
Effect of nitrogen or phosphorus starvation on pigment composition of cultured Heterocapsa sp

Role of a VPS41 homologue in starvation response,intracellular survival and virulence of Cryptococcusneoformans
Xiaoguang Liu,1 Guowu Hu,1 John Panepinto1 andPeter R. Williamson1,2*1Section of Infectious Diseases, Department ofMedicine, University of Illinois at Chicago, Collegeof Medicine, Chicago, IL, USA.2Jesse Brown VA Medical Center, Chicago, IL, USA.
Summary
Previous studies have demonstrated an important rolefor the vacuole in the virulence of the fungus Crypto-coccus and studies in yeast have implicated the vacu-olar protein Vps41 in copper loading of proteins suchas iron transporters. However, our studies found thata cryptococcal vps41D strain displayed wild-typegrowth on media containing iron and copper chelatorsand normal activity of the copper-containing virulencefactor laccase as well as almost normal growth at 37°Cand wild-type production of the virulence factorcapsule. Despite these attributes, the vps41D mutantstrain showed a dramatic attenuation of virulence inmice and co-incubation of mutant cells with the mac-rophage cell line, J774.16, resulted in a dramatic lossin viability of the vps41D mutant strain at 10 h com-pared with wild-type and complemented strains.Closer examination revealed that the vps41D mutantdisplayed a dramatic loss in viability after nutrientstarvation which was traced to a failure to undergo G2arrest, but there was no defect in the formation ofautophagic or proteolytic vesicles. Our results indi-cate that VPS41 plays a key role in regulating starva-tion response in this pathogenic organism and thatdefects in cell cycle arrest are associated with attenu-ated pathogenic fitness in mammalian hosts.
Introduction
Cryptococcus neoformans is a major fungal pathogenprimarily infecting immunocompromised hosts, especiallypatients with impaired cellular immunity related to HIVinfection (Perfect et al., 1998). It is generally believed thatinhalation of desiccated yeasts or basidiospores from the
environment is the route for pulmonary C. neoformansinfection. The alveolar macrophages (AMs) thereforeserve as one of the first lines of anticryptococcal defencethrough phagocytosis and subsequent killing of theingested yeast cells. An effective immune response of theimmunocompetent host normally results in clearance ofintracellular yeast cells or containment of the fungus in alatent state (Feldmesser et al., 2001a). However, crypto-coccal infection of hosts with impaired cellular immunitysuch as AIDS patients results in intracellular growth of thefungus following phagocytosis by AMs and eventuallyleads to the lysis of macrophages and release of replicatedinfectious yeast cells (Feldmesser et al., 2001b). Dissemi-nation of cryptococcal cells to the brain causes life-threatening cryptococcal meningitis, the predominantmanifestations in the clinical setting associated withcryptococcal infection of AIDS patients (Casadevall andPerfect, 1998).
The ability of C. neoformans to elaborate a variety ofvirulence factors forms the foundation of a rich repertoire ofpathogenic strategies as an opportunistic pathogen andenables it to survive in both extracellular and intracellularenvironments. For example, production of laccase, a keyenzyme involved in lignin degradation and in vitro pigmentformation, was shown in vitro to interfere with hydroxylradical production by host macrophages and thus likelycontributes to cryptococcal survival inside macrophage(Liu et al., 1999a). Moreover, melanized cryptococcal cellsexhibit more resistance to antibody-mediated phagocyto-sis and the killing effects of murine macrophages thannon-melanized cells (Wang et al., 1995), although theformation of significant amounts of melanin during in vivopathogenesis has been called into question (Liu et al.,1999b). A second virulence factor of C. neoformans is theproduction of an extensive polysaccharide capsule whichinhibits phagocytosis and promotes survival after phago-cytosis (Bulmer and Sans, 1967; Fromtling and Shadomy,1982). Accumulation of polysaccharide-containingvesicles in the cytoplasm of infected macrophages hasbeen hypothesized to augment intracellular survival by theencapsulated cryptococcal cells (Tucker and Casadevall,2002). Furthermore, the ability to grow at host tempera-tures allows C. neoformans either to proliferate or to estab-lish latent infection in mammalian hosts, serving as a
Accepted 21 June, 2006. *For correspondence. E-mail [email protected];Tel. (+1) 312 996 6070; Fax (+1) 312 413 1657.
Molecular Microbiology (2006) 61(5), 1132–1146 doi:10.1111/j.1365-2958.2006.05299.xFirst published online 25 July 2006
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd

fundamental component of the pathogenic pathway of thisorganism (Odom et al., 1997). Additional genes that havebeen shown to contribute to the intracellular and extracel-lular survival of cryptoccal cells, by inhibiting phagocytosisor by counteracting the oxidative and nitrosative stressesof macrophages, include AOX1 (Akhter et al., 2003),URE1, PLB1, SOD1 (Cox et al., 2000; 2001; 2003), APP1(Luberto et al., 2001), FHB1 (de Jesus-Berrios et al.,2003), TSA1 and TRX1 (Missall et al., 2004; Missall andLodge, 2005).
Previously, a vph1D mutant was isolated from a screen-ing for laccase-deficient mutations using an insertionalmutagenesis approach (Erickson et al., 2001). The VPH1gene encodes a component of the vesicular proton pumprequired for post-translational modification of proteins.Importantly, loss of VPH1 function resulted in defects inexpression of four virulence traits: laccase, capsule,urease and growth at 37°C, suggesting an important rolefor normal vesicular/vacuolar functions in the expression ofvirulence factors in C. neoformans. In addition, studies ofvacuolar mutants of C. neoformans has shed light on therole of these proteins in the biosynthesis of metalo-enzymes. For example, defective vesicular acidificationresulted in defects in metalation of the copper-containingvirulence factor, laccase, which could be reconstituted withexogenous copper (Zhu et al., 2002). Similarly, the vesicu-lar chloride channel, CLC1, was found to be important inlaccase activity and has also been implicated in efficientmetalation of copper-containing factors, such as Fet3 inSaccharomyces cerevisiae (Zhu and Williamson, 2003). Tofurther test the potential role of cryptococcal vesicularfunction in virulence expression, we deleted a cryptococcalVPS41 homologue, which functions in both the alkalinephosphate (ALP) and carboxypeptidase Y (CPY) sortingpathways and has been implicated in copper loading of theiron transporter, Fet3 in yeast (Radisky et al., 1997).
In this study, we report that although VPS41 is dispens-able for normal growth of C. neoformans even under con-ditions of low iron availability and plays no role inexpression of several important metal-dependent viru-lence factors such as laccase, capsule and urease, it isessential for virulence and survival in macrophages. Fur-thermore, the VPS41 mutation was associated with adefect in tolerance to nutrient starvation, related to aninability to arrest in G2. These studies thus provide geneticsupport for a role for adaptation to nutrient starvation as acomponent of the virulence composite of pathogenic fungi.
Results
Isolation of the VPS41 gene and creation of a vps41deletion mutant
A BLAST search of the Duke serotype A genome databasewith the protein sequence of the S. cerevisiae VPS41
gene identified a single cryptococcal VPS41 homologue(E = 3.9 e-56; http://cneo.genetics.duke.edu/) and a ~6 kbgenomic fragment encompassing the whole VPS41 genewas PCR-amplified, cloned and sequenced (as detailedin Experimental procedures). To accurately predict theprotein sequence of VPS41, a 3245 bp cDNA fragmentcontaining the full open reading frame (ORF) of VPS41was isolated from total cryptococcal RNA by reversetranscription polymerase chain reaction (RT-PCR)amplification. Sequence comparison of the genomic DNAof VPS41 and its cDNA revealed that the full length of thecryptococcal VPS41 gene contains 13 introns. Sequenceanalysis indicated that the full ORF of VPS41 is predictedto encode a hydrophilic protein of 1038 amino acids.Alignment with the Saccharomyces Vps41 proteinshowed that the two share distinct conserved domains,namely, an N-terminal Domain I, a domain required forbinding of S. cerevisiae Vps41p to the C-terminal coiledcoil containing domain of Apl5p (Darsow et al., 2001), andtwo clathrin heavy-chain repeat (CHCR) domains involv-ing in homo-oligomerization and a RING-H2 motif presentin higher eukaryotes but not Vps41 from S. cerevisiae(Ward et al., 2001). However, the overall shared identity ofthe two proteins is moderate [27.58% (246/892)]. South-ern blot analysis indicated that, as in S. cerevisiae, theC. neoformans genome contains only one copy of theVPS41 gene (data not shown).
To determine the potential role of the VPS41 gene in thevirulence of C. neoformans, we disrupted the VPS41 locususing a 1.6 kb URA5 fragment to replace the 447 bpEcoRV/BglII fragment of the wild-type locus via a biolistictransformation approach. Of 125 transformants obtained,three vps41D deletion mutants were detected based oninitial PCR screening. Southern blot analysis showed thatall three mutants contained a specific integration of theURA5 marker into the targeted genomic region of theVPS41 gene. One of the deletion mutants, designed asvps41D, was used for experiments in this study to evaluatethe functional roles of the VPS41 gene in C. neoformans.
To complement the vps41D mutant, a cloned ~6 kbgenomic fragment containing the full ORF of VPS41 plus~800 bp of the 5′ promoter region was transformed intothe vps41D mutant cells by electroporation using the 2 kbHgR gene, which confers hygromycin-B resistance, as aselection marker and genomic insertion verified by South-ern blot hybridization using uncut genomic DNA.
VPS41-related virulence phenotypes in C. neoformans
As VPS41 has been implicated in intact function of thecopper-dependent iron transporter, Fet3 in Saccharomy-ces, we investigated whether VPS41 deletion resulted in areduction in activity of the virulence factor, laccase, whichis also a multicopper oxidase of the same family as
VPS41 of Cryptococcus 1133
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 61, 1132–1146
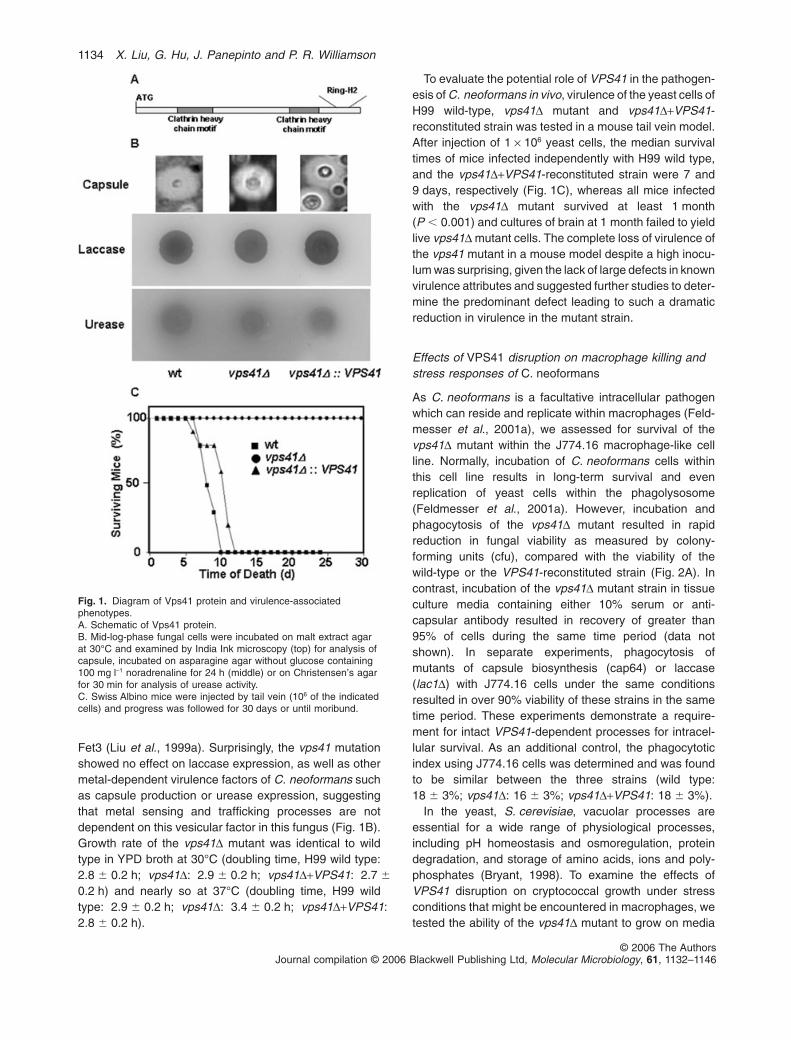
Fet3 (Liu et al., 1999a). Surprisingly, the vps41 mutationshowed no effect on laccase expression, as well as othermetal-dependent virulence factors of C. neoformans suchas capsule production or urease expression, suggestingthat metal sensing and trafficking processes are notdependent on this vesicular factor in this fungus (Fig. 1B).Growth rate of the vps41D mutant was identical to wildtype in YPD broth at 30°C (doubling time, H99 wild type:2.8 � 0.2 h; vps41D: 2.9 � 0.2 h; vps41D+VPS41: 2.7 �
0.2 h) and nearly so at 37°C (doubling time, H99 wildtype: 2.9 � 0.2 h; vps41D: 3.4 � 0.2 h; vps41D+VPS41:2.8 � 0.2 h).
To evaluate the potential role of VPS41 in the pathogen-esis of C. neoformans in vivo, virulence of the yeast cells ofH99 wild-type, vps41D mutant and vps41D+VPS41-reconstituted strain was tested in a mouse tail vein model.After injection of 1 ¥ 106 yeast cells, the median survivaltimes of mice infected independently with H99 wild type,and the vps41D+VPS41-reconstituted strain were 7 and9 days, respectively (Fig. 1C), whereas all mice infectedwith the vps41D mutant survived at least 1 month(P � 0.001) and cultures of brain at 1 month failed to yieldlive vps41D mutant cells. The complete loss of virulence ofthe vps41 mutant in a mouse model despite a high inocu-lum was surprising, given the lack of large defects in knownvirulence attributes and suggested further studies to deter-mine the predominant defect leading to such a dramaticreduction in virulence in the mutant strain.
Effects of VPS41 disruption on macrophage killing andstress responses of C. neoformans
As C. neoformans is a facultative intracellular pathogenwhich can reside and replicate within macrophages (Feld-messer et al., 2001a), we assessed for survival of thevps41D mutant within the J774.16 macrophage-like cellline. Normally, incubation of C. neoformans cells withinthis cell line results in long-term survival and evenreplication of yeast cells within the phagolysosome(Feldmesser et al., 2001a). However, incubation andphagocytosis of the vps41D mutant resulted in rapidreduction in fungal viability as measured by colony-forming units (cfu), compared with the viability of thewild-type or the VPS41-reconstituted strain (Fig. 2A). Incontrast, incubation of the vps41D mutant strain in tissueculture media containing either 10% serum or anti-capsular antibody resulted in recovery of greater than95% of cells during the same time period (data notshown). In separate experiments, phagocytosis ofmutants of capsule biosynthesis (cap64) or laccase(lac1D) with J774.16 cells under the same conditionsresulted in over 90% viability of these strains in the sametime period. These experiments demonstrate a require-ment for intact VPS41-dependent processes for intracel-lular survival. As an additional control, the phagocytoticindex using J774.16 cells was determined and was foundto be similar between the three strains (wild type:18 � 3%; vps41D: 16 � 3%; vps41D+VPS41: 18 � 3%).
In the yeast, S. cerevisiae, vacuolar processes areessential for a wide range of physiological processes,including pH homeostasis and osmoregulation, proteindegradation, and storage of amino acids, ions and poly-phosphates (Bryant, 1998). To examine the effects ofVPS41 disruption on cryptococcal growth under stressconditions that might be encountered in macrophages, wetested the ability of the vps41D mutant to grow on media
Fig. 1. Diagram of Vps41 protein and virulence-associatedphenotypes.A. Schematic of Vps41 protein.B. Mid-log-phase fungal cells were incubated on malt extract agarat 30°C and examined by India Ink microscopy (top) for analysis ofcapsule, incubated on asparagine agar without glucose containing100 mg l-1 noradrenaline for 24 h (middle) or on Christensen’s agarfor 30 min for analysis of urease activity.C. Swiss Albino mice were injected by tail vein (106 of the indicatedcells) and progress was followed for 30 days or until moribund.
1134 X. Liu, G. Hu, J. Panepinto and P. R. Williamson
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 61, 1132–1146

containing hydrogen peroxide or the redox cycling agentmenadione, the latter of which tests for the sensititivy ofa cell to superoxide radicals (Narasipura et al., 2003).Compared with H99 wild type, and the reconstitutedvps41D+VPS41 strain, no observable differences in growthwere observed for the vps41D mutant (Fig. 2C), suggest-ing that the function of VPS41 is dispensable for survivaland growth in the presence of oxidative stresses. Thevps41D mutant strain was also found to be insensitive toosomotic and ionic stress indicated by wild-type growth onmedia containing 1.2 M NaCl or 1.8 M sorbitol (data notshown). In addition, as VPS41 deletion in S. cerevisiaeproduces growth defects on low-iron medium (Radiskyet al., 1997), conditions that are believed to exist within theintracellular phagolysosome (Fang et al., 2000; Chunget al., 2004), we next investigated whether VSP41 isrequired for normal cryptococcal growth in iron- or copper-deficient medium at 30°C and 37°C. H99 wild type, vps41Dmutant and the reconstituted vps41D+VPS41 strain wereallowed to grow on YPD agar with addition of either the ironchelator 3-(2-Pyridyl)-5,6-diphenyl-1,2,4-triazine-p,p′-disulfonic acid, monosodium (500 mM) or the copper chela-tor Bathocuproinedisulphonic acid, disodium (1 mM).Again, no defects in growth were detected for the vps41Dmutant and its growth on both conditions was comparable
to that of wild-type and reconstituted vps41D+VPS41strains (Fig. 2C). Thus, unlike that of its counterpart inS. cerevisiae, VPS41 function is not required for C. neo-formans to grow on iron-deficient medium, suggesting thatcryptococal cells may use a different iron transportpathway or that there are other gene(s) possessing over-lapping function with VPS41 in this pathogenic fungus.Finally, we assessed the functional role of VPS41 inC. neoformans in response to nutrient starvation condi-tions as previous studies showing a role for the glyoxylatepathway and gluconeogenesis in fungal pathogenesissuggest an importance for nutrient deprivation in protectionfrom fungal pathogens (Lorenz and Fink, 2001; Panepintoet al., 2005). The wild-type, vps41D mutant and reconsti-tuted vps41D+VPS41 strains were cultured at 37°C withshaking (250 r.p.m. min-1) in sterile water supplementedwith YNB without amino acids and ammonium sulphate forvarious time periods. As shown in Fig. 2B, the vps41Dmutant strain exhibited severe defects in response tonutrient starvation characterized by an 80% decline inviability at the 10 h point, compared with a viability of84.5% and 97.4% for the wild-type and reconstitutedvps41D+VPS41 strains, respectively, over the same timeperiod (P � 0.001). Interestingly, the cryptococcal vps41Dmutant survival defect could not be complemented by
Fig. 2. Incubation of VPS41 mutants inmacrophages and starvation medium.A. Cells were opsonized with anti-capsularantibody and incubated with J774macrophages, washed extensively to removenon-phagocytosed cells and, at the indicatedtimes, wells were washed with watercontaining 0.01% SDS and inoculated onYPD for 3 days and cfu measured andnormalized to time zero.B. Indicated log-phase cells were incubated inYNB without amino acids and ammoniumsulphate for the indicated times, followed byremoval of aliquots and inoculation on YPDagar to determine cfu.C. Indicated log-phase cells were suspendedat a constant concentration (106 ml-1) serialdiluted and inoculated on YPD with theindicated additions as described inExperimental procedures at 30°C andvisualized at 30 h. Panels represent a 1000¥dilution series.
VPS41 of Cryptococcus 1135
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 61, 1132–1146

expression of a VPS41 open reading frame from S. cerevi-siae under the C. neoformans promoter, suggesting arequirement for unique cryptococcal sequences such asthe C-terminus RING-H2 motif described above. Insummary, the above results indicate that the function ofVPS41 is essential for cryptococcal survival under starva-tion conditions and represents a new phenotypic associa-tion for this gene in eukaryotes.
VPS41-associated cellular function under nutrientstarvation conditions
Previous studies in Saccharomyces have implicatedVPS41 in intact vacuolar morphology (Radisky et al.,
1997), and recently the Vps41 protein has been localizedto the yeast vacuole (LaGrassa TJ, 2005), a phenotypeshared by the cryptococcal homologue as demonstratedby colocalization of a Vps41-green fluorescent protein(GFP) construct with the vacuolar marker, FM4-64 (Fig.3A). In addition, smaller vesicular structures have beenobserved in yeast vps41D mutants, but the role of nutrientstarvation in this phenomenon has not been studied.Thus, to investigate the role of VPS41 during nutrient-replete as well as under starvation conditions, and toassess this role in C. neoformans, we used a coumarindye, 7-amino-4-chloromethyl-coumarin (CMAC), whichlocalizes to acidic vesicular compartments (Roberts et al.,1991; Shoji et al., 2006). As shown in Fig. 3B, during log
Fig. 3. Visualization Vps41-GFP andvacuoles of Cryptococcus neoformans underlog-phase and starvation conditions.A. Cryptococcus neoformans vps41D cellsexpressing a VPS41-GFP construct wereincubated under starvation conditions,incubated with FM4-64 and examined byepifluorescence or under bright field (BF).B. Cells in either log phase or after incubationin YNB without amino acids and ammoniumsulphate for 3 h at 37°C (starvation) wereincubated with7-amino-4-chloromethyl-coumarin andvisualized by epifluorescence or by Nomarskioptics as described in Experimentalprocedures (1000¥).
1136 X. Liu, G. Hu, J. Panepinto and P. R. Williamson
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 61, 1132–1146

phase, nutrient-replete conditions, both wild-type andvps41D mutant C. neoformans cells were observed toform numerous small acidic vesicles observed by bothCMAC and DIC imaging. However, under nutrient starva-tion conditions, wild-type cells formed larger, more typicalvacuolar structures, whereas the vps41D mutant cellsretained the small vesicular structures characteristic of logphase. Reconstitution with wild-type VPS41 or a Vps41-GFP construct restored the ability to form large acidicvacuoles during nutrient starvation (Fig. 3A and B). Thisfailure to form appropriately sized vacuole compartmentsin the mutant strain could thus be due to a primary defectin vacuole formation or a failure to signal events leading tostarvation-appropriate phenotypes.
During formation of large vacuolar structures duringnutrient starvation, studies in yeast have shown that cellsform smaller cytosolic autophagic vesicules which engulfcellular elements, transporting them into the vacuole forprogrammatic digestion, thus allowing survival underthese conditions (Klionsky, 2004). In contrast, cells defec-tive in autophagic processes show defects in tolerance tonutrient starvation (Klionsky, 2004) and genes involved inautophagy have recently been shown to be upregulatedby phagocytosed C. neoformans (Fan et al., 2005). Thus,we investigated autophagic properties of the vps41Dmutant to identify processes that may be involved in cryp-tococcal macrophage survival. As shown in Fig. 4A, panela, wild-type cryptococcal cells starved by incubation inYNB without amino acids and ammonium sulphate for 3 hat 37°C formed typical small granular bodies that can belabelled with the specific autophagic marker, Atg8. Atg8 ishighly conserved among eukaryotes (Lang et al., 1998;Wu et al., 2006) and includes a single cryptococcal homo-logue (http://www.tigr.org) that is 93% identical to Atg8from S. cerevisiae. Similarly, during starvation conditions,the vps41D mutant cells also formed Atg8-labelledautophagic digestive vesicles (Fig. 4A, panel d). In con-trast, log-phase, nutrient-replete growth conditionsresulted in repressed production of Atg8-labelled autoph-agic vesicles in wild-type C. neoformans (data not shown)as described in yeast (Lang et al., 1998). To furtherconfirm that vesicles observed during starvation showedintact proteolytic function required for autophagy, the fluo-rescent dye markers, 7-amino-4-chloromethylcoumarin,L-arginine amide (CMAC-Arg) and 7-amino-4-chloromethylcoumarin, L-alanyl-L-proline amide (CMAC-Ala-Pro), were used that require proteolytic cleavageby amino-peptidase or dipeptidyl-peptidase enzymes,respectively, to activate fluorescence (Suarez-Rendueleset al., 1981; Achstetter et al., 1983; Roberts et al., 1991).As shown in Fig. 4B, in log-phase cells and under starva-tion conditions, both wild-type and vps41D mutant cellsform small proteolytic vesicles which colocalize to thevesicular membrane marker, MDY-64. Similar results
were observed using the dipeptidyl peptidase substrate,corroborating the presence of intact proteolytic vesicles inthe vps41D mutant strains (data not shown).
Deletion of VPS41 results in failure of G2 arrest duringnutrient starvation
Cell cycle arrest is a prominent response to nutrient dep-rivation of higher eukaryotes and has been observed bylaser scanning cytometry in C. neoformans cells arrestedin stationary-phase and oxygen deprivation (Ohkusuet al., 2004). Initial studies with the vps41D mutantshowed that upon incubation in nutrient-deprived condi-tions in YNB without amino acids and ammonium sulphatefor 24 h at 37°C, the budding index of the mutant waselevated (31.6%) compared with that of wild-type cells(15.8%) incubated under the same conditions (P �
0.0001). This suggested that VPS41 may have a role incell cycle control under starvation conditions, as abnor-malities in the budding index have been shown to corre-late with abnormalities in cell cycle (Zettel et al., 2003). Incontrast, the budding indexes of the two strains in logphase were found to be similar (H99 wild type: 69.7%versus vps41D: 74.4; P � 0.10). As cell cycle arrest is animportant requirement for survival under nutrient-deprivedconditions (Werner-Washburne et al., 1996), we usedlaser scanning cytometry to assess for successful arrestin vps41D cells. As shown in Fig. 5A, wild-type C. neo-formans cells in log phase are distributed almost equallybetween G1 and G2, indicated by a similar proportion ofcells with 1C DNA and 2C DNA. Cell size of G2 cells werealso greater than that of G1 cells as indicated by a ratio offorward scatter of G2/G1 of approximately 1.4 for wild-type cells. (Peaks greater than 2C are most likely due toadherent, budding cells.) However, in response to nutrientstarvation, wild-type cells arrested in G2, resulting inalmost all cells found in this phase of the cell cycle asdescribed previously for C. neoformans (Ohkusu et al.,2004; Fig. 5A). As cells with a 2C DNA content can alsobe due to cells arrested in mitosis, we visualized nuclei by4′,6-diaminidino-2-phenylindole dilactate (DAPI) stainingto distinguish the stages. As shown in Fig. 5B, wild-typecells were found to arrest with a well-formed, large nucleitypical for G2 arrest. In contrast, we failed to identify anymitotic cells in over 200 cells counted. Similar to thewild-type cells, the vps41D mutant cells were also found tobe distributed between G1 and G2 during log phase.(Ratio of forward scatter G2/G1 was again approximately1.4. More tailing of the G2 peak is most likely due toslower release of buds in the vps41D cells.) However,after 24 h of nutrient starvation, the vps41D cells contin-ued to be distributed between G1 and G2 in amountssimilar to that of log phase (Fig. 5A). DAPI staining alsoshowed two populations of cells, one containing larger
VPS41 of Cryptococcus 1137
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 61, 1132–1146

nuclei typical of G2 arrest, and a population with typicalearly postmitotic nuclei, which are small and condensed(Straube et al., 2003). However, mutant cells were stillmetabolically active after incubation in starvation condi-tions, as shown by metabolic processing of FUN 1 dye toa fluorescent vesicular product as previously described(Martinez and Casadevall, 2005). Flow cytometry-derivedDNA distributions of starved vps41D mutant were notaltered by addition of 1 mg ml-1 rapamycin, an inhibitor ofTOR in C. neoformans (Cruz et al., 2001), suggesting thatthe VPS41 gene acts either downstream of or is indepen-
dent of TOR under these starvation conditions. Overall,these data suggest that VPS41 may play a role in effec-tive cell cycle control during nutrient deprivation andprovide an explanation for the mutant’s inability to toleratenutrient starvation conditions encountered in the intracel-lular environment and the mammalian host.
Discussion
Targeted deletion of VPS41 was initiated as an effort tofurther evaluate the potential role of vacuolar functions in
Fig. 4. Immunofluorescent localization ofAtg8-containing autophagic and proteolyticvesicles.A. Cells in mid-log phase were incubated inYNB without amino acids and ammoniumsulphate for 3 h at 37°C, and methanol wasfixed and incubated with an anti-Atg8antibody, followed with an anti-rabbitAlexafluor 594 antibody (panels a and e) oranti-rabbit Alexafluor 594 alone (panels c andg) and examined for epifluorescence. Panelsb, d, f, h represents equivalent bright fieldviews. (1000¥).B. Log-phase cells were either harvested orincubated in YNB without amino acids andammonium sulphate for 3 h at 37°C andincubated 7-amino-4-chloromethylcoumarin,L-arginine amide (CMAC-Arg) and MDY-64 asdescribed in Experimental procedures andexamined by epifluorescence or by Nomarskioptics as indicated (1000¥).
1138 X. Liu, G. Hu, J. Panepinto and P. R. Williamson
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 61, 1132–1146

laccase expression and virulence of C. neoformans. Inter-estingly, many of the functions of proteins localized tovacuoles also play roles in biosynthetic machinery withinsecretory vesicles. For example, in both Saccharomycesand Cryptococcus, deletion of genes encoding compo-nents of the vacuolar chloride channel and proton pump
have impacts on the bio-incorporation of copper cofactorsinto the iron transporter Fet3 and the virulence factorlaccase respectively (Davis-Kaplan et al., 1998; Zhu andWilliamson, 2003; Zhu et al., 2003). As Vps41 has alsobeen implicated to play a role in the metalation of Fet3 inSaccharomyces with growth defects on iron or copper-
Fig. 5. VPS41 mutants: visualization andcontent of DNA, and metabolic activitymeasured by FUN 1.A. Cells in mid-log phase were eitherharvested or incubated in YNB without aminoacids and ammonium sulphate for 24 h at37°C, washed, fixed and incubated withpropidium iodide and subjected to laserscanning cytometry as described inExperimental procedures.B and C. (B) Cells incubated in YNB withoutamino acids and ammonium sulphate wereincubated with DAPI or (C) FUN 1 andexamined by epifluorescence (1000¥).
VPS41 of Cryptococcus 1139
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 61, 1132–1146

limiting media (Radisky et al., 1997), we investigated itsrole in the expression of laccase. Structurally, the crypto-coccal Vps41 protein is highly similar to the Saccharomy-ces homologue, but the cryptococcal homologue alsocontains a vesicle-binding RING-H2 motif more typical ofhigher eukaryotes such as mammalian cells (Ward et al.,2001), suggesting that there could be divergent functionsof VPS41 between ascomycete and basidiomycete fungi.Such a structural divergence in VPS41 may explain thelack of a role in copper homeostasis in C. neoformans;alternatively, there may be redundant pathways for copperinsertion in basidiomycete fungus.
Despite the normal expression of virulence factors inthe vps41D mutant and a minimal effect on growth rate at37°C, a dramatic attenuation in virulence was observedusing a mouse model with survival of all inoculated miceand clearance of organisms despite using a high inoculum(106 cfu) of fungi. To investigate this in more detail, fungalcells were incubated with a J774.16 macrophage cell lineto study phagocytosis and killing. While phagocytosis wassimilar to wild type for the mutant strain, a dramatic rate ofkilling was observed within hours of phagocyosis that wasmore pronounced than that observed for mutants of twoprincipal virulence factors of C. neoformans, capsule andlaccase. This dramatic rate of killing is unusual for a viablecryptococcal mutant and could not be accounted for bythe slight reduction in growth at 37°C or the slower budrelease observed during flow cytometry. Indeed, previousstudies of wild-type C. neoformans have shown success-ful replication within J774.16 cells and even acapsularstrains show significant survival within a 24 or 48 h period(Mukherjee et al., 1996). Thus, rapid killing of the vps41Dcells over a period of a few hours suggests that VPS41 isinvolved in expression of an additional phenotype essen-tial for intracellular survival, a property that has recentlybeen implicated in both acute and chronic cryptococcalinfections (Feldmesser et al., 2001a). Expression of suchcritical phenotypes appears to be key to the ability of apathogen to survive in the host, a property recentlydescribed as pathogenic fitness (Panepinto and William-son, 2006).
Previous studies have suggested that macrophage celllines derived from BALB/c mice such as J774 containdefects in the membrane transporter NRamp, which isrequired for wild-type oxidative burst (Barton et al., 1995)and is involved in anticryptococcal killing (Blasi et al.,2001). Thus, it may be less likely for an oxidative killingmechanism to be primarily responsible for the rapiddemise of the vps41D strain in this cell line, and, indeed,the vps41D mutant in vitro did not appear to show alteredsurvival in response to oxidative stresses such as hydro-gen peroxide or menadione, previously shown to be fun-gicidal for C. neoformans (Narasipura et al., 2003; deJesus-Berrios et al., 2003). Instead, the cryptococcal
vps41D mutant cells displayed a defect in tolerance tonutrient starvation with a rapid rate of demise similar tothat observed after phagocytosis by the macrophage cellline. This represents a new role for Vps41 proteins, andmay explain previous data showing that class C complexvacuolar genes such as VPS41 and VPS39 are hypersen-sitive to the TOR inhibitor rapamycin whose pathway isinvolved in tolerance to nutrient deprivation by activationof cell cycle arrest and autophagy (Xie et al., 2005).Indeed, these data may shine light on a recent finding thatVPS41 was identified in a haplo-insufficient screen formutants defective in the filamentation, suggesting therelationship between nutrient-induced cell cycle arrestand filamentation (Uhl et al., 2003).
These data led to further investigation of starvation-related phenotypes in the cryptococcal vps41D mutantwhich suggested several defects in phenotypes importantto survival under nutrient-deprived conditions. Forexample, in wild-type C. neoformans, log-phase cellsdemonstrated numerous small staining vesicles whichaccumulated the acidic CMAC fluorescent dye, and,under starvation conditions, developed larger, moretypical vacuoles similar to other yeasts. However, thevps41D mutant failed to demonstrate enlarged vacuolesupon nutrient starvation, typical for the aberrant mor-phology described in class C vacuolar mutants ofS. cerevisiae (Radisky et al., 1997). This defect could bedue to an inherent defect in enlargement of vacuolesduring the transition to nutrient starvation conditions, or itcould be due to a defect in either nutrient sensing orexecution of processes associated with cell cycle arrest. Asecond starvation-associated pathway, the formation ofAtg8-labelled autophagic vesicles (Noda and Ohsumi,1998; Kamada et al., 2004), appeared to be intact duringnutrient starvation, and formation of proteolytic competentvesicles required for autophagic digestion also appearedintact. However, we were not able to study proteolyticmaturation of specific VPS41-associated proteases suchas CPY due to a lack of appropriate homologues inC. neoformans.
Surprisingly, investigation of cell cycle in the VPS41mutants demonstrated that the VPS41 gene may have arole in cell cycle control mechanisms during starvationthat may help to explain the poor survival of the mutantstrain both in vitro and after macrophage phagocytosis.G2 arrest upon nutrient deprivation has been previouslydescribed under stress conditions and stationary phase(Ohkusu et al., 2001), and thus may play an important rolein nutrient conservation in the intracellular habitat. Failureof successful cell cycle arrest results in rapid depletion ofcellular nutrient stores and has been associated with poorsurvival under nutrient-depleted conditions in yeast (Iidaand Yahara, 1984). An alternative explanation may be thatthe mutant cells simply shut down and die during nutrient
1140 X. Liu, G. Hu, J. Panepinto and P. R. Williamson
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 61, 1132–1146

starvation, but the retention of starvation-specific meta-bolic events such as expression of enzymatically activelaccase, production of autophagic vesicles, as well asmetabolic conversion of the FUN 1 fluoresecent dye,make this explanation less likely. However, a specific roleof VPS41 in cell cycle control will require the identificationof the specific pathways and target genes involved and isthe subject of active investigation. Interestingly, addition ofrapamycin to mutants during starvation failed to restorewild-type cell cycle arrest as reported previously in yeast(Zaragoza et al., 1998), suggesting that VPS41 inC. neoformans may act downstream of or act indepen-dently of TOR under these conditions.
These data thus suggest a role of VPS41 in tolerance tonutrient stress expected to occur within the macrophageintracellular environment. Despite the need for host cellsto obtain basic nutrients in its own growth and survival,pathogens appear to be denied these basic tools for lifeduring intracellular phagocytosis. Thus, in addition to aneed for the pathogen to obtain nutrients under theadverse host environment, it appears that an essentialtool for pathogen survival is also to prevent growth bymechanisms such as cell cycle arrest under conditionsthat may temporarily deprive the fungal pathogen of nec-essary substrates required for growth.
Experimental procedures
Fungal strains, plasmids and media
Cryptococcus neoformans ATCC 208821 (H99) was a gen-erous gift of J. Perfect. Strain H99FOA19, a uracil aux-otrophic mutant derived from H99 (Erickson et al., 2001), wasemployed as a recipient strain for creating targeted genedeletion. The strains were grown in YPD media (2% glucose,1% yeast extract, 2% Bacto-peptone) or YPD agar medium(adding 2% agar) or incubated in yeast nitrogen base(1.7 g l-1 YNB, without amino acids and ammonium sulphate,Becton, Dickinson, MD) media for starvation experiments.Asparagine minimum selective media for transformant selec-tion and for detection of laccase production were previouslydescribed (Zhu and Williamson, 2003). Plasmids pPM8 andpCIP containing the URA5 gene are a kind gift of K.J. Kwon-Chung. Plasmids containing the hygromycin resistance genewas a gift of G. Cox.
Cell lines and culture media.
J774.16 (ATCC) is a murine macrophage-like cell line derivedfrom a reticulum sarcoma. Cells were maintained at 37°C in10% CO2 in DMEM (Invitrogen, Rockville, MD) that wassupplemented with 10% heat-killed fetal calf serum (FCS)(Harlan, Indianapolis), Cellgrow 1¥ non-essential aminoacids (Mediatech, Haerndon, VA), Cellgrow 100 mg ml-1
penicillin/streptomycin and 10% NCTC-109 medium(Invitrogen). Cell lines were used between 5 and 15passages.
Disruption and complementation of VPS41
To make the deletion construct, a 2.5 kb genomic fragmentencompassing the major part of VPS41 ORF was PCR-amplified using primer pairs of VPS41F1 (5′-GCTCCAGAAGAGCACGGATG-3′) VPS41R1 (5′-GTGTGACATGTTTGCGTATG-3′) and subsequently cloned into TOPO TAvector (Invitrogen). The resultant plasmid was digested withEcoRV and BglII and the 447 bp EcoRV/BglII fragment wasreplaced by a 1.6 kb fragment of a URA5 transformationmaker described previously (Varma et al., 1992) to generatethe deletion construct plasmid. The final disruption allele witha 1.6 kb URA5 marker flanked on either side by a 500 bpDNA sequence homologous to genomic regions of theVPS41 gene was PCR-amplified and purified by spin column(Invitrogen) and introduced into H99FOA cells via a biolisticapproach (Cox et al., 1996). Transformants were screenedfor potential VPS41 deletion mutant by a PCR approachusing one set of primers, designed to detect disruptionevents, including a primer just outside of the transformingvector and an opposing primer based on the URA5sequence. The specific disruption of the VPS41 gene in can-didate mutants was verified by using a set of primers outsidethe sequences of the knockout construct and by Southernblot analysis. To complement the vps41D mutant strain, a6 kb genomic fragment encompassing the full ORF of VPS41plus ~800 bp of the 5′ promoter region was PCR-amplifiedusing a primer set of VPS41FC (5′-TCGAGCTTGGACAGCAGAG-3′) and VPS41RC (5′-ATTCACTGCGCACACGTCTGC-3′) and then cloned into a modified Blue-script SK vector (Stratagene) containing the 2 kbhygromycin-B resistance gene under the control of a crypto-coccal actin promoter (Cox et al., 1996) to generate thecomplementation construct, which was introduced intovps41D mutant cells by electroporation as previouslydescribed (Erickson et al., 2001) and transformants wereselected on hygromycin-containing YEPD agar plates(200 m ml-1).
Macrophage killing assay
The J774.16 macrophage-like cell line (ATCC) was used toevaluate the ability of the vps41D mutant to grow insidemacrophages by a previously described method (Tucker andCasadevall, 2002). Cells were allowed to grow for 5–7 daysin Dulbecco’s modified Eagle medium (Cellgro, Herndon, VA)supplemented 10% FCS, 100 mg ml-1 Cellgro penicillin-streptomycin at 37°C in the presence of 5% CO2, and thenharvested from monolayers using 0.25% trypsin and thenumber of cells were counted with a hemocytometer. Themacrophage concentration was adjusted to 105 cells ml-1 and100 ml of the macrophage suspension was added to eachwell of a 96-well plate. The cells were primed with murinegamma interferon (IFNg) (Sigma) at a concentration of50 U ml-1 and were incubated at 37°C, 5% CO2 overnight.Yeast cell suspensions (107 ml-1) of H99, vps41D and thecomplement were prepared from fresh cultures and antibody18BF (Y1) (a generous gift of A. Casadevall) added(10 mg ml-1) and incubated at 37°C for 1 h. To each well in the96-well plate, 104 antibody-treated cryptococcal cells wereadded plus 50 units of IFNg (Sigma) and 1 mg of LPS and
VPS41 of Cryptococcus 1141
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 61, 1132–1146

incubated at 37°C 5% CO2. The macrophage and yeast mix-tures were harvested at various time points (1, 3, 10 h) andextracellular yeast cells were removed by washing withphosphate-buffered saline (PBS) three times and lysed with0.1% SDS in water. The collected yeast cells were finallywashed with and suspended in PBS and plated onchloramphenicol-containing YPD agar for colony counts. Allexperiments were performed in triplicate.
Macrophage phagocytosis assay
Macrophage phagocytosis assays were conducted accordingto the method of Tucker and Casadevall (2002). Briefly,18–24 h before assays, macrophages were plated at adensity of 1–3 ¥ 105 cells per ml in culture media as above.Log-phase cryptococcal cells were washed three times inPBS and counted in a haemacytometer. Yeast cells wereadded to cultured cells at a multiplicity of infection of 1:1 in thepresence of 10 mg ml-1 monoclonal antibody 18B7 (a gener-ous gift of A. Casadevall) and IFNg and LPS added asdescribed above. Phagocytosis was allowed to proceed at37°C for 1 h in 10% CO2, and extracellular yeast cells wereremoved by three successive washes with regular culturefeeding medium without LPS or IFNg, then stained with DiffQuick and phagocytosis assessed by counting 100macrophages. Five randomly selected fields were counted.The phagocytosis index was expressed as a mean percent-age of AMs attaching to or ingesting one or more organismsper 100 Ams � standard error (SE) as previously defined (Liuet al., 1999a).
Starvation stress response assay
To assess the role of VPS41 in starvation stress response,yeast cells of overnight cultures in YPD liquid medium ofH99 wild-type, vps41D mutant and the reconstitutedvps41D+VPS41 strains were collected and resuspended,respectively, to a concentration of concentration of 108
cells ml-1, in 15 ml of YNB (without amino acids and ammo-nium sulphate) in a 50 ml tube and then cultivated at 37°Cwith shaking of 250 r.p.m. min-1. At the indicated time point,100 ml of each strain suspension was taken out and used fordetermining cell viability based on cfu counting on YPD agarusing 10-fold serial dilutions.
Virulence studies
Capsule was assayed by India Ink microscopy after incu-bation on malt extract agar for 5 days at 30°C, laccase byincubation on noradrenaline asparagines agar and ureaseactivity measured on Christensen’s media as described pre-viously (Panepinto et al., 2005). The virulence of thevps41D mutant, reconstituted vps41D and wild-type strainswere evaluated in a previously described mouse menin-goencephalitis model (Salas et al., 1996). In brief, eachyeast strain was allowed to grow for 2 days on YPD agar at30°C, and cells were collected, washed once with sterilewater and suspended in sterile PBS attaining a concentra-tion of 1 ¥ 107 cells ml-1. For each yeast strain, 100 ml of acell suspension (containing 106 cells) was tail vein-injected
into 10 NIH-Swiss Albino female mice of 6–10 weeks old.Inoculated mice were monitored twice daily for survival for30 days. Mice showing an inability to reach food or waterwere sacrificed.
Antibody staining and Western blotting
The method of Tucker and Casadevall (2002) and Eksi et al.(2004) was adapted as follows. Cells were grown to mid-logphase and either harvested or washed twice in sterile waterand incubated for the indicated time in YNB without aminoacids and ammonium sulphate at 37°C. Cells were fixed in3% formaldehyde for 1 h at 4°C, washed extensively, andthen subjected to spheroplasting using 40 mg ml-1 of lysingenzyme from Trichoderma harzianum (Amersham) in 1 Msorbitol, 10 mM sodium citrate, pH 5.8, as described (Varmaand Kwon-Chung, 1991) for 4 h at 30°C. Cells were thenwashed in 1 M sorbitol, 10 mM sodium citrate buffer, anddiluted 1:8 in PBS and dried on microscope slides, fixed in100% anhydrous methanol at -60°C and dried. To quenchautofluorescence, cells were incubated in 0.1 M glycine for10 min. Cells were then incubated with a solution of 1:500dilution of rabbit anti-yeast Atg8 (Abcam, Cambridge) in PBScontaining 1 mg ml-1 bovine serum albumin (Sigma) at 4°Cfor 1 h, followed by extensive washing in PBS, then incuba-tion for 1 h at 4°C with 1:1000 Alexafluor 594 chicken anti-rabbit antibody followed by three washes with PBS andimmunofluorescence examined using an Olympus IX-70microscope. Western blots using cryptococcal cell extract,anti-Atg8 antibody and a mouse anti-rabbit horseradish per-oxidase antibody (Sigma) was performed to demonstrateimmunoreactivity of cryptococcal Atg8 as described (Zhuet al., 2001).
Construction of pORA-XK cryptococcal shuttle vector
Because of limitations regarding size and the presence ofunique restriction sites in available vectors, a C. neoformansshuttle vector was constructed for high-copy episomal reten-tion and expression, based on the pPM8 shuttle vectorderived previously (Mondon et al., 2000). The origin of repli-cation from pBluescript SK (Stratagene) was PCR-amplifiedusing primers: ORI-NotI-1000S, 5′-GCCGCCGCGGCCGCGGGGAGAGGCGGTTTG-3′ and ORI-XbaI-1942 A, 5′-GCCGCCTCTAGATTGATTTAAAACTTCATTTTTA-3′, anddigested with NotI, XbaI. Plasmid pPM8 (a generous gift ofK.J. Kwon-Chung) was then digested with NotI, XbaI, and twobands, the first corresponding to a Telomere/Kanamycinsequence and a second corresponding to the STABsequence was gel-purified and the three fragments ligatedand transformed into Escherichia coli. After confirmation byrestriction mapping, the resultant plasmid was digested withXbaI and a double-stranded oligonuclotide was produced bytemperature annealing and phosphorylation with T4 kinaseusing two oligonucleotides: RNAi-MCS-S, 5′-TATGGATCCGAATTCGAGCTCTAGACTCGA-3′ and RNAi-MCS-A,5′-GATCTCGAGTCTAGAGCTCGAATTCGGATCCA-3′, andwere ligated and transformed into E. coli cells, and the result-ant 3 kb plasmid, pORA-XK, recovered, and verified bysequencing. For transformation into C. neoformans, a 1.3 kb
1142 X. Liu, G. Hu, J. Panepinto and P. R. Williamson
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 61, 1132–1146

PCR-amplified fragment of the URA5 gene described previ-ously (Erickson et al., 2001) was ligated into the KpnI site ofpORA-SK to produce pORAS-KU, then a fragment of theEF1-alpha terminator was PCR-amplified from H99 genomicDNA using primers: EF1-T-Age-A, 5′-GCCGCCACCGGTTGAGACACCTTCACCTTGAT-3′ and EF1-T-RI-S, 5′-GCCGCCGAATTCTCGCTTCTGTAGAGCCCAT-3′, digestedwith AgeI and EcoRI and ligated into compatible sites ofpORA-SKU to produce pORA-SKUT. Next, a fragment of theActin promoter was PCR-amplified from H99 genomic DNAusing primers: ACT1-ATG-R1, 5′-CGCCGAATTCATAGACATGTTGGGCGAGTTTAC-3′ and ACT1-Bgl-S, 5′-GCCGCCAGATCTCCATTGCGCATGTACTCGC-3′, digested withBglII and EcoRI and ligated into compatible sites to producethe pORA-KUTAP C. neoformans expression vector andwas sequence verified. Transformation of C. neoformansH99FOA19 was performed by electroporation after lineariza-tion with I-SceI and C. neoformans transformants were foundto contain pORAS-KUTAP with a copy number of approxi-mately 15–20 measured by the ratio of the intensity of thegenomic to episomal URA5 band on uncut Southern blots,and was comparable to equivalent ratios obtained in trans-formants of the same cells using pPM8.
Localization of Vps41, vacuole and proteolytic vesiclesof C. neoformans
To facilitate cellular localization studies in C. neoformans, asynthetic copy of GFP was constructed that utilized codonbias from C. neoformans. To produce this, a codon use tablewas determined for two highly expressed proteins inC. neoformans, SSA1 and EF1a, using annotated sequencefrom the H99 cryptococcal sequencing project http://www.broad.mit.edu and MacVector software. Synthetic overlap-ping oligonucleotides were then designed and synthesizedand PCR amplification was used to produce the syntheticgene according to the method of Gao et al. (2003) which wasintroduced into a puc19 vector and verified by sequencing(GenBank DQ474233) and was inserted into an EcoRI siteupstream of a 500 bp EF1-alpha terminator within pORAS-KUTAP using MunI sites at the 3′-end of the GFP to maintainthe unique upstream EcoRI site for further use. A genomiccopy of VPS41 spanning 1.5 kb upstream of the ATG startcodon and terminating at amino acid 1038 in frame with wasthen introduced upstream of the Cneo-GFP construct previ-ously inserted into pORA-S-KUTAP using BglII, EcoRI sites(which removes the ACT1 promoter) and the construct veri-fied by sequencing. The cryptococcal shuttle vector was thenlinearized with I-SceI and transformed into a vps41D ura5–strain produced by standard methods (Varma et al., 1992).Expression of VPS41 was verified by Northern blot andcomplementation was verified by restoration of wild-typevacuolar morphology by FM4-64 staining described below.Expression of the VPS41 gene from S. cerevisiae wasperformed by insertion of a PCR-amplified copy of the genefrom S. cerevisiae strain BJ-3505 (Kodak) using primers:1, 5′-CAAGAATTCAATGACTACAGATAATCATCAG and 1,CAAGAATTCATGACTACAGATAATCATCAG into pORAS-KUTAP after digestion with EcoRI and ligation into compatiblesites, then verified by sequencing. The plasmid was thenlinearized with I-SceI and transformed into a vps41D ura5
strain produced as described (Varma et al., 1992) andexpression verified by Northern blot.
The vacuolar fluorescent stains, CMAC, CMAC-Arg,CMAC-Ala-Pro and MDY-64 (all from Invitrogen), were usedto localize and determine the function of vacuolar proteases.Fungal cells were incubated under the described conditions,then incubated according to the manufacturer’s directions,then embedded in 2% low-molecular-weight agarose under acoverslip, allowed 10 min to settle at 42°C, then cooled to 4°Con a Perkin Elmer PCR machine. Cells were then examinedby epifluorescence and Nomarski or bright field optics usingan Olympus IX-70 inverted microscope at optical wave-lengths recommended for each dye according to themanufacturer.
Budding index and determination of cellular DNAcontent
Budding index was determined similar to the method of Zettelet al. (2003). Briefly, cells were harvested, vortexed for 30 sand the proportion of adherent buds measured. Methods forthe determination of cellular DNA content was adapted fromthat by Ohkusu et al. (2001). Briefly, yeast cells were grown tomid-log phase and either harvested or washed twice in steriledistilled water and incubated for 24 h in YNB without aminoacids and ammonium sulphate at 37°C in an orbital shaker.Harvested cells were washed once with sterile distilled waterand resuspended in ice-cold 70% ethanol for fixation andstored at 4°C. Aliquots (1 ml) of the fixed cells were centri-fuged in a microcentrifuge at 13 000 g for 30 s and resus-pended in 1 ml of PBS and allowed to rehydrate for 5 min.Cells were centrifuged again and resuspended in 1 ml ofpropidium iodide (PI) staining solution (0.1% Triton X-100,0.2 mg ml-1 DNAse-free RNAse, 5 mg ml-1 PI in PBS) andincubated at room temperature for 30 min. Stained cells werekept on ice until flow cytometry was performed. DNA contentwas examined in each cell using a laser-scanning-BeckmanCoulter 500 cytometer with peak versus interval used to gateour aggregates with a 590 dichoic and a 610 fixed band pathfilter. DNA of cells were visualized by epifluorescence usingthe soft agar embedding technique described above afterincubation with 1 mg ml-1 DAPI (Sigma-Aldrich) in PBS,pH 7.0. Assessment for metabolic activity was performed byincubating log-phase cells in YNB without amino acids andammonium sulphate for 1.5 h at 30°C, followed by a 30 minincubation in the presence of 20 mM FUN 1 (Invitrogen) andobserved for red staining vesicles according to the manufac-turer’s directions.
Statistics
Statistical significance of mouse survival times was assessedby Kruskall–Wallis analysis (ANOVA on Ranks). Pairwiseanalyses were performed post hoc by using Dunn’sprocedure. Budding index comparisons were performed by achi-square two-sided test using a 2 ¥ 2 contingency table.
Acknowledgements
This work was supported, in part, by United States PublicHealth Service Grants NIH AI49371 and AI45995 (to P.R.W.).
VPS41 of Cryptococcus 1143
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 61, 1132–1146

We also thank TIGR (supported by NIH Grant 1 UO1AI48594) and the Fungal Genomic Initiative for sequenceinformation for the C. neoformans genome.
References
Achstetter, T., Ehmann, C., and Wolf, D. (1983) Proteolysis ineucaryotic cells: aminopeptidases and dipeptidyl ami-nopeptidases of yeast revisited. Arch Biochem Biophys226: 292–305.
Akhter, S., McDade, H.C., Gorlach, J.M., Heinrich, G., Cox,G.M., and Perfect, J.R. (2003) Role of alternative oxidasegene in pathogenesis of Cryptococcus neoformans. InfectImmun 71: 5794–5802.
Barton, C., Whitehead, S., and Blackwell, J. (1995) Nramptransfection transfers Ity/Lsh/Bcg-related pleiotropic effectson macrophage activation: influence on oxidative burst andnitric oxide pathways. Mol Med 1: 267–279.
Blasi, E., Colombari, B., Mucci, A., Cossarizza, A., Radzioch,D., Boelaert, J.R., and Neglia, R. (2001) Nramp1 geneaffects selective early steps in macrophage-mediated anti-cryptococcal defense. Med Microbiol Immunol (Berl) 189:209–216.
Bryant, N. (1998) Vacuole biogenesis in Saccharomyces cer-evisiae: protein transport to the yeast vacuole. Micro MolBiol Rev 62: 230–247.
Bulmer, G.S., and Sans, M.D. (1967) Cryptococcusneoformans. II. Phagocytosis by human leukocytes. J Bac-teriol 94: 1480–1483.
Casadevall, A., and Perfect, J. (1998) Cryptococcusneoformans. Washington, DC: American Society for Micro-biology Press.
Chung, J., Haile, D., and Wessling-Resnick, M. (2004)Copper-induced ferroportin-1 expression in J774 macroph-ages is associated with increased iron efflux. Proc NatlAcad Sci USA 101: 2700–2705.
Cox, G.M., Toffaletti, D.L., and Perfect, J.R. (1996) Dominantselection system for use in Cryptococcus neoformans. JMed Vet Mycol 34: 385–391.
Cox, G.M., Mukherjee, J., Cole, G.T., Casadevall, A., andPerfect, J.R. (2000) Urease as a virulence factor in experi-mental cryptococcosis. Infect Immun 68: 443–448.
Cox, G.M., McDade, H.C., Chen, S.C., Tucker, S.C., Got-tfredsson, M., Wright, L.C., et al. (2001) Extracellular phos-pholipase activity is a virulence factor for Cryptococcusneoformans. Mol Microbiol 39: 166–175.
Cox, G.M., Harrison, T.S., McDade, H.C., Taborda, C.P.,Heinrich, G., Casadevall, A., and Perfect, J.R. (2003)Superoxide dismutase influences the virulence of Crypto-coccus neoformans by affecting growth withinmacrophages. Infect Immun 71: 173–180.
Cruz, M.C., Goldstein, A.L., Blankenship, J., Del Poeta, M.,Perfect, J.R., McCusker, J.H., et al. (2001) Rapamycin andless immunosuppressive analogs are toxic to Candidaalbicans and Cryptococcus neoformans via FKBP12-dependent inhibition of TOR. Antimicrob AgentsChemother 45: 3162–3170.
Darsow, T., Katzmann, D., Cowles, C., and Emr, S. (2001)Vps41p function in the alkaline phosphatase pathwayrequires homo-oligomerization and interaction with
AP-3 through two distinct domains. Mol Biol Cell 12:37–51.
Davis-Kaplan, S., Askwith, C., Bengtzen, A., Radisky, D., andKaplan, J. (1998) Chloride is an allosteric effector of copperassembly for the yeast multicopper oxidase Fet3p: anunexpected role for intracellular chloride channels. ProcNatl Acad Sci USA 95: 13641–13645.
Eksi, S., Czesny, B., Greenbaum, D., Bogyo, M., and Will-iamson, K. (2004) Targeted disruption of Plasmodium fal-ciparum cysteine protease, falcipain 1, reduces oocystproduction, not erythrocytic stage growth. Mol Microbiol 53:243–250.
Erickson, T., Liu, L., Gueyikian, A., Zhu, X., Gibbons, J., andWilliamson, P.R. (2001) Multiple virulence factors of Cryp-tococcus neoformans are dependent on VPH1. Mol Micro-biol 42: 1121–1131.
Fan, W., Kraus, P., Boily, M., and Heitman, J. (2005) Cryp-tococcus neoformans gene expression during murine mac-rophage infection. Eukaryot Cell 4: 1420–1433.
Fang, C.T., Hung, C.C., Chang, S.C., Hsueh, P.R., Chang,Y.L., Chen, M.Y., and Luh, K.T. (2000) Pulmonary infectionin human immunodeficiency virus-infected patients inTaiwan. J Formos Med Assoc 99: 123–127.
Feldmesser, M., Tucker, S., and Casadevall, A. (2001a) Intra-cellular parasitism of macrophages by Cryptococcusneoformans. Trends Microbiol 9: 273–278.
Feldmesser, M., Kress, Y., and Casadevall, A. (2001b)Dynamic changes in the morphology of Cryptococcus neo-formans during murine pulmonary infection. Microbiology147: 2355–2365.
Fromtling, R.A., and Shadomy, H.J. (1982) Immunity in cryp-tococcosis: an overview. Mycopathologia 77: 183–190.
Gao, X., Yo, P., Keith, A., Ragan, T., and Harris, T. (2003)Thermodynamically balanced inside-out (TBIO) PCR-based gene synthesis: a novel method of primer design forhigh-fidelity assembly of longer gene sequences. NucleicAcid Res 31: e143.
Iida, H., and Yahara, I. (1984) Specific early-G1 blocksaccompanied with stringent response in Saccharomycescerevisiae lead to growth arrest in resting state similar tothe G0 of higher eucaryotes. J Cell Biol 98: 1185–1193.
de Jesus-Berrios, M., Liu, L., Nussbaum, J.C., Cox, G.M.,Stamler, J.S., and Heitman, J. (2003) Enzymes that coun-teract nitrosative stress promote fungal virulence. Curr Biol13: 1963–1968.
Kamada, Y., Sekito, T., and Ohsumi, Y. (2004) Autophagy inyeast: a TOR-mediated response to nutrient starvation.Curr Top Microbiol Immunol 279: 73–84.
Klionsky, D. (2004) The molecular machinery of autophagy:unanswered questions. J Cell Sci 118: 7–18.
LaGrassa TJ, U.C. (2005) The vacuolar kinase Yck3 main-tains organelle fragmentation by regulating the HOPS teth-ering complex. J Cell Biol 168: 401–414.
Lang, T., Schaeffeler, E., Bernreuther, D., Bredschneider, M.,Wolf, D., and Thumm, M. (1998) Aut2p and Aut7p, twonovel microtubule-associated proteins are essential fordelivery of autophagic vesicles to the vacuole. EMBO J 17:3597–3607.
Liu, L., Tewari, R.P., and Williamson, P.R. (1999a) Laccaseprotects Cryptococcus neoformans from antifungal activityof alveolar macrophages. Infect Immun 67: 6034–6039.
1144 X. Liu, G. Hu, J. Panepinto and P. R. Williamson
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 61, 1132–1146

Liu, L., Wakamatsu, K., Ito, S., and Williamson, P.R. (1999b)Catecholamine oxidative products, but not melanin, areproduced by Cryptococcus neoformans during neuro-pathogenesis in mice. Infect Immun 67: 108–112.
Lorenz, M., and Fink, G. (2001) The glyoxylate cycle isrequired for fungal virulence. Nature 412: 83–86.
Luberto, C., Toffaletti, D.L., Wills, E.A., Tucker, S.C., Casa-devall, A., Perfect, J.R., et al. (2001) Roles for inositol-phosphoryl ceramide synthase 1 (IPC1) in pathogenesis ofC. neoformans. Genes Dev 15: 201–212.
Martinez, L.R., and Casadevall, A. (2005) Specific antibodycan prevent fungal biofilm formation and this effect corre-lates with protective efficacy. Infect Immun 73: 6350–6362.
Missall, T.A., and Lodge, J.K. (2005) Thioredoxin reductaseis essential for viability in the fungal pathogen Cryptococ-cus neoformans. Eukaryot Cell 4: 487–489.
Missall, T.A., Pusateri, M.E., and Lodge, J.K. (2004) Thiolperoxidase is critical for virulence and resistance to nitricoxide and peroxide in the fungal pathogen, Cryptococcusneoformans. Mol Microbiol 51: 1447–1458.
Mondon, P., Chang, Y.C., Varma, A., and Kwon-Chung, K.J.(2000) A novel episomal shuttle vector for transformation ofCryptococcus neoformans with the ccdB gene as a positiveselection marker in bacteria. FEMS Microbiol Lett 187:41–45.
Mukherjee, S., Feldmesser, M., and Casadevall, A. (1996)J774 murine macrophage-like cell interactions with Cryp-tococcus neoformans in the presence and absence ofopsonins. J Infect Dis 173: 1222–1231.
Narasipura, S.D., Ault, J.G., Behr, M.J., Chaturvedi, V., andChaturvedi, S. (2003) Characterization of Cu, Zn superox-ide dismutase (SOD1) gene knock-out mutant of Crypto-coccus neoformans var. gattii: role in biology and virulence.Mol Microbiol 47: 1681–1694.
Noda, T., and Ohsumi, Y. (1998) Tor, a phosphatidylinositolkinase homologue, controls autophagy in yeast. J BiolChem 273: 3963–3966.
Odom, A., Muir, S., Lim, E., Toffaletti, D.L., Perfect, J., andHeitman, J. (1997) Calcineurin is required for virulence ofCryptococcus neoformans. EMBO J 16: 2576–2589.
Ohkusu, M., Raclavsky, V., and Takeo, K. (2001) Deficit inoxygen causes G(2) budding and unbudded G(2) arrest inCryptococcus neoformans. FEMS Microbiol Lett 204:29–32.
Ohkusu, M., Raclavsky, V., and Takeo, K. (2004) Inducedsynchrony in Cryptococcus neoformans after release fromG2-arrest. Antonie Van Leeuwenhoek 85: 37–44.
Panepinto, J., and Williamson, P.R. (2006) Intersection offungal fitness and virulence in Cryptococcus neoformans.FEMS Yeast Res (in press).
Panepinto, J., Liu, L., Ramos, J., Zhu, X., Valyi-Nagy, T.,Eksi, S., et al. (2005) The DEAD-box RNA helicase Vad1regulates multiple virulence-associated genes in Crypto-coccus neoformans. J Clin Invest 115: 632–641.
Perfect, J.R., Wong, B., Chang, Y.C., Kwon-Chung, K.J., andWilliamson, P.R. (1998) Cryptococcus neoformans: viru-lence and host defences. Med Mycol 36 (Suppl. 1): 79–86.
Radisky, D., Snyder, W., Emr, S., and Kaplan, J. (1997)Characterization of VPS41, a gene required for vacuolartrafficking and high-affinity iron transport in yeast. Proc NatlAcad Sci USA 94: 5662–5666.
Roberts, C., Raymond, C., Yamashiron, C., and Stevens, T.(1991) Methods for studying the yeast vacuole. MethEnzymol 194: 644–661.
Salas, S.D., Bennett, J.E., Kwon-Chung, K.J., Perfect, J.R.,and Williamson, P.R. (1996) Effect of the laccase geneCNLAC1, on virulence of Cryptococcus neoformans. J ExpMed 184: 377–386.
Shoji, J., Arioka, M., and Kitamoto, K. (2006) Vacuolar mem-brane dynamics in the filamentous fungus Aspergillusoryzae. Eukaryot Cell 5: 411–421.
Straube, A., Brill, B., Oakley, R., Horio, T., and Steinberg, G.(2003) Microtubule organization requires cell cycle depen-dent nucleation at dispersed cytoplasmic sites, polar andperinuclear MTOCs in the plant pathogen Ustilago maydis.Mol Biol Cell 14: 642–657.
Suarez-Rendueles, M., Schwenche, J., Garcia Alvarez, N.,and Gascon, S. (1981) A new X-prolyl-dipeptidyl ami-nopeptidase from yeast associated with a particulatefraction. FEBS Lett 131: 296–300.
Tucker, S.C., and Casadevall, A. (2002) Replication of Cryp-tococcus neoformans in macrophages is accompanied byphagosomal permeabilization and accumulation of vesiclescontaining polysaccharide in the cytoplasm. Proc Natl AcadSci USA 99: 3165–3170.
Uhl, M., Biery, M., Craig, N., and Johnson, A. (2003)Haploinsufficiency-based large-scale forward geneticanalysis of filamentous growth in the diploid human fungalpathogen C. albicans. EMBO J 22: 2668–2678.
Varma, A., and Kwon-Chung, K.J. (1991) Rapid method toextract DNA from Cryptococcus neoformans. J Clin Micro-biol 29: 810–812.
Varma, A., Edman, J.C., and Kwon-Chung, K.J. (1992)Molecular and genetic analysis of URA5 transformants ofCryptococcus neoformans. Infect Immun 60: 1101–1108.
Wang, Y., Aisen, P., and Casadevall, A. (1995) Cryptococcusneoformans melanin and virulence: mechanism of action.Infect Immun 63: 3131–3136.
Ward, D., Radisky, D., Scullion, M., Tuttle, M., Vaughn, M.,and Kaplan, J. (2001) hVPS41 is expressed in multipleisoforms and can associate with vesicles through aRING-H2 finger motif. Exp Cell Res 267: 126–134.
Werner-Washburne, M., Braun, E., Crawford, M., and Peck,V. (1996) Stationary phase in Saccharomyces cerevisiae.Mol Microbiol 19: 1159–1166.
Wu, J., Dang, Y., Su, W., Liu, C., Ma, H., Shan, Y., et al.(2006) Molecular cloning and characterization of rat LC3Aand LC3B – two novel markers of autophagosome.Biochem Biophys Res Commun 339: 437–442.
Xie, M., Jin, F., Hwang, H., Hwang, S.K., Anand, V.K.,Duncan, M., and Huang, J. (2005) Insights into TOR func-tion and rapamycin response: chemical genomic profilingby using a high-density cell array method. PNAS 102:7215–7220.
Zaragoza, D., Ghavidel, A., Heitman, J., and Schultz, M.(1998) Rapamycin induces the G0 program of transcrip-tional repression in yeast by interfering with the TOR sig-naling pathway. Mol Cell Biol 18: 4463–4470.
Zettel, M., Garza, L., Cass, A., Myhre, R., Haizlip, L., Osadebe,S., et al. (2003) The budding index of Saccharomycescerevisiae deletion strains identifies genes important for cellcycle progression. FEMS Microbiol Lett 27: 253–258.
VPS41 of Cryptococcus 1145
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 61, 1132–1146

Zhu, X., and Williamson, P.R. (2003) A CLC-type chloridechannel gene is required for laccase activity and virulencein Cryptococcus neoformans. Mol Microbiol 50: 1271–1281.
Zhu, X., Gibbons, J., Garcia-Rivera, J., Casadevall, A., andWilliamson, P.R. (2001) Laccase of Cryptococcus neofor-mans is a cell wall-associated virulence factor. InfectImmun 69: 5589–5596.
Zhu, X., Gibbon, J., Zhang, S., and Williamson, P. (2002)Copper-mediated reversal of defective laccase in a vph1avirulent mutant of Cryptococcus neoformans. Mol Micro-biol 47: 1007–1014.
Zhu, X., Gibbons, J., Zhang, S., and Williamson, P.R. (2003)Copper-mediated reversal of defective laccase in aDeltavph1 avirulent mutant of Cryptococcus neoformans.Mol Microbiol 47: 1007–1014.
1146 X. Liu, G. Hu, J. Panepinto and P. R. Williamson
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 61, 1132–1146
Copyright © 2022 FDOKUMEN




















