAR Disintegrin Durissin 1-73 [P68521]
651.9 3 GDWNDDTCTGQSADCPR679.3 3 LRPGAQCADGLCCDQCR
GEECDCGSPANPCCDAA 7442 Disintegrin Durissin 3-73 [P68521]EECDCGSPANPCCDAAT 7383 Disintegrin Durissin 4-73 [P6852]EAGEECDCGSPANPCCD 7135 Disintegrin Durissin 1-68 [P68521]
7 Blocked 639.6 2 ZQWPPGHHIPP Bradykinin-potentiating peptide [P0C7J9]8,28 GIECDCGSIENPCCYA 8-9 kDa9/1 ∼Disintegrin atrolysin-e [P34182]9 GCYCGAGGQGWPQDAS 8261, 8968 Crotoxin acid chain [P08878] Chain A 41-
RQEDGEIVCGEDDPC Chain B (83-IVCGEDDPCGTQICE Chain B (89-ENCQGESQPC Chain C (129-138)
29 GCYCGAGGQGWPQDAS 8-9 kDa9/1 Crotoxin acid chain [P08878] Chain A 41-RQEDGEIVCGEDDPC Chain B (83-
30 GCYCGAGGQGWPQDAS 8-9 kDa9/1 Crotoxin acid chain [P08878] Chain A 41-CYCGAGGQGWPQDA Chain A (42-RQEDGEIVCGEDDPC Chain B (83-
10 GCYCGAGGQGWPQDAS 8777 Crotoxin acid chain [P08878] Chain A 41-112ENCQGESQPC Chain C (129-138)
11 GCYCGAGGQGWPQDAS 9226 Crotoxin acid chain [P08878] Chain A 41-115ENCQGESQPC Chain C (129-138)
12 HLLQFNKMIKFETRKNAI- 14186 649.9 2 YGYMFYPDSR Crotoxin basic chain 1 [P62022]-PFYAFYGCYCGWGGRG 753.3 2 CCFVHDCCYGK
871.9 2 GTWCEEQICECDR720.3 3 NAIPFYAFYGCYCGWGGR
13 VVGGHPCNINEHRSL 34 kDa9 698.3 2 AAYPEYGLPATSR Serine proteinase ∼ catroxase I [Q8QHK3]13* SLVQFNKMIKFETRK 14 kDa9/1 ∼Crotoxin basic chain 2 [P24027]13-32 SLVQFETLIMKIAGR 14 kDa9/1 PLA2 [∼ P00623] Crotalus adamanteus14 VVGGDECNINEHR 34 kDa9 698.3 2 AAYPEYGLPATSR Serine proteinase ∼ catroxase I [Q8QHK3]
IFGGRPCNINEHRFL 27 kDa9 698.3 2 AAYPEYGLPATSR Serine proteinase ∼ catroxase I [Q8QHK3]641.3 2 INLLNYSVCR714.9 3 LLDDAVCQPPYPELPATSR Serine proteinase ∼ catroxase II [Q8QHK2]
14* SLVQFNKMIKFETRK 14 kDa1/1 ∼ Crotoxin basic chain 2 [P24027]15 SLVQFETLIMKIAGRSGL- 14-16 kDa9/1 475.8 2 YWLFPPK PLA2 [∼ P00623] Crotalus adamanteus
-LWYSAYG 533.3 2 DNIPSYDNK753.3 2 CCFVHDCCYGK
16 VIGGDECNINEHRFL 27 kDa9 698.3 2 AAYPEYGLPATSR Serine proteinase ∼ catroxase I [Q8QHK3]714.9 3 LLDDAVCQPPYPELPATSR Serine proteinase ∼ catroxase II [Q8QHK2]
17 VIGGDECNINEHRFL 26 kDa9 568.8 2 SVQFDKEQRR Serine proteinase ∼[Q58G94] C. d. terrificus647.8 2 XNXXDYEVCR874.6 2 XXCAGVXEGGXDTCHR
18,19 AXDRNPLEECFRETD 56 kDa9/1 624.3 3 NNPGLLEYPVKPSEEGK L-amino acid oxidase [∼P56742]678.1 3 LNEFSQENENAWYFIK612.2 2 DWYANLGPMR619.2 2 SAAQLYVESLR519.3 2 FWEDDGIR532.3 2 NPLEECFR485.6 2 VQVHFNAR557.8 2 VIEIQQNDR891.1 2 EDIQTFCRPSMIQR748.9 3 FDEIVGGMDQLPTSMYEAIK
20,21 NPEHQRYVELFIVVD 26 kDa9/1 874.1 2 VAVTMTHELGHNLGIR PI-metalloproteinase [∼ ABA42116]555.3 2 TXNSFGEWR448.3 2 YSEXNXR
22 Blocked 989/481 kDa 684.8 2 LYCFPNSPENK PIII-metalloproteinase (dimeric) [JC7530]471.8 2 FQFNR906.5 2 QGAQCAEGLCCDQCR867.8 2 MYDIVNVITPIYHR
23-26 Blocked 52 kDa1/1 275.3 2 CADGK PIII-metalloproteinase [Q2QA02]354.7 2 SGTECR359.8 2 TDXXTR401.3 2 GMVXPGTK417.3 2 NNDDXDK467.8 2 ZKYNPFR526.3 2 GNYYGYCR
Venomics of the Tropical Rattlesnake Complex research articles
Journal of Proteome Research • Vol. 9, No. 1, 2010 531

Whole immunoglobulins were purified by caprylic acid pre-cipitation.51 IgG concentration was determined spectrophoto-metrically using an extinction coefficient (ε) of 1.4 for a 1 mg/mL IgG or F(ab′)2 fragment concentration at 280 nm using a 1cm light path length cuvette.52
Isolation and Characterization of Venom Proteins. Venomproteins were separated by reverse-phase HPLC as de-scribed.44-47 Isolated protein fractions were subjected toN-terminal sequence analysis (using a Procise instrument,Applied Biosystems, Foster City, CA, USA) following the manu-facturer’s instructions. Amino acid sequence similarity searcheswere performed against the available databanks using theBLAST program53 implemented in the WU-BLAST2 searchengine at http://www.bork.embl-heidelberg.de. The molecularmasses of the purified proteins were determined by SDS-PAGE(on 12 or 15% polyacrylamide gels), MALDI-TOF mass spec-trometry (using an Applied Biosystems Voyager-DE Pro instru-ment), and electrospray ionization (ESI) mass spectrometryusing an Applied Biosystems QTrap 2000 mass spectrometer54
operated in Enhanced Multiple Charge mode in the range m/z600-1700. Protein bands of interest were excised from Coo-massie Brilliant Blue-stained SDS-PAGE gels and subjected toautomated reduction, alkylation, and in-gel digestion withsequencing grade porcine pancreas trypsin (Promega) as previ-ously reported.44-47,55-59 Doubly- or triply charged ions ofselected peptides from the MALDI-TOF mass fingerprintspectra were sequenced by CID-MS/MS22-25 using an AppliedBiosystem’s QTrap 2000.
Antivenomics: Immunodepletion of Venom Proteins byPolyvalent Antivenoms. We have coined the term “antive-nomics” for the identification of venom proteins bearingepitopes recognized by an antivenom using proteomic tech-niques.23,24,44-47 Briefly, two milligrams of whole venom weredissolved in 70 µL of 20 mM phosphate buffer, pH 7.0, mixedwith 4 mg of purified polyvalent ICP antivenom IgGs or ABCF(ab′)2 fragments, and incubated with gentle stirring for 1 h at37 °C. IgG concentration was determined spectrophotometri-
cally as above. Thereafter, 6 mg of rabbit antihorse IgGantiserum (Sigma) in 350 µL of 20 mM phosphate buffer, pH7.0, were added, and the mixture was incubated for another1 h at 37 °C. Immunocomplexes were precipitated by centrifu-gation at 13 000 rpm for 30 min in an Eppendorf centrifugeand the supernatant was submitted to reverse-phase separationas described for the isolation of venom proteins. HPLC-fractions were characterized as described above. Controlsamples were subjected to the same procedure except thatantivenom IgGs were not included in the reaction mixture.
Results and Discusion
General Considerations and Aims. The C. durissus complexrepresents a wide-ranging species and an ill-defined numberof subspecies are currently recognized (http://www.reptile-database.org): C. d. durissus (the Guiana rattlesnake, previouslyrecognized as C. d. dryinas); C. d. cumanensis (Venezuelanrattlesnake; dry lowlands of Venezuela and Colombia); C. d.ruruima (slopes of Mount Roraima and Mount Cariman-Peruin Venezuela); C. d. marajoensis (restricted to the savannas ofIlha de Marajo at the mouth of the Amazon and TocantinsRivers in the State of Para, Brazil); C. d. terrificus (Brazil southof the Amazonian forests, extreme southeastern Peru, Bolivia,Paraguay, Uruguay, northern Argentina); C. d. unicolor (ArubaIsland rattlesnake, a dwarfed snake inhabiting the Island ofAruba off the coast of Venezuela); and C. d. trigonicus (Rupu-nuni savannas in southwestern Guyana). The validity of thelatter taxon is questionable,15 and the subspecies previouslyknown as C. d. cascavella (dry Caatinga region, Brazil) and C.d. collilineatus have been synonymized with C. d. terrificusfollowing Wuster et al. (2005).16
The very extensive range of the Central and South Americanrattlesnakes raises an interesting opportunity to investigateintragenus and inter(sub)specific venom variations, as numer-ous studies have demonstrated a significant variability in thecomposition and pharmacology of viperid venoms,60 including
Table 1. Continued
peptide ion
HPLC fraction Cs- N-terminal sequence molecular mass m/z Z MS/MS-derived sequence protein family
615.8 2 DNSPGQNNPCK649.9 2 MFYSNEDEHK760.8 3 YMYXHVAXVGXEXWSNEDK766.4 2 VXGXAYVGSMCHPK801.3 2 MYEXANTVNDXYR776.2 2 VCSNGHCVDVATAY885.4 2 SGSQCGHGDCCEQCK684.6 3 XTVKPEAGYTXNAFGEWR926.3 3 ASMSECDPAEHCTGQSSECPADVFHK
25,26 NPEHQRYVELFIVVD 24-26 kDa9/1 553.6 2 YNSDLNIIR PI-metalloproteinase [Q1ZZ79]664.3 2 YIELVIVADHR605.6 2 VTALPKGAVQPK793.6 2 GASLCIMRPGLTPGR622.3 3 IENDADSTASISACNGLK853.6 2 HDNAQLLTAIVLDKR804.6 2 VHEIVNFINGFYR
27 N.D. 56 kDa9/1 776.2 2 VCSNGHCVDVATAY PIII-metalloproteinase31,33 SLVQFETLIMKIAGR 16 kDa9/1 PLA2 [∼ P00623] Crotalus adamanteus32 VIGGDECNINEHRFL 289/321 kDa Serine proteinase34 VIGGDECNINEHRFL 289/321 kDa Serine proteinase
SLVQFNKMIKFETRK 16 kDa9/1 ∼Crotoxin basic chain 2 [P24027]
a Underlined numbers correspond to fractions found only in the baby snakes. Fraction numbers labeled with an asterisk indicate proteins only found inadult snakes in this particular fraction. X, Ile or Leu. Z, pyrrolidone carboxylic acid. Unless other stated, for MS/MS analyses, cysteine residues werecarbamidomethylated; Molecular masses of native proteins were determined by electrospray-ionization (( 0.02%) or MALDI-TOF (( 0.2%) massspectrometry. Apparent molecular masses were also determined by SDS-PAGE of non-reduced (9) and reduced (1) samples (insert); n.p., non peptidicmaterial found. N.D., not determined. Asterisk-labeled protein species in fractions 13 and 14 were exclusively found in the venom of neonates.
research articles Calvete et al.
532 Journal of Proteome Research • Vol. 9, No. 1, 2010

those from C. d. terrificus,37 C. d. ruruima,43,49 and C. d.cumanensis,61,62 and this situation has broad clinical andtherapeutic implications (discussed below). On the other hand,investigations aimed at understanding the molecular bases ofvenom variability as a plausible selective trait underlyingevolutionary patterns in this group of snakes, which may havealso clinical implications, have been few and the literature isoften confusing.63 With the aim of gaining a deeper insight intotheir toxin composition we have carried out a comparativeproteomic study of the venoms of adult and neonate C. simusand C. durissus subspecies. Ophidian accidents involving
subspecies of C. durissus are usually serious and frequently fatalin the absence of specific and adequate treatment. A furtheraim of this study was to investigate the spectrum of recognitionof Crotalus toxins, and thus the potential therapeutic useful-ness, of a polyvalent antivenom generated in Costa Rica againsta mixture of the venoms of Bothrops asper, Crotalus simus, andLachesis stenophrys.
Defining Ontogenetic Changes in the Venom Proteomeof Crotalus simus. The crude venoms pooled from adults and6-week old C. simus specimens were fractionated by reverse-phase HPLC (Figure 1A and B), followed by analysis of eachchromatographic fraction by SDS-PAGE (Figure 1A and B,inserts), N-terminal sequencing, and molecular mass determi-
Figure 2. Characterization of the venom proteome of Crotalusdurissus terrificus from Brazil. Venom proteins were separatedby reverse-phase HPLC as in Figure 1. Fractions were collectedmanually and submitted to N-terminal sequencing and moleculardetermination by SDS-PAGE, ESI, and MALDI mass spectrometry.The results are shown in Table 2. (Inset) SDS-PAGE gels showingthe protein composition of the reverse-phase HPLC separatedvenom protein fractions and run under nonreduced (upperpanels) and reduced (lower panels) conditions. Molecular massmarkers (in kDa) are indicated at the side of each gel. Proteinbands were excised and characterized by mass fingerprinting andCID-MS/MS of selected doubly or triply charged peptide ions(Table 2).
Figure 3. Characterization of the venom proteome of Crotalusdurissus cumanensis from Venezuela. Venom proteins wereseparated by reverse-phase HPLC and analyzed as described inthe legend of Figure 1. The results are shown in Table 3. (Inset)SDS-PAGE of the reverse-phase HPLC separated venom proteinsrun under nonreduced (upper panels) and reduced (lower panels)conditions. Molecular mass markers (in kDa) are indicated at theside of each gel. Protein bands were excised and characterizedby mass fingerprinting and CID-MS/MS of selected doubly ortriply charged peptide ions (Table 3).
Figure 4. Characterization of the venom proteome of Crotalusdurissus ruruima from Brazil. Venom proteins were separatedby reverse-phase HPLC and analyzed as described in the legendof Figure 1. (Inset) SDS-PAGE of the reverse-phase HPLC sepa-rated venom proteins run under nonreduced (upper panels) andreduced (lower panels) conditions. Molecular mass markers (inkDa) are indicated at the side of each gel. Protein bands wereexcised and characterized by mass fingerprinting and CID-MS/MS of selected doubly or triply charged peptide ions. The resultsare shown in Table 4.
Figure 5. Characterization of the venom proteome of Crotalusdurissus durissus (dryinas) from French Guiana. Venom proteinswere separated by reverse-phase HPLC and analyzed as de-scribed in the legend of Figure 1. (Inset) SDS-PAGE of the reverse-phase HPLC separated venom proteins run under nonreduced(upper panels) and reduced (lower panels) conditions. Molecularmass markers (in kDa) are indicated at the side of each gel.Protein bands were excised and characterized by mass finger-printing and CID-MS/MS of selected doubly or triply chargedpeptide ions. The results are shown in Table 5.
Venomics of the Tropical Rattlesnake Complex research articles
Journal of Proteome Research • Vol. 9, No. 1, 2010 533

nation by MALDI-TOF and ESI mass spectrometry (Table 1).Pure protein fractions were straightforwardly assigned byBLAST analysis to a known protein family. Protein fractionsshowing heterogeneous or blocked N-termini were analyzedby SDS-PAGE and the bands of interest were excised andsubjected to automated reduction, carbamidomethylation, andin-gel tryptic digestion. The resulting tryptic peptides were thenanalyzed by MALDI-TOF mass fingerprinting followed byautomated or de novo amino acid sequence determination ofselected doubly- and triply charged peptide ions by collision-induced dissociation tandem mass spectrometry (Table 1).
In line with previous reports by Gutierrez and colleagues27,31
who observed marked differences in the electrophoretic andpharmacological activity patterns between the venoms of
newborn and adult C. simus from Costa Rica, comparison ofthe HPLC separation profiles of pooled venoms of adultspecimens (Figure 1A) with those of neonate snakes (Figure1B) revealed prominent protein expression changes. Supportingthe view that venom proteomes are mainly composed ofproteins belonging to a few protein families,22-24,64,65 thevenom proteome of adult C. simus comprises toxins from 7protein families (Figure 6A and Table 6). A large proportion(71.7%) of these toxins are metalloproteinases (SVMPs), two-thirds of which belong to the PIII class, which are among themost potent hemorrhagic toxins.66,67 The other toxin familiesare represented by more modest amounts (3.5-8% of the totalvenom proteins). In marked contrast, the venom of 6-week oldC. simus is predominantly made of crotoxin (55.9%) and serine
Table 2. Assignment of the Reverse-Phase Fractions of Crotalus durissus terrificus Venom, Isolated as in Figure 2, to ProteinFamilies by N-Terminal Edman Sequencing, Mass Spectrometry, and Collision-Induced Fragmentation by nESI-MS/MS of SelectedPeptide Ions from In-Gel Digested Protein Bands (Separated by SDS-PAGE, Figure 2 Inset)a
peptide ion
HPLC fraction Cdt- N-terminal sequence molecular mass m/z Z MS/MS-derived sequence protein family
1 TPPAGPDGGPR 1021.6 Bradykinin-inhibitory peptide [Q90Y12]2-4 YKQCHKKGGHCFPKE 4783 Myotoxin Crotamine 3 [O73799]
4886 Myotoxin Crotamine 1 [P01475]5112 Myotoxin Crotamine 4 [P24334]
5 SSYGCYCGAGGQGWPQDAS 9430 Crotoxin acid chain [P08878]A-chain (38-76)-SS-B-chain (84-118)-SS-C-chain (125-138)
6 SSYGCYCGAGGQGWPQDAS 9647 Crotoxin acid chain [P08878]A-chain (38-78-SS-B-chain (84-118)-SS-C-chain (125-138)
7 SSYGCYCGAGGQGWPQDAS 9810 Crotoxin acid chain [P08878]A-chain (38-78-SS-B-chain (84-119-SS-C-chain (125-138)
8 SSYGCYCGAGGQGWPQDAS 9910 Crotoxin acid chain [P08878]A-chain (38-80SS-B-chain (84-119-SS-C-chain (125-138)
10073 Crotoxin acid chain [P08878]A-chain (38-81)-SS-B-chain (84-119)-SS-C-chain (125-138)
5-8 Blocked 1656.87 ZWSQRWPHLEIPP Bradykinin-potentiating peptide Cdt-1a [Q90Y12]9 HLLQFNKMIKFETRKNAI- 14186 Crotoxin basic chain 1 [P62022]
-PFYAFYGCYCGWGGRG10 IFGGHPCNINEHRFLALVYS 31 kDa9,1 Serine proteinase [∼ Q8UUK2]11 VVGGHPCNINEHRSLVVLFS 37.5, 39 kDa1 698.3 2 AAYPEYGLPATSR Serine proteinase ∼ catroxase I
641.3 2 INLLNYSVCR [Q8QHK3]10-18 N.D. 16.5 kDa1,9 649.9 2 YGYMFYPDSR ∼Crotoxin basic chain 1 [P62022]
871.9 2 GTWCEEQICECDR720.3 3 NAIPFYAFYGCYCGWGGR475.8 2 YWLFPPK PLA2[∼ P00623] Crotalus adamanteus
12 VIGGDECNINEHRFL 39 kDa1 698.3 2 AAYPEYGLPATSR Serine proteinase ∼ catroxase I [Q8QHK3]33 kDa1 714.9 3 LLDDAVCQPPYPELPATSR Serine proteinase ∼ catroxase II [Q8QHK2]
13 (V/I)IGGDECNINEHRFL 34 kDa1 565.3 2 FXVAXYTFR Serine proteinase756.3 2 VIGGDECNINEHR770.7 2 IIGG(D-oMe)ECNINEHR
14 VIGGRPCNINEHR 33 kDa1 568.8 2 SVQFDKEQRR Serine proteinase [Q58G94]647.8 2 XNXXDYEVCR874.6 2 ILCAGVLEGGIDTCHR
15-17 AXDRNPLEECFRETD 56 kDa1,9 935.2 2 NNPGLLEYPVKPSEEGK L-amino acid oxidase [∼P56742]612.2 2 DWYANLGPMR619.2 2 SAAQLYVESLR519.3 2 FWEDDGIR532.3 2 NPLEECFR485.6 2 VQVHFNAR557.8 2 VIEIQQNDR
97 kDa1 577.9 2 EGNHYGYCRKEQNTK PIII-metalloproteinase16 GLHCPSDWYYYDQH 14-16 kDa1 C-type lectin convulxin-R [O93426]
GFCCPSHWSSYDRY C-type lectin convulxin-� [O93427]17 N.D. 989/481 kDa 684.8 2 LYCFPNSPENK PIII-metalloproteinase Crotastasin [Q076D1]
906.5 2 QGAQCAEGLCCDQCR (dimeric)867.8 2 MYDIVNVITPIYHR514.9 2 DHQEFLIK501.9 2 NMPQCILK577.9 2 EGNHYGYCRKEQNTK602.4 2 NNNGYCYNGK864.3 2 NPCNIYYSPNDEDK
18 N.D. 56 kDa1,9 697.8 2 AAYPEYGLPATSR Serine proteinase Gyroxin-like [Q8QHK3]556.9 2 TLCAGIQQGGK
29,33 kDa1,9 728.4 2 THFLIYVGVHDR Serine proteinase Gyroxin-like B1_4874.3 2 ILCAGVLEGGIDTCNR [B0FXM2]773.8 3 NNEHIAPLSLPSSPPSVGSVCR
a X, Ile or Leu. Z, pyrrolidone carboxylic acid. Unless other stated, for MS/MS analyses, cysteine residues were carbamidomethylated; molecular masses of native proteins weredetermined by electrospray-ionization ((0.02%) or MALDI-TOF ((0.2%) mass spectrometry. Apparent molecular masses were determined by SDS-PAGE of non-reduced (9) andreduced (1) samples (insert); n.p., non peptidic material found.
research articles Calvete et al.
534 Journal of Proteome Research • Vol. 9, No. 1, 2010

proteinases (36%), whereas SVMPs, exclusively of class PIII,represent only 2% of the total venom proteins (Figure 6B, Table6). Neonate and adult C. simus share about 50% of their venomproteome (Figure 1, Table 1), although the concentration ofmost shared toxins (particularly of the crotoxin acid and basicchains (peaks 9-12, Figure 1) and the PIII-SVMP Q2QA02eluted in peak 24) strongly depart between the venoms. Thetoxin profiles correlate well with the overall biochemicalcharacteristics and pharmacological profile of venoms from
adult (hemorrhagic) and newborn (neurotoxic) C. simus.26
Further, the high amount of serine proteinases in the venomof 6-week old C. simus may account for the higher plasmacoagulant (in vitro) and defibrinating (in vivo) activities ofnewborn versus adult C. simus venoms reported by Saravia andcolleagues.27 These effects are likely due to the action ofthrombin-like serine proteinases.68
Stage-specific venom proteins differentially expressed duringontogenetic development have been reported for Bothrops atrox
Table 3. Assignment of the Reverse-Phase Fractions of Crotalus durissus cumanensis Venom, Isolated as in Figure 3, to ProteinFamilies by N-Terminal Edman Sequencing, Mass Spectrometry, and Collision-Induced Fragmentation by nESI-MS/MS of SelectedPeptide Ions from In-Gel Digested Protein Bands (Separated by SDS-PAGE, Figure 3 inset)a
peptide ion
HPLC fraction Cdc- N-terminal sequence molecular mass m/z Z MS/MS-derived sequence protein family
1 TPPAGPDGGPR 1021.5 Bradykinin-inhibitory peptide [Q2PE51]2,3 n.p.4 GEECDCGSPANPCCDAA 7100 Da9 ∼ Disintegrin Durissin 3-70 [P68521]5 EAGEECDCGSPANPCCD 7642 Da9 651.9 3 GDWNDDTCTGQSADCPR ∼ Disintegrin Durissin 1-73 [P68521]
679.3 3 LRPGAQCADGLCCDQCR6 GEECDCGSPANPCCDAA 7442 Da9 ∼ Disintegrin Durissin 3-73 [P68521]7,8 YKKCHKKGGHCFPKE 4786 Da9 610.9 2 ICIPPSSDFGK ∼ Myotoxin Crotamine 3 [O73799]
4889 Da9 ∼ Myotoxin Crotamine 1 [P01475]5111 Da9 ∼ Myotoxin Crotamine 4 [P24334]
9 GCYCGAGGQGWPQDAS 9.3-9.8 kDa9 Crotoxin acid chain [P08878] Chain A 41-RQEDGEIVCGEDDPC Chain B (83-ENCQGESQPC Chain C (129-138)
10 GCYCGAGGQGWPQDAS 9356 Da9 Crotoxin acid chain [P08878] Chain A 41-RQEDGEIVCGEDDPC Chain B (83-ENCQGESQPC Chain C (129-138)
11 {GCYCGAGGQGWPQDAS 9615 Da9 Crotoxin acid chain [P08878] Chain A 41-RQEDGEIVCGEDDPC Chain B (83-ENCQGESQPC Chain C (129-138)
12 HLLQFNKMIKFETRKNAI- 14186 Da9 649.9 2 YGYMFYPDSR Crotoxin basic chain 1 [P62022]-PFYAFYGCYCGWGGRG 753.3 2 CCFVHDCCYGK
871.9 2 GTWCEEQICECDR720.3 3 NAIPFYAFYGCYCGWGGR
13 SLLDFEMMIIKVAKK 13805 Da9 670.8 2 DNIPSYNNNYK PLA2[∼ AAO93138]753.4 2 CCFVHDCCYGK828.9 2 DVCECDGVAAVCFR
37 kDa9 594.8 2 IMGWGTISPTK ∼ Serine proteinase Catroxase I [Q8QHK3]698.3 2 AAYPEYGLPATSR
33 kDa9 641.3 2 INLLNYSVCR ∼ Serine proteinase Catroxase I [Q8QHK3]698.3 2 AAYPEYGLPATSR
14,15 V(I/V)GGHPCNINEHRSL 27 kDa9 605.9 2 XNXXDHAVCR Serine proteinase715.9 2 SXPSSPPSVGS(435.2)
16,17 V(I/V)GGDECNINEHRFL 27 kDa9 543.9 2 VPNEDEQTR ∼ Serine proteinase Catroxase II [Q8QHK2]750.3 2 VVGGDECNINEHR715.3 3 LLDDAVCQPPYPELPATSR696.1 2 TLPDVPYCANIK
17 NPEHQRYVELFIVVD 23 kDa9 555.3 2 TXNSFGEWR PI-metalloproteinase [∼ ABA42116]874.1 2 VAVTMTHELGHNLGIR
18 AXDRNPLEECFRETD 58980 Da9 624.3 3 NNPGLLEYPVKPSEEGK L-amino acid oxidase [∼P56742]612.2 2 DWYANLGPMR619.2 2 SAAQLYVESLR519.3 2 FWEDDGIR532.3 2 NPLEECFR485.6 2 VQVHFNAR557.8 2 VIEIQQNDR688.2 3 DCADIVINDLSLIHQLPK
19 Blocked 1159/601 kDa 766.4 2 VXGXAYVGSMCHPK Dimeric PIII-metalloproteinase678.3 2 (SG)TYFWSSPTPK
19,20 NPEHQRYVELFIVVD 24-26 kDa9/1 553.6 2 YNSDLNIIR PI-metalloproteinase [Q1ZZ79]664.3 2 YIELVIVADHR605.6 2 VTALPKGAVQPK
20 DCPSGWSSYEGHCYK 229/14-161 kDa 653.6 2 KGISYIWIAVR C-type lectin-like [∼ DQ464258]446.8 2 TWDDAER
21,22 Blocked 989/491 kDa 684.8 2 LYCFPNSPENK PIII-metalloproteinase (dimeric) [∼Q076D1]471.8 2 FQFNR906.5 2 QGAQCAEGLCCDQCR867.8 2 MYDIVNVITPIYHR
23 Blocked 52 kDa9/1 526.3 2 GNYYGYCR PIII-metalloproteinase [Q2QA02]801.3 2 MYEXANTVNDXYR885.4 2 SGSQCGHGDCCEQCK684.6 3 XTVKPEAGYTXNAFGEWR926.3 3 ASMSECDPAEHCTGQSSECPADVFHK
a X, Ile or Leu. Unless other stated, for MS/MS analyses, cysteine residues were carbamidomethylated; molecular masses of native proteins were determined byelectrospray-ionization ((0.02%) or MALDI-TOF ((0.2%) mass spectrometry. Apparent molecular masses were determined by SDS-PAGE of non-reduced (9) and reduced (1)samples (inset); n.p., non peptidic material found. M and m, denote, respectively, major and minor products within the same HPLC fraction.
Venomics of the Tropical Rattlesnake Complex research articles
Journal of Proteome Research • Vol. 9, No. 1, 2010 535

from the Brazilian Amazon69 and B. asper from the Caribbeanand the Pacific versants of Costa Rica.55 In these taxa, P-III classmetalloproteinases and serine proteinases appear to be moreabundant in juvenile specimens, while metalloproteinases fromclass P-I exhibit higher concentration in adult venoms. Further,the venom of adult B. asper is more complex and contains adistinct ratio of K49/D49 PLA2 molecules than that of the neo-nate,55 suggesting that the requirement for the venom to im-mobilize prey and initiate digestion may change with the size (age)of the snake. In addition, maintenance of venom complexity islikely driven by adaptive detoxification responses of the prey.70,71
The ontogenic shift described here for C. simus venom follows asomewhat different trend: drastic reduction in serine proteinaseand neurotoxic PLA2 expression concominantly with an elevatedsecretion of metalloproteinases and only a slight increase invenom complexity. The qualitative and quantitative adjustmentsin the composition of the venom proteome linked to the animal’sbody development appears to be linked to a shift in the feedinghabits of certain juvenile versus adult snakes, from cold-blooded(frogs and lizards) to warm-blooded (mammals) prey.72 Shifts invenom lethality for certain kinds of prey follow this generaltrend.73 This ontogenetic mechanism does not apply to C. simus,because this species feeds from its birth primarily on small rodents(Liomys salvini and Sigmodon hispidus among the most frequent)and lizards (Ctenosaura similis).15,74 Such a conspicuous venom
change may be related to variations in the size of prey by snakesof different ages and the variable requirements for immobilizingand digesting them. It has also been hypothesized that thesignificance of venom adaptation to specific diets represents atrade-off between the metabolic cost of venom production andincreasing foraging efficiency.75,76 Further, it has been docu-mented that venoms from neonate Neartic Crotalus snakes exhibita higher lethal potency to lizards than adult venoms.77 Gibbs andMackessy78 have recently provided strong evidence that toxicityto mammals represents a major axis along which venom evolutionhas occurred among Sistrurus rattlesnakes. However, the notionthat reciprocal coevolutionary “arm’s race” between snakes andtheir prey can potentially influence venom composition79,80 hasbeen questioned arguing that ecological and evolutionary factorsin addition to selection in relation to diet may be involved.81-83
The ontogenetic venom change observed in C. simus may haverepresented a phenotypic hallmark during the adaptation ofancestral diverging snake populations to new ecological nichesor competition for resources in existing ones.
Venom Proteomes of Neotropical Rattlesnakes: Com-position-Activity Correlations Point to Neurotoxicity Asan Adaptive Paedomorphic Trend along Crotalus Dispersalin South America. The venom proteomes of the South Ameri-can rattlesnakes Crotalus durissus terrificus (Figure 2), C. d.cumanensis (Figure 3), C. d. ruruima (Figure 4), and C. d.
Table 4. Assignment of the Reverse-Phase Fractions of Crotalus durissus ruruima Venom, Isolated as in Figure 4, to ProteinFamilies by N-Terminal Edman Sequencing, Mass Spectrometry, and Collision-Induced Fragmentation by nESI-MS/MS of SelectedPeptide Ions from In-Gel Digested Protein Bands (Separated by SDS-PAGE, Shown as Inserted Panel in Figure 4)a
peptide ion
HPLC fraction Cdc- N-terminal sequence molecular mass m/z Z MS/MS-derived sequence protein family
1 TPPAGPDGGPR 1021.5 Bradykinin-inhibitory peptide [Q2PE51]2,3 n.p.4,5 YKQCHKKGGHCFPKE 4-5 kDa1,9 Myotoxin Crotamine6-8 SSYGCYCGAGGQGWP 9.3-9.8 kDa1 Crotoxin acid chain [P08878]9 HLLQFNKMIKFETRK 15 kDa9 Crotoxin basic chain 1 [P62022]10 24 kDa9
SLLQFNKMIKFETRK 15 kDa9 PLA2 [∼ P62022]11 VVGGDECNINEHRSL 31 kDa9 Serine proteinase
SLLDFEMMIIKVAKK 13.8 kDa 670.8 2 DNIPSYNNNYK PLA2 [∼ AAO93138]753.4 2 CCFVHDCCYGK828.9 2 DVCECDGVAAVCFR
12 VVGGDECNINEHRSL 31 kDa9 715.9 2 SXPSSPPSVGS(435.2) Serine proteinase13 VIGGDECNINEHRFL 31 kDa9 714.8 3 LLDDAVCQPPYPELPATSR Serine proteinase ∼ catroxase II [Q8QHK2]14,15 VIGGDECNINEHRFL 30 kDa9 874.6 2 ILCAGVLEGGIDTCHR Serine proteinase [∼Q58G94]
647.8 2 XNXXDYEVCR15 Heterogenous 1159/581 kDa 766.4 2 VXGXAYVGSMCHPK Dimeric PIII-metalloproteinase
678.3 2 (SG)TYFWSSPTPK97 kDa1 577.9 2 EGNHYGYCRKEQNTK PIII-metalloproteinase56 kDa1,9 935.2 2 NNPGLLEYPVKPSEEGK L-amino acid oxidase [∼P56742]
612.2 2 DWYANLGPMR619.2 2 SAAQLYVESLR519.3 2 FWEDDGIR532.3 2 NPLEECFR485.6 2 VQVHFNAR557.8 2 VIEIQQNDR
16-19 DCPSGWSSYEGHCYK 26-28 kDa9/14-16 kDa1 602.9 2 HWFSRPCSK C-type lectin-like [∼ DQ464258]18 N.D. 989/481 kDa 684.8 2 LYCFPNSPENK PIII-metalloproteinase Crotastasin [Q076D1]
906.5 2 QGAQCAEGLCCDQCR (dimeric)514.9 2 DHQEFLIK501.9 2 NMPQCILK577.9 2 EGNHYGYCRKEQNTK602.4 2 NNNGYCYNGK864.3 2 NPCNIYYSPNDEDK
19 N.D. 1109/14-161 kDa 534.9 2 YYVWIGLR C-type lectin-like Crotocetin-2 [Q719L9]664.6 3 ETEFLQWYNTDCEEK730.6 3 DFDCPSGWSAYDQYCYR703.2 3 GFCCPLGWSSYEGHCYK C-type lectin-like Crotocetin-1 [Q719L8]773.8 3 GLHCPSDWYYYDQHCYR C-type lectin-like ∼[AAQ11363]
N.D. 56 kDa1,9 697.8 2 AAYPEYGLPATSR Serine proteinase Gyroxin-like [Q8QHK3]
a X, Ile or Leu. Unless other stated, for MS/MS analyses, cysteine residues were carbamidomethylated; molecular masses of native proteins were determined byelectrospray-ionization ((0.02%) or MALDI-TOF ((0.2%) mass spectrometry. Apparent molecular masses were also determined by SDS-PAGE of non-reduced (9) and reduced (1)samples (insert); n.p., non peptidic material found. M and m, denote, respectively, major and minor products within the same HPLC fraction.
research articles Calvete et al.
536 Journal of Proteome Research • Vol. 9, No. 1, 2010

durissus (Figure 5) were investigated applying the proteomicapproach described above for C. simus, and the proteinsidentified are listed in Tables 2-5, respectively. Their overalltoxin composition (Figure 6, Table 6) correlates well with thesigns and symptoms evoked by poisoning by these species.Notably, the venoms of the Neotropical Crotalus subspeciessampled belong to one of two distinct phenotypes, whichbroadly correspond to type I (high levels of SVMPs and lowtoxicity, LD50 >1 µg/g mouse body weight) and type II (lowmetalloproteinase activity and high toxicity, LD50 <1 µg/g mousebody weight) venoms defined by Mackessy,71 who has dem-onstrated an inverse relationship between toxicity and metal-loprotease activity in the crude venoms of 25 taxa of rattle-snakes.71 Here, we show that Middle and South AmericanCrotalus venoms also follow this trend (Table 6). Thus, exceptfor the high amounts (13%) of crotamine, the venom of C. d.cumanensis exhibits the hemorrhagic (PIII-SVMP-rich) toxinprofile found in the venom of adult C. simus (compare panelsA and D in Figure 6). The occurrence of crotamine in C. d.cumanensis venom explains its distinct ability from C. simusto induce spastic paralysis via hind limb hyperextension in miceabout 30 min following venom injection.84 However, the lowcrotoxin concentration of C. d. cumanensis venom is at oddswith the marked neurotoxicity reported for this species.27 Theoccurrence of different populations of C. d. cumanensis ex-pressing distinct venom phenotypes (A. Marlene Vargas, Uni-versidad Central deVenezuela, personal communication) could
be a plausible explanation for this discrepancy. The hypothesisof intraspecific crotoxin variation would have evolutionaryimplications, that is, pointing to C. d. cumanensis as a transitiontaxon between the northern hemorrhagic and the southernneurotoxic venom phenotypes. Similarly, the occurrence ofconsiderable intrapopulational variability in the pharmacologi-cal activities of C. d. ruruima venoms43,49 may explain thequalitative and quantitative diversity of symptoms observed inpatients bitten by this subspecies, even in the same locality,and points to the Roraima region as a transitional territory fortype I and type II venom phenotypes.
The venoms of C. durissus subspecies terrificus (Figure 6C),ruruima (Figure 6E), and durissus (Figure 6F) show crotoxin-rich compositions (Table 6), in line with the marked neurotoxiccharacter of envenomations by these subspecies.25 C.d terrificusand C. d. durissus also contain large amounts (18-19%) ofcrotamine, whereas C. d. ruruima expresses a 10× lowerconcentration of this toxin in its venom. Crotamine appearsto represent a unique Crotalus (and thus recent) innovation ofa protein fold acting on Na+ channels.85 Among North Ameri-can snakes, crotamine has been detected only in some Sistrurusand Crotalus venoms,86 and in South America different C. d.terrificus and C. d. ruruima populations exhibit variable expres-sion of crotamine in their venoms.43,49,87 The toxin appears tobe widely distributed among rattlesnake species in the NewWorld but may vary qualitatively and quantitatively by geo-graphical region in several species and subspecies. The geo-
Table 5. Assignment of the Reverse-Phase Fractions of Crotalus durissus durissus (Formerly C. d. dryinas) Venom, Isolated As inFigure 5, to Protein Families by N-Terminal Edman Sequencing, Mass Spectrometry, and Collision-Induced Fragmentation bynESI-MS/MS of Selected Peptide Ions from In-Gel Digested Protein Bands (Separated by SDS-PAGE, Shown as Inserted Panel inFigure 4)a
peptide ion
HPLC fraction Cdd- N-terminal sequence molecular mass m/z Z MS/MS-derived sequence protein family
1 TPPAGPDGGPR 1021.6 511.1 2 Bradykinin-inhibitory peptide [Q2PE51]2,3 n.p.4,5 YKRCHKKGGHCFPKE 4806.8, 4911.2 Myotoxin Crotamine6-9 GCYCGAGGQGWPQDA 9552, 9891 Crotoxin acid chain [P08878] Chain A 41-
SSYGCYCGAGGQGWPQ Chain A 38-RQEDGEIVCGEDDPC Chain B (83-ZFSPENCQGESQPC Chain C (125-138)
10 HLLQFNKMIKFETRK 14201.8 649.9 2 YGYMFYPDSR Crotoxin basic chain 1 [P62022]753.3 2 CCFVHDCCYGK592.8 2 WDIYPYSLK871.9 2 GTWCEEQICECD
11 N.D. 289/141 kDa 592.8 2 WDIYPYSLK PLA212 VGGHPCNINEHRSLV 29 kDa1,9 502.3 2 DKDXMXPR Serine proteinase
552.3 2 VXNEDEQTR12-17 SLGQFNKMIKFETRKN 14.5 kDa1,9 649.9 2 YGYMFYPDSR ∼ Crotoxin basic chain 2 [P24027]
753.3 2 CCFVHDCCYGK592.8 2 WDIYPYSLK871.9 2 GTWCEEQICECD
13 VIGGDECNINEHRFL 269/30,331 kDa 565.3 2 FXVAXYTFR Serine proteinase715.4 2 SXPSSPPSVGSVCR647.8 2 XNXXDYEVCR
14 VIGGDECNINEHRFL 289/33,361 kDa 756.6 2 VIGGDECNINEHR Serine proteinase728.8 2 SXPSNPPSVGSVCR
15 VIGGDECNINEHRFL 269/331 kDa 756.6 2 VIGGDECNINEHR Serine proteinase604.8 2 FXVAXYDVXR
GLHCPSDWYYYDQH 1109/14.51 kDa C-type lectin ∼convulxin-R [O93426]GFCCPSHWSSYDRY C-type lectin ∼convulxin-� [O93427]
16 AXDRNPLEECFRETD 549/591 kDa 612.2 2 DWYANLGPMR L-amino acid oxidase [∼P56742]619.2 2 SAAQLYVESLR519.3 2 FWEDDGIR532.3 2 NPLEECFR485.6 2 VQVHFNAR557.8 2 VIEIQQNDR
17 N.D. 989/481 kDa 538.9 3 TCRDPCCDAATCK PIII-metalloproteinase [∼S41607]837.9 3 NPCNIYYSPNDEDKGMVLPGTK PIII-metalloproteinase [∼JC7530]684.7 2 LYCFPNSPENK
a X, Ile or Leu. Z, pyrrolidone carboxylic acid. Unless other stated, for MS/MS analyses, cysteine residues were carbamidomethylated; Molecular masses of native proteins weredetermined by electrospray- ionization ((0.02%) or MALDI-TOF ((0.2%) mass spectrometry. Apparent molecular masses were also determined by SDS-PAGE of nonreduced (9) andreduced (1) samples (insert); n.p., non peptidic material found.
Venomics of the Tropical Rattlesnake Complex research articles
Journal of Proteome Research • Vol. 9, No. 1, 2010 537
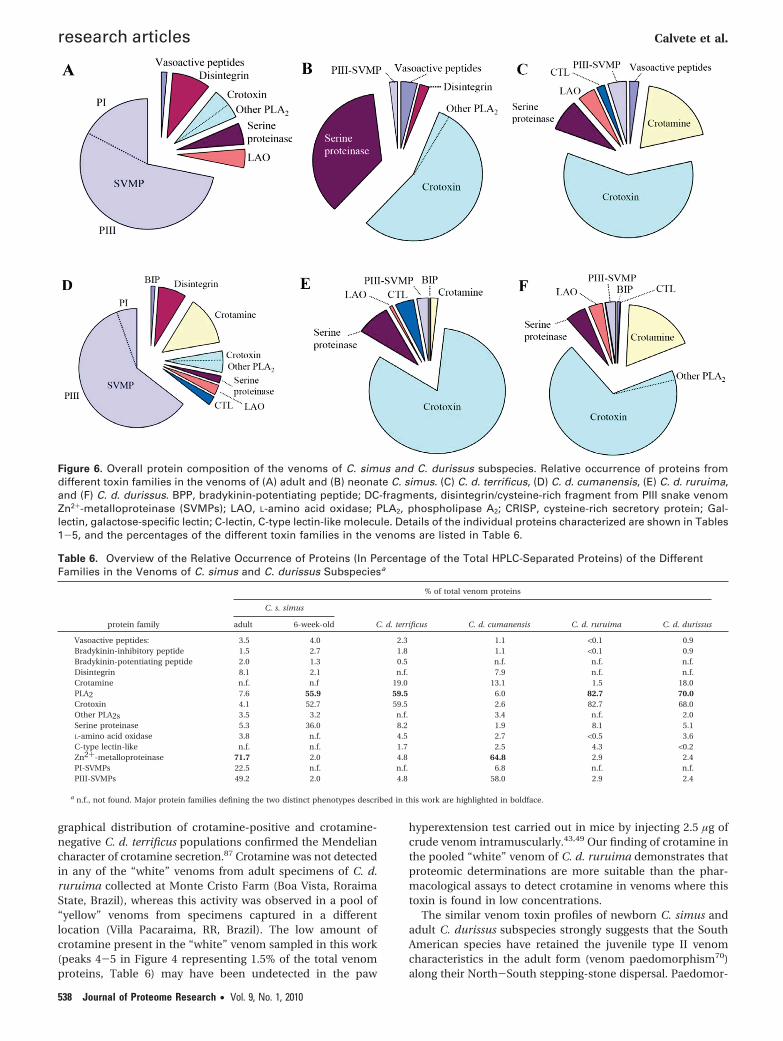
graphical distribution of crotamine-positive and crotamine-negative C. d. terrificus populations confirmed the Mendeliancharacter of crotamine secretion.87 Crotamine was not detectedin any of the “white” venoms from adult specimens of C. d.ruruima collected at Monte Cristo Farm (Boa Vista, RoraimaState, Brazil), whereas this activity was observed in a pool of“yellow” venoms from specimens captured in a differentlocation (Villa Pacaraima, RR, Brazil). The low amount ofcrotamine present in the “white” venom sampled in this work(peaks 4-5 in Figure 4 representing 1.5% of the total venomproteins, Table 6) may have been undetected in the paw
hyperextension test carried out in mice by injecting 2.5 µg ofcrude venom intramuscularly.43,49 Our finding of crotamine inthe pooled “white” venom of C. d. ruruima demonstrates thatproteomic determinations are more suitable than the phar-macological assays to detect crotamine in venoms where thistoxin is found in low concentrations.
The similar venom toxin profiles of newborn C. simus andadult C. durissus subspecies strongly suggests that the SouthAmerican species have retained the juvenile type II venomcharacteristics in the adult form (venom paedomorphism70)along their North-South stepping-stone dispersal. Paedomor-
Figure 6. Overall protein composition of the venoms of C. simus and C. durissus subspecies. Relative occurrence of proteins fromdifferent toxin families in the venoms of (A) adult and (B) neonate C. simus. (C) C. d. terrificus, (D) C. d. cumanensis, (E) C. d. ruruima,and (F) C. d. durissus. BPP, bradykinin-potentiating peptide; DC-fragments, disintegrin/cysteine-rich fragment from PIII snake venomZn2+-metalloproteinase (SVMPs); LAO, L-amino acid oxidase; PLA2, phospholipase A2; CRISP, cysteine-rich secretory protein; Gal-lectin, galactose-specific lectin; C-lectin, C-type lectin-like molecule. Details of the individual proteins characterized are shown in Tables1-5, and the percentages of the different toxin families in the venoms are listed in Table 6.
Table 6. Overview of the Relative Occurrence of Proteins (In Percentage of the Total HPLC-Separated Proteins) of the DifferentFamilies in the Venoms of C. simus and C. durissus Subspeciesa
% of total venom proteins
C. s. simus
protein family adult 6-week-old C. d. terrificus C. d. cumanensis C. d. ruruima C. d. durissus
Vasoactive peptides: 3.5 4.0 2.3 1.1 <0.1 0.9Bradykinin-inhibitory peptide 1.5 2.7 1.8 1.1 <0.1 0.9Bradykinin-potentiating peptide 2.0 1.3 0.5 n.f. n.f. n.f.Disintegrin 8.1 2.1 n.f. 7.9 n.f. n.f.Crotamine n.f. n.f 19.0 13.1 1.5 18.0PLA2 7.6 55.9 59.5 6.0 82.7 70.0Crotoxin 4.1 52.7 59.5 2.6 82.7 68.0Other PLA2s 3.5 3.2 n.f. 3.4 n.f. 2.0Serine proteinase 5.3 36.0 8.2 1.9 8.1 5.1L-amino acid oxidase 3.8 n.f. 4.5 2.7 <0.5 3.6C-type lectin-like n.f. n.f. 1.7 2.5 4.3 <0.2Zn2+-metalloproteinase 71.7 2.0 4.8 64.8 2.9 2.4PI-SVMPs 22.5 n.f. n.f. 6.8 n.f. n.f.PIII-SVMPs 49.2 2.0 4.8 58.0 2.9 2.4
a n.f., not found. Major protein families defining the two distinct phenotypes described in this work are highlighted in boldface.
research articles Calvete et al.
538 Journal of Proteome Research • Vol. 9, No. 1, 2010

phosis was first proposed by Walter Garstang in 1922.88 Thedriving force behind paedomorphism is often competition orpredation pressure. Per instance, increased procoagulant activ-ity of B. alcatraz venom has been hypothesized to represent apaedomorphic character.89 B. alcatraz has been isolated in theAlcatrazes Archipelago, 35 Km off the Atlantic coast of SaoPaulo, southeastern Brazil, since approximately 10 000 yearsago.90 Retention of juvenile characters in B. alcatraz appearsto be associated with a diet based on ectotherms (mainlycentipedes), owing to the absence of small mammalian preyon the islands.89 Moreover, the observation that the venomsof Brazilian, Ecuadorian, and Peruvian B. atrox exhibit the samePIII-SVMP-rich phenotype found in juvenile B. asper from CostaRica suggested that paedomorphism may have represented aselective trend during the trans-Amazonian southward expan-sion of B. atrox through the Andean Corridor.47
The underlying mechanisms for the paedomorphic trend inSouth American rattlesnake venoms (neoteny?, progenesis?,postdisplacement?) remains elusive. The interplay of differentcosts and benefits may explain the success of this phenomenonacross variable environments. However, comparison of thevenom toxin profiles of Central and South American Crotalus
taxa reveals a clear trend toward increasing concentration ofcrotoxin and crotamine along a North-South axis coincidentwith the dispersal route of Crotalus in South America (Figure7). Further, the increased concentration of neurotoxins in SouthAmerican rattlesnakes strongly argues that the gain of neuro-toxicity and lethal venom activities to mammals (type I to typeII venom shift71) may have represented the key axis along whichoverall venom toxicity has evolved during Crotalus durissusinvasion of South America. This hypothesis is in line with theprediction that the extremely high toxicity exhibited by Mojave-neurotoxin-positive North American rattlesnake venoms is atransitory populational phenomenon associated with novelprey bases91 and is supported by mapping a “lethal neurotox-icity coefficient” (LNC), which we define as the ratio betweenthe average LD50 of the venom and the crotoxin + crotamineconcentration, along the phylogeography pattern of the Neo-tropical rattlesnakes (Figure 7). The calculated LNC values forthe Crotalus species with known venom LD50s26,43,61,92 are: C.simus (adult), ∼ 201; C. simus (neonate), ∼ 2.1; C. d. cuman-ensis, 27.4; C. d. ruruima, 1.5; C. d. terrificus, 0.96. Further, thefact that venoms from newborn and adult C. d. terrificus, aspecies which feeds on rodents (Cavia aperea, Rattus norvegicusand Mus sp.) from birth,93 exhibit very similar electrophoreticpatterns and no ontogenetic difference could be detected intheir lethality to mice94 strongly suggests that venom-inducingrapid prey death may have represented an important featurein the evolution of the prey-capture strategy of Neotropicalrattlesnakes.
In Crotalus, venom type does not appear to be related tophylogeny, as neither basal nor derived taxa within any cladegroupings show one type preferentially.71 In species showingontogenetic shift from type II to type I, venom may conferselective advantage by ensuring rapid immobilization of theprey allowing venom lytic components, such as metallopro-teinases, to predigest efficiently and sufficiently large preyavoiding thereby the possibilty of putrefaction via gut floraactivity.71 On the other hand, as has been put forward forarthropod-feeding Echis76 and vertebrate-feeding snakes,75,95-97
shaping venom composition toward increased venom toxicityto natural prey species (type I to type II shift) could be adaptivein terms of metabolic energy savings, as less venom is requiredto achieve the same effect. As a whole, our data point tocrotoxin and crotamine, respectively, as paedomorphic andinnovative adaptive traits and suggest that achieving sexualmaturity while maintaining increased neurotoxicity may haveconferred evolutionary fitness to the C. durissus ancestorpopulation, thus fueling the paedomorphic trend.
Antivenomics of Crotalus simus and C. durissus Subspecies.The proteomic characterization of Middle and South Americanrattlesnake venoms reported here provides a molecular groundfor understanding (a) the failure of the polyvalent antivenomgenerated in Costa Rica using the venom of adult specimensof C. simus in the immunization mixture to neutralize theneurotoxicity and rhabdomyolysis induced by young CentralAmerican rattlesnake venoms26,29 and (b) the ineffectivenessof an anticrotalic antivenom produced at Instituto Butantan(Sao Paulo, Brazil) against C. d. terrificus venom to neutralizethe hemorrhagic activity of C. simus and C. d. cumanensisvenoms.27 The immunization mixtures used in Costa Rica andBrazil may simply lack, respectively, immunogenic concentra-tions of neurotoxins (crotamine and crotoxin) and hemorrha-gins (PIII-SVMPs), respectively. To assess this assumption wehave investigated the immunodepleting ability of (i) a Costa
Figure 7. Neurotoxicity as an adaptative trait. Phylogeographicaldistribution of the Central American, Crotalus simus, and Neo-tropical, Crotalus durissus, rattlesnakes. The relative concentra-tion of neurotoxic PLA2, crotoxin, and crotamine in the venomsis color-coded (see also Table 6). The gain of neurotoxicity-associated lethal venom activity to mammals, which is hypoth-esized to have occurred through an adaptive paedomorphic trait,appears to represent a major mode of venom evolution duringCrotalus invasion of, and speciation in, South America. Arrowsindicating estimated dates for the gradual dispersal along theCentral American Isthmus, followed by more rapid dispersal intoand across South America along a central Amazonian corridorduring the middle Pleistocene have been adapted from Wusteret al.16 Intraperitoneal median lethal dose ranges (LD50) reportedin the literature26,43,61,91 are expressed in µg/kg. LD50 data for C.d. durissus (formerly dryinas) are not available in the literature.The two-headed arrow at the left lower corner of the figurerepresents the proposed paedomorphic trend toward increasingconcentration of neurotoxins (crotoxin and crotamine), concomi-tantly with an inverse relationship between lethality (LD50) andmetalloprotease activity, along the North-South axis coincidentwith the dispersal route of Crotalus in South America. Thepaedomorphic trend was quantitated by a LNC (lethal neurotox-icity coefficient define as the ratio between the average LD50 ofthe venom and the crotoxin + crotamine concentration) alongthe phylogeography pattern of the Neotropical rattlesnakes.
Venomics of the Tropical Rattlesnake Complex research articles
Journal of Proteome Research • Vol. 9, No. 1, 2010 539
Rican polyvalent antivenom (generated against a mixture ofvenoms from Bothrops asper, Crotalus simus, and Lachesisstenophrys) and (ii) an antibothropic and anticrotalic (ABC)antivenom raised in Venezuela against a mixture of the venomsof Bothrops colombiensis and C. d. cumanensis, toward the
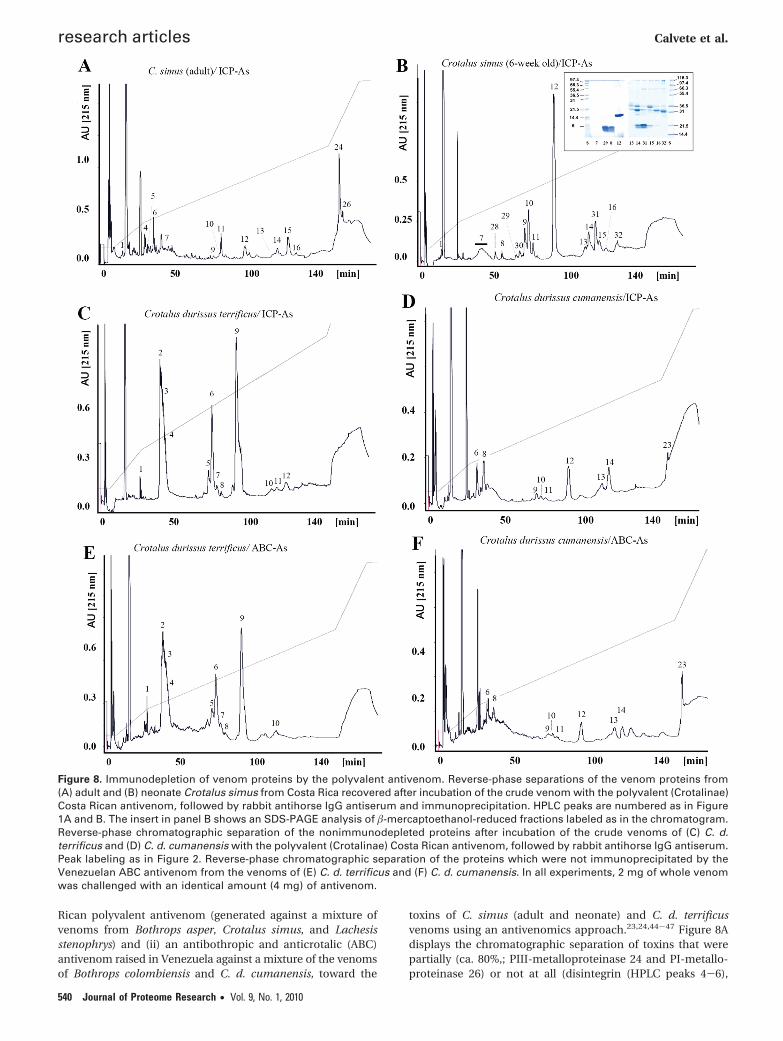
toxins of C. simus (adult and neonate) and C. d. terrificusvenoms using an antivenomics approach.23,24,44-47 Figure 8Adisplays the chromatographic separation of toxins that werepartially (ca. 80%,; PIII-metalloproteinase 24 and PI-metallo-proteinase 26) or not at all (disintegrin (HPLC peaks 4-6),
Figure 8. Immunodepletion of venom proteins by the polyvalent antivenom. Reverse-phase separations of the venom proteins from(A) adult and (B) neonate Crotalus simus from Costa Rica recovered after incubation of the crude venom with the polyvalent (Crotalinae)Costa Rican antivenom, followed by rabbit antihorse IgG antiserum and immunoprecipitation. HPLC peaks are numbered as in Figure1A and B. The insert in panel B shows an SDS-PAGE analysis of �-mercaptoethanol-reduced fractions labeled as in the chromatogram.Reverse-phase chromatographic separation of the nonimmunodepleted proteins after incubation of the crude venoms of (C) C. d.terrificus and (D) C. d. cumanensis with the polyvalent (Crotalinae) Costa Rican antivenom, followed by rabbit antihorse IgG antiserum.Peak labeling as in Figure 2. Reverse-phase chromatographic separation of the proteins which were not immunoprecipitated by theVenezuelan ABC antivenom from the venoms of (E) C. d. terrificus and (F) C. d. cumanensis. In all experiments, 2 mg of whole venomwas challenged with an identical amount (4 mg) of antivenom.
research articles Calvete et al.
540 Journal of Proteome Research • Vol. 9, No. 1, 2010

bradykinin-potentiating peptide (7), crotoxin acid (9-11) andbasic (12) subunits, and serine proteinases (13, 14, and 16)immunodepleted from the venom of adult C. simus by theCosta Rican polyvalent antivenom. The negligible titer of thisantivenom toward crotoxin was consistently confirmed inexperiments with venoms of newborn C. simus (Figure 8B) andadult C. durissus terrificus (Figure 8C). In addition, the CostaRican antivenom did not deplete crotamine from C. d. terrificusvenom. In line with its immunoreactivity, this antivenomquantitatively depletes the C-type lectin-like molecules, L-amino acid oxidase, and SVMPs from the venom of C. d.cumanensis, though it did not show any affinity for disintegrindurissin (6), crotamine (8), crotoxin acid (9-11) and basic (12)subunits, and some PLA2 (13) and serine proteinase (14)molecules (Figure 8D). The Venezuelan ABC antivenom pre-sented identical immunodepleting activity as the antivenomfrom Costa Rica toward the venoms of C. d. terrificus (Figure8E) and C. d. cumanensis, except for a slightly greater affinityfor crotamine and crotoxin (Figure 8F). The observed immu-noreactivity patterns of the ICP and the ABC polyvalentantivenoms is in line with our hypothesis that the immuniza-tion mixtures used to generate them did not contain thenecessary amounts of crotamine and crotoxin to elicit high-affinity antibodies. Our antivenomic outcome also explains whythe Costa Rican antivenom effectively neutralizes the hemor-rhagic activity of adult C. simus and C. cumanensis venoms27
but does not protect against the neurotoxicity produced byadult C. durissus and newborn C. simus envenoming.27
Concluding Remarks: Learning from Evolution to ImproveAntivenom Effectiveness. Snakebite envenoming constitutes ahighly relevant public health issue on a global basis, althoughit has been systematically neglected by health authorities inmany parts of the world.98-100 Envenomation is a pathologymainly affecting young agricultural workers living in villagesfar from health care centers in low-income countries of Africa,Asia, and Latin America, so it must be regarded as a “neglectedtropical disease”.98 In Central America, bites by C. simusconstitute a low percentage of the total number of cases. InCosta Rica, for instance, from 477 snakebite cases recorded in1979, only 5 of them (1.01%) corresponded to rattlesnakebites.101 In Brazil, however, 9% of the 26 688 snakebite casesreported to have occurred between 2000 and 2007 wereinflicted by Crotalus durissus ssp,102 and the case fatality rateranged from 0.8 to 2.53%.34,103 Subspecies of C. durissus areresponsible for ca. 5-10% of snakebite cases in other SouthAmerican countries, such as Colombia, French Guiana, Ven-ezuela, Bolivia, Paraguay, and Argentina.33,34,104 In Peru, rattle-snake bites are very rare and occur only in the southernDepartment of Puno.105
The growing concern of the relevance of the snakebitepathology has prompted an international initiative, under theauspices of the World Health Organization106 and involvingmany participants, aimed at coordinating actions at a globallevel to confront this neglected, though treatable, healthproblem. One of the key aspects of this initiative is theimprovement of the quality, quantity, and access of antiven-oms. In this respect, our comparative antivenomic study of theICP and ABC polyvalent antivenoms strongly suggests thepossibility of using any of these antivenoms for the manage-ment of snakebites by adult C. simus and C. d. cumanensis fromthose populations exhibiting the hemorrhagic venom pheno-type. On the other hand, the identification of evolutionarytrends among tropical Crotalus, as reported here, may impact
the choice of venoms for immunization to produce an effectivepan-American anti-Crotalus antivenom. Such antivenom mustprimarily neutralize the toxic actions of four major toxin groups,PIII-SVMPs, crotoxin, crotamine, and thrombin-like serineproteinases. This assertion is also supported by studies showingthat antibodies generated against a single P-III metallopro-teinase from C. simus neutralized 69% of the hemorrhageinduced by the whole venom,107 and antibodies against cro-toxin from C. d. terrificus neutralized the lethal activity of thevenom.108,109 The decreasing LNC associated with increasingneurotoxin (crotoxin + crotamine) content, and the concomi-tantly inverse relationship between neurotoxicity and metal-loproteinase activity along the North-South Crotalus durissusinvasion of South America, strongly suggest that a pan-American anti-Crotalus antivenom might be achievable byhyperimmunizing with a mixture of southern and northernvenoms, that is, from adult C. d. terrificus and adult C. simus(or a Neartic species, i.e. C. atrox110). We are currently testingthis hypothesis in our laboratories.
Acknowledgment. This study has been financed bygrant BFU2007-61563 from the Ministerio de Educacion yCiencia, Madrid, projects from the Vicerrectorıa deInvestigacion, Universidad de Costa Rica (741-A7-611),CRUSA-CSIC (2007CR0004), and CYTED (206AC0281).Travelling between Spain and Costa Rica was financed byAcciones Integradas 2006CR0010 between CSIC and theUniversity of Costa Rica (UCR). We gratefully acknowledgethe assistance of the personnel of the serpentarium of theInstituto Clodomiro Picado, San Jose, Costa Rica, and toJose Luis Yrausquin, Herpetologist from the Research Unit,Department of Animal Health, Veterinary SciencesProgramme, Universidad Nacional Experimental Franciscode Miranda, Coro, Venezuela, for snake management andvenom collection.
References(1) Bullock, T. H.; Cowles, R. B. Physiology of an infrared receptor:
the facial pit of pit vipers. Science 1952, 115, 541–543.(2) Bakken, G. S.; Krochmal, A. R. The imaging properties and
sensitivity of the facial pit of pitvipers as determined by opticaland heat-transfer analysis. J. Exp. Biol. 2007, 210, 2801–2810.
(3) Castoe, T. A.; Daza, J. M.; Smith, E. N.; Sasa, M.; Kuch, U.;Campbell, J. A.; Chippindale, P. T.; Parkinson, C. L. Comparativephylogeography of pitvipers suggests a consensus of ancientMiddle American highland biogeography. J. Biogeogr. 2009, 36,88–103.
(4) Castoe, T. A.; Parkinson, C. L. Bayesian mixed models and thephylogeny of pitvipers (Viperidae: Serpentes). Mol. Phylogenet.Evol. 2006, 39, 91–110.
(5) Knight, A.; Styler, D.; Pelikan, S.; Campbell, J. A.; Densmore, L. D.;Mindrell, D. P. Choosing among hypotheses of rattlesnakephylogeny: a best-fit rate test for DNA sequence data. SystematicsBiol. 1993, 42, 356–367.
(6) Parkinson, C. L. Molecular systematics and biogeographicalhistory of pitvipers as determined by mitochondrial ribosomalDNA sequences. Copeia 1999, 1999, 576–586.
(7) Murphy, R. W.; Fu, J.; Lathrop, A.; Feltham, J. V.; Kovac, V.Phylogeny of the rattlesnakes (Crotalus and Sistrurus) inferredfrom sequences of five mitochondrial DNA genes In Biology ofthe Vipers; Hoggren, M., Schuett, G. W., Greene, H., Douglas,M. E., Eds.; Eagle Mountain Publishing: Eagle Mountain, UT,2002; pp 69-92.
(8) Gloyd, H. K. The Rattlesnakes, Genera Sistrurus and Crotalus: AStudy in Zoogeography and Evolution; Special Publication No. 4;Chicago Academy of Sciences: Chicago, 1940; pp 1-270.
(9) Brattstrom, B. H. Evolution of the pitvipers. Trans. San Diego Soc.Nat. History 1964, 13, 185–268.
(10) Place, A. J.; Abramson, C. I. A quantitative analysis of the ancestralarea of rattlesnakes. J. Herpetol. 2004, 38, 152–156.
Venomics of the Tropical Rattlesnake Complex research articles
Journal of Proteome Research • Vol. 9, No. 1, 2010 541

(11) Greene, H. Snakes: The Evolution of Mystery in Nature; Univ. ofCalifornia Press: Berkeley, 1997.
(12) Parkinson, C. L.; Campbell, J. A.; Chippindale, P. Multigenephylogenetic analysis of pitvipers, with comments on theirbiogeography. In Biology of the Vipers; Hoggren, M., Schuett,G. W., Greene, H., Douglas, M. E. , Eds.; Eagle MountainPublishing: Eagle Mountain, UT, 2002; pp 93-110.
(13) Klauber, L. M. Classification, distribution and biology of thevenomous snakes of northern Mexico, the United States andCanada. In Crotalus and Sistrurus; Bucherl, W., Buckley, E. E. ,Eds; Venomous animals and their venoms, Vol. 2, Venomousvertebrates.Academic Press: New York, 1971; pp 115-156.
(14) Klauber, L. M. Rattlesnakes: Their Habitats, Life Histories, andInfluence on Mankind. 2nd ed.; University of California Press:Berkeley, 1997.
(15) Campbell, J. A.; Lamar, W. W. The Venomous Reptiles of theWestern Hemisphere; Comstock Publishing Associates: Ithaca, NY,2004.
(16) Wuster, W.; Ferguson, J. E.; Quijada-Mascarenas, J. A.; Pook, C. E.;Salomao, M. G.; Thorpe, R. S. Tracing an invasion: landbridges,refugia and the phylogeography of the Neotropical rattlesnake(Serpentes: Viperidae: Crotalus durissus. Mol. Ecol. 2005, 14,1095–1108.
(17) Quijada-Mascarenas, J. A.; Ferguson, J. E.; Pook, C. E.; Salomao,M. G.; Thorpe, R. S.; Wuster, W. Phylogeographic patterns oftrans-Amazonian vicariants and Amazonian biogeography: theNeotropical rattlesnake (Crotalus durissus complex) as an ex-ample. J. Biogeogr. 2007, 34, 1296–1312.
(18) Lewontin, R. C. The genetic basis of evolutionary change; ColumbiaUniversity Press: New York, 1974.
(19) Orr, H. A.; Coyne, J. A. The genetics of adaptation - A reassess-ment. Am. Nat. 1992, 140, 725–742.
(20) Golding, G. B.; Dean, A. M. The structural basis of molecularadaptation. Mol. Biol. Evol. 1998, 15, 355–369.
(21) Greene, H. W. Dietary correlates of the origin and radiation ofsnakes. Am. Zool. 1983, 23, 431–441.
(22) Calvete, J. J.; Juarez, P.; Sanz, L. Snake venomics. Strategy andapplications. J. Mass Spectrom. 2007, 42, 1405–1414.
(23) Calvete, J. J.; Sanz, L.; Angulo, Y.; Lomonte, B.; Gutierrez, J. M.Venoms, venomics, antivenomics. FEBS Lett. 2009, 583, 1736–1743.
(24) Gutierrez, J. M.; Lomonte, B.; Leon, G.; Alape-Giron, A.; Flores-Dıaz, M.; Sanz, L.; Angulo, Y.; Calvete, J. J. Snake venomics andantivenomics: proteomic tools in the design and control ofantivenoms for the treatment of snakebite envenoming. J. Pro-teomics 2009, 72, 165–182.
(25) Warrell, D. A. Snakebites in Central and South America: Epide-miology, Clinical Features, and Clinical Management. In TheVenomous Reptiles of the Western Hemisphere; Campbell, J. A.;Lamar, W. W., Eds.; Comstock Publishing Associates: Ithaca, NY,2004; pp 709-761.
(26) Gutierrez, J. M.; Dos Santos, M. C.; Furtado, M. F.; Rojas, G.Biochemical and pharmacological similarities between the ven-oms of newborn Crotalus durissus durissus and adult Crotalusdurissus terrificus rattlesnakes. Toxicon 1991, 29, 1273–1277.
(27) Saravia, P.; Rojas, E.; Arce, V.; Guevara, C.; Lopez, J. C.; Chaves,E.; Velasquez, R.; Rojas, G.; Gutierrez, J. M. Geographic andontogenic variability in the venom of the neotropical rattlesnakeCrotalus durissus: Pathophysiological and therapeutic implica-tions. Rev. Biol. Trop. 2002, 50, 337–346.
(28) Bolanos, R.; Marın, O.; Mora-Medina, E.; Alfaro, E. A. El accidenteofıdico por cascabela (Crotalus durissus durissus) en Costa Rica.Acta Med. Costarric. 1981, 24, 211–214.
(29) Gutierrez, J. M. Snakebite envenomation in Central America InHandbook of venoms and toxins of reptiles; Mackessy, S. P., Ed.;CRC Press: Boca Raton, 2009; pp 491-507.
(30) Bon, C. Multicomponent neurotoxic phospholipases A2 In Venomphospholipase A2 enzymes: structure, function and mechanism(Kini, R. M. , ed.), Wiley: Chichester, 1997; pp 269-28.
(31) Lomonte, B.; Gene, J. A.; Gutierrez, J. M.; Cerdas, L. Estudiocomparativo de los venenos de serpiente cascabel (Crotalusdurissus durissus) de ejemplares adultos y recien nacidos. Toxicon1983, 21, 379–384.
(32) Azevedo-Marques, M. M.; Cupo, P.; Coimbra, T. M.; Hering, S. E.;Rossi, M. E.; Laure, C. J. Myonecrosis, myoglobinuria and acuterenal failure induced by South American rattlesnake (Crotalusdurissus terrificus) envenomation in Brazil. Toxicon 1985, 23, 631–636.
(33) Otero, R. Manual de Diagnostico y Tratamiento del AccidenteOfıdico; Editorial Universidad de Antioquia: Medellın, 1994.
(34) Fan, H. W.; Cardoso, J. L. Clinical toxicology of snake bites SouthAmerica. In Handbook of Clinical Toxicology of Animal Venomsand Poisons; Meier, J., White, J., Eds.; CRC Press: Boca Raton,FL, 1995; pp 667-688.
(35) Sanchez, E. F.; Freitas, T. V.; Ferreira-Alves, D. L.; Velarde, D. T.;Diniz, M. R.; Cordeiro, M. N.; Agostini-Cotta, G.; Diniz, C. R.Biological activities of venoms from South American snakes.Toxicon 1992, 30, 95–103.
(36) Santoro, M. L.; Sousa-e-Silva, M. C. C.; Goncalves, L. R. C.;Almeida-Santos, S. M.; Cardoso, D. F.; Laporta-Ferreira, I. L.; Saiki,M.; Peres, C. A.; Sano-Martins, I. S. Comparison of the biologicalactivities in venoms from three subspecies of the South Americanrattlesnake (Crotalus durissus terrificus, C. durissus casca6ella andC. durissus collilineatus) . Comp. Biochem. Physiol. C 1999, 122,61–73.
(37) Rosenfeld, G. Symptomatology, pathology, and treatment of snakebites in South America In Crotalus and Sistrurus; Bucherl, W.,Buckley, E. E., eds; Venomous animals and their venoms. Vol. 2,Venomous vertebrates. Academic Press: New York, 1971; pp 345-384.
(38) Azevedo-Marques, M. M.; Hering, S. E.; Cupo, P. Acidentecrotalico. In Animais Peconhentos no Brasil. Biologıa, Clınica eTerapeutica dos Acidentes, 2nd ed.; Cardoso, J. L. C., Franca,F. O. S., Wen, F. H., Malaque, C. M. S., Haddad, V., Eds.; Sarvier:Sao Paulo, 2009; pp 108-115.
(39) Yoshida-Kanashiro, E.; Navarrete, L. F.; Rodriguez-Acosta, A. Onthe unusual hemorrhagic and necrotic activities caused by therattlesnake (Crotalus durissus cumanensis) in a Venezuelanpatient. Rev. Cubana Med. Tropical 2003, 55, 38–40.
(40) Vital Brazil, O. Pharmacology of crystalline crotoxin. II. Neuro-muscular blocking action. Mem. Inst. Butantan 1966, 33, 981–992.
(41) Chang, C. C.; Tseng, H. Effect of crotamine, a toxin of SouthAmerican rattlesnake venom, on the sodium channel of murineskeletal muscle. Br. J. Pharmacol. 1978, 63, 551–559.
(42) Oguiura, N.; Boni-Mitak, M.; Radis-Baptista, G. New view oncrotamine, a small basic polypeptide myotoxin from SouthAmerican rattlesnake venom. Toxicon 2005, 46, 363–370.
(43) Dos-Santos, M. C.; Ferreira, L. C. L.; Dias da Silva, W.; Furtado,M. F. D. Caracterizacion de las actividades biologicas de losvenenos ‘amarillo’ y ‘blanco’ de Crotalus durissus ruruimacomparados con el veneno de Crotalus durissus terrificus. Toxicon1993, 31, 1459–1469.
(44) Lomonte, B.; Escolano, J.; Fernandez, J.; Sanz, L.; Angulo, Y.;Gutierrez, J. M.; Calvete, J. J. Snake venomics and antivenomicsof the arboreal neotopical pitvipers Bothriechis lateralis andBothriehis schlegelii. J. Proteome Res. 2008, 7, 2445–2457.
(45) Gutierrez, J. M.; Sanz, L.; Escolano, J.; Fernandez, J.; Lomonte,B.; Angulo, Y.; Rucavado, A.; Warrell, D. A.; Calvete, J. J. Snakevenomic of the Lesser Antillean pit vipers Bothrops caribbaeusand Bothrops lanceolatus: correlation with toxicological activitiesand immunoreactivity of a heterologous antivenom. J. ProteomeRes. 2008, 7, 4396–3408.
(46) Calvete, J. J.; Borges, A.; Segura, A.; Flores-Dıaz, M.; Alape-Giron,A.; Gutierrez, J. M.; Diez, N.; De Sousa, L.; Kiriakos, D.; Sanchez,E.; Faks, J. G.; Escolano, J.; Sanz, L. Snake venomics andantivenomics of Bothrops colombiensis, a medically importantpitviper of the Bothrops atrox-asper complex endemic to Ven-ezuela: Contributing to its taxonomy and snakebite management.J. Proteomics 2009, 72, 227–240.
(47) Nunez, V.; Cid, P.; Sanz, L.; De La Torre, P.; Angulo, Y.; Lomonte,B.; Gutierrez, J. M.; Calvete, J. J. Snake venomics and antivenom-ics of Bothrops atrox venoms from Colombia and the Amazonregions of Brazil, Peru and Ecuador suggest the occurrence ofgeographic variation of venom phenotype by a trend towardspaedomorphism. J. Proteomics 2009, 73, 57–78.
(48) Savage, J. M.; Campbell, J. A.; Lamar, W. W. On names forNeotropical rattlesnakes. Herpetol. Rev. 2005, 36, 369–371.
(49) Dos-Santos, M. C.; Borges de Assis, E.; Moreira, T. D.; Pinheiro,J.; Fortes-Dias, C. L. Individual venom variability in Crotalusdurissus ruruima snakes, a subspecies of Crotalus durissus fromthe Amazonian region. Toxicon 2005, 46, 958–961.
(50) Angulo, Y.; Estrada, R.; Gutierrez, J. M. Clinical and laboratoryalterations in horses during immunization with snake venomsfor the production of polyvalent (Crotalinae) antivenom. Toxicon1997, 35, 81–90.
(51) Rojas, G.; Jimenez, J. M.; Gutierrez, J. M. Caprylic acid fraction-ation of hyperimmune horse plasma: description of a simpleprocedure for antivenom production. Toxicon 1994, 32, 59–67.
(52) Fasman, D. G. Practical Handbook of Biochemistry and MolecularBiology; CRC Press: Boston, 1992.
research articles Calvete et al.
542 Journal of Proteome Research • Vol. 9, No. 1, 2010

(53) Altschul, S. F.; Madden, T. L.; Schaffer, A. A.; Zhang, J.; Zhang,Z.; Miller, W.; Lipman, D. J. Gapped BLAST and PSI-BLAST: anew generation of protein database search programs. NucleicAcids Res. 1997, 25, 3389–3402.
(54) Le Blanc, J. C.; Hager, J. W.; Ilisiu, A. M.; Hunter, C.; Zhong, F.;Chu, I. Unique scanning capabilities of a new hybrid linear iontrap mass spectrometer (QTRAP) used for high sensitivity pro-teomics applications. Proteomics 2003, 3, 859–869.
(55) Alape-Giron, A.; Sanz, L.; Escolano, J.; Flores-Dıaz, M.; Madrigal,M.; Sasa, M.; Calvete, J.J. Snake venomics of the lanceheadpitviper Bothrops asper: Geographic, individual, and ontogeneticvariations. J. Proteome Res. 2008, 7, 3556–3571.
(56) Angulo, Y.; Escolano, J.; Lomonte, B.; Gutierrez, J. M.; Sanz, L.;Calvete, J. J. Snake venomics of Central American pitvipers. Cluesfor rationalizing the distinct envenomation profiles of Atropoidesnummifer and Atropoides picadoi. J. Proteome Res. 2008, 7, 708–719.
(57) Sanz, L.; Escolano, J.; Ferretti, M.; Biscoglio, M. J.; Rivera, E.;Crescenti, E. J.; Angulo, Y.; Lomonte, B.; Gutierrez, J. M.; Calvete,J. J. Snake venomics of the South and Central American Bush-masters. Comparison of the toxin composition ofLachesis mutagathered from proteomic versus transcriptomic analysis. J. Pro-teomics 2008, 71, 46–60.
(58) Sanz, L.; Ayvazyan, N.; Calvete, J. J. Snake venomics of theArmenian mountain vipers Macrovipera lebetina obtusa andVipera raddei. J. Proteomics 2008, 71, 198–209.
(59) Wagstaff, S. C.; Sanz, L.; Juarez, P.; Harrison, R. A.; Calvete, J. J.Combined snake venomics and venom gland transcriptomicanalysis of the ocellated carpet viper, Echis ocellatus. J. Proteomics2009, 71, 609–623.
(60) Chippaux, J. P.; Williams, V.; White, J. Snake venom variability:methods of study, results and interpretation. Toxicon 1991, 29,1279–1303.
(61) Aguilar, I.; Guerrero, B.; Salazar, A. M.; Giron, M. E.; Perez, J. C.;Sanchez, E. E.; Rodrıguez-Acosta, A. Individual venom variabilityin the South American rattlesnake Crotalus durissus cumanensis.Toxicon 2007, 50, 214–224.
(62) Salazar, A, M.; Aguilar, I.; Guerrero, B.; Giron, M. E.; Lucena, S.;Sanchez, E. E.; Rodrıguez-Acosta, A. Intraspecies differences inhemostatic venom activities of the South American rattlesnakes,Crotalus durissus cumanensis, as revealed by a range of proteaseinhibitors. Blood Coagulation Fibrinolysis 2008, 19, 525–530.
(63) Vanzolini, P. E.; Calleffo, M. E. V. A taxonomic bibliography ofthe South American snakes of the Crotalus durissus complex(Serpentes, Viperidae). An. Acad. Bras. Cienc. 2002, 74, 37–83.
(64) Fry, B. G.; Wuster, W. Assembling an arsenal: origin and evolutionof the snake venom proteome inferred from phylogenetic analysisof toxin sequences. Mol. Biol. Evol. 2004, 21, 870–883.
(65) Fry, B. G. From genome to “venome”: molecular origin andevolution of the snake venom proteome inferred from phyloge-netic analysis of toxin sequences and related body proteins.Genome Res. 2005, 15, 403–420.
(66) Fox, J. W.; Serrano, S. M. Structural considerations of the snakevenom metalloproteinases, key members of the M12 reprolysinfamily of metalloproteinases. Toxicon 2005, 45, 969–985.
(67) Gutierrez, J. M.; Rucavado, A.; Escalante, T.; Dıaz, C. Hemorrhageinduced by snake venom metalloproteinases: biochemical andbiophysical mechanisms involved in microvessel damage. Toxi-con 2005, 45, 997–1011.
(68) Raw, I.; Rocha, M. C.; Esteves, M. I.; Kamiguti, A. S. Isolation andcharacterization of a thrombin-like enzyme from the venom ofCrotalus durissus terrificus. Braz. J. Med. Biol. Res. 1986, 19, 333–338.
(69) Guercio, R. A. P.; Shevchenko, A.; Shevchenko, A.; Lopez-Lozano,J. L.; Paba, J.; Sousa, M. V.; Ricart, C. A. O. Ontogenetic variationsin the venom proteome of the Amazonian snake Bothrops atrox.Proteome Sci. 2006, 4, 11.
(70) Mackessy, S. P.; Williams, K.; Ashton, K. G. Ontogenetic variationin venom composition and diet of Crotalus oreganus concolor: acase of venom paedomorphosis? Copeia 2003, 2003, 769–782.
(71) Mackessy, S. P. Venom Composition in Rattlesnakes: Trends andBiological Significance In The Biology of Rattlesnakes; Hayes,W. K., Beaman, K. R., Cardwell, M. D., Bush, S. P., Eds.; LomaLinda University Press: Loma Linda, CA, 2008; pp 495-510.
(72) Saldarriaga, M. M.; Otero, R.; Nunez, V.; Toro, M. F.; Dıaz, A.;Gutierrez, J. M. Ontogenetic variability of Bothrops atrox andBothrops asper snake venoms from Colombia. Toxicon 2003, 42,405–411.
(73) Marques, O. A. V.; Martins, M.; Sazima, I. A new insular speciesof pitviper from Brazil, with comments on evolutionary biology
and conservation of the Bothrops jararaca group (Serpentes,Viperidae). Herpetologica 2002, 58, 303–312.
(74) Solorzano, A. Serpientes de Costa Rica/Snakes of Costa Rica; INBio:Santo Domingo de Heredia, Costa Rica, 2004; pp 677-682.
(75) McCue, M. D. Cost of producing venom in three North Americanpitviper species. Copeia 2006, 2006, 818–825.
(76) Baslow, A.; Pook, C. E.; Harrison, R. A.; Wuster, W. Coevolutionof diet and prey-specific venom activity supports the role ofselection in snake venom evolution. Proc. R. Soc. B 2009, 276,2443–2449.
(77) Mackessy, S. P. Venom ontogeny in the Pacific rattlesnakesCrotalus viridis helleri and C.v. oreganus. Copeia 1988, 1988, 92–101.
(78) Gibbs, H. L.; Mackessy, S. P. Functional basis of a molecularadaptation: prey-specific toxic effects of venom from Sistrurusrattlesnakes. Toxicon 2009, 53, 672–679.
(79) Daltry, J. C.; Wuster, W.; Thorpe, R. S. Diet and snake venomevolution. Nature 1996, 379, 537–540.
(80) Mackessy, S. P.; Sixberry, N. M.; Heyborne, W. H.; Fritts, T. Venomof the Brown Treesnake, Boiga irregularis: ontogenetic shifts andtaxa-specific toxicity. Toxicon 2006, 47, 537–548.
(81) Sasa, M. Diet and snake venom evolution: Can local selectionalone explain intraspecific venom variation? Toxicon 1999, 37,249–252.
(82) Wuster, W.; Daltry, J. C.; Thorpe, R. S. Can diet explain intraspe-cific venom variation? Reply to Sasa. Toxicon 1999, 37, 253–258.
(83) Mebs, D. Toxicity in animals. Trends in evolution? Toxicon 2001,39, 87–96.
(84) Mancin, A. C.; Soares, A. M.; Andriao-Escarso, S. H.; Faca, V. M.;Greene, L. J.; Zuccolotto, S.; Pela, I. R.; Giglio, J. R. The analgesicactivity of crotamine, a neurotoxin from Crotalus durissus terri-ficus (South American rattlesnake) venom: a biochemical andpharmacological study. Toxicon 1998, 36, 1927–1937.
(85) Mouhat, S.; Jouirou, B.; Mosbah, A.; De Waard, M.; Sabatier, J. M.Diversity of folds in animal toxins acting on ion channels.Biochem. J. 2004, 378, 717–726.
(86) Bober, M. A.; Glenn, J. L.; Straight, R. C.; Ownby, C. L. Detectionof myotoxin alpha-like proteins in various snake venoms. Toxicon1988, 26, 665–673.
(87) Schenberg, S. Geographical pattern of crotamine distribution inthe same rattlesnake subspecies. Science 1959, 129, 1361–1363.
(88) Garstang, W. The Theory of Recapitulation: a critical Re-statementof the Biogenetic Law. Linn. Soc. Jour. Zool. 1922, XXXV, 81–101.
(89) Marques, O. A. V.; Martins, M.; Sazima, I. A new insular speciesof pitviper from Brazil, with comments on evolutionary biologyand conservation of the Bothrops jararaca group (Serpentes,Viperidae). Herpetologica 2002, 58, 303–312.
(90) Grazziotin, F. G.; Monzel, M.; Echeverrigaray, S.; Bonatto, S. L.Phylogeography of the Bothrops jararaca complex (Serpentes:Viperidae): past fragmentation and island colonization in theBrazilian Atlantic forest. Mol. Ecol. 2006, 15, 3969–3682.
(91) Powell, R. L.; Lieb, C. S. Perspectives on venom evolution inCrotalus. In The Biology of Rattlesnakes; Hayes, W. K., Beaman,K. R., Cardwell, M. D., Bush, S. P., Eds.; Loma Linda UniversityPress: Loma Linda, CA, 2008; pp 551-556.
(92) Rangel-Santos, A.; Dos-Santos, E. C.; Lopes-Ferreira, M.; Lima,C.; Cardoso, D. F.; Mota, I. A comparative study of biologicalactivities of crotoxin and CB fraction of venoms from Crotalusdurissus terrificus, Crotalus durissus cascavella and Crotalusdurissus collilineatus. Toxicon 2004, 43, 801–810.
(93) Salomao, M. G.; Samtos, S. M. A.; Puorto, G. Activity patternofCrotalus durissus (Viperidae, Crotalinae): Feeding, reproductionand snakebite. Stud. Neotrop. Fauna Environ. 1995, 30, 101–106,1995.
(94) Furtado, M. F. D.; Santos, M. C.; Kamiguti, A. S. Age-relatedbiological activity of South American rattlesnake (Crotalus duris-sus terrificus) venom. J. Venom. Anim. Toxins incl. Trop. Dis. 2009,9, 186–201.
(95) Pawlak, J.; Mackessy, S. P.; Fry, B. G.; Bhatia, M.; Mourier, G.;Fruchart-Gaillard, C.; Servent, D.; Menez, R.; Stura, E.; Menez,A.; Kini, R. M. Denmotoxin, a three-finger toxin from the colubridsnake Boiga dendrophila (mangrove catsnake) with bird-specificactivity. J. Biol. Chem. 2006, 281, 29030–29041.
(96) Pawlak, J.; Mackessy, S. P.; Sixberry, N. M.; Stura, E. A.; Le Du,M. H.; Menez, R.; Foo, C. A.; Menez, A.; Nirthanan, S.; Kini, R. M.Iriditoxin, a novel covalently-linked heterodimeric three-fingertoxin with high taxon-specific neurotoxicity. FASEB J. 2009, 23,534–545.
(97) Starkov, V. G.; Osipov, A. V.; Utkin, Y. N. Toxicity of venoms fromvipers of Pelias group to crickets Gryllus assimilis and its relationto snake entomophagy. Toxicon 2007, 49, 995–1001.
Venomics of the Tropical Rattlesnake Complex research articles
Journal of Proteome Research • Vol. 9, No. 1, 2010 543

(98) Gutierrez, J. M.; Theakston, R. D. G.; Warrell, D. A. Confrontingthe neglected problem of snake bite envenoming: The need fora global partnership. PLoS Med. 2006, 3, e150.
(99) Gutierrez, J. M.; Higashi, H. G.; Wen, F. H.; Burnouf, T. Strength-ening antivenom production in Central and South Americanpublic laboratories: Report of a workshop. Toxicon 2007, 49, 30–35.
(100) Stock, R. P.; Massougbodji, A.; Alagon, A.; Chippaux, J.-P. Bringingantivenoms to Sub-Saharan Africa. Nat. Biotechnol. 2007, 2, 173–177.
(101) Bolanos, R. Las serpientes venenosas de Centroamerica y elproblema del ofidismo. Primera parte. Aspectos zoologicos,epidemiologicos y biomedicos. Rev. Costarric. Cien. Med. 1982,3, 165–184.
(102) De Oliveira, R. C. ; Wen, F. H.; Sifuentes, D. N. Epidemiologiados accidentes poe animais peconhentos. In Animais Peconhentosno Brasil. Biologia, Clınica e Terapeutica dos Acidentes, 2nd ed.;Cardoso, J. L. C., Franca, F. O. S., Wen, F. H., Malaque, C. M. S.,Haddad, V., Eds.; Sarvier: Sao Paulo, 2009; pp 6-21.
(103) Jorge, M. T.; Ribeiro, L. A. Epidemiology and clinical picture ofaccidental bites by the South American rattlesnake (Crotalusdurissus. Rev. Inst. Med. Trop. Sao Paulo 1992, 34, 347–354.
(104) Chippaux, J. P. Les envenimations ophidiennes en GuyaneFrancaise. Med. Trop. 2002, 62, 177–184.
(105) Zavaleta, A.; Salas, M. C. Ofidismo: envenenamiento por mord-edura de serpientes. In Emergencias en Medicina Interna; Vil-
laverde, J. R., Leon-Barua, R., Vidal-Neira, L., Losno-Garcıa, R.,Eds.; Lima, 1996; pp 241-260.
(106) World Health Organization Rabies and envenomings. A neglectedpublic health issue: Report of a consultative meeting; WHO:Geneva, 2007; Available: http://www.who.int/bloodproducts/animal_sera/Rabies.pdf.
(107) Azofeifa-Cordero, A; Arce-Estrada, V.; Flores-Dıaz, M.; Alape-Giron, A. Immunization with cDNA of a novel P-III type metal-loproteinase from the rattlesnake Crotalus durissus durissus elicitsantibodies which neutralize 69% of the hemorrhage induced bythe whole venom. Toxicon 2008, 52, 302–308.
(108) Rodriguez, J. P.; De Marzi, M.; Marunak, S.; Malchiodi, E. L.; Leiva,L. C.; Acosta, O. Rabbit IgG antibodies against phospholipase A2from Crotalus durissus terrificus neutralize the lethal activity ofthe venom. Med. (B. Aires) 2006, 66, 512–516.
(109) Beghini, D. G.; Hernandez-Oliveira, S.; Rodrigues-Simioni, L.;Novello, J. C.; Hyslop, S.; Marangoni, S. Anti-sera raised in rabbitsagainst crotoxin and phospholipase A2 from Crotalus durissuscascavella venom neutralize the neurotoxicity of the venom andcrotoxin. Toxicon 2004, 44, 141–148.
(110) Calvete, J. J.; Fasoli, E.; Sanz, L.; Boschetti, E.; Righetti, P. G.Exploring the venom proteome of the western diamondbackrattlesnake, Crotalus atrox, via snake venomics and combinatorialpeptide ligand library approaches. J. Proteome Res. 2009, 8,3055–3067.
PR9008749
research articles Calvete et al.
544 Journal of Proteome Research • Vol. 9, No. 1, 2010
Copyright © 2022 FDOKUMEN




















