
Matrilysin Cleavage of Corneal Collagen Type XVIII NC1 Domain and Generation of a 28-kDa Fragment
Upload
independentCategory
view
2download
0
h t . J. Cancer: 55,10-18 (1993) Publication of the International Union Against Cancer Publication de I'Union Internationale Contre le Cancer
0 1993 Wiley-Liss, Inc.
ROLE AND REGULATION OF EXPRESSION OF 92-kDa TYPE-IV
CELL LINES OF THE ORAL CAVITY COLLAGENASE (MMP-9) IN 2 INVASIVE SQUAMOUS-CELL-CARCINOMA
Jose JUAREZ~, Gary CLAY MAN^, Motowo NAKAJIMA3, Kenneth K. TANABE4, Hideyuki SAYAS, Garth L. NICOLSON~ and Douglas BoYD'.~ Departments of 'Tumor Biology and 2Head and Neck Surgery, M.D. Anderson Cancer Center, Houston TX 77030, USA; 31nstitute of Applied Microbiology, University of Tokyo, Japan; and Departments of 4General Surgery and sNeuro-Oncologv, M.D. Anderson Cancer Center, Houston TX 77030, USA.
The present study was undertaken to determine the role of the metalloproteinase MMP-9 in the invasive phenotype of squamous-cell carcinoma of the oral cavity and the regulation of i ts expression. Zymographic analysis of conditioned medium from 2 highly invasive squamous-cell-carcinoma cell lines indi- cated large amounts of an enzyme which was indistinguishable, in size (92 kDa) from the MMP-9 pro-enzyme. Conversion of the 92-kDa gelatinase into a lower-molecular-weight species (84 kDa), identical in size to the activated gelatinase, was evident when both cell lines, which are avid secreton of urokinase, were cultured in the presence of plasminogen. Penetration of an extracellular-matrix-coated filter was dramatically reduced in the presence of the collagenase inhibitor, tissue inhibitor of metalloproteinase-2, suggesting a critical role for MMP-9 in the invasive process. lmmunohistochemical studies demonstrating the presence of MMP-9 in tumor cells of resected squamous-cell cancers suggested that secretion of this collagenase by cells in vitro was reflective of the in vivo setting. Since several phorbol- ester response elements are present in the MMP-9 promoter, we determined the role of protein-kinase-C pathways in the regulation of MMP-9 expression in cultured SCC. Treatment of cells with PMA resulted in a more-than-20-fold increase in the level of protein and mRNA. Conversely, culturing of cells in the presence of the protein-kinase-C inhibitor, calphostin-C, led to a dose-dependent decrease in the amount of MMP-9 mRNA and protein, suggesting that the constitutive expression of this collagenase reflects activation of this signal transduction path- way. In summary, our data suggest that, for a sub-population of squamous-cell carcinomas, secreted MMP-9 is an important determinant of the invasive phenotype, and that the expression of this metalloproteinase is regulated by protein-kinase-C pathways. o 1993 Wiley-Liss, Inc.
Disease control and survival among patients with advanced squamous-cell carcinoma of the oral cavity (SCCOC) has not improved since surgery and radiotherapy were first combined to treat the disease. The poor prognosis of those patients with advanced disease most often reflects the spread of the disease to local, regional and distant sites, events that are poorly controlled by combined surgery/irradiation. Moreover, at- tempts to control local disease utilizing extensive surgical resection results in several undesirable effects, including impair- ment of key physiological functions such as speech and swallowing, as well as having adverse cosmetic effects. There- fore, studies addressing the mechanism(s) underlying the invasiveness of SCCOC may lead to the development of novel treatments that attenuate the invasiveness of these tumors and consequently improve the quality of life and survival of patients afflicted with the disease.
The process of tumor invasion by SCCOC involves degrada- tion of the underlying basement membrane, which is largely made up of type-IV collagen (Hirota et al., 1990; Tryggvason et al., 1987). For SCCOC, hydrolysis of this collagen is a probable prerequisite for acquisition of an invasive phenotype (Naka- jima et al., 1987; Tryggvason et al., 1987). This can be accomplished by the collagenases MMP-2 (72-kDa type-IV collagenase), MMP-3 (stromelysin) and MMP-9 (92-kDa type-IV collagenase), which are encoded by separate genes
(Bernhard et al., 1990; Garbisa et al., 1987; Schultz et al., 1988). These hydrolases have been implicated in tumor invasion/ metastasis because (i) their release correlates well with the invasive and metastatic phenotype in diverse tumor types (Bernhard et al., 1990); (ii) inhibition of the expression or activity of these enzymes leads to reduced invasive and metastatic capacity (Schultz et al., 1988); and (iii) expression of an anti-sense cDNA to the type-IV-collagenase inhibitor TIMP promotes invasion and metastatic behavior in 3T3 cells (Khokha et al., 1989).
At present, the proteolytic mechanism(s) responsible for breach of the basement membrane by SCCOC are poorly understood. Kusukawa et al. (1992) concluded that both MMP-2 and MMP-3 played a role in this process, since both of these metalloproteinases were present in conditioned medium from 2 cultured SCCOC cell lines. However, to date there have been no studies addressing the potential role of the 92-kDa type-IV collagenase (MMP-9) in invasion by SCCOC. We present data herein indicating a role for MMP-9 in the invasive phenotype as well as evidence for a role of protein kinase C pathways in the regulation of expression of MMP-9 in SCCOC.
METHODS Material
Transwell plates (12-pm pore size) were purchased from Costar (Cambridge, MA). Matrigel was provided by Collabora- tive Research (Bedford, MA). The peptide LGRFQTFEG- DLKWHH was prepared by the peptide-synthesizing facility at M.D. Anderson Cancer Center and was determined to be more than 80% pure by HPLC. Immunohistochemical re- agents were obtained from Biogenics (San Ramon, CA). Plasmin was obtained from American Diagnostic (Greenwich, CT). TIMP-2 was kindly provided by Dr. Stetler-Stevenson at the NIH. PMA (phorbol 12-myristate 13-acetate) was pur- chased from Sigma (St. Louis, MO). Calphostin-C was ob- tained from Calbiochem (La Jolla, CA).
Cell culture The SCCOC cancer cell line UM-SCC-1, derived from the
oral cavity, was obtained from Dr. T. Carey at the University of Michigan. MDA-TU-138 were established at this institution
6To whom correspondence and reprint requests should be ad- dressed, at Department of Tumor Biology, Box 108, The University of Texas M.D. Anderson Cancer Center, 1515 Holcombe Blvd, Houston, TX 77030, USA. Fax: (713) 794 0209.
Abbreviations: EGF, epidermal growth factor; GAPDH, glyceralde- hyde phosphate dehydrogenase; MMP-9,92-kDa type-IV collagenase; MTT, 3-[4,5-dimethylthiazol-2-y1]-2,5-diphenyltetrazolium bromide; PMA, phorbol 12-myristate 13-acetate; SF, serum-free; SCCOC, squamous-cell carcinoma of the oral cavity; TIMP-2, type-2 tissue inhibitor of metalloproteinase.
Received: January 27,1993 and in revised form March 29,1993.

MMP-9 EXPRESSION IN SQUAMOUS-CELL CARCINOMA 11
35
30
- 25 u z 20 0 2 1 5 t
v
2 10
from a highly invasive tumor of the oral cavity (Clayman et al., 1993). All cells were maintained in 5% (v/v) serum-containing medium in McCoy’s 5A medium. For the collection of condi- tioned medium, UM-SCC-1 cells grown to 90% confluence, were changed to fresh McCoy’s 5A medium supplemented with 4 pg/ml transferrin, 10 ng/ml E G F and 5 pg/ml insulin (serum-free medium). Conditioned medium was collected 24 hr later and cellular debris was removed by centrifugation at
Experimental invasion Invasion assays were carried out as described by Schlechte et
al. (1990). Briefly, Matrigel was diluted in cold S F medium to a protein concentration of 1.0 mg/ml and supplemented with 25 p,g/ml plasminogen. The preparation was added to the porous filters (pore size 12 pm) and allowed to gel at 37°C. Cells were dispersed with Ca2+ /MgZ+-free medium containing 5 mM EDTA and 1.5 X lo5 cells dispensed into the Transwell insert, which was subsequently placed into the outer well, the latter also containing culture medium. The cells were incubated at 37°C for 1 day, after which both compartments were supple- mented with 0.2 mg/ml of the vital stain MTT and incubated at 37°C. The amount of vital stain converted to water-insoluble crystals is representative of the number of cells (Mossman, 1983). Crystals on the lower and upper aspects of the filter were removed with filter paper and dissolved in dimethylsulfox- ide. Color intensity of the solubilized crystals was determined spectrophotometrically at 570 nm.
Zymographic assays for collagenase (gelatinase) Conditioned medium was concentrated 20- to 30-fold and
equal amounts, based on cell numbers determined a t the time of harvest, were denatured under non-reducing conditions. Samples were electrophoresed in a 7.5% SDS-PAGE gel containing 0.1% (w/v) type-A porcine skin gelatin. The gel was
2OOga, ’
-
:
\
-
washed for 3 hr a t room temperature in a solution containing 2.5% (v/v) Triton X-100, 50 mM Tris-HC1, p H 7.5, and subsequently transferred to a buffer containing 10 mM CaCI2, 0.15 M NaCI, 50 mM Tris-HCI, p H 7.5, and incubated at 37°C overnight. The gel was stained for 45 min with 0.25% (w/v) Coomassie blue in 45% (v/v) methanol/l% (v/v) acetic acid and de-stained in 10% acetic acid (v/v)/25% methanol (viv).
MMP- 9 probe mRNA was extracted from MCF-7 cells and cDNA gener-
ated with reverse transcriptase. The MMP-9 cDNA was amplified by PCR (32 cycles of denaturation at 94°C for 1 min, annealing at 60°C for 1 min and extension at 72°C for 1 min) using 25 U/ml Taq DNA polymerase and the oligonucleotide primers (5’ GACCTCTGCAAGGCTTTCAA-3’ and 5’ CAC- TGGGGTGGAATGTGTCT-3’) that flank nucleotides 1178 to 1699 of the published MMP-9 sequence (Huhtala et al., 1991). This amplified region contains no sequence homology with any known rnetalloproteinases. The PCR amplification product was cloned into pCRlOOO (Invitrogen, San Diego, CA) and the identity of the product was confirmed by DNA sequence analysis.
Northern blotting The level of steady-state transcript for MMP-9 was deter-
mined by Northern analysis (Boyd et al., 1989). Total cellular R N A was extracted from 90%-confluent cultures using 5.0 M guanidinium isothiocyanate and purified on a cesium chloride cushion (5.7 M) by centrifugation at 150,000 g for 20 hr. Purified R N A was electrophoresed in a 1.5% (wiv) formalde- hyde-agarose gel and transferred to Nytran modified nylon by capillary action using 10 x SSC. The Northern blot was probed at 42°C with a random primed radiolabelled 520-base pair (extending from 1178 to 1699) cDNA specific for MMP-9
.i 0 4 10 20 40 40 ( n M ) 0
-TIMP-2- E z 0 V 0
0 * 3
2 s
FIGURE 1 - Matrigel invasion by cultured SCCOC is inhibited by TIMP-2 which is directed at the catalytic site of the metalloprotein- ase. UM-SCC-1 cells (1.5 x lo5) were cultured on polycarbonate filters (12-pm pore size) coated with plasminogen-impregnated Matrigel. TIMP-2 was added both to the upper and the lower chambers 3 hr after plating. Cultures were maintained for 24 hr and assayed for invasion by metabolism of the vital stain, MTT. Invasion was defined as the accumulation of cells on the lower aspect of the membrane and in the outer well, as a function of the total number of cells. The data are shown as average values t S.D. for 3 experiments.
FIGURE 2 - Zymographic analysis of collagenase (gelatinolytic) activity in conditioned medium derived from cultured SCCOC cell lines. Conditioned medium derived from the indicated cell lines was concentrated 25- to 30-fold and equal amounts, based on cell number equivalents, electrophoresed (non-reducing conditions) in a 7.5% SDS-PAGE gel containing 0.1% (wiv) elatin. The gel was subsequently treated for 3 hr with 2.5% (v/vT Triton X-100, incubated overnight in a solution containing 10 mM CaC12 and stained with 0.25% (w/v) Coomassie blue. Gelatinolytic (collage- nase) activity is indicated by the white zones where the gelatin has been hydrolyzed. Molecular weights are indicated to the left. The data are representative of more than 5 determinations.
12 JUAREZ ET AL.
mRNA (Huhtala et al., 1991). Stringency washes were under- taken at 65°C using 0.50 x SSC in the presence of 1.0% (wiv) SDS. Loading efficiencies were checked by reprobing the blot with a radioactive cDNA which hybridizes with the glyceralde- hyde-3-phosphate dehydrogenase mRNA (GAPDH).
Immunohistochemical staining of tumor sections for MMP-9 Tissue sections (4 pm) were cut from paraffin blocks,
rehydrated and treated with normal goat serum. The sections were incubated with a 1 : l O O dilution of rabbit anti-serum raised against the peptide LGRFQTFEGDLKWHH present in the published sequence of MMP-9 (Wilhelm et al., 1989) or with anti-serum which had been pre-adsorbed with 10 pg/ml of the peptide antigen. Subsequently, the sections were incu- bated with biotinylated anti-rabbit IgG and alkaline phospha- tase conjugated to streptavidin. Immunoreactive complexes were visualized with fast red chromogen. Washes with PBS were performed between steps.
Western blotting Western blotting was carried out as described by Hollas et al.
(1991). Equal amounts of conditioned medium (based on cell number), which was concentrated 20- to 30 fold, was dena- tured at 100°C in 2 x SDS gel-loading buffer containing 100
mMTris.HC1 (pH 6.8), 2.3% (w/v) SDS, 0.2% (wiv) bromophe- no1 blue, and 20% (v/v) glycerol. The denatured material was electrophoresed in a 7.5% polyacrylamide gel and the resolved proteins transferred to a nitrocellulose filter. The filter was blocked with PBS containing 0.05% (v/v) Tween 20 (PBS/ Tween), and 3% (wiv) BSA and incubated either with a 1500 dilution of rabbit anti-serum to MMP-9 or with the anti-serum that had been pre-adsorbed with 25 pg/ml of the immunizing peptide (LGRFQTFEGDLKWHH) a t room temperature for 1 hr. Reactive bands were visualized by incubating the filter with 5 x lo4 dpm/ml of radioactive protein A. The filter was washed extensively with PBSITween and exposed to X-ray film.
RESULTS Invasive cultured SCCOC cells secrete large amounts of MMP-9
To investigate the function of MMP-9 in invasion, we made use of 2 cultured SCCOC cell lines, UM-SCC-1 and MDA-TU- 138, derived from highly invasive tumors in vivo. Both cell lines were very active in invading extracellular matrix coated filters, in vitro (Fig. l ) , similarly to their behavior in vivo. In a 24-hr period, over 25% of the cells penetrated the barrier. Higher
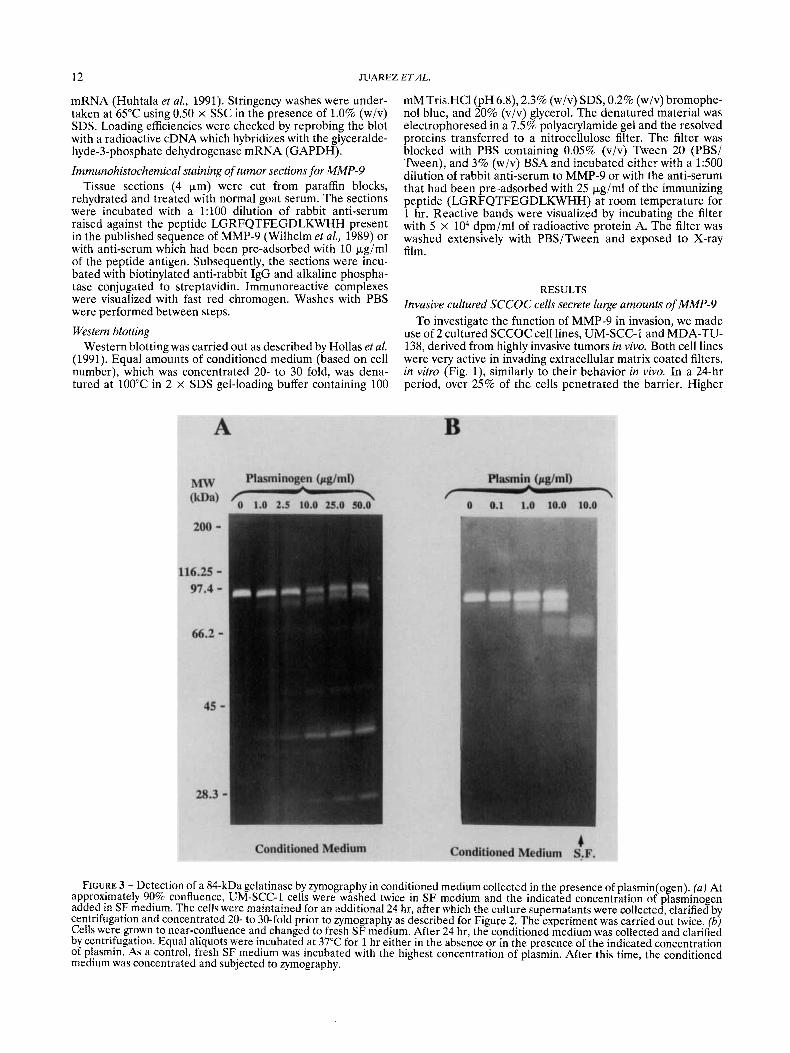
FIGURE 3 - Detection of a 84-kDa gelatinase by zymography in conditioned medium collected in the presence of plasmin(o en) (a) At approximately 90% confluence, UM-SCC-1 cells were washed twice in SF medium and the indicated concentration of pfasminogen added in SF medium. The cells were maintained for an additional 24 hr, after which the culture supernatants were collected, clarified by centrifugation and concentrated 20- to 30-fold prior to zymography as described for Figure 2. The experiment was carried out twice. (b) Cells were grown to near-confluence and changed to fresh SF medium. After 24 hr, the conditioned medium was collected and clarified by centrifugation. Equal aliquots were incubated at 37°C for 1 hr either in the absence or in the presence of the indicated concentration of plasmin. As a control, fresh SF medium was incubated with the highest concentration of plasmin. After this time, the conditioned medium was concentrated and subjected to zymography.

MMP-9 EXPRESSION IN SQUAMOUS-CELL CARCINOMA 13
rates of invasion were observed with longer incubation peri- ods; however, due to the fragility of the Matrigel layer, the assays were terminated after 1 day.
Zymographic analysis of conditioned medium from both cell lines grown on plastic indicated gelatinase (collagenase) activ- ity; that on the basis of its molecular weight (92 kDa) was indistinguishable from the 92-kDa type-IV collagenase (MMP-9) (Davis and Martin, 1990; Wilhelm et al., 1989) (Fig. 2). No other type of collagenase, including the 72-kDa type-IV Collagenase, MMP-2, could be detected in either SCCOC cell h e , whether grown on plastic or on Matrigel (Fig. 2). The inability to detect MMP-2 did not reflect technical limitations, since a molecule of this size was detected in conditioned medium derived from fibroblasts established from a primary tumor of the oral cavity (Fig. 2).
Although secreted MMP-9 derived from cells grown on plastic was predominantly in the pro-enzyme form, as evident from its molecular weight (92 kDa), analysis of conditioned medium collected from the in vitro invasion assays indicated an 84-kDa species (data not shown) identical in size to the activated metalloproteinase (Davis and Martin, 1990; Wilhelm et al., 1989). Since the in v i m invasion assays are routinely performed in the presence of plasminogen (Schlechte et al., 1990), and other investigators have reported activation of collagenase by plasmin (Salo et al., 1982), we conducted
experiments to determine whether generation of the 84-kDa species was plasminogen-dependent. Conditioned medium was collected from UM-SCC-1 cells grown on plastic for 24 hr, with varying concentrations of plasminogen, covering the physiological range (Collen and Verstraete, 1975), and sub- jected to zymography. Whereas the only activity detected in conditioned medium collected from UM-SSC-1 cells cultured alone was that of the 92-kDa species, inclusion of plasminogen in the culture medium led to the appearance of an 84-kDa species. The amount of the 84-kDa species increased as a function of the input level of plasminogen, this increase being paralleled by a decrease in the level of 92-kDa gelatinase (Fig. 3a). Parallel experiments using plasminogen collected in SF medium (without cells) ruled out the possibility that the 84-kDa band was a consequence of the direct hydrolysis of the gelatin in the zymogram by plasmin(ogen). Generation of the 84-kDa gelatinase could also be achieved when conditioned medium, collected in the absence of plasminogen, was subse- quently treated with plasmin (Fig. 3b). Since the SCCOC cell lines used in these studies are avid secretors of urokinase (UM-SCC-1 and MDA-TU-138 cells secrete approximately 750 ng/(106 cells.24 hr); Clayman et al., 1993) it is likely that tumor-derived urokinase converts the circulating zymogen plasminogen into plasmin, the latter being capable of process- ing the 94-kDa species into the 84-kDa gelatinase.
FIGURE 4 - Immunohistochemical localization of MMP-9 in resected SCCOC. De-paraffinized tissue sections (4 pm) were incubated with 1:lOO dilution of a rabbit anti-serum to MMP-9 (a,c) for 1 hr, or with the anti-serum which had been pre-adsorbed for 1 hr with 20 pgirnl of the peptide LGRFQTFEGDLKWHH (b). Immunoreactive complexes were visualized using alkaline phosphatase conjugated to streptavadin and fast red chromogen as substrate. Washes were performed with PBS. s, stroma; t, tumor cell. Scale bar, 150 ym. These data are representative of SCCOC tissue sections derived from 4 patients.
14 JUAREZ ETAL.
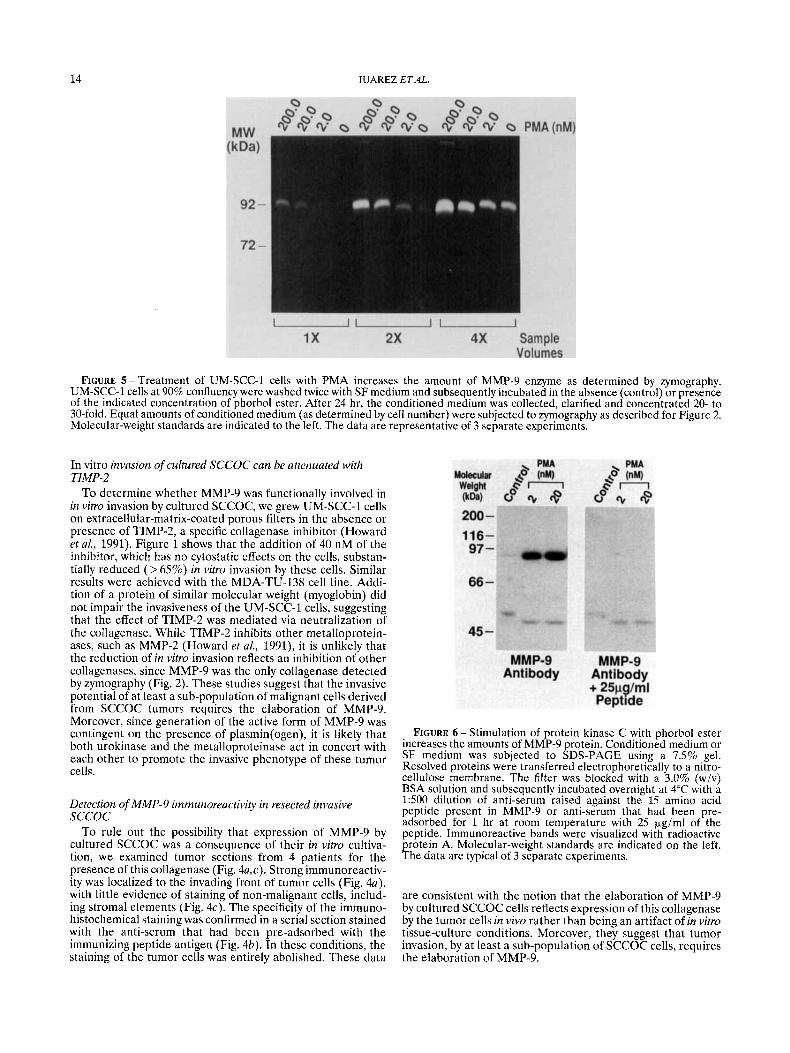
FIGURE 5 -Treatment of UM-SCC-1 cells with PMA increases the amount of MMP-9 enzyme as determined by zymography. UM-SCC-1 cells at 90% confluency were washed twice with SF medium and subsequently incubated in the absence (control) or presence of the indicated concentration of phorbol ester. After 24 hr, the conditioned medium was collected, clarified and concentrated 20- to 30-fold. Equal amounts of conditioned medium (as determined by cell number) were subjected to zymography as described for Figure 2. Molecular-weight standards are indicated to the left. The data are representative of 3 separate experiments.
In vitro invasion of cultured SCCOC can be attenuated with TZMP-2
To determine whether MMP-9 was functionally involved in in vitro invasion by cultured SCCOC, we grew UM-SCC-1 cells on extracellular-matrix-coated porous filters in the absence or presence of TIMP-2, a specific collagenase inhibitor (Howard et al., 1991). Figure 1 shows that the addition of 40 nM of the inhibitor, which has no cytostatic effects on the cells, substan- tially reduced ( > 65%) in vitro invasion by these cells. Similar results were achieved with the MDA-TU-138 cell line. Addi- tion of a protein of similar molecular weight (myoglobin) did not impair the invasiveness of the UM-SCC-1 cells, suggesting that the effect of TIMP-2 was mediated via neutralization of the collagenase. While TIMP-2 inhibits other metalloprotein- ases, such as MMP-2 (Howard et al., 1991), it is unlikely that the reduction of in vitro invasion reflects an inhibition of other collagenases, since MMP-9 was the only collagenase detected by zymography (Fig. 2). These studies suggest that the invasive potential of at least a sub-population of malignant cells derived from SCCOC tumors requires the elaboration of MMP-9. Moreover, since generation of the active form of MMP-9 was contingent on the presence of plasmin(ogen), it is likely that both urokinase and the metalloproteinase act in concert with each other to promote the invasive phenotype of these tumor cells.
Detection of MMP-9 immunoreactivity in resected invasive SCCOC
To rule out the possibility that expression of MMP-9 by cultured SCCOC was a consequence of their in vitro cultiva- tion, we examined tumor sections from 4 patients for the presence of this collagenase (Fig. 4a,c). Strong immunoreactiv- ity was localized to the invading front of tumor cells (Fig. 4u), with little evidence of staining of non-malignant cells, includ- ing stromal elements (Fig. 4c). The specificity of the immuno- histochemical staining was confirmed in a serial section stained with the anti-serum that had been pre-adsorbed with the immunizing peptide antigen (Fig. 4h). In these conditions, the staining of the tumor cells was entirely abolished. These data
FIGURE 6 - Stimulation of protein kinase C with phorbol ester increases the amounts of MMP-9 protein. Conditioned medium or SF medium was subjected to SDS-PAGE using a 7.5% gel. Resolved proteins were transferred electrophoretically to a nitro- cellulose membrane. The filter was blocked with a 3.0% (w/v) BSA solution and subsequently incubated overnight at 4°C with a 1500 dilution of anti-serum raised against the 15 amino acid peptide present in MMP-9 or anti-serum that had been pre- adsorbed for 1 hr at room temperature with 25 kg/ml of the peptide. Immunoreactive bands were visualized with radioactive protein A. Molecular-weight standards are indicated on the left. The data are typical of 3 separate experiments.
are consistent with the notion that the elaboration of MMP-9 by cultured SCCOC cells reflects expression of this collagenase by the tumor cells in vivo rather than being an artifact of in vitro tissue-culture conditions. Moreover, they suggest that tumor invasion, by at least a sub-population of SCCOC cells, requires the elaboration of MMP-9.

MMP-9 EXPRESSION IN SQUAMOUS-CELL CARCINOMA 15
FIGURE 7 - Stimulation of protein kinase C with phorbol ester increases the steady-state level of mRNA encoding the metalloprotein- ase. UM-SCC-1 cells were grown to near-confluence and treated either with a range of PMA concentrations for 24 hr (a) or with 20 nM phorbol ester for varying times (b). The RNA was extracted, resolved in a 1.5% (w/v) agarose-formaldehyde gel and transferred to a nylon filter. The filter was probed at 42°C for 18 hr with a radiolabelled 0.5-kb cDNA specific for the MMP-9 transcript. Stringency washes were conducted at 65°C using 0.5 x SSC/O.75% (wiv) SDS. Equalities in loading were checked with GAPDH. The experiments were carried out on 2 separate occasions.
Induction of MMP-9 by treatment of cultured SCCOC with phorbol ester
Since the MMP-9 promoter sequence contains the phorbol- ester response elements AP-1, AP-2-like, and NFKB-like motifs in the 5' flanking region (Huhtala et al., 1991; Sen and Baltimore, 1986; Lee et al., 1987), we speculated that MMP-9 expression in SCCOC cells might be regulated by protein- kinase-C pathways. As a first step to testing this hypothesis, we treated UM-SCC-1 cells with varying concentrations of the phorbol ester, PMA, and analyzed the conditioned medium for MMP-9, either by zyrnography or by Western blotting (Figs. 5 and 6). Both assays indicated an increase in the amount of MMP-9 brought about by phorbol ester. Laser densitometry of the Western blot indicated a 30-fold induction of MMP-9 by the phorbol ester. PMA also induced the amount of MMP-9 secreted by MDA-TU-138 cells as determined by zymography (data not shown).
To determine whether the increase in the amount of MMP-9 protein was a result of a more abundant transcript. UM-SCC-1 cells were treated with varying concentrations of phorbol ester for 24 hr, or 20 nM of the agent for varying times, and the R N A was subjected to Northern blotting using a cDNA specific for the MMP-9 message (Fig. 7a,b). Treatment of the UM-SCC-1 cells with phorbol ester led to a dose- and time-dependent increase in the amount of MMP-9 transcript. A minimum of 4 hr was required for the induction of the metalloproteinase mRNA with an 18-hr treatment resulting in a 20- to 30-fold increase (as determined by laser densitometry) in the amount of mRNA (Fig. 7b). The magnitude of this change in MMP-9 mRNA was unaltered by growth of the cells on Matrigel. Thus, the 30-fold increase in the amount of MMP-9 protein, ob- served in Western blotting (Fig. 6), can be entirely accounted for by a parallel increase in level of mRNA encoding the metalloproteinase. Whether this increase in the level of steady-state mRNA reflects transcriptional activation or a more stable message is unclear a t present and will be the subject of future investigations.
Experiments were carried out to determine whether the induction in steady-state level of mRNA encoding the metallo- proteinase by PMA required de novo protein synthesis. To address this, UM-SCC-1 cells were treated with the protein- synthesis inhibitor, cycloheximide, the phorbol ester, PMA, or a combination of both of these agents. R N A was extracted under each condition and blotted for MMP-9 transcript. It is
FIGURE 8-Treatment of UM-SCC-1 cells with a protein- synthesis inhibitor abolishes the MMP-9-inductive effect of phor- bol ester. UM-SCC-1 cells were incubated at 90% confluence either in the presence or in the absence of cycloheximide (20 Fgiml) for 1 hr. After this time, PMA was added (final concentra- tion 20 nM) to the cells cultured with or without the protein- synthesis inhibitor. RNA was extracted 6 hr later and subjected to Northern blotting for MMP-9 transcript as described for Figure 7. Equalities in loading were checked by re-probing the blot with GAPDH. The data are typical of 2 separate experiments.
apparent from Figure 8 that treatment of the cells with cycloheximide alone caused an increase in the steady-state level of MMP-9 message similar to that brought about by the phorbol ester. However, when UM-SCC-1 cells were treated with PMA, in the presence of the protein-synthesis inhibitor, the inductive effect of the phorbol ester was entirely abolished. These data could be explained in 2 ways. First, the inductive effect of PMA on MMP-9 mRNA levels could require de novo protein synthesis with the cycloheximide-induced elevation in transcript reflecting the reduction of a labile repressor of the gene. Alternatively, the treatment of UM-SCC-1 cells either
16 JUAREZ ETAL.
with phorbol ester or with the protein-synthesis inhibitor may result in decreased production of an RNAase responsible for the turnover of the MMP-9 transcript, culminating in an increase in the steady-state levels of this message. Clearly, further studies will be required to discriminate between these 2 possibilities. Treatment of UM-SCC-1 cells with the protein-kinase-C inhibitor, calphostin-C, results in a reduction in the constitutive expression of MMP-9
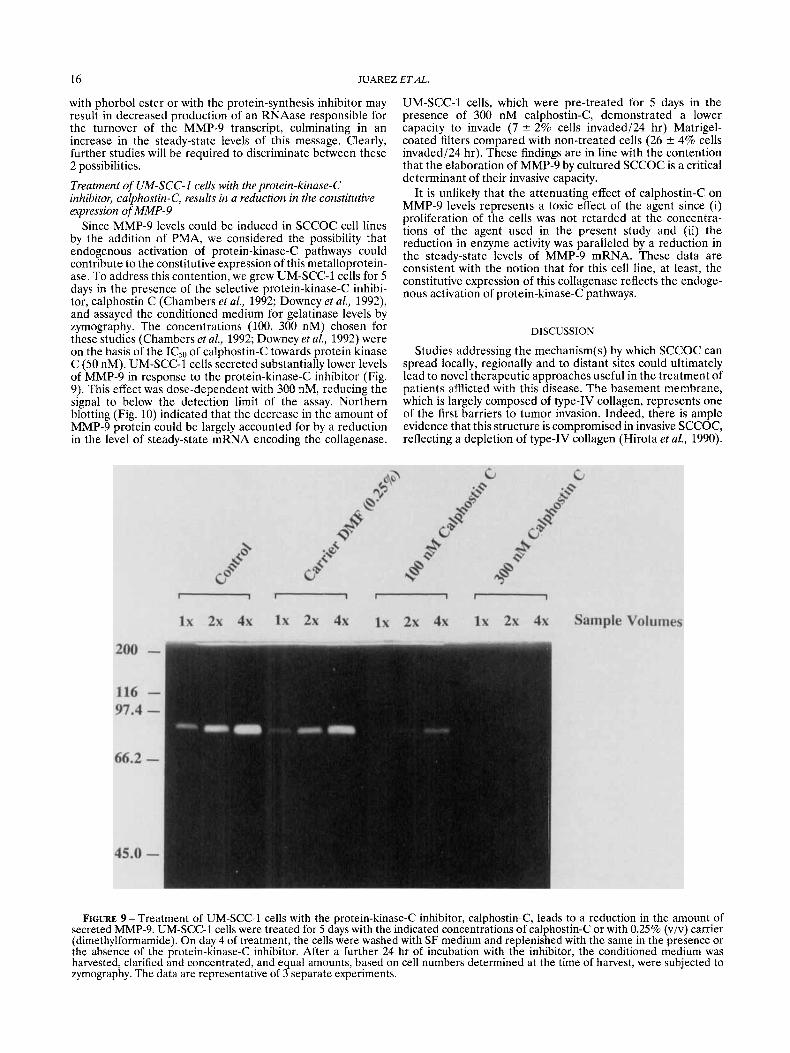
Since MMP-9 levels could be induced in SCCOC cell lines by the addition of PMA, we considered the possibility that endogenous activation of protein-kinase-C pathways could contribute to the constitutive expression of this metalloprotein- ase. To address this contention, we grew UM-SCC-1 cells for 5 days in the presence of the selective protein-kinase-C inhibi- tor, calphostin C (Chambers et al., 1992; Downey et al., 1992), and assayed the conditioned medium for gelatinase levels by zymography. The concentrations (100, 300 nM) chosen for these studies (Chambers et al., 1992; Downey et al., 1992) were on the basis of the IC50 of calphostin-C towards protein kinase C (50 nM). UM-SCC-1 cells secreted substantially lower levels of MMP-9 in response to the protein-kinase-C inhibitor (Fig. 9). This effect was dose-dependent with 300 nM, reducing the signal to below the detection limit of the assay. Northern blotting (Fig. 10) indicated that the decrease in the amount of MMP-9 protein could be largely accounted for by a reduction in the level of steady-state mRNA encoding the collagenase.
UM-SCC-1 cells, which were pre-treated for 5 days in the presence of 300 nM calphostin-C, demonstrated a lower capacity to invade (7 2 2% cells invaded/24 hr) Matrigel- coated filters compared with non-treated cells (26 rfr 4% cells invaded124 hr). These findings are in line with the contention that the elaboration of MMP-9 by cultured SCCOC is a critical determinant of their invasive capacity.
It is unlikely that the attenuating effect of calphostin-C on MMP-9 levels represents a toxic effect of the agent since (i) proliferation of the cells was not retarded at the concentra- tions of the agent used in the present study and (ii) the reduction in enzyme activity was paralleled by a reduction in the steady-state levels of MMP-9 mRNA. These data are consistent with the notion that for this cell line, at least, the constitutive expression of this collagenase reflects the endoge- nous activation of protein-kinase-C pathways.
DISCUSSION
Studies addressing the rnechanism(s) by which SCCOC can spread locally, regionally and to distant sites could ultimately lead to novel therapeutic approaches useful in the treatment of patients afflicted with this disease. The basement membrane, which is largely composed of type-IV collagen, represents one of the first barriers to tumor invasion. Indeed, there is ample evidence that this structure is compromised in invasive SCCOC, reflecting a depletion of type-IV collagen (Hirota et aL, 1990).
FIGURE 9 -Treatment of UM-SCC-1 cells with the protein-kinase-C inhibitor, calphostin-C, leads to a reduction in the amount of secreted MMP-9. UM-SCC-1 cells were treated for 5 days with the indicated concentrations of calphostin-C or with 0.25% (viv) carrier (dimethylformamide). On day 4 of treatment, the cells were washed with SF medium and replenished with the same in the presence or the absence of the protein-kinase-C inhibitor. After a further 24 hr of incubation with the inhibitor, the conditioned medium was harvested, clarified and concentrated, and equal amounts, based on cell numbers determined at the time of harvest, were subjected to zymography. The data are representative of 3 separate experiments.

ON IN SQUAMOUS-CELL CARCINOMA 17
population of tumor cells derived from SCCOC, an understand- ing of the mechanism(s) by which its expression is regulated could ultimately lead to novel anti-invasive agents that act by reducing the levels of this metalloproteinase. The published sequence of the MMP-9 promoter (Huhtala et al., 1991)
FIGURE 10-Treatment of UM-SCC-1 cells with the protein- kinase-C inhibitor, calphostin-C, leads to a reduction in the amount of steady-state mRNA encoding MMP-9. The cells were treated as described for Figure 9. RNA was extracted and blotted for MMP-9 mRNA as described for Figure 7. The experiment was carried out on 3 separate occasions.
Consequently, we undertook a study to determine the role of the type-IV collagen-degrading MMP-9 in the invasive pheno- type demonstrated by invasive SCCOC. Several findings in our study indicated a critical role for MMP-9 in the invasive phenotype demonstrated by at least a sub-population of SCCOC tumor cells. Thus, (i) tumor cells in invasive SCCOC stained positive for the metalloproteinase, (ii) 2 independent SCCOC cell lines that were highly invasive in vitro expressed high levels of the collagenase, and (iii) the ability of cultured SCCOC, which elaborate MMP-9, to penetrate an extracellular- matrix-coated filter could be largely inhibited by TIMP-2, a specific collagenase inhibitor. Since only 2 SCCOC cell lines were employed in the present study, caution should be exercised in assigning a role for this metalloproteinase in the invasive phenotype demonstrated by the different types of cancer cells seen in any one SCCOC malignancy.
Indeed, studies by Muller et al. (1991) and Kusukawa et al. (1992) suggested that MMP-2 and MMP-3 was required for invasion by SCCOC. Interestingly, we could find no evidence for a contribution of these collagenases in the invasive pheno- type demonstrated by either UM-SCC-1 or MDA-TU-138 cells. Thus, we were unable to detect activities at 72 or 57 kDa in zymograms using conditioned medium derived from either of the invasive SCCOC cell lines, UM-SCC-1 or MDA-TU- 138. This did not reflect technical limitations, since we were successful in detecting MMP-2 (or an activity which was indistinguishable from it in terms of molecular weight) in conditioned medium derived from fibroblasts. These divergent results between laboratories could very well reflect the heterog- enous nature of SCCOC, insofar as the expression of these coIlagenases may be restricted to a sub-population of cells present in the malignancy.
If in fact the expression of MMP-9 is a key factor in determining the invasive phenotype of at least a sub-
includes identical or relatedmotifs for the transcription factors AP-1, AP-2 and NF-KB, all of which have been identified as effectors of the protein-kinase-C signal-transduction pathway (Sen and Baltimore, 1986; Lee et af., 1987). With this in mind, we determined the role of protein-kinase-C pathways in the regulation of MMP-9 expression. Stimulation of this pathway with phorbol ester led to a dramatic induction in the amount of the metalloproteinase, this being a consequence of a more abundant MMP-9 transcript. The observation of an induction of MMP-9 mRNA/protein levels by phorbol ester led us to speculate that the constitutive expression of this collagenase in SCCOC could partly reflect endogenous activation of the protein-kinase-C signal-transduction pathway. Consistent with this notion was our finding of MMP-9 down-regulation upon treatment with the protein-kinase-C inhibitor calphostin-C. These data suggest that for UM-SCC-1 cells, activation of the protein-kinase-C signal-transduction pathway is responsible, at least in part, for the constitutive expression of this metallo- proteinase.
Endogenous activation of protein-kinase-C signal-transduc- tion pathways is elicited by several growth factors, including TNFa and TGF-a, subsequent to their binding to their receptor at the cell surface (Presta et al., 1991; Schutze et af., 1992; Kudlow and Bjorge, 1990). Interestingly, other investiga- tors have shown induction of type-IV collagenase by these same growth factors (Unemori et af., 1991; Turksen et al., 1991). Thus, there is a possibility that the constitutive produc- tion of MMP-9 in UM-SCC-1 cells might reflect the autocrine stimulation of these cells by these or other cytokines which act through the protein-kinase-C signal-transduction pathway.
In contrast to UM-SCC-1 cells, we were unable to down- regulate the expression of the collagenase in MDA-TU-138 cells with the protein-kinase-C inhibitor. Thus, for MDA-TU- 138 cells, it is unlikely that autocrine mechanisms, which activate PKC pathways, are responsible for the elevated expression of this metalloproteinase. These findings could be explained in 2 ways. Firstly, in MDA-TU-138 cells, the signal- transduction pathway may be activated down-stream of the protein-kinase-C enzyme. Such a lesion would be resistant to the action of calphostin-C, which is confined to a blockade of the protein-kinase-C enzyme. Alternatively, the role of protein- kinase-C pathways in the regulation of MMP-9 expression may be restricted to a sub-population of SCCOC malignant cells from which the UM-SCC-1 cell line was derived.
In conclusion, our study provides evidence for a critical role for 92-kDa type-IV collagenase (MMP-9) in the invasive phenotype demonstrated by at least a sub-population of tumor cells derived from SCCOC and, in addition, argues for a role of protein-kinase-C pathways in the constitutive expression of this metalloproteinase.
ACKNOWLEDGEMENTS
This work was supported by NIH grants CA 51539 and RR 5511 and a grant (3438) from the Council for Tobacco Research to D.B and CA 44352 and the National Foundation for Cancer Research to G.L.N. We thank Mrs. N. Steward for excellent technical help.
REFERENCES
BERNHARD, E.J., MUSCHEL, R.J. and HUGHES, E.N., Mr-92,000 gelati- nase release correlates with the metastatic phenotype in transformed rat embryo cells. Cancer Res., 50,3872-3877 (1990). BOYD, D., ZIOBER, B., CHAKRABARTY, S. and BRATTAIN, M.G.,
Examination of urokinase proteinitranscript levels and their relation- ship with laminin degradation in cultured colon carcinoma. Cancer Res., 49,816-820 (1989). CHAMBERS, T.C., ZHENG, B. and Kuo, J.F., Regulation by phorbol

18 JUAREZ ETAL.
ester and protein-kinase-C inhibitors and by a protein-phosphatase inhibitor (okadaic acid) of P-glycoprotein phosphorylation and relation- ship to drug accumulation in multidrug-resistant human Kl3 cells. Mol. Pharmacol., 41,1008-1015 (1992). CLAYMAN, G., WANG, S.W., NICOLSON, G.L., EL-NAGGAR, A., MAZAR, A,, HENKIN, J., BLASI, F., GOEPFERT, H. and BOYD, D., Regulation of urokinase-type plasminogen activator expression in squamous-cell carcinoma of the oral cavity. Int. J. Cancer, 54,73-80 (1993). COLLEN, D. and VERSTRAETE, M., Molecular biology of human plasminogen. 11. Metabolism in physiological and some, pathological conditions in man. Thrombosis Diathesis Haemovhagica, 34, 403 (1975). DAVIS, G.E. and MARTIN, B.M., A latent Mr-94,000 gelatin-degrading metalloprotease induced during differentiation of HL-60 promyelo- cytic leukemia cells: a member of the collagenase family of enzymes. CancerRes., 50,1113-1120 (1990). DOWNEY, G.P., CHAN, C.K., LEA, P., TAKAI, A. and GRINSTEIN, S., Phorbol-ester-induced actin assembly in neutrophils: role of protein kinase C. J. Cell Biol., 116,695-706 (1992). GARBISA, S., POZZATTI, R., MUSHCEL, R.J., SAFFIOTTI, U., BALLIN, M., GOLDFARB, R., KHOURY, G. and LIOTTA, L., Secretion of type-IV collagenase protease and metastatic phenotype: induction by transfec- tion with c-Ha-ras but not c-Ha-ras plus Ad2-Ela. Cancer Res., 47, 1523-1528 (1987). HIROTA, J., YONEDA, K. and OSAKI, T., Basement-membrane type-IV collagen in oral squamous-cell carcinoma. Head and Neck, 12,400-405 (1990). HOLLAS, W., BLASI, F. and BOYD, D., Role of the urokinase receptor in facilitating extracellular-matrix invasion by cultured colon cancer. CancerRes., 51,3690-3695 (1991). HOWARD, E.W., BULLEN, E. and BANDA, M.J., Preferential inhibition of 72- and 92-kDa gelatinases by tissue inhibitor of metalloprotein- ases-2.J. biol. Chem., 266,13070-13075 (1991). HUHTALA, P., TUUITILA, A., CHOW, L., LOHI, J., KESKI-OJA, J. and TRYGGVASON, K., Complete structure of the human gene for 92-kDa type-IV collagenase. J. biol. Chem., 266,16485-16490 (1991). KHOKHA, R., WATERHOUSE, P., YAGEL, S., LALA, P., OVERALL, C., NORTON. G. and DENHARDT. D.. Anti-sense RNA-induced reduction in murine TIMP levels confers oncogenicity on Swiss 3T3 cells. Science, 243,947-950 (1989). KUDLOW, J.E. and BJORGE, J.D., TGF-a in normal physiology. Sem. Cancer Biol., 1,293-302 (1990). KUSUKAWA, J., SASAGURI, Y., SHIMA, I., KAMEYAMA, T. and MORI- MATSU, M., Production of matrix metalloproteinase 2 (gelatinasei type-IV collagenase) and 3 (stromelysin) by cultured oral squamous- cell carcinoma. J. oral Pathol. Med., 21,221-224 (1992). LEE, W., MITCHELL, P. and TJIAN, R., Purified transcription factor
AP-1 interacts with TPA-inducible enhancer elements. Cell, 49, 741-752 (1987). MOSSMAN, T., Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytoxicity assays. J . immunol. Methods, 65,55-63 (1983). MULLER, D., BREATHNACH, R., ENGELMANN, A,, MILLON, R., BRON- NER, G., FLESCH, H., DUMONT, P., EBER, M. and ABECASSIS, J., Expression of collagenase-related metalloproteinase genes in human lung or head and neck tumours. Int. J. Cancer, 48,550-556 (1991). NAKAJIMA, M., IRMURA, T. and NICOLSON, G.L., Basement-membrane- degradative enzymes and tumor metastasis. Cancer Bulletin, 39, 142- 149 (1987). PRESTA, M., TIBERIO, L., RUSNATI, M., DELL'ERA, P. and RAGNOTTI, G., Basic fibroblast growth factor requires a long-lasting activation of protein kinase C to induce cell proliferation in transformed fetal bovine aortic endothelial cells. Cell Regulation, 2,719-726 (1991). SALO, T., LIOTTA, L., KESKI-OJA, J., TURPEENNIEMI-HUJANEN, T. and TRYGGVASON, K., Secretion of basement-collagen-degrading enzyme and plasminogen activator by transformed cells role in metastasis. Int. J. Cancer, 30,669-673 (1982). SCHLECHTE, W., BRATTAIN, M. and BOYD, D., Invasion of extracellular matrix b cultured colon cancer: dependence on receptor display. Cancer d m m u n . , 2,173-179 (1990). SCHULTZ, R.M., SILBERMAN, S., PERSKY, B., BAJKOSKI, AS . and CARMICHAEL, D.F., Inhibition by human-recombinant-tissue inhibitor of metalloproteinases of human amnion invasion and lung coloniza- tion by murine B16-FlO melanoma cells. Cancer Res., 48, 5539-5545 (1988). SCHUTZ, S., MACHLEIDT, T. and KRONKE, M., Mechanisms of tumor- necrosis-factor action. Sem. Oncol., 19, 16-24 (1992). SEN, R. and BALTIMORE, D., Inducibility of k-immunoglobin-enhancer- binding protein NF-kb by a post-translational mechanism. CeH, 47, 921-928 (1986). TRYGGVASON, K., HOYHTYA, M. and SALO, T., Proteolytic degradation of extracellular matrix in tumor invasion. Biochim. biophys. Acta, 907,
TURKSEN, K., CHOI, Y. and FUCHS, E., Transforming growth factor alpha induces collagen degradation and cell migration in differentiat- ing human epidermal raft cultures. Cell Regulation, 2,613-625 (1991). UNEMORI, E.N., HIBBS, M.S. and AMENTO, E.P., Constitutive expres- sion of a 92-kDa gelatinase (type-IV collagenase) by rheumatoid synovial fibroblasts and its induction in normal human fibroblasts by inflammatory cytokines. J. din. Invest., 88, 1656-1662 (1991). WILHELM, S.M., COLLIER, I.E., MARMER, B.L., EISEN, A.Z., GRANT, G. and GOLDBERG, G., SV40-transformed human lung fibroblasts secrete a 92-kDa type-IV collagenase which is identical to that secreted by normal human macrophages. J. biol. Chem, 264,17213-17221 (1989).
191-217 (1987).
Copyright © 2022 FDOKUMEN




















