
should Australia still be considered free from Hepatozoon canis
38
RESEARCH REPOSITORY This is the author’s final version of the work, as accepted for publication following peer review but without the publisher’s layout or pagination. The definitive version is available at: https://doi.org/10.1016/j.ijpara.2018.05.002 Greay, T.L., Barbosa, A.D., Rees, R.L., Paparini, A., Ryan, U.M., Oskam, C.L. and Irwin, P.J. (2018) An Australian dog diagnosed with an exotic tick-borne infection: Should Australia still be considered free from Hepatozoon canis? International Journal for Parasitology http://researchrepository.murdoch.edu.au/id/eprint/41691/ Copyright: © 2018 Australian Society for Parasitology It is posted here for your personal use. No further distribution is permitted.
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of should Australia still be considered free from Hepatozoon canis

RESEARCH REPOSITORY
This is the author’s final version of the work, as accepted for publication following peer review but without the publisher’s layout or pagination.
The definitive version is available at:
https://doi.org/10.1016/j.ijpara.2018.05.002
Greay, T.L., Barbosa, A.D., Rees, R.L., Paparini, A., Ryan, U.M., Oskam, C.L. and Irwin, P.J. (2018) An Australian dog diagnosed with an exotic tick-borne infection: Should Australia
still be considered free from Hepatozoon canis? International Journal for Parasitology
http://researchrepository.murdoch.edu.au/id/eprint/41691/
Copyright: © 2018 Australian Society for Parasitology
It is posted here for your personal use. No further distribution is permitted.

Accepted Manuscript
Current Opinion
An Australian dog diagnosed with an exotic tick-borne infection: should Aus-tralia still be considered free from Hepatozoon canis?
Telleasha L. Greay, Amanda D. Barbosa, Robert L. Rees, Andrea Paparini, UnaM. Ryan, Charlotte L. Oskam, Peter J. Irwin
PII: S0020-7519(18)30163-2DOI: https://doi.org/10.1016/j.ijpara.2018.05.002Reference: PARA 4079
To appear in: International Journal for Parasitology
Received Date: 15 February 2018Revised Date: 22 May 2018Accepted Date: 28 May 2018
Please cite this article as: Greay, T.L., Barbosa, A.D., Rees, R.L., Paparini, A., Ryan, U.M., Oskam, C.L., Irwin,P.J., An Australian dog diagnosed with an exotic tick-borne infection: should Australia still be considered free fromHepatozoon canis?, International Journal for Parasitology (2018), doi: https://doi.org/10.1016/j.ijpara.2018.05.002
This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customerswe are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, andreview of the resulting proof before it is published in its final form. Please note that during the production processerrors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.

1
Current Opinion
An Australian dog diagnosed with an exotic tick-borne infection: should Australia still be
considered free from Hepatozoon canis?
Telleasha L Greaya,b, Amanda D Barbosaa,c, Robert L Reesd, Andrea Paparinia, Una M Ryana, Charlotte L
Oskama, Peter J Irwina,*
aVector and Waterborne Pathogens Research Group, School of Veterinary and Life Sciences, Murdoch
University, Perth, Western Australia, Australia
bWestern Australian State Agricultural Biotechnology Centre, Murdoch University, Perth, Western
Australia, Australia
cCAPES Foundation, Ministry of Education of Brazil, Brasília – DF 70040-020, Brazil
dFaculty of Veterinary and Agricultural Sciences, The University of Melbourne, Melbourne, Victoria,
Australia
*Corresponding author.
School of Veterinary and Life Sciences, College of Veterinary Medicine, 90 South Street, Murdoch, Western
Australia, 6150, Australia.
E-mail address: [email protected]

2
Abstract
Recent molecular and sero-surveillance studies of the tick-borne pathogen Hepatozoon canis have identified
new hosts, potential vector species, and have revealed that H. canis is more widespread than previously
thought. We report the first diagnosed case of canine hepatozoonosis in Australia from a Maremma
Sheepdog in Sarina, Queensland. Hepatozoon canis was detected with blood smear examination and 18S
rRNA sequencing. It is unknown when or how the organism was introduced into Australia, which raises
questions about border biosecurity policies and the H. canis infection status of its potential vectors and hosts
in Australia. Surveillance for this pathogen is required to determine whether H. canis has established in
Australia.
Keywords: Hepatozoon canis, Australia, dogs, Ixodes holocyclus, Rhipicephalus sanguineus, 18S rRNA

3
1. Introduction
Hepatozoon canis (Adeleorina: Hepatozoidae) is an apicomplexan blood parasite that causes
hepatozoonosis and was first described in dogs from India in 1905 (James, 1905). Hepatozoon canis
infections have been most frequently reported in domestic dogs (Canis lupus familiaris) and are usually
transmitted by ingestion of infected vector ticks (Baneth et al., 2001). Although autochthonous H. canis
infections have been reported in almost every continent, this has not been the case in Australia until this
report. Herein, we provide a brief review of H. canis, describe the diagnosis of H. canis infection in the
Australian dog by blood smear examination and 18S rRNA (18S) sequence analysis, and discuss the
potential sources of acquisition and directions for future studies to determine whether H. canis has
established in Australia.
1.1. Canine hepatozoonosis is a well characterised disease with few treatment options that have uncertain
efficacy
Subclinical or mild hepatozoonosis is most commonly observed in H. canis-infected dogs and is
usually associated with low parasitaemias; <800 gametocytes/μL of blood or <1-5% of infected neutrophils
(Baneth and Weigler, 1997; Baneth, 2011). If untreated, latent H. canis infections can persist for long
periods of time without causing severe illness (Baneth and Weigler, 1997). However, severe and life-
threatening illnesses manifest when high parasitaemia develops (>800 gametocytes/μL of blood; almost
100% of peripheral blood neutrophils may be infected) (Baneth, 2011). Clinical signs associated with high
parasitaemia include anaemia, cachexia, emaciation, fever and lethargy (Baneth et al., 1995; Baneth and
Weigler, 1997; Karagenc et al., 2006; Mundim et al., 2008). Hepatozoon canis meronts can infect bone
marrow, kidneys, liver, lungs, lymph nodes, periosteum and the spleen, while gamonts infect leukocytes
(Baneth et al., 1995; Baneth and Weigler, 1997; Marchetti et al., 2009). Clinical biochemical abnormalities
include hyperproteinaemia, hypoalbuminaemia, polyclonal gammopathy, and increased serum creatine
kinase and alkaline phosphatase activities (Baneth, 2011). Concomitant infections may influence the
pathogenesis of H. canis and the development of high parasitaemia; co-infections that have been associated
with high H. canis parasitaemia include anaplasmosis (Andersson et al., 2013), babesiosis (McCully et al.,
1975; Cardoso et al., 2010), canine distemper (McCully et al., 1975), ehrlichiosis (Kruzeniski et al., 2013;
Baneth et al., 2015), filariasis (Ramos et al., 2014), heartworm disease (Craig et al., 1978), leishmaniasis

4
(Rioux et al., 1964), parvoviral enteritis (Baneth et al., 1997; Baneth and Weigler, 1997) and toxoplasmosis
(Harmelin et al., 1992). Similarly, immune system immaturity in pups, immune system disorders and
immunosuppression induced by chemotherapy or immunosuppressive drugs may also lead to the
development of severe canine hepatozoonosis in H. canis-positive dogs (Craig, 1990; Baneth et al., 1997).
Imidocarb dipropionate (5-6 mg/kg every 14 days) is used to treat canine hepatozoonosis (Baneth
and Weigler, 1997), but the therapeutic efficacy is inconsistent and will not necessarily result in a
parasitological cure (Ogunkoya et al., 1981; Sasanelli et al., 2010; De Tommasi et al., 2014). Similarly, the
efficacy of toltrazuril is inconsistent (Krampitz and Haberkorn, 1988; Macintire et al., 1997; Tort et al.,
2007) and results from studies on combination therapies with emodepside/toltrazuril (Procox® suspension
for dogs (Altreuther et al., 2011)) and clindamycin (De Tommasi et al., 2014), and imidocarb
dipropionate/toltrazuril (Pasa et al., 2011) showed no statistically significant difference in clinical efficacy
compared with monotherapies. However, Pasa et al. (2011) reported a better overall clinical response with
imidocarb dipropionate/toltrazuril compared with imidocarb dipropionate treatment alone (Pasa et al., 2011).
1.2. Vertical and vectorial transmission of H. canis has been established; could predation also be a
transmission route?
Sexual development and sporogony occur in the definitive hosts of H. canis, hard ticks (Acari:
Ixodidae) (Baneth et al., 2007). Ticks infected with mature H. canis oocysts are ingested by intermediate
hosts (e.g. domestic dogs) (Baneth et al., 2001) and the sporozoites are released into the gastrointestinal tract.
After penetrating the gut wall, the sporozoites infect mononuclear cells and travel via blood and lymph to
haemolymphatic tissues, and other tissues such as the liver, kidneys and lungs, where meront stages (micro-
and macromerozoites) develop. Mature meronts can then infect monocytes and neutrophils to form gamonts,
which are ingested by the definitive host during blood feeding. The gamonts are released into the tick gut
where the male and female gamonts undergo gametogenesis to form gametes. Following fertilisation and
sporogony, oocysts form in the haemocoel that contains infective sporozoites, which are ingested by an
intermediate host (Baneth et al., 2001, 2007). The entire life cycle can be completed in 81 days (Baneth et
al., 2007). There is also evidence of intrauterine transmission of H. canis in domestic dogs (Murata et al.,
1993) and red foxes (Vulpes vulpes) (Hodzic et al., 2018). Transstadial transmission of H. canis by the
brown dog tick, Rhipicephalus sanguineus sensu lato (s.l.), has also been reported recently in Turkey, from

5
larva to nymph and from nymph to adult, but there was no evidence of transovarial transmission (Aktas and
Ozubek, 2017). Although predation is a transmission route for a close relative of H. canis, Hepatozoon
americanum (Johnson et al., 2009; Allen et al., 2011), this has not yet been demonstrated for H. canis, but
remains a possibility considering that monozoic cysts form in tissues (Baneth and Shkap, 2003; Baneth,
2011).
1.3. Hepatozoon canis has been detected in a variety of mammals, but are they intermediate or incidental
hosts?
Hepatozoon canis has been most widely reported in domestic dogs and, increasingly, in red foxes
(Table 1). Compared with dogs, a higher prevalence (ranging from 12-95%) of H. canis has been observed in
red fox populations in Austria (Duscher et al., 2014), Bosnia and Herzegovina (Hodzic et al., 2015), the
Czech Republic (Mitkova et al., 2016), Germany (Najm et al., 2014), Poland (Karbowiak et al., 2010) and
Slovakia (Miterpakova et al., 2017). It has been proposed that the predatory feeding behaviour of foxes may
explain the higher prevalence of H. canis in foxes than in dogs, as there would be a greater potential for the
ingestion of infected ticks (Gabrielli et al., 2010; Miterpakova et al., 2017) and, if predation is a viable H.
canis transmission route, infected prey. Hepatozoon canis has also been isolated from a variety of other
mammals including carnivores, marsupials (in South America), rodents and ungulate species (Table 1). This
indicates that either H. canis lacks host specificity, or that other Hepatozoon spp. have been misidentified as
H. canis due to a lack of interspecific differences in morphology. Whether these hosts play a role in the
transmission of H. canis, or are incidental hosts, remains to be elucidated.
1.4. A better understanding of H. canis vector competence in putative vectors and the R. sanguineus species
complex is required
Rhipicephalus sanguineus s.l. is the most widely studied vector of H. canis (Baneth et al., 2007) and
is distributed throughout the world in tropical, subtropical and temperate regions (Dantas-Torres, 2010).
Recent studies have found that R. sanguineus s.l. consists of a species complex (R. sanguineus group ticks)
(Dantas-Torres et al., 2013), which may explain the differences in R. sanguineus s.l. vector competency for a
number of tick-borne pathogens (Dantas-Torres and Otranto, 2015). For example, H. canis has been detected
in Rhipicephalus sp. I from Israel, Rhipicephalus sp. II (temperate or southern lineage) from Italy, Portugal

6
and Spain, and Rhipicephalus sp. III from India (Latrofa et al., 2014). However, H. canis has not been
detected in the southeastern lineage of R. sanguineus s.l. in Romania (Andersson et al., 2017; Chitimia-
Dobler et al., 2017), and the tropical (or northern) lineage of R. sanguineus s.l. does not appear to be a major
H. canis vector in Brazil (Forlano et al., 2005; Gomes et al., 2010; Demoner et al., 2013). Hepatozoon canis
is also transmitted by Amblyomma ovale in Brazil (Forlano et al., 2005; Rubini et al., 2009; Demoner et al.,
2013), and oocysts have been observed in the haemocoel of putative vectors Haemaphysalis longicornis and
Haemaphysalis flava from Japan (Murata et al., 1995), and Rhipicephalus microplus from Brazil (de
Miranda et al., 2011). Recently, successful H. canis sporogonic development and infection of canine
leukocytes was demonstrated in Rhipicephalus turanicus, which occurs in Africa, and in Mediterranean and
dry regions of Asia and Europe (Giannelli et al., 2017b).
Hepatozoon canis has been detected in several other tick species, including Amblyomma sculptum
(Demoner et al., 2013; Melo et al., 2016), Dermacentor marginatus, (Hornok et al., 2013; Aktas, 2014;
Karasartova et al., 2018), Dermacentor reticulatus (Hornok et al., 2013; Najm et al., 2014), Haemaphysalis
concinna (Hornok et al., 2013; Andersson et al., 2017), Haemaphysalis parva (Orkun and Nalbantoğlu,
2018), Haemaphysalis punctata (Andersson et al., 2017), Haemaphysalis sulcata (Aktas, 2014), Ixodes
canisuga, Ixodes hexagonus (Najm et al., 2014) and Ixodes ricinus (Gabrielli et al., 2010; Reye et al., 2010;
Aktas, 2014; Najm et al., 2014; Hamsikova et al., 2016; Andersson et al., 2017). The vector competence of
these species for H. canis is unknown, with many collected from H. canis-infected animals. Hornok et al.
(2013) found that the genotype of H. canis identified in D. marginatus and D. reticulatus ticks was identical
to the genotype detected in the host dog’s blood, suggesting that the presence of H. canis in the ticks may be
due to the H. canis-infected blood meal (Hornok et al., 2013). However, the H. canis genotype detected in H.
concinna ticks differed from the host-derived H. canis genotype and was also detected in a H. concinna tick
removed from a H. canis-negative dog (Hornok et al., 2013). A recent study showed that although H. canis
has been detected in A. sculptum (reported as Amblyomma cajennense), it was not a competent vector of H.
canis (Demoner et al., 2013; Melo et al., 2016). Although H. canis has been detected in questing I. ricinus
(Reye et al., 2010; Hamsikova et al., 2016), which indicates that transstadial transmission occurs, there are
no studies at present that have identified successful sporogony in I. ricinus (Gabrielli et al., 2010; Giannelli
et al., 2013).

7
1.5. The geographical distribution of H. canis has expanded: an emerging disease or an increase in tick-
borne pathogen surveillance?
Autochthonous H. canis infections have been reported from more than 60 countries (Fig. 1;
Supplementary Table S1) in Africa, Asia, Europe, the Middle East, and in North and South America. It has
only recently been discovered that H. canis is endemic to countries such as Germany, where in the last
decade the first autochthonous cases were reported in domestic dogs (Gaertner et al., 2008), a high
prevalence (45%) of the parasite was detected in red foxes, and was also detected in ticks (D. reticulatus, I.
canisuga, I. hexagonus and I. ricinus) (Najm et al., 2014). Similarly, H. canis was recently considered to be
endemic in Austria, with a prevalence of 30-58% in red foxes (Duscher et al., 2014; Hodzic et al., 2018), in
Slovakia, with a prevalence of 0.1% in questing I. ricinus, 1% in dogs and 17% in red foxes (Hamsikova et
al., 2016; Miterpakova et al., 2017), and the Czech Republic, with a prevalence ranging from 0.04% in I.
ricinus, to 50% in dogs and 95% in red foxes (Hamsikova et al., 2016; Mitkova et al., 2016). The recent
identification of H. canis in countries previously considered to be non-endemic could be due to the incursion
of infected canids and ticks to non-endemic regions (Majlathova et al., 2007), but could also be due to a lack
of prior tick-borne pathogen surveillance using sensitive molecular techniques. For example, the first
molecular survey for Hepatozoon spp. in Mexico in 2014 revealed a high prevalence (63%) of H. canis in
domestic dogs (Jarquín-Díaz et al., 2016). In Ethiopia, there has been only one infection reported in C.
adustus (side-striped jackal) (Maia et al., 2014a); similarly, one report in V. pallida (pale fox) in Senegal,
(Maia et al., 2014a), one positive questing I. ricinus female in Luxembourg (Reye et al., 2010), and one
report in a domestic dog with an unknown travel history in Ukraine (Hamel et al., 2013).
2. First report of H. canis infection in a dog in Australia
During a molecular survey of ticks (n = 711) for apicomplexan parasites in 2017, H. canis DNA was
detected in an engorged Ixodes holocyclus (paralysis tick) female that was removed from a 3-month-old
Maremma Sheepdog from Sarina, Queensland (Qld, Australia)) in 2014 (Greay et al., 2018). The dog, born
in the city of Rockhampton, Qld, 200 km south west of Sarina, was located and the owner consented to a
blood test to evaluate the health and parasite status of the dog.

8
2.1. Clinical and laboratory findings: asymptomatic with mild haematological and biochemistry
abnormalities
Clinical examination and history indicated that the dog was asymptomatic and had no significant
medical history other than tick paralysis at the original time of the tick removal and collection. Blood was
collected by routine venepuncture and transported from Sarina, Qld to Perth, Western Australia (WA) for
analysis. The blood sample was moderately haemolysed on arrival; haematological abnormalities included a
mild eosinophilia (1.44 x 109/L; reference interval (RI) 0.10-1.25 x 109/L) and reduced mean corpuscular
haemoglobin concentration (315 g/L; RI 320-360 g/L), the latter finding attributed to the delay between
collection and analysis (7 days). Serum biochemistry revealed elevations in creatinine kinase (CK) (785 U/L;
RI 47-22 U/L) and aspartate transaminase (AST) (154 U/L; RI 10-60 U/L) activities, cholesterol (9.5
mmol/L; RI 3.30-6.90 mmol/L), potassium (5.7 mmol/L; RI 3.4-5.3 mmol/L), total protein (85 g/L; RI 56-80
g/L), albumin (49 g/L; RI 24-38 g/L) and phosphate (3.47 mmol/L; RI 0.80-2.20 mmol/L) concentrations,
and a decrease in serum calcium concentration (1.84 mmol/L; RI 2.20-2.80 mmol/L). Total bilirubin and
alkaline phosphatase activity could not be measured due to haemolysis, and the elevations in CK and AST
activities, hyperkalaemia, and an unmeasurable blood glucose concentration were also attributed to
prolonged storage prior to analysis.
2.2. Hepatozoon canis and heartworm morphologically identified by blood smear examination
Multiple blood smears were made from a drop of fresh peripheral blood, air-dried and fixed with
methanol prior to transport. The smears were then stained with Wright-Giemsa and coverslip mounted using
DPX (Merck Pty. Limited, Kilsyth, Victoria, Australia). Stained blood smears were examined under a light
microscope at 400-1,000x magnification. Red and white blood cell morphology was normal, however
ellipsoid-shaped intracellular inclusions approximately 10 μm in length, consistent with H. canis gamonts
(Baneth et al., 2007), were detected very occasionally in the cytoplasm of neutrophils (<1%) (Fig. 2).
Additionally, microfilariae, presumed to be Dirofilaria immitis, were observed.
2.3. Identity of H. canis confirmed with 18S sequence analysis
For molecular confirmation of H. canis in the blood sample, DNA was extracted using a DNeasy
Blood & Tissue Kit (Qiagen, Hilden, Germany) following the manufacturer’s recommendations. The 18S

9
gene was amplified using conventional PCR with apicomplexan 18SApiF/18SApiR primers (Greay et al.,
2018), Hepatozoon spp. HepF300/Hep900 primers (Ujvari et al., 2004) and Hepatozoon spp.
HEMO1/HEMO2 primers (Perkins and Keller, 2001) that target approximately 300 bp, 600 bp and 900 bp of
18S, respectively. The PCR, Sanger sequencing and sequence analysis methodologies were carried out as
described in Greay et al. (2018). The sequences obtained with the 18SApiF/18SApiR primer set (303 bp,
GenBank® accession no. MG076961) and the HepF300/Hep900 and HEMO1/HEMO2 primer sets
(concatenated to produce a 1,413 bp sequence, MG062866) were 100% identical to the H. canis sequences
from the I. holocyclus tick removed from the dog (MG062865 and MG758124). Aside from the I. holocyclus
sequences, the 1,413 bp sequence was most similar (99.9%), with two single nucleotide polymorphisms
(SNPs), to two different H. canis genotypes from the Czech Republic, isolated from a golden jackal
(KX712124), and from Spain, isolated from a domestic dog (AY150067). Therefore, the 1,413 bp H. canis
sequence obtained in the present study represents a novel H. canis genotype.
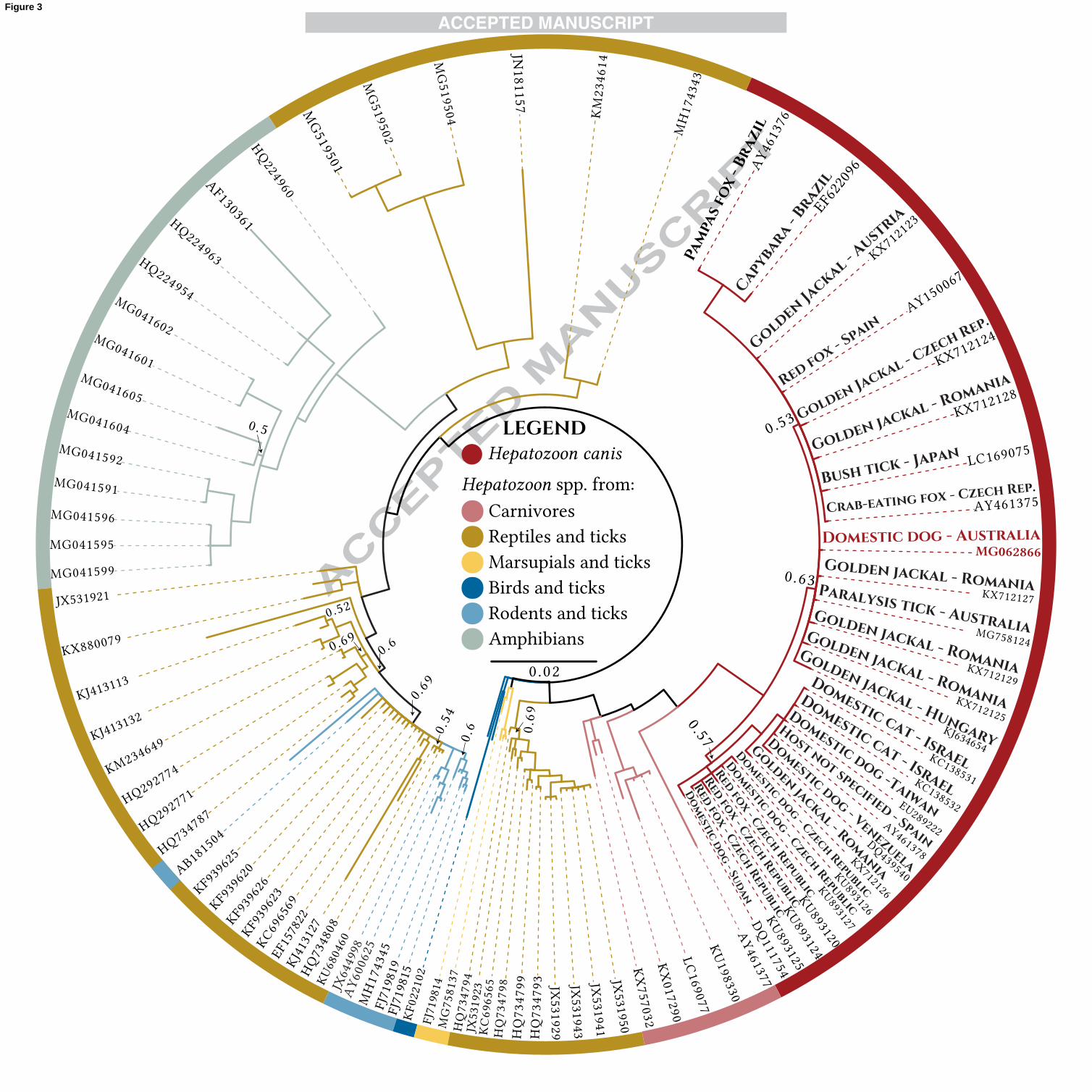
A Bayesian phylogenetic tree was constructed using the MrBayes plugin v3.2.6 (Huelsenbeck and
Ronquist, 2001) in Geneious v10.2.2 (http://www.geneious.com, Kearse et al. 2012) with long 18S
Hepatozoon spp. sequences (>1 kb) from GenBank. The phylogenetic tree shows that all H. canis isolates in
the alignment form a clade distinct from other Hepatozoon spp. (Fig. 3), and these H. canis sequences have
an average pairwise genetic distance (percent similarity) of 99.4% (S.D. = 0.3%), with intraspecific genetic
distances of ≤1.7%; the high genetic distances are caused by H. canis isolates from a capybara (EF622096)
and pampas fox (AY461376) from Brazil, and from a golden jackal from Hungary (KJ634654) (pairwise
percent identities calculated with a Kimura distance matrix (Kimura, 1980), Supplementary Table S2). The
H. canis sequence from the Australian dog groups with the H. canis clade with strong support (posterior
probability (PP) = 1.0) and has an average pairwise percent similarity of 99.6% (S.D. = 0.3%) to the other H.
canis isolates in the alignment. The three major H. canis subclades have very low support (PP 0.53-0.63) and
the isolates do not form distinct groups based on host species or geography (Fig. 3). The H. canis sequence
from the Australian dog groups with a clade of H. canis genotypes from golden jackals from Hungary
(KJ634654) and Romania (KX712125; KX712127; KX712129), but with low support (PP = 0.63). The
polytomy present in the tree suggests that the topology of phylogenetic reconstructions of H. canis genotypes
needs to be resolved with additional genetic characterisation.

10
3. Potential sources of H. canis acquisition in Australia
The identification of H. canis reported in the present study was unanticipated because Australia is
currently considered to be free of this canine pathogen. The route by which the dog was infected is unknown,
but there are two main possibilities: vertical and vector-borne transmission.
3.1. Vertical transmission from infected dam to pup?
Although there was no international travel history for this individual, which means that the infection
of the dog is autochthonous, the origin and international travel history of the dam are unknown. Thus, it is
possible that the dam may have been H. canis-positive and imported into Australia from a H. canis-endemic
country and vertically (transplacentally) transmitted the infection to the pup (Murata et al., 1993). It is also
possible that other prior female generations of this infected dog (e.g. the grandmother) were infected.
Unfortunately, the current location and H. canis infection status of the mother, grandmother, and other
individuals related to the infected dog are not known.
3.2. Potential tick-borne transmission
It is also possible that the dog could have been infected by ingesting an infected tick in Australia.
Both R. sanguineus s.l. and the putative H. canis vector H. longicornis occur in Australia (Barker and
Walker, 2014), and both species are distributed in the region where this dog was living. Although H. canis
DNA was not found in R. sanguineus s.l. (0/183; 95% confidence interval (CI), 0-2) or H. longicornis (0/75;
95% CI, 0-4.8) in the apicomplexan survey of ticks in Australia (Greay et al., 2018), the prevalence of H.
canis in ticks collected from dogs in endemic areas overseas has been reported to be as low as 1.5% (4/267)
(Dantas-Torres et al., 2012). Furthermore, only six R. sanguineus s.l. ticks from Sarina, Qld were tested
during the previous study by Greay et al. (2018). Regardless of the route of infection, we hypothesise that the
dog subsequently developed a parasitaemia and it seems likely that H. canis was present in the I. holocyclus
tick as a result of the ingestion of H. canis-infected blood from the dog.
4. Is H. canis endemic to Australia?
Further surveillance for H. canis in Australia is required to determine if the pathogen is more
widespread than this single case report. If H. canis is non-endemic and this case represents an isolated

11
incursion, this emphasises the need for more stringent animal biosecurity policies for the importation of
domestic dogs (and cats), which currently do not require H. canis testing as a veterinary procedure prior to
importation (http://www.agriculture.gov.au/cats-dogs/). Given this, it would not be surprising if H. canis has
established in Australia, especially with the popularity of international travel by people and their pets, and
the fact that two of its tick vectors, discussed previously, are well-established in Australia. It should be noted
that veterinary authorities have been notified about this case of H. canis infection.
4.1. Potential hosts include domestic and feral dogs and cats, dingoes, dingo-wild dog hybrids and foxes
How did this parasite come to Australia, a continent with unique fauna and flora, and isolated
geographically from the rest of the world for millennia, and why has it not been discovered sooner?
Domestic dogs were imported into Australia only in the last 230 years, during European colonisation, and it
is possible that H. canis-infected dogs arrived during this time. It is also possible that H. canis has been
present for much longer, brought onto the continent with feral or wild dogs, ancestors of today’s Australian
dingo (Canis dingo), which are estimated to have arrived in Australia approximately 4,000-18,000 years ago
with early traders from southeastern Asia (Savolainen et al., 2004; Oskarsson et al., 2012). A third possibility
is that H. canis entered Australia in European red foxes, which were introduced more recently (~160 years
ago) for recreational hunting and have become distributed throughout most of Australia, except in Tasmania
and far northern tropical regions of the Northern Territory, Qld and WA (Clarke et al., 2000). Given the high
prevalence of H. canis in foxes and wild canids in other countries, particularly in countries that were
originally thought to be H. canis-free, surveillance for H. canis in the population of wild canid species in
Australia may be fruitful in determining whether the parasite is, in fact, endemic and simply not recognised
to date. It would also be reasonable to examine domestic and feral cats (Felis catus) for H. canis (Criado-
Fornelio et al., 2009b; Díaz-Regañón et al., 2017; Giannelli et al., 2017a). It is curious that the parasite has
not been observed in Australia before, if indeed it is endemic, since thousands of canine blood samples are
examined every day in veterinary practices and clinical pathology laboratories. Over-reliance on automated
haematological analysers, without routine examination of blood smears, coupled with the typically low
parasitaemia, may explain why H. canis has not been reported in Australia before now.

12
4.2. The genotypes and thus vector competence of the ‘main’ H. canis vector, R. sanguineus s.l., are
unconfirmed in Australia
Importantly, also, the genotypes of R. sanguineus s.l. in Australia require further investigation for
the presence of H. canis and for its molecular species identification; only 50 R. sanguineus s.l. from New
South Wales have been genetically identified before and were found to belong to the tropical (northern)
lineage (Dantas-Torres et al., 2013). Although the brown dog tick is distributed mostly in tropical and
subtropical regions in Australia (Seddon, 1951; Roberts, 1965) R. sanguineus s.l. has also been recorded
south of this historical distribution range, in dry and Mediterranean climatic zones (Greay et al., 2016).
Therefore, other R. sanguineus group ticks that are adapted to these climates could also be present in
Australia. A summary of the potential transmission routes, hosts and vectors are presented in Fig. 4.
4.3. Surveillance and diagnostic strategies
Hepatozoon canis infections can be diagnosed with blood smear examination for intracytoplasmic
gamonts, histopathological examination for meronts and monozoic cysts in tissues (Baneth and Shkap,
2003), serological testing for anti-H. canis antibodies using an IFAT (Shkap et al., 1994) or an ELISA
(Gonen et al., 2004), and PCR followed by Sanger sequencing of the 18S gene. PCR is a much more
sensitive diagnostic method than blood smear examination alone, with studies reporting a ~3-5 fold increase
in positive cases detected with PCR compared with blood smear examination (Karagenc et al., 2006; Otranto
et al., 2011), and PCR is more sensitive than histopathology (Cardoso et al., 2014). At present, the only
available genetic material for comparisons of this parasite are 18S sequences, and there are a variety of 18S
primers that can be used for H. canis detection (Modry et al., 2017). However, there is no current consensus
on the most sensitive PCR method. PCR assays that target shorter fragments of the 18S gene are more
sensitive than assays that target longer fragments. Primers that can be used with conventional PCR that target
the shortest regions of 18S and are specific for the genus Hepatozoon are the HepF/HepR primers (Inokuma
et al., 2002) and the Hep-1/Hep-4 primers ((Criado-Fornelio et al., 2006) corrected Hep-1 primer sequence
reported in (Modry et al., 2017)), which target 670 bp and 660 bp, respectively. However, the detection
limits and sensitivity of these assays require assessment. Theoretically, the real-time fluorescence resonance
energy transfer (FRET) PCR and nested PCR assays should be more sensitive than the HepF/HepR and Hep-
1/Hep-4 assays. A FRET PCR assay developed for the detection of H. canis and Babesia canis vogeli that

13
targets 278 bp of the 18S gene has reported a high sensitivity in one study (100%) and a detection limit of
150 copies of positive control H. canis plasmid. The specificity of the primers for H. canis genotypes has
only been assessed thus far by bioinformatic analysis, and the primers may hybridise with H. americanum
(Kongklieng et al., 2015). There is also a more sensitive FRET PCR (limit of detection seven copies per mL)
that targets 198 bp of the 18S gene of H. canis and H. americanum (Li et al., 2008), but the specificity of
these FRET PCR assays for the different H. canis genotypes should also be experimentally tested. A less
specific FRET PCR assay, that is haemogregarine-specific, could be used if the previously mentioned assays
do not detect all H. canis genotypes, as it can differentiate between mixed infections of Hepatozoon spp.,
Schellackia spp. and Lankesterella spp. by melt curve analysis, and has a greater sensitivity (a detection limit
of five copies of Hepatozoon plasmid DNA) (Maia et al., 2014b). Alternatively, a nested PCR with
Hepatozoon-specific internal primers that amplify 620 bp of the 18S gene
(H14Hepa18SFw/H14Hepa18SRv) described by Hodzic et al. (2015) could be used, but the sensitivity and
detection limit of the assay are unknown and have not been compared with the FRET PCR assays. A
comparitive study on the detection limits, specificity and sensitivity of these PCR assays for the different H.
canis genotypes is required to determine which PCR assay is most sensitive and accurate for H. canis
detection.
While PCR can detect false negatives from serological tests that are due to acute infections, PCR
may fail to identify chronically infected individuals. The IFAT or ELISA assays have similar sensitivity and
specificity, and are useful for the diagnosis of chronic H. canis infections (except in rare cases when
antibodies levels are below the detection limit) (Shkap et al., 1994; Gonen et al., 2004; Karagenc et al.,
2006). Both tests are also more sensitive than microscopy, but the ELISA has several technical advantages
over the IFAT (easier to perform, higher throughput and more rapid results (Gonen et al., 2004)). We
recommend that the most thorough screening methodology for H. canis should be used for surveillance
purposes; this would employ an ELISA or sensitive PCR assay (e.g. the haemogregarine FRET PCR assay
by Maia et al. (2014b)) for initial screening, with negative samples subsequently screened with the
alternative test to detect false negatives from acute or chronic infections. A longer 18S gene fragment should
then be amplified and sequenced for all positive samples (e.g. 1,760 bp with the Hepatozoon spp. HAM-
1/HPF-2 primers (Criado-Fornelio et al., 2006)) for genotyping and phylogenetic analyses, as longer 18S
sequences provide better species resolution.

14
To maximize the chance of finding positive cases if H. canis infection is locally established, risk-
based sampling strategies should be used that target the region where the infected dog was born
(Rockhampton, Qld). Dogs that were in contact with the index case, dogs in local shelters and pounds, wild
Canidae (red foxes, dingoes, wild dogs, dingo-wild dog hybrids) and ticks (R. sanguineus s.l. and H.
longicornis) should be targeted, particularly in regions where these potential host and vector species overlap
(Fig. 5). Since splenic tissues have a higher sensitivity for H. canis detection compared with blood samples
(Duscher et al., 2014; Hodzic et al., 2018), opportunistic testing of spleens in foxes that have been culled
ethically for pest control may provide a more accurate estimation of the prevalence of H. canis in Australian
fox populations.
5. Conclusions
Molecular surveillance strategies for tick-borne pathogens have greatly improved our understanding
of their epidemiology and have recently changed our perspectives on the global occurrence of H. canis. If
this parasite is endemic to Australia, appropriate diagnosis and treatment by veterinarians is of critical
importance to the health and welfare of Australian dogs and cats. Alternatively, if H. canis is not established,
more stringent animal importation biosecurity policies are required to prevent future incursions of the
pathogen into Australia, which would reduce the risk of the pathogen becoming endemic.
Acknowledgements
We thank Murdoch University (Australia), Bayer Animal Health (Germany; 130100050), Bayer
Australia (130100050) and the Australian Research Council (130100050) for financially supporting our
research. We gratefully acknowledge the assistance of staff at the Sarina Veterinary Clinic (Australia) for
their contribution to our original tick survey, and for sampling the dog during the current investigation. We
would also like to thank Ms. Frances Brigg at the Western Australian State Agricultural Biotechnology
Centre (Australia) for her assistance with Sanger sequencing.

15
References
Aktas, M., 2014. A survey of ixodid tick species and molecular identification of tick-borne pathogens. Vet.
Parasitol. 200, 276-283.
Aktas, M., Ozubek, S., 2017. Transstadial transmission of Hepatozoon canis by Rhipicephalus sanguineus
(Acari: Ixodidae) in field conditions. J. Med. Entomol. 54, 1044-1048.
Alencar, N.X., Kohayagawa, A., Santarem, V.A., 1997. Hepatozoon canis infection of wild carnivores in
Brazil. Vet. Parasitol. 70, 279-282.
Allen, K.E., Johnson, E.M., Little, S.E., 2011. Hepatozoon spp infections in the United States. Vet. Clin. N.
Am.: Small Anim. Pract. 41, 1221-1238.
Altreuther, G., Gasda, N., Adler, K., Hellmann, K., Thurieau, H., Schimmel, A., Hutchens, D., Krieger, K.J.,
2011. Field evaluations of the efficacy and safety of Emodepside plus toltrazuril (Procox® oral
suspension for dogs) against naturally acquired nematode and Isospora spp. infections in dogs.
Parasitol. Res. 109 Suppl 1, S21-28.
Andersson, M., Turcitu, M.A., Stefanache, M., Tamba, P., Barbuceanu, F., Chitimia, L., 2013. First evidence
of Anaplasma platys and Hepatozoon canis co-infection in a dog from Romania - a case report.
Ticks Tick Borne Dis. 4, 317-319.
Andersson, M.O., Tolf, C., Tamba, P., Stefanache, M., Radbea, G., Rubel, F., Waldenström, J., Dobler, G.,
Chițimia-Dobler, L., 2017. Babesia, Theileria, and Hepatozoon species in ticks infesting animal
hosts in Romania. Parasitol. Res. 116, 2291-2297.
Andre, M.R., Adania, C.H., Teixeira, R.H., Vargas, G.H., Falcade, M., Sousa, L., Salles, A.R., Allegretti,
S.M., Felippe, P.A., Machado, R.Z., 2010. Molecular detection of Hepatozoon spp. in Brazilian and
exotic wild carnivores. Vet. Parasitol. 173, 134-138.
Baneth, G., 2011. Perspectives on canine and feline hepatozoonosis. Vet. Parasitol. 181, 3-11.

16
Baneth, G., Aroch, I., Presentey, B., 1997. Hepatozoon canis infection in a litter of Dalmatian dogs. Vet.
Parasitol. 70, 201-206.
Baneth, G., Harmelin, A., Presentey, B.Z., 1995. Hepatozoon canis infection in two dogs. J. Am. Vet. Med.
Assoc. 206, 1891-1894.
Baneth, G., Harrus, S., Gal, A., Aroch, I., 2015. Canine vector-borne co-infections: Ehrlichia canis and
Hepatozoon canis in the same host monocytes. Vet. Parasitol. 208, 30-34.
Baneth, G., Samish, M., Alekseev, E., Aroch, I., Shkap, V., 2001. Transmission of Hepatozoon canis to dogs
by naturally-fed or percutaneously-injected Rhipicephalus sanguineus ticks. J. Parasitol. 87, 606-
611.
Baneth, G., Samish, M., Shkap, V., 2007. Life cycle of Hepatozoon canis (Apicomplexa: Adeleorina:
Hepatozoidae) in the tick Rhipicephalus sanguineus and domestic dog (Canis familiaris). J.
Parasitol. 93, 283-299.
Baneth, G., Shkap, V., 2003. Monozoic cysts of Hepatozoon canis. J. Parasitol. 89, 379-381.
Baneth, G., Weigler, B., 1997. Retrospective case-control study of hepatozoonosis in dogs in Israel. J. Vet.
Intern. Med. 11, 365-370.
Barker, S.C., Walker, A.R., 2014. Ticks of Australia. The species that infest domestic animals and humans.
Zootaxa 3816.
Cardoso, L., Cortes, H.C., Eyal, O., Reis, A., Lopes, A.P., Vila-Vicosa, M.J., Rodrigues, P.A., Baneth, G.,
2014. Molecular and histopathological detection of Hepatozoon canis in red foxes (Vulpes vulpes)
from Portugal. Parasit. Vectors 7, 113.
Cardoso, L., Yisaschar-Mekuzas, Y., Rodrigues, F.T., Costa, A., Machado, J., Diz-Lopes, D., Baneth, G.,
2010. Canine babesiosis in northern Portugal and molecular characterization of vector-borne co-
infections. Parasit. Vectors 3, 27.

17
Chitimia-Dobler, L., Langguth, J., Pfeffer, M., Kattner, S., Kupper, T., Friese, D., Dobler, G., Guglielmone,
A.A., Nava, S., 2017. Genetic analysis of Rhipicephalus sanguineus sensu lato ticks parasites of
dogs in Africa north of the Sahara based on mitochondrial DNA sequences. Vet. Parasitol. 239, 1-6.
Clarke, G., Grosse, S., Matthews, M., Catling, P., Baker, B., Hewitt, C., Crowther, D., Sadlier, S., 2000.
Environmental pest species in Australia. Australia: state of the environment, second technical paper
series (biodiversity). Department of the Environment and Heritage, Canberra.
Clarke, L.L., Ballweber, L.R., Allen, K., Little, S.E., Lappin, M.R., 2014. Prevalence of select vector-borne
disease agents in owned dogs of Ghana. J. South African Vet. Assoc. 85, 996.
Conceicao-Silva, F.M., Abranches, P., Silva-Pereira, M.C., Janz, J.G., 1988. Hepatozoonosis in foxes from
Portugal. J. Wildl. Dis. 24, 344-347.
Craig, T., Smallwood, J., Knauer, K., McGrath, J., 1978. Hepatozoon canis infection in dogs: clinical,
radiographic, and hematologic findings. J. Am. Vet. Med. Assoc. 173, 967-972.
Craig, T.M., 1990. Hepatozoonosis, in: Greene, C.E. (Ed.), Clinical microbiology and infectious diseases of
the dog and cat, 2nd ed. WB Saunders, Philadelphia, PA, pp. 778-785.
Criado-Fornelio, A., Buling, A., Casado, N., Gimenez, C., Ruas, J., Wendt, L., da Rosa-Farias, N., Pinheiro,
M., Rey-Valeiron, C., Barba-Carretero, J.C., 2009a. Molecular characterization of arthropod-borne
hematozoans in wild mammals from Brazil, Venezuela and Spain. Acta Parasitol. 54, 187-193.
Criado-Fornelio, A., Buling, A., Pingret, J.L., Etievant, M., Boucraut-Baralon, C., Alongi, A., Agnone, A.,
Torina, A., 2009b. Hemoprotozoa of domestic animals in France: prevalence and molecular
characterization. Vet. Parasitol. 159, 73-76.
Criado-Fornelio, A., Ruas, J.L., Casado, N., Farias, N.A., Soares, M.P., Muller, G., Brumt, J.G., Berne,
M.E., Buling-Sarana, A., Barba-Carretero, J.C., 2006. New molecular data on mammalian
Hepatozoon species (Apicomplexa: Adeleorina) from Brazil and Spain. J. Parasitol. 92, 93-99.

18
da Silva, M.R.L., Fornazari, F., Demoner, L.d.C., Teixeira, C.R., Langoni, H., O’Dwyer, L.H., 2017.
Didelphis albiventris naturally infected with Hepatozoon canis in southeastern Brazil. Ticks Tick
Borne Dis. 8, 878-881.
Dantas-Torres, F., Latrofa, M.S., Annoscia, G., Giannelli, A., Parisi, A., Otranto, D., 2013. Morphological
and genetic diversity of Rhipicephalus sanguineus sensu lato from the New and Old Worlds. Parasit.
Vectors 6, 213.
Dantas-Torres, F., Latrofa, M.S., Weigl, S., Tarallo, V.D., Lia, R.P., Otranto, D., 2012. Hepatozoon canis
infection in ticks during spring and summer in Italy. Parasitol. Res. 110, 695-698.
Dantas-Torres, F., Otranto, D., 2015. Further thoughts on the taxonomy and vector role of Rhipicephalus
sanguineus group ticks. Vet. Parasitol. 208, 9-13.
de Azevedo Gomes, L., Moraes, L.A., Figueira Aguiar, D.C., Tavares Dias, H.L., Sardinha Ribeiro, A.S.,
Piram do Couto Rocha, H., Teixeira Nunes, M.R., Costa Gonçalves, E., 2018. Genetic diversity of
Hepatozoon spp. in Hydrochoerus hydrochaeris and Pecari tajacu from eastern Amazon. Ticks Tick
Borne Dis. 9, 314-318.
de Miranda, R.L., de Castro, J.R., Olegario, M.M., Beletti, M.E., Mundim, A.V., O'Dwyer, L.H., Eyal, O.,
Talmi-Frank, D., Cury, M.C., Baneth, G., 2011. Oocysts of Hepatozoon canis in Rhipicephalus
(Boophilus) microplus collected from a naturally infected dog. Vet. Parasitol. 177, 392-396.
De Tommasi, A.S., Giannelli, A., de Caprariis, D., Ramos, R.A., Di Paola, G., Crescenzo, G., Dantas-Torres,
F., Baneth, G., Otranto, D., 2014. Failure of imidocarb dipropionate and toltrazuril/emodepside plus
clindamycin in treating Hepatozoon canis infection. Vet. Parasitol. 200, 242-245.
Demoner, L.d.C., Rubini, A.S., dos Santos Paduan, K., Metzger, B., de Paula Antunes, J.M.A., Martins, T.F.,
Mathias, M.I.C., O’Dwyer, L.H., 2013. Investigation of tick vectors of Hepatozoon canis in Brazil.
Ticks Tick Borne Dis. 4, 542-546.

19
Dezdek, D., Vojta, L., Curkovic, S., Lipej, Z., Mihaljevic, Z., Cvetnic, Z., Beck, R., 2010. Molecular
detection of Theileria annae and Hepatozoon canis in foxes (Vulpes vulpes) in Croatia. Vet.
Parasitol. 172, 333-336.
Díaz-Regañón, D., Villaescusa, A., Ayllón, T., Rodríguez-Franco, F., Baneth, G., Calleja-Bueno, L., García-
Sancho, M., Agulla, B., Sainz, Á., 2017. Molecular detection of Hepatozoon spp. and Cytauxzoon sp.
in domestic and stray cats from Madrid, Spain. Parasit. Vectors 10, 112.
Dubey, J.P., Bwangamoi, O., 1994. Microbesnoitia leoni Bwangamoi, 1989, from the African lion (Panthera
leo) redetermined as a junior synonym of Hepatozoon canis (James, 1905) Wenyon, 1926. J.
Parasitol. 80, 333-334.
Duscher, G.G., Fuehrer, H.P., Kubber-Heiss, A., 2014. Fox on the run--molecular surveillance of fox blood
and tissue for the occurrence of tick-borne pathogens in Austria. Parasit. Vectors 7, 521.
Duscher, G.G., Kubber-Heiss, A., Richter, B., Suchentrunk, F., 2013. A golden jackal (Canis aureus) from
Austria bearing Hepatozoon canis -import due to immigration into a non-endemic area? Ticks Tick
Borne Dis. 4, 133-137.
Ebani, V.V., Rocchigiani, G., Nardoni, S., Bertelloni, F., Vasta, V., Papini, R.A., Verin, R., Poli, A.,
Mancianti, F., 2017. Molecular detection of tick-borne pathogens in wild red foxes (Vulpes vulpes)
from Central Italy. Acta Trop. 172, 197-200.
Farkas, R., Solymosi, N., Takacs, N., Hornyak, A., Hornok, S., Nachum-Biala, Y., Baneth, G., 2014. First
molecular evidence of Hepatozoon canis infection in red foxes and golden jackals from Hungary.
Parasit. Vectors 7, 303.
Forlano, M., Scofield, A., Elisei, C., Fernandes, K.R., Ewing, S.A., Massard, C.L., 2005. Diagnosis of
Hepatozoon spp. in Amblyomma ovale and its experimental transmission in domestic dogs in Brazil.
Vet. Parasitol. 134, 1-7.

20
Gabrielli, S., Kumlien, S., Calderini, P., Brozzi, A., Iori, A., Cancrini, G., 2010. The first report of
Hepatozoon canis identified in Vulpes vulpes and ticks from Italy. Vector Borne Zoonotic Dis. 10,
855-859.
Gaertner, S., Just, F.T., Pankraz, A., 2008. Hepatozoon canis infections in two dogs from Germany.
Kleintierpraxis 53, 81-87.
Giannelli, A., Latrofa, M.S., Nachum-Biala, Y., Hodzic, A., Greco, G., Attanasi, A., Annoscia, G., Otranto,
D., Baneth, G., 2017a. Three different Hepatozoon species in domestic cats from southern Italy.
Ticks Tick Borne Dis. 8, 721-724.
Giannelli, A., Lia, R.P., Annoscia, G., Buonavoglia, C., Lorusso, E., Dantas-Torres, F., Baneth, G., Otranto,
D., 2017b. Rhipicephalus turanicus, a new vector of Hepatozoon canis. Parasitology 144, 730-737.
Giannelli, A., Ramos, R.A., Dantas-Torres, F., Mencke, N., Baneth, G., Otranto, D., 2013. Experimental
evidence against transmission of Hepatozoon canis by Ixodes ricinus. Ticks Tick Borne Dis. 4, 391-
394.
Gomes, P.V., Mundim, M.J., Mundim, A.V., de Avila, D.F., Guimaraes, E.C., Cury, M.C., 2010. Occurrence
of Hepatozoon sp. in dogs in the urban area originating from a municipality in southeastern Brazil.
Vet. Parasitol. 174, 155-161.
Gonen, L., Strauss-Ayali, D., Shkap, V., Vincent-Johnson, N., Macintire, D.K., Baneth, G., 2004. An
enzyme-linked immunosorbent assay for antibodies to Hepatozoon canis. Vet. Parasitol. 122, 131-
139.
Greay, T.L., Oskam, C.L., Gofton, A.W., Rees, R.L., Ryan, U.M., Irwin, P.J., 2016. A survey of ticks (Acari:
Ixodidae) of companion animals in Australia. Parasit. Vectors 9, 1-10.
Greay, T.L., Zahedi, A., Krige, A.-S., Owens, J.M., Rees, R.L., Ryan, U.M., Oskam, C.L., Irwin, P.J., 2018.
Endemic, exotic and novel apicomplexan parasites detected during a national study of ticks from
companion animals in Australia. Parasit. Vectors 11, 197.

21
Guindon, S., Dufayard, J.F., Lefort, V., Anisimova, M., Hordijk, W., Gascuel, O., 2010. New algorithms and
methods to estimate maximum-likelihood phylogenies: assessing the performance of PhyML 3.0.
Syst. Biol. 59, 307-321.
Hamel, D., Silaghi, C., Zapadynska, S., Kudrin, A., Pfister, K., 2013. Vector-borne pathogens in ticks and
EDTA-blood samples collected from client-owned dogs, Kiev, Ukraine. Ticks Tick Borne Dis. 4,
152-155.
Hamsikova, Z., Silaghi, C., Rudolf, I., Venclikova, K., Mahrikova, L., Slovak, M., Mendel, J., Blazejova, H.,
Berthova, L., Kocianova, E., Hubalek, Z., Schnittger, L., Kazimirova, M., 2016. Molecular detection
and phylogenetic analysis of Hepatozoon spp. in questing Ixodes ricinus ticks and rodents from
Slovakia and Czech Republic. Parasitol. Res. 115, 3897-3904.
Harmelin, A., Dubey, J.P., Yakobson, B., Nyska, A., Orgad, U., 1992. Concurrent Hepatozoon canis and
Toxoplasma gondii infections in a dog. Vet. Parasitol. 43, 131-136.
Harris, D.J., Pereira, A., Halajian, A., Luus-Powell, W.J., Kunutu, K.D., 2017. Screening for Hepatozoon
parasites in gerbils and potential predators in South Africa. J. South African Vet. Assoc. 88, e1-e4.
Hodzic, A., Alic, A., Fuehrer, H.P., Harl, J., Wille-Piazzai, W., Duscher, G.G., 2015. A molecular survey of
vector-borne pathogens in red foxes (Vulpes vulpes) from Bosnia and Herzegovina. Parasit. Vectors
8, 88.
Hodzic, A., Mrowietz, N., Cezanne, R., Bruckschwaiger, P., Punz, S., Habler, V.E., Tomsik, V., Lazar, J.,
Duscher, G.G., Glawischnig, W., Fuehrer, H.P., 2018. Occurrence and diversity of arthropod-
transmitted pathogens in red foxes (Vulpes vulpes) in western Austria, and possible vertical
(transplacental) transmission of Hepatozoon canis. Parasitology 145, 335-344.
Hornok, S., Tanczos, B., Fernandez de Mera, I.G., de la Fuente, J., Hofmann-Lehmann, R., Farkas, R., 2013.
High prevalence of Hepatozoon-infection among shepherd dogs in a region considered to be free of
Rhipicephalus sanguineus. Vet. Parasitol. 196, 189-193.

22
Huelsenbeck, J.P., Ronquist, F., 2001. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics
17, 754-755.
Imre, M., Dudu, A., Ilie, M.S., Morariu, S., Imre, K., Darabus, G., 2015. Molecular survey of Hepatozoon
canis in red foxes (Vulpes vulpes) from Romania. J. Parasitol. 101, 490-491.
Inokuma, H., Okuda, M., Ohno, K., Shimoda, K., Onishi, T., 2002. Analysis of the 18S rRNA gene sequence
of a Hepatozoon detected in two Japanese dogs. Vet. Parasitol. 106, 265-271.
James, S.P., 1905. On a parasite found in the white corpuscles of the blood of dogs. Office of the
superintendant of government printing, India.
Jarquín-Díaz, V.H., Barbachano-Guerrero, A., Maldonado-Rodríguez, R., Vásquez-Aguilar, A.A., Aguilar-
Faisal, J.L., 2016. First molecular evidence of Hepatozoon canis in domestic dogs and ticks in
fragmented rainforest areas in Mexico. Vet. Parasitol. (Amst) 6, 4-8.
Johnson, E.M., Panciera, R.J., Allen, K.E., Sheets, M.E., Beal, J.D., Ewing, S.A., Little, S.E., 2009.
Alternate pathway of infection with Hepatozoon americanum and the epidemiologic importance of
predation. J. Vet. Intern. Med. 23, 1315-1318.
Karagenc, T.I., Pasa, S., Kirli, G., Hosgor, M., Bilgic, H.B., Ozon, Y.H., Atasoy, A., Eren, H., 2006. A
parasitological, molecular and serological survey of Hepatozoon canis infection in dogs around the
Aegean coast of Turkey. Vet. Parasitol. 135, 113-119.
Karasartova, D., Gureser, A.S., Gokce, T., Celebi, B., Yapar, D., Keskin, A., Celik, S., Ece, Y., Erenler,
A.K., Usluca, S., Mumcuoglu, K.Y., Taylan-Ozkan, A., 2018. Bacterial and protozoal pathogens
found in ticks collected from humans in Corum province of Turkey. PLoS Negl. Trop. Dis. 12,
e0006395.
Karbowiak, G., Majláthová, V., Hapunik, J., Pet'ko, B., Wita, I., 2010. Apicomplexan parasites of red foxes
(Vulpes vulpes) in northeastern Poland. Acta Parasitol. 55, 210-214.
Katoh, K., Misawa, K., Kuma, K.-i., Miyata, T., 2002. MAFFT: a novel method for rapid multiple sequence
alignment based on fast Fourier transform. Nucleic Acids Res. 30, 3059-3066.

23
Kearse, M., Moir, R., Wilson, A., Stones-Havas, S., Cheung, M., Sturrock, S., Buxton, S., Cooper, A.,
Markowitz, S., Duran, C., Thierer, T., Ashton, B., Meintjes, P., Drummond, A., 2012. Geneious
Basic: an integrated and extendable desktop software platform for the organization and analysis of
sequence data. Bioinformatics 28, 1647-1649.
Kelly, P., Marabini, L., Dutlow, K., Zhang, J., Loftis, A., Wang, C., 2014. Molecular detection of tick-borne
pathogens in captive wild felids, Zimbabwe. Parasit. Vectors 7, 514.
Kimura, M., 1980. A simple method for estimating evolutionary rates of base substitutions through
comparative studies of nucleotide sequences. J. Mol. Evol. 16, 111-120.
Kistler, W.M., Brown, J.D., Allison, A.B., Nemeth, N.M., Yabsley, M.J., 2014. First report of
Angiostrongylus vasorum and Hepatozoon from a red fox (Vulpes vulpes) from West Virginia, USA.
Vet. Parasitol. 200, 216-220.
Kongklieng, A., Intapan, P.M., Boonmars, T., Thanchomnang, T., Janwan, P., Sanpool, O., Lulitanond, V.,
Taweethavonsawat, P., Chungpivat, S., Maleewong, W., 2015. Detection of Babesia canis vogeli and
Hepatozoon canis in canine blood by a single-tube real-time fluorescence resonance energy transfer
polymerase chain reaction assay and melting curve analysis. J. Vet. Diag. Invest. 27, 191-195.
Krampitz, H.E., Haberkorn, A., 1988. Experimental treatment of Hepatozoon infections with the
anticoccidial agent Toltrazuril. J. Vet. Med. B Infect. Dis. Vet. Public Health 35, 131-137.
Kruzeniski, S.J., Tam, F.M., Burgess, H.J., 2013. Pathology in practice. Ehrlichia-Hepatozoon coinfection in
a dog. J. Am. Vet. Med. Assoc. 243, 1705-1707.
Laird, M., 1959. Malayan Protozoa 2. Hepatozoon Miller (Sporozoa : Coccidia), with an unusual host record
for H. canis (James). J. Protozool. 6, 316-319.
Latrofa, M.S., Dantas-Torres, F., Giannelli, A., Otranto, D., 2014. Molecular detection of tick-borne
pathogens in Rhipicephalus sanguineus group ticks. Ticks Tick Borne Dis. 5, 943-946.

24
Li, Y., Wang, C., Allen, K.E., Little, S.E., Ahluwalia, S.K., Gao, D., Macintire, D.K., Blagburn, B.L.,
Kaltenboeck, B., 2008. Diagnosis of canine Hepatozoon spp. infection by quantitative PCR. Vet.
Parasitol. 157, 50-58.
Macintire, D.K., Vincent-Johnson, N., Dillon, A.R., Blagburn, B., Lindsay, D., Whitley, E.M., Banfield, C.,
1997. Hepatozoonosis in dogs: 22 cases (1989-1994). J. Am. Vet. Med. Assoc. 210, 916-922.
Maia, J.P., Alvares, F., Boratynski, Z., Brito, J.C., Leite, J.V., Harris, D.J., 2014a. Molecular assessment of
Hepatozoon (Apicomplexa: Adeleorina) infections in wild canids and rodents from north Africa,
with implications for transmission dynamics across taxonomic groups. J. Wildl. Dis. 50, 837-848.
Maia, J.P., Harris, D.J., Carranza, S., Gómez-Díaz, E., 2014b. A comparison of multiple methods for
estimating parasitemia of hemogregarine hemoparasites (Apicomplexa: Adeleorina) and its
application for studying infection in natural populations. PLoS One 9, e95010.
Majlathova, V., Hurnikova, Z., Majlath, I., Petko, B., 2007. Hepatozoon canis infection in Slovakia:
imported or autochthonous? Vector Borne Zoonotic Dis. 7, 199-202.
Marchetti, V., Lubas, G., Baneth, G., Modenato, M., Mancianti, F., 2009. Hepatozoonosis in a dog with
skeletal involvement and meningoencephalomyelitis. Vet. Clin. Pathol. 38, 121-125.
Margalit Levi, M., Nachum-Biala, Y., King, R., Baneth, G., 2018. A survey of Babesia spp. and Hepatozoon
spp. in wild canids in Israel. Parasit. Vectors 11, 150.
McCully, R.M., Basson, P.A., Bigalke, R.D., De Vos, V., Young, E., 1975. Observations on naturally
acquired hepatozoonosis of wild carnivores and dogs in the Republic of South Africa. Onderstepoort
J. Vet. Res. 42, 117-133.
Melo, A.L., Witter, R., Martins, T.F., Pacheco, T.A., Alves, A.S., Chitarra, C.S., Dutra, V., Nakazato, L.,
Pacheco, R.C., Labruna, M.B., Aguiar, D.M., 2016. A survey of tick-borne pathogens in dogs and
their ticks in the Pantanal biome, Brazil. Med. Vet. Entomol. 30, 112-116.

25
Miterpakova, M., Komjati-Nagyova, M., Hurnikova, Z., Vichova, B., 2017. Retrospective molecular study
on canine hepatozoonosis in Slovakia - Does infection risk for dogs really exist? Ticks Tick Borne
Dis. 8, 567-573.
Mitková, B., Hrazdilová, K., D’Amico, G., Duscher, G.G., Suchentrunk, F., Forejtek, P., Gherman, C.M.,
Matei, I.A., Ionică, A.M., Daskalaki, A.A., Mihalca, A.D., Votýpka, J., Hulva, P., Modrý, D., 2017.
Eurasian golden jackal as host of canine vector-borne protists. Parasit. Vectors 10, 183.
Mitkova, B., Hrazdilova, K., Steinbauer, V., D'Amico, G., Mihalca, A.D., Modry, D., 2016. Autochthonous
Hepatozoon infection in hunting dogs and foxes from the Czech Republic. Parasitol. Res. 115, 4167-
4171.
Modry, D., Beck, R., Hrazdilová, K., Baneth, G., 2017. A review of methods for detection of Hepatozoon
infection in carnivores and arthropod vectors. Vector Borne Zoonotic Dis. 17, 66-72.
Mundim, A.V., Morais, I.A.d., Tavares, M., Cury, M.C., Mundim, M.J.S., 2008. Clinical and hematological
signs associated with dogs naturally infected by Hepatozoon sp. and with other hematozoa: A
retrospective study in Uberlândia, Minas Gerais, Brazil. Vet. Parasitol. 153, 3-8.
Murata, T., Inoue, M., Tateyama, S., Taura, Y., Nakama, S., 1993. Vertical transmission of Hepatozoon
canis in dogs. J. Vet. Med. Sci. 55, 867-868.
Murata, T., Inoue, M., Taura, Y., Nakama, S., Abe, H., Fujisaki, K., 1995. Detection of Hepatozoon canis
oocyst from ticks collected from the infected dogs. J. Vet. Med. Sci. 57, 111-112.
Najm, N.A., Meyer-Kayser, E., Hoffmann, L., Pfister, K., Silaghi, C., 2014. Hepatozoon canis in German
red foxes (Vulpes vulpes) and their ticks: molecular characterization and the phylogenetic
relationship to other Hepatozoon spp. Parasitol. Res. 113, 2679-2685.
Ogunkoya, A.B., Adeyanju, J.B., Aliu, Y.O., 1981. Experiences with the use of Imizol in treating canine
blood parasites in Nigeria. J. Small Anim. Prac. 22, 775-777.
Orkun, Ö., Nalbantoğlu, S., 2018. Hepatozoon canis in Turkish red foxes and their ticks. Vet. Parasitol.
(Amst) 13, 35-37.

26
Oskarsson, M.C., Klutsch, C.F., Boonyaprakob, U., Wilton, A., Tanabe, Y., Savolainen, P., 2012.
Mitochondrial DNA data indicate an introduction through Mainland Southeast Asia for Australian
dingoes and Polynesian domestic dogs. Proc R Soc B 279.
Otranto, D., Dantas-Torres, F., Weigl, S., Latrofa, M.S., Stanneck, D., Decaprariis, D., Capelli, G., Baneth,
G., 2011. Diagnosis of Hepatozoon canis in young dogs by cytology and PCR. Parasit. Vectors 4,
55.
Pasa, S., Voyvoda, H., Karagenc, T., Atasoy, A., Gazyagci, S., 2011. Failure of combination therapy with
imidocarb dipropionate and toltrazuril to clear Hepatozoon canis infection in dogs. Parasitol. Res.
109, 919-926.
Penzhorn, B.L., Netherlands, E.C., Cook, C.A., Smit, N.J., Vorster, I., Harrison-White, R.F., Oosthuizen,
M.C., 2018. Occurrence of Hepatozoon canis (Adeleorina: Hepatozoidae) and Anaplasma spp.
(Rickettsiales: Anaplasmataceae) in black-backed jackals (Canis mesomelas) in South Africa.
Parasit. Vectors 11, 158.
Perkins, S.L., Keller, A.K., 2001. Phylogeny of nuclear small subunit rRNA genes of hemogregarines
amplified with specific primers. J. Parasitol. 87, 870-876.
Ramos, R.A., Giannelli, A., Lia, R.P., Brianti, E., Tarallo, V.D., Breitshwerdt, E.B., Dantas-Torres, F.,
Stanneck, D., Otranto, D., 2014. Incidence of Cercopithifilaria bainae in dogs and probability of co-
infection with other tick-borne pathogens. PLoS One 9, e88198.
Reye, A.L., Hubschen, J.M., Sausy, A., Muller, C.P., 2010. Prevalence and seasonality of tick-borne
pathogens in questing Ixodes ricinus ticks from Luxembourg. Appl. Environ. Microbiol. 76, 2923-
2931.
Rioux, J.A., Golvan, Y.J., Houin, R., 1964. A case of mixed infestation with Hepatozoon canis (James 1905)
and Leishmania "canis" in a dog from Sète (Hérault). Ann. Parasitol. Hum. Comp. 39, 131-135.
Roberts, F., 1965. The taxonomic status of the species of the genera Rhipicephalus Koch and Boophilus
Curtice (Acarina: Ixodidae) occurring in Australia. Aust. J. Zool. 13, 491-524.

27
Rubini, A.S., Paduan, K.S., Martins, T.F., Labruna, M.B., O'Dwyer, L.H., 2009. Acquisition and
transmission of Hepatozoon canis (Apicomplexa: Hepatozoidae) by the tick Amblyomma ovale
(Acari: Ixodidae). Vet. Parasitol. 164, 324-327.
Sasanelli, M., Paradies, P., Greco, B., Eyal, O., Zaza, V., Baneth, G., 2010. Failure of imidocarb
dipropionate to eliminate Hepatozoon canis in naturally infected dogs based on parasitological and
molecular evaluation methods. Vet. Parasitol. 171, 194-199.
Savolainen, P., Leitner, T., Wilton, A.N., Matisoo-Smith, E., Lundeberg, J., 2004. A detailed picture of the
origin of the Australian dingo, obtained from the study of mitochondrial DNA. Proc. Natl. Acad. Sci.
U.S.A 101, 12387-12390.
Seddon, H., 1951. Diseases of domestic Animals in Australia. Part 3. Tick and Mite Infestations. Service
Publications. Department of Health, Australia. Veterinary Hygiene.
Shkap, V., Baneth, G., Pipano, E., 1994. Circulating antibodies to Hepatozoon canis demonstrated by
immunofluorescence. J. Vet. Diag. Invest. Inc 6, 121-123.
Starkey, L.A., Panciera, R.J., Paras, K., Allen, K.E., Reiskind, M.H., Reichard, M.V., Johnson, E.M., Little,
S.E., 2013. Genetic diversity of Hepatozoon spp. in coyotes from the south-central United States. J.
Parasitol. 99, 375-378.
Tolnai, Z., Sreter-Lancz, Z., Sreter, T., 2015. Spatial distribution of Anaplasma phagocytophilum and
Hepatozoon canis in red foxes (Vulpes vulpes) in Hungary. Ticks Tick Borne Dis. 6, 645-648.
Tort, G., Petetta, L., Favre, M., Mas, J., Robles, A., 2007. First description of an outbreak of hepatozoonoses.
Veterinaria Argentina 235, 399.
Ujvari, B., Madsen, T., Olsson, M., 2004. High prevalence of Hepatozoon spp. (Apicomplexa,
Hepatozoidae) infection in water pythons (Liasis fuscus) from tropical Australia. J. Parasitol. 90,
670-672.

28
Legend to Figures
Fig. 1. Hepatozoon canis country occurrence map. Countries that have reported autochthonous infections are
considered ‘endemic’ (red); some countries have reported H. canis infection(s), but are currently considered
‘non-endemic’ (yellow); and countries that have not reported H. canis infections are considered non-endemic
(grey). A list of countries and references is provided in Supplementary Table S1.
Fig. 2. Light micrograph of a Giemsa-stained blood film showing an ellipsoidal-shaped gamont of
Hepatozoon canis (arrow) within a neutrophil at 1,000x magnification.
Fig. 3. Bayesian phylogenetic tree of a 1,165 bp (without gaps) alignment of 18S Hepatozoon spp. sequences
aligned using the MAFFT program (Katoh et al., 2002). Values at nodes indicate Bayesian posterior
probabilities. The most suitable nucleotide substitution model, HKY85+G+I, was determined by the program
PhyML (Guindon et al., 2010) using the Bayesian information criterion (BIC). A 1,100,000 Markov chain
Monte Carlo (MCMC) length and a burn-in length of 10,000 with a subsampling frequency of 200 were
used. The outgroup, Hemolivia stellata (GenBank® accession number KP881349), and posterior
probabilities ≥0.7 are not shown.
Fig. 4. Potential hosts, vectors and transmission routes of Hepatozoon canis in Australia. Red fox
distribution adapted from Clarke et al. (2000). Rhipicephalus sanguineus sensu lato (s.l.) distribution is
adapted from Roberts (1965). K YA, thousand years ago.

29
Table 1. Vertebrate host records of Hepatozoon canis excluding domestic dogs (Canis lupus familiaris)
Scientific Name Common Name Country References
Artiodactyla
Ovis aries Sheep Mexico Jarquín-Díaz et al. (2016) Pecari tajacu Collared peccary Brazil de Azevedo Gomes et al. (2018)
Carnivora: Caniformia
Canis adustus Side-striped jackal
Ethiopia Maia et al. (2014a)
Canis aureus Golden jackal Algeria Austria Duscher et al. (2013); Mitková et
al. (2017) Czech Republic
Mitková et al. (2017)
Hungary Farkas et al. (2014) Israel Margalit Levi et al. (2018) Mauritania Maia et al. (2014a) Romania Mitková et al. (2017)
Canis latrans Coyote USA Starkey et al. (2013) Canis mesomelas Black-backed
jackal South Africa McCully et al. (1975); Penzhorn et
al. (2018) Cerdocyon thous Crab-eating fox Brazil Alencar et al. (1997); Criado-
Fornelio et al. (2006) Chrysocyon brachyurus Maned wolf Andre et al. (2010) Lycalopex gymnocercus Pampas fox Criado-Fornelio et al. (2006) Pseudalopex vetulus Hoary fox Andre et al. (2010) Vulpes chama Cape fox South Africa Harris et al. (2017)
Vulpes pallida Pale fox Mauritania Maia et al. (2014a) Niger
Senegal Vulpes rueppellii Rüppell’s fox Mauritania
Morocco Vulpes vulpes Red fox Austria Hodzic et al. (2018)
Bosnia and Herzegovina
Hodzic et al. (2015)
Croatia Dezdek et al. (2010) Germany Najm et al. (2014) Hungary Farkas et al. (2014); Tolnai et al.
(2015) Israel Margalit Levi et al. (2018) Italy Gabrielli et al. (2010); Ebani et al.
(2017) Japan Maia et al. (2014a) Morocco Poland Karbowiak et al. (2010) Portugal Conceicao-Silva et al. (1988);
Clarke et al. (2014) Romania Imre et al. (2015)
Slovakia Majlathova et al. (2007) Spain Criado-Fornelio et al. (2006) Tunisia Maia et al. (2014a) Turkey Orkun and Nalbantoğlu (2018) USA Kistler et al. (2014)
Vulpes zerda Fennec fox Mauritania Maia et al. (2014a) Morocco Western Sahara

30
Carnivora: Feliformia
Crocuta crocuta Hyena South Africa McCully et al. (1975) Felis catus Domestic cat France Criado-Fornelio et al. (2009b)
Italy Giannelli et al. (2017a)
Spain Díaz-Regañón et al. (2017)
Panthera leo Lion Kenya Dubey and Bwangamoi (1994) Zimbabwe Kelly et al. (2014)
Paradoxurus hermaphroditus Asian palm civet Malaysia Laird (1959)
Didelphimorphia
Didelphis albiventris White-eared opossum
Brazil da Silva et al. (2017)
Rodentia
Hydrochoerus hydrochaeris Capybara Brazil Criado-Fornelio et al. (2009a); de Azevedo Gomes et al. (2018)

31
'Highlights'
Hepatozoon canis was discovered in a dog for the first time in Australia
The vector competence of Ixodes holocyclus for H. canis is unknown
The H. canis sequence represents a novel genotype
Possible sources of acquisition are discussed

Photograph by Koja Niska, www.flickr.com
Should Australia still be considered free from Hepatozoon canis?*Graphical Abstract (for review)

cape verde
mauritius
Map designed by Free Vector Maps (www.freevectormaps.com)
Australia
CHINA
MALAYSIA
THAILAND
CAMBODIA
INDIA
IRAN
USA
MEXICO
PAKISTAN
IRAQ
NICARAGUA
COSTA RICA
COLOMBIA
VENEZUELA
BRAZIL
Argentina
SOUTHAFRICA
NAMIBIA
ANGOLA
ZIMBABWE
ZAMBIA
KENYA
ETHIOPIA
SUDAN
EGYPTALGERIA
WESTERN SAHARA
MOROCCO
GHANA
NIGERIA
NIGER
SPAINPORTUGAL
FRANCE
GERMANY
LUXEMBOURG
ITALY
POLAND
CZECH REP.
AUSTRIA
SLOVAKIA
HUNGARY
ROMANIA
BULGARIA
SERBIA
CROATIA
kosovo
bosnia and herzegovina
albania
greece turkey
ukraine
senegal
japansouthkorea
israelpalestine
cyprustunisia
malta
qatar
haiti
grenada
mauritania
singapore
Philippines
sri lanka
jordan
saint kitts and nevis
taiwan
Autochthonous H. canis infections reported (endemic)
H. canis infections reported, but occurrence not endemic
H. canis infections not reported (non-endemic)
LEGEND
Figure 1

Figure 2Click here to download high resolution image
0.02
KU893126
KJ413132
MG041596
KM234649
KX712125
MG041604
AF130361
AY4
6137
6
JX531921
HQ292771
AB181504
MG
519501
LC169077M
H17
4345
JX64
4998
KX880079
KM
2346
14
JN181157
MG041601
KF939
620
EF62
2096
KU
893125
MG041595
KX
757032
AY461375
KU893127
HQ224963
KC138531
AY150067
FJ71
9815
FJ71
9819
MG041591
KJ413113
MG758124
KJ4
1312
7 DQ
111754
KC69
6569
JX53
1923
HQ292774
HQ224954
KX712123
FJ71
9814AY
6006
25
HQ734787
KC138532
KF0
2210
2
KC
6965
65
KX712124
KF9396
25
HQ
7348
08
EF15
7822
MG
519504
AY461378
MG041602
KF939
623
MG041605
KX712126
MG041592
MG
519502
KU
893124
LC169075
EU289222
MG
7581
37
HQ
224960
HQ
7347
94
MG062866
KJ634654
AY461377
DQ439540
KX712128
KU893120
KX712127
KF939
626
MG041599
KX712129M
H17
4343
0 .69
0.53
0.6 0.
69
0 .60.69
0.5
0.57
0.52
0.63
0.54
Hepatozoon canisLEGEND
Reptiles and ticks Marsupials and ticks
Carnivores
Birds and ticksRodents and ticksAmphibians
Hepatozoon spp. from:
Do
mestic d
og - Sud
an
Red fox - C
zech Republic
Red fox - Czech Republic
Red fox - Czech Republic
Domestic dog - Czech Republic
Golden jackal - Romania
Domestic dog - Venezuela
Host not specified - Spain
Domestic dog -Taiwan
Domestic cat - Israel
Domestic cat - Israel
Golden jackal - Hungary
Golden jackal - Romania
Golden jackal - Romania
Paralysis tick - Australia
Golden jackal - Romania
Domestic dog - Australia
Crab-eating fox - Czech Rep.Bush tick - JapanGolden jackal - Romania
Golden Jackal - Czech Rep.
Red fox - Spain
Golden Jackal - A
ustria
Capyba
ra - Brazi
l
Pam
pas
fox
- Bra
zil
Domestic dog - Czech Republic
KU
6804
60
HQ
7347
98H
Q73
4799
HQ
7347
93 JX531929
JX531941
JX531943
JX531950
KX
017290
KU
198330
Figure 3

~160 YA
Red foxes(Vulpes vulpes)
~4K - 18K YA
Brown dog ticks(Rhipicephalus sanguineus s.l.)
230-18K YA?~230 YA
Dingoes(Canis dingo)
DINGO-WILD DOG HYBRIDS(Canis lupus familiaris)
Domestic and ‘wild’ dogs(Canis lupus familiaris)
R. sanguineus s.l. distribution
Red fox distribution
Overlapping R. sanguineus s.l. and red fox distribution
Rockhampton
Sarina
Autochthonous or non- autochthonous infection
of H. canis in dam? 2017
First H. canis infection diagnosed in Australia in a
Maremma Sheepdog
Intrauterine transmission?
Transm
itted by vector?
Vector images designed by Freepik (www.freepik.com). Fonts from www.dafont.com designed by DaFont (Linux Libertine), Scott Simpson (Mermaid) and Natanael Gama (Cinzel)
Figure 4




















