
Seasonal variation of fatty acids from drip water in Heshang Cave, central China
7
Seasonal variation of fatty acids from drip water in Heshang Cave, central China Xiuli Li a , Canfa Wang b , Junhua Huang b , Chaoyong Hu a , Shucheng Xie a,⇑ a Key Laboratory of Biogeology and Environmental Geology, Ministry of Education, China University of Geosciences, Wuhan 430074, China b State Key Laboratory of Geological Processes and Mineral Resources, China University of Geosciences, Wuhan 430074, China article info Article history: Received 20 April 2010 Accepted 22 December 2010 Available online 28 December 2010 Editorial handled by R. Fuge abstract In order to investigate how lipids in cave water respond to seasonal climate change, drip water samples were collected from 2006 to 2008 in Heshang Cave, central China for fatty acid analysis. These lipids are abundant in the drip water. Their compositions are dominated by lower-molecular-weight nC16:0, nC18:0 and nC14:0 acids, together with mono-unsaturated nC18:1, nC16:1 and nC14:1. Analysis of one water sample revealed marked differences between the dissolved and particulate fractions. The dissolved fraction contains total fatty acids one order of magnitude higher than that of the particulate fraction. The distributional patterns of the fatty acids suggest that microbes living in the overlying soils and/or ground- water system contribute most fatty acids to the drip water. This 2-a monitoring experiment reveals that the abundance of mono-unsaturated fatty acids relative to the saturated homologues (nC16:1/nC16:0 and nC18:1/nC18:0) relate inversely to the changes of synchronous external air temperature. Higher values occur under cold conditions (winter/spring), while lower values appear in warm intervals (summer). Fur- ther studies are needed to elucidate the dynamic processes by which the external temperature affects fatty acids in drip water and to confirm the potential application of fatty acid ratios such as nC16:1/ nC16:0 and nC18:1/nC18:0 in paleotemperature reconstructions. Ó 2010 Elsevier Ltd. All rights reserved. 1. Introduction Stalagmites are one of the most important terrestrial archives for paleoenvironmental reconstructions (e.g. Hu et al., 2008b; Wang et al., 2008). Most previous studies have focused on the car- bonate O isotopic signals, which are interpreted as the indicators of monsoon strength (e.g. Dykoski et al., 2005; Wang et al., 2005, 2008). Recently, lipid biomarkers trapped in the stalagmite have also attracted much attention (Xie et al., 2003, 2005; Blyth et al., 2006, 2007; Huang et al., 2008). These trace molecules can provide information about the source and the diagenetic processes affect- ing organic matter in speleothems. However, compared to studies of other types of sediments (e.g. peat, lacustrine and marine sedi- ments), reports on lipid biomarkers in stalagmites are still quite limited. This situation exists partly due to the low abundance and complex origin of lipids in stalagmites (Blyth et al., 2006; Huang et al., 2008). In addition, research is needed to elucidate how lipid biomarkers are transported in groundwater and then preserved in stalagmites and how lipid distributions respond to modern climate changes. Process monitoring in modern caves is a perfect approach to deal with the above problems. To date, researchers have paid much attention to the physical processes of modern caves (e.g. Genty and Deflandre, 1998; McDonald et al., 2004, 2007; Baldini et al., 2006; Hu et al., 2008a). Some studies also describe the bulk organic matter (e.g. Baker et al., 1997; Baker and Barnes, 1998; Baker and Genty, 1999; Birdwell and Engel, 2010), but no study concerning lipids in drip water has yet been reported. Here results are presented of a nearly 2-a monitoring of fatty acids in drip water collected from Heshang Cave, Hubei Province, central China, with the aim of investigating how lipids in drip water respond to seasonal climate change. 2. Materials and methods Heshang Cave (30°27 0 N, 110°25 0 E, 294 m a.s.l.) is located on the south bank of the Qing River, which is a major branch of the Yangtze River in Hubei Province. Annual climate in this region is strongly monsoonal. In boreal summer, the average temperature is 28 °C, with rainfall peaking in July at 270 mm month 1 (Hu et al., 2008b). In boreal winter, temperature drops below 10 °C, and rainfall is less than 50 mm month 1 . This cave is overlain by 400 m of Cambrian dolomite. For a detailed description of Heshang cave, please refer to Hu et al. (2008a). The drip water was collected just under the former growing site of the HS-4 stalagmite studied by Hu et al. (2008b). Eleven water samples were collected during a 2-a period (November 2006– November 2008). Water was collected in a 4 L glass container. Concentrated HCl was added to restrain microbial activities during 0883-2927/$ - see front matter Ó 2010 Elsevier Ltd. All rights reserved. doi:10.1016/j.apgeochem.2010.12.007 ⇑ Corresponding author. Fax: +86 27 67885096. E-mail address: [email protected] (S. Xie). Applied Geochemistry 26 (2011) 341–347 Contents lists available at ScienceDirect Applied Geochemistry journal homepage: www.elsevier.com/locate/apgeochem
-
Upload
independent -
Category
Documents
-
view
7 -
download
0
Transcript of Seasonal variation of fatty acids from drip water in Heshang Cave, central China

Applied Geochemistry 26 (2011) 341–347
Contents lists available at ScienceDirect
Applied Geochemistry
journal homepage: www.elsevier .com/ locate/apgeochem
Seasonal variation of fatty acids from drip water in Heshang Cave, central China
Xiuli Li a, Canfa Wang b, Junhua Huang b, Chaoyong Hu a, Shucheng Xie a,⇑a Key Laboratory of Biogeology and Environmental Geology, Ministry of Education, China University of Geosciences, Wuhan 430074, Chinab State Key Laboratory of Geological Processes and Mineral Resources, China University of Geosciences, Wuhan 430074, China
a r t i c l e i n f o
Article history:Received 20 April 2010Accepted 22 December 2010Available online 28 December 2010Editorial handled by R. Fuge
0883-2927/$ - see front matter � 2010 Elsevier Ltd. Adoi:10.1016/j.apgeochem.2010.12.007
⇑ Corresponding author. Fax: +86 27 67885096.E-mail address: [email protected] (S. Xie).
a b s t r a c t
In order to investigate how lipids in cave water respond to seasonal climate change, drip water sampleswere collected from 2006 to 2008 in Heshang Cave, central China for fatty acid analysis. These lipids areabundant in the drip water. Their compositions are dominated by lower-molecular-weight nC16:0,nC18:0 and nC14:0 acids, together with mono-unsaturated nC18:1, nC16:1 and nC14:1. Analysis of onewater sample revealed marked differences between the dissolved and particulate fractions. The dissolvedfraction contains total fatty acids one order of magnitude higher than that of the particulate fraction. Thedistributional patterns of the fatty acids suggest that microbes living in the overlying soils and/or ground-water system contribute most fatty acids to the drip water. This 2-a monitoring experiment reveals thatthe abundance of mono-unsaturated fatty acids relative to the saturated homologues (nC16:1/nC16:0 andnC18:1/nC18:0) relate inversely to the changes of synchronous external air temperature. Higher valuesoccur under cold conditions (winter/spring), while lower values appear in warm intervals (summer). Fur-ther studies are needed to elucidate the dynamic processes by which the external temperature affectsfatty acids in drip water and to confirm the potential application of fatty acid ratios such as nC16:1/nC16:0 and nC18:1/nC18:0 in paleotemperature reconstructions.
� 2010 Elsevier Ltd. All rights reserved.
1. Introduction
Stalagmites are one of the most important terrestrial archivesfor paleoenvironmental reconstructions (e.g. Hu et al., 2008b;Wang et al., 2008). Most previous studies have focused on the car-bonate O isotopic signals, which are interpreted as the indicators ofmonsoon strength (e.g. Dykoski et al., 2005; Wang et al., 2005,2008). Recently, lipid biomarkers trapped in the stalagmite havealso attracted much attention (Xie et al., 2003, 2005; Blyth et al.,2006, 2007; Huang et al., 2008). These trace molecules can provideinformation about the source and the diagenetic processes affect-ing organic matter in speleothems. However, compared to studiesof other types of sediments (e.g. peat, lacustrine and marine sedi-ments), reports on lipid biomarkers in stalagmites are still quitelimited. This situation exists partly due to the low abundanceand complex origin of lipids in stalagmites (Blyth et al., 2006;Huang et al., 2008). In addition, research is needed to elucidatehow lipid biomarkers are transported in groundwater and thenpreserved in stalagmites and how lipid distributions respond tomodern climate changes. Process monitoring in modern caves isa perfect approach to deal with the above problems. To date,researchers have paid much attention to the physical processesof modern caves (e.g. Genty and Deflandre, 1998; McDonald
ll rights reserved.
et al., 2004, 2007; Baldini et al., 2006; Hu et al., 2008a). Somestudies also describe the bulk organic matter (e.g. Baker et al.,1997; Baker and Barnes, 1998; Baker and Genty, 1999; Birdwelland Engel, 2010), but no study concerning lipids in drip waterhas yet been reported.
Here results are presented of a nearly 2-a monitoring of fattyacids in drip water collected from Heshang Cave, Hubei Province,central China, with the aim of investigating how lipids in dripwater respond to seasonal climate change.
2. Materials and methods
Heshang Cave (30�270N, 110�250E, 294 m a.s.l.) is located on thesouth bank of the Qing River, which is a major branch of theYangtze River in Hubei Province. Annual climate in this region isstrongly monsoonal. In boreal summer, the average temperatureis 28 �C, with rainfall peaking in July at 270 mm month�1 (Huet al., 2008b). In boreal winter, temperature drops below 10 �C,and rainfall is less than 50 mm month�1. This cave is overlain by�400 m of Cambrian dolomite. For a detailed description ofHeshang cave, please refer to Hu et al. (2008a).
The drip water was collected just under the former growing siteof the HS-4 stalagmite studied by Hu et al. (2008b). Eleven watersamples were collected during a 2-a period (November 2006–November 2008). Water was collected in a 4 L glass container.Concentrated HCl was added to restrain microbial activities during

Fig. 1. The m/z 117 mass chromatograph of water sample collected in November 2008. i: iso-; a: anteiso-.
Fig. 2. Histograms of lower-molecular-weight fatty acids in water samples collected during warm and cold periods.
342 X. Li et al. / Applied Geochemistry 26 (2011) 341–347

X. Li et al. / Applied Geochemistry 26 (2011) 341–347 343
the field trip. To extract lipids, the water sample was passedthrough a C18 SPE column (Supelco, USA). The organics retainedin the column were then eluted by dichloromethane (Absolv,TEDIA, USA) and concentrated using rotary evaporation underreduced pressure. In order to minimize contamination, all glass-ware was soaked in a decontamination solution, rinsed with ultrapurified water, and heated for 12 h at 500 �C.
After being derivatized with BSTFA (N,O-bis(trimethylsilyl)trifluoacetamide), the extracted fatty acids were analyzed on aHewlett Packard 6890 gas chromatograph coupled to a HewlettPackard 5973 mass selective detector (GC–MS). The oven temper-ature was ramped from 70 �C to 210 �C at a rate of 10 �C min�1, andthen ramped to 300 �C (held 30 min) at a rate of 3 �C min�1. Theindividual components of the lipid fractions were identified by acombination of GC relative retention times and mass spectral data(NIST02).
The conventional shorthand notation is employed to describefatty acid compounds in the text as follows, the prefix a indicatesanteiso- (i: iso-; n: normal), the number before the colon specifiesthe number of C atoms, and the number after the colon gives thenumber of double bonds.
3. Results and discussion
3.1. Distributions of fatty acids in cave drip water
In all water samples, fatty acid distributions include straight-chain (nC10:0–nC30:0), branched (iC14:0–iC26:0, aC15:0 andaC17:0) and a mono-unsaturated series (nC14:1–nC18:1; Figs. 1and 2, Table 1). The short-chain homologues (nC10–nC20), domi-nated by nC16:0, show a strong predominance over their long-chaincounterparts (nC21–nC30). nC22:0 and nC24:0 are the most
Table 1Quantitative data (ng/L) of short-chain fatty acids in drip water samples collected from H
2006 2007
November January April May July September
C10:0 83.1 6.3 22.44 0.9 0.8 0.6C11:0 25.4 2.4 5.2 0.7 0.4 ndC12:0 447.6 57.4 56.4 29.9 2.6 0.7C13:0 42.6 4.4 15.4 4.9 0.6 ndiC14:0 91.1 11.9 24.2 11.9 0.9 0.3C14:1 81.6 7.3 20.0 1.9 0.2 0.1C14:0 1150.3 168.6 232.1 159.1 9.5 3.1iC15:0 23.9 4.2 10.2 11.4 0.9 0.1aC15:0 76.6 11.5 41.1 1.4 1.1 ndC15:1 41.4 6.1 16.1 nd 0.2 ndC15:0 525.5 75.8 134.3 54.7 4.3 1.9iC16:0 69.4 16.6 15.3 9.6 0.9 0.5C16:1 463.6 90.3 nd 12.6 1.8 1.0C16:1 39.3 21.5 106.2 3.5 0.7 0.6C16:0 7941.3 569.3 561.6 353.9 35.5 15.4iC17:0 21.0 5.7 9.6 2.2 0.3 0.4aC17:0 17.7 9.6 5.7 nd 0.3 ndC17:1 65.8 14.8 13.4 nd nd ndC17:0 339.9 18.1 35.8 11.2 1.3 0.8C18:2 32.4 nd nd nd nd ndC18:1 373.8 96.0 41.7 8.7 2.7 1.4C18:1 14.7 13.6 4.2 2.3 0.6 0.3C18:0 1680.9 99.3 152.0 91.6 10.3 6.0C19:0 40.0 nd nd nd 0.2 ndC20:0 114.3 nd 12.9 nd 0.8 ndC16:1/C16:0 0.1 0.2 0.2 0.0 0.1 0.1C18:1/C18:0 0.3 1.1 0.3 0.1 0.3 0.3C14:1/C14:0 0.1 0.0 0.1 0.0 0.0 0.0Unsaturated% 8.4 19.2 13.8 3.7 7.5 9.7Branched% 3.3 7.2 10.7 6.0 7.1 6.2
a Dissolved fraction.b Particulate fraction.
abundant long-chain homologues (Fig. 1). The dominance oflower-molecular-weight homologues suggests that microbes arethe major sources of fatty acids in the cave drip water (Meyersand Ishiwatari, 1993; Johns et al., 1994; Zegouagh et al., 1996).The distributions of fatty acids in drip water are similar to thosein the overlying soils and the stalagmites of Heshang Cave (Xieet al., 2005; Huang et al., 2007, 2008). In all these samples, lower-molecular-weight fatty acids are the dominant components. Thissimilarity suggests that the overlying soil is an important sourcefor fatty acids in drip water and stalagmites in Heshang Cave.
The percentages of total mono-unsaturated fatty acids (mainlynC16:1, nC18:1 and nC14:1 in relative abundance) vary between3.9% and 27.5%, averaging 12.3% (Table 1). The higher values occurin winter/spring water samples (>10%; January, April, November,December 2007 and November 2008), while lower values occurin early summer (May, July 2007 and July 2008). Straight-chainmono-unsaturated fatty acids are abundant in microalgae(Zegouagh et al., 1996; Zou et al., 2006). Hence, microalgae livingin the percolating water may contribute fatty acids to the dripwater. In addition, odd-numbered mono-unsaturated fattyacids (C15:1 and C17:1) occur in water samples (Fig. 1). Thesecompounds are abundant in SO4-reducing bacteria, especiallyDesulfobulbus and Desulfovibrio (Parkes, 1987; Zegouagh et al.,1996; Rontani and Volkman, 2005). The occurrence of odd-numbered mono-unsaturated fatty acids in the drip water possiblyoriginates from the soil SO4-reducing bacteria.
3.2. Distribution of fatty acids in dissolved and particulate fractions
The water sample collected in July 2008 was separated intoparticulate (>0.7 lm) and dissolved (<0.7 lm) fractions usinga Whatman GF/F glass microfiber binder free filter. Although
eshang Cave.
2008
November December May July (Da) July (Pb) November
2.1 1.4 0.2 53.6 3.2 49.60.5 0.1 nd 28.9 nd 8.31.0 5.9 0.8 165.8 8.3 70. 9nd 3.7 0.3 25.4 1.4 17.80.6 9.5 0.6 53.4 4.6 42.31.6 11.0 0.2 55.5 3.2 23.76.0 74.2 6.8 749.6 40.2 293.50.3 3.0 0.2 23.1 1.8 9.51.2 14.2 0.7 82.4 5.2 36.50.6 7.8 0.1 23.3 2.3 16.13.6 50.6 3.1 293.5 24.5 172.30.5 5.9 0.3 62.0 6.1 34.8nd nd nd 304.2 32.8 142.25.5 87.3 0.9 nd 3.9 8.720.7 184.4 17.6 2734.9 214.1 821.10.3 2.9 nd 41.1 4.0 3.30.2 3.0 nd 9.3 0.7 9.40.9 12.2 0.1 49.2 5.9 16.12.5 9.4 0.4 84.9 8.1 45.7nd nd nd nd 1.6 4.50.2 2.5 nd 93.0 36.4 86.323 31.0 nd nd 1.9 6.09.2 20.4 1.8 1027.6 50.3 246.6nd 1.5 nd 7.1 1.4 6.3nd 2.8 nd 20.8 4.0 12.80.3 0.5 0.1 0.1 0.2 0.20.3 1.6 0.0 0.1 0.8 0.40.3 0.1 0.0 0.1 0.1 0.118.3 27.5 3.9 8.6 17.8 14.56.0 11.4 6.7 4.6 4.6 9.1
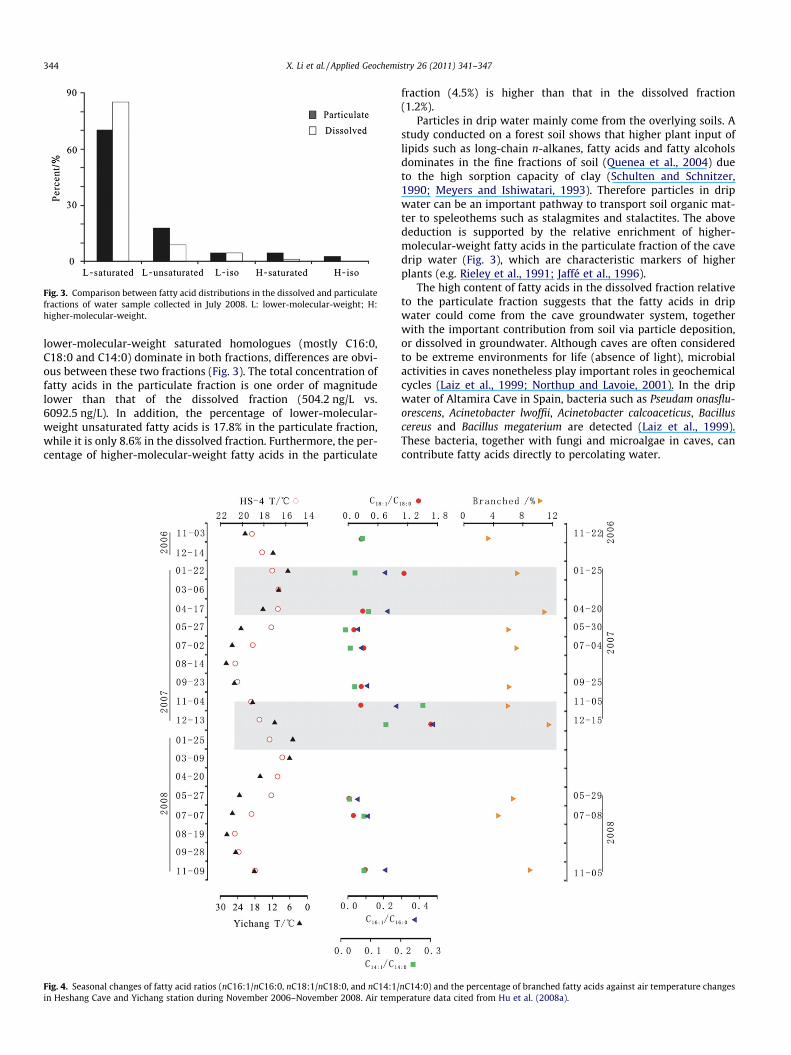
Fig. 3. Comparison between fatty acid distributions in the dissolved and particulatefractions of water sample collected in July 2008. L: lower-molecular-weight; H:higher-molecular-weight.
344 X. Li et al. / Applied Geochemistry 26 (2011) 341–347
lower-molecular-weight saturated homologues (mostly C16:0,C18:0 and C14:0) dominate in both fractions, differences are obvi-ous between these two fractions (Fig. 3). The total concentration offatty acids in the particulate fraction is one order of magnitudelower than that of the dissolved fraction (504.2 ng/L vs.6092.5 ng/L). In addition, the percentage of lower-molecular-weight unsaturated fatty acids is 17.8% in the particulate fraction,while it is only 8.6% in the dissolved fraction. Furthermore, the per-centage of higher-molecular-weight fatty acids in the particulate
Fig. 4. Seasonal changes of fatty acid ratios (nC16:1/nC16:0, nC18:1/nC18:0, and nC14:1/in Heshang Cave and Yichang station during November 2006–November 2008. Air temp
fraction (4.5%) is higher than that in the dissolved fraction(1.2%).
Particles in drip water mainly come from the overlying soils. Astudy conducted on a forest soil shows that higher plant input oflipids such as long-chain n-alkanes, fatty acids and fatty alcoholsdominates in the fine fractions of soil (Quenea et al., 2004) dueto the high sorption capacity of clay (Schulten and Schnitzer,1990; Meyers and Ishiwatari, 1993). Therefore particles in dripwater can be an important pathway to transport soil organic mat-ter to speleothems such as stalagmites and stalactites. The abovededuction is supported by the relative enrichment of higher-molecular-weight fatty acids in the particulate fraction of the cavedrip water (Fig. 3), which are characteristic markers of higherplants (e.g. Rieley et al., 1991; Jaffé et al., 1996).
The high content of fatty acids in the dissolved fraction relativeto the particulate fraction suggests that the fatty acids in dripwater could come from the cave groundwater system, togetherwith the important contribution from soil via particle deposition,or dissolved in groundwater. Although caves are often consideredto be extreme environments for life (absence of light), microbialactivities in caves nonetheless play important roles in geochemicalcycles (Laiz et al., 1999; Northup and Lavoie, 2001). In the dripwater of Altamira Cave in Spain, bacteria such as Pseudam onasflu-orescens, Acinetobacter lwoffii, Acinetobacter calcoaceticus, Bacilluscereus and Bacillus megaterium are detected (Laiz et al., 1999).These bacteria, together with fungi and microalgae in caves, cancontribute fatty acids directly to percolating water.
nC14:0) and the percentage of branched fatty acids against air temperature changeserature data cited from Hu et al. (2008a).

Fig. 5. Correlation analysis between fatty acid ratios/proportion and internal/external air temperature records (HS-4/Yichang station). Air temperature data cited from Huet al. (2008a).
X. Li et al. / Applied Geochemistry 26 (2011) 341–347 345
3.3. Seasonal variations of fatty acid ratios
Fatty acid ratios such as nC16:1/nC16:0 and nC18:1/nC18:0have been used for paleoenvironmental reconstructions (e.g.Kawamura et al., 1987; Xie et al., 2005; Zhou et al., 2005). The 2-a monitoring experiment reveals that these mono-unsaturatedfatty acid ratios exhibit obvious seasonal variations. In samples col-lected in the cold winter and spring (e.g. January and November2007), these molecules are present in relatively high amounts,and the ratios nC16:1/nC16:0 and nC18:1/nC18:0 are greater than0.2 and 0.6, respectively (Fig. 4). In samples collected in thesummer, these two ratios fall below 0.2 and 0.6 (e.g. May 2007and 2008), respectively. Over the 21 months, the ratios nC16:1/nC16:0 and nC18:1/nC18:0 shift quite similarly. The difference isthat higher values of ratio nC16:1/nC16:0 occur over a widerinterval. The nC14:1/nC14:0 ratio shifts to generally follow thenC16:1/nC16:0 and nC18:1/nC18:0 ratios (Fig. 4). However, the
highest value of nC14:1/nC14:0 occurs in November 2007, aheadof the maximum of the ratios nC16:1/nC16:0 and nC18:1/nC18:0.In addition, values of nC14:1/nC14:0 in spring of 2007 are only alittle higher than the time-adjacent water samples. The above dif-ferences among three ratios may result from source differences ofthese mono-unsaturated homologues in the drip water of HeshangCave. Short-chain branched fatty acids are characteristic biomark-ers of bacteria (see Kaneda, 1991 for a review), and their composi-tions also show a seasonal variation generally comparable with thethree ratios of mono-unsaturated fatty acids (Fig. 4).
The correlation analysis is conducted to discriminate the rela-tionship between unsaturated fatty acid ratios and the internaland external air temperature. Ratios of nC16:1/nC16:0 andnC18:1/nC18:0 both show a strong negative relationship (R2 = 0.5–0.6) with the external temperature recorded in Yichang meteoricstation, whereas these two ratios display no relationship with inter-nal temperature recorded in HS-4 site (Fig. 5). A 3-a monitoring in

346 X. Li et al. / Applied Geochemistry 26 (2011) 341–347
Heshang Cave conducted by Hu et al. (2008a) demonstrated that thetemperature immediately outside Heshang Cave closely followedthat of the nearest meterological station at Changyang County, Yi-chang City and the internal temperature lagged behind the externaltemperature by about 10 days. The above correlation analysis re-sults suggest that in situ cave microbes are probably not the predom-inant source for C16 and C18 acids in drip water collected in HeshangCave. Other inputs such as the overlying soil microbes would be theappropriate main source for these acid compounds. In contrast, theratio nC14:1/nC14:0 and the percentage of branched fatty acids dis-play no relationship with both the internal and external tempera-ture records (Fig. 5), indicating that these acids perhaps havemuch more complex inputs from both the cave and soils.
Fatty acids are a very important component in the cell mem-branes of most bacteria. The fluidity of membranes composed ofstraight-chain fatty acids is adjusted by the proportion of mono-unsaturated fatty acids (Kaneda, 1991). In order to adapt to coldconditions, microbes synthesize more unsaturated fatty acids tomaintain the fluidity of cell membranes (Suutari and Laakso,1994). Therefore, the close relationship between fatty acid ratiosnC16:1/nC16:0 and nC18:1/nC18:0 and external air temperaturemay result from a response of some microbial community in thesystem (e.g. soil microbes) to temperature changes.
For both ratios nC16:1/nC16:0 and nC18:1/nC18:0, the correla-tions with external air temperature are only moderately strong(Fig. 5), revealing that factors other than temperature (such assource difference, flow rate and rainfall amount, time lag) can alsoaffect the distributions of fatty acids in dripping waters. It isnoticeable that these non-temperature processes occurring fromthe soil to drip water could affect the absolute abundance of theindividual fatty acids, the relative abundance (the ratio) used heremight exclude, to some extent, some of these processes. For exam-ple, more microbes might multiply in the drip water in less wetseasons, which in turn promote the abundance of both saturatedand unsaturated fatty acids, but the fatty acid ratios might stay un-changed if the temperature remains unchanged. Nevertheless,these non-temperature factors need to be fully considered to de-velop a quantitative proxy based on lipids such as fatty acids indripping waters in future studies. The preliminary results indicatethat the unsaturated fatty acid ratios might have the potential totrace the change in paleotemperature. More quantitative dataare, however, required to gain a final paleotemperature proxy infuture studies.
The above correlation analysis confirms that some fatty acid ra-tios can strongly correlate with the external air temperature. None-theless, the current data available would not be able to provide theexploration of the dynamic processes to build a bridge betweenfatty acids in cave drip water and the synchronous external airtemperature. For example, in cave systems, drip rates usually lag,though it is hard to quantify the exact duration, the local rainfall.However, there is no obvious lead or lag between temperaturechanges and fatty acid ratios during the sampling interval(Fig. 4), though water samples were absent during the spring of2008. A continuous and high resolution sampling strategy isneeded in the future to identify the dynamic processes leading tothe observation of the relationship between fatty acids in dripwater and outside temperature changes.
4. Conclusions
Fatty acids are abundant in the drip water samples collectedfrom Heshang Cave, central China. The distributions are dominatedby lower-molecular-weight nC16:0, nC18:0 and nC14:0, togetherwith mono-unsaturated nC18:1, nC16:1 and nC14:1, suggestingthat microbes are the dominant source for these fatty acids in drip
water. The obvious difference between the dissolved and particu-late fractions in the water sample collected in July 2008 revealsthat soil microbes are an important source for drip water fattyacids, which can be transported into the cave system by adsorptiononto particles. The importance of soil microbes is further sup-ported by the obvious different correlation between fatty acid ra-tios (nC16:1/nC16:0 and nC18:1/nC18:0) and internal andexternal air temperature.
The mono-unsaturated fatty acid ratios (nC16:1/nC16:0 andnC18:1/nC18:0) display an obvious negative relationship with thesynchronous external air temperature but not with the internaltemperature. Higher values of the above ratios occur in cold condi-tions (winter/spring), while lower values appear in warm intervals(summer). Future work is needed to identify the dynamic pro-cesses which bridge the linkage of the drip water fatty acids andthe external air temperature, and to validate the potential applica-tion of fatty acid ratios such as nC16:1/nC16:0 and nC18:1/nC18:0in paleotemperature reconstructions in stalagmites.
Acknowledgements
This work was supported by the 973 project (Grant No.2011CB808800), the National Natural Science Foundation of China(Grant No. 40930210), the 111 project (Grant No. B08030) and theSpecial Fund for Basic Scientific Research of Central Colleges, ChinaUniversity of Geosciences (CUG090103). We thank Xiong Wang,Fushan Li, Ze Zhang and Jiaoyang Ruan for their help during samplecollection and preparation and data analysis. Professor Philip A.Meyers, University of Michigan, is acknowledged for assistance inimproving the English writing of an early draft. Dr. Marcelo Bernar-des and an anonymous reviewer are thanked for constructivecomments.
References
Baker, A., Barnes, W.L., 1998. Comparison of the luminescence properties of watersdepositing flowstone and stalagmites at Lower Cave, Bristol. Hydrol. Process. 12,1447–1459.
Baker, A., Genty, D., 1999. Fluorescence wavelength and intensity variations of cavewaters. J. Hydrol. 217, 19–34.
Baker, A., Barnes, W., Smart, P.L., 1997. Variations in the discharge and organicmatter content of stalagmite drip waters in Lower Cave, Bristol. Hydrol. Process.11, 1541–1555.
Baldini, J.U.L., McDermott, F., Fairchild, I.J., 2006. Spatial variability in cave dripwater hydrochemistry: implications for stalagmite paleoclimate records. Chem.Geol. 235, 390–404.
Birdwell, J.E., Engel, A.S., 2010. Characterization of dissolved organic matter in caveand spring waters using UV–Vis absorbance and fluorescence spectroscopy.Org. Geochem.. doi:10.1016/j.orggeochem.2009.11.002.
Blyth, A.J., Farrimond, Paul, Jones, Martin, 2006. An optimized method for theextraction and analysis of lipid biomarkers from stalagmites. Org. Geochem. 37,882–890.
Blyth, A.J., Asfawossen, A., Baker, A., Gulliver, P., Leng, M.J., Genty, D., 2007. A newapproach to detecting vegetation and land-use change using high-resolutionlipid biomarker records in stalagmites. Quatern. Res. 68, 314–324.
Dykoski, C.A., Edwards, R.L., Cheng, H., Yuan, D., Cai, Y., Zhang, M., Lin, Y., Qing, J., An,Z., Revenaugh, J., 2005. A high-resolution, absolute-dated Holocene anddeglacial Asian monsoon record from Dongge Cave, China. Earth Planet. Sci.Lett. 233, 71–86.
Genty, D., Deflandre, G., 1998. Drip flow variations under a stalactite of the PereNoel cave (Belgium). Evidence of seasonal variations and air pressureconstraints. J. Hydrol. 211, 208–232.
Hu, C., Henderson, G.M., Huang, J., Chen, Z., Johnson, K.R., 2008a. Report of a three-year monitoring programme at Heshang Cave, Central China. Int. J. Speleol. 37,143–151.
Hu, C., Henderson, G.M., Huang, J., Xie, S., Sun, Y., Johnson, K.R., 2008b.Quantification of Holocene Asian monsoon rainfall from spatially separatedcave records. Earth Planet. Sci. Lett. 266, 221–232.
Huang, X., Pu, Y., Cui, J., Wu, Y., Li, X., Huang, J., Hu, C., Xie, S., 2007. Distributions offatty acids in the drip water at Heshang Cave, Hubei Province and theirecological implications. Quatern. Sci. 27, 401–407 (in Chinese).
Huang, X., Cui, J., Pu, Y., Huang, J., Blyth, A.J., 2008. Identifying ‘‘free’’ and ‘‘bound’’lipid fractions in stalagmite samples: an example from Heshang Cave, SouthernChina. Appl. Geochem. 23, 2589–2595.

X. Li et al. / Applied Geochemistry 26 (2011) 341–347 347
Jaffé, R., Elismé, T., Cabrera, A.C., 1996. Organic geochemistry of seasonally floodedrain forest soils: molecular composition and early diagenesis of lipidcomponents. Org. Geochem. 25, 9–17.
Johns, R.B., Brady, B.A., Butler, M.S., Dembitsky, V.M., Smith, J.D., 1994. Organicgeochemistry and geochemical studies of Inner Great Barrier Reef sediments-1.Identification of terrigenous and marine sourced inputs. Org. Geochem. 21,1027–1035.
Kaneda, T., 1991. Iso- and anteiso-fatty acids in bacteria: biosynthesis, function, andtaxonomic significance. Microbiol. Rev. 55, 288–302.
Kawamura, K., Ishiwatari, R., Ogura, K., 1987. Early diagenesis of organic matter inthe water column and sediments: microbial degradation and resynthesis oflipids in Lake Haruna. Org. Geochem. 11, 251–264.
Laiz, L., Groth, I., Gonzalez, I., Saiz-Jimenez, C., 1999. Microbiological study of thedripping waters in Altamira Cave (Santillana del Mar, Spain). J. Microbiol.Methods 36, 129–138.
Drysdale, R., McDonald, J., Hill, D., 2004. The 2002–2003 El Nino recorded inAustralian cave drip waters: implications for reconstructing rainfall historiesusing stalagmites. Geophys. Res. Lett. 31, L22202. doi:10.1029/2004GL020859.
McDonald, J., Drysdale, R., Hill, D., Chisari, R., Wong, H., 2007. The hydrochemicalresponse of cave drip waters to sub-annual and inter-annual climate variability,Wombeyan Caves, SE Australia. Chem. Geol. 244, 605–623.
Meyers, P.A., Ishiwatari, R., 1993. Lacustrine organic geochemistry: an overview ofindicators of organic matter sources and diagenesis in lake sediments. Org.Geochem. 20, 867–900.
Northup, D.E., Lavoie, K.H., 2001. Geomicrobiology of caves: a review. Geomicrobiol.J. 18, 199–220.
Parkes, R.J., 1987. Analysis of microbial communities within sediments usingbiomarkers. In: Fletcher, M., Gray, T.R.G., Jones, J.G. (Eds.), Ecology of MicrobialCommunities. Cambridge University Press, London.
Quenea, K., Derenne, S., Largeau, C., Rumpel, C., Mariotti, A., 2004. Variation in lipidrelative abundance and composition among different particle size fractions of aforest soil. Org. Geochem. 35, 1335–1370.
Rieley, G., Collier, R.J., Jones, D.M., Eglinton, G., 1991. The biogeochemistry ofEllesmere Lake, U.K. – 1: source correlation of leaf wax inputs to thesedimentary lipid record. Org. Geochem. 17, 901–912.
Rontani, J.-F., Volkman, J.K., 2005. Lipid characterization of coastal hypersalinecyanobacterial mats from the Camargue (France). Org. Geochem. 36, 251–272.
Schulten, H.-R., Schnitzer, M., 1990. Aliphatics in soil organic matter in fine-clayfractions. Soil Sci. Soc. Am. J. 54, 98–105.
Suutari, M., Laakso, S., 1994. Microbial fatty acids and thermal adaptation. Crit. Rev.Microbiol. 20, 285–328.
Wang, Y., Cheng, H., Edwards, R.L., He, Y., Kong, X., An, Z., Wu, J., Kelly, M.J., Dykoski,C.A., Li, X., 2005. The Holocene Asian monsoon: links to solar changes and northAtlantic climate. Science 308, 854–857.
Wang, Y., Cheng, H., Edwards, R.L., Kong, X., Shao, X., Chen, S., Wu, J., Jiang, X., Wang,X., An, Z., 2008. Millennial- and orbital-scale changes in the East Asian monsoonover the past 224,000 years. Nature 451, 1090–1093.
Xie, S., Yi, Y., Huang, J., Hu, C., Cai, Y., Collins, M., Baker, A., 2003. Lipid distribution ina subtropical southern China stalagmite as a record of soil ecosystem responseto palaeoclimate change. Quatern. Res. 60, 340–347.
Xie, S., Huang, J., Wang, H., Yi, Y., Hu, C., Cai, Y., Cheng, H., 2005. Distributions of fattyacids in a stalagmite related to paleoclimate change at Qingjiang in Hubei,southern China. Sci. Chin. Ser. D Earth Sci. 48, 1463–1469.
Zegouagh, Y., Derenne, S., Largeau, C., Saliot, A., 1996. Organic matter sources andearly diagenetic alternations in Arctic surface sediments (Lena River delta andLaptev Sea, Eastern Siberia) – 1. Analysis of the carboxylic acids released viasequential treatments. Org. Geochem. 24, 841–857.
Zhou, W., Xie, S., Meyers, P.A., Zheng, Y., 2005. Reconstruction of late glacial andHolocene climate evolution in southern China from geolipids and pollen in theDingnan peat sequence. Org. Geochem. 36, 1272–1284.
Zou, L., Sun, M.-Y., Guo, L., 2006. Temporal variations of organic carbon inputs intothe upper Yukon River: evidence from fatty acids and their stable carbonisotopic compositions in dissolved, colloidal and particulate phases. Org.Geochem. 37, 944–956.




















