
Seasonal variation of snow microbial community structure in the East Rongbuk glacier, Mt. Everest
11
ARTICLES 1476 Chinese Science Bulletin Vol. 51 No. 12 June 2006 Chinese Science Bulletin 2006 Vol. 51 No. 12 1476 1486 DOI: 10.1007/s11434-006-1476-7 Seasonal variation of snow microbial community structure in the East Rongbuk glacier, Mt. Everest LIU Yongqin 1 , YAO Tandong 1,2 , KANG Shichang 1,2 , JIAO Nianzhi 3 , ZENG Yonghui 3 , SHI Yang 3 , LUO Tingwei 2 , JING Zhefang 2 & HUANG Shijun 3 1. Institute of Tibetan Plateau Research, Chinese Academy of Sciences, Beijing 100085, China; 2. Key Laboratory of Cryosphere and Environment, Chinese Academy of Sciences, Lanzhou 730000, China; 3. National Key Laboratory for Marine Environmental Sciences, Xiamen University, Xiamen 361005, China Correspondence should be addressed to Liu Yongqin (email: yqliu@ itpcas.ac.cn) Received December 28, 2005; accepted April 17, 2006 Abstract The bacterial diversity and abundance in the snow of East Rongbuk glacier, Mt. Everest were examined through 16S rRNA gene clone library and flow cytometry approaches. In total, 35 16S rRNA gene sequences were obtained, which belong to α, β, γ-Proteobacteria, Actinobacteria, Firmicutes, CFB, Cyanobacteria, Eukaryotic chloroplast, and TM7 candidate phylum respectively. γ-Proteobacteria was the dominant bacterial group in this region, while the genera Acinetobacter and Leclercia were domi- nant on the genus level. The community structure varied seasonally. The bacterial abundance in sum- mer snow was higher than that in winter. Moreover, the snow bacterial community structures in both seasons were diverse, with not only common species but season-specific species. The common species most likely originated from the Tibet Plateau. Bacteria in summer snow are affiliated with marine environ- ment, whereas bacteria in winter snow are closely related to more diverse environments and show the feature of resistance to cold. Seasonal variations of abundance and bacterial diversity were most proba- bly due to the seasonal characteristics of climate and atmospheric circulation in Mt. Everest. Keywords: Mt. Everest, east Rongbuk glacier, bacterial diversity and abundance, seasonal variation. Mt. Everest, the zenith of the world, plays an impor- tant role in the global climate change. During the last decade, some studies on glacier and snow have been undertaken in this region. The climate and environment changes in the past hundred years have been researched, and the impact of anthropogenic pollution on this high-elevation remote regions has been evaluated [1―6] . However, most of the studies focused on chemical or physical characters of ice and snow, little is touched on microorganism. In 1975, fungi and algae have been studied in this region by the Chinese Academy of Sciences through the expedition on Mt. Everest [7,8] . Xiong et al. isolated yeasts from two snow samples in Xixiabangma Peak, Himalaya [9] and revealed that the number of bacteria were 6.9×10 6 and 17.9×10 6 cells/mL −1 , respectively. Baghela et al. [10] studied proteolytic bacteria in the cold Gangotri glacier, Western Himalaya, India. Yoshitaka et al. [11] examined the snow algae in a Himalayan glacier and the changes of algal biomass and community structure along the altitude gradient. In recent years, a lot of work on microorganisms in the ice core and their relationship with the climatic and environmental changes have been carried out on the Tibet Plateau. Yao et al. [12] , Xiang et al. [13] and Zhang et al. [14] surveyed the relationship between the bacteria in Malan ice core and the past climate and environment, and found that the bacterial concentration negatively correlated with temperature and positively with the distribution of dirty layers. In addition, their work revealed that the layer distribution of bacterial community in the ice core can reflect the microbial response to the climatic and envi- ronmental conditions at the time of deposition. A re- search on the bacteria isolated from the different depths of Muztag Ata glacier showed that the bacterial abun- dance was closely related to dust and, responding to the environmental change, the distribution pattern of iso- lates in different depths varied with the high G+C and CFB being the dominant groups [15] . During the fourth expedition on Mt. Everest of the Chinese Academy of Sciences from April to June 2005, we studied the native bacterial communities in snow in the col of East Rongbuk (ER) glacier (6500 m a.s.l.). This work is the first report of bacterial community in the north slope of Mt. Everest. Here we use flow cy- tometry to enumerate the total number of microbial cells, and construct 16S rRNA gene clone libraries us- ing PCR method. Snow physical, chemical and other environmental variables are measured as well to have a better understanding of the relationship between the seasonal change of bacterial community structure and
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Seasonal variation of snow microbial community structure in the East Rongbuk glacier, Mt. Everest

ARTICLES
1476 Chinese Science Bulletin Vol. 51 No. 12 June 2006
Chinese Science Bulletin 2006 Vol. 51 No. 12 1476 1486 DOI: 10.1007/s11434-006-1476-7
Seasonal variation of snow microbial community structure in the East Rongbuk glacier, Mt. Everest LIU Yongqin1, YAO Tandong1,2, KANG Shichang1,2, JIAO Nianzhi3, ZENG Yonghui3, SHI Yang3, LUO Tingwei2, JING Zhefang2 & HUANG Shijun3
1. Institute of Tibetan Plateau Research, Chinese Academy of Sciences, Beijing 100085, China;
2. Key Laboratory of Cryosphere and Environment, Chinese Academy of Sciences, Lanzhou 730000, China;
3. National Key Laboratory for Marine Environmental Sciences, Xiamen University, Xiamen 361005, China
Correspondence should be addressed to Liu Yongqin (email: yqliu@ itpcas.ac.cn) Received December 28, 2005; accepted April 17, 2006
Abstract The bacterial diversity and abundance in the snow of East Rongbuk glacier, Mt. Everest were examined through 16S rRNA gene clone library and flow cytometry approaches. In total, 35 16S rRNA gene sequences were obtained, which belong to α, β, γ-Proteobacteria, Actinobacteria, Firmicutes, CFB, Cyanobacteria, Eukaryotic chloroplast, and TM7 candidate phylum respectively. γ-Proteobacteria was the dominant bacterial group in this region, while the genera Acinetobacter and Leclercia were domi-nant on the genus level. The community structure varied seasonally. The bacterial abundance in sum-mer snow was higher than that in winter. Moreover, the snow bacterial community structures in both seasons were diverse, with not only common species but season-specific species. The common species most likely originated from the Tibet Plateau. Bacteria in summer snow are affiliated with marine environ-ment, whereas bacteria in winter snow are closely related to more diverse environments and show the feature of resistance to cold. Seasonal variations of abundance and bacterial diversity were most proba-bly due to the seasonal characteristics of climate and atmospheric circulation in Mt. Everest.
Keywords: Mt. Everest, east Rongbuk glacier, bacterial diversity and abundance, seasonal variation.
Mt. Everest, the zenith of the world, plays an impor-tant role in the global climate change. During the last
decade, some studies on glacier and snow have been undertaken in this region. The climate and environment changes in the past hundred years have been researched, and the impact of anthropogenic pollution on this high-elevation remote regions has been evaluated[1―6]. However, most of the studies focused on chemical or physical characters of ice and snow, little is touched on microorganism.
In 1975, fungi and algae have been studied in this region by the Chinese Academy of Sciences through the expedition on Mt. Everest[7,8]. Xiong et al. isolated yeasts from two snow samples in Xixiabangma Peak, Himalaya[9] and revealed that the number of bacteria were 6.9×106 and 17.9×106 cells/mL−1, respectively. Baghela et al.[10] studied proteolytic bacteria in the cold Gangotri glacier, Western Himalaya, India. Yoshitaka et al.[11] examined the snow algae in a Himalayan glacier and the changes of algal biomass and community structure along the altitude gradient. In recent years, a lot of work on microorganisms in the ice core and their relationship with the climatic and environmental changes have been carried out on the Tibet Plateau. Yao et al.[12], Xiang et al.[13] and Zhang et al.[14] surveyed the relationship between the bacteria in Malan ice core and the past climate and environment, and found that the bacterial concentration negatively correlated with temperature and positively with the distribution of dirty layers. In addition, their work revealed that the layer distribution of bacterial community in the ice core can reflect the microbial response to the climatic and envi-ronmental conditions at the time of deposition. A re-search on the bacteria isolated from the different depths of Muztag Ata glacier showed that the bacterial abun-dance was closely related to dust and, responding to the environmental change, the distribution pattern of iso-lates in different depths varied with the high G+C and CFB being the dominant groups[15].
During the fourth expedition on Mt. Everest of the Chinese Academy of Sciences from April to June 2005, we studied the native bacterial communities in snow in the col of East Rongbuk (ER) glacier (6500 m a.s.l.). This work is the first report of bacterial community in the north slope of Mt. Everest. Here we use flow cy-tometry to enumerate the total number of microbial cells, and construct 16S rRNA gene clone libraries us-ing PCR method. Snow physical, chemical and other environmental variables are measured as well to have a better understanding of the relationship between the seasonal change of bacterial community structure and

ARTICLES
www.scichina.com www.springerlink.com 1477
the environment.
1 Materials and methods
1.1 Sample collection and the measurement of envi-ronmental variables
A 1.7 m deep snow pit was dug in the ER Glacier on the northern slope of Mt. Everest (27°59′N, 86°55′E) (Fig. 1). Snow samples were collected continuously at 15 cm depth interval for microbiological analysis, and at 5 cm interval for the determination of stable oxygen isotope ratios, and the concentration of major dissolv- ing ions and metal elements. Extreme care was taken at all times during the whole sampling processes to ensure the minimal contamination. For example, non-particul- ating suits, polyethylene gloves and masks were always pulled on. Pre-washed high-density polyethylene HDPE containers were used as sample scoops. The bottles filled with ultra-pure waters were used as negative con- trol which were opened during samples collection and handled in the same way as snow samples. The in situ temperature was −11.6�. Snow samples were kept fro-zen during the transport process.
Snow melt water were subjected to flow cytometry analysis (Beckman Coulter, Epics Altra II)in the Na- tional Key Laboratory for Marine Environmental Sci- ences, Xiamen University for counting the total number of bacteria. Analysis of δ18O was performed in the Key Laboratory of Cryosphere and Environment, the Chi- nese Academy of Sciences, using an MAT-252 delta-
plus mass spectrometer, and results are expressed as the relative deviation of heavy isotope content of standard mean ocean water (SMOW). The major chemical spe- cies were measured in the laboratory of Institute of Ti- bet an Plateau Research, the Chinese Academy of Sc- iences, via suppressed ion chromatography (Dionex 320 series instruments). Metal elements were detected in the Center for Analysis and Test of Institute of Soil Science, the Chinese Academy of Sciences.
1.2 DNA extraction, PCR amplification and 16S rRNA gene clone library construction
Approximately 500 1000 mL of each snow sample was melted at 4�, and filtered through a 0.22 μm filter (Millipore). The membrane was incubated with 1 mL GTE buffer (25 mmol/L Tris, 10 mmol/L EDTA, 50 mmol/L Sucrose, 20 mg/mL, pH 8.0) at 37� for 2 h. Then, proteinase K, NaCl and sodium dodecyl sulfate were added at the final concentrations of 0.2 mg/mL, 0.7 mol/L and 1%, respectively. The mixture was incu- bated at 65� for 1.5 h. After extraction with phe- nol-chloroform-isoamyl alcoholl (25:24:1) and chlo- roform-isoamyl alcohol (24:1), DNA in the aqueous phase was precipitated with an equal volume of iso- propanol overnight at −20�. To prevent contamination in any step of the procedure, a negative control was established by filtering 1 L of autoclaved deionized water with the same method as described above. After centrifugation and washing with 75% ethanol, the DNA
Fig. 1. Sample location, East Rongbuk glacier in Mt. Everest (6500 m a.s.l.).

ARTICLES
1478 Chinese Science Bulletin Vol. 51 No. 12 June 2006
was dissolved in TE buffer. DNA preparations from both the sample and the negative control were used as templates to amplify the bacterial 16S rDNA genes. Primers were 27F (forward, 5′-AGA GTT TGA TCM TGG CTC AG-3′) and 1392R (reverse, 5′-ACG GGC GGT GTG TRC-3′). The PCR program was as follows: after an initial incubation at 94 for 5 min, 30 cycles were run with 94 for 1 min, 56 for 1 min and 72 for 1.5 min, followed by a final extension at 72 for 8 min.
The PCR products were purified using Agarose gel DNA purification kit (TaKaRa Biotechnology Co., Da- lian, China) and ligated into pGEM-T vector (TaKaRa Biotechnology Co., Dalian, China), then transformed into E. coli DH5α. The presence of inserts was checked by ampicillin resistance selection and colony PCR.
For each clone library, 75―100 clones were selected randomly for reamplificaton and restriction digest by enzymes Hha I and Afa I. The digested fragments were visualized on a 3% agarose gel, and different clones were discriminated according to the RFLP patterns. One clone of each RFLP type was sequenced.
Genetic diversity obtained by RFLP analysis was subjected to statistical analysis including the following indexes: (1) Taxa, the total numbers of RFLP pattern in each library; (2) Individuals, the total clone numbers examined; (3) Coverage which is derived from the equation Coverage=1−(N/Individuals), N is the number of clones that occurred only once[16]; (4) Diversity in- dexes (Dominance, Evenness, Shannon, Simpson), cal- culated using the statistical program PAST (http://folk. uio.no/ohammer/past).
1.3 Sequencing and phylogenetic analysis
All sequences obtained were checked for chimeric artifacts by the CHIMERA_CHECK[17] program. The nearest neighbors were retrieved from the NCBI (http://www.ncbi.nih.gov/BLAST) and Ribosomal Database Project 9.0 (http://rdp.cme.msu.edu) databases though the BLAST search.
Sequences and their neighbors in the database were aligned using the program ClustalX 1.80. The neighbor- joining analysis was performed with the program Phylip 3.63 (http://evolution.genetics.washington.edu/ phylip.html). The neighbor-joining tree was constructed from the distance matrix corrected with Kimura’s two- parameter model. Bootstrap values were obtained with 100 resamplings. The 16S rDNA sequences from ar- chaea Methanothermus fervidus was used as outgroup.
The program DOTUR was used to determine the af- filiation of each sequence on the genus level (http:// www.plantpath.wisc.edu/fac/joh/dotur.html).
The nucleotide sequences of partial 16S rRNA genes have been deposited in the GenBank database with the accession numbers: DQ323081―DQ323115.
2 Results
2.1 Stable oxygen isotope ratios and seasonal differ-ence of snowpit
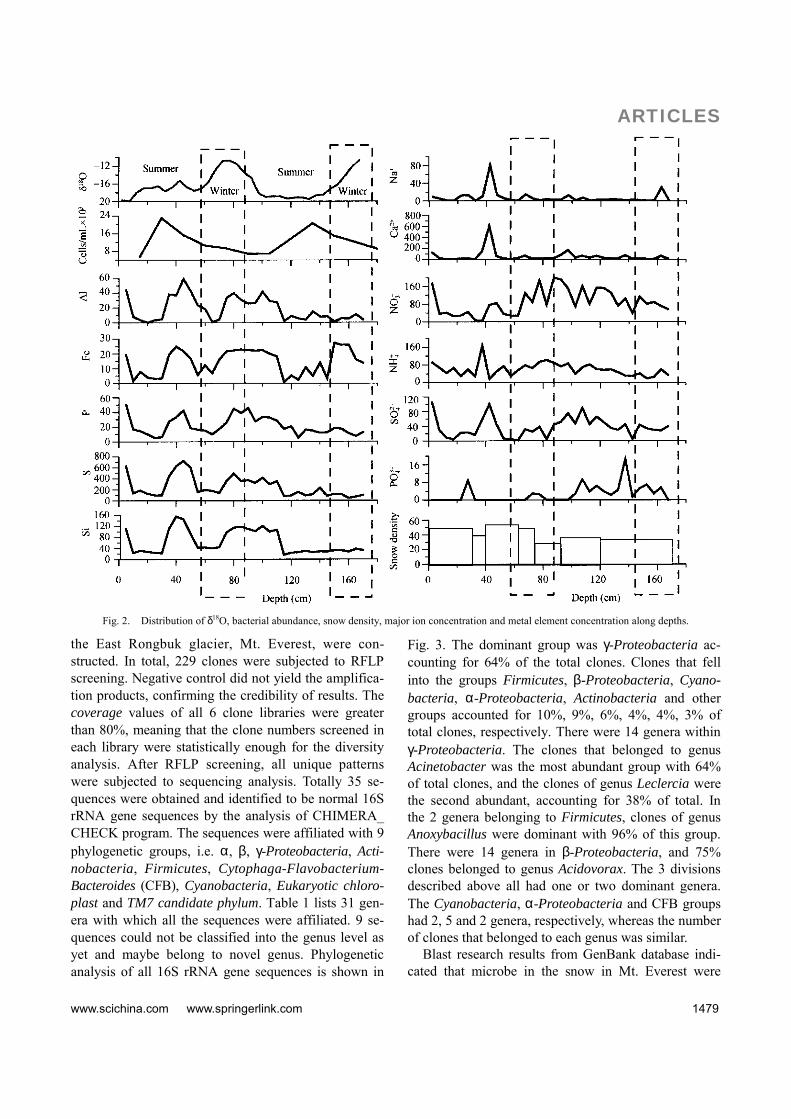
Stable oxygen isotope ratios of precipitation were controlled by the changes in condensation level, con-densation temperature and moisture in Tibetan Pla-teau[18]. Strong monsoon activity in the southern Ti-betan Plateau results in high precipitation rates and more depleted heavy isotopes, which was described as a precipitation “amount effect”[19,20]. For Himalayan precipitation, δ18O was enriched during the winter dry season and depleted during the wet summer season. Therefore, the corresponding precipitation seasons of different layers of snow pit could be determined. Our results show that the 1.7 m depth snow pit spans two years (Fig. 2). The snow deposition of the two parts with a higher value of δ 18O (layer 62 98 cm, layer 158 170 cm) occurred in winter, and that of another two parts with lower δ 18O (layer 0 62 cm, layer 98158 cm) occurred in summer. However, the snowfall in the field that belongs to the winter of 2004 2005 was not found. The δ 18O results also show the feature that all the top layers of snow pit are of lower values. Major dissolved ions such as Ca2+ and Mg2+ had the same character, but the concentrations of metal elements were high. It was deduced that little snowfall in winter between 2004 and 2005 caused the low dissolved ion concentration, but the dry precipitation resulted in the high metal element value.
2.2 Bacterial abundance in the snow
Bacterial abundance in the snow of Mt. Everest ranged from 5.7×103 to 2.3×104 cells/mL. Populations were maximal at the 15―30 cm depth of snow, and the lowest abundance occurred in the top of snow pit with 0―15 cm depth. The results show that the deposition was stronger in summer than in winter.
2.3 Bacterial 16S rRNA gene clone libraries and phylogenetic analysis
Six snow bacterial 16S rRNA gene clone libraries in
ARTICLES
www.scichina.com www.springerlink.com 1479
Fig. 2. Distribution of δ18O, bacterial abundance, snow density, major ion concentration and metal element concentration along depths.
the East Rongbuk glacier, Mt. Everest, were con-structed. In total, 229 clones were subjected to RFLP screening. Negative control did not yield the amplifica-tion products, confirming the credibility of results. The coverage values of all 6 clone libraries were greater than 80%, meaning that the clone numbers screened in each library were statistically enough for the diversity analysis. After RFLP screening, all unique patterns were subjected to sequencing analysis. Totally 35 se-quences were obtained and identified to be normal 16S rRNA gene sequences by the analysis of CHIMERA_ CHECK program. The sequences were affiliated with 9 phylogenetic groups, i.e. α, β, γ-Proteobacteria, Acti-nobacteria, Firmicutes, Cytophaga-Flavobacterium- Bacteroides (CFB), Cyanobacteria, Eukaryotic chloro-plast and TM7 candidate phylum. Table 1 lists 31 gen-era with which all the sequences were affiliated. 9 se-quences could not be classified into the genus level as yet and maybe belong to novel genus. Phylogenetic analysis of all 16S rRNA gene sequences is shown in
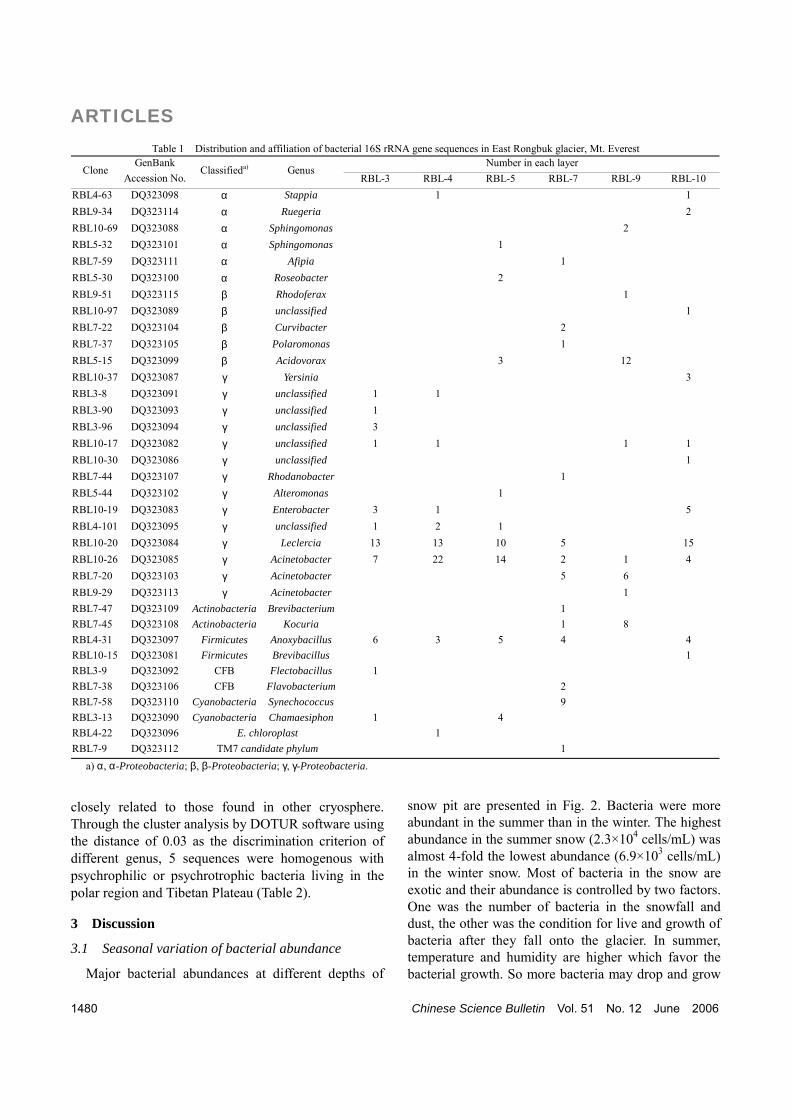
Fig. 3. The dominant group was γ-Proteobacteria ac- counting for 64% of the total clones. Clones that fell into the groups Firmicutes, β-Proteobacteria, Cyano-bacteria, α-Proteobacteria, Actinobacteria and other groups accounted for 10%, 9%, 6%, 4%, 4%, 3% of total clones, respectively. There were 14 genera within γ-Proteobacteria. The clones that belonged to genus Acinetobacter was the most abundant group with 64% of total clones, and the clones of genus Leclercia were the second abundant, accounting for 38% of total. In the 2 genera belonging to Firmicutes, clones of genus Anoxybacillus were dominant with 96% of this group. There were 14 genera in β-Proteobacteria, and 75% clones belonged to genus Acidovorax. The 3 divisions described above all had one or two dominant genera. The Cyanobacteria, α-Proteobacteria and CFB groups had 2, 5 and 2 genera, respectively, whereas the number of clones that belonged to each genus was similar.
Blast research results from GenBank database indi-cated that microbe in the snow in Mt. Everest were
ARTICLES
1480 Chinese Science Bulletin Vol. 51 No. 12 June 2006
Table 1 Distribution and affiliation of bacterial 16S rRNA gene sequences in East Rongbuk glacier, Mt. Everest Number in each layer
Clone GenBank
Accession No. Classifieda) Genus
RBL-3 RBL-4 RBL-5 RBL-7 RBL-9 RBL-10 RBL4-63 DQ323098 α Stappia 1 1 RBL9-34 DQ323114 α Ruegeria 2 RBL10-69 DQ323088 α Sphingomonas 2 RBL5-32 DQ323101 α Sphingomonas 1 RBL7-59 DQ323111 α Afipia 1 RBL5-30 DQ323100 α Roseobacter 2 RBL9-51 DQ323115 β Rhodoferax 1 RBL10-97 DQ323089 β unclassified 1 RBL7-22 DQ323104 β Curvibacter 2 RBL7-37 DQ323105 β Polaromonas 1 RBL5-15 DQ323099 β Acidovorax 3 12 RBL10-37 DQ323087 γ Yersinia 3 RBL3-8 DQ323091 γ unclassified 1 1 RBL3-90 DQ323093 γ unclassified 1 RBL3-96 DQ323094 γ unclassified 3 RBL10-17 DQ323082 γ unclassified 1 1 1 1 RBL10-30 DQ323086 γ unclassified 1 RBL7-44 DQ323107 γ Rhodanobacter 1 RBL5-44 DQ323102 γ Alteromonas 1 RBL10-19 DQ323083 γ Enterobacter 3 1 5 RBL4-101 DQ323095 γ unclassified 1 2 1 RBL10-20 DQ323084 γ Leclercia 13 13 10 5 15 RBL10-26 DQ323085 γ Acinetobacter 7 22 14 2 1 4 RBL7-20 DQ323103 γ Acinetobacter 5 6 RBL9-29 DQ323113 γ Acinetobacter 1 RBL7-47 DQ323109 Actinobacteria Brevibacterium 1 RBL7-45 DQ323108 Actinobacteria Kocuria 1 8 RBL4-31 DQ323097 Firmicutes Anoxybacillus 6 3 5 4 4 RBL10-15 DQ323081 Firmicutes Brevibacillus 1 RBL3-9 DQ323092 CFB Flectobacillus 1 RBL7-38 DQ323106 CFB Flavobacterium 2 RBL7-58 DQ323110 Cyanobacteria Synechococcus 9 RBL3-13 DQ323090 Cyanobacteria Chamaesiphon 1 4 RBL4-22 DQ323096 E. chloroplast 1 RBL7-9 DQ323112 TM7 candidate phylum 1
a) α, α-Proteobacteria; β, β-Proteobacteria; γ, γ-Proteobacteria.
closely related to those found in other cryosphere. Through the cluster analysis by DOTUR software using the distance of 0.03 as the discrimination criterion of different genus, 5 sequences were homogenous with psychrophilic or psychrotrophic bacteria living in the polar region and Tibetan Plateau (Table 2).
3 Discussion
3.1 Seasonal variation of bacterial abundance
Major bacterial abundances at different depths of
snow pit are presented in Fig. 2. Bacteria were more abundant in the summer than in the winter. The highest abundance in the summer snow (2.3×104 cells/mL) was almost 4-fold the lowest abundance (6.9×103 cells/mL) in the winter snow. Most of bacteria in the snow are exotic and their abundance is controlled by two factors. One was the number of bacteria in the snowfall and dust, the other was the condition for live and growth of bacteria after they fall onto the glacier. In summer, temperature and humidity are higher which favor the bacterial growth. So more bacteria may drop and grow

ARTICLES
www.scichina.com www.springerlink.com 1481
Fig. 3. Phylogenetic analysis of bacterial 16S rRNA gene sequences in the East Rongbuk glacier Mt. Everest. The phylogenetic tree was constructed using the neighbor joining algorithm. Archaeal Methanothermus fervidus was used as outgroup. Bootstrap values (100 replicates) greater than 70% are indicated above the branches. Scale bar represents the 5% nucleotide substitution percentage. (a) α, β-Proteobacteria, TM7 candidate phylum; (b) γ-Proteobacteria; (c) Cytophaga-Flavobacterium-Bacteroides (CFB), Actinobacteria, Firmicutes, Cyanobacteria; (d) E. chloroplast.
in glacier during summer and the bacterial population with high abundance in summer snow can form ac-cordingly. Christner’s study[21] showed that the Sajama ice which was deposited ~12000 years ago under cool, wet climate conditions contained more recoverable bacteria than modern deposited ice at the same location during a warmer dryer period. Wet climatic conditions increase the vegetation density and productivity and the
concentration of large airborne particles, such as pollen that transport microorganisms. Such conditions also decrease the desiccation rate[22]. The sources of mois-ture in Mt. Everest are different between winter and summer. During summer, the south Asia monsoon trav-els through India and Nepal with vast vegetation cover, bringing a large amount of vapor from Indian Ocean and transporting more bacteria as well. In situ growth

ARTICLES
1482 Chinese Science Bulletin Vol. 51 No. 12 June 2006
Table 2 Similarity comparison between bacteria in Mt. Everest and other cryosphere Bacteria in Mt. Everest Sequence Cryosphere Similarity RBL9-34 AF479378 Taylor Dome glacier ice Antarctic 98% RBL9-51 AJ440986 Antarctic lake 98% AY315172 Glacier ice and sediment 99% RBL7-37 AY315177 Glacier ice and sediment 99% RBL7-22 AY315181 Glacier ice and sediment 99% RBL3-16 AF076162 Antarctic lake ice cover 98% RBL7-44 AY218596 Antarctic sediment 96% RBL10-19 AY439272 Greenland ice core 94% RBL10-20 AY439272 Greenland ice core 95% RBL5-32 AF395036 Antarctic Vostok lake accreted ice 96% RBL3-13 AF076162 Antarctic lake ice cover 98% RBL7-45 AY439260 Greenland ice core 99% RBL7-59 AY169423 Greenland ice core 96%
also increases the bacteria abundance in summer snow. A study of bacterial flora and biomass in mountain snows from the Tateyama Mountains in Japan showed that the biomass of three psychrotrophic bacteria Cry- obacterium psychrophilum, Variovorax paradoxus and Janthinobacterium lividum increased largely during the melting season from March to October[23]. Microorgan- isms in Malan ice core in Tibetan Plateau were more abundant in the cooler period[13]. But the cool and warm periods in the history are the comparative con- cepts on a long-term scale, which are not in accordance with the temperature variations between summer and winter. Malan is located on the north of the plateau, near the desert. During the cool period, dust can trans- port more bacteria, and therefore result in the bacterial population with high abundance.
3.2 Relationship between bacterial abundance and snow physical-chemical conditions
There was no obvious relationship between bacterial abundance and density of snow. Between the depths of 40―50 cm, the bacteria were most abundant with the snow density of 39 kg/m3, while from 75 to 95 cm where the snow density was 33 kg/m3, the bacteria number was the lowest. Similar bacterial population sturctures (in 50 75 and 95 120 cm) did not corre-spond to similar snow density. Bacterial abundance showed no correlation with chemical characters as well. As shown in Fig. 2, the depth with higher nutrition (higher concentration of 3
4PO − , 4NO− , +4NH ) did not
contain more bacteria. This indicated that nutrition was not the determining factor of bacterial abundance.
3.3 Seasonal variation of bacterial community struc-ture
Bacterial community structure varied in difference snow layers (Table 3). γ-Proteobacteria occurred in 6
layers, Firmicutes and β-Proteobacteria in 5 layers, and other groups in different layers. Our results were simi-lar to that from Malan and Muztag ice cores where bacterial distribution showed layered feature[13,15], and indicated that bacteria could reflect the environmental changes.
Statistical comparison results among different clone libraries are listed in Table 4. The diversity of bacteria in the snow layer deposited in winter (RBL-7) was the highest, and that of summer deposited snow layer (RBL-4) was the lowest. Layer RBL-7 (winter snow) had the least abundance and the highest diversity of bacteria. Layer RBL-9 (summer snow) had the most abundant bacterial population (20689 cells/m3) among all 6 layers, but the Shannon value was much lower than that of RBL-7. Similar to the bacterial abundance, the diversity was also influenced by many factors. The source and transport of moisture and dust were the two most important factors. Bacteria carried by westerlies in winter were distinct from those transported by mon- soon in summer. The adaptive abilities of bacteria to the low temperature, oxygen-deficient and intense ra-diation influenced the kinds of bacteria in snows as well. More abundance and low diversity in summer snow were possibly caused by all of the above reasons.
Community structure of bacteria in snow in Mt. Ev-erest varied seasonally. 9 genera only occurred in win-ter snow, 14 genera only in summer snow, and the two seasons shared 8 common genera. Acinetobacter, Le-clercia and Anoxybacillus were the main common gen-era, accounting for 61% of total clones.
Genus Acinetobacter are ubiquitous organism in soil and water. They have been found at density exceeding 105 cells/L. Many strains have the ability to degrade a wide range of organic compounds. Acinetobacters are capable of degrading many aromatic compounds toxic to most microorganisms, such as salicylate, halogenated
ARTICLES
www.scichina.com www.springerlink.com 1483
Table 3 Bacterial distribution in each snow layers in East Rongbuk glacier, Mt. Everesta)
Act α β γ CFB Cyano Firm Euk TM7 RBL-3 30 1 1 6 RBL-4 1 40 3 1 RBL-5 3 3 26 4 5 RBL-7 2 1 3 13 2 9 4 1 RBL-9 8 2 13 9 RBL-10 3 1 29 5
Percentage in total clones 4.37% 4.37% 8.73% 64.19% 1.31% 6.11% 10.04% 0.44% 0.44%
a) Act, Actinobacteria; α, α-Proteobacteria; β, β-Proteobacteria; γ, γ-Proteobacteria; Cyano, Cyanobacteria; Firm, Firmicutes; Euk, E. chloroplast; TM7, TM7 candidate phylum.
Table 4 Statistical analysis of the 6 bacterial 16S rRNA gene clone libraries RBL-3 RBL-4 RBL-5 RBL-7 RBL-9 RBL-10 Taxa 11 9 9 13 8 11 Individuals 38 45 41 35 32 38 Coverage 84.2% 88.8% 92.6% 82.8% 87.5% 86.8% Dominance 0.1925 0.3314 0.21 0.1347 0.2461 0.2078 Shannon 1.945 1.45 1.805 2.253 1.635 1.942 Simpson 0.8075 0.6686 0.79 0.8653 0.7539 0.7922 Evenness 0.636 0.4739 0.6756 0.7322 0.641 0.6337
aromatics and phenol (the Prokaryotes. http://141.150. 157.117:8080/prokPUB/index.htm). Regarding their abundance in the snow, more acinetobacters were transported to glacier and deposited in snow, and dis-tributed in all 6 layers of winter and summer snow with the largest numbers. This genus was also found in other environments in cryosphere. The clone RBL9-11 was 98% similar to Acinetobacter calcoaceticue which is isolated from Taylor Dome ice core, Antarctic[21] with respect to 16S rRNA gene sequence.
Genus Leclercia, a kind of enteric bacterium, dis-tributes widespread in a number of different habitats such as drinking water, plant, humans sputum, blood, urine and stool. Leclercia adecarboxylata was isolated from subsurface soil in an oily-sludge storage pit at the Digboi oil refinery, India[24] with ability of degrading two- and three-benzene-ring polycyclic aromatic hy-drocarbons. Clones that belonged to genus Leclercia distributed in 5 layers, and the number of clones was the second largest. Clones closely related to organisms that are associated with human feces or animal gastro-intestinal tracts were found in Greenland ice core[25]. The clones of genus Leclercia that exists in snow in Mt. Everest possibly originated from yak feces. Yak is the main transport tool in this region, and can reach the camp (6500 m a.s.l). Yak feces are used as fuel by in-habitants in Tibet after being insolated and stored in the courtyard everywhere. The bacteria capable of degrad-
ing organic compound, such as CFB group, were iso-lated from Greenland ice core, Antarctic soil, Malan ice core and the Tibetan Plateau as well[13,14,25,26]. Xie et al.[27] identified extractable organic compounds from petroleum residues in a snow pit atan altitude of 7000 m a.s.l. in Himalayas. This indicated that the remote middle-upper troposphere was penetrated by anthropo-genic pollutants from India and Middle East. Studies on the same snow pit of our research showed that the poly-cyclic aromatic hydrocarbons were present in every layers, and concentrations were similar to those in Arctic and Alps regions (unpublished, Wang Xiaoping). The dominant species in the snow with degrading ability indicated that microorganism represented the consistent information with other environmental index. Whether it is the response of microbe to the environ-ments and what is the connection between them still need to be deeply studied in the future.
Bacteria of genus Anoxybacillus distributed in 5 lay-ers (10% of total). Genus Anoxybacillus exist in hot spring and are thermophilic and endospore-forming bacterium. In Greenland ice core, clones 1―71 and 1―43 were closely related to thermophile Thermus scoto-ductus, originally isolated from a geothermal heating system in southern Iceland[25], indicating that thermo-phile could exist in extreme cool environments after transport and deposition. Genus Anoxybacillus possibly originated from hot spring in plateau.

ARTICLES
1484 Chinese Science Bulletin Vol. 51 No. 12 June 2006
Besides the above 3 genera, there existed genus Sphingomonas, genus Acidovorax, genus Chamaesi-phon, and an unclassified genus that belongs to γ-Pro-teobacteria in winter and summer snow. Most species of genus Sphingomonas could endure intense radiation and desiccation and survive low-nutrient or starvation conditions. Sphingomonas existed in Antarctic soil, Vostok ice core and plant[29,30]. Acidovorax is phy- topathogenic bacterium, and some strains were isolated from salt marsh[31]. Cyanobacteria is a diverse group of oxygenic photosynthetic prokaryotes, exhibiting versa-tile physiology and wide ecological tolerance that contributes to their competitive success over broad spec-trum of environments, both planktonic and benthic. They are dominant in a broad spectra of terrestrial habitats, including deserts, where they are found as en-doliths in the antarctic dry valleys, cold deserts, hot deserts, tropical rain forests, and mangrove swamps (the Prokaryotes. http://141.150.157.117:8080/prokPUB/
index.htm). There were 14 genera that only occurred in the
summer snow, i.e. genera Stappia and Ruegeria of the subphylum α-Proteobacteria, genus Rhodoferax and an unclassified genus of the subphylum β-Proteobacteria, genera Yersinia, Enterobacter and 5 unclassified genera of the subphylum γ-Proteobacteria, genus Brevibacillus of the phylum Firmicute, genus Flectobacillus in CFB group, and a group affiliated with E. chloroplast. Ruegeria atlantica was a marine strain capable of de-grading dimethylsulfide (DMS) and dimethylsulfonio-propionate (DMSP), an organic sulfur compound pro-duced in abundance by marine algae[32]. Stappia is a new genus including marine species of genus Agrobac-terium in which Stappia stellulata is a type strain. Stap-pia marina was isolated from tidal flat area. All known strains of this genus are able to oxidize carbon monox-ide (CO) and possess the gene for CO dehydrogenase (coxL)[33]. Rhodoferax ferrireducens isolated from sediment in bay is a psychrotolerant, facultatively anaerobic bacterium that oxidizes acetate with the re-duction of Fe (III)[34]. All these genera were related to the marine environment. Because the precipitation mainly come from Bengal and Arabian during summer in Mt. Everest, bacteria attached to the moisture exhibit the “marine character”.
E. chloroplast originated from algae[35], which means that algae exist in the snow of this region. Yo-shitaka et al.[11] measured the number of snow algae in glacier (5430 m a.s.l) at the south slope of Himalayas
and the abundance reached 0.9×103 cells/mL. However, snow algae in the north slope, especially in the snow at col (6350 m a.s.l.), is waiting for further studies. Clone RBL4-22 is the first report of 16S rRNA gene sequence related to snow algae in this region.
Concerning the season-specific species, we found that nine genera only existed in winter. They were gen-era Afipia and Roseobacter (within α-Proteobacteria), genera Curvibacter and Ploaromonas (within β-Proteo- bacteria), genera Rhodanobacter and Alteromonas (within γ-Proteobacteria), genus Brevibacterium (with- in Actinobacteria), genus Flavobacterium (within CFB group), genus Synechococcus (within Cyanobacteria), and candidate phylum TM7. Species of genus Afipia was mainly isolated from lymph nodes of patients with cat scratch disease, and also obtained from hospital sewage (the Prokaryotes. http://141.150.157.117:8080/ prokPUB/index.htm)[36]. Members of the Roseobacter group are ubiquitous in marine environments. Investi-gation of Roseobacter group in world ocean showed they were highest abundance in the polar regions[37]. Polaromonas vacuolata is a psychrophilic, marine, gas vacuolate bacterium isolated from Pamler, Antarc-tica[38]. Bacteria of genus Alteromonas are heterotrophs, chemorganotrophs, psychrotrophs, slightly or moder-ately halophilic. Members of Alteromonas-like bacteria are isolated from a variety of biotopes, i.e. sediments, Antarctic seawater, sea ice, deepsea hydrothermal vent, etc. (the Prokaryotes. http://141.150.157.117:8080/prok- PUB/index.htm)[39]. Candidate division TM7 is one of several newly described bacterial divisions exclusively characterized by environmental sequence data. TM7 partial 16S rDNA sequences were obtained from PCR clonal studies of a peat bog (one sequence), a mature forest soil (two sequences), and two sequencing batch reactor sludges (seven sequences)[40]. The sequences obtained from the snow in this work added more mem-bers to this group. Genus Flavobacterium existed in Malan and Guliya ice core, Tibetan Plateau[13,21]. Mi-crobe in the winter snow correlated with both marine and terrestrial environment. It was possible that they were westerly transported to the middle of Himalayas with terrestrial dust from aridity and semi-aridity region and marine moisture from North Atlantic during winter. Multiplex source resulted in the high diversity of bacte-ria in winter snow and strong relatedness to both ma-rine and terrestrial environments. Bacteria in winter snow exhibited more cold-resistant character than those in summer. It indicated that bacteria were selectively

ARTICLES
www.scichina.com www.springerlink.com 1485
eliminated during transport process, and only those that could endure low temperature survived and were de-posited in the glacier.
4 Conclusion Abundant and diverse bacteria, correlating with ma-
rine and terrestrial environments, exist in the snow in Mt. Everest. Population and community structure of bacteria vary seasonally. Bacteria in summer snow are more abundant than in winter snow. Diversity of snow bacteria was high with γ-Proteobacteria as the domi-nant group. Total 35 partial 16S rRNA gene fragments were sequenced, which fell into the groups α, β, γ- Pro-teobacteria, Actinobacteria, Firmicutes, CFB, Cya- nobacteria, E. chloroplast, and TM7 candidate phylum. Bacterial community structures in winter and summer snow are diverse. There are both common species and particular species of each season. Acinetobacter and Leclercia are dominant genera, possibly the indigenous bacteria of the Tibetan Plateau. More microbes in summer snow originated from marine environment, whereas bacteria in winter snow were of more multi-plex environmental origins and showed more cold- resistant features. On the whole, community structure and population of bacteria do not show close related-ness with environment variables. Physical characters and concentrations of nutrition are not the determining factor of bacterial abundance and diversity. Seasonal variation of abundance and diversity of microbes are also influenced by the seasonal feature of climate and the atmospheric circulation in Mt. Everest.
Acknowledgements This work was supported by the Min-istry of Science and Technology of the People’s Republic of China (Grant No. 2005CB422004), the National Natural Science Foundation of China (Grant Nos. 40121101 & 40401054) and the Innovation Program of the Chinese Academy of Sciences (Grant No. KZCX3-SW-339).
References
1 Qin D, Mayewski P A, Wake C P, et al. Evidence for recent climate change from ice cores in the central Himalayas. Ann Glaciol, 2000, 31: 153―158
2 Qin D, Hou S, Zhang D, et al. Preliminary results from the chemi-cal records of an 80.4 m ice core recovered from East Rongbuk Glacier, Qomolangma (Mount Everest). Ann Glaciol, 2002, 35: 278―284
3 Reng J, Qin D, Kang S, et al. Glacier variations and climate warm-ing and drying in the central Himalayas. Chin Sci Bull, 49(1): 65―69
4 Hou S, Qin D, Wake C P, et al. Changes in net accumulation from ice core records in Mt. Everest and its climatological significance. Chin Sci Bull,1999, 44(21): 2336―2342
5 Kang S, Qin D, Mayewski P A, et al. Climatic and environmental records from the Far East Rongbuk ice core, Mt. Qomolangma (Everest). Episodes, 2001, 24(3): 176―181
6 Kang S, Mayewski P A, Qin D, et al. Twentieth century increase of atmospheric ammonia recorded in Mt. Everest ice core. J Geophys Res, 2002, 107(D21), 45957
7 Expedition of Qinghai-Tibet Plateau, Chinese Academy of Science. Tibet Algae (in Chinese). Beijing: Science press, 1992. 509
8 Expedition of Qinghai-Tibet Plateau, Chinese Academy of Science. Tibet Flugi (in Chinese). Beijing: Science Press, 1992. 226
9 Xiong K, Tang X, Xie S. Yeast isolated from snow at Tibet. J Mi-crobio (in Chinese), 1999, 19(2): 58―62
10 Baghela V S, Tripathia R D, Ramtekeb P W, et al. Psychrotrophic proteolytic bacteria from cold environment of Gangotri glacier, Western Himalaya, India. Enzyme Microb Technol, 2005, 36: 654―659
11 Yoshitaka Y, Shiro K, Shuji O. A community of snow algae on a Himalayan glacier: Change of algal biomass and community struc-ture with altitude. Arct Alp Res, 1997, 29(1): 126―137
12 Yao T, Xiang S, Zhang X, et al. Microbiological characeteristics recorded by Manlan and Puruogangri ice core. Quaternary Sci (in Chinese), 2003, 23: 193―199
13 Xiang S, Yao T, An L, et al. Change of bacterial community in the Malan Ice Core and its relation to climate and environment. Chin Sci Bull, 2004, 49(17): 1869―1875
14 Zhang X, Ma X, Yao T, et al. Diversity of 16S rDNA and environ-mental factor influencing microorganisms in Malan ice core. Chin Sci Bull, 2003, 48(11): 1146―1150
15 Xiang S, Yao T, An L, et al. 16S rRNA sequences and differences in bacteria isolated from the Muztag Ata Glacier at increasing depths. Sci China Ser D-Earth Sci, 2005, 35(3): 252―262
16 Kemp P F, Aller J Y. Bacterial diversity in aquatic and other envi-ronments: What 16S rDNA libraries can tell us. FEMS Microbiol Ecol, 2004, 47:161―177
17 Maidak B L, Cole J R, Lilburn T G, et al. The RDP-II (Ribosomal Database Project). Nucleic Acids Res, 2001, 29: 173―174
18 Tian L, Yao T, Schuster P F, et al. δ18O concentrations in recent precipitation and ice cores on the Tibetan Plateau. J Geophys Res, 2003, 108(D9), 4293
19 Tian L, Yao T, Zhang X. δ18O in precipitation and moisture sources upon the Tibetan Plateau. Cryosphere, 1996, 21, 33―39
20 Tian L, Yao T, White J W, et al. Westerly moisture transport to the middle of Himalayas revealed from the high deuterium excess. Chin Sci Bull, 2005, 50(10): 1026―1030
21 Christner B C, Mosley-Thompson E, Thompson L G, et al. Recov-ery and identification of viable bacteria immured in glacial ice.

ARTICLES
1486 Chinese Science Bulletin Vol. 51 No. 12 June 2006
Icarus, 2000, 144(2): 479―485 22 Liu K, Yao Z, Thompson L G. A pollen record of Holocene climate
changes from the Dunde ice cap, Qinghai-Tibetan Plateau. Geology, 1998, 26: 135―138
23 Takabiro S, Koji M, Kazunari U, et al. Seasonal change in bacterial flora and biomass in mountain snow from the Tateyama Mountains, Japan, analyzed by 16S rRNA gene sequencing and Real-Time PCR. Appl Environ Microb, 2005, 71(1): 123―130
24 Sarma P M, Bhattacharya D, Krishnan S. Degradation of polycyclic aromatic hydrocarbons by a newly discovered enteric bacterium, Le-clercia adecarboxylata. Appl Environ Microb, 2004, 3163―3166
25 Peter P S, Miteva V I, Brenchley J E. Phylogenetic analysis of an-aerobic psychrophilic enrichment cultures obtained from a Greenl- and glacier ice core. Appl Environ Microb, 2003, 69(4): 2153―2160
26 David J S, Jackie M A, Caroline E B, et al. Hydrocarbon contami-nation changes the bacterial diversity of soil from around Scott Base, Antarctica. FEMS Microbiol Ecol, 2005, 53: 141―155
27 Xie S, Yao T, Kang S, et al. Geochemical analyses of a Himalayan snowpit profile: Implications for atmospheric pollution and climate. Org Geochem, 2000, 31: 15―23
28 Ali O B, Sabriye D, Zihni D. Anoxybacillus gonensis sp. nov., a moderately thermophilic, xylose-utilizing, endospore-forming bac-terium. Int J Syst Evol Micr, 2003, 53: 1315―1320
29 Takeuchi M, Hamana K, Hiraishi A. Proposal of the genus Sphingo-monas sensu stricto and three new genera, Sphingobium Novosphin-gobium and Sphingopyxis on the basis of phylogenetic and chemo-taxonomic analyses. Int J Syst Evol Micr, 2001, 51: 1405―1417
30 Hans J B, Ewald B M, Denner S B. Sphingomonas aurantiaca sp. nov., Sphingomonas aerolata sp. nov. and Sphingomonas faeni sp. nov. air- and dustborne and Antarctic, orangepigmented, psychro-tolerant bacteria, and emended description of the genus Sphingo-monas. Int J Syst Evol Micr, 2003, 53: 1253―1260
31 Gardan L, Dauga C, Prior P. Acidovorax anthurii sp. nov., a new
phytopathogenic bacterium which causes bacterial leaf-spot of an-thurium. Int J Syst Evol Micr, 2000, 50: 235―246
32 Moran M A, González J M, Kiene R P. Linking a bacterial taxon to sulfur cycling in the sea: Studies of the marine Roseobacter group. Geomicrobiology Journal, 2003, 20: 375―388
33 Byung-Chun K, Ja Ryeong P, Jin-Woo B. Stappia marina sp. nov., a marine bacterium isolated from the Yellow Sea. Int J Syst Evol Micr, 2006, 56:75―79
34 Finneran K T, Johnsen C V, Lovley D R. Rhodeferax ferrireducens sp. nov., a psychrotolerant, facultatively anaerobic bacterium that oxidizes acetate with the reduction of FE(III). Int J Syst Evol Micr, 2003, 53: 669―673
35 Crump B C, Hobbie J E. Synchrony and seasonality in bacterio-plankton communities of two temperate rivers. Limnol Oceanogr, 2005, 50 (6) : 1718―1729
36 Brenner D J, Hollis D G, Moss C W, et al. Proposal of Afipia gen. nov., with Afipia felis sp. nov. (formerly the cat scratch disease ba-cillus), Afipia clevelandensis sp. nov. (formerly the Cleveland Clinic Foundation strain), Afipia broomeae sp. nov., and three un-named genospecies. J Clin Microbiol, 1991, 29: 2450―2460
37 Natascha S, Meinhard S, Thorsten B. A newly discovered Roseo-bacter cluster in temperate and polar oceans. Nature, 2004, 427(29): 445―448
38 Irgens R L, Gosink J J, Staley J. Polaromonas vacuolata gen. nov., sp. nov., a psychrophilic, marine, gas vacuolate bacterium from Antarctica. Int J Syst Evol Micr, 1996, 46(3): 822―826
39 Raguenes G, Peres A, Ruimy R, et al. Alteromonas infernus sp. nov., a new polysaccharide producing bacterium isolated from a deepsea hydrothermal vent. J Appl Bacteriol, 1997, 82: 422―430
40 Hugenholtz P, Tyson G W, Webb R I. Investigation of Candidate Division TM7, a recently recognized major lineage of the domain bacteria with no known pure-culture representatives. Appl Environ Microb, 2001, 67: 411―419




















