
Saxicolous and riparian vegetation of a piedmont in central-western Argentina
13
Journal of Arid Environments (1999) 42: 305–317 Article No. jare.1999.0517 Available online at http://www.idealibrary.com on Saxicolous and riparian vegetation of a piedmont in central-western Argentina E. Martinez Carretero* Botany and Plant Sociology, Argentine Institute of Arid Lands Researches (IADIZA), CC 507, 5500 Mendoza, Argentina (Received 31 July 1998, accepted 24 March 1999) Four plant communities were determined through floristical and non-metric analysis: a Dolichlasium lagascae community in crevices on sunny rock out- crops; a Cercidium praecox ssp. glaucum community on sunny slopes; an Artemisia mendozana}Adesmia trijuga community on shady slopes; and a Eupatorium buniifolium community on dry riverbanks. The discriminant func- tion analysis showed that the first group corresponds to the sunny rock outcrops discriminated by Na content (r"0·53, p(0·05), the second group is for sunny slopes discriminated by electric conductivity and the third for shady slopes}riparian discriminated by the (Ca#Mg) content (r"0·54, p(0·05). A pattern of primary succession on outcrop surfaces is proposed. ( 1999 Academic Press Keywords: arid; outcrop; saxicolous vegetation; xerosere Introduction An outstanding feature of saxicolous vegetation is its occurrence in environments where microtopography plays a major role. There is much information concerning the com- position and succession of outcrop plant communities, though most of it deals with granite outcrops (Baker, 1956; Burbank & Platt, 1964; Shure & Ragsdale, 1977; Funes & Cabido, 1995; Wiser et al., 1996). Only a few ecological and floristic studies on saxicolous vegetation have been made in Argentina, generally because of their limited extent and frequent inaccessibility. Frangi (1975) described saxicolous vegetation for the Sierra de Tandil, Cabido et al. (1990) for granite outcrops of the Sierras de Co H rdoba and Funes & Cabido (1995) analysed local and regional variability in the saxicolous vegetation of the Sierras Grandes in Co H rdoba Province. Roig (1976, 1989) studied the plant communities of the piedmont in the Province of Mendoza, and described saxicolous communities present on shady (Adesmia trijuga community) and sunny slopes (Astericium glaucum Hieron. et Wolff community). Martinez Carretero (1985, 1987) and Martinez Carretero & Roig (1994) studied the ecological aspects of these communities, particularly those related to water erosion and soil characteristics. Associated with the Andean tectonics that began during the Upper Tertiary and continued until the Lower Quaternary, the rock outcrops of the Andean piedmont are primarily composed of consolidated Triassic sandstone (Barton & Rodriguez, 1989) *(E-mail: mcarrete@lab.cricyt.edu.ar). 0140-1963/99/080305#13 $30.00/0 ( 1999 Academic Press
Transcript of Saxicolous and riparian vegetation of a piedmont in central-western Argentina

Journal of Arid Environments (1999) 42: 305–317Article No. jare.1999.0517Available online at http://www.idealibrary.com on
Saxicolous and riparian vegetation of a piedmont incentral-western Argentina
E. Martinez Carretero*
Botany and Plant Sociology, Argentine Institute of Arid Lands Researches(IADIZA), CC 507, 5500 Mendoza, Argentina
(Received 31 July 1998, accepted 24 March 1999)
Four plant communities were determined through floristical and non-metricanalysis: a Dolichlasium lagascae community in crevices on sunny rock out-crops; a Cercidium praecox ssp. glaucum community on sunny slopes; anArtemisia mendozana}Adesmia trijuga community on shady slopes; and aEupatorium buniifolium community on dry riverbanks. The discriminant func-tion analysis showed that the first group corresponds to the sunny rockoutcrops discriminated by Na content (r"0·53, p(0·05), the second groupis for sunny slopes discriminated by electric conductivity and the third forshady slopes}riparian discriminated by the (Ca#Mg) content (r"0·54,p(0·05). A pattern of primary succession on outcrop surfaces is proposed.
( 1999 Academic Press
Keywords: arid; outcrop; saxicolous vegetation; xerosere
Introduction
An outstanding feature of saxicolous vegetation is its occurrence in environments wheremicrotopography plays a major role. There is much information concerning the com-position and succession of outcrop plant communities, though most of it deals withgranite outcrops (Baker, 1956; Burbank & Platt, 1964; Shure & Ragsdale, 1977; Funes& Cabido, 1995; Wiser et al., 1996). Only a few ecological and floristic studies onsaxicolous vegetation have been made in Argentina, generally because of their limitedextent and frequent inaccessibility. Frangi (1975) described saxicolous vegetation forthe Sierra de Tandil, Cabido et al. (1990) for granite outcrops of the Sierras de CoH rdobaand Funes & Cabido (1995) analysed local and regional variability in the saxicolousvegetation of the Sierras Grandes in CoH rdoba Province. Roig (1976, 1989) studied theplant communities of the piedmont in the Province of Mendoza, and describedsaxicolous communities present on shady (Adesmia trijuga community) and sunnyslopes (Astericium glaucum Hieron. et Wolff community). Martinez Carretero(1985, 1987) and Martinez Carretero & Roig (1994) studied the ecological aspects ofthese communities, particularly those related to water erosion and soil characteristics.
Associated with the Andean tectonics that began during the Upper Tertiary andcontinued until the Lower Quaternary, the rock outcrops of the Andean piedmont areprimarily composed of consolidated Triassic sandstone (Barton & Rodriguez, 1989)
*(E-mail: [email protected]).
0140-1963/99/080305#13 $30.00/0 ( 1999 Academic Press

306 E. MARTINEZ CARRETERO
which is usually friable and cemented with calcium carbonate. This intense tectonicactivity resulted in Triassic and Tertiary strata dipping in opposite directions to thegeneral slope (Duffar, 1983).
Two types of rock surfaces can be recognized in connection with outcrops, namelyexposed rocks and associated down-slopes with materials resulting from in situ weather-ing of the rock surface. Low plant cover on these rocks, restricted either to crevices(cracks and fissures) or to shallow substrate accumulated on level parts of slopes, alongwith climatic aridity and marked topographical differences, determines the land-scape of these outcrops.
Outcrop vegetation is therefore an interesting topic for study given the criticalenvironments in which it grows (shallow soil, low soil moisture, high soil warmingparticularly on exposed rocks and sunny north-facing slopes). On the other hand, mostoutcrops are dissected by dry rivers (wadis), where close relationships between riparianand saxicolous communities can be found.
This paper describes the plant communities associated with 12 outcrops to gainfurther knowledge on the floristic relationships between saxicolous and riparian plantcommunities, and to propose a pattern of primary succession on outcrop surfaces ina geographic area that is poorly understood, both in Argentina and elsewhere in theworld.
Methods
Study area
The piedmont area (Fig. 1) shows a structure of folds and faults, with winding axesstretching mostly in a NNE}SSW direction (Barton & Rodriguez, 1989). Being anancient eroded area, highly dissected from west to east, it exhibits alternate north– andsouth-facing slopes with gradients between 303 and 453, where extensive Tertiary rockoutcrops have been left exposed by water erosion. The Andean piedmont exhibitsa great number of rock outcrops, some extending over 100}200 m. Soils are indicated asEntisols or Lithic on the rock outcrops, and Torripsament is typical of the riverbanks(Masotta & Berra, 1996).
Climatic data are shown in Fig. 2. Mean annual rainfall in 196 mm, 72% of whichoccurs in summer (October}March) (Servicio MeteoroloH gico Nacional, 1958). Accord-ing to the pluviothermal equation (Emberger, 1971) the area reaches the valueQ"19·36, which corresponds to arid bioclimatic conditions (Martinez Carretero,1985).
All observations were made on the piedmont locality of Sierras de Uspallata, 10 kmwest of Mendoza, at an altitude of 1400 m. The dominant vegetation is a Larreadivaricata Cav. mature community (Larreetea divaricatae-cuneifoliae Roig 1989; pied-mont shrubby vegetation).
Sampling and data analysis
A total of 43 floristic releveH s were carried out on 12 Triassic outcrops (PotrerillosFormation), on their adjacent slopes, and in the dry rivers that dissect their surface(Table 1). Only the most contrasting and floristically dissimilar exposure were con-sidered, namely the shady and sunny slopes with inclinations of 40}453. The outcropswere fully sampled following the crevices. All vascular plants were recorded and coverwas estimated using the Braun-Blanquet scale (r41%; #"1}10%; 1"11}20%;2"21}40%; 3"41}60%; 4"61}80%; 5"'81% ). The life-form of each specieswas recorded in each community and is indicated in Table 1.
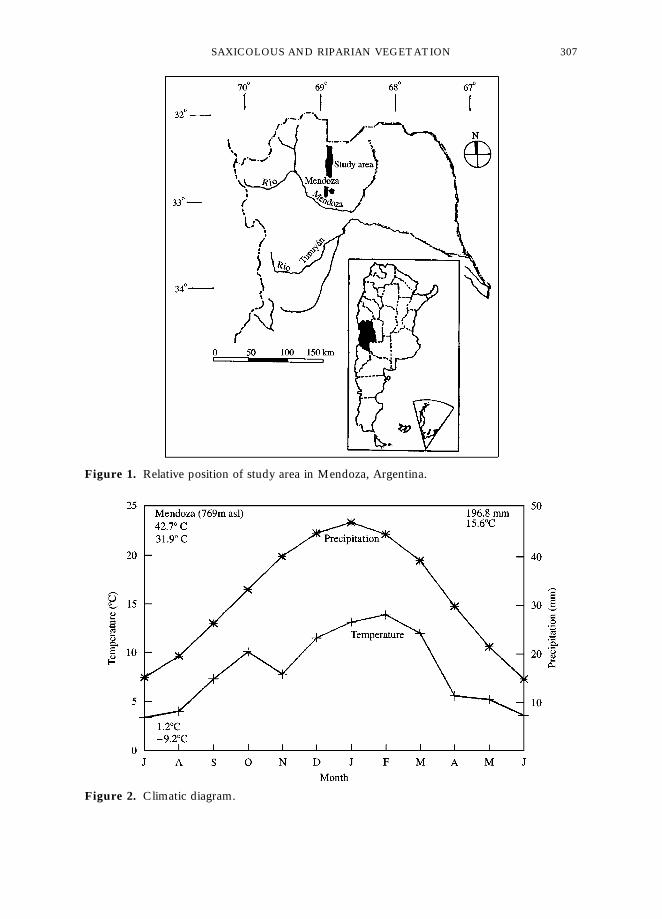
Figure 1. Relative position of study area in Mendoza, Argentina.
Figure 2. Climatic diagram.
SAXICOLOUS AND RIPARIAN VEGETATION 307

Table 1. Table of releveHs: (a ) Dolichlasium lagascae community; (b) Cer-cidium praecox community; (c) Eupatorium buniifolium community;(d) Artemisia mendozana community. Lf"life-form
308 E. MARTINEZ CARRETERO
A total of 48 soils samples, four per outcrop, were collected from the materialaccumulated in crevices and in riverbanks: (i) on sunny rock outcrops in cracks up to5 cm wide and 5}8 cm deep; (ii) on sunny removed slopes in fissures up to 10 cm wideand 10}12 cm deep; and (iii) on shady slopes in fissures up to 10}15 cm wide and20}25 cm deep. All samples weighted 250}300 g. Soil parameters, i.e. moisture (as percent dry weight), pH, (Ca#Mg) (EDTA tritation method), Na content (photometry offlame method), organic matter (Kjeldahl method) and electrical conductivity, weredetermined for each sample (Table 2). Depth and width of crevices were measured witha measuring tape.
Data were subjected to cluster analysis and non-metric multi-dimensional scalinganalysis (Beals, 1984). Each dominance}abundance value was replaced with the corre-sponding mean cover value in the Braun-Blanquet cover abundance scale (r,#"2·5;1"5; 2"17·5; 3"37·5; 4"62·5; 5"87·5), and transformed by the logarithmicfunction. The similarity matrix was calculated using Pearson’s correlation. The com-plete linkage method was used to produce the classification dendrogram. The relation-ship between soil parameters and each community was analysed using the discriminantfunction analysis (Wildi & Orloci, 1983). The correlation between soil parameters andreleveH s was calculated using Pearson’s correlation analysis.

Tab
le2.
Soi
lpa
ram
eter
sin
crev
ices
ofro
ckou
tcro
psan
dsu
nny
slop
es,
and
shad
ysl
opes
and
rive
rban
ks
pHN
a(C
a#M
g)H3
Org
anic
Mat
ter
EC
(me
l~1)
(%)
(g%
g)(%
)(k
Scm
~1)
Roc
kyou
tcro
p6·
94$
0·03
40·9$
1·08
43·3$
1·10
0·44
$0·
060·
22$
0·03
8311
$10
3S
unny
slop
e7·
11$
0·08
36·9
4$1·
1022
·34$
1·90
0·74
$0·
100·
19$
0·02
3961
$18
9S
hady
slop
e7·
08$
0·07
0·36
$0·
018·
76$
0·40
2·83
$0·
160·
82$
0·04
258$
22R
iver
bank
7·66
$0·
210·
45$
0·06
2·97
$0·
103·
88$
0·08
0·67
4$0·
1032
9$25
SAXICOLOUS AND RIPARIAN VEGETATION 309

310 E. MARTINEZ CARRETERO
Results
In general the outcrop crevices and rivers that dissect these communities show a lownumber of species: 15 in the D. lagascae community (average five species per plot), 23 inthe C. praecox community (six per plot), 29 in the E. buniifolium community (seven perplot) and 23 in the A. mendozana community (seven per plot). The species richnessdecreases from riparian to sunny-facing slopes and with shallow soil.
Soils accumulated in cracks of rock outcrops formed by in situ weathering, and onsunny slopes show high values of electric conductivity: 8311 kSiemens cm~1 and3961 kSiemens cm~1, respectively. Soils of shady slopes and riparian soils have higherorganic matter contents (Table 2). All of them differ statistically (p(0·05). Thesoils of riverbanks have more plant cover and accumulate litter from upper communities.
Classification and ordination
The releveH table (Table 1) was derived from the dendrogram constructed by clusteranalysis. The cluster analysis (Fig. 3) distinguished two major groups at a 0·2 similaritylevel. One of them comprises north-facing slopes and rock outcrops, the other includesshady slopes (south-facing) and riparian vegetation. At higher similarity levels, fourclusters corresponding to four communities can be clearly recognized.
A. Dolichlasium lagascae community, established in crevices of sunny rock outcrops(Cluster 1)
This community shows the smallest number of species and the lowest plant cover((10%). Dolichlasium lagascae, a species endemic to these rocks on sunny slopes, isdominant, and found associated with Stipa scirpea, Astericium glaucum and Chuquiragaerinacea Don., amongst others. Stipa psittacorum Speg., strongly associated with shadyslopes (Astericietum glauci, Roig, 1976), appears in cracks of up to 5 cm width and in soilaccumulations on sunny slopes.
B. Cercidium praecox subsp. glaucum community, on sunny slopes (Cluster 2)
This community occurs on strongly water-eroded, north-facing slopes, in crevices widerthan 8 cm. Cercidium praecox is the dominant species, its roots stretching out several
Figure 3. Clustering of releveH s. Communities are indicated in Table 1.

SAXICOLOUS AND RIPARIAN VEGETATION 311
meters along rock crevices, thus exhibiting a great capacity for holding and settlingaccumulated loose soil. Mentzelia parvifolia Urban. et Gilg. ex Kurtz, Porophylumlanceolatum (Spreng.) DC. and Hyalis argentea v. argentea Don. ex Hook. et Arn. are alsopresent, particularly on slopes with little weathered material. This community attainsa mean cover of 20}25%. Some species common at low altitude, in the warmer belt ofLarrea cuneifolia Cav. (Roig, 1989), such as Zuccagnia punctata Cav. and Opuntiasulphurea (S.D.) Bock., are able to grow at this altitude, mainly on north-facing slopesand on sites where the soil is deeper.
C. Eupatorium buniifolium community, on dry riverbanks (Cluster 3)
This community belongs to that described by Roig (1989) as Senecio-Proustietea (ripar-ian vegetation of dry rivers). Its component species develop best in narrow, shallowriverbeds, where sporadic surface runoff provides the banks with moisture. Suchwater availability is evidenced by an increased plant cover ('80% on average) andspecies richness.
Eupatorium buniifolium is the dominant species, associated with E. patens Don. exHook. et Arn., Baccharis salicifolia (R. Ap.) Pearson, Senecio gilliesianus Hieron. andSalvia gilliesii Benth., amongst others. Where the dry rivers dissect highly saline rockoutcrops, Tessaria absinthioides (Hook. et Arn.) DC. and T. dodonaefolia (Hook. et Arn.)DC. are co-dominant. Several species of this community, such as E. buniifolium, Salviagilliesii, Proustia cuneifolia Don. and Schinus fasciculatus (Griseb.) Johnston, also occur inadjacent xeric environments, i.e. on rock ledges, in fissures wider than 8 cm and in dryrivulets on sunny slopes.
Some species belonging to higher, cooler and moister vegetation belts (shrubbycommunity of Diostea scoparia Gill. et Hook-Colliguaja integerrima Gill. ex Hook. etArn.), such as D. scoparia and Senecio filaginoides DC., reach these lower belts alongriverbanks or down shady slopes. A subgroup (releveH 20 and 7) can be clearly distin-guished within this group because of the presence of species such as Larrea divaricataCav., Tricomaria usillo Gill. ex Hook. et Arn. and Stipa eriostachya H.B.K., which arecharacteristic of the zonal community of L. divaricata (Larreetum divaricatae-cuneifoliae,Roig, 1989) where the outcrops occur.
D. Artemisia mendozana community, on shady slopes (Cluster 4)
Artemisia mendozana predominates, along with Adesmia trijuga which is readily recog-nized as the co-dominant species. This community belongs to that described by Roig(1976) as Adesmietum trijugae. These slopes show deeper soils, which allow greaterspecies richness, higher plant cover (45}50% on average) and increased water-holdingcapacity leading to a decrease in surface runoff and soil loss. Adesmia trijuga attainshigher cover on rock outcrops of shady slopes, occurring mostly in crevices of theunderlying consolidated material. Stipa psittacorum is a characteristic species of thiscommunity, accompanied by Stevia gilliesii Hook. et Arn., Monnina dyctiocarpa Griseb.and Erioneuron pilosum (Buckley) Nash.
In the non-metric analysis, 55% of the variance is explained by four axes. Axis Irepresents a moisture gradient along which the composition of vegetation varies.Axes II and III show a change in topographic position from riparian to outcrops (Fig. 4).
From the co-ordinate matrix for each variable (species), and assuming that co-ordinate values represent the relative importance of each species on each variation axis(only considering the three first axes), those species showing closer correspondence witheach habitat can be determined. Sunny-slope species, such as Cercidium praecox, Ment-zelia parvifolia, Menodora decemfida (Gill. ex Hook et Arn.) Gray and Opuntia sulphurea,appear with high values on axis I. Helenium donianum (Hook. et Arn.) Cabrera, Plantago

Figure 4. Ordination of variables (species) on the three first variation axes. Species are num-bered according to Table 1. (m) riparian species; (n) shady slope species; (d) sunny rocksspecies; (s) sunny slope species.
312 E. MARTINEZ CARRETERO
patagonica Jacq., Junellia aspera (Gill. et Hook.) Mold., Relbunium richardianum (Gill. exHook. et Arn.) Hicken, Stevia gilliesii and Monnina dyctiocarpa have high values on axisII, and Eupatorium buniifolium, Oxybaphus ovatus Vahl, Tessaria absinthiodes and Clema-tis denticulata Velloso have high values on axis III.
The discriminant function analysis performed on the survey, taking edaphic factorsinto consideration, shows that the first discriminant function corresponds to rockoutcrops discriminated by Na content. The second function is for sunny slopes dis-criminated by electric conductivity and the third function is for riparian shady slopesdiscriminated by the (Ca#Mg) content and moisture.

SAXICOLOUS AND RIPARIAN VEGETATION 313
The correlation between releveH s, considering their components scores, and soilparameters show r"0·54 (p'0·05) for the (Ca#Mg) content and shady slopes,r"0·53 (p'0·05) for the Na content and rock outcrops and r"0·55 (p'0·05) andr"0·37 (p'0·05) for organic matter and moisture, respectively, with riparian releveH s.
Xerosere in crevices of rock outcrops and consolidated sunny slopes
The floristic relationships among ecologically different communities, shown inTable 1 and in Fig. 3, are revealed by the occurrence of riparian species such asE. buniifolium, E. patens, O. ovatus, S. gilliesianus and Salvia gilliesii in crevices of rockoutcrops and adjacent slopes. Such crevices also behave as small dry rivers, there-by achieving higher moisture levels than the rock outcrops. Riparian species find inthe rock and slope crevices the necessary moisture to become established, and behavetherefore as pioneer plants. The habitat is, none the less, ecologically unfavourable, soboth tall and small trees (nanophanerophytes), such as Proustia cuneifolia and Schinusfasciculatus, of riverbanks can only develop as shrubs (chamaephytes) in crevices(Table 1).
The strong water erosion that affects these crevices, especially on sharp slopes,gradually increases their size until they become deep enough to sustain riparian vegeta-tion with a high floristic richness. Only one species behaves as a colonizer in each habitat:D. lagascae in rock outcrop cracks and C. praecox in sunny slope fissures, owing to thelow soil moisture and depth. When crevices reach further development, (soils becomedeeper and moisture availability increases) other riparian species occur (i.e. releveH s 12and 13). Both D. lagascae and C. praecox communities are not persistent and are replacedthrough physical and biological weathering by different riparian and shady slopesspecies assemblages. Therefore, both original communities will be replaced in the futureby the riparian community (Fig. 5).
In this case, a set of species characterisitc of a syntaxonomical unit (Senecio}Proustietea, riparian vegetation) occurs in another unit (Astericio}Cercidietea, a vegeta-tion of rock and sunny slope crevices), revealing local changes in the rock environment(Fig. 6). These species form either an ecological group (Ellemberg, 1954; Le HoueH rou,1959, 1969) that, though riparian, occurs in crevices of rocks and sunny slopes owing totheir higher moisture and organic matter content (i.e. Group A, Fig. 6).
Discussion
Soil depth and soil moisture gradients on outcrops elsewhere are frequently related and itis considered that they represent successional sequences (Oosting & Anderson, 1937;Sharitz & McCormick, 1973; Collins et al., 1989), as well as combinations of moisture,aspect and slope (Fuls et al., 1992). The relevance of soil depth, moisture gradients andfloristic composition in this work are in agreement with the above mentioned literature.
In Mendoza, the studied vegetation of the piedmont rock outcrops becomes estab-lished only in cracks. On slopes, vegetation occurs not only in fissures but also on theshallow soil accumulated on top of the consolidated material. On sunny slopes, thematerial weathered in situ reaches a few centimeters in depth, and vegetation occursmostly in crevices. Shady slopes, moister and with a thicker layer of accumulated organicmaterial, occasionally show signs of an incipient soil development, and exhibit increasedplant cover and richness of species.
Triassic material usually attain a high level of sodium soluble salts as they weather, anddetritus accumulates in outcrop or slope crevices. Moreover, as lixiviation is low in thesehabitats because of the scarce rainfall and extreme aridity, rock crevices and sunnyslopes reach Na values of 40·9 and 36·9 me l~1, respectively.

Fig
ure
5.P
rop
osed
prim
ary
succ
essi
onon
rock
outc
rop
san
dsu
nny
slop
es.
314 E. MARTINEZ CARRETERO

Figure 6. Ecological soil groups: (A) Riparian group on sunny slopes and crevices of rocks; (B)saxicolous group on riverbanks of wadis. (j)"Nanophanerophyte; (h)"Chamaephyte. Barsize corresponds to the absolute cover value.
SAXICOLOUS AND RIPARIAN VEGETATION 315
Increased moisture and soil depth in riverbanks and on shady slopes determinea greater lixiviation of soluble salts, and hence smaller values of electrical conductivity:329 kSiemens cm~1 and 258 kSiemens cm~1, respectively. On the other hand, the veryshallow substrate present in the crevices of rocks and sunny slopes brings about a greatersurface runoff that prevents litter accumulation which, in addition to high radiation,accounts for the low content of organic matter. Roig & Marone (1982) calculated thatthe total solar radiation received by sunny and shady slopes of the piedmont in Mendozawas of 320 and 185 cal cm~2 day~1, respectively, on 21 July 1982. Intermediate valuesare reported for riverbanks. For this reason, marked differences in soil warming,

316 E. MARTINEZ CARRETERO
and therefore in soil moisture, can be obtained. Le HoueH rou et al. (1993) calculated theglobal solar radiation for different exposures and slopes below 363N, concludingthat sunny slopes show three times as much potential evapo-transpiration as shadyslopes.
At this scale, plant}plant interactions are important on the Mendoza piedmontoutcrops. The floristic relationships found in plant communities occurring on sunnyoutcrops and riverbanks, as well as the changes caused by water erosion to the depth,length and width of crevices, have led to the proposed pattern of primary succession(xerosere) on outcrops surfaces.
The author would like to thank F. Roig (IADIZA), A Diblasi (Mathematics Lab., CRICYT)and H. Le HoueH rou (CNRS, France) for their helpful comments, and E. Soler for reviewing theEnglish version.
References
Baker, W.B. (1956). Some interesting plants on the granite outcrops of Georgia. Georgia MineralNewsletter, 9: 10}19.
Barton, M. & Rodriguez, E. (1989). GeologmHa del piedemonte de la Sierra de Uspallata al W de laciudad de Mendoza. In: Roig, F. (Ed.), Curso Latinoamericano de DeteccioH n y Control de laDesertificacioH n, pp. 154}160. Mendoza: UNEP-IADIZA. 364 pp.
Beals, E.W. (1984). Bray-Curtis ordination: an effective strategy for analysis of multivariateecological data. Advances in Ecological Research, 14: 1}55.
Burbank, M.O. & Platt, R.B. (1964). Granite outcrop communities of the piedmont plateau inGeorgia. Ecology, 45: 292}306.
Cabido, M., Acosta, A. & Diaz, S. (1990). The vascular flora and vegetation of granitic outcropsin the upper CoH rdoba mountains, Argentina. Phytocoenologia, 19: 267}281.
Collins, S.L., Mitchell, G.S. & Klahr, S.C. (1989). Vegetation environment relationships in a rockoutcrop community in Southern Oklahoma. American Midland Naturalist, 122: 339}348.
Duffar, E. (1983). Carta hidrogeomorfoloH gica en la cuenca del Arroyo Maure. Deserta,7: 219}245.
Ellenberg, H. (1954). Landwirtschaftliche Pflanzensociologie. I. Unkrautgemeinschaften als Zieger fuK rKlima und Boden. Stuttgart: Verlag E. Ulmer, 141 pp.
Emberger, L. (1971). Travaux de Botanique et D’Ecologie. BoletmHn de la Sociedad Hongrie: Mason& CIE. 520 pp.
Frangi, J. (1975). Sinopsis de las comunidades vegetales y el medio de las Sierras de Tandil(Provincia de Buenos Aires). Boletin de la Sociedad Argentia de BotaH nica, 16: 293}319.
Fuls, E.R., Bredenkamp, G.J. & van Rooyen, N. (1992). Plant communities of the rocky outcropsof the northern Orange Free State, South Africa. Vegetatio, 103: 79}92.
Funes, G. & Cabido, M. (1995). Variabilidad local y regional de la vegetacioH n rupicola de lasSierras Grandes de CoH rdoba, Argentina. Kurtziana, 24: 173}188.
Le HoueH rou, H.N. (1959). Recherches ecologiques et floristiques sur las vegetation de la TunisieMeridionale. Premiere partie: Les Milieu Naturels, la Vegetation. Universite D’Alger. Institut deRecherches Sahariennes. Memoire No.6.
Le HoueH rou, H.N. (1969). La vegetation de la Tunisie steppique. (avec reference aux vegetationsanalogues d’Algerie, de Lybie et du Maroc). Premiere these. Annales de l’Institut National de laRecherche Agronomique de Tunisie, 42: 625 pp.
Le Houerou, H.N., Popov, G.F. & See, L. (1993). Agro-bioclimatic Classification of Africa.Agrometeorology Series Working Paper, Number 6, Rome, Italy: FAO. 204 pp.
Martinez Carretero, E. (1985). La vegetacioH n de la Reserva Natural Divisadero Largo (Mendoza,Argentina). Documents Phytosociologiques N.S. IX: 25}50. (In Italian.)
Martinez Carretero, E. (1987). FormacioH n geoloH gica y vegetacioH n en la cuenca del DivisaderoLargo, Mendoza. Parodiana, 5: 73}88.
Martinez Carretero, E. & Roig, F. (1994). Asarina antirrhiniflora (Scrophulariacae), enredaderaanfitroH pical de la Reserva Divisadero largo, Mendoza. Multequina, 3: 3}7.
Mosatta, H. & Berra, A. (1996). Los suelos de la cuenca del Arroyo Maure, Provincia deMendoza. Multequina, 5: 57}76.

SAXICOLOUS AND RIPARIAN VEGETATION 317
Oosting, H.J. & Anderson, L.E. (1937). The vegetation of a barefaced cliff in western NorthCarolina. Ecology, 18: 290}292.
Roig, F. (1976). Las Comunidades Vegetales del Piedemonte de la Precordillera de Mendoza.ECOSUR, 3: 1}45.
Roig, F. (1989). Ensayo de deteccioH n y control de la desertificacioH n en el W de la ciudad deMendoza, desde el punto de vista de la vegetacioH n. In: Roig, F. (Ed.), DeteccioH n y Control de laDesertificacioH n. Conferencias, trabajos y resultados del Curso LatinoameH ricano, pp. 196}232.Mendoza: CONICET}IADIZA}UNEP. 364 pp.
Roig, F. & Marone, J. (1982). Influencia de la radiacioH n solar en la distribucioH n de lascomunidades vegetales de la Precordillera de Mendoza. Serie CientnH fica, 27: 9}13.
Sharitz, R.R. & McCormick, J.F. (1973). Population dynamics of two competing annual plantspecies. Ecology, 54: 723}740.
Servicio MeteoroloH gico Nacional (1958). Estadisticas ClimatoloHgicas 1901}1950. Pub. B1 (1),Bs. As.
Shure, D.J. & Ragsdale, H.L. (1977). Patterns of primary succesion on granite outcrop surfaces.Ecology, 58: 993}1006.
Wildi, O. & Orloci, L. (1983). Management and Multivariate Analysis of Vegetation Data. Birmen-sdorf, Switzerland. Swiss Federal Institute of Forestry Research. 139 pp.
Wiser, S.K., Peet, R.K. & White, P.S. (1996). High-elevation rock outersop vegetation of theSouthern Appalachian Mountains. Journal of Vegetation Sciences, 7: 703}722.




















