
Role of calcium and heat treatments in alleviating physiological changes induced by mechanical...
13
Postharvest Biology and Technology 34 (2004) 155–167 Role of calcium and heat treatments in alleviating physiological changes induced by mechanical damage in plum Mar´ ıa Serrano a , Domingo Mart´ ınez-Romero b , Salvador Castillo b , Fabián Guillén b , Daniel Valero b,∗ a Department of Applied Biology, University Miguel Hernández, Ctra. Beniel km. 3,2, 03312 Orihuela, Alicante, Spain b Department of Food Technology, University Miguel Hernández, Ctra. Beniel km. 3,2, 03312 Orihuela, Alicante, Spain Received 11 March 2004; accepted 9 May 2004 Abstract Plum (Prunus salicina Lindl. cv. Blackstar) fruit were harvested at the pre-climacteric stage of ripeness. After being treated with calcium (calcium chloride at 1 mM) or heat at 45 ◦ C (hot water dip immersion for 10 min), fruit were mechanically damaged at three equatorial zones with a given force of 50 N and then held at 20 ◦ C under controlled conditions. Results revealed that both treatments led to a reduction of the mechanical damage, and in turn alleviated the physiological responses that occurred in mechanically damaged plums. A reduction of “wound-induced” ethylene and respiration rates, which were detected just a few hours following mechanical damage, was observed for treated plums. During storage, mechanical damage advanced the climacteric ethylene and respiration peaks, while a delay in those plums previously treated either with calcium or heat was found. Thereafter, control plums exhibited an accumulation of free spermidine and abscisic acid in mechanically damaged flesh, which could be considered a physiological response to mechanical stress. These changes were minimised in calcium- and heat-treated damaged plums, indicating that these treatments would induce a significant resistance to mechanical damage in plum. © 2004 Elsevier B.V. All rights reserved. Keywords: Abscisic acid; Calcium; Ethylene; Heat; Mechanical damage; Plum; Polyamines 1. Introduction Mechanical damage is considered as a type of stress that occurs during harvesting and postharvest manip- ulation of fruit. This stress is accompanied by phys- iological and morphological changes affecting fruit quality (Miller, 1992). The development of mechanical damage results in cell wall rupture and loss of tissue in- ∗ Corresponding author. Tel.: +34 96 674 9743; fax: +34 96 674 9677. E-mail address: [email protected] (D. Valero). tegrity, such as has been observed in peaches (Vergano et al., 1991), tomatoes (Thiagu et al., 1993), pears and apples (Garc´ ıa et al., 1995), papayas (Quintana and Paull, 1993), and mandarins (Valero et al., 1998a), among others. In addition, some of the modifications that occur naturally during fruit ripening are acceler- ated after the mechanical damage, with severe con- sequences from the point of view of marketing and consumption. Moreover, mechanical damage causes physiological changes in ethylene, polyamines (PAs) and abscisic acid (ABA) (Mart´ ınez-Romero et al., 2004). 0925-5214/$ – see front matter © 2004 Elsevier B.V. All rights reserved. doi:10.1016/j.postharvbio.2004.05.004
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Role of calcium and heat treatments in alleviating physiological changes induced by mechanical...

Postharvest Biology and Technology 34 (2004) 155–167
Role of calcium and heat treatments in alleviating physiologicalchanges induced by mechanical damage in plum
Marıa Serranoa, Domingo Martınez-Romerob, Salvador Castillob,Fabián Guillénb, Daniel Valerob,∗
a Department of Applied Biology, University Miguel Hernández, Ctra. Beniel km. 3,2, 03312 Orihuela, Alicante, Spainb Department of Food Technology, University Miguel Hernández, Ctra. Beniel km. 3,2, 03312 Orihuela, Alicante, Spain
Received 11 March 2004; accepted 9 May 2004
Abstract
Plum (Prunus salicina Lindl. cv. Blackstar) fruit were harvested at the pre-climacteric stage of ripeness. After being treatedwith calcium (calcium chloride at 1 mM) or heat at 45◦C (hot water dip immersion for 10 min), fruit were mechanically damagedat three equatorial zones with a given force of 50 N and then held at 20◦C under controlled conditions. Results revealed thatboth treatments led to a reduction of the mechanical damage, and in turn alleviated the physiological responses that occurredin mechanically damaged plums. A reduction of “wound-induced” ethylene and respiration rates, which were detected just afew hours following mechanical damage, was observed for treated plums. During storage, mechanical damage advanced theclimacteric ethylene and respiration peaks, while a delay in those plums previously treated either with calcium or heat was found.Thereafter, control plums exhibited an accumulation of free spermidine and abscisic acid in mechanically damaged flesh, whichcould be considered a physiological response to mechanical stress. These changes were minimised in calcium- and heat-treateddamaged plums, indicating that these treatments would induce a significant resistance to mechanical damage in plum.© 2004 Elsevier B.V. All rights reserved.
Keywords: Abscisic acid; Calcium; Ethylene; Heat; Mechanical damage; Plum; Polyamines
1. Introduction
Mechanical damage is considered as a type of stressthat occurs during harvesting and postharvest manip-ulation of fruit. This stress is accompanied by phys-iological and morphological changes affecting fruitquality (Miller, 1992). The development of mechanicaldamage results in cell wall rupture and loss of tissue in-
∗ Corresponding author. Tel.:+34 96 674 9743;fax: +34 96 674 9677.
E-mail address: [email protected] (D. Valero).
tegrity, such as has been observed in peaches (Verganoet al., 1991), tomatoes (Thiagu et al., 1993), pears andapples (Garcıa et al., 1995), papayas (Quintana andPaull, 1993), and mandarins (Valero et al., 1998a),among others. In addition, some of the modificationsthat occur naturally during fruit ripening are acceler-ated after the mechanical damage, with severe con-sequences from the point of view of marketing andconsumption. Moreover, mechanical damage causesphysiological changes in ethylene, polyamines (PAs)and abscisic acid (ABA) (Martınez-Romero et al.,2004).
0925-5214/$ – see front matter © 2004 Elsevier B.V. All rights reserved.doi:10.1016/j.postharvbio.2004.05.004

156 M. Serrano et al. / Postharvest Biology and Technology 34 (2004) 155–167
Ethylene plays a key role in the fruit responseagainst mechanical stress, with stress increasingethylene levels immediately after mechanical dam-age (Maness et al., 1992). This ethylene is so-called“wound ethylene”, which was found to be proportionalto impact intensity in peaches (Martınez-Romeroet al., 2000). Thereafter, mechanical damage increasedethylene production and advanced the climacteric inapricots (Martınez-Romero et al., 2002).
In recent years, research on PAs has shown themto be involved in a wide array of processes in plants,ranging from triggering organogenesis to protectingagainst stress (Kakkar and Sawhney, 2002). In thissense, an accumulation of putrescine (Put), spermi-dine (Spd) and/or spermine (Spm) has been found inplants and fruit as a response to several stresses, suchas acidity, salinity, osmotic, mineral deficiency, CO2and chilling injury (see review byBouchereau et al.,1999). However, little is known about the changes inPA levels in response to external mechanical bruis-ing of fruit. Put accumulation was found in woundedgreen banana, probably through the action of woundethylene (Yoza et al., 1996). Accordingly, increasesin Spd and/or Spm have been found in mechanicallydamaged lemon, apricot, peach and plum, suggestingthat these PAs could be used as indicators or mark-ers of the mechanical stress (Martınez-Romero et al.,1999, 2000, 2002; Pérez-Vicente et al., 2002). In ad-dition to the free forms, PA conjugates occur as bothwater soluble and insoluble forms. The physiologicalrole of these conjugated amines is not well known,but they may act as storage forms of PAs, from whichthe free bases may be released during growth or theseconjugates may be transported as and when required(Martin-Tanguy, 1997).
In both climacteric and non-climacteric fruit, suchas lemons and apricots, an increase in ABA dur-ing storage has been shown, with concentrationsbeing significantly higher in mechanically dam-aged fruit, probably a response to mechanical stress(Martınez-Romero et al., 1999, 2002), as has been re-ported to occur against chilling injury stress (Serranoet al., 1997, 1998).
Calcium and heat treatments applied before stor-age have been shown to protect fruit against stress.Thus, exogenous calcium chloride treatment reducedchilling injury in squash (Wang, 1994) and mechani-cal damage in lemon (Martınez-Romero et al., 1999).
The same effects have been found after heat treatment(González-Aguilar et al., 1997; Lurie, 1998). In plum,the beneficial effect of calcium or heat treatment interms of prolonging storability, delaying the ripeningprocess and maintaining fruit quality has been recentlyreported (Valero et al., 2002). Since one of the maineffects of both treatments was a net increase in fruitfirmness, we wondered if these treatments could in-duce a greater resistance to mechanical damage. Thus,the role of postharvest treatments with calcium or heaton reducing the mechanical damage and the inducedphysiological responses during storage, focusing onthe metabolism of ethylene, polyamines and ABA, wasstudied.
2. Materials and methods
2.1. Plant material
Plum (Prunus salicina Lindl. cv. ‘Blackstar’) fruitwere harvested at the commercial ripening stage andmanually picked to avoid incidence of mechanicaldamage. In the laboratory, plums were selected by vi-sual external colour and weight, and then randomisedin one lot of 10 fruit (to establish fruit properties atharvest) and four lots of 50 fruit to perform treat-ments. One lot was treated with calcium (1 mMCaCl2) through pressure-infiltration with fruit in 5 lof solution, containing Tween-20 (2 g l−1), and ap-plying a pressure of 0.2 bar for 8 min. Another lotwas heat-treated by placing the fruit in a controlledhot water dip at 45◦C for 10 min, which was foundto be the optimum for plum. The remaining lots weretreated with distilled water with the same conditionsas for calcium treatment, and served as controls. Fruitwere placed on Kraft paper and allowed to dry beforestorage at 20◦C in a temperature-controlled chamberin permanent darkness and with a relative humidityof 90%. The following day, mechanical damage wasinflicted in calcium- and heat-treated plums, and inone of the control lot, while the fourth lot servedas a non-damaged control. Then, 10 fruit for eachtreatment were sampled to measure ethylene pro-duction and respiration rate during 4 h following theapplication of mechanical damage. Ten plums of theremaining lots were sampled after 4, 7, 11 and 14days of storage. They were weighed and the ethy-lene and CO2 production determined. The damaged

M. Serrano et al. / Postharvest Biology and Technology 34 (2004) 155–167 157
tissue (three flesh cylinders of 1 cm diameter for eachfruit) was individually frozen in liquid N2, milledand then stored at−20◦C until polyamines (free,conjugated-soluble and cell wall-bound) and ABAwere analysed. The same procedure was made fornon-damaged control fruit.
2.2. Mechanical damage application
For each fruit, the diameter (mm) was measured andthen mechanical damage with a compression force of50 N was applied, using a steel sphere of 20 mm diam-eter. The compression probe was mounted on a Uni-versal Assay Machine Lloyd, model LR5K (Lloyd In-struments, Segensworth, UK). Force was applied at arate of 20 mm min−1 until the 50 N force was achievedafter contacting the fruit, then held for 10 s. For eachfruit, three impact forces were applied at three differ-ent zones of the equatorial plane separated by 120◦.A bevelled holder prevented bruising of the oppositeside. Fruit deformation was measured in mm.
2.3. Ethylene and CO2 production rates
Ethylene and CO2 production were measured byplacing each fruit in a 0.5 l glass jar hermeticallysealed with a rubber stopper for 1 h. One ml ofthe holder atmosphere was withdrawn with a gassyringe, and the ethylene was quantified using aGC (Hewlett-PackardTM 5890A, Wilmington, DE)equipped with a FID detector. Results were expressedas nl g−1 h−1. For respiration rate determination, an-other sample of 1 ml of the same atmosphere was with-drawn and CO2 quantified using a GC (ShimadzuTM
GC 14A, Kyoto, Japan) with a thermal conductivitydetector. Results were expressed as mg kg−1 h−1.
2.4. Free, conjugated-soluble, and cell wall-boundpolyamine analyses
For each fruit, 2 g of frozen tissue was extractedwith 10 ml of 5% cold perchloric acid. The ho-mogenate was then centrifuged (24,000× g, 20 min,4◦C). A 2 ml aliquot from the supernatant served todetermine free polyamines. For conjugated-solublepolyamine analysis, a 2 ml aliquot was taken from thesupernatant, and then hydrolysed with 2 ml 12N HClat 116◦C during 16 h. For cell wall-bound polyamines,
the pellet was re-dissolved in 2 ml of perchloric acid,and then hydrolysed as above. Hydrolysed polyamines(conjugated-soluble and cell wall-bound) were fil-tered through a glass-wool filter, immediately driedunder N2, and then re-dissolved in 2 ml of perchloricacid. Polyamines (free, conjugated-soluble, and cellwall-bound) were benzoylated, and analysed by HPLCaccording toSerrano et al. (1997). A relative calibra-tion procedure was used to determine the polyaminesin the samples, using 1,6 hexanediamine as the inter-nal standard and polyamine concentrations rangingfrom 1 to 500 nM. The calibration curves were:y= 0.048x − 250.71,r2 = 0.99 (for Put);y = 0.039x− 291.63, r2 = 0.99 (for Spd); andy = 0.045x −180.39,r2 = 0.99 (for Spm). Results were expressedas nmol g−1 FW, and represent the mean± S.E. ofduplicate measurements in each of ten fruit.
2.5. Abscisic acid determination
Abscisic acid was extracted from 0.5 g frozen tis-sue with 10 ml solution of 80% acetone containingbutylated hydroxytoluene at 100 mg l−1 and citricacid at 0.5 g l−1. The extracts were centrifuged for5 min at 3000× g and the supernatant was dilutedwith a 50 mM Tris buffer (pH= 7.8) containing1 mM MgCl2 and 150 mM NaCl, and then quantifiedby an enzyme-linked immunosorbent assay (ELISA)(Weiler, 1980), using an IgG monoclonal antibody(Idexx, Inc., San Bruno, CA, USA). Synthesis ofABA–bovine serum albumin conjugate was carriedout as described byWeiler (1980). ABA content wasestimated from the standard curve prepared for eachparticular plate using a spectrophotometer StarFax2100 (Awareness Technology, Inc., Palm City, FL,USA). The absorbance was fixed at 405 nm. Extrac-tions were made independently from each fruit, andfor each extract, three dilutions were prepared in du-plicate and at least two of them fell on the standardcurve. All manipulations were carried out in dimlight. The ABA levels were consistent with the appro-priate dilution. No interference from impurities wasdetected when ABA standards were added to dilutedextracts of fruit.
2.6. Statistical design
A two-factor ANOVA was made to determine theeffect of treatments and storage time on ethylene, CO2,

158 M. Serrano et al. / Postharvest Biology and Technology 34 (2004) 155–167
polyamines and ABA. Mean comparisons were per-formed using the Student–Newman–Keuls (SNK) testto examine differences (P < 0.05) between treatmentsand storage time. All analyses were performed withSAS’s personal computer software package version6.03 (SAS Institute, 1988).
3. Results
3.1. Effect of treatments on mechanical damage
All fruit were mechanically damaged with the sameload (50 N), but response of fruit tissue was dependenton treatment. Thus, deformation caused by the forcethat induced mechanical damage was significantlyhigher in control plums (6.33± 0.24 mm) than inheat-treated (5.44± 0.27 mm) or calcium-treated ones(4.64± 0.14 mm), which showed the lowest deforma-tion. In addition, damaged control plums exhibited ahigher intensity of flesh browning compared to thosetreated either with calcium or heat (data not shown).
3.2. Ethylene production and respiration rates
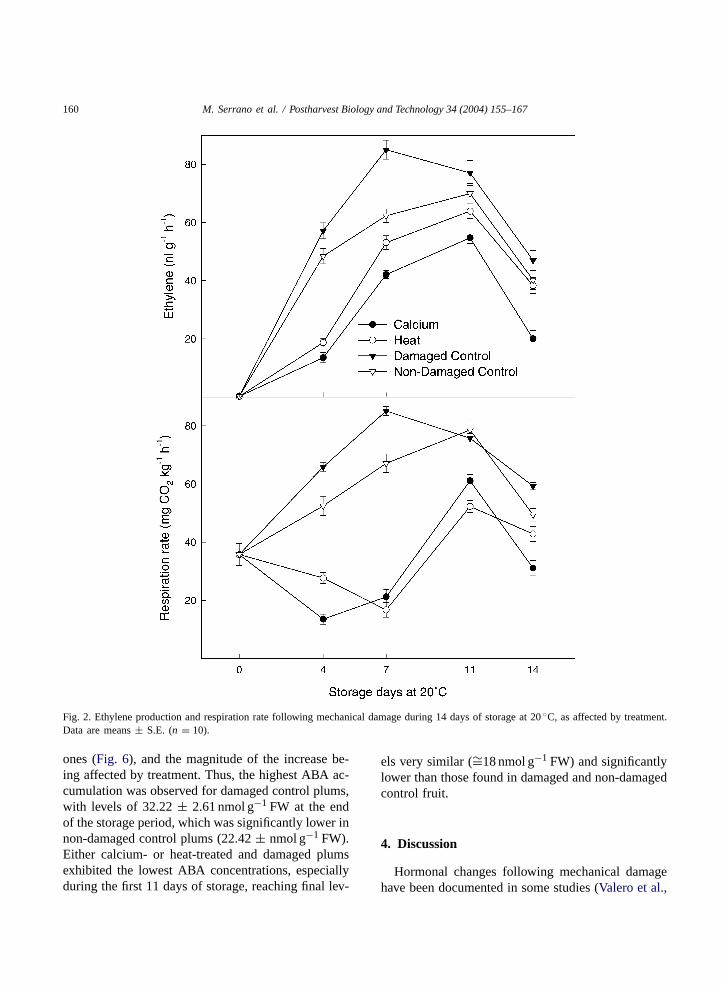
Ethylene production and respiration rates increasedfollowing mechanical damage within the first 4 h(Fig. 1), reaching peaks of 1.23± 0.04 nl g−1 h−1 forethylene and 57.76± 1.36 mg CO2 kg−1 h−1 for res-piration in damaged control plums, while no increasewas detected for non-damaged control fruit. In thosefruit that were previously treated either with calciumor heat, the increases of both ethylene productionand respiration rates were significantly lower than indamaged control fruit, especially in calcium-treatedplums. Similarly, during prolonged storage at 20◦C,the same behaviour was detected (Fig. 2), with thehighest ethylene production and respiration rates be-ing found in damaged control plums, with levels of85.02 ± 3.28 nl g−1 h−1 and 85.08± 1.57 mg CO2kg−1 h−1, respectively, at the maximum climactericpeak, which was reached at day 7. These maximawere achieved at day 11 of storage in plum fromthe remaining treatments, with levels of 69.94±3.50 nl g−1 h−1 for ethylene and 78.58± 3.12 mgCO2 kg−1 h−1 for respiration rate in non-damagedcontrol plums, and significantly lower in damagedcalcium- and heat-treated plums.
3.3. Weight loss
All fruit (damaged and non-damaged) showed a sig-nificant increase in weight loss during storage (Fig. 3).The final weight losses were: 16.53± 1.15% in dam-aged control fruit and 13.88± 1.04% in non-damagedcontrol fruit. For those plums treated with calcium orheat before being mechanically damaged, a signifi-cant reduction in weight loss was observed, with finallosses of 8.19± 0.96% and 9.65± 0.92%, respec-tively.
3.4. Free, conjugated-soluble and cell wall-boundpolyamines
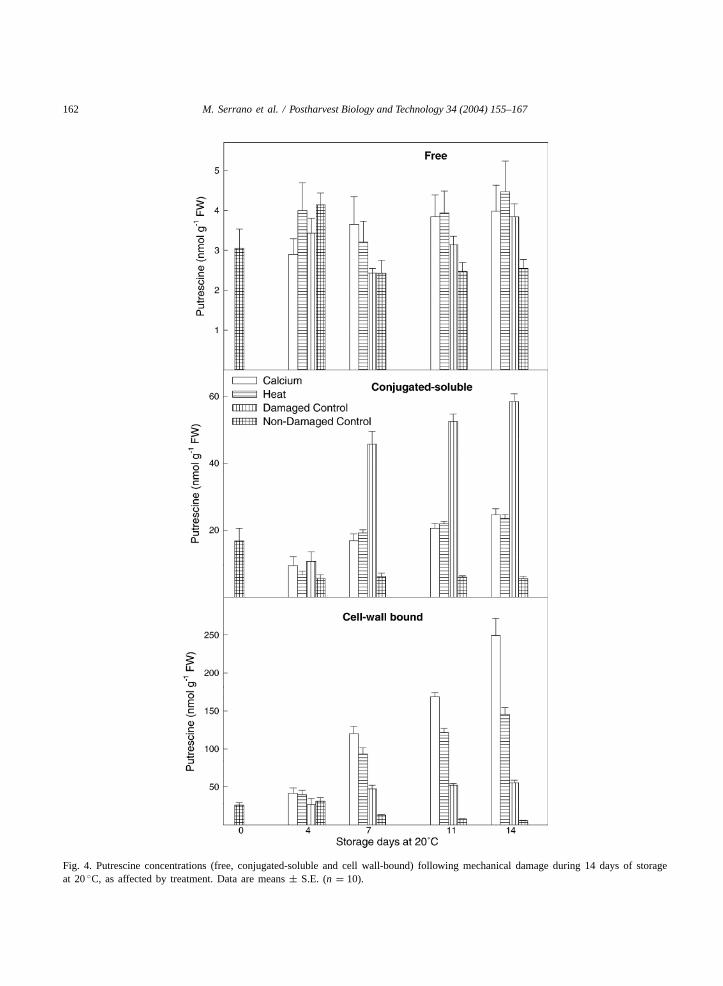
Free Put levels at harvest were 3.05± 0.48 nmolg−1 FW and did not significantly change duringstorage at 20◦C, nor by mechanical damage or thepostharvest treatment applied (Fig. 4). Conversely,an accumulation of conjugated-soluble Put was ob-served in mechanically damaged control fruit, reach-ing levels of 58.47± 2.24 nmol g−1 FW at day 14 ofstorage. This effect was reduced in those fruit thatwere previously treated with calcium or heat (whichreached a final Put concentration ca. 24 nmol g−1 FW),while in non-damaged control fruit, a significantdiminution was observed, reaching levels of 2.55± 0.22 nmol g−1 FW at the end of storage. Cellwall-bound Put increased significantly over storage incalcium- or heat-damaged plum, reaching the great-est concentrations after 14 days of storage, 249.23±22.78 nmol g−1 FW and 145.41± 8.85 nmol g−1 FW,respectively. This increase was also detected indamaged control plums but with lower magnitude(55.42 ± nmol g−1 FW). However, in non-damagedcontrol plums, cell wall-bound Put diminished dur-ing storage and the lowest concentrations (5.41±0.55 nmol g−1 FW) were found.
Mechanical damage induced an accumulation offree Spd, especially in damaged control plums, withconcentrations of 388.48± 37.57 nmol g−1 FW atday 7 of storage. However, in those damaged plumspreviously treated with calcium or heat, this increasewas significantly lower, while in non-damaged con-trol plums, free Spd decreased over storage andthe lowest levels were found (Fig. 5). Conversely,in calcium-treated damaged plum, a sharp increasein conjugated-soluble Spd was found over storage,

M. Serrano et al. / Postharvest Biology and Technology 34 (2004) 155–167 159
Fig. 1. Ethylene production and respiration rate following mechanical damage during 4 h of storage at 20◦C, as affected by treatment.Data are means± S.E. (n = 10).
reaching levels of 65.47± 4.68 nmol g−1 FW after 14days, while in heat-treated damaged fruit, a sharp in-crease was observed for cell wall-bound Spd, with lev-els of 55.63± 2.47 nmol g−1 FW at the end of storage.In control plums, for both damaged and non-damagedones, a reduction of both conjugate-soluble and cellwall-bound Spd was shown during storage, especiallyfor those plums that were not mechanically injured.Spm levels in ‘Blackstar’ plum were very low and no
changes were observed during storage, independentof whether fruit were treated and/or damaged (datanot shown).
3.5. Abscisic acid levels
ABA levels increased progressively over storagefrom the initial concentrations (2.99± 0.31 nmol g−1
FW) for all fruit, both damaged and non-damaged
160 M. Serrano et al. / Postharvest Biology and Technology 34 (2004) 155–167
Fig. 2. Ethylene production and respiration rate following mechanical damage during 14 days of storage at 20◦C, as affected by treatment.Data are means± S.E. (n = 10).
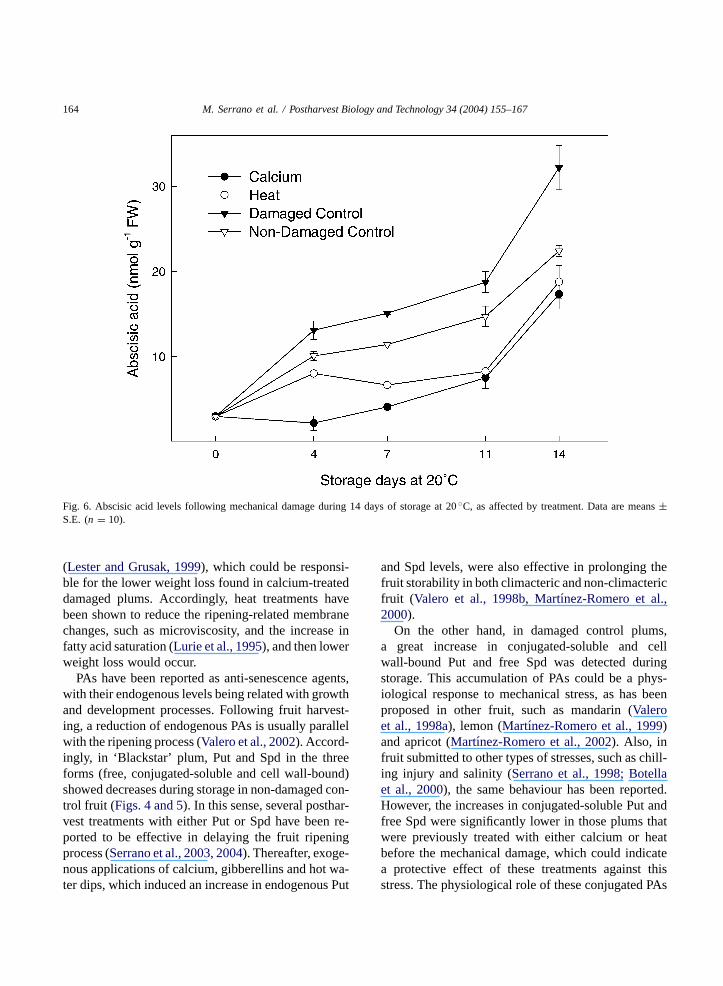
ones (Fig. 6), and the magnitude of the increase be-ing affected by treatment. Thus, the highest ABA ac-cumulation was observed for damaged control plums,with levels of 32.22± 2.61 nmol g−1 FW at the endof the storage period, which was significantly lower innon-damaged control plums (22.42± nmol g−1 FW).Either calcium- or heat-treated and damaged plumsexhibited the lowest ABA concentrations, especiallyduring the first 11 days of storage, reaching final lev-
els very similar (∼=18 nmol g−1 FW) and significantlylower than those found in damaged and non-damagedcontrol fruit.
4. Discussion
Hormonal changes following mechanical damagehave been documented in some studies (Valero et al.,

M. Serrano et al. / Postharvest Biology and Technology 34 (2004) 155–167 161
Fig. 3. Weight loss change following mechanical damage during 14 days of storage at 20◦C, as affected by treatment. Data are means±S.E. (n = 10).
1998a; Martınez-Romero et al., 1999, 2000, 2002).However, whether these changes act as a protectivemechanism against the mechanical stress or are a fruitphysiological response to the mechanical damage isstill unclear. In the present work, ‘Blackstar’ plum ex-hibited a net increase in ethylene production follow-ing mechanical damage, which was detectable after afew hours of storage (Fig. 1). This ethylene productionis so-called “wound-inducible” ethylene and it seemsto be a general response of plant tissues to injuriesthrough an activation of ACC synthase and ACC ox-idase (Kato et al., 2000; Karakurt and Huber, 2003).However, in both calcium- and heat-treated damagedplums, this increase in wound ethylene was signif-icantly lower, especially for calcium-treated plums.Similarly, respiration rate increased also as a con-sequence of the mechanical damage within the firsthours of storage in control fruit, this response beingalleviated in calcium- or heat-treated damaged plums(Fig. 1). This higher respiration rate has been attributedto the fact that in injured tissues, the oxygen diffusesmore rapidly into the interior of the cells, increasingtheir metabolic activity (Watada et al., 1996). Also,the ethylene action has been reported as responsiblefor the high respiration rate in wounded plant tissues(Brecht, 1995). Thus, both treatments were effectivein protecting fruit tissue against mechanical damage
since lower physiological responses were obtained.This effect could be attributed to the higher fruit firm-ness and the lower fruit deformation found in treatedfruit when mechanical damage was performed.
In addition, during prolonged storage, the climac-teric peak of both ethylene production and respira-tion rate occurred earlier and reached higher valuesin damaged control fruit than in non-damaged ones(Fig. 2), showing an advancement of the ripening pro-cess, as has been reported in damaged apricot and plum(Martınez-Romero et al., 2002; Pérez-Vicente et al.,2002). However, in those fruit that were treated ei-ther with calcium or heat before mechanical damage,a significant reduction of ethylene production and res-piration rate was detected, with values even lower thanthose observed for non-damaged control fruit. An-other positive effect of the treatments was the reducedweight loss compared with that found in control plums,both damaged and non-damaged (Fig. 3). Weight lossis a consequence of fruit dehydration due to changesin surface transfer resistance to water vapour, in respi-ration rate, and the occurrence of small fissures con-necting the internal and external atmospheres (Woods,1990). In this sense, calcium dip applications haveshown to be effective in terms of membrane function-ality and integrity maintenance, with lower losses ofphospholipids and proteins and reduced ion leakage
162 M. Serrano et al. / Postharvest Biology and Technology 34 (2004) 155–167
Fig. 4. Putrescine concentrations (free, conjugated-soluble and cell wall-bound) following mechanical damage during 14 days of storageat 20◦C, as affected by treatment. Data are means± S.E. (n = 10).

M. Serrano et al. / Postharvest Biology and Technology 34 (2004) 155–167 163
Fig. 5. Spermidine concentrations (free, conjugated-soluble and cell wall-bound) following mechanical damage during 14 days of storageat 20◦C, as affected by treatment. Data are means± S.E. (n = 10).
164 M. Serrano et al. / Postharvest Biology and Technology 34 (2004) 155–167
Fig. 6. Abscisic acid levels following mechanical damage during 14 days of storage at 20◦C, as affected by treatment. Data are means±S.E. (n = 10).
(Lester and Grusak, 1999), which could be responsi-ble for the lower weight loss found in calcium-treateddamaged plums. Accordingly, heat treatments havebeen shown to reduce the ripening-related membranechanges, such as microviscosity, and the increase infatty acid saturation (Lurie et al., 1995), and then lowerweight loss would occur.
PAs have been reported as anti-senescence agents,with their endogenous levels being related with growthand development processes. Following fruit harvest-ing, a reduction of endogenous PAs is usually parallelwith the ripening process (Valero et al., 2002). Accord-ingly, in ‘Blackstar’ plum, Put and Spd in the threeforms (free, conjugated-soluble and cell wall-bound)showed decreases during storage in non-damaged con-trol fruit (Figs. 4 and 5). In this sense, several posthar-vest treatments with either Put or Spd have been re-ported to be effective in delaying the fruit ripeningprocess (Serrano et al., 2003, 2004). Thereafter, exoge-nous applications of calcium, gibberellins and hot wa-ter dips, which induced an increase in endogenous Put
and Spd levels, were also effective in prolonging thefruit storability in both climacteric and non-climactericfruit (Valero et al., 1998b, Martınez-Romero et al.,2000).
On the other hand, in damaged control plums,a great increase in conjugated-soluble and cellwall-bound Put and free Spd was detected duringstorage. This accumulation of PAs could be a phys-iological response to mechanical stress, as has beenproposed in other fruit, such as mandarin (Valeroet al., 1998a), lemon (Martınez-Romero et al., 1999)and apricot (Martınez-Romero et al., 2002). Also, infruit submitted to other types of stresses, such as chill-ing injury and salinity (Serrano et al., 1998; Botellaet al., 2000), the same behaviour has been reported.However, the increases in conjugated-soluble Put andfree Spd were significantly lower in those plums thatwere previously treated with either calcium or heatbefore the mechanical damage, which could indicatea protective effect of these treatments against thisstress. The physiological role of these conjugated PAs

M. Serrano et al. / Postharvest Biology and Technology 34 (2004) 155–167 165
is not well known, but several hypotheses have beenclaimed. The main conjugates are through bindingwith cinnamic acids, and could serve as a pool of freePAs when necessary (Martin-Tanguy, 1997). Also,conjugations of PAs could act as a protective mecha-nism to abiotic stresses (Bouchereau et al., 1999), andthen calcium treatment would alleviate the mechanicaldamage through the accumulation of conjugated PAs.
Thereafter, the great accumulation of conjugated-soluble Spd in those damaged plums that were previ-ously treated with calcium, as compared to damagedcontrol fruit, could be a consequence of the displace-ment of the Spd bound at the cell-wall level by cal-cium ions. Thus, calcium could compete with Spd inthe linkage to the negative groups of carboxylic acidsof pectic substances (Valero et al., 2002). It couldbe suggested that higher exogenous calcium appliedwould lead to a high liberation of bound Spd, and thengreater levels of conjugated-soluble Spd would be ex-pected. However, this does not seem to occur betweenPut and Ca2+, since cell wall-bound Put levels werehigher in damaged calcium-treated plum than in dam-aged control ones. These findings are in agreementwith Messiaen et al. (1997), who postulated that ionselectivity of the cell walls for PAs followed the se-quence: Spm4+ > Spd3+ ≈ Ca2+ > Pu2+.
In heat-treated damaged plums, an accumulation incell wall-bound Put and Spd occurred during stor-age. In this sense, heat treatments have been found toincrease pectinmethylesterase (PME) activity (Javeriet al., 1991), which cleaves the methoxyl groups frommethylated galacturonic acid residues in pectin, withgeneration of non-methylated pectic acids, which con-tain newly available carboxyl groups to bind withmore cations, such as Put or Spd. Accordingly, it hasbeen also observed that exogenous calcium signifi-cantly stimulated PME activity (Leiting and Wicker,1997), which could similarly explain the increases inboth cell wall-bound Put and Spd. In addition, sinceincreases in cell wall-bound PAs have been reported asa mechanism of protection against stress (Bouchereauet al., 1999; Pérez-Vicente et al., 2002), the higherincreases found in cell wall-bound Put and Spd, forboth calcium- and heat-treated plums, could indicate astimulation of defence mechanisms induced by thesetreatments.
The plant hormone ABA plays a predominant rolein the conversion of environmental signals from sev-
eral stresses into changes in plant gene expression.An increase in ABA levels has been obtained uponwounding, not only in damaged zones, but also in thesurrounding non-damaged tissues (Herde et al., 1999).Accordingly, in ‘Blackstar’ plum, ABA concentrationincreased during storage, with this increase being sig-nificantly higher in control-damaged fruit than in thosepreviously treated with calcium or heat. Therefore,ABA accumulation could be considered a physiolog-ical response of plum tissues to mechanical damage.The higher ABA concentrations found in damagedcontrol plums were correlated with the accumulationof free Spd, the predominant PA. In this sense, thepresence of ABA has been shown to increase the PAvalues and to modulate the activities of the enzymesresponsible for Put synthesis, ornithine and argininedescarboxylase (Arusisano et al., 1993). However, inother types of stress, such as chilling injury, it has beenpostulated that ABA and PAs play independent rolesas protective agents against cold-induced membranedamage, with ABA being the major mediator in theresponse to this stress (Kim et al., 2002). Thus, morestudies are necessary to gain a better knowledge aboutthe interaction between ABA and PAs.
In conclusion, calcium or hot water treatments in-duced a significant resistance to mechanical damage inplum, and in turn alleviated the physiological changesthat usually occurred in damaged tissues, such as in-creases in ethylene production, in respiration rate, andin the concentrations of PAs and ABA. The accumu-lations of free Spd and ABA could be considered asphysiological markers of mechanical damage, one ofthe less known types of stress.
Acknowledgements
CYCYT has funded this work. Project ALI99-1058-C02-02.
References
Arusisano, N., Bertani, A., Mattana, M., Reggiani, R., 1993.Abscisic acid induced stress-like polyamine pattern in wheatseedlings, and its reversal by potassium ions. Physiol. Plant.89, 687.
Botella, M.A., Del Amor, F., Amorós, A., Serrano, M.,Martınez, V., Cerdá, A., 2000. Polyamine, ethylene and

166 M. Serrano et al. / Postharvest Biology and Technology 34 (2004) 155–167
other physico-chemical parameters in tomato (Lycopersiconesculentum) fruits as affected by salinity. Physiol. Plant. 109,428–434.
Bouchereau, A., Aziz, A., Larher, F., Martin-Tanguy, J., 1999.Polyamines and environmental challenges: recent development.Plant Sci. 140, 103–125.
Brecht, J.K., 1995. Physiology of lightly processed fruits andvegetables. HortScience 30, 18–22.
Garcıa, J.L., Ruız-Altisent, M., Barreiro, P., 1995. Factorsinfluencing mechanical properties and bruise susceptibility ofapples and pears. J. Agric. Eng. Res. 61, 11–18.
González-Aguilar, G.A., Zacarıas, L., Mulas, M., Lafuente, M.T.,1997. Temperature and duration of water dips influence chillinginjury, decay and polyamine content in ‘Fortune’ mandarins.Postharvest Biol. Technol. 12, 61–69.
Herde, O., Cortés, H.P., Wasternack, C., Willmitzer, L., Fisahn,J., 1999. Electric signalling andPin2 gene expression ondifferent abiotic stimuli depend on a distinct threshold levelof endogenous abscisic acid in several abscisic acid-deficienttomato mutants. Plant Physiol. 119, 213–218.
Javeri, H., Toledo, R., Wicker, L., 1991. Vacuum infusion ofcitrus pectinmethylesterase and calcium effects on firmness ofpeaches. J. Food Sci. 56, 739–742.
Kakkar, R.K., Sawhney, V.K., 2002. Polyamine research in plants– a changing perspective. Physiol. Plant. 116, 281–292.
Karakurt, Y., Huber, D.J., 2003. Activities of several membraneand cell-wall hydrolases, ethylene biosynthetic enzymes,and cell-wall polyuronide degradation during low-temperaturestorage of intact and fresh-cut papaya (Carica papaya) fruit.Postharvest Biol. Technol. 28, 219–229.
Kato, M., Hayakawa, Y., Hyodo, H., Ikoma, Y., Yano, M.,2000. Wound-induced ethylene synthesis and expressionof 1-aminocyclopropane-1-carboxylate (ACC) synthase, ACCoxidase, phenylalanine ammonia-lyase, and peroxidase inwounded mesocarp tissue ofCucurbita maxima. Plant CellPhysiol. 41, 440–447.
Kim, T.E., Kim, S.K., Han, T.J., Lee, J.S., Chang, S.C., 2002.ABA and polyamines act independently in primary leaves ofcold-stressed tomato (Lycopersicon esculentum). Physiol. Plant.115, 370–376.
Leiting, V.A., Wicker, L., 1997. Inorganic cations and polyaminesmoderate pectinesterase activity. J. Food Sci. 62, 253–255.
Lester, G.E., Grusak, M.A., 1999. Postharvest application ofcalcium and magnesium to honeydew and netted muskmelons:effects on tissue ion concentrations, quality and senescence. J.Am. Soc. Hort. Sci. 124, 545–552.
Lurie, S., 1998. Postharvest heat treatments. Postharvest Biol.Technol. 14, 257–269.
Lurie, S., Othman, S., Borochov, A., 1995. Effects of heat treatmenton plasma membrane of apple fruit. Postharvest Biol. Technol.5, 29–38.
Maness, N.O., Brusewitz, G.H., McCollum, T.G., 1992.Impact bruise resistance comparison among peach cultivars.HortScience 27, 1008–1011.
Martin-Tanguy, J., 1997. Conjugated polyamines and reproductivedevelopment: biochemical, molecular and physiologicalapproaches. Physiol. Plant. 100, 675–688.
Martınez-Romero, D., Serrano, M., Carbonell, A., Burgos, L.,Riquelme, F., Valero, D., 2002. Effects of postharvestputrescine treatment on extending shelf life and reducingmechanical damage in apricot. J. Food Sci. 67, 1706–1712.
Martınez-Romero, D., Serrano, M., Carbonell, A., Castillo, S.,Riquelme, F., Valero, D., 2004. Mechanical damage during fruitpost-harvest handling: technical and physiological implications,in: Dris, R., Jain, S.M. (Ed.), Production Practises andQuality Assessment of Food Crops, vol. 3. Quality Handlingand Evaluation. Kluwer Academic Publishers, Dordrecht,pp. 233–252.
Martınez-Romero, D., Valero, D., Serrano, M., Riquelme, F., 1999.Effects of postharvest putrescine and calcium treatments onreducing mechanical damage and polyamines and ABA levelsduring lemon storage. J. Sci. Food Agric. 79, 1589–1595.
Martınez-Romero, D., Valero, D., Serrano, M., Burló, F., Carbonell,A., Burgos, L., Riquelme, F., 2000. Exogenous polyamines andgibberellic acid effects on peach (Prunus persica L.) storabilityimprovement. J. Food Sci. 65, 288–294.
Messiaen, J., Cambier, P., Van Custem, P., 1997. Polyamines andpectins. I. Ion exchange and selectivity. Plant Physiol. 113,387–395.
Miller, A.R., 1992. Physiology, biochemistry and detectionof bruising (mechanical stress) in fruits and vegetables.Postharvest News Inform. 3, 53–58.
Pérez-Vicente, A., Martınez-Romero, D., Carbonell, A., Serrano,M., Riquelme, F., Guillén, F., Valero, D., 2002. Role ofpolyamines in extending shelf life and the reduction ofmechanical damage during plum (Prunus salicina Lindl.)storage. Postharvest Biol. Technol. 25, 25–32.
Quintana, M.E.G., Paull, R.E., 1993. Mechanical injury duringpostharvest handling of ‘Solo’ papaya fruit. J. Am. Soc. Hort.Sci. 118, 618–622.
SAS Institute, SAS/STAT User’s Guide, Release 6.03, SAS Inst.,Cary, NC, 1988.
Serrano, M., Martınez-Madrid, M.C., Pretel, M.T., Riquelme, F.,Romojaro, F., 1997. Modified atmosphere packaging minimizesincreases in putrescine and abscisic acid levels caused bychilling injury in pepper fruit. J. Agric. Food Chem. 45, 1668–1672.
Serrano, M., Martınez-Madrid, M.C., Romojaro, F., Riquelme,F., 1998. CO2 treatment of zucchini squash reduceschilling-induced physiological changes. J. Agric. Food Chem.46, 2465–2468.
Serrano, M., Martınez-Romero, D., Guillén, F., Valero, D., 2003.Effects of exogenous putrescine on improving shelf life of fourplum cultivars. Postharvest Biol. Technol. 30, 259–271.
Serrano, M., Martınez-Romero, D., Zuzunaga, M., Riquelme,F., Valero, D., 2004. Calcium, polyamine and gibberellintreatments to improve postharvest fruit quality, in: Dris, R.,Jain, S.M. (Eds.), Production Practises and Quality Assessmentof Food Crops, vol. 4. Postharvest Treatment and Technology.Kluwer Academic Publishers, Dordrecht, pp. 55–68.
Thiagu, R., Chand, N., Ramana, K.V.R., 1993. Evolution ofmechanical characteristics of tomatoes of two varieties duringripening. J. Sci. Food Agric. 62, 175–183.

M. Serrano et al. / Postharvest Biology and Technology 34 (2004) 155–167 167
Valero, D., Martınez-Romero, D., Serrano, M., Riquelme,F., 1998a. Polyamine response to external mechanicalbruising in two mandarin cultivars. HortScience 33, 1220–1223.
Valero, D., Martınez-Romero, D., Serrano, M., Riquelme, F.,1998b. Postharvest gibberellin and heat treatment effects onpolyamines, abscisic acid and firmness in lemons. J. Food Sci.63, 611–615.
Valero, D., Martınez-Romero, D., Serrano, M., 2002. The role ofpolyamines in the improvement of shelf life of fruits. TrendsFood Sci. Technol. 13, 228–234.
Vergano, P.J., Testin, R.F., Newall, J.W.C., 1991. Distinguishingamong bruises in peaches caused by impact, vibration, andcompression. J. Food Qual. 14, 285–289.
Wang, C.Y., 1994. Combined treatment of heat shock and lowtemperature conditioning reduces chilling injury in zucchinisquash. Postharvest Biol. Technol. 4, 65–73.
Watada, A.E., Ko, N.P., Minott, D.A., 1996. Factors affectingquality of fresh-cut horticultural products. Postharvest Biol.Technol. 9, 115–125.
Weiler, E.W., 1980. Radioimmunoassay for the differential anddirect analysis of free and conjugated abscisic in plant extracts.Planta 148, 262–272.
Woods, J.L., 1990. Moisture loss from fruits and vegetables.Postharvest News Inform. 1, 195–199.
Yoza, K., Takeda, Y., Sekiya, K., Nogata, Y., Ohta, H., 1996.Putrescine accumulation in wounded green banana fruit.Phytochemistry 42, 331–334.




















