
Comparative in vitro antimicrobial effect of Sarcocephalus ...

Jurnal
AGROTROPTKAISSN 0216-7662
Telah Diakreditasi
VolumeX Nomor I Juni 2005

Regenerasi In-Vitro Tomat Hibrida Kingkong
Menggunakan Eksplan Kotiledon Dan Hipokotil
(In-vitro Regeneration on Kingkong Hybrid Tomato
Using Cotyledon and Hypocotyls)
Sholeh Avivi1)
, Slameto2)
, dan Ully Herawati3)
1) Author for correspondence,
Jurusan Budidaya Pertanian
Fakultas Pertanian Universitas Jember
Jl. Kalimantan 23 Jember 68121
e-mail: [email protected]
Telp/Fax: (0331)335055
HP: 08123569092
2) Jurusan Budidaya Pertanian,
Fakultas Pertanian, Universitas Jember
3)Alumni Fakultas Pertanian, Universitas Jember

2
Regenerasi In-Vitro Tomat Hibrida Kingkong
Menggunakan Eksplan Kotiledon Dan Hipokotil
(In-vitro Regeneration on Kingkong Hybrid Tomato
Using Cotyledons and Hypocotyls)
ABSTRAK
Penelitian ini ditujukan untuk menemukan teknik perbanyakan bibit tomat
dengan bantuan zat pengatur tumbuh BAP, IAA dan NAA pada media propagasi
in vitro. Penelitian dilakukan dalam dua tahap yaitu, induksi tunas dan induksi
akar. Konsentrasi BAP yang digunakan pada tahap I adalah 1, 2, dan 3 ppm dalam
media MS + vitamin B5 + 0,2 ppm IAA. Eksplan yang digunakan adalah
kotiledon, pangkal kotiledon, dan hipokotil. Konsentrasi NAA yang digunakan
pada tahap II adalah 0,1; 0,2; dan 0,3 ppm dalam media MS dengan memakai
eksplan tunas yang dihasilkan tahap I. Hasil penelitian menunjukkan bahwa
eksplan kotiledon dan 1 ppm BAP menghasilkan jumlah tunas dan daun paling
banyak (rata-rata 9,8 dan 3,6). Sedangkan 0,1 ppm NAA paling baik dapat
menginduksi akar (jumlah akar dan panjang akar berturut-turut 20,1 dan 2,8 cm).
Dengan demikian teknologi perbanyakan bibit tomat terbaik adalah eksplan
kotiledon yang ditanam menggunakan media MS + 0,2 ppm IAA + 1 ppm BAP
untuk induksi tunas, dan untuk induksi perakaran paling baik digunakan media
MS + 0,1 ppm NAA.
Kata Kunci: eksplan, BAP, NAA, IAA, Lycopersicum esculentum.
ABSTRACT
The objectives of this research were to find the micropropagation
technique using Benzylamino purin (BAP), Indole Acetic Acid (IAA), and
Naphthaleneacetic acid (NAA). There were two steps in this study, shoot induction
and root induction. At the first step, BAP concentrations used were 1, 2, and 3
ppm. All of them were supplemented with 0.2 ppm IAA in MS medium + B5
vitamin and combined with three types of explants. The explants were cotyledon,
cotyledon base, and hypocotyls. At the second step, NAA concentrations used were
0.1, 0.2, and 0.3 ppm. The result showed cotyledon explants and 0.2 ppm IAA + 1

3
ppm BAP had the highest number of shoot and leave (9.80 and 3.60, respectively).
NAA 0.1 ppm significantly affected the root induction of tomato (the number of
root and root length were 20.14 and 2.79 cm, respectively). Therefore, it could be
concluded that the best regeneration method in this study was the application 0.2
ppm IAA + 1 ppm BAP in the MS medium and the cotyledon as the explants for
shoot induction. In addition, 0.1 ppm NAA in the MS medium could be
recommended as the best root growth regulator.
Key words: explants, BAP, NAA, IAA, Lycopersicum esculentum.
PENDAHULUAN
Komoditas tomat telah dijadikan wahana wirausahatani yang cukup
menggiurkan, terutama jenis-jenis tomat yang mempunyai reputasi kualitas
ekspor. Peluang menembus pasar ekspor, tidak terlepas dari kehadiran tomat
hibrida; seperti tomat varietas Kingkong (TW-372) yang merupakan tomat hibrida
hasil introduksi dari Taiwan. Tomat Kingkong ini memiliki beberapa keunggulan
seperti buahnya berukuran besar (+ 150 g per buah), tahan terhadap penyakit layu
bakteri, tahan terhadap suhu panas, dan rasanya masam sehingga sangat cocok
untuk bahan industri saus (Trubus, 1993).
Faktor benih memegang peran penting dalam pertanian. Data tahun 1999
menyebutkan bahwa petani Indonesia hanya memakai sekitar 13% benih
hortikultura berkualitas sedangkan sisanya dipenuhi dengan berbagai macam cara
misalnya dengan mengulang benih. Benih berkualitas tadi 90% masih dipenuhi
dengan impor sehingga Indonesia menjadi pasar benih yang potensial (Kompas,
2001).
Tingginya harga benih tomat hibrida yang tidak diikuti dengan upaya
pengadaan benih yang efisien, menyebabkan perlunya dicarikan alternatif untuk
pengadaan bibit, misalnya melalui teknik kultur jaringan. Diharapkan dengan
teknik kultur jaringan dapat diperoleh bibit tomat dengan mutu yang sama dengan
bibit tomat yang diperoleh dari biji hibrida. Hal ini disebabkan melalui kultur

4
jaringan dapat diperbanyak bibit yang berasal dari seluruh bagian tanaman, yang
mengandung DNA yang sama dengan induknya.
Nogueira et al. (2001) menggunakan media MS dalam regenerasi tunas
eksplan kotiledon tomat cv. Santa Clara. Nirmala (1998) juga menggunakan
media MS untuk induksi kalus dari eksplan kotiledon tomat varietas Kemir.
Sabapathi and Gupta (1986) menggunakan media MS untuk mengetahui pengaruh
faktor genetik terhadap induksi tunas dari eksplan hipokotil pada beberapa
varietas tomat. Penelitian yang dilakukan oleh Taylor dan El-Kheir (1993) pada
kultur akar kecambah tomat cv. VFN8 juga menggunakan media dasar MS.
Arrillaga et al. (2001) dalam penelitiannya mengunakan eksplan leaflet dan
kotiledon tomat liar (Lycopersicon cheesmanii). Locy (1983) dalam Arillaga et al.
(2001) menyatakan bahwa eksplan dari hipokotil dan daun tomat liar L.
cheesmanii memiliki potensial rendah dalam pembentukan organ (organogenesis).
Masih sedikit peneliti yang melaporkan teknik perbanyakan bibit tomat
melalui kultur jaringan. Bahkan hingga saat ini belum ada yang melaporkan
teknik perbanyakan in-vitro bibit tomat hibrida yang beredar di Indonesia mulai
dari teknik pemilihan jenis eksplan yang digunakan, teknis perbanyakan tunasnya,
hingga teknik perakaran. Rekayasa paket teknologi perbanyakan bibit tomat dalam
penelitian ini meliputi pemilihan eksplan terbaik, teknik perbanyakan tunas, dan
perakaran secara in-vitro.
BAHAN DAN METODE
Penelitian dilaksanakan di Laboratorium Kultur Jaringan, Jurusan BDP,
FAPERTA, UNEJ mulai bulan Agustus 2002 sampai dengan Januari 2003.
Bahan yang digunakan meliputi benih tomat varietas hibrida Kingkong,
agar sebagai pemadat, sukrosa, IAA, BAP, NAA, media MS (Murashige and
Skoog), Vitamin B5, desinfektan (Clorox 10%, Benlate, Agrept, Alkohol 70%,
Betadine, dan Aquadest steril), dan bahan lain yang mendukung penelitian ini.
Alat yang digunakan meliputi peralatan gelas (botol kultur, gelas ukur, beaker
glass, erlenmeyer, dan petridish), timbangan analitik, pH meter, autoclave,

5
Laminar Air Flow (LAF), peralatan diseksi (pinset, gunting, dan skalpel), stirer,
lampu spiritus, rak kultur, dan alat pendukung lainnya.
Penelitian Tahap I: Induksi tunas; menggunakan Rancangan Acak Lengkap
(RAL) secara faktorial, terdiri atas 2 faktor dan 5 ulangan. Faktor yang digunakan
adalah: faktor pertama, konsentrasi BAP (B) yang terdiri 3 taraf yaitu B1 = 1 ppm;
B2 = 2 ppm; dan B3 = 3 ppm yang masing-masing ditambahkan dengan 0,2 ppm
IAA. Faktor kedua, macam eksplan (E) yang terdiri 3 taraf yaitu E1 = kotiledon, E2
= pangkal kotiledon, dan E3 = hipokotil.
Penelitian Tahap II: Induksi akar; menggunakan Rancangan Acak Lengkap
(RAL) dengan 7 ulangan dan eksplan berupa eksplan tunas dari tahap I. Faktor
yang digunakan adalah konsentrasi NAA (N) yang terdiri 3 taraf yaitu N1 = 0,1
ppm; N2 = 0,2 ppm; dan N3 = 0,3 ppm. Data dianalisis dengan sidik ragam, jika
menunjukkan berbeda nyata dilanjutkan dengan Uji Duncan pada taraf 5%.
HASIL DAN PEMBAHASAN
Kedinian Pembentukan Tunas
Perlakuan yang diberikan untuk parameter kedinian pembentukan tunas,
menunjukkan respon berbeda nyata baik pada faktor media perlakuan (B) maupun
macam eksplan (E). Namun, faktor interaksi (BE) menunjukkan respon berbeda
tidak nyata. Pertumbuhan tunas yang berasal dari eksplan kotiledon dan
kecambah tomat yang merupakan asal dari eksplan kotiledon disajikan pada
Gambar 1.
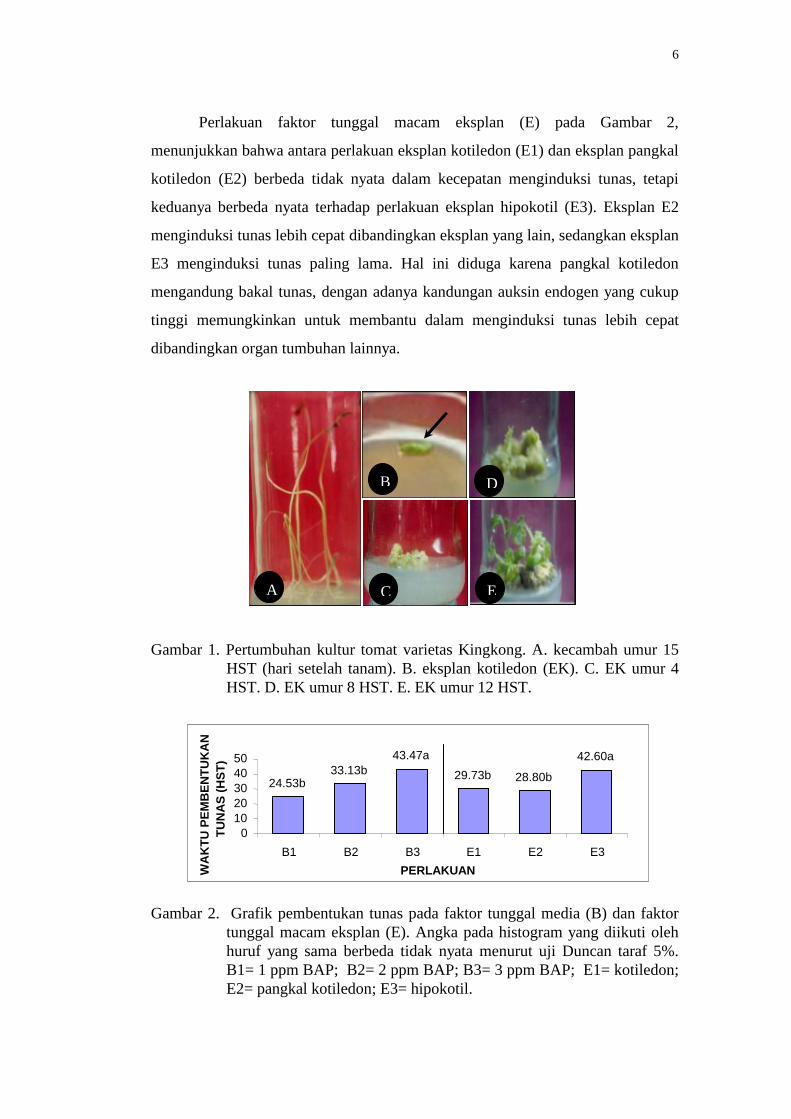
Pada Gambar 2. tampak bahwa perlakuan faktor media (B) menunjukkan
berbeda tidak nyata antara perlakuan media MS + 0,2 ppm IAA + 1 ppm BAP
(B1) dan media MS + 0,2 ppm IAA + 2 ppm BAP (B2), tetapi keduanya
menunjukkan berbeda nyata terhadap media MS + 0,2 ppm IAA + 3 ppm BAP
(B3). Media B1 memiliki rata-rata kedinian menginduksi tunas paling cepat
dibandingkan media lainnya sedangkan media B3 paling lama dalam menginduksi
tunas. Ini berarti perbandingan/nisbah konsentrasi BAP dan IAA yang terlalu
tinggi dapat menghambat awal tumbuhnya tunas pada eksplan kultur tomat.
6
Perlakuan faktor tunggal macam eksplan (E) pada Gambar 2,
menunjukkan bahwa antara perlakuan eksplan kotiledon (E1) dan eksplan pangkal
kotiledon (E2) berbeda tidak nyata dalam kecepatan menginduksi tunas, tetapi
keduanya berbeda nyata terhadap perlakuan eksplan hipokotil (E3). Eksplan E2
menginduksi tunas lebih cepat dibandingkan eksplan yang lain, sedangkan eksplan
E3 menginduksi tunas paling lama. Hal ini diduga karena pangkal kotiledon
mengandung bakal tunas, dengan adanya kandungan auksin endogen yang cukup
tinggi memungkinkan untuk membantu dalam menginduksi tunas lebih cepat
dibandingkan organ tumbuhan lainnya.
Gambar 1. Pertumbuhan kultur tomat varietas Kingkong. A. kecambah umur 15
HST (hari setelah tanam). B. eksplan kotiledon (EK). C. EK umur 4
HST. D. EK umur 8 HST. E. EK umur 12 HST.
Gambar 2. Grafik pembentukan tunas pada faktor tunggal media (B) dan faktor
tunggal macam eksplan (E). Angka pada histogram yang diikuti oleh
huruf yang sama berbeda tidak nyata menurut uji Duncan taraf 5%.
B1= 1 ppm BAP; B2= 2 ppm BAP; B3= 3 ppm BAP; E1= kotiledon;
E2= pangkal kotiledon; E3= hipokotil.
A
B
C
D
E
24.53b 33.13b
43.47a
29.73b 28.80b
42.60a
0 10 20 30 40 50
B1 B2 B3 E1 E2 E3
PERLAKUAN
TU
NA
S (
HS
T)
WA
KT
U P
EM
BE
NT
UK
AN

7
Umumnya sitokinin paling banyak terdapat di organ muda (biji, daun,
buah) dan di ujung akar sedangkan auksin banyak disintesis di ujung
batang/kuncup, daun muda, dan ujung akar (Salisbury dan Ross, 1995). Eksplan
E1 (kotiledon = daun muda) memang tidak memiliki bakal tunas, namun dengan
adanya kandungan sitokinin dan auksin endogen yang cukup dapat menginduksi
tunas lebih cepat dibandingkan E3 dengan cara mensintesis cadangan makanan
yang tersimpan dalam organ tersebut. Sintesis bahan makanan yang terjadi akibat
adanya auksin dapat digunakan sebagai sumber tenaga dalam pertumbuhan
(Hendaryono dan Wijayani, 1994).
Eksplan E3 menginduksi tunas paling lama. Ini dikarenakan sitokinin dan
auksin endogen mungkin tidak disintesis di dalam hipokotil sehingga kandungan
sitokinin dan auksinnya lebih sedikit, padahal sitokinin berperan dalam
menginduksi terbentuknya tunas. Untuk itu, perlu adanya tambahan sitokinin
eksogen dalam jumlah yang cukup. Brown et al. (1997) menyatakan bahwa jika
perbandingan antara IAA dan kinetin sebesar 1:3 maka tidak akan terbentuk organ
tunas melainkan akan terjadi perbesaran sel yang disebut kalus. Duzyaman et al.
(2000) mengatakan bahwa media dengan konsentrasi kinetin sebesar 1.2 M dapat
menginduksi tunas tertinggi eksplan hipokotil, kotiledon 15 hari, dan leaflet pada
tomat.
Jumlah Tunas
Perlakuan yang diberikan untuk parameter jumlah tunas, menunjukkan
respon berbeda nyata baik pada faktor media perlakuan (B) maupun macam
eksplan (E). Namun, faktor interaksi (BE) menunjukkan respon berbeda tidak
nyata.
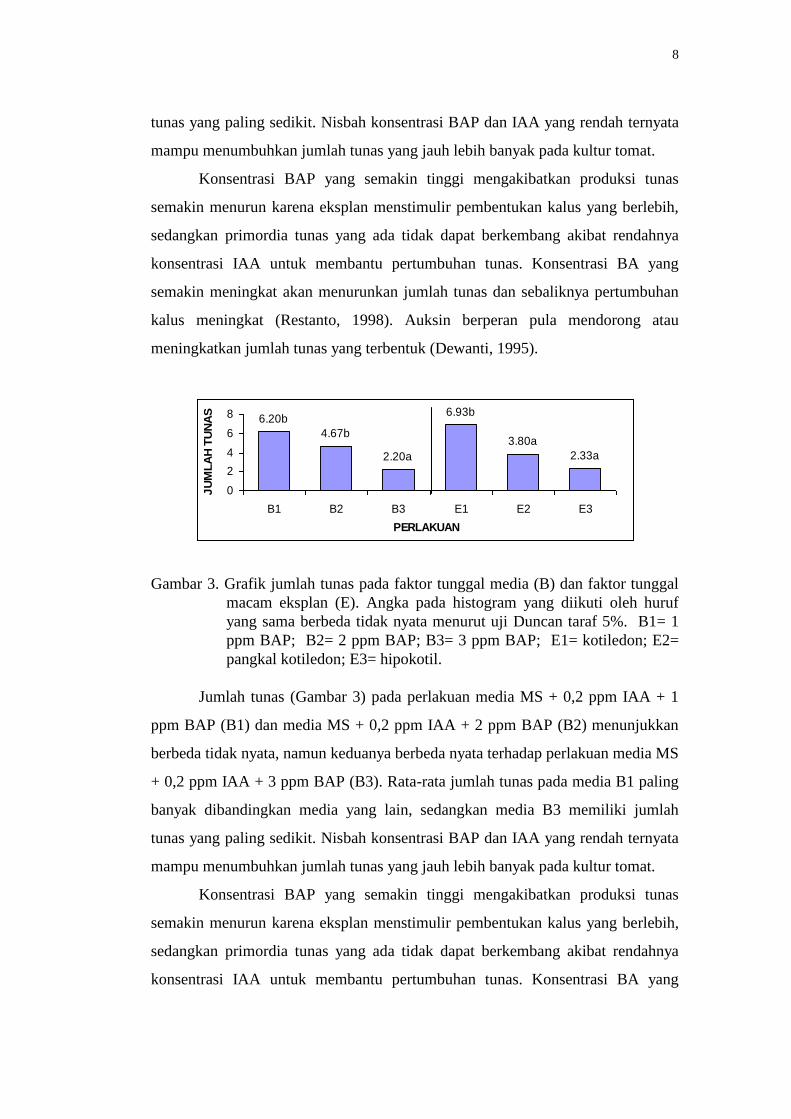
Jumlah tunas (Gambar 3) pada perlakuan media MS + 0,2 ppm IAA + 1
ppm BAP (B1) dan media MS + 0,2 ppm IAA + 2 ppm BAP (B2) menunjukkan
berbeda tidak nyata, namun keduanya berbeda nyata terhadap perlakuan media MS
+ 0,2 ppm IAA + 3 ppm BAP (B3). Rata-rata jumlah tunas pada media B1 paling
banyak dibandingkan media yang lain, sedangkan media B3 memiliki jumlah
8
6.20b
4.67b
2.20a
6.93b
3.80a
2.33a
0
2
4
6
8
B1 B2 B3 E1 E2 E3
PERLAKUAN
JU
ML
AH
TU
NA
S
tunas yang paling sedikit. Nisbah konsentrasi BAP dan IAA yang rendah ternyata
mampu menumbuhkan jumlah tunas yang jauh lebih banyak pada kultur tomat.
Konsentrasi BAP yang semakin tinggi mengakibatkan produksi tunas
semakin menurun karena eksplan menstimulir pembentukan kalus yang berlebih,
sedangkan primordia tunas yang ada tidak dapat berkembang akibat rendahnya
konsentrasi IAA untuk membantu pertumbuhan tunas. Konsentrasi BA yang
semakin meningkat akan menurunkan jumlah tunas dan sebaliknya pertumbuhan
kalus meningkat (Restanto, 1998). Auksin berperan pula mendorong atau
meningkatkan jumlah tunas yang terbentuk (Dewanti, 1995).
Gambar 3. Grafik jumlah tunas pada faktor tunggal media (B) dan faktor tunggal
macam eksplan (E). Angka pada histogram yang diikuti oleh huruf
yang sama berbeda tidak nyata menurut uji Duncan taraf 5%. B1= 1
ppm BAP; B2= 2 ppm BAP; B3= 3 ppm BAP; E1= kotiledon; E2=
pangkal kotiledon; E3= hipokotil.
Jumlah tunas (Gambar 3) pada perlakuan media MS + 0,2 ppm IAA + 1
ppm BAP (B1) dan media MS + 0,2 ppm IAA + 2 ppm BAP (B2) menunjukkan
berbeda tidak nyata, namun keduanya berbeda nyata terhadap perlakuan media MS
+ 0,2 ppm IAA + 3 ppm BAP (B3). Rata-rata jumlah tunas pada media B1 paling
banyak dibandingkan media yang lain, sedangkan media B3 memiliki jumlah
tunas yang paling sedikit. Nisbah konsentrasi BAP dan IAA yang rendah ternyata
mampu menumbuhkan jumlah tunas yang jauh lebih banyak pada kultur tomat.
Konsentrasi BAP yang semakin tinggi mengakibatkan produksi tunas
semakin menurun karena eksplan menstimulir pembentukan kalus yang berlebih,
sedangkan primordia tunas yang ada tidak dapat berkembang akibat rendahnya
konsentrasi IAA untuk membantu pertumbuhan tunas. Konsentrasi BA yang

9
semakin meningkat akan menurunkan jumlah tunas dan sebaliknya pertumbuhan
kalus meningkat (Restanto, 1998). Auksin berperan pula mendorong atau
meningkatkan jumlah tunas yang terbentuk (Dewanti, 1995).
Dalam hal menghasilkan tunas, perlakuan eksplan kotiledon (E1)
menghasilkan jumlah yang berbeda nyata terhadap eksplan pangkal kotiledon (E2)
dan hipokotil (E3), sedangkan eksplan E2 berbeda tidak nyata terhadap E3
(Gambar 3). Kotiledon merupakan bagian dari kecambah tanaman dikotil yang
merupakan tempat penyimpanan cadangan makanan (seperti karbohidrat, lemak,
dan protein). Pada kecambah yang utuh, cadangan makanan yang terdapat dalam
kotiledon diserap guna disintesis untuk menghasilkan energi bagi pertumbuhan
dan perkembangan tanaman itu. Kotiledon juga disebut sebagai daun muda
tumbuhan dikotil, yang membentuk klorofil dan melakukan proses fotosintesis
dengan bantuan cahaya (Salisbury dan Ross, 1995). Fungsi kotiledon inilah yang
diduga menjadi faktor utama dapat memproduksi jumlah tunas lebih banyak
dibandingkan organ yang lain. Hendaryono dan Wijayani (1994) berpendapat
bahwa, kenaikan sintesa bahan-bahan makanan dapat digunakan sebagai sumber
tenaga dalam pertumbuhan.
Tinggi Tunas
Perlakuan yang diberikan untuk parameter tinggi tunas menunjukkan
respon berbeda nyata pada faktor tunggal media perlakuan (B), faktor tunggal
macam eksplan (E), dan faktor interaksi (BE).
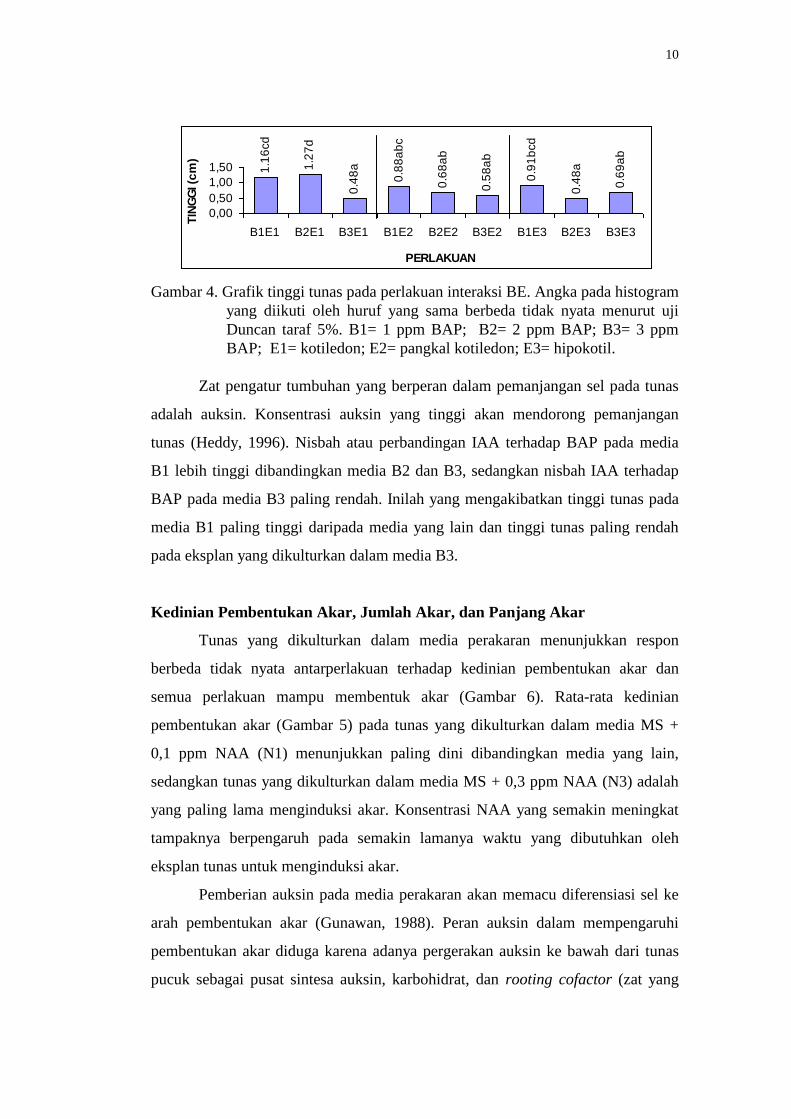
Perlakuan media MS + 0,2 ppm IAA + 2 ppm BAP dengan eksplan
kotiledon (B2E1) menunjukkan rata-rata tinggi tunas yang paling tinggi
dibandingkan dengan perlakuan yang lain (Gambar 4). Perlakuan eksplan
kotiledon dalam media MS + 0,2 ppm IAA + 1 ppm BAP (B1E1) menunjukkan
respon berbeda tidak nyata terhadap B2E1 dan kedua perlakuan tersebut berbeda
nyata terhadap eksplan kotiledon dalam media MS + 0,2 ppm IAA + 3 ppm BAP
(B3E1). Perlakuan B3E1 memiliki rata-rata tinggi tunas yang paling rendah
termasuk juga perlakuan eksplan hipokotil dalam media MS + 0,2 ppm IAA + 2
ppm BAP (B2E3) dibandingkan dengan perlakuan yang lain (Gambar 4).
10
0.6
9a
b
0.4
8a
0.9
1b
cd
0.5
8a
b
0.6
8a
b
0.8
8a
bc
0.4
8a1
.27
d
1.1
6cd
0,00
0,50
1,00
1,50
B1E1 B2E1 B3E1 B1E2 B2E2 B3E2 B1E3 B2E3 B3E3
PERLAKUAN
TIN
GG
I (c
m)
Gambar 4. Grafik tinggi tunas pada perlakuan interaksi BE. Angka pada histogram
yang diikuti oleh huruf yang sama berbeda tidak nyata menurut uji
Duncan taraf 5%. B1= 1 ppm BAP; B2= 2 ppm BAP; B3= 3 ppm
BAP; E1= kotiledon; E2= pangkal kotiledon; E3= hipokotil.
Zat pengatur tumbuhan yang berperan dalam pemanjangan sel pada tunas
adalah auksin. Konsentrasi auksin yang tinggi akan mendorong pemanjangan
tunas (Heddy, 1996). Nisbah atau perbandingan IAA terhadap BAP pada media
B1 lebih tinggi dibandingkan media B2 dan B3, sedangkan nisbah IAA terhadap
BAP pada media B3 paling rendah. Inilah yang mengakibatkan tinggi tunas pada
media B1 paling tinggi daripada media yang lain dan tinggi tunas paling rendah
pada eksplan yang dikulturkan dalam media B3.
Kedinian Pembentukan Akar, Jumlah Akar, dan Panjang Akar
Tunas yang dikulturkan dalam media perakaran menunjukkan respon
berbeda tidak nyata antarperlakuan terhadap kedinian pembentukan akar dan
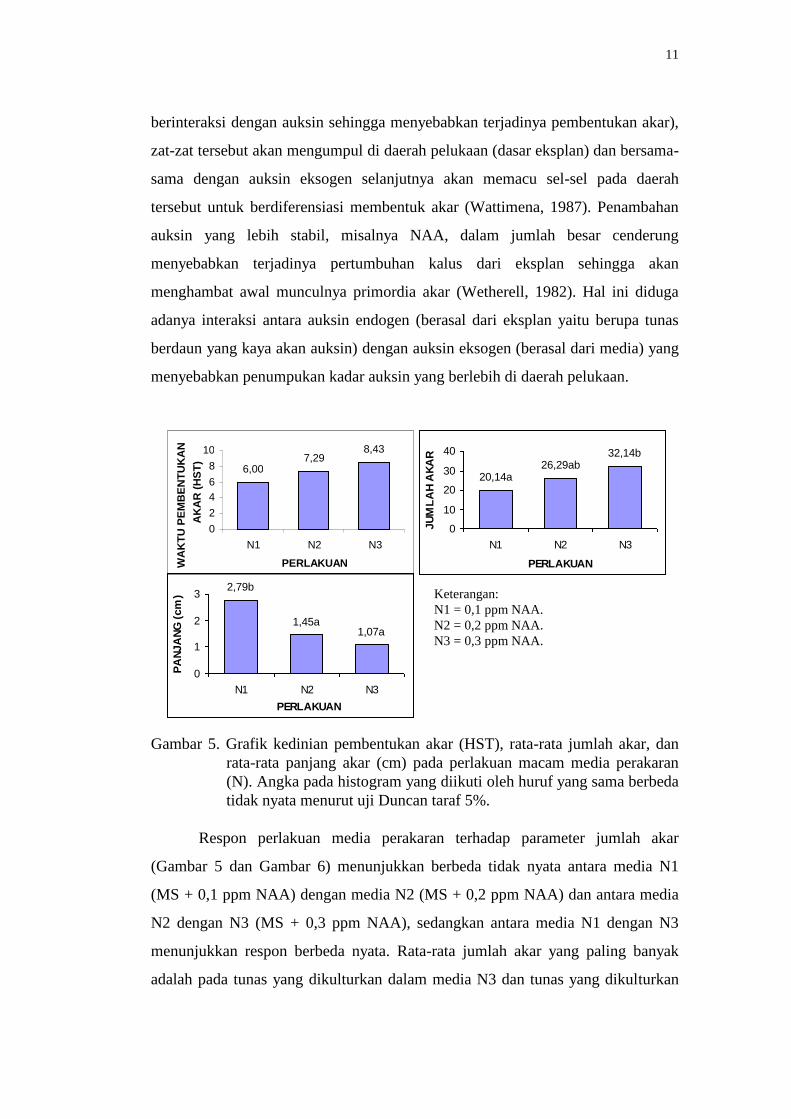
semua perlakuan mampu membentuk akar (Gambar 6). Rata-rata kedinian
pembentukan akar (Gambar 5) pada tunas yang dikulturkan dalam media MS +
0,1 ppm NAA (N1) menunjukkan paling dini dibandingkan media yang lain,
sedangkan tunas yang dikulturkan dalam media MS + 0,3 ppm NAA (N3) adalah
yang paling lama menginduksi akar. Konsentrasi NAA yang semakin meningkat
tampaknya berpengaruh pada semakin lamanya waktu yang dibutuhkan oleh
eksplan tunas untuk menginduksi akar.
Pemberian auksin pada media perakaran akan memacu diferensiasi sel ke
arah pembentukan akar (Gunawan, 1988). Peran auksin dalam mempengaruhi
pembentukan akar diduga karena adanya pergerakan auksin ke bawah dari tunas
pucuk sebagai pusat sintesa auksin, karbohidrat, dan rooting cofactor (zat yang
11
32,14b26,29ab
20,14a
0
10
20
30
40
N1 N2 N3
PERLAKUAN
JU
ML
AH
AK
AR
2,79b
1,45a1,07a
0
1
2
3
N1 N2 N3
PERLAKUAN
PA
NJA
NG
(cm
)
berinteraksi dengan auksin sehingga menyebabkan terjadinya pembentukan akar),
zat-zat tersebut akan mengumpul di daerah pelukaan (dasar eksplan) dan bersama-
sama dengan auksin eksogen selanjutnya akan memacu sel-sel pada daerah
tersebut untuk berdiferensiasi membentuk akar (Wattimena, 1987). Penambahan
auksin yang lebih stabil, misalnya NAA, dalam jumlah besar cenderung
menyebabkan terjadinya pertumbuhan kalus dari eksplan sehingga akan
menghambat awal munculnya primordia akar (Wetherell, 1982). Hal ini diduga
adanya interaksi antara auksin endogen (berasal dari eksplan yaitu berupa tunas
berdaun yang kaya akan auksin) dengan auksin eksogen (berasal dari media) yang
menyebabkan penumpukan kadar auksin yang berlebih di daerah pelukaan.
Gambar 5. Grafik kedinian pembentukan akar (HST), rata-rata jumlah akar, dan
rata-rata panjang akar (cm) pada perlakuan macam media perakaran
(N). Angka pada histogram yang diikuti oleh huruf yang sama berbeda
tidak nyata menurut uji Duncan taraf 5%.
Respon perlakuan media perakaran terhadap parameter jumlah akar
(Gambar 5 dan Gambar 6) menunjukkan berbeda tidak nyata antara media N1
(MS + 0,1 ppm NAA) dengan media N2 (MS + 0,2 ppm NAA) dan antara media
N2 dengan N3 (MS + 0,3 ppm NAA), sedangkan antara media N1 dengan N3
menunjukkan respon berbeda nyata. Rata-rata jumlah akar yang paling banyak
adalah pada tunas yang dikulturkan dalam media N3 dan tunas yang dikulturkan
Keterangan:
N1 = 0,1 ppm NAA.
N2 = 0,2 ppm NAA.
N3 = 0,3 ppm NAA.
7,29 8,43
6,00
0
2
4 6
8
10
N1 N2 N3
PERLAKUAN WA
KT
U P
EM
BE
NT
UK
AN
AK
AR
(H
ST
)

12
dalam media N1 menghasilkan rata-rata jumlah akar yang paling sedikit. Kondisi
tersebut menggambarkan adanya hubungan yang sesuai antara jumlah akar dengan
konsentrasi NAA dalam media, yaitu dengan semakin meningkatnya konsentrasi
NAA yang diberikan dalam media akan semakin banyak jumlah akar yang
dihasilkan oleh eksplan.
Namun, hal ini tidak berlaku pada parameter panjang akar, dimana
semakin tinggi konsentrasi NAA dalam media akan menghambat proses
pemanjangan akar. Ini terlihat dari Gambar 5, tunas yang dikulturkan dalam media
N1 memiliki rata-rata panjang akar lebih panjang dibandingkan media N2 dan
tunas yang dikulturkan dalam media N3 memiliki rata-rata panjang akar lebih
pendek dibandingkan dengan media yang lain. Berdasarkan hasil di atas
menimbulkan dugaan bahwa terdapat reaksi yang bertolak belakang/berlawanan
antara jumlah akar dengan pemanjangan akar terhadap konsentrasi NAA yang
diberikan dalam media perakaran.
Gambar 6. Perbandingan panjang akar pada induksi akar. A. media N1 (0,1 ppm
NAA), B. media N2 (0,2 ppm NAA), dan C. media N3 (0,3 ppm
NAA).
KESIMPULAN
Dari penelitian ini dapat ditarik kesimpulan sebagai berikut:
1. Interaksi antara 0,2 ppm IAA + 1 ppm BAP dengan eksplan kotiledon paling
baik pengaruhnya terhadap induksi tunas kultur in-vitro tomat varietas
Kingkong. Perlakuan ini menghasilkan tunas sebanyak 9,80 per eksplan
dengan rata-rata tinggi tunas 1,16 cm. Sedangkan media MS yang
mengandung 0,1 ppm NAA berpengaruh paling baik terhadap induksi akar
B A C

13
tomat varietas Kingkong, yakni sebanyak 20,14 akar per eksplan dengan rata-
rata panjang akar 2,79 cm.
2. Metode regenerasi terbaik untuk tomat hibrida varietas Kingkong adalah
media MS+0,2 ppm IAA+1 ppm BAP untuk induksi tunas, dengan eksplan
kotiledon sebagai eksplan terbaik, dan penambahan 0,1 ppm NAA dalam
media untuk induksi perakaran.
UCAPAN TERIMAKASIH
Terimakasih diucapkan kepada Dra. Erna Rochiyati, MSi atas saran dan
masukannya sebagai penyelaras bahasa.
DAFTAR PUSTAKA
Arrillaga, I.; C. Gisbert; E. Sales; L. Roig; and V. Moreno. 2001. In-vitro Plant
Regeneration and Gene Transfer in The Wild Tomato Lycopersicon
cheesmanii. Journal of Horticultural Science & Biotechnology 76 (4) : 413-
418.
Brown, S C. 1997. Plant Regeneration using Tissue Culture and Agrobacterium
tumefaciens Transformation. In Partial Fulfillment of Plant Biotechnology.
Roger Williams University. (www. Earthlink.net.)
Dewanti, P. 1995. Pengaruh Konsentrasi Kinetin dan Macam Eksplan terhadap
Hasil Kultur In-vitro Tembakau (Nicotiana tabaccum L.). Laporan
Penelitian (Tidak dipublikasikan). Lembaga Penelitian Universitas Jember,
Jember.
Duzyaman, E A A. Tanriseiver ang G. Gunver. 2000. Comparative Studies on
Regeneration of Different Tissues of Tomato In Vitro. Symposium on
Cultivate of Solanaceae in Wild Winter Climates. (www. Actahort. Org.).
Gunawan, L. W. 1988. Teknik Kultur Jaringan. Laboratorium Kultur Jaringan
PAU Bioteknologi Tanaman IPB, Bogor.
Heddy, S. 1996. Hormon Tumbuhan. PT Raja Grafindo Persada, Jakarta.
Hendaryono, D. P. S. dan A. Wijayani. 1994. Teknik Kultur Jaringan. Kanisius,
Yogyakarta.

14
Kompas. 2001. Darajingga, Tomat Indo Temuan Putri Indonesia (online).
http://www.kompas.com/kompas-cetak/0110/28/iptek/dara22.htm, diakses
pada 25 Pebruari 2002.
Nirmala, R. 1998. Pengaruh 2,4-D dan Kombinasi NAA dengan Kinetin Terhadap
Pertumbuhan dan Perkecambahan Kalus Tomat (Lycopersicon esculentum
MILL.) Varietas Kemir. Frontir-Journal of Mulawarman University
(online). http://unmul.ac.id/dat/pub/frontir/ratna.pdf, diakses pada 10 Maret
2002.
Nogueira, F. T. S.; M. C. Costa; M. L. Figueira; W. C. Otoni; and F. L. Finger.
2001. In-vitro Regeneration of ‘Santa Clara’ Tomato Plantlets and Its
Natural Mutant ‘Firme’. Cienc. Agrotec : 63-71 (online). http://www.
editorauflagr/revista/25-1/art08.pdf, diakses pada 25 Pebruari 2002.
Restanto, D. P. 1998. Studi Regenerasi Kedelai secara In-vitro. Laporan
Penelitian (Tidak dipublikasikan). Lembaga Penelitian Universitas Jember,
Jember.
Sabapathi, B. R. and N. Gupta. 1986. Genotypic Differences in Shoot Forming
Capacity of Cultured Hypocotyl Explants of Tomato. Report of The Tomato
Genetics Cooperative (online). http://ars-genome.cornell.edu/tgc/news-
letters/vol36/ v36p26.html, diakses pada 25 Pebruari 2002.
Salisbury, F. B. dan C. W. Ross. 1995. Fisiologi Tumbuhan (edisi bahasa
Indonesia). Penerbit ITB, Bandung.
Taylor, B. H. and R. A. El-Kheir. 1993. Induction of Lateral Root Initiation by
Auxin in Tomato Seedling Roots. Report of the Tomato Genetics
Cooperative : 46-48 (online). http://ars-genome.cornell.edu/tgc/newsletter/
vol43/p46.html, diakses pada 19 Pebruari 2002.
Trubus. 1993. Tomat Kingkong yang Menggiurkan. Trubus (Kumpulan Kliping
Tomat). Pusat Informasi Pertanian, Jakarta.
Wattimena, G. A. 1987. Zat Pengatur Tumbuh Tanaman. Laboratorium Kultur
Jaringan Tanaman PAU Bioteknologi IPB, Bogor.
Wetherell, D. F., 1982, Pengantar Propagasi Tanaman secara In-vitro (edisi
bahasa Indonesia). IKIP Semarang Press, Semarang.
Copyright © 2022 FDOKUMEN




















