
Evolutionary and ecological consequences of interspecific hybridization in cladocerans

Original article
Seasonal succession of cladocerans and phytoplanktonand their interactions in a shallow eutrophic lake (Lake Vela, Portugal)
N. Abrantes *, S.C. Antunes, M.J. Pereira, F. Gonçalves
Departamento de Biologia, Universidade de Aveiro, 3810-193 Aveiro, Portugal
Received 17 June 2004; accepted 22 July 2005
Available online 16 September 2005
Abstract
Lake Vela is a polymictic shallow lake exhibiting some characteristics typical of an advanced trophic state, namely, the permanently turbidwater, the reduction in biodiversity, and the recurrent occurrence of Cyanobacteria blooms, which occasionally lead to large fish kills. Thisstudy was carried out in order to understand the seasonal variation of Lake Vela’s phytoplankton and cladoceran communities and theirinteractions under advanced eutrophic conditions. When comparing our results with general models of plankton succession observed in othertemperate and tropical lakes we found some coherence between several seasonal events. Our results suggest that phytoplankton is mainlyregulated by nutrients (“bottom-up” effect). However, the warm mediterranean temperatures had an important role in the phytoplanktonsuccession, being responsible for the rapid and intense Cyanobacteria development in spring and summer. Our work also demonstrated thatphytoplankton is one of the main factors responsible for the seasonal structure of the community of cladocerans, which are well related tochanges in algae diversity and abundance, being the Cyanobacteria having major impact. Moreover, the occurrence of a massive Cyanobac-teria bloom during the study, which induced anoxia and consequent fish kill, enhanced the structuring role that fish have on the cladoceranseasonal succession and the effects of this episode in the normal seasonal succession of plankton.© 2005 Elsevier SAS. All rights reserved.
Keywords: Eutrophication; Cladoceran; Phytoplankton; Trophic relationships; Shallow lake
1. Introduction
The relationship between zooplankton and phytoplanktonis an important base of the food web in lakes. The abundanceand diversity of phytoplankton and zooplankton vary in fresh-water bodies according to their limnological features andtrophic state (Jensen et al., 1994; Jeppessen et al., 2000; 2002;2003). When nutrient loading is low, lakes present a clearwa-ter phase with low algae biomass and a dense community ofsubmerged macrophytes. On the other hand, the increase ofnutrient loads in lakes may be responsible for an increase ofthe phytoplankton biomass. As the primary producers com-munity undergoes changes, the structure of communities alongthe trophic web is subsequently affected. The phytoplanktonincrease leads to the high turbidity of water, which in turn,may contribute to the depletion of the submerged macro-phytes community (Freedman, 1989; Caraco, 1993; Klinge
et al., 1995). This fact enables changes in community struc-ture, as it reduces plant surface area for algal fixation, thehabitat and refuge for zooplankton, macroinvertebrates andfish, as well as impairing the slow-growing biomass of mac-rophytes, which is responsible for long term storage of nutri-ents by removing them from water column and sediment(McDougal and Goldsborough, 1996). Moreover, some algaetaxa that respond more swiftly than others to increase in nutri-ents, as Cyanobacteria, are favoured. It is known that Cyano-bacteria are low quality food for zooplankton, due to theirfilamentous or colonial structure, low digestibility and toxinproduction, inducing limitations to the growth and reproduc-tion of zooplanktonic organisms (e.g. Arnold, 1971; Haney,1987; Lampert, 1987; Reinikainen et al., 1994; Repka, 1996;1997; DeMott, 1999; DeMott et al., 2001; Rohrlack et al.,2001; Ghadouani et al., 2003). Ultimately, the increase ofnutrient concentrations can promote the massive growth ofphytoplankton, which eventually die and are decomposed bymicroorganisms that are responsible for the elevated rates oforganic matter decomposition and the subsequent anoxia,
* Corresponding author. Fax: +351 2 3437 0777.E-mail address: [email protected] (N. Abrantes).
Acta Oecologica 29 (2006) 54–64
www.elsevier.com/locate/actoec
1146-609X/$ - see front matter © 2005 Elsevier SAS. All rights reserved.doi:10.1016/j.actao.2005.07.006

leading to large kills of fish and other aquatic life (Brönmarkand Weisner, 1992). Zooplankton represents the channel oftransmission of the energy flux from the primary producersto the top consumers, because they include the primary her-bivores and are important diet items for fish and other preda-tors. In particular, cladocerans are an important constituentof most lake zooplankton communities, due to their capabil-ity to filter all particles within a given size range, includingbacteria, protozoa, detritus and algae (Lampert, 1978; Infanteand Litt, 1985; De Bernardi et al., 1987; Hessen and Ander-sen, 1990; Brendelberger, 1991; Müller-Navarra and Lampert,1996; Greenwood et al., 1999; Hülsmann, 2001). Hence, theseorganisms are faced with a great diversity of resources of dif-ferent quality and since their capacity of selection is limited,the quality and quantity of food are determinants in the lifehistory of cladocerans (Lampert, 1978; Müller-Navarra andLampert, 1996; Greenwood et al., 1999; Hülsmann, 2001).On the other hand, in addition to nutrient conditions, it isknown that the zooplankton can affect the composition andabundances of phytoplankton species, both by direct grazing(Shapiro, 1995) and by altering the nutrient conditions throughnutrient regeneration (Carney and Elser, 1990).
Lake Vela is a shallow polymictic lake situated in the Westcoast of Portugal, which is located in the Mediterranean-iberoatlantic group of the Mediterranean region, where theclimate is influenced by the proximity of the Atlantic Ocean.Due to Lake Vela’s strategical-geographic position it is con-sidered as a protected zone with high conservational interest.Thereby, it was classified as a first level protection naturalarea and it benefits the protection statute defined by the Bió-topos Project of Corine Programme 85/338/CEE, of 27 June(Farinha and Trindade, 1994). However, Lake Vela water-shed shows intense agricultural and livestock activities on theEast site. Due to the sandy nature of soils and the small depthof the groundwater (Fernandes, 1999), the drainage basin ofLake Vela is potentially exposed to loads of nutrients gener-ated by these activities. Thus, this lake is classified aseutrophic since 1960 (Nauwerck, 1960), showing high N andP levels typical of eutrophic systems (Barros, 1994; Pereira,1997, Fernandes, 1999; Abrantes, 2002; Antunes et al., 2003).The increase of phytoplankton biomass, with the presence ofalgal blooms, the depletion of submerged macrophytes, theincrease in turbidity, and a subsequent reduction in biodiver-sity are some of the characteristics associated with theeutrophication process in Lake Vela, that could compromiseits wildlife and recreational use.
Although general hypotheses and patterns of plankton suc-cession were well described in temperate dimictic lakes (Som-mer et al., 1986), there is still very few studies assessing plank-ton succession patterns in shallow lakes with Mediterraneanclimate, polymictic thermal regime and extreme eutrophicconditions associated to Cyanobacteria blooms. Thus, thisstudy constitutes a complement for previous works, namelyworks on plankton seasonal succession, as PEG model. Fur-thermore, to contribute for the management of the Lake Velasystem and the activities in its drainage basin it is important
to understand the behaviour and linkages between the trophiclevels under advanced trophic conditions. Thus, we aim tostudy the structure and the seasonal changes of the primaryproducers and herbivorous cladocerans and their interdepen-dency in the eutrophic polymictic Lake Vela.
2. Material and methods
2.1. Study description
LakeVela, located in the littoral-centre of Portugal (40°5´N8°8´W, 45 m elevation), is a temperate shallow lake (meandepth = 0.9 m, maximum depth = 2.4 m) with 70 ha of flood-able surface area and a total volume of 70 × 104 m3. It is sur-rounded in the East side by agricultural fields and some humansettlements and, in the West side, by a forest road, which isseparated from the lake by pine trees (Pinus pinaster) andacacias (Acacia spp.). Due to the regular nutrient inputs, thelake is highly eutrophic. It is characterised by a permanentlyturbid water, without a spring clear water phase, and algalblooms are frequently observed, namely in the warmer sea-sons. The phytoplankton is typically dominated by Cyano-bacteria, Bacillariophyceae (e.g. Cyclotella spp.) and coc-coid Chlorophyta (Barros, 1994). The macrophyte communityis characterised by the dominance of Cladium mariscus,Myriophyllum verticillatum, Nymphaea alba and species fromPoaceae family (Pereira, 1997). However, nearly 80–90% ofthe flooded lake bed does not contain any vegetation(Fernandes, 1999). The major crustacean species recorded areDaphnia longispina, Ceriodaphnia pulchella and Bosminalongirostris (Antunes et al., 2003). Fish are also abundant,with prevalent species such as: Lepomis gibbosus, Gambusiaholbrooki and Micropterus salmoides, which are all intro-duced species.
2.2. Physico-chemical analysis
Samples were collected bimonthly near surface (0.4 m inaverage) in the pelagic zone of the lake, over a 1-year period.For chemical analysis (alkalinity, hardness and nutrients),chlorophyll a and suspended solids, water samples were col-lected with a 1.5 l plastic bottle and transported to the lab inthe dark, at 4 °C. Until further processing, the samples forchemical analysis were preserved at –20 °C. Chlorophyll awas measured after collection on glass fibre filters (WhatmanGF/C) and extracted in 90% acetone in the dark, followingA.P.H.A. (1995). Likewise, for suspended solids analysis,samples were filtered through preweighed filters (WhatmanGF/C) and the solids (volatile and fixed) were calculatedaccording to A.P.H.A. (1995). Hardness and alkalinity weredetermined through buret titration methods. Ammonia wasdetermined by Nessler method, nitrites by a colorimetricmethod, nitrates by cadmium reduction method and solublereactive phosphorous (SRP) by the ascorbic acid method, fol-lowing A.P.H.A. (1995) procedures. Simultaneously to field
55N. Abrantes et al. / Acta Oecologica 29 (2006) 54–64

sampling, temperature, pH (WTW 340-A pH meter), dis-solved oxygen (WTW OXI 320 oxygen meter) and conduc-tivity (WTW LF 330 conductivity meter) were recorded sub-superficially.
2.3. Biological parameters
For phytoplankton a 1 l sample were preserved in lugol(1% v/v) for later identification and enumeration. The iden-tification of phytoplankton species was made by observationthrough a light microscope using different references forCyanobacteria (Geitler, 1932; Komárek and Anagnostidis,1999), Bacillariophyceae (Krammer and Lange-Bertalot,1986–1991) and Chlorophyta (Chlorococcales) (Komárek andFott, 1983). The enumeration was performed according toLund et al. (1958) with at least 400 cells counted. For cla-doceran analysis, three replicate samples were collected andconcentrated by filtration through a 55 µm-mesh screen untilsignificant clogging occurred. Each nylon screen was imme-diately placed in 4% sugar formalin for zooplankton preser-vation (Haney and Hall, 1973). In the laboratory, zooplank-ton samples were sorted out and cladoceran taxa wereidentified through a binocular stereoscope, using the keys ofScourfield and Harding (1966); Amoros (1984a, 1984b), andenumerated.
2.4. Statistical analysis
A canonical correspondence analysis or CCA (Ter Braak,1995) was used to ascertain the relationships between zoop-lankton, phytoplankton and environmental variables along thestudy period. CCA extracts synthetic gradients from the bioticand environmental matrices and the explanatory variables arequantitatively represented by arrows in graphical biplots. Thearrow direction indicates positive or negative correlations andtheir length is relative to the importance of the explanatoryvariable in the ordination. A Monte Carlo permutation testwas used to assess the significance of the relationshipsbetween the biotic data and the explanatory variables. Varia-tion partitioning was performed according to the methoddeveloped by Borcard et al. (1992), which permits to singleout the respective effects of the environmental factors. In ourwork, we considered two sets of explanatory variables: biotic(Bio) and abiotic (Abio), being included in Bio the phy-toplankton and cladoceran communities and in Abio thephysico-chemical factors. Bio and Abio variables wereobtained by CCA after forward selection of variables. Totalvariations in phytoplankton and cladocerans were portionedas follows: Bio|Abio: fraction of total variation explained bybiotic factors (Bio) alone, not shared with abiotic factors(Abio); BioxAbio: fraction of total variation explained by theinteraction of Bio and Abio; Abio|Bio: fraction of total varia-tion explained by abiotic factors (Abio) alone, not shared withbiotic factors (Bio); x: fraction of total variation unexplainedby the abiotic and biotic factors.
3. Results
3.1. Physical and chemical characteristics
Water temperature was lower in autumn–winter seasons(10.3 °C recorded in November), and higher in spring-summer seasons (29.4 °C recorded in July) (Fig. 1). Oxygen
Fig. 1. Variation of physical and chemical characteristics of Lake Vela waterduring the studied period.
56 N. Abrantes et al. / Acta Oecologica 29 (2006) 54–64
levels were relatively stable during the study period, howevercomplete oxygen depletion occurred in May 29, which cor-responded to a large fish kill. In general, during the study, pHand conductivity did not show significant variations. Sus-pended solids presented their lower values in winter and beganto increase in spring, reaching their higher values in summerand early autumn. They were mainly constituted by volatileorganic materials. All analysed nutrients (nitrites, nitrates,SRP and ammonia) showed a similar trend, with highest con-centrations in the period from December to April, which isexplained by the adjacent soil leaching, promoted by rain.Nevertheless, ammonia showed an increase in May 29, cor-responding to the fish kill. Alkalinity did not shown consid-erable changes during the sampling period, while hardnessshowed a substantial decrease after August (Fig. 1).
3.2. Seasonal succession of phytoplankton and cladocerancommunities
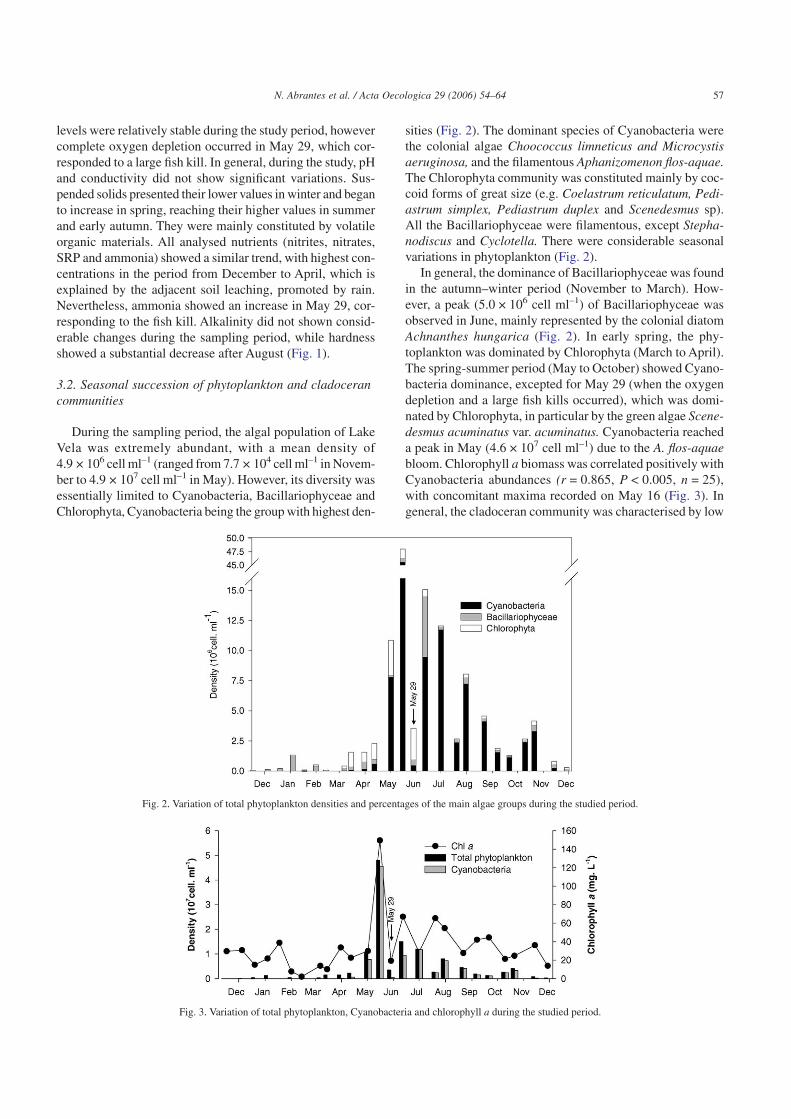
During the sampling period, the algal population of LakeVela was extremely abundant, with a mean density of4.9 × 106 cell ml–1 (ranged from 7.7 × 104 cell ml–1 in Novem-ber to 4.9 × 107 cell ml–1 in May). However, its diversity wasessentially limited to Cyanobacteria, Bacillariophyceae andChlorophyta, Cyanobacteria being the group with highest den-
sities (Fig. 2). The dominant species of Cyanobacteria werethe colonial algae Choococcus limneticus and Microcystisaeruginosa, and the filamentous Aphanizomenon flos-aquae.The Chlorophyta community was constituted mainly by coc-coid forms of great size (e.g. Coelastrum reticulatum, Pedi-astrum simplex, Pediastrum duplex and Scenedesmus sp).All the Bacillariophyceae were filamentous, except Stepha-nodiscus and Cyclotella. There were considerable seasonalvariations in phytoplankton (Fig. 2).
In general, the dominance of Bacillariophyceae was foundin the autumn–winter period (November to March). How-ever, a peak (5.0 × 106 cell ml–1) of Bacillariophyceae wasobserved in June, mainly represented by the colonial diatomAchnanthes hungarica (Fig. 2). In early spring, the phy-toplankton was dominated by Chlorophyta (March to April).The spring-summer period (May to October) showed Cyano-bacteria dominance, excepted for May 29 (when the oxygendepletion and a large fish kills occurred), which was domi-nated by Chlorophyta, in particular by the green algae Scene-desmus acuminatus var. acuminatus. Cyanobacteria reacheda peak in May (4.6 × 107 cell ml–1) due to the A. flos-aquaebloom. Chlorophyll a biomass was correlated positively withCyanobacteria abundances (r = 0.865, P < 0.005, n = 25),with concomitant maxima recorded on May 16 (Fig. 3). Ingeneral, the cladoceran community was characterised by low
Fig. 2. Variation of total phytoplankton densities and percentages of the main algae groups during the studied period.
Fig. 3. Variation of total phytoplankton, Cyanobacteria and chlorophyll a during the studied period.
57N. Abrantes et al. / Acta Oecologica 29 (2006) 54–64

diversity and elevated abundances. The major cladoceran taxarecorded during the study period were D. longispina, Bosminalongirostris, Ceriodaphnia pulchella and Chydoridae. In gen-eral, D. longispina and Bosmina longirostris were the domi-nant cladocerans in Lake Vela. However, the highest abun-dances were observed for B. longirostris and Chydoridae(Fig. 4). Abundances and composition of cladocerans variedsignificantly through the seasons (Fig. 4). D. longispina pre-sented their maximum abundance in August and September(318 ind. l–1), being the dominant species in the winter (fromJanuary to March) and in early autumn (from September toNovember). Ceriodaphnia pulchella presented very low den-sities along the year. Nevertheless, they attained a peak inMay 16 (28 ind. l–1), coincident with the maximum densityof total phytoplankton. High positive correlations wereobserved between C. pulchella and Cyanobacteria densities(r = 0.864, P < 0.005, n = 25). During the first study months,where D. longispina and C. pulchella presented very lowdensities, Chydoridae and B. longirostris reached very highdensities (≈1000–1500 ind. l–1) (Fig. 4). Bosmina longiros-tris became the dominant species in the summer, attainingtheir highest density in August (1551 ind. l–1). Chydoridaealso reached high densities in the summer, but less pro-nounced than B. longirostris. Practically, no cladocerans wereobserved in the May 29 (due to the anoxia and large fish kills),including C. pulchella, which attained a peak 2 weeks ear-lier, in May 16.
3.3. CCA and variation partitioning analysis
The results of the CCA applied to the environmental fac-tors (physical and chemical variables and cladoceran com-munity) indicate that only the abiotic factors significantlyinfluence the phytoplankton community (P ≤ 0.005; MonteCarlo test of first constrained axis), explaining 31% of thetotal variance (Table 1 and Figs. 5 and 6). The most discrimi-nant physico-chemical variables were temperature, volatilesolids, hardness, SRP and pH (Fig. 5), nevertheless, cleartrends are difficult to extract. Provided by the variation par-titioning analysis, the pure contribution of the physical andchemical variables (Abio|Bio) was large (30%) and signifi-cant (P ≤ 0.005) while the pure contribution of cladocerans(Bio|Abio) was lower (13%) and not significant (P = 0.095).The total variation, explained by the abiotic and biotic fac-tors and their interaction, was relatively high with a total of44% (Fig. 7a). The CCA and partial CCA analysis applied tothe environmental factors allowed to ascertain that thepure contribution of the physico-chemical parameters(Abio|Bio = 1%) and the phytoplankton (Bio|Abio = 4%) didnot explain the cladocerans abundance and diversity (Fig. 7b).However, the variation shared by (BioxAbio) was 24%, sug-gesting an interaction between physico-chemical factors andphytoplankton. Thus, we intend to perform a CCA of cla-docerans with respect to phytoplankton and physico-chemicalvariables, which was explaining 29% of the total variance
Fig. 4. Variation of total cladocerans densities and percentages of the main species during the studied period.
Table 1Summary of CCA analysis between phytoplankton and different variables for the studied period
Physico-chemical factors Cladoceran communityAxis 1 Axis 2 Axis 1 Axis 2
Eigenvalues 0.200 0.105 0.084 0.055Species–environment correlations 0.954 0.956 0.746 0.837Cumulative percentage variance of species data 17.6 26.8 7.3 12.2of species–environment relation 26.7 40.7 39.6 65.7Sum of all unconstrained eigenvalues 1.14 1.14Sum of all canonical eigenvalues 0.751 0.211Variance explained 31% 14%
58 N. Abrantes et al. / Acta Oecologica 29 (2006) 54–64

and showed a statistical significance (P ≤ 0.005) (Table 2).Clear trends are difficult to extract, mainly because of theproximity of the species scores (Fig. 8). However, ammoniawas the most discriminant variable, which is probably relatedto the Chl a peak and subsequent oxygen depletion on theoccasion of the fish kill (May 29).
4. Discussion
Lake Vela is a eutrophic shallow lake with high nutrientlevels (mainly nitrates and phosphates) and its cladocerancommunity presents low diversity and higher abundances.Cyanobacteria blooms are frequent, especially A. flos-aquaeand M. aeruginosa, and fish kills have occurred in the lastyears. Submerged macrophytes are usually abundant, plank-tivorous fish now dominate, and Chl a presents high valuesduring the warmer months. This description is in agreementwith the findings of Jeppesen et al. (1999) on eutrophic tur-bid lakes.
In eutrophic lakes the role of nutrients versus zooplanktongrazing in controlling phytoplankton continues to be a con-troversial question among limnologists. Sommer et al. (1986)
present a general model (the PEG model) for the seasonalplanktonic patterns. According to these authors, with nutri-ent availability and increased light, phytoplankton productiv-ity increases in spring and supports growth of cladoceran her-bivore populations; the herbivore populations increaseexponentially and eventually exceed food availability anddecline; in some systems, planktivores fish allied to the foodlimited accelerates the decline of crustacean herbivores and areplacement of larger species by smaller species in the sum-mer; autumn destratification, resulting in nutrient replenish-ment and a deterioration of the effective underwater light cli-mate, permits the development of edible phytoplanktonspecies and a second peak in zooplankton biomass (appliedto stratifying lakes only); a reduction of light energy resultsin decline of algal biomass to the winter minimum, whichassociated to low winter temperatures restrict herbivores pro-duction and some species produce resting stages.
The CCA performed on the data revealed that the phy-toplankton abundances and diversity is mainly regulated byabiotic factors, including temperature and nutrients availabil-ity. In respect to the cladocerans, CCA suggests that theirdistribution was, in part, influenced by an interaction betweenphysico-chemical factors and phytoplankton. Therefore, we
Fig. 5. Spatial ordination resulting from CCA of phytoplankton with respect to physical and chemical parameters. cyAPFL – A. flos-aquae; cyMIAE – M.aeruginosa; dACDE – Achnanthes delicatula; dACHU – Achnanthes hungarica; dACLA -Achnanthes lanceolata; dACMI – Achnanthes minutissima; dAUGA– Aulacoseira granulata var. angustissima; dCRHA – Craticula halophila; dCYOC – Cyclotella ocellata; dCYME – Cyclotella meneghiniana; dCYHE –Cymbella helvetica; dLUVE – Luticola ventricosa; dDIOV – Diploneis ovalis; dFRCO – Fragilaria construens; dGOAF – Gomphonema affıne; dNARY –Navicula rynchocephala; dNIGR – Nitzschia gracilis; dPIBO – Pinnularia borealis; dRHGI – Rhopalodia gibba; dSEPU – Sellaphora pupula; dSUAN –Surirella angusta; chLASU – Lagerheimia subsalsa; chPEDU – Pediastrum duplex var. duplex; chPESS – Pediastrum simplex var. simplex; chPETE –Pedistrum tetras; chSACU – S. acuminatus var. acuminatus; chSCGU – Scenedesmus gutwinskii var. heterospina; chSCHE – Scenedesmus heterospina;chSCOP – Scenedesmus opoliensis var. monoensis; chSCSE – Scenedesmus semicristatus; chSCSP – Scenedesmus spinosus.
59N. Abrantes et al. / Acta Oecologica 29 (2006) 54–64

believe that physico-chemical factors do not influence directlythe cladocerans community, but control the phytoplanktongrowth, which in turn influence the dynamics of cladocerans.The drainage basin of Lake Vela shows at the East side smallrural properties characterised by an intense agricultural activ-ity, as well as intensive livestock breeding. Thus, the run-offinto the Lake Vela has a high nutrient content, which explainsthe high values recorded for nutrients from winter to earlyspring. As a consequence, we observed an increase of phy-toplankton in early spring. According to PEG model, a springproduction of small fast-growing algae such as Crypto-
phyceae and small centric Bacillariophyceae develops. In ourwork, which is corroborated by previous studies in Lake Vela(Barros, 1994; Fernandes, 1999), was observed that the springgrowth of algae is firstly dominated by Chlorophyta, beingreplaced by Cyanobacteria. The warm temperatures in spring,typical of Mediterranean areas, that enables a faster growthof Cyanobacteria, possibly allied with cladoceran grazingpressure in small edible phytoplankton, promote their domi-nance and the appearance of blooms in Lake Vela. As alreadystated, the presence of Cyanobacteria leads to a decrease inDaphnia densities, because Cyanobacteria interfere with fil-
Fig. 6. Spatial ordination resulting from CCA of phytoplankton with respect to cladoceran community. cyAPFL – A. flos-aquae; dAUDI – Aulacoseira distans;dCYOC – Cyclotella ocellata; dCYHE – Cymbella helvetica; dFRBR – Fragilaria brevistriata; dFRCO – Fragilaria construens; dFRPI – Fragilaria pinnata;dGOGR – Gomphonema gracile; dGOPA – Gomphonema parvulum; dNARY – Navicula rynchocephala; dNIGR – Nitzschia gracilis; dNIPL – Nitzschiapaleacea; chTECA – Tetraedron caudatum.
Fig. 7. Relative importance of the pure environmental fraction (Bio|Abio and Abio|Bio), the variation shared by Bio and Abio (BioxAbio) and the unexplainedvariation (x) for phytoplankton (A) and cladocerans abundances (B).
60 N. Abrantes et al. / Acta Oecologica 29 (2006) 54–64

tration, have low digestibility and also because they releasetoxins (e.g. Arnold, 1971; Haney, 1987; Lampert, 1987; Reini-kainen et al., 1994; Repka, 1996; 1997; DeMott, 1999;DeMott et al., 2001; Rohrlack et al., 2001; Ghadouani et al.,2003). This evidence can explain the non observed spring peakof zooplankton in Lake Vela (also non reported in previouslimnological works in Lake Vela), which is largely docu-mented in other works (e.g. Sommer et al., 1986; Saunders etal., 1999) as responsible for the reduction of phytoplanktonand the consequent clear water phase. In general, smaller cla-docerans are less affected by Cyanobacteria (Berzins andPejler, 1989; Radwan and Popiolek, 1989; Pinel-Alloul,
1993). Frequently there is the replacement of large speciesby small ones, when the Cyanobacteria are dominant(Lampert, 1982). When comparing the different populationsof cladocerans during the study period, it was evident thatDaphnia was the most affected species by the spring Cyano-bacteria blooms, leading to a drastic reduction of its abun-dance. On the other hand, simultaneously to the peak ofCyanobacteria (due to the A. flos-aquae bloom), the smallcladoceran C. pulchella reached their highest density everrecorded during the study, showing a positive significantlycorrelation with Cyanobacteria densities. This fact could bealso related to the efficient capture and consumption of bac-teria by C. pulchella in the presence of inedible algae(DeMott, 1989). PEG model also reports the replacement oflarger cladoceran species by smaller ones, but this occur-rence is seen most distinctly during the summer (Sommer etal., 1986).
The large amount of unexplicated variation (71%) obtainedin our study, suggests that cladoceran community structuremay not be regulated only by “bottom up” forces such asfood availability or nutrients. Predation is often an importantfactor conditioning zooplankton structure (e.g. Christoffer-sen et al., 1993). In summer, zooplankters are subjected tovery high predatory pressure, especially because ofYOY fish,and this pressure is size-selective, with major impact in larger-size cladocerans—Daphnia.
Table 2Summary of CCA analysis between cladocerans and environmental varia-bles (phytoplankton and abiotic factors jointly) for the studied period
Phytoplankton andphysico-chemical factors
Axis 1 Axis 2Eigenvalues 0.230 0.058Species–environment correlations 1 1Cumulative percentage variance
of species data 72.2 90.5of species–environment relation 72.2 90.6
Sum of all unconstrained eigenvalues 0.319Sum of all canonical eigenvalues 0.319Variance explained 29%
Fig. 8. Spatial ordination resulting from CCA of cladocerans with respect to explanatory variables (phytoplankton and physico-chemical variables). CyAPFL –Aphanizomenon flosaquae; cyMIAE – M. aeruginosa; chBOBR – Botriococcus braunii; chLASU – Lagerheimia subsalsa; chMOCO – Monoraphidiumcontortum; chPEPO – Pediastrum boryanum; chSACU – S. acuminatus var. acuminatus; chSCAC – Scenedesmus acutus var. acutus; chSCSP – Scenedesmusspinosus; dACDE – Achnanthes delicatula; dACMA – Achnanthes minutissima var. affınis; dAMLY – Amphora lybica; dAUGA – Aulacoseira granulata var.angustissima; dCYHE – Cymbella helvetica; dCYST – Cyclotella stelligera; dDETE – Denticula tenuis; dDIOV – Diploneis ovalis; dGOCL – Gomphoneisclevei; dNACA – Navicula capitata.
61N. Abrantes et al. / Acta Oecologica 29 (2006) 54–64

The massive growth of Cyanobacteria that occurred in Maywas responsible for the large kill of fish and other aquaticlife, as a consequence of complete oxygen depletion.Althoughthe extent of the fish kill was not studied, it seems to havebeen drastic enough to diminish the predatory pressure oncladocerans. Under the same conditions, Daphnia is a bettercompetitor than Ceriodaphnia and it has been experimen-tally shown that Daphnia can suppress small species, espe-cially in eutrophic systems, where the competitive effects aremore pronounced (Vanni, 1986). Thus, the major fish killsobserved in May 29 can explain the summer Daphnia peak.However, the decrease of Cyanobacteria after this occur-rence, can also explain the increase of D. longispina, andconsequently the reduction of C. pulchella densities. Previ-ous studies on Lake Vela, in which the fish kills were notrecorded, have shown a dominance of D. longispina and B.longirostris in the beginning of the summer, but they werereplaced by C. pulchella during the rest of the season (Vas-concelos, 1990 (only studied the zooplankton community dur-ing the summer; Barros, 1994).
Nutrient limitation is a common characteristic during sum-mer stratification (Sommer et al., 1986). Lake Vela, althoughit is a non-stratified lake, also presents a limitation of nutri-ents in summer, namely a phosphorous limitation (Barros,1994; Fernandes, 1999). This absence is associated to the highgrowth of phytoplankton that leads to the nutrient depletion.As referred by Sommer et al. (1986) the competition for phos-phate leads to a replacement of green algae by large Bacilla-riophyceae and later the nitrogen depletion favours a shift tonitrogen-fixing of filamentous Cyanobacteria. Lake Vela pre-sents Cyanobacteria dominance in summer (Barros, 1994;Fernandes, 1999), being the A. flos-aquae (a nitrogen-fixingalgae) and M. aeruginosa the most representative species inour study.A new zooplankton peak typically occurs in autumn,as a consequence of the nutrient replenishment and the sub-sequent development of edible phytoplankton species (e.g.Sommer et al., 1986; Saunders et al., 1999). Lake Vela pre-sents a gradual transition between summer and winter, with-out the autumn destratification event and the consequentrelease of nutrients. However, in our study we observed aDaphnia peak in the beginning of autumn, which could berelated to the continue decrease of Cyanobacteria and adecrease in predation still associated to the May 29 fish kill.Barros (1994) presents in her work an autumnal peak of cla-docerans, although with C. pulchella dominance, which ispossibly related to the fish predation pressure on major cla-docerans that persists in early autumn. In winter, the reduc-tion of light energy results in phytoplankton biomass decreaseto the minimum, which is associated to cool winter tempera-tures and restricts zooplankton production rates and, for somespecies, stimulates diapause (Sommer et al., 1986). Our study,as well as the work developed by Barros (1994) in Lake Vela,showed low abundances of cladocerans during the winter. Thedecline of edible algae biomass in winter, that only starts toincrease after the typical winter precipitation and the conse-quent run-off and nutrient replenishment, can explain this
event. In addition, the role of temperature, which influencesthe metabolic rates of cladocerans (e.g. Burns, 1969; Gossand Bunting, 1983; Moore et al., 1996; Giebelhausen andLampert, 2001), is also an important factor that can explaintheir lower winter densities. It is possible to observe the pres-ence of resting eggs produced by D. longispina in Lake Vela.However, this occurrence was not only associated with thelow temperatures, being observed the presence of resting eggsalong the year as a consequence of several stressing factors.
5. Conclusion
From the present study we conclude that the seasonal pat-tern of the phytoplankton and cladoceran communities is moreor less coherent with the general model of plankton seasonalsuccession described for other temperate lakes. Phytoplank-ton in Lake Vela is mainly regulated by physico-chemical fac-tors, namely nutrients. However, the high mediterranean tem-peratures had an important role in the plankton succession,being responsible for the exponential Cyanobacteria devel-opment in spring and summer. Moreover, the present workdemonstrated that phytoplankton is one of the main factorsresponsible for the succession of cladoceran community,which are well related to changes in algae composition anddensity, the Cyanobacteria being the group with major impact.Nevertheless, the consequences of fish kills, such as the onerecorded here, enhanced the structuring role that fish have onthe cladoceran seasonal dynamics, and that any irregular eventin the system has effects on the normal seasonal successionof plankton. It is important to consider that this occurrencehas been regularly observed in Lake Vela and its effects onthe zooplanktonic community are not yet fully understood. Itshould be considered that the changes shown by organisms,in general, do not occur as a response to a single factor, but toa complexity of interactions generated by a large range offactors. Thus, further information on competitive interac-tions within plankton, toxic cyanobacteria, anti-predatordefences, or YOY fish, should be included in such models inorder to understand the intricate trophic relationships ofaquatic systems Wetzel, 1993.
Acknowledgments
This research was supported by a grant from Science andTechnology Foundation of the Portuguese Ministry of Sci-ence and Higher Education. We are grateful to Bruno Castroand Catarina Marques for helpful suggestions.
References
A.P.H.A., 1995. Standard Methods for the Examination of Water andWastewater—19th Edition. American Public Health Association, Wash-ington.
62 N. Abrantes et al. / Acta Oecologica 29 (2006) 54–64

Abrantes, N., 2002. Dinâmica populacional de Ceriodaphnia pulchella(Crustacea, Cladocera). M.Sc. thesis. University of Aveiro. Departmentof Biology, Aveiro.
Amoros, C., 1984. Introduction pratique à la systématique des organismesdes eaux continentales françaises. Crustacés cladocères. Bull. Mens.Soc. Linn. Lyon. 3, 72–107 (a).
Amoros, C., 1984. Introduction pratique à la systématique des organismesde eaux continentales françaises. Crustacés cladocères (suite). Bull.Mens. Soc. Linn. Lyon. 4, 120–144 (b).
Antunes, S.C., Abrantes, N., Gonçalves, F., 2003. Seasonal variation of theabiotic parameters and the cladoceran assemblage of Lake Vela. Ann.Limnol. 39, 255–264.
Arnold, D.E., 1971. Ingestion, assimilation, survival, and reproduction byDaphnia pulex fed seven species of blue-green algae. Limnol. Oceanogr.16, 906–920.
Barros, P., 1994. Implicações ecotoxicológicas de cianobactérias emcladóceros. M.Sc. thesis. University of Coimbra. Faculty of Sciences andTechnology, Coimbra.
Berzins, B., Pejler, B., 1989. Rotifers occurrence and trophic degree. Hydro-biologia 182, 1373–1403.
Borcard, D., Legendre, P., Drapeau, P., 1992. Partialling out the spatialcomponent of ecological variation. Ecology 73, 1045–1055.
Brendelberger, H., 1991. Filter mesh size of cladocerans predicts retentionefficiently for bacteria. Limnol. Oceanogr. 36, 884–894.
Brönmark, C., Weisner, S.E.B., 1992. Indirect effects of fish communitystructure on submerged vegetation in shallow, eutrophic lakes: an alter-native mechanism. Hydrobiologia 243/244, 239–301.
Burns, C.W., 1969. Relation between filtering rate, temperature and bodysize in four species of Daphnia. Limnol. Oceanogr. 14, 693–700.
Caraco, N.F., 1993. Disturbance of phosphorus cycle: a case of indirecteffects of human activity. Trends Ecol. Evol. 8, 51–54.
Carney, H.J., Elser, J.J., 1990. Strength of zooplankton-phytoplankton cou-pling in relation to lake trophic state. In: Tizler, M.M., Serruya, C. (Eds.),Large Lakes: Ecological Structures and Functions. Springer-Verlag,New York, pp. 615–631.
Christoffersen, K., Riemann, B., Klysner, A., Sondergaard, M., 1993. Poten-tial role of fish predation and natural populations of zooplankton instructuring a plankton community in eutrophic lake water. Limnol.Oceanogr. 38, 561–573.
De Bernardi, R., Guissani, G., Manca, M., 1987. Cladocera: predators andprey. Hydrobiologia 145, 225–243.
DeMott, W.R., 1989. The role of competition in zooplankton succession. In:Sommer, U. (Ed.). Plankton Ecology, Berlin, pp. 195–252.
DeMott, W.R., 1999. Foraging strategies and growth inhibition in fivedaphnids feeding on mixture of a toxic cyanobacterium and green alga.Freshwater Biol. 42, 263–274.
DeMott, W.R., Gulati, R.D., Van Donk, E., 2001. Daphnia food limitation inthree hypereutrophic Dutch lakes: evidence for exclusion of large-bodied species by interfering filaments of cyanobacteria. Limnol.Oceanogr. 46, 2054–2060.
Farinha, J.C., Trindade, A., 1994. Contribuição para o inventário e caracter-ização de zonas húmidas em Portugal Continental. Publ. MedWet/Inst.Cons. Natureza, Lisboa.
Fernandes, M.J.S., 1999. Modelação e simulação nas lagoas de Quiaios.Ph.D. thesis. University of Algarve, Faro.
Freedman, B., 1989. Environmental Ecology. The impacts of pollution andothers stresses on ecosystem structure and function. Academic Press Inc,California.
Geitler, L., 1932. Dr. L. Rabenhost’s Kryptogamen-Flora von Deutschland,Österreich und der Schweiz. Cyanophyceae von Europa. AkademischeVerlagsgesellschaft m. b. H., Leipzig. Johnson Reprint Corporation,New York.
Ghadouani, A., Pinel-Alloul, B., Prepas, E.E., 2003. Effects of experimen-tally induced cyanobacterial blooms on crustacean zooplankton commu-nities. Freshwater Biol. 48, 363–381.
Giebelhausen, B., Lampert, W., 2001. Temperature reaction norms ofDaphnia: the effect of food concentration. Freshwater Biol. 46, 281–289.
Goss, B.L., Bunting, D.L., 1983. Daphnia development and reproduction:responses to temperature. J. Therm. Biol. 8, 375–380.
Greenwood, T.L., Green, J.D., Hicks, B.J., Chapman, A., 1999. Seasonalabundance of small cladocerans in lake Mangakaware, Waikato, NewZealand. N. Z. J. Mar. Freshwat. Res. 33, 399–415.
Haney, J.F., 1987. Field studies on zooplankton–cyanobacteria interactions.N. Z. J. Mar. Freshwat. Res. 21, 467–475.
Haney, J.F., Hall, D.J., 1973. Sugar-coated Daphnia: a preservation tech-nique for Cladocera. Limnol. Oceanogr. 18, 331–333.
Hessen, D.O., Andersen, T., 1990. Bacteria as source of phosphorus forzooplankton. Hydrobiologia 206, 217–223.
Hülsmann, S., 2001. Reproductive potential of Daphnia galeata in relationto food conditions: implications of a changing size-structure of thepopulation. Hydrobiologia 442, 241–252.
Infante, A., Litt, A.H., 1985. Differences between two species of Daphnia inthe use of 10 species of algae in Lake Washington. Limnol. Oceanogr. 30,1053–1059.
Jensen, J.P., Jeppesen, E., Olrik, K., Kristensen, P., 1994. Impact of nutrientsand physical factors on the shift from cyanobacterial to chlorophytedominance in shallow Danish lakes. Can. J. Fish. Aquat. Sci. 51, 1692–1699.
Jeppesen, E., Jensen, J.P., Søndergaard, M., Lauridsen, T., 1999. Trophicdynamics in turbid and clearwater lakes with special emphasis on therole of zooplankton for water clarity. Hydrobiologia 408/409, 217–231.
Jeppesen, E., Jensen, J.P., Søndergaard, M., Lauridsen, T., Landkildehus, F.,2000. Trophic structure, species richness and biodiversity in Danishlakes: changes along a phosphorus gradient. Freshwater Biol. 45, 201–213.
Jeppesen, E., Jensen, J.P., Søndergaard, M., 2002. Response of phytoplank-ton, zooplankton and fish to re-oligotrophication: an 11-year study of23 Danish lakes. Aquatic Ecosystems Health & Management 5, 31–43.
Jeppesen, E., Jensen, J.P., Jensen, C., Faafeng, B., Brettum, P., Hessen, D.,Søndergaard, M., Lauridsen, T., Christoffersen, K., 2003. The impact ofnutrient state and lake depth on top down control in the pelagic zone oflakes: study of 466 lakes from the temperate zone to the Arctic. Ecosys-tems 6, 313–325.
Klinge, M., Grimm, M.P., Hosper, S.H., 1995. Eutrophication and ecologi-cal rehabilitation of Dutch lakes: presentation of a new conceptual modelframework. Wat. Sci. and Tech. 31, 207–218.
Komárek, J., Anagnostidis, K., 1999. Cyanoprokariota—1. Teil Choococ-cales. In: Ettl, H., Gärtner, G., Heynig, H., Mollenhauer, D. (Eds.),Sübwasserflora von Mitteleuropa, Band 19/1. G. Fischer Verlag. Lübeck,Ulm, Jena, Stuttgart, pp. 1–548.
Komárek, J., Fott, B., 1983. Das Phytoplankton des Süsswassers, Systematikund Biologie. 7. Teil, 1. Hälfte, Chlorophyceae (Grünalgen). Ordnung:Chlorococcales. Scweizerbart’sche Verlagsbuchhandlung, Stuttgart.
Krammer, K., Lange-Bertalot, H., 1986–1991. Bacillariophyceae. (Vol. 2)(1–4). In: Ettl, H., Gärtner, G., Heynig, H., Mollenhauer, D. (Eds.),Sübwasserflora von Mitteleuropa, Band 2/(14). G. Fischer Verlag.Lübeck, Jena, Stuttgart, pp. 1–876 (pp. 1–596, pp. 1–577, pp. 1–437).
Lampert, W., 1978. Field study on the dependence of the fecundity ofDaphnia spec. on food concentration. Oecologia 36, 363–369.
Lampert, W., 1982. Further studies on the inhibitory effect of the toxicblue-green Microcystis aeruginosa on the filtering rate of zooplankton.Arch. Hydrobiol. 95, 20–220.
Lampert, W., 1987. Laboratory studies as zooplankton–cyanobacteria inter-actions. N. Z. J. Mar. Freshwat. Res. 21, 483–490.
Lund, J.W.G., Kipling, C., Le Cren, E.D., 1958. The inverted microscopemethod of estimating algal numbers and the statistical basis of estima-tions by counting. Hidrobiologia 11, 143–170.
McDougal, R.L., Goldsborough, L.G., 1996. The effect of macrophyteexclusion and inorganic nutrient addition on the algal communities in aprairie wetland. UFS (Delta Marsh). Annual Report 31, 19–23.
Moore, M.V., Folt, C.F., Stemberger, R.S., 1996. Consequences of elevatedtemperatures for zooplankton assemblages in temperate lakes. Arch.Hydrobiol. 135, 289–319.
63N. Abrantes et al. / Acta Oecologica 29 (2006) 54–64

Müller-Navarra, D., Lampert, W., 1996. Seasonal patterns of food limitationin Daphnia galeata: separating food quantity and food quality effects. J.Plankton Res. 18, 1137–1157.
Nauwerck, A., 1960. Zur Systematik und Ökologie portugiesischer Plank-tonalgen. Bol. Soc. Brot. XI, 7–56.
Pereira, R., 1997. Plano de Ordenamento e Gestão das Lagoas das Braças eda Vela (CentroLitoral). M.Sc. thesis. University of Coimbra. Faculty ofSciences and Technology, Coimbra.
Pinel-Alloul, B., 1993. Zooplankton community structure in hardwatereutrophic and hypereutrophic lakes of Alberta. Water Sci. Technol. 27,353–361.
Radwan, S., Popiolek, B., 1989. Percentage of rotifers in spring zooplanktonin lakes of different trophy. Hydrobiologia 186/187, 235–238.
Reinikainen, M., Ketola, M., Walls, M., 1994. Effects of the concentrationsof toxic Microcystis aeruginosa and an alternative food on the survival ofDaphnia pulex. Limnol. Oceanogr. 39, 424–432.
Repka, S., 1996. Inter-and intraspecific differences in Daphnia life historiesin response to two food sources: the green alga Scenedesmus and thefilamentous cyanobacterium Oscillatoria. J. Plankton Res. 18, 1213–1224.
Repka, S., 1997. Effects of food type on the life history of Daphnia clonesfrom lakes differing in trophic state. I. Daphnia galeata feeding onScenedesmus and Oscillatoria. Freshwater Biol. 37, 675–683.
Rohrlack, T., Dittmann, E., Borner, T., Christoffersen, K., 2001. Effects ofcell-bound microcystins on survival and feeding of Daphnia spp. Appl.Environ. Microbiol. 67, 3523–3529.
Saunders, P.A., Porter, K.G., Taylor, B.E., 1999. Population dynamics ofDaphnia spp. and implications for trophic interactions in a small,monomictic lake. J. Plankton Res. 21, 1823–1845.
Scourfield, D.J., Harding, J.P., 1966. A key to the British freshwaterCladocera—third ed. Scientific Publication no. 5. Freshwater BiologicalAssociation, Ambleside.
Shapiro, J., 1995. Lake restoration by biomanipulation—a personal view.Environmental Review 3, 83–93.
Sommer, U., Gliwicz, M., Lampert, W., Duncan, A., 1986. The PEG-modelof seasonal succession of planktonic events in fresh waters. Arch. Hydro-biol. 106, 433–471.
Ter Braak, C.J.F., 1995. Ordination. In: Jongman, R.H.G., Ter Braak, C.J.F.,Tongeran, O.F.R. (Eds.), Data Analysis in Community and LandscapeEcology. Cambridge University Press, Cambridge, pp. 91–631.
Vanni, M.J., 1986. Competition in zooplankton communities: suppression ofsmall species by Daphnia pulex. Limnol. Oceanogr. 31, 1039–1056.
Vasconcelos, V.M., 1990. Ecotoxicologia de cianobactérias: impacte dosseus “blooms” nas comunidades animais e sua aplicação na gestão daqualidade da água. Provas de Aptidão Pedagógica e Científica. Univer-sity of Porto. Faculty of Sciences, Porto.
Wetzel, R.G., 1993. Limnologia. Fundação Calouste Gulbenkian, Lisboa.
64 N. Abrantes et al. / Acta Oecologica 29 (2006) 54–64
Copyright © 2022 FDOKUMEN




















