
Iron-based granules in body of bumblebees
13
1 23 BioMetals An International Journal on the Role of Metal Ions in Biology, Biochemistry and Medicine ISSN 0966-0844 Biometals DOI 10.1007/s10534-014-9805-9 Iron-based granules in body of bumblebees Petr Jandacka, Barbora Kasparova, Yvonna Jiraskova, Katerina Dedkova, Katerina Mamulova-Kutlakova & Jana Kukutschova
Transcript of Iron-based granules in body of bumblebees

1 23
BioMetalsAn International Journal on the Role ofMetal Ions in Biology, Biochemistry andMedicine ISSN 0966-0844 BiometalsDOI 10.1007/s10534-014-9805-9
Iron-based granules in body of bumblebees
Petr Jandacka, Barbora Kasparova,Yvonna Jiraskova, Katerina Dedkova,Katerina Mamulova-Kutlakova & JanaKukutschova

1 23
Your article is protected by copyright and all
rights are held exclusively by Springer Science
+Business Media New York. This e-offprint is
for personal use only and shall not be self-
archived in electronic repositories. If you wish
to self-archive your article, please use the
accepted manuscript version for posting on
your own website. You may further deposit
the accepted manuscript version in any
repository, provided it is only made publicly
available 12 months after official publication
or later and provided acknowledgement is
given to the original source of publication
and a link is inserted to the published article
on Springer's website. The link must be
accompanied by the following text: "The final
publication is available at link.springer.com”.

Iron-based granules in body of bumblebees
Petr Jandacka • Barbora Kasparova • Yvonna Jiraskova •
Katerina Dedkova • Katerina Mamulova-Kutlakova •
Jana Kukutschova
Received: 3 June 2014 / Accepted: 20 October 2014
� Springer Science+Business Media New York 2014
Abstract The paper deals with the presence of iron-
based granules in body parts of bumblebees. Two
groups of bumblebees were collected from their
natural habitat, industrial landscape, and from a
breeding station. Detection of the magnetic particles
was performed by a vibratory magnetometer and their
morphology and elemental composition was analysed
by scanning electron microscopy with EDX micro-
analysis. By means of the EDX spectra, wild bumble-
bees were found to have many magnetic and non-
magnetic particles on their body, containing Fe, O, Al,
Si, Bi, Mg, K, and Ni, likely having origin in the
industrial pollution of the environment. In the case of
bred bumblebees the presence of iron-rich granules,
which occurred more abundantly in subsurface tissues
on the head and wings, was observed. Phase analysis
based on X-ray diffraction shows that iron-based
granules contain magnetite and wuestite and Moss-
bauer spectroscopy admits a superparamagnetic form
of these minerals. Magnetoreception, i.e. the sensory
function of these granules, is discussed within the
paper.
Keywords Magnetoreception � Bumblebee �Biomineralisation � Iron granules
Introduction
Bumblebees (Bombidae sp.) have recently been the
subject of numerous studies in part due to the
increased interest in their protection. They belong to
the insect Hymenoptera order and to the Apidae
family, so their relatives are e.g. honeybees (Apis
mellifera). The honeybee species exhibit excellent
orientation in their habitats and one of the cues for this
property could be a magnetoreception based on
presence of iron-containing particles in their body.
Since the iron-based magnetoreception was already
studied in detail on bodies of honeybees, it could be
possible to expect a very similar occurrence and
significance of iron-based particles in the bodies of
bumblebees (Hsu and Chan 2011; Hsu et al. 2007; Hsu
and Li 1994).
The presence of iron-based magnetic structures in
many living organisms including human beings was
P. Jandacka (&)
IT4Innovations Centre and Institute of Physics,
VSB - Technical University of Ostrava, 17. listopadu
15/2172, 708 33 Ostrava, Czech Republic
e-mail: [email protected]
B. Kasparova � K. Dedkova � K. Mamulova-Kutlakova �J. Kukutschova
Nanotechnology Centre, VSB - Technical University
of Ostrava, 17. listopadu 15/2172, 708 33 Ostrava,
Czech Republic
Y. Jiraskova
Institute of Physics of Materials, Academy of Sciences
of the Czech Republic, Zizkova 22, 616 62 Brno,
Czech Republic
123
Biometals
DOI 10.1007/s10534-014-9805-9
Author's personal copy

previously described by some research studies (Safar-
ik and Safarikova 2002). Their role is still subject to
discussion, but one of their functions could be
magnetoreception, which may be connected with the
movement of magnetic crystals in tissues. Proven
magnetoreception exhibits magnetotactic bacteria via
magnetite or greigite magnetosomes arranged in
crystal chains. These bacterial cells passively rotate
in a water environment in the direction of geomagnetic
field lines (Bazylinski and Frankel 2004; Faivre and
Schuler 2008). Magnetite structures were also
detected in the pigeon upper beak, and iron-containing
otoliths and intracellular ferritin corpuscles were
found in the inner ear of birds (Fleissner et al. 2003;
Hanzlik et al. 2000; Harada 2008; Lauwers et al.
2013).
In insects, the magnetic crystals were detected via
magnetic precipitation methods and magnetic reso-
nance in body parts of ants (Acosta-Avalos et al. 1999;
Wajnberg et al. 2005). In ant’s antennae, their main
sensory organ, ferro/ferri-magnetic crystals likely
composed of magnetite/maghemite, hematite, goethite
and others minerals were found (de Oliveira et al.
2010; Wajnberg et al. 2004). By means of magneti-
sation of honeybees, it was observed that their bodies
exhibit magnetisation (Gould et al. 1978). The study of
composition of honeybees’ magnetic structures was
addressed by Kuterbach and Walcott (Kuterbach and
Walcott 1986), who discovered that particles isolated
from the cells of honeybees’ abdomen contain iron.
Their subsequent Mossbauer spectroscopic analysis
showed that at least 95 % of the present iron is in the
form of hydrous iron oxide, not magnetite. The
ferromagnetic material was also detected using a
magnetic resonance method (El-Jaick et al. 2001). Hsu
and Li (1994) discovered that the abdomen cells
contain trophocytes rich in clusters of superparamag-
netic particles connected with their cell membrane,
thus reaching a hypothesis of the magnetosensoric
function of superparamagnetic trophocytes (Hsu et al.
2007). Subsequently they found that the alternative
external magnetic field induces size fluctuations of
superparamagnetic clusters. These size fluctuations
lead to the increase of the Ca2? concentration in cells,
which influences neural processes around cells. Gen-
erally, magnetic crystals may not in fact play
magnetoreception role in insects, though presence of
magnetite crystals in ant’s antennae makes sense (de
Oliveira et al. 2010). Currently, the main effort of
research on magnetoreception is focused on the
evaluation of the radical-pair/cryptochrome hypothe-
sis. This hypothesis is based on photochemical reac-
tions inside the honeybees’ eyes influenced by
geomagnetic field (Valkova and Vacha 2012).
In bumblebees (Bombus sp.), the iron biominerals
were already discovered in abdominal cells (Hsu
2004; Walcott 1985). According to electron micros-
copy, the granules are approximately 1 lm in size,
often with spherical shape and associated with the
membranes, and located namely under the cuticle on
the ventral side of the abdomen. Elemental analysis
revealed that these granules contain Fe, P and Ca.
From the point of view of magnetoreception, the
orientation of bumblebees in the environment is based
mainly on their optical and olfactory abilities, but their
good orientation abilities in a total darkness with
limited olfactory information available indicate that
bumblebees use a magnetic compass that is indepen-
dent of light (Chittka et al. 1999).
The aim of this study is to detect, by means of
magnetic measurements and several experimental
techniques, whether bodies of bumblebees contain
magnetic particles or aggregates, as a potential
component of their geomagnetic sensor.
Experimental
Bumblebee samples
Bumblebee species Bombus terrestris, and its individ-
ual body parts, were used for the study. The first group
of bumblebees was wild, caught specifically in a post-
mining landscape in Ostrava, Czech Republic. The
death was caused by inhalation of ethyl acetate.
Drying proceeded under normal conditions (at room
temperature and 40–60 % humidity). The second
group were bumblebees of a special breed from the
Research Institute for Fodder Crops (VUP) in Brno-
Troubsko, Czech Republic. Bumblebees were bred
there at 25 �C and 50–60 % humidity. They were fed
by sugar solution and pollen, which is collected from
honeybees. The age of the bred bumblebees for the
purpose of the measurements was about 3 months.
The three following casts were available: queens,
workers (female) and males in a number of 10 of each.
Their death was induced by freezing.
Biometals
123
Author's personal copy

Magnetic model substances
Fresh equine spleen ferritin served as a model
substance (at concentration of 53 ± 3 mg/mL of
saline solution, \0.9 % of NaCl, product number:
F4503) and superparamagnetic magnetite (at concen-
tration of 5.0 ± 0.3 mg/mL of toluene, product num-
ber: 700320) were provided by Sigma Aldrich. The
size of superparamagnetic magnetite crystals, having a
size range of 5–18 nm, was measured using the laser
analyser Malvern Zetasizer NanoZS (type ZEN3600).
Measurement techniques
To confirm the presence of magnetic granules, a
combination of four experimental methods was per-
formed—vibrating sample magnetometry (VSM),
scanning electron microscopy (SEM), Mossbauer
spectroscopy and X-ray powder diffraction (XRPD).
The magnetometry serves as an input method for
detection of magnetic particles in the body parts of
bumblebees (head, thorax, abdomen and rest body-
parts—limbs and wings). The data from magnetic
measurements were processed in Matlab 7.0. Physical
quantities and their units are presented in the SI unit
system.
Magnetic measurements were performed using a
vibrating magnetometer VSM EV9 Microsense (mag-
netic moment resolution 10-10 A m2, field resolution
10-4 T). Magnetic measurements were carried out at
normal as well as variable temperatures. Bumblebee
bodies were attached to the glass holder with paraffin.
This glass holder with paraffin exhibited a diamag-
netic signal of its own, therefore the holder was
measured separately and then the value of its diamag-
netic signal was subtracted from the total signal value
(holder, paraffin, and sample). The ferritin was
magnetised in liquid form in a cylindrical glass
container sealed with paraffin. Without ferritin, these
components exhibit a diamagnetic moment indepen-
dent of temperature. Superparamagnetic magnetite
was measured in dry form on diamagnetic glass of the
sample holder. Before the measurement, the holder
was soaked into the toluene-magnetite solution and
when the toluene evaporated, dry magnetite remained
on the glass surface. The temperature measurements
were performed using temperature regulation of N2
gas flow in the range of -165 to 0 �C, i.e. 108–273 K,
which is the limit for the VSM system that was used. A
very slow regime of 5 K/min was selected to ensure
the smallest difference possible between temperatures
of the sensor and samples. Uncertainties of tempera-
ture measurements are ±1 �C.
In order to determine morphology of the magnetic
structures and measure their elemental composition,
the electron microscopy SEM QUANTA 450 FEG by
FEI was used. The microscopic images were obtained
in back scattered electron (BSE) and scattered electron
(SE) regimes with subsequent EDX microanalysis of
revealed granules. For the purposes of this measure-
ment, bumblebees were coated (three times) with gold
and palladium and then they were placed on a carbon
tape and attached to a small brass target. The
microscopy was performed under magnification rang-
ing from 600 to 60009 while using a voltage of
15–20 kV.
The transmission Mossbauer spectrum was mea-
sured at room temperature using 57Co(Rh) source
moving in a constant acceleration mode. The velocity
scale was calibrated using a thin foil of a-Fe and the
isomer shift was taken with respect to its RT
Mossbauer spectrum. The measured spectrum was
evaluated within the transmission integral approach
using the program CONFIT (Zak and Jiraskova 2006).
Generally, the crystalline components can be repre-
sented by single-, double-, and/or six-line Lorentzian
subspectra determined by discrete values of hyperfine
parameters: isomer shift(s), quadrupole, and magnetic
splitting(s), corresponding to paramagnetic and/or
ferromagnetic phases.
The XRPD patterns were recorded under CoKairradiation (k = 1.789 A) using the Bruker D8
Advance diffractometer (Bruker AXS) equipped with
a fast position sensitive detector VANTEC 1. Mea-
surements were carried out in the reflection mode, and
the powder sample was placed on a rotational holder.
Phase composition was evaluated using database PDF
2 Release 2004 (International Centre for Diffraction
Data).
Results
Wild bumblebees
Magnetisation of wild bumblebees bodies exhibits
magnetic hysteresis reflecting the presence of ferro/
ferri-magnetic crystals (Fig. 1). Magnetic curves are
Biometals
123
Author's personal copy

influenced by the diamagnetic character of the tissue.
SEM revealed presence of various granules situated
directly on the surface of their bodies, especially on
their hairs, usually in the form of granules of
approximately the size of 1–10 lm. Examples of
EDX analysis results are presented in Fig. 2. The
presence of iron in the granules was clearly confirmed.
Detected silicates (containing mostly Si, Al, Ca, K)
probably come from dust particles and from plants
with which the wild bumblebees come into contact.
Less expected was the presence of Bi, Sb, Ni and Cr.
These elements are most likely associated with
environmental pollution (air pollution) of the area
where the wild bumblebees lived and may come e.g.
from pyrometallurgical plant or might be released by
road traffic, which can be inferred from some studies
(Kim et al. 2007; Kukutschova et al. 2011; Maher et al.
2008; Muxworthy et al. 2003; Rodriguez-Germade
et al. 2014).
Breed bumblebees
Magnetic and chemical characterisation
In this phase of the research, it was apparent that it was
necessary to work with a control group of bumblebees
that had not been exposed to much environmental
pollution. Magnetisation of these clean bodies from
breed bumblebees exhibits paramagnetic or diamag-
netic magnetisation (Figs. 3, 4, 5, 6, 7, 8) affected by
diamagnetic tissue. The same results were obtained for
all three groups (queens, workers and males). The
paramagnetic signal is stronger in the case of head and
wings than in the case of thorax and abdomen. SEM
observations using BSE and SE modes revealed, that
in subsurface layers in hair roots on wings (in front
edge of the upper wings) and on the head, iron-based
granules can be found (Figs. 4, 6). From the EDX
spectra (Figs. 4, 6), it can be assumed that these
granules may be based on compounds of Fe, Mg, Si, P,
Ca, O, K or Cr. The EDX measurement is in
accordance with the results of magnetic measurements
except for the fact that magnetic structures and their
signal are stronger in lighter weight parts (head, wing),
because they are not shielded by the diamagnetic
signal of the tissue. In the case of males the approx-
imate weight percentage ratio of caput/thorax/abdo-
men/rest of the body (wings and legs) is 11/29/62/\1.
Workers’ ratio is 22/51/28/\1, and queens’ ratio
10/42/47/\1.
Mineralogical composition of iron-based granules
To explain an importance of detected iron-based
granules we focused on those in wings since they have
a better volume ratio granules/tissue. It seems to be
better to use surface methods that allow for aiming the
subsurface granules and analyse them directly, as
provided by Raman microscopy, which enables a point
phase analysis of a single granule approximately down
to 500 nm in size. However, this method did not
provide us any results due to unsuccessful focusing of
granules and insufficient energy of the laser that has to
screen the cuticle. Based on such a failure, volume
methods for determination of mineralogical composi-
tion of iron-based granules in the wings had to be used.
Firstly, the magnetic method was selected for the
estimation of mineral composition of the discovered
granules. The aim was to evaluate whether thermo-
magnetic curves for superparamagnetic magnetite or
ferritin (equine spleen ferritin was used as a model
substance) have a similar progress as thermomagnetic
curves from the wings of bumblebees. Constant field
of 0.02 T was chosen as it represents an approximate
linear boundary of ferritin magnetisation (Brem et al.
2006), for magnetite it is 0.2 T (Jandacka et al. 2013),
where the measured data can then be fit to Curies law
(Kittel 2004). All three measurements are influenced
by a diamagnetic moment b (A m2) independent of the
Fig. 1 Magnetisation of selected body parts of wild bumble-
bees. Diamagnetic moment of the glass holder and paraffin
presented in figure is subtracted from total moments. A fit is
represented by equation m = -4.14 9 10-7 9 B -
7.91 9 10-9, R2 = 0.999. Residual diamagnetic moment from
tissue has an impact on the ferromagnetic moment
Biometals
123
Author's personal copy

Fig. 2 Examples of SEM images of particles (designated by white arrows) situated on surface hairs of wild bumblebees and their EDX
spectra. Samples were coated with Au and Pd and put on carbon tape fixed on a brass sample holder
Biometals
123
Author's personal copy

Fig. 3 Magnetisation of breed bumblebee—head. Diamagnetic
moment (glass holder and paraffin) is subtracted from total
magnetic moments. Residual diamagnetic moment from tissue
has an impact on the paramagnetic moment
Fig. 4 SEM images of bred bumblebee—head (queen) with
subsurface iron granule (white points in photo) displayed in
BSED and SE regimes. Through the combination of these
regimes, it is possible to distinguish Fe-containing granules
located in subsurface structures of hairs. EDX spectra of such
particles are presented. Samples were coated with Au and Pd
and put on carbon tape fixed on a brass sample holder
Fig. 5 Magnetisation of bred bumblebee—wing. Diamagnetic
moment is subtracted from total magnetic moments. Residual
diamagnetic moment from tissue has an impact on the
paramagnetic moment. Note worker means female
Biometals
123
Author's personal copy

Fig. 6 SEM images of subsurface Fe-containing granules on a
wing (white points in photo) in bred bumblebee sample (queen)
displayed in BSE and SE regimes. EDX spectra of such granules
are presented. Samples were coated with Au and Pd and put on
carbon tape fixed on a brass sample holder
Fig. 7 Magnetisation of bred bumblebee—thorax. Diamag-
netic moment of glass holder and paraffin is subtracted from
total magnetic moments. Note worker means female
Fig. 8 Magnetisation of bred bumblebee—abdomen. Diamag-
netic moment of glass holder and paraffin is subtracted from
total magnetic moments. Note worker means female
Biometals
123
Author's personal copy
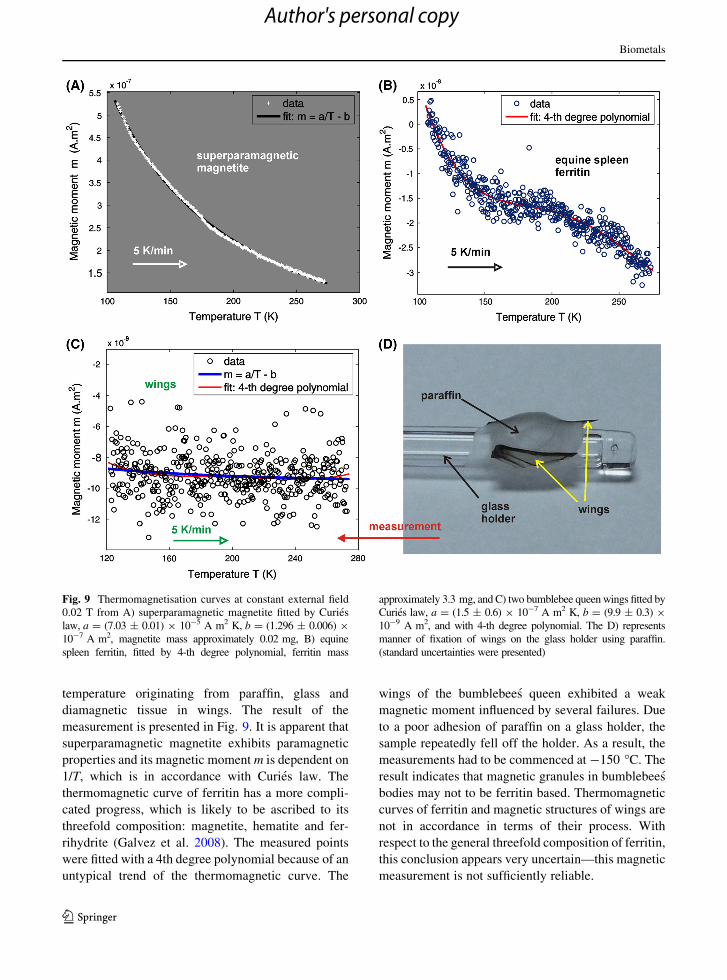
temperature originating from paraffin, glass and
diamagnetic tissue in wings. The result of the
measurement is presented in Fig. 9. It is apparent that
superparamagnetic magnetite exhibits paramagnetic
properties and its magnetic moment m is dependent on
1/T, which is in accordance with Curies law. The
thermomagnetic curve of ferritin has a more compli-
cated progress, which is likely to be ascribed to its
threefold composition: magnetite, hematite and fer-
rihydrite (Galvez et al. 2008). The measured points
were fitted with a 4th degree polynomial because of an
untypical trend of the thermomagnetic curve. The
wings of the bumblebees queen exhibited a weak
magnetic moment influenced by several failures. Due
to a poor adhesion of paraffin on a glass holder, the
sample repeatedly fell off the holder. As a result, the
measurements had to be commenced at -150 �C. The
result indicates that magnetic granules in bumblebees
bodies may not to be ferritin based. Thermomagnetic
curves of ferritin and magnetic structures of wings are
not in accordance in terms of their process. With
respect to the general threefold composition of ferritin,
this conclusion appears very uncertain—this magnetic
measurement is not sufficiently reliable.
Fig. 9 Thermomagnetisation curves at constant external field
0.02 T from A) superparamagnetic magnetite fitted by Curies
law, a = (7.03 ± 0.01) 9 10-5 A m2 K, b = (1.296 ± 0.006) 9
10-7 A m2, magnetite mass approximately 0.02 mg, B) equine
spleen ferritin, fitted by 4-th degree polynomial, ferritin mass
approximately 3.3 mg, and C) two bumblebee queen wings fitted by
Curies law, a = (1.5 ± 0.6) 9 10-7 A m2 K, b = (9.9 ± 0.3) 9
10-9 A m2, and with 4-th degree polynomial. The D) represents
manner of fixation of wings on the glass holder using paraffin.
(standard uncertainties were presented)
Biometals
123
Author's personal copy

57Fe Mossbauer spectrometry is highly sensitive to
the short-range interaction in iron-containing solids
and it is also being frequently used in investigation of
various iron-based oxides and/or iron-based compos-
ites down to nano-scale objects. The bumblebee
sample for the Mossbauer measurement was prepared
from approximately six main bumblebee wings in
such a way that the upper parts of the wings have filled
the measured area as much as possible (see inset in
Fig. 10). The Mossbauer measurement has resulted in
an asymmetric doublet seen in Fig. 10. It represents
iron-based species yielding either superparamagnetic
behaviour of the nanometre size particles or paramag-
netic objects—the paramagnetic behaviour is consis-
tent with the VSM measurements. The spectrum can
be analyzed either by a broadened double-line spec-
trum, the lines of which have various intensities or by
two double-line subcomponents as depicted in Fig. 10,
both characterized again by certain asymmetry. The57Fe Mossbauer hyperfine parameters for both indi-
vidual doublets are summarized in Table 1. They do
not reflect the parameters, isomer shift and quadrupole
splitting, usually presented for Fe2? and Fe3? ions in
the iron oxides. Therefore, the phase composition(s) of
the measured iron-based object(s) in the wings, similar
to the asymmetry of the Mossbauer spectrum, are
difficult to clarify conclusively from the present
measurement. The Mossbauer measurements in the
applied magnetic field and/or low temperatures could
be helpful in future investigations.
XRPD measurement was performed on 6 pairs of
dry wings in a powder form, prepared via drying in
34 �C for 2 h and via pulverising of them in agate
mortar. This method detected three mineral phases in
the wings, see Fig. 11. The first is crystalline silicon
oxide, SiO2, and the others are magnetite, Fe3O4, and
wuestite, FeO. All of them probably create the
granules.
It is apparent that only partial success was reached
in mineralogical analysis of our samples. XRPD
method revealed that iron-based granules are com-
posed of magnetite and wuestite crystals, however
Mossbauer spectroscopy did not confirm such results
clearly, since content of Fe2? and Fe3? ions, typical
for magnetite and wuestite, is not obvious. This
ambiguity may be induced by sensitivity of Mossbauer
spectroscopy to measurement environment of iron
oxides, which is tissue of wings in this case. On the
other hand, the presence of magnetite/wuestite in
superparamagnetic forms is not excluded. We have to
admit, that an uncertainty of measurement was entered
via different form of samples (XRPD = dry powder,
Mossbauer spectroscopy = fresh wings), since the
drying and milling processes on wings may cause
recrystallisation of the granular crystals. Therefore,
the mineralogical analysis is not final and needs to be
continued in future.
Fig. 10 Mossbauer spectrum of the bumblebees wings com-
posite into a sample (inset) 10 mm in diameter
Fig. 11 XRPD spectrum of bumblebee wings fragmentised to
powder form after drying
Table 1 57Mossbauer hyperfine parameters; isomer shift, d,
quadrupole splitting, D, asymmetry, D, and areas of sub-
spectra, A
d (mm/s) D (mm/s) D (-) A (%)
Doublet 1 (grey) 0.113 (07) 0.605 (23) 1.52 77.5
Doublet 2 (pattern) 0.090 (30) 0.215 (68) 1.44 22.5
Biometals
123
Author's personal copy

Discussion: function of the iron-based granules
To the best of our knowledge, the presence of
magnetic iron-based granules in bumblebee wings
and head was not yet observed. The results achieved
within this study are different to similar studies on
honeybees, mentioned in introduction section and it is
not consistent with the study of Walcott (1985) on
bumblebees, who revealed iron-rich granules contain-
ing P and Ca in the abdomen cuticle. We did not detect
such particles in this bodypart by means of magnetic
and other techniques.
Very recently, similar discovery was done by
Lauwers et al. (2013). They detected iron-rich gran-
ules in the inner avian ears, in cuticular plates of
receptor hair cells. This finding resembles our discov-
ery of iron-based granules in the roots of surface hairs
of bumblebees. The authors had a difficulty determin-
ing the mineral composition of the iron-rich granules,
but by means of electron diffraction, they concluded
that granules are probably composed of ferritin
molecules. The similarity between their and our
results shows that iron may be significant element
for normal evolution of surface hairs, thus the iron-
based granules in bumblebees may not be connected
with magnetosensation.
In spite of the fact that presence of strongly
magnetic magnetite in bumblebee wings was not
definitely evidenced, one can consider a sense of
magnetic granules in oscillating wings, in front edge
of the upper wings, close to head. These granules are
magnetised in a geomagnetic field and perform a
periodic motion with a frequency of approximately
220 Hz when a bumblebee flies. This motion gener-
ates an electromagnetic induction and this effect might
provide them magnetic compass information via a
head sensor (see Fig. 12). This hypothesis could be
evaluated by the behavioural experiment, when the
granules would be somehow removed from the wings.
Conclusions
In the surface tissue of the studied bumblebees, in the
roots of surface hairs, on the wings and on the head,
iron-based granules exhibiting paramagnetic magnet-
isation are located. The presence of these granules was
detected with the help of magnetometry and scanning
electron microscopy. XRPD analysis shows that
granules contain iron minerals magnetite and wuestite,
likely in superparamagnetic form according to Moss-
bauer spectroscopy. In this phase, the exact function of
these structures is very difficult to be determined, but it
can be stated that there are three options available:
directional magnetoreception (magnetic compass),
metabolic reservoir of iron for a proper function of
surface hairs and metabolic waste deposition. The
considered magnetoreception functioning of granules
located in the wings could be based on electromag-
netic induction generated by periodic motion of wings.
In further analyses, the mineralogical measure-
ments have to be done in samples from tens/hundred of
freshly killed bumblebee bodies where iron-based
granules will be present in sufficient amount, for better
reliability of analyses. For future, we do not recom-
mend to prepare the samples via drying and separation
processes, since in this case the validity of results is
threatened.
Acknowledgments This paper was elaborated in the framework
of the projects ‘‘IT4 Innovations Centre of Excellence’’, Reg. No.
CZ.1.05/1.1.00/02.0070, and Nanotechnology—the basis for
Fig. 12 Presentation of
hypothetic
magnetoreception principle
based on electromagnetic
induction activated during
bumblebee flight
Biometals
123
Author's personal copy

international cooperation project, Reg. No. CZ.1.07/2.3.00/20.0074
financed by the Structural Funds of EU and from the state budget of
the Czech Republic, the project Detection and characterization
ultrafine particles in living systems and their influence on
environment provided within Students Grant competition, Reg.
no. SP 2014/76, and finally Grant P108/11/1350.
References
Acosta-Avalos D, Wajnberg E, Oliveira PS, Leal I, Farina M,
Esquivel DM (1999) Isolation of magnetic nanoparticles from
Pachycondyla marginata ants. J Exp Biol 202:2687–2692
Bazylinski DA, Frankel RB (2004) Magnetosome formation in
prokaryotes. Nat Rev Microbiol 2:217–230. doi:10.1038/
Nrmicro842
Brem F, Stamm G, Hirt AM (2006) Modeling the magnetic
behavior of horse spleen ferritin with a two-phase core
structure. J Appl Phys 99. doi:10.1063/1.2206101
Chittka L, Williams NM, Rasmussen H, Thomson JD (1999)
Navigation without vision: bumblebee orientation in
complete darkness. Proc R Soc Lond B Biol Sci 266:45–50
de Oliveira JF, Wajnberg E, de Souza Esquivel DM, Weinkauf
S, Winklhofer M, Hanzlik M (2010) Ant antennae: are they
sites for magnetoreception? J R Soc Interface 7:143–152.
doi:10.1098/rsif.2009.0102
El-Jaick LJ, Acosta-Avalos D, De Souza DM, Wajnberg E,
Linhares MP (2001) Electron paramagnetic resonance
study of honeybee Apis mellifera abdomens. Eur Biophys J
29:579–586
Faivre D, Schuler D (2008) Magnetotactic bacteria and magneto-
somes. Chem Rev 108:4875–4898. doi:10.1021/Cr078258w
Fleissner G, Holtkamp-Rotzler E, Hanzlik M, Winklhofer M,
Fleissner G, Petersen N, Wiltschko W (2003) Ultrastruc-
tural analysis of a putative magnetoreceptor in the beak of
homing pigeons. J Comp Neurol 458:350–360
Galvez N et al (2008) Comparative structural and chemical
studies of ferritin cores with gradual removal of their iron
contents. J Am Chem Soc 130:8062–8068. doi:10.1021/
Ja800492z
Gould JL, Kirschvink JL, Deffeyes KS (1978) Bees have mag-
netic remanence. Science 201:1026–1028. doi:10.1126/
science.201.4360.1026
Hanzlik M, Heunemann C, Holtkamp-Rotzler E, Winklhofer M,
Petersen N, Fleissner G (2000) Superparamagnetic mag-
netite in the upper beak tissue of homing pigeons. Bio-
metals 13:325–331
Harada Y (2008) The relation between the migration function of
birds and fishes and their lagenal function. Acta Oto-Lar-
yngol 128:432–439. doi:10.1080/00016480701724920
Hsu CY (2004) The processes of iron deposition in the common
hornet (vespa affinis). Biol Cell 96:529–537. doi:10.1016/j.
biolcel.2004.05.001
Hsu CY, Chan YP (2011) Identification and localization of
proteins associated with biomineralization in the iron
deposition vesicles of honeybees (Apis mellifera). PLoS
One 6. doi:10.1371/journal.pone.0019088
Hsu CY, Ko FY, Li CW, Fann K, Lue JT (2007) Magnetore-
ception System in Honeybees (Apis mellifera). PLoS One.
doi:10.1371/journal.pone.0000395
Hsu CY, Li CW (1994) Magnetoreception in honeybees. Sci-
ence 265:95–97. doi:10.1126/science.265.5168.95
Jandacka P, Alexa P, Pistora J, and Trojkova J (2013) Hypo-
thetical superparamagnetic magnetometer in a pigeon’s
upper beak probably does not work. Eur Phys J E Soft
Matter 36. doi:10.1140/Epje/I2013-13040-1
Kim W, Doh SJ, Park YH, Yun ST (2007) Two-year magnetic
monitoring in conjunction with geochemical and electron
microscopic data of roadside dust in Seoul Korea. Atmos
Environ 41:7627–7641. doi:10.1016/j.atmosenv.2007.05.
050
Kittel C (2004) Introduction to solid state physics. Wiley, New
York
Kukutschova J et al (2011) On airborne nano/micro-sized wear
particles released from low-metallic automotive brakes.
Environ Pollut 159:998–1006. doi:10.1016/j.envpol.2010.
11.036
Kuterbach DA, Walcott B (1986) Iron containing cells in the
honeybee (Apis mellifera). 1. Adult Morphology and
Physiology. J Exp Biol 126:375–387
Lauwers M et al (2013) An iron-rich organelle in the cuticular
plate of avian hair cells. Curr Biol 23:924–929. doi:10.
1016/j.cub.2013.04.025
Maher BA, Moore C, Matzka J (2008) Spatial variation in
vehicle-derived metal pollution identified by magnetic and
elemental analysis of roadside tree leaves. Atmos Environ
42:364–373. doi:10.1016/j.atmosenv.2007.09.013
Muxworthy AR, Matzka M, Davila AF, Petersen N (2003)
Magnetic signature of daily sampled urban atmospheric
particles. Atmos Environ 37:4163–4169. doi:10.1016/
S1352-2310(03)00500-4
Rodriguez-Germade I, Mohamed KJ, Rey D, Rubio B, Garcia A
(2014) The influence of weather and climate on the reli-
ability of magnetic properties of tree leaves as proxies for
air pollution monitoring. Sci Total Environ 468:892–902.
doi:10.1016/j.scitotenv.2013.09.009
Safarik I, Safarikova M (2002) Magnetic nanoparticles and
biosciences. Monatsh Chem 133:737–759. doi:10.1007/
s007060200047
Valkova T, Vacha M (2012) How do honeybees use their
magnetic compass? Can they see the North? Bull Entomol
Res 102:461–467. doi:10.1017/S0007485311000824
Wajnberg E, Alves OC, Harada AY, de Esquivel DMS (2005)
Brazilian ants diversity and the local geomagnetic field: a
ferromagnetic resonance study. Biometals 18:595–602.
doi:10.1007/s10534-005-2995-4
Wajnberg E, Cernicchiaro G, de Souza Esquivel DM (2004)
Antennae: the strongest magnetic part of the migratory ant.
Biometals 17:467–470. doi:10.1023/B:Biom.0000029443.
93732.62
Walcott B (1985) The cellular localization of particulate iron.
In: Kirschvink JL, Jones DS, MacFadden BJ (eds) Mag-
netite biomineralization and magnetoreception in organ-
isms: a new biomagnetism. Plenum Press, New York,
pp 417–438
Zak T, Jiraskova Y (2006) CONFIT: mossbauer spectra fitting
program. Surf Interface Anal 38:710–714. doi:10.1002/
Sia.2285
Biometals
123
Author's personal copy




















