
MAMMALIAN EPIDERMAL BARRIER LAYER LIPIDS: COMPOSITION AND INFLUENCE ON STRUCTURE

03 (2007) 703–714www.elsevier.com/locate/ydbio
Developmental Biology 3
Runx3 regulates dendritic epidermal T cell development
Eilon Woolf a, Ori Brenner b, Dalia Goldenberg a, Ditsa Levanon a, Yoram Groner a,⁎
a Department of Molecular Genetics, The Weizmann Institute of Science, Rehovot 76100, Israelb Department of Veterinary Resources, The Weizmann Institute of Science, Rehovot 76100, Israel
Received for publication 26 June 2006; revised 22 November 2006; accepted 5 December 2006Available online 8 December 2006
Abstract
The Runx3 transcription factor regulates development of T cells during thymopoiesis and TrkC sensory neurons during dorsal root ganglianeurogenesis. It also mediates transforming growth factor-β signaling in dendritic cells and is essential for development of skin Langerhans cells.Here, we report that Runx3 is involved in the development of skin dendritic epidermal T cells (DETCs); an important component of tissueimmunoregulation. In developing DETCs, Runx3 regulates expression of the αEβ7 integrin CD103, known to affect migration and epithelialretention of DETCs. It also regulates expression of IL-2 receptor β (IL-2Rβ) that mediates cell proliferation in response to IL-2 or IL-15. In theabsence of Runx3, the reduction in CD103 and IL-2Rβ expression on Runx3−/− DETC precursors resulted in impaired cell proliferation andmaturation, leading to complete lack of skin DETCs in Runx3−/− mice. The data demonstrate the requirement of Runx3 for DETCs developmentand underscore the importance of CD103 and IL-2Rβ in this process. Of note, while Runx3−/− mice lack both DETCs and Langerhans cells, thetwo most important components of skin immune surveillance, the mice did not develop skin lesions under pathogen-free (SPF) conditions.© 2006 Elsevier Inc. All rights reserved.
Keywords: Runx3-deficient mice; Transcription regulation; Skin DETCs; Vg3 T cell development; CD103 integrin; IL-2Rβ
Introduction
Developing T lymphocytes are divided into subclassesaccording to surface expression of T cell receptors (TCR) αβ orγδ (Pennington et al., 2005). Most γδ T cells reside in theepithelial layers of tissues such as skin and intestinal epithelium(Hayday et al., 2001). In mice, skin associated intraepithelialγδ T cells have a marked dendritic morphology and are knownas DETCs. These intraepithelial γδ T cells are thought torecognize stress-induced and pathogen-derived antigens and toplay an important role in the first line of epithelial immunedefense (Kabelitz et al., 2005). The unique specificity ofDETCs is conferred by an invariant skin-specific TCRcomposed of Vγ3/Vδ1 TCR chains lacking junctional diversity(Hayday and Tigelaar, 2003).
Vγ3 cells, the precursors to skin resident DETCs (Payer etal., 1991), appear in thymus at embryonal days (E) 14–16(Havran and Allison, 1988; Ito et al., 1989; Lafaille et al., 1989).
⁎ Corresponding author. Fax: +972 8 9344108.E-mail address: [email protected] (Y. Groner).
0012-1606/$ - see front matter © 2006 Elsevier Inc. All rights reserved.doi:10.1016/j.ydbio.2006.12.005
Immature Vγ3 thymocytes with high heat-stable antigen(HSAhigh) develop into mature HSAmed/low cells (Leclercq etal., 1993), which subsequently migrate to the skin. From E16on, the relative number of Vγ3 cells in the thymus decreasesand becomes nearly undetectable at late stages of fetaldevelopment (Havran and Allison, 1990). Skin DETCs appearin fetal epidermis at E17–E19 (Havran and Allison, 1988) andmaintain their round shape until birth; their typical dendriticmorphology is acquired only postnatally as the density of DETCclusters in the skin increases into adulthood (Payer et al., 1991).Cultured mature Vγ3 thymocytes require IL-2 for proliferation,but IL-2-deficient mice have normal numbers of Vγ3 cells(Leclercq et al., 1990b). This occurrence was attributed to invivo activity of IL-15 which can also support the proliferation ofmature Vγ3 thymocytes (Leclercq et al., 1996). IL-2 and IL-15are both ligands of the IL-2 receptor β (IL-2Rβ), expressed onmature Vγ3 thymocytes (Leclercq et al., 1995). In IL-15 and IL-2Rβ-deficient mice, the number of mature Vγ3 thymocytes isreduced and adult skin Vγ3 DETCs are absent (De Creus et al.,2002; Kawai et al., 1998). Thus, signaling through IL-2Rβplays a critical role in Vγ3 T cell proliferation and survival.

704 E. Woolf et al. / Developmental Biology 303 (2007) 703–714
The αEβ7 integrin CD103 is mainly expressed by intraepithe-lial lymphocytes (Kilshaw and Baker, 1988) and confersspecificity for the epithelial ligand E-cadherin (Cepek et al.,1994). The interaction of CD103 with E-cadherin is assumed tomediate the epithelial retention of mucosal T lymphocytes andcould explain the low number of mucosal T cells in CD103-deficient mice (Schon et al., 1999). CD103 is also expressed onmost Vγ3 DETCs and may affect their immigration and retentionin epidermis, consistent with the marked reduction of DETCs inCD103-deficient mice (Schon et al., 2002). Interestingly, bothCD103-deficient andTCRδ-deficientmice spontaneously developinflammatory skin lesions (Girardi et al., 2002; Schon et al., 2000).The αEβ7 integrin CD103 is a heterodimer composed of twochains (αE and β7) each encoded by a separate gene (Robinson etal., 2001). In vitro studies indicated that surface expression ofCD103 on activated T cells is regulated by TGFβ (Smith et al.,1994) as is the transcription of the αE gene (Robinson et al., 2001).In vivo, TGFβ signaling is required for efficient induction ofCD103 expression by graft-infiltrating CD8 effector T cells (El-Asady et al., 2005;Wang et al., 2004). Finally, both TGFβ and thetranscription factor Runx3 are required for CD103 expression onthe CD4 T cell line RLM-11-1 (Grueter et al., 2005) and eitherknockdown or knockout (KO) of Runx3 markedly reduced thefrequency of CD103 expressingCD8+ Tcells (Grueter et al., 2005;Yarmus et al., 2006). Hence, Runx3 positively regulates theexpression of CD103 during T cell development.
Mammalian Runx3 is one of three proteins comprising theRUNT domain family of transcription factors RUNX (Cameronand Neil, 2004; de Bruijn and Speck, 2004; Levanon and Groner,2004; van Wijnen et al., 2004). In the thymus where the threemembers Runx1, Runx2 and Runx3 are highly expressed(Bangsow et al., 2001; Taniuchi et al., 2002; Vaillant et al.,2002), the expression of Runx1 and Runx3 is confined to thecortex and medulla, respectively (Woolf et al., 2003). Studies inKO mice have delineated several cell-autonomous functions ofRunx3. Loss of Runx3 is associated with defects in neurogenesis,thymopoiesis and dendritic cell (DC) function and withdevelopment of colitis, gastritis and asthma like features (Levanonet al., 2002; Fainaru et al., 2004;Woolf et al., 2003; Brenner et al.,2004; Fainaru et al., 2005; Inoue et al., 2002; Levanon et al., 2003;Taniuchi et al., 2002). In neurogenesis and thymopoiesis, Runx3was required for development and survival of dorsal root gangliaTrkC neurons (Levanon et al., 2002; Inoue et al., 2002) and forsilencing the CD4 gene during T cell development (Ehlers et al.,2003; Woolf et al., 2003; Taniuchi et al., 2002). In the DCcompartment, Runx3 functions as a component of the TGFβsignaling pathway (Fainaru et al., 2004; Fainaru et al., 2005) andin its absence TGFβ-dependent development of skin Langerhans(LC) cells was abrogated, as was TGFβ-dependent inhibition ofDC maturation (Fainaru et al., 2004; Fainaru et al., 2005).
Given the important functions of Runx3 in development ofepidermal LC (Fainaru et al., 2004; Fainaru et al., 2005) andspecifications of CD8-lineage T cells (Ehlers et al., 2003; Woolfet al., 2003; Taniuchi et al., 2002), we have investigated thefunction of Runx3 in biogenesis of Vγ3 DETCs. We show thatRunx3 is required for the proper development of embryonalVγ3 thymocytes and when lost their maturation and prolifer-
ation in the thymus are impaired, resulting in a completeabsence of skin DETCs in adult KO mice. How does Runx3regulate DETCs development? We found that in developingVγ3 DETCs Runx3 regulates the expression of both CD103and IL-2Rβ that are markedly reduced on the KO Vγ3thymocytes. While the diminished level of IL-2Rβ results inimpaired IL-2- and IL-15-dependent proliferation of the KOVγ3 thymocytes, the reduced level of CD103 may affect cellmigration and epithelial retention, leading to lack of skinDETCs in the KO mice. As Runx3 is essential for the properdevelopment of LCs (Fainaru et al., 2004) and DETCs (thisreport), the two main components of skin immunity (Borkowskiet al., 1996; Romani et al., 2003; Sharp et al., 2005b), it wasinteresting to note that although cutaneous abnormalities havebeen described in mice deficient in either LCs or DETCs(Girardi and Hayday, 2005; Kaplan et al., 2005; Schon et al.,2000; Sharp et al., 2005a), Runx3 KO mice neither developedspontaneous skin lesions, under SPF conditions, nor did theyexhibit enhanced contact hypersensitivity (CHS). Usingtransgenic mice bearing diphtheria toxin receptor in theLangerin gene, two groups ablated skin LCs and demonstratedthat these cells are not essential for CHS (Bennett et al., 2005;Kissenpfennig et al., 2005b; Kissenpfennig and Malissen,2006). Thus, the cutaneous phenotype of Runx3 KO micesupports the notion that dermal DCs are sufficient for inductionof T cell response to epicutaneous application of haptens.
Materials and methods
Mice and histology
Runx3 KO mice were generated as described previously (Levanon et al.,2002) and bred on several genetic backgrounds including ICR, MF1, C57BL/6and FVB. When crossed onto a C57BL/6 or FVB background, homozygotesRunx3 KO did not survive postnatally (Levanon et al., 2002). Therefore, WTmice and KO littermates of ICR and MF1 backgrounds were used for all theexperiments. Mice were bred and maintained in a pathogen-free facility. TheInstitutional Animal Care and Use Committee (IACUC) of The WeizmannInstitute approved all mouse experiments. Analysis of KO mice (n=5) and WTlittermates (n=4) including males and females was conducted as previouslydescribed (Levanon et al., 2002). Samples of dorsal interscapular skin and rightfore- or hindlimb paws were collected fixed in 10% neutral-buffered formalinand embedded in paraffin. Section were cut at 4 μm and stained withhematoxylin and eosin (Levanon et al., 2002).
Flow cytometry and cell sorting
Suspension of fetal thymocytes was obtained by teasing the thymic lobeswith fine forceps and passing through a nylon mesh. Suspension of epidermalcells was prepared from ear skin of adult mice by treatment with trypsin asdescribed (Borkowski et al., 1996). Cells (1–10×105) were resuspended in PBSsupplemented with 2% fetal calf serum, 0.05% NaN3 and 5 to 25 mM EDTA.After pre-incubation with anti-FcγII/III receptor mAb (2.4G2) to avoid non-specific binding of mAb, saturating amounts of mAbs were added. Cells wereincubated for 30 min at 4 °C, washed, resuspended in FACS buffer andanalyzed. When necessary, cells were incubated with secondary reagents for30 min on ice. In most cases TO-PRO3-iodide (Molecular Probes, Eugene, OR,USA) was added prior to analysis to specifically label the non-viable cells. Flowcytometry was performed by using a FACSCalibur (Becton Dickinson)equipped with CELLQUEST software (Becton Dickinson). Preparative cellsorting of CD8β+ splenocytes was performed using FACS Vantage cell sorter(BD Biosciences). Monoclonal Abs used for staining and for in vitro T cell

705E. Woolf et al. / Developmental Biology 303 (2007) 703–714
stimulation were CD4-FITC (GK1.5), CD8β-PE (H35-17.2), TCR-Vγ3 (F536),CD103-biotin (M290), TCRγδ (GL3), CD16/32 (2.4G2), CD3-purified andCD3-biotin (145-2C11) (all from BD; Pharmingen). CD3-APC (145-2C11) andIL-2Rβ (TM-β1) were from eBiosciences.
Immunofluorescence staining of epidermal sheets
Epidermal sheets were prepared from either embryo skin or adult ear byincubation in 0.5 M ammonium thiocyanate for 20 min at 37 °C (Juhlin andShelley, 1977). Acetone-fixed epidermal sheets were then incubated with biotin-conjugated CD3 mAb for 1 h at 37 °C, rinsed in PBS, mounted in bufferedglycerol containing 0.3 M 1,4-diazabicyclo [2.2.2] octane (Sigma Chemical,USA) and examined by an Olympus IX71 fluorescent microscope.
Stimulation of T cells
For monitoring up-regulation of CD103 in T cells, cells were released fromspleen and erythrocytes removed by treatment with ACK buffer (0.15 M NH4Cl;1 mM KHCO3; 0.1 mM Na2 EDTA). Splenocytes were depleted of adherentcells, passed through a nylon mesh and 2×106 cells/ml were stimulated with5 μg/ml plate bound anti-CD3 mAb and 2 ng/ml recombinant murine IL-2(PeproTech Rehovot Israel) in the absence or presence of 5 ng/ml recombinanthuman TGF-β1 or TGF-β2 (PeproTech Israel). Cell were cultured for 4 days inRPMI 1640 supplemented with 10% heat-inactivated FCS (Gibco, USA), 100 U/ml penicillin-streptomycin, 2 mM L-glutamine and 50 μM2-ME, in a humidified5% CO2 at 37 °C. For FACS analysis, cells were stained with CD4, CD8β andCD103mAbs. For monitoring Runx3 protein level in naïve vs. stimulated CD4 Tcells, splenocytes were stimulated with 2 μg/ml Con-A (Sigma, USA) in thepresence of IL-2 and TGF-β1 for 3 days after which CD4+ and CD4+CD103+
cells were sorted using the FACS Vantage and Runx3 protein was determined aspreviously described (Woolf et al., 2003; Fainaru et al., 2004). IL-2 stimulation offetal thymocytes was performed as described (Leclercq et al., 1990b).
Measurement of apoptosis
Apoptosis was analyzed by quantifying exposed phosphatidylserine residueson external cell membrane (Vermes et al., 1995). Cells were stained with Vγ3-PE mAb, washed and stained with Aposcreen Annexin-V FITC (SouthernBiotech, USA) according to the manufacture's instructions. Prior to analysis byflow cytometry, cells were counterstained with 2 μM TO-PRO3-iodide(Molecular Probes, Eugene, OR, USA).
RT-PCR
RNA was isolated using EZ-RNA total RNA isolation kit (Biologicalindustries, Israel). cDNA was generated using Superscript II RNaseH− reversetranscriptase (Invitrogen) and PCR products were derived using the sets ofoligonucleotides and PCR conditions indicated below: αE (sense) 5′-CCAGAAGGCCAAAAATTTCA-3′ (anti-sense) 5′-TCAGCAGC-GACTCCTTTTCCGCTT-3′; 60 °C; 30,25 cycles, yielding a 197-bp product.β-Actin (sense) 5′-GATGACGATATCGCTGCGCTG-3′ (anti-sense) 5′-GTAC-GACCAGAGGCATACAGG-3′; 55 °C; 19,17 cycles, yielding a 430-bpproduct. Vγ3-Jγ1 RT-PCR was performed as described (Minagawa et al.,2001). The primers used to detect Runx3 transcripts encoding the p33 and p46proteins are as follows: for the p33 transcript, forward AGTCCTTGCCACTGT-CAGC, reverse GTGAGGCTCTGCAGCGTAG. Size of PCR product 570 bp.For the p46 transcript, forward GGCAAGATGGGCGAGAACAC, reverseCGTAGGGAAGGAGCGGTCA. Size of PCR product 670 bp.
Results
Runx3 KO mice lack skin DETCs
Skin immunity depends to a large extent on residentLangerhans cells (LCs) and Vγ3 TCR-expressing DETCs.
The requirment for Runx3 in the development of skin LC wasdemonstrated (Fainaru et al., 2004), but its role in the biogenesisof DETCs was not investigated. We examined epidermal sheetsof embryos and adult WT and KO mice by in situ fluorescentstaining with anti-CD3 antibodies (Ab). Skin of WT E18embryos and 8-week old adult mice contained round anddendritic shaped DETCs, respectively (Fig. 1A). In contrast, noDETCs were detected in either embryo or adult Runx3 KOmice(Fig. 1A). Further analysis of epidermal cells by flow cytometryusing anti CD3, CD103 and Vγ3 Abs verified the completeabsence of DETCs in Runx3 KO epidermis (Fig. 1B).Epidermal sheets of adult WT and KO mice were alsoimmunostained with Langerin (CD207); a C type lectinspecifically expressed by LCs (Valladeau et al., 2002). Nostaining was detected in KO epidermis, whereas abundant LCswere stained in WT (Fig. 1C), confirming the previousobservation (Fainaru et al., 2004) that skin epidermis ofRunx3 KO mice completely lack LCs.
DETCs uniformly express non-diverse γδ TCR, composedof Vγ3-Jγ1 and Vδ1-Dδ2-Jδ2 chains (Asarnow et al., 1988,1989), with invariant junctional sequences identical to TCR ofearly fetal thymocytes (Lafaille et al., 1989). Vγ3-Jγ1transcripts were recently also detected in the gut of E18mouse embryo (Minagawa et al., 2001). Using this information,we next evaluated the presence of DETCs in skin and gut of WTand KO mice using RT-PCR. Vγ3-Jγ1 transcripts were notdetected in either epidermis or gut of Runx3 KO E18 embryos(Fig. 1D), reconfirming the complete absence of peripheral,non-thymic, Vγ3 T cells in the KO mice.
Since a reduction in the number of DETCs is associated withinflammatory skin lesions (Schon et al., 2000, 2002; Girardi etal., 2002), we examined histologically skin samples from adultKO and WT mice bred on two different genetic backgrounds(Brenner et al., 2004; Levanon et al., 2002), MF1 and ICR.Macroscopic and microscopic evaluation revealed no signs ofan inflammatory process in any of the mice examined (Fig. 1E).Additionally, skin IHC analysis using TdT-mediated dUTP nickend labeling (TUNEL) showed no increase in apoptosis in KOepidermis (not shown). Nevertheless, there was a strikingdifference in skin thickness primarily due to absence ofsubcutaneous fat in the KO mice (Fig. 1E). This occurrence isrelated to the markedly reduced body weight of the KO mice(Brenner et al., 2004; Levanon et al., 2002). The epidermis wasof similar thickness in both WT and KO mice (Fig. 1E).
We next used IHC and Western blotting analysis ofepidermal differentiation markers to further examine theepidermis of Runx3 KO mice. Comparison revealed thatdistribution and morphological pattern of the spinous layermarker K10 and the granular layer marker Loricrin were similarin WT and KO mice (Fig. 1F). These results completelyconform to the recently reported data that show no major changebetween Runx3 KO and WT in the overall morphology of theskin and its appendages (Raveh et al., 2005). Skin sections ofRunx3 KO and WTwere also stained with anti-Runx3 Ab (Fig.1G). Runx3 expression was detected in WT dermis in the hairdermal papillae and in isolated cells in the epidermis. Thesescattered Runx3-positive cells, which did not express

Fig. 1. Analysis of Runx3 KO skin shows complete absence of DETCs and LCs, but no structural or morphological changes, as compared to WT. (A) Lack ofepidermal DETCs in E18 embryos (top) and adult Runx3 KOmice (bottom). Epidermal sheets of E18 dorsal skin or 8 weeks old ear lobes fromWTand KOmice weresubjected to immunofluorescence using CD3-biotin Ab/Cy3-streptavidin. Fetal DETCs have a round shape and acquire the characteristic dendritic morphology inadult. (B) Flow cytometry analysis of epidermal cell suspension reveals the absence of Vγ3+CD3+CD103+ DETCs in Runx3 KOmice. Epidermal cell suspension of 8-week-old mice were stained with anti HSA, Vγ3 and CD103 Abs. Dot plot analysis depicts the distribution of CD103 vs. Vγ3 in gated HSAlow-positive cells. (C) Lackof epidermal LCs in adult Runx3 KO mice. Epidermal sheets of 8 weeks old ear lobes from WT and KO mice were subjected to immunofluorescence using rat antiLangerin Ab (1:200, secondary Ab: anti rat Cy3) (Kissenpfennig et al., 2005a). (D) Absence of Vγ3 RNA in epidermis of Runx3 KO mice. RNA obtained from WTand KO epidermis (left panel) or from gut of E18 WTand KO embryos (right panel) was analyzed for presence of the Vγ3-Jγ1 transcript by RT-PCR. (E) Histologicalanalysis of skin from WT and KO mice revealed no inflammatory lesions. Sections of dorsal interscapular skin of WT (left panel) and KO (right panel) stained withhematoxylin and eosin. Ep—epidermis, De—dermis, Sub Fat—subcutaneous fat (magnifications: ×10 and ×25). (F) IHC analysis of epidermal differentiationmarkers (left panel) and Western blotting (right panel). Shown are the spinous layer marker K10 and the granular layer marker Loricrin. (G) Two days old back skinsections from WT and Runx3 KO mice were analyzed using Runx3 Ab (Levanon et al., 2001). No Runx3-specific staining is observed in the KO section.
706 E. Woolf et al. / Developmental Biology 303 (2007) 703–714
cytokeratins (Raveh et al., 2005), are most probably LCs andDETCs. Consistent with this interpretation are the observationsthat these cells first appear at birth, after which their densityincreases (Raveh et al., 2005) and are MHCII and CD3 positive,respectfully. No Runx3-specific staining was observed insections of Runx3 KO skin (Fig. 1G).
Runx3 is required for the development of embryonal Vγ3thymocytes
The complete absence of peripheral Vγ3 DETCs in Runx3KO mice raised the possibility that Runx3 plays a critical role inthe development of Vγ3 thymocytes during fetal development;

707E. Woolf et al. / Developmental Biology 303 (2007) 703–714
hence, we examined whether Runx3 is expressed in Vγ3thymocytes. Immunostaining of WT, FACS sorted, E17.5 Vγ3thymocytes revealed the expression of Runx3 in a largeproportion of Vγ3 thymocytes (Fig. 2A).
Immature TCR Vγ3 thymocytes, with a Vγ3low/HSAhigh
phenotype, differentiate into mature Vγ3high/HSAlow cellsduring the last few days before birth (Leclercq et al., 1993).To examine the development of fetal Vγ3 thymocytes in WTand KO mice, E16.5 and E18.5 thymocytes were stained withHSA and Vγ3 Abs and analyzed by FACS. Compared to WT, amarked reduction in the proportion of Vγ3high cells wasobserved among E16.5 KO thymocytes, with a concomitant risein the proportion of Vγ3med and Vγ3low cells (Fig. 2B). Of note,the Vγ3med/Vγ3high ratio was 7-fold higher in the KOcompared to WT (0.6 in WT Vs 4.2 in KO), and almost allVγ3high cells in the KO displayed an HSAmed/high phenotype(Fig. 2B) indicating a defect in the maturation process of KOVγ3 thymocytes. Supporting this conclusion is the observationthat even at a more advanced developmental stage (E18.5), theproportion of Vγ3high cells in the KO remained low and the
Fig. 2. Impaired development of Vγ3 thymocytes in Runx3 KO mice. (A)Runx3 is expressed in Vγ3 fetal thymocytes. E17 thymocytes were stained withHSA-biotin and Vγ3-PE, followed by streptavidin-APC and Vγ3+ cells weresorted by FACS. Isolated HSAlowVγ3+ thymocytes were placed on cover slipand stained with a rabbit anti-Runx3 polyclonal Ab followed with donkey anti-Rabbit Cy2 (green). Substantial fraction of cells displayed coexpression ofcytoplasmic/membranal Vγ3-PE (stained red) and nuclear Runx3-CY2 (stainedgreen). (B) Maturation of Runx3 KO Vγ3 thymocytes is impaired. Freshlyprepared cell suspensions of E16.5 (upper panel) and E18.5 (lower panel)thymocytes, were double stained with HSA-FITC and Vγ3-PE and analyzed byFACS. Vγ3med and Vγ3high subsets are gated and percentages of eachpopulation are shown. Results are representative of three to four independentexperiments.
average cell-HSA level was higher than in WT (Fig. 2B). Theseresults indicate that Runx3 is required for the properdevelopment of embryonal Vγ3 thymocytes and when Runx3is lost their maturation in the thymus is impaired.
Runx3 regulates CD103 expression in Vγ3 thymocytes
The αEβ7 integrin CD103 is expressed on a substantialportion of Vγ3 thymocytes and on the majority of skin residentVγ3 cells (Lefrancois et al., 1994). Whereas CD103-deficientmice exhibit a marked reduction in skin DETCs (Schon et al.,2002), not much is known about the role of CD103 indevelopment of thymic Vγ3+ DETC precursors during fetalthymopoiesis. Because CD103 expression in CD8 T cells isregulated by Runx3 (Grueter et al., 2005; Yarmus et al., 2006),we compared the expression of CD103 on developing Vγ3 fetalthymocytes of WT and Runx3 KO mice. E16.5 Vγ3med KOthymocytes exhibited reduced CD103 expression, reflected in alower mean-fluorescence-intensity (MFI) (Fig. 3A). This lowerCD103 expression was more pronounced in Vγ3high thymo-cytes, where both the MFI and the proportion of CD103expressing cells were significantly reduced in the KO (Fig. 3A).At E18.5, both Vγ3med and Vγ3high KO thymocytes exhibitedreduced expression of CD103 as compared to WT (Fig. 3B).These data indicate that Runx3 is required for CD103expression in developing Vγ3 thymocytes.
Since Runx3 functions as a component of the TGFβsignaling pathway (Shi and Stavnezer, 1998; Fainaru et al.,2004, 2005), and TGFβ can induce CD103 expression on Tlymphocytes (Kilshaw and Murant, 1991; Parker et al., 1992;Robinson et al., 2001; Smith et al., 1994), we evaluated theability of TGFβ1 and TGFβ2 to induce expression of CD103 incultured fetal thymocytes. Analysis of incubated E16.5 WTthymocytes showed that both TGFβ1 and TGFβ2 inducedCD103 expression on Vγ3+ fetal thymocytes (Fig. 4A). Wenext asked whether Runx3 transcriptionally regulates CD103expression in response to TGFβ using cultured primary CD4+ Tcells from WT and Runx3 KO mice. We employed CD4+
splenocytes because contrary to CD8 T cells, only a smallfraction (<6%) of TCRαβ CD4+ T cells normally expressCD103. WT and Runx3 KO splenocytes were cultured in thepresence or absence of TGFβ1. After 4 days in culture withTGFβ, more than 40% of WT CD4+ T cells expressed CD103whereas no such expression was observed on KO cells (Fig.4B). As Runx3 expression in naïve CD4+ splenocytes is low(Taniuchi et al., 2002; Woolf et al., 2003), we examined whetherT cell activation induces Runx3 expression in CD4+ spleno-cytes. Western blot analysis clearly demonstrated that in vitrostimulation of CD4+ splenocytes resulted in up-regulation ofRunx3 expression in both CD4+CD103+ and CD4+CD103−
splenocytes (Fig. 4C). Interestingly, this enhanced Runx3expression was mainly due to transcriptional activation of thegene P1 promoter (Bangsow et al., 2001), reflected by thesubstantial amount of a truncated transcript, encoding theRunx3 p33 protein, which was detected in activated KO CD4+
splenocytes (Fig. 4C) (Fainaru et al., 2004). The impact ofRunx3 on TGFβ2-mediated expression of CD103 was also

Fig. 3. Diminished CD103 expression in Runx3 KO Vγ3 fetal thymocytes. Cell suspensions of WT and KO E16.5 and E18.5 thymocytes were triple stained withHSA-FITC, Vγ3-PE and CD103-biotin Abs, followed by reaction with streptavidin PE-Cy5 and analyzed by FACS. Results are representative of three to fourindependent experiments. (A) CD103 expression in E16.5 gated Vγ3med (upper) and Vγ3high (lower) thymocytes is depicted. Percentages of CD103 expressing cellsare shown. Mean fluorescence intensities (MFI) of CD103 are indicated at the right corners of histograms. (B) CD103 expression in E18.5 Vγ3 thymocytes wasanalyzed as in A.
708 E. Woolf et al. / Developmental Biology 303 (2007) 703–714
evaluated in CD8+ splenocytes. After 5 days in culture,expression of CD103 was up-regulated in WT but not in KOCD8+ splenocytes (Fig. 4D). When taken together these data areconsistent with the conclusion that Runx3 mediates the well-documented TGFβ-induced expression of CD103 in cultured Tcells and that both TGFβ1 and TGFβ2 promote this induction.
Finally, we asked whether Runx3 directly regulates CD103transcription. As mentioned earlier, CD103 is a heterodimercomposed of the αE and β7 chains each encoded by a separategene. As the β7 chain is highly expressed in T cells, the limitingfactor in CD103 expression is the transcription of the aE gene(Robinson et al., 2001). We used RT-PCR to monitor aEtranscription in sorted CD8αβ+ T cells of WT and Runx3 KOmice. In comparison to WT, transcription of αE in KO CD8+ Tcells was greatly diminished (Fig. 4E), attesting to the function ofRunx3 as a transcription regulator of CD103 in T cells. AsCD103-deficient mice exhibit a marked reduction in skin DETCs(Schon et al., 2002), it is conceivable that the decreased CD103expression in Runx3 KO T cells contributed to the completeabsence of peripheral, non-thymic, Vγ3 T cells in the KO mice.
Diminished expression of IL-2Rβ on Runx3 KO Vγ3thymocytes is associated with a lower IL-2/IL-15-mediatedcell-proliferation and a higher apoptosis
Several studies have suggested a role for IL-2 and IL-15signaling, through the IL-2Rβ chain, in Vγ3 Tcell development
and DETCs localization in the skin. First, both IL-2 and IL-15play a role in the development of Vγ3 T cells (De Creus et al.,2002; Leclercq et al., 1990b). Second, mature fetal Vγ3thymocytes and Vγ3 DETCs express the IL-2Rβ chain and inutero treatment with anti-IL-2Rβ Ab abrogates DETCsappearance in the adult skin (Kawai et al., 1998; Tanaka etal., 1992). Third, both IL-2 and IL-15 interact with receptorcomplexes that contain the γc, IL-2Rβ chain and a specific IL-2R or IL-15Rα chain (de Jong et al., 1996; Giri et al., 1994).Fourth, IL-2Rβ-deficient mice exhibit a moderate reduction ofmature Vγ3 cells in fetal thymus and a complete lack of skinDETCs in adult mice (Kawai et al., 1998). Consequently, weexamined whether the expression level of IL-2Rβ ondeveloping Runx3 KO fetal thymocytes was altered.
FACS analysis of E18.5 Vγ3 thymocytes revealed asubstantially lower IL-2Rβ level on KO compared to WTcells (Fig. 5A). This lower IL-2Rβ expression posed thequestion as to whether it affects the response of KO Vγ3thymocytes to IL-2. Leclercq et al. (1990b) have shown thatE17 to E19 Vγ3 thymocytes preferentially expand during invitro stimulation by IL-2. We therefore cultured E18.5 WT andRunx3 KO thymocytes in the presence of IL-2 and monitoredthe increase in the HSAlow Vγ3+ cell population between 24and 72 h of incubation (Fig. 5B). We observed a 4 times higherincrease in WT HSAlow Vγ3+ thymocytes compared to KOthymocytes (Fig. 5B) (28±3-fold increased of WT, compared to7.2±0.75-fold of KO). The results indicate that Runx3 impacts

Fig. 4. Runx3 mediates TGFβ-dependent expression of CD103 in cultured primary T cells. (A) TGFβ up-regulates CD103 expression in E16 Vγ3+ thymocytes. WTE16 thymocytes were cultured for 5 days with anti-TCRγδ Ab in the presence of IL-2 and IL-7 without or with TGFβ1 or TGFβ2. Cell suspensions were processedand analyzed by FACS as detailed in Fig. 3B. CD103 expression in gated Vγ3+ thymocytes is shown. Results are representative of three to four independentexperiments. (B) Runx3 is required for TGFβ-dependent expression of CD103 in primary CD4+ T cells. Splenocytes from WT and Runx3 KO mice were depleted ofadherent cells and stimulated with plate bound anti-CD3 and IL-2 in the presence or absence of TGFβ1 (5 ng/ml). After 4 days, cells were harvested and stained withCD4, CD8β and CD103 Abs and analyzed by FACS. To-Pro3 was added immediately before analysis and dead cells were gated out. CD4+ cells were gated andmonitored for CD103 expression. Results are representative of five independent experiments. ((−) TGFb—solid line; (+) TGFb—interrupted line). (C) Expression ofRunx3 in activated CD4+ and CD4+CD103+ T cells. Left panel: WT splenocytes were cultured and stimulated with ConA, IL-2 and TGFβ1. After 3 days, cells wereFACS sorted into CD4+ and CD4+CD103+ T cell subsets proteins extracted and analyzed by Western blots using polyclonal Rabbit anti-Runx3 and anti-GAPDH Abs.A marked stimulation of the two Runx3 protein bands of 48 and 46 kDa was evident in activated CD4+ and CD4+CD103+ splenocytes compared to non-activated naïvecells. Right panel: activated WT and KO CD4+ splenocytes were FACS sorted; RNAwas derived and analyzed by RT-PCR. The WT and KO RT-PCR products aredesignated Runx3 p46 and p33, respectively. (D) Runx3 regulates TGFβ2-mediated expression of CD103 in CD8+ T cells. WT and Runx3 KO splenocytes werecultured for 5 days in the presence of anti-CD3 Ab and IL-2 and in the presence or absence of TGF-β2 (5 ng/ml). Cells were stained with CD4, CD8β and CD103 Absand CD103 expression was determined in gated CD8β+ cells. Results are representative of three independent experiments. (E) Runx3 transcriptionally regulates theexpression of the αE gene. FACS-sorted CD8β+ splenocytes were obtained and extracted RNA used in an RTPCR reaction to detect αE mRNA. (197 bp; 32 and 29cycles). The corresponding semi-quantitative β-actin RT-PCR is shown (490 bp; 20 and 16 cycles).
709E. Woolf et al. / Developmental Biology 303 (2007) 703–714
on IL-2Rβ expression and in the absence of Runx3, IL-2Rβlevel on KO Vγ3 thymocytes diminished resulting in a lowerproliferation in response to IL-2. This conclusion was supportedby the finding that the proliferation rate of KO Vγ3 thymocytesin response to IL-15 was similarly lower compared to WT (WT15±2.7; KO 2±0.8; Fig. 5C).
IL-2 not only stimulates in vitro proliferation of Vγ3thymocytes, but it can also rescue mature Vγ3 thymocytes fromapoptosis (Leclercq et al., 1995). We asked whether culturedRunx3 KO Vγ3 thymocytes, that express lower levels of IL-2Rβ, display increased apoptosis during IL-2 stimulation ascompared to WT thymocytes (Figs. 5D and E). At 24 h afterseeding, KO cultures contained twice as many apoptotic Vγ3cells compared to WT (KO—53±4.6%; WT—25±5.7%) (Fig.5D), and this preponderance of apoptotic Vγ3 cells among the
KO thymocytes increased at 36 h in culture (KO—50±2.1%;WT—14±2.1%) (Fig. 5E). Together these data show that inRunx3 KO Vγ3 thymocytes, the impaired expression of IL-2Rβ is associated with diminished effects of IL-2 and IL-15 oncell proliferation and apoptosis.
Discussion
During T cell development, Runx3 is highly expressed indouble positive CD4+/CD8+ thymocytes and in mature CD8+ Tcells (Taniuchi et al., 2002; Woolf et al., 2003), acting as bothpositive and negative regulator of target genes (Yarmus et al.,2006). In CD4+/CD8+ thymocytes, Runx3 negatively regulatesCD4 expression whereas in mature CD8+ T cells it positivelyregulates cell growth-promoting genes. Accordingly, in Runx3-
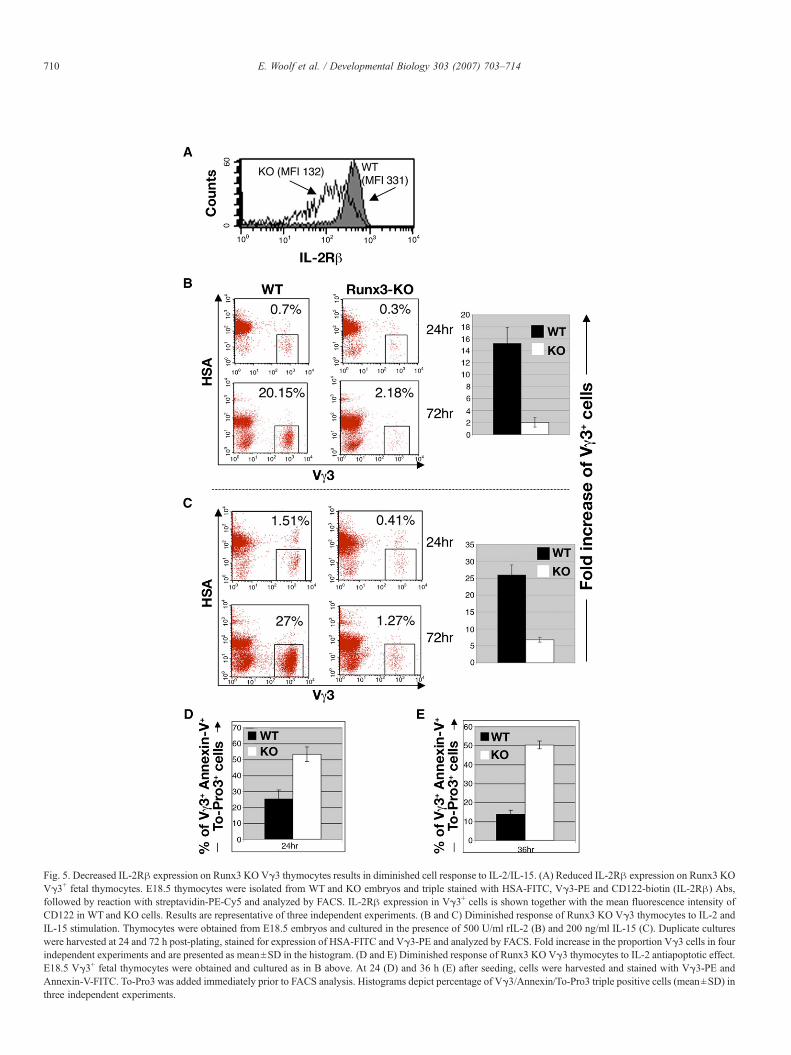
Fig. 5. Decreased IL-2Rβ expression on Runx3 KO Vγ3 thymocytes results in diminished cell response to IL-2/IL-15. (A) Reduced IL-2Rβ expression on Runx3 KOVγ3+ fetal thymocytes. E18.5 thymocytes were isolated from WT and KO embryos and triple stained with HSA-FITC, Vγ3-PE and CD122-biotin (IL-2Rβ) Abs,followed by reaction with streptavidin-PE-Cy5 and analyzed by FACS. IL-2Rβ expression in Vγ3+ cells is shown together with the mean fluorescence intensity ofCD122 in WT and KO cells. Results are representative of three independent experiments. (B and C) Diminished response of Runx3 KO Vγ3 thymocytes to IL-2 andIL-15 stimulation. Thymocytes were obtained from E18.5 embryos and cultured in the presence of 500 U/ml rIL-2 (B) and 200 ng/ml IL-15 (C). Duplicate cultureswere harvested at 24 and 72 h post-plating, stained for expression of HSA-FITC and Vγ3-PE and analyzed by FACS. Fold increase in the proportion Vγ3 cells in fourindependent experiments and are presented as mean±SD in the histogram. (D and E) Diminished response of Runx3 KO Vγ3 thymocytes to IL-2 antiapoptotic effect.E18.5 Vγ3+ fetal thymocytes were obtained and cultured as in B above. At 24 (D) and 36 h (E) after seeding, cells were harvested and stained with Vγ3-PE andAnnexin-V-FITC. To-Pro3 was added immediately prior to FACS analysis. Histograms depict percentage of Vγ3/Annexin/To-Pro3 triple positive cells (mean±SD) inthree independent experiments.
710 E. Woolf et al. / Developmental Biology 303 (2007) 703–714

711E. Woolf et al. / Developmental Biology 303 (2007) 703–714
deficient mice CD4 silencing is abrogated and CD8+ T cellproliferation is diminished (Ehlers et al., 2003; Taniuchi et al.,2002; Woolf et al., 2003; Yarmus et al., 2006). Here we showthat Runx3 is expressed in Vγ3+ thymocytes and when absentthe skin population of Vγ3 DETCs largely diminished,underscoring the importance of Runx3 function for properdevelopment of Vγ3+ DETCs. Interestingly, Runx3 is alsoessential for development of LCs, as Runx3 KO mice lack skin-resident LCs (Fainaru et al., 2004). We have recently reportedthe existence of a CD3+ cell population in epidermal sheets ofRunx3 KO mice (Fainaru et al., 2004). However, furtheranalysis that culminated in the present study revealed that theobserved staining with anti CD3 antibodies was actually a non-specific absorption, which was mistakenly interpreted as DETC.As further demonstrated in Fig. 1G, this correction does notaffect the conclusion of the Fainaru et al. (2004) study that theepidermis of Runx3 KO mice is devoid of LCs. Hence, Runx3-deficient mice constitute a unique mouse model in which bothLCs and DETCs are missing.
Given the fact that Vγ3+ DETCs and LCs are importantcomponents of skin immunity (Kabelitz et al., 2005; Romani etal., 2003; Sharp et al., 2005b), it is worth noting that Runx3 KOmice neither developed skin lesions nor did they displayincreased epidermal apoptosis or contact hypersensitivity (Fig.1 and unpublished observations). This finding is intriguing asother mouse mutants that lack either DETCs or LCs exhibitedskin inflammation (Girardi et al., 2002; Schon et al., 2000),increased epidermal apoptosis (Sharp et al., 2005a) or enhancedcontact hypersensitivity (CHS) (Kaplan et al., 2005), respec-tively. On the other hand, the lack of enhanced CHS in ourRunx3 KO mice is consistent with recent reports demonstratingthat LCs are not essential for CHS (Bennett et al., 2005;Kissenpfennig et al., 2005b; Kissenpfennig and Malissen,2006). Thus, the cutaneous phenotype of Runx3 KO micesupports the notion that CHS is mediated mainly by dermal DC(Bennett et al., 2005; Kissenpfennig et al., 2005b; Kissenpfen-nig and Malissen, 2006). However, more studies will berequired to define the physiological factors underlying the lackof skin lesions in Runx3 KO mice because in addition to itsimportance for DCs, LCs and DETCs development andfunction, Runx3 also regulates T cells differentiation andhomeostasis (Fainaru et al., 2004; Taniuchi et al., 2002; Woolfet al., 2003).
The complete absence of skin Vγ3+ DETCs in Runx3 KOmice incited us to examine whether the absence of Runx3impacted on early stages of Vγ3 thymocyte development. Wefound that in KO fetuses, the maturation of Vγ3 thymocyteswas impaired resulting in a marked reduction in the proportionof Vγ3high/HSAlow cells in the thymus. But what is theunderlying cause for this reduced proportion of Vγ3high/HSAlow fetal thymocytes in the KO mice?
Integrins play a pivotal role in thymocyte development(Schmeissner et al., 2001), by influencing stage-specific cellgrowth and differentiation (Kutlesa et al., 2002). The integrinCD103 mediates T cell adhesion to epithelial cells throughbinding to its only known ligand E-cadherin, which isselectively expressed on epithelial cells (Cepek et al., 1994).
As CD103 is expressed on 40–60% of Vγ3+ thymocytes and onthe majority of skin-resident Vγ3+ cells, it was suggested thatCD103 plays a role in thymic development and subsequentmigration of Vγ3 cells (Lefrancois et al., 1994). Indeed,CD103-deficient mice exhibit a marked reduction in epidermalVγ3 DETCs (Schon et al., 2002). We find, as was previouslyreported for CD8+ T cells (Grueter et al., 2005; Yarmus et al.,2006), that Runx3 mediates TGFβ-induced expression ofCD103 in Vγ3+ thymocytes and in activated CD4+ T cells.Specifically, in developing Vγ3 thymocytes, Runx3 positivelyregulates CD103 expression during the transition fromimmature Vγ3med/HSAhigh to mature Vγ3high/HSAmed/low
thymocytes.It was previously reported that expression of CD103 in
cultured peripheral T cells is induced by TGFβ (Kilshaw andMurant, 1990; Parker et al., 1992; Robinson et al., 2001). Wefound, as did Grueter et al. (2005), that this induction ofCD103 expression in cultured T cells was mediated by Runx3.Of note, in naïve CD4+ T cells that express low level ofRunx3 (Fig. 4C and Taniuchi et al., 2002; Woolf et al., 2003),expression of Runx3 is up-regulated upon cell activation,concomitantly with TGFβ-dependent induction of CD103.This finding suggests that during Vγ3 T cell maturation invivo, Runx3 mediates the expression of CD103 in response toTGFβ signaling. It also raises the possibility that in fetalthymus either TGFβ1 and/or TGFβ2 can induce CD103expression, as both are produced in thymic epithelium (Lai etal., 1997; Takahama et al., 1994). This notion is supported byour finding that both TGFβ1 and TGFβ2 can induce CD103expression in cultured fetal Vγ3 thymocytes and by theobservation that TGFβ1-deficient mice are not devoid of skinDETCs (Borkowski et al., 1996), probably due to compen-sation by TGFβ2. No such compensation could occur in theKO mice because in the absence of Runx3, the KO Vγ3thymocytes respond neither to TGFβ1 nor to TGFβ2. As aresult, expression of CD103 diminishes leading to impairedmaturation of Vγ3 precursors. CD103 could also be involvedin Vγ3 cell proliferation, as E-cadherin expressing thymicepithelial cells are capable of inducing proliferation ofCD8+CD103+ but not of CD8+CD103− T cells (Kutlesa etal., 2002). Thus, diminished CD103 expression in KO Vγ3thymocytes could affect cell proliferation and thereby impactson DETCs development. Indeed, Runx3 KO CD8+ T cells, inwhich CD103 expression was diminished (Grueter et al.,2005; Yarmus et al., 2006), display a largely reducedproliferation rate (Taniuchi et al., 2002; Woolf et al., 2003).These data underscore the importance of CD103 for thedevelopment of DETC precursors in fetal thymus. But doesthe reduction in CD103 on Runx3 KO Vγ3 thymocytes fullyexplain the complete absence of skin DETCs in the KO mice?
Fetal Vγ3 DETC precursors depend on IL-2/IL-15 signalingthrough the IL-2Rβ for their development. When IL-2/IL-15signaling is lost, as in IL-2Rβ or IL-15-deficient mice, a markedreduction in mature Vγ3+ fetal thymocytes and skin DETCsoccurs (De Creus et al., 2002; Kawai et al., 1998). We show thatin Runx3 KO Vγ3 thymocytes the decreased expression of IL-2Rβwas associated with a lower response of the KO cells to IL-

712 E. Woolf et al. / Developmental Biology 303 (2007) 703–714
2 and IL-15 stimulation. It is possible that this lower response ofthe KO Vγ3 thymocytes to IL-2/IL-15 stimulation is, in part,due to the lower CD103 expression on these cells. Moreover,signaling through the IL-2Rβ and γ chains is also important formaintaining the viability of mature Vγ3 thymocytes in culture(De Creus et al., 2002; Leclercq et al., 1990a,b). Indeed, whencultured in the presence of IL-2, the proportion of apoptoticcells among KO Vγ3 thymocytes was significantly highercompared to WT. Additionally, as thymic epithelial cells at E14to E18 express E-cadherin (Lee et al., 1994), it is possible thatdiminished CD103-E-cadherin interactions in the KO contrib-ute to the apparent lower IL-2/IL-15 responsiveness. Finally, asIL-2Rβ expression on Vγ3 fetal thymocytes correlates withexpression of the skin homing chemokine receptor CCR10 onthese cells (Xiong et al., 2004), it is possible that the skin-homing capability of KO Vγ3 thymocytes was also compro-mised, which may further contribute to the complete absence ofskin DETCs in Runx3 KO mice.
In summary, we demonstrated that Runx3 plays an essentialrole in the development of DETC precursors in fetal thymus. Inthe absence of Runx3, expression of CD103 and IL-2Rβ in fetalVγ3 thymocytes diminished, which compromised the signalingthrough these surface receptors. As a result, the maturation andproliferation of DETC precursors in the thymus decreased andtheir apoptosis rate increased, leading to a complete lack of skinDETCs in the KO mice.
Acknowledgments
We thank Judith Chermesh and Rafi Saka for help in animalhusbandry, Dorit Nathan, Tamara Berkuzki for technicalassistance, Uri Gat and Eli Raveh for skin analysis, MadelonNoordegraafand for immunostaining of Langerin and JosephLotem and Steffen Jung for helpful discussions and insightfulcomments. The work was supported by grants from the PhilipMorris USA Inc. and Philip Morris International and the IsraelScience Foundation.
References
Asarnow, D.M., Kuziel, W.A., Bonyhadi, M., Tigelaar, R.E., Tucker, P.W.,Allison, J.P., 1988. Limited diversity of gamma delta antigen receptor genesof Thy-1+ dendritic epidermal cells. Cell 55, 837–847.
Asarnow, D.M., Goodman, T., LeFrancois, L., Allison, J.P., 1989. Distinctantigen receptor repertoires of two classes of murine epithelium-associated Tcells. Nature 341, 60–62.
Bangsow, C., Rubins, N., Glusman, G., Bernstein, Y., Negreanu, V.,Goldenberg, D., Lotem, J., Ben-Asher, E., Lancet, D., Levanon, D., Groner,Y., 2001. The RUNX3 gene-sequence, structure and regulated expression.Gene 279, 221–232.
Bennett, C.L., van Rijn, E., Jung, S., Inaba, K., Steinman, R.M., Kapsenberg,M.L., Clausen, B.E., 2005. Inducible ablation of mouse Langerhans cellsdiminishes but fails to abrogate contact hypersensitivity. J. Cell Biol. 169,569–576.
Borkowski, T.A., Letterio, J.J., Farr, A.G., Udey, M.C., 1996. A role forendogenous transforming growth factor beta 1 in Langerhans cell biology:the skin of transforming growth factor beta 1 null mice is devoid ofepidermal Langerhans cells. J. Exp. Med. 184, 2417–2422.
Brenner, O., Levanon, D., Negreanu, V., Golubkov, O., Fainaru, O., Woolf, E.,Groner, Y., 2004. Loss of Runx3 function in leukocytes is associated with
spontaneously developed colitis and gastric mucosal hyperplasia. Proc. Natl.Acad. Sci. U. S. A. 101, 16016–16021.
Cameron, E.R., Neil, J.C., 2004. The Runx genes: lineage-specific oncogenesand tumor suppressors. Oncogene 23, 4308–4314.
Cepek, K.L., Shaw, S.K., Parker, C.M., Russell, G.J., Morrow, J.S., Rimm, D.L.,Brenner, M.B., 1994. Adhesion between epithelial cells and T lymphocytesmediated by E-cadherin and the alpha E beta 7 integrin. Nature 372,190–193.
de Bruijn, M.F., Speck, N.A., 2004. Core-binding factors in hematopoiesis andimmune function. Oncogene 23, 4238–4248.
De Creus, A., Van Beneden, K., Stevenaert, F., Debacker, V., Plum, J., Leclercq,G., 2002. Developmental and functional defects of thymic and epidermal Vgamma 3 cells in IL-15-deficient and IFN regulatory factor-1-deficient mice.J. Immunol. 168, 6486–6493.
de Jong, J.L., Farner, N.L., Widmer, M.B., Giri, J.G., Sondel, P.M., 1996.Interaction of IL-15 with the shared IL-2 receptor beta and gamma csubunits. The IL-15/beta/gamma c receptor-ligand complex is less stablethan the IL-2/beta/gamma c receptor-ligand complex. J. Immunol. 156,1339–1348.
Ehlers, M., Laule-Kilian, K., Petter, M., Aldrian, C.J., Grueter, B., Wurch, A.,Yoshida, N., Watanabe, T., Satake, M., Steimle, V., 2003. Morpholinoantisense oligonucleotide-mediated gene knockdown during thymocytedevelopment reveals role for Runx3 transcription factor in CD4 silencingduring development of CD4−/CD8+ thymocytes. J. Immunol. 171,3594–3604.
El-Asady, R., Yuan, R., Liu, K., Wang, D., Gress, R.E., Lucas, P.J., Drachenberg,C.B., Hadley, G.A., 2005. TGF-{beta}-dependent CD103 expression byCD8(+) T cells promotes selective destruction of the host intestinalepithelium during graft-versus-host disease. J. Exp. Med. 201, 1647–1657.
Fainaru, O., Woolf, E., Lotem, J., Yarmus, M., Brenner, O., Goldenberg, D.,Negreanu, V., Bernstein, Y., Levanon, D., Jung, S., Groner, Y., 2004. Runx3regulates mouse TGF-beta-mediated dendritic cell function and its absenceresults in airway inflammation. EMBO J. 23, 969–979.
Fainaru, O., Shseyov, D., Hantisteanu, S., Groner, Y., 2005. Acceleratedchemokine receptor 7-mediated dendritic cell migration in Runx3 knockoutmice and the spontaneous development of asthma-like disease. Proc. Natl.Acad. Sci. U. S. A. 102, 10598–10603.
Girardi, M., Hayday, A.C., 2005. Immunosurveillance by gammadelta T cells:focus on the murine system. Chem. Immunol. Allergy 86, 136–150.
Girardi, M., Lewis, J., Glusac, E., Filler, R.B., Geng, L., Hayday, A.C., Tigelaar,R.E., 2002. Resident skin-specific gammadelta T cells provide local,nonredundant regulation of cutaneous inflammation. J. Exp. Med. 195,855–867.
Giri, J.G., Ahdieh, M., Eisenman, J., Shanebeck, K., Grabstein, K., Kumaki, S.,Namen, A., Park, L.S., Cosman, D., Anderson, D., 1994. Utilization of thebeta and gamma chains of the IL-2 receptor by the novel cytokine IL-15.EMBO J. 13, 2822–2830.
Grueter, B., Petter, M., Egawa, T., Laule-Kilian, K., Aldrian, C.J., Wuerch, A.,Ludwig, Y., Fukuyama, H., Wardemann, H., Waldschuetz, R., Moroy, T.,Taniuchi, I., Steimle, V., Littman, D.R., Ehlers, M., 2005. Runx3 regulatesintegrin alphaE/CD103 and CD4 expression during development of CD4−/CD8+ T cells. J. Immunol. 175, 1694–1705.
Havran, W.L., Allison, J.P., 1988. Developmentally ordered appearance ofthymocytes expressing different T-cell antigen receptors. Nature 335,443–445.
Havran, W.L., Allison, J.P., 1990. Origin of Thy-1+ dendritic epidermal cells ofadult mice from fetal thymic precursors. Nature 344, 68–70.
Hayday, A., Tigelaar, R., 2003. Immunoregulation in the tissues by gammadeltaT cells. Nat. Rev., Immunol. 3, 233–242.
Hayday, A., Theodoridis, E., Ramsburg, E., Shires, J., 2001. Intraepitheliallymphocytes: exploring the Third Way in immunology. Nat. Immunol. 2,997–1003.
Inoue, K., Ozaki, S., Shiga, T., Ito, K., Masuda, T., Okado, N., Iseda, T.,Kawaguchi, S., Ogawa, M., Bae, S.C., Yamashita, N., Itohara, S., Kudo, N.,Ito, Y., 2002. Runx3 controls the axonal projection of proprioceptive dorsalroot ganglion neurons. Nat. Neurosci. 5, 946–954.
Ito, K., Bonneville, M., Takagaki, Y., Nakanishi, N., Kanagawa, O., Krecko,E.G., Tonegawa, S., 1989. Different gamma delta T-cell receptors are

713E. Woolf et al. / Developmental Biology 303 (2007) 703–714
expressed on thymocytes at different stages of development. Proc. Natl.Acad. Sci. U. S. A. 86, 631–635.
Juhlin, L., Shelley, W.B., 1977. New staining techniques for the Langerhans cell.Acta Derm. Venereol. 57, 289–296.
Kabelitz, D., Marischen, L., Oberg, H.H., Holtmeier, W., Wesch, D., 2005.Epithelial defence by gamma delta T cells. Int. Arch. Allergy Immunol. 137,73–81.
Kaplan, D.H., Jenison, M.C., Saeland, S., Shlomchik, W.D., Shlomchik, M.J.,2005. Epidermal Langerhans cell-deficient mice develop enhanced contacthypersensitivity. Immunity 23, 611–620.
Kawai, K., Suzuki, H., Tomiyama, K., Minagawa, M., Mak, T.W., Ohashi, P.S.,1998. Requirement of the IL-2 receptor beta chain for the development ofVgamma3 dendritic epidermal T cells. J. Invest. Dermatol. 110, 961–965.
Kilshaw, P.J., Baker, K.C., 1988. A unique surface antigen on intraepitheliallymphocytes in the mouse. Immunol. Lett. 18, 149–154.
Kilshaw, P.J., Murant, S.J., 1990. A new surface antigen on intraepitheliallymphocytes in the intestine. Eur. J. Immunol. 20, 2201–2207.
Kilshaw, P.J., Murant, S.J., 1991. Expression and regulation of beta 7(beta p)integrins on mouse lymphocytes: relevance to the mucosal immune system.Eur. J. Immunol. 21, 2591–2597.
Kissenpfennig, A., Malissen, B., 2006. Langerhans cells—revisiting theparadigm using genetically engineered mice. Trends Immunol. 27,132–139.
Kissenpfennig, A., Ait-Yahia, S., Clair-Moninot, V., Stossel, H., Badell, E.,Bordat, Y., Pooley, J.L., Lang, T., Prina, E., Coste, I., Gresser, O., Renno, T.,Winter, N., Milon, G., Shortman, K., Romani, N., Lebecque, S., Malissen,B., Saeland, S., Douillard, P., 2005a. Disruption of the Langerin/CD207gene abolishes Birbeck granules without a marked loss of Langerhans cellfunction. Mol. Cell. Biol. 25, 88–99.
Kissenpfennig, A., Henri, S., Dubois, B., Laplace-Builhe, C., Perrin, P., Romani,N., Tripp, C.H., Douillard, P., Leserman, L., Kaiserlian, D., Saeland, S.,Davoust, J., Malissen, B., 2005b. Dynamics and function of Langerhanscells in vivo: dermal dendritic cells colonize lymph node areas distinct fromslower migrating Langerhans cells. Immunity 22, 643–654.
Kutlesa, S., Wessels, J.T., Speiser, A., Steiert, I., Muller, C.A., Klein, G., 2002.E-cadherin-mediated interactions of thymic epithelial cells with CD103+thymocytes lead to enhanced thymocyte cell proliferation. J. Cell Sci. 115,4505–4515.
Lafaille, J.J., DeCloux, A., Bonneville, M., Takagaki, Y., Tonegawa, S., 1989.Junctional sequences of T cell receptor gamma delta genes: implications forgamma delta T cell lineages and for a novel intermediate of V-(D)-J joining.Cell 59, 859–870.
Lai, Z.W., Hundeiker, C., Gleichmann, E., Esser, C., 1997. Cytokine geneexpression during ontogeny in murine thymus on activation of the arylhydrocarbon receptor by 2,3,7,8-tetrachlorodibenzo-p-dioxin. Mol. Pharma-col. 52, 30–37.
Leclercq, G., De Smedt, M., Plum, J., 1990a. Direct evidence for partial targetspecificity in lymphokine-activated killer thymocytes. Scand. J. Immunol.32, 373–379.
Leclercq, G., De Smedt, M., Tison, B., Plum, J., 1990b. Preferential proliferationof T cell receptor V gamma 3-positive cells in IL-2-stimulated fetalthymocytes. J. Immunol. 145, 3992–3997.
Leclercq, G., Plum, J., Nandi, D., De Smedt, M., Allison, J.P., 1993.Intrathymic differentiation of V gamma 3 T cells. J. Exp. Med. 178,309–315.
Leclercq, G., De Smedt, M., Plum, J., 1995. Cytokine dependence of V gamma3 thymocytes: mature but not immature V gamma 3 cells requireendogenous IL-2 and IL-7 to survive-evidence for cytokine redundancy.Int. Immunol. 7, 843–851.
Leclercq, G., Debacker, V., de Smedt, M., Plum, J., 1996. Differential effects ofinterleukin-15 and interleukin-2 on differentiation of bipotential T/naturalkiller progenitor cells. J. Exp. Med. 184, 325–336.
Lee, M.G., Sharrow, S.O., Farr, A.G., Singer, A., Udey, M.C., 1994. Expressionof the homotypic adhesion molecule E-cadherin by immature murinethymocytes and thymic epithelial cells. J. Immunol. 152, 5653–5659.
Lefrancois, L., Barrett, T.A., Havran, W.L., Puddington, L., 1994. Develop-mental expression of the alpha IEL beta 7 integrin on T cell receptor gammadelta and T cell receptor alpha beta T cells. Eur. J. Immunol. 24, 635–640.
Levanon, D., Groner, Y., 2004. Structure and regulated expression ofmammalian RUNX genes. Oncogene 23, 4211–4219.
Levanon, D., Brenner, O., Negreanu, V., Bettoun, D., Woolf, E., Eilam, R.,Lotem, J., Gat, U., Otto, F., Speck, N., Groner, Y., 2001. Spatial andtemporal expression pattern of Runx3 (Aml2) and Runx1 (Aml1) indicatesnon-redundant functions during mouse embryogenesis. Mech. Dev. 109,413–417.
Levanon, D., Bettoun, D., Harris-Cerruti, C., Woolf, E., Negreanu, V., Eilam, R.,Bernstein, Y., Goldenberg, D., Xiao, C., Fliegauf, M., Kremer, E., Otto, F.,Brenner, O., Lev-Tov, A., Groner, Y., 2002. The Runx3 transcription factorregulates development and survival of TrkC dorsal root ganglia neurons.EMBO J. 21, 3454–3463.
Levanon, D., Glusman, G., Bettoun, D., Ben-Asher, E., Negreanu, V., Bernstein,Y., Harris-Cerruti, C., Brenner, O., Eilam, R., Lotem, J., Fainaru, O.,Goldenberg, D., Pozner, A., Woolf, E., Xiao, C., Yarmus, M., Groner, Y.,2003. Phylogenesis and regulated expression of the RUNT domaintranscription factors RUNX1 and RUNX3. Blood Cells, Mol. Dis. 30,161–163.
Minagawa, M., Ito, A., Shimura, H., Tomiyama, K., Ito, M., Kawai, K., 2001.Homogeneous epithelial gamma delta T cell repertoire of the skin is shapedthrough peripheral selection. J. Dermatol. Sci. 25, 150–155.
Parker, C.M., Cepek, K.L., Russell, G.J., Shaw, S.K., Posnett, D.N., Schwarting,R., Brenner, M.B., 1992. A family of beta 7 integrins on human mucosallymphocytes. Proc. Natl. Acad. Sci. U. S. A. 89, 1924–1928.
Payer, E., Elbe, A., Stingl, G., 1991. Circulating CD3+/T cell receptor V gamma3+ fetal murine thymocytes home to the skin and give rise to proliferatingdendritic epidermal T cells. J. Immunol. 146, 2536–2543.
Pennington, D.J., Silva-Santos, B., Hayday, A.C., 2005. Gammadelta T celldevelopment-having the strength to get there. Curr. Opin. Immunol. 17,108–115.
Raveh, E., Cohen, S., Groner, Y., Gat, U., 2005. Runx3 is involved in hair shapedetermination. Dev. Dyn. 233, 1478–1487.
Robinson, P.W., Green, S.J., Carter, C., Coadwell, J., Kilshaw, P.J., 2001.Studies on transcriptional regulation of the mucosal T-cell integrinalphaEbeta7 (CD103). Immunology 103, 146–154.
Romani, N., Holzmann, S., Tripp, C.H., Koch, F., Stoitzner, P., 2003.Langerhans cells—dendritic cells of the epidermis. APMIS 111, 725–740.
Schmeissner, P.J., Xie, H., Smilenov, L.B., Shu, F., Marcantonio, E.E., 2001.Integrin functions play a key role in the differentiation of thymocytes invivo. J. Immunol. 167, 3715–3724.
Schon, M.P., Arya, A., Murphy, E.A., Adams, C.M., Strauch, U.G., Agace,W.W., Marsal, J., Donohue, J.P., Her, H., Beier, D.R., Olson, S.,Lefrancois, L., Brenner, M.B., Grusby, M.J., Parker, C.M., 1999. MucosalT lymphocyte numbers are selectively reduced in integrin alpha E(CD103)-deficient mice. J. Immunol. 162, 6641–6649.
Schon, M.P., Schon, M., Warren, H.B., Donohue, J.P., Parker, C.M., 2000.Cutaneous inflammatory disorder in integrin alphaE (CD103)-deficientmice. J. Immunol. 165, 6583–6589.
Schon, M.P., Schon, M., Parker, C.M., Williams, I.R., 2002. Dendritic epidermalT cells (DETC) are diminished in integrin alphaE(CD103)-deficient mice.J. Invest. Dermatol. 119, 190–193.
Sharp, L.L., Jameson, J.M., Cauvi, G., Havran, W.L., 2005a. Dendriticepidermal T cells regulate skin homeostasis through local production ofinsulin-like growth factor 1. Nat. Immunol. 6, 73–79.
Sharp, L.L., Jameson, J.M., Witherden, D.A., Komori, H.K., Havran, W.L.,2005b. Dendritic epidermal T-cell activation. Crit. Rev. Immunol. 25,1–18.
Shi, M.J., Stavnezer, J., 1998. CBF alpha3 (AML2) is induced by TGF-beta1 tobind and activate the mouse germline Ig alpha promoter. J. Immunol. 161,6751–6760.
Smith, T.J., Ducharme, L.A., Shaw, S.K., Parker, C.M., Brenner, M.B., Kilshaw,P.J., Weis, J.H., 1994. Murine M290 integrin expression modulated by mastcell activation. Immunity 1, 393–403.
Takahama, Y., Letterio, J.J., Suzuki, H., Farr, A.G., Singer, A., 1994. Earlyprogression of thymocytes along the CD4/CD8 developmental pathway isregulated by a subset of thymic epithelial cells expressing transforminggrowth factor beta. J. Exp. Med. 179, 1495–1506.
Tanaka, T., Takeuchi, Y., Shiohara, T., Kitamura, F., Nagasaka, Y., Hamamura,

714 E. Woolf et al. / Developmental Biology 303 (2007) 703–714
K., Yagita, H., Miyasaka, M., 1992. In utero treatment with monoclonalantibody to IL-2 receptor beta-chain completely abrogates development ofThy-1+ dendritic epidermal cells. Int. Immunol. 4, 487–491.
Taniuchi, I., Osato, M., Egawa, T., Sunshine, M.J., Bae, S.C., Komori, T., Ito, Y.,Littman, D.R., 2002. Differential requirements for Runx proteins in CD4repression and epigenetic silencing during T lymphocyte development. Cell111, 621–633.
Vaillant, F., Blyth, K., Andrew, L., Neil, J.C., Cameron, E.R., 2002. Enforcedexpression of Runx2 perturbs T cell development at a stage coincident withbeta-selection. J. Immunol. 169, 2866–2874.
Valladeau, J., Clair-Moninot, V., Dezutter-Dambuyant, C., Pin, J.J., Kissenpfennig,A., Mattei, M.G., Ait-Yahia, S., Bates, E.E., Malissen, B., Koch, F., Fossiez, F.,Romani, N., Lebecque, S., Saeland, S., 2002. Identification of mouse Langerin/CD207 in Langerhans cells and some dendritic cells of lymphoid tissues. J.Immunol. 168, 782–792.
van Wijnen, A.J., Stein, G.S., Gergen, J.P., Groner, Y., Hiebert, S.W., Ito, Y.,Liu, P., Neil, J.C., Ohki, M., Speck, N., 2004. Nomenclature for Runt-related(RUNX) proteins. Oncogene 23, 4209–4210.
Vermes, I., Haanen, C., Steffens-Nakken, H., Reutelingsperger, C., 1995. Anovel assay for apoptosis. Flow cytometric detection of phosphatidylserineexpression on early apoptotic cells using fluorescein labelled Annexin V.J. Immunol. Methods 184, 39–51.
Wang, D., Yuan, R., Feng, Y., El-Asady, R., Farber, D.L., Gress, R.E., Lucas,P.J., Hadley, G.A., 2004. Regulation of CD103 expression by CD8+ T cellsresponding to renal allografts. J. Immunol. 172, 214–221.
Woolf, E., Xiao, C., Fainaru, O., Lotem, J., Rosen, D., Negreanu, V., Bernstein,Y., Goldenberg, D., Brenner, O., Berke, G., Levanon, D., Groner, Y., 2003.Runx3 and Runx1 are required for CD8 T cell development duringthymopoiesis. Proc. Natl. Acad. Sci. U. S. A. 100, 7731–7736.
Xiong, N., Kang, C., Raulet, D.H., 2004. Positive selection of dendriticepidermal gammadelta T cell precursors in the fetal thymus determinesexpression of skin-homing receptors. Immunity 21, 121–131.
Yarmus, M., Woolf, E., Bernstein, Y., Fainaru, O., Negreanu, V., Levanon, D.,Groner, Y., 2006. Groucho/transducin-like Enhancer-of-split (TLE)-depen-dent and -independent transcriptional regulation by Runx3. Proc. Natl.Acad. Sci. U. S. A. 103, 7384–7389.
Copyright © 2022 FDOKUMEN




















