
Tectonic control of cave development: a case study of the Bystra Valley in the Tatra Mts., Poland

Biologia, Bratislava, 61/Suppl. 18: S203—S212, 2006 S203
Seasonal dynamics of chironomids in the profundal zoneof a mountain lake (Ľadové pleso, the Tatra Mountains, Slovakia)
Jolana Tátosová1 & Evžen Stuchlík2
1Institute for Environmental Studies, Faculty of Science, Charles University in Prague, Benátská 2, CZ-12801 Prague 2,Czech Republic; e-mail: [email protected] Station, Institute for Environmental Studies, Charles University in Prague, P.O. Box 47, CZ-38801 Blatná,Czech Republic; e-mail: [email protected]
Abstract: The profundal community of Ľadové pleso (an oligotrophic high mountain seepage lake at an altitude of 2,057m with a max. depth of 18 m and an ice-cover period from October – July) was studied from December 2000 – October2001. Chironomidae, the most significant part of the studied community, are represented by four taxa and dominated byMicropsectra radialis Goetghebuer, 1939 and Pseudodiamesa nivosa (Goethgebuer, 1928). These two species showed a 1-yearlife cycle. The total densities of chironomids varied from 0 to 5,927 ind. m−2; no chironomids, or very low densities, werefound during the winter/spring period, probably due to low oxygen concentrations in the medial part of the lake. These lowoxygen concentrations probably caused the relocation of larvae from the medial part of the sedimentary area at the sametime.
Key words: Non-biting midges, Chironomidae, life history, distribution, migration, environmental parameters, Slovakia.
Introduction
High mountain glacial lakes represent a very special en-vironment for water organisms because of their low av-erage annual temperature, oligotrophic character andthe minor impact of human activities. These specialproperties aroused interest in lakes in the High TatraMountains (Mts), although the accessibility of lakes wasdifficult, which especially complicated the investigationof the profundal sediments. The first investigation ofprofudnal fauna was carried out in the 1930s by Hraběand Zavřel. In contrast to lowland lakes or ponds, thefauna of the deepest part of high mountain lakes wasvery poor and was usually formed only by oligochaetesand the larvae of chironomids (Hrabě, 1939, 1942; Za-vřel, 1937). Later, the study of chironomids was con-nected with research of trophic status changes in someTatra lakes (Ertlová, 1964), and since the 1980s thechironomid fauna has been studied mainly with an em-phasis on the process of acidification (Ertlová, 1987;Tátosová, 2002; Bitušík et al., 2006). The samplingof chironomid larvae is often an important part of sys-tematic limnological research because of their very sen-sitive reaction to the amount and quality of availablefood, as well as temperature, concentration of dissolvedoxygen, and pH (Sæther, 1979; Raddum & Sæther,1981). Not only their abundances or taxonomic compo-sition, but also their life history, can reflect inclement
conditions of the mountain climate (Armitage et al.,1995). Ľadové pleso was chosen as the key lake in theTatra Mts for the Fifth Framework Program of Euro-pean Union: project EMERGE, which made possiblesystematic investigations of biota life cycles and sea-sonal variations in lake water chemistry.This paper summarizes results of the first complete
round-year study of chironomids in the profundal zoneof an oligotrophic high mountain Tatra lake. The mainaim of the presented study is to describe the populationdynamics of chironomids in Ľadové pleso in relation toenvironmental factors and phytoplankton production.
Study site
Ľadové pleso (49◦18′41′′ N, 20◦16′29′′ E) is located in theVeľká Studená dolina valley on the southern slope of theHigh Tatra Mts at 2,057 m a.s.l. The lake area is 1.72 ha,catchment area 12.3 ha, and maximum depth 18 m. Granitedominates in the catchment, and bare rocks cover 85% ofits area (KOPÁČEK et al., 2006). The lake has no visibleinflow or outflow, and the lake water level oscillates in-depthby more than 5 m during the year because of its seepagecharacter (TUREK, 2002; KŘEČEK et al., 2006). Majority ofthe lake bottom consists of rocks, and fine-grained sedimentis localized in the deepest part of the lake (Fig. 1). Thereare no fish in the lake.
Ľadové pleso is situated at high elevation, which influ-ences the duration of ice-cover and average annual tempera-

S204 J. Tátosová & E. Stuchlík
Fig. 1. Bathymetry of Ľadové pleso. The stars � indicate the coring sites and the arrows show the shift of the sample sites during theinvestigated period. The circles ⊗ indicate the positions of the emergence traps. The triangle marks � the place of the sedimentarytraps location.
ture. Despite its location and generally oligotrophic charac-ter, this lake is one of the most productive lakes in the HighTatra Mts, probably due to its seepage character (FOTTet al., 1987). This lake remained non-acidified during thepeak of acidification in this area (STUCHLÍK et al., 1985;FOTT et al., 1994; KOPÁČEK et al., 2000); nevertheless,a temporary and partial acidification of the upper part ofthe water column (to a depth of ∼5 m) has been repeatedlyrecorded at the end of the snow/ ice melting period, whenpH dropped below 6 in this part of the lake water volume(DARGOCKÁ et al., 1997; KNESLOVÁ et al., 1997; TUREK,2002).
Methods
Three sampling stations were chosen in the profundal zoneat depths varying from 15 to 18 m. Sites A and C weresituated at the edges of the sedimentary area, site B in themiddle of this area (Fig. 1). This location of sample sites waschosen so that the spatial distribution of the chironomid lar-vae would be recorded. The sample sites were moved slightlyin a clockwise direction at each sampling in order not totake samples from the same places and to obtain samplesfrom the whole sedimentary area. Sediment was obtainedby a Kajak corer with a sampling area of 28 cm2. Four coresamples were taken at each site, seven times in the periodfrom December 2000 to October 2001. In total, 84 sampleswere taken and processed; each sample was sieved througha 100 µm polypropylene mesh in the shape of a planktonnet (DAVIS, 1984) and stored in 4% formalin. Animals weresorted by hand in the laboratory, and head capsules werephotographed and measured using LUCIA software (Olym-pus C&S). They were then divided into four instar groups
based on size groups formed from the capsule width andlength measurements (Tab. 1).
Six emergence traps were installed above different lakedepths (Fig. 1). Traps with fixing solution could not be usedin Ľadové pleso because of concurrent analyses of organicpollutants in the lake water. The “live” emergence trapsused instead require daily control that was not possible atthis site, therefore the time of the trap exposition variedand the results are not suitable for the inference of chirono-mid biomass production. These installed traps were usedwith the aim to obtain chironomid imagoes for more reli-able identification.
Vertical stratification of physical and chemical param-eters (temperature, pH and dissolved oxygen) was measuredin situ by a Hydrolab H2O multi-parameter probe and datalogger Surveyor 3, (Hydrolab, USA) in 2 week intervals. Ver-tical samples for analyses of chlorophyll-a and total volumeof seston were taken 9 times from September 2000 to Octo-ber 2001, and during the winter period surface and bottomsamples were also taken on the following dates: 15 March,6 April, 11 May and 20 June. The water samples for de-termination of chlorophyll-a were filtered through What-man GF/C glass fiber filters, and after hot extraction ina 5 : 1 mixture of acetone : methanol (PECHAR, 1987)analyzed fluorometrically on a Turner TD-700 (Turner,USA). For more details of the procedure see FOTT et al.(1999). Samples were analyzed for total volume of seston(TVP3.3-16800, mm3 L−1) by filtration through a 40 µmmesh and determination with a Coulter Counter model ZBwith a tube of 70 µm aperture size (DARGOCKÁ et al., 1997).
The amount of particulate matter accumulated at thelake bottom was taken using a sediment trap, which wassuspended at a depth of 13 m (Fig. 1). The trap was formedby four 50 cm long tubes with a diameter of 6 cm. Durations
Seasonal dynamics of chironomids in Ľadové pleso S205
Table 1. Measured parameters of larval head capsules of M. radialis and P. nivosa.
Width (µm) Length (µm)
n Mean Min Max SE n Mean Min Max SE
M. radialisInstar: 1 5 79.26 64.28 96.06 11.83 6 83.36 74.74 93.25 7.50
2 50 131.75 104.65 154.48 9.80 49 134.43 108.32 150.73 11.713 84 208.27 163.15 240.94 15.17 87 214.10 174.48 242.03 13.564 122 318.51 251.64 376.91 21.20 124 327.51 260.85 388.94 24.20
P. nivosaInstar: 1 1 181.66 181.66 181.66 1 182.68 182.68 182.68
2 6 282.40 261.48 301.08 16.67 6 306.19 293.73 329.58 12.863 8 492.33 429.24 551.15 44.32 9 526.36 451.03 589.73 45.394 14 765.56 668.86 859.74 48.63 14 903.04 760.38 1100.42 82.78
Key: n – number of measurements; Max – maximum, Min – minimum, SE – standard error.
Table 2. Time intervals of the sedimentary traps exposure.
Start of End of Trap Duration ofan exposure an exposure depth an exposure (days)
8.12.2000 14.2.2001 13 6815.2.2001 23.5.2001 13 9724.5.2001 30.6.2001 13 373.7.2001 2.8.2001 13 303.8.2001 29.8.2001 13 2631.8.2001 27.9.2001 13 2730.9.2001 26.10.2001 13 26
of exposure are summarized in Table 2. TPV was analyzedfrom this material by the method described above.
Results
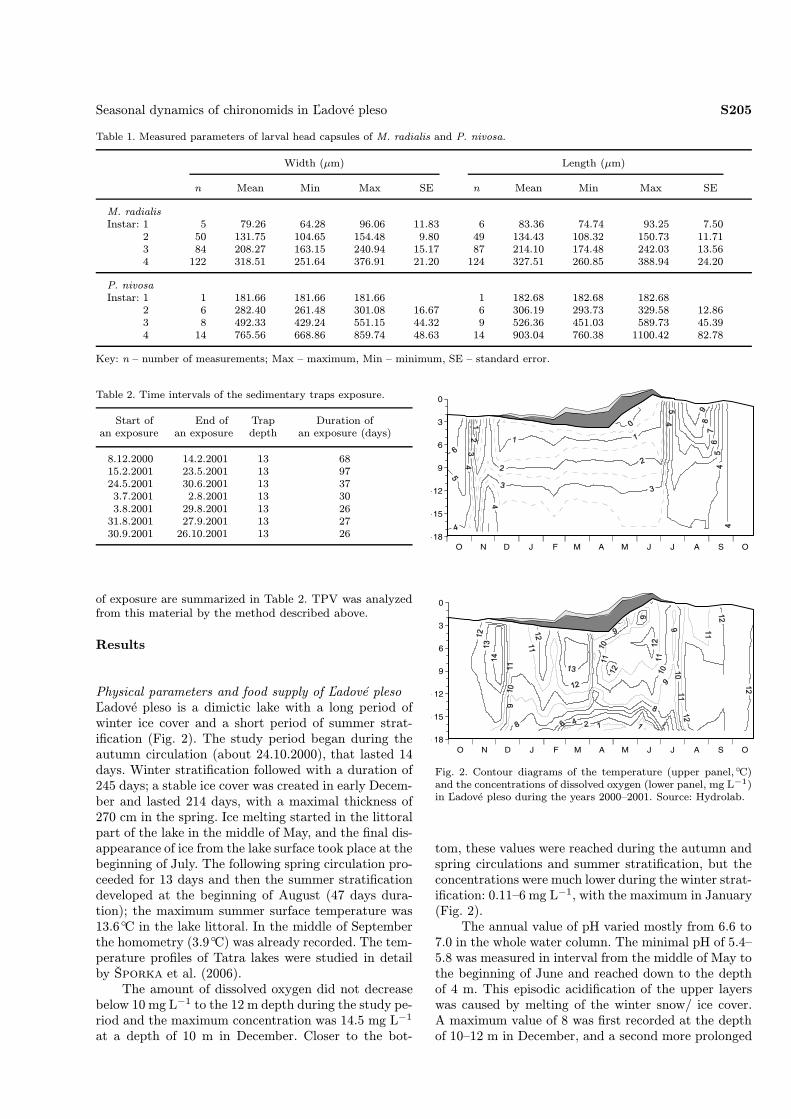
Physical parameters and food supply of Ľadové plesoĽadové pleso is a dimictic lake with a long period ofwinter ice cover and a short period of summer strat-ification (Fig. 2). The study period began during theautumn circulation (about 24.10.2000), that lasted 14days. Winter stratification followed with a duration of245 days; a stable ice cover was created in early Decem-ber and lasted 214 days, with a maximal thickness of270 cm in the spring. Ice melting started in the littoralpart of the lake in the middle of May, and the final dis-appearance of ice from the lake surface took place at thebeginning of July. The following spring circulation pro-ceeded for 13 days and then the summer stratificationdeveloped at the beginning of August (47 days dura-tion); the maximum summer surface temperature was13.6◦C in the lake littoral. In the middle of Septemberthe homometry (3.9◦C) was already recorded. The tem-perature profiles of Tatra lakes were studied in detailby Šporka et al. (2006).The amount of dissolved oxygen did not decrease
below 10 mg L−1 to the 12 m depth during the study pe-riod and the maximum concentration was 14.5 mg L−1
at a depth of 10 m in December. Closer to the bot-
-18
-15
-12
-9
-6
-3
0
O N D J F M A M J J A S O
-18
-15
-12
-9
-6
-3
0
O N D J F M A M J J A S O
Fig. 2. Contour diagrams of the temperature (upper panel,◦C)and the concentrations of dissolved oxygen (lower panel, mg L−1)in Ľadové pleso during the years 2000–2001. Source: Hydrolab.
tom, these values were reached during the autumn andspring circulations and summer stratification, but theconcentrations were much lower during the winter strat-ification: 0.11–6 mg L−1, with the maximum in January(Fig. 2).The annual value of pH varied mostly from 6.6 to
7.0 in the whole water column. The minimal pH of 5.4–5.8 was measured in interval from the middle of May tothe beginning of June and reached down to the depthof 4 m. This episodic acidification of the upper layerswas caused by melting of the winter snow/ ice cover.A maximum value of 8 was first recorded at the depthof 10–12 m in December, and a second more prolonged
S206 J. Tátosová & E. Stuchlík
Ch
iro
no
mid
ae [
ind
.m-2
]
0
1000
2000
3000
4000
5000
6000
7000
Chironomidae
Date
IX X XI XII I II III IV V VI VII VIII IX X XI
IX X XI XII I II III IV V VI VII VIII IX X XI
Ch
loro
ph
yll-
a [
µg
. L-1
]
0
5
10
15
20
Chlorophyll-a (0 m)Chlorophyll-a (5 m)Chlorophyll-a (8 m)Chlorophyll-a (bottom)
Ch
iro
no
mid
ae [
ind
. m-2
]
0
1000
2000
3000
4000
5000
6000
7000
Chironomidae
Date
IX X XI XII I II III IV V VI VII VIII IX X XI
IX X XI XII I II III IV V VI VII VIII IX X XI
TP
V [
mm
3 . L-1
]
0,0
0,5
1,0
1,5
2,0
2,5
3,0
TPV (0 m)TPV (5 m)TPV (8 m)TPV (bottom)
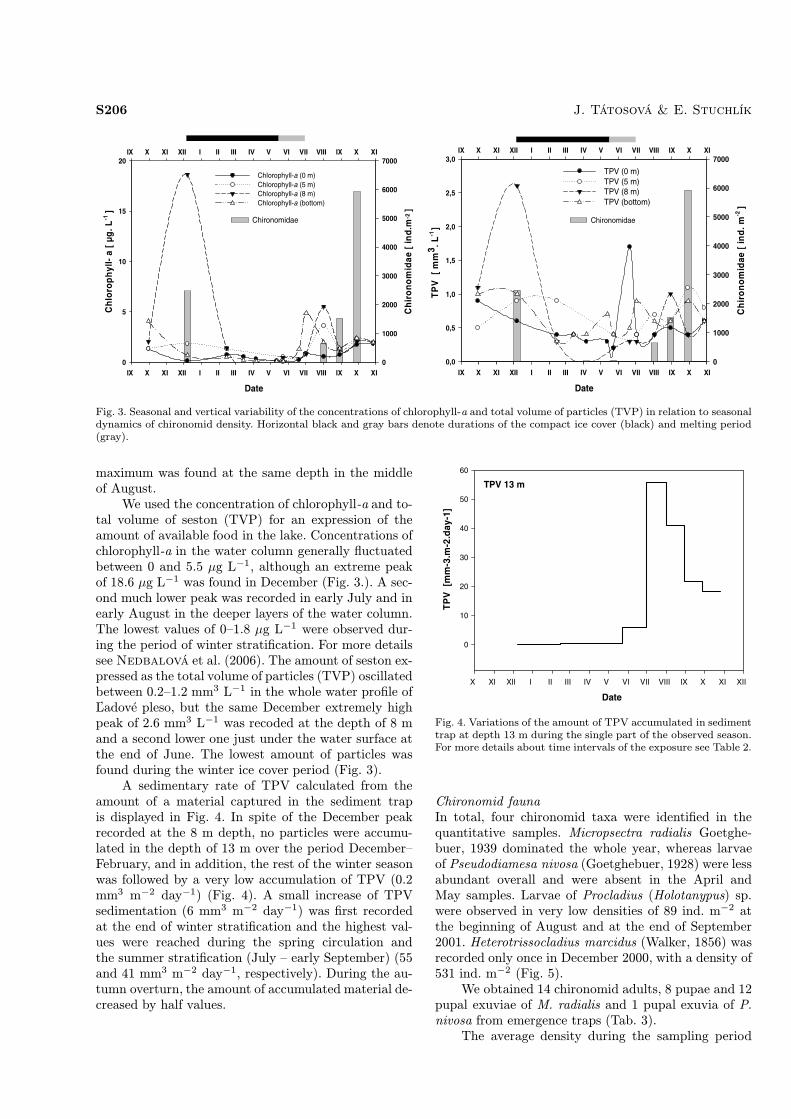
Fig. 3. Seasonal and vertical variability of the concentrations of chlorophyll-a and total volume of particles (TVP) in relation to seasonaldynamics of chironomid density. Horizontal black and gray bars denote durations of the compact ice cover (black) and melting period(gray).
maximum was found at the same depth in the middleof August.We used the concentration of chlorophyll-a and to-
tal volume of seston (TVP) for an expression of theamount of available food in the lake. Concentrations ofchlorophyll-a in the water column generally fluctuatedbetween 0 and 5.5 µg L−1, although an extreme peakof 18.6 µg L−1 was found in December (Fig. 3.). A sec-ond much lower peak was recorded in early July and inearly August in the deeper layers of the water column.The lowest values of 0–1.8 µg L−1 were observed dur-ing the period of winter stratification. For more detailssee Nedbalová et al. (2006). The amount of seston ex-pressed as the total volume of particles (TVP) oscillatedbetween 0.2–1.2 mm3 L−1 in the whole water profile ofĽadové pleso, but the same December extremely highpeak of 2.6 mm3 L−1 was recoded at the depth of 8 mand a second lower one just under the water surface atthe end of June. The lowest amount of particles wasfound during the winter ice cover period (Fig. 3).A sedimentary rate of TPV calculated from the
amount of a material captured in the sediment trapis displayed in Fig. 4. In spite of the December peakrecorded at the 8 m depth, no particles were accumu-lated in the depth of 13 m over the period December–February, and in addition, the rest of the winter seasonwas followed by a very low accumulation of TPV (0.2mm3 m−2 day−1) (Fig. 4). A small increase of TPVsedimentation (6 mm3 m−2 day−1) was first recordedat the end of winter stratification and the highest val-ues were reached during the spring circulation andthe summer stratification (July – early September) (55and 41 mm3 m−2 day−1, respectively). During the au-tumn overturn, the amount of accumulated material de-creased by half values.
X XI XII I II III IV V VI VII VIII IX X XI XII
0
10
20
30
40
50
60
TP
V [
mm
-3.m
-2.d
ay-1
]
Date
TPV 13 m
Fig. 4. Variations of the amount of TPV accumulated in sedimenttrap at depth 13 m during the single part of the observed season.For more details about time intervals of the exposure see Table 2.
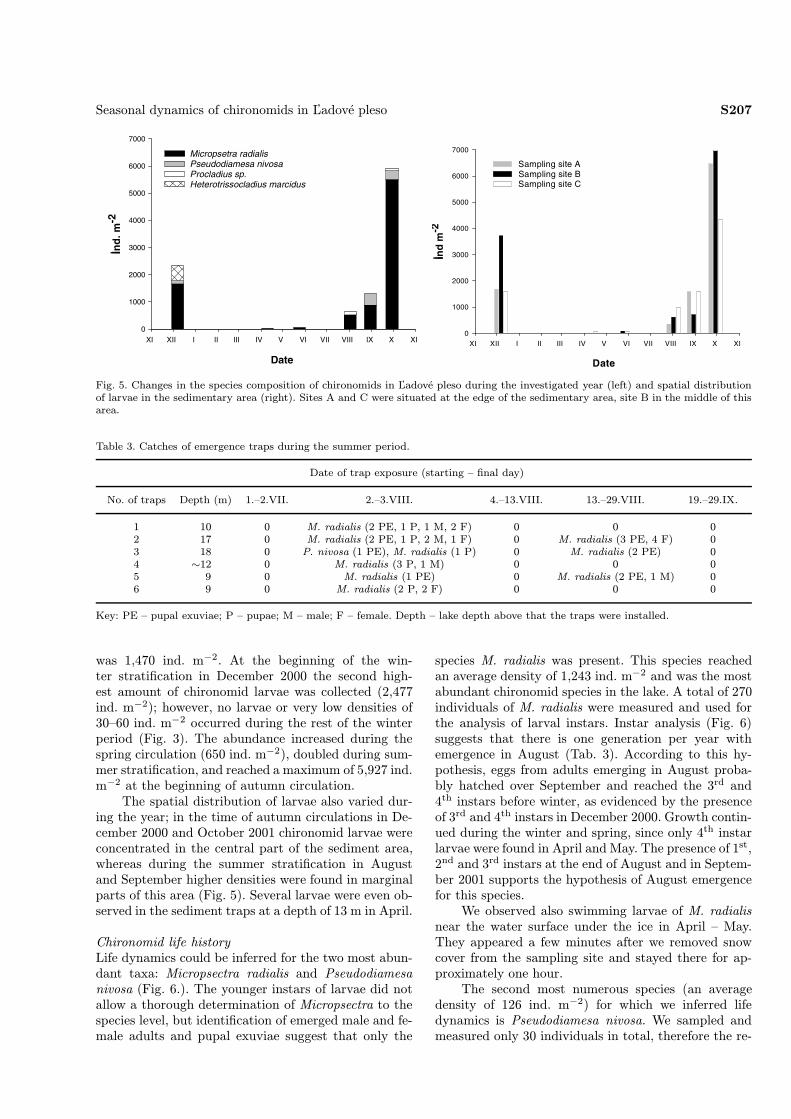
Chironomid faunaIn total, four chironomid taxa were identified in thequantitative samples. Micropsectra radialis Goetghe-buer, 1939 dominated the whole year, whereas larvaeof Pseudodiamesa nivosa (Goetghebuer, 1928) were lessabundant overall and were absent in the April andMay samples. Larvae of Procladius (Holotanypus) sp.were observed in very low densities of 89 ind. m−2 atthe beginning of August and at the end of September2001. Heterotrissocladius marcidus (Walker, 1856) wasrecorded only once in December 2000, with a density of531 ind. m−2 (Fig. 5).We obtained 14 chironomid adults, 8 pupae and 12
pupal exuviae of M. radialis and 1 pupal exuvia of P.nivosa from emergence traps (Tab. 3).The average density during the sampling period
Seasonal dynamics of chironomids in Ľadové pleso S207
Date
XI XII I II III IV V VI VII VIII IX X XI
ind
m- 2
0
1000
2000
3000
4000
5000
6000
7000
Sampling site A Sampling site B Sampling site C
Date
XI XII I II III IV V VI VII VIII IX X XI
ind
. m- 2
0
1000
2000
3000
4000
5000
6000
7000
Micropsetra radialisPseudodiamesa nivosa Procladius sp. Heterotrissocladius marcidus
Fig. 5. Changes in the species composition of chironomids in Ľadové pleso during the investigated year (left) and spatial distributionof larvae in the sedimentary area (right). Sites A and C were situated at the edge of the sedimentary area, site B in the middle of thisarea.
Table 3. Catches of emergence traps during the summer period.
Date of trap exposure (starting – final day)
No. of traps Depth (m) 1.–2.VII. 2.–3.VIII. 4.–13.VIII. 13.–29.VIII. 19.–29.IX.
1 10 0 M. radialis (2 PE, 1 P, 1 M, 2 F) 0 0 02 17 0 M. radialis (2 PE, 1 P, 2 M, 1 F) 0 M. radialis (3 PE, 4 F) 03 18 0 P. nivosa (1 PE), M. radialis (1 P) 0 M. radialis (2 PE) 04 ∼12 0 M. radialis (3 P, 1 M) 0 0 05 9 0 M. radialis (1 PE) 0 M. radialis (2 PE, 1 M) 06 9 0 M. radialis (2 P, 2 F) 0 0 0
Key: PE – pupal exuviae; P – pupae; M – male; F – female. Depth – lake depth above that the traps were installed.
was 1,470 ind. m−2. At the beginning of the win-ter stratification in December 2000 the second high-est amount of chironomid larvae was collected (2,477ind. m−2); however, no larvae or very low densities of30–60 ind. m−2 occurred during the rest of the winterperiod (Fig. 3). The abundance increased during thespring circulation (650 ind. m−2), doubled during sum-mer stratification, and reached a maximum of 5,927 ind.m−2 at the beginning of autumn circulation.The spatial distribution of larvae also varied dur-
ing the year; in the time of autumn circulations in De-cember 2000 and October 2001 chironomid larvae wereconcentrated in the central part of the sediment area,whereas during the summer stratification in Augustand September higher densities were found in marginalparts of this area (Fig. 5). Several larvae were even ob-served in the sediment traps at a depth of 13 m in April.
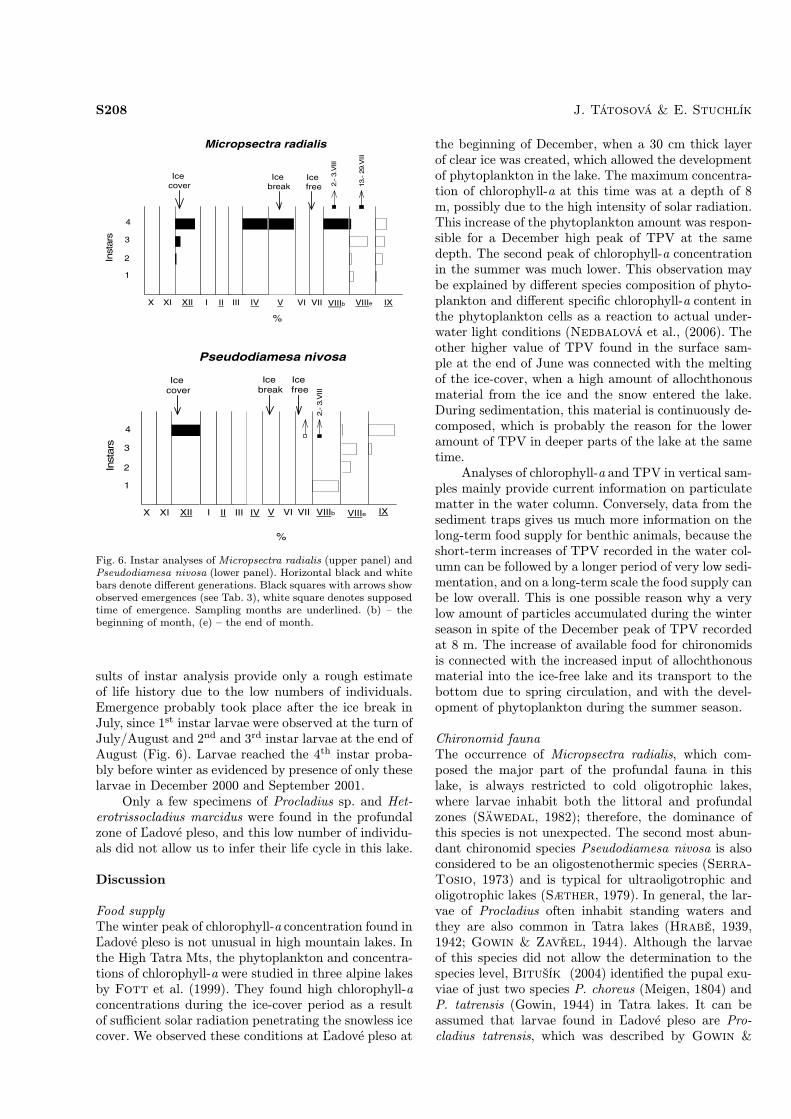
Chironomid life historyLife dynamics could be inferred for the two most abun-dant taxa: Micropsectra radialis and Pseudodiamesanivosa (Fig. 6.). The younger instars of larvae did notallow a thorough determination of Micropsectra to thespecies level, but identification of emerged male and fe-male adults and pupal exuviae suggest that only the
species M. radialis was present. This species reachedan average density of 1,243 ind. m−2 and was the mostabundant chironomid species in the lake. A total of 270individuals of M. radialis were measured and used forthe analysis of larval instars. Instar analysis (Fig. 6)suggests that there is one generation per year withemergence in August (Tab. 3). According to this hy-pothesis, eggs from adults emerging in August proba-bly hatched over September and reached the 3rd and4th instars before winter, as evidenced by the presenceof 3rd and 4th instars in December 2000. Growth contin-ued during the winter and spring, since only 4th instarlarvae were found in April and May. The presence of 1st,2nd and 3rd instars at the end of August and in Septem-ber 2001 supports the hypothesis of August emergencefor this species.We observed also swimming larvae of M. radialis
near the water surface under the ice in April – May.They appeared a few minutes after we removed snowcover from the sampling site and stayed there for ap-proximately one hour.The second most numerous species (an average
density of 126 ind. m−2) for which we inferred lifedynamics is Pseudodiamesa nivosa. We sampled andmeasured only 30 individuals in total, therefore the re-
S208 J. Tátosová & E. StuchlíkIn
star
s
Micropsectra radialis
1
2
3
4
%
XII II IVIII VI VIII V VIIIb VIIIe IXXIX
Ice cover
Ice break
Ice free 2.
- 3.
VIII
13.- 2
9.V
IIIPseudodiamesa nivosa
1
2
3
4
Inst
ars
Ice cover
%
Ice break
VI VIIV VIIIb VIIIe IX
Ice free
XII II IVIIIIXIX
2.- 3.
VIII
Fig. 6. Instar analyses of Micropsectra radialis (upper panel) andPseudodiamesa nivosa (lower panel). Horizontal black and whitebars denote different generations. Black squares with arrows showobserved emergences (see Tab. 3), white square denotes supposedtime of emergence. Sampling months are underlined. (b) – thebeginning of month, (e) – the end of month.
sults of instar analysis provide only a rough estimateof life history due to the low numbers of individuals.Emergence probably took place after the ice break inJuly, since 1st instar larvae were observed at the turn ofJuly/August and 2nd and 3rd instar larvae at the end ofAugust (Fig. 6). Larvae reached the 4th instar proba-bly before winter as evidenced by presence of only theselarvae in December 2000 and September 2001.Only a few specimens of Procladius sp. and Het-
erotrissocladius marcidus were found in the profundalzone of Ľadové pleso, and this low number of individu-als did not allow us to infer their life cycle in this lake.
Discussion
Food supplyThe winter peak of chlorophyll-a concentration found inĽadové pleso is not unusual in high mountain lakes. Inthe High Tatra Mts, the phytoplankton and concentra-tions of chlorophyll-a were studied in three alpine lakesby Fott et al. (1999). They found high chlorophyll-aconcentrations during the ice-cover period as a resultof sufficient solar radiation penetrating the snowless icecover. We observed these conditions at Ľadové pleso at
the beginning of December, when a 30 cm thick layerof clear ice was created, which allowed the developmentof phytoplankton in the lake. The maximum concentra-tion of chlorophyll-a at this time was at a depth of 8m, possibly due to the high intensity of solar radiation.This increase of the phytoplankton amount was respon-sible for a December high peak of TPV at the samedepth. The second peak of chlorophyll-a concentrationin the summer was much lower. This observation maybe explained by different species composition of phyto-plankton and different specific chlorophyll-a content inthe phytoplankton cells as a reaction to actual under-water light conditions (Nedbalová et al., (2006). Theother higher value of TPV found in the surface sam-ple at the end of June was connected with the meltingof the ice-cover, when a high amount of allochthonousmaterial from the ice and the snow entered the lake.During sedimentation, this material is continuously de-composed, which is probably the reason for the loweramount of TPV in deeper parts of the lake at the sametime.Analyses of chlorophyll-a and TPV in vertical sam-
ples mainly provide current information on particulatematter in the water column. Conversely, data from thesediment traps gives us much more information on thelong-term food supply for benthic animals, because theshort-term increases of TPV recorded in the water col-umn can be followed by a longer period of very low sedi-mentation, and on a long-term scale the food supply canbe low overall. This is one possible reason why a verylow amount of particles accumulated during the winterseason in spite of the December peak of TPV recordedat 8 m. The increase of available food for chironomidsis connected with the increased input of allochthonousmaterial into the ice-free lake and its transport to thebottom due to spring circulation, and with the devel-opment of phytoplankton during the summer season.
Chironomid faunaThe occurrence of Micropsectra radialis, which com-posed the major part of the profundal fauna in thislake, is always restricted to cold oligotrophic lakes,where larvae inhabit both the littoral and profundalzones (Sawedal, 1982); therefore, the dominance ofthis species is not unexpected. The second most abun-dant chironomid species Pseudodiamesa nivosa is alsoconsidered to be an oligostenothermic species (Serra-Tosio, 1973) and is typical for ultraoligotrophic andoligotrophic lakes (Sæther, 1979). In general, the lar-vae of Procladius often inhabit standing waters andthey are also common in Tatra lakes (Hrabě, 1939,1942; Gowin & Zavřel, 1944). Although the larvaeof this species did not allow the determination to thespecies level, Bitušík (2004) identified the pupal exu-viae of just two species P. choreus (Meigen, 1804) andP. tatrensis (Gowin, 1944) in Tatra lakes. It can beassumed that larvae found in Ľadové pleso are Pro-cladius tatrensis, which was described by Gowin &

Seasonal dynamics of chironomids in Ľadové pleso S209
Zavřel (1944), who found pupae and imagoes onlyfrom Tatra lakes situated above the tree line. The lastrecorded species, Heterotrissocladius marcidus, is theleast cold stenothermic member of this genus, but itis still restricted to relatively cold waters (Sæther,1975) and together with Procladius are the most com-mon taxa in the High Tatra Mts (Zavřel, 1937), eventhough its larvae occur in very low densities (Hrabě,1939). Paleolimnological studies of Tatra lakes supportthat H. marcidus is a stable but not numerous compo-nent of the chironomid fauna (Stuchlík et al., 2002,Šporka et al., 2002; Kubovčík et al., 2003).Chironomid taxa known from the profundal part
of Ľadové pleso are also found in other high mountainlakes in Europe. For example, the dominant species inĽadové pleso Micropsectra radialis was the only speciesfound in the profundal part of high mountain Lago diLatte Lake in the Alps (Cameron et al., 1997).M. radi-alis together with Heterotrissocladius marcidus, formedthe profundal chironomid assemblage in Lake Redo inthe Pyrenees, and together with Corynoneura arcticawere the only species in the deepest part of Lake LaCaldera (at 3,050 m a.s.l.) in the Sierra Nevada Mts.(SE Spain) (Rieradevall & Prat, 1999). Similar chi-ronomid compositions are found in deep high mountainlakes above the tree line in Austria (Bretschko, 1974).Not only a similar species composition but also low di-versity in general is known for high mountain and sub-arctic lakes. Low numbers of chironomid taxa as wererecorded in Ľadové pleso were also found in the profun-dal of several north Norwegian lakes (Aagaard, 1986);for example, in the similarly deep lakes (about 20 m)Austerdalsvatn and Haukvatn, Heterotrissocladius sub-pilosus was the only species recorded in the profun-dal part. Only Procladius sagittalis was collected in thePyrenean Aguilo Lake (Cameron et al., 1997). Boththe species composition and low diversity reflect thespecific nature of the altitude and latitude of extremelylocated lakes.The abundance of chironomid larvae usually does
not reach very high values in high mountain lakesmainly due to the low productivity of these lakes. Theaverage chironomid abundance of 1,470 ind. m−2 inthe profundal zone of Ľadové pleso is similar to thatrecorded by Brundin (1956) in the arctic ultraolig-otrophic Lake Kattejaure in northern Sweden, and Lin-degaard & Mæhl (1992) found the same density of1,400 ind. m−2 in the profundal zone of arctic Lake95 in South Greenland. In the profundal part of thementioned north Norwegian lakes, the total number ofchironomids didn’t exceed 1,000 ind. m−2 (Aagaard,1986). Similarly, high mountain lakes in other parts ofCentral Europe have shown chironomid densities of thismagnitude. For example Steinbock (1955) recordedfrom 300 to 2,800 ind. m−2 in eight Austrian moun-tain lakes situated from 2,000 to 2,800 m a.s.l., andBretschko (1974) found 2,100 ind. m−2 of chirono-mids in the Vorderer Finstertaler See (2,237 m a.s.l.).
Seasonal variationsThe variability of chironomid abundance was consider-able during the study period (from 0 to 5,927 ind. m−2),with the lowest densities recorded within the periodof the winter stratification (November – the beginningof July), when the concentration of dissolved oxygenas well as the supply of available food were very low(in spite of the high concentrations of chlorophyll-a atthe early winter stratification in December). These win-ter minima of both parameters are in close relation-ship. As mentioned above, the high concentrations ofchlorophyll-a recorded in December (18.6 µg L−1) arecommon in mountain lakes (Fott et al., 1999) andthey occur when compact ice cover without snow isformed, which transmits enough light. The sedimenta-tion and subsequent decomposition of high amounts ofphytoplankton can then cause a decline in the oxygencontent at the bottom of oligotrophic lakes. This lowwinter oxygen concentration probably caused the mi-gration of larvae from the sediment to the upper layerof the water column, as evidenced by the observationof swimming larvae, and relocation of larvae from thesedimentary area to the upper part of the lake bottom.Combined, these effects are probably the reason suchlow winter densities of chironomids were found in thislake. This migrational behavior of chironomid larvaeis one of many adaptations to low oxygen conditions(Heinis & Crommentuijn, 1992). Such behavior ofchironomids in a lowland Spanish lake has been pub-lished by Prat & Rieradevall (1995), but it has notbeen described in a mountain lake before.At the end of the winter stratification the increased
amount of the seston coming from the melting ice coverentered the lake and the dissolved oxygen concentra-tions at the lake bottom increased during the follow-ing spring circulation. Chironomid fauna responded tothese events with a slight increase in their abundancefrom 59 to 649 ind. m−2 at the beginning of August.This rise of abundance probably occurred due to re-versed migration of chironomids to the sedimentaryarea as evidenced by the presence of only overwintering4th instars of the dominant species Micropsectra radi-alis. Also, the spatial chironomid distribution showedhigher abundances in the marginal part of the sedimen-tary area then in the central part during this time pe-riod. To confirm this migrational hypothesis, a detailedstudy would be necessary with the possibility to takesamples from the sublittoral part of the lake bottom,which is composed of large boulders.The stable higher concentrations of chlorophyll-a,
TPV and dissolved oxygen and good temperature con-ditions following the period of summer stratificationestablished suitable conditions for the development ofthe chironomid fauna that reached the maximum abun-dance (5,927 ind. m−2) at the end of September, whenthe chironomid populations were composed of individ-uals of the new generation.Also, the presence of single species changed dur-

S210 J. Tátosová & E. Stuchlík
ing the investigated season. As mentioned above, larvaeof M. radialis are typical dwellers of cold oligotrophiclakes, where they inhabit both the littoral and profun-dal zones (Sawedal, 1982). Therefore, their stable oc-currence in the profundal part of Ľadové pleso duringthe whole year was expected. The species Pseudodi-amesa nivosa is also a typical inhabitant of ultraoligo-trophic and oligotrophic lakes, has been often found inthe littoral part of Tatra lakes (Hrabě, 1939, 1942;Ertlová, 1987); we also recorded them in abundancein the littoral of Ľadové pleso. The absence of theselarvae in the profundal zone during winter was presum-ably a result of the reaction to worsened oxygen con-ditions as well as to insufficient food supply. P. nivosaas a predator usually preys on smaller chironomids andother small organisms. As the littoral part of lakes istypically colonized by invertebrates more than the deepprofundal zone, it is possible to assume that larvae ofP. nivosa migrated from the profundal zone to the lit-toral during the winter period, where they stayed untilthe time of their emergence in July. This could also beone reason why we found so few specimens of the 1st
instar in the profundal at the beginning of August.Even though the larvae of Procladius are common
in Tatra lakes (Zavřel, 1937), very low densities andonly the sporadic presence of this species were recordedin Ľadové pleso. There is one possible explanation: asBrooks & Birks (2001) published, the temperatureoptimum of this species is about 11◦C, which is a tem-perature that was measured in Ľadové pleso only overa very short time in the summer and only in the lit-toral part of the lake. Ľadové pleso probably lies on theborder of the distribution area of this species, and itsdensities are affected by inter-annual air temperaturevariations.
Life historiesWe inferred life cycles for the two most abundant chi-ronomid taxa in Ľadové pleso. Preliminary results ofinstar analyses suggest that there is one generationper year, with emergence of P. nivosa in the Julyand M. radialis in the August. According to publisheddata, this hypothesis could be correct. Moore (1979)found that chironomids in the subarctic Great SlaveLake were all univoltine, and the same was reportedby Wiederholm et al. (1977) from a shallow subarcticlake in northern Sweden. Also, chironomids from otherhigh mountain lakes in C Europe are univoltine, as ob-served by Pechlaner et al. (1972) in Vorderer Finster-taler See (2237 m a.s.l.) and Laville & Gaini (1974)in Lake Port Biehl (Central Pyrenees, 2285 m a.s.l.).Through the use of ash free dry weight measured
for individual larvae, Lindegaard & Mæhl (1992)discovered two different cohorts of Micropsetra withemergence in June and September, which belonged,however, to the different univoltine species M. brun-dini and M. groenlandica. On the other hand, it isknown that some species from the cold Holarctic re-
gion need more than one year to complete their devel-opment (Armitage et al., 1995). For instance,Welch(1976) found a long life cycle of 3 years for Heterotris-socladius oliveri in the high arctic Lake Char. In sub-arctic Lake Thingvallavatn in Greenland, Lindegaard(1992) assumed a 2-year life cycle for Chironomus is-landicus, whose 4th instar larvae of both younger andolder generations were distinguished by their averagelarval weight. Even though we didn’t weigh collectedanimals to confirm or disprove either the bivoltine orsemivoltine life cycle of Micropsectra, that only 4th in-star larvae of M. radialis were found in the ice-coverperiod and that there was only the one August periodof theM. radialis emergence suggest that there was onlyone generation of this species. In addition, the ice freeperiod of Ľadové pleso lasts 5–6 months, which meansa relatively long growth season for chironomids everyyear.Because of the disappearance of Pseudodiamesa
nivosa larvae from the profundal part of the lake wehave no information about their winter development.We assume that P. nivosa migrated from the profun-dal zone to the littoral due to worsening life conditions,where they probably stayed until the time of their emer-gence in July. We found a few specimens of the 1st in-star in the profundal at the beginning of August, whichsuggests that a majority of the population lived in thelittoral after hatching. As was found by Lindegaard(1992), the growth of the littoral population of thisspecies can be very fast after hatching. This fast growthrate in the littoral zone and relatively long ice-free pe-riod can support the univoltine life cycle of this species;however, a more detailed study is necessary to confirmthis hypothesis.
Acknowledgements
We wish to thank P. BITUŠÍK for identifying the Microp-sectra species and for revision of the identified chironomidtaxa. We also wish to thank our colleagues for technical as-sistance during field work and D. HARDEKOPF for linguisticcorrection of the manuscript. This study was enabled by theFP 5 EC project EMERGE (EVK1-CT 1999-00032, address:www.mountain-lakes.org).
References
AAGAARD, K. 1986. The chironomid fauna of North Norwegianlakes, with a discussion on methods of community classifica-tion. Holarctic Ecology 9: 1–12.
ARMITAGE, P.D., CRASTON, P.S. & PINDER, L.V.C. 1995. TheChironomidae: Biology and ecology of non-biting midges.Chapman & Hall, London, 572 pp.
BITUŠÍK, P. 2004. Chironomids (Diptera: Chironomidae) of themountain lakes in the Tatra Mts. (Slovakia). A review.Dipterologica Bohemoslovaca 12, Acta Fac. Ecol., Zvolen 12,Suppl. 1: 25–33.
BITUŠÍK, P., SVITOK, M., KOLOŠTA, P. & HUBKOVÁ, M. 2006.Classification of the Tatra Mountain lakes (Slovakia) usingchironomids (Diptera, Chironomidae). Biologia, Bratislava61, Suppl. 18: S191–S201.

Seasonal dynamics of chironomids in Ľadové pleso S211
BRETSCHKO, G. 1974. The chironomid fauna of high alpine lake(Vorderer Finstertaler See, Tyrol, Austria, 2237 m a.s.l.). En-tomol. Tidskr., Supplement 95: 22–33.
BROOKS, S.J. & BIRKS, H.J.B. 2001. Chironomid-inferred airtemperatures from Lateglacial and Holocene sites in north-west Europe: progress and problems. Quaternary Science Re-views 20: 1723–1741
BRUNDIN, L. 1956. Die bodenfaunistischen Seetypen und ihreAnwendbarkeit auf der Sudhalbkugel. Zugleich eine Theorieder produktionsbiologischen Bedeutung der glazialen Erosion.Rep. Inst. Freshwater Res. Drottingholm 37: 192–235.
CAMERON, N., FJELLHEIM, A., RIERADEVALL, M., RADDUM,G.G., SCHNELL, O., FOTT, J., STUCHLÍK, E., ČERNÝ, M.& KOPÁČEK, J. 1997. Contemporary biology, pp. 1–60. In:WATHNE, B.M., PATRICK, S. & CAMERON, N. (eds) AL:PE –Acidification of Mountain Lakes: Palaeolimnology and Ecol-ogy. Part 2 – Remote Mountain Lakes as Indicators of AirPollution and Climate Change, Norwegian Institute for Wa-ter Research Report No. 3638, 1997.
DARGOCKÁ, J., KNESLOVÁ, P. & STUCHLÍK, E. 1997. Phyto-plankton of several high mountain lakes in different stage ofacidification. Štúdie TANAP 2: 41–62.
DAVIS, I.J. 1984. Sampling aquatic insect emergence, pp. 161–227. In: DOWNING, J.A. & RIGLER, F.H. (eds) A manual onmethods for the assessment of secondary productivity in freshwaters, Blackwell Scientific Publications, Oxford, England.
ERTLOVÁ, E. 1964. Príspevok k poznaniu zoobentosu Poprad-ského plesa. Biologia, Bratislava 19: 666–674.
ERTLOVÁ, E. 1987. Chironomids (Chironomidae, Diptera) of thelittoral of the selected lakes in the High Tatras. Acta Fac.Rerum Nat. Univ. Comen., Zool. 29: 53–66.
FOTT, J., BLAŽO, M., STUCHLÍK, E. & STRUNECKÝ, O. 1999.Phytoplankton in three Tatra Mountain lakes with differentacidification status. J. Limnol. 52: 107–116.
FOTT, J., PRAŽÁKOVÁ, M., STUCHLÍK, E., & STUCHLÍKOVÁ Z.1994. Acidification of lakes in Šumava (Bohemia) and in theHigh Tatra Mountains (Slovakia). Hydrobiologia, 274: 37–47.
FOTT, J., STUCHLÍK, E. & STUCHLÍKOVÁ, Z. 1987. Acidifica-tion of lakes in Czechoslovakia, pp. 77–79. In: MOLDAN, B.& PAČES, T. (eds) Extended abstracts of the Internationalworkshop on geochemistry and monitoring in representativebasins, Geological Survey, Prague.
GOWIN, F. & ZAVŘEL, J. 1944. Nový Procladius z VysokýchTater. Folia Entomol. 7: 8–90.
HEINIS, F. & CROMMENTUIJN, T. 1992. Behavioural responsesto changing oxygen concentrations of deposit feeding chi-ronomind larvae (Diptera) of littoral and profundal habitats.Arch. Hydrobiol. 124: 173–185.
HRABĚ, S. 1939. Bentická zvířena tatranských jezer. SborníkKlubu Přírodovědců v Brně 22: 1–13.
HRABĚ, S. 1942. O bentické zvířeně jezer ve Vysokých Tatrách.Bohemica 25: 123–177.
KNESLOVÁ, P., DARGOCKÁ, J. & STUCHLÍK, E. 1997. Zooplank-ton of eight the Tatra Mountain lake in different stage ofacidification. Štúdie TANAP 2: 123–134.
KOPÁČEK, J., STUCHLÍK, E. & HARDEKOPF, D. 2006. Chemi-cal composition of the Tatra Mountain lakes: Recovery fromacidification. Biologia, Bratislava 61, Suppl. 18: S21–S33.
KOPÁČEK, J., STUCHLÍK, E., STRAŠKRABOVÁ, V. & PŠENÁKO-VÁ, P. 2000. Factors governing nutrient status of mountainlakes in the Tatra Mountains. Freshwater Biol. 43: 369–383.
KŘEČEK, J., TUREK, J., LJUNGREN, E., STUCHLÍK, E. &ŠPORKA, F. 2006. Hydrological processes in small catchmentsof mountain headwater lakes: The Tatra Mountains. Biologia,Bratislava 61, Suppl. 18: S1–S10.
KUBOVČÍK, V., BETÁK, M. & FEČKANINOVÁ, G. 2003. Sub-fosilná fauna pakomárov (Diptera: Chironomidae) Ľadovéhoplesa (Vysoké Tatry, Slovensko), pp. 201–203. In: BITUŠÍK,P. & NOVIKMEC, M. (eds) Proc. 13th Conference of SlovakLimnol. Soc. & Czech Limnol. Soc., Banská Štiavnica, June2003, Acta Facultatis Ecologiae, Zvolen 10, Suppl. 1.
LAVILLE, H. & GAINI, N. 1974. Phénologie et cycles biologiquesdes chironomides de la zone littorale (0–7 m) du lac de Port-Bielh (Pyrénées centrales). Entomol. Tidskr., Suppl. 95: 139–155.
LINDEGAARD, C. 1992. Zoobenthos ecology of Thingvallavatn:vertical distribution, abundance, population dynamics andproduction. Oikos 64: 257–304.
LINDEGAARD, C. & MÆHL P. 1992. Abundance, populationdynamics and production of Chironomidae (Diptera) in anultraoligotrophic lake in South Greenland. Netherlands J.Aquat. Ecol. 26 (2–4): 297–308.
MOORE, J.W. 1979. Some factors influencing the distribution,seasonal abundance and feeding of subarctic Chironomidae(Diptera). Arch. Hydrobiol. 85: 302–325.
NEDBALOVÁ, L., STUCHLÍK, E. & STRUNECKÝ, O. 2006. Phyto-plankton of a mountain lake (Ľadové pleso, the Tatra Moun-tains, Slovakia): Seasonal development and first indications ofa response to decreased acid deposition. Biologia, Bratislava61, Suppl. 18: S91–S100.
PECHAR, L. 1987. Use on an acetone : methanol mixturefor the extraction and spectrophotometric determination ofchlorophyll-a in phytoplankton. Arch. Hydrobiol. Suppl. 78:99–117.
PECHLANER, R., BRETSCHKO, G., GOLLMANN, P., PFEIFER, H.,TILZER, M. & WEISSENBACH, H.P. 1972. The productionprocesses in two high-mountain lakes (Vorderer and HintererFinstertaler See, Kuthai, Austria), pp. 239–269. In: KAJAK,Z. & HILLBRICHT-ILKOWSKA, A. (eds) Productivity prob-lems of freshwaters, PWN Pol. Sci. Publ., Warsawa.
PRAT, N. & RIERADEVALL, M. 1995. Life cycle and productionof Chironomidae from the karstic Lake Banyoles (NE Spain).Freshwater Biol. 33: 511–524.
RADDUM, G.G. & SÆTHER, O.A. 1981. Chironomid communi-ties in Norwegian lakes with different degrees of acidification.Verh. Int. Verein. Limnol. 21: 399–405.
RIERADEVALL, M. & PRAT, N. 1999. Chironomidae from highmountain lakes in Spain and Portugal, pp. 605–613. In: HOF-FRICHTER, O. (ed.) Late 20th century research on Chirono-midae: An Anthology from the 13th International Symposiumon Chironomidae, Shaker Verlag, Aachen.
SAWEDAL, L. 1982. Taxonomy, morphology, phylogenetic re-lationships and distribution of Micropsectra Kieffer, 1909(Diptera: Chironomidae). Entomol. Scand. 13: 371–400.
SÆTHER, O.A. 1975. Nearctic and Palaearctic Heterotrissocla-dius (Diptera: Chironomidae). Bull. Fish. Res. Board Can.139: 27–36.
SÆTHER, O.A. 1979. Chironomid communities as indicators oflake typology. Verh. Int. Verein. Limnol. 19: 3127–3133.
SERRA-TOSIO, B. 1973. Ecologie et biogeography des Diamesinid’Europe (Diptera, Chironomidae). Trav. Lab. Hydrobiol.Piscic. Univ. Grenoble 63: 5–175.
STEINBOCK, O. 1955. Uber die Verhaltnisse in der Hochge-birgsseen. Mem. Inst. Ital. Idrobiol., Suppl. 8: 311–343.
STUCHLÍK, E., APPLEBY, P., BITUŠÍK, P., CURTIS, C., FOTT, J.,KOPÁČEK, J., PRAŽÁKOVÁ, M., ROSE, N., STRUNECKÝ, O.& WRIGHT, R.F. 2002. Reconstruction of long-term changesin lake water chemistry, zooplankton and benthos of a small,acidified high-mountain lake: Magic modeling and paleolim-nological analysis. Water Air Soil Poll.: Focus 2: 127–138.
STUCHLÍK, E., STUCHLÍKOVÁ, Z., FOTT, J., RŮŽIČKA, L. &VRBA, J. 1985. Effect of acid precipitation on waters of theTANAP territory. Zborník TANAP 26: 173–211.
ŠPORKA, F., LIVINGSTONE, D.M., STUCHLÍK, E., TUREK, J. &GALAS, J. 2006. Water temperatures and ice cover in lakesof the Tatra Mountains. Biologia, Bratislava 61, Suppl. 18:S77–S90.
ŠPORKA, F., ŠTEFKOVÁ, E., BITUŠÍK, P., THOMPSON, A.R.,AGUSTI-PANAREDA, A., APPLEBY, P., GRYTNES, J.A., KA-MENIK, C., KRNO, I., LAMI, A., ROSE, N. & SHILLAND, N.E.2002. The paleolimnological analysis of sediment from highmountain lake Nižné Terianske pleso in the High Tatras (Slo-vakia). J. Paleolimnol. 28: 95–109.

S212 J. Tátosová & E. Stuchlík
TÁTOSOVÁ, J. 2002. Makrozoobentos profundálu jezer v oblastiVysokých Tater. Thesis, Faculty of Science, Charles Univer-sity in Prague, 66 pp.
TUREK, J. 2002. Hydrologický režim vysokohorských jezerv oblasti Vysokých Tater. Thesis, Faculty of Science, CharlesUniversity in Prague, 82 pp.
WELCH, H.E. 1976. Ecology of Chironomidae (Diptera) in a polarlake. J. Fish. Res. Board Can. 33: 227–247.
WIEDERHOLM, T., DANELL, K. & SJOBERG, K. 1977. Emergenceof chironomids from a small man-made lake in northern Swe-den. Norw. J. Entomol. 24: 99–105.
ZAVŘEL, J. 1937. Orthocladiinen aus der Hohen Tatra. Verh. Int.Verein. Limnol. 7: 483–496.
Received September 2, 2005Accepted May 9, 2006
Copyright © 2022 FDOKUMEN




















