
Surgical treatment for secundum atrial septal defects in patients >40 years old
Upload
independentCategory
view
0download
0
Exp Brain Res (1992) 88:78-90
Experimental Brain Research © Springer-Verlag 1992
Septal cholinergic afferents regulate expression of brain-derived neurotrophic factor and t-nerve growth factor mRNA in rat hippocampus Nils Lindefors 1, Patrik Ernfors 2, Torkel Falkenberg 1, and H~kan Persson 2
1 Department of Pharmacology and z Medical Chemistry, Laboratory of Molecular Neurobiology, Karolinska Institutet, P.O. Box 60 400, S-104 01 Stockholm, Sweden
Received May 31, 1991 / Accepted August 28, 1991
Summary. In situ hybridization was used to study the expression of members of the nerve growth factor family of trophic factors in rat hippocampus following stimula- tion of afferent cholinergic and glutamatergic pathways with quisqualate. A transient increase in brain-derived neurotrophic factor (BDNF) and t-nerve growth factor (NGF) mRNA expression in the hippocampus was seen 4 h after a quisqualate injection into the medial septal nucleus. Both BDNF and NGF mRNA levels increased more than 4-fold in the granule layer of the dentate gyrus and for BDNF mRNA also in the pyramidal cells of CA1, while the levels of BDNF mRNA in CA3 increased 2-fold. The increase in BDNF and NGF mRNA levels were completely prevented by pretreatment with systemic injections of either scopolamine or diazepam. A quis- qualate injection into the entorhinal cortex, containing glutamatergic afferents to the hippocampus, resulted in a 15-, 5- and 17-fold increase in the expression of BDNF mRNA in the ipsilateral granule cells, CA3 and CA1 pyramidal cells, respectively. Similar increases were also seen in the hippocampus contralateral to the injections. In contrast, the level of NGF mRNA did not increase significantly in any of the subfields in the hippocampus. The increase in BDNF mRNA after cortex injections was attenuated by diazepam but not by scopolamine. These findings imply that increased activity in afferent choliner- gic and glutamatergic pathways to the hippocampus dif- ferentially regulate expression of the NGF family of neurotrophic factors in the hippocampus.
Key words: BDNF - NGF - Cholinergic - Hippocampus - In situ hybridization - mRNA
Introduction
The most extensive cholinergic pathways in the rat brain arise from cholinergic cell bodies in the basal forebrain
Offprint requests to: N. Lindefors
(McGeer et al. 1984). The rostromedial cells (medial septum and vertical limb of the diagonal band) are mid- line nuclei and project primarily to both sides of the hippocampus while more caudal cells (horizontal limb of the diagonal band and nucleus basalis) divide into a separate cell group for each hemisphere and project mostly to the ispilateral amygdala and to ipsilateral re- gions of the neocortex. Much attention has recently been paid to the basal forebrain cholinergic system since these neurons undergo a profound degeneration in senile de- mentia of the Alzheimer type that results in cholinergic deficiency in the target area of these neurons (Bartus et al. 1982; Whitehouse et al. 1982; Collerton 1986). More- over, a strong correlation between the reduction in cho- linergic marker activity and severity of the dementia exists (Perry et al. 1978; Wilcock et al. 1982). Damage to the hippocampal formation produces an amnesic syn- drome both in man (Milner et al. 1968), monkey (Mish- kin 1982), and rat (O'Keefe and Nadel 1978; Olton et al. 1979) and from these and other studies the hippocampus has been strongly implicated in learning and memory. Transection of the fimbria-fornix, containing the cholin- ergic afferents from the medial septum to the hippocam- pus results in behavioural deficits similar to those seen after removal of the whole hippocampus (O'Keefe and Nadel 1978; Olton et al. 1979) indicating that the medial septum chotinergic afferents are of major importance for the function of the hippocampal neuronal network.
Considerable support has been obtained for the hypothesis that neuronal survival is dependent on, and regulated by, neurotrophic factors synthesized in limited amounts in the target-field of sensitive neurons (Barde 1989). Most data supporting this concept has come from extensive studies on t-nerve growth factor (NGF), which in the peripheral nervous system supports the develop- ment and maintenance of sympathetic and neural crest- derived sensory neurons (Levi-Montalchini and Angelet- ti 1968 ; Thoenen and Barde 1980). More recently, NGF has also been shown to be present in the brain (Kor- sching et al. 1985; Whittemore et al. 1986; Shelton and Reichardt 1986; Goedert et al. 1986), where NGF sup-

79
ports basal forebrain cholinergic neurons. Thus, these neurons have NGF receptors (Richardson et al. 1986; Taniuchi et al. 1986) and retrogradely transport NGF (Schwab et al. 1979; Seiter and Schwab 1984). Moreover, the levels of both NGF mRNA and protein correlate with the density of cholinergic innervation in the adult rat brain (Korsching et al. 1985; Whittemore et at. 1986; Shelton and Reichardt 1986; Goedert et al. 1986; Ayer- LeLievre et al. 1988) and basal forebrain cholinergic neu- rons can be prevented from degenerating after transec- tion of the septo-hippocampal pathway by either chronic infusion of exogenous NGF (Hefti et al. 1986; Williams et al. 1986; Kromer 1987) or implantation of genetically modified cells producing recombinant NGF (Rosenberg et al. 1988). Furthermore, implantation of recombinant NGF-producing cells into acetylcholine denervated cere- bral cortex results in a marked increase in the survival of, and fiber outgrowth from, grafts of fetal basal forebrain cholinergic neurons (Ernfors et al. 1989).
Indirect evidence for the presence of other neuro- trophic factors in the brain has been obtained from several reports that demonstrate an enhanced survival of neurons in vitro using various tissue extracts (Tomozowa and Appel 1986) or co-cultures between different neuro- nal populations (Prochiantz et al. 1979). This was direct- ly demonstrated by the purification and biochemical characterization of brain-derived neurotrophic factor (BDNF) from pig brain (Barde et al. 1982). The subse- quent cloning of the BDNF gene (Leibrock et al. 1989) revealed a strong amino acid similarity to NGF and suggested the possible existence of a gene family of neu- rotrophic factors. In support of this, a third structurally related factor, hippocampus derived neurotrophic factor (HDNF; Ernfors et al. 1990) also known as neurotroph- in-3 (NT-3; Hohn et al. 1990; Maisonpierre et al. 1990; Rosenthal et al. 1990) and NGF-2 (Kaisko et al. 1990) has recently been described. In the brain, mRNAs for all three proteins have been shown to be expressed in neu- rons (Ayer-Lelievre et al. 1988; Gall and Isackson 1989; Ernfors et al. 1990a, b; Hofer et al. 1990; Wetmore et al. 1990; Philips et al. t990) although the distribution of cells expressing the different factors in the adult rat brain is unique for each. Interestingly, however, the highest levels of mRNA for each factor in the brain is found in the hippocampus where NGF is expressed primarily in neurons scattered throughout the pyramidal cell layer and stratum oriens as well as in the hilar region of the dentate gyrus (Gall and Isackson 1989; Ernfors et al. 1990b). BDNF is most highly expressed in pyramidal neurons in the CA3 and CA2 regions of the hippocampus as well as in granule neurons of the dentate gyrus and neurons in the hilus, with lower levels in CA1 (Ernfors et al. 1990a, 1990b; Hofer et at. 1990; Wetmore et al. 1990). In contrast, NT-3 /HDNF (here called NT-3 for simplicity) mRNA expressing neurons in the adult rat hippocampus are confined to the medial part of CA1, and to CA2 and the granule layer of the dentate gyrus (Ernfors et al. 1990a, b; Philips et al. 1990). The distribu- tion of BDNF and NT-3 mRNA-expressing neurons within the hippocampus mostly persist during postnatal brain development (Friedman et al. 1991a, 1991b) sug-
gesting that the two factors are not only important to maintain a proper neuronal innervation in the hippocam- pus but also play a role in the formation of synaptic contacts and neuronal maturation during brain develop- ment. The neurons that require BDNF or NT-3 for their development and survival remain to be identified, though BDNF has been shown to influence retinal ganglion cells (Johnson et al. t986; Hofer and Barde 1988) and, in similarity to NGF, basal forebrain cholinergic neurons (Alderson et al. 1990).
In addition to the septo-hippocampal system, other neuronal pathways innervate the hippocampus. One of the major afferent systems, the cortico-hippocampal af- ferent system, uses glutamate as transmitter. The amino acid glutamate is used as an excitatory neurotransmitter in most parts of the central nervous system (Fonnum 1984) and the effects of glutamate are mediated by at least three types of receptors, the N-methyl-D-aspartate, NMDA; the kainate and the quisqualate or (z-amino-3- hydroxy-5-methylisoxazole-4-propionic acid, AMPA re- ceptor (Monaghan et al. 1989). The majority of the corti- cal afferents to the hippocampus run in the perforant path from the entorhinal cortex, bilaterally to the dentate gyrus and to the pyramidal layer (Swanson et al. 1987) and this pathway is glutamatergic (White et al. 1977). Much simplified, postsynaptic to these cortical afferents are a sequence of intrahippocampal neurons containing granule cell mossy fibers from the dentate gyrus to CA3 and postsynaptic to the mossy fibers are pyramidal cell Schaffer collaterals from CA3 to ipsi- and contralateral CA1. Both of these pathways appear to use glutamate as their major neurotransmitter (Swanson et al. 1987). It is also known that non-NMDA receptor stimulation is followed by increased expression of BDNF and NGF mRNA in neurons of the hippocampus (Zafra et al. 1990). It was thus considered of interest to examine the possibility of transsynaptic activation of NGF or BDNF mRNAs following the activation of neuronal afferents to the hippocampus in vivo.
In this study we have activated septo-hippocampal and cortico-hippocampal neurons by the injection of the glutamate analog quisqualate into the medial septal nu- cleus and the entorhinal cortex, respectively. The ex- pression of the NGF family of neurotrophic factors was then analysed in the hippocampus by quantitative in situ hybridization. The results indicate that expression of both BDNF and NGF mRNA, but not NT-3 mRNA, in the hippocampus are regulated by presynaptic activa- tion of cholinergic neurotransmission. Evidence was also obtained that activation of glutamatergic afferents from the entorhinal cortex strongly activates hippocampal BDNF mRNA expression, whereas the expression of NGF mRNA in the hippocampus was only moderately affected by this treatment and NT-3 mRNA expression appeared to be unaffected.

80
Material and methods
Animal treatment and tissue preparation
Male Sprague-Dawley rats (b.w. 270 g) were anaesthetized with halothane and mounted in a stereotaxic frame. The scull was ex- posed and holes were drilled according to the position of the injec- tion cannula. The cannula consisted of a 1.0 gl Hamilton syringe held by the micromanipulator of the stereotaxic instrument.
Animals were divided into two major groups, one group re- ceived injections into the medial septal nucleus and the second group into the entorhinal cortex. For injections into the medial septal nucleus, the syringe was inserted at an angle of 10 ° relative to the vertical and implanted 0.7 mm rostral and 5.6 mm ventral to Bregma in the midline, as measured from the surface of the scull. These rats were subdivided into four subgroups. Group I (n = 12) was injected with phosphate buffered saline (PBS), group II (n = 12) was injected with 0.5 gl of 0.025 M quisqualate and group III (n = 4) was injected with quisqualate after pretreatment with scopolamine (25 mg/kg s.c. 1 h before the intracerebral injection and an addition- al injection directly following the intracerebral injection). Group IV (n=4) was injected with quisquatate after pretreatment with diazepam (1.5 mg/kg i.p. 1 h before intracerebral injection with an additional injection directly following the intracerebral injection). The animals in groups I and II were decapitated 1, 4 or 20 h after the intracerebral injections and the animals in groups III and IV were decapitated 4 h after the injections.
For entorhinal cortex injections, the syringe was inserted verti- cally and implanted 7.8 mm caudal, 5.5 mm lateral (left side) and 7.8 mm ventral to Bregma. These rats were similarly divided into four subgroups, group V (n = 4) was injected with PBS, group VI (n=4) with 0.5 gl of 0.025 M quisqualate acid, group VII (n=4) with quisqualate acid after scopolamine pretreatment group VIII (n = 4) with quisqualate acid after diazepam pretreatment. Other animals received the systemic scopolamine (n=4) or diazepam injections only (n = 4), to determine the effects of these drugs alone, and additional animals received intracerebral quisqualate injections into the caudate-putamen (n=4) to determine anatomically un- specific effects due to drug diffusion. The animals in groups V to VIII and animals receiving injections into the caudate-putamen were decapitated 4 h after the imracerebral injections, while animals receiving systemic injections only were decapitated 5 h after the injections.
Tissue sections of hippocampus (16 grn) were cut using a cryos- tate (Leitz, Wetzlar, West Germany) and thaw-mounted onto poly- L-lysine coated slides (50 gg/ml). The location of the injection cannula was verified during cryostat cutting.
In situ hybridization
After cryostat sectioning, the tissues were fixed in 10% formalin in phosphate buffered saline for 30 min, rinsed twice for 4 rain in PBS, and delipidated in a graded series of ethanol, including a 5 rain incubation with chloroform. The sections were then airdried. The hybridization cocktail contained 50% formamide, 4 x SSC (saline sodium citrate buffer; 1 x SSC is 0.15 M NaC1, 0.015 M sodium citrate pH 7,0), 1 x Dendhardts solution, 1% Sarcosyl, 0.02M Na3PO4 (pH 7,0) 10% Dextran sulphate, 0.3 gg/ml yeast tRNA, 0.06 M dithiothrietol (DTT) and 0.1 mg/ml sheared salmon sperm DNA. For BDNF mRNA hybridization a 50-mer oligonucleotide complementary to nucleotides 650-699 of rat BDNF mRNA (Hofer et al. 1990) was used. For NGF mRNA hybridization a 50-mer oligonucleotide complementary to nucleotides 869 to 918 of rat NGF mRNA (Whittemore et al. 1988) was used. For NT-3 mRNA hybridization a 50-met oligonucleotide complmnentary to nucleo- tides 667 to 716 of rat NT-3 mRNA (Ernfors et al. 1990b) was used. The oligonucleotides were 3'-end labeled with u-35S-dATP using terminal deoxyribonucleotidyt transferase (International Biotech.
Inc., New Haven, Connecticut) to a specific activity of 1 x 109 cpm/gg. The labeled probes were purified on a Nensorb column (DuPont, Wilmington, DE) prior to use. Following hybridization for 16 h at 42 ° C, the sections were rinsed 5 times for 15 min in 1 x SSC at 55 ° C and rinsed in autoclaved water for 5 min, and then dehydrated through a series of graded alcohol and airdried. The sections were exposed to X-ray film (Amersham 13-max) for 14 days and to photographic emulsion (Kodak NTB3) for 40 days. After having been developed, the slides were stained with cresyl violet followed by microscopic analysis. Several criteria for the specificity of the hybridizations were used, including hybridization with un- related oligonucleotide probes, sense probes as negative controls, as well as with cDNA probes for BDNF and NGF mRNAs showing a similar hybridization pattern as the oligonucleotide probes used (Ernfors et al. 1990a).
The autoradiograms from the in situ hybridizations were analyzed with a Microcomputer Imaging Device (Imaging Research Inc., Canada). Three tissue sections from each brain were used for measurements and optical density in each anatomical region was determined in triplicate in each section. Optical density values were determined and converted to mRNA levels using external standards exposed to the same film. Optical density values were measured in an interval with linear correlation to tissue levels of radioactive isotope.
Statistics
Data are presented as mean~S.E.M. Statistical analysis of data from from in situ experiments using ANOVA with Scheffe's post- comparative F-test.
Results
Experimental rational controls for hybridization specificity and behaviourat effects of quisqualate injections
In the r a t bra in , the h ighest levels o f the three k n o w n me mbe r s o f the N G F fami ly o f t roph ic fac tors ( N G F , B D N F , N T - 3 ) a re f o u n d wi th in the h i p p o c a m p u s (Ern- fors et al. 1990a, b ; H o h n et al. 1990; M a i s o n p i e r r e et al., 1990; Phi l l ips et al. 1990). Septa l chol inerg ic neurons innerva te the h i p p o c a m p u s a n d these neu rons are k n o w n to depend u p o n N G F for their survival b o t h in vi t ro (Heft i et al. 1984; M a r t i n e z et al. 1987) a n d in vivo (Hef t i 1986; Wi l l i ams et al. 1986; K r o m e r 1987; Ern fo r s et al. 1989). Recent ly , septal chol inerg ic neurons have also been shown to r e s p o n d in v i t ro to B D N F (Atde r son et al. 1990). In this r e p o r t we have examined i f an enhanced n e u r o n a l ac t iv i ty in septa l chol inerg ic neu rons affects express ion o f m R N A s for the th ree n e u r o t r o p h i c fac tors o f the N G F fami ly in the h i p p o c a m p u s . To answer this ques t ion , septa l chol inerg ic ne u rons were ac t iva t ed by in jec t ion o f the g l u t a m a t e a n a l o g qu i squa la t e in to the media l septa l nucleus and the levels o f the n e u r o t r o p h i c fac to r m R N A s in the h i p p o c a m p u s were then ana lyzed at different t imes af ter the in jec t ion by in si tu hybr id i za - t ion. In the do r sa l h i p p o c a m p u s o f n o n - o p e r a t e d , con t ro l an imals , B D N F m R N A label ing was seen in g ranu le cells o f the den ta t e gyrus , in h i la r cells, in the p y r a m i d a l cells o f C A 2 and CA3 a n d to a lesser ex tent a lso in C A t . Dens i t ome t r i c ana lys is o f X - r a y fi lms revealed a 4-fold
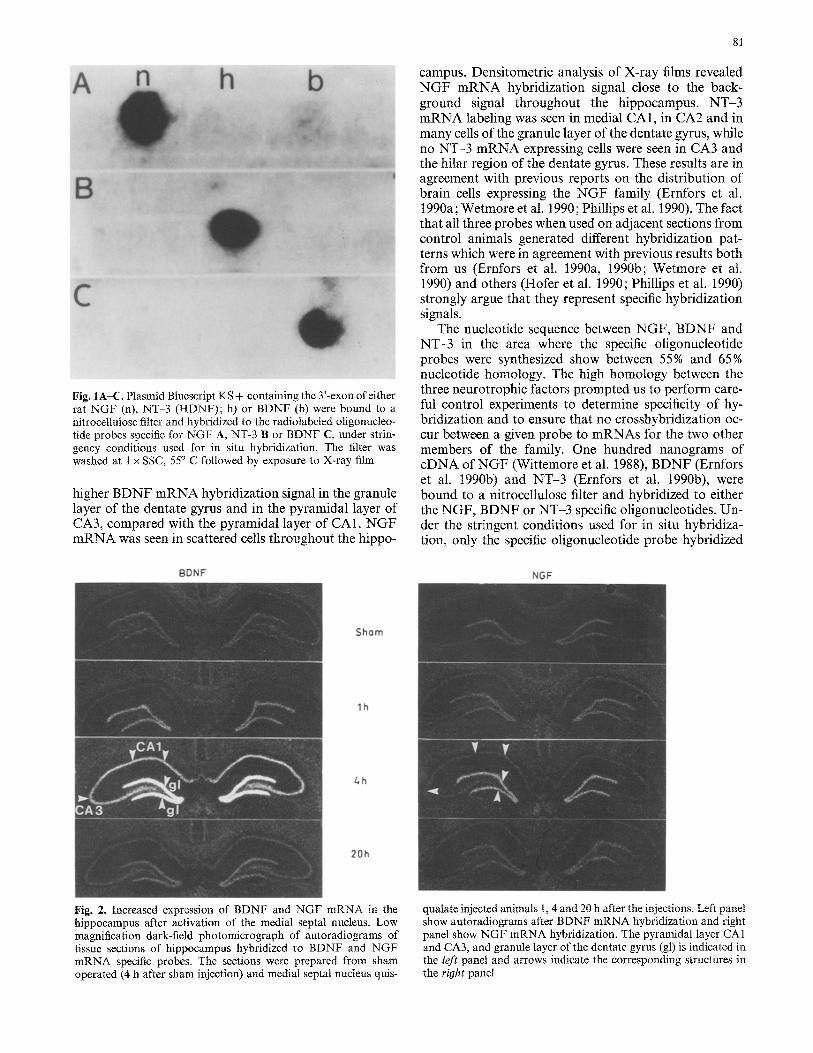
Fig. 1A-C. Plasmid Bluescript KS + containing the 3'-exon of either rat NGF (n), NT-3 (HDNF); h) or BDNF (b) were bound to a nitrocellulose filter and hybridized to the radiolabeled oligonucleo- tide probes specific for NGF A, NT-3 B or BDNF C, under strin- gency conditions used for in situ hybridization. The filter was washed at 1 x SSC, 55 ° C followed by exposure to X-ray film
higher B D N F m R N A hybridization signal in the granule layer of the dentate gyrus and in the pyramidal layer of CA3, compared with the pyramidal layer of CA1. N G F m R N A was seen in scattered cells throughout the hippo-
BDNF
81
campus. Densitometric analysis of X-ray films revealed N G F m R N A hybridization signal close to the back- ground signal throughout the hippocampus. N T - 3 m R N A labeling was seen in medial CA1, in CA2 and in many cells of the granule layer of the dentate gyrus, while no NT-3 m R N A expressing cells were seen in CA3 and the hilar region of the dentate gyrus. These results are in agreement with previous reports on the distribution of brain cells expressing the N G F family (Ernfors et al. t990a; Wetmore et al. 1990; Phillips et al. 1990). The fact that all three probes when used on adjacent sections from control animals generated different hybridization pat- terns which were in agreement with previous results both from us (Ernfors et al. 1990a, 1990b; Wetmore et at. 1990) and others (Hofer et al. 1990; Phillips et al. 1990) strongly argue that they represent specific hybridization signals.
The nucleotide sequence between NGF, B D N F and NT-3 in the area where the specific oligonucleotide probes were synthesized show between 55% and 65% nucleotide homology. The high homology between the three neurotrophic factors prompted us to perform care- ful control experiments to determine specificity of hy- bridization and to ensure that no crosshybridization oc- cur between a given probe to mRNAs for the two other members of the family. One hundred nanograms of cDNA of N G F (Wittemore et al. 1988), BDNF (Ernfors et al. 1990b) and N T - 3 (Ernfors et al. t990b), were bound to a nitrocellulose filter and hybridized to either the N G F , BDNF or NT-3 specific oligonucleotides. Un- der the stringent conditions used for in situ hybridiza- tion, only the specific oligonucleotide probe hybridized
NGF
Sham
lh
4h
20h
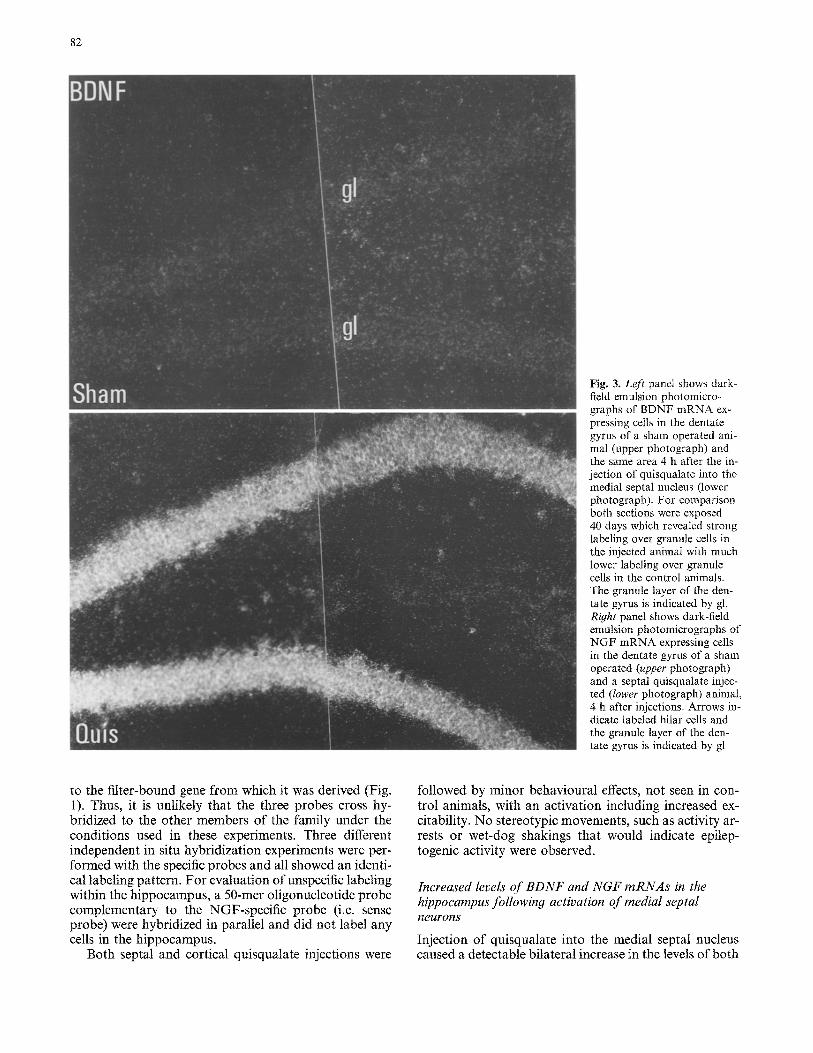
Fig. 2. Increased expression of BDNF and NGF mRNA in the hippocampus after activation of the medial septat nucleus. Low magnification dark-field photomicrograph of autoradiograms of tissue sections of hippocampus hybridized to BDNF and NGF mRNA specific probes. The sections were prepared from sham operated (4 h after sham injection) and medial septal nucleus quis-
qualate injected animals 1, 4 and 20 h after the injections. Left panel show autoradiograms after BDNF mRNA hybridization and right panel show NGF mRNA hybridization. The pyramidal layer CA1 and CA3, and granule layer of the dentate gyrus (gl) is indicated in the left panel and arrows indicate the corresponding structures in the right panel
82
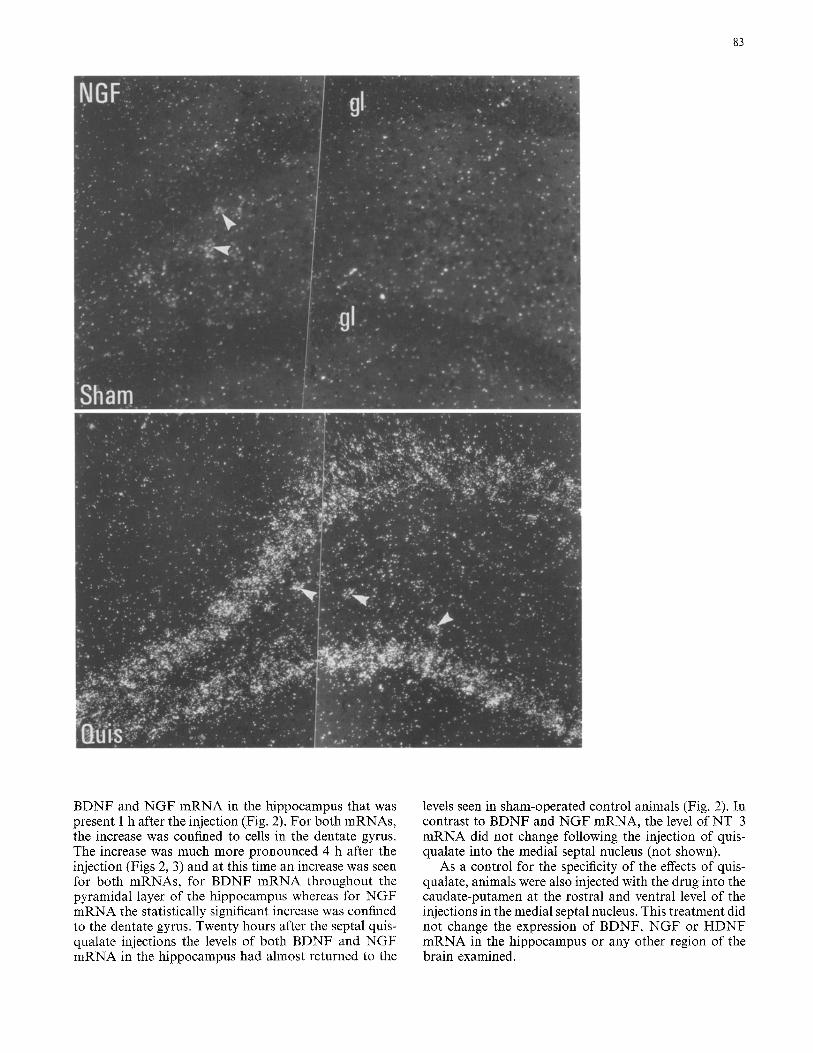
Fig. 3. Left panel shows dark- field emulsion photomicro- graphs of BDNF mRNA ex~ pressing cells in the dentate gyrus of a sham operated ani- mal (upper photograph) and the sm:ae area 4 h after the in- jection of quisqualate into the medial septal nucleus (lower photograph). For comparison both sections were exposed 40 days which revealed strong labeling over granule cells in the injected animal with much lower labeling over granule cells in the control animals. The granule layer of the den- tate gyrus is indicated by gl. R~ht panel shows dark-field emulsion photomicrographs of NGF mRNA expressing cells in the dentate gyrus of a sham operated (upper photograph) and a septal quisqualate injec- ted (lower photograph) animal, 4 h after injections. Arrows in- dicate labeled hilar cells and the granule layer of the den- tate gyrus is indicated by gl
to the filter-bound gene f rom which it was derived (Fig. I). Thus, it is unlikely that the three probes cross hy- bridized to the other members of the family under the conditions used in these experiments. Three different independent in situ hybridization experiments were per- formed with the specific probes and all showed an identi- cal labeling pattern. For evaluation of unspecific labeling within the hippocampus, a 50-mer oligonucleotide probe complementary to the NGF-specific probe (i.e. sense probe) were hybridized in parallel and did not label any cells in the hippocampus.
Both septal and cortical quisqualate injections were
followed by minor behavioural effects, not seen in con- trol animals, with an activation including increased ex- citability. No stereotypic movements , such as activity ar- rests or wet-dog shakings that would indicate epilep- togenic activity were observed.
Increased levels of BDNF and NGF mRNAs in the hippocampus following activation of medial septal neurons
Injection of quisqualate into the medial septal nucleus caused a detectable bilateral increase in the levels of both
83
BDNF and NGF mRNA in the hippocampus that was present 1 h after the injection (Fig. 2). For both mRNAs, the increase was confined to cells in the dentate gyrus. The increase was much more pronounced 4 h after the injection (Figs 2, 3) and at this time an increase was seen for both mRNAs, for BDNF mRNA throughout the pyramidal layer of the hippocampus whereas for NGF mRNA the statistically significant increase was confined to the dentate gyrus. Twenty hours after the septal quis- qualate injections the levels of both BDNF and NGF mRNA in the hippocampus had almost returned to the
levels seen in sham-operated control animals (Fig. 2). In contrast to BDNF and NGF mRNA, the level of NT 3 mRNA did not change following the injection of quis- qualate into the medial septal nucleus (not shown).
As a control for the specificity of the effects of quis- qualate, animals were also injected with the drug into the caudate-putamen at the rostral and ventral level of the injections in the medial septal nucleus. This treatment did not change the expression of BDNF, NGF or HDNF mRNA in the hippocampus or any other region of the brain examined.
84
BDNF NGF
Sham
(~uis
Gtuis+ Scop
Quis+ Diaz
700
6190
Z 5OO E [a. Z 4OO
3130
O
ta 200
100
4t
[ ] Sham [] Quis [ ] Quis + Scop
[ ] Quis + Diaz
Z
Z
O
D G CA 3 CA 1
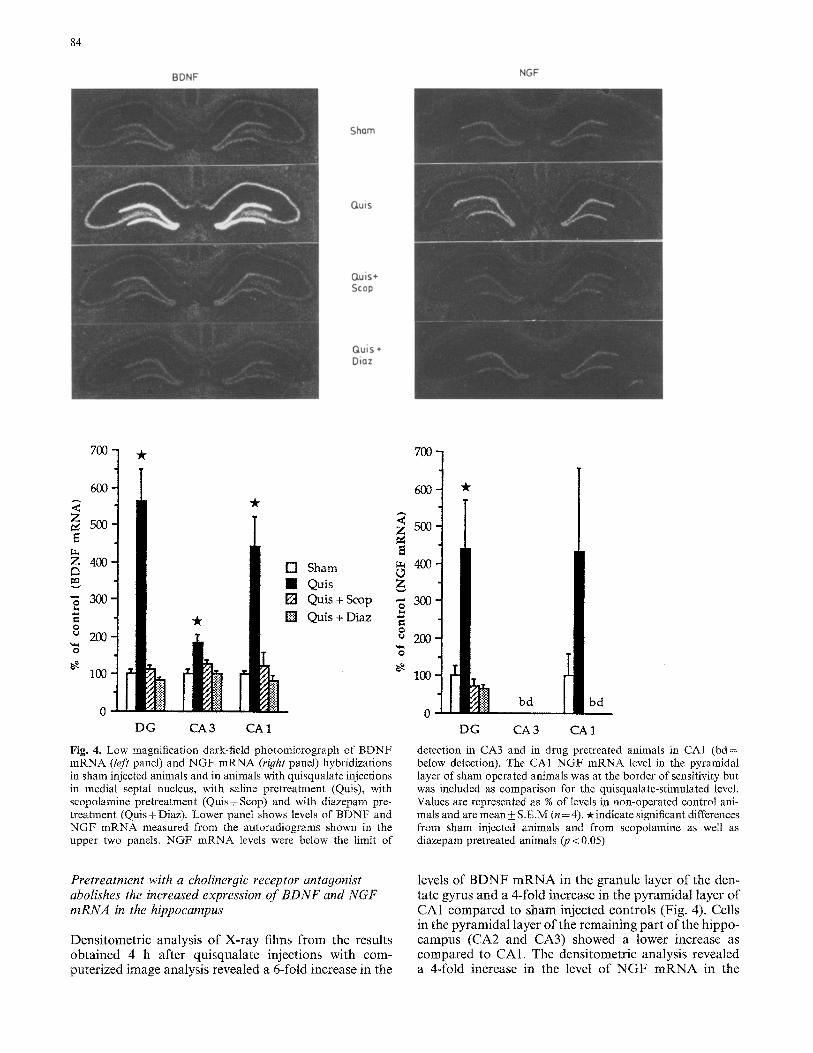
Fig. 4. Low" magnification dark-field photomicrograph of BDNF mRNA (left panel) and NGF mRNA (right panel) hybridizations in sham injected animals and in animals with quisqualate injections in medial septal nucleus, with saline pretreatment (Quis), with scopolamine pretreatment (Quis+Scop) and with diazepam pre- treatment (Quis+ Diaz). Lower panel shows levels of BDNF and NGF mRNA measured from the autoradiograms shown in the upper two panels. NGF mRNA levels were below the limit of
700
600
500
400
300
200
100
"k
bd t bd
D G CA 3 CA 1
detection in CA3 and in drug pretreated animals in CAI (bd= below detection). The CA1 NGF mRNA level in the pyramidal layer of sham operated animals was at the border of sensitivity but was included as comparison for the quisqualate-stimulated level. Values are represented as % of levels in non-operated control ani- mals and are mean + S.E.M (n = 4). -x indicate significant differences from sham injected animals and from scopolamine as well as diazepam pretreated animals (p < 0.05)
Pretreatment with a cholinergic receptor antagonist abolishes the increased expression of BDNF and NGF mRNA in the hippocampus
Densitometric analysis of X-ray films from the results obtained 4 h after quisqualate injections with com- puterized image analysis revealed a 6-fold increase in the
levels of B D N F m R N A in the granule layer of the den- tate gyrus and a 4-fold increase in the pyramidal layer of CA1 compared to sham injected controls (Fig. 4). Cells in the pyramidal layer of the remaining part of the hippo- campus (CA2 and CA3) showed a lower increase as compared to CA1. The densitometric analysis revealed a &fold increase in the level of N G F m R N A in the
BDNF
85
Sham
Quis
Gtuis÷ Stop
Quis+ Diaz
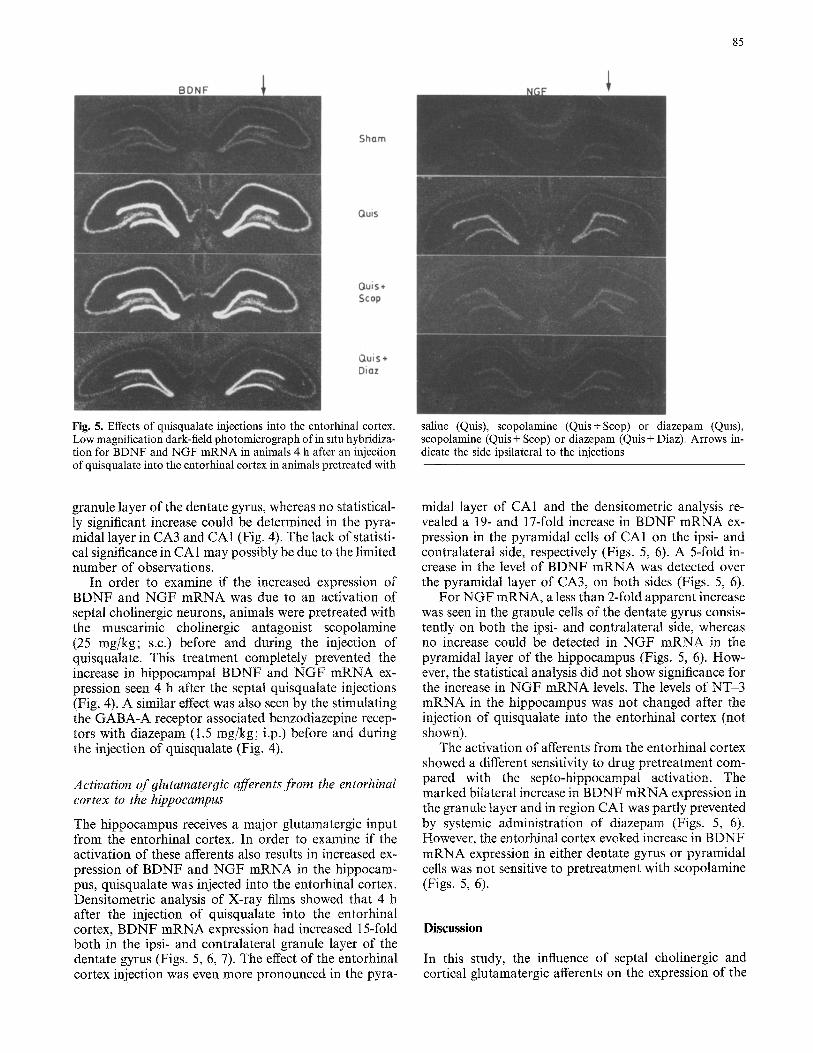
Fig. 5. Effects of quisqualate injections into the entorhinal cortex. Low magnification dark-field photomicrograph of in situ hybridiza- tion for BDNF and NGF mRNA in animals 4 h after an injection of quisqualate into the entorhinal cortex in animals pretreated with
saline (Quis), scopolamine (Quis+Scop) or diazepam (Quis), scopolamine (Quis + Scop) or diazepam (Quis + Diaz). Arrows in- dicate the side ipsilateral to the injections
granule layer of the dentate gyrus, whereas no statistical- ly significant increase could be determined in the pyra- midal layer in CA3 and CA1 (Fig. 4). The lack of statisti- cal significance in CA 1 may possibly be due to the limited number of observations.
In order to examine if the increased expression of BDNF and NGF mRNA was due to an activation of septal cholinergic neurons, animals were pretreated with the muscarinic cholinergic antagonist scopolamine (25 mg/kg; s.c.) before and during the injection of quisqualate. This treatment completely prevented the increase in hippocampal BDNF and NGF mRNA ex- pression seen 4 h after the septat quisqualate injections (Fig. 4). A similar effect was also seen by the stimulating the GABA-A receptor associated benzodiazepine recep- tors with diazepam (1.5 mg/kg; i.p.) before and during the injection of quisqualate (Fig. 4).
Activation of glutamatergic afferents from the entorhinal cortex to the hippocampus
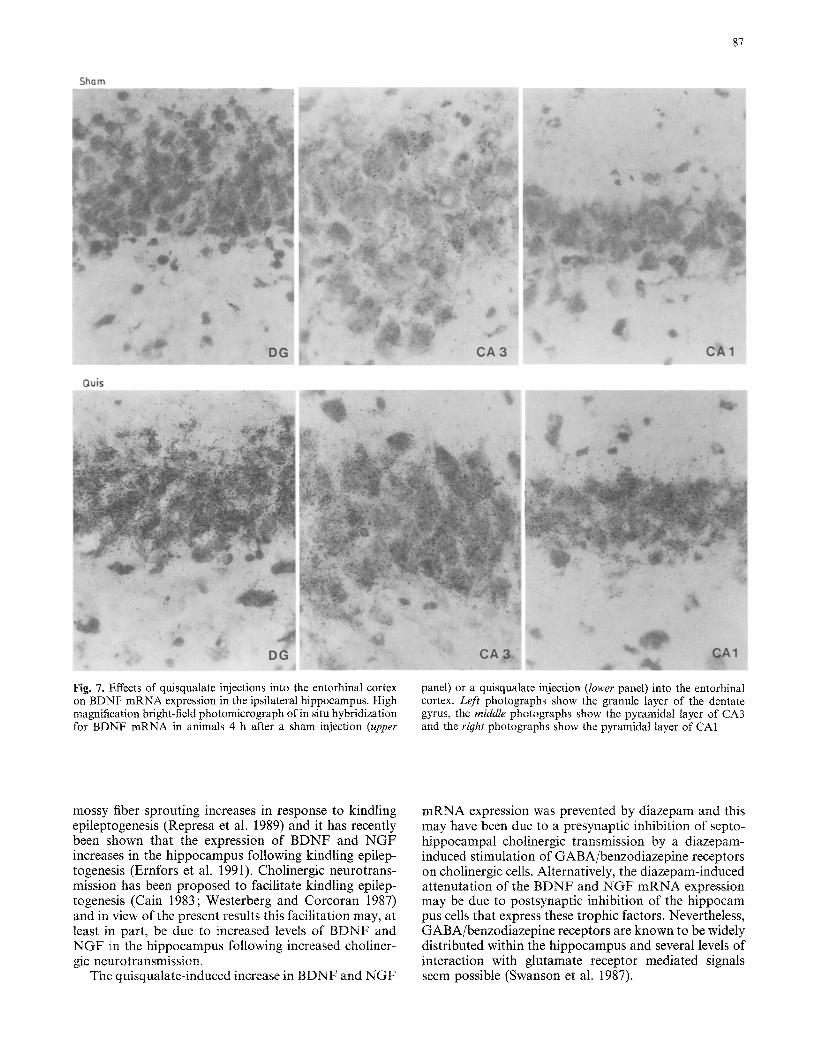
The hippocampus receives a major glutamatergic input from the entorhinal cortex. In order to examine if the activation of these afferents also results in increased ex- pression of BDNF and NGF mRNA in the hippocam- pus, quisqualate was injected into the entorhinal cortex. Densitometric analysis of X-ray films showed that 4 h after the injection of quisqualate into the entorhinal cortex, BDNF mRNA expression had increased 15-fold both in the ipsi- and contralateral granule layer of the dentate gyrus (Figs. 5, 6, 7). The effect of the entorhinal cortex injection was even more pronounced in the pyra-
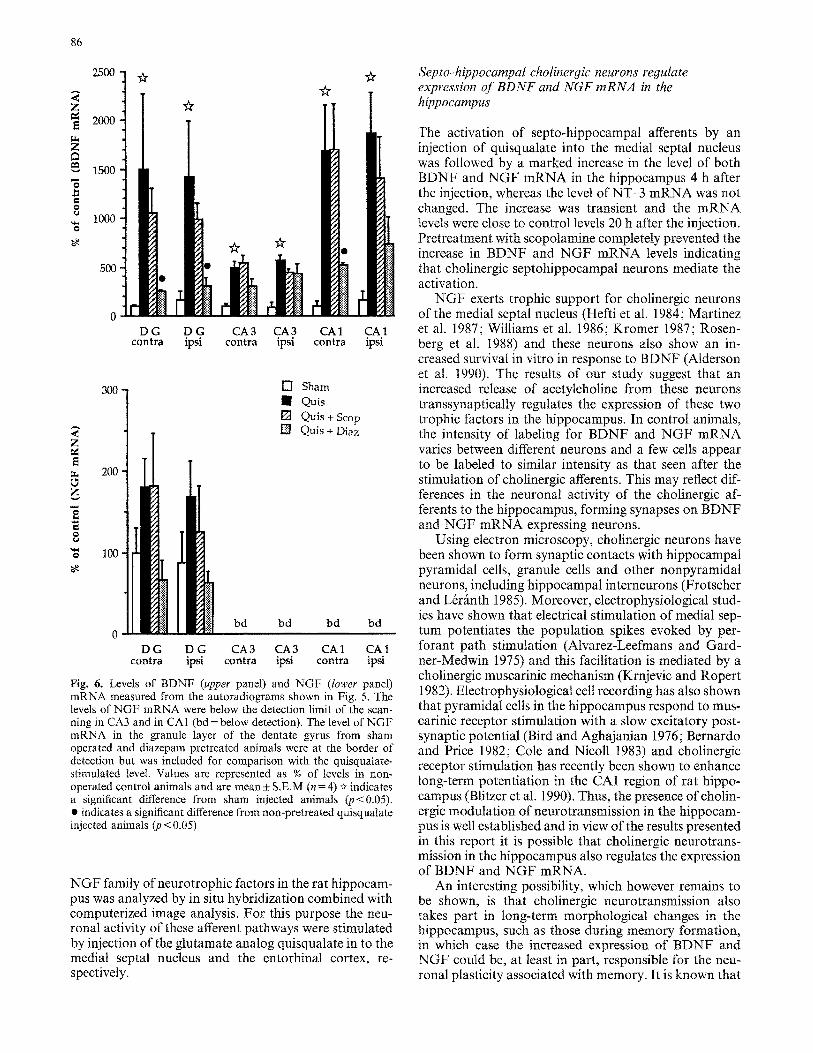
midal layer of CA1 and the densitometric analysis re- vealed a 19- and 17-fold increase in BDNF mRNA ex- pression in the pyramidal cells of CA1 on the ipsi- and contralateral side, respectively (Figs. 5, 6). A 5-fold in- crease in the level of BDNF mRNA was detected over the pyramidal layer of CA3, on both sides (Figs. 5, 6).
For NGF mRNA, a less than 2-fold apparent increase was seen in the granule cells of the dentate gyrus consis- tently on both the ipsi- and contralateral side, whereas no increase could be detected in NGF mRNA in the pyramidal layer of the hippocampus (Figs. 5, 6). How- ever, the statistical analysis did not show significance for the increase in NGF mRNA levels. The levels of NT-3 mRNA in the hippocampus was not changed after the injection of quisqualate into the entorhinal cortex (not shown).
The activation of afferents from the entorhinal cortex showed a different sensitivity to drug pretreatment com- pared with the septo-hippocampal activation. The marked bilateral increase in BDNF mRNA expression in the granule layer and in region CA1 was partly prevented by systemic administration of diazepam (Figs. 5, 6). However, the entorhinal cortex evoked increase in BDNF mRNA expression in either dentate gyrus or pyramidal cells was not sensitive to pretreatment with scopolamine (Figs. 5, 6).
Discussion
In this study, the influence of septal cholinergic and cortical glutamatergic afferents on the expression of the
86
2500 ] ~. ~r
2000
1500
1000
.~. ~ •
•
o D G D G CA3 CA3 CA1 CA1
contra ip~ contra ipsi contra ipsi
3 0 0 -
A
~u 200 , o z
O
'~ 100
bd
[ ] Sham I Quis [] Quis +Scop
Quis + Diaz
bd bd bd 0
D G D G CA 3 CA 3 CA 1 CA 1 contra ipsi contra ipsi contra ipsi
Fig. 6. Levels of BDNF (upper panel) and NGF (lower panel) mRNA measured from the autoradiograms shown in Fig. 5. The levels of NGF mRNA were below the detection limit of the scan- ning in CA3 and in CA1 (bd= below detection). The level of NGF mRNA in the granule layer of the dentate gyrus from sham operated and diazepam pretreated animals were at the border of detection but was included for comparison with the quisqualate- stimulated level. Values are represented as % of levels in non- operated control animals and are mean ± S.E.M (n= 4) ~ indicates a significant difference from sham injected animals (p<0.05). • indicates a significant difference from non-pretreated quisqualate injected animals (4) < 0.05)
NGF family of neurotrophic factors in the rat hippocam- pus was analyzed by in situ hybridization combined with computerized image analysis. For this purpose the neu- ronal activity of these afferent pathways were stimulated by injection of the glutamate analog quisqualate in to the medial septal nucleus and the entorhinat cortex, re- spectively.
Septo~hippocampal cholinergic neurons regulate expression of BDNF and NGF mRNA in the hippoeampus
The activation of septo-hippocampal afferents by an injection of quisqualate into the medial septal nucleus was followed by a marked increase in the level of both BDNF and NGF mRNA in the hippocampus 4 h after the injection, whereas the level of NT-3 mRNA was not changed. The increase was transient and the mRNA levels were close to control levels 20 h after the injection. Pretreatment with scopolamine completely prevented the increase in BDNF and NGF mRNA levels indicating that cholinergic septohippocampal neurons mediate the activation.
NGF exerts trophic support for cholinergic neurons of the medial septal nucleus (Heftiet al. 1984; Martinez et al. 1987; Williams et al. 1986; Kromer 1987; Rosen- berg et al. 1988) and these neurons also show an in- creased survival in vitro in response to BDNF (Alderson et al. 1990). The results of our study suggest that an increased release of acetylcholine from these neurons transsynapticatly regulates the expression of these two trophic factors in the hippocampus. In control animals, the intensity of labeling for BDNF and NGF mRNA varies between different neurons and a few cells appear to be labeled to similar intensity as that seen after the stimulation of cholinergic afferents. This may reflect dif- ferences in the neuronal activity of the cholinergic af- ferents to the hippocampus, forming synapses on BDNF and NGF mRNA expressing neurons.
Using electron microscopy, cholinergic neurons have been shown to form synaptic contacts with hippocampal pyramidal cells, granule cells and other nonpyramidal neurons, including hippocampal interneurons (Frotscher and L&~inth 1985). Moreover, electrophysiological stud- ies have shown that electrical stimulation of medial sep- tum potentiates the population spikes evoked by per- forant path stimulation (Alvarez-Leefmans and Gard- ner-Medwin 1975) and this facilitation is mediated by a cholinergic muscarinic mechanism (Krnjevic and Ropert 1982). Electrophysiological cell recording has also shown that pyramidal cells in the hippocampus respond to mus- carinic receptor stimulation with a slow excitatory post- synaptic potential (Bird and Aghajanian 1976; Bernardo and Price 1982; Cole and Nicoll 1983) and cholinergic receptor stimulation has recently been shown to enhance long-term potentiation in the CA1 region of rat hippo- campus (Blitzer et al. 1990). Thus, the presence of cholin- ergic modulation of neurotransmission in the hippocam- pus is well established and in view of the results presented in this report it is possible that cholinergic neurotrans- mission in the hippocampus also regulates the expression of BDNF and NGF mRNA.
An interesting possibility, which however remains to be shown, is that cholinergic neurotransmission also takes part in long-term morphological changes in the hippocampus, such as those during memory formation, in which case the increased expression of BDNF and NGF could be, at least in part, responsible for the neu- ronal plasticity associated with memory. It is known that
Sham
guis
87
Fig. 7. Effects of quisqualate injections into the entorhinal cortex on BDNF mRNA expression in the ipsilateral hippocampus. High magnification bright-field photomicrograph of in situ hybridization for BDNF mRNA in animals 4 h after a sham injection (upper
panel) or a quisqualate injection (lower panel) into the entorhinal cortex. Left photographs show the granule layer of the dentate gyrus, the middle photographs show the pyramidal layer of CA3 and the right photographs show the pyramidal layer of CA1
mossy fiber sprouting increases in response to kindling epileptogenesis (Represa et al. 1989) and it has recently been shown that the expression of BDNF and NGF increases in the hippocampus following kindling epilep- togenesis (Ernfors et al. 1991). Cholinergic neurotrans- mission has been proposed to facilitate kindling epilep- togenesis (Cain 1983; Westerberg and Corcoran 1987) and in view of the present results this facilitation may, at least in part, be due to increased levels of BDNF and NGF ]in the hippocampus following increased choliner- gic neurotransmission.
The quisqualate-induced increase in BDNF and NGF
mRNA expression was prevented by diazepam and this may have been due to a presynaptic inhibition of septo- hippocampal cholinergic transmission by a diazepam- induced stimulation of GABA/benzodiazepine receptors on cholinergic cells. Alternatively, the diazepam-induced attenutation of the BDNF and NGF mRNA expression may be due to postsynaptic inhibition of the hippocam pus cells that express these trophic factors. Nevertheless, GABA/benzodiazepine receptors are known to be widely distributed within the hippocampus and several levels of interaction with glutamate receptor mediated signals seem possible (Swanson et al. 1987).

88
Entorhinal cortex afferents differentially regulate expression of the NGF family in the hippocampus
A quisqualate-mediated activation of hippocampal af- ferents from the entorhinal cortex differently affected the expression of the members of the NGF family of trophic factors in the hippocampus, particularly in the pyramidal cells. In CA1, BDNF mRNA levels increased 15-fold on both the ipsi- and contralateral side, respectively and in CA3 the BDNF mRNA increase was 5-fold on both sides, while the levels of NGF and NT-3 mRNA were not changed compared to control animals. The response in the granule cells of the dentate gyrus also differed between the three factors with a marked 19- and 17-fold increase in BDNF mRNA levels on the ipsi- and con- tralateral side, respectively while NGF mRNA levels increased less than a 2-fold (not statistically significant but consistent both ipsi- and contralateral to the injec- tions) with no change in the level of NT-3 mRNA. These findings indicate that glutamatergic transmission in the hippocampus differentially regulate expression of the NGF family of trophic factors in the hippocampus in which case release of glutamate from terminals of the perforant pathway markedly increases BDNF mRNA expression with a much less effect on the expression of NGF mRNA. The activation of BDNF mRNA ex- pression is probably due to a non-NMDA receptor stim- ulation by glutamate released from cortical afferent nerve terminals, since non-NMDA but not NMDA receptor stimulation is known to increase the levels of BDNF mRNA in hippocampal neurons (Zafra et al. 1990). The observed increase in BDNF and NGF mRNA was not prevented by pretreatment with the muscarinic antag- onist scopolamine, consistent with the fact that the per- forant pathway is glutamatergic (White et al. 1977). However, pretreatment with the benzodiazepine receptor agonist diazepam attenuated the cortical afferent stimu- lation of both BDNF and NGF mRNA expression in the granule layer of dentate gyrus and the stimulation of BDNF mRNA expression in the pyramidal layer of CA 1, possibly due to activation of presynaptic and/or post- synaptic inhibitory benzodiazepine receptors.
Further a,sTects of transsynaptic activation of BDNF mRNA expression in the hippocampus
The fact that the glutamatergic cortical afferent activa- tion produced a much stronger and more widely spread hippocampal activation of BDNF mRNA expression compared to the expression of NGF and NT-3 mRNAs indicates that BDNF may be more important for neu- ronal interactions between hippocampal neurons and perforant pathway afferents. On the other hand, the septo-hippocampal cholinergic activation produced a similar activation of BDNF and NGF mRNA expression both in the granule layer of the dentate gyrus and in the pyramidal layer of the remaining part of the hippocam- pus. In this context it is interesting to note that both BDNF and NGF have been shown to increase the sur- vival of basal forebrain cholinergic neurons in vitro (A1-
derson et al. 1990). By extension, the lack of response in NT-3 mRNA expression by either the medial septum or the entorhinal cortex quisqulate injections could indicate that NT-3 is not important for neuronal interaction with these afferent pathways. It would therefore be of interest to investigate the role of different monoaminergic af- ferents or peptidergic interneurons in the regulation of NT-3 mRNA expression. However, it can not be ruled out that, in contrast to NGF and BDNF, the expression of NT-3 mRNA is not regulated by neuronal activation.
The observation of behavioural effects by the quis- qualate injections including increased excitability but no stereotypic movements provide no evidence of stimula- tion-induced epileptogenic activity, although this possi- bility can not be excluded. In earlier experiments, how- ever, including direct injections of a non-NMDA agonist (kainic acid) into the hippocampus we have seen clear evidence of epileptogenic activity with e.g. stereotypic forepaw movements and increased salivation (Ballarin et al. 1991) combined with a stronger increase in BDNF mRNA levels in the hippocampus, compared with the increase in BDNF mRNA levels seen in the present study. Thus it seem probable that transsynaptic choliner- gic and glutamatergic activation of BDNF mRNA ex- pression in hippocampus is possible also using stimula- tion intensities not producing seizures.
The transsynaptic cholinergic and glutamatergic stim- ulation of BDNF and NGF mRNA expression described in this study opens up the possibility to selectively in- crease the levels of these two trophic factors by pharma- cological treatments which may facilitate neuronal plas- ticity and perhaps prevent degeneration of specific pop- ulations of neurons in the central nervous system.
Acknowledgements. This work was supported by grams from The Swedish Medical Research Council (grant no. 8653), from Ake Wibergs Stiftelse, Lot and Hans Ostermans fond and The Swedish Natural Science Research Council.
References
Alderson RF, Alterman AL, Barde Y-A, Lindsay RM (1990) Brain- derived neurotrophic factor increases survival and differentiated functions of rat septal chotinergic neurons in culture. Neuron 5: 297-306
Alvarez-Leefmans FJ, Gardner-Medwin AR (1975) Influences of the septum on the hippocampal dentate area which are unac- companied by field potentials. J Physiol 249: 14~-16P
Ayer-LeLi~vre C, Olson L, Ebendal T, Seiger A, Persson H (1988) Expression of the 13-nerve growth factor in hippocampal neu- rons. Science 240:1339-1341
Ballarin M, Ernfors P, Lindefors N, Persson H (1991) Rapid induc- tion of mRNA tbr brain derived neurotrophic factor following neuronal excitation by kainic acid. Exp NeuroI (in press)
Barde Y-A, Edgar D, Thoenen H (1982) Purification of a new neu- rotrophic factor from mammalian brain. EMBO J 1:549-553
Barde Y-A (1989) Trophic factors and survival. Neuron 2:1525-1534
Bartus RT, Dean RL, Beer B, Lippa AS (1982) The cholinergic hypothesis of geriatric memory dysfunction. Science 217:408-4t4
Bernardo LS, Price DA (1982) Cholinergic pharmacology of mam- malian hippocampal pyramidal cells. Neuroscience 7:1703-1712

89
Bird SJ, Aghajanian GK (1976) The cholinergic pharmacology of hippocampal pyramidal cells: a microiontophoretic study. Neuropharmacology 15: 273-282
Blizer RD, Gil O, Landau EM (t990) Cholinergic stimulation enhances long-term potentation in the CA1 region of rat hippo- campus. Neurosci Lett 119: 201-210
Cain DP (1983) Bidirectional transfer of electrical and carbachol kindling. Brain Res 260:135-138
Cole AE, Nicoll RA (1983) Acetylcholine mediates a slow synaptic potential in hippocampal pyramidal cells. Science 221 : 1299-1301
Cole AE, Nicoll RA (1984) The pharmacology of cholinergic ex- citatory responses in hippocampal pyramidal cells. Brain Res 305 : 283-290
Collerton D (1986) Cholinergic function and intellectual decline in Alzheimer's disease. Neuroscience 19:1-28
Ernfors P, Ebendal T, Olson L, Mouton P, StrSmberg I, Pers- son H (1989) A celt line producing recombinant nerve growth factor evokes growth responses in intrinsic and grafted central cholinergic neurons. Proc Natl Acad Sci USA 86:4756-4760
Ernfors P, Wetmore C, Olson L, Persson H (1990a) Identification of cells in rat brain and peripheral tissues expressing mRNA for members of the nerve growth factor family. Neuron 5:511-526
Ernfors P, Ibafiez CF, Ebendal T, Olson L, Persson H (1990b) Molecular cloning and neurotrophic activities of a protein with structural similarities to nerve growth factor: developmental and topographical expression in the brain. Proc Natl Acad Sci USA 87:5454-5458
Ernfors P, Bengtzon J, Kokaia Z, Persson H, Lindval O (1991) Increased levels of messenger RNAs for neu~otrophic factors in the brain during kindling epileptogenesis. Neuron (in press)
Fische W, Wictorin K, BjSrklund A, Williams LR, Varon S, Gage FH (1987) Amelioration of cholinergic atrophy and spatial memory impairment in aged rats by nerve growth factor. Nature (London) 329: 65-68
Fonnum F (1984) Glutamate: a neurotransmitter in mammalian brain. J Neurochem 42: 1-11
Friedman W, Ernfors P, Persson H (1991 b) Transient and persistent expression of HDNF/NT-3 in the rat brain during postnatal development. J Neurosci (in press)
Frotscher M, Ldr~inth C (t985) Cholinergic innervation of the rat hippocampus as revealed by choline acetyltransferase immuno- histochemistry: a combined light and electron microscopic study. J Comp Neurol 239:237-246
Gall CM, Isackson PJ (1989) Limbic seizures increase neuronal production of messenger RNA for nerve growth factor. Science 245: 758-761
Goedert M, Fine A, Hunt SP, Ullrich A (1986) Nerve growth factor mRNA in peripheral and central rat tissue and in the human central nervous system: lesion effects in the rat brain and levels in Alzheimer's disease. Mol Brain Res t:85-92
Hagg T, Vahlsing HL, Manthorpe M, Varon S (1990) Nerve growth factor infusion into the denervated adult rat hippocampal formation promotes its cholinergic reinnervetion. J Neurosci 10:3087-3092
Hefti F (1986) Nerve growth thctor promotes survival of septal cholinergic neurons after fimbrial transections. J Neurosci 6:2155-2162
Hohn A, Leibrock J, Bailey K, Barde Y-A (1990) Identification and characterization of a novel member of the nerve growth factor/ brain derived neurotrophic factor family. Nature (London) 344:339-341
Hofer MM, Barde Y-A (1988) Brain-derived neurotrophic factor prevents neuronal cell death in vivo. Nature (London) 33 t : 261-262
Hofer MM, Pagliusi SR, Hohn A, Leibrock J, Barde YA (1990) Regional distribution of brain-derived neurotrophic factor in the adult mouse brain. EMBO J 9:2459-2464
Johnson JE, Barde Y-A, Schwab M, Thoenen H (1986) Brain- derived neurotrophic factor supports the survival of cultured rat ganglion cells. J Neurosci 6:3031-3038
Kaisho Y, Yoshimura K, Nakahama K (1990) Cloning and ex- pression of a cDNA encoding a novel human neurotrophic factor. FEBS Lett 266:187-191
Korsching S, Auburger G, Heumann R, Scott J, Thoenen H (1985) Levels of nerve growth factor and its mRNA in the central nervous system of the rat correlate with cholinergic innervation. EMBO J 4:1389-1393
Krejevic K, Ropert N (1981) Septo-hippocampal pathway modu- lates hippocampal activity by a cholinergic mechanism. Can J Physiol Pharmacol 59:911-914
Krejevic K, Ropert N (1982) Electrophysiological and pharmaco- logical characteristics of facilitation of hippocampal population spikes by stimulation of the medial septum. Neuroscience 7:2165-2183
Kromer LF (1987) Nerve growth factor treatment after brain injury prevents neuronal death. Science 235: 214-216
Leibrock J, Lottspeich F, Hohn A, Hofer M, Hengerer B, Ma- siakowski P, Thoenen H, Barde YA (1989) Molecular cloning and expression of brain-derived neurotrophic factor. Nature (London) 341:149-152
Martinez H J, Dreyfus CF, Jonakait GM, Black IB (1987) Nerve growth factor selectively increases cholinergic markers but not neuropeptides in rat basal forebrain culture. Brain Res 412:295-301
McGeer PL, McGeer EG, Peng JH (1984) Choline acetyltransferase purification and immunohistochemical localization. Life Sci 34: 2319-2338
Levi-Montalchini R, Angeletti PU (1968) The nerve growth factor. Physiol Rev 48: 534-569
Maisonpierre PC, Belluscio L, Squinto S, Ip NY, Furth ME, Lind- say RM, Yancopoulos GD (1990) Neurotrophin-3: a neuro- trophic factor related to NGF and BDNF. Science 247:1446-1451
Milner B, Corkin S, Teuber H-L (1968) Further analysis of the hippocampal amenestic syndrome: 14-year follow-up study of H.M. Neuropsychology 6:215~34
Mishkin M (1982) A memory system in the monkey. Philos Trans R Soc London 298:85-95
MoNey WC, Rutkowsky JL, Tennekoon GI, Buchanan K, Johnston MV (1985) Choline acetyltransferase activity in stri- atum of rats increased by nerve growth factor. Science 229: 284-287
Monaghan DT, Bridges RJ, Cotman CW (1989) The excitatory amino acid receptors. Ann Rev Pharmacol Toxicol 29:365-401
O'Keefe J, Nadel L (1978) The hippocampus as a cognitive map. Clarendon, Oxford
Olton DS, Becker JT, Handelman GE (1979) Hippocampus, space and memory. Behav Brain Sci 2:313-365
Perry EK, Tomilson BE, Blessed G, Bergman K, Gibson PH, Perry RH (1978) Correlation of cholinergic abnormalities with senile plaques and mental test scores in senile dementia. Br Med J 2:1457-1459
Philips HD, Hains JM, Laramee GR, Rosenthal A, Winslow JW (t990) Widespred expression of BDNF but not NT-3 by target areas of basal forebrain cholinergic neurons. Science 250: 290-294
Prochiantz A, DiPortio U, Kato A, Breger B, Glowinski J (1979) In vitro maturation of mesencephalic dopaminergic neurons form mouse embryos is enhanced in presence of their striatal target ceils. Proc Natl Acad Sci USA 76:5387-5391
Radeke MJ, Misko TP, Hsu C, Herzenberg LA, Shooter EM (1987) Gene transfer and molec~dar cloning of the rat nerve growth factor receptor. Nature 325:593-597
Represa A, Ire Gall La Salle G, Ben-Ari Y (1989) Hippocampal plasticity in the kindling model of epilepsiy in rats. Neurosci Lett 99:345-350
Richardson PM, Verge Issa VMK, Riopelle RJ (1986) Distribution of neuronal receptors for the nerve growth factor in the rat. J Neurosci 6: 2312-2321
Rosenberg MB, Friedmann T, Robertson RC, Tuszynski M, Wolff JA, Bl'eakefield XO, Gage FH (1988) Grafting genetically

90
modified cells to the damaged brain: restorative effects of NGF expression. Science 242:1575-1578
Rosenthal A, Goeddel DV, Nguyen T, Lewis M, Shih A, Laramee GR, Nikolics K, Winslow JW (1990) Primary structure and biological activity of a novel human neurotrophic factor. Neuron 4:767-773
Schwab ME, Otten U, Agid Y, Thoenen H (1979) Nerve growth factor (NGF) in the CNS: absence of specific retrograde axonal transport and tyrosine hydroxylase induction in locus coeruleus and substantia nigra. Brain Res 168:473~483
Shetton DL, Reichardt LF (1986) Studies on the expression of the [3-nerve growth factor (NGF) gene in the central nervous sys- tem: level and regional distribution of NGF mRNA suggest that NGF functions as a trophic factor for several distinct popula- tions of neurons. Proc Natl Acad Sci USA 83:2714-2718
Seiter M, Schwab ME (1984) Specific retrograde transport of nerve groth factor (NGF) from neocortex to nucleus basalis in the rat. Brain Res 300:33-39
Swanson LW, K6hler C, Bj6rklund A (1987) The limbic region. I. The septo-hippocampal system. In: (Bj6rklund A, H6kfelt T, Swanson LW (eds) Handbook of chemical anatomy, Vol 5. Integrated systems of the CNS, Part I. Elseiver, Amsterdam, pp 125-277
Taniuchi M, Schweitzer JB, Johnson EJ (1986) Nerve growth factor receptor molecules in rat brain. Proc Natl Acad Sci USA 83 : 1950--1954
Thoenen H, Barde Y-A (1980) Physiology of nerve growth factor. Physiol Rev 60:1284-1335
Tomozowa Y, Appel SH (1986) Soluble striatal extracts enhance development of mesencephalic dopaminergic neurons in vitro. Brain Res 399:111-124
Westerberg V, Corcoran ME (1987) Antagonism of central but not peripheral cholinergic receptors retards amygdala kindling in rats. Exp Neurol 95:194-206
Wetmore C, Ernfors P, Persson H, Olson L (1990) Localization of brain-derived neurotrophic factor mRNA to neurons in the brain by in situ hybridization. Exp Neuron 109:141 152
White WF, Nadler JV, Hamberger A, Cotman CW, Cummings JT (1977) Glutamate as transmitter of hippocampal perforant path. Nature (London) 270:356-357
Whitehouse PJ, Price DL, Struble RG, Clark AW, Coyle JT, De- Long MR (1982) Alzheimer's disease and senile dementia: loss of cholinergic neurons in the basal forebrain. Science 215:1237-1239
Whittemore SR, Ebendal T, L/irkfors L, Olson L, Seiger A, Str6m- berg I, Persson H (1986) Developmental and and regional ex- pression of nerve gowth factor messenger RNA and protein in the rat central nervus system. Proc Natl Acad Sci USA 83:817-821
Whittemore SR, Friedman, PL, Larhammar D, Persson H, Gonzalez-Carvajal M, Holets VR (1988) Rat [3-nerve growth factor sequence and site of sythesis in the audlt hippocampus. J Neurosci Res 20:403-410
Wilcock GK, Esiri MM, Bowen DM, Smith CC (1982) Alzheimer's disease. Correlation of cortical acetyltransferase activity with the severity of dementia and and histological abnormalities. J Neurol Sci 57:40-417
Williams LR, Varon S, Peterson GM, Wictorin K, Fischer W, Bj6rklund A, Gage FH (1986) Continous infusion of nerve growth factor prevents basal forebrain neuronal cell death after fimbria fornix transection. Proc Natl Acad Sci USA 83:9231-9235
Zafra F, Hengerer B, Leibrock J, Thoenen H, Lindholm D (1990) Activity dependent regulation of BDNF and NGF mRNAs in the hippocampus is mediated by non-NMDA glutamate recep- tors. EMBO J 9:3545 3550
Copyright © 2022 FDOKUMEN





![Results of Ventricular Septal Myectomy and Hypertrophic Cardiomyopathy (from Nationwide Inpatient Sample [1998–2010])](https://static.fdokumen.com/doc/165x107/632e4970f835cf7c7c0a2906/results-of-ventricular-septal-myectomy-and-hypertrophic-cardiomyopathy-from-nationwide.jpg)














