
Salmonellosis outbreak linked to the consumption of fried ice-cream
Upload
khangminh22Category
view
0download
0
Nutrients 2020, 12, 974; doi:10.3390/nu12040974 www.mdpi.com/journal/nutrients
Commentary
Potential Adverse Public Health Effects Afforded by
the Ingestion of Dietary Lipid Oxidation Product
Toxins: Significance of Fried Food Sources
Martin Grootveld 1,*, Benita C. Percival 1, Justine Leenders 1 and Philippe B. Wilson 1
1 Leicester School of Pharmacy, De Montfort University, The Gateway, Leicester LE1 9BH, UK;
[email protected] B.C.P.); [email protected] (J.L.);
[email protected] (P.B.W.)
* Correspondence: [email protected]; Tel.: +44-(0)116-250-6443
Received: 07 October 2019; Accepted: 13 March 2020; Published: 1 April 2020
Abstract: Exposure of polyunsaturated fatty acid (PUFA)-rich culinary oils (COs) to high
temperature frying practices generates high concentrations of cytotoxic and genotoxic lipid
oxidation products (LOPs) via oxygen-fueled, recycling peroxidative bursts. These toxins, including
aldehydes and epoxy-fatty acids, readily penetrate into fried foods and hence are available for
human consumption; therefore, they may pose substantial health hazards. Although previous
reports have claimed health benefits offered by the use of PUFA-laden COs for frying purposes,
these may be erroneous in view of their failure to consider the negating adverse public health threats
presented by food-transferable LOPs therein. When absorbed from the gastrointestinal (GI) system
into the systemic circulation, such LOPs may significantly contribute to enhanced risks of chronic
non-communicable diseases (NCDs), e.g. cancer, along with cardiovascular and neurological
diseases. Herein, we provide a comprehensive rationale relating to the public health threats posed
by the dietary ingestion of LOPs in fried foods. We begin with an introduction to sequential lipid
peroxidation processes, describing the noxious effects of LOP toxins generated therefrom. We
continue to discuss GI system interactions, the metabolism and biotransformation of primary lipid
hydroperoxide LOPs and their secondary products, and the toxicological properties of these agents,
prior to providing a narrative on chemically-reactive, secondary aldehydic LOPs available for
human ingestion. In view of a range of previous studies focused on their deleterious health effects
in animal and cellular model systems, some emphasis is placed on the physiological fate of the more
prevalent and toxic α,β-unsaturated aldehydes. We conclude with a description of targeted
nutritional and interventional strategies, whilst highlighting the urgent and unmet clinical need for
nutritional and epidemiological trials probing relationships between the incidence of NCDs, and
the frequency and estimated quantities of dietary LOP intake.
Keywords: lipid oxidation products; lipid hydroperoxides; aldehyde toxins; frying oils; fried foods;
cytogenicity/gentoxicity/mutagenicity; cancer; atherosclerosis; acrolein; cooking oil fumes;
maximum human dietary intake (MHDI)
1. Introduction
An increasingly large proportion of the human population consuming Western World diets
frequently ingest oxidised/peroxidised lipids, and the possibility that regular ingestion of such agents
may be deleterious to human health has recently attracted a large amount of high-impacting research
interest and focus [1–5].

Nutrients 2020, 12, 974 2 of 49
Such lipid oxidation products (LOPs), which include cytotoxic and genotoxic aldehydes, along
with their lipid hydroperoxide precursors, epoxy-fatty acids, and many other secondary or even
tertiary LOPs [6,7], arise from the peroxidative deterioration of unsaturated fatty acids (UFAs),
particularly polyunsaturated fatty acids (PUFAs), and are commonly encountered in UFA-rich
culinary oils (COs), e.g. refined, non-genetically-engineered natural corn, sunflower or soybean oils,
when exposed to high temperature frying practices at ca. 180 °C, or when stored at ambient
temperatures for prolonged durations [8–11] (Figure 1).
(a)
(b)
Figure 1. (a) Simplified reaction scheme for the peroxidation of a linoleic acid substrate molecule
present in a culinary oil linoleoylglycerol species (H represents a hydrogen atom); the conjugated
hydroperoxydiene (CHPD) species shown is one of the cis,trans-CHPD classification. (b) Molecular
structures of aldehydes arising from the fragmentation of lipid hydroperoxides (HPMs and CHPDs).

Nutrients 2020, 12, 974 3 of 49
n-Nonanal and trans-2-decenal arise from the fragmentation of oleoylglycerol-derived HPMs; n-
hexanal, trans-2-octenal and trans,trans-deca-2,4-dienal from the fragmentation of linolenoylglycerol-
derived CHPDs; and propanal, acrolein, trans-2-pentenal and trans,trans-hepta-2,4-adienal from
linolenoylglycerol-derived CHPD fragmentation. cis,trans-Deca-2,4-dienal may arise from the
thermally-induced isomerism of its trans,trans-isomer [12].
Aldehydes act as potent toxins since they are extremely chemically-reactive [1,3,7,13]. Indeed,
they cause damage to critically important biomolecules such as DNA: since they are powerful
electrophilic alkylating agents, the -unsaturated classes of these aldehydes readily alkylate DNA
base adducts, and this generally but not exclusively serves to explain their now established
mutagenic, genotoxic and carcinogenic properties. Higher concentrations of this reactive aldehyde
are effective in potently suppressing a wide range of cellular processes, which leads to indiscriminant
cellular damage and ultimately apoptosis [14].
This Commentary paper focuses on the very wide range of potential public health threats
presented by both primary LOPs (lipid hydroperoxides) and their secondary fragmentation products
(aldehydes, etc.). Primarily, Section 2 provides an extensive comprehensive review of all possible
dietary sources of LOPs, and includes subsections focused on estimates of individual dietary
aldehyde intake, most especially the molecular nature and contents of those detectable in fried foods,
along with estimated risk assessments of their consumption by humans. Section 3 delineates the GI
system interactions, in vivo absorption, metabolism and biotransformation, toxicological properties
and potential adverse health effects of these agents respectively. Data available has provided
powerful evidence that only secondary LOPs (particularly aldehydes and epoxy-acids), and not their
primary hydroperoxide precursors, are transferred to foods during high-temperature frying
practices, and that these toxins have sufficient longevity therein [12], a factor which renders them
freely available for ingestion by human populations. Section 3 begins with a full evaluation of the
biomolecular pathways and probable physiological fates of aldehydic LOPs and their conjugated
hydroperoxydiene (CHPD) precursors; potential associations of the fractional contents of different
classes of aldehydes in fried foods and those of human blood plasma are also explored for the first
time. Since many previous investigations have focused on the potential roles of dietary LOPs and
their fried food sources as major risk factors for the induction and development of atherosclerosis
and its cardiovascular disease sequelae, and cancer, a review of these involvements and their adverse
health implications are provided in Sections 4 and 5, respectively (acrolein, crotonaldehyde and
trans,trans-2,4-decadienal as inhaled or ingested carcinogens represent special cases for
consideration). Subsequently, Section 6 discusses potential mechanisms available for the toxicities
and associated adverse health effects of dietary aldehydes, with a critical consideration of the
concentrations of these agents available in the GI system, the systemic circulation and elsewhere in
vivo. Finally, Section 7 explores targeted nutrition and potential interventional strategies for
diminishing the amounts of dietary LOPs available in the human diet, and which hopefully will
provide effective barriers to health risks posed by their ingestion; alternative ‘anti-aldehyde’
prophylactic or therapeutic strategies are also discussed. This section also considers the performance
of further, more intense research investigations to establish, optimize and validate maximum human
daily intake (MHDI) values for a full range of such dietary aldehydes, rather than relying on the very
limited data currently available. Throughout the text, reference to a series of examples of the
multicomponent analysis of secondary aldehydic LOPs in COs and foods (fried or otherwise) is made
in the Figures provided. The urgent requirement for future clinical feeding trials or epidemiological
investigations focused on explorations of relationships between the incidence and/or severity of
chronic non-communicable human diseases (NCDs), and the frequency and levels of dietary LOP
intake, is stressed in the Conclusions section. In view of the focus of this Commentary on lipid
hydroperoxides and their aldehydic chain-cleavage products, the in vivo absorption and toxicities of
dietary epoxy-fatty acids (FAs) are summarized in section S1 of the Supplementary Materials section.
2. Systematic Review of Major Points

Nutrients 2020, 12, 974 4 of 49
2.1. Lipid peroxidation process: mechanistic considerations and relative susceptibilities of acylglycerol FAs
The peroxidation of UFAs during high-temperature frying practices (ca. 180 °C) represents a
complex oxidative deterioration process involving chemically-reactive free radical species (i.e.
reactive chemical species with one or more unpaired electrons), and similarly-reactive singlet oxygen
(1O2). For PUFAs, primarily this process involves the heat- and/or light-induced loss of a hydrogen
atom (H●) from relatively weak bis-allylic-CH2- function carbon-hydrogen (C-H) bonds to generate a
resonance-stabilised pentadienyl carbon-centred radical (L●), which then reacts with atmospheric
dioxygen (O2) to form a reactive peroxyl radical (LOO●) (Figure 1(a)). Structurally, these PUFA-
specific bis-allylic-CH2- functions may be viewed as being ‘sandwiched’ between two strongly
electron-withdrawing carbon-carbon double bonds (>C=C<), and this explains the weakness of their
C-H bonds, which facilitates the abstraction of H● therefrom. Once formed, LOO● radicals can then
continue to react with another, adjacent PUFA molecule to generate an additional L● radical, the
unpaired oxygen-centred electron of the peroxyl radical being converted to a more stable lipid
hydroperoxide (LOOH) species, which for PUFAs are known as conjugated hydroperoxydienes
(CHPDs). Hence, this process is known as an autocatalytic, self-propagating chain reaction, which
unless terminated by the donation of another H● from a suitable lipid-soluble chain-breaking
antioxidant (e.g. phenolic agents such as alpha-tocopherol (α-TOH)), will continue relentlessly until
all PUFAs have been consumed. In view of its autocatalytic nature, plots of aldehydic LOP
concentrations generated against time usually appear as S-shaped curves, i.e. as sigmoidal
relationships [3,12].
Once formed, CHPDs fragment to a wide range of degradation products (secondary LOPs),
particularly at high frying temperatures, and these include extremely toxic aldehydes in particular
[1,7,8]. Further CHPD deterioration products include alcohols, ketones, oxo-acids, alkanes and
alkenes [1,7,15–17], in addition to epoxy-fatty acids such as 9,10-epoxy-12-octadecenoate, which is
also known as leukotoxin [18].
However, monounsaturated fatty acids (MUFAs), which produce corresponding
hydroperoxymonoenes (HPMs) in the same manner, are much more resistant to peroxidation than
PUFAs since they only have mono-allylic-CH2- functions, with stronger C-H bonds than those of the
bis-allylic-CH2- fucntions in PUFAs. Hence, MUFAs give rise to lower or much lower levels of both
primary and secondary LOPs when heated in this manner, with a much less broader range of
secondary aldehydic LOP classifications than those derived from PUFAs, and generally only after
prolonged exposures at standard frying temperatures [13]. For example, thermal stressing of MUFA-
rich olive oil generates much lower levels of aldehydes than those observed with PUFA-rich
sunflower or corn oils, and are predominantly limited to only trans-2-alkenals and longer-chain n-
alkanals. As expected, saturated fatty acids (SFAs) are virtually completely resistant to peroxidative
damage, even at high frying temperatures. Therefore, the order of toxic LOP production in culinary
oils is PUFAs >> MUFAs >>>>> SFAs, and hence PUFA-rich culinary oils represent the riskiest choice
for use as frying media, especially when exposed to repeated frying episodes [2,3,12]. Indeed, the
relative oxidative susceptibilities of these lipid classes are 1:100:1,200:2,500 for 18-carbon chain length
fatty acids containing 0:1:2:3 >C=C< functions respectively [8]. Moreover, the rate of fragmentation of
CHPDs or HPMs to the above series of smaller molecular LOPs also increases with increasing FA
unsaturation status, i.e. it is in the order linolenoyl- > linoleoyl- >>> oleoylglycerols [9].
2.2. Dietary sources of LOPs
An extensive review of the dietary availability of LOPs is provided in Section S2 of the
Supplementary Materials. These comprise outlines of the adverse generation and analysis of LOPs in
red meat, chicken and poultry (Section S2.1); fish products (Section S2.2); dairy products (Section
S2.3); grain products (Section S2.4); fruits and vegetables (Section S2.5); and alcoholic beverages
(Section S2.6). Moreover, a full outline of the use of aldehydes as food flavouring agents is presented
in Section S2.7, and their deleterious generation in thermoplastic food packaging materials is
summarised in Section S2.8.

Nutrients 2020, 12, 974 5 of 49
A review of estimations of dietary aldehyde intake by humans, and the essential considerations
required for such criteria, is provided in this section. Used and reused culinary frying oils, and more
especially foods which have been fried therein and therefore have uptaken variable levels of such
LOP-containing media, e.g. potato chips, chicken portions and beef patties, etc., serve as rich and
very important dietary sources of these toxins (especially aldehydes) in view of their frequent human
consumption in Western diets. Therefore, this area is covered in full detail below (Section 2.3). Section
2.4 outlines the exposure of human populations to fried food sources of dietary LOPs, along with
rational estimates of their ingestions (including sub-sections focused on acrolein and 4-hydroxy-
trans-2-nonenal/4-hydroxy-trans-2-hexenal (HNE/HHE)). Moreover, section 2.5 provides
information on the risk assessment of environmental aldehydes, with special reference to the
computation of margin of exposure (MOE) values.
Wang et. al. [19] conducted an extensive review of the availability of human exposure to
environmental aldehydes from various water, food, tobacco cigarette and ambient air sources (Table
1). Of the foods listed, dietary acrolein intake from vegetables, donuts, cheese and red wine are very
high, the latter providing as much as 3.8 g/kg of this aldehyde alone! Similarly, vinegar and coffee
are very rich sources of acetaldehyde (1.06 g/kg) and furfural (255 mg/kg) respectively, whereas anise
contains a staggeringly high content of anisaldehyde (25 g/kg).
Table 1. Sources of environmental aldehydes (adapted from [19], with permission). References for
these data are provided in the Supplementary Materials section of [19].
Source Aldehyde(s) Concentration
Water Surface water (irrigation canal) acrolein 20–200 mg/L
Non – carbonated bottled water
Formaldehyde,
acetaldehyde, nonanal,
and methyl glyoxal
1.7–57.5 mg/L
Carbonated bottled water
Formaldehyde,
acetaldehyde, nonanal,
and methyl glyoxal
3.9–197 mg/L
Ground and surface water Formaldehyde and
acetaldehyde 4.5–12 mg/L
Ozone – purified water Formaldehyde and
acetaldehyde 2–20 mg/L
Foods Fruits acrolein 10–50 mg/kg
Vegetables acrolein 10–590 mg/kg
Donuts acrolein 100–900 mg/kg
Codfish fillets acrolein 100 mg/kg
Cheese acrolein 290–1300 mg/kg
Red wine acrolein 3800 mg/kg
Vinegar acetaldehyde 1.06 gm/kg
Wheaten bread butanal 51 mg/kg
Coffee furfural 255 mg/kg
Bread propanal 31 mg/kg
banana 2–hexenal 2 mg/kg
Heated butter 2–pentenal 6 mg/kg
Heated butter 2,4–nonadienal 1.5 mg/kg
Vanilla vanillin 23 g/kg
Lime, peel oil citral 130 g/kg
Anise anisaldehyde 25 g/kg
Tangerine, peel oil 2,4–decadienal 500 mg/kg
Heated lard acrolein 109 mg/L
Sunflower oil acrolein 163 mg/L
Cigarettes Mainstream acrolein 10–140 mg/cigarette
Mainstream crotonaldehyde 18.5 mg/cigarette
Mainstream acetaldehyde 619 mg/cigarette

Nutrients 2020, 12, 974 6 of 49
Total aldehyde 777 mg/cigarette
Side – stream acrolein 100–1700 mg/cigarette
Ambient Urban air acrolein 0002–0.035 mg/m3
Smoky interiors acrolein 0.01–0.05 mg/m3
However, in Table 1, only estimated acrolein concentration values were provided for heated lard
and sunflower oil, these values being 109 and 163 mg/L repectively, and therefore our determined
concentrations of all other alkenals (predominantly trans-2-alkenals such as trans-2-octenal) and the
range of all other aldehyde classifications (including relatively predominant trans,trans-alka-2,4-
dienals, along with lower levels of substituted alkenals and n-alkanals) will undoubtedly provide an
additional and significant contribution to these environmental aldehyde levels, most notably for
consumers with a high incidence of fried food intake. Indeed, the only mention of fried food sources
in Table 1 is codfish fillets, which contained an acrolein content of 100 mg/kg. Notwithstanding,
heated butter was noted to provide low levels of 2-pentenal and 2,4-nonadienal, but this was the only
other peroxidised UFA source of aldehydes mentioned.
Wang et. al. [19] also provided estimates of the mean daily consumption of acrolein form a range
of food sources, and these were cheese (40 µg); donuts (380 µg); codfish fillets (10 µg); wine (1,520
µg); fruits (15 µg); vegetables (250 µg); potatoes (150 µg); and edible cooking oils (10 µg). However,
the latter value will be critically dependent on frying oil sources (those with relatively high levels of
ω-3 fatty acids yielding higher levels of this aldehyde), and also their use and reuse status. From these
8 classes of foods, the average human daily consumption of acrolein alone was estimated to be 2.35
mg. However, since at that time it was known that this LOP was detectable in 35 food classes, the
estimated maximal daily food consumption level in [19] was 5.0 mg/day. Moreover, exposure from
the smoking of tobacco cigarette products (50–100 µg per cigarette) was estimated, which is
equivalent to an additional 1.0–2.0 mg for a human smoking 20 cigarettes per day.
Of the above unsaturated aldehyde estimate, that for the maximal daily human exposure of
acrolein from a combination of food and water sources alone was found to be 0.1 mg/kg BW, along
with an equivalent quantity estimated from tobacco smoking, if appropriate [19].
Consideration of only the estimated non-smoking human contribution, this value in itself is
already 200-fold greater than the ADI value of 0.5 µg/kg BW specified for this aldehyde by the
Australian Government Department of Health (AGDH) [20], which is clearly a major cause for
concern. Dietary sources and estimated dietary intakes of acetaldehyde and formaldehyde, both of
which are also generated in the lipid peroxidation process (from the degradation of secondary
aldehydic LOPs, e.g. MDA for the latter [12]), are provided in Section S3 of the Supplementary
Materials. Moreover, comparative evaluations of the dietary availability for ingestion of aldehydic
LOPS with those of the process toxins acrylamide, monochloropropanediol (MCPD) adduct toxins,
and trans-FAs are discussed in Section S4 (Supplementary Materials).
2.3. Fried food sources of LOPs
Very high concentrations of LOPs, particularly secondary aldehydic ones, are generated during
such processes in view of the autocatalytic, self-propagating nature of this singlet oxygen (1O2)-
catalysed and lipid peroxyl radical (LOO)-mediated peroxidation process [14–17]. Indeed, the total
-unsaturated aldehyde concentration measured in PUFA-rich COs such as sunflower oil
thermally-stressed for a period of 90 min. according to laboratory-simulated shallow frying episodes
(LSSFEs) can reach values as high as 50 mmol/kg [12]. These unsaturated aldehydes are more toxic
than the saturated classes of these compounds which are also generated, and adversely represent 70–
75% of the total aldehyde remaining in COs heated in this manner. Notwithstanding, the very high
total levels of aldehydes often found in used UFA-rich frying oils only represent those retained after
the loss of substantial amounts of them through volatilisation processes, so that they also represent
components of very harmful cooking oil fumes. Health hazards arising from the human inhalation
of such aldehyde-laden fumes in poorly-ventilated kitchen areas are also discussed herein.

Nutrients 2020, 12, 974 7 of 49
Our research group’s extensive historic investigations of the oxidative deterioration of PUFA-
rich COs during standard frying practices, and their availability for uptake into fried foods such as
potato chips and crisp snacks, etc. available for human ingestion, have been accessible to the scientific
community since 1994 [6]. However, since that time, major advances in the development of
analytical/bioanalytical techniques for the investigation of patterns and concentrations of LOPs in
both food, and biofluid and solid biopsy samples, have been made. Indeed, earlier problems and
complications experienced with the high-resolution NMR analysis of such samples, including
sensitivity issues, have now been largely overcome via the application of newly-developed pulse
sequences, for example. The aldehydic-CHO function regions of the 600 MHz 1H-NMR spectra of a
commercially-available corn oil product exposed to laboratory-simulated shallow frying episodes
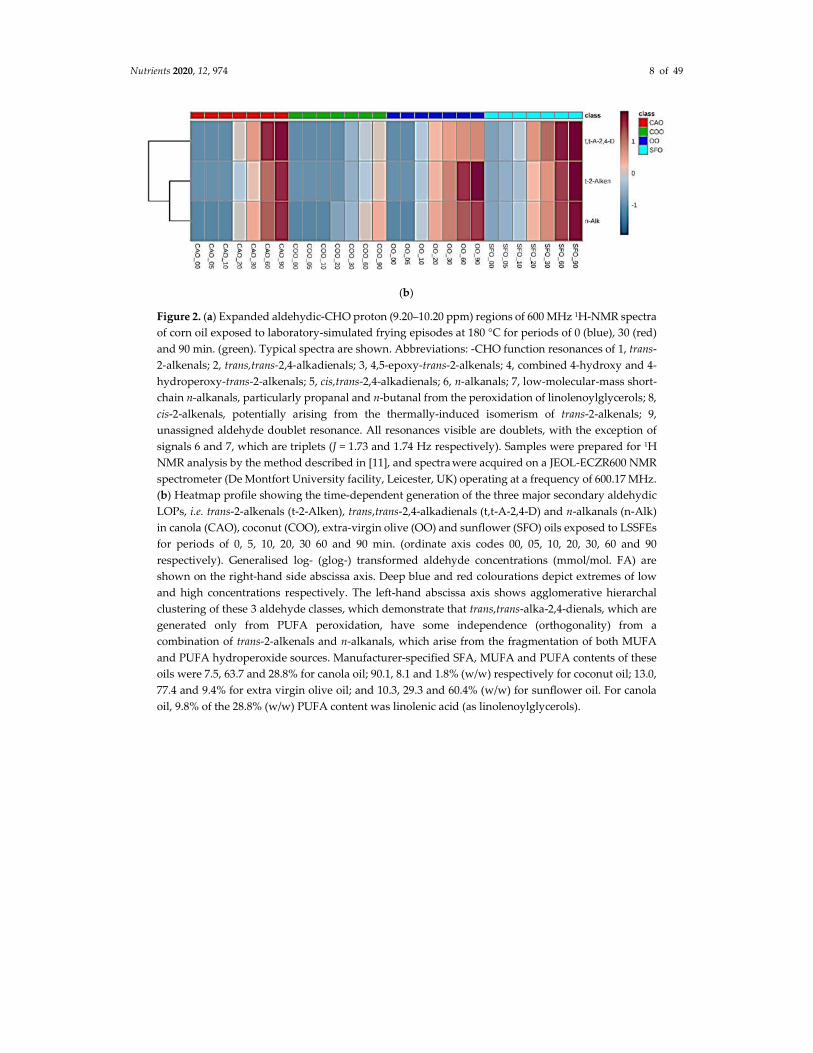
(LSSFEs) for periods of 0, 30 and 90 min. are shown in Figure 2, together with a heatmap diagram
displaying the critical dependence of the concentrations of three major aldehydic LOPs generated in
four different COs (of variable SFA, MUFA and PUFA contents) on increasing LSSFE time-points.
These data clearly demonstrate that the heating period-dependent levels of CO aldehydes generated
are high in PUFA-rich oils (corn and sunflower oils), intermediate in MUFA-rich ones (canola oil),
and much lower in SFA-laden coconut oil. More recently, we have confirmed passage of these
secondary LOP toxins from thermally-stressed frying oils into foods fried therein (Figure 3), and have
estimated their contents, which are consistently and considerably greater than those of acrylamide
and monochloro-propanediol (MCPD) adducts [3,14] (Section S4). Indeed, samples of repeatedly-
used frying oils collected from domestic kitchens, fast-food retail outlets and restaurants have
confirmed the generation of aldehydic and further LOP toxins at high concentrations during ‘on-site’
frying practices [3,14]. Our original research studies have been repeated, replicated, and further
exemplified by many research laboratories globally, e.g. [9]. Encouragingly, it now appears that these
highly important public health concerns are appreciated and respected by food science, nutrition and
associated clinical researchers.
(a)
Nutrients 2020, 12, 974 8 of 49
(b)
Figure 2. (a) Expanded aldehydic-CHO proton (9.20–10.20 ppm) regions of 600 MHz 1H-NMR spectra
of corn oil exposed to laboratory-simulated frying episodes at 180 °C for periods of 0 (blue), 30 (red)
and 90 min. (green). Typical spectra are shown. Abbreviations: -CHO function resonances of 1, trans-
2-alkenals; 2, trans,trans-2,4-alkadienals; 3, 4,5-epoxy-trans-2-alkenals; 4, combined 4-hydroxy and 4-
hydroperoxy-trans-2-alkenals; 5, cis,trans-2,4-alkadienals; 6, n-alkanals; 7, low-molecular-mass short-
chain n-alkanals, particularly propanal and n-butanal from the peroxidation of linolenoylglycerols; 8,
cis-2-alkenals, potentially arising from the thermally-induced isomerism of trans-2-alkenals; 9,
unassigned aldehyde doublet resonance. All resonances visible are doublets, with the exception of
signals 6 and 7, which are triplets (J = 1.73 and 1.74 Hz respectively). Samples were prepared for 1H
NMR analysis by the method described in [11], and spectra were acquired on a JEOL-ECZR600 NMR
spectrometer (De Montfort University facility, Leicester, UK) operating at a frequency of 600.17 MHz.
(b) Heatmap profile showing the time-dependent generation of the three major secondary aldehydic
LOPs, i.e. trans-2-alkenals (t-2-Alken), trans,trans-2,4-alkadienals (t,t-A-2,4-D) and n-alkanals (n-Alk)
in canola (CAO), coconut (COO), extra-virgin olive (OO) and sunflower (SFO) oils exposed to LSSFEs
for periods of 0, 5, 10, 20, 30 60 and 90 min. (ordinate axis codes 00, 05, 10, 20, 30, 60 and 90
respectively). Generalised log- (glog-) transformed aldehyde concentrations (mmol/mol. FA) are
shown on the right-hand side abscissa axis. Deep blue and red colourations depict extremes of low
and high concentrations respectively. The left-hand abscissa axis shows agglomerative hierarchal
clustering of these 3 aldehyde classes, which demonstrate that trans,trans-alka-2,4-dienals, which are
generated only from PUFA peroxidation, have some independence (orthogonality) from a
combination of trans-2-alkenals and n-alkanals, which arise from the fragmentation of both MUFA
and PUFA hydroperoxide sources. Manufacturer-specified SFA, MUFA and PUFA contents of these
oils were 7.5, 63.7 and 28.8% for canola oil; 90.1, 8.1 and 1.8% (w/w) respectively for coconut oil; 13.0,
77.4 and 9.4% for extra virgin olive oil; and 10.3, 29.3 and 60.4% (w/w) for sunflower oil. For canola
oil, 9.8% of the 28.8% (w/w) PUFA content was linolenic acid (as linolenoylglycerols).

Nutrients 2020, 12, 974 9 of 49
(a)
(b)
Figure 3. 1H-NMR Analysis of Aldehydic LOPs in C2HCl3 Extracts of Fast-Food Restaurant Fried Food
Samples. (a) and (b), Expanded aldehydic-CHO proton (9.40–9.90 ppm) regions of the 400 MHz 1H
NMR spectra of C2HCl3 extracts of fried potato chip and chicken (batter portion) servings purchased
from fast-food restaurants, which contain trans-2-alkenal, trans,trans-2,4-alkadienal, 4,5-epoxy-trans-
2-alkenal, combined 4-hydroxy-/4-hydroperoxy-trans-2-alkenal, cis,trans-2,4-alkadienal and n-alkanal
aldehydic LOP resonances in (a), and trans-2-alkenal, trans,trans-2,4-alkadienal and n-alkanal
resonances in (b). Typical spectra are shown. Typically, no aldehydic LOPs were 1H NMR-detectable
in the corresponding meat portion of the fried chicken sample corresponding to the batter extract
spectrum shown in (b). Samples were extracted and prepared for 1H NMR analysis by the method
described in [11], and spectra were acquired on a 400 MHz Bruker Avance NMR spectrometer
equipped with a QNP probe, and operating at 399.93 MHz (De Montfort University facility, Leicester,
UK). Abbreviations: as Figure 1, with F representing formaldehyde in (b).
2.4. Exposure of human populations to fried food sources of dietary aldehydes: rational estimates of their
dietary ingestion from these sources
Unfortunately, global governmental health recommendations for the maximum acceptable
human daily intakes (MHDIs) of aldehydes (i.e. those which are considered to be an acceptable intake

Nutrients 2020, 12, 974 10 of 49
that may be ingested daily throughout an entire lifetime without these agents presenting any
appreciable risk to human health) are either extremely limited or completely unavailable.
However, for acrolein (which is generated from the peroxidation of linolenoylyglycerols, or
alternatively from direct oxidation of glycerol liberated from triacylglycerol backbones via hydrolysis
reactions), the AGDH specified that this value was only 0.5 µg per kg of body weight, i.e. a total of
only 35 g for a mean human body weight of 70 kg [20]. Therefore, the observation that much higher
contents of trans-2-alkenals, trans,trans-2,4-alkadienals and n-alkanals than this limit are present in
fried potato chip servings purchased from a range of fast food ‘take-away/take-out’ restaurants [12]
(Figure 3), indicates a critically-important public health concern.
Using the assumption that all the trans-2-alkenals generated in the frying oils employed by these
outlets is trans-2-octenal (the predominant homologue of this aldehyde classification derived from
the fragmentation of linoleoylglycerol hydroperoxides), its estimated content in what is described as
a ‘large’ 154 g portion of this frequently-consumed fried food is 2.4 mg, a value which is ca. 70-fold
larger than that of this acceptable daily human intake limit for its lower homologue acrolein (which
corresponds to ca. 30-fold greater for its acrolein mass-equivalent figure of 1.04 mg). Parallel estimates
for the most predominant trans,trans-2,4-alkadienal and n-alkanal (n-hexanal) agents produced from
such linoleoylglycerol peroxidation sources were 3.8 and 1.9 mg (acrolein mass-equivalent values of
1.4 and 1.1 mg), respectively, within such a 154 g potato chip portion, and therefore the total aldehyde
content of this typical fast-food source, or at least for that fried in a hypothetical vegetable-derived
frying oil containing 100% (w/w) linoleoylglycerols, is (2.4 + 3.8 + 1.9) mg = 8.1 mg, with acrolein
mass-adjusted values of (1.0 + 1.4 + 1.1) mg = 3.5 mg, of which ca. 70–75% (w/w) are the more toxic
-unsaturated classes. However, it should be noted that the value computed here is estimated from
the consumption of a single staple fried food serving, and also that the above aldehydes are only
three possible, albeit three of the most prevalent, classes of aldehydic LOPs detectable, out of a total
of 10 or more of these generated in UFA-rich culinary vegetable oils during or following standard
frying practices [3,14].
Similarly, assuming that all aldehydes are the most prevalent ones arising from the
fragmentation of oleoylglycerol hydroperoxide (HPM) precursors, estimated potato chip portion
contents of trans-2-alkenal and n-alkanal toxins generated are 2.9 and 2.8 mg respectively (trans,trans-
2,4-alkadienals only arise from the fragmentation of PUFA-derived hydroperoxides) [13]. However,
since oleoylglycerol peroxidation reaction rates are much slower than those of PUFAs, as are the rates
of fragmentation of their HPMs to aldehydes and further products (i.e. it has a substantially lower
peroxidative susceptibility index (PSI) value [1,16]), much lower levels of LOPs are therefore
generated from such sources, and this explains why MUFA-rich oils such as olive oil are relatively
highly resistant to thermally-induced oxidation during standard frying episodes. Indeed,
peroxidative lag-phases for MUFA-rich cooking oils are much longer than those observed with oils
which contain high PUFA contents, e.g. sunflower or corn oils (Figure 2(b)). Several minor (lower
content level) classes of cytotoxic/genotoxic aldehydes are also detectable in fried potato chip samples
(e.g., cis,trans-2,4-alkadienals and formaldehyde), and this also adds significantly to the dietary
aldehydic LOP load.
Importantly, the above estimates pertain to only one 154 g-sized fried potato chip single meal
serving, and those of 300 g, or more are also quite common in the Western diet.
Recently, Grootveld et. al. [3,14] demonstrated that, in addition to the unsaturation status of
frying oils (reflected by their PSI values, which soar with increasing PUFA content), along with a
range of other factors such as frying methods (i.e. deep vs. shallow frying practices), frying
temperatures and durations, for example, the uptake of aldehydic LOP-containing culinary frying
oils (monitored as total lipids through high-resolution 1H-NMR analysis) was a critical determinant
of the aldehyde contents of fried potato chip products. However, the relative molecular content ratios
of trans-2-alkenals, trans,trans-2,4-alkadienals and n-alkanals of these products did not reflect those
present in their frying oil sources (which were also analyzed using 1H-NMR analysis in a C2HCl3
medium), with higher than expected n-alkanal contents. These results are best rationalized in terms
of the higher level of reactivities of the -unsaturated aldehyde classes with potato proteins, amino

Nutrients 2020, 12, 974 11 of 49
acids and further aldehyde-consuming biomolecules, over those of the saturated ones, or more
specifically, the ability of these classes of aldehydes to engage in Michael addition reactions, unlike
their saturated counterparts.
Interestingly, no further updates to the above AGDH ADI value of 0.5 µg per kg of BW, for
acrolein were made in their updated Edition 1/2017 (current as of 31st March 2017). Neither CHPDs,
nor HPMs, were detectable in these C2HCl3 fried food product sample extracts, and this observation
is presumably ascribable to their reactions with potato chip electron donors such as the amino acid
L-cysteine and/or other thiol function-containing biomolecules therein to form less toxic conjugated
hydroxydiene species, and/or their catalytic deterioration to further aldehydes and other secondary
LOP fragmentation products [16]. Indeed, formic acid, a product arising from the degradation of
MDA, was also detectable in these extracts. Therefore, from this investigation, it appears that
commonly-fried food products act as poor sources of CHPDs and HPMs, but nevertheless are rich in
their aldehydic degradation products.
Of further pertinence, shallow-frying episodes generate much higher levels of frying oil LOPs
than deep-frying ones in view of the greater surface area of the oil in the former case, i.e. its much
greater exposure to atmospheric O2 required for the lipid peroxidation process [1,7,14] (Figure 1). For
deep-frying practices, LOPs are only formed on the O2-exposed and -richer surface environment of
the oils employed for this purpose, and subject to a sufficient level of oil admixing/homogenisation
during such processes, are then markedly diluted through their dissipation throughout the much
larger volume bulk oil medium. Interestingly, Totani et. al. [21] found that oxidation was active at the
oil/air interface of bubbles produced by foods being fried in a canola-soybean oil blend according to
deep-frying practices. Marked decreases in the O2 content of these oil blends commenced at a
temperature of 120 °C; however, on being allowed to cool at ambient temperature, a slow restoral of
these pre-diminished O2 levels was found in oil blends pre-heated at a frying temperature of 180oC.
Therefore, it appears that intermittent cooling periods involved in the repeated use of frying oils
during recycling frying episodes facilitate their absorption of atmospheric O2.
2.4.1. Acrolein
Acrolein is generated during the frying, cooking or processing of lipid-containing foods [22–24],
especially those rich in ω-3 FAs such as oily fish products, and also artefactually peroxidised dietary
fish oil provisions or supplements in general [25]. An estimated mean concentration of acrolein of
0.51 mmol/kg was found in samples of five types of cooking oil heated to 80oC and aerated for a
period of 20 hr. [26]. Notably, in view of its high volatility, this unsaturated aldehyde was detected
in emissions arising from n = 4 heated cooking oil products in China [27] at levels varying from 49
µg/L in peanut oil to 392 µg/L in rapeseed oil (the latter oil has a relatively high content of the ω-3
FA linolenic acid (as linolenoylglycerols), one major PUFA source of this aldehyde). It should also be
noted that selected ingredients present in commercially-available breading systems and batter can
also give rise to acrolein in fried food matrices [24].
2.4.2. HNE and HHE
Estimates of the concentrations of HNE alone in French fry samples collected from n = 6 U.S.
fast-food restaurants [28] were found to range from 8 to 32 g/100 g (0.51 to 2.05 mol/kg), values
corresponding to 12–50 g for a standard ‘large’ sized 154 g serving. Moreover, assuming a mean
frying oil uptake of 12% (w/w) (range 1-33% (w/w) [3]), our laboratory’s 1H-NMR-based estimate of
the mean HNE content of 154 g potato chip portions is ca. 30 µg, a value which is in very good
agreement with those found in [28] (assuming no chemical reactions of this LOP with potato chip
biomolecules, e.g. proteins and amino acids, which is, however, unlikely). Furthermore, these
determined values are not dissimilar to the above Korean estimates. Our estimates have also
confirmed that HNE accounts for only ≤ 1% of the total molar -unsaturated aldehyde content of
fried potato chips (relative amounts of 4-hydroxy-trans-2-alkenals found in thermally-stressed
sunflower, corn and canola frying oils were <10% of the total measured [12]). Moreover, HNE levels
previously determined in sunflower oil were found to be ca. 350 and 430 mol./L when it was

Nutrients 2020, 12, 974 12 of 49
thermally-stressed at 190 °C for prolonged 17.5 and 20.0 hr. durations, respectively [29]. These large
differences observed between the HNE contents of fried potato chips and the oils in which they are
fried are presumably explicable by the higher reactivity of 4-hydroxy-trans-2-alkenals with HNE-
scavenging potato chip biomolecules than that of trans-2-alkenals and n-alkanals, and/or an enhanced
level of their degradation or further oxidation therein when expressed relative to those for the other
aldehyde classes detectable. Human exposure to 4-hydroxy-trans-2-alkenals in vegetable frying oils, fish and shellfish in
Korean diets has been previously assessed using GC/MS/SIM as an analytical strategy, along with
National Health and Nutrition Survey data to evaluate dietary intake patterns [30]. From these
results, the combined HNE and HHE exposure was estimated to be only 16.1 µg per day
(approximately 75% of which was HNE), i.e. 0.3 g/kg for a mean Korean human body weight of 60
kg. Notwithstanding, the risks posed to humans could not be determined, despite the known
toxicological actions of these aldehydes. However, on consideration of their basal tissue
concentrations, the researchers involved concluded that the dietary availability of such agents may
not present a significant human health risk.
2.5. Risk assessments of aldehyde intake in humans: Estimated margin of exposure (MOE) values
Acceptable daily intake (ADI) is a very important parameter for the evaluation of risks to
humans presented by dietary and environmental toxins, for example, and represents the maximum
amount of a chemical substance that can be ingested on a daily basis throughout an entire lifetime
with no appreciable health risk. For food additives or contaminants, this ADI parameter is usually
computed and then employed to determine its risk status via comparisons of it to mean and
associated confidence interval values for estimated human exposure and/or intake levels. However,
for food contaminants and additives, the ADI may also be termed the tolerable daily intake value
(TDI) value.
ADI values are usually obtained from the lowest no observed effect level (NOAEL), which is
derived from long-term in vivo animal model investigations. Hence, such ADI indices arise from the
application of a safety or uncertainty factor to the NOAEL value of the most sensitive testing species.
This safety factor, which is most commonly 100, is applied in view of the requirement to allow for
‘between-species’ differences and variabilities, and also those featured in their toxicokinetic and
toxicodynamic properties. As an example, and for the purpose of comparative evaluations with
dietary aldehydes, in 2010 Tardiff et. al. [31] performed a safety evaluation of ingested acrylamide
using a ‘state-of-the-art’ physiologically-based toxicokinetic model, and TDI (ADI) values for this
food toxin was found to be 40 µg/kg BW per day (equivalent to 2.8 mg for a 70 kg BW human), but
for cancer only 2.6 and 16 µg/kg BW per day for this agent and its glycidamide metabolite respectively
(equivalent to only 182 µg and 1.12 mg/day respectively for a mean 70 kg BW human). The margin
of exposure (MOE) values (equation 1) of aldehydes, and LOPs in general, should be employed in
risk determinations, since these consider the benchmark dose lower confidence limit (BMDL10), a
parameter which represents the lower 95% confidence interval limit of the amount (dose) of an
aldehyde to give rise to the occurrence of a toxic effect when expressed relative to that of a control:
MOE = BMDL10 (µg/kg BW/day)/EDI (µg/kg BW/day) (1)
For acrolein, acetaldehyde and formaldehyde, these BMDL10 values are 360 [32], 5,600 [33] and
2,800 µg/kg BW/day [34] respectively. These values have been previously documented by Ferreira et.
al. [35], and Peterle et. al. [36].
On the basis of these figures, estimated MOE values are 360/33.6 = 10.7 for acrolein; 5,600/137 =
40.9 (Europe) and 5,600/274 = 20.4 (USA) for acetaldehyde; and 2,800/(21–200) = 14 to 133 for
formaldehyde. EDI values for acrolein, acetaldehyde and formaldehyde were obtained from Wang
et. al. [19], [37] and [38] respectively. MOE values which are lower than a value of 10,000 indicate a
potential WHO-defined health risk [39].
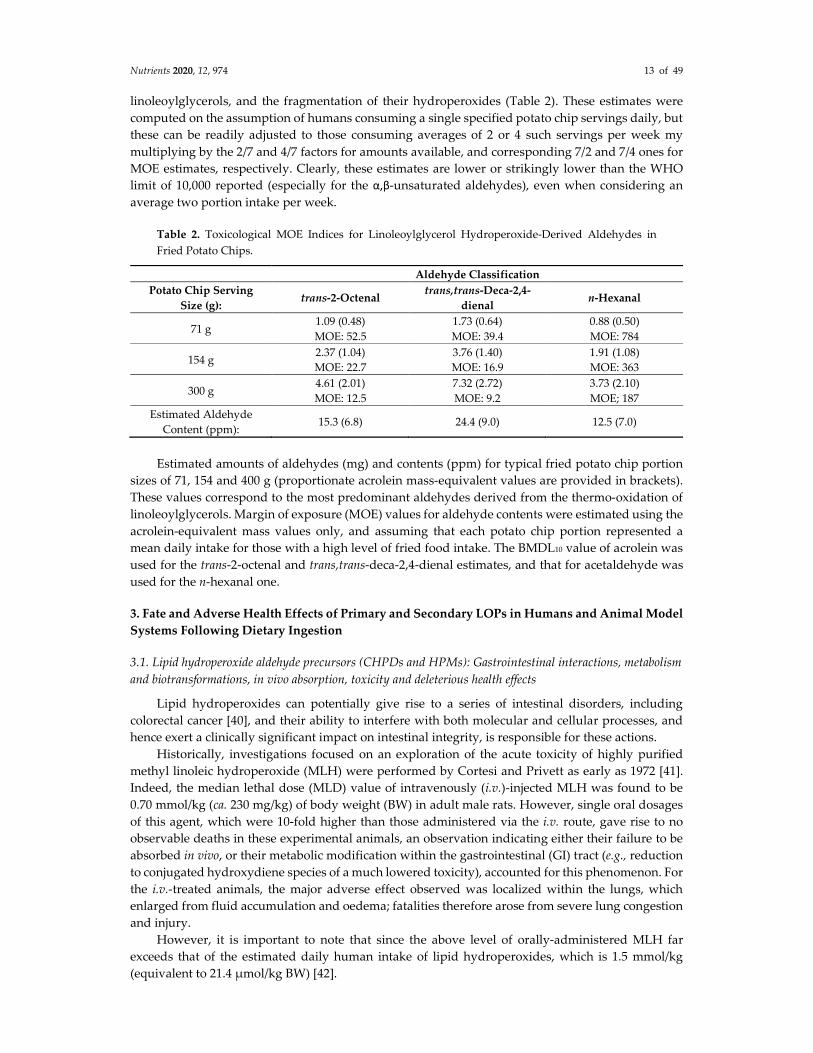
For the purpose of this Commentary paper, we have also estimated MOE values for the potato
chip contents of the most predominant aldehydes derived from the thermo-oxidation of
Nutrients 2020, 12, 974 13 of 49
linoleoylglycerols, and the fragmentation of their hydroperoxides (Table 2). These estimates were
computed on the assumption of humans consuming a single specified potato chip servings daily, but
these can be readily adjusted to those consuming averages of 2 or 4 such servings per week my
multiplying by the 2/7 and 4/7 factors for amounts available, and corresponding 7/2 and 7/4 ones for
MOE estimates, respectively. Clearly, these estimates are lower or strikingly lower than the WHO
limit of 10,000 reported (especially for the α,β-unsaturated aldehydes), even when considering an
average two portion intake per week.
Table 2. Toxicological MOE Indices for Linoleoylglycerol Hydroperoxide-Derived Aldehydes in
Fried Potato Chips.
Aldehyde Classification
Potato Chip Serving
Size (g): trans-2-Octenal
trans,trans-Deca-2,4-
dienal n-Hexanal
71 g 1.09 (0.48)
MOE: 52.5
1.73 (0.64)
MOE: 39.4
0.88 (0.50)
MOE: 784
154 g 2.37 (1.04)
MOE: 22.7
3.76 (1.40)
MOE: 16.9
1.91 (1.08)
MOE: 363
300 g 4.61 (2.01)
MOE: 12.5
7.32 (2.72)
MOE: 9.2
3.73 (2.10)
MOE; 187
Estimated Aldehyde
Content (ppm): 15.3 (6.8) 24.4 (9.0) 12.5 (7.0)
Estimated amounts of aldehydes (mg) and contents (ppm) for typical fried potato chip portion
sizes of 71, 154 and 400 g (proportionate acrolein mass-equivalent values are provided in brackets).
These values correspond to the most predominant aldehydes derived from the thermo-oxidation of
linoleoylglycerols. Margin of exposure (MOE) values for aldehyde contents were estimated using the
acrolein-equivalent mass values only, and assuming that each potato chip portion represented a
mean daily intake for those with a high level of fried food intake. The BMDL10 value of acrolein was
used for the trans-2-octenal and trans,trans-deca-2,4-dienal estimates, and that for acetaldehyde was
used for the n-hexanal one.
3. Fate and Adverse Health Effects of Primary and Secondary LOPs in Humans and Animal Model
Systems Following Dietary Ingestion
3.1. Lipid hydroperoxide aldehyde precursors (CHPDs and HPMs): Gastrointestinal interactions, metabolism
and biotransformations, in vivo absorption, toxicity and deleterious health effects
Lipid hydroperoxides can potentially give rise to a series of intestinal disorders, including
colorectal cancer [40], and their ability to interfere with both molecular and cellular processes, and
hence exert a clinically significant impact on intestinal integrity, is responsible for these actions.
Historically, investigations focused on an exploration of the acute toxicity of highly purified
methyl linoleic hydroperoxide (MLH) were performed by Cortesi and Privett as early as 1972 [41].
Indeed, the median lethal dose (MLD) value of intravenously (i.v.)-injected MLH was found to be
0.70 mmol/kg (ca. 230 mg/kg) of body weight (BW) in adult male rats. However, single oral dosages
of this agent, which were 10-fold higher than those administered via the i.v. route, gave rise to no
observable deaths in these experimental animals, an observation indicating either their failure to be
absorbed in vivo, or their metabolic modification within the gastrointestinal (GI) tract (e.g., reduction
to conjugated hydroxydiene species of a much lowered toxicity), accounted for this phenomenon. For
the i.v.-treated animals, the major adverse effect observed was localized within the lungs, which
enlarged from fluid accumulation and oedema; fatalities therefore arose from severe lung congestion
and injury.
However, it is important to note that since the above level of orally-administered MLH far
exceeds that of the estimated daily human intake of lipid hydroperoxides, which is 1.5 mmol/kg
(equivalent to 21.4 µmol/kg BW) [42].

Nutrients 2020, 12, 974 14 of 49
In an earlier study [43], daily i.v. injections of a more realistic, lower dose of MLH (50 mg/day),
or its continuous infusion at a rate of 206 g/min., to experimental rabbits, was found to markedly
diminish their -TOH stores. Following 10–14 days, the injected animals displayed significant fatty
degeneration and necrosis of the liver, along with creatinuria, and an accelerated muscular
incoordination over those of an untreated control group. Although the creatinuria was circumvented
by the oral administration of very high doses of -TOH (100 mg/day), glutathione peroxidase-
replenishing selenite exerted no blocking influence on both creatinuria and liver lesion incidence. For
the MLH-infused group of animals, an elevated erythrocyte fragility and substantial creatinuria were
observed. Therefore, chronic administration of these low MLH doses gave rise to a rapid
consumption/degeneration of endogenous -TOH, together with an increased incidence of deficiency
symptoms for this antioxidant.
A further study explored the metabolic transformations of orally-administered lipid
hydroperoxides in carp both in vitro and in vivo [44]. Analysis of methyl ester reaction product
derivatives arising from equilibration of 13-hydroperoxy-cis-9,trans-11-octadecadienoic acid with
carp ‘acetone powder’ in vitro revealed that methyl 13-oxo-cis-9,trans-11-octadecadienate, methyl 13-
hydroxy-cis-9,trans-11-octadeca-dienoate, methyl 11-hydroxy-trans-12,13-epoxy-9-cis-octadecenoate,
and methyl 9-hydroxy-trans-12,13-epoxy-trans-10-octadecenoate were the four major metabolites
identified, i.e. one oxodiene and one hydroxydiene species from redox routes, and two hydroxy-
epoxy acids. Oral administration of U-14C-labeled MLH to carp at a level of 0.10 mL/100 g, equivalent
to a very high dose of 2.68 mmol./kg, demonstrated that the predominant metabolites found in
selected organs were hydroxy-octadecadienoate and oxo-octadecadienoate, with ca. 8% of the dosed 14C radiolabel remaining in the body following a 24 hr. period. Since hydroperoxy-octadecadienoates
were found to be absent from carp organ lipid profiles, these data indicate that linoleate-derived
CHPDs (the most common dietary CHPDs) are firstly intestinally redox-transformed to their
corresponding hydroxy- and oxo-adducts, and are then absorbed into the fish circulatory system
where they have access to essential organs and tissues. Hence, these observations suggest that
although CHPDs are not absorbed in vivo, their intestinal primary redox metabolism products are, at
least in carp. These observations are supported by further experimental animal system investigations
described below.
Additionally, Kanazawa and Ashida [45] studied the catabolic fate of linoleic acid
hydroperoxide in a rat GI system in order to determine the molecular nature of LOPs derived from
this source, and which are absorbed into the systemic circulation. Low, albeit perhaps dietarily-
relevant concentrations of this primary LOP (6.5 or 18 mol/L) failed to penetrate into the intestines
(presumably because of its rapid biochemical consumption prior to reaching this site), whereas
higher doses (200 or 800 mol/L) partially leached into this environment. In 14C radiolabel
investigations, products generated therefrom comprised conjugated hydroxydienes (~ 4%), epoxy-
ketones (~ 10%), aldehydes (~ 2.4%), and ~ 13% unidentified 14C-labelled species, along with 27% of
the unmodified peroxide substrate. However, gastric tissue took up 25% of the label, and ca. 6% was
found in the intestinal lumen and tissue as degraded aldehydes. Administration of an aldehyde
mixture dose gave rise to the accumulation of significant amounts of HNE (Section S5), and the less
toxic saturated aldehyde n-hexanal in the liver after a 15 hr. duration (both these aldehydes are
known to specifically arise from the fragmentation of linoleic acid hydroperoxides). Therefore,
evidence for the degradation of linoleic acid hydroperoxide to aldehydes in the stomach was
provided by this study, and the researchers involved concluded that such secondary aldehydic LOPs
are partially absorbed into the circulation.
Fortunately, the human intestinal system is set up with a battery of defense mechanisms to
counter the toxicological onslaught of CHPDs from both endogenous and dietary sources, along with
other ROS. Such defense barriers include peroxide-scavenging catalase, superoxide dismutase (SOD)
and, most importantly for lipid hydroperoxides, the hydroperoxide-neutralising electron-donor thiol
compound glutathione (GSH) and its peroxidase enzymes (GPx) [46–48]. Intriguingly, the GI tissue
network is the only one which has the ability to express all four classes of GPx enzymes
simultaneously, and the sole expression of GI-GPx in this system has indicated that it may be

Nutrients 2020, 12, 974 15 of 49
exclusively targeted to protect against the adverse in vivo absorption of dietary lipid hydroperoxides,
and peroxides in general [49].
Interestingly, GI GPx blocks the shuttling of lipid hydroperoxides in CaCo-2 cells [50], which are
of much value to intestinal absorption investigations since they differentiate to form a polarized
epithelial cell monolayer serving as a physical and biochemical hurdle to low-molecular-mass
molecules and ions. Kanner and Lapidot [51] were the first to demonstrate that ingested PUFAs were
peroxidised within the gut, and for this purpose they investigated free radical-mediated processes
taking place in the stomach’s acidotic environment which could, in principle, promote the generation
of CHPDs from these precursors, along with the concomitant oxidation of further dietary substrates.
Their results suggested that human gastric fluid serves as a highly appropriate ‘bioreactor’ matrix for
accelerating the peroxidation of dietary PUFAs and additional dietary constituents, and also the
potential harmful actions of ingested CHPD LOPs. Moreover, they also found that such localized
stomach-based oxidation was completely suppressed by the inclusion of plant-derived dietary chain-
breaking antioxidants, an observation which demonstrates the protective actions of such agents, and
their beneficial health effects in vivo.
Similarly, Tullberg et. al. [52] explored the oxidation of cod liver oil lipids during GI digestion,
using models involving standardised digestion protocol-matched human digestive juice, and porcine
bile and digestive juice media; fish oil mixed with water at a level of 0.13 mg/mL was employed as
an initial meal. Malondialdehyde (MDA), HNE and 4-hydroxy-trans-2-hexenal (HHE) were analysed
in these digests (using liquid chromatography/atmospheric pressure chemical ionization-mass
spectrometry), as were free fatty acids (FAs) by gas chromatography-mass spectrometry (GC-MS); HHE
specifically arises from the peroxidation of -3 FA sources. Results acquired showed that although
aldehydic LOP generation was low during gastric digestion, it was enhanced in the duodenal digestive
process. Aldehyde generation was accelerated when using human digestive juices over that found using
the porcine system. Free FA liberation was only detectable during the intestinal phase of the protocol, and
this parameter attained values of up to ca. 30%.
Interestingly, stable hydroxymonoenes and conjugated hydroxydienes generated from the GI-
based reduction of lipid hydroperoxides are also available in the human diet, and, like aldehydes,
can also be absorbed from the gut into the systemic circulation [53]. Therefore, there is no shortage of
controversy regarding their measurement in biofluids and tissues as biomarkers of ‘oxidative stress’
in vivo.
However, despite these considerations, it appears that fried foods, which represent one major
source of dietary LOPs, contain little or no lipid hydroperoxide precursors of aldehydes [12], which
as noted above also exert a range of toxicological effects when administered via the i.v. route in animal
model studies [41], and also in in vitro evaluations. Such secondary aldehydic LOPs are more stable
than CHPDs and HPMs when introduced into complex food and biological matrices (the latter
including human biofluids and tissues), which both contain relatively high levels of many LOP-
reactive scavenging agents, including hydroperoxide function-reducing electron donors, and
aldehyde-consuming amine and thiol functions present in a wide variety of biomolecules of both
low- and high-molecular-mass. Moreover, in vivo, enzymes available for the redox interconversion of
hydroperoxides to hydroxydienes, along with the oxidation and/or reduction of aldehydes to their
corresponding carboxylic acid and alcohol adducts, respectively, are readily available. Additionally,
such aldehydes may reversibly react with food alcohols and/or carbohydrates to from hemiacetals
and acetals.
3.2. Secondary aldehydic LOPs: Dietary ingestion, gastrointestinal fate, in vivo absorption, metabolism and
toxicological effects
The molecular nature, toxicities and health hazards potentially presented by aldehydic LOP
toxins have been previously explored in some detail, as have analytical strategies available for their
determination and monitoring, e.g. in fried food sources, and human/animal biofluids and tissues,
for probing their in vivo absorption, biodistribution, metabolism and urinary excretion (an example
of the 1H NMR analysis of aldehydes, specifically LOPs and vanillin, in a typical non-fried food
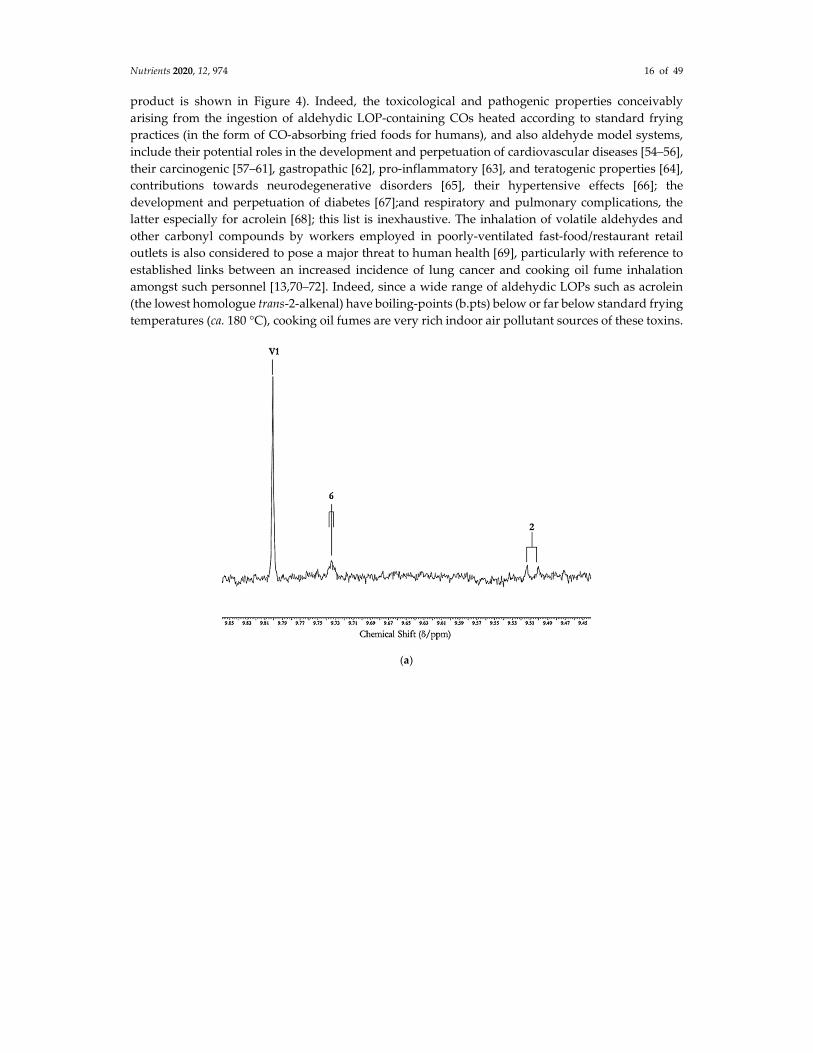
Nutrients 2020, 12, 974 16 of 49
product is shown in Figure 4). Indeed, the toxicological and pathogenic properties conceivably
arising from the ingestion of aldehydic LOP-containing COs heated according to standard frying
practices (in the form of CO-absorbing fried foods for humans), and also aldehyde model systems,
include their potential roles in the development and perpetuation of cardiovascular diseases [54–56],
their carcinogenic [57–61], gastropathic [62], pro-inflammatory [63], and teratogenic properties [64],
contributions towards neurodegenerative disorders [65], their hypertensive effects [66]; the
development and perpetuation of diabetes [67];and respiratory and pulmonary complications, the
latter especially for acrolein [68]; this list is inexhaustive. The inhalation of volatile aldehydes and
other carbonyl compounds by workers employed in poorly-ventilated fast-food/restaurant retail
outlets is also considered to pose a major threat to human health [69], particularly with reference to
established links between an increased incidence of lung cancer and cooking oil fume inhalation
amongst such personnel [13,70–72]. Indeed, since a wide range of aldehydic LOPs such as acrolein
(the lowest homologue trans-2-alkenal) have boiling-points (b.pts) below or far below standard frying
temperatures (ca. 180 °C), cooking oil fumes are very rich indoor air pollutant sources of these toxins.
(a)

Nutrients 2020, 12, 974 17 of 49
(b)
(c)

Nutrients 2020, 12, 974 18 of 49
(d)
(e)
Figure 4. 600 MHz 1D 1H and 2D 1H-1H correlation spectroscopy (COSY) NMR spectral profiles of a
C2HCl3 extract of a commercially-available chocolate hazelnut spread product. (a) Expanded 9.40–
9.90 ppm region of a 1D spectrum of this extract showing an intense –CHO function resonance arising
from the flavouring agent vanillin (abbreviated V1), along with 1H-NMR-detectable traces of
trans,trans-2,4-alkadienals (2) and long-chain n-alkanals (6). (b) Expanded 5.655–6.670 (F1 axis) and
9.364–9.660 ppm (F2 axis) region of a 1H-1H COSY spectrum acquired on this extract, revealing
connectivities between the C1-CHO and C2-CH=CH- resonances of trans,trans-2,4-alkadienals. (c)
Expanded 2.230–2.670 (F1 axis) and 9.668–9.797 ppm (F2 axis) region of the 1H-1H COSY spectrum
shown in (b), showing differential molecular couplings between one major (A) and one relatively
minor (A1) long-chain n-alkanal species. (d) and (e), Expanded 5.7–8.2 and 3.4–4.2 ppm regions of the
1D spectrum shown in (a) respectively, with resonances ascribable to the C5H/C6H (V2) and C2H
(V3) aromatic, and C3-OCH3 (V4) protons of vanillin indicated. DV represents a tentative assignment
to the C3-OCH3 function of divanillin, a vanillin oxidation product. Further abbreviations: -OOH,
lipid hydroperoxide-OOH function resonance; CHCl3, residual chloroform; X, residual chloroform 13C satellite.

Nutrients 2020, 12, 974 19 of 49
3.2.1. In vivo absorption of and metabolic/biotransformation routes for aldehydic LOPs
The GI tract is continually exposed to toxic aldehydes, and subsequent to digestion they are
absorbed into the lymphatic system, or directly into the systemic circulation [73]. Indeed, in 1998, our
laboratory demonstrated that typical trans-2-alkenals generated during the thermal stressing of
PUFA-containing frying oils (trans-2-pentenal and -nonenal) are indeed absorbed from the gut into
the systemic circulation in vivo, then metabolised by a process involving the primary addition of GSH
across their electrophilic carbon-carbon double bonds, and finally excreted in the urine as C-3
mercapturate alcohol derivatives, i.e. as N-acetyl-S-(3-hydroxypentyl)-L-cysteine and -(3-hydroxy-
nonyl)-L-cysteine derivatives, respectively, in experimental rats [73]. However, the administered
levels of these aldehydes were as high as 10 and 100 mg/kg. Generation of these metabolites also
involves reduction of their chemically-reactive aldehyde/aldehyde hydrate (-CHO/-CH(OH)2)
functions to primary alcohol species via the actions of hepatic alcohol dehydrogenase. These results
were consistent with the findings made in [50], which provided evidence for the at least partial
absorption of such aldehydes into the circulation.
However, it should also be noted that this study found that at a 16 hr. post-dosing time-point,
approximately 15% of the orally-administered dose of trans-2-nonenal was oxidatively transformed
to its corresponding carboxylic acid metabolite within the stomach [73].
Consistently, following the subcutaneous injection of the simplest trans-2-alkenal acrolein to
rats, N-acetyl-S-(3-hydroxypropyl)-L-cysteine was detected and isolated as a key urinary excretion
product [74], and these results ae also fully consistent with our 1H NMR-based urinary metabolic
screening investigations [73], including the hepatic metabolic reduction of the aldehyde functions to
alcohol derivatives. However, for these experiments, acrolein was administered by the subcutaneous
injection of a 1% (v/v) solution in arachis (peanut) oil into the lumbar region; the vehicle may itself
have served as a source of aldehydic LOPs, especially if allowed to peroxidise during periods of
storage or solution preparation.
In a scientifically elegant and highly informative early study published in 1985, McGirr et. al.
[75] found that a significantly high proportion of dietary MDA is covalently linked to dietary
proteins, and an acid-labile urinary metabolite (the N-acetyl derivative of the lysine-MDA enaminal
N-(2-propenal) lysine) was detectable in experimental rats following oral administration of a serum
albumin-MDA adduct at a level of 2 mg MDA equivalents/kg BW. Furthermore, this compound was
also demonstrated to be a major urinary metabolite of this dialdehyde administered as its sodium
enolate salt via stomach intubation. Elevated concentrations of this metabolite were excreted by rats
fed a diet rich in highly-peroxidisable cod liver oil. However, these researchers were also able to
identify low levels of this metabolite in the urine of fasted rats, and this observation provided
evidence that it is also formed as a product derived from the in vivo peroxidation of PUFAs, in
addition to its ingestion as a dietary LOP (such as those formed during high temperature frying
practices in the human diet), or alternatively, through the prolonged storage of PUFA-containing
foods. Injection of MDA as its sodium enolate salt to fasted animals markedly increased its urinary
concentration, as expected. In view of the acid lability of N-(2-propenal) lysine, it is possible that free
MDA may be liberated from this primary Schiff base product, and perhaps also from more prevalent
dietary protein lysyl residue adducts, in the GI tract (particularly the stomach), so that it may be
ingested into the systemic circulation as a free (non-adducted) agent.
One recent key investigation appears to have resolved the longstanding critical question
regarding whether there is some clinically-significant in vivo absorption of 4-hydroxy-trans-2-
alkenals, potentially one of the most toxic classes of -unsaturated aldehydes available in human
dietary sources [76]. Details of this study are provided in Section S5 (Supplementary Materials).
Since HNE is universally considered to represent a very important secondary LOP, its metabolic
fate has been extensively investigated. An exhaustive review of the roles of 4-HNE in health and
disease is provided in [77], including a detailed evaluation of its metabolic and biotransformation
products. However, important examples of studies of its metabolic fate both in vivo and in vitro are
also provided in section S5 of the Supplementary Materials. Interestingly. HNE-modified proteins

Nutrients 2020, 12, 974 20 of 49
also appear to be key features of metabolic diseases, and hence offer potential to serve as effective
biomarkers for such conditions [78].
3.2.2. Associations between dietary fried food aldehyde concentration patterns and those of human
blood plasma: Potential tracking of dietary LOPS in vivo?
In 2000, Mak et. al. [79] determined a total of 22 individual aldehydes in circulating arterial blood
plasma samples collected from n = 8 patients with congestive heart failure (CHF), along with those
from an equivalent number of age-matched participants with normal left ventricle (LV) function, i.e.
non-CHF controls. Aldehydes were determined via a GC/MS bioanalytical strategy, and these
included long- and short-chain n-alkanals, trans-2-alkenals, 4-hydroxy-trans-2-alkenals, trans,trans-
2,4-alkadienals, MDA and the dietary flavouring agent furfural. Mean plasma concentrations, or
ranges for the mean aldehyde concentration values of specific structural homologues within each
class, are provided in Table 3 for both control and CHF groups, as are full ranges for the individual
sampling values found in n = 36 samples of potato chips collected from fast-food/take-away
restaurant outlets.
Table 3. Mean concentrations, or concentration ranges of these mean values (nmol/L) of aldehydes
determined in the blood plasma of n = 8 patients with congestive heart failure (CHF) and n = 8 age-
matched normal LV function controls by a GC-MS technique (adapted from [79]).
Non-CHF Controls
(nmol/L)
CHF Disease
(nmol/L)
Fried Potato Chips
(μmol/kg)
Long-Chain n-Alkanals (7) 69–573 42–339 19–560
Short-Chain n-Alkanals (1) 67 91 nd
trans-2-Alkenals (4) 106–527 163–874 0–430
4-Hydroxy-trans-2-Alkenals
(4) 33–211 16–434 0.5–2.1 [68]
trans,trans-2,4-Alkadienals
(2) 152–180 148–420 0–443
Malondialdehyde (MDA) 96 101 0–6 *
Furfural 2,450 4,060 nd
The bracketed numbers in the first (molecular classification) column refer to the number of aldehydes
included for each classification specified for the blood plasma samples analysed. Also listed are the
ranges of contents (µmol/kg) found for samples of fried potato chips (or French fries) purchased from
fast-food/take-out restaurants (long- and short-chain n-alkanals, trans-2-alkenals, and trans,trans-alka-
2,4-dienals were determined by our 1H-NMR analysis approach, but those for 4-hydroxy-trans-2-
alkenals are those reported in Ref. [68] using an LC-MS method. However, both 4-hydroxy-trans-2-
alkenals and furfural are also readily 1H-NMR-detectable and quantifiable. * MDA was specifically
determined by a modification of the method outlined in [6], which involved the reaction of
thiobarbituric acid (TBA) with this dialdehyde to form a pink/red chromophoric derivative, but only
subsequent to its relatively specific extraction into an aqueous medium (mean ± SD first extraction
efficacy: 78 ± 2%).
The blood plasma results acquired in [79] demonstrated that CHF patients had significantly
higher levels of total aldehydes, together with a range of unsaturated ones (specifically, trans-2-
alkenals and 4-hydroxy-trans-2-alkenals, the latter including HHE and HNE), and furfural.
Conversely, the normal LV function control group involved had significantly higher levels of n-
alkanals over those of the CHF patients. Furthermore, the dietary flavouring agent furfural was by
far the most predominant aldehyde present, i.e. 37 and 44% of the total aldehydes determined in
control subjects and CHF participants respectively) and was found be significantly upregulated in
the latter. Furfural is not a LOP, but in addition to its potential genotoxic and carcinogenic properties
[73], this food flavourant has been shown to give rise to the accumulation of ROS and cellular damage
in Saccharomyces cerevisiae [80].

Nutrients 2020, 12, 974 21 of 49
However, aldehydes of the 2,4-alkadienal class monitored in these samples only featured
trans,trans-hepta- and trans,trans-2,4-nonadienals, and the only other di-unsaturated aldehyde
monitored was trans,trans-2,6-nonadienal. Moreover, cis- and trans-deca-4-enals were measured as a
combined sum. Additionally, this study was complicated by (1) the very high incidences of
comorbidities in the male participants recruited to it (mean within-group ages ca. 60 years),
specifically diabetes, hypertension, and hyperchloesterolemia in both groupings, and (2) medical
therapies received by them, i.e. β-blockers, nitrates, ACE inhibitors and calcium channel blockers in
both groups, and additionally diuretics in the CHF one. Notably, all vitamin supplements were
withheld from participants for a minimum duration of 7 days prior the study, and all oral medications
were withheld on the morning of the investigation.
From these results, we therefore elected to perform a comparative statistical evaluation of these
blood plasma LOP profiles in terms of the mean molar levels of different classes of aldehydes
determined therein expressed as a proportion of the total LOP-relevant aldehyde concentration found
in the samples analysed, i.e. those within the above control and CHF groups, to those of the same
mean molar ratios of the aldehyde classification contents found in frequently-consumed fried potato
chip samples collected from fast-food restaurants (Table 2 and Table 3), specifically those fried in
commonly-utilized vegetable oil frying media, as noted in Section 2.3 above. The use of molecular
ratio variables for this exercise is, however, quite fortuitous, since they are expected to be less
sensitive to the potential influences of a range of latent generic variables such as participant BMIs,
ages, etc.
For this purpose, blood plasma levels of furfural were excluded from the computation of
proportionate aldehyde contents since it is not a LOP, and nor was it detectable in any of the fried
potato chip samples analysed by 1H-NMR analysis Unfortunately, it was also not possible to compute
the relative proportions of alka-2,4-dienals in the above two blood plasma groups, since trans,trans-
2,4-decadienal, the major trans,trans-2,4-alkadienal arising from the peroxidative deterioration of
linoleoylglycerols (Section S2), was not determined in [79], and neither was HHE, the major 4-
hydroxy-trans-2-alkenal derived from the decomposition of CHPDs generated from the oxidation of
-3 FAs, e.g. -linolenoylglycerols. Short-chain aldehyde concentrations provided in this report were
those for n-butanal only – since this was the only such analyte included, these values were also
removed from the dataset prior to statistical analysis, although they do remain valuable, since such
aldehydes predominantly arise from the peroxidation of -3 FAs [12]. Therefore, each proportionate
aldehyde class considered comprised those of n-alkanals, trans-2-alkenals, 4-hydroxy-trans-2-
alkenals and MDA only, and all proportions computed represented the concentrations of each of
these LOPs divided by the sum total of them, plus those of all possible alkadienals found. In view of
these limitations, results obtained from these comparative evaluations should be treated with some
caution.
The mean relative proportions (ratios) of the concentrations of long-chain n-alkanals:trans-2-
alkenals:4-hydroxy-trans-2-alkenals:MDA in these three groups of samples were compared and
statistically tested for any significant differences between them. Expressed as percentages of the total
aldehydes detectable (minus contributions from furfural), these ratios were: 40:31:0.20:0.60 for fried
potato chips (mean percentages for a newly-acquired 1H-NMR dataset, n = 36); 46:30:9:2 for normal
LV function (control) subject blood plasma; and 26:39:17:2 for CHF patient blood plasma. Direct
comparison of these proportions for the potato chip sample profiles with those of the control blood
plasma group showed that although the trans-2-alkenal and, to a lesser extent, long-chain n-alkanal
values were quite similar for this comparison, those of 4-hydroxy-trans-2-alkenals were much
elevated in the latter, and these data indicate that, in addition to post-ingestional, aldehyde class-
dependent modifying factors such as differential rates and extents of their absorption, metabolism,
chemical reactivity, protein adduct formation and biodistribution, etc. between each aldehyde class
considered, this aldehyde classification appears to arise from in vivo peroxidation processes.
Moreover, although the proportionate MDA levels remained small for both these groups, such mean
values were elevated approximately 4-fold in the normal LV function blood plasma one.

Nutrients 2020, 12, 974 22 of 49
However, a further major consideration is the dietary availability of all aldehydes considered,
i.e. what proportion of them are ‘free’ and what are constituted as adducts with food proteins (as
noted for MDA [75]), alternative biomacromolecules, or low-molecular-mass nutrient metabolites
such as free amino acids?; such adducts may represent latent sources of these toxins, which may be
liberated within the GI system, for example. Notably, our laboratory determines the ‘free’, non-
adducted form of these aldehydes in fried food products, and hence our estimated values (Table 2
and Table 3) will presumably represent underestimates of the total taken up from COs during frying
practices.
A permutation testing strategy performed via partial redundancy analysis (PRDA) on the log10-
transformed proportionate aldehyde level dataset (involving 104 permutations) revealed that
aldehyde classification-conditioned differences observed between the three sample groups were
statistically significant (p = 0.049), as indeed were those ‘between-aldehyde classifications’ (p = 0.009),
the latter being expected, of course (the log10-transformation was required to counteract within-
sample negative correlations between proportionate/percentage variables). These significant
differences were clearly manifested by 4-hydroxy-trans-2-alkenals and MDA being much greater in
the normal LV function (control) blood plasma profiles over those of fried potato chips. However,
they also arise from the CHF blood plasma group having upregulated proportionate trans-2-alkenal
and 4-hydroxy-trans-2-alkenal levels (over both the control plasma and potato chip serving groups),
and significantly higher proportionate MDA concentrations than the fried potato chip group. This
significant ‘’between-sample group’ effect observed is also explicable by the large differences
observed between the proportionate levels of total n-alkanals between the CHF group and the two
others compared.
Therefore, the observation of very similar fractional aldehyde contents of both n-alkanals and
trans-2-alkenals in the large potato chip and smaller control blood plasma sampling groups may serve
to indicate that such LOPs are dietary-derived. If this is the case, then the in vivo ‘conservation’ of
their proportionate levels may also reflect the overall lower, albeit differential chemical/biochemical
reactivities of these classes of aldehydes than those of 4-hydroxy-trans-2-alkenals and MDA,
following their ingestion by humans. Of particular note, in vivo, n-alkanals serve as substrates for
pyruvate dehydrogenase, but -unsaturated aldehydes are not affected by this enzyme [81].
Moreover, as noted above, unsaturated aldehydes readily take part in Michael addition reactions
with GSH to form their primary detoxification GSH conjugate products [82,83], but n-alkanals clearly
do not (although they may form Schiff base adducts with the terminal amino function of this
tripeptide). However, the rather substantial differences observed between these two groups’
proportionate 4-hydroxy-trans-2-alkenal and MDA concentrations certainly indicate, but do not
confirm, that such toxins may be generated from in vivo lipid peroxidation processes.
Proportionate total 4-hydroxy-trans-2-alkenal levels in the CHF blood plasma group were also
significantly greater than those of the normal LV function control group (ca. 2-fold), but this
observation was reversed for long-chain n-alkanal concentrations, the latter results being consistent
with our proportionate levels estimated in fried potato chips. The markedly elevated proportionate
value of CHF blood plasma levels of the former class of aldehydes may have been expected in view
of an enhanced level of in vivo oxidative stress associated with this condition. However, if this was
the case, why was it that the mean level of furfural, a non-LOP dietary flavouring agent, was also
significantly increased from 2.45 µmol/L in the control group to 4.06 µmol/L in the CHF one (p <
0.01)? Possibly these differences are also partially explicable by differing dietary regimens between
these two groups, perhaps an increased level of aldehyde-loaded fried food consumption and/or an
enhanced furfural intake in the latter (this flavouring agent is readily absorbed subsequent to
administration by any route [84])? Study participants were not fasted for a minimum required period
prior to the collection of blood samples in this study, so it certainly appears that such aldehydes may
at least partially arise from such dietary, or perhaps alternative exogenous sources. Further possible
limitations of the study reported in [79] are detailed in Section S6 (Supplementary Materials).
However, the authors of [79] also suggested that differences in the aldehyde profiles between
their two groups may arise from those between the FA compositions of their diets [73], i.e. with a

Nutrients 2020, 12, 974 23 of 49
possible higher peroxidised PUFA and therefore aldehyde content of those received by the CHF one,
but they also indicated that such systematic dietary variations between them were unlikely.
In the control group of participants, the order of decreasing total blood plasma total trans-2-
alkenal concentrations were (peroxidised FA source(s) in brackets, with L, -Ln, -Ln, O, Po and Ar
representing predominant linoleoyl-, -linolenoyl-, -linolenoyl-, oleoyl-, palmitoleoyl- and
arachidonylglycerols respectively) trans-2-octenal (L) > trans-2-hexenal (-Ln - minor aldehydic LOP,
but also a major dietary flavouring agent (section 3.1)) > trans-2-heptenal (O and L) > trans-2-nonenal
(Po, Ar and -Ln, but also a food flavouring agent); that for n-alkanals was n-heptanal (L) > -nonanal
(O) > -octanal (O) > -hexanal (Ar/L, but also derived from the decomposition of trans,trans-2,4-
decadienal [11]); and that for 4-hydroxy-trans-2-alkenals was HHE (-Ln) > HNE (L and Ar) >>> 4-
hydroxy-trans-2-decenal (L) ≈ 4-hydroxy-trans-2-octenal (unknown peroxidised FA sources). As
expected, for the 4-hydroxy-trans-2-alkenals determined, the most predominant ones were those
arising from the sequential peroxidation of α-linolenoyl- (HHE) and linoleoyl-/arachidonoylglycerols
(HNE).
Using the somewhat broad assumption that a highly significant fraction of at least some of these
blood plasma aldehyde levels arise from dietary sources, it should be considered that those found
therein represent only residual concentrations, i.e. what remains following their in vivo consumption
through their metabolic fate in the GI system, in vivo absorption and then further metabolism
thereafter in organs such as the liver, along with any biotransformation of them in human blood
plasma and other environments, e.g. the generation of protein carbonyl species from the reaction of
-unsaturated aldehydes with plasma proteins such as human serum albumin and gamma-
globulins, and additional Schiff base products arising from the reactions of all possible aldehydes
with free primary and secondary amine functions present in selected biomolecules, for example.
‘Between-aldehyde class’ differences in the rates and extents of their consumption will also account
for those observed between their relative blood plasma concentrations. In principle, since -
unsaturated aldehydes are more chemically-reactive than saturated ones [82], should we perhaps
expect higher n-alkanal:trans-2-alkenal ratios in human blood plasma than what is found in fried food
products? Indeed, this ratio is already significantly > 1 in fried potato chip samples (mean ± SEM 1.39
± 0.10 for our dataset (Table 2); 95% confidence intervals 1.18-1.61, p < 0.01) than it is in the culinary
oil sources of these aldehydes, and this has been attributed to their differential levels of reactivity
with potato proteins, amino acids and further biomolecules between these two classes of LOP toxins
[3,14]. This was indeed the case in the above normal LV function group plasma profiles explored, the
ratio being 1.53; however, this difference observed was found not to be statistically significant from
that found in fried potato chips (one sample t-test). Notwithstanding, it may be conjectured that these
proportionately lower control group blood plasma trans-2-alkenal levels may also arise from a higher
level of reactivity of them in vivo. Interestingly, the fractional blood plasma MDA aldehydic LOP
content in these control participants (only 2%) was found to be ca. 3-fold greater than that observed
in potato chip samples (0.60%). Nevertheless, this observation again confirms that MDA represents
only a minor secondary LOP.
Overall, also important is the observation that the total unsaturated aldehyde content of normal
LV function patient blood plasma is significantly greater than that of saturated aldehydes, and this is
indeed also the case for estimated weight percentage human dietary intakes of these toxins by
humans in Refs. [19,85,86], i.e. an unsaturated:saturated aldehyde ratio of 5:2 in mg/kg units [85,86].
Corresponding weight percentage (ppm) values for fried potato chip samples were found to be a very
similar value of ca. 3:1 [12].
A further study performed by Ogihara et. al. in 1999 [87], which determined the blood plasma
concentrations of secondary aldehydic LOPs in premature infants with and without chronic lung
disease (CLD), was, however, limited to only 3 long-chain n-alkanals and 4 trans-2-alkenals, together
with HNE. Full details of this study are available in Section S7 of the Supplementary Materials.
However, it is clear that further research investigations targeted on dietary patterns, human
intake, GI fate, absorption, biodistribution and further metabolism of such dietary LOPs are required
in order to ratify potential relationships between their dietary availabilities and those detected in

Nutrients 2020, 12, 974 24 of 49
human biofluids and tissues. Although not simply conceivable for all patient and age-matched
control groups investigated in human trials, it is also thoroughly recommended that for future clinical
studies focused on explorations of oxidative stress in vivo (particularly the in vivo generation of LOPs
such as reactive aldehydes), all participants involved should be fasted for a sufficient minimal time
period prior to the collection of biofluid or biopsy samples for analysis. Such an approach will
presumably overcome any interferences or confounding effects arising from dietary LOP sources.
4. Atherosclerosis and its Cardiovascular Disease Sequelae
Covalent structural modification of lysyl and further selected amino acid residues of the apo-B
protein composite of low-density-lipoprotein (LDL) by various aldehydes (including trans-2-
alkenals, 4-hydroxy-trans-2-alkenals, acrolein and malondialdehyde, but not exclusively so) affords
its uptake by macrophages to form foam cells, which in turn give rise to artery-blocking fatty streaks.
Indeed, Staprans et. al. [56] discovered that feeding an oxidized lipid-rich diet to New Zealand white
rabbits culminated in a 100% increase in fatty streak lesions within the aorta over those fed an
unoxidized lipid control diet. Interestingly, rabbits receiving the oxidized lipid diet were found to
have a >100% increase in total cholesterol in the pulmonary artery (predominantly as cholesteryl
esters). HNE- and MDA-modified proteins have been previously identified in atherosclerotic lesions
using immunological methods and techniques (reviewed in [88]). Such aldehydes, which can also be
generated in vivo, are also implicated in a range of pathologies arising from or linked to
atherosclerosis, such as their cardiovascular disease sequelae, and complications arising from poorly-
controlled types 1 and 2 diabetes.
Major aldehydic LOPs associated with the chemopathology and pathobiology of atherosclerotic
oxidant injury have included trans-2-alkenals, 4-hydroxy-trans-2-alkenals such as HNE, MDA and 4-
ketoaldehydes [86], although it should be noted that the most predominant ones present in fried food
sources are trans-2-alkenals, trans,trans-2,4-alkadienals and n-alkanals (section 3.1 [3,14]). Moreover,
those arising from dietary sources also include 4,5-epoxy-trans-2-alkenals, cis,trans-alka-2,4-dienals
and hydroperoxy- and hydroxy-trans-2-alkenals, for example. As delineated above, all these
aldehydes readily react with proteins to form relatively stable protein-aldehyde adducts via Schiff
base and Michael addition reactions, and these potential biomarker species may be identified and
determined in in vitro models involving LDL, and in biosamples collected from animals in models of
atherosclerosis, together with those from human patients with potentially enhanced risks, or clinical
signs and symptoms of this disorder. Moreover, some aldehydes have been shown to induce
intracellular oxidative stress assaults, and also activate stress signaling pathways that exert effects on
cellular responses to extracellular stimulating agents [89].
Earlier investigations, which focused on exploring the pathological roles of aldehyde-modified
proteins have demonstrated that LDL-aldehyde adducts have an enhanced recognition by
macrophages, and the uptake of these species is therefore significantly increased in these cells [90].
Moreover, Steinberg et. al. [91,92] first recognized that aldehyde adducts of the apolipoprotein B (Apo
B) component of LDL transforms this lipoprotein to a pro-atherogenic form which is readily taken
up by macrophages to generate foam cells. Further studies which focused on the Apo B moiety of
‘oxidised’ LDL, featured MDA as Nε-(2-propanal)-lysine adducts [93] and 1-amino-3-iminopropene
MDA-lysine cross-links [94]; acrolein adducts, including N-(3-methylpyridinium) lysine [95] and 3-
formyl-3,4-dehydropiperidine species [96,97]; and HNE-derivatized adducts, e.g. enaminal class
HNE-histidine and HNE-lysine species [98].
More recently, Tamamizu-Kato et. al. [99] demonstrated that acrolein markedly impaired the
functional integrity of Apo E, an exchangeable anti-atherogenic apolipoprotein when present at a 10-
fold molar excess, along with heparin-, lipid- and lipid-receptor-binding; experiments were
performed using recombinant Apo E, and immuno-blotting employing an acrolein-lysine-specific
antibody. These studies are fully consistent with the detection of acrolein in atherosclerotic lesions
[96], and acrolein-modified LDL was also found to induce the generation of foam cells from
macrophages [100].

Nutrients 2020, 12, 974 25 of 49
Furthermore, it has been demonstrated that reactive aldehydes suppress mitochondrial
respiration [101], modify ion-channel conductance pathways [102], and diminish myofilament
sensitivity and cardiac contraction [103]. Intriguingly, Wang et. al. [19] found that acrolein effectively
propagates myocardial ischaemic injury and suppresses nitric oxide (NO●)-induced cardioprotection
in mice by a mechanism involving attenuation of protein kinase Cϵ (PKCϵ) signal transduction. In
2011, Ishmail et. al. [104] found that long-term oral exposure to acrolein, at a level consistent with the
human intake of unsaturated aldehydes, gave rise to a dilated cardiomyopathy phenotype in C57BL/6
mice, and from these studies concluded that corresponding effects in humans may be induced by
their exposure to this aldehyde. Therefore, human exposure to environmental/dietary sources of
acrolein and other α,β-unsaturated aldehydes may provide a rational foundation for heart failure.
Since acrolein induces myotube atrophy in vitro, and acrolein-inhalable cigarette smoking serve
as major risk factors for skeletal muscle deterioration (atrophy), a very recent investigation [105]
focused on the mechanism of this phenomenon discovered that low doses of this aldehyde
significantly inhibited myogenic differentiation in vitro, a process which may occur through
suppression of the serine-threonine protein kinase (Akt) signalling pathway. Mice with or without
glycerol-induced muscle injury were exposed to 2.5 and 5 mg/kg BW/day acrolein in distilled water
via the oral route for 4 weeks in order to investigate its effects on muscle wasting and regeneration.
Acrolein’s ability to induce muscle wasting was confirmed in this animal model system, and muscle
regeneration was also found to be retarded. Hence, these data are fully consistent with acrolein’s
potential role in the pathogenesis of myogenesis and disease-linked myopathy. At the cellular level,
exposure to acrolein exerts a wide range of toxic effects, e.g. membrane damage, immune
dysfunction, endoplasmic reticulum stress, and mitochondrial disruption, along with oxidative stress
and protein and DNA adduction [106].
Relationships between the consumption of fried foods in the diet and the risks of cardiovascular
and related diseases have recently been extensively reviewed by Gadariju et. al. [107], to which
readers are referred for further information. This review outlines current evidence available on
associations between human cardiovascular diseases, hypertension, diabetes and obesity, and
estimates of the fried food consumption of population cohorts. However, data acquired in this survey
of many publications focused on this topic were predominantly based on questionnaires to capture
fried food intake information, and the study experimental designs involved were limited to case-
control and cohort investigations. However, on the basis of this (Ref. [107]) review, there is convincing
evidence available to support clear linkages between the risks of these non-communicable chronic
diseases and an increased frequency of fried food consumption, i.e. ≥ 4 times per week.
In the context of metabolic syndrome cluster conditions, further investigations have focused on
the effects of diets containing high contents of oxidised frying oils on the development and/or
progression of type 2 diabetes, and those conducted by Chiang et. al. [67] have demonstrated that
such diets can give rise to lowered levels of insulin secretion and hence glucose intolerance. The
mechanism for these chemopathological impairments appears to involve an oxidative damage-
mediated alteration of glucose metabolism, a process which affects the secretion of insulin by the
pancreatic islets. However, such effects were shown to be circumvented by -TOH supplementation,
an observation supporting the role of this chain-breaking lipid-soluble antioxidant in consuming
highly toxic primary LOO radical species, which in turn suppresses the degradation of any lipid
hydroperoxides formed to biochemically-reactive aldehydes in vivo.
5. Mutagenicity, genotoxicity and carcinogenicity of secondary aldehydic LOPs, and potentially
their dietary/fried food sources
There is now a long and expansive history of research work which has focused on these area for
more than 40 years or so. Indeed, much valuable information is available in this area, with acrolein,
MDA and HNE being the most widely investigated, although there are some limitations with direct
comparative evaluations of these results in view of the wide diversity of cell lines tested and their
tissular sources. Notwithstanding, recent developments in the areas of epigenetic effects, i.e. histon

Nutrients 2020, 12, 974 26 of 49
modification and DNA methylation, are very encouraging e.g. [108]. LOP genotoxicity is extensively
reviewed in [109], and the mutagenicities of carbonyl compounds was fully established in the 1980’s. Moreover, much evidence currently available indicates that aldehydes act as carcinogens [108–
112], or in some cases at least have variable degrees of carcinogenic potential. Moreover, a working
group of the International Agency for Research on Cancer (IARC) found that aldehyde-containing
emissions arising from high temperature frying episodes are “probably carcinogenic to humans
(Group 2A)” [111]. The study performed in [112] has provided a high level of evidence that oxidative
stress mediated by -unsaturated aldehydes significantly contributes towards cytotoxic and
genotoxic cell damage, and these effects are, of course, critically dependent on the structural nature
of the agents tested in this manner. Feron et. al. [113] performed an overall assessment of the cancer
risk status of a range of dietary aldehydes, and from this concluded that although acetaldehyde,
crotonaldehyde and furfural do represent dietary risk factors, this was not the case for acrolein,
formaldehyde, citral and vanillin. However, it was not possible to evaluate this risk factor for MDA,
glycidaldehyde (an acrolein metabolite), benzaldehyde, cinnamaldehyde and anisaldehyde in view
of unavailability of sufficient data. These researchers also concluded that dietary-sourced aldehydes
should be screened for their mutagenic, cytotoxic and cytogenic activities, and emphasised that such
screenings should be prioritised on the basis of their degree of human exposure and expected
mechanisms of action, the latter of which is now a rapidly expanding field of study. Notwithstanding,
this evaluation review [113] is now very dated, and since that time (1991) there have been major
advances in research information and data available on both the dietary availability of these
aldehydes, their in vivo absorption and biodistribution, along with their mechanisms of action and
target organs, in addition to that based on their exertion of deleterious toxicological and carcinogenic
effects.
Attack of endogenous DNA bases by chemically-reactive aldehydic LOPs can represent very
important aetiologies of cancer and human genetic diseases in general. Indeed, such structurally-
modified DNA adducts arising therefrom can give rise to frameshift mutations [114]. MDA
represents a relatively minor aldehydic LOP arising from the peroxidation of -3 FAs containing ≥ 3
carbon-carbon double bonds such as eicosapentaenoic (EPA) and docosahexaenoic acid (DHA)
acylglycerol derivatives (containing five and six double bond units respectively), and which are
prevalent constituents of marine oils.
As an illustration of the pattern of aldehydic LOPS generated from the peroxidation of -3 FAs,
Figure 5 shows two-dimensional (2D) 600 MHz 1H-1H correlation spectroscopy (COSY) NMR profiles
of a commercially-available sample of cod liver oil which was exposed to a prolonged (90 min.)
episode of thermal stressing at a standard frying temperature of 180oC in order to explore its
peroxidative resistivity. Although CLO products are predominantly employed as dietary
supplements and not for frying or cooking purposes, this exposure represents an extreme for the
oxidation of ω-3 FAs and further PUFAs therein during increasing periods of storage and exposure
to atmospheric O2 at ambient temperature, and is also a more analytically-specific approach than
alternative, high temperature-dependent methods for monitoring the oxidative susceptibility of such
oils products since it has the ability to determine the molecular nature and levels of a wide range of
LOPs simultaneously. Data acquired demonstrates the major advantages offered by this technique,
specifically the ability to distinguish between, and electronically integrate resonances arising from a
variety of aldehydes and aldehyde classes, including acrolein and 4,5-epoxy-trans-2-alkenals.
Moreover, this analytical approach was also able to provide much valuable information regarding
the molecular natures of five or more saturated aldehyde classifications, including both long- and
short-chain ones (the latter including -3 FA hydroperoxide-derived propanal).

Nutrients 2020, 12, 974 27 of 49
(a)
(b)

Nutrients 2020, 12, 974 28 of 49
(c)
Figure 5. (a) 5.7–10.2 ppm regions of single-pulse (1D) and two-dimensional (2D) 1H-1H COSY spectra
of a commercial cod liver oil product exposed to a LSSFE for a period of 90 min. at 180 °C, with 1H
chemical shift scales (ppm) on the F1 (ordinate) and F2 (abscissa) axes. (b) Expanded 5.98–6.47 (F1
axis) and 9.36–9.73 ppm (F2 axis) region of the above 1H-1H COSY spectrum, revealing linkages
between the C1-CHO, C2-CH=CH- (and in some cases C3-CH=CH-) resonances of trans-2-alkenals,
trans,trans-2,4-alkadienals, cis,trans-2,4-alkadienals, acrolein and 4,5-epoxy-trans-2-alkenals (1, 2, 5,
Acr and Epox, respectively). A1 represents a connectivity between the = 9.484 and 6.119 ppm
resonances, and is tentatively assigned to a trans-2-alkenal classification with a significantly different
carbon chain length range than that giving rise to the characteristic 9.480 ppm signal. (c) Expanded
2.33–2.94 (F1 axis) and 9.68-9.91 ppm (F2 axis) region of the 1H-1H COSY spectrum shown in (a),
exhibiting clear distinctions between connectivities arising from three long-chain (A, A1 and B) and
one short-chain (D) n-alkanal classification. C represents the 1H-1H correlation for the -CHO and -
CH2 function protons of 4-oxo-n-alkanals. Samples were prepared for 1H-NMR analysis by the
method described in [11], and spectra were acquired on the NMR facility described in Figure 2.
Notwithstanding, such FAs are only present at relatively low levels in most vegetable-based
COs, which are rich in -6 linoleoyl-, but only limited or deplete in -3 linolenoylglycerols, such as
sunflower oil with ≤ 0.20-0.30% (w/w) [12]. However, relatively higher concentrations of -3 FAs are
present in canola and soybean oils, which have approximately 10% and 7% (w/w) of them
respectively. Linseed oil is a notable exception, but this oil is far too dangerous to employ for standard
shallow- or deep-frying episodes in view of its highly explosive nature, although it is still quite
commonly employed for traditional Chinese wok cooking purposes! Fortunately, Belgium and
France have regulations which sensibly limit the amount of this FA for use in frying oils to only 2%
(w/w) [115], as have some other countries such as Chile.
-3 FAs, e.g. -linolenic acid-containing acylglycerols in rapeseed, soybean and linseed oils, and
those containing EPA and DHA in marine oils, are more susceptible to peroxidation than the
linoleoylglycerols predominant in many vegetable oils, and also give rise to a differential pattern of
aldehydic LOPs following peroxidation (e.g. acrolein, 4-oxo-trans-2-alkenals and low-molecular-mass
n-alkanals such as propanal). Therefore, distinction of their 1 and 2D 1H NMR profiles from those of
-6- and -9-rich vegetable COs is a relatively facile process. As expected, 1H-1H COSY linkages for
acrolein were not detectable in spectra acquired on thermally-stressed samples of sunflower oil (data
not shown). Interestingly, 4-hydroxy-trans-2-alkenals were readily 1H NMR-detectable in thermally-

Nutrients 2020, 12, 974 29 of 49
stressed linoleoylglycerol-rich vegetable oils, together with oleoylglycerol-rich palm oil
(predominantly as HNE, with 1H NMR signals located at = 9.59 (d, C1-CHO), 6.82 (dd, C3-CH=CH-
), 6.31 (dd, C2-CH=CH-) and 4.53 ppm (m, C4-CHOH) [12]), but not in heated marine oils, in which
the corresponding aldehyde generated from fragmentation of such FA hydroperoxides is HHE.
Below, the potential or proven mutagenic, genotoxic and carcinogenic actions of MDA, trans-2-
alkenals in general, 4,5-epoxy-trans-2-alkenals, and acetaldehyde and formaldehyde, are reviewed
and discussed, as are those of acrolein, the latter with special reference to the exposure of humans to
wok cooking episodes. Associations cancer risk and fried food intake levels are also described.
Section S5 (Supplementary Materials) provides an outline of the toxicological properties and
potential adverse health effects of HNE, and its lower HHE homologue.
5.1. MDA
Early studies involving an E. coli mutagenesis system revealed that MDA is indeed mutagenic
in cells which feature active DNA-repair systems, and these results indicated that this aldehyde had
the ability to induce inter-strand cross-linking (fluorescent products were detected from such reaction
systems) [116]. Much later, this dialdehyde was found to react with the DNA base adduct guanine to
form the exocyclic adduct, pyrimido(1,2-)purin-10(3H)-one derivative (M1G) [117], and therefore
when MDA is absorbed in vivo [75], it has the ability to generate this derivative. The M1G adduct has
been detected in selected healthy human tissues, including colorectal mucosa [118], and it induces
sequence-dependent frameshift mutations and base pair substitutions in bacteria and in mammalian
cells. This finding suggests a potential role for the M1G lesion in the induction of mutations commonly
related to human diseases. Another early study [119] found that administration of MDA as it enolate
anion sodium salt (throughout a 0.1–10.0 g/g/day dosage level range) to mice in drinking water for
a duration of 12 months caused dose-related hyperplastic and neoplastic alterations to liver nuclei,
but no gross hepatic tumours were generated. However, addition of MDA to the medium of cultured
rat skin fibroblasts gave rise to nuclear abnormalities at added concentrations of only 1.0 µmol/L,
despite a cellular uptake of only 4%. MDA/(3-hydroxy acrolein) have been shown to exert cancer-
initiating activities in female Swiss mice [120,121].
5.2. trans-2-Alkenals
The mutagenicities of 2-hexenal, -heptenal, -octenal and -nonenal have been previously
evaluated in bacterial systems [122], and each of these was found to exert significant effects at µmol/L
concentrations; since 2-hexenal occurs naturally in a range of foods, it has received particular focus
in such investigations. Testing of these aldehydes, along with 2-pentenal, in V79 Chinese hamster
cells at added levels of 3.0–300 µmol/L demonstrated that all of them gave rise to a dose-dependent
enhancement in 6-thioguanine-resistant mutant frequency effect, which increased with increasing
molecular size [123]. Furthermore, 2-nonenal at doses of only 0.10 and 10 µmol/L was found to give
rise to notable sister chromatid exchanges (SCEs), although no chromosomal aberrations, nor
micronuclei, were observed in these studies. A further report, focused on trans-2-hexenal
detoxification and its DNA adduct formation in humans [124], is also featured in Section S8 of the
Supplementary Materials. However, although estimates of the mean daily intake of trans-2-hexenal
for a ‘normal’ diet are 4.2 mg per mean 70 kg BW human, that for a trans-2-hexenal-rich diet is as
much as 42–147 mg/day [125], a range substantially greater than the 12.5 mg 95th percentile intake
estimate of Ref. [124].
In 2005, Nadasi et. al. [125] evaluated the potential carcinogenicity of trans-2-hexenal, for which
humans have a dietary pattern-dependent continuous intake, and for this purpose monitored Ha-ras
and p53 gene expression alterations, together with tumour development in mice and rats following
its administration. For short-term experiments, this study involved CBA/Ca(H-2K), AKR/J(H-2K) and
C3He-mg(H-2K) mice (6–8 weeks old) and Long-Evans, Wistar and Fischer 344 rats (6 females and 6
males of each strain), each receiving 3 x 50 mg/kg BW trans-2-hexenal in a corn oil vehicle orally (age-
matched controls received the same volume of unspiked corn oil). Animals were autopsied 24, 48 and
72 hr. following administration of the aldehyde or its vehicle alone. However, in a long-term study,

Nutrients 2020, 12, 974 30 of 49
mice and rats received 150 mg/kg body weight of trans-2-hexenal in total intraperitoneally (i.p.), i.e.
50 mg/kg on the 1st, 8th and 15th days of the investigation, and were then autopsied following an 18-
month survival period. Any developed tumours were removed and 5-µm formalin-fixed, and
paraffin-embedded sections were routinely stained by haematoxylin/eosin, and then examined by
light microscopy. In the short-term study, no gene alterations were noted 24–72 hr. post-
administration. However, ca. 14% of the 72 mice and rats within the long-term study were found to
develop malignant tumours at the 18-month time-point follow-up evaluation. Therefore, despite
exerting no effects on the expression of both onco- and suppressor genes, this reactive -
unsaturated aldehyde displayed a significant carcinogenic potential, which is potentially explicable
by its epigenetic effects, i.e. they appear to be non-genotoxic carcinogens; in general, consistencies
between the genotoxic and carcinogenic effects of compounds is only ca. 90% [125]. This may serve
to explain the lack of genotoxic risk found for this aldehyde reported in [124].
A further noteworthy point is that since trans-2-hexenal arises from other food sources such as
fruits, especially bananas [125], its overall total daily human intake is expected to be inflated by a
consideration of fried food consumption, which may sometimes exceed more than one fried meal per
day, and also possibly the human consumption of serving portions of fried food sources of this -
unsaturated aldehyde greater than 154 g. However, this agent is not one of the more predominant
ones derived from the peroxidation of UFAs [12].
A study which reported comparisons of the cytotoxic and mutagenic properties of the natural
product 2-cyclohexene-1-one with those of a range of dietary aldehydes is discussed in section S9
(Supplementary Materials).
5.3. Acrolein and Chinese wok cooking
Early investigations demonstrated that both i.p. and intravesicular administration of acrolein to
rats gave rise to anomalous levels of cellular proliferation and hyperplasia of bladder urothelium and
epithelium [126]. Furthermore, a greater incidence of an abnormal DNA (2-deoxyadenosine)-acrolein
adduct has been found in liver [127], oral [128] and bladder cancers [129]. Further information
regarding the carcinogenic potential of acrolein as an inhaled or ingested toxin are outlined in section
S10 (Supplementary Materials)
One epidemiological study reported in 2013 found an elevated incidence of lung cancer in non-
smoking Chinese women who cooked/fried food at very high temperatures using a traditional
Chinese-style wok process, and hence had a high level of exposure to cooking oil fumes arising
therefrom [130]. Supporting laboratory evidence for this hypothesis was provided by the observation
that these women participants excreted significantly higher creatinine (Cn)-normalised
concentrations of acrolein and crotonaldehyde (the latter the next higher trans-2-alkenal homologue
from acrolein) as their mercapturate metabolites. No differences were found between these groups
for corresponding urinary levels of benzene mercapturate. In their conclusion, the authors therefore
recommended that domestic kitchen proprietors/users should act to alleviate human exposure to
toxic and carcinogenic cooking oil fumes generated during traditional wok cooking styles by
ensuring that these areas are sufficiently ventilated at the sites involved. A similar recommendation
should, of course, also apply to all commercial/restaurant cooking sites, albeit to a greater, more
expansive extent. Highly detailed reviews of the molecular mechanisms featured in acrolein toxicity
is provided in [106] and [131]. Information regarding the potential carcinogenic properties of
trans,trans-2,4-decadienal present in linoleoylglycerol-rich cooking oil fumes is available in section
S11 (Supplementary Materials). A related epidemiological study performed in 2002 [132] is related in
the Supplementary Materials (section S10). Notably, wok frying practices are also expected to
markedly promote the oxidative deterioration of PUFAs, since these stir-frying approaches generally
use only small volumes of oil per food portion (say, 10–40 mL), and therefore the oil surface area is
very large, and exposure to atmospheric O2 is maximised. Moreover, trace catalytic transition metal
ions from fried foods will promote both the peroxidation of UFAs, and also the breakdown of CHPDs
and HPMs to aldehyde fragments, etc. Similarly, adventitious trace metal ions such as Cu(II) derived
from the wok metal alloy material itself may provide catalytic sources for these processes.

Nutrients 2020, 12, 974 31 of 49
Intriguingly, when applied dermally to mice and rats, acrolein’s glycidaldehyde metabolite
exerts carcinogenic properties [106,133]. Additionally, acrolein is a major lung cancinogen present in
cigarette smoke [134]. Vinyl chloride, which is structurally related to acrolein, has been identified as
a carcinogen in both animals and humans [135]. In cell culture experiments, acrolein can exert
cytotoxic properties at concentrations of < 0.1 µmol/L [136].
Major health threats posed by aldehydes such as acrolein and crotonaldehyde present in
cigarette smoke are also worthy of much consideration; however, this aspect of aldehyde toxicology
is beyond the scope of this work. Notwithstanding, it is important to note that reliable estimates of
the amounts of ingestible aldehydes available in single, average-sized servings of fried potato chips
are not too dissimilar to those derived from the smoking of a mean daily allocation of 25 cigarettes
[137].
5.4. Crotonaldehyde
In 1986, Chung et. al. [138] orally-administered crotonaldehyde (the next higher α,β-unsaturated
homologue of acrolein), which is mutagenic without metabolic activation [139], to F344 rats in
drinking water at concentrations of either 0.60 or 6.00 mmol/L for a 113-week period, and
histologically evaluated liver tumours in these groups against an untreated age-matched control one.
At the lower dose level, crotonaldehyde was found to induce neoplastic lesions in the liver in 9/27
rats; a further 9 had neoplastic nodules, and 2 had hepatocellular carcinomas. At the higher dose
level, however, this aldehyde gave rise to severe liver damage in 43% of these animals, with the
remaining 57% developing abnormal liver cell foci. Although results acquired also indicated that
crotonaldehyde was a weaker tumorigen than the established carcinogen N-nitrosopyrrolidine
(NPYR), they provided strong evidence for its carcinogenicity. Indeed, the incidence of liver tumours
in rats treated with crotonaldehyde and NPYR at equivalent doses (0.60 mmol/L) was 87 and 33%
respectively. α,β-Unsaturated aldehyde concentrations of 0.60 and 6.00 mmol/L are not at all
dissimilar to those of total trans-2-alkenals found in vegetable-based culinary oils exposed to high-
temperature frying durations [12]. In fact, the higher level is lower than those typically determined
in repeatedly-used frying oils. Much further information focused on the carcinogenicity of aldehydes
may be found in [58] and [140].
5.5. 4,5-Epoxy-trans-2-alkenals
In 2017, the FGE.19 EFSA Panel on Food Contact Materials, Enzymes, Flavourings and
Processing Aids concluded that ‘4,5-epoxydec-2-(trans)-enal (FL-no: 16.071) does raise a safety
concern with respect to genotoxicity and, therefore, it cannot be evaluated according to the
Procedure.’ [141]. As noted in our studies [12], 4,5-Epoxy-trans-2-alkenals represent ca. 10 molar % of
the total α,β-unsaturated aldehyde contents of PUFA-rich corn or sunflower oils when thermally-
stressed according to laboratory-simulated shallow frying episodes at 180 °C.
5.6. Acetaldehyde and formaldehyde
Salaspuro [142] found that aldehyde and alcohol dehydrogenase gene polymorphisms (ALDH2
and ADH respectively) are associated with excessive acetaldehyde exposure, and substantially
increase cancer risk in alcohol drinkers, observations which strongly supports the hypothesis that
this saturated aldehyde represents a local carcinogen in oesophageal and gastric cancers.
Interestingly, acetaldehyde can be classified as a tertiary LOP, since it arises from the deterioration of
isomeric alka-2,4-dienals [143], or 2,3- or 4,5-epoxyaldehydes [144,145] during high temperature
frying episodes. Section S3 (Supplementary Materials) provides information on dietary sources and
estimated dietary intakes of acetaldehyde and formaldehyde, most notably alcoholic beverages for
the former. A further study involved human cells collected from patients with a faulty copy of the
BRCA2 breast cancer gene to investigate mechanisms associated with aldehyde-mediated cancer
induction [146], and the investigators found that formaldehyde exposure leads to the degradation of
cellular BRCA2 protein. In those with one faulty copy of its gene (approximately 1 in 100 humans),

Nutrients 2020, 12, 974 32 of 49
this process reduces this protein’s concentration below that which is deemed sufficient for efficient
DNA repair; this process therefore facilitates the induction of cancer.
The International Agency for Research on Cancer (IARC) has classified formaldehyde, another
known aldehydic LOP, as a human carcinogen [147]. Moreover, in 2011, the National Toxicology
Program, an interagency program of the Department of Health and Human Services also classified
formaldehyde as a known human carcinogen in its 12th Report on Carcinogens [148].
5.7. Impact of fried food intake on cancer risks in humans
Particularly notable are epidemiological studies focused on the impact of fried food intake on
cancer risk. In 2013, Stott-Miller et. al. [149] explored links between the male human consumption of
fried foods and prostate cancer risk, and following a review of dietary intake data from more than
3,000 participants, found that this condition was more prevalent amongst those who frequently
consumed deep-fried foods, particularly French fries, fried chicken, fried fish and doughnuts. These
results provide strong evidence for a relationship between fried food intake and prostate cancer risk;
results were found to be more highly significant for a more aggressive disease status. Additionally, a
meta-analysis of published data [150] found that greater fried food intakes induced an estimated 35%
enhancement of prostate cancer risk.
Knecht et. al. [151] found evidence for a positive association between the human intake of fried
meat and combined breast, endometrium and ovarian cancers in women, i.e. female hormone-related
cancers. Moreover, Bosetti et. al. [152] investigated the role of fried food intake on laryngeal cancer
risk in a case-controlled study focused in Italy and Switzerland (> 500 and 1,200 cases and negative
hospital controls respectively), and discovered a significantly elevated risk for participants who had
high consumption rates of fried potatoes (odds ratio 1.9), meat (1.6), fish (3.1) and eggs (1.9).
Both genotoxic and carcinogenic risks linked to the ingestion of repeatedly-boiled sunflower oil
were investigated by Srivastava et. al. [153], and this study found that its oral administration to Wistar
rats resulted in a dose-dependent induction of aberrant cells and micronuclei; such dosing also
depleted antioxidant enzyme availabilities. Moreover, this treatment also influenced hepatic foci,
along with significant decreases in liver mass.
Woutersen et. al. [154] reviewed both animal model and epidemiologic studies focused on the
effects of dietary fat consumption on the risks of breast, colorectal, pancreatic, and prostate cancers,
and found that its increasing intake exerted a significant influence on prostaglandin and leukotriene
biosynthetic routes, and that these properties represented a universal mechanism for such adverse
effects. These researchers also reported that the 50% lethal dose (LD50) values for acrolein in rabbits
and mice were 7 and 40 mg/kg, i.e. there is a wide ‘between-species’ sensitivity to this α,β-
unsaturated aldehyde.
6. Potential Mechanisms for the Toxicity and Health Effects of Dietary Aldehydes
Saturated aldehydes (both short- and long-chain) act as ‘hard’ electrophiles, exerting their toxic
actions through chemical reactions with the primary and secondary amine functions, for example
that of protein lysyl or histidyl residue side-chains. However, -unsaturated aldehydes and
additional alkenals, along with -oxoaldehydes, act as ‘soft’ electrophiles which preferentially react
with ‘softer’ thiol/thiolate functions within protein, peptide (critically GSH) and free cysteine
residues. In general, chemopathological mechanisms available for the toxicity of LOPs can be defined
as either direct or indirect. Direct mechanisms include the formation of adducts with biomolecules,
e.g. reactions of aldehydes with biochemically-critical proteins and DNA in vivo (i.e. adduction
routes), whereas indirect mechanisms may involve the albeit secondary triggering of mitochondrial,
oxidative, and/or endoplasmic reticulum stress, with special reference to their associations with
human diseases and any target tissues and organs affected.
Recently, Xie et. al. [155] suggested a mechanism for the cytotoxicities of the two different classes
of aldehydes, and these involved protein and/or DNA damage. They stipulated the importance of
DNA repair processes as means for protection against damage provoked by less toxic saturated
aldehydes, but not by the unsaturated ones, and hence surmised that inactivation of cells by the more

Nutrients 2020, 12, 974 33 of 49
toxic latter classes occurs via protein adduct formation. Furthermore, this review suggested that DNA
inter-strand crosslinks, but not DNA-protein crosslinks, nor double-strand DNA breakages, are
critical factors for DNA damage resulting from aldehyde attack. In addition, it appears that aldehyde
cytotoxicity, which is DNA damage-independent, is mediated by the loss of intracellular GSH, which
readily traps -unsaturated aldehydes via Michael addition reactions. However, there are only very
low, sub-micromolar levels of aldehydes available in vivo (i.e. residual concentrations following their
metabolism, or protein/DNA adduct formation), for example those found in blood plasma [79].
Hence, thermodynamically, such diminished concentrations have the ability to only chemically
consume an absolute maximum of their equivalent level of intracellular GSH (for 1:1 aldehyde-GSH
conjugates), which represents only a very small fraction of the large intracellular pools of this
scavenging thiol available in vivo, and which are often much higher millimolar concentrations, e.g.
5.5–6.0 mmol/L in whole human blood, which is almost exclusively intracellular [156]. Therefore, it
is conceivable that such low (micromolar or sub-micromolar concentrations) of aldehydes have the
ability to ‘prime’ cells for a cascade of such damaging events following their uptake.
However, this is certainly not expected to be the case within the GI system, where
[aldehyde]:[thiol/GSH] concentration ratios are expected to be much greater (the fate of aldehydes
therein is outlined in section 3.2). However, aldehyde metabolites such as mercapturate conjugates
of the -unsaturated classes (both with and/or without their aldehydic functions reduced or
oxidised to corresponding alcohol or carboxylate anion, respectively), potentially serve as valuable
biomarkers of human exposure to these dietary toxins in biofluids such as human urine and blood
plasma, as may be protein-conjugated aldehyde adducts in the latter. Indeed, blood plasma
concentrations of both the low- and high-molecular-mass classes of these biotransformation products
are likely to be present at much higher levels than the reactive aldehydic precursors themselves in
such biofluids, which renders their detection and quantification more responsive to some lower
sensitivity bioanalytical techniques. Moreover, mercapturate metabolites are also valuable for the
reliable tracking of these aldehydic LOPs in vivo [117]. Interestingly, conceivably higher level thiol-
unsaturated aldehyde Michael addition products in vivo, may also serve as latent sources of these
reactive and more highly toxic LOPs [157]. It is also anticipated that the bulk of circulating -
unsaturated aldehydes may be covalently bound to the cysteine-34 residue of human serum albumin,
since this protein represents the major source of thiol/thiolate anion in this biofluid, and is present at
a concentration of 30 mg/mL (approximately 0.5 mmol/L, with a near-equivalent thiol concentration).
However, alternative albumin amino acid-aldehyde conjugates have been observed for added
acrolein in in vitro experiments, and these involve the covalent modification of its histidyl and lysyl
residues [158].
A series of proteomic-based investigations have confirmed that unsaturated aldehydes inhibit
functionally-important cellular enzyme activities by specifically targeting active-site cysteinyl
residues therein, or more specifically their thiol/thiolate functions. For example, the impairment of
glutathione S-transferase P1-1 (GSTP1-1) activity via Michael-type adduct generation at its Cys-47
residue by a whole range of -unsaturated aldehydes and ketones (a critical factor for
consideration, since this protein detoxifies xenobiotics through glutathione conjugation processes);
suppression of mitochondrial sirtuin 3 (SIRT3) function by reacting with its Cys-280 residue; and
inhibition of glyceraldehyde 3-phosphate dehydrogenase (GAPDH) activity by acrolein via its ability
to form a Michael addition product at its active-site Cys-152 residue (reviewed in [159]).
Notwithstanding, oxidation of thioredoxin 1 is also considered to represent an important mechanistic
factor for consideration.
Aldehydes readily diffuse across cellular membranes in view of their amphiphilic structures,
and therefore have the capacity to covalently react with and hence modify the structure of
biomolecules located within the cytoplasm and nucleus, for example. Hence, such damage may occur
far from their site of generation if formed in vivo [160], and if produced or available extracellularly,
they have the capacity to interact with adjacent cells located remotely from sites of UFA peroxidation;
in such cases, it appears that plasma membrane proteins serve as the primary targets for aldehyde
attack and adduct formation [161]. Such remote attack may be enabled and/or facilitated by the prior

Nutrients 2020, 12, 974 34 of 49
generation of Michael addition adducts with GSH or other endogenous thiols, which represent latent
sources of these aldehydes [157]. Moreover, both endogenous and exogenous (dietary derived)
aldehydic LOPs react with nuclear proteins, and hence attenuate protein expression via chemical
reactions with transcription factors [162].
Pathologically-significant enzymatic targets for aldehydes critically depend on the cell type
involved, and also on the pattern and distribution of such aldehyde-reactive proteins available, along
with cell-penetrating aldehyde levels. Aldehydic protein adduct generation will undoubtedly have
differential physiological ramifications for different protein targets with differing cellular functions,
along with the precise molecular structure of the aldehyde toxin itself. Moreover, the abilities of such
secondary LOPs to reach these targets will also be critically influenced by the intracellular availability
of low-molecular-mass aldehyde scavengers such as thiols (predominantly GSH and L-cysteine),
other scavenging free amino acids such as L-histidine and L-lysine, and biomolecules with free
primary or secondary amine functions (e.g., the secondary metabolite dimethylamine). Intriguingly,
when present at low (non-toxic) concentrations, selected aldehydes (especially HNE) have the ability
to interfere with signal transduction processes, block cellular proliferation and adhesion, and
promote angiogenesis, differentiation and/or apoptosis in cancer cell lines by exerting an influence
on the modulation of gene expression through the formation of covalently-modified protein and/or
DNA adducts [163,164]. These effects appear to represent an example of an aldehyde-triggered
cellular ‘priming’ process, which requires only very low levels of these induction agents.
Although the biomolecular mechanisms of unsaturated aldehyde toxicity are directly linked to
their abilities to form adducts at functionally-important, regulatory cysteine residues in key enzymes,
and hence abrogate their functions, it appears that the onset of toxicological and associated adverse
health effects are not controlled by the impairment of a single protein. Indeed, much data available
now indicates that such toxins effectively suppress an electrophile-responsive proteome, which
comprises cysteine-directed cell-specific proteins. Such considerations are markedly complicated by
the wide range of aldehydes and aldehyde classes identified in both fried foods and their thermally-
stressed CO sources. In addition to their differential dietary availability and intakes, such proteome
inhibition will be strongly mediated by toxicokinetic parameters, including their GI reactivity and
fate, the rate and extent of their in vivo absorption, subsequent metabolism and biodistribution, etc.,
which, in turn, are determined by their physicochemical characteristics such as electrophilicity and
water solubility, etc. Also important is their ability to access such cysteinyl protein residue targets,
which can be limited for at least some unsaturated aldehydes which have structural steric hindrance
[155,159].
Fortunately, powerful batteries of aldehyde-metabolising enzymes are available for the rapid
consumption and removal of aldehyde toxins in the cardiovascular system [165]. However, gene
polymorphisms, which modify the efficacies and extents of aldehyde removal, may significantly
contribute towards human susceptibilities to exposure to these agents, and allelic variations in the
highly polymorphic glutathione S-transferase P has major catalytic function consequences regarding
its abilities to metabolise aldehydes, e.g. acrolein and crotonaldehyde, effectively. Similarly,
polymorphisms in aldehyde-reducing aldo-keto reductases, and/or aldehyde-oxidising aldehyde
dehydrogenases and cytochrome P450s may also have deleterious health implications. Recent
evidence has indicated that a large number of the genes encoding for aldehyde-metabolising enzymes
are triggered by selected natural plant products, for example diallyl-disulphide and -trisulphide, and
dithiol-2-thione (present in garlic and cruciferous vegetables respectively), and hence these agents
and their plant sources may serve to offer humans protection against dietary aldehydic onslaughts
[165].
6.1. Important considerations for HNE and HNE
In 2019, Sottero et. al. [166] critically reviewed the adverse health effects of secondary aldehydic
LOPs and oxysterols; however, their considerations of the former were limited to HHE and HNE,
which are formed in only relatively low quantities during the thermal-stressing of linolenoyl- and
linoleoylglycerols respectively, in culinary frying oils and consequently fried foods, when expressed

Nutrients 2020, 12, 974 35 of 49
relative to those of the more predominant α,β-unsaturated aldehydes (trans-2-alkenals and
trans,trans-alka-2,4-dienals). Such 4-hydroxy-alkenals readily diffuse from the gut into the blood
circulatory system, as do trans-2-alkenals [73] and many additional aldehyde classes.
Subsequent to the digestion of ω-3 FA-rich foods, significant concentrations of HHE were found
to be generated in both static and dynamic in vitro systems modelling gastric and intestinal digestion
[2–4]. Further evidence for the in vivo absorption of dietary sources of both HHE and HNE has been
provided by Awada et. al. in 2012 [76], who demonstrated that the former aldehyde accumulated in
mouse blood after the feeding of these animals with high fat, ω-3 FA-containing diets for a period of
8 weeks. Similarly, this study revealed that mice which were originally fed with HHE had transient
elevations in its plasma level. Moreover, enterocytes treated with that this LOP increased its
barolateral medium concentrations [76], and taken together, these results provide evidence for its
intestinal absorption. Additionally, radioactivity and radiolabelled HNE metabolites have been
detected in the urine, faeces, intestinal contents and major organs of rats following the oral
administration of tritiated HNE to these animals.
The persistence of α,β-unsaturated aldehydes, including HHE and HNE, following the in vitro
digestion of food sources of them, particularly within the lipidic phases of digestion products, was
confirmed by GC-MS analyses, and this again indicates that they are bioavailable to the GI tract for
absorption [167] (further details on this are available in the Supplementary Materials, Section S5).
However, aldehydes detectable in the blood and tissues of raw fish may also arise from the actions
of lipoxygenases [168].
6.1.1. In vivo generation of HNE/HHE from high and sustained dietary supplementation with ω-3
PUFAs?
In vitro models have been developed in order to evaluate the adverse in vivo generation of HHE
and HNE from dietary ω-3 FA sources, and these effectively mimic the digestion of vegetable cooking
and marine oils [2]. Notably, the in vitro digestion of metmyoglobin-containing fish oil emulsions
gave rise to the production of both HHE and HNE (ca. 2 and 7 µM, respectively) within this matrix
[169]. Moreover, for salmon loin and minced beef of equivalent lipid contents, the formation of both
these aldehydes was determined in such digestive fluids following the gastric and intestinal phases,
and the maximal digestive fluid HNE level observed was ca. 2 µM for both food classes, whereas
intestinal digestion of salmon oil gave rise to a higher concentration of HHE than that observed for
minced beef (3.5 versus 2 µM) [170].
6.1.2. Influence of CO consumption on blood plasma levels of HNE and HHE
Hence, it is of much importance to determine the concentrations of dietary-derived aldehydes
in human peripheral blood following the dietary ingestion of cooking oil acylglycerol PUFAs, both
unheated and those subjected to increasing numbers of high-temperature frying episodes. Indeed,
one interesting study conducted by Calzada et. al. [171] involved calculation of the plasma levels of
healthy adult participants who were supplemented with dietary DHA (200–1,600 mg/day)
throughout a 14-day period. Although no change in HHE concentrations were observed at doses of
200 and 400 mg/day, progressively significant elevations in these values were observed the higher
doses administered (800 and 1,600 mg/day), and these reached 60 and 87 nmol/L respectively.
However, it appears that these researchers did not perform essential quality checks on the
peroxidation status of DHA samples administered in these investigations. Moreover, as noted above,
the TBARS test featured in these studies has major artefactual and interference issues associated with
its use as a means to determine secondary LOP levels, and the heating stage involved in the form of
the test employed in this study was 96 °C for a 60 min. period, which is more than sufficient to
peroxidise PUFAs in analytical samples.
Although it is presumed that such HHE may arise from the in vivo peroxidation of fish oil DHA,
it is, of course, conceivable that this secondary LOP was also present in the samples administered to
the above experimental animals or humans; unfortunately, it certainly appears that the fish oil diets
used for these experiments were not tested for aldehydic and other LOPs prior to their

Nutrients 2020, 12, 974 36 of 49
administration, and therefore HHE may itself have been directly administered along with fish oil
EPA and DHA. HHE is a product of DHA and not EPA peroxidation [172], and this observation is
consistent with that of Nagagawa et. al.’s [173].
HNE is detectable at sub-micromolar levels in human cells, tissues and biofluids, and its ‘free’,
non-adducted concentrations in human plasma is 3–125 nM; such values have been shown to be
markedly enhanced (0.1–1.0 µM) in human disease such as coronary and peripheral artery diseases
[174], and rheumatoid arthritis [175].
However, as noted throughout here, 4-hydroxy-trans-2-alkenal levels present in used
linoleoylglycerol-rich culinary frying oils are always ≤ 10% of their total aldehyde content, and less
so in potato chip samples [12], a likely consequence of the increased reactivity of this and indeed
other classes of α,β-unsaturated aldehydes towards food biomolecules such as proteins, peptides and
amino acids, along with acetal/hemiacetal-forming carbohydrates and alcohols, over those of less
reactive saturated aldehydes.
6.1.3. Haem oxygenase-1 expression and dietary marine oil supplementation: potential beneficial
role of HHE
Since haem oxygenase-1 (HO-1) exerts protective actions against a range of diseases, the role of
ω-3 FAs, which are involved in the induction of its expression both in vitro and in vivo, is of much
interest. Intriguingly, Nagagawa et. al. [173] examined the ability of dietary supplementation of fish
oil on the pattern of FAs and their peroxidation products (specifically HHE and HNE) on HO-1
expression within an extremely wide range of tissues (including liver and kidney) and the blood
plasma of C57BL/6 mice, and found that both HHE concentration and HO-1 expression were
upregulated following institution of this dietary regimen. Such changes were correlated with
corresponding increases in DHA but not EPA levels. Overall, these results were proposed to be
consistent with the hypothesis that DHA-derived HHE actually induces HO-1 expression, and that
this aldehyde may be responsible for the HO-1-mediated protective effects exerted by dietary marine
oils when generated from it in vivo.
6.1.4. Cell signalling by HHE and HNE
Briefly, studies focused on the potential involvements of both HHE and HNE in cellular
signalling processes have indicated their roles in the great majority of signal transduction pathways
[176,177], including redox homeostasis, and mediation of key transcription factor activities, e.g. those
of nuclear factor-κB (NF-κB), nuclear erythroid-related factor (Nrf2) and activator protein 1 (AP-1)
[176–179]. Undoubtedly, their high level of reactivities with thiol(ate) and amine functions to form
Michael addition (both) and Schiff base (the latter only) adducts is of crucial importance here.
6.1.5. Aldehydes as the dominant carcinogens present in cigarette smoke
Finally, a very recent observation of much significance has provided a high level of evidence
that aldehydes represent the dominant carcinogens present in tobacco smoke which give rise to DNA
damage, inhibit DNA repair in tobacco smoke carcinogenesis and also prevent many other tobacco
smoke procarcinogens (including 4-(methylnitrosamine)1-(3-pyridyl)-1-butanone and polyaromatic
hydrocarbons) from becoming DNA-damaging agents [180]. On the basis of these results, the authors
of this paper proposed that toxic aldehydes represent the dominant tobacco smoke carcinogens. As
noted in [12], the aldehyde contents of a typical large size serving of restaurant fried potato chips are
not very dissimilar to those available for inhalation during the smoking of a 20–25 allocation of
tobacco cigarettes [137].
7. Targeted Nutrition and Potential Interventional Routes for Eliminating or Alleviating Health
Risks Associated with Dietary LOP Intake in Humans
Potential strategies for alleviating or circumventing health hazards presented by dietary LOPs
represents a widespread area for careful consideration. These include viable means for the determent

Nutrients 2020, 12, 974 37 of 49
of LOP generation in PUFA-rich frying oils such as their prior supplementation with heat-resistant
lipid-soluble antioxidants, or the removal of these toxins via the treatment of repetitively-used COs
with selected LOP-targeted filtration aid materials. However, a major drawback to this antioxidant
fortification approach is that many studies have provided evidence that naturally-occurring or higher
concentrations of plant-derived chain-breaking antioxidants such as - or γ-TOH, or synthetic ones
such as butylated hydroxytoluene (BHT), are only poorly effective in this context in view of the
extremely high level of repetitive thermally-damaging peroxidative recycling bursts often
encountered during standard high-temperature frying periods that COs are often exposed to [1].
Moreover, at least some of these antioxidants are thermally unstable, and they may also be
significantly volatilised at these temperatures (ca. 180 °C) [9,73]. In view of these findings, the future
availability of more powerful and more thermally-stable antioxidants, including a range of unusual
molecules which are not normally considered to act in this capacity (natural or otherwise), may
indeed develop into a productive area for future development.
The employment of currently-available dimethylpolysiloxane polymers, which are surfactants
and anti-foaming agents which also limit exposure of culinary oil surfaces to atmospheric O2 required
for peroxidation, may also be effective for inhibiting LOP generation during deep-frying episodes;
the future customised design and synthesis of more efficient or composite-function derivatives of
these may therefore promote frying oil safety, along with an extension of the frying reuse periods of
CO products. Additionally, technological approaches available involving methylcellulose or
alternative ‘barrier’ agents, which block the uptake of LOP-loaded used PUFA-rich oils by foods fried
therein [181], may also offer a solution to this critically important public health issue. Indeed, fried
potato chip aldehyde toxin contents are strongly and positively correlated with their total lipid
contents, i.e. they are related to the extent of frying oil uptake in this food LOP source [3].
Interestingly, our research work has also revealed that CO LOPs are predominantly present in the
external batter layer of battered fried foods such as chicken or fish, with little or none detectable in
the food component itself (Figure 3), and so this appears to represent a novel means of protecting
fried foods against LOP uptake and human intake, if only consumers were prepared to remove the
battered covering of such foods prior to eating! Unfortunately, this battered layer tends to serve as a
very palatable, savoury and attractive component of such fried meat and fish food products.
However, a notable and highly plausible prophylactic approach is the dietary supplementation
of human fried food consumers (especially those with poor diets, or a high consumption rate of such
foods) with suitable aldehyde-trapping therapies. For example, the amino acid L-cysteine, which is
equipped with an aldehyde-consuming side-chain thiol/thiolate function [164], and/or suitable chain-
terminating antioxidants, although it should be noted that the latter interventional action will only
serve to potentially terminate the further generation of directly-ingested lipid hydroperoxides in the
GI system, and hence block their degradation to more stable aldehydes and other fragmentation
product toxins. Alternatively, prior fortification of this bioenvironment with relatively high levels of
such ingested antioxidants may effectively impair the peroxidation of ingested UFAs which can be
triggered therein [51,52].
One further possible anti-aldehyde strategy involves the antihypertensive drug hydralazine,
which reacts with acrolein and crotonaldehyde to form stable reaction products, for example (1E)-
acrylaldehyde phthalazin-1-ylhydrazone (E-APH) and (1Z)-acrylaldehyde phthalazin-1-ylhydrazone
(Z-APH) from acrolein, and (1E,2E)-but-2-enal phthalazin-1-ylhydrazone (E-BPH) and (1Z,2E)-but-2-
enal phthalazin-1-ylhydrazone (Z-BPH) from crotonaldehyde [182]. This drug is similarly reactive
towards other 2-alkenals, as is its structural analogue dihydralazine. Hydralazine therefore blocks the
cellular toxicity exerted by acrolein, and other 2-alkenals arising as secondary LOPs, and in 2011 Leung et.
al. [183] found that this treatment (described as an ‘anti-acrolein’ initiative, but not exclusively limited to
trapping only this 2-alkenal) significantly diminished myelin damage and improved behavioural outcome
in an experimental mouse model system of autoimmune encephalomyelitis.
Alternatively, plausible targeted manipulations of human levels and activities of aldehyde-
neutralising enzymes could also serve as a means for combating the deleterious exposures to
aldehyde toxins. Indeed, an improved understanding of the biomolecular mechanisms involved in

Nutrients 2020, 12, 974 38 of 49
the induction or stimulation of such enzymes with, for example, selected sulphur-containing natural
plant products [165], may provide valuable information regarding specific therapeutic targets which,
when activated, may offer an enhanced level of cellular protection against the adverse health effects
of exogenous aldehydes.
Notwithstanding, perhaps the best strategic protective approach is for consumers, together with
restaurant and fast-food outlet proprietors, to simply employ COs with only limited PUFA contents
for frying and cooking purposes. Notwithstanding, the avoidance of fried meals cooked in PUFA-
rich oils is not easily achievable when consumers dine in restaurants or purchase take-out fast food
products (especially if they request such frying oil identity information from restaurant staff). Such a
development will serve as the most palpable, easily instigated, and consumer-controllable approach
for directly avoiding or minimizing aldehyde-mediated adverse health effects. One recent study
demonstrated that a MUFA-rich algal frying oil, which contained ca. 90% oleoylglycerols and only ~
5% (w/w) PUFAs, generated only very low levels of aldehydic toxins when exposed to both actual
and laboratory-simulated frying episodes (deep- and shallow-frying processes respectively) [12].
From all the studies reviewed here, evaluation of the possible health-threatening effects and
disease risks of dietary LOPs realistically remains a dauntingly complex task, since these
considerations should be made with special reference to recommended maximum human daily
intake (MHDI) values for these hazardous agents, i.e. those stipulated by relevant regulatory health
authorities and organizations. However, currently documented values are either very limited to
selected aldehydic LOPs such as acrolein, outdated, or even inconsistent between regulatory bodies,
i.e. very few are available. Similar considerations also apply to outdated or unrealistic estimates for
the MHDI values of such toxins, either from dietary or other sources. One approach employed to
date, however, is the determination of an ‘acrolein-adjusted’ MHDI index, which has been employed
to relate potential values for higher molecular weight 2-alkenals to that available for its lowest
homologue class member, acrolein [3,14]: this value is simply determined by dividing the molecular
mass of acrolein by those of higher 2-alkenals (e.g., linoleoylglycerol hydroperoxide-derived trans-2-
octenal), and then multiplying this fractional ratio by the MHDI value of acrolein itself. Currently,
the authors do not consider this approach to be completely satisfactory for trans- and cis-2-alkenals,
and certainly not so for alternative aldehydic LOP classes such as n-alkanals, for example. Hence, the
future consideration, establishment and ratification of many currently unavailable MHDIs for LOPs
of known molecular identities also represent major demands for action. Consumer concerns
regarding the nutritional and health properties of their foods strongly warrant such requirements
[184].
In view of the above considerations, optimizations of combinations of food processing methods
for eliminating or reducing the content of undesirable LOPs will be facilitated, together with
corresponding assessments of the safety of fried and convenience foods, with special reference to the
ever-changing consumer lifestyles of the global population.
Finally, as a further critically important factor, the multitude of previous investigational
scientific reports available which focus on the possible beneficial health effects of dietary PUFAs
should be thoroughly revisited, particularly with regard to those featuring feeding trials with human
participants, or other related cohort epidemiological or meta-analysis studies. On reflection, it
certainly appears that many of these previously conducted studies may be flawed, since the
researchers involved have predominantly neglected the potentially substantial confounding adverse
health effects associated with the intake of LOPs such as aldehydes, which were undoubtedly present
or even prevalent in the oils or diets originally explored in such investigations.
8. Conclusions
Heating of culinary frying oils at temperatures associated with standard frying practices gives
rise to the generation of very high concentrations of cytotoxic and genotoxic aldehydic LOPs from
thermally-promoted, self-propagating oxygen-fuelled recycling peroxidative assaults occurring
therein. These toxins penetrate into and hence are ‘carried’ by foods fried in such media, and therefore
are available for human ingestion. Since the repeated dietary consumption of such LOPs, especially

Nutrients 2020, 12, 974 39 of 49
the -unsaturated classes, may pose serious and chronic hazards to humans, the development of
strategies for overcoming these threats is of paramount importance. Future clinical feeding trial or
epidemiological investigations focused on explorations of the relationships between the incidence
and/or severity of selected human diseases (such as coronary heart disease, cancer, etc.), and the
frequency and level of dietary LOP ingestion, may therefore serve to decipher and clarify the nature
of such relationships. Similarly, previously available reports that PUFA-laden cooking oils are
‘beneficial’ or ‘safe’ for human consumption after being employed for frying or alternative high
temperature cooking purposes may be erroneous and inaccurate, since they predominantly fail to
monitor or even consider any LOPs therein, nor the major public health threats posed by their human
ingestion.
Following their in vivo ingestion, blockage of the activities and functional status of one or more
intracellular protective enzymes at critical active-site cysteinyl residues appears to represent the most
important mechanism for the cyto- and genotoxicities of unsaturated aldehydes. In general,
aldehydes readily cross cell membranes and enter intracellular environments where they may exert
such damaging actions. An analysis of the fractional concentrations of four classes of aldehydic LOPs
in human blood plasma, a study performed here for the first time (Section 3.2.2), demonstrated that
their mean n-alkanal:trans-2-alkenal ratio was similar to that observed in a fried potato chip dataset,
and this may indicate that such aldehyde classes are at least partially dietary-derived, although there
are, of course, many limitations to this form of evaluation. However, proportionate circulating levels
of 4-hydroxy-trans-2-alkenals (including HNE and HHE) and MDA were found to be significantly
much greater than those present in this commonly-consumed fried food source, and again allowing
for the above limitations, these data suggest that these secondary LOPs arise from in vivo peroxidation
episodes. In principle, secondary aldehydic LOPs ingested by humans have the ability to provoke
further cellular ROS generation in vivo, a phenomenon which, in turn, may stimulate further
aldehydic LOP generation and hence amplify and perpetuate any deleterious health effects inducible.
The World Health Organisation (WHO) has indeed identified concerns with the toxicological
and genotoxic potentials of aldehydes [185]. As an example, in 2002 they reported that there were >
30 epidemiological case-control studies focused on populations exposed to formaldehyde (also a
known LOP [186]), and their cancer incidence [187]. Whilst identifying significant concerns on the
inhalation of aldehydes and linked respiratory tract carcinomas, this report makes little reference to
data available in terms of risks arising from food sources. However, non-respiratory tract cancers
were detected in populations exposed to the inhalation of this aldehyde, i.e. multiple myelomas,
pancreatic, colon and brain cancers, amongst others. Furthermore, the European Union Scientific
Committee on Consumer Safety reported on the ingestion of acetaldehyde and its carcinogenicity,
reproductive toxicity and genotoxicity [187]; the cancer risk status of this LOP is detailed in Section
S3 (Supplementary Materials).
Similarly, in view of a potential role of tt-DDE in human carcinogenesis, and its widespread
occurrence in food products, there remain increasing concerns regarding potential associations
between dienaldehyde exposure and the development of human cancers [188]. These concerns are
now strongly supported by the detection of this aldehyde and other -unsaturated ones in fried
foods and thermally-stressed (used) CO sources of this toxin [12], along with high levels detectable
in fumes generated from linoleoylglycerol-rich cooking oils [189]. Additionally, a large amount of
experimental evidence acquired from animal model system investigations have revealed powerful
associations between reproductive and developmental toxicities and exposure to formaldehyde
(extensively reviewed in Ref. [190]); such experiments have involved a range of exposure routes and
dose levels, in different species.
Notably, since the potential adverse health effects of the low content food process contaminants
acrylamide and MCPD derivatives have attracted much significant attention (both in scientific
publications and the media), why is it that toxic aldehydic LOPs, which are present in fried foods at
much greater concentrations, are not receiving a similar level of consideration? The authors have a
high level of public health concern regarding this issue, not least because it is much more widespread,
i.e. it is shared by many other researchers engaged in this increasingly important research area. Is it

Nutrients 2020, 12, 974 40 of 49
not the right time for health authorities and governmental food standards agencies to warn the public
about these very important health threats?
Critical factors which are most likely to play key roles in determining the nature and level of
dietary LOP intake in humans, e.g., shallow versus deep-frying processes, and particularly their
permeation into fried foods available for human consumption such as potato chips, beef patties,
battered chicken portions, etc. should be further investigated. The availability for human
consumption of high, toxicologically-significant (up to 25 ppm for each class) levels of the
predominant classes of toxic aldehydes in servings of fried foods collected directly from fast-food
retail outlets/restaurants, including ubiquitous, globally-accessible large chain ones, should also be
considered in detail. Moreover, the rigorous establishment of currently unavailable ADIs and MHDIs
for the extensive number of dietary aldehydic LOPs is also a key future prospect. Such requirements
are of much importance in view of consumer stakeholder concerns regarding the nutritional and
health properties, both positive and negative, prospectively offered by contemporary foods and
dietary patterns worldwide.
Supplementary Materials: The following are available online at www.mdpi.com/xxx/s1. Section S1: Epoxy-Fatty
Acid (Epoxy-FA) LOPs: In Vivo Absorption and Toxicities; Section S2: Overview of Dietary Sources of LOPs;
Section S3: Food Sources and Cancer Risks of Acetaldehyde and Formaldehyde; Section S4: Comparisons of the
Dietary Availability and Ingestion of Aldehydic LOPs to those of trans-Fatty Acids, Acrylamide and
Monochloro-Propanediol (MCPD) CO/Fried Food/Lipid Product Toxins; Section S5: In vivo Absorption,
Metabolic Fate, Toxicology, and Adverse Health Effects of HNE and HHE; Section S6: Further Limitations of the
Mak et. al. Study (Ref. [79]); Section S7: Blood Plasma Aldehyde Concentrations in Infants with Chronic Lung
Disease; Section S8: DNA Adduct Formation with and Detoxification of trans-2-Hexenal; Section S9: Cytotoxic
and Mutagenic Potential of the Natural Product 2-Cyclohexene-1-one Evaluated against a Range of Dietary
Aldehydes; Section S10: Acrolein as an Inhaled or Ingested Toxin of Carcinogenic Potential: a Special Case for
Consideration; Section S11: Dietary Sources, Inhalation/Ingestion, Cytotoxicity and Genotoxicity of t,t-DDE
Author Contributions: M. G. had the original concept for this work and all supporting experiments described
therein. B.C.P. performed experiments involving the thermal stressing of CLO products, the time-dependent
collection and analysis of LSSFE samples, sample preparation and 1H-NMR spectral data acquisition, together
with NMR data processing, analysis, and interpretation. B.C.P., J.L. and P.B.W. generated and/or finalised all
manuscript Figures. M.G. contributed towards all these work-tasks, together with the full literature review
involved, the statistical analysis of experimental data, and also preparation and finalisation of the manuscript.
P.B.W., B.C.P. and J.L. all reviewed and edited the manuscript, and B.C.P. also contributed towards the
interpretation and assignment of 1H-NMR spectra. M.G. also fully designed and supervised the experimental
components of the investigations. P.B.W. and M.G. designed and produced the graphical abstract. All authors
contributed towards manuscript preparation and development. All authors have read and agreed to the
published version of the manuscript.
Funding: This research received no external funding.
Acknowledgments: All authors are very grateful to the Dave Wetzel and Jie Zhang of Green Pastures Products
Inc. (NE, USA), and Sally Fallon of the Weston A. Price Foundation (DC, USA) for the provision of culinary and
marine oil samples for NMR analysis and further investigations, and also for stimulating discussions. We also
thank the International Trade Centre, United Nations-WTO (Geneva, Switzerland) for non-financial support.
B.C.P is very grateful to De Montfort University, Leicester, UK for the provision of a fees-waiver PhD scholarship
bursary. J. L. is very grateful to Cancer Research UK (CRUK) for providing funding for her post-doctoral research
fellowship based at DMU.
Conflicts of Interest: None of the authors declare any conflicts of interest.
Ethical Approval: This article does not contain any investigations with human participants or experimental
animals performed by any of the authors.
References
1. Grootveld, M.; Ruiz-Rodado, V.; Silwood, C.J.L. Detection, monitoring and deleterious health effects of
lipid oxidation products generated in COs during thermal stressing episodes. Inform Am. Oil Chem. Soc.
2014, 25, 614–624.

Nutrients 2020, 12, 974 41 of 49
2. Grootveld, M.; Silwood, C.J.L.; Addis, P.; Claxson, A.W.D.; Bonet Serra, B.; Viana, M. Health effects of
oxidised heated oils. Foodserv. Res. Internat. 2001, 13, 39–53.
3. Grootveld, M.; Percival, B.C.; Grootveld, K.L. Chronic non-communicable disease risks presented by lipid
oxidation products in fried foods. Hepatobil. Surg. Nutr. 2018, 7, 305–312.
4. Estévez, M.; Li, Z.; Soladoye, O.P.; Van-Hecke, T. Chapter Two - Health Risks of Food Oxidation. Advanc.
Food Nutrit. Res. 2017, 82, 45–81.
5. Jackson, V.; Penumetcha, M. Dietary oxidised lipids, health consequences and novel food technologies that
thwart food lipid oxidation: An update. Intern. J. Food Sci. Technol. 2019, 54, 1981–1988.
6. Frankel, E.N. Volatile lipid oxidation-products. Prog. Lipid Res. 1983, 22, 1–33.
7. Dobarganes, M.C.; Perez-Camino, M.C. Fatty acid composition: A useful tool for the determination of
alteration level in heated fats. Rev. Franc. Corps Gras. 1988, 35, 67–70.
8. Claxson, A.W.D.; Hawkes, G.E.; Richardson, D.P.; Naughton, D.P.; Haywood, R.M.; Chander, C.L.;
Atherton, M.; Lynch, E.J.; Grootveld, M.C. Generation of lipid peroxidation products in culinary oils and
fats during episodes of thermal stressing: A high field 1H NMR study. FEBS Lett. 1994, 355, 81–90.
9. Haywood, R.M.; Claxson, A.W.D.; Hawkes, G.E.; Richardson, D.P.; Naughton, D.P.; Coumbarides, G.;
Hawkes, J.; Lynch, E.J.; Grootveld, M.C. Detection of aldehydes and their conjugated hydroperoxydiene
precursors in thermally-stressed culinary oils and fats: Investigations using high resolution proton NMR
spectroscopy. Free Rad. Res. 1995, 22, 441–482.
10. Silwood, C.J.L.; Grootveld, M. Application of high-resolution two-dimensional 1H and 13C nuclear magnetic
resonance techniques to the characterization of lipid oxidation products in autoxidized
linoleoyl/linolenoyglycerols. Lipids 1999, 34, 741–756.
11. Martınez-Yusta, A.; Goicoechea, E.; Guillen, M.D. A Review of thermo-oxidative degradation of food lipids
studied by 1H NMR spectroscopy: Influence of degradative conditions and food lipid nature. Compr. Rev.
Food Sci. Food Saf. 2014, 13, 838–859.
12. Esterbauer, H. Cytotoxicity and genotoxicity of lipid-oxidation products. Am. J. Clin. Nutr. 1993, 57, 779S–
786S, doi:10.1093/ajcn/57.5.779S
13. Wang, L.; Xiang, Z.; Stevanovic, S.; Ristovski, Z.; Salimi, F.; Gao, J.; Wang, H.; Li, L. Role of Chinese cooking
emissions on ambient air quality and human health. Sci. Total Environ. 2017, 589, 173–181.
14. Moumtaz, S.; Percival, B.C.; Parmar, D.; Grootveld, K.L.; Jansson, P.; Grootveld, M. Toxic aldehyde
generation in and food uptake from COs during frying practices: Peroxidative resistance of a
monounsaturate-rich algae oil. Sci. Rep. 2019, 9, 4125.
15. Choe, E.; Min, DB. Mechanisms and factors for edible oil oxidation. Comprehen. Rev. Food Sci. Food Saf. 2006,
5, 169–186.
16. Frankel, E.N. Lipid Oxidation; The Oily Press: Dundee, Scotland, UK, 1998; p. 9.
17. Frankel, E.N. Frying fats. In Lipid Oxidation; Frankel, E.N., Ed.; The Oily Press: Dundee, Scotland, UK, 2005;
pp. 227–248.
18. Neff, W.E.; Byrdwell, W.C. Characterization of model triacylglycerol (triolein, trilinolein and trilinolenin)
autoxidation products via high-performance liquid chromatography coupled with atmospheric pressure
chemical ionization mass spectrometry. J. Chromatogr. A 1998, 818, 169–186.
19. Wang, G.W.; Guo, Y.; Vondriska, T.M.; Zhang, J.; Zhang, S.; Tsai, L.L.; Zong, N.C.; Bolli, R.; Bhatnagar, A.;
Prabhu, S.D. Acrolein consumption exacerbates myocardial ischemic injury and blocks nitric oxide-
induced PKCepsilon signaling and cardioprotection. J. Mol. Cell. Cardiol. 2008, 44, 1016–1022.
20. Acceptable Daily Intakes for Agricultural and Veterinary Chemicals; Current as of 31 March 2016, Australian
Government Department of Health (Commonwealth of Australia, 2016); The Office of Chemical Safety,
Department of Health: Canberra, ACT, Australia, 2016.
21. Totani, N.; Yawata, M.; Mori, T.; Hammond, E.G. Oxygen content and oxidation in frying oil. J. Oleo Sci.
2013, 62, 989–995.
22. Beauchamp, R.; Andjelkovich, D.; Klingerman, A.; Morgan, K.; Heck, H. A critical review of the literature
on acrolein toxicity. Crit. Rev. Toxicol. 1985, 14, 309–380.
23. Hirayama, T.; Yamaguchi, M.; Nakata, T.; Okumura, M.; Yamazaki, T.; Watanabe, T.; Fukui, S. Formation
of acrolein by the autooxidation of unsaturated fatty acid methyl esters. Eisei Kagaku 1989, 35, 303–306.
24. Lane, R.; Smathers, J. Monitoring aldehyde production during frying by reversed-phase liquid
chromatography. J. Assoc. Off. Anal. Chem. 1991, 74, 957–960.

Nutrients 2020, 12, 974 42 of 49
25. Albert, B.; Derraik, J.; Cameron-Smith, D.; Hofman, P.L.; Tumanov, S.; Villas-Boas, S.G.; Garg, M.L.;
Cutfield, W.S. Fish oil supplements in New Zealand are highly oxidised and do not meet label content of
n-3 PUFA. Sci. Rep. 2015, 5, 7928, doi:10.1038/srep07928.
26. Hirayama, T.; Miura, S.; Mori, Y.; Ueta, M.; Tagami, E.; Yoshizawa, T.; Watanabe, T. High-performance
liquid chromatographic determination of 2-alkenals in oxidized lipid as their 7-amino-6-methylquinoline
derivatives. Chem. Pharm. Bull. 1991, 39, 1253–1257.
27. Shields, P.G.; Xu, G.X.; Blot, W.J.; Trivers, G.E.; Weston, A.; Pellizzari, E.D.; Qu, Y.H.; Harris, C. C Volatile
emissions of wok cooking oil. Proc. Am. Assoc. Cancer Res. 1993, 34, 118.
28. Csallany, A.S.; Han, I.; Shoeman, D.W.; Chen, C. 4-Hydroxynonenal (HNE), a toxic aldehyde in French fries
from fast food restaurants. Free Rad. Biol. Med. 2012, 53, S250. doi:10.1016/j.freeradbiomed.2012.08.043.
29. Guillén, M.D.; Uriarte, P.S. Aldehydes contained in edible oils of a very different nature after prolonged
heating at frying temperature: Presence of toxic oxygenated α,β-unsaturated aldehydes. Food Chem. 2012,
131, 915–926.
30. Suhr, J.; Kwan, H. Estimation of daily exposure to 4-hydroxy-2-alkenals in Korean foods containing n-3
and n-6 polyunsaturated fatty acids Food Add. Contamin. 2003, 22, 701–708.
31. Tardiff, R.G.; Gargas, M.L.; Kirman, C.R.; Carson, M.L.; Sweeney, L.M. Estimation of safe dietary intake
levels of acrylamide for humans. Food Chem Toxicol. 2010, 48, 658–667.
32. Agency for Toxic Substances and Disease Registry. Toxicological Profile for Acrolein (August). CAS#: 107-
02-8. Available online: https://www.atsdr.cdc.gov/toxprofiles/tp.asp?id=557&tid=102 (accessed on 10
October 2019).
33. Lachenmeier, D.W.; Kanteres, F.; Rehm, J. Carcinogenicity of acetaldehyde in alcoholic beverages: Risk
assessment outside ethanol metabolism. Addiction 2009, 104, 533–550, doi:10.1111/j.1360-0443.2009.02516.x.
34. Monakhova, Y.B.; Jendral, J.A.; Lachenmeier, D.W. The margin of exposure to formaldehyde in alcoholic
beverages. Arch. Indust. Hyg. Toxicol. 2012, 63, 227–237, doi:10.2478/10004-1254-63-2012-2201.
35. Ferreira, D.C.; Nicolli, K.P.; Souza-Silva, É.A.; Manfroi, V.; Zini, C.A.; Welke, J.E. Carbonyl compounds in
different stages of vinification and exposure risk assessment through Merlot wine consumption. Food
Additiv. Contamin. 2018, 35, 2315–2331. doi:10.1080/19440049.2018.1539530.
36. Peterle, G.P.; Hernandes, K.C.; Schmidt, L.M.; Hoffmann, J.B.; Zini, C.A.; Welke, J.E. Exposure risk to
carbonyl compounds and furfuryl alcohol through the consumption of sparkling wines. Ciência Rural 2019,
49, e20180986. doi:10.1590/0103-8478cr20180986/
37. WHO Food Additives Series 40, 49th Meeting of JECFA. 1998. Available online:
www.inchem.org/documents/jecfa/jecmono/v040je17.htm (accessed on 3 September 2019).
38. Chapter 5.8 Formaldehyde. Air Quality Guidelines, 2nd ed.; WHO Regional Office for Europe: Copenhagen,
Denmark, 2001.
39. World Health Organization (WHO). Human Health Risk Assessment Toolkit: Chemical. 2010. Available
online: http://www.who.int/ipcs/methods/harmonization/areas/ra_toolkit/en/ (accessed on 18 February
2020).
40. Sottero, B.; Rossin, D.; Poli, G.; Biasi, F. Lipid oxidation products in the pathogenesis of inflammation-
related gut diseases. Curr. Med. Chem. 2018, 25, 1311–1326. doi:10.2174/0929867324666170619104105
41. Cortesi, R.; Privett, O.S. Toxicity of fatty ozonides and peroxides. Lipids 1972, 7, 715–721.
doi:10.1007/BF02533120.
42. Wolff, S.P.; Nourooz-Zadeh, I. Hypothesis: UK consumption of dietary lipid hydroperoxides—A possible
contributory factor to atherosclerosis. Atherosclerosis 1996, 119, 261–263.
43. Kokatnur, M.G.; Bergan, J.G.; Draper, H.H. Chronic toxicity of methyl linoleate hydroperoxide for the
rabbit. Proc. Soc. Exp. Biol. Med. 1966, 123, 254–258. doi:10.3181/00379727-123-31459.
44. Hata, K.; Fujimoto, K.; Kaneda, T. Absorption of lipid hydroperoxides in carp. Nihon-suisan-gakkai-shi
1986, 52, 677–684. doi:10.2331/suisan.52.677.
45. Kanazawa, K.; Ashida, H. Dietary hydroperoxides of linoleic acid decompose to aldehydes in stomach
before being absorbed into the body. Biochimica et Biophysica Acta (BBA) Lipids Lipid Metab. 1998, 1393, 349–
361.
46. Hartz, J.W.; Funakoshi, S.; Deutsch, H.F. The levels of superoxide dismutase and catalase in human tissue
as determined immunochemically. Clin. Chim. Acta 1973, 46, 125–132.

Nutrients 2020, 12, 974 43 of 49
47. Marklund, S.L.; Westman, N.G.; Lundgren, E.; Roos, G. Copper- and zinc-containing superoxide dismutase,
manganese-containing superoxide dismutase, catalase, glutathione peroxidase in normal and neoplastic
human cell lines and normal human tissues. Cancer Res. 1982, 42, 1955–1961.
48. Ogasawara, T.; Hoensch, H.; Ohnhaus, E.E. Distribution of glutathione and its related enzymes in small
intestinal mucosa of rats. Arch. Toxicol. 1985, 8, 110–112.
49. Chu, F.F.; Doroshow, J.H.; Esworthy, R.S. Expression, characterisation, and tissue distribution of a new
cellular selenium-dependent glutathione peroxidase, GSHPx-GI. J. Biol. Chem. 1993, 268, 2571–2578.
50. Wingler, K.; Muller, C.; Schmehl, K.; Florian, S.; Brigelius-Flohe, R. Gastrointestinal glutathione peroxidase
prevents transport of lipid hydroperoxides in CaCo-2 cells. Gastroenterology 2000, 119, 420–430.
51. Kanner, J.; Lapidot, T. The stomach as a bioreactor: Dietary lipid peroxidation in the gastric fluid and the
effects of plant-derived antioxidants. Free Rad. Biol. Med. 2001, 31, 1388–1395.
52. Tullberg, C.; Larsson, K.; Carlsson, N.-G.; Comi, I.; Scheers, N.; Vegarud, G.; Undeland, I. Formation of
reactive aldehydes (MDA, HHE, HNE) during the digestion of cod liver oil: Comparison of human and
porcine in vitro digestion models. Food and Funct. 2016, 3, 1401–1412.
53. Wilson, R.; Lyall, K.; Smyth, L.; Fernie, C.E.; Riemersma, R.A. Dietary hydroxy fatty acids are absorbed in
humans: Implications for the measurement of 'oxidative stress' in vivo. Free Rad. Biol. Med. 2002, 32, 162–
168.Bottom
54. Penumetcha, M.; Khan, N.; Parthasarathy, S. Dietary oxidized fatty acids: An atherogenic risk? J. Lipid Res.
2000, 41, 1473–1480.
55. Kritchevsky, D.; Tepper, S.A. Cholesterol vehicle in experimental atherosclerosis. 9. Comparison of heated
corn oil and heated olive oil. J. Atheroscler. Res. 1967, 7, 647–651.
56. Staprãns, I.; Rapp, J.H.; Pan, X.-M.; Hardman, D.A.; Feingold, K.R. Oxidized lipids in the diet accelerate the
development of fatty streaks in cholesterol-fed rabbits. Arterioscler. Thromb. Vasc. Biol. 1996, 16, 533–538.
57. Soffritti, M.; Belpoggi, F.; Lambertini, L.; Lauriola, M.; Padovani, M.; Maltoni, C. Results of long-term
experimental studies on the carcinogenicity of formaldehyde and acetaldehyde in Rats. Ann. N. Y. Acad.
Sci. 2002, 982, 87–105.
58. Stavridis, J.C. Toxicity and carcinogenicity of aldehydes. In Oxidation: The Cornerstone of
Carcinogenesis. Oxidation and Tobacco Smoke Carcinogenesis. A Relationship Between Cause and Effect; Stavridis,
J.C., Ed.; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2007; pp. 161–173,
doi:10.1007/978-1-4020-6704-4_11.
59. Emerit, I. Reactive oxygen species, chromosome mutation, and cancer: Possible role of clastogenic factors
in carcinogenesis. Free Rad. Biol. Med. 1994, 36, 99–109.
60. Benigni, R.; Passerini, L.; Rodomonte, A. Structure–activity relationships for the mutagenicity and
carcinogenicity of simple and α-β unsaturated aldehydes. Environ. Mol. Mutagen. 2003, 42, 136–143.
doi:10.1002/em.10190.
61. Young, S.-C.; Chang, L.W.; Lee, H.-L.; Tsai, L.-H.; Liu, V.-C.; Lisn, P. DNA damage induced by trans,trans-
2,4-decadienal (tt-DD), a component of cooking oil fume, in human bronchial epithelial cells. Environ. Mol.
Mutagen. 2010, 51, 315–321.
62. Jayaraj, A.P.; Rees, K.R.; Tovey, F.E.I.; White, J.S. A molecular basis of peptic ulceration due to diet. Br. J.
Exp. Path. 1986, 67, 149–155.
63. Benedetti, A.; Ferrali, A.; Casini, A.F.; Peiri, S.; Comporti, M. Foot edema induced by carbonyl compounds
originating from the peroxidation of microsomal lipids. Biochem. Pharmacol. 1980, 29, 121–124.
64. Indart, A.; Viana, M.; Grootveld, M.; Silwood, C.; Sanchez-Vera, I.; Bonet Serra, B. Teratogenic actions of
thermally-stressed COs in rats. Free Rad. Res. 2002, 36, 1051–1058.
65. Long, E.K.; Murphy, T.C.; Leiphon, L.J.; Watt, J.; Morrow, J.D.; Milne, G.L.; Howard, J.R.H.; Picklo, M.J., Sr.
Trans-4-hydroxy-2-hexenal is a neurotoxic product of docosahexaenoic (22:6; n-3) acid oxidation. J.
Neurochem. 2008, 105, 714–724.
66. Leong, X.-F.; Mustafa, M.R.; Das, S.; Jaarin, K. Association of elevated blood pressure and impaired
vasorelaxation in experimental Sprague-Dawley rats fed with heated vegetable oil. Lipids Health Dis. 2010,
9, 66.
67. Chiang, Y.F.; Shaw, H.M.; Yang, M.F.; Huang, C.Y.; Hsieh, C.H.; Chao, P.M. Dietary oxidised frying oil
causes oxidative damage of pancreatic islets and impairment of insulin secretion, effects associated with
vitamin E deficiency. Br. J. Nutr. 2011, 105, 1311–1319. doi:10.1017/S0007114510005039.
68. Bein, K.; Leikauf, G. Acrolein—A pulmonary hazard. Molec. Nutr. Food Res. 2011, 55, 1342–1360.

Nutrients 2020, 12, 974 44 of 49
69. Lee, T.; Gany, F. Cooking oil fumes and lung cancer: A review of the literature in the context of the U.S.
population. J. Immigr. Minor Health 2013, 15, 646–652.
70. Xue, Y.; Jiang, Y.; Jin, S.; Li, Y. Association between cooking oil fume exposure and lung cancer among
Chinese nonsmoking women: A meta-analysis. Onco. Targets Ther. 2016, 9, 2987–2992.
doi:10.2147/OTT.S100949.
71. Yin, Z.; Cui, Z.; Guan, P.; Li, X.; Wu, W.; Ren, Y.; He, Q.; Zhou, B. Interaction between polymorphisms in
Pre-MiRNA genes and cooking oil fume exposure on the risk of lung cancer in Chinese non-smoking female
population. PLoS One. 2015, 10(6), e0128572.
72. Yu, I.T.S.; Chiu, Y.-L.; Au, J.S.K.; Wong, T.-W.; Tang, J-L. Dose-response relationship between cooking
fumes exposures and lung cancer among Chinese nonsmoking women. Cancer Res. 2006, 66, 4961–4967.
73. Grootveld, M.; Atherton, M.D.; Sheerin, A.N.; Hawkes, J.; Blake, D.R.; Richens, T.E.; Silwood, C.J.L.; Lynch,
E.; Claxson, A.W.D. In vivo absorption, metabolism, and urinary excretion of ,-unsaturated aldehydes
in experimental animals. Relevance to the development of cardiovascular diseases by the dietary ingestion
of thermally stressed polyunsaturate-rich culinary oils. J. Clin. Invest. 1998, 101, 1210–1218.
74. Kaye, C.M. Biosynthesis of mercapturic acids from allyl alcohol, allyl esters and acrolein. Blochem. J. 1973,
134, 1093–1101.
75. McGirr, L.G.; Hadley, M.; Draper, H.H. Identification of N alpha-acetyl-epsilon-(2-propenal)lysine as a
urinary metabolite of malondialdehyde. J. Biol. Chem. 1985, 260, 15427–15431.
76. Awada, M.; Soulage, C.O.; Meynier, A.; Debard, C.; Plaisancié, P.; Benoit, B.; Picard, G.; Loizon, E.;
Chauvin, M.-A.; Estienne, M.; et al. Dietary oxidized n-3 PUFA induce oxidative stress and inflammation:
Role of intestinal absorption of 4-HHE and reactivity in intestinal cells. J. Lipid Res. 2012, 53, 2069–2080.
doi:10.1194/jlr.M026179.
77. Csala, M.; Kardon, T.; Legeza, B.; Lizák, B.; Mandl, J.; Margittai, É.; Puskás, F.; Száraz, P.; Szelényi, P.;
Bánhegyi, G. On the role of 4-hydroxynonenal in health and disease. Biochimica et Biophysica Acta 2015, 1852,
826–838.
78. Srivastava, S.; Dixit, B.L.; Cai, J.; Sharma, S.; Hurst, H.E.; Bhatnagar, A.; Srivastava, S.K. Metabolism of lipid
peroxidation product, 4-hydroxynonenal (HNE) in rat erythrocytes: Role of aldose reductase. Free Rad. Biol.
Med. 2000, 29, 642–651.
79. Mak, S.; Lehotay, D.C.; Yazdanpanah, M.; Azevedo, E.R.; Liu, P.P.; Newton, G.E. Unsaturated aldehydes
including 4-OH-nonenal are elevated in patients with congestive heart failure. J. Card. Fail. 2000, 6, 109–114.
80. Allen, S.A.; Clark, W.; McCaffery, J.M.; Cai, Z.; Lanctot, A.; Slininger, P.J.; Liu, Z.L.; Gorsich, S.W. Furfural
induces reactive oxygen species accumulation and cellular damage in Saccharomyces cerevisiae. Biotechnol.
Biofuels 2010, 3, 2. doi:10.1186/1754-6834-3-2
81. Montgomery, J.A.; Jette, M.; Huot, S. Desrosiers C: Acyloin production from aldehydes in the perfused rat
heart: The potential role of pyruvate dehydrogenase. Biochem. J. 1993, 294, 727–733.
82. Witz, G. Biological interactions of α,β-unsaturated aldehydes. Free Rad. Biol. Med. 1988, 7, 333–349.
83. Srivastava, S.; Chandra, A.; Wang, L.F.; Seifert, W.E.; Dague, B.B.; Ansari, N.H.; Srivastava, S.D.; Bhatnagar,
A. Metabolism of the lipid peroxidation product, 4-hydroxy-trans-2-nonenal, in isolated perfused rat heart.
J. Biol. Chem. 1998, 273, 10893–10900.
84. Kroes, R. WHO Food Additives Series: 42. Furfural. Prepared by the Fifty-first Meeting of the Joint
FAO/WHO Expert Committee on Food Additives (JECFA). World Health Organization, Geneva, 1999.
International Program on Chemical Safety (IPCS) INCHEM Home.
85. Conklin, D.J.; Barski, O.A.; Lesgards, J.F.; Juvan, P.; Rezen, T.; Rozman, D.; Prough, R.A.; Vladykovskaya,
E.; Liu, S.; Srivastava, S. Acrolein consumption induces systemic dyslipidemia and lipoprotein modification.
Toxicol. Appl. Pharmacol. 2010, 243, 1–12.
86. Conklin, D. J.; Bhatnagar, A. Chapter 6.26 - Aldehydes and Cardiovascular Disease; In Comprehensive
Toxicology Reference Work (2nd Edn.); Ed: McQueen, C. A., Ed.; Elsevier, Amsterdam, The Netherlands.
2010, 6, 489–512.
87. Ogihara, T.; Hirano, K.; Morinobu, T.; Kim, H.-S.; Hiroi, M.; Ogihara, H.; Tamai, H. Raised concentrations
of aldehyde lipid peroxidation products in premature infants with chronic lung disease. Arch. Dis. Child
Fetal Neonatal Ed. 1999, 80, F21–F25.
88. Levitan, I.; Volkov, S.; Subbaiah, P.V. Oxidized LDL: Diversity, patterns of recognition, and
pathophysiology. Antioxid. Redox Signal. 2010, 13, 39–75. doi:10.1089/ars.2009.2733.

Nutrients 2020, 12, 974 45 of 49
89. Uchida, K. Role of reactive aldehyde in cardiovascular diseases. Free Rad. Biol. Med. 2000, 28, 1685–1696,
doi:10.1016/S0891-5849(00)00226-4.
90. Hoff, H.F.; O'Neil, J.; Wu, Z.; Hoppe, G.; Salomon, R.L. Phospholipid hydroxyalkenals. Biological and
chemical properties of specific oxidized lipids present in atherosclerotic lesions. Arterioscler. Thromb. Vasc.
Biol. 2003, 23, 275–282. doi:10.1161/01.ATV.0000051407.42536.73
91. Steinberg, D.; Parthasarathy, S.; Carew, T.E.; Khoo, J.C.; Witztum, J.L. Modification of low-density
lipoprotein that increases its atherogenecity. N. Engl. J. Med. 1989, 320, 915–924.
92. Steinberg, D. Role of oxidized LDL and antioxidants in atherosclerosis. Adv. Exp. Med. Biol. 1995, 369, 39–
48. doi:10.1007/978-1-4615-1957-7_5
93. Uchida, K.; Sakai, K.; Itakura, K.; Osawa, T.; Toyokuni, S. Protein modification by lipid peroxidation
products: foration of malondialdehyde-derived Ne-(2-propenal)lysine in proteins. Arch. Biochem. Biophys.
1997, 346, 45–52. doi:10.1006/abbi.1997.0266
94. Requena, J.R.; Fu, M.X.; Ahmed, M.U.; Jenkins, A.J.; Lyons, T.J.; Baynes, J.W.; Thorpe, S.R. Quantification
of malondialdehyde and 4-hydroxynonenal adducts to lysine residues in native and oxidized human low-
density lipoprotein. Biochem. J. 1997, 322, 317–325.
95. Obama, T.; Kato, R.; Masuda, Y.; Takahashi, K.; Aiuchi, T.; Itabe, H. Analysis of modified apolipoprotein
B-100 structures formed in oxidized low-density lipoprotein using LC-MS/MS. Proteomics 2007, 7, 2132–
2141. doi:10.1002/pmic.200700111
96. Uchida, K.; Kanematsu, M.; Sakai, K.; Matsuda, T.; Hattori, N.; Mizuno, Y.; Suzuki, D.; Miyata, T.; Noguchi,
N.; Niki, E.; et al. Protein-bound acrolein: Potential markers for oxidative stress. Proc. Natl. Acad. Sci. USA
1998, 95, 4882–4887. doi:10.1073/pnas.95.9.4882.
97. Uchida, K.; Kanematsu, M.; Morimitsu, Y.; Osawa, T.; Noguchi, N.; Niki, E. Acrolein is a product of lipid
peroxidation reaction: Ormation of acrolein and its conjugate with lysine residues in oxidized low-density
lipoprotein. J. Biol. Chem. 1998, 273, 16058–16066. doi:10.1074/jbc.273.26.16058.
98. Uchida, K.; Toyokuni, S.; Nishikawa, K.; Kawokishi, S.; Oda, H.; Hiai, H.; Stadtman, E.R. Michael addition-
type 4-hydroxy-2-nonenal adducts in modified low density lipoproteins: Arkers for atherosclerosis.
Biochemistry 1994, 33, 12487–12494. doi:10.1021/bi00207a016
99. Tamamizu-Kato, S.; Wong, J.Y.; Jairam, V.; Uchida, K. Modification by acrolein, a component of tobacco
smoke and age-related oxidative stress, mediates functional impairment of human apolipoprotein E.
Biochemistry 2007, 46, 8392–8400. doi:10.1021/bi700289k
100. Watanabe, K.; Nakazato, Y.; Saiki, R.; Igarashi, K.; Kitada, M.; Ishii, I. Acrolein-conjugated low-density
lipoprotein induces macrophage foam cell formation. Atherosclerosis 2013, 227, 51–57.
doi:10.1016/j.atherosclerosis.2012.12.020.
101. 102. Humphries, K.M.; Szweda, L.I. Selective inactivation of alpha-ketoglutarate dehydrogenase and
pyruvate dehydrogenase: Reaction of lipoic acid with 4-hydroxy-2-nonenal. Biochemistry 1998, 37, 15835–
15841.
102. 101. Bhatnagar, A. Electrophysiological effects of 4-hydroxynonenal, an aldehydic product of lipid
peroxidation, on isolated rat ventricular myocytes. Circ. Res. 1995, 76, 293–304.
103. Luo, J.; Hill, B.G.; Gu, Y.; Cai, J.; Srivastava, S.; Bhatnagar, A.; Prabhu, S.D. Mechanisms of acrolein-induced
myocardial dysfunction: Implications for environmental and endogenous aldehyde exposure. Am. J.
Physiol. Heart Circ. Physiol. 2007, 293, H3673–H3684.
104. Ismahil, M.A.; Hamid, T.; Haberzettl, P.; Gu, Y.; Chandrasekar, B.;, Srivastava, S.; Prabhu, S.D. Chronic oral
exposure to the aldehyde pollutant acrolein induces dilated cardiomyopathy. Am. J. Physiology. Heart Circ.
Physiol. 2011, 301, H2050–H2060. doi:10.1152/ajpheart.00120.2011
105. Chen, H.J.; Wang, C.C.; Chan, D.C.; Chiu, C.Y.; Yang, R.S.; Liu, S.H. Adverse effects of acrolein, a
ubiquitous environmental toxicant, on muscle regeneration and mass. J. Cachexia Sarcopenia Muscle 2019,
10, 165–176. doi:10.1002/jcsm.12362
106. Moghe, A.; Ghare, S.; Lamoreau, B.; Mohammad, M.; Barve, S.; McClain, C.; Joshi-Barve, S. Molecular
mechanisms of acrolein toxicity: Relevance to human disease. Toxicolog. Sci. 2015, 143, 242–255.
doi:10.1093/toxsci/kfu233.
107. Gadiraju, T.V.; Patel, Y.; Gaziano, J.M.; Djoussé, L. Fried food consumption and cardiovascular health: A
review of current evidence. Nutrients 2015, 7, 8424–8430, doi:10.3390/nu7105404.
108. Cho, Y.; Song, M.; Kim, T.S.; Ryu, J.C. DNA Methylome analysis of saturated aliphatic aldehydes in
pulmonary toxicity. Sci. Rep. 2018, 8, 10497, doi:10.1038/s41598-018-28813-z.

Nutrients 2020, 12, 974 46 of 49
109. Eckl, P.M.; Bresgen, N. Review Article: Genotoxicity of lipid oxidation compounds. Free Radic. Biol. Med.
2017, 111, 244–252.
110. Dung, C. H.; Wu, S. C.; Yen, G. C. Genotoxicity and oxidative stress of the mutagenic compounds formed
in fumes of heated soybean oil, sunflower oil and lard. Toxicol In Vitro. 2006, 20, 439-447.
111. Straif, K.; Baan, R.; Grosse, Y.; Secretan, B.; El Ghissassi, F.; Cogliano, V. Carcinogenicity of household solid
fuel combustion and of high-temperature frying. Lancet Oncol. 2006, 7, 977–978.
112. Janzowski, C.; Glaab, V.; Mueller, C.; Straesser, U.; Kamp, H.G.; Eisenbrand, G. α,β-Unsaturated carbonyl
compounds: Induction of oxidative DNA damage in mammalian cells. Mutagenesis 2003, 18, 465–470.
doi:10.1093/mutage/geg018.
113. Feron, V.J.; Til, H.P.; deVrijer, F.; Woutersen, R.A.; Cassee, F.R.; van Bladeren, P.J. Aldehydes: Occurrence,
carcinogenic potential, mechanism of action and risk assessment. Mutat. Res. 1991, 259, 363–385.
114. Van der Veen, L.A.; Hashim, M.F.; Shyr, Y.; Marnett, L.J. Induction of frameshift and base pair substitution
mutations by the major DNA adduct of the endogenous carcinogen malondialdehyde. Proc. Natl. Acad. Sci.
USA 2003, 100, 14247–14252. doi:10.1073/pnas.2332176100.
115. Firestone, D. Worldwide regulation of frying fats and oils. Inform Am. Oil Chem. Soc. 1993, 4, 1366–1371.
116. Yonei, S.; Furui, H. Lethal and mutagenic effects of malondialdehyde, a decomposition product of
peroxidized lipids, on Escherichia coli with different DNA-repair capacities. Mutat. Res./Genet. Toxicol.
1981, 88, 23–32.
117. Seto, H.; Okuda, T.; Takesue, T.; Ikemura, T. Reaction of malonaldehyde with nucleic acid. I. Formation of
fluorescent pyrimido(1,2-a)purin-10(3H)-one nucleosides. Bull. Chem. Soc. Jpn. 1983, 56, 1799–1802.
118. Leuratti, C.; Watson, M.A.; Deag, E.J.; Welch, A.; Singh, R.; Gottschalg, E.; Marnett, L.J.; Atkin, W.; Day,
N.E.; Shuker, D.E.G.; et al. Detection of malondialdehyde DNA adducts in human colorectal mucosa.
Cancer Epidemiol. Biomark. Prev. 2002, 11, 267–273.
119. Draper, H.H.; McGirr, L.G.; Hadley, M. The metabolism of malondialdehyde. Lipids 1986, 21, 305.
doi:10.1007/BF02536418.
120. National Toxicology Program. Toxicology and Carcinogenesis Studies of Malonaldehyde, Sodium Salt; Technical
Report; National Institutes of Health: Research Triangle Park, NC, USA, 1988; Volume 331; pp. 5–13.
121. Basu, A.K.; Marne, Â.L.J. Molecular requirements for the mutagenicity of malondialdehyde and related
acroleins. Cancer Res. 1984, 44, 2848–2854.
122. Marnett, L.J.; Hurd, H.K.; Hollstein, M.C.; Levin, D.E.; Esterbauer, H.; Ames, B.N. Naturally occurring
carbonyl compounds are mutagens Salmonella tester strain TA104. Mutat. Res./Fundam. Mol. Mech.
Mutagen. 1985, 148, 25–34.
123. Canonero, R.; Martelli, A.; Marinari, U.M.; Brambilla, G. Mutation induction in Chinese hamster lung V79
cells by five alk-2-enals produced by lipid peroxidation. Mutat. Res. Lett. 1990, 244, 153–156.
124. Kiwamoto, R.; Spenkelink, A.; Rietjens, I.M.C.M.; Punt, A. A physiologically based in silico model for trans-
2-hexenal detoxification and DNA adduct formation in human including interindividual variation
indicates efficient detoxification and a negligible genotoxicity risk. Toxicokinet. Metab. 2013, 87, 1725–1737.
125. Nadasi, E.; Varjas, T.; Pajor, L.; Ember, I. Carcinogenic potential of trans-2-hexenal is based on epigenetic
effect. In Vivo 2005, 19, 559–562.
126. Cohen, S.M.; Garland, E.M.; St John, M.; Okamura, T.; Smith, R.A. Acrolein initiates rat urinary bladder
carcinogenesis. Cancer Res. 1992, 52, 3577–3581.
127. Kawai, Y.; Furuhata, A.; Toyokuni, S.; Aratani, Y.; Uchida, K. Formation of acrolein-derived 2’-
deoxyadenosine adduct in an iron-induced carcinogenesis model. J. Biol. Chem. 2003, 278, 50346–50354.
128. Nath, R. G.; Ocando, J. E.; Guttenplan, J. B.; Chung, F-L. N2-Propanodeoxyguanosine adducts: Potential
new biomarkers of smoking-induced DNA damage in human oral tissue. Cancer Res. 1998, 58(4), 581-584;
129. Lee, H.W.; Wang, H.T.; Weng, M.W.; Hu, Y.; Chen, W.S.; Chou, D.; Liu, Y.; Donin, N.; Huang, W.C.; Lepor,
H.; et al. Acrolein- and 4-aminobiphenyl-DNA adducts in human bladder mucosa and tumor tissue and
their mutagenicity in human urothelial cells. Oncotarget 2014, 5, 3526–3540.
130. Hecht, S.S.; Koh, W.P.; Wang, R.; Chen, M.; Carmella, S.G.; Murphy, S.E.; Yuan, J.M. Elevated levels of
mercapturic acids of acrolein and crotonaldehyde in the urine of Chinese women in Singapore who
regularly cook at home. PLoS ONE 2015, 10, e0120023. doi:10.1371/journal.pone.0120023.
131. Stevens J. F.; Maier, C. S. Acrolein: sources, metabolism, and biomolecular interactions relevant to human
health and disease. Mol.. Nutr. Food Res.. 2008, 52, 7–25.

Nutrients 2020, 12, 974 47 of 49
132. Metayera, C.; Wangb, Z.; Kleinermana, R.A.; Wangc, L.; Brennera, A.V.; Cuib, H.; Caob, J.; Lubin, J.H.
Cooking oil fumes and risk of lung cancer in women in rural Gansu, China. Lung Canc. 2002, 35, 111–117.
133. Beland, F. A.; Olson, G. R.; Mendoza, M. C.; Marques, M. M.; Doerge, D. R. Carcinogenicity of glycidamide
in B6C3F 1 mice and F344/N rats from a two-year drinking water exposure. Food Chem. Toxicol. 2015, 86,
104-115.
134. Feng, Z.; Hu, W.; Hu, Y.; Tang, M.-S. Acrolein is a major cigarette-related lung cancer agent: Preferential
binding at p53 mutational hotspots and inhibition of DNA repair. Proc. Natl. Acad. Sci. USA 2006, 103,
15404–15409. doi:10.1073/pnas.0607031103
135. Singer, B.; Muriseli, H. (Eds.) The Role of Cyclic Nucleic Acid Adducts in Carcinogenesis and Mutagenesis; IARC
Scientific Publications, Volume 70; International Agency for Research on Cancer: Lyon, France, 1986.
136. Smith, R.A.; Cohen, S.M.; Lawson, T.A. Acrolein mutagenicity in the V79 assay. Carcinogenesis (Lond.) 1990,
11, 497–498.
137. van Andel, I.; Sleijffers, A.; Schenk, E.; Rambali, E.; Wolterink, G.; van der Werken, G.; van Aerts, L. A. G.
J. M.; Vleeming, W.; van Amsterdam, J. G. C. Adverse Health Effects of Cigarette Smoke: Aldehydes
Crotonaldehyde, Butyraldehyde, Hexanal, and Malonaldehyde; RIVM report 340630002/2006; 2006. Nutrition,
Health Protection and Prevention Department, Ministry of Health, Welfare and Sports (VWS) and of the
Food and Consumer Product Safety Authority (VWA). Bilthoven, The Netherlands.
138. Chung, F.-L.; Tanaka, T.; Hecht, S.S. Induction of liver tumors in F344 rats by crotonaldehyde. Cancer Res.
1986, 46, 1285–1289.
139. Lutz, D.; Eder, E.; Neudecker, T.; Henschler, D. Structure-mutagenicity relationships in unsaturated
carbonylic compounds and their corresponding allylic alcohol. Mutat. Res. 1982, 93, 305–315.
140. Laskar, A. A.; Younus, H. (2018): Aldehyde toxicity and metabolism:the role of aldehyde dehydrogenases
in detoxification, drug resistance and carcinogenesis, Drug Metab. Revs., 2018, 1-24. DOI:
10.1080/03602532.2018.1555587
141. Silano, V.; Bolognesi, C.; Castle, L.; Cravedi, J.-P.; Engel, K.-H.; Fowler, P.; Franz, R.; Grob, K.; Husøy, T.;
Karenlampi, S.; et al. Scientific Opinion on Flavouring Group Evaluation 226 Revision 1 (FGE.226 Rev1):
consideration of genotoxicity data on -unsaturated aldehyde from chemical subgroup 1.1.1(b) of FGE.19
EFSA Panel on Food Contact Materials, Enzymes, Flavourings and Processing Aids (CEF). ADOPTED: 4
EFSA J. 2017, doi:10.2903/j.efsa.2017.4847.
142. Salaspuro, M. Acetaldehyde and gastric cancer. J. Digest. Dis. 2011, 12, 51–59.
143. Matthews, R.F.; Scanlan, R.A.; Libbey, L.M. Autoxidation products of 2,4-decadienal. J. Am. Oil Chem. Soc.
1971, 48, 745–747.
144. Zhang, Q.; Qin, W.; Lin, D.; Shen, Q.; Saleh, A.S.M. The changes in the volatile aldehydes formed during
the deep-fat frying process. J. Food Sci. Technol. 2015, 52, 7683–7696. doi:10.1007/s13197-015-1923-z.
145. Andrikopoulos, N.K.; Boskou, G.; Dedoussis, G.V.Z.; Chiou, A.; Tzamtzis, V.A.; Papathanasiou, A. A
Quality assessment of frying oils and fats from 63 restaurants in Athens, Greece. Food Serv. Techn. 2003, 3,
49–59.
146. Tan, S.L.W.; Chadha, S.; Liu, Y.; Gabasova, E.; Perera, D.; Ahmed, K.; Constantinou, S.; Renaudin, X.; Lee,
M.; Aebersold, R.; et al. A class of environmental and endogenous toxins induces BRCA2
haploinsufficiency and genome instability. Cell 2017, doi:10.1016/j.cell.2017.05.010.
147. International Agency for Research on Cancer (June 2004). IARC Monographs on the Evaluation of
Carcinogenic Risks to Humans Volume 88 (2006): Formaldehyde, 2-Butoxyethanol and 1-tert-
Butoxypropan-2-ol. Available online: http://monographs.iarc.fr/ENG/Monographs/vol88/index.phpExit
Disclaimer (accessed on 10 June 2011).
148. National Toxicology Program. Report on Carcinogens, Twelfth Edition. Department of Health and Human
Services, Public Health Service, National Toxicology Program. Available online:
http://ntp.niehs.nih.gov/go/roc12 (accessed on 10 June 2011).
149. Stott-Miller, M.; Neuhouser, M.L.; Stanford, J.L. Consumption of deep-fried foods and risk of prostate
cancer. Prostate 2013, 73, 960–969. doi:10.1002/pros.22643.
150. Lippi, G.; Mattiuzzi, C. Fried food and prostate cancer risk: Systematic review and meta-analysis. Int. J.
Food Sci. Nutr. 2015, 66, 587–589.
151. Knekt, P.; Steineck, G.; Järvinen, R.; Hakulinen, T.; Aromaa, A. Intake of fried meat and risk of cancer: A
follow-up study in Finland. Int. J. Cancer 1994, 59, 756–760. doi:10.1002/ijc.2910590608.

Nutrients 2020, 12, 974 48 of 49
152. Bosetti, C.; Talamini, R.; Levi, F.; Negri, E.; Franceschi, S.; Airoldi, L.; La Vecchia, C. Fried foods: A risk
factor for laryngeal cancer? Br. J. Cancer 2002, 87, 1230–1233.
153. Srivastava, S.M.; Singh, M.; George, J.; Bhui, K.; Shukla, Y. Genotoxic and carcinogenic risks associated with
the consumption of repeatedly boiled sunflower oil. J. Agric. Food Chem. 2010, 58, 11179–11186.
154. Woutersen, R.A.; Appel, M.J.; van Garderen-Hoetmer, A.; Wijnands, M. V. Dietary fat and carcinogenesis.
Mutat. Res. 1999, 443, 111–127.
155. Xie, M.Z.; Shoulkamy, M.I.; Salem, A.M.H.; Oba, S.; Goda, M.; Nakano, T.; Ide, H. Aldehydes with high
and low toxicities inactivate cells by damaging distinct cellular targets. Mutat. Res./Fund. Molec. Mechan.
Mutagenesis 2016, 786, 41–51.
156. Richie, J.P., Jr.; Kleinman, W.; Marina, P.; Abraham, P.; Wynder, E.L.; Muscat, J.E. Blood iron, glutathione,
and micronutrient levels and the risk of oral cancer. Nutr. Cancer 2008, 60, 474–482.
doi:10.1080/01635580801956477.
157. Esterbauer, H.; Ertl, A.; Scholz, N. The reaction of cysteine with α,β-unsaturated aldehydes. Tetrahedron
1976, 32, 285–289.
158. Gan, J.C.; Oandasan, A.; Ansari, G.A.S. In vitro covalent modification of serum albumin by acrolein.
Chemosphere 1991, 23, 939–947. doi:10.1016/0045-6535(91)90098-X.
159. LoPachin, R.M.; Gavin, T. Molecular mechanisms of aldehyde toxicity: A chemical perspective. Chem. Res.
Toxicol. 2014, 27, 1081–1091. dx.doi.org/10.1021/tx5001046
160. Negre-Salvayre, A.; Vieira, O.; Escargueil-Blanc, I.; Salvayre, R. Oxidized LDL and 4-hydroxynonenal
modulate tyrosine kinase receptor activity. Mol. Aspects Med. 2003, 24, 251–261. doi:10.1016/S0098-
2997(03)00020-7.
161. Pizzimenti, S.; Ciamporcero, E.; Daga, M.; Pettazzoni, P.; Arcaro, A.; Cetrangolo, G.; Minelli, R.; Dianzani,
C.; Lepore, A.; Gentile, F.; et al. Interaction of aldehydes derived from lipid peroxidation and membrane
proteins. Front. Physiol. 2013, 4, 242. doi:10.3389/fphys.2013.00242.
162. Jacobs, A.T.; Marnett, L.J. Systems analysis of protein modification and cellular responses induced by
electrophile stress. Acc. Chem. Res. 2010, 43, 673–683, doi:10.1021/ar900286y.
163. Gentile, F.; Pizzimenti, S.; Arcaro, A.; Pettazzoni, P.; Minelli, R.; D'Angelo, D.; Mamone, G.; Ferrant, P.;
Toaldo, C.; Cetrangolo, G.; et al. Exposure of HL-60 human leukaemic cells to 4-hydroxynonenal promotes
the formation of adduct(s) with alpha-enolase devoid of plasminogen binding activity. Biochem. J. 2009, 422,
285–294. doi:10.1042/BJ20090564.
164. Barrera, G. Oxidative stress and lipid peroxidation products in cancer progression and therapy. ISRN
Oncol. 2012, 2012, 137289, doi:10.5402/2012/137289.
165. Conklin, D.J.; Bhatnagar, A. 6.26—Aldehydes and Cardiovascular Disease. In Comprehensive Toxicology 2nd
ed.; McQueen, C.A., Ed.; Elsevier: Amsterdam, The Netherlands, 2010; pp. 489–512, doi:10.1016/B978-0-08-
046884-6.00723-5.
166. Sottero, B.; Leonarduzzi, G.; Testa, G.; Gargiulo, S.; Poli, G.; Biasi, F. Lipid Oxidation Derived Aldehydes
and Oxysterols Between Health and Disease. Eur. J. Lipid Sci. Technol. 2019, 12, 1700047.
doi:10.1002/ejlt.201700047
167. Goicoechea, E.; Van Twillert, K.; Duits, M.; Brandon, E.D.; Kootstra, P.R.; Blokland, M.H.; Guillén, M.D.
Use of an in vitro digestion model to study the bioaccessibility of 4-hydroxy-2-nonenal and related
aldehydes present in oxidized oils rich in omega-6 acyl groups. J. Agric. Food Chem. 2008, 56, 8475–8483.
doi:10.1021/jf801212k.
168. EFSA Panel on Biological Hazards (BIOHAZ). Scientific Opinion on Fish Oil for Human Consumption.
EFSA J. 2010, 8, 1874.
169. Kenmogne-Domguia, H.B.; Moisan, S.; Viau, M.; Genot, C.; Meynier, A. The initial characteristics of marine
oil emulsions and the composition of the media inflect lipid oxidation during in vitro gastrointestinal
digestion. Food Chem. 2014, 152, 146–154. doi:10.1016/j.foodchem.2013.11.096.
170. Steppeler, C.; Haugen, J.E.; Rødbotten, R.; Kirkhus, B. Formation of malondialdehyde, 4-hydroxynonenal,
and 4-hydroxyhexenal during in vitro digestion of cooked beef, pork, chicken, and salmon. J. Agric. Food
Chem. 2016, 64, 487–496. doi:10.1021/acs.jafc.5b04201.
171. Calzada, C.; Colas, R.; Guillot, N.; Guichardant, M.; Laville, M.; Véricel, E.; Lagarde, M. Subgram daily
supplementation with docosahexaenoic acid protects low-density lipoproteins from oxidation in healthy
men. Atherosclerosis 2010, 8, 467–472. doi:10.1016/j.atherosclerosis.2009.07.049.

Nutrients 2020, 12, 974 49 of 49
172. Van Kuijk, F.J.; Holte, L.L.; Dratz, E.A. 4-hydroxyhexenal: A lipid peroxidation product derived from
oxidized docosahexaenoic acid. Biochim. Biophys. Acta 1990, 1043, 116–118.
173. Nakagawa, F.; Morino, K.; Ugi, S.; Ishikado, A.; Kondo, K.; Sato, D.; Konno, S.; Nemoto, K.-I.; Kusunoki,
C.; Sekine, O.; et al. 4-Hydroxy hexenal derived from dietary n-3 polyunsaturated fatty acids induces anti-
oxidative enzyme heme oxygenase-1 in multiple organs. Biochem. Biophys. Res. Commun. 2014, 443, 991–996.
174. Tsikas, D.; Rothmann, S.; Schneider, J.Y.; Gutzki, F.M.; Beckmann, B.; Frölich, J.C. Simultaneous GC-MS/MS
measurement of malondialdehyde and 4-hydroxy-2-nonenal in human plasma: Effects of long-term L-
arginine administration. Anal. Biochem. 2017, 524, 31–44. doi:10.1016/j.ab.2016.08.009.
175. Gindzienska-Sieskiewicz, L.W.; Jarocka-Karpowicz, E.; Andrisic, I.; Sierakowski, L.; Zarkovic, S.; Waeg, N.;
Skrzydlewska, G. The onset of lipid peroxidation in rheumatoid arthritis: Consequences and monitoring.
Free Radic. Res. 2016, 50, 304–313. doi:10.3109/10715762.2015.1112901.
176. Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling
mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 360, 438.
doi:10.1155/2014/360438.
177. Zhang, H.; Forman, H.J. 4-hydroxynonenal-mediated signaling and aging. Free Radic. Biol. Med. 2017, 111,
219–225. doi:10.1016/j.freeradbiomed.2016.11.032.
178. Łuczaj, W.; Skrzydlewska, G.A. Antioxidants and HNE in redox homeostasis. Free Radic. Biol. Med. 2017,
111, 87–101. doi:10.1016/j.freeradbiomed.2016.11.033.
179. Dianzani, M.U. 4-hydroxynonenal from pathology to physiology. Mol. Aspects Med. 2003, 24, 263–272.
180. Weng, M.-W.; Lee, H.-W.; Park, S.-Y.; Hu, Y.; Wang, H.-T.; Chen, L.-C.; Rom, W.N.; Huang, W.C.; Lepro,
H.; Wu, X.-R.; et al. Aldehydes are the predominant forces inducing DNA damage and inhibiting DNA
repair in tobacco smoke carcinogenesis. Proc. Natl. Acad. Sci. USA 2018, 115, E6152–E6161,
doi:10.1073/pnas.1804869115
181. Mirzael, H.O.; Salehi, F.; Garovni, H.; Farhadpour, F.J. Role of MC compounds at less of oil absorption of
French fries potato. Nutr. Health Food Eng. 2015, 2, 00075.
182. Kaminskas, L.M.; Pyke, S.M.; Burcham, P.C. Reactivity of hydrazinophthalazine drugs with the lipid
peroxidation products acrolein and crotonaldehyde. Org. Biomol. Chem. 2004, 2, 2578–2584.
183. Leung, G.; Sun, W.; Zheng, L.; Brookes, S.; Tully, M.; Shi, R. Anti-acrolein treatment improves behavioral
outcome and alleviates myelin damage in experimental autoimmune encephalomyelitis mouse.
Neuroscience. 2011, 173, 150–155, doi:10.1016/j.neuroscience.2010.11.018.
184. National Research Council (US) Committee on Technological Options to Improve the Nutritional
Attributes of Animal Products. 4. Consumer Concerns and Animal Product Options. In Designing Foods:
Animal Product Options in the Marketplace; National Academies Press (US): Washington, DC, USA, 1988;
Available online: https://www.ncbi.nlm.nih.gov/books/NBK218169/ (accessed on 2 July 2019).
185. World Health Organization. Formaldehyde: Concise International Chemical Assessment Document 40;
Wissenschaftliche Verlagsgesellschaft: Geneva, Switzerland, 2002.
186. Shibamoto, T. Analytical methods for trace levels of reactive carbonyl compounds formed in lipid
peroxidation systems. J. Pharm. Biomed. Anal. 2006, 41, 12–25, doi:10.1016/j.jpba.2006.01.047.
187. SCCS (Scientific Committee on Consumer Safety). Opinion on Acetaldehyde; 18 September 2012—revision 11
December 2012; European Commission: Brussels, Belgium, 2012; doi:10.2772/8968, ISSN 1831-4767, ISBN
978-92-79-30778-2.
188. National Toxicology Program; P.H.S. National Institutes of Health; U.S. Department of Health and Human
Services. 2,4-Decadienal CAS No. 25152–84–5. Testing Status of Agents at NTP; 1993. National Toxicology
Program, Research Triangle Park, NC, USA.
189. Wu, S.C.; Yen, G.C.; Sheu, F. Mutagenicity and identification of mutagenic compounds of fumes obtained
from heating peanut oil. J. Food Prot. 2001, 64, 240–245.
190. Duong, A.; Steinmaus, C.; McHale, C.M.; Vaughan, C.P.; Zhang, L. Reproductive and developmental
toxicity of formaldehyde: A systematic review. Mutat. Res. 2011, 728, 118–138,
doi:10.1016/j.mrrev.2011.07.003
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access
article distributed under the terms and conditions of the Creative Commons Attribution
(CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright © 2022 FDOKUMEN




















