
CSE 505 Lecture 6: Interrupts in 8086 - Shahadat Hussain ...

10.1128/MCB.25.6.2450-2462.2005.
2005, 25(6):2450. DOI:Mol. Cell. Biol. Sung Key JangWoo Jae Kim, Sung Hoon Back, Vit Kim, Incheol Ryu and Signaling under Stress ConditionsGranules Interrupts Tumor Necrosis Factor Sequestration of TRAF2 into Stress
http://mcb.asm.org/content/25/6/2450Updated information and services can be found at:
These include:
REFERENCEShttp://mcb.asm.org/content/25/6/2450#ref-list-1at:
This article cites 50 articles, 23 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on Septem
ber 24, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from
on Septem
ber 24, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

MOLECULAR AND CELLULAR BIOLOGY, Mar. 2005, p. 2450–2462 Vol. 25, No. 60270-7306/05/$08.00�0 doi:10.1128/MCB.25.6.2450–2462.2005Copyright © 2005, American Society for Microbiology. All Rights Reserved.
Sequestration of TRAF2 into Stress Granules Interrupts TumorNecrosis Factor Signaling under Stress Conditions
Woo Jae Kim,† Sung Hoon Back,† Vit Kim, Incheol Ryu, and Sung Key Jang*National Research Laboratory, Postech Biotech Center, Department of Life Science, Division of Molecular and Life Sciences,
Pohang University of Science and Technology, Pohang, Kyungbuk, Korea
Received 26 August 2004/Returned for modification 14 September 2004/Accepted 24 November 2004
The cellular stress response (SR) is a phylogenetically conserved protection mechanism that involvesinhibition of protein synthesis through recruitment of translation factors such as eIF4G into insoluble stressgranules (SGs) and blockade of proinflammatory responses by interruption of the signaling pathway fromtumor necrosis factor alpha (TNF-�) to nuclear factor-�B (NF-�B) activation. However, the link between thesetwo physiological phenomena has not been clearly elucidated. Here we report that eIF4GI, which is a scaffoldprotein interacting with many translation factors, interacts with TRAF2, a signaling molecule that plays a keyrole in activation of NF-�B through TNF-�. These two proteins colocalize in SGs during cellular exposure tostress conditions. Moreover, TRAF2 is absent from TNFR1 complexes under stress conditions even afterTNF-� treatment. This suggests that stressed cells lower their biological activities by sequestration of trans-lation factors and TRAF2 into SGs through a protein-protein interaction.
Virtually all organisms, from prokaryotes to humans, arecapable of mounting the stress response (SR) for cellular sur-vival under stress conditions. Pretreatment of an organism witha sublethal stress can protect that organism from subsequentlethal shock responses to a more severe stress. For instance,high-temperature preconditioning reduces acute lung injury,septic shock, and liver damage caused by ischemia reperfusioninjury in mice, resulting in a dramatic increase in survival rate(19, 30, 40). These phenomena are attributed to the inhibitionof the proinflammatory nuclear factor-�B (NF-�B) signaltransduction pathway by hyperthermia preconditioning (30, 31,44). Hsp70 was suggested to be responsible for heat-inducedprotection against tumor necrosis factor alpha (TNF-�)-in-duced lethal inflammatory shock (32, 38, 46, 47), althoughinhibition of TNF signaling was also demonstrated in the ab-sence of Hsp70 expression in cells preconditioned with heat(30, 44) or sodium arsenite (SA) treatments (18).
At the cellular level, discrete cytoplasmic foci called stressgranules (SGs) are formed in environmentally stressed eukary-otic cells (1, 2, 21, 23, 24, 26, 29, 36, 41), perhaps through theaction of translation factor eIF2� (24, 29). In heat-, thapsigar-gin-, or arsenite-treated cells, the SGs contain most of thecomponents of the translational 48S preinitiation complex (i.e.,small, but not large, ribosomal subunits, eIF3, eIF4E, eIF4G,eIF2, and eIF2B), other RNA-binding proteins such as T-cell-restricted intracellular antigens-1 (TIA-1) and T-cell-restrictedintracellular antigen-related protein (TIAR), and untranslatedmRNAs (1, 2, 23, 24, 26). As a consequence, mRNA transla-tion is generally inhibited under stress conditions (14, 43).However, the relationship between the translational blockage
and the anti-inflammatory response has not been clearly elu-cidated in previous work (13). Here we demonstrate a molec-ular link between these two physiological phenomena.
MATERIALS AND METHODS
Plasmid construction. To obtain the specific cDNA sequence of eIF4GI,cDNA corresponding to amino acids (aa) 1 to 370 of eIF4GI was amplified fromChang liver cDNA library (6) by nested PCR using four primers: 5�CAAGCGACACAAATGAACAC3� and 5�CTCTGGGCTTGACTCCACCTC3� for thefirst round of PCR and 5�AGGTTACCCGGGAACCATGAACACGCCTTCTCAGC3� and 5�AGGTTACTGCAGTTACAGATCTTCAGATGGGAC3� forthe second round of PCR. The amplified cDNA fragment and plasmid pSK(�)were digested with SmaI and PstI and then ligated together to construct pSK-eIF4GI(1–370). The sequence of eIF4GI was confirmed by sequencing. To con-struct pSK-eIF4GI(full length), pSK-eIF4GI(1-370) was treated with EcoRI. Togenerate eIF4GI, a full-length eIF4GI clone, plasmids pSK-eIF4GI (1-370) andpSK-HFCI (provided by Robert E. Rhoads, Louisiana State University) weretreated with EcoRI and then ligated together. To construct pGBT9-eIF4GI(182-589), pSK-HFCI treated with XmaI-Klenow was inserted into pGBT9 treatedwith SmaI. To construct pGBT9-eIF4GI(182-415), pGBT9-eIF4GI(182-589) wastreated with AflII-BamHI-Klenow and then self-ligated. To construct pGBT9-eIF4GI(415-589), pGBT9-eIF4GI(182-589) was treated with AflII-Klenow-SalIand the inserted into pGBT9 treated with BamHI-Klenow-SalI. To constructpRSET-A-eIF4GI(182-549), pSK-HFCI was treated with EcoRI-EcoNI-Klenowand then inserted into pRSET-A treated with NheI-Klenow-PvuI.
To construct the plasmid pGBT9-eIF4GI series used in the Y2H assays shownin Fig. 1D, cDNAs corresponding to the indicated amino acids were amplifiedfrom pSK-eIF4GI by PCRs using proper pairs of primers. The amplified cDNAfragments and pGBT9 were digested with EcoRI and BamHI and then ligatedtogether to generate a series of plasmids. The clones were confirmed by DNAsequencing. To construct pEBG-eIF4GI�C(1–509), pSK-eIF4GI(full length)was treated with XmaI-EcoNI-Klenow and then inserted into pEBG-Mnk1 (pro-vided by Robert J. Schneider, New York University) treated with BamHI-Kle-now. To construct pEBG-eIF4GI(full length), pSK-eIF4GI(full length) wastreated with XbaI-HindIII-Klenow and then inserted into pEBG-Mnk1 treatedwith BamHI-Klenow. To construct pEBG, pEBG-Mnk1 was treated withBamHI-Klenow and self-ligated. To construct pEBG-eIF4GI(293–303), PCRwas performed with oligonucleotides 5�CGGGATCCCGATACAAATGGCCGTAGAAGCC3� and 5�CCATCGATGGCTAGATGGGGGTTGAGGCTTCTA3�. The amplified DNA fragment and pEBG were digested with BamHI-ClaI andthen ligated together. To construct pEBG-eIF4GI(293-303) mut, PCR was per-formed with the two oligonucleotides 5�CGGGATCCCGATACAAATGTCTGTAGAAGAA3� and 5�CCATCGATGGCTAGATGGGGGTTGATTCTTCTA3�. The amplified DNA fragment and pEBG were digested with BamHI-ClaI and
* Corresponding author. Mailing address: NRL, PBC, Departmentof Life Science, Division of Molecular and Life Sciences, PohangUniversity of Science and Technology, Hyoja-Dong San 31, Pohang,Kyungbuk 790-784, Korea. Phone: 82 54 279 2298. Fax: 82 54 279 8009.E-mail: [email protected].
† W.J.K. and S.H.B. contributed equally to the work.
2450
on Septem
ber 24, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from
then ligated together. Note that the underlined regions in the oligonucleotidescan anneal each other. To construct pEGFP-CI-eIF4GI, pSK-eIF4GI(fulllength) was treated with XbaI-KpnI-Klenow and then inserted into pEGFP-CItreated with HindIII-Klenow.
To obtain a cDNA clone of eIF4GII, nested PCR was performed using a
Chang liver cDNA library (6). Four primers, 5�ACGCTGGTATACCCTCAAAC3� and 5�TGATAATTTTTCGCTAACTCC3� for the first round of PCR and5�AGGTTACCCGGGAACCATGAATTCACAACCTCAAAC3� and 5�AGGTTACCCGGGTTAATTAATTAGGTTAGTACTAG3� for the second round ofPCR, were used to generate nucleotides encoding amino acids 1 to 357 of
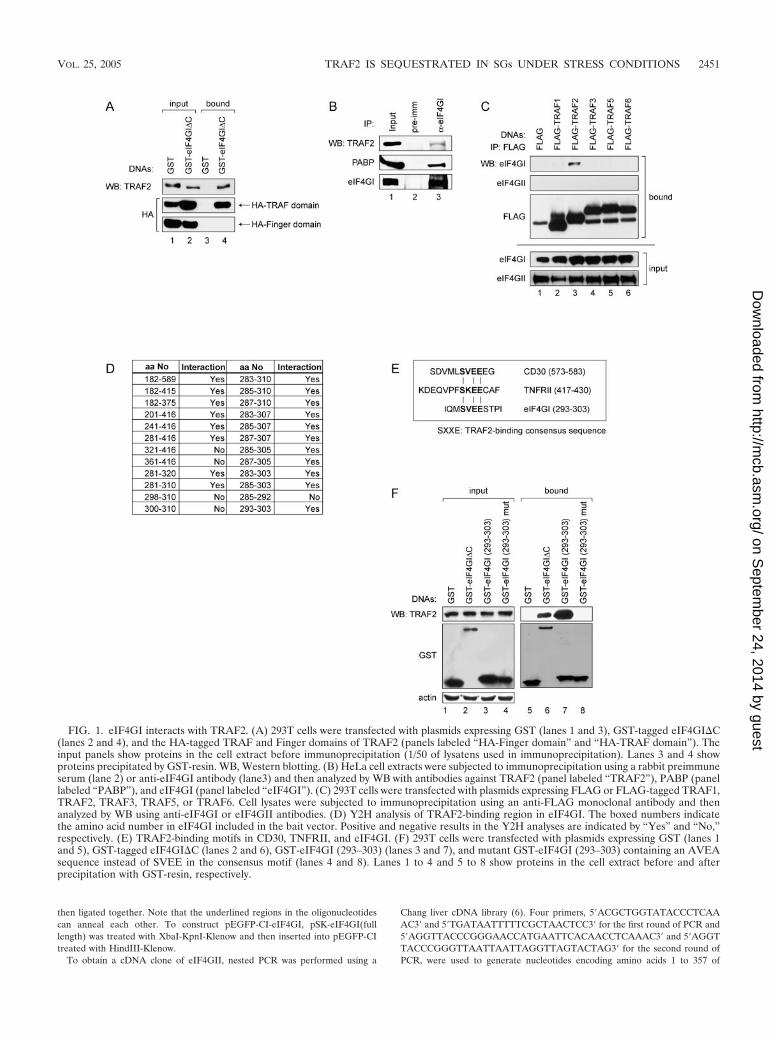
FIG. 1. eIF4GI interacts with TRAF2. (A) 293T cells were transfected with plasmids expressing GST (lanes 1 and 3), GST-tagged eIF4GI�C(lanes 2 and 4), and the HA-tagged TRAF and Finger domains of TRAF2 (panels labeled “HA-Finger domain” and “HA-TRAF domain”). Theinput panels show proteins in the cell extract before immunoprecipitation (1/50 of lysatens used in immunoprecipitation). Lanes 3 and 4 showproteins precipitated by GST-resin. WB, Western blotting. (B) HeLa cell extracts were subjected to immunoprecipitation using a rabbit preimmuneserum (lane 2) or anti-eIF4GI antibody (lane3) and then analyzed by WB with antibodies against TRAF2 (panel labeled “TRAF2”), PABP (panellabeled “PABP”), and eIF4GI (panel labeled “eIF4GI”). (C) 293T cells were transfected with plasmids expressing FLAG or FLAG-tagged TRAF1,TRAF2, TRAF3, TRAF5, or TRAF6. Cell lysates were subjected to immunoprecipitation using an anti-FLAG monoclonal antibody and thenanalyzed by WB using anti-eIF4GI or eIF4GII antibodies. (D) Y2H analysis of TRAF2-binding region in eIF4GI. The boxed numbers indicatethe amino acid number in eIF4GI included in the bait vector. Positive and negative results in the Y2H analyses are indicated by “Yes” and “No,”respectively. (E) TRAF2-binding motifs in CD30, TNFRII, and eIF4GI. (F) 293T cells were transfected with plasmids expressing GST (lanes 1and 5), GST-tagged eIF4GI�C (lanes 2 and 6), GST-eIF4GI (293–303) (lanes 3 and 7), and mutant GST-eIF4GI (293–303) containing an AVEAsequence instead of SVEE in the consensus motif (lanes 4 and 8). Lanes 1 to 4 and 5 to 8 show proteins in the cell extract before and afterprecipitation with GST-resin, respectively.
VOL. 25, 2005 TRAF2 IS SEQUESTRATED IN SGs UNDER STRESS CONDITIONS 2451
on Septem
ber 24, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

eIF4GII. The amplified cDNA and plasmid pSK(�) were digested with SmaIand then ligated together to construct pSK-eIF4GII(1-357). 5� CTGTCACTGCAGCTAGCGAC3� and 5�CCGGAAATTCACTGTGTTAC3� were used to gen-erate nucleotides encoding aa 209 to 834 of eIF4GII, which was digested withNheI and NcoI and then ligated into NheI-NcoI-digested pSK(�) to constructpSK-eIF4GII(209-834). 5�GGCTTACGCAAACATGTGTC3� and 5�TCCCCGCGGGGATTAGTTATCCTCAGACTC3� were used to generate nucleotidesencoding aa 808 to 1585 of eIF4GII, which was digested with NcoI and KspI andthen ligated into NcoI-KspI-digested pSK(�) to construct pSK-eIF4GII(808-1585). To construct a full-length eIF4GII clone, pSK-eIF4GII, nucleotides gen-erated from both NheI-NcoI-digested pSK-eIF4GII(209-834) and NcoI-KspI-digested pSK-eIF4GII(808-1585) were inserted into NheI-KspI-digested pSK-eIF4GII(1-357). The clone was confirmed by DNA sequencing. To constructpRSET-A-eIF4GII(197-545), nested PCR was performed using pSK-eIF4GII.5�CGCGGATCCGCGGGGACTGTGGAGAGCGCTCATC3� and 5�CCGGAATTCCGGTCAATCTTGTTCAGATTCAAGTAC3� were used to generatenucleotides encoding aa 197 to 545 of eIF4GII, which was digested with BamHIand EcoRI and then inserted into BamHI-EcoRI-digested pRSET-A.
Plasmid pAD/GAL4-2-1-TRAF2 is the original clone isolated from the Changliver cDNA library. To construct pTM1-TRAF2, pAD/GAL4-2-1-TRAF2 wastreated with BamHI-SalI-Klenow and then inserted into pTM1 treated withStuI-NcoI-Klenow. To construct pCMV-HA-TRAF2, pTM1-TRAF2 was treatedwith NcoI-SalI-Klenow and then inserted into pCMV-HA treated with EcoRI-SmaI-Klenow. To construct pAD/GAL4-2-1-Finger domain, pAD/GAL4-2-1-TRAF2 was treated with BamHI-Klenow and then self-ligated. To constructpAD/GAL4-2-1-TRAF domain, pSK-TRAF domain was treated with BamHI-EcoRV and then inserted into pAD/GAL4-2-1-TRAF2 treated with SalI-Kle-now-BamHI. To construct pSK-TRAF domain, pAD/GAL4-2-1-TRAF2 wastreated with DraIII-BstXI-T4 polymerase and then inserted into pSK treatedwith SmaI. pRK-FLAG-TRAF1, pRK-FLAG-TRAF2, pRK-FLAG-TRAF3,pRK-FLAG-TRAF5, and pRK-FLAG-TRAF6 were provided by D. V. Goeddelof Tularik Inc. To construct pDsRed-TRAF2, pRK-FLAG-TRAF2 was treatedwith HindIII-Klenow-BamHI and then inserted into pDsRed-CI treated withXbaI-Klenow-BamHI.
Antibodies and chemicals. Antibody against eIF4GI and eIF4GII was pre-pared in our lab (see below). TRAF2 was purchased from BD Pharmingen,FLAG was purchased from Sigma, actin was purchased from ICN, hsp27 waspurchased from StressGen, and phospho-eIF2� was purchased from Cell Sig-naling Technology. Antibodies against hemagglutinin (HA), TIA-1, TNFR1,lamin A/C, hsp70, eIF3�, eIF4AII, DAP5, p65, and RIP were purchased fromSanta Cruz. Antibodies against PABP and PTB4 were gifts of Gideon Dreyfussof Howard Hughes Medical Institute, and glutathione S-transferase (GST) an-tibody was a gift of Sung Ho Ryu of POSTECH. Human recombinant TNF-� waspurchased from Roche. Puromycin, sodium arsenite, cadmium chloride, magne-sium chloride, hydrogen peroxide, and carbonyl cyanide p-trifluoromethoxyphe-nylhydrazone (FCCP) were purchased from Sigma.
Cell cultures and transient transfection. 293T, HeLa, and COS-7 cells weregrown in the presence of 6.0% CO2 in Dulbecco’s modified Eagle’s medium(Gibco) supplemented with antibiotics (penicillin, 100 U/ml; streptomycin, 100�g/ml) and 10% fetal bovine serum (HyClone) at 37°C. Transient transfectionwas performed as described elsewhere (6). Heat shock of cells was carried out at44°C for indicated times unless otherwise stated.
Y2H screening. Yeast two-hybrid (Y2H) screening was performed by Panbio-net Corp. (www.panbionet.com) using the yeast strain PBN204 (MAT� ura3-52his3-200 ade2-101 trp-901 leu2-3,112 gal4� gal80� ura3::kanMX6-pGAL1-URA3pGAL1-lacZ ade2::pGAL2-ADE2).
Protein expression and purification. Plasmid pRSET-A-eIF4GI(182–549) andpRSET-A-eIF4GII(197–545) was transformed into Escherichia coli strainBL21(DE3) pLysS (Novagen) by the use of heat shock techniques. Cultivation ofE. coli and purification of His-tagged eIF4GI and eIF4GII proteins [His-eIF4GI(182–549) and His-eIF4GII(197–545)] were performed similarly to a methoddescribed elsewhere (5). The proteins were eluted from Ni-nitrilotriacetic acidcolumns with a 60-to-200 mM imidazole gradient. Additional Mono-Q columnchromatography was performed to further purify proteins. The proteins wereeluted from columns with a 240-to-500 mM NaCl gradient.
Preparation of anti-eIF4GI and -eIF4GII polyclonal antibodies. A New Zea-land white rabbit was immunized with 100 �g of the purified recombinant proteinHis-eIF4GI(182–549) and His-eIF4GII(197–545) by subcutaneous injection withcomplete Freud’s adjuvant before and after a 14-day interval.
Fluorescence microscopy. After transfection of DNAs, cells were grown oncoverslips coated with 0.2% gelatin for 48 h and then washed three times withphosphate-buffered saline (PBS). The cells were fixed with 3.5% (wt/vol) para-formaldehyde (Sigma) at room temperature (RT) for 12 min. After being washed
three times with PBS, the cells were permeabilized with 0.1% Triton X-100 at RTfor 2 min and then washed three times with PBS. The samples were soaked inblocking solution (PBS containing 1% bovine serum albumin) for 30 min at RTand then incubated with primary antibodies for 1 h at RT. After being washedwith PBS, the samples were treated with Hoechst 33258 for 2 min at RT andwashed again with PBS three times. Samples were treated with rhodaminetetramethyl isocyanate-conjugated and/or fluorescein isothiocyanate-conjugatedsecondary antibodies (Jackson ImmunoResearch Laboratories, Inc.) for 1 h atRT. Finally, the coverslips were washed three times with PBS, placed on a glassslide, and then sealed with transparent nail polish. The fluorescent images werecaptured with a cooled charge-coupled device camera and a Zeiss (Jena, Ger-many) Axioplan microscope. Data were processed using Adobe Photoshop soft-ware (Mountain View, Calif.). The microscopic observations of proteins wereperformed as described elsewhere (6).
GST pull-down assay. 293T cells transfected with DNAs were lysed by soakingin Triton lysis buffer (0.1% Triton X-100, 25 mM HEPES [pH 7.5], 150 mMNaCl, 10% glycerol, 1 mM dithiothreitol, 0.1 mM EDTA, 2 mM sodium or-thovanadate, 1 �g of aprotinin/ml, 1 �g of antipain/ml, 1 �g of bestatin/ml, 1 �gof pepstatin A/ml, 1 mM phenylmethylsulfonyl fluoride). Lysates were clarifiedby centrifugation at 14,000 g at 4°C for 15 min and then incubated with 20 �lof glutathione-Sepharose 4B at 4°C for 4 h. Precipitated proteins were washedthree times with the lysis buffer, resolved by sodium dodecyl sulfate-polyacryl-amide gel electrophoresis, and then transferred to a nitrocellulose membrane.
Immunoprecipitation. 293T cells and HeLa cells were lysed using the Tritonlysis buffer. The lysates were clarified by centrifugation at 14,000 g for 15 min.Anti-FLAG monoclonal antibody (4 �g), rabbit polyclonal serum against eIF4GI(20 �l), or rabbit preimmune serum (20 �l) was incubated with 20 �l of proteinG- or protein A-agarose overnight in 1 ml of PBS at 4°C. Lysates were preclearedwith 10 �l of protein G- or A-agarose at 4°C for 3 h. Then antibodies conjugatedwith protein G- or A-agarose were incubated with the precleared lysates at 4°Cfor 4 h. Precipitates were washed three times with lysis buffer and analyzed bysodium dodecyl sulfate-polyacrylamide gel electrophoresis. Immunoprecipitationof TNFR1 complex was performed as described elsewhere (49). Briefly, HeLacells were lysed at 4°C for 45 min in 1 ml of lysis buffer (20 mM Tris-HCl [pH7.5], 150 mM NaCl, 1% Triton X-100, 1 mM EDTA, 30 mM NaF, 2 mM sodiumpyrophosphate, 1 �g of aprotinin/ml, 1 �g of antipain/ml, 1 �g of bestatin/ml, 1�g of pepstatin A/ml, 1 mM phenylmethylsulfonyl fluoride). Cellular debris wasremoved by centrifugation performed twice at 10,000 g for 5 min. The celllysates were precleared with 50 �l of protein A-Sepharose beads, incubated at4°C for 2 h with 2 �g of anti-p55 TNF receptor monoclonal antibody, and thenmixed with 40 �l of protein G-Sepharose slurry (Pharmacia) (1:1 with PBS) andincubated for another 2 h. The Sepharose beads were washed twice with 1 ml oflysis buffer, twice with 1 ml of high-salt (1 M NaCl) lysis buffer, and twice morewith 1 ml of lysis buffer.
Luciferase assay. Reporter plasmid pNF-�B-Luc (Stratagene) (0.5 �g) wastransfected into 293T cells by electroporation together with indicated expressionvectors and transfection control plasmid pRLCMV-Luc (Promega) (0.1 �g).Luciferase assays were performed with a dual luciferase assay kit (Promega) perthe manufacturer’s instructions. Firefly luciferase activity values were normalizedby Renilla luciferase activity values that reflect transfection efficiency and generalcellular activities.
RESULTS
eIF4GI interacts with TNF-� receptor-associated factor 2(TRAF2). To understand the molecular basis of the stress re-sponse, we first sought to identify proteins interacting witheIF4GI, which is a component of SGs and a scaffold proteinrequired for the assembly of translational initiation factors (10,16, 27, 35). Yeast two-hybrid (Y2H) analysis was performedusing the N-terminal part of eIF4GI (aa 182 to 589) accordingto the largest eIF4G identified by Byrd et al. (10) as bait, sinceno protein was previously known to interact with this region.The Y2H results showed that human TNF-� receptor associ-ated factor 2 (TRAF2) interacted with the N-terminal part ofeIF4GI (17 out of 23 positive clones obtained by Y2H analy-sis). To determine the regions required for this interaction,Y2H analysis was further performed using serially deletedeIF4GI and TRAF2 gene sequences in the bait and prey vec-
2452 KIM ET AL. MOL. CELL. BIOL.
on Septem
ber 24, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

tors, respectively. The TRAF domain of TRAF2 (about 200amino acids at the C-terminal end) and the N-terminal regionof eIF4GI (aa 182 to 415) were found to be required for theinteraction (data not shown). The TRAF domain is well con-served among TRAF family members and is essential for theformation of TRAF homo- and heterodimers (9). When a Y2Hanalysis was performed using TRAF2 and the N-terminal partof eIF4GII (aa 1 to 357), which is an eIF4GI isoform sharingabout 50% amino acid identity (17), no protein-protein inter-action was detected (data not shown). These results indicatethat eIF4GI but not eIF4GII specifically interacts withTRAF2, likely because the two differ at the TRAF2-bindingsite of eIF4GI (17) (see below). The interaction betweenTRAF2 and eIF4GI was further confirmed by biochemicalmethods. A plasmid expressing the N-terminal region ofeIF4GI (aa 1 to 549) fused with GST (GST-eIF4GI�C) wastransfected into 293T cells, and a GST pull-down assay wasperformed (Fig. 1). Endogenous TRAF2 was pulled down byGST-eIF4GI�C (aa 1 to 549) (Fig. 1A; lane 4 on the top panel)but not by GST alone (Fig. 1A; lane 3 on the top panel). Toconfirm the interaction between eIF4GI and the TRAF do-main of TRAF2 in mammalian cells, plasmids expressing theHA-tagged TRAF or Finger domains of TRAF2 were cotrans-fected with plasmids expressing GST-eIF4GI�C. When cellextracts were precipitated with glutathione-Sepharose 4B, theHA-TRAF domain was coprecipitated, but the HA-Finger do-main was not (Fig. 1A; lane 4 on the middle and the bottompanels).
To confirm the interaction between endogenous eIF4GI andTRAF2 under normal conditions (37°C), HeLa cell extractswere immunoprecipitated with an anti-eIF4GI antibody (Fig.1B). Endogenous TRAF2 was coprecipitated, albeit weakly(see below), by the anti-eIF4GI antibody (Fig. 1B, lane 3 onthe top panel), and the poly(A)-binding protein (PABP), whichis known to make a complex with eIF4GI, was also coprecipi-tated by the anti-eIF4GI antibody (Fig. 1B; lane 3 on themiddle panel). These results indicate that endogenous TRAF2and eIF4GI make a complex in HeLa cell under normal con-ditions.
There are many TRAF2-related proteins belonging to theTRAF family. All members of the TRAF family have a con-served TRAF domain that mediates protein-protein interac-tions (9, 20). Accordingly, we examined whether the TRAFdomains of various TRAF2 homologues interact with eIF4GI.GST-eIF4GI�C was coexpressed with FLAG-tagged TRAF1,TRAF2, TRAF3, TRAF5, or TRAF6, and GST pull-downassays were performed. Interestingly, only TRAF2 coprecipi-tated with eIF4GI (data not shown). Conversely, 293T cellswere transfected with constructs expressing FLAG-taggedTRAF1, TRAF2, TRAF3, TRAF5, and TRAF6, and immu-noprecipitation was performed with an anti-FLAG monoclo-nal antibody. Endogenous eIF4GI was coprecipitated by theanti-FLAG antibody when FLAG-TRAF2 was ectopically ex-pressed in the cells (Fig. 1C; lane 3 on the first panel). Inter-estingly, TRAF1, the TRAF family protein showing the highesthomology with TRAF2 (4, 9), showed weak binding witheIF4GI (Fig. 1C; lane 2 on the first panel), perhaps suggestingthat TRAF1 interacts weakly with eIF4GI. Alternatively,TRAF1 might have coprecipitated with eIF4GI via indirectbinding through the TRAF1-TRAF2 interaction (4, 9). eIF4GI
did not coprecipitate with TRAF3, TRAF5, or TRAF6 (Fig.1C; lanes 4 to 6 on the first panel). Interaction betweeneIF4GII and TRAF2 was not observed by Y2H analysis (datanot shown). Coincident with this, eIF4GII was not coprecipi-tated with any of TRAF members by immunoprecipitation(Fig. 1C; lanes 1 to 6 on the second panel). Taken together,these results indicate that eIF4GI specifically interacts withTRAF2.
We then used the Y2H system to further analyze theTRAF2-binding region of eIF4GI (aa 182 to 415) (Fig. 1D).Y2H analysis revealed that 11 amino acids of eIF4GI (aa 293to 303) not present in eIF4GII were sufficient for the interac-tion with TRAF2. This seems to explain why eIF4GII does notinteract with TRAF2. Interestingly, the TRAF2-interactingmotif SXXE, which is known to be essential for interactionwith TRAF2 (37), resides in the center of the 11 amino acids.The same SXXE motif sequence (SVEE) is found both ineIF4GI and in CD30, which is also known to interact withTRAF2 (8) (bold characters in Fig. 1E). An artificial mutationin the consensus sequence of eIF4GI (from SVEE to AVEA)hampered the interaction between eIF4GI and TRAF2 (Fig.1F), indicating that the SVEE sequence plays a key role in theinteraction between eIF4GI and TRAF2.
TRAF2 migrates to stress granules (SGs) under heat stressconditions. It is known that under heat stress conditions, TNFsignaling is blocked (30, 31, 44) and mRNA translation isgenerally inhibited by the sequestration of translation factorsinto SGs (1, 2, 12, 23, 24, 26, 29). However, the link betweenthese two physiological phenomena has not been clearly elu-cidated. To investigate the relationship between translationand TNF signaling, the subcellular localization patterns of en-dogenous eIF4GI and TRAF2 proteins were investigated byimmunocytochemistry under normal (37°C) and heat stress(44°C) conditions. A polyclonal antibody against eIF4GI and amonoclonal antibody against TRAF2 were used in the immu-nocytochemical analyses (Fig. 2). At 37°C, eIF4GI was local-ized mainly in the cytoplasm and partially in the nucleus ofHeLa cell as previously described by McKendrick et al. (33),whereas TRAF2 proteins showed a punctate pattern in thecytoplasm elsewhere (4, 15). Our subcellular localization dataindicated that most of the TRAF2 and eIF4GI proteins dis-tributed to separate regions of the cell at 37°C (Fig. 2A[“Mock”]). As the localization pattern of eIF4GI was previ-ously shown to change under stress conditions (1, 12, 29), wefurther analyzed the subcellular distribution of eIF4GI andTRAF2 under heat stress conditions. Following cellular incu-bation at 44°C for 0.5 h, some of the eIF4GI proteins werefound in distinct regions of the cell known as SGs localizedaround the nucleus (Fig. 2A [“Heat 0.5 h”]), as previouslydescribed (1, 2, 12, 21, 23, 24, 26, 29, 36, 41). Interestingly, theTRAF2 proteins also migrated to the SGs following heat treat-ment. A large portion of the eIF4GI and cytoplasmic TRAF2proteins colocalized in the SGs after heat treatment for 0.5 h,as shown by the yellow dots in the merged image, althoughsome of the eIF4GI and TRAF2 proteins, especially nuclearTRAF2 (34), remained in distinct regions, as shown by red andgreen signals, respectively (Fig. 2A [“Heat 0.5 h”]). Most of thecytoplasmic eIF4GI and TRAF2 proteins coincided with eachother in the cells heat treated for longer than 1 h (Fig. 2A[“Heat 1 h” and “Heat 3 h”]; also see the enlarged portion on
VOL. 25, 2005 TRAF2 IS SEQUESTRATED IN SGs UNDER STRESS CONDITIONS 2453
on Septem
ber 24, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from
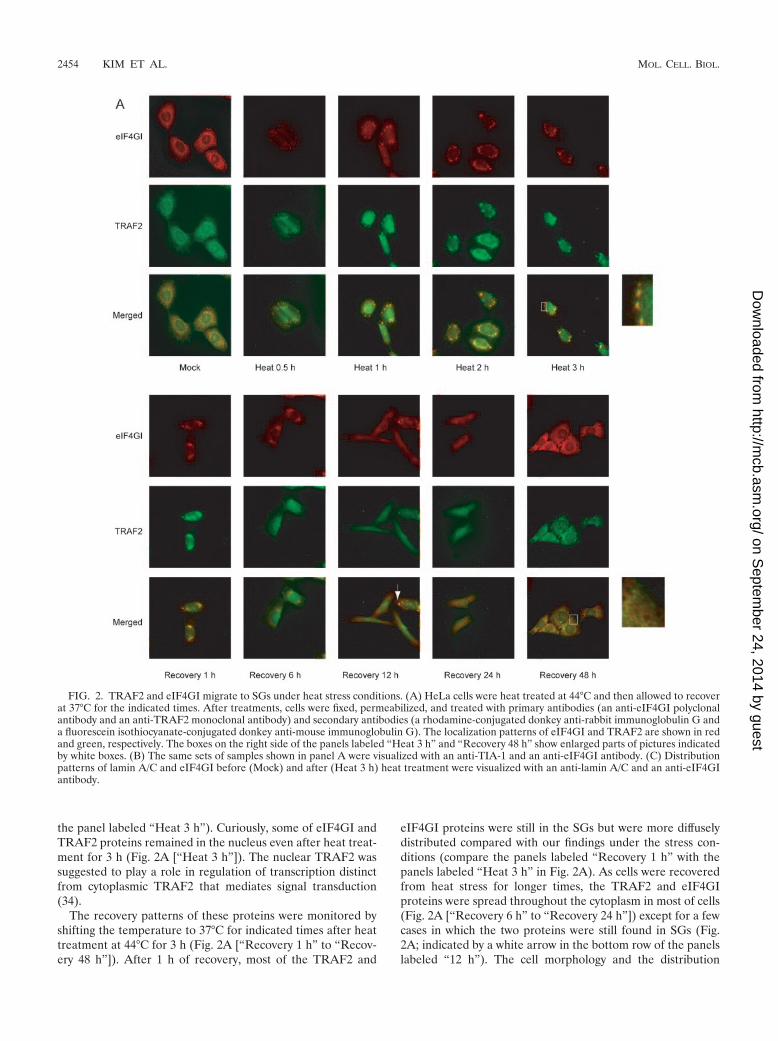
the panel labeled “Heat 3 h”). Curiously, some of eIF4GI andTRAF2 proteins remained in the nucleus even after heat treat-ment for 3 h (Fig. 2A [“Heat 3 h”]). The nuclear TRAF2 wassuggested to play a role in regulation of transcription distinctfrom cytoplasmic TRAF2 that mediates signal transduction(34).
The recovery patterns of these proteins were monitored byshifting the temperature to 37°C for indicated times after heattreatment at 44°C for 3 h (Fig. 2A [“Recovery 1 h” to “Recov-ery 48 h”]). After 1 h of recovery, most of the TRAF2 and
eIF4GI proteins were still in the SGs but were more diffuselydistributed compared with our findings under the stress con-ditions (compare the panels labeled “Recovery 1 h” with thepanels labeled “Heat 3 h” in Fig. 2A). As cells were recoveredfrom heat stress for longer times, the TRAF2 and eIF4GIproteins were spread throughout the cytoplasm in most of cells(Fig. 2A [“Recovery 6 h” to “Recovery 24 h”]) except for a fewcases in which the two proteins were still found in SGs (Fig.2A; indicated by a white arrow in the bottom row of the panelslabeled “12 h”). The cell morphology and the distribution
FIG. 2. TRAF2 and eIF4GI migrate to SGs under heat stress conditions. (A) HeLa cells were heat treated at 44°C and then allowed to recoverat 37°C for the indicated times. After treatments, cells were fixed, permeabilized, and treated with primary antibodies (an anti-eIF4GI polyclonalantibody and an anti-TRAF2 monoclonal antibody) and secondary antibodies (a rhodamine-conjugated donkey anti-rabbit immunoglobulin G anda fluorescein isothiocyanate-conjugated donkey anti-mouse immunoglobulin G). The localization patterns of eIF4GI and TRAF2 are shown in redand green, respectively. The boxes on the right side of the panels labeled “Heat 3 h” and “Recovery 48 h” show enlarged parts of pictures indicatedby white boxes. (B) The same sets of samples shown in panel A were visualized with an anti-TIA-1 and an anti-eIF4GI antibody. (C) Distributionpatterns of lamin A/C and eIF4GI before (Mock) and after (Heat 3 h) heat treatment were visualized with an anti-lamin A/C and an anti-eIF4GIantibody.
2454 KIM ET AL. MOL. CELL. BIOL.
on Septem
ber 24, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

FIG
.2—
Continued.
VOL. 25, 2005 TRAF2 IS SEQUESTRATED IN SGs UNDER STRESS CONDITIONS 2455
on Septem
ber 24, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from
pattern of TRAF2 proteins returned to normal 48 h postrecov-ery, when the distinct punctate patterns of TRAF2 proteinscould be observed (Fig. 2A; see in particular the enlargedportion on the panels labeled “Recovery 48 h”).
The identity of the granules formed by heat stress was con-
firmed by immunocytochemistry with pairs of antibodiesagainst eIF4GI versus TIA-1 (Fig. 2B), eIF4GI versus eIF3�(data not shown), and eIF4GI versus eIF4AII (data not shown)(1), which are known to migrate to SGs during heat stress.TIA-1 proteins were localized mostly in the nucleus at 37°C,
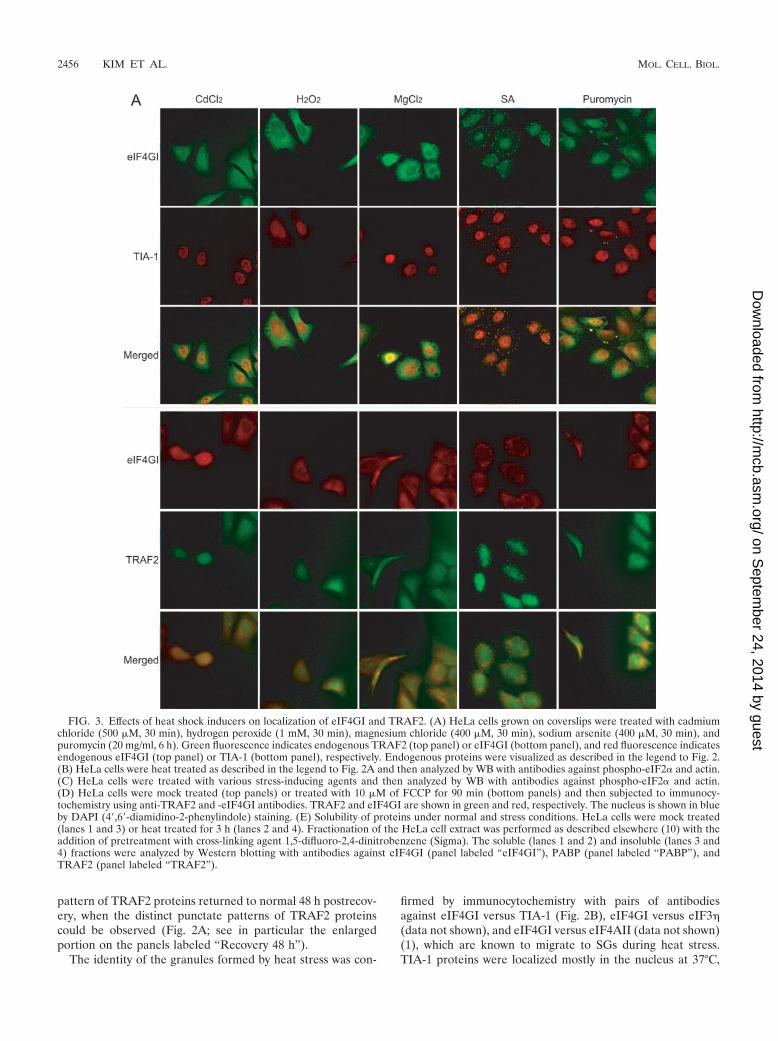
FIG. 3. Effects of heat shock inducers on localization of eIF4GI and TRAF2. (A) HeLa cells grown on coverslips were treated with cadmiumchloride (500 �M, 30 min), hydrogen peroxide (1 mM, 30 min), magnesium chloride (400 �M, 30 min), sodium arsenite (400 �M, 30 min), andpuromycin (20 mg/ml, 6 h). Green fluorescence indicates endogenous TRAF2 (top panel) or eIF4GI (bottom panel), and red fluorescence indicatesendogenous eIF4GI (top panel) or TIA-1 (bottom panel), respectively. Endogenous proteins were visualized as described in the legend to Fig. 2.(B) HeLa cells were heat treated as described in the legend to Fig. 2A and then analyzed by WB with antibodies against phospho-eIF2� and actin.(C) HeLa cells were treated with various stress-inducing agents and then analyzed by WB with antibodies against phospho-eIF2� and actin.(D) HeLa cells were mock treated (top panels) or treated with 10 �M of FCCP for 90 min (bottom panels) and then subjected to immunocy-tochemistry using anti-TRAF2 and -eIF4GI antibodies. TRAF2 and eIF4GI are shown in green and red, respectively. The nucleus is shown in blueby DAPI (4�,6�-diamidino-2-phenylindole) staining. (E) Solubility of proteins under normal and stress conditions. HeLa cells were mock treated(lanes 1 and 3) or heat treated for 3 h (lanes 2 and 4). Fractionation of the HeLa cell extract was performed as described elsewhere (10) with theaddition of pretreatment with cross-linking agent 1,5-difluoro-2,4-dinitrobenzene (Sigma). The soluble (lanes 1 and 2) and insoluble (lanes 3 and4) fractions were analyzed by Western blotting with antibodies against eIF4GI (panel labeled “eIF4GI”), PABP (panel labeled “PABP”), andTRAF2 (panel labeled “TRAF2”).
2456 KIM ET AL. MOL. CELL. BIOL.
on Septem
ber 24, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from
but a part of them migrated to the SGs in the cytoplasm wheresome of eIF4G proteins migrated after heat treatment for0.5 h. Cytoplasmic eIF4GI and TIA-1 were almost completelycolocalized at the SGs after heat treatment for 3 h. SGs wereclearly observed by 6 h postrecovery, as seen by eIF4GI andTRAF2 results (panels labeled “Recovery 1 h” and “Recovery6 h” in Fig. 2B). SGs disappeared from most of cells by 12 hpostrecovery, and eIF4GI and TIA-1 showed normal distribu-tion patterns 48 h postrecovery.
To confirm that the integrity of cells remained intact under
the heat stress conditions, immunocytochemical analyses wereperformed with antibodies against nuclear lamin A/C andeIF4GI (Fig. 2C). The nuclei of cells remained intact by 3 hpost-heat treatment, as indicated by the distribution patternsof lamin A/C that were confined to the nuclei (Fig. 2C; panels“lamin A/C”). The lamin A/C and eIF4GI proteins were local-ized at distinct regions 3 h post-heat treatment (panels labeled“Heat 3 h” in Fig. 2C). These results clearly demonstrate thatTRAF2 and eIF4GI proteins congregate in SGs under heatstress conditions and that these proteins return to separate
FIG. 3—Continued.
VOL. 25, 2005 TRAF2 IS SEQUESTRATED IN SGs UNDER STRESS CONDITIONS 2457
on Septem
ber 24, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

subcellular regions upon cellular return to the nonstress tem-perature of 37°C.
Migration of TRAF2 to SGs was further investigated byobserving distribution patterns of TRAF2, eIF4GI, and TIA-1in the cells treated with various stress-inducing agents (heavymetal ions cadmium chloride [CdCl2] and SA, oxidative stressinducer hydrogen peroxide [H2O2], concentrated divalent ionmagnesium chloride [MgCl2], and polysome destabilizer puro-mycin) (22, 28) (Fig. 3). SA, a well-known heat shock protein(HSP) inducer (1), is often used to induce SG formation (26).SGs were detected in SA-treated cells (Fig. 3A; panels labeled“SA”), as described by Kedersha and Anderson (23). In thesecells, cytoplasmic eIF4GI and TIA-1 were colocalized to theSGs. Cytoplasmic TRAF2 was also localized to the SGs, asindicated by colocalization of eIF4GI and TRAF2 on the pan-els labeled “SA” in Fig. 3A. Curiously, puromycin treatmentinduced SG formation in some, but not all, cells where cyto-plasmic eIF4GI and TIA-1 were colocalized (Fig. 3A; panelslabeled “Puromycin”). Cytoplasmic TRAF2 also migrated tothe SGs in the cells containing SGs (Fig. 3A; panels labeled“Puromycin”). Other stress-inducing chemicals (CdCl2, H2O2,and MgCl2) did not induce SG formation or TRAF2 migration.These data suggest that stress per se does not induce SGformation or TRAF2 migration.
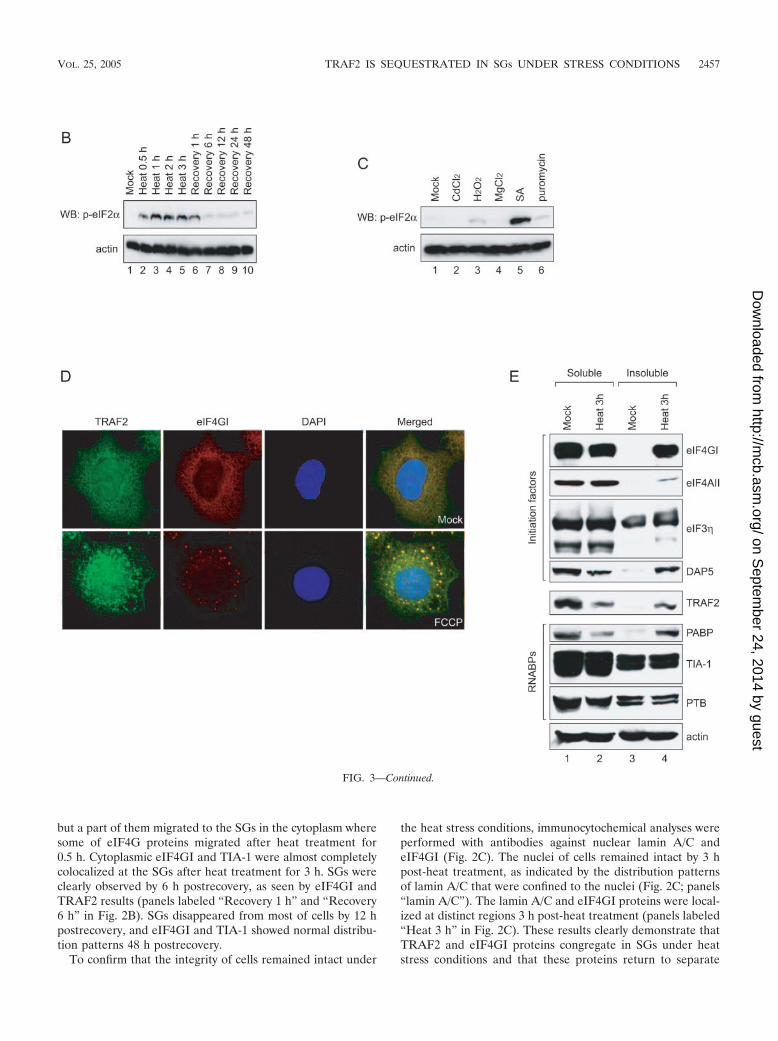
SG formation was suggested to be related to the phosphor-ylation of eIF2� subunits and/or to the reduced availability ofeIF2�-GTP-tRNAi
Met (23, 24, 26). Therefore, we monitoredphosphorylation levels of eIF2� subunits under various stressconditions (Fig. 3B and C). Phosphorylation of eIF2� subunitswas strongly induced by heat treatment and remained at highlevel by 1 h postrecovery (lanes 2 to 6 in Fig. 3B). The phos-phorylation level of eIF2� subunits returned to normal after6 h postrecovery (lanes 7 to 10 in Fig. 3B). SA, a strong SGformer, strongly induced phosphorylation of eIF2� subunits(Fig. 3C, lane 5). Puromycin, a weak SG former, weakly in-duced phosphorylation of eIF2� subunits (Fig. 3C, lane 6).H2O2 did not induce SG formation, even though it inducedweak phosphorylation of eIF2� subunits, in similarity to puro-mycin results (Fig. 3C, lane 3). The molecular basis of thediscrepancy in SG formation by the weak inducers of the eIF2�phosphorylation (puromycin and H2O2) remains to be eluci-dated. Other stress-inducing chemicals (CdCl2 and MgCl2) didnot induce phosphorylation of eIF2� (Fig. 3C, lanes 2 and 4).These results suggest that the phosphorylation of eIF2� sub-units is related to SG formation and that weak phosphorylationof eIF2� subunits occasionally induces SG formation.
The effect of eIF2� phosphorylation on the migration ofTRAF2 to SGs was further investigated by using an ionophoreFCCP that induces SG formation without phosphorylation ofeIF2� (24). As shown on the bottom panel in Fig. 3D, TRAF2and eIF4G1 comigrated to SGs upon treatment with FCCP.This indicates that the formation of SG, rather than the phos-phorylation of eIF2�, is required for the colocalization ofTRAF2 and eIF4G1 to SGs irrespective of the causes of SGformation. This phenomenon provides an explanation for thelong-lasting enigma of how the depletion of the mitochondrialelectron transport eliminates the cytotoxic and gene-inductiveeffects of TNF (42).
The migration of TRAF2 to SGs was further investigated bymeasuring the solubility of TRAF2 and translation factors un-
der normal and heat stress conditions. Upon heat treatmentfor 3 h, large proportions of translation factors eIF4GI,eIF4AII, and DAP5 were accumulated in the insoluble fractionof cell extracts where components of SGs are enriched (Fig.3E; lanes 4 in panels labeled “eIF4GI,” “eIF4AII,” and“DAP5”), as previously shown by Cuesta et al. (12). Similarly,a large proportion of TRAF2 proteins were also enriched inthe insoluble fraction upon heat treatment (Fig. 3E, lane 4 onpanel TRAF2). Conversely, the level of TRAF2 in solublefraction was reduced when cells were treated with heat (Fig.3E; lane 2 on panel labeled “TRAF2”). Redistribution ofTRAF2 to the insoluble subcellular components has been ob-served in the cells containing constitutively active receptors (3)and the cells treated with agents blocking mitochondrial activ-ity (11). The distribution pattern of TRAF2 during heat treat-ment was very similar to that of PABP that augments transla-tion through an interaction with the poly(A) tail of mRNA(compare panels labeled “TRAF2” and “PABP” in Fig. 3E).On the other hand, solubility of other RNA-binding proteins(TIA-1 and PTB) was not changed by the heat treatment (pan-els labeled “TIA-1” and “PTB”). It is noteworthy that thesolubility of TIA-1, which partially migrates to SGs, was notchanged during heat treatment. The dynamic nature of TIA-1in the SGs is likely to contribute to the maintenance of solu-bility of TIA-1 during heat treatment (25).
Taken together, these localization and solubility datastrongly indicate that TRAF2, a key TNF-signaling molecule,is sequestered into SGs under SG-forming conditions.
TRAF2 is not associated with the TNFR1 complex underheat stress conditions. Following treatment with TNF-�, cellsform a complex called the “TNFR1-mediated-signal transduc-tion complex,” which is composed of TNFR1, adaptorTRADD, kinase RIP1, and TRAF2 (49). To determinewhether TRAF2 exists in the TNFR1 complex when cells wereactivated by TNF-�, immunoprecipitation was performed withmonoclonal antibody against TNFR1. Prior to treatment withTNF-�, TRAF2 was weakly coprecipitated with TNFR1 (Fig.4A [lane 1 on panel labeled “TRAF2”]). TRAF2 was stronglycoimmunoprecipitated with TNFR1 when cells were treatedTNF-� at 37°C, as previously described by Zhang et al. (49)(Fig. 4A [lane 2 on panel labeled “TRAF2”]), but TRAF2 wasnot coimmunoprecipitated with TNFR1 with or withoutTNF-� treatment when cells were cultivated at 44°C for 3 h(Fig. 4 [lanes 4 and 3 on panels labeled “bound”]). The level ofTNFR1 on the cell surfaces remained the same even under theheat stress conditions, as analyzed by fluorescence-activatedcell sorter (data not shown). Together, these results indicatethat TRAF2, which is associated with SGs under stress condi-tions, cannot participate in TNFR1 complex formation. Thereduced levels of TRAF2 in the input panels (Fig. 4A [lanes 3and 4 on panel labeled “TRAF2”]) are attributed to insolubil-ity of TRAF2 under stress conditions (Fig. 3D); only solublefractions can be analyzed in immunoprecipitation assays.
TNFR1 complex is composed of several adaptor proteinssuch as TRADD, TRAF2, RIP, and FADD. An immunocyto-chemical experiment was performed to investigate whetherthese proteins are also associated with SGs under heat shockconditions. RIP, a receptor-interacting protein critical for ac-tivation of transcription factor NF-�B (49), was not observed atSGs during heat shock (Fig. 4B, bottom panels). This indicates
2458 KIM ET AL. MOL. CELL. BIOL.
on Septem
ber 24, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from
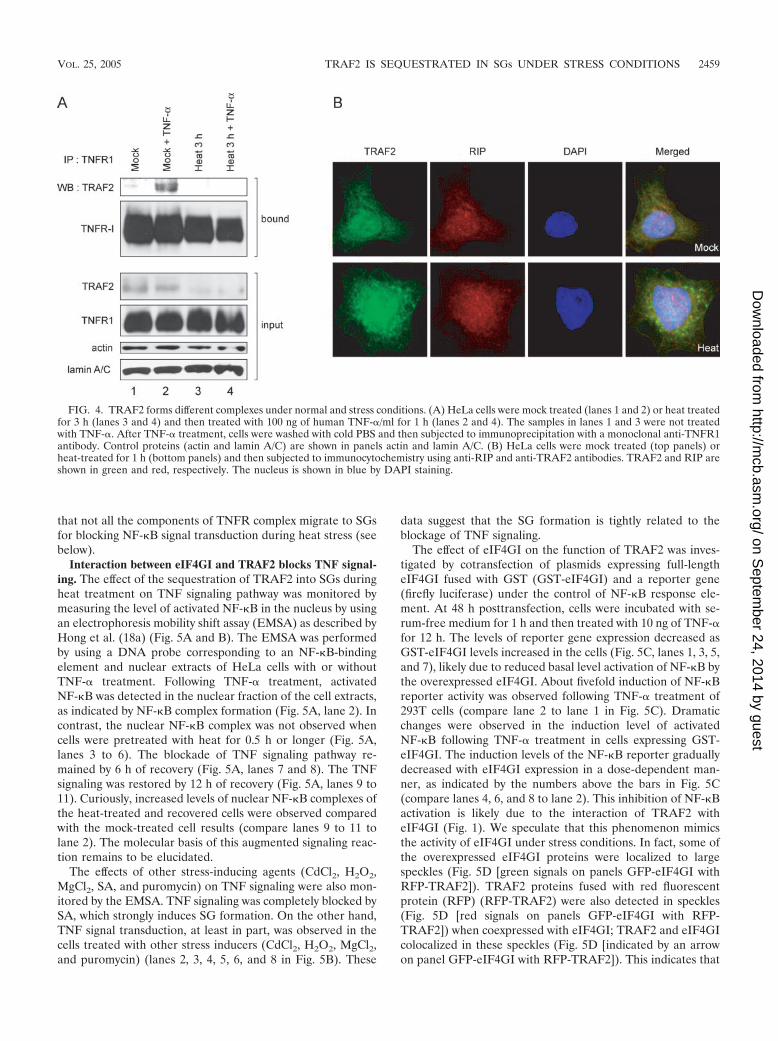
that not all the components of TNFR complex migrate to SGsfor blocking NF-�B signal transduction during heat stress (seebelow).
Interaction between eIF4GI and TRAF2 blocks TNF signal-ing. The effect of the sequestration of TRAF2 into SGs duringheat treatment on TNF signaling pathway was monitored bymeasuring the level of activated NF-�B in the nucleus by usingan electrophoresis mobility shift assay (EMSA) as described byHong et al. (18a) (Fig. 5A and B). The EMSA was performedby using a DNA probe corresponding to an NF-�B-bindingelement and nuclear extracts of HeLa cells with or withoutTNF-� treatment. Following TNF-� treatment, activatedNF-�B was detected in the nuclear fraction of the cell extracts,as indicated by NF-�B complex formation (Fig. 5A, lane 2). Incontrast, the nuclear NF-�B complex was not observed whencells were pretreated with heat for 0.5 h or longer (Fig. 5A,lanes 3 to 6). The blockade of TNF signaling pathway re-mained by 6 h of recovery (Fig. 5A, lanes 7 and 8). The TNFsignaling was restored by 12 h of recovery (Fig. 5A, lanes 9 to11). Curiously, increased levels of nuclear NF-�B complexes ofthe heat-treated and recovered cells were observed comparedwith the mock-treated cell results (compare lanes 9 to 11 tolane 2). The molecular basis of this augmented signaling reac-tion remains to be elucidated.
The effects of other stress-inducing agents (CdCl2, H2O2,MgCl2, SA, and puromycin) on TNF signaling were also mon-itored by the EMSA. TNF signaling was completely blocked bySA, which strongly induces SG formation. On the other hand,TNF signal transduction, at least in part, was observed in thecells treated with other stress inducers (CdCl2, H2O2, MgCl2,and puromycin) (lanes 2, 3, 4, 5, 6, and 8 in Fig. 5B). These
data suggest that the SG formation is tightly related to theblockage of TNF signaling.
The effect of eIF4GI on the function of TRAF2 was inves-tigated by cotransfection of plasmids expressing full-lengtheIF4GI fused with GST (GST-eIF4GI) and a reporter gene(firefly luciferase) under the control of NF-�B response ele-ment. At 48 h posttransfection, cells were incubated with se-rum-free medium for 1 h and then treated with 10 ng of TNF-�for 12 h. The levels of reporter gene expression decreased asGST-eIF4GI levels increased in the cells (Fig. 5C, lanes 1, 3, 5,and 7), likely due to reduced basal level activation of NF-�B bythe overexpressed eIF4GI. About fivefold induction of NF-�Breporter activity was observed following TNF-� treatment of293T cells (compare lane 2 to lane 1 in Fig. 5C). Dramaticchanges were observed in the induction level of activatedNF-�B following TNF-� treatment in cells expressing GST-eIF4GI. The induction levels of the NF-�B reporter graduallydecreased with eIF4GI expression in a dose-dependent man-ner, as indicated by the numbers above the bars in Fig. 5C(compare lanes 4, 6, and 8 to lane 2). This inhibition of NF-�Bactivation is likely due to the interaction of TRAF2 witheIF4GI (Fig. 1). We speculate that this phenomenon mimicsthe activity of eIF4GI under stress conditions. In fact, some ofthe overexpressed eIF4GI proteins were localized to largespeckles (Fig. 5D [green signals on panels GFP-eIF4GI withRFP-TRAF2]). TRAF2 proteins fused with red fluorescentprotein (RFP) (RFP-TRAF2) were also detected in speckles(Fig. 5D [red signals on panels GFP-eIF4GI with RFP-TRAF2]) when coexpressed with eIF4GI; TRAF2 and eIF4GIcolocalized in these speckles (Fig. 5D [indicated by an arrowon panel GFP-eIF4GI with RFP-TRAF2]). This indicates that
FIG. 4. TRAF2 forms different complexes under normal and stress conditions. (A) HeLa cells were mock treated (lanes 1 and 2) or heat treatedfor 3 h (lanes 3 and 4) and then treated with 100 ng of human TNF-�/ml for 1 h (lanes 2 and 4). The samples in lanes 1 and 3 were not treatedwith TNF-�. After TNF-� treatment, cells were washed with cold PBS and then subjected to immunoprecipitation with a monoclonal anti-TNFR1antibody. Control proteins (actin and lamin A/C) are shown in panels actin and lamin A/C. (B) HeLa cells were mock treated (top panels) orheat-treated for 1 h (bottom panels) and then subjected to immunocytochemistry using anti-RIP and anti-TRAF2 antibodies. TRAF2 and RIP areshown in green and red, respectively. The nucleus is shown in blue by DAPI staining.
VOL. 25, 2005 TRAF2 IS SEQUESTRATED IN SGs UNDER STRESS CONDITIONS 2459
on Septem
ber 24, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from
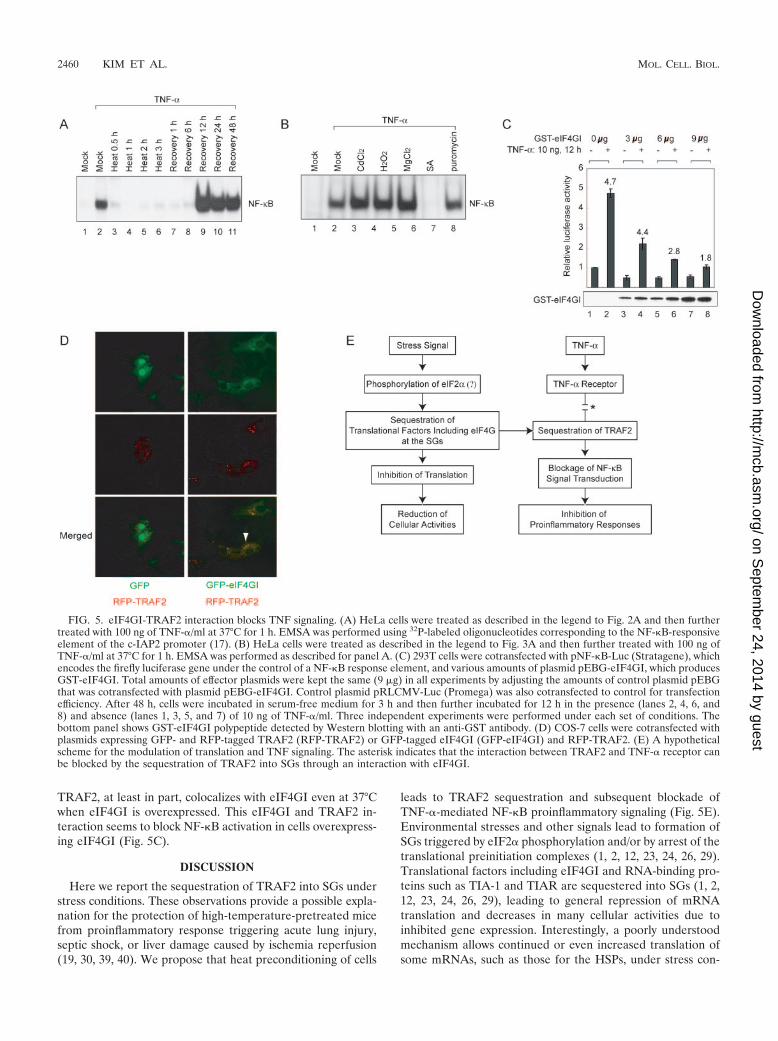
TRAF2, at least in part, colocalizes with eIF4GI even at 37°Cwhen eIF4GI is overexpressed. This eIF4GI and TRAF2 in-teraction seems to block NF-�B activation in cells overexpress-ing eIF4GI (Fig. 5C).
DISCUSSION
Here we report the sequestration of TRAF2 into SGs understress conditions. These observations provide a possible expla-nation for the protection of high-temperature-pretreated micefrom proinflammatory response triggering acute lung injury,septic shock, or liver damage caused by ischemia reperfusion(19, 30, 39, 40). We propose that heat preconditioning of cells
leads to TRAF2 sequestration and subsequent blockade ofTNF-�-mediated NF-�B proinflammatory signaling (Fig. 5E).Environmental stresses and other signals lead to formation ofSGs triggered by eIF2� phosphorylation and/or by arrest of thetranslational preinitiation complexes (1, 2, 12, 23, 24, 26, 29).Translational factors including eIF4GI and RNA-binding pro-teins such as TIA-1 and TIAR are sequestered into SGs (1, 2,12, 23, 24, 26, 29), leading to general repression of mRNAtranslation and decreases in many cellular activities due toinhibited gene expression. Interestingly, a poorly understoodmechanism allows continued or even increased translation ofsome mRNAs, such as those for the HSPs, under stress con-
FIG. 5. eIF4GI-TRAF2 interaction blocks TNF signaling. (A) HeLa cells were treated as described in the legend to Fig. 2A and then furthertreated with 100 ng of TNF-�/ml at 37°C for 1 h. EMSA was performed using 32P-labeled oligonucleotides corresponding to the NF-�B-responsiveelement of the c-IAP2 promoter (17). (B) HeLa cells were treated as described in the legend to Fig. 3A and then further treated with 100 ng ofTNF-�/ml at 37°C for 1 h. EMSA was performed as described for panel A. (C) 293T cells were cotransfected with pNF-�B-Luc (Stratagene), whichencodes the firefly luciferase gene under the control of a NF-�B response element, and various amounts of plasmid pEBG-eIF4GI, which producesGST-eIF4GI. Total amounts of effector plasmids were kept the same (9 �g) in all experiments by adjusting the amounts of control plasmid pEBGthat was cotransfected with plasmid pEBG-eIF4GI. Control plasmid pRLCMV-Luc (Promega) was also cotransfected to control for transfectionefficiency. After 48 h, cells were incubated in serum-free medium for 3 h and then further incubated for 12 h in the presence (lanes 2, 4, 6, and8) and absence (lanes 1, 3, 5, and 7) of 10 ng of TNF-�/ml. Three independent experiments were performed under each set of conditions. Thebottom panel shows GST-eIF4GI polypeptide detected by Western blotting with an anti-GST antibody. (D) COS-7 cells were cotransfected withplasmids expressing GFP- and RFP-tagged TRAF2 (RFP-TRAF2) or GFP-tagged eIF4GI (GFP-eIF4GI) and RFP-TRAF2. (E) A hypotheticalscheme for the modulation of translation and TNF signaling. The asterisk indicates that the interaction between TRAF2 and TNF-� receptor canbe blocked by the sequestration of TRAF2 into SGs through an interaction with eIF4GI.
2460 KIM ET AL. MOL. CELL. BIOL.
on Septem
ber 24, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

ditions. This stress-specific regulation of gene expression al-lows cells to recover from stress damage and minimizes thepotential risks from aberrant gene expression during stressconditions (1, 14, 43). We found that the TRAF2, a signalingmolecule, is also sequestered into the SGs, possibly through aprotein-protein interaction with eIF4GI (Fig. 1). EndogenouseIF4GI interacts with TRAF2 under normal conditions, albeitweakly (Fig. 1B and C), and overexpressed TRAF2 andeIF4GI partially colocalize at 37°C (Fig. 5D), suggesting thateIF4GI interacts weakly with TRAF2 even under normal con-ditions. The interaction between eIF4GI and TRAF2 seems tobe strengthened during stress conditions, as indicated by thestrong colocalization (Fig. 2) and coenrichment of these pro-teins in the insoluble fraction (Fig. 3D) under heat stress con-ditions. The sequestration of TRAF2 into SGs results in theblockage of TRAF2-mediated signaling processes such as TNFsignaling, which subsequently reduces activation of NF-�B(Fig. 4 and 5A to C). As a consequence, proinflammatoryresponses, which are responsible for lethal stress responsessuch as septic shock (7, 45, 48), are nullified when organismsare preconditioned with sublethal stresses.
Sequestration of TRAF2 into SGs breaks the positive-feed-back loop of proinflammatory response through TNF-� andNF-�B. This prevents propagation of the inflammatory re-sponse to neighboring cells and tissues and protects organismsfrom systemic inflammation. The sequestration of TRAF2 tothe SGs may not be the only mechanism functioning in theblockage of the positive-feedback loop of proinflammatory re-sponse. The sequestration of TRAF2 and other safety checkingmechanisms are likely to function in balancing pro- and anti-inflammatory responses in a concerted manner. Investigationsinto the mechanism of sequestration of TRAF2 into SGs mayprovide a clue for a better understanding of the protection oforganisms from environmental stresses and may help research-ers to find novel approaches for protecting against damagingproinflammatory responses.
ACKNOWLEDGMENTS
We thank David V. Goeddel of Tularik Inc. (San Francisco) forproviding the FLAG-tagged TRAF1, TRAF2, TRAF3, TRAF5, andTRAF6 plasmid vectors. We also thank Sung Ho Ryu (Division ofMolecular and Life Sciences, Pohang University of Science and Tech-nology) and Jee Hee Youn (Department of Anatomy, College ofMedicine, Hanyang University) for their valuable suggestions and crit-ical review of the manuscript.
This work was supported in part by grants NRL (M10204000018-03J0000-01410) and MCBRG(M10106000056-03B4500-01010) fromMOST, grants 02-PJ2-PG1-CH16-0002 and 405-VN02-0702-0008 fromKHIDI, grant NCRC(R15-2004-033-01001-0) from KRF, and a grantfrom POSCO.
REFERENCES
1. Anderson, P., and N. Kedersha. 2002. Stressful initiations. J. Cell Sci. 115:3227–3234.
2. Anderson, P., and N. Kedersha. 2002. Visibly stressed: the role of eIF2,TIA-1, and stress granules in protein translation. Cell Stress Chaperones7:213–221.
3. Arch, R. H., R. W. Gedrich, and C. B. Thompson. 2000. Translocation ofTRAF proteins regulates apoptotic threshold of cells. Biochem. Biophys.Res. Commun. 272:936–945.
4. Arron, J. R., Y. Pewzner-Jung, M. C. Walsh, T. Kobayashi, and Y. Choi.2002. Regulation of the subcellular localization of tumor necrosis factorreceptor-associated factor (TRAF)2 by TRAF1 reveals mechanisms ofTRAF2 signaling. J. Exp. Med. 196:923–934.
5. Back, S. H., J. E. Kim, J. Rho, B. Hahm, T. G. Lee, E. E. Kim, J. M. Cho, and
S. K. Jang. 2000. Expression and purification of an active, full-length hepa-titis C viral NS4A. Protein Expr. Purif. 20:196–206.
6. Back, S. H., Y. K. Kim, W. J. Kim, S. Cho, H. R. Oh, J.-E. Kim, and S. K.Jang. 2002. Translation of polioviral mRNA is inhibited by cleavage ofpolypyrimidine tract-binding proteins executed by polioviral 3Cpro. J. Virol.76:2529–2542.
7. Barnes, P. J., and M. Karin. 1997. Nuclear factor-kappaB: a pivotal tran-scription factor in chronic inflammatory diseases. N. Engl. J. Med. 336:1066–1071.
8. Boucher, L. M., L. E. Marengere, Y. Lu, S. Thukral, and T. W. Mak. 1997.Binding sites of cytoplasmic effectors TRAF1, 2, and 3 on CD30 and othermembers of the TNF receptor superfamily. Biochem. Biophys. Res. Com-mun. 233:592–600.
9. Bradley, J. R., and J. S. Pober. 2001. Tumor necrosis factor receptor-asso-ciated factors (TRAFs). Oncogene 20:6482–6491.
10. Byrd, M. P., M. Zamora, and R. E. Lloyd. 2002. Generation of multipleisoforms of eukaryotic translation initiation factor 4GI by use of alternatetranslation initiation codons. Mol. Cell. Biol. 22:4499–4511.
11. Chandel, N. S., P. T. Schumacker, and R. H. Arch. 2001. Reactive oxygenspecies are downstream products of TRAF-mediated signal transduction.J. Biol. Chem. 276:42728–42736.
12. Cuesta, R., G. Laroia, and R. J. Schneider. 2000. Chaperone hsp27 inhibitstranslation during heat shock by binding eIF4G and facilitating dissociationof cap-initiation complexes. Genes Dev. 14:1460–1470.
13. DeMeester, S. L., T. G. Buchman, and J. P. Cobb. 2001. The heat shockparadox: does NF-kappaB determine cell fate? FASEB J. 15:270–274.
14. Duncan, R. F., and J. W. Hershey. 1989. Protein synthesis and proteinphosphorylation during heat stress, recovery, and adaptation. J. Cell Biol.109:1467–1481.
15. Feng, X., M. L. Gaeta, L. A. Madge, J. H. Yang, J. R. Bradley, and J. S.Pober. 2001. Caveolin-1 associates with TRAF2 to form a complex that isrecruited to tumor necrosis factor receptors. J. Biol. Chem. 276:8341–8349.
16. Gingras, A. C., B. Raught, and N. Sonenberg. 1999. eIF4 initiation factors:effectors of mRNA recruitment to ribosomes and regulators of translation.Annu. Rev. Biochem. 68:913–963.
17. Gradi, A., H. Imataka, Y. V. Svitkin, E. Rom, B. Raught, S. Morino, and N.Sonenberg. 1998. A novel functional human eukaryotic translation initiationfactor 4G. Mol. Cell. Biol. 18:334–342.
18. Hershko, D. D., B. W. Robb, E. S. Hungness, G. Luo, and P. O. Hasselgren.2002. Arsenite stabilizes IkappaBalpha and prevents NF-kappaB activationin IL-1 beta-stimulated Caco-2 cells independent of the heat shock response.J. Cell Biochem. 84:687–698.
18a.Hong, S. Y., W. H. Yoon, J. H. Park, S. G. Kang, J. H. Ahn, and T. H. Lee.2000. Involvement of two NF-�B binding elements in tumor necrosis factoralpha-, CD40-, and Epstein-Barr virus latent membrane protein 1-mediatedinduction of the cellular inhibitor of apoptosis protein 2 gene. J. Biol. Chem.275:18022–18028.
19. Hotchkiss, R., I. Nunnally, S. Lindquist, J. Taulien, G. Perdrizet, and I.Karl. 1993. Hyperthermia protects mice against the lethal effects of endo-toxin. Am. J. Physiol. 265:R1447–R1457.
20. Inoue, J., T. Ishida, N. Tsukamoto, N. Kobayashi, A. Naito, S. Azuma, andT. Yamamoto. 2000. Tumor necrosis factor receptor-associated factor(TRAF) family: adapter proteins that mediate cytokine signaling. Exp. CellRes. 254:14–24.
21. Ivanov, P. A., E. M. Chudinova, and E. S. Nadezhdina. 2003. Disruption ofmicrotubules inhibits cytoplasmic ribonucleoprotein stress granule forma-tion. Exp. Cell Res. 290:227–233.
22. Jacquier-Sarlin, M. R., and B. S. Polla. 1996. Dual regulation of heat-shocktranscription factor (HSF) activation and DNA-binding activity by H2O2:role of thioredoxin. Biochem. J. 318:187–193.
23. Kedersha, N., and P. Anderson. 2001. Stress granules: sites of mRNA triagethat regulate mRNA stability and translatability. Biochem. Soc. Trans. 30:963–969.
24. Kedersha, N., S. Chen, N. Gilks, W. Li, I. J. Miller, J. Stahl, and P. Ander-son. 2002. Evidence that ternary complex (eIF2-GTP-tRNA(i)(Met))-defi-cient preinitiation complexes are core constituents of mammalian stressgranules. Mol. Biol. Cell 13:195–210.
25. Kedersha, N., M. R. Cho, W. Li, P. W. Yacono, S. Chen, N. Gilks, D. E.Golan, and P. Anderson. 2000. Dynamic shuttling of TIA-1 accompanies therecruitment of mRNA to mammalian stress granules. J. Cell Biol. 151:1257–1268.
26. Kedersha, N. L., M. Gupta, W. Li, I. Miller, and P. Anderson. 1999. RNA-binding proteins TIA-1 and TIAR link the phosphorylation of eIF-2 alpha tothe assembly of mammalian stress granules. J. Cell Biol. 147:1431–1442.
27. Keiper, B. D., W. Gan, and R. E. Rhoads. 1999. Protein synthesis initiationfactor 4G. Int. J. Biochem. Cell Biol. 31:37–41.
28. Kim, D., S. Somji, S. H. Garrett, M. A. Sens, D. Shukla, and D. A. Sens. 2001.Expression of hsp 27, hsp 60, hsc 70, and hsp 70 by immortalized humanproximal tubule cells (HK-2) following exposure to heat shock, sodiumarsenite, or cadmium chloride. J. Toxicol. Environ. Health Part A 63:475–493.
29. Kimball, S. R., R. L. Horetsky, D. Ron, L. S. Jefferson, and H. P. Harding.
VOL. 25, 2005 TRAF2 IS SEQUESTRATED IN SGs UNDER STRESS CONDITIONS 2461
on Septem
ber 24, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

2003. Mammalian stress granules represent sites of accumulation of stalledtranslation initiation complexes. Am. J. Physiol. Cell Physiol. 284:C273–C284.
30. Malhotra, V., T. Eaves-Pyles, K. Odoms, G. Quaid, T. P. Shanley, and H. R.Wong. 2002. Heat shock inhibits activation of NF-kappaB in the absence ofheat shock factor-1. Biochem. Biophys. Res. Commun. 291:453–457.
31. Malhotra, V., and H. R. Wong. 2002. Interactions between the heat shockresponse and the nuclear factor-kappaB signaling pathway. Crit. Care Med.30:S89–S95.
32. Marber, M. S., R. Mestril, S. H. Chi, M. R. Sayen, D. M. Yellon, and W. H.Dillmann. 1995. Overexpression of the rat inducible 70-kD heat stress pro-tein in a transgenic mouse increases the resistance of the heart to ischemicinjury. J. Clin. Investig. 95:1446–1456.
33. McKendrick, L., E. Thompson, J. Ferreira, S. J. Morley, and J. D. Lewis.2001. Interaction of eukaryotic translation initiation factor 4G with thenuclear cap-binding complex provides a link between nuclear and cytoplas-mic functions of the m(7) guanosine cap. Mol. Cell. Biol. 21:3632–3641.
34. Min, W., J. R. Bradley, J. J. Galbraith, S. J. Jones, E. C. Ledgerwood, andJ. S. Pober. 1998. The N-terminal domains target TNF receptor-associatedfactor-2 to the nucleus and display transcriptional regulatory activity. J. Im-munol. 161:319–324.
35. Morino, S., H. Imataka, Y. V. Svitkin, T. V. Pestova, and N. Sonenberg. 2000.Eukaryotic translation initiation factor 4E (eIF4E) binding site and themiddle one-third of eIF4GI constitute the core domain for cap-dependenttranslation, and the C-terminal one-third functions as a modulatory region.Mol. Cell. Biol. 20:468–477.
36. Nover, L., K. D. Scharf, and D. Neumann. 1983. Formation of cytoplasmicheat shock granules in tomato cell cultures and leaves. Mol. Cell. Biol.3:1648–1655.
37. Park, Y. C., V. Burkitt, A. R. Villa, L. Tong, and H. Wu. 1999. Structural basisfor self-association and receptor recognition of human TRAF2. Nature 398:533–538.
38. Plumier, J. C., B. M. Ross, R. W. Currie, C. E. Angelidis, H. Kazlaris, G.Kollias, and G. N. Pagoulatos. 1995. Transgenic mice expressing the humanheat shock protein 70 have improved post-ischemic myocardial recovery.J. Clin. Investig. 95:1854–1860.
39. Ryan, A. J., S. W. Flanagan, P. L. Moseley, and C. V. Gisolfi. 1992. Acuteheat stress protects rats against endotoxin shock. J. Appl. Physiol. 73:1517–1522.
40. Saad, S., M. Kanai, M. Awane, Y. Yamamoto, T. Morimoto, W. Isselhard, T.Minor, H. Troidl, K. Ozawa, and Y. Yamaoka. 1995. Protective effect of heatshock pretreatment with heat shock protein induction before hepatic warmischemic injury caused by Pringle’s maneuver. Surgery 118:510–516.
41. Scharf, K. D., H. Heider, I. Hohfeld, R. Lyck, E. Schmidt, and L. Nover.1998. The tomato Hsf system: HsfA2 needs interaction with HsfA1 forefficient nuclear import and may be localized in cytoplasmic heat stressgranules. Mol. Cell. Biol. 18:2240–2251.
42. Schulze-Osthoff, K., R. Beyaert, V. Vandevoorde, G. Haegeman, and W.Fiers. 1993. Depletion of the mitochondrial electron transport abrogates thecytotoxic and gene-inductive effects of TNF. EMBO J. 12:3095–3104.
43. Scorsone, K. A., R. Panniers, A. G. Rowlands, and E. C. Henshaw. 1987.Phosphorylation of eukaryotic initiation factor 2 during physiological stresseswhich affect protein synthesis. J. Biol. Chem. 262:14538–14543.
44. Shanley, T. P., M. A. Ryan, T. Eaves-Pyles, and H. R. Wong. 2000. Heatshock inhibits phosphorylation of I-kappaBalpha. Shock 14:447–450.
45. Tang, G., Y. Minemoto, B. Dibling, N. H. Purcell, Z. Li, M. Karin, and A.Lin. 2001. Inhibition of JNK activation through NF-kappaB target genes.Nature 414:313–317.
46. Trost, S. U., J. H. Omens, W. J. Karlon, M. Meyer, R. Mestril, J. W. Covell,and W. H. Dillmann. 1998. Protection against myocardial dysfunction aftera brief ischemic period in transgenic mice expressing inducible heat shockprotein 70. J. Clin. Investig. 101:855–862.
47. Van Molle, W., B. Wielockx, T. Mahieu, M. Takada, T. Taniguchi, K.Sekikawa, and C. Libert. 2002. HSP70 protects against TNF-induced lethalinflammatory shock. Immunity 16:685–695.
48. Wang, C. Y., M. W. Mayo, R. G. Korneluk, D. V. Goeddel, and A. S. Baldwin,Jr. 1998. NF-kappaB antiapoptosis: induction of TRAF1 and TRAF2 andc-IAP1 and c-IAP2 to suppress caspase-8 activation. Science 281:1680–1683.
49. Zhang, S. Q., A. Kovalenko, G. Cantarella, and D. Wallach. 2000. Recruit-ment of the IKK signalosome to the p55 TNF receptor: RIP and A20 bindto NEMO (IKKgamma) upon receptor stimulation. Immunity 12:301–311.
2462 KIM ET AL. MOL. CELL. BIOL.
on Septem
ber 24, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from
Copyright © 2022 FDOKUMEN




















