
Spatially discrete metapopulation models with directional dispersal
Upload
independentCategory
view
1download
0
RESEARCH ARTICLE
Sacrificing patches for linear habitat elements enhancesmetapopulation performance of woodland birdsin fragmented landscapes
Peter Schippers AElig Carla J Grashof-Bokdam AElig Jana Verboom AElig Johannes M Baveco AEligRene Jochem AElig Henk A M Meeuwsen AElig Marjolein H C Van Adrichem
Received 23 July 2008 Accepted 29 November 2008 Published online 19 December 2008
Springer Science+Business Media BV 2008
Abstract It is generally assumed that large patches
of natural habitat are better for the survival of
species than the same amount of habitat in smaller
fragments or linear elements like hedges and tree
rows We use a spatially explicit individual-based
model of a woodland bird to explore this hypothesis
We specifically ask whether mixtures of large small
and linear habitat elements are better for population
performance than landscapes that consist of only
large elements With equal carrying capacity meta-
populations perform equally or better in heterogeneous
landscape types that are a mix of linear large and
small habitat elements We call this increased
metapopulation performance of large and small
elements lsquolsquosynergyrsquorsquo These mixed conditions are
superior because the small linear elements facilitate
dispersal while patches secure the population in
the long run because they have a lower extinction
risk The linear elements are able to catch and
guide dispersing animals which results in higher
connectivity between patches leading to higher
metapopulation survival Our results suggest that
landscape designers should not always seek to
conserve and create larger units but might better
strive for more variable landscapes with mixtures of
patch sizes and shapes This is especially important
when smaller units play a key role in connecting
patches and dispersal through the matrix is poor
Keywords SLOSS Woodland birds Linear elements Hedgerows Patch size Metapopulation Dispersal Synergy Landscape design
Introduction
Habitat patches in fragmented and intensively used
landscapes can be considered as islands in a sea of
non habitat (Bender et al 2003 Wiens 1995) In
these landscapes the amount of habitat size of habitat
patches inter-patch distance and habitat configura-
tion can be regarded as key factors determining
species survival and diversity (Ricketts 2001
Thomas and Kunin 1999 Verboom et al 2001
Tischendorf et al 2003 Schultz and Crone 2005)
Since in many countries space is scarce there is an
increasing need for knowledge of how to optimize the
design of landscapes and nature reserves from a
biodiversity conservation perspective (Lomolino
1994 Kingsland 2002 McCarthy et al 2005 2006)
P Schippers (amp) C J Grashof-Bokdam J Verboom J M Baveco R Jochem H A M Meeuwsen M H C Van Adrichem
Alterra Landscape Centre Wageningen University and
Research Centre PO Box 47 NL-6700AA Wageningen
The Netherlands
e-mail peterschipperswurnl
123
Landscape Ecol (2009) 241123ndash1133
DOI 101007s10980-008-9313-9
Already since 1986 a debate is going on that is
commonly known as the SLOSS debate (Single
Large Or Several Small Patterson and Atmar 1986)
This debate is concentrating on the question whether
many small habitats elements are better than a
single large patch for species conservation pur-
poses given the same total amount of habitat area
(Patterson and Atmar 1986 Baz and GarciaBoyero
1996 Etienne and Heesterbeek 2000 Kingsland
2002 Ovaskainen 2002) It is generally assumed
that a single large patch is superior when a single
species is considered because in a large patch the
overall population is less prone to dispersal mortal-
ity Allee-effects and demographic stochasticity
(MacArthur and Wilson 1967 Verboom et al
2001 Vos et al 2001 Ovaskainen 2002) Compared
to this the lsquoSeveral Smallrsquo configuration is inferior
because of the larger loss of animals during
dispersal the relatively high extinction probability
of small populations due to demographic stochas-
ticity and Allee-effects The situation becomes more
complicated when we compare landscapes that have
Few Large patches with landscapes that have Many
Small (FLOMS) patches (Etienne and Heesterbeek
2000 Ovaskainen 2002) because in the landscape
that consists of a few large patches dispersal
limitation might play a key role
In many intensively used landscapes however
landscapes consist of mixtures of large and small of
habitat patches and of linear habitat Because land is
scarce a design of a sustainable habitat configuration
is essential to improve the conservation potential of
the landscape (Van Apeldoorn et al 1998) Existing
theory favors large patches (eg key patches
Verboom et al 2001) suggesting that all effort
should be put into conservation enlargement
and improvement of large nature areas in a habitat
network We challenge this viewpoint here by
exploring the hypothesis that synergy is possible
between large and small patches Therefore a logical
next step is to evaluate mixtures of large and small
habitat patches and compare their performance with
landscapes that consist of only small or large patches
Here we might expect advantage synergy from
mixed landscape configurations because these land-
scapes might have the best of both (Grashof-Bokdam
et al 2008) In many landscapes linear elements like
hedges and tree rows are important small elements
(Haddad 1999 Chardon et al 2003) Moreover these
elements which are often considered as part of the
matrix may play a key role in the dispersal survival
and reproduction in many species (VanDorp et al
1997 Ricketts 2001 Grashof-Bokdam and van
Langevelde 2005) Knowledge about how these
linear elements function and under what conditions
they facilitate population survival will provide insight
in the use of these elements in landscape planning
and their role in the ecology of species
To investigate the possible added value of mixed
landscapes we explore the population dynamics of a
woodland bird in artificial landscapes in which the
landscape configuration is defined as the percentage
of landscape coverage of large small and linear
elements We specifically ask is there an ecological
advantage (synergy) possible in landscapes which
consist of a mix of small patches linear elements and
large habitat patches compared to landscapes that
consist of only one type Furthermore we want to
understand the mechanisms and prerequisites that
make this lsquosynergyrsquo possible To answer these
questions we adapt the individual-based model
METAPHOR (Verboom et al 2001 Vos et al
2001) and use the model to evaluate the survival
and populations dynamics in different artificial land-
scapes that have the same total amount of habitat
using a woodland bird in open landscapes as an
example
Materials and methods
We use a detailed spatially explicit individual-based
model (METAPHOR) that simulates the dynamics of
a metapopulation (Verboom et al 2001 Vos et al
2001) The model keeps track of every individual in
space and time We use the model to simulate the
population dynamics of a territorial bird (parameter-
ized as the middle spotted woodpecker) living in
patchy landscapes with hedgerows having 25 ha
territories Six kinds of individuals were distin-
guished juveniles territory owners and floaters
(non territorial adults) of both sexes The model
simulates a number of life cycle events every year
reproduction dispersal survival (mortality) aging
and completing pairs (Fig 1) To model the dynamics
in linear elements the habitat area is explicitly
divided into territories that have a sufficient amount
of habitat to support a pair and their offspring
1124 Landscape Ecol (2009) 241123ndash1133
123
Population dynamics of the model
At the start of a year there are only floaters and adults
present in a territory (Fig 1) The adults may
reproduce if paired The offspring the number of
juveniles per female surviving their nesting and
fledging period is drawn from a Poisson distribution
with the parameter k representing the expected
number of surviving offspring and its variance Only
juveniles are allowed to disperse to other territories
The next step in the model is the survival of the
different stages The survival probability in a territory
(i) is made up by a background survival affected by
environmental variability Here the environment is
the same for all territories in any year the environ-
mental variation is perfectly correlated for all
territories and both sexes The survival probability
for a specific territory (Psi) is a Normal distributed
random variable with mean Ps and standard devia-
tion SDs
Psifrac14 Ps thorn et SDs
The standard deviation SDs corresponds to the
natural variation in the survival rate (environmental
stochasticity) It is obtained from a random variable
et with a standard normal distribution We assume
that floaters and juveniles have a lower survival than
the territorial adults (Table 1) Subsequently the
juveniles become adult floaters Floaters may become
territorial adults when a residential adult of the same
sex dies and a territory becomes available (Fig 1)
Artificial landscape configurations
In this paper we focus on Western-European agricul-
tural landscapes where the coverage of woody
elements is typically in the order of magnitude of
10 (Tellerıa et al 2003) Therefore we choose our
artificial landscapes with 10 of woody habitat
Definitions of large and small woodland elements are
based on Dutch nature policy where nature areas
should be larger than 5 ha whereas smaller units were
A A
J
F
S
RD
D
S FC
1 year
Counting
R Reproduction
Dispersal
S SurvivalCJ
Completing pairs
S J
F
A = adults (having a territory)J = juvenilesF = floaters (adults without territory)
AA Aging
Fig 1 Simulation scheme
of stages and life cycle
events of a woodland bird
during a year Life stages Aterritorial adult F floater
adult that does not
reproduce J juvenile
Events R reproduction Ddispersal S survival Aaging C completing pairs
floater replace an adult of
his own sex that has died
Table 1 Parameters of a woodpecker type bird used in the
model
Description Symbol Value Unit
Population dynamics
Survival of territorial adults Psa 078 Year-1
Standard dev aldult survival SDsa 01 Year-1
Survival of juviniles and floaters Psj 065 Year-1
Standard dev Juv and floaters SDsj 02 Year-1
Number of offspring k 3 Year-1
Territorial area requirement T 25 ha
Dispersal
Number of steps N 1000 Year-1
Standard deviation walking angle SDD 5 8
Step size St 10 m
Catchment radius of territory R 89 m
Probability of reflexion
Habitat- [ non habitat Ph 095
Non habitat- [ habitat Pn 0
Landscape Ecol (2009) 241123ndash1133 1125
123
considered to be part of the matrix (Grashof-Bokdam
et al 2008) Therefore we define three landscape
elements small patches of one territory that are
25 ha and linear elements that are 25 ha but have a
width of 25 m and large patches of four territories
that are 10 ha So here we simulate the matrix
structure explicitly To avoid confusion however we
will use the term matrix to describe the non habitat
part of the landscape The total area of the artificial
landscapes is 6 9 6 km
We use a replacement approach to construct
different landscapes We start with a landscape with
only large elements subsequently we replace large
elements by small patches andor linear elements
keeping the same total amount of habitat This results
in seven landscape types all having 10 of habitat
area but differing with respect to the total abundance
of the different landscape elements (Table 2) We
choose the maximal connection version of linear
elements and patches of the landscape as our default
landscape Here linear elements are as much as
possible physically connected to the patches (Fig 2)
In the simulations and results section we will evaluate
what the effect is of this choice Additionally we
define the precise location of the territories To do
this we first overlay the landscape with a large cloud
of random points and check around every point
whether there is enough habitat (25 ha) to support a
territory We continue this procedure until we have
144 territories in all the landscape types As a result
we get landscapes havening all 10 of habitat and
90 non-habitat with scattered territories located in
the 10 habitat Animals are allocated to a territory
when they do not disperse Dispersing animals are
allowed to move freely in all directions in this
landscape in both habitat and non-habitat according
to the rules described in the next section until they are
within a critical radius from a new territory and are
allocated to this territory
Random walk dispersal
We simulate the dispersal between territories using
SmallSteps a separate movement module (Baveco
2006) This module simulates the dispersal of the
juveniles from a territory in which they were born to
a territory in which they settle The animal movement
in the model is in fact a correlated random walk
(Schippers et al 1996 Tischendorf and Wissel 1997
Tischendorf et al 1998 Haddad 1999) In this model
a dispersing animal has a coordinate in a continuous
xndashy space and a walking direction between 0 and
360 Each time step a new direction (Dt) relative to
the old direction (Dt-1) is calculated
Dt frac14 modethDt1 thorn et SDD 2pTHORN
where et is a number drawn from a standard normal
distribution SDD is the standard deviation of the
walking direction lsquolsquomodrsquorsquo is a modulus function with
2p (rad) as the basis number Next the dispersing
animal takes a step in this new direction with a step
size (St) of 10 m subsequently the new position is
calculated A dispersing animal is allowed to take
1000 steps during dispersal If during dispersal in
habitat an animal meets non habitat it has a proba-
bility to be reflected Ph of 95 (Table 1) Reflecting
means that the walking direction is mirrored against
the habitat edge When a dispersing animal however
enter habitat the animal continues its path Animals
continue dispersing until they find a new territory that
is not occupied by a territorial adult of the same sex
If an animal has not found another suitable patch after
Table 2 Habitat cover
( of total) of different
landscapes
Landscape
description
Habitat large
patches
Habitat small
patches
Habitat linear
elements
Matrix
Large 10 0 0 90
Mix-P 5 5 0 90
Mix-L 5 0 5 90
Mixed-LP 5 25 25 90
Small-P 0 10 0 90
Small-L 0 0 10 90
Small-LP 0 5 5 90
1126 Landscape Ecol (2009) 241123ndash1133
123
1000 steps and is not located in habitat the dispersing
animal dies Animals that arrive in habitat after 1000
steps but not in a vacant territory are allocated to the
nearest territory Animals that arrive in a new
territory become floaters and may reproduce when
the territorial adult of the same sex dies Animals that
encounter the landscape edge are reflected These
rules cause that dispersing animals first disperse in
the habitat patch of their territory checking neighbor
territories before they start dispersing through the
matrix and might find another habitat patch and
territories in there
Parameterization and calibration
We use demographic parameters of the middle
spotted woodpecker (Pettersson 1985 Kosenko and
Kaigorodova 2001 Kosinski and Ksit 2006) whereas
the dispersal distance is based on the work of Muller
(1982 Table 1) Demographic parameters were cal-
ibrated to meet criteria published by Verboom et al
(2001) for medium sized birds
Evaluating synergy
Synergy occurs when mixtures of large and small
habitat patches perform better with respect to survival
than landscapes with patches of the same type In this
study we did 250 simulations in the default land-
scapes each run starting with one reproductive unit
ie an adult male and female in every territory We
evaluated metapopulation survival and the mean
number of adult individuals between year 50 and
150 to avoid transient behavior due to the initiali-
zation state (lsquolsquoburn-inrsquorsquo) Since there is a strong
correlation between the mean number of individuals
and survival we present only the survival results in
our graphs Of every survival fraction the 95
confidence interval is calculated using a v-test We
present our results as a replacement series in which
we replace large patches by smaller elements keeping
the same amount of habitat ultimately resulting in the
landscape with small elements only (Fig 3) Monot-
onously increasing relations (small- [ mixed- [large) support the more traditional hypothesis that
Large
Large
Large
Small-P Mix-P
Small-L
Small-PL
Mix-L
Mix-PL
only small mixed only largereplacing
Small toLarge
Small andLinear toLarge
Linear tolarge
Fig 2 Default landscapes
of 6 9 6 km lines are
linear elements of
6000 9 25 m smallsquares are small forest
patches of 25 ha that can
contain one territory Largesquares are large patches of
10 ha that contain four
territories
Landscape Ecol (2009) 241123ndash1133 1127
123
larger patches are always better an optimum rela-
tionship supports our alternative hypothesis that
synergy occurs
Simulations and results
Reference simulations
We evaluate metapopulation survival in the seven
default landscapes for the species parameterized
according to Table 1 When we replace half of the
large patches by small elements these mixed land-
scapes performe best if these small elements were all
linear elements (L Fig 4) Here metapopulation
survival in mixed landscapes was higher than in the
landscape with only large patches indicating syn-
ergy If half of the large patches is replaced by a mix
of small patches and linear elements (LP) metapop-
ulation survival is the same as in the landscapes with
only large patches This indicates that mixed land-
scapes with linear elements perform equally or better
than the landscape with only large patches Mixed
landscapes without linear elements (P) or landscapes
with only small elements however always performe
worse compared to the other landscapes Furthermore
we see that a landscape with only small elements that
contained both linear elements and small patches
(LP) performed much better than landscapes with
only small elements that contained only linear
elements (L) or only small patches (P) These results
suggest that replacing a part of the small patches by
linear elements also enhances population survival
Effects of physical connections
The results suggest that linear elements are respon-
sible for the synergy due to the fact that they connect
patches with each other and guide dispersers To
investigate this effect we create three alternative
landscapes in which the linear elements are not
connected to the patches
If linear elements are not connected to the large
patches number of individuals and survival decrease
in the mixed landscapes with linear elements (L and
LP) (compare Figs 4 5) This indicates that linear
connections are responsible for the synergy found in
the reference conditions Survival in landschapes
Large
Mix-L
Mix-LP
Mix-P
Small-L
Small-LP
Small-P
S
urvi
val o
f met
apop
ulat
ion
Only Mix Onlysmall largeelements patches
L-line
P-line
LP-line
Fig 3 Explanatory graph that relates individual simulation
results to synergy If mixed landscapes perform better than
landscapes with only small and large elements there is synergy
L linear elements present as small elements P small patches
present as small elements LP both linear elements and small
patches present as small elements
0
02
04
06
08
1
only small mix only large
surv
ivin
g m
etap
opul
atio
ns
L
P
LP
Fig 4 Fraction of surviving metapopulations of default
simulations of seven different landscapes that differ with
respect to the percentage of large and small habitat patches or
linear habitat elements error bars represent the 95 confi-
dence inteval Only small landscapes that only consist of small
elements mix landscape that are a mix of large and small
elements only large is a landscape that has only large patches
L linear elements present as small elements P small patches
present as small elements LP both linear elements and small
patches present as small elements
1128 Landscape Ecol (2009) 241123ndash1133
123
with only small elements however is still higher in
landscapes with small patches and linear elements
(LP) than in landscapes with only small patches
Effects of decreasing the dispersal through
the matrix
To investigate whether linear elements are indeed
crucial for synergy we decrease the dispersal effi-
ciency through the matrix by increasing the standard
deviation of the turning angle (SDD) from 5 to 20 and
40 This change results in a decrease of the realized
dispersal distance As a result 90 of the animals
move less than resp 29 and 15 km in the matrix
whereas in the reference situation this was 84 km In
these simulations we use the seven default landscapes
again
As a result of the decreased dispersal efficiency
through the matrix the metapopulation survival in
landscapes without linear elements (P) decrease
strongly (Fig 6a b) resulting in a clear synergy
pattern for the linear replacement lines (L) and
linear combined with small patches replacement
lines (LP Fig 6) Replacing large patches by
smaller patches does not help to compensate for
the loss of dispersal efficiency through the matrix
The landscapes containing only small patches the
decreased dispersal efficiency even caused extinc-
tion in a simualtion for the most inefficient dispersal
class These results indicate that if dispersal through
the matrix is difficult or avoided by a species linear
elements are crucial
Discussion
Main results
It is generally assumed that larger patches of natural
habitat are better for the survival of species than the
same amount of habitat in smaller fragments
(MacArthur and Wilson 1967 Verboom et al 2001
Vos et al 2001) Our results suggest that replacing a
part of the larger units by smaller linear elements
enhance population performance with respect to
population level and survival Without any large
patches however we see a serious decrease in
performance of the population indicating that large
patches are still very important The explanation for
this synergy is that the large patches secure survival
of the population whereas the linear units enhance
inter-patch dispersal by serving as corridors Addi-
tionally we see that populations in landscapes with
small patches also benefit from the availability of
linear elements Also here we see synergy to the
extent that a mixture of linear elements and small
patches performs better than landscapes consisting of
solely linear elements or small patches
In this study linear elements have a double
function they can serve as a corridor but also as
habitat In real landscapes corridors may not be wide
or qualitatively good enough to support both func-
tions Small patches may serve as stepping stones
because the mean inter-patch distance decreases with
increasing number of patches (Etienne and Heester-
beek 2000) However if we reduce the area of the
patches with a certain factor the average inter-patch
distance (centre to centre) is reduced with only the
square root of this factor Consequently replacing
large patches by smaller patches is inferior to
replacing large patches by linear elements
0
02
04
06
08
1
only small mix only large
surv
ing
met
apop
ulat
ions
L
LP
Fig 5 Fraction of surviving metapopulations of simulations
in which the linear elements were not connected to the large
patches of five different landscapes that differ with respect to
the percentage of large and small habitat patches or linear
habitat elements error bars represent the 95 confidence
inteval Only small landscapes that only consist of small
elements mix landscape that are a mix of large and small
elements only large is a landscape that has only large patches
L linear elements present as small elements LP both linear
elements and small patches present as small elements
Landscape Ecol (2009) 241123ndash1133 1129
123
Mechanisms favoring large or small patches
Seven processes are put forward in literature that
determines metapopulation performance in response
to the subdivision of habitat
1 Population extinction larger patches have a
higher carrying capacity and therefore a lower
extinction risk due to Allee-effects and demo-
graphic stochasticity than smaller patches
(Shaffer 1981 Soule 1987)
2 Colonization When dispersal potential is weak
and the dispersal mortality in the matrix is high
however the lsquoSeveral Smallrsquo approach might be
superior (Etienne and Heesterbeek 2000) The
extreme case is when the inter-patch distances of
the large patches are too large for recolonization
whereas the species is able to bridge the smaller
inter-patch distance between the small patches
Here the landscape configuration with few large
patches is actually more fragmented than the
landscape with many small patches (Flather and
Bevers 2002)
3 Finding a mate it seems likely that especially
monogamous animals that live in a landscape
with many small patches have a higher risk to
stay single This lowers the reproduction (Engen
et al 2003)
4 Density dependent population growth in a more
fragmented landscape animals encounter more
density dependent effects because some patches
are at or above carrying capacity while others are
empty Here individuals are not optimally utiliz-
ing the resources This lowers the growth rate of
fragmented populations
5 Patch edge effects if a species utilizes patch
edges more than the patch core several small
patches might be superior over a few large
patches because the amount of edges and
ecotones is larger when the landscape is more
fragmented On the other hand if core habitat is
preferred eg because the influence of the
Fig 6 The effect of increasing the turning angle of dispersing
birds from 5 to 20 and 40 degrees resulting in lower matrix
permeability The figure depict the fraction of surviving
metapopulations in the default landscape and the error bars
represent the 95 confidence interval Only small landscapes
that only consist of small elements mix landscape that are a
mix of large and small elements only large is a landscape that
has only large patches L linear elements present as small
elements P small patches present as small elements LP both
linear elements and small patches present as small elements
c
40 degrees
0
02
04
06
08
1
surv
ivin
g m
etap
opul
atio
ns
L
P
LP
20 degrees
0
02
04
06
08
1
only small mix only large
only small mix only large
only small mix only large
surv
ivin
g m
etap
opul
atio
ns
L
P
LP
5 degrees
0
02
04
06
08
1
surv
ivin
g m
etap
opul
atio
ns
L
P
LP
1130 Landscape Ecol (2009) 241123ndash1133
123
matrix makes the edges suboptimal larger ele-
ments are better
6 Spreading of risk if populations fluctuate due to
local environmental factors subdivision may
save parts of the metapopulation from events
such as fire or other adverse conditions When
species like a predator or a disease are taken into
account a fragmented landscape may be superior
Here predator or disease might also suffers from
isolation leaving their prey or host untouched in
patches that are not yet colonized by the predator
or disease (Baz and GarciaBoyero 1996 Lang-
lois et al 2001 Tscharntke et al 2002) Another
special case is habitat heterogeneity among
patches which makes eg wet patches more
suitable in dry years and dry patches in wet years
(Boer 1981) Although the spreading of risk
might also occur in larger elements it generally
favors population subdivision
7 Round off effects in fragmented landscapes
specific patch carrying capacity might not be
whole even numbers For instance if we sub-
stitute a patch with a carrying capacity of ten by
four fragments with carrying capacities of 25
these patches together can only support eight
breeding pairs
In our simulation study we do not incorporate all
these seven processes We take extinction (due to
demographic stochasticity and Allee-effects) coloni-
zation (dispersal success) pair formation and density
dependent population growth into account We do not
incorporate patch edge effects (when habitat quality
differs between patch core and patch edge) spreading
of risk (local disturbances patch heterogeneity and
antagonistic species) and round off effects These
three missing factors have an evident effect while
round off effects will favor large patch landscapes
and spreading of risk will favor fragmented land-
scapes the effect of taking into account edge effects
will depend on a preference for patch core or edge
habitat but in either case a straightforward relation is
expected Therefore our results may be slightly
modulated by taking into account these aspects but
our conclusions should be robust since some factors
are in favor of small patches and others are in favor of
large patches an optimum configuration is expected
somewhere between the two extreme cases with only
large or only small patches
Climate change
In the face of climate change there might be a few
extra considerations the habitat of species in climatic
terms is no longer stationary so species should be
able to track their climatic window Consequently the
global survival of a species is no longer only
dependent on the survival locally but also on the
ability of species to move and follow the climate
suitability window (Tischendorf et al 2003 Travis
2003 Saether et al 2004 Thomas et al 2004) This
underpins the idea that under climate change condi-
tions heterogeneous landscapes with many linear
elements might be even more superior for species
survival because they both facilitate animal movement
en survival Whereas large patches are necessary to
buffer the population against increasingly stochastic
weather events and provide dispersers connectivity by
linear elements should allow populations to shift
Generality of the results
We did a series of simulations with a spatially
explicit individual-based population model simulat-
ing the population dynamics of a territorial woodland
birds in open agricultural landscapes Our results
indicate that replacing patches by linear elements that
can serve as habitat enhances metapopulation perfor-
mance Clearly we can not extrapolate these results to
conclusions for other species and spatial configura-
tions We think however that our conclusions are
likely to be valid for other species and conditions if
(1) the carrying capacity of the habitat is not affected
by the replacement (2) the dispersal trough habitat is
more efficient and less risky than dispersal through
matrix and (3) if spatial configurations are such that
patches are relatively isolated so that dispersal
limitation is an important factor
Implications for sustainable landscape design
and nature conservation
Since in many countries space is scarce there is an
increasing need for knowledge about how to optimize
the design of landscapes nature reserves and habitat
networks from a biodiversity conservation perspec-
tive (Lomolino 1994 Kingsland 2002 McCarthy
Landscape Ecol (2009) 241123ndash1133 1131
123
et al 2005 McCarthy et al 2006) Our results
indicate that landscape designers and nature conser-
vationists should reconsider the value of small linear
elements that often are considered as part of the
matrix and not always seek to create larger units but
may better create more variable landscapes with
mixtures of patch sizes and shapes This is especially
important when smaller units play a key role in
physically connecting large patches when dispersal
through the matrix is poor or dispersal distances are
smaller than inter-patch distances On the other hand
the smaller units should be large enough to have a
habitat function for the species considered ie
reproduction should be possible there
Acknowledgments This research was financed by the
program lsquolsquoVernieuwend Ruimtegebruikrsquorsquo of Habiforum and
co-financed by the Dutch Ministry of Agriculture Nature and
Food Quality
References
Baveco JM (2006) Small steps movement model manual
Version 10 Alterra-Wageningen UR Wageningen p 19
Baz A GarciaBoyero A (1996) The SLOSS dilemma a
butterfly case study Biodivers Conserv 5493ndash502 doi
101007BF00056393
Bender DJ Tischendorf L Fahrig L (2003) Using patch isolation
metrics to predict animal movement in binary landscapes
Landscape Ecol 1817ndash39 doi101023A10229372
26820
PJd Boer (1981) On the survival of populations in a hetero-
geneous and variable environment Oecologia 5039ndash53
doi101007BF00378792
Chardon JP Adriaensen F Matthysen E (2003) Incorporating
landscape elements into a connectivity measure a case
study for the Speckled wood butterfly (Pararge aegeria L)
Landscape Ecol 18561ndash573 doi101023A10260625
30600
Engen S Lande R Seather BE (2003) Demographic stochas-
ticity and allee effects in populations with two sexes
Ecology 842378ndash2386 doi10189002-0123
Etienne RS Heesterbeek JAP (2000) On optimal size and
number of reserves for metapopulation persistence
J Theor Biol 20333ndash50 doi101006jtbi19991060
Flather CH Bevers M (2002) Patchy reaction-diffusion
and population abundance the relative importance of
habitat amount and arrangement Am Nat 15940ndash56 doi
101086324120
Grashof-Bokdam CJ Chardon P Vos CC Foppen RPB
Wallisdevries MF VanDerVeen M Meeuwsen HAM
(2008) The synergistic effect of combining woodlands
and green veining for biodiversity Landscape Ecol doi
101007s10980-008-9274-2
Grashof-Bokdam CJ van Langevelde F (2005) Green veining
landscape determinants of biodiversity in European
agricultural landscapes Landscape Ecol 20417ndash439
doi101007s10980-004-5646-1
Haddad NM (1999) Corridor and distance effects on interpatch
movements a landscape experiment with butterflies Ecol
Appl 9612ndash622 doi1018901051-0761(1999)009[0612
CADEOI]20CO2
Kingsland SE (2002) Creating a science of nature reserve
design perspectives from history Environ Model Assess
761ndash69 doi101023A1015633830223
Kosenko SM Kaigorodova EY (2001) Effect of habitat frag-
mentation on distribution density and breeding
performance of the middle spotted woodpecker Dend-
rocopos medius (Alves Picidae) in Nerussa-Desna
Polesye Zool Zh 8071ndash78
Kosinski Z Ksit P (2006) Comparative reproductive biology of
middle spotted woodpeckers Dendrocopos medius and
great spotted woodpeckers D-major in a riverine forest
Bird Study 53237ndash246
Langlois JP Fahrig L Merriam G Artsob H (2001) Landscape
structure influences continental distribution of hantavirus
in deer mice Landscape Ecol 16255ndash266 doi101023
A1011148316537
Lomolino MV (1994) An evaluation of alternative strategies
for building networks of nature-reserves Biol Conserv
69243ndash249 doi1010160006-3207(94)90423-5
MacArthur RH Wilson EO (1967) The theory of island bio-
geography Princeton University Press Princeton
McCarthy MA Thompson CJ Possingham HP (2005) Theory
for designing nature reserves for single species Am Nat
165250ndash257 doi101086427297
McCarthy MA Thompson CJ Williams NSG (2006) Logic for
designing nature reserves for multiple species Am Nat
167717ndash727 doi101086503058
Muller W (1982) Die Besiedlung der Eichenwalder im Kanton
Zurich durch den Mittelspecht Dendrocopus medius Orn
Beob 79105ndash119
Ovaskainen O (2002) Long-term persistence of species and the
SLOSS problem J Theor Biol 218419ndash433
Patterson BD Atmar W (1986) Nested subsets and the structure
of insular mammalian faunas and archipelagoes Biol J
Linn Soc Lond 2865ndash82 doi101111j1095-83121986
tb01749x
Pettersson B (1985) Extinction of an isolated population of the
middle spotted woodpecker Dendrocopos-Medius (L) in
Sweden and its relation to general theories on extinction
Biol Conserv 32335ndash353 doi1010160006-3207(85)
90022-9
Ricketts TH (2001) The matrix matters Effective isolation in
fragmented landscapes Am Nat 15887ndash99 doi101086
320863
Saether BE Sutherland WJ Engen S (2004) Climate influences
on avian population dynamics In Moller AP Fielder W
Berthold P (eds) Birds and climate change pp 185ndash209
Schippers P Verboom J Knaapen JP vanApeldoorn RC
(1996) Dispersal and habitat connectivity in complex
heterogeneous landscapes an analysis with a GIS-based
random walk model Ecography 1997ndash106 doi101111
j1600-05871996tb00160x
Schultz CB Crone EE (2005) Patch size and connectivity
thresholds for butterfly habitat restoration Conserv Biol
19887ndash896 doi101111j1523-1739200500462x
1132 Landscape Ecol (2009) 241123ndash1133
123
Shaffer ML (1981) Minimum population sizes for species con-
servation Bioscience 31131ndash134 doi1023071308256
Soule M (1987) Viable populations for conservation
Cambridge University Press Cambridge
Tellerıa JL Baquero R Santos T (2003) Effects of forest
fragmentation on European birds implications of regional
differences in species richness J Biogeogr 30621ndash628
doi101046j1365-2699200300960x
Thomas CD Cameron A Green RE Bakkenes M Beaumont LJ
Collingham YC Erasmus BFN de Siqueira MF Grainger
A Hannah L Hughes L Huntley B van Jaarsveld AS
Midgley GF Miles L Ortega-Huerta MA Peterson AT
Phillips OL Williams SE (2004) Extinction risk from
climate change Nature 427145ndash148 doi101038nature
02121
Thomas CD Kunin WE (1999) The spatial structure of pop-
ulations J Anim Ecol 68647ndash657 doi101046j1365-
2656199900330x
Tischendorf L Bender DJ Fahrig L (2003) Evaluation of patch
isolation metrics in mosaic landscapes for specialist vs
generalist dispersers Landscape Ecol 1841ndash50 doi
101023A1022908109982
Tischendorf L Irmler U Hingst R (1998) A simulation
experiment on the potential of hedgerows as movement
corridors for forest carabids Ecol Modell 106107ndash118
doi101016S0304-3800(97)00186-5
Tischendorf L Wissel C (1997) Corridors as conduits for small
animals attainable distances depending on movement
pattern boundary reaction and corridor width Oikos
79603ndash611 doi1023073546904
Travis JMJ (2003) Climate change and habitat destruction a
deadly anthropogenic cocktail Proc R Soc Lond B Biol
Sci 270467ndash473 doi101098rspb20022246
Tscharntke T Steffan-Dewenter I Kruess A Thies C (2002)
Contribution of small habitat fragments to conservation of
insect communities of grassland-cropland landscapes
Ecol Appl 12354ndash363
Van Apeldoorn RC Knaapen JP Schippers P Verboom J
Engen van H Meeuwsen H (1998) Landscape planning
and conservation biology simulation models as tools to
evaluate scenarios for badger in the Netherlands Land-
scape Urban Plan 4157ndash69 doi101016S0169-2046(97)
00058-3
VanDorp D Schippers P Groenendael van JM (1997)
Migration rates of grassland plants along corridors in
fragmented landscapes assessed with a cellular automa-
tion model Landscape Ecol 1239ndash50 doi101007
BF02698206
Verboom J Foppen R Chardon P Opdam P Luttikhuizen P
(2001) Introducing the key patch approach for habitat
networks with persistent populations an example for
marshland birds Biol Conserv 10089ndash101 doi101016
S0006-3207(00)00210-X
Vos CC Verboom J Opdam PFM Ter Braak CJF (2001)
Toward ecologically scaled landscape indices Am Nat
15724ndash41 doi101086317004
Wiens JA (1995) Landscape mosaics and ecological theory In
Hansson L Fahrig L Merriam G (eds) Mosaic landscapes
and ecological processes Chapman and Hall London
pp 1ndash26
Landscape Ecol (2009) 241123ndash1133 1133
123

Already since 1986 a debate is going on that is
commonly known as the SLOSS debate (Single
Large Or Several Small Patterson and Atmar 1986)
This debate is concentrating on the question whether
many small habitats elements are better than a
single large patch for species conservation pur-
poses given the same total amount of habitat area
(Patterson and Atmar 1986 Baz and GarciaBoyero
1996 Etienne and Heesterbeek 2000 Kingsland
2002 Ovaskainen 2002) It is generally assumed
that a single large patch is superior when a single
species is considered because in a large patch the
overall population is less prone to dispersal mortal-
ity Allee-effects and demographic stochasticity
(MacArthur and Wilson 1967 Verboom et al
2001 Vos et al 2001 Ovaskainen 2002) Compared
to this the lsquoSeveral Smallrsquo configuration is inferior
because of the larger loss of animals during
dispersal the relatively high extinction probability
of small populations due to demographic stochas-
ticity and Allee-effects The situation becomes more
complicated when we compare landscapes that have
Few Large patches with landscapes that have Many
Small (FLOMS) patches (Etienne and Heesterbeek
2000 Ovaskainen 2002) because in the landscape
that consists of a few large patches dispersal
limitation might play a key role
In many intensively used landscapes however
landscapes consist of mixtures of large and small of
habitat patches and of linear habitat Because land is
scarce a design of a sustainable habitat configuration
is essential to improve the conservation potential of
the landscape (Van Apeldoorn et al 1998) Existing
theory favors large patches (eg key patches
Verboom et al 2001) suggesting that all effort
should be put into conservation enlargement
and improvement of large nature areas in a habitat
network We challenge this viewpoint here by
exploring the hypothesis that synergy is possible
between large and small patches Therefore a logical
next step is to evaluate mixtures of large and small
habitat patches and compare their performance with
landscapes that consist of only small or large patches
Here we might expect advantage synergy from
mixed landscape configurations because these land-
scapes might have the best of both (Grashof-Bokdam
et al 2008) In many landscapes linear elements like
hedges and tree rows are important small elements
(Haddad 1999 Chardon et al 2003) Moreover these
elements which are often considered as part of the
matrix may play a key role in the dispersal survival
and reproduction in many species (VanDorp et al
1997 Ricketts 2001 Grashof-Bokdam and van
Langevelde 2005) Knowledge about how these
linear elements function and under what conditions
they facilitate population survival will provide insight
in the use of these elements in landscape planning
and their role in the ecology of species
To investigate the possible added value of mixed
landscapes we explore the population dynamics of a
woodland bird in artificial landscapes in which the
landscape configuration is defined as the percentage
of landscape coverage of large small and linear
elements We specifically ask is there an ecological
advantage (synergy) possible in landscapes which
consist of a mix of small patches linear elements and
large habitat patches compared to landscapes that
consist of only one type Furthermore we want to
understand the mechanisms and prerequisites that
make this lsquosynergyrsquo possible To answer these
questions we adapt the individual-based model
METAPHOR (Verboom et al 2001 Vos et al
2001) and use the model to evaluate the survival
and populations dynamics in different artificial land-
scapes that have the same total amount of habitat
using a woodland bird in open landscapes as an
example
Materials and methods
We use a detailed spatially explicit individual-based
model (METAPHOR) that simulates the dynamics of
a metapopulation (Verboom et al 2001 Vos et al
2001) The model keeps track of every individual in
space and time We use the model to simulate the
population dynamics of a territorial bird (parameter-
ized as the middle spotted woodpecker) living in
patchy landscapes with hedgerows having 25 ha
territories Six kinds of individuals were distin-
guished juveniles territory owners and floaters
(non territorial adults) of both sexes The model
simulates a number of life cycle events every year
reproduction dispersal survival (mortality) aging
and completing pairs (Fig 1) To model the dynamics
in linear elements the habitat area is explicitly
divided into territories that have a sufficient amount
of habitat to support a pair and their offspring
1124 Landscape Ecol (2009) 241123ndash1133
123
Population dynamics of the model
At the start of a year there are only floaters and adults
present in a territory (Fig 1) The adults may
reproduce if paired The offspring the number of
juveniles per female surviving their nesting and
fledging period is drawn from a Poisson distribution
with the parameter k representing the expected
number of surviving offspring and its variance Only
juveniles are allowed to disperse to other territories
The next step in the model is the survival of the
different stages The survival probability in a territory
(i) is made up by a background survival affected by
environmental variability Here the environment is
the same for all territories in any year the environ-
mental variation is perfectly correlated for all
territories and both sexes The survival probability
for a specific territory (Psi) is a Normal distributed
random variable with mean Ps and standard devia-
tion SDs
Psifrac14 Ps thorn et SDs
The standard deviation SDs corresponds to the
natural variation in the survival rate (environmental
stochasticity) It is obtained from a random variable
et with a standard normal distribution We assume
that floaters and juveniles have a lower survival than
the territorial adults (Table 1) Subsequently the
juveniles become adult floaters Floaters may become
territorial adults when a residential adult of the same
sex dies and a territory becomes available (Fig 1)
Artificial landscape configurations
In this paper we focus on Western-European agricul-
tural landscapes where the coverage of woody
elements is typically in the order of magnitude of
10 (Tellerıa et al 2003) Therefore we choose our
artificial landscapes with 10 of woody habitat
Definitions of large and small woodland elements are
based on Dutch nature policy where nature areas
should be larger than 5 ha whereas smaller units were
A A
J
F
S
RD
D
S FC
1 year
Counting
R Reproduction
Dispersal
S SurvivalCJ
Completing pairs
S J
F
A = adults (having a territory)J = juvenilesF = floaters (adults without territory)
AA Aging
Fig 1 Simulation scheme
of stages and life cycle
events of a woodland bird
during a year Life stages Aterritorial adult F floater
adult that does not
reproduce J juvenile
Events R reproduction Ddispersal S survival Aaging C completing pairs
floater replace an adult of
his own sex that has died
Table 1 Parameters of a woodpecker type bird used in the
model
Description Symbol Value Unit
Population dynamics
Survival of territorial adults Psa 078 Year-1
Standard dev aldult survival SDsa 01 Year-1
Survival of juviniles and floaters Psj 065 Year-1
Standard dev Juv and floaters SDsj 02 Year-1
Number of offspring k 3 Year-1
Territorial area requirement T 25 ha
Dispersal
Number of steps N 1000 Year-1
Standard deviation walking angle SDD 5 8
Step size St 10 m
Catchment radius of territory R 89 m
Probability of reflexion
Habitat- [ non habitat Ph 095
Non habitat- [ habitat Pn 0
Landscape Ecol (2009) 241123ndash1133 1125
123
considered to be part of the matrix (Grashof-Bokdam
et al 2008) Therefore we define three landscape
elements small patches of one territory that are
25 ha and linear elements that are 25 ha but have a
width of 25 m and large patches of four territories
that are 10 ha So here we simulate the matrix
structure explicitly To avoid confusion however we
will use the term matrix to describe the non habitat
part of the landscape The total area of the artificial
landscapes is 6 9 6 km
We use a replacement approach to construct
different landscapes We start with a landscape with
only large elements subsequently we replace large
elements by small patches andor linear elements
keeping the same total amount of habitat This results
in seven landscape types all having 10 of habitat
area but differing with respect to the total abundance
of the different landscape elements (Table 2) We
choose the maximal connection version of linear
elements and patches of the landscape as our default
landscape Here linear elements are as much as
possible physically connected to the patches (Fig 2)
In the simulations and results section we will evaluate
what the effect is of this choice Additionally we
define the precise location of the territories To do
this we first overlay the landscape with a large cloud
of random points and check around every point
whether there is enough habitat (25 ha) to support a
territory We continue this procedure until we have
144 territories in all the landscape types As a result
we get landscapes havening all 10 of habitat and
90 non-habitat with scattered territories located in
the 10 habitat Animals are allocated to a territory
when they do not disperse Dispersing animals are
allowed to move freely in all directions in this
landscape in both habitat and non-habitat according
to the rules described in the next section until they are
within a critical radius from a new territory and are
allocated to this territory
Random walk dispersal
We simulate the dispersal between territories using
SmallSteps a separate movement module (Baveco
2006) This module simulates the dispersal of the
juveniles from a territory in which they were born to
a territory in which they settle The animal movement
in the model is in fact a correlated random walk
(Schippers et al 1996 Tischendorf and Wissel 1997
Tischendorf et al 1998 Haddad 1999) In this model
a dispersing animal has a coordinate in a continuous
xndashy space and a walking direction between 0 and
360 Each time step a new direction (Dt) relative to
the old direction (Dt-1) is calculated
Dt frac14 modethDt1 thorn et SDD 2pTHORN
where et is a number drawn from a standard normal
distribution SDD is the standard deviation of the
walking direction lsquolsquomodrsquorsquo is a modulus function with
2p (rad) as the basis number Next the dispersing
animal takes a step in this new direction with a step
size (St) of 10 m subsequently the new position is
calculated A dispersing animal is allowed to take
1000 steps during dispersal If during dispersal in
habitat an animal meets non habitat it has a proba-
bility to be reflected Ph of 95 (Table 1) Reflecting
means that the walking direction is mirrored against
the habitat edge When a dispersing animal however
enter habitat the animal continues its path Animals
continue dispersing until they find a new territory that
is not occupied by a territorial adult of the same sex
If an animal has not found another suitable patch after
Table 2 Habitat cover
( of total) of different
landscapes
Landscape
description
Habitat large
patches
Habitat small
patches
Habitat linear
elements
Matrix
Large 10 0 0 90
Mix-P 5 5 0 90
Mix-L 5 0 5 90
Mixed-LP 5 25 25 90
Small-P 0 10 0 90
Small-L 0 0 10 90
Small-LP 0 5 5 90
1126 Landscape Ecol (2009) 241123ndash1133
123
1000 steps and is not located in habitat the dispersing
animal dies Animals that arrive in habitat after 1000
steps but not in a vacant territory are allocated to the
nearest territory Animals that arrive in a new
territory become floaters and may reproduce when
the territorial adult of the same sex dies Animals that
encounter the landscape edge are reflected These
rules cause that dispersing animals first disperse in
the habitat patch of their territory checking neighbor
territories before they start dispersing through the
matrix and might find another habitat patch and
territories in there
Parameterization and calibration
We use demographic parameters of the middle
spotted woodpecker (Pettersson 1985 Kosenko and
Kaigorodova 2001 Kosinski and Ksit 2006) whereas
the dispersal distance is based on the work of Muller
(1982 Table 1) Demographic parameters were cal-
ibrated to meet criteria published by Verboom et al
(2001) for medium sized birds
Evaluating synergy
Synergy occurs when mixtures of large and small
habitat patches perform better with respect to survival
than landscapes with patches of the same type In this
study we did 250 simulations in the default land-
scapes each run starting with one reproductive unit
ie an adult male and female in every territory We
evaluated metapopulation survival and the mean
number of adult individuals between year 50 and
150 to avoid transient behavior due to the initiali-
zation state (lsquolsquoburn-inrsquorsquo) Since there is a strong
correlation between the mean number of individuals
and survival we present only the survival results in
our graphs Of every survival fraction the 95
confidence interval is calculated using a v-test We
present our results as a replacement series in which
we replace large patches by smaller elements keeping
the same amount of habitat ultimately resulting in the
landscape with small elements only (Fig 3) Monot-
onously increasing relations (small- [ mixed- [large) support the more traditional hypothesis that
Large
Large
Large
Small-P Mix-P
Small-L
Small-PL
Mix-L
Mix-PL
only small mixed only largereplacing
Small toLarge
Small andLinear toLarge
Linear tolarge
Fig 2 Default landscapes
of 6 9 6 km lines are
linear elements of
6000 9 25 m smallsquares are small forest
patches of 25 ha that can
contain one territory Largesquares are large patches of
10 ha that contain four
territories
Landscape Ecol (2009) 241123ndash1133 1127
123
larger patches are always better an optimum rela-
tionship supports our alternative hypothesis that
synergy occurs
Simulations and results
Reference simulations
We evaluate metapopulation survival in the seven
default landscapes for the species parameterized
according to Table 1 When we replace half of the
large patches by small elements these mixed land-
scapes performe best if these small elements were all
linear elements (L Fig 4) Here metapopulation
survival in mixed landscapes was higher than in the
landscape with only large patches indicating syn-
ergy If half of the large patches is replaced by a mix
of small patches and linear elements (LP) metapop-
ulation survival is the same as in the landscapes with
only large patches This indicates that mixed land-
scapes with linear elements perform equally or better
than the landscape with only large patches Mixed
landscapes without linear elements (P) or landscapes
with only small elements however always performe
worse compared to the other landscapes Furthermore
we see that a landscape with only small elements that
contained both linear elements and small patches
(LP) performed much better than landscapes with
only small elements that contained only linear
elements (L) or only small patches (P) These results
suggest that replacing a part of the small patches by
linear elements also enhances population survival
Effects of physical connections
The results suggest that linear elements are respon-
sible for the synergy due to the fact that they connect
patches with each other and guide dispersers To
investigate this effect we create three alternative
landscapes in which the linear elements are not
connected to the patches
If linear elements are not connected to the large
patches number of individuals and survival decrease
in the mixed landscapes with linear elements (L and
LP) (compare Figs 4 5) This indicates that linear
connections are responsible for the synergy found in
the reference conditions Survival in landschapes
Large
Mix-L
Mix-LP
Mix-P
Small-L
Small-LP
Small-P
S
urvi
val o
f met
apop
ulat
ion
Only Mix Onlysmall largeelements patches
L-line
P-line
LP-line
Fig 3 Explanatory graph that relates individual simulation
results to synergy If mixed landscapes perform better than
landscapes with only small and large elements there is synergy
L linear elements present as small elements P small patches
present as small elements LP both linear elements and small
patches present as small elements
0
02
04
06
08
1
only small mix only large
surv
ivin
g m
etap
opul
atio
ns
L
P
LP
Fig 4 Fraction of surviving metapopulations of default
simulations of seven different landscapes that differ with
respect to the percentage of large and small habitat patches or
linear habitat elements error bars represent the 95 confi-
dence inteval Only small landscapes that only consist of small
elements mix landscape that are a mix of large and small
elements only large is a landscape that has only large patches
L linear elements present as small elements P small patches
present as small elements LP both linear elements and small
patches present as small elements
1128 Landscape Ecol (2009) 241123ndash1133
123
with only small elements however is still higher in
landscapes with small patches and linear elements
(LP) than in landscapes with only small patches
Effects of decreasing the dispersal through
the matrix
To investigate whether linear elements are indeed
crucial for synergy we decrease the dispersal effi-
ciency through the matrix by increasing the standard
deviation of the turning angle (SDD) from 5 to 20 and
40 This change results in a decrease of the realized
dispersal distance As a result 90 of the animals
move less than resp 29 and 15 km in the matrix
whereas in the reference situation this was 84 km In
these simulations we use the seven default landscapes
again
As a result of the decreased dispersal efficiency
through the matrix the metapopulation survival in
landscapes without linear elements (P) decrease
strongly (Fig 6a b) resulting in a clear synergy
pattern for the linear replacement lines (L) and
linear combined with small patches replacement
lines (LP Fig 6) Replacing large patches by
smaller patches does not help to compensate for
the loss of dispersal efficiency through the matrix
The landscapes containing only small patches the
decreased dispersal efficiency even caused extinc-
tion in a simualtion for the most inefficient dispersal
class These results indicate that if dispersal through
the matrix is difficult or avoided by a species linear
elements are crucial
Discussion
Main results
It is generally assumed that larger patches of natural
habitat are better for the survival of species than the
same amount of habitat in smaller fragments
(MacArthur and Wilson 1967 Verboom et al 2001
Vos et al 2001) Our results suggest that replacing a
part of the larger units by smaller linear elements
enhance population performance with respect to
population level and survival Without any large
patches however we see a serious decrease in
performance of the population indicating that large
patches are still very important The explanation for
this synergy is that the large patches secure survival
of the population whereas the linear units enhance
inter-patch dispersal by serving as corridors Addi-
tionally we see that populations in landscapes with
small patches also benefit from the availability of
linear elements Also here we see synergy to the
extent that a mixture of linear elements and small
patches performs better than landscapes consisting of
solely linear elements or small patches
In this study linear elements have a double
function they can serve as a corridor but also as
habitat In real landscapes corridors may not be wide
or qualitatively good enough to support both func-
tions Small patches may serve as stepping stones
because the mean inter-patch distance decreases with
increasing number of patches (Etienne and Heester-
beek 2000) However if we reduce the area of the
patches with a certain factor the average inter-patch
distance (centre to centre) is reduced with only the
square root of this factor Consequently replacing
large patches by smaller patches is inferior to
replacing large patches by linear elements
0
02
04
06
08
1
only small mix only large
surv
ing
met
apop
ulat
ions
L
LP
Fig 5 Fraction of surviving metapopulations of simulations
in which the linear elements were not connected to the large
patches of five different landscapes that differ with respect to
the percentage of large and small habitat patches or linear
habitat elements error bars represent the 95 confidence
inteval Only small landscapes that only consist of small
elements mix landscape that are a mix of large and small
elements only large is a landscape that has only large patches
L linear elements present as small elements LP both linear
elements and small patches present as small elements
Landscape Ecol (2009) 241123ndash1133 1129
123
Mechanisms favoring large or small patches
Seven processes are put forward in literature that
determines metapopulation performance in response
to the subdivision of habitat
1 Population extinction larger patches have a
higher carrying capacity and therefore a lower
extinction risk due to Allee-effects and demo-
graphic stochasticity than smaller patches
(Shaffer 1981 Soule 1987)
2 Colonization When dispersal potential is weak
and the dispersal mortality in the matrix is high
however the lsquoSeveral Smallrsquo approach might be
superior (Etienne and Heesterbeek 2000) The
extreme case is when the inter-patch distances of
the large patches are too large for recolonization
whereas the species is able to bridge the smaller
inter-patch distance between the small patches
Here the landscape configuration with few large
patches is actually more fragmented than the
landscape with many small patches (Flather and
Bevers 2002)
3 Finding a mate it seems likely that especially
monogamous animals that live in a landscape
with many small patches have a higher risk to
stay single This lowers the reproduction (Engen
et al 2003)
4 Density dependent population growth in a more
fragmented landscape animals encounter more
density dependent effects because some patches
are at or above carrying capacity while others are
empty Here individuals are not optimally utiliz-
ing the resources This lowers the growth rate of
fragmented populations
5 Patch edge effects if a species utilizes patch
edges more than the patch core several small
patches might be superior over a few large
patches because the amount of edges and
ecotones is larger when the landscape is more
fragmented On the other hand if core habitat is
preferred eg because the influence of the
Fig 6 The effect of increasing the turning angle of dispersing
birds from 5 to 20 and 40 degrees resulting in lower matrix
permeability The figure depict the fraction of surviving
metapopulations in the default landscape and the error bars
represent the 95 confidence interval Only small landscapes
that only consist of small elements mix landscape that are a
mix of large and small elements only large is a landscape that
has only large patches L linear elements present as small
elements P small patches present as small elements LP both
linear elements and small patches present as small elements
c
40 degrees
0
02
04
06
08
1
surv
ivin
g m
etap
opul
atio
ns
L
P
LP
20 degrees
0
02
04
06
08
1
only small mix only large
only small mix only large
only small mix only large
surv
ivin
g m
etap
opul
atio
ns
L
P
LP
5 degrees
0
02
04
06
08
1
surv
ivin
g m
etap
opul
atio
ns
L
P
LP
1130 Landscape Ecol (2009) 241123ndash1133
123
matrix makes the edges suboptimal larger ele-
ments are better
6 Spreading of risk if populations fluctuate due to
local environmental factors subdivision may
save parts of the metapopulation from events
such as fire or other adverse conditions When
species like a predator or a disease are taken into
account a fragmented landscape may be superior
Here predator or disease might also suffers from
isolation leaving their prey or host untouched in
patches that are not yet colonized by the predator
or disease (Baz and GarciaBoyero 1996 Lang-
lois et al 2001 Tscharntke et al 2002) Another
special case is habitat heterogeneity among
patches which makes eg wet patches more
suitable in dry years and dry patches in wet years
(Boer 1981) Although the spreading of risk
might also occur in larger elements it generally
favors population subdivision
7 Round off effects in fragmented landscapes
specific patch carrying capacity might not be
whole even numbers For instance if we sub-
stitute a patch with a carrying capacity of ten by
four fragments with carrying capacities of 25
these patches together can only support eight
breeding pairs
In our simulation study we do not incorporate all
these seven processes We take extinction (due to
demographic stochasticity and Allee-effects) coloni-
zation (dispersal success) pair formation and density
dependent population growth into account We do not
incorporate patch edge effects (when habitat quality
differs between patch core and patch edge) spreading
of risk (local disturbances patch heterogeneity and
antagonistic species) and round off effects These
three missing factors have an evident effect while
round off effects will favor large patch landscapes
and spreading of risk will favor fragmented land-
scapes the effect of taking into account edge effects
will depend on a preference for patch core or edge
habitat but in either case a straightforward relation is
expected Therefore our results may be slightly
modulated by taking into account these aspects but
our conclusions should be robust since some factors
are in favor of small patches and others are in favor of
large patches an optimum configuration is expected
somewhere between the two extreme cases with only
large or only small patches
Climate change
In the face of climate change there might be a few
extra considerations the habitat of species in climatic
terms is no longer stationary so species should be
able to track their climatic window Consequently the
global survival of a species is no longer only
dependent on the survival locally but also on the
ability of species to move and follow the climate
suitability window (Tischendorf et al 2003 Travis
2003 Saether et al 2004 Thomas et al 2004) This
underpins the idea that under climate change condi-
tions heterogeneous landscapes with many linear
elements might be even more superior for species
survival because they both facilitate animal movement
en survival Whereas large patches are necessary to
buffer the population against increasingly stochastic
weather events and provide dispersers connectivity by
linear elements should allow populations to shift
Generality of the results
We did a series of simulations with a spatially
explicit individual-based population model simulat-
ing the population dynamics of a territorial woodland
birds in open agricultural landscapes Our results
indicate that replacing patches by linear elements that
can serve as habitat enhances metapopulation perfor-
mance Clearly we can not extrapolate these results to
conclusions for other species and spatial configura-
tions We think however that our conclusions are
likely to be valid for other species and conditions if
(1) the carrying capacity of the habitat is not affected
by the replacement (2) the dispersal trough habitat is
more efficient and less risky than dispersal through
matrix and (3) if spatial configurations are such that
patches are relatively isolated so that dispersal
limitation is an important factor
Implications for sustainable landscape design
and nature conservation
Since in many countries space is scarce there is an
increasing need for knowledge about how to optimize
the design of landscapes nature reserves and habitat
networks from a biodiversity conservation perspec-
tive (Lomolino 1994 Kingsland 2002 McCarthy
Landscape Ecol (2009) 241123ndash1133 1131
123
et al 2005 McCarthy et al 2006) Our results
indicate that landscape designers and nature conser-
vationists should reconsider the value of small linear
elements that often are considered as part of the
matrix and not always seek to create larger units but
may better create more variable landscapes with
mixtures of patch sizes and shapes This is especially
important when smaller units play a key role in
physically connecting large patches when dispersal
through the matrix is poor or dispersal distances are
smaller than inter-patch distances On the other hand
the smaller units should be large enough to have a
habitat function for the species considered ie
reproduction should be possible there
Acknowledgments This research was financed by the
program lsquolsquoVernieuwend Ruimtegebruikrsquorsquo of Habiforum and
co-financed by the Dutch Ministry of Agriculture Nature and
Food Quality
References
Baveco JM (2006) Small steps movement model manual
Version 10 Alterra-Wageningen UR Wageningen p 19
Baz A GarciaBoyero A (1996) The SLOSS dilemma a
butterfly case study Biodivers Conserv 5493ndash502 doi
101007BF00056393
Bender DJ Tischendorf L Fahrig L (2003) Using patch isolation
metrics to predict animal movement in binary landscapes
Landscape Ecol 1817ndash39 doi101023A10229372
26820
PJd Boer (1981) On the survival of populations in a hetero-
geneous and variable environment Oecologia 5039ndash53
doi101007BF00378792
Chardon JP Adriaensen F Matthysen E (2003) Incorporating
landscape elements into a connectivity measure a case
study for the Speckled wood butterfly (Pararge aegeria L)
Landscape Ecol 18561ndash573 doi101023A10260625
30600
Engen S Lande R Seather BE (2003) Demographic stochas-
ticity and allee effects in populations with two sexes
Ecology 842378ndash2386 doi10189002-0123
Etienne RS Heesterbeek JAP (2000) On optimal size and
number of reserves for metapopulation persistence
J Theor Biol 20333ndash50 doi101006jtbi19991060
Flather CH Bevers M (2002) Patchy reaction-diffusion
and population abundance the relative importance of
habitat amount and arrangement Am Nat 15940ndash56 doi
101086324120
Grashof-Bokdam CJ Chardon P Vos CC Foppen RPB
Wallisdevries MF VanDerVeen M Meeuwsen HAM
(2008) The synergistic effect of combining woodlands
and green veining for biodiversity Landscape Ecol doi
101007s10980-008-9274-2
Grashof-Bokdam CJ van Langevelde F (2005) Green veining
landscape determinants of biodiversity in European
agricultural landscapes Landscape Ecol 20417ndash439
doi101007s10980-004-5646-1
Haddad NM (1999) Corridor and distance effects on interpatch
movements a landscape experiment with butterflies Ecol
Appl 9612ndash622 doi1018901051-0761(1999)009[0612
CADEOI]20CO2
Kingsland SE (2002) Creating a science of nature reserve
design perspectives from history Environ Model Assess
761ndash69 doi101023A1015633830223
Kosenko SM Kaigorodova EY (2001) Effect of habitat frag-
mentation on distribution density and breeding
performance of the middle spotted woodpecker Dend-
rocopos medius (Alves Picidae) in Nerussa-Desna
Polesye Zool Zh 8071ndash78
Kosinski Z Ksit P (2006) Comparative reproductive biology of
middle spotted woodpeckers Dendrocopos medius and
great spotted woodpeckers D-major in a riverine forest
Bird Study 53237ndash246
Langlois JP Fahrig L Merriam G Artsob H (2001) Landscape
structure influences continental distribution of hantavirus
in deer mice Landscape Ecol 16255ndash266 doi101023
A1011148316537
Lomolino MV (1994) An evaluation of alternative strategies
for building networks of nature-reserves Biol Conserv
69243ndash249 doi1010160006-3207(94)90423-5
MacArthur RH Wilson EO (1967) The theory of island bio-
geography Princeton University Press Princeton
McCarthy MA Thompson CJ Possingham HP (2005) Theory
for designing nature reserves for single species Am Nat
165250ndash257 doi101086427297
McCarthy MA Thompson CJ Williams NSG (2006) Logic for
designing nature reserves for multiple species Am Nat
167717ndash727 doi101086503058
Muller W (1982) Die Besiedlung der Eichenwalder im Kanton
Zurich durch den Mittelspecht Dendrocopus medius Orn
Beob 79105ndash119
Ovaskainen O (2002) Long-term persistence of species and the
SLOSS problem J Theor Biol 218419ndash433
Patterson BD Atmar W (1986) Nested subsets and the structure
of insular mammalian faunas and archipelagoes Biol J
Linn Soc Lond 2865ndash82 doi101111j1095-83121986
tb01749x
Pettersson B (1985) Extinction of an isolated population of the
middle spotted woodpecker Dendrocopos-Medius (L) in
Sweden and its relation to general theories on extinction
Biol Conserv 32335ndash353 doi1010160006-3207(85)
90022-9
Ricketts TH (2001) The matrix matters Effective isolation in
fragmented landscapes Am Nat 15887ndash99 doi101086
320863
Saether BE Sutherland WJ Engen S (2004) Climate influences
on avian population dynamics In Moller AP Fielder W
Berthold P (eds) Birds and climate change pp 185ndash209
Schippers P Verboom J Knaapen JP vanApeldoorn RC
(1996) Dispersal and habitat connectivity in complex
heterogeneous landscapes an analysis with a GIS-based
random walk model Ecography 1997ndash106 doi101111
j1600-05871996tb00160x
Schultz CB Crone EE (2005) Patch size and connectivity
thresholds for butterfly habitat restoration Conserv Biol
19887ndash896 doi101111j1523-1739200500462x
1132 Landscape Ecol (2009) 241123ndash1133
123
Shaffer ML (1981) Minimum population sizes for species con-
servation Bioscience 31131ndash134 doi1023071308256
Soule M (1987) Viable populations for conservation
Cambridge University Press Cambridge
Tellerıa JL Baquero R Santos T (2003) Effects of forest
fragmentation on European birds implications of regional
differences in species richness J Biogeogr 30621ndash628
doi101046j1365-2699200300960x
Thomas CD Cameron A Green RE Bakkenes M Beaumont LJ
Collingham YC Erasmus BFN de Siqueira MF Grainger
A Hannah L Hughes L Huntley B van Jaarsveld AS
Midgley GF Miles L Ortega-Huerta MA Peterson AT
Phillips OL Williams SE (2004) Extinction risk from
climate change Nature 427145ndash148 doi101038nature
02121
Thomas CD Kunin WE (1999) The spatial structure of pop-
ulations J Anim Ecol 68647ndash657 doi101046j1365-
2656199900330x
Tischendorf L Bender DJ Fahrig L (2003) Evaluation of patch
isolation metrics in mosaic landscapes for specialist vs
generalist dispersers Landscape Ecol 1841ndash50 doi
101023A1022908109982
Tischendorf L Irmler U Hingst R (1998) A simulation
experiment on the potential of hedgerows as movement
corridors for forest carabids Ecol Modell 106107ndash118
doi101016S0304-3800(97)00186-5
Tischendorf L Wissel C (1997) Corridors as conduits for small
animals attainable distances depending on movement
pattern boundary reaction and corridor width Oikos
79603ndash611 doi1023073546904
Travis JMJ (2003) Climate change and habitat destruction a
deadly anthropogenic cocktail Proc R Soc Lond B Biol
Sci 270467ndash473 doi101098rspb20022246
Tscharntke T Steffan-Dewenter I Kruess A Thies C (2002)
Contribution of small habitat fragments to conservation of
insect communities of grassland-cropland landscapes
Ecol Appl 12354ndash363
Van Apeldoorn RC Knaapen JP Schippers P Verboom J
Engen van H Meeuwsen H (1998) Landscape planning
and conservation biology simulation models as tools to
evaluate scenarios for badger in the Netherlands Land-
scape Urban Plan 4157ndash69 doi101016S0169-2046(97)
00058-3
VanDorp D Schippers P Groenendael van JM (1997)
Migration rates of grassland plants along corridors in
fragmented landscapes assessed with a cellular automa-
tion model Landscape Ecol 1239ndash50 doi101007
BF02698206
Verboom J Foppen R Chardon P Opdam P Luttikhuizen P
(2001) Introducing the key patch approach for habitat
networks with persistent populations an example for
marshland birds Biol Conserv 10089ndash101 doi101016
S0006-3207(00)00210-X
Vos CC Verboom J Opdam PFM Ter Braak CJF (2001)
Toward ecologically scaled landscape indices Am Nat
15724ndash41 doi101086317004
Wiens JA (1995) Landscape mosaics and ecological theory In
Hansson L Fahrig L Merriam G (eds) Mosaic landscapes
and ecological processes Chapman and Hall London
pp 1ndash26
Landscape Ecol (2009) 241123ndash1133 1133
123
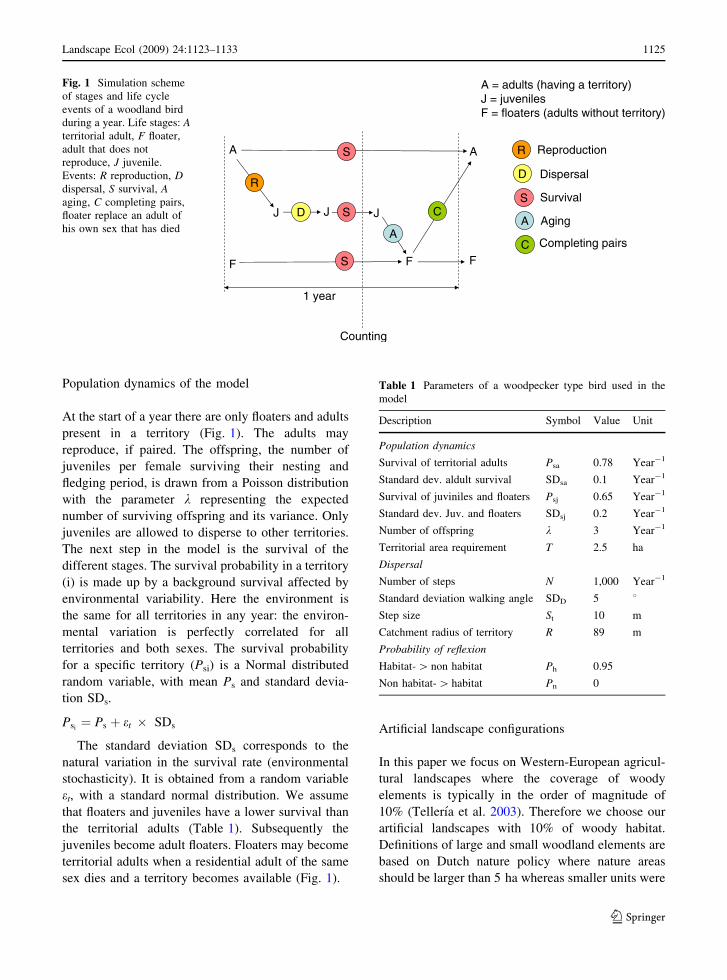
Population dynamics of the model
At the start of a year there are only floaters and adults
present in a territory (Fig 1) The adults may
reproduce if paired The offspring the number of
juveniles per female surviving their nesting and
fledging period is drawn from a Poisson distribution
with the parameter k representing the expected
number of surviving offspring and its variance Only
juveniles are allowed to disperse to other territories
The next step in the model is the survival of the
different stages The survival probability in a territory
(i) is made up by a background survival affected by
environmental variability Here the environment is
the same for all territories in any year the environ-
mental variation is perfectly correlated for all
territories and both sexes The survival probability
for a specific territory (Psi) is a Normal distributed
random variable with mean Ps and standard devia-
tion SDs
Psifrac14 Ps thorn et SDs
The standard deviation SDs corresponds to the
natural variation in the survival rate (environmental
stochasticity) It is obtained from a random variable
et with a standard normal distribution We assume
that floaters and juveniles have a lower survival than
the territorial adults (Table 1) Subsequently the
juveniles become adult floaters Floaters may become
territorial adults when a residential adult of the same
sex dies and a territory becomes available (Fig 1)
Artificial landscape configurations
In this paper we focus on Western-European agricul-
tural landscapes where the coverage of woody
elements is typically in the order of magnitude of
10 (Tellerıa et al 2003) Therefore we choose our
artificial landscapes with 10 of woody habitat
Definitions of large and small woodland elements are
based on Dutch nature policy where nature areas
should be larger than 5 ha whereas smaller units were
A A
J
F
S
RD
D
S FC
1 year
Counting
R Reproduction
Dispersal
S SurvivalCJ
Completing pairs
S J
F
A = adults (having a territory)J = juvenilesF = floaters (adults without territory)
AA Aging
Fig 1 Simulation scheme
of stages and life cycle
events of a woodland bird
during a year Life stages Aterritorial adult F floater
adult that does not
reproduce J juvenile
Events R reproduction Ddispersal S survival Aaging C completing pairs
floater replace an adult of
his own sex that has died
Table 1 Parameters of a woodpecker type bird used in the
model
Description Symbol Value Unit
Population dynamics
Survival of territorial adults Psa 078 Year-1
Standard dev aldult survival SDsa 01 Year-1
Survival of juviniles and floaters Psj 065 Year-1
Standard dev Juv and floaters SDsj 02 Year-1
Number of offspring k 3 Year-1
Territorial area requirement T 25 ha
Dispersal
Number of steps N 1000 Year-1
Standard deviation walking angle SDD 5 8
Step size St 10 m
Catchment radius of territory R 89 m
Probability of reflexion
Habitat- [ non habitat Ph 095
Non habitat- [ habitat Pn 0
Landscape Ecol (2009) 241123ndash1133 1125
123
considered to be part of the matrix (Grashof-Bokdam
et al 2008) Therefore we define three landscape
elements small patches of one territory that are
25 ha and linear elements that are 25 ha but have a
width of 25 m and large patches of four territories
that are 10 ha So here we simulate the matrix
structure explicitly To avoid confusion however we
will use the term matrix to describe the non habitat
part of the landscape The total area of the artificial
landscapes is 6 9 6 km
We use a replacement approach to construct
different landscapes We start with a landscape with
only large elements subsequently we replace large
elements by small patches andor linear elements
keeping the same total amount of habitat This results
in seven landscape types all having 10 of habitat
area but differing with respect to the total abundance
of the different landscape elements (Table 2) We
choose the maximal connection version of linear
elements and patches of the landscape as our default
landscape Here linear elements are as much as
possible physically connected to the patches (Fig 2)
In the simulations and results section we will evaluate
what the effect is of this choice Additionally we
define the precise location of the territories To do
this we first overlay the landscape with a large cloud
of random points and check around every point
whether there is enough habitat (25 ha) to support a
territory We continue this procedure until we have
144 territories in all the landscape types As a result
we get landscapes havening all 10 of habitat and
90 non-habitat with scattered territories located in
the 10 habitat Animals are allocated to a territory
when they do not disperse Dispersing animals are
allowed to move freely in all directions in this
landscape in both habitat and non-habitat according
to the rules described in the next section until they are
within a critical radius from a new territory and are
allocated to this territory
Random walk dispersal
We simulate the dispersal between territories using
SmallSteps a separate movement module (Baveco
2006) This module simulates the dispersal of the
juveniles from a territory in which they were born to
a territory in which they settle The animal movement
in the model is in fact a correlated random walk
(Schippers et al 1996 Tischendorf and Wissel 1997
Tischendorf et al 1998 Haddad 1999) In this model
a dispersing animal has a coordinate in a continuous
xndashy space and a walking direction between 0 and
360 Each time step a new direction (Dt) relative to
the old direction (Dt-1) is calculated
Dt frac14 modethDt1 thorn et SDD 2pTHORN
where et is a number drawn from a standard normal
distribution SDD is the standard deviation of the
walking direction lsquolsquomodrsquorsquo is a modulus function with
2p (rad) as the basis number Next the dispersing
animal takes a step in this new direction with a step
size (St) of 10 m subsequently the new position is
calculated A dispersing animal is allowed to take
1000 steps during dispersal If during dispersal in
habitat an animal meets non habitat it has a proba-
bility to be reflected Ph of 95 (Table 1) Reflecting
means that the walking direction is mirrored against
the habitat edge When a dispersing animal however
enter habitat the animal continues its path Animals
continue dispersing until they find a new territory that
is not occupied by a territorial adult of the same sex
If an animal has not found another suitable patch after
Table 2 Habitat cover
( of total) of different
landscapes
Landscape
description
Habitat large
patches
Habitat small
patches
Habitat linear
elements
Matrix
Large 10 0 0 90
Mix-P 5 5 0 90
Mix-L 5 0 5 90
Mixed-LP 5 25 25 90
Small-P 0 10 0 90
Small-L 0 0 10 90
Small-LP 0 5 5 90
1126 Landscape Ecol (2009) 241123ndash1133
123
1000 steps and is not located in habitat the dispersing
animal dies Animals that arrive in habitat after 1000
steps but not in a vacant territory are allocated to the
nearest territory Animals that arrive in a new
territory become floaters and may reproduce when
the territorial adult of the same sex dies Animals that
encounter the landscape edge are reflected These
rules cause that dispersing animals first disperse in
the habitat patch of their territory checking neighbor
territories before they start dispersing through the
matrix and might find another habitat patch and
territories in there
Parameterization and calibration
We use demographic parameters of the middle
spotted woodpecker (Pettersson 1985 Kosenko and
Kaigorodova 2001 Kosinski and Ksit 2006) whereas
the dispersal distance is based on the work of Muller
(1982 Table 1) Demographic parameters were cal-
ibrated to meet criteria published by Verboom et al
(2001) for medium sized birds
Evaluating synergy
Synergy occurs when mixtures of large and small
habitat patches perform better with respect to survival
than landscapes with patches of the same type In this
study we did 250 simulations in the default land-
scapes each run starting with one reproductive unit
ie an adult male and female in every territory We
evaluated metapopulation survival and the mean
number of adult individuals between year 50 and
150 to avoid transient behavior due to the initiali-
zation state (lsquolsquoburn-inrsquorsquo) Since there is a strong
correlation between the mean number of individuals
and survival we present only the survival results in
our graphs Of every survival fraction the 95
confidence interval is calculated using a v-test We
present our results as a replacement series in which
we replace large patches by smaller elements keeping
the same amount of habitat ultimately resulting in the
landscape with small elements only (Fig 3) Monot-
onously increasing relations (small- [ mixed- [large) support the more traditional hypothesis that
Large
Large
Large
Small-P Mix-P
Small-L
Small-PL
Mix-L
Mix-PL
only small mixed only largereplacing
Small toLarge
Small andLinear toLarge
Linear tolarge
Fig 2 Default landscapes
of 6 9 6 km lines are
linear elements of
6000 9 25 m smallsquares are small forest
patches of 25 ha that can
contain one territory Largesquares are large patches of
10 ha that contain four
territories
Landscape Ecol (2009) 241123ndash1133 1127
123
larger patches are always better an optimum rela-
tionship supports our alternative hypothesis that
synergy occurs
Simulations and results
Reference simulations
We evaluate metapopulation survival in the seven
default landscapes for the species parameterized
according to Table 1 When we replace half of the
large patches by small elements these mixed land-
scapes performe best if these small elements were all
linear elements (L Fig 4) Here metapopulation
survival in mixed landscapes was higher than in the
landscape with only large patches indicating syn-
ergy If half of the large patches is replaced by a mix
of small patches and linear elements (LP) metapop-
ulation survival is the same as in the landscapes with
only large patches This indicates that mixed land-
scapes with linear elements perform equally or better
than the landscape with only large patches Mixed
landscapes without linear elements (P) or landscapes
with only small elements however always performe
worse compared to the other landscapes Furthermore
we see that a landscape with only small elements that
contained both linear elements and small patches
(LP) performed much better than landscapes with
only small elements that contained only linear
elements (L) or only small patches (P) These results
suggest that replacing a part of the small patches by
linear elements also enhances population survival
Effects of physical connections
The results suggest that linear elements are respon-
sible for the synergy due to the fact that they connect
patches with each other and guide dispersers To
investigate this effect we create three alternative
landscapes in which the linear elements are not
connected to the patches
If linear elements are not connected to the large
patches number of individuals and survival decrease
in the mixed landscapes with linear elements (L and
LP) (compare Figs 4 5) This indicates that linear
connections are responsible for the synergy found in
the reference conditions Survival in landschapes
Large
Mix-L
Mix-LP
Mix-P
Small-L
Small-LP
Small-P
S
urvi
val o
f met
apop
ulat
ion
Only Mix Onlysmall largeelements patches
L-line
P-line
LP-line
Fig 3 Explanatory graph that relates individual simulation
results to synergy If mixed landscapes perform better than
landscapes with only small and large elements there is synergy
L linear elements present as small elements P small patches
present as small elements LP both linear elements and small
patches present as small elements
0
02
04
06
08
1
only small mix only large
surv
ivin
g m
etap
opul
atio
ns
L
P
LP
Fig 4 Fraction of surviving metapopulations of default
simulations of seven different landscapes that differ with
respect to the percentage of large and small habitat patches or
linear habitat elements error bars represent the 95 confi-
dence inteval Only small landscapes that only consist of small
elements mix landscape that are a mix of large and small
elements only large is a landscape that has only large patches
L linear elements present as small elements P small patches
present as small elements LP both linear elements and small
patches present as small elements
1128 Landscape Ecol (2009) 241123ndash1133
123
with only small elements however is still higher in
landscapes with small patches and linear elements
(LP) than in landscapes with only small patches
Effects of decreasing the dispersal through
the matrix
To investigate whether linear elements are indeed
crucial for synergy we decrease the dispersal effi-
ciency through the matrix by increasing the standard
deviation of the turning angle (SDD) from 5 to 20 and
40 This change results in a decrease of the realized
dispersal distance As a result 90 of the animals
move less than resp 29 and 15 km in the matrix
whereas in the reference situation this was 84 km In
these simulations we use the seven default landscapes
again
As a result of the decreased dispersal efficiency
through the matrix the metapopulation survival in
landscapes without linear elements (P) decrease
strongly (Fig 6a b) resulting in a clear synergy
pattern for the linear replacement lines (L) and
linear combined with small patches replacement
lines (LP Fig 6) Replacing large patches by
smaller patches does not help to compensate for
the loss of dispersal efficiency through the matrix
The landscapes containing only small patches the
decreased dispersal efficiency even caused extinc-
tion in a simualtion for the most inefficient dispersal
class These results indicate that if dispersal through
the matrix is difficult or avoided by a species linear
elements are crucial
Discussion
Main results
It is generally assumed that larger patches of natural
habitat are better for the survival of species than the
same amount of habitat in smaller fragments
(MacArthur and Wilson 1967 Verboom et al 2001
Vos et al 2001) Our results suggest that replacing a
part of the larger units by smaller linear elements
enhance population performance with respect to
population level and survival Without any large
patches however we see a serious decrease in
performance of the population indicating that large
patches are still very important The explanation for
this synergy is that the large patches secure survival
of the population whereas the linear units enhance
inter-patch dispersal by serving as corridors Addi-
tionally we see that populations in landscapes with
small patches also benefit from the availability of
linear elements Also here we see synergy to the
extent that a mixture of linear elements and small
patches performs better than landscapes consisting of
solely linear elements or small patches
In this study linear elements have a double
function they can serve as a corridor but also as
habitat In real landscapes corridors may not be wide
or qualitatively good enough to support both func-
tions Small patches may serve as stepping stones
because the mean inter-patch distance decreases with
increasing number of patches (Etienne and Heester-
beek 2000) However if we reduce the area of the
patches with a certain factor the average inter-patch
distance (centre to centre) is reduced with only the
square root of this factor Consequently replacing
large patches by smaller patches is inferior to
replacing large patches by linear elements
0
02
04
06
08
1
only small mix only large
surv
ing
met
apop
ulat
ions
L
LP
Fig 5 Fraction of surviving metapopulations of simulations
in which the linear elements were not connected to the large
patches of five different landscapes that differ with respect to
the percentage of large and small habitat patches or linear
habitat elements error bars represent the 95 confidence
inteval Only small landscapes that only consist of small
elements mix landscape that are a mix of large and small
elements only large is a landscape that has only large patches
L linear elements present as small elements LP both linear
elements and small patches present as small elements
Landscape Ecol (2009) 241123ndash1133 1129
123
Mechanisms favoring large or small patches
Seven processes are put forward in literature that
determines metapopulation performance in response
to the subdivision of habitat
1 Population extinction larger patches have a
higher carrying capacity and therefore a lower
extinction risk due to Allee-effects and demo-
graphic stochasticity than smaller patches
(Shaffer 1981 Soule 1987)
2 Colonization When dispersal potential is weak
and the dispersal mortality in the matrix is high
however the lsquoSeveral Smallrsquo approach might be
superior (Etienne and Heesterbeek 2000) The
extreme case is when the inter-patch distances of
the large patches are too large for recolonization
whereas the species is able to bridge the smaller
inter-patch distance between the small patches
Here the landscape configuration with few large
patches is actually more fragmented than the
landscape with many small patches (Flather and
Bevers 2002)
3 Finding a mate it seems likely that especially
monogamous animals that live in a landscape
with many small patches have a higher risk to
stay single This lowers the reproduction (Engen
et al 2003)
4 Density dependent population growth in a more
fragmented landscape animals encounter more
density dependent effects because some patches
are at or above carrying capacity while others are
empty Here individuals are not optimally utiliz-
ing the resources This lowers the growth rate of
fragmented populations
5 Patch edge effects if a species utilizes patch
edges more than the patch core several small
patches might be superior over a few large
patches because the amount of edges and
ecotones is larger when the landscape is more
fragmented On the other hand if core habitat is
preferred eg because the influence of the
Fig 6 The effect of increasing the turning angle of dispersing
birds from 5 to 20 and 40 degrees resulting in lower matrix
permeability The figure depict the fraction of surviving
metapopulations in the default landscape and the error bars
represent the 95 confidence interval Only small landscapes
that only consist of small elements mix landscape that are a
mix of large and small elements only large is a landscape that
has only large patches L linear elements present as small
elements P small patches present as small elements LP both
linear elements and small patches present as small elements
c
40 degrees
0
02
04
06
08
1
surv
ivin
g m
etap
opul
atio
ns
L
P
LP
20 degrees
0
02
04
06
08
1
only small mix only large
only small mix only large
only small mix only large
surv
ivin
g m
etap
opul
atio
ns
L
P
LP
5 degrees
0
02
04
06
08
1
surv
ivin
g m
etap
opul
atio
ns
L
P
LP
1130 Landscape Ecol (2009) 241123ndash1133
123
matrix makes the edges suboptimal larger ele-
ments are better
6 Spreading of risk if populations fluctuate due to
local environmental factors subdivision may
save parts of the metapopulation from events
such as fire or other adverse conditions When
species like a predator or a disease are taken into
account a fragmented landscape may be superior
Here predator or disease might also suffers from
isolation leaving their prey or host untouched in
patches that are not yet colonized by the predator
or disease (Baz and GarciaBoyero 1996 Lang-
lois et al 2001 Tscharntke et al 2002) Another
special case is habitat heterogeneity among
patches which makes eg wet patches more
suitable in dry years and dry patches in wet years
(Boer 1981) Although the spreading of risk
might also occur in larger elements it generally
favors population subdivision
7 Round off effects in fragmented landscapes
specific patch carrying capacity might not be
whole even numbers For instance if we sub-
stitute a patch with a carrying capacity of ten by
four fragments with carrying capacities of 25
these patches together can only support eight
breeding pairs
In our simulation study we do not incorporate all
these seven processes We take extinction (due to
demographic stochasticity and Allee-effects) coloni-
zation (dispersal success) pair formation and density
dependent population growth into account We do not
incorporate patch edge effects (when habitat quality
differs between patch core and patch edge) spreading
of risk (local disturbances patch heterogeneity and
antagonistic species) and round off effects These
three missing factors have an evident effect while
round off effects will favor large patch landscapes
and spreading of risk will favor fragmented land-
scapes the effect of taking into account edge effects
will depend on a preference for patch core or edge
habitat but in either case a straightforward relation is
expected Therefore our results may be slightly
modulated by taking into account these aspects but
our conclusions should be robust since some factors
are in favor of small patches and others are in favor of
large patches an optimum configuration is expected
somewhere between the two extreme cases with only
large or only small patches
Climate change
In the face of climate change there might be a few
extra considerations the habitat of species in climatic
terms is no longer stationary so species should be
able to track their climatic window Consequently the
global survival of a species is no longer only
dependent on the survival locally but also on the
ability of species to move and follow the climate
suitability window (Tischendorf et al 2003 Travis
2003 Saether et al 2004 Thomas et al 2004) This
underpins the idea that under climate change condi-
tions heterogeneous landscapes with many linear
elements might be even more superior for species
survival because they both facilitate animal movement
en survival Whereas large patches are necessary to
buffer the population against increasingly stochastic
weather events and provide dispersers connectivity by
linear elements should allow populations to shift
Generality of the results
We did a series of simulations with a spatially
explicit individual-based population model simulat-
ing the population dynamics of a territorial woodland
birds in open agricultural landscapes Our results
indicate that replacing patches by linear elements that
can serve as habitat enhances metapopulation perfor-
mance Clearly we can not extrapolate these results to
conclusions for other species and spatial configura-
tions We think however that our conclusions are
likely to be valid for other species and conditions if
(1) the carrying capacity of the habitat is not affected
by the replacement (2) the dispersal trough habitat is
more efficient and less risky than dispersal through
matrix and (3) if spatial configurations are such that
patches are relatively isolated so that dispersal
limitation is an important factor
Implications for sustainable landscape design
and nature conservation
Since in many countries space is scarce there is an
increasing need for knowledge about how to optimize
the design of landscapes nature reserves and habitat
networks from a biodiversity conservation perspec-
tive (Lomolino 1994 Kingsland 2002 McCarthy
Landscape Ecol (2009) 241123ndash1133 1131
123
et al 2005 McCarthy et al 2006) Our results
indicate that landscape designers and nature conser-
vationists should reconsider the value of small linear
elements that often are considered as part of the
matrix and not always seek to create larger units but
may better create more variable landscapes with
mixtures of patch sizes and shapes This is especially
important when smaller units play a key role in
physically connecting large patches when dispersal
through the matrix is poor or dispersal distances are
smaller than inter-patch distances On the other hand
the smaller units should be large enough to have a
habitat function for the species considered ie
reproduction should be possible there
Acknowledgments This research was financed by the
program lsquolsquoVernieuwend Ruimtegebruikrsquorsquo of Habiforum and
co-financed by the Dutch Ministry of Agriculture Nature and
Food Quality
References
Baveco JM (2006) Small steps movement model manual
Version 10 Alterra-Wageningen UR Wageningen p 19
Baz A GarciaBoyero A (1996) The SLOSS dilemma a
butterfly case study Biodivers Conserv 5493ndash502 doi
101007BF00056393
Bender DJ Tischendorf L Fahrig L (2003) Using patch isolation
metrics to predict animal movement in binary landscapes
Landscape Ecol 1817ndash39 doi101023A10229372
26820
PJd Boer (1981) On the survival of populations in a hetero-
geneous and variable environment Oecologia 5039ndash53
doi101007BF00378792
Chardon JP Adriaensen F Matthysen E (2003) Incorporating
landscape elements into a connectivity measure a case
study for the Speckled wood butterfly (Pararge aegeria L)
Landscape Ecol 18561ndash573 doi101023A10260625
30600
Engen S Lande R Seather BE (2003) Demographic stochas-
ticity and allee effects in populations with two sexes
Ecology 842378ndash2386 doi10189002-0123
Etienne RS Heesterbeek JAP (2000) On optimal size and
number of reserves for metapopulation persistence
J Theor Biol 20333ndash50 doi101006jtbi19991060
Flather CH Bevers M (2002) Patchy reaction-diffusion
and population abundance the relative importance of
habitat amount and arrangement Am Nat 15940ndash56 doi
101086324120
Grashof-Bokdam CJ Chardon P Vos CC Foppen RPB
Wallisdevries MF VanDerVeen M Meeuwsen HAM
(2008) The synergistic effect of combining woodlands
and green veining for biodiversity Landscape Ecol doi
101007s10980-008-9274-2
Grashof-Bokdam CJ van Langevelde F (2005) Green veining
landscape determinants of biodiversity in European
agricultural landscapes Landscape Ecol 20417ndash439
doi101007s10980-004-5646-1
Haddad NM (1999) Corridor and distance effects on interpatch
movements a landscape experiment with butterflies Ecol
Appl 9612ndash622 doi1018901051-0761(1999)009[0612
CADEOI]20CO2
Kingsland SE (2002) Creating a science of nature reserve
design perspectives from history Environ Model Assess
761ndash69 doi101023A1015633830223
Kosenko SM Kaigorodova EY (2001) Effect of habitat frag-
mentation on distribution density and breeding
performance of the middle spotted woodpecker Dend-
rocopos medius (Alves Picidae) in Nerussa-Desna
Polesye Zool Zh 8071ndash78
Kosinski Z Ksit P (2006) Comparative reproductive biology of
middle spotted woodpeckers Dendrocopos medius and
great spotted woodpeckers D-major in a riverine forest
Bird Study 53237ndash246
Langlois JP Fahrig L Merriam G Artsob H (2001) Landscape
structure influences continental distribution of hantavirus
in deer mice Landscape Ecol 16255ndash266 doi101023
A1011148316537
Lomolino MV (1994) An evaluation of alternative strategies
for building networks of nature-reserves Biol Conserv
69243ndash249 doi1010160006-3207(94)90423-5
MacArthur RH Wilson EO (1967) The theory of island bio-
geography Princeton University Press Princeton
McCarthy MA Thompson CJ Possingham HP (2005) Theory
for designing nature reserves for single species Am Nat
165250ndash257 doi101086427297
McCarthy MA Thompson CJ Williams NSG (2006) Logic for
designing nature reserves for multiple species Am Nat
167717ndash727 doi101086503058
Muller W (1982) Die Besiedlung der Eichenwalder im Kanton
Zurich durch den Mittelspecht Dendrocopus medius Orn
Beob 79105ndash119
Ovaskainen O (2002) Long-term persistence of species and the
SLOSS problem J Theor Biol 218419ndash433
Patterson BD Atmar W (1986) Nested subsets and the structure
of insular mammalian faunas and archipelagoes Biol J
Linn Soc Lond 2865ndash82 doi101111j1095-83121986
tb01749x
Pettersson B (1985) Extinction of an isolated population of the
middle spotted woodpecker Dendrocopos-Medius (L) in
Sweden and its relation to general theories on extinction
Biol Conserv 32335ndash353 doi1010160006-3207(85)
90022-9
Ricketts TH (2001) The matrix matters Effective isolation in
fragmented landscapes Am Nat 15887ndash99 doi101086
320863
Saether BE Sutherland WJ Engen S (2004) Climate influences
on avian population dynamics In Moller AP Fielder W
Berthold P (eds) Birds and climate change pp 185ndash209
Schippers P Verboom J Knaapen JP vanApeldoorn RC
(1996) Dispersal and habitat connectivity in complex
heterogeneous landscapes an analysis with a GIS-based
random walk model Ecography 1997ndash106 doi101111
j1600-05871996tb00160x
Schultz CB Crone EE (2005) Patch size and connectivity
thresholds for butterfly habitat restoration Conserv Biol
19887ndash896 doi101111j1523-1739200500462x
1132 Landscape Ecol (2009) 241123ndash1133
123
Shaffer ML (1981) Minimum population sizes for species con-
servation Bioscience 31131ndash134 doi1023071308256
Soule M (1987) Viable populations for conservation
Cambridge University Press Cambridge
Tellerıa JL Baquero R Santos T (2003) Effects of forest
fragmentation on European birds implications of regional
differences in species richness J Biogeogr 30621ndash628
doi101046j1365-2699200300960x
Thomas CD Cameron A Green RE Bakkenes M Beaumont LJ
Collingham YC Erasmus BFN de Siqueira MF Grainger
A Hannah L Hughes L Huntley B van Jaarsveld AS
Midgley GF Miles L Ortega-Huerta MA Peterson AT
Phillips OL Williams SE (2004) Extinction risk from
climate change Nature 427145ndash148 doi101038nature
02121
Thomas CD Kunin WE (1999) The spatial structure of pop-
ulations J Anim Ecol 68647ndash657 doi101046j1365-
2656199900330x
Tischendorf L Bender DJ Fahrig L (2003) Evaluation of patch
isolation metrics in mosaic landscapes for specialist vs
generalist dispersers Landscape Ecol 1841ndash50 doi
101023A1022908109982
Tischendorf L Irmler U Hingst R (1998) A simulation
experiment on the potential of hedgerows as movement
corridors for forest carabids Ecol Modell 106107ndash118
doi101016S0304-3800(97)00186-5
Tischendorf L Wissel C (1997) Corridors as conduits for small
animals attainable distances depending on movement
pattern boundary reaction and corridor width Oikos
79603ndash611 doi1023073546904
Travis JMJ (2003) Climate change and habitat destruction a
deadly anthropogenic cocktail Proc R Soc Lond B Biol
Sci 270467ndash473 doi101098rspb20022246
Tscharntke T Steffan-Dewenter I Kruess A Thies C (2002)
Contribution of small habitat fragments to conservation of
insect communities of grassland-cropland landscapes
Ecol Appl 12354ndash363
Van Apeldoorn RC Knaapen JP Schippers P Verboom J
Engen van H Meeuwsen H (1998) Landscape planning
and conservation biology simulation models as tools to
evaluate scenarios for badger in the Netherlands Land-
scape Urban Plan 4157ndash69 doi101016S0169-2046(97)
00058-3
VanDorp D Schippers P Groenendael van JM (1997)
Migration rates of grassland plants along corridors in
fragmented landscapes assessed with a cellular automa-
tion model Landscape Ecol 1239ndash50 doi101007
BF02698206
Verboom J Foppen R Chardon P Opdam P Luttikhuizen P
(2001) Introducing the key patch approach for habitat
networks with persistent populations an example for
marshland birds Biol Conserv 10089ndash101 doi101016
S0006-3207(00)00210-X
Vos CC Verboom J Opdam PFM Ter Braak CJF (2001)
Toward ecologically scaled landscape indices Am Nat
15724ndash41 doi101086317004
Wiens JA (1995) Landscape mosaics and ecological theory In
Hansson L Fahrig L Merriam G (eds) Mosaic landscapes
and ecological processes Chapman and Hall London
pp 1ndash26
Landscape Ecol (2009) 241123ndash1133 1133
123

considered to be part of the matrix (Grashof-Bokdam
et al 2008) Therefore we define three landscape
elements small patches of one territory that are
25 ha and linear elements that are 25 ha but have a
width of 25 m and large patches of four territories
that are 10 ha So here we simulate the matrix
structure explicitly To avoid confusion however we
will use the term matrix to describe the non habitat
part of the landscape The total area of the artificial
landscapes is 6 9 6 km
We use a replacement approach to construct
different landscapes We start with a landscape with
only large elements subsequently we replace large
elements by small patches andor linear elements
keeping the same total amount of habitat This results
in seven landscape types all having 10 of habitat
area but differing with respect to the total abundance
of the different landscape elements (Table 2) We
choose the maximal connection version of linear
elements and patches of the landscape as our default
landscape Here linear elements are as much as
possible physically connected to the patches (Fig 2)
In the simulations and results section we will evaluate
what the effect is of this choice Additionally we
define the precise location of the territories To do
this we first overlay the landscape with a large cloud
of random points and check around every point
whether there is enough habitat (25 ha) to support a
territory We continue this procedure until we have
144 territories in all the landscape types As a result
we get landscapes havening all 10 of habitat and
90 non-habitat with scattered territories located in
the 10 habitat Animals are allocated to a territory
when they do not disperse Dispersing animals are
allowed to move freely in all directions in this
landscape in both habitat and non-habitat according
to the rules described in the next section until they are
within a critical radius from a new territory and are
allocated to this territory
Random walk dispersal
We simulate the dispersal between territories using
SmallSteps a separate movement module (Baveco
2006) This module simulates the dispersal of the
juveniles from a territory in which they were born to
a territory in which they settle The animal movement
in the model is in fact a correlated random walk
(Schippers et al 1996 Tischendorf and Wissel 1997
Tischendorf et al 1998 Haddad 1999) In this model
a dispersing animal has a coordinate in a continuous
xndashy space and a walking direction between 0 and
360 Each time step a new direction (Dt) relative to
the old direction (Dt-1) is calculated
Dt frac14 modethDt1 thorn et SDD 2pTHORN
where et is a number drawn from a standard normal
distribution SDD is the standard deviation of the
walking direction lsquolsquomodrsquorsquo is a modulus function with
2p (rad) as the basis number Next the dispersing
animal takes a step in this new direction with a step
size (St) of 10 m subsequently the new position is
calculated A dispersing animal is allowed to take
1000 steps during dispersal If during dispersal in
habitat an animal meets non habitat it has a proba-
bility to be reflected Ph of 95 (Table 1) Reflecting
means that the walking direction is mirrored against
the habitat edge When a dispersing animal however
enter habitat the animal continues its path Animals
continue dispersing until they find a new territory that
is not occupied by a territorial adult of the same sex
If an animal has not found another suitable patch after
Table 2 Habitat cover
( of total) of different
landscapes
Landscape
description
Habitat large
patches
Habitat small
patches
Habitat linear
elements
Matrix
Large 10 0 0 90
Mix-P 5 5 0 90
Mix-L 5 0 5 90
Mixed-LP 5 25 25 90
Small-P 0 10 0 90
Small-L 0 0 10 90
Small-LP 0 5 5 90
1126 Landscape Ecol (2009) 241123ndash1133
123
1000 steps and is not located in habitat the dispersing
animal dies Animals that arrive in habitat after 1000
steps but not in a vacant territory are allocated to the
nearest territory Animals that arrive in a new
territory become floaters and may reproduce when
the territorial adult of the same sex dies Animals that
encounter the landscape edge are reflected These
rules cause that dispersing animals first disperse in
the habitat patch of their territory checking neighbor
territories before they start dispersing through the
matrix and might find another habitat patch and
territories in there
Parameterization and calibration
We use demographic parameters of the middle
spotted woodpecker (Pettersson 1985 Kosenko and
Kaigorodova 2001 Kosinski and Ksit 2006) whereas
the dispersal distance is based on the work of Muller
(1982 Table 1) Demographic parameters were cal-
ibrated to meet criteria published by Verboom et al
(2001) for medium sized birds
Evaluating synergy
Synergy occurs when mixtures of large and small
habitat patches perform better with respect to survival
than landscapes with patches of the same type In this
study we did 250 simulations in the default land-
scapes each run starting with one reproductive unit
ie an adult male and female in every territory We
evaluated metapopulation survival and the mean
number of adult individuals between year 50 and
150 to avoid transient behavior due to the initiali-
zation state (lsquolsquoburn-inrsquorsquo) Since there is a strong
correlation between the mean number of individuals
and survival we present only the survival results in
our graphs Of every survival fraction the 95
confidence interval is calculated using a v-test We
present our results as a replacement series in which
we replace large patches by smaller elements keeping
the same amount of habitat ultimately resulting in the
landscape with small elements only (Fig 3) Monot-
onously increasing relations (small- [ mixed- [large) support the more traditional hypothesis that
Large
Large
Large
Small-P Mix-P
Small-L
Small-PL
Mix-L
Mix-PL
only small mixed only largereplacing
Small toLarge
Small andLinear toLarge
Linear tolarge
Fig 2 Default landscapes
of 6 9 6 km lines are
linear elements of
6000 9 25 m smallsquares are small forest
patches of 25 ha that can
contain one territory Largesquares are large patches of
10 ha that contain four
territories
Landscape Ecol (2009) 241123ndash1133 1127
123
larger patches are always better an optimum rela-
tionship supports our alternative hypothesis that
synergy occurs
Simulations and results
Reference simulations
We evaluate metapopulation survival in the seven
default landscapes for the species parameterized
according to Table 1 When we replace half of the
large patches by small elements these mixed land-
scapes performe best if these small elements were all
linear elements (L Fig 4) Here metapopulation
survival in mixed landscapes was higher than in the
landscape with only large patches indicating syn-
ergy If half of the large patches is replaced by a mix
of small patches and linear elements (LP) metapop-
ulation survival is the same as in the landscapes with
only large patches This indicates that mixed land-
scapes with linear elements perform equally or better
than the landscape with only large patches Mixed
landscapes without linear elements (P) or landscapes
with only small elements however always performe
worse compared to the other landscapes Furthermore
we see that a landscape with only small elements that
contained both linear elements and small patches
(LP) performed much better than landscapes with
only small elements that contained only linear
elements (L) or only small patches (P) These results
suggest that replacing a part of the small patches by
linear elements also enhances population survival
Effects of physical connections
The results suggest that linear elements are respon-
sible for the synergy due to the fact that they connect
patches with each other and guide dispersers To
investigate this effect we create three alternative
landscapes in which the linear elements are not
connected to the patches
If linear elements are not connected to the large
patches number of individuals and survival decrease
in the mixed landscapes with linear elements (L and
LP) (compare Figs 4 5) This indicates that linear
connections are responsible for the synergy found in
the reference conditions Survival in landschapes
Large
Mix-L
Mix-LP
Mix-P
Small-L
Small-LP
Small-P
S
urvi
val o
f met
apop
ulat
ion
Only Mix Onlysmall largeelements patches
L-line
P-line
LP-line
Fig 3 Explanatory graph that relates individual simulation
results to synergy If mixed landscapes perform better than
landscapes with only small and large elements there is synergy
L linear elements present as small elements P small patches
present as small elements LP both linear elements and small
patches present as small elements
0
02
04
06
08
1
only small mix only large
surv
ivin
g m
etap
opul
atio
ns
L
P
LP
Fig 4 Fraction of surviving metapopulations of default
simulations of seven different landscapes that differ with
respect to the percentage of large and small habitat patches or
linear habitat elements error bars represent the 95 confi-
dence inteval Only small landscapes that only consist of small
elements mix landscape that are a mix of large and small
elements only large is a landscape that has only large patches
L linear elements present as small elements P small patches
present as small elements LP both linear elements and small
patches present as small elements
1128 Landscape Ecol (2009) 241123ndash1133
123
with only small elements however is still higher in
landscapes with small patches and linear elements
(LP) than in landscapes with only small patches
Effects of decreasing the dispersal through
the matrix
To investigate whether linear elements are indeed
crucial for synergy we decrease the dispersal effi-
ciency through the matrix by increasing the standard
deviation of the turning angle (SDD) from 5 to 20 and
40 This change results in a decrease of the realized
dispersal distance As a result 90 of the animals
move less than resp 29 and 15 km in the matrix
whereas in the reference situation this was 84 km In
these simulations we use the seven default landscapes
again
As a result of the decreased dispersal efficiency
through the matrix the metapopulation survival in
landscapes without linear elements (P) decrease
strongly (Fig 6a b) resulting in a clear synergy
pattern for the linear replacement lines (L) and
linear combined with small patches replacement
lines (LP Fig 6) Replacing large patches by
smaller patches does not help to compensate for
the loss of dispersal efficiency through the matrix
The landscapes containing only small patches the
decreased dispersal efficiency even caused extinc-
tion in a simualtion for the most inefficient dispersal
class These results indicate that if dispersal through
the matrix is difficult or avoided by a species linear
elements are crucial
Discussion
Main results
It is generally assumed that larger patches of natural
habitat are better for the survival of species than the
same amount of habitat in smaller fragments
(MacArthur and Wilson 1967 Verboom et al 2001
Vos et al 2001) Our results suggest that replacing a
part of the larger units by smaller linear elements
enhance population performance with respect to
population level and survival Without any large
patches however we see a serious decrease in
performance of the population indicating that large
patches are still very important The explanation for
this synergy is that the large patches secure survival
of the population whereas the linear units enhance
inter-patch dispersal by serving as corridors Addi-
tionally we see that populations in landscapes with
small patches also benefit from the availability of
linear elements Also here we see synergy to the
extent that a mixture of linear elements and small
patches performs better than landscapes consisting of
solely linear elements or small patches
In this study linear elements have a double
function they can serve as a corridor but also as
habitat In real landscapes corridors may not be wide
or qualitatively good enough to support both func-
tions Small patches may serve as stepping stones
because the mean inter-patch distance decreases with
increasing number of patches (Etienne and Heester-
beek 2000) However if we reduce the area of the
patches with a certain factor the average inter-patch
distance (centre to centre) is reduced with only the
square root of this factor Consequently replacing
large patches by smaller patches is inferior to
replacing large patches by linear elements
0
02
04
06
08
1
only small mix only large
surv
ing
met
apop
ulat
ions
L
LP
Fig 5 Fraction of surviving metapopulations of simulations
in which the linear elements were not connected to the large
patches of five different landscapes that differ with respect to
the percentage of large and small habitat patches or linear
habitat elements error bars represent the 95 confidence
inteval Only small landscapes that only consist of small
elements mix landscape that are a mix of large and small
elements only large is a landscape that has only large patches
L linear elements present as small elements LP both linear
elements and small patches present as small elements
Landscape Ecol (2009) 241123ndash1133 1129
123
Mechanisms favoring large or small patches
Seven processes are put forward in literature that
determines metapopulation performance in response
to the subdivision of habitat
1 Population extinction larger patches have a
higher carrying capacity and therefore a lower
extinction risk due to Allee-effects and demo-
graphic stochasticity than smaller patches
(Shaffer 1981 Soule 1987)
2 Colonization When dispersal potential is weak
and the dispersal mortality in the matrix is high
however the lsquoSeveral Smallrsquo approach might be
superior (Etienne and Heesterbeek 2000) The
extreme case is when the inter-patch distances of
the large patches are too large for recolonization
whereas the species is able to bridge the smaller
inter-patch distance between the small patches
Here the landscape configuration with few large
patches is actually more fragmented than the
landscape with many small patches (Flather and
Bevers 2002)
3 Finding a mate it seems likely that especially
monogamous animals that live in a landscape
with many small patches have a higher risk to
stay single This lowers the reproduction (Engen
et al 2003)
4 Density dependent population growth in a more
fragmented landscape animals encounter more
density dependent effects because some patches
are at or above carrying capacity while others are
empty Here individuals are not optimally utiliz-
ing the resources This lowers the growth rate of
fragmented populations
5 Patch edge effects if a species utilizes patch
edges more than the patch core several small
patches might be superior over a few large
patches because the amount of edges and
ecotones is larger when the landscape is more
fragmented On the other hand if core habitat is
preferred eg because the influence of the
Fig 6 The effect of increasing the turning angle of dispersing
birds from 5 to 20 and 40 degrees resulting in lower matrix
permeability The figure depict the fraction of surviving
metapopulations in the default landscape and the error bars
represent the 95 confidence interval Only small landscapes
that only consist of small elements mix landscape that are a
mix of large and small elements only large is a landscape that
has only large patches L linear elements present as small
elements P small patches present as small elements LP both
linear elements and small patches present as small elements
c
40 degrees
0
02
04
06
08
1
surv
ivin
g m
etap
opul
atio
ns
L
P
LP
20 degrees
0
02
04
06
08
1
only small mix only large
only small mix only large
only small mix only large
surv
ivin
g m
etap
opul
atio
ns
L
P
LP
5 degrees
0
02
04
06
08
1
surv
ivin
g m
etap
opul
atio
ns
L
P
LP
1130 Landscape Ecol (2009) 241123ndash1133
123
matrix makes the edges suboptimal larger ele-
ments are better
6 Spreading of risk if populations fluctuate due to
local environmental factors subdivision may
save parts of the metapopulation from events
such as fire or other adverse conditions When
species like a predator or a disease are taken into
account a fragmented landscape may be superior
Here predator or disease might also suffers from
isolation leaving their prey or host untouched in
patches that are not yet colonized by the predator
or disease (Baz and GarciaBoyero 1996 Lang-
lois et al 2001 Tscharntke et al 2002) Another
special case is habitat heterogeneity among
patches which makes eg wet patches more
suitable in dry years and dry patches in wet years
(Boer 1981) Although the spreading of risk
might also occur in larger elements it generally
favors population subdivision
7 Round off effects in fragmented landscapes
specific patch carrying capacity might not be
whole even numbers For instance if we sub-
stitute a patch with a carrying capacity of ten by
four fragments with carrying capacities of 25
these patches together can only support eight
breeding pairs
In our simulation study we do not incorporate all
these seven processes We take extinction (due to
demographic stochasticity and Allee-effects) coloni-
zation (dispersal success) pair formation and density
dependent population growth into account We do not
incorporate patch edge effects (when habitat quality
differs between patch core and patch edge) spreading
of risk (local disturbances patch heterogeneity and
antagonistic species) and round off effects These
three missing factors have an evident effect while
round off effects will favor large patch landscapes
and spreading of risk will favor fragmented land-
scapes the effect of taking into account edge effects
will depend on a preference for patch core or edge
habitat but in either case a straightforward relation is
expected Therefore our results may be slightly
modulated by taking into account these aspects but
our conclusions should be robust since some factors
are in favor of small patches and others are in favor of
large patches an optimum configuration is expected
somewhere between the two extreme cases with only
large or only small patches
Climate change
In the face of climate change there might be a few
extra considerations the habitat of species in climatic
terms is no longer stationary so species should be
able to track their climatic window Consequently the
global survival of a species is no longer only
dependent on the survival locally but also on the
ability of species to move and follow the climate
suitability window (Tischendorf et al 2003 Travis
2003 Saether et al 2004 Thomas et al 2004) This
underpins the idea that under climate change condi-
tions heterogeneous landscapes with many linear
elements might be even more superior for species
survival because they both facilitate animal movement
en survival Whereas large patches are necessary to
buffer the population against increasingly stochastic
weather events and provide dispersers connectivity by
linear elements should allow populations to shift
Generality of the results
We did a series of simulations with a spatially
explicit individual-based population model simulat-
ing the population dynamics of a territorial woodland
birds in open agricultural landscapes Our results
indicate that replacing patches by linear elements that
can serve as habitat enhances metapopulation perfor-
mance Clearly we can not extrapolate these results to
conclusions for other species and spatial configura-
tions We think however that our conclusions are
likely to be valid for other species and conditions if
(1) the carrying capacity of the habitat is not affected
by the replacement (2) the dispersal trough habitat is
more efficient and less risky than dispersal through
matrix and (3) if spatial configurations are such that
patches are relatively isolated so that dispersal
limitation is an important factor
Implications for sustainable landscape design
and nature conservation
Since in many countries space is scarce there is an
increasing need for knowledge about how to optimize
the design of landscapes nature reserves and habitat
networks from a biodiversity conservation perspec-
tive (Lomolino 1994 Kingsland 2002 McCarthy
Landscape Ecol (2009) 241123ndash1133 1131
123
et al 2005 McCarthy et al 2006) Our results
indicate that landscape designers and nature conser-
vationists should reconsider the value of small linear
elements that often are considered as part of the
matrix and not always seek to create larger units but
may better create more variable landscapes with
mixtures of patch sizes and shapes This is especially
important when smaller units play a key role in
physically connecting large patches when dispersal
through the matrix is poor or dispersal distances are
smaller than inter-patch distances On the other hand
the smaller units should be large enough to have a
habitat function for the species considered ie
reproduction should be possible there
Acknowledgments This research was financed by the
program lsquolsquoVernieuwend Ruimtegebruikrsquorsquo of Habiforum and
co-financed by the Dutch Ministry of Agriculture Nature and
Food Quality
References
Baveco JM (2006) Small steps movement model manual
Version 10 Alterra-Wageningen UR Wageningen p 19
Baz A GarciaBoyero A (1996) The SLOSS dilemma a
butterfly case study Biodivers Conserv 5493ndash502 doi
101007BF00056393
Bender DJ Tischendorf L Fahrig L (2003) Using patch isolation
metrics to predict animal movement in binary landscapes
Landscape Ecol 1817ndash39 doi101023A10229372
26820
PJd Boer (1981) On the survival of populations in a hetero-
geneous and variable environment Oecologia 5039ndash53
doi101007BF00378792
Chardon JP Adriaensen F Matthysen E (2003) Incorporating
landscape elements into a connectivity measure a case
study for the Speckled wood butterfly (Pararge aegeria L)
Landscape Ecol 18561ndash573 doi101023A10260625
30600
Engen S Lande R Seather BE (2003) Demographic stochas-
ticity and allee effects in populations with two sexes
Ecology 842378ndash2386 doi10189002-0123
Etienne RS Heesterbeek JAP (2000) On optimal size and
number of reserves for metapopulation persistence
J Theor Biol 20333ndash50 doi101006jtbi19991060
Flather CH Bevers M (2002) Patchy reaction-diffusion
and population abundance the relative importance of
habitat amount and arrangement Am Nat 15940ndash56 doi
101086324120
Grashof-Bokdam CJ Chardon P Vos CC Foppen RPB
Wallisdevries MF VanDerVeen M Meeuwsen HAM
(2008) The synergistic effect of combining woodlands
and green veining for biodiversity Landscape Ecol doi
101007s10980-008-9274-2
Grashof-Bokdam CJ van Langevelde F (2005) Green veining
landscape determinants of biodiversity in European
agricultural landscapes Landscape Ecol 20417ndash439
doi101007s10980-004-5646-1
Haddad NM (1999) Corridor and distance effects on interpatch
movements a landscape experiment with butterflies Ecol
Appl 9612ndash622 doi1018901051-0761(1999)009[0612
CADEOI]20CO2
Kingsland SE (2002) Creating a science of nature reserve
design perspectives from history Environ Model Assess
761ndash69 doi101023A1015633830223
Kosenko SM Kaigorodova EY (2001) Effect of habitat frag-
mentation on distribution density and breeding
performance of the middle spotted woodpecker Dend-
rocopos medius (Alves Picidae) in Nerussa-Desna
Polesye Zool Zh 8071ndash78
Kosinski Z Ksit P (2006) Comparative reproductive biology of
middle spotted woodpeckers Dendrocopos medius and
great spotted woodpeckers D-major in a riverine forest
Bird Study 53237ndash246
Langlois JP Fahrig L Merriam G Artsob H (2001) Landscape
structure influences continental distribution of hantavirus
in deer mice Landscape Ecol 16255ndash266 doi101023
A1011148316537
Lomolino MV (1994) An evaluation of alternative strategies
for building networks of nature-reserves Biol Conserv
69243ndash249 doi1010160006-3207(94)90423-5
MacArthur RH Wilson EO (1967) The theory of island bio-
geography Princeton University Press Princeton
McCarthy MA Thompson CJ Possingham HP (2005) Theory
for designing nature reserves for single species Am Nat
165250ndash257 doi101086427297
McCarthy MA Thompson CJ Williams NSG (2006) Logic for
designing nature reserves for multiple species Am Nat
167717ndash727 doi101086503058
Muller W (1982) Die Besiedlung der Eichenwalder im Kanton
Zurich durch den Mittelspecht Dendrocopus medius Orn
Beob 79105ndash119
Ovaskainen O (2002) Long-term persistence of species and the
SLOSS problem J Theor Biol 218419ndash433
Patterson BD Atmar W (1986) Nested subsets and the structure
of insular mammalian faunas and archipelagoes Biol J
Linn Soc Lond 2865ndash82 doi101111j1095-83121986
tb01749x
Pettersson B (1985) Extinction of an isolated population of the
middle spotted woodpecker Dendrocopos-Medius (L) in
Sweden and its relation to general theories on extinction
Biol Conserv 32335ndash353 doi1010160006-3207(85)
90022-9
Ricketts TH (2001) The matrix matters Effective isolation in
fragmented landscapes Am Nat 15887ndash99 doi101086
320863
Saether BE Sutherland WJ Engen S (2004) Climate influences
on avian population dynamics In Moller AP Fielder W
Berthold P (eds) Birds and climate change pp 185ndash209
Schippers P Verboom J Knaapen JP vanApeldoorn RC
(1996) Dispersal and habitat connectivity in complex
heterogeneous landscapes an analysis with a GIS-based
random walk model Ecography 1997ndash106 doi101111
j1600-05871996tb00160x
Schultz CB Crone EE (2005) Patch size and connectivity
thresholds for butterfly habitat restoration Conserv Biol
19887ndash896 doi101111j1523-1739200500462x
1132 Landscape Ecol (2009) 241123ndash1133
123
Shaffer ML (1981) Minimum population sizes for species con-
servation Bioscience 31131ndash134 doi1023071308256
Soule M (1987) Viable populations for conservation
Cambridge University Press Cambridge
Tellerıa JL Baquero R Santos T (2003) Effects of forest
fragmentation on European birds implications of regional
differences in species richness J Biogeogr 30621ndash628
doi101046j1365-2699200300960x
Thomas CD Cameron A Green RE Bakkenes M Beaumont LJ
Collingham YC Erasmus BFN de Siqueira MF Grainger
A Hannah L Hughes L Huntley B van Jaarsveld AS
Midgley GF Miles L Ortega-Huerta MA Peterson AT
Phillips OL Williams SE (2004) Extinction risk from
climate change Nature 427145ndash148 doi101038nature
02121
Thomas CD Kunin WE (1999) The spatial structure of pop-
ulations J Anim Ecol 68647ndash657 doi101046j1365-
2656199900330x
Tischendorf L Bender DJ Fahrig L (2003) Evaluation of patch
isolation metrics in mosaic landscapes for specialist vs
generalist dispersers Landscape Ecol 1841ndash50 doi
101023A1022908109982
Tischendorf L Irmler U Hingst R (1998) A simulation
experiment on the potential of hedgerows as movement
corridors for forest carabids Ecol Modell 106107ndash118
doi101016S0304-3800(97)00186-5
Tischendorf L Wissel C (1997) Corridors as conduits for small
animals attainable distances depending on movement
pattern boundary reaction and corridor width Oikos
79603ndash611 doi1023073546904
Travis JMJ (2003) Climate change and habitat destruction a
deadly anthropogenic cocktail Proc R Soc Lond B Biol
Sci 270467ndash473 doi101098rspb20022246
Tscharntke T Steffan-Dewenter I Kruess A Thies C (2002)
Contribution of small habitat fragments to conservation of
insect communities of grassland-cropland landscapes
Ecol Appl 12354ndash363
Van Apeldoorn RC Knaapen JP Schippers P Verboom J
Engen van H Meeuwsen H (1998) Landscape planning
and conservation biology simulation models as tools to
evaluate scenarios for badger in the Netherlands Land-
scape Urban Plan 4157ndash69 doi101016S0169-2046(97)
00058-3
VanDorp D Schippers P Groenendael van JM (1997)
Migration rates of grassland plants along corridors in
fragmented landscapes assessed with a cellular automa-
tion model Landscape Ecol 1239ndash50 doi101007
BF02698206
Verboom J Foppen R Chardon P Opdam P Luttikhuizen P
(2001) Introducing the key patch approach for habitat
networks with persistent populations an example for
marshland birds Biol Conserv 10089ndash101 doi101016
S0006-3207(00)00210-X
Vos CC Verboom J Opdam PFM Ter Braak CJF (2001)
Toward ecologically scaled landscape indices Am Nat
15724ndash41 doi101086317004
Wiens JA (1995) Landscape mosaics and ecological theory In
Hansson L Fahrig L Merriam G (eds) Mosaic landscapes
and ecological processes Chapman and Hall London
pp 1ndash26
Landscape Ecol (2009) 241123ndash1133 1133
123

1000 steps and is not located in habitat the dispersing
animal dies Animals that arrive in habitat after 1000
steps but not in a vacant territory are allocated to the
nearest territory Animals that arrive in a new
territory become floaters and may reproduce when
the territorial adult of the same sex dies Animals that
encounter the landscape edge are reflected These
rules cause that dispersing animals first disperse in
the habitat patch of their territory checking neighbor
territories before they start dispersing through the
matrix and might find another habitat patch and
territories in there
Parameterization and calibration
We use demographic parameters of the middle
spotted woodpecker (Pettersson 1985 Kosenko and
Kaigorodova 2001 Kosinski and Ksit 2006) whereas
the dispersal distance is based on the work of Muller
(1982 Table 1) Demographic parameters were cal-
ibrated to meet criteria published by Verboom et al
(2001) for medium sized birds
Evaluating synergy
Synergy occurs when mixtures of large and small
habitat patches perform better with respect to survival
than landscapes with patches of the same type In this
study we did 250 simulations in the default land-
scapes each run starting with one reproductive unit
ie an adult male and female in every territory We
evaluated metapopulation survival and the mean
number of adult individuals between year 50 and
150 to avoid transient behavior due to the initiali-
zation state (lsquolsquoburn-inrsquorsquo) Since there is a strong
correlation between the mean number of individuals
and survival we present only the survival results in
our graphs Of every survival fraction the 95
confidence interval is calculated using a v-test We
present our results as a replacement series in which
we replace large patches by smaller elements keeping
the same amount of habitat ultimately resulting in the
landscape with small elements only (Fig 3) Monot-
onously increasing relations (small- [ mixed- [large) support the more traditional hypothesis that
Large
Large
Large
Small-P Mix-P
Small-L
Small-PL
Mix-L
Mix-PL
only small mixed only largereplacing
Small toLarge
Small andLinear toLarge
Linear tolarge
Fig 2 Default landscapes
of 6 9 6 km lines are
linear elements of
6000 9 25 m smallsquares are small forest
patches of 25 ha that can
contain one territory Largesquares are large patches of
10 ha that contain four
territories
Landscape Ecol (2009) 241123ndash1133 1127
123
larger patches are always better an optimum rela-
tionship supports our alternative hypothesis that
synergy occurs
Simulations and results
Reference simulations
We evaluate metapopulation survival in the seven
default landscapes for the species parameterized
according to Table 1 When we replace half of the
large patches by small elements these mixed land-
scapes performe best if these small elements were all
linear elements (L Fig 4) Here metapopulation
survival in mixed landscapes was higher than in the
landscape with only large patches indicating syn-
ergy If half of the large patches is replaced by a mix
of small patches and linear elements (LP) metapop-
ulation survival is the same as in the landscapes with
only large patches This indicates that mixed land-
scapes with linear elements perform equally or better
than the landscape with only large patches Mixed
landscapes without linear elements (P) or landscapes
with only small elements however always performe
worse compared to the other landscapes Furthermore
we see that a landscape with only small elements that
contained both linear elements and small patches
(LP) performed much better than landscapes with
only small elements that contained only linear
elements (L) or only small patches (P) These results
suggest that replacing a part of the small patches by
linear elements also enhances population survival
Effects of physical connections
The results suggest that linear elements are respon-
sible for the synergy due to the fact that they connect
patches with each other and guide dispersers To
investigate this effect we create three alternative
landscapes in which the linear elements are not
connected to the patches
If linear elements are not connected to the large
patches number of individuals and survival decrease
in the mixed landscapes with linear elements (L and
LP) (compare Figs 4 5) This indicates that linear
connections are responsible for the synergy found in
the reference conditions Survival in landschapes
Large
Mix-L
Mix-LP
Mix-P
Small-L
Small-LP
Small-P
S
urvi
val o
f met
apop
ulat
ion
Only Mix Onlysmall largeelements patches
L-line
P-line
LP-line
Fig 3 Explanatory graph that relates individual simulation
results to synergy If mixed landscapes perform better than
landscapes with only small and large elements there is synergy
L linear elements present as small elements P small patches
present as small elements LP both linear elements and small
patches present as small elements
0
02
04
06
08
1
only small mix only large
surv
ivin
g m
etap
opul
atio
ns
L
P
LP
Fig 4 Fraction of surviving metapopulations of default
simulations of seven different landscapes that differ with
respect to the percentage of large and small habitat patches or
linear habitat elements error bars represent the 95 confi-
dence inteval Only small landscapes that only consist of small
elements mix landscape that are a mix of large and small
elements only large is a landscape that has only large patches
L linear elements present as small elements P small patches
present as small elements LP both linear elements and small
patches present as small elements
1128 Landscape Ecol (2009) 241123ndash1133
123
with only small elements however is still higher in
landscapes with small patches and linear elements
(LP) than in landscapes with only small patches
Effects of decreasing the dispersal through
the matrix
To investigate whether linear elements are indeed
crucial for synergy we decrease the dispersal effi-
ciency through the matrix by increasing the standard
deviation of the turning angle (SDD) from 5 to 20 and
40 This change results in a decrease of the realized
dispersal distance As a result 90 of the animals
move less than resp 29 and 15 km in the matrix
whereas in the reference situation this was 84 km In
these simulations we use the seven default landscapes
again
As a result of the decreased dispersal efficiency
through the matrix the metapopulation survival in
landscapes without linear elements (P) decrease
strongly (Fig 6a b) resulting in a clear synergy
pattern for the linear replacement lines (L) and
linear combined with small patches replacement
lines (LP Fig 6) Replacing large patches by
smaller patches does not help to compensate for
the loss of dispersal efficiency through the matrix
The landscapes containing only small patches the
decreased dispersal efficiency even caused extinc-
tion in a simualtion for the most inefficient dispersal
class These results indicate that if dispersal through
the matrix is difficult or avoided by a species linear
elements are crucial
Discussion
Main results
It is generally assumed that larger patches of natural
habitat are better for the survival of species than the
same amount of habitat in smaller fragments
(MacArthur and Wilson 1967 Verboom et al 2001
Vos et al 2001) Our results suggest that replacing a
part of the larger units by smaller linear elements
enhance population performance with respect to
population level and survival Without any large
patches however we see a serious decrease in
performance of the population indicating that large
patches are still very important The explanation for
this synergy is that the large patches secure survival
of the population whereas the linear units enhance
inter-patch dispersal by serving as corridors Addi-
tionally we see that populations in landscapes with
small patches also benefit from the availability of
linear elements Also here we see synergy to the
extent that a mixture of linear elements and small
patches performs better than landscapes consisting of
solely linear elements or small patches
In this study linear elements have a double
function they can serve as a corridor but also as
habitat In real landscapes corridors may not be wide
or qualitatively good enough to support both func-
tions Small patches may serve as stepping stones
because the mean inter-patch distance decreases with
increasing number of patches (Etienne and Heester-
beek 2000) However if we reduce the area of the
patches with a certain factor the average inter-patch
distance (centre to centre) is reduced with only the
square root of this factor Consequently replacing
large patches by smaller patches is inferior to
replacing large patches by linear elements
0
02
04
06
08
1
only small mix only large
surv
ing
met
apop
ulat
ions
L
LP
Fig 5 Fraction of surviving metapopulations of simulations
in which the linear elements were not connected to the large
patches of five different landscapes that differ with respect to
the percentage of large and small habitat patches or linear
habitat elements error bars represent the 95 confidence
inteval Only small landscapes that only consist of small
elements mix landscape that are a mix of large and small
elements only large is a landscape that has only large patches
L linear elements present as small elements LP both linear
elements and small patches present as small elements
Landscape Ecol (2009) 241123ndash1133 1129
123
Mechanisms favoring large or small patches
Seven processes are put forward in literature that
determines metapopulation performance in response
to the subdivision of habitat
1 Population extinction larger patches have a
higher carrying capacity and therefore a lower
extinction risk due to Allee-effects and demo-
graphic stochasticity than smaller patches
(Shaffer 1981 Soule 1987)
2 Colonization When dispersal potential is weak
and the dispersal mortality in the matrix is high
however the lsquoSeveral Smallrsquo approach might be
superior (Etienne and Heesterbeek 2000) The
extreme case is when the inter-patch distances of
the large patches are too large for recolonization
whereas the species is able to bridge the smaller
inter-patch distance between the small patches
Here the landscape configuration with few large
patches is actually more fragmented than the
landscape with many small patches (Flather and
Bevers 2002)
3 Finding a mate it seems likely that especially
monogamous animals that live in a landscape
with many small patches have a higher risk to
stay single This lowers the reproduction (Engen
et al 2003)
4 Density dependent population growth in a more
fragmented landscape animals encounter more
density dependent effects because some patches
are at or above carrying capacity while others are
empty Here individuals are not optimally utiliz-
ing the resources This lowers the growth rate of
fragmented populations
5 Patch edge effects if a species utilizes patch
edges more than the patch core several small
patches might be superior over a few large
patches because the amount of edges and
ecotones is larger when the landscape is more
fragmented On the other hand if core habitat is
preferred eg because the influence of the
Fig 6 The effect of increasing the turning angle of dispersing
birds from 5 to 20 and 40 degrees resulting in lower matrix
permeability The figure depict the fraction of surviving
metapopulations in the default landscape and the error bars
represent the 95 confidence interval Only small landscapes
that only consist of small elements mix landscape that are a
mix of large and small elements only large is a landscape that
has only large patches L linear elements present as small
elements P small patches present as small elements LP both
linear elements and small patches present as small elements
c
40 degrees
0
02
04
06
08
1
surv
ivin
g m
etap
opul
atio
ns
L
P
LP
20 degrees
0
02
04
06
08
1
only small mix only large
only small mix only large
only small mix only large
surv
ivin
g m
etap
opul
atio
ns
L
P
LP
5 degrees
0
02
04
06
08
1
surv
ivin
g m
etap
opul
atio
ns
L
P
LP
1130 Landscape Ecol (2009) 241123ndash1133
123
matrix makes the edges suboptimal larger ele-
ments are better
6 Spreading of risk if populations fluctuate due to
local environmental factors subdivision may
save parts of the metapopulation from events
such as fire or other adverse conditions When
species like a predator or a disease are taken into
account a fragmented landscape may be superior
Here predator or disease might also suffers from
isolation leaving their prey or host untouched in
patches that are not yet colonized by the predator
or disease (Baz and GarciaBoyero 1996 Lang-
lois et al 2001 Tscharntke et al 2002) Another
special case is habitat heterogeneity among
patches which makes eg wet patches more
suitable in dry years and dry patches in wet years
(Boer 1981) Although the spreading of risk
might also occur in larger elements it generally
favors population subdivision
7 Round off effects in fragmented landscapes
specific patch carrying capacity might not be
whole even numbers For instance if we sub-
stitute a patch with a carrying capacity of ten by
four fragments with carrying capacities of 25
these patches together can only support eight
breeding pairs
In our simulation study we do not incorporate all
these seven processes We take extinction (due to
demographic stochasticity and Allee-effects) coloni-
zation (dispersal success) pair formation and density
dependent population growth into account We do not
incorporate patch edge effects (when habitat quality
differs between patch core and patch edge) spreading
of risk (local disturbances patch heterogeneity and
antagonistic species) and round off effects These
three missing factors have an evident effect while
round off effects will favor large patch landscapes
and spreading of risk will favor fragmented land-
scapes the effect of taking into account edge effects
will depend on a preference for patch core or edge
habitat but in either case a straightforward relation is
expected Therefore our results may be slightly
modulated by taking into account these aspects but
our conclusions should be robust since some factors
are in favor of small patches and others are in favor of
large patches an optimum configuration is expected
somewhere between the two extreme cases with only
large or only small patches
Climate change
In the face of climate change there might be a few
extra considerations the habitat of species in climatic
terms is no longer stationary so species should be
able to track their climatic window Consequently the
global survival of a species is no longer only
dependent on the survival locally but also on the
ability of species to move and follow the climate
suitability window (Tischendorf et al 2003 Travis
2003 Saether et al 2004 Thomas et al 2004) This
underpins the idea that under climate change condi-
tions heterogeneous landscapes with many linear
elements might be even more superior for species
survival because they both facilitate animal movement
en survival Whereas large patches are necessary to
buffer the population against increasingly stochastic
weather events and provide dispersers connectivity by
linear elements should allow populations to shift
Generality of the results
We did a series of simulations with a spatially
explicit individual-based population model simulat-
ing the population dynamics of a territorial woodland
birds in open agricultural landscapes Our results
indicate that replacing patches by linear elements that
can serve as habitat enhances metapopulation perfor-
mance Clearly we can not extrapolate these results to
conclusions for other species and spatial configura-
tions We think however that our conclusions are
likely to be valid for other species and conditions if
(1) the carrying capacity of the habitat is not affected
by the replacement (2) the dispersal trough habitat is
more efficient and less risky than dispersal through
matrix and (3) if spatial configurations are such that
patches are relatively isolated so that dispersal
limitation is an important factor
Implications for sustainable landscape design
and nature conservation
Since in many countries space is scarce there is an
increasing need for knowledge about how to optimize
the design of landscapes nature reserves and habitat
networks from a biodiversity conservation perspec-
tive (Lomolino 1994 Kingsland 2002 McCarthy
Landscape Ecol (2009) 241123ndash1133 1131
123
et al 2005 McCarthy et al 2006) Our results
indicate that landscape designers and nature conser-
vationists should reconsider the value of small linear
elements that often are considered as part of the
matrix and not always seek to create larger units but
may better create more variable landscapes with
mixtures of patch sizes and shapes This is especially
important when smaller units play a key role in
physically connecting large patches when dispersal
through the matrix is poor or dispersal distances are
smaller than inter-patch distances On the other hand
the smaller units should be large enough to have a
habitat function for the species considered ie
reproduction should be possible there
Acknowledgments This research was financed by the
program lsquolsquoVernieuwend Ruimtegebruikrsquorsquo of Habiforum and
co-financed by the Dutch Ministry of Agriculture Nature and
Food Quality
References
Baveco JM (2006) Small steps movement model manual
Version 10 Alterra-Wageningen UR Wageningen p 19
Baz A GarciaBoyero A (1996) The SLOSS dilemma a
butterfly case study Biodivers Conserv 5493ndash502 doi
101007BF00056393
Bender DJ Tischendorf L Fahrig L (2003) Using patch isolation
metrics to predict animal movement in binary landscapes
Landscape Ecol 1817ndash39 doi101023A10229372
26820
PJd Boer (1981) On the survival of populations in a hetero-
geneous and variable environment Oecologia 5039ndash53
doi101007BF00378792
Chardon JP Adriaensen F Matthysen E (2003) Incorporating
landscape elements into a connectivity measure a case
study for the Speckled wood butterfly (Pararge aegeria L)
Landscape Ecol 18561ndash573 doi101023A10260625
30600
Engen S Lande R Seather BE (2003) Demographic stochas-
ticity and allee effects in populations with two sexes
Ecology 842378ndash2386 doi10189002-0123
Etienne RS Heesterbeek JAP (2000) On optimal size and
number of reserves for metapopulation persistence
J Theor Biol 20333ndash50 doi101006jtbi19991060
Flather CH Bevers M (2002) Patchy reaction-diffusion
and population abundance the relative importance of
habitat amount and arrangement Am Nat 15940ndash56 doi
101086324120
Grashof-Bokdam CJ Chardon P Vos CC Foppen RPB
Wallisdevries MF VanDerVeen M Meeuwsen HAM
(2008) The synergistic effect of combining woodlands
and green veining for biodiversity Landscape Ecol doi
101007s10980-008-9274-2
Grashof-Bokdam CJ van Langevelde F (2005) Green veining
landscape determinants of biodiversity in European
agricultural landscapes Landscape Ecol 20417ndash439
doi101007s10980-004-5646-1
Haddad NM (1999) Corridor and distance effects on interpatch
movements a landscape experiment with butterflies Ecol
Appl 9612ndash622 doi1018901051-0761(1999)009[0612
CADEOI]20CO2
Kingsland SE (2002) Creating a science of nature reserve
design perspectives from history Environ Model Assess
761ndash69 doi101023A1015633830223
Kosenko SM Kaigorodova EY (2001) Effect of habitat frag-
mentation on distribution density and breeding
performance of the middle spotted woodpecker Dend-
rocopos medius (Alves Picidae) in Nerussa-Desna
Polesye Zool Zh 8071ndash78
Kosinski Z Ksit P (2006) Comparative reproductive biology of
middle spotted woodpeckers Dendrocopos medius and
great spotted woodpeckers D-major in a riverine forest
Bird Study 53237ndash246
Langlois JP Fahrig L Merriam G Artsob H (2001) Landscape
structure influences continental distribution of hantavirus
in deer mice Landscape Ecol 16255ndash266 doi101023
A1011148316537
Lomolino MV (1994) An evaluation of alternative strategies
for building networks of nature-reserves Biol Conserv
69243ndash249 doi1010160006-3207(94)90423-5
MacArthur RH Wilson EO (1967) The theory of island bio-
geography Princeton University Press Princeton
McCarthy MA Thompson CJ Possingham HP (2005) Theory
for designing nature reserves for single species Am Nat
165250ndash257 doi101086427297
McCarthy MA Thompson CJ Williams NSG (2006) Logic for
designing nature reserves for multiple species Am Nat
167717ndash727 doi101086503058
Muller W (1982) Die Besiedlung der Eichenwalder im Kanton
Zurich durch den Mittelspecht Dendrocopus medius Orn
Beob 79105ndash119
Ovaskainen O (2002) Long-term persistence of species and the
SLOSS problem J Theor Biol 218419ndash433
Patterson BD Atmar W (1986) Nested subsets and the structure
of insular mammalian faunas and archipelagoes Biol J
Linn Soc Lond 2865ndash82 doi101111j1095-83121986
tb01749x
Pettersson B (1985) Extinction of an isolated population of the
middle spotted woodpecker Dendrocopos-Medius (L) in
Sweden and its relation to general theories on extinction
Biol Conserv 32335ndash353 doi1010160006-3207(85)
90022-9
Ricketts TH (2001) The matrix matters Effective isolation in
fragmented landscapes Am Nat 15887ndash99 doi101086
320863
Saether BE Sutherland WJ Engen S (2004) Climate influences
on avian population dynamics In Moller AP Fielder W
Berthold P (eds) Birds and climate change pp 185ndash209
Schippers P Verboom J Knaapen JP vanApeldoorn RC
(1996) Dispersal and habitat connectivity in complex
heterogeneous landscapes an analysis with a GIS-based
random walk model Ecography 1997ndash106 doi101111
j1600-05871996tb00160x
Schultz CB Crone EE (2005) Patch size and connectivity
thresholds for butterfly habitat restoration Conserv Biol
19887ndash896 doi101111j1523-1739200500462x
1132 Landscape Ecol (2009) 241123ndash1133
123
Shaffer ML (1981) Minimum population sizes for species con-
servation Bioscience 31131ndash134 doi1023071308256
Soule M (1987) Viable populations for conservation
Cambridge University Press Cambridge
Tellerıa JL Baquero R Santos T (2003) Effects of forest
fragmentation on European birds implications of regional
differences in species richness J Biogeogr 30621ndash628
doi101046j1365-2699200300960x
Thomas CD Cameron A Green RE Bakkenes M Beaumont LJ
Collingham YC Erasmus BFN de Siqueira MF Grainger
A Hannah L Hughes L Huntley B van Jaarsveld AS
Midgley GF Miles L Ortega-Huerta MA Peterson AT
Phillips OL Williams SE (2004) Extinction risk from
climate change Nature 427145ndash148 doi101038nature
02121
Thomas CD Kunin WE (1999) The spatial structure of pop-
ulations J Anim Ecol 68647ndash657 doi101046j1365-
2656199900330x
Tischendorf L Bender DJ Fahrig L (2003) Evaluation of patch
isolation metrics in mosaic landscapes for specialist vs
generalist dispersers Landscape Ecol 1841ndash50 doi
101023A1022908109982
Tischendorf L Irmler U Hingst R (1998) A simulation
experiment on the potential of hedgerows as movement
corridors for forest carabids Ecol Modell 106107ndash118
doi101016S0304-3800(97)00186-5
Tischendorf L Wissel C (1997) Corridors as conduits for small
animals attainable distances depending on movement
pattern boundary reaction and corridor width Oikos
79603ndash611 doi1023073546904
Travis JMJ (2003) Climate change and habitat destruction a
deadly anthropogenic cocktail Proc R Soc Lond B Biol
Sci 270467ndash473 doi101098rspb20022246
Tscharntke T Steffan-Dewenter I Kruess A Thies C (2002)
Contribution of small habitat fragments to conservation of
insect communities of grassland-cropland landscapes
Ecol Appl 12354ndash363
Van Apeldoorn RC Knaapen JP Schippers P Verboom J
Engen van H Meeuwsen H (1998) Landscape planning
and conservation biology simulation models as tools to
evaluate scenarios for badger in the Netherlands Land-
scape Urban Plan 4157ndash69 doi101016S0169-2046(97)
00058-3
VanDorp D Schippers P Groenendael van JM (1997)
Migration rates of grassland plants along corridors in
fragmented landscapes assessed with a cellular automa-
tion model Landscape Ecol 1239ndash50 doi101007
BF02698206
Verboom J Foppen R Chardon P Opdam P Luttikhuizen P
(2001) Introducing the key patch approach for habitat
networks with persistent populations an example for
marshland birds Biol Conserv 10089ndash101 doi101016
S0006-3207(00)00210-X
Vos CC Verboom J Opdam PFM Ter Braak CJF (2001)
Toward ecologically scaled landscape indices Am Nat
15724ndash41 doi101086317004
Wiens JA (1995) Landscape mosaics and ecological theory In
Hansson L Fahrig L Merriam G (eds) Mosaic landscapes
and ecological processes Chapman and Hall London
pp 1ndash26
Landscape Ecol (2009) 241123ndash1133 1133
123
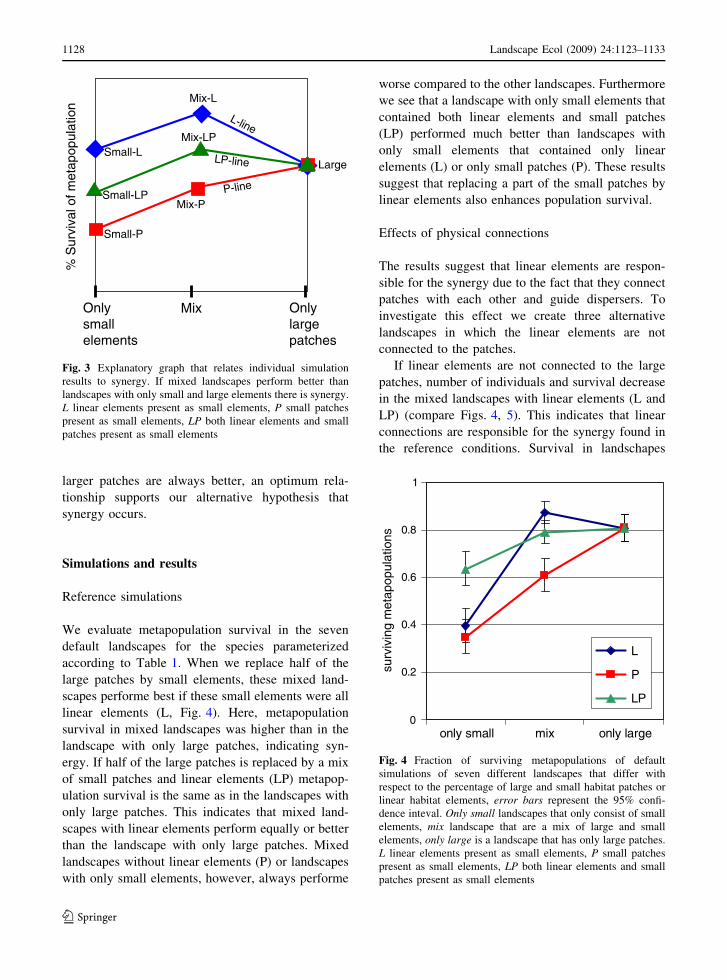
larger patches are always better an optimum rela-
tionship supports our alternative hypothesis that
synergy occurs
Simulations and results
Reference simulations
We evaluate metapopulation survival in the seven
default landscapes for the species parameterized
according to Table 1 When we replace half of the
large patches by small elements these mixed land-
scapes performe best if these small elements were all
linear elements (L Fig 4) Here metapopulation
survival in mixed landscapes was higher than in the
landscape with only large patches indicating syn-
ergy If half of the large patches is replaced by a mix
of small patches and linear elements (LP) metapop-
ulation survival is the same as in the landscapes with
only large patches This indicates that mixed land-
scapes with linear elements perform equally or better
than the landscape with only large patches Mixed
landscapes without linear elements (P) or landscapes
with only small elements however always performe
worse compared to the other landscapes Furthermore
we see that a landscape with only small elements that
contained both linear elements and small patches
(LP) performed much better than landscapes with
only small elements that contained only linear
elements (L) or only small patches (P) These results
suggest that replacing a part of the small patches by
linear elements also enhances population survival
Effects of physical connections
The results suggest that linear elements are respon-
sible for the synergy due to the fact that they connect
patches with each other and guide dispersers To
investigate this effect we create three alternative
landscapes in which the linear elements are not
connected to the patches
If linear elements are not connected to the large
patches number of individuals and survival decrease
in the mixed landscapes with linear elements (L and
LP) (compare Figs 4 5) This indicates that linear
connections are responsible for the synergy found in
the reference conditions Survival in landschapes
Large
Mix-L
Mix-LP
Mix-P
Small-L
Small-LP
Small-P
S
urvi
val o
f met
apop
ulat
ion
Only Mix Onlysmall largeelements patches
L-line
P-line
LP-line
Fig 3 Explanatory graph that relates individual simulation
results to synergy If mixed landscapes perform better than
landscapes with only small and large elements there is synergy
L linear elements present as small elements P small patches
present as small elements LP both linear elements and small
patches present as small elements
0
02
04
06
08
1
only small mix only large
surv
ivin
g m
etap
opul
atio
ns
L
P
LP
Fig 4 Fraction of surviving metapopulations of default
simulations of seven different landscapes that differ with
respect to the percentage of large and small habitat patches or
linear habitat elements error bars represent the 95 confi-
dence inteval Only small landscapes that only consist of small
elements mix landscape that are a mix of large and small
elements only large is a landscape that has only large patches
L linear elements present as small elements P small patches
present as small elements LP both linear elements and small
patches present as small elements
1128 Landscape Ecol (2009) 241123ndash1133
123
with only small elements however is still higher in
landscapes with small patches and linear elements
(LP) than in landscapes with only small patches
Effects of decreasing the dispersal through
the matrix
To investigate whether linear elements are indeed
crucial for synergy we decrease the dispersal effi-
ciency through the matrix by increasing the standard
deviation of the turning angle (SDD) from 5 to 20 and
40 This change results in a decrease of the realized
dispersal distance As a result 90 of the animals
move less than resp 29 and 15 km in the matrix
whereas in the reference situation this was 84 km In
these simulations we use the seven default landscapes
again
As a result of the decreased dispersal efficiency
through the matrix the metapopulation survival in
landscapes without linear elements (P) decrease
strongly (Fig 6a b) resulting in a clear synergy
pattern for the linear replacement lines (L) and
linear combined with small patches replacement
lines (LP Fig 6) Replacing large patches by
smaller patches does not help to compensate for
the loss of dispersal efficiency through the matrix
The landscapes containing only small patches the
decreased dispersal efficiency even caused extinc-
tion in a simualtion for the most inefficient dispersal
class These results indicate that if dispersal through
the matrix is difficult or avoided by a species linear
elements are crucial
Discussion
Main results
It is generally assumed that larger patches of natural
habitat are better for the survival of species than the
same amount of habitat in smaller fragments
(MacArthur and Wilson 1967 Verboom et al 2001
Vos et al 2001) Our results suggest that replacing a
part of the larger units by smaller linear elements
enhance population performance with respect to
population level and survival Without any large
patches however we see a serious decrease in
performance of the population indicating that large
patches are still very important The explanation for
this synergy is that the large patches secure survival
of the population whereas the linear units enhance
inter-patch dispersal by serving as corridors Addi-
tionally we see that populations in landscapes with
small patches also benefit from the availability of
linear elements Also here we see synergy to the
extent that a mixture of linear elements and small
patches performs better than landscapes consisting of
solely linear elements or small patches
In this study linear elements have a double
function they can serve as a corridor but also as
habitat In real landscapes corridors may not be wide
or qualitatively good enough to support both func-
tions Small patches may serve as stepping stones
because the mean inter-patch distance decreases with
increasing number of patches (Etienne and Heester-
beek 2000) However if we reduce the area of the
patches with a certain factor the average inter-patch
distance (centre to centre) is reduced with only the
square root of this factor Consequently replacing
large patches by smaller patches is inferior to
replacing large patches by linear elements
0
02
04
06
08
1
only small mix only large
surv
ing
met
apop
ulat
ions
L
LP
Fig 5 Fraction of surviving metapopulations of simulations
in which the linear elements were not connected to the large
patches of five different landscapes that differ with respect to
the percentage of large and small habitat patches or linear
habitat elements error bars represent the 95 confidence
inteval Only small landscapes that only consist of small
elements mix landscape that are a mix of large and small
elements only large is a landscape that has only large patches
L linear elements present as small elements LP both linear
elements and small patches present as small elements
Landscape Ecol (2009) 241123ndash1133 1129
123
Mechanisms favoring large or small patches
Seven processes are put forward in literature that
determines metapopulation performance in response
to the subdivision of habitat
1 Population extinction larger patches have a
higher carrying capacity and therefore a lower
extinction risk due to Allee-effects and demo-
graphic stochasticity than smaller patches
(Shaffer 1981 Soule 1987)
2 Colonization When dispersal potential is weak
and the dispersal mortality in the matrix is high
however the lsquoSeveral Smallrsquo approach might be
superior (Etienne and Heesterbeek 2000) The
extreme case is when the inter-patch distances of
the large patches are too large for recolonization
whereas the species is able to bridge the smaller
inter-patch distance between the small patches
Here the landscape configuration with few large
patches is actually more fragmented than the
landscape with many small patches (Flather and
Bevers 2002)
3 Finding a mate it seems likely that especially
monogamous animals that live in a landscape
with many small patches have a higher risk to
stay single This lowers the reproduction (Engen
et al 2003)
4 Density dependent population growth in a more
fragmented landscape animals encounter more
density dependent effects because some patches
are at or above carrying capacity while others are
empty Here individuals are not optimally utiliz-
ing the resources This lowers the growth rate of
fragmented populations
5 Patch edge effects if a species utilizes patch
edges more than the patch core several small
patches might be superior over a few large
patches because the amount of edges and
ecotones is larger when the landscape is more
fragmented On the other hand if core habitat is
preferred eg because the influence of the
Fig 6 The effect of increasing the turning angle of dispersing
birds from 5 to 20 and 40 degrees resulting in lower matrix
permeability The figure depict the fraction of surviving
metapopulations in the default landscape and the error bars
represent the 95 confidence interval Only small landscapes
that only consist of small elements mix landscape that are a
mix of large and small elements only large is a landscape that
has only large patches L linear elements present as small
elements P small patches present as small elements LP both
linear elements and small patches present as small elements
c
40 degrees
0
02
04
06
08
1
surv
ivin
g m
etap
opul
atio
ns
L
P
LP
20 degrees
0
02
04
06
08
1
only small mix only large
only small mix only large
only small mix only large
surv
ivin
g m
etap
opul
atio
ns
L
P
LP
5 degrees
0
02
04
06
08
1
surv
ivin
g m
etap
opul
atio
ns
L
P
LP
1130 Landscape Ecol (2009) 241123ndash1133
123
matrix makes the edges suboptimal larger ele-
ments are better
6 Spreading of risk if populations fluctuate due to
local environmental factors subdivision may
save parts of the metapopulation from events
such as fire or other adverse conditions When
species like a predator or a disease are taken into
account a fragmented landscape may be superior
Here predator or disease might also suffers from
isolation leaving their prey or host untouched in
patches that are not yet colonized by the predator
or disease (Baz and GarciaBoyero 1996 Lang-
lois et al 2001 Tscharntke et al 2002) Another
special case is habitat heterogeneity among
patches which makes eg wet patches more
suitable in dry years and dry patches in wet years
(Boer 1981) Although the spreading of risk
might also occur in larger elements it generally
favors population subdivision
7 Round off effects in fragmented landscapes
specific patch carrying capacity might not be
whole even numbers For instance if we sub-
stitute a patch with a carrying capacity of ten by
four fragments with carrying capacities of 25
these patches together can only support eight
breeding pairs
In our simulation study we do not incorporate all
these seven processes We take extinction (due to
demographic stochasticity and Allee-effects) coloni-
zation (dispersal success) pair formation and density
dependent population growth into account We do not
incorporate patch edge effects (when habitat quality
differs between patch core and patch edge) spreading
of risk (local disturbances patch heterogeneity and
antagonistic species) and round off effects These
three missing factors have an evident effect while
round off effects will favor large patch landscapes
and spreading of risk will favor fragmented land-
scapes the effect of taking into account edge effects
will depend on a preference for patch core or edge
habitat but in either case a straightforward relation is
expected Therefore our results may be slightly
modulated by taking into account these aspects but
our conclusions should be robust since some factors
are in favor of small patches and others are in favor of
large patches an optimum configuration is expected
somewhere between the two extreme cases with only
large or only small patches
Climate change
In the face of climate change there might be a few
extra considerations the habitat of species in climatic
terms is no longer stationary so species should be
able to track their climatic window Consequently the
global survival of a species is no longer only
dependent on the survival locally but also on the
ability of species to move and follow the climate
suitability window (Tischendorf et al 2003 Travis
2003 Saether et al 2004 Thomas et al 2004) This
underpins the idea that under climate change condi-
tions heterogeneous landscapes with many linear
elements might be even more superior for species
survival because they both facilitate animal movement
en survival Whereas large patches are necessary to
buffer the population against increasingly stochastic
weather events and provide dispersers connectivity by
linear elements should allow populations to shift
Generality of the results
We did a series of simulations with a spatially
explicit individual-based population model simulat-
ing the population dynamics of a territorial woodland
birds in open agricultural landscapes Our results
indicate that replacing patches by linear elements that
can serve as habitat enhances metapopulation perfor-
mance Clearly we can not extrapolate these results to
conclusions for other species and spatial configura-
tions We think however that our conclusions are
likely to be valid for other species and conditions if
(1) the carrying capacity of the habitat is not affected
by the replacement (2) the dispersal trough habitat is
more efficient and less risky than dispersal through
matrix and (3) if spatial configurations are such that
patches are relatively isolated so that dispersal
limitation is an important factor
Implications for sustainable landscape design
and nature conservation
Since in many countries space is scarce there is an
increasing need for knowledge about how to optimize
the design of landscapes nature reserves and habitat
networks from a biodiversity conservation perspec-
tive (Lomolino 1994 Kingsland 2002 McCarthy
Landscape Ecol (2009) 241123ndash1133 1131
123
et al 2005 McCarthy et al 2006) Our results
indicate that landscape designers and nature conser-
vationists should reconsider the value of small linear
elements that often are considered as part of the
matrix and not always seek to create larger units but
may better create more variable landscapes with
mixtures of patch sizes and shapes This is especially
important when smaller units play a key role in
physically connecting large patches when dispersal
through the matrix is poor or dispersal distances are
smaller than inter-patch distances On the other hand
the smaller units should be large enough to have a
habitat function for the species considered ie
reproduction should be possible there
Acknowledgments This research was financed by the
program lsquolsquoVernieuwend Ruimtegebruikrsquorsquo of Habiforum and
co-financed by the Dutch Ministry of Agriculture Nature and
Food Quality
References
Baveco JM (2006) Small steps movement model manual
Version 10 Alterra-Wageningen UR Wageningen p 19
Baz A GarciaBoyero A (1996) The SLOSS dilemma a
butterfly case study Biodivers Conserv 5493ndash502 doi
101007BF00056393
Bender DJ Tischendorf L Fahrig L (2003) Using patch isolation
metrics to predict animal movement in binary landscapes
Landscape Ecol 1817ndash39 doi101023A10229372
26820
PJd Boer (1981) On the survival of populations in a hetero-
geneous and variable environment Oecologia 5039ndash53
doi101007BF00378792
Chardon JP Adriaensen F Matthysen E (2003) Incorporating
landscape elements into a connectivity measure a case
study for the Speckled wood butterfly (Pararge aegeria L)
Landscape Ecol 18561ndash573 doi101023A10260625
30600
Engen S Lande R Seather BE (2003) Demographic stochas-
ticity and allee effects in populations with two sexes
Ecology 842378ndash2386 doi10189002-0123
Etienne RS Heesterbeek JAP (2000) On optimal size and
number of reserves for metapopulation persistence
J Theor Biol 20333ndash50 doi101006jtbi19991060
Flather CH Bevers M (2002) Patchy reaction-diffusion
and population abundance the relative importance of
habitat amount and arrangement Am Nat 15940ndash56 doi
101086324120
Grashof-Bokdam CJ Chardon P Vos CC Foppen RPB
Wallisdevries MF VanDerVeen M Meeuwsen HAM
(2008) The synergistic effect of combining woodlands
and green veining for biodiversity Landscape Ecol doi
101007s10980-008-9274-2
Grashof-Bokdam CJ van Langevelde F (2005) Green veining
landscape determinants of biodiversity in European
agricultural landscapes Landscape Ecol 20417ndash439
doi101007s10980-004-5646-1
Haddad NM (1999) Corridor and distance effects on interpatch
movements a landscape experiment with butterflies Ecol
Appl 9612ndash622 doi1018901051-0761(1999)009[0612
CADEOI]20CO2
Kingsland SE (2002) Creating a science of nature reserve
design perspectives from history Environ Model Assess
761ndash69 doi101023A1015633830223
Kosenko SM Kaigorodova EY (2001) Effect of habitat frag-
mentation on distribution density and breeding
performance of the middle spotted woodpecker Dend-
rocopos medius (Alves Picidae) in Nerussa-Desna
Polesye Zool Zh 8071ndash78
Kosinski Z Ksit P (2006) Comparative reproductive biology of
middle spotted woodpeckers Dendrocopos medius and
great spotted woodpeckers D-major in a riverine forest
Bird Study 53237ndash246
Langlois JP Fahrig L Merriam G Artsob H (2001) Landscape
structure influences continental distribution of hantavirus
in deer mice Landscape Ecol 16255ndash266 doi101023
A1011148316537
Lomolino MV (1994) An evaluation of alternative strategies
for building networks of nature-reserves Biol Conserv
69243ndash249 doi1010160006-3207(94)90423-5
MacArthur RH Wilson EO (1967) The theory of island bio-
geography Princeton University Press Princeton
McCarthy MA Thompson CJ Possingham HP (2005) Theory
for designing nature reserves for single species Am Nat
165250ndash257 doi101086427297
McCarthy MA Thompson CJ Williams NSG (2006) Logic for
designing nature reserves for multiple species Am Nat
167717ndash727 doi101086503058
Muller W (1982) Die Besiedlung der Eichenwalder im Kanton
Zurich durch den Mittelspecht Dendrocopus medius Orn
Beob 79105ndash119
Ovaskainen O (2002) Long-term persistence of species and the
SLOSS problem J Theor Biol 218419ndash433
Patterson BD Atmar W (1986) Nested subsets and the structure
of insular mammalian faunas and archipelagoes Biol J
Linn Soc Lond 2865ndash82 doi101111j1095-83121986
tb01749x
Pettersson B (1985) Extinction of an isolated population of the
middle spotted woodpecker Dendrocopos-Medius (L) in
Sweden and its relation to general theories on extinction
Biol Conserv 32335ndash353 doi1010160006-3207(85)
90022-9
Ricketts TH (2001) The matrix matters Effective isolation in
fragmented landscapes Am Nat 15887ndash99 doi101086
320863
Saether BE Sutherland WJ Engen S (2004) Climate influences
on avian population dynamics In Moller AP Fielder W
Berthold P (eds) Birds and climate change pp 185ndash209
Schippers P Verboom J Knaapen JP vanApeldoorn RC
(1996) Dispersal and habitat connectivity in complex
heterogeneous landscapes an analysis with a GIS-based
random walk model Ecography 1997ndash106 doi101111
j1600-05871996tb00160x
Schultz CB Crone EE (2005) Patch size and connectivity
thresholds for butterfly habitat restoration Conserv Biol
19887ndash896 doi101111j1523-1739200500462x
1132 Landscape Ecol (2009) 241123ndash1133
123
Shaffer ML (1981) Minimum population sizes for species con-
servation Bioscience 31131ndash134 doi1023071308256
Soule M (1987) Viable populations for conservation
Cambridge University Press Cambridge
Tellerıa JL Baquero R Santos T (2003) Effects of forest
fragmentation on European birds implications of regional
differences in species richness J Biogeogr 30621ndash628
doi101046j1365-2699200300960x
Thomas CD Cameron A Green RE Bakkenes M Beaumont LJ
Collingham YC Erasmus BFN de Siqueira MF Grainger
A Hannah L Hughes L Huntley B van Jaarsveld AS
Midgley GF Miles L Ortega-Huerta MA Peterson AT
Phillips OL Williams SE (2004) Extinction risk from
climate change Nature 427145ndash148 doi101038nature
02121
Thomas CD Kunin WE (1999) The spatial structure of pop-
ulations J Anim Ecol 68647ndash657 doi101046j1365-
2656199900330x
Tischendorf L Bender DJ Fahrig L (2003) Evaluation of patch
isolation metrics in mosaic landscapes for specialist vs
generalist dispersers Landscape Ecol 1841ndash50 doi
101023A1022908109982
Tischendorf L Irmler U Hingst R (1998) A simulation
experiment on the potential of hedgerows as movement
corridors for forest carabids Ecol Modell 106107ndash118
doi101016S0304-3800(97)00186-5
Tischendorf L Wissel C (1997) Corridors as conduits for small
animals attainable distances depending on movement
pattern boundary reaction and corridor width Oikos
79603ndash611 doi1023073546904
Travis JMJ (2003) Climate change and habitat destruction a
deadly anthropogenic cocktail Proc R Soc Lond B Biol
Sci 270467ndash473 doi101098rspb20022246
Tscharntke T Steffan-Dewenter I Kruess A Thies C (2002)
Contribution of small habitat fragments to conservation of
insect communities of grassland-cropland landscapes
Ecol Appl 12354ndash363
Van Apeldoorn RC Knaapen JP Schippers P Verboom J
Engen van H Meeuwsen H (1998) Landscape planning
and conservation biology simulation models as tools to
evaluate scenarios for badger in the Netherlands Land-
scape Urban Plan 4157ndash69 doi101016S0169-2046(97)
00058-3
VanDorp D Schippers P Groenendael van JM (1997)
Migration rates of grassland plants along corridors in
fragmented landscapes assessed with a cellular automa-
tion model Landscape Ecol 1239ndash50 doi101007
BF02698206
Verboom J Foppen R Chardon P Opdam P Luttikhuizen P
(2001) Introducing the key patch approach for habitat
networks with persistent populations an example for
marshland birds Biol Conserv 10089ndash101 doi101016
S0006-3207(00)00210-X
Vos CC Verboom J Opdam PFM Ter Braak CJF (2001)
Toward ecologically scaled landscape indices Am Nat
15724ndash41 doi101086317004
Wiens JA (1995) Landscape mosaics and ecological theory In
Hansson L Fahrig L Merriam G (eds) Mosaic landscapes
and ecological processes Chapman and Hall London
pp 1ndash26
Landscape Ecol (2009) 241123ndash1133 1133
123

with only small elements however is still higher in
landscapes with small patches and linear elements
(LP) than in landscapes with only small patches
Effects of decreasing the dispersal through
the matrix
To investigate whether linear elements are indeed
crucial for synergy we decrease the dispersal effi-
ciency through the matrix by increasing the standard
deviation of the turning angle (SDD) from 5 to 20 and
40 This change results in a decrease of the realized
dispersal distance As a result 90 of the animals
move less than resp 29 and 15 km in the matrix
whereas in the reference situation this was 84 km In
these simulations we use the seven default landscapes
again
As a result of the decreased dispersal efficiency
through the matrix the metapopulation survival in
landscapes without linear elements (P) decrease
strongly (Fig 6a b) resulting in a clear synergy
pattern for the linear replacement lines (L) and
linear combined with small patches replacement
lines (LP Fig 6) Replacing large patches by
smaller patches does not help to compensate for
the loss of dispersal efficiency through the matrix
The landscapes containing only small patches the
decreased dispersal efficiency even caused extinc-
tion in a simualtion for the most inefficient dispersal
class These results indicate that if dispersal through
the matrix is difficult or avoided by a species linear
elements are crucial
Discussion
Main results
It is generally assumed that larger patches of natural
habitat are better for the survival of species than the
same amount of habitat in smaller fragments
(MacArthur and Wilson 1967 Verboom et al 2001
Vos et al 2001) Our results suggest that replacing a
part of the larger units by smaller linear elements
enhance population performance with respect to
population level and survival Without any large
patches however we see a serious decrease in
performance of the population indicating that large
patches are still very important The explanation for
this synergy is that the large patches secure survival
of the population whereas the linear units enhance
inter-patch dispersal by serving as corridors Addi-
tionally we see that populations in landscapes with
small patches also benefit from the availability of
linear elements Also here we see synergy to the
extent that a mixture of linear elements and small
patches performs better than landscapes consisting of
solely linear elements or small patches
In this study linear elements have a double
function they can serve as a corridor but also as
habitat In real landscapes corridors may not be wide
or qualitatively good enough to support both func-
tions Small patches may serve as stepping stones
because the mean inter-patch distance decreases with
increasing number of patches (Etienne and Heester-
beek 2000) However if we reduce the area of the
patches with a certain factor the average inter-patch
distance (centre to centre) is reduced with only the
square root of this factor Consequently replacing
large patches by smaller patches is inferior to
replacing large patches by linear elements
0
02
04
06
08
1
only small mix only large
surv
ing
met
apop
ulat
ions
L
LP
Fig 5 Fraction of surviving metapopulations of simulations
in which the linear elements were not connected to the large
patches of five different landscapes that differ with respect to
the percentage of large and small habitat patches or linear
habitat elements error bars represent the 95 confidence
inteval Only small landscapes that only consist of small
elements mix landscape that are a mix of large and small
elements only large is a landscape that has only large patches
L linear elements present as small elements LP both linear
elements and small patches present as small elements
Landscape Ecol (2009) 241123ndash1133 1129
123
Mechanisms favoring large or small patches
Seven processes are put forward in literature that
determines metapopulation performance in response
to the subdivision of habitat
1 Population extinction larger patches have a
higher carrying capacity and therefore a lower
extinction risk due to Allee-effects and demo-
graphic stochasticity than smaller patches
(Shaffer 1981 Soule 1987)
2 Colonization When dispersal potential is weak
and the dispersal mortality in the matrix is high
however the lsquoSeveral Smallrsquo approach might be
superior (Etienne and Heesterbeek 2000) The
extreme case is when the inter-patch distances of
the large patches are too large for recolonization
whereas the species is able to bridge the smaller
inter-patch distance between the small patches
Here the landscape configuration with few large
patches is actually more fragmented than the
landscape with many small patches (Flather and
Bevers 2002)
3 Finding a mate it seems likely that especially
monogamous animals that live in a landscape
with many small patches have a higher risk to
stay single This lowers the reproduction (Engen
et al 2003)
4 Density dependent population growth in a more
fragmented landscape animals encounter more
density dependent effects because some patches
are at or above carrying capacity while others are
empty Here individuals are not optimally utiliz-
ing the resources This lowers the growth rate of
fragmented populations
5 Patch edge effects if a species utilizes patch
edges more than the patch core several small
patches might be superior over a few large
patches because the amount of edges and
ecotones is larger when the landscape is more
fragmented On the other hand if core habitat is
preferred eg because the influence of the
Fig 6 The effect of increasing the turning angle of dispersing
birds from 5 to 20 and 40 degrees resulting in lower matrix
permeability The figure depict the fraction of surviving
metapopulations in the default landscape and the error bars
represent the 95 confidence interval Only small landscapes
that only consist of small elements mix landscape that are a
mix of large and small elements only large is a landscape that
has only large patches L linear elements present as small
elements P small patches present as small elements LP both
linear elements and small patches present as small elements
c
40 degrees
0
02
04
06
08
1
surv
ivin
g m
etap
opul
atio
ns
L
P
LP
20 degrees
0
02
04
06
08
1
only small mix only large
only small mix only large
only small mix only large
surv
ivin
g m
etap
opul
atio
ns
L
P
LP
5 degrees
0
02
04
06
08
1
surv
ivin
g m
etap
opul
atio
ns
L
P
LP
1130 Landscape Ecol (2009) 241123ndash1133
123
matrix makes the edges suboptimal larger ele-
ments are better
6 Spreading of risk if populations fluctuate due to
local environmental factors subdivision may
save parts of the metapopulation from events
such as fire or other adverse conditions When
species like a predator or a disease are taken into
account a fragmented landscape may be superior
Here predator or disease might also suffers from
isolation leaving their prey or host untouched in
patches that are not yet colonized by the predator
or disease (Baz and GarciaBoyero 1996 Lang-
lois et al 2001 Tscharntke et al 2002) Another
special case is habitat heterogeneity among
patches which makes eg wet patches more
suitable in dry years and dry patches in wet years
(Boer 1981) Although the spreading of risk
might also occur in larger elements it generally
favors population subdivision
7 Round off effects in fragmented landscapes
specific patch carrying capacity might not be
whole even numbers For instance if we sub-
stitute a patch with a carrying capacity of ten by
four fragments with carrying capacities of 25
these patches together can only support eight
breeding pairs
In our simulation study we do not incorporate all
these seven processes We take extinction (due to
demographic stochasticity and Allee-effects) coloni-
zation (dispersal success) pair formation and density
dependent population growth into account We do not
incorporate patch edge effects (when habitat quality
differs between patch core and patch edge) spreading
of risk (local disturbances patch heterogeneity and
antagonistic species) and round off effects These
three missing factors have an evident effect while
round off effects will favor large patch landscapes
and spreading of risk will favor fragmented land-
scapes the effect of taking into account edge effects
will depend on a preference for patch core or edge
habitat but in either case a straightforward relation is
expected Therefore our results may be slightly
modulated by taking into account these aspects but
our conclusions should be robust since some factors
are in favor of small patches and others are in favor of
large patches an optimum configuration is expected
somewhere between the two extreme cases with only
large or only small patches
Climate change
In the face of climate change there might be a few
extra considerations the habitat of species in climatic
terms is no longer stationary so species should be
able to track their climatic window Consequently the
global survival of a species is no longer only
dependent on the survival locally but also on the
ability of species to move and follow the climate
suitability window (Tischendorf et al 2003 Travis
2003 Saether et al 2004 Thomas et al 2004) This
underpins the idea that under climate change condi-
tions heterogeneous landscapes with many linear
elements might be even more superior for species
survival because they both facilitate animal movement
en survival Whereas large patches are necessary to
buffer the population against increasingly stochastic
weather events and provide dispersers connectivity by
linear elements should allow populations to shift
Generality of the results
We did a series of simulations with a spatially
explicit individual-based population model simulat-
ing the population dynamics of a territorial woodland
birds in open agricultural landscapes Our results
indicate that replacing patches by linear elements that
can serve as habitat enhances metapopulation perfor-
mance Clearly we can not extrapolate these results to
conclusions for other species and spatial configura-
tions We think however that our conclusions are
likely to be valid for other species and conditions if
(1) the carrying capacity of the habitat is not affected
by the replacement (2) the dispersal trough habitat is
more efficient and less risky than dispersal through
matrix and (3) if spatial configurations are such that
patches are relatively isolated so that dispersal
limitation is an important factor
Implications for sustainable landscape design
and nature conservation
Since in many countries space is scarce there is an
increasing need for knowledge about how to optimize
the design of landscapes nature reserves and habitat
networks from a biodiversity conservation perspec-
tive (Lomolino 1994 Kingsland 2002 McCarthy
Landscape Ecol (2009) 241123ndash1133 1131
123
et al 2005 McCarthy et al 2006) Our results
indicate that landscape designers and nature conser-
vationists should reconsider the value of small linear
elements that often are considered as part of the
matrix and not always seek to create larger units but
may better create more variable landscapes with
mixtures of patch sizes and shapes This is especially
important when smaller units play a key role in
physically connecting large patches when dispersal
through the matrix is poor or dispersal distances are
smaller than inter-patch distances On the other hand
the smaller units should be large enough to have a
habitat function for the species considered ie
reproduction should be possible there
Acknowledgments This research was financed by the
program lsquolsquoVernieuwend Ruimtegebruikrsquorsquo of Habiforum and
co-financed by the Dutch Ministry of Agriculture Nature and
Food Quality
References
Baveco JM (2006) Small steps movement model manual
Version 10 Alterra-Wageningen UR Wageningen p 19
Baz A GarciaBoyero A (1996) The SLOSS dilemma a
butterfly case study Biodivers Conserv 5493ndash502 doi
101007BF00056393
Bender DJ Tischendorf L Fahrig L (2003) Using patch isolation
metrics to predict animal movement in binary landscapes
Landscape Ecol 1817ndash39 doi101023A10229372
26820
PJd Boer (1981) On the survival of populations in a hetero-
geneous and variable environment Oecologia 5039ndash53
doi101007BF00378792
Chardon JP Adriaensen F Matthysen E (2003) Incorporating
landscape elements into a connectivity measure a case
study for the Speckled wood butterfly (Pararge aegeria L)
Landscape Ecol 18561ndash573 doi101023A10260625
30600
Engen S Lande R Seather BE (2003) Demographic stochas-
ticity and allee effects in populations with two sexes
Ecology 842378ndash2386 doi10189002-0123
Etienne RS Heesterbeek JAP (2000) On optimal size and
number of reserves for metapopulation persistence
J Theor Biol 20333ndash50 doi101006jtbi19991060
Flather CH Bevers M (2002) Patchy reaction-diffusion
and population abundance the relative importance of
habitat amount and arrangement Am Nat 15940ndash56 doi
101086324120
Grashof-Bokdam CJ Chardon P Vos CC Foppen RPB
Wallisdevries MF VanDerVeen M Meeuwsen HAM
(2008) The synergistic effect of combining woodlands
and green veining for biodiversity Landscape Ecol doi
101007s10980-008-9274-2
Grashof-Bokdam CJ van Langevelde F (2005) Green veining
landscape determinants of biodiversity in European
agricultural landscapes Landscape Ecol 20417ndash439
doi101007s10980-004-5646-1
Haddad NM (1999) Corridor and distance effects on interpatch
movements a landscape experiment with butterflies Ecol
Appl 9612ndash622 doi1018901051-0761(1999)009[0612
CADEOI]20CO2
Kingsland SE (2002) Creating a science of nature reserve
design perspectives from history Environ Model Assess
761ndash69 doi101023A1015633830223
Kosenko SM Kaigorodova EY (2001) Effect of habitat frag-
mentation on distribution density and breeding
performance of the middle spotted woodpecker Dend-
rocopos medius (Alves Picidae) in Nerussa-Desna
Polesye Zool Zh 8071ndash78
Kosinski Z Ksit P (2006) Comparative reproductive biology of
middle spotted woodpeckers Dendrocopos medius and
great spotted woodpeckers D-major in a riverine forest
Bird Study 53237ndash246
Langlois JP Fahrig L Merriam G Artsob H (2001) Landscape
structure influences continental distribution of hantavirus
in deer mice Landscape Ecol 16255ndash266 doi101023
A1011148316537
Lomolino MV (1994) An evaluation of alternative strategies
for building networks of nature-reserves Biol Conserv
69243ndash249 doi1010160006-3207(94)90423-5
MacArthur RH Wilson EO (1967) The theory of island bio-
geography Princeton University Press Princeton
McCarthy MA Thompson CJ Possingham HP (2005) Theory
for designing nature reserves for single species Am Nat
165250ndash257 doi101086427297
McCarthy MA Thompson CJ Williams NSG (2006) Logic for
designing nature reserves for multiple species Am Nat
167717ndash727 doi101086503058
Muller W (1982) Die Besiedlung der Eichenwalder im Kanton
Zurich durch den Mittelspecht Dendrocopus medius Orn
Beob 79105ndash119
Ovaskainen O (2002) Long-term persistence of species and the
SLOSS problem J Theor Biol 218419ndash433
Patterson BD Atmar W (1986) Nested subsets and the structure
of insular mammalian faunas and archipelagoes Biol J
Linn Soc Lond 2865ndash82 doi101111j1095-83121986
tb01749x
Pettersson B (1985) Extinction of an isolated population of the
middle spotted woodpecker Dendrocopos-Medius (L) in
Sweden and its relation to general theories on extinction
Biol Conserv 32335ndash353 doi1010160006-3207(85)
90022-9
Ricketts TH (2001) The matrix matters Effective isolation in
fragmented landscapes Am Nat 15887ndash99 doi101086
320863
Saether BE Sutherland WJ Engen S (2004) Climate influences
on avian population dynamics In Moller AP Fielder W
Berthold P (eds) Birds and climate change pp 185ndash209
Schippers P Verboom J Knaapen JP vanApeldoorn RC
(1996) Dispersal and habitat connectivity in complex
heterogeneous landscapes an analysis with a GIS-based
random walk model Ecography 1997ndash106 doi101111
j1600-05871996tb00160x
Schultz CB Crone EE (2005) Patch size and connectivity
thresholds for butterfly habitat restoration Conserv Biol
19887ndash896 doi101111j1523-1739200500462x
1132 Landscape Ecol (2009) 241123ndash1133
123
Shaffer ML (1981) Minimum population sizes for species con-
servation Bioscience 31131ndash134 doi1023071308256
Soule M (1987) Viable populations for conservation
Cambridge University Press Cambridge
Tellerıa JL Baquero R Santos T (2003) Effects of forest
fragmentation on European birds implications of regional
differences in species richness J Biogeogr 30621ndash628
doi101046j1365-2699200300960x
Thomas CD Cameron A Green RE Bakkenes M Beaumont LJ
Collingham YC Erasmus BFN de Siqueira MF Grainger
A Hannah L Hughes L Huntley B van Jaarsveld AS
Midgley GF Miles L Ortega-Huerta MA Peterson AT
Phillips OL Williams SE (2004) Extinction risk from
climate change Nature 427145ndash148 doi101038nature
02121
Thomas CD Kunin WE (1999) The spatial structure of pop-
ulations J Anim Ecol 68647ndash657 doi101046j1365-
2656199900330x
Tischendorf L Bender DJ Fahrig L (2003) Evaluation of patch
isolation metrics in mosaic landscapes for specialist vs
generalist dispersers Landscape Ecol 1841ndash50 doi
101023A1022908109982
Tischendorf L Irmler U Hingst R (1998) A simulation
experiment on the potential of hedgerows as movement
corridors for forest carabids Ecol Modell 106107ndash118
doi101016S0304-3800(97)00186-5
Tischendorf L Wissel C (1997) Corridors as conduits for small
animals attainable distances depending on movement
pattern boundary reaction and corridor width Oikos
79603ndash611 doi1023073546904
Travis JMJ (2003) Climate change and habitat destruction a
deadly anthropogenic cocktail Proc R Soc Lond B Biol
Sci 270467ndash473 doi101098rspb20022246
Tscharntke T Steffan-Dewenter I Kruess A Thies C (2002)
Contribution of small habitat fragments to conservation of
insect communities of grassland-cropland landscapes
Ecol Appl 12354ndash363
Van Apeldoorn RC Knaapen JP Schippers P Verboom J
Engen van H Meeuwsen H (1998) Landscape planning
and conservation biology simulation models as tools to
evaluate scenarios for badger in the Netherlands Land-
scape Urban Plan 4157ndash69 doi101016S0169-2046(97)
00058-3
VanDorp D Schippers P Groenendael van JM (1997)
Migration rates of grassland plants along corridors in
fragmented landscapes assessed with a cellular automa-
tion model Landscape Ecol 1239ndash50 doi101007
BF02698206
Verboom J Foppen R Chardon P Opdam P Luttikhuizen P
(2001) Introducing the key patch approach for habitat
networks with persistent populations an example for
marshland birds Biol Conserv 10089ndash101 doi101016
S0006-3207(00)00210-X
Vos CC Verboom J Opdam PFM Ter Braak CJF (2001)
Toward ecologically scaled landscape indices Am Nat
15724ndash41 doi101086317004
Wiens JA (1995) Landscape mosaics and ecological theory In
Hansson L Fahrig L Merriam G (eds) Mosaic landscapes
and ecological processes Chapman and Hall London
pp 1ndash26
Landscape Ecol (2009) 241123ndash1133 1133
123

Mechanisms favoring large or small patches
Seven processes are put forward in literature that
determines metapopulation performance in response
to the subdivision of habitat
1 Population extinction larger patches have a
higher carrying capacity and therefore a lower
extinction risk due to Allee-effects and demo-
graphic stochasticity than smaller patches
(Shaffer 1981 Soule 1987)
2 Colonization When dispersal potential is weak
and the dispersal mortality in the matrix is high
however the lsquoSeveral Smallrsquo approach might be
superior (Etienne and Heesterbeek 2000) The
extreme case is when the inter-patch distances of
the large patches are too large for recolonization
whereas the species is able to bridge the smaller
inter-patch distance between the small patches
Here the landscape configuration with few large
patches is actually more fragmented than the
landscape with many small patches (Flather and
Bevers 2002)
3 Finding a mate it seems likely that especially
monogamous animals that live in a landscape
with many small patches have a higher risk to
stay single This lowers the reproduction (Engen
et al 2003)
4 Density dependent population growth in a more
fragmented landscape animals encounter more
density dependent effects because some patches
are at or above carrying capacity while others are
empty Here individuals are not optimally utiliz-
ing the resources This lowers the growth rate of
fragmented populations
5 Patch edge effects if a species utilizes patch
edges more than the patch core several small
patches might be superior over a few large
patches because the amount of edges and
ecotones is larger when the landscape is more
fragmented On the other hand if core habitat is
preferred eg because the influence of the
Fig 6 The effect of increasing the turning angle of dispersing
birds from 5 to 20 and 40 degrees resulting in lower matrix
permeability The figure depict the fraction of surviving
metapopulations in the default landscape and the error bars
represent the 95 confidence interval Only small landscapes
that only consist of small elements mix landscape that are a
mix of large and small elements only large is a landscape that
has only large patches L linear elements present as small
elements P small patches present as small elements LP both
linear elements and small patches present as small elements
c
40 degrees
0
02
04
06
08
1
surv
ivin
g m
etap
opul
atio
ns
L
P
LP
20 degrees
0
02
04
06
08
1
only small mix only large
only small mix only large
only small mix only large
surv
ivin
g m
etap
opul
atio
ns
L
P
LP
5 degrees
0
02
04
06
08
1
surv
ivin
g m
etap
opul
atio
ns
L
P
LP
1130 Landscape Ecol (2009) 241123ndash1133
123
matrix makes the edges suboptimal larger ele-
ments are better
6 Spreading of risk if populations fluctuate due to
local environmental factors subdivision may
save parts of the metapopulation from events
such as fire or other adverse conditions When
species like a predator or a disease are taken into
account a fragmented landscape may be superior
Here predator or disease might also suffers from
isolation leaving their prey or host untouched in
patches that are not yet colonized by the predator
or disease (Baz and GarciaBoyero 1996 Lang-
lois et al 2001 Tscharntke et al 2002) Another
special case is habitat heterogeneity among
patches which makes eg wet patches more
suitable in dry years and dry patches in wet years
(Boer 1981) Although the spreading of risk
might also occur in larger elements it generally
favors population subdivision
7 Round off effects in fragmented landscapes
specific patch carrying capacity might not be
whole even numbers For instance if we sub-
stitute a patch with a carrying capacity of ten by
four fragments with carrying capacities of 25
these patches together can only support eight
breeding pairs
In our simulation study we do not incorporate all
these seven processes We take extinction (due to
demographic stochasticity and Allee-effects) coloni-
zation (dispersal success) pair formation and density
dependent population growth into account We do not
incorporate patch edge effects (when habitat quality
differs between patch core and patch edge) spreading
of risk (local disturbances patch heterogeneity and
antagonistic species) and round off effects These
three missing factors have an evident effect while
round off effects will favor large patch landscapes
and spreading of risk will favor fragmented land-
scapes the effect of taking into account edge effects
will depend on a preference for patch core or edge
habitat but in either case a straightforward relation is
expected Therefore our results may be slightly
modulated by taking into account these aspects but
our conclusions should be robust since some factors
are in favor of small patches and others are in favor of
large patches an optimum configuration is expected
somewhere between the two extreme cases with only
large or only small patches
Climate change
In the face of climate change there might be a few
extra considerations the habitat of species in climatic
terms is no longer stationary so species should be
able to track their climatic window Consequently the
global survival of a species is no longer only
dependent on the survival locally but also on the
ability of species to move and follow the climate
suitability window (Tischendorf et al 2003 Travis
2003 Saether et al 2004 Thomas et al 2004) This
underpins the idea that under climate change condi-
tions heterogeneous landscapes with many linear
elements might be even more superior for species
survival because they both facilitate animal movement
en survival Whereas large patches are necessary to
buffer the population against increasingly stochastic
weather events and provide dispersers connectivity by
linear elements should allow populations to shift
Generality of the results
We did a series of simulations with a spatially
explicit individual-based population model simulat-
ing the population dynamics of a territorial woodland
birds in open agricultural landscapes Our results
indicate that replacing patches by linear elements that
can serve as habitat enhances metapopulation perfor-
mance Clearly we can not extrapolate these results to
conclusions for other species and spatial configura-
tions We think however that our conclusions are
likely to be valid for other species and conditions if
(1) the carrying capacity of the habitat is not affected
by the replacement (2) the dispersal trough habitat is
more efficient and less risky than dispersal through
matrix and (3) if spatial configurations are such that
patches are relatively isolated so that dispersal
limitation is an important factor
Implications for sustainable landscape design
and nature conservation
Since in many countries space is scarce there is an
increasing need for knowledge about how to optimize
the design of landscapes nature reserves and habitat
networks from a biodiversity conservation perspec-
tive (Lomolino 1994 Kingsland 2002 McCarthy
Landscape Ecol (2009) 241123ndash1133 1131
123
et al 2005 McCarthy et al 2006) Our results
indicate that landscape designers and nature conser-
vationists should reconsider the value of small linear
elements that often are considered as part of the
matrix and not always seek to create larger units but
may better create more variable landscapes with
mixtures of patch sizes and shapes This is especially
important when smaller units play a key role in
physically connecting large patches when dispersal
through the matrix is poor or dispersal distances are
smaller than inter-patch distances On the other hand
the smaller units should be large enough to have a
habitat function for the species considered ie
reproduction should be possible there
Acknowledgments This research was financed by the
program lsquolsquoVernieuwend Ruimtegebruikrsquorsquo of Habiforum and
co-financed by the Dutch Ministry of Agriculture Nature and
Food Quality
References
Baveco JM (2006) Small steps movement model manual
Version 10 Alterra-Wageningen UR Wageningen p 19
Baz A GarciaBoyero A (1996) The SLOSS dilemma a
butterfly case study Biodivers Conserv 5493ndash502 doi
101007BF00056393
Bender DJ Tischendorf L Fahrig L (2003) Using patch isolation
metrics to predict animal movement in binary landscapes
Landscape Ecol 1817ndash39 doi101023A10229372
26820
PJd Boer (1981) On the survival of populations in a hetero-
geneous and variable environment Oecologia 5039ndash53
doi101007BF00378792
Chardon JP Adriaensen F Matthysen E (2003) Incorporating
landscape elements into a connectivity measure a case
study for the Speckled wood butterfly (Pararge aegeria L)
Landscape Ecol 18561ndash573 doi101023A10260625
30600
Engen S Lande R Seather BE (2003) Demographic stochas-
ticity and allee effects in populations with two sexes
Ecology 842378ndash2386 doi10189002-0123
Etienne RS Heesterbeek JAP (2000) On optimal size and
number of reserves for metapopulation persistence
J Theor Biol 20333ndash50 doi101006jtbi19991060
Flather CH Bevers M (2002) Patchy reaction-diffusion
and population abundance the relative importance of
habitat amount and arrangement Am Nat 15940ndash56 doi
101086324120
Grashof-Bokdam CJ Chardon P Vos CC Foppen RPB
Wallisdevries MF VanDerVeen M Meeuwsen HAM
(2008) The synergistic effect of combining woodlands
and green veining for biodiversity Landscape Ecol doi
101007s10980-008-9274-2
Grashof-Bokdam CJ van Langevelde F (2005) Green veining
landscape determinants of biodiversity in European
agricultural landscapes Landscape Ecol 20417ndash439
doi101007s10980-004-5646-1
Haddad NM (1999) Corridor and distance effects on interpatch
movements a landscape experiment with butterflies Ecol
Appl 9612ndash622 doi1018901051-0761(1999)009[0612
CADEOI]20CO2
Kingsland SE (2002) Creating a science of nature reserve
design perspectives from history Environ Model Assess
761ndash69 doi101023A1015633830223
Kosenko SM Kaigorodova EY (2001) Effect of habitat frag-
mentation on distribution density and breeding
performance of the middle spotted woodpecker Dend-
rocopos medius (Alves Picidae) in Nerussa-Desna
Polesye Zool Zh 8071ndash78
Kosinski Z Ksit P (2006) Comparative reproductive biology of
middle spotted woodpeckers Dendrocopos medius and
great spotted woodpeckers D-major in a riverine forest
Bird Study 53237ndash246
Langlois JP Fahrig L Merriam G Artsob H (2001) Landscape
structure influences continental distribution of hantavirus
in deer mice Landscape Ecol 16255ndash266 doi101023
A1011148316537
Lomolino MV (1994) An evaluation of alternative strategies
for building networks of nature-reserves Biol Conserv
69243ndash249 doi1010160006-3207(94)90423-5
MacArthur RH Wilson EO (1967) The theory of island bio-
geography Princeton University Press Princeton
McCarthy MA Thompson CJ Possingham HP (2005) Theory
for designing nature reserves for single species Am Nat
165250ndash257 doi101086427297
McCarthy MA Thompson CJ Williams NSG (2006) Logic for
designing nature reserves for multiple species Am Nat
167717ndash727 doi101086503058
Muller W (1982) Die Besiedlung der Eichenwalder im Kanton
Zurich durch den Mittelspecht Dendrocopus medius Orn
Beob 79105ndash119
Ovaskainen O (2002) Long-term persistence of species and the
SLOSS problem J Theor Biol 218419ndash433
Patterson BD Atmar W (1986) Nested subsets and the structure
of insular mammalian faunas and archipelagoes Biol J
Linn Soc Lond 2865ndash82 doi101111j1095-83121986
tb01749x
Pettersson B (1985) Extinction of an isolated population of the
middle spotted woodpecker Dendrocopos-Medius (L) in
Sweden and its relation to general theories on extinction
Biol Conserv 32335ndash353 doi1010160006-3207(85)
90022-9
Ricketts TH (2001) The matrix matters Effective isolation in
fragmented landscapes Am Nat 15887ndash99 doi101086
320863
Saether BE Sutherland WJ Engen S (2004) Climate influences
on avian population dynamics In Moller AP Fielder W
Berthold P (eds) Birds and climate change pp 185ndash209
Schippers P Verboom J Knaapen JP vanApeldoorn RC
(1996) Dispersal and habitat connectivity in complex
heterogeneous landscapes an analysis with a GIS-based
random walk model Ecography 1997ndash106 doi101111
j1600-05871996tb00160x
Schultz CB Crone EE (2005) Patch size and connectivity
thresholds for butterfly habitat restoration Conserv Biol
19887ndash896 doi101111j1523-1739200500462x
1132 Landscape Ecol (2009) 241123ndash1133
123
Shaffer ML (1981) Minimum population sizes for species con-
servation Bioscience 31131ndash134 doi1023071308256
Soule M (1987) Viable populations for conservation
Cambridge University Press Cambridge
Tellerıa JL Baquero R Santos T (2003) Effects of forest
fragmentation on European birds implications of regional
differences in species richness J Biogeogr 30621ndash628
doi101046j1365-2699200300960x
Thomas CD Cameron A Green RE Bakkenes M Beaumont LJ
Collingham YC Erasmus BFN de Siqueira MF Grainger
A Hannah L Hughes L Huntley B van Jaarsveld AS
Midgley GF Miles L Ortega-Huerta MA Peterson AT
Phillips OL Williams SE (2004) Extinction risk from
climate change Nature 427145ndash148 doi101038nature
02121
Thomas CD Kunin WE (1999) The spatial structure of pop-
ulations J Anim Ecol 68647ndash657 doi101046j1365-
2656199900330x
Tischendorf L Bender DJ Fahrig L (2003) Evaluation of patch
isolation metrics in mosaic landscapes for specialist vs
generalist dispersers Landscape Ecol 1841ndash50 doi
101023A1022908109982
Tischendorf L Irmler U Hingst R (1998) A simulation
experiment on the potential of hedgerows as movement
corridors for forest carabids Ecol Modell 106107ndash118
doi101016S0304-3800(97)00186-5
Tischendorf L Wissel C (1997) Corridors as conduits for small
animals attainable distances depending on movement
pattern boundary reaction and corridor width Oikos
79603ndash611 doi1023073546904
Travis JMJ (2003) Climate change and habitat destruction a
deadly anthropogenic cocktail Proc R Soc Lond B Biol
Sci 270467ndash473 doi101098rspb20022246
Tscharntke T Steffan-Dewenter I Kruess A Thies C (2002)
Contribution of small habitat fragments to conservation of
insect communities of grassland-cropland landscapes
Ecol Appl 12354ndash363
Van Apeldoorn RC Knaapen JP Schippers P Verboom J
Engen van H Meeuwsen H (1998) Landscape planning
and conservation biology simulation models as tools to
evaluate scenarios for badger in the Netherlands Land-
scape Urban Plan 4157ndash69 doi101016S0169-2046(97)
00058-3
VanDorp D Schippers P Groenendael van JM (1997)
Migration rates of grassland plants along corridors in
fragmented landscapes assessed with a cellular automa-
tion model Landscape Ecol 1239ndash50 doi101007
BF02698206
Verboom J Foppen R Chardon P Opdam P Luttikhuizen P
(2001) Introducing the key patch approach for habitat
networks with persistent populations an example for
marshland birds Biol Conserv 10089ndash101 doi101016
S0006-3207(00)00210-X
Vos CC Verboom J Opdam PFM Ter Braak CJF (2001)
Toward ecologically scaled landscape indices Am Nat
15724ndash41 doi101086317004
Wiens JA (1995) Landscape mosaics and ecological theory In
Hansson L Fahrig L Merriam G (eds) Mosaic landscapes
and ecological processes Chapman and Hall London
pp 1ndash26
Landscape Ecol (2009) 241123ndash1133 1133
123

matrix makes the edges suboptimal larger ele-
ments are better
6 Spreading of risk if populations fluctuate due to
local environmental factors subdivision may
save parts of the metapopulation from events
such as fire or other adverse conditions When
species like a predator or a disease are taken into
account a fragmented landscape may be superior
Here predator or disease might also suffers from
isolation leaving their prey or host untouched in
patches that are not yet colonized by the predator
or disease (Baz and GarciaBoyero 1996 Lang-
lois et al 2001 Tscharntke et al 2002) Another
special case is habitat heterogeneity among
patches which makes eg wet patches more
suitable in dry years and dry patches in wet years
(Boer 1981) Although the spreading of risk
might also occur in larger elements it generally
favors population subdivision
7 Round off effects in fragmented landscapes
specific patch carrying capacity might not be
whole even numbers For instance if we sub-
stitute a patch with a carrying capacity of ten by
four fragments with carrying capacities of 25
these patches together can only support eight
breeding pairs
In our simulation study we do not incorporate all
these seven processes We take extinction (due to
demographic stochasticity and Allee-effects) coloni-
zation (dispersal success) pair formation and density
dependent population growth into account We do not
incorporate patch edge effects (when habitat quality
differs between patch core and patch edge) spreading
of risk (local disturbances patch heterogeneity and
antagonistic species) and round off effects These
three missing factors have an evident effect while
round off effects will favor large patch landscapes
and spreading of risk will favor fragmented land-
scapes the effect of taking into account edge effects
will depend on a preference for patch core or edge
habitat but in either case a straightforward relation is
expected Therefore our results may be slightly
modulated by taking into account these aspects but
our conclusions should be robust since some factors
are in favor of small patches and others are in favor of
large patches an optimum configuration is expected
somewhere between the two extreme cases with only
large or only small patches
Climate change
In the face of climate change there might be a few
extra considerations the habitat of species in climatic
terms is no longer stationary so species should be
able to track their climatic window Consequently the
global survival of a species is no longer only
dependent on the survival locally but also on the
ability of species to move and follow the climate
suitability window (Tischendorf et al 2003 Travis
2003 Saether et al 2004 Thomas et al 2004) This
underpins the idea that under climate change condi-
tions heterogeneous landscapes with many linear
elements might be even more superior for species
survival because they both facilitate animal movement
en survival Whereas large patches are necessary to
buffer the population against increasingly stochastic
weather events and provide dispersers connectivity by
linear elements should allow populations to shift
Generality of the results
We did a series of simulations with a spatially
explicit individual-based population model simulat-
ing the population dynamics of a territorial woodland
birds in open agricultural landscapes Our results
indicate that replacing patches by linear elements that
can serve as habitat enhances metapopulation perfor-
mance Clearly we can not extrapolate these results to
conclusions for other species and spatial configura-
tions We think however that our conclusions are
likely to be valid for other species and conditions if
(1) the carrying capacity of the habitat is not affected
by the replacement (2) the dispersal trough habitat is
more efficient and less risky than dispersal through
matrix and (3) if spatial configurations are such that
patches are relatively isolated so that dispersal
limitation is an important factor
Implications for sustainable landscape design
and nature conservation
Since in many countries space is scarce there is an
increasing need for knowledge about how to optimize
the design of landscapes nature reserves and habitat
networks from a biodiversity conservation perspec-
tive (Lomolino 1994 Kingsland 2002 McCarthy
Landscape Ecol (2009) 241123ndash1133 1131
123
et al 2005 McCarthy et al 2006) Our results
indicate that landscape designers and nature conser-
vationists should reconsider the value of small linear
elements that often are considered as part of the
matrix and not always seek to create larger units but
may better create more variable landscapes with
mixtures of patch sizes and shapes This is especially
important when smaller units play a key role in
physically connecting large patches when dispersal
through the matrix is poor or dispersal distances are
smaller than inter-patch distances On the other hand
the smaller units should be large enough to have a
habitat function for the species considered ie
reproduction should be possible there
Acknowledgments This research was financed by the
program lsquolsquoVernieuwend Ruimtegebruikrsquorsquo of Habiforum and
co-financed by the Dutch Ministry of Agriculture Nature and
Food Quality
References
Baveco JM (2006) Small steps movement model manual
Version 10 Alterra-Wageningen UR Wageningen p 19
Baz A GarciaBoyero A (1996) The SLOSS dilemma a
butterfly case study Biodivers Conserv 5493ndash502 doi
101007BF00056393
Bender DJ Tischendorf L Fahrig L (2003) Using patch isolation
metrics to predict animal movement in binary landscapes
Landscape Ecol 1817ndash39 doi101023A10229372
26820
PJd Boer (1981) On the survival of populations in a hetero-
geneous and variable environment Oecologia 5039ndash53
doi101007BF00378792
Chardon JP Adriaensen F Matthysen E (2003) Incorporating
landscape elements into a connectivity measure a case
study for the Speckled wood butterfly (Pararge aegeria L)
Landscape Ecol 18561ndash573 doi101023A10260625
30600
Engen S Lande R Seather BE (2003) Demographic stochas-
ticity and allee effects in populations with two sexes
Ecology 842378ndash2386 doi10189002-0123
Etienne RS Heesterbeek JAP (2000) On optimal size and
number of reserves for metapopulation persistence
J Theor Biol 20333ndash50 doi101006jtbi19991060
Flather CH Bevers M (2002) Patchy reaction-diffusion
and population abundance the relative importance of
habitat amount and arrangement Am Nat 15940ndash56 doi
101086324120
Grashof-Bokdam CJ Chardon P Vos CC Foppen RPB
Wallisdevries MF VanDerVeen M Meeuwsen HAM
(2008) The synergistic effect of combining woodlands
and green veining for biodiversity Landscape Ecol doi
101007s10980-008-9274-2
Grashof-Bokdam CJ van Langevelde F (2005) Green veining
landscape determinants of biodiversity in European
agricultural landscapes Landscape Ecol 20417ndash439
doi101007s10980-004-5646-1
Haddad NM (1999) Corridor and distance effects on interpatch
movements a landscape experiment with butterflies Ecol
Appl 9612ndash622 doi1018901051-0761(1999)009[0612
CADEOI]20CO2
Kingsland SE (2002) Creating a science of nature reserve
design perspectives from history Environ Model Assess
761ndash69 doi101023A1015633830223
Kosenko SM Kaigorodova EY (2001) Effect of habitat frag-
mentation on distribution density and breeding
performance of the middle spotted woodpecker Dend-
rocopos medius (Alves Picidae) in Nerussa-Desna
Polesye Zool Zh 8071ndash78
Kosinski Z Ksit P (2006) Comparative reproductive biology of
middle spotted woodpeckers Dendrocopos medius and
great spotted woodpeckers D-major in a riverine forest
Bird Study 53237ndash246
Langlois JP Fahrig L Merriam G Artsob H (2001) Landscape
structure influences continental distribution of hantavirus
in deer mice Landscape Ecol 16255ndash266 doi101023
A1011148316537
Lomolino MV (1994) An evaluation of alternative strategies
for building networks of nature-reserves Biol Conserv
69243ndash249 doi1010160006-3207(94)90423-5
MacArthur RH Wilson EO (1967) The theory of island bio-
geography Princeton University Press Princeton
McCarthy MA Thompson CJ Possingham HP (2005) Theory
for designing nature reserves for single species Am Nat
165250ndash257 doi101086427297
McCarthy MA Thompson CJ Williams NSG (2006) Logic for
designing nature reserves for multiple species Am Nat
167717ndash727 doi101086503058
Muller W (1982) Die Besiedlung der Eichenwalder im Kanton
Zurich durch den Mittelspecht Dendrocopus medius Orn
Beob 79105ndash119
Ovaskainen O (2002) Long-term persistence of species and the
SLOSS problem J Theor Biol 218419ndash433
Patterson BD Atmar W (1986) Nested subsets and the structure
of insular mammalian faunas and archipelagoes Biol J
Linn Soc Lond 2865ndash82 doi101111j1095-83121986
tb01749x
Pettersson B (1985) Extinction of an isolated population of the
middle spotted woodpecker Dendrocopos-Medius (L) in
Sweden and its relation to general theories on extinction
Biol Conserv 32335ndash353 doi1010160006-3207(85)
90022-9
Ricketts TH (2001) The matrix matters Effective isolation in
fragmented landscapes Am Nat 15887ndash99 doi101086
320863
Saether BE Sutherland WJ Engen S (2004) Climate influences
on avian population dynamics In Moller AP Fielder W
Berthold P (eds) Birds and climate change pp 185ndash209
Schippers P Verboom J Knaapen JP vanApeldoorn RC
(1996) Dispersal and habitat connectivity in complex
heterogeneous landscapes an analysis with a GIS-based
random walk model Ecography 1997ndash106 doi101111
j1600-05871996tb00160x
Schultz CB Crone EE (2005) Patch size and connectivity
thresholds for butterfly habitat restoration Conserv Biol
19887ndash896 doi101111j1523-1739200500462x
1132 Landscape Ecol (2009) 241123ndash1133
123
Shaffer ML (1981) Minimum population sizes for species con-
servation Bioscience 31131ndash134 doi1023071308256
Soule M (1987) Viable populations for conservation
Cambridge University Press Cambridge
Tellerıa JL Baquero R Santos T (2003) Effects of forest
fragmentation on European birds implications of regional
differences in species richness J Biogeogr 30621ndash628
doi101046j1365-2699200300960x
Thomas CD Cameron A Green RE Bakkenes M Beaumont LJ
Collingham YC Erasmus BFN de Siqueira MF Grainger
A Hannah L Hughes L Huntley B van Jaarsveld AS
Midgley GF Miles L Ortega-Huerta MA Peterson AT
Phillips OL Williams SE (2004) Extinction risk from
climate change Nature 427145ndash148 doi101038nature
02121
Thomas CD Kunin WE (1999) The spatial structure of pop-
ulations J Anim Ecol 68647ndash657 doi101046j1365-
2656199900330x
Tischendorf L Bender DJ Fahrig L (2003) Evaluation of patch
isolation metrics in mosaic landscapes for specialist vs
generalist dispersers Landscape Ecol 1841ndash50 doi
101023A1022908109982
Tischendorf L Irmler U Hingst R (1998) A simulation
experiment on the potential of hedgerows as movement
corridors for forest carabids Ecol Modell 106107ndash118
doi101016S0304-3800(97)00186-5
Tischendorf L Wissel C (1997) Corridors as conduits for small
animals attainable distances depending on movement
pattern boundary reaction and corridor width Oikos
79603ndash611 doi1023073546904
Travis JMJ (2003) Climate change and habitat destruction a
deadly anthropogenic cocktail Proc R Soc Lond B Biol
Sci 270467ndash473 doi101098rspb20022246
Tscharntke T Steffan-Dewenter I Kruess A Thies C (2002)
Contribution of small habitat fragments to conservation of
insect communities of grassland-cropland landscapes
Ecol Appl 12354ndash363
Van Apeldoorn RC Knaapen JP Schippers P Verboom J
Engen van H Meeuwsen H (1998) Landscape planning
and conservation biology simulation models as tools to
evaluate scenarios for badger in the Netherlands Land-
scape Urban Plan 4157ndash69 doi101016S0169-2046(97)
00058-3
VanDorp D Schippers P Groenendael van JM (1997)
Migration rates of grassland plants along corridors in
fragmented landscapes assessed with a cellular automa-
tion model Landscape Ecol 1239ndash50 doi101007
BF02698206
Verboom J Foppen R Chardon P Opdam P Luttikhuizen P
(2001) Introducing the key patch approach for habitat
networks with persistent populations an example for
marshland birds Biol Conserv 10089ndash101 doi101016
S0006-3207(00)00210-X
Vos CC Verboom J Opdam PFM Ter Braak CJF (2001)
Toward ecologically scaled landscape indices Am Nat
15724ndash41 doi101086317004
Wiens JA (1995) Landscape mosaics and ecological theory In
Hansson L Fahrig L Merriam G (eds) Mosaic landscapes
and ecological processes Chapman and Hall London
pp 1ndash26
Landscape Ecol (2009) 241123ndash1133 1133
123

et al 2005 McCarthy et al 2006) Our results
indicate that landscape designers and nature conser-
vationists should reconsider the value of small linear
elements that often are considered as part of the
matrix and not always seek to create larger units but
may better create more variable landscapes with
mixtures of patch sizes and shapes This is especially
important when smaller units play a key role in
physically connecting large patches when dispersal
through the matrix is poor or dispersal distances are
smaller than inter-patch distances On the other hand
the smaller units should be large enough to have a
habitat function for the species considered ie
reproduction should be possible there
Acknowledgments This research was financed by the
program lsquolsquoVernieuwend Ruimtegebruikrsquorsquo of Habiforum and
co-financed by the Dutch Ministry of Agriculture Nature and
Food Quality
References
Baveco JM (2006) Small steps movement model manual
Version 10 Alterra-Wageningen UR Wageningen p 19
Baz A GarciaBoyero A (1996) The SLOSS dilemma a
butterfly case study Biodivers Conserv 5493ndash502 doi
101007BF00056393
Bender DJ Tischendorf L Fahrig L (2003) Using patch isolation
metrics to predict animal movement in binary landscapes
Landscape Ecol 1817ndash39 doi101023A10229372
26820
PJd Boer (1981) On the survival of populations in a hetero-
geneous and variable environment Oecologia 5039ndash53
doi101007BF00378792
Chardon JP Adriaensen F Matthysen E (2003) Incorporating
landscape elements into a connectivity measure a case
study for the Speckled wood butterfly (Pararge aegeria L)
Landscape Ecol 18561ndash573 doi101023A10260625
30600
Engen S Lande R Seather BE (2003) Demographic stochas-
ticity and allee effects in populations with two sexes
Ecology 842378ndash2386 doi10189002-0123
Etienne RS Heesterbeek JAP (2000) On optimal size and
number of reserves for metapopulation persistence
J Theor Biol 20333ndash50 doi101006jtbi19991060
Flather CH Bevers M (2002) Patchy reaction-diffusion
and population abundance the relative importance of
habitat amount and arrangement Am Nat 15940ndash56 doi
101086324120
Grashof-Bokdam CJ Chardon P Vos CC Foppen RPB
Wallisdevries MF VanDerVeen M Meeuwsen HAM
(2008) The synergistic effect of combining woodlands
and green veining for biodiversity Landscape Ecol doi
101007s10980-008-9274-2
Grashof-Bokdam CJ van Langevelde F (2005) Green veining
landscape determinants of biodiversity in European
agricultural landscapes Landscape Ecol 20417ndash439
doi101007s10980-004-5646-1
Haddad NM (1999) Corridor and distance effects on interpatch
movements a landscape experiment with butterflies Ecol
Appl 9612ndash622 doi1018901051-0761(1999)009[0612
CADEOI]20CO2
Kingsland SE (2002) Creating a science of nature reserve
design perspectives from history Environ Model Assess
761ndash69 doi101023A1015633830223
Kosenko SM Kaigorodova EY (2001) Effect of habitat frag-
mentation on distribution density and breeding
performance of the middle spotted woodpecker Dend-
rocopos medius (Alves Picidae) in Nerussa-Desna
Polesye Zool Zh 8071ndash78
Kosinski Z Ksit P (2006) Comparative reproductive biology of
middle spotted woodpeckers Dendrocopos medius and
great spotted woodpeckers D-major in a riverine forest
Bird Study 53237ndash246
Langlois JP Fahrig L Merriam G Artsob H (2001) Landscape
structure influences continental distribution of hantavirus
in deer mice Landscape Ecol 16255ndash266 doi101023
A1011148316537
Lomolino MV (1994) An evaluation of alternative strategies
for building networks of nature-reserves Biol Conserv
69243ndash249 doi1010160006-3207(94)90423-5
MacArthur RH Wilson EO (1967) The theory of island bio-
geography Princeton University Press Princeton
McCarthy MA Thompson CJ Possingham HP (2005) Theory
for designing nature reserves for single species Am Nat
165250ndash257 doi101086427297
McCarthy MA Thompson CJ Williams NSG (2006) Logic for
designing nature reserves for multiple species Am Nat
167717ndash727 doi101086503058
Muller W (1982) Die Besiedlung der Eichenwalder im Kanton
Zurich durch den Mittelspecht Dendrocopus medius Orn
Beob 79105ndash119
Ovaskainen O (2002) Long-term persistence of species and the
SLOSS problem J Theor Biol 218419ndash433
Patterson BD Atmar W (1986) Nested subsets and the structure
of insular mammalian faunas and archipelagoes Biol J
Linn Soc Lond 2865ndash82 doi101111j1095-83121986
tb01749x
Pettersson B (1985) Extinction of an isolated population of the
middle spotted woodpecker Dendrocopos-Medius (L) in
Sweden and its relation to general theories on extinction
Biol Conserv 32335ndash353 doi1010160006-3207(85)
90022-9
Ricketts TH (2001) The matrix matters Effective isolation in
fragmented landscapes Am Nat 15887ndash99 doi101086
320863
Saether BE Sutherland WJ Engen S (2004) Climate influences
on avian population dynamics In Moller AP Fielder W
Berthold P (eds) Birds and climate change pp 185ndash209
Schippers P Verboom J Knaapen JP vanApeldoorn RC
(1996) Dispersal and habitat connectivity in complex
heterogeneous landscapes an analysis with a GIS-based
random walk model Ecography 1997ndash106 doi101111
j1600-05871996tb00160x
Schultz CB Crone EE (2005) Patch size and connectivity
thresholds for butterfly habitat restoration Conserv Biol
19887ndash896 doi101111j1523-1739200500462x
1132 Landscape Ecol (2009) 241123ndash1133
123
Shaffer ML (1981) Minimum population sizes for species con-
servation Bioscience 31131ndash134 doi1023071308256
Soule M (1987) Viable populations for conservation
Cambridge University Press Cambridge
Tellerıa JL Baquero R Santos T (2003) Effects of forest
fragmentation on European birds implications of regional
differences in species richness J Biogeogr 30621ndash628
doi101046j1365-2699200300960x
Thomas CD Cameron A Green RE Bakkenes M Beaumont LJ
Collingham YC Erasmus BFN de Siqueira MF Grainger
A Hannah L Hughes L Huntley B van Jaarsveld AS
Midgley GF Miles L Ortega-Huerta MA Peterson AT
Phillips OL Williams SE (2004) Extinction risk from
climate change Nature 427145ndash148 doi101038nature
02121
Thomas CD Kunin WE (1999) The spatial structure of pop-
ulations J Anim Ecol 68647ndash657 doi101046j1365-
2656199900330x
Tischendorf L Bender DJ Fahrig L (2003) Evaluation of patch
isolation metrics in mosaic landscapes for specialist vs
generalist dispersers Landscape Ecol 1841ndash50 doi
101023A1022908109982
Tischendorf L Irmler U Hingst R (1998) A simulation
experiment on the potential of hedgerows as movement
corridors for forest carabids Ecol Modell 106107ndash118
doi101016S0304-3800(97)00186-5
Tischendorf L Wissel C (1997) Corridors as conduits for small
animals attainable distances depending on movement
pattern boundary reaction and corridor width Oikos
79603ndash611 doi1023073546904
Travis JMJ (2003) Climate change and habitat destruction a
deadly anthropogenic cocktail Proc R Soc Lond B Biol
Sci 270467ndash473 doi101098rspb20022246
Tscharntke T Steffan-Dewenter I Kruess A Thies C (2002)
Contribution of small habitat fragments to conservation of
insect communities of grassland-cropland landscapes
Ecol Appl 12354ndash363
Van Apeldoorn RC Knaapen JP Schippers P Verboom J
Engen van H Meeuwsen H (1998) Landscape planning
and conservation biology simulation models as tools to
evaluate scenarios for badger in the Netherlands Land-
scape Urban Plan 4157ndash69 doi101016S0169-2046(97)
00058-3
VanDorp D Schippers P Groenendael van JM (1997)
Migration rates of grassland plants along corridors in
fragmented landscapes assessed with a cellular automa-
tion model Landscape Ecol 1239ndash50 doi101007
BF02698206
Verboom J Foppen R Chardon P Opdam P Luttikhuizen P
(2001) Introducing the key patch approach for habitat
networks with persistent populations an example for
marshland birds Biol Conserv 10089ndash101 doi101016
S0006-3207(00)00210-X
Vos CC Verboom J Opdam PFM Ter Braak CJF (2001)
Toward ecologically scaled landscape indices Am Nat
15724ndash41 doi101086317004
Wiens JA (1995) Landscape mosaics and ecological theory In
Hansson L Fahrig L Merriam G (eds) Mosaic landscapes
and ecological processes Chapman and Hall London
pp 1ndash26
Landscape Ecol (2009) 241123ndash1133 1133
123

Shaffer ML (1981) Minimum population sizes for species con-
servation Bioscience 31131ndash134 doi1023071308256
Soule M (1987) Viable populations for conservation
Cambridge University Press Cambridge
Tellerıa JL Baquero R Santos T (2003) Effects of forest
fragmentation on European birds implications of regional
differences in species richness J Biogeogr 30621ndash628
doi101046j1365-2699200300960x
Thomas CD Cameron A Green RE Bakkenes M Beaumont LJ
Collingham YC Erasmus BFN de Siqueira MF Grainger
A Hannah L Hughes L Huntley B van Jaarsveld AS
Midgley GF Miles L Ortega-Huerta MA Peterson AT
Phillips OL Williams SE (2004) Extinction risk from
climate change Nature 427145ndash148 doi101038nature
02121
Thomas CD Kunin WE (1999) The spatial structure of pop-
ulations J Anim Ecol 68647ndash657 doi101046j1365-
2656199900330x
Tischendorf L Bender DJ Fahrig L (2003) Evaluation of patch
isolation metrics in mosaic landscapes for specialist vs
generalist dispersers Landscape Ecol 1841ndash50 doi
101023A1022908109982
Tischendorf L Irmler U Hingst R (1998) A simulation
experiment on the potential of hedgerows as movement
corridors for forest carabids Ecol Modell 106107ndash118
doi101016S0304-3800(97)00186-5
Tischendorf L Wissel C (1997) Corridors as conduits for small
animals attainable distances depending on movement
pattern boundary reaction and corridor width Oikos
79603ndash611 doi1023073546904
Travis JMJ (2003) Climate change and habitat destruction a
deadly anthropogenic cocktail Proc R Soc Lond B Biol
Sci 270467ndash473 doi101098rspb20022246
Tscharntke T Steffan-Dewenter I Kruess A Thies C (2002)
Contribution of small habitat fragments to conservation of
insect communities of grassland-cropland landscapes
Ecol Appl 12354ndash363
Van Apeldoorn RC Knaapen JP Schippers P Verboom J
Engen van H Meeuwsen H (1998) Landscape planning
and conservation biology simulation models as tools to
evaluate scenarios for badger in the Netherlands Land-
scape Urban Plan 4157ndash69 doi101016S0169-2046(97)
00058-3
VanDorp D Schippers P Groenendael van JM (1997)
Migration rates of grassland plants along corridors in
fragmented landscapes assessed with a cellular automa-
tion model Landscape Ecol 1239ndash50 doi101007
BF02698206
Verboom J Foppen R Chardon P Opdam P Luttikhuizen P
(2001) Introducing the key patch approach for habitat
networks with persistent populations an example for
marshland birds Biol Conserv 10089ndash101 doi101016
S0006-3207(00)00210-X
Vos CC Verboom J Opdam PFM Ter Braak CJF (2001)
Toward ecologically scaled landscape indices Am Nat
15724ndash41 doi101086317004
Wiens JA (1995) Landscape mosaics and ecological theory In
Hansson L Fahrig L Merriam G (eds) Mosaic landscapes
and ecological processes Chapman and Hall London
pp 1ndash26
Landscape Ecol (2009) 241123ndash1133 1133
123
Copyright © 2022 FDOKUMEN












![Some Paradoxes of Human Rights: Fragmented Refractions in Neoliberal Times [2011]](https://static.fdokumen.com/doc/165x107/63151cd2fc260b71020fd9db/some-paradoxes-of-human-rights-fragmented-refractions-in-neoliberal-times-2011.jpg)







