
Identification of a cytoskeleton-associated 120 kDa RNA-binding protein in developing rice seeds
Upload
independentCategory
view
0download
0
Bacillus thuringiensis Protein Cry6B (BGSC ID 4D8) is Toxicto Larvae of Hypera postica
Anil Sharma • Suresh Kumar • Raj K. Bhatnagar
Received: 24 February 2010 / Accepted: 10 August 2010
� Springer Science+Business Media, LLC 2010
Abstract Insecticidal proteins produced by strains of
Bacillus thuringenesis are specific toward target pests. One
of the Bt proteins, Cry 1Ac has been used successfully for
controlling crop predation by polyphagous pests Heli-
coverpa armigera. Structurally, Bt proteins consist of three
domains; domain I and III are fairly homologous in various
Bt proteins while domain II is hypervariable. The hyper-
variable domain II is believed to be responsible for speci-
ficity toward target pest. Successful deployment of Bt
proteins requires knowledge of its specificity toward the
insect. Various Bt proteins have been characterized for
activity against coleopteran pests. Some Bt proteins of
class Cry6 have been found to be active against potato
weevil. We have evaluated the activity of Cry6B protein
(BGSC-4D8) against lucerne weevil, Hypera postica,
which is a major pest of forage crop Medicago sativa.
Results revealed that the purified Cry6B protein is signif-
icantly active against the coleopteran pest with LC50 value
280 ng/ll. The leaves coated with the purified Cry6 toxin
were three times less damaged as compared with the neg-
ative control.
Introduction
Medicago sativa L. (lucerne or alfalfa) is one of the most
important forage legumes used as cattle feed [14]. Alfalfa
is well known for its nutritionally rich biomass production.
The record average yield of alfalfa is 10 tons/acre without
irrigation, while 24 tons/acre with irrigation (United States
Department of Agriculture, 1998). It is highly palatable and
nutritious to the cattle as it is enriched with protein, cal-
cium, phosphorus, and vitamins (A, B, and D). It is the
third important forage crop in India following Sorghum
(Sorghum bicolor) and Berseem (Trifolium alexandrinum).
One of the rampant problems in lucerne cultivation is the
damage caused by lucerne weevil (Hypera postica Gyll.,
Coleoptera), particularly in the Northern India. Larva of
weevil ravages phyto-phagously, reducing productivity of
the crop. Larvae cause skeletonization of leaves, stunted
plant growth and up to 20–30% reduction in green fodder
yield [10]. Severe infestation may even ruin entire stand of
the crop. No usable source of the insect resistance in
M. sativa gene pool is available so far. Screening of the
germplasm of Medicago led to identification of a few
Medicago species showing resistance to lucerne weevil [4].
Inter-specific incompatibility among the Medicago species
is the major limitation for conventional breeding methods.
In the absence of conventional breeding, genetic transfor-
mation of lucerne plants with Bt gene is a promising
approach to control the damage caused by lucerne weevil.
Bacillus thuringiensis Serovar. Kurstaki is a Gram-
positive bacterium, which produces proteinaceous inclu-
sions during sporulation. The inclusions are composed of
proteins known as ICPs, Cry proteins, or d-endotoxins,
which are encoded by cry genes. Cry proteins are highly
toxic to a wide variety of important agricultural and health
related insect pests as well as other invertebrates. Due to
A. Sharma � R. K. Bhatnagar (&)
Insect Resistance Group, International Center for Genetic
Engineering and Biotechnology, Aruna Asaf Ali Marg,
New Delhi 110067, India
e-mail: [email protected]
S. Kumar
Division of Crop Improvement, Indian Grassland and Fodder
Research Institute, Jhansi 284003, India
123
Curr Microbiol
DOI 10.1007/s00284-010-9749-4

their high specificity and their safety for the environment,
ICPs are valuable alternative to chemical pesticides for
control of insect pests in agriculture and forestry, and in the
home. It is believed that the rational application of
B. thuringiensis toxins will provide a variety of alternatives
for insect control and for coping with the problem of insect
resistance to pesticides.
Cry toxins have specific activities against species of the
orders Lepidoptera (Moth and butterflies), Diptera (Flies
and Mosquitoes) and Coleoptera (Beetles and Weevils).
Several Bt proteins (Cry6A, Cry3, and that from many
other strains of Bt) have been identified to be effective
against Coleopteran insects, in particular against lucerne
weevil [9]. Here, we report expression, purification, and
toxicity of codon-optimized Cry6B insecticidal protein.
The cry6b gene with codon bias for expression in Medi-
cago sativa was expressed in E. coli and screened for
activity against lucerne weevil, Hypera postica. The puri-
fied protein was active against the larvae of the weevil at
LC50 280 ng/ll.
Materials and Methods
Codon Optimization of cry6b According to M. sativa
Sequence of a delta-endotoxin cry6b gene (GenBank:
L07024.1) from Bacillus thuringiensis was optimized
according to codon usage of Medicago sativa and the gene
was synthesized commercially from GENEART (Regens-
berg) with BamH1 and Sac1 restriction endonuclease sites
attached at the 50 and 30 end of open reading frame. Codon
selection and % frequency of occurrence of a codon are
given in Table 1.
Cloning and Expression of the cry6b gene in Bacteria
The cry6b gene was cloned into pQE30 expression vector
at BamH1 and Sac1 sites down stream the T5 promoter and
was transformed into competent E. coli M15 cells by heat-
shock method. Expression of the Cry6B protein was
induced by 1 mM IPTG. The expression of the recombi-
nant Cry6B was checked by SDS-PAGE and western blot
analysis using anti-His antibodies. The Cry6B was
expressed in both soluble as well as in aggregated forms.
The recombinant Cry6B protein containing 6-histidine
residue-tag was purified from E. coli M15 cells using
affinity chromatography on Ni-NTA column (Qiagen). For
the purpose, the supernatant was passed through the Ni-
NTA resin at pH 8.0 to allow the ‘‘His’’ tag of the protein
to bind to the resin. The resin was washed with the wash
buffer (50 mM tris, 300 mM NaCl and 30 mM Immidaz-
ole) to elute out non-specific proteins. ‘‘His’’ containing
recombinant Cry6B was eluted with elution buffer con-
taining 50 mM tris buffer pH 8.0, 300 mM NaCl and
300 mM Immidazole. Different eluted fractions were
checked on SDS-PAGE gels for the purified Cry6B.
Cry6B-positive eluted fractions were pooled, dialyzed
overnight against 10 mM Tris–Cl (pH 8.0) and concen-
trated using 10 kDa centricon membrane. The quantity of
the concentrated protein was estimated using Bradford’s
reagent [2].
Table 1 Codon optimization of gene encoding Bacillus thuringiensis insecticidal protein cry6b as per codon usage of Medicago sativa
Codon Amino acid Freq. Codon Amino acid Freq. Codon Amino acid Freq. Codon Amino cid Freq.
UUU F 7 UCU S 10 UAU Y 11 UGU C 3
UUC F 5 UCC S 5 UAC Y 9 UGC C 2
UUA L 5 UCA S 9 UAA * UGA *
UUG L 11 UCG S 2 UAG * UGG W 8
CUU L 12 CCU P 5 CAU H 5 CGU R 2
CUC L 6 CCC P 2 CAC H 4 CGC R 1
CUA L 4 CCA P 4 CAA Q 20 CGA R 1
CUG L 4 CCG P 1 CAG Q 11 CGG R 1
AUU I 21 ACU T 10 AAU N 25 AGU S 7
AUC I 11 ACC T 6 AAC N 19 AGC S 5
AUA I 9 ACA T 9 AAA K 18 AGA R 3
AUG M 8 ACG T 2 AAG K 19 AGG R 2
GUU V 9 GCU A 12 GAU D 26 GGU G 7
GUC V 3 GCC A 4 GAC D 10 GGC G 3
GUA V 3 GCA A 10 GAA E 17 GGA G 7
GUG V 4 GCG A 2 GAG E 12 GGG G 2
GC content: 43.7%
A. Sharma et al.: Cry6B is Toxic to Hypera postica
123

Raising of Polyclonal Antisera
For raising polyclonal antisera against the purified cry6B
toxin, 1 mg/ml of protein was mixed with equal volume of
complete Freund’s adjuvant, emulsified and injected 100 ll
each intra-dermally at 10 different places of a New Zealand
rabbit [11]. The rabbit was boosted with Cry6B plus
incomplete Fruend’s adjuvant after 21 days of injection.
Serum was collected after 12 days of first booster. Anti-
Cry6B antibody titer in rabbit blood was measured by
western blotting against purified Cry6B protein using serial
dilutions of the antibodies ranging from 1:1,000 to
1:20,000.
Bioassay Against Weevil on Leaves Coated with ICPs
Purified Cry6B insecticidal crystal protein (ICP) was used
for insect bioassay, along with controls using neonate lar-
vae of the lucerne weevil (during January–March, 2009) to
assess the bio-efficacy of the insecticidal protein. For the
purpose, the Cry6B protein was expressed in E. coli and the
protein was isolated from the bacterial culture using protein
extraction and purification kit (Medox-Bio) followed by
washing with crystal wash buffers. The wet paste of the
250 ng/ll of Cry6B protein with 0.6% of gelatin was
coated on both the sides of trifoliate leaves of lucerne. The
average area of a normal M. sativa leaf is nearly
12.37 sq cm. The leaf was placed on a moist Whatman
filter paper and five neonate larvae were released on each
trifoliate leaf coated with the protein in a 90 mm petri
plate. The petriplate was covered with its lid to restrict run-
away of the larvae. Weight of the leaf material was taken
before and after 2 days of setting up the bioassays to
estimate the loss incurred to the leaves. Similarly, the
insects were also weighed before and after 2 days of the
application of the ICP on leaf to estimate the growth rate of
the larvae. Mortality of the larvae was recorded after
5 days of incubation. The leaf damage, larval growth, and
mortality were compared with corresponding Cry1Ac and
uncoated leaves as negative control.
For calculation of lethal concentrations (LC50) values, a
range of purified Cry6B toxin from 100 to 1,000 ng was
prepared and was coated on trifoliate leaf of M. sativa. Five
larvae were released on each leaf as described above.
Mortality was recorded on fifth day of the release of larvae.
Data of all the independent experiments were pooled to
calculate the LC50 value using Probit analysis. Each bio-
assay experiment was independently conducted thrice in
three replications. Further to plot the survival ship curves,
the mortality of the larvae were daily recorded till ninth
day of start of experiment till all the Cry6B treated insects
were dead and control larvae started pupating.
Statistical Analysis
The data obtained from the bioassay were found to be nor-
mally distributed. The normally distributed data were ana-
lyzed using one-way analysis of variance (ANOVA) and
HSD Tukey test to check the significance of the differences
in the mean values of mortality, growth rate, and leaf damage
using VassarStats: Statistical Computation Website—
http://faculty.vassar.edu/lowry/VassarStats.html. The sur-
vivalship curves were plotted using Microsoft Excel and the
significance in the difference is calculated using Log Rank
Test on http://bioinf.wehi.edu.au/software/russell/logrank/
index.html site. LC50 and LC90 values were manually cal-
culated using Log Probit analysis.
Results
Codon Optimization of cry6b gene According
to M. sativa
The codon of 1.5 kb cry6b gene was optimized according
to the codon usage table of M. sativa. For the choice of
codon used in M. sativa, care was taken to distribute pre-
dominately used codons evenly. The overall codon usage
and their relative abundance are described in Table 1. The
synthesized gene corresponds to 52 kDa insecticidal pro-
tein. The sequence of the modified cry6b gene is given in
Fig. 1.
Cloning and Expression of the cry6b gene in E. coli
and Evaluation of Activity Against Lucerne Weevil
The codon-optimized cry6b was cloned into expression
vector, pQE30, and transformed into E.coli M15 cells. The
transformed cells were induced with IPTG to express the
protein. The recombinant Cry6B protein was accumulated
in soluble and insoluble fractions of the cells (Fig. 2a,b).
Since recombinant Cry6B was expressed with a 6xHis tag,
the latter was used to purify the protein using affinity
chromatography on Ni-NTA columns (Fig. 2c). The puri-
fied protein was dialyzed and concentrated. Polyclonal
antiserum was raised against the purified protein in New
Zealand rabbit.
The purified protein was employed for toxicity against
lucerne weevil. Different concentrations of purified Cry6B,
ranging from 100 to 1,000 ng/ll were coated on M. sativa
leaves and five neonate larvae were released on each tri-
foliate leaf. The bioassay results indicated significant dif-
ference in leaf damage and insect mortality (Tables 2, 3,
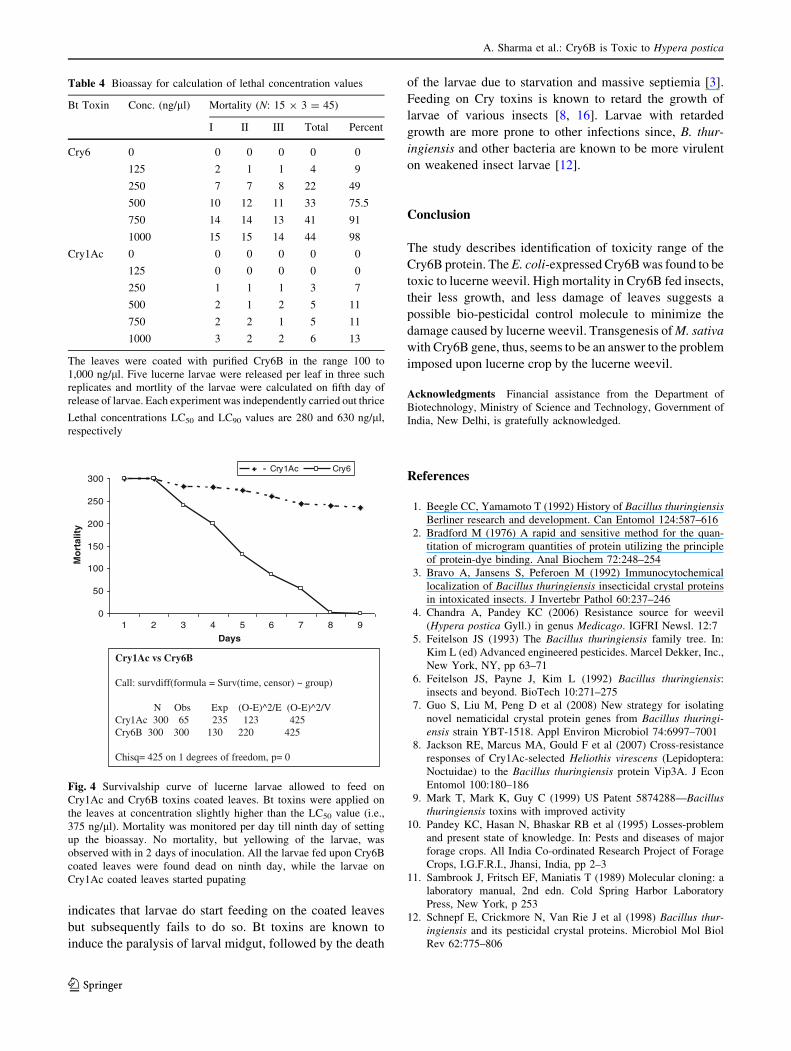
Figs. 3, 4) caused by the insect. The LC50 and LC90 values
of the Cry6B were calculated to be *280 and 630 ng/ll,
respectively (Table 4). Significantly less insect mortality
A. Sharma et al.: Cry6B is Toxic to Hypera postica
123

Fig
.1
Nu
cleo
tid
ese
qu
ence
of
mo
difi
edcr
y6b
gen
e.T
he
nu
cleo
tid
ese
qu
ence
of
cry
6b
gen
eo
fB
aci
llu
sth
uri
ng
ien
sis
was
cod
on
op
tim
ized
acco
rdin
gto
cod
on
usa
ge
of
Med
ica
go
sati
vaan
d
Bam
H1
and
Sac
1si
tes
(sh
ad
ed)
are
app
end
edb
efo
rean
daf
ter
the
seq
uen
ce
A. Sharma et al.: Cry6B is Toxic to Hypera postica
123

and consequently high foliage damage was observed in
leaves coated with Bt protein, Cry1Ac, or buffer control.
Based on visual observations, it was estimated that
uncoated leaves and Cry1Ac coated leaves were four to
five times more damaged than those coated with Cry6B
protein (ANOVA; P [ 0001). Similarly, significant dif-
ference between treatment and control group was observed
in growth rate and of larval mortality (Table 3). The
growth of the larvae that were forced to feed on leaves
coated with Cry6B toxin was one-third as compared to the
larvae that were fed upon Cry1Ac coated or uncoated
leaves (ANOVA; P [ 001). On fifth day of experiment,
nearly 5-folds more larvae were dead on leaves that were
pre-coated with Cry6B as compared with the larvae on
leaves coated with Cry1Ac. Whereas, the mortality on cry6B
coated leaves was[9-fold more as compared with the larvae
on uncoated leaves (ANOVA; P [ 0.0001). The survival
ship curves show significant difference in the survival of the
weevil larvae upon treatment with Cry6B protein as com-
pared with the control Cry1Ac treatment (Fig. 4).
Discussion
Leading bio-rational pesticide, Bacillus thuringiensis, is a
ubiquitous Gram-positive, spore-forming bacterium that
forms a parasporal crystal during the stationary phase of its
growth cycle. B. thuringiensis was initially characterized as
an insect pathogen, and its insecticidal activity was
attributed to the parasporal crystals. Bt strains are spe-
cialized in killing certain target insects rather than
becoming generalist killers of all classes of insect. This
observation led to the development of bio-insecticides
based on B. thuringiensis for the control of certain insect
Vector pQE30 + Cry6alone Unind Ind
A
53kDa
Suptt PelletUnind Ind Uunind Ind
B
53kDa
Flow Wash ElutionThrough 1 2 1 2 4 6 8 10
C
53kDa
Fig. 2 Expression and
purification of Cry6B protein.
The Cry6B protein was cloned
and expressed using pQE30
vector and E. coli M15 host
cells. The protein can be seen as
53 kDa polypeptide band on
SDS-PAGE Gel (a) and on
western blot developed by anti-
His antibodies (b). The
insecticidal protein was purified
using Ni-NTA column
chromatography (c)
A. Sharma et al.: Cry6B is Toxic to Hypera postica
123

species among the orders Lepidoptera, Diptera, Coleoptera,
Hymenoptera, Homoptera, Orthoptera, and Mallophaga
and against nematodes, mites, and protozoa [1, 5, 6].
B. thuringiensis is already a useful alternative or supple-
ment to synthetic chemical pesticide application in com-
mercial agriculture, forest management, and mosquito
control. It is also a key source of genes for transgenic
expression to provide pest resistance in plants.
Bacillus thuringiensis Cry6 subfamily contains two
proteins, Cry6A and Cry6B. These are nearly 50% identi-
cal to each other, but they are unrelated by protein
sequence to the main family of Bt crystal toxins. Delta-
endotoxin proteins Cry6A and Cry6B were identified as
anti-nematocidal agent, of which anti-nematocidal activity
of Cry6B was not found to be reproducible [7, 15]. Both
Cry6 family toxins have been described in patent on
Table 2 Leaf damage caused by the lucerne weevil during insect bioassay using ICPs
Treatment Replication Weight of trifoliate leaf (mg) Damage (%)
Initial Wt. Wt. 2 days after
inoculation
No ICP I 2,938 2,262 676 (23.01) 717.66 ± 40.1
(23.66)II 3,004 2,283 721 (24.00)
III 3,153 2,397 756 (23.98)
Cry1Ac I 2,689 2,286 403 (14.98) 533 ± 115.3
(15.33)II 3,825 3,252 573 (14.98)
III 3,888 3,265 623 (16.02)
Cry6 I 2,701 2,545 156 (5.77) 134 ± 35.6
(4.27)II 3,100 3,007 93 (3.00)
III 3,810 3,657 153 (4.02)
ANOVA summary for loss of larval weight
Source Sum of squares Degree of freedom Mean of squares F value P value
Treatment (Between Groups) 533969.556 2 266984.778 49.53 0.00019
Error 32342 6 5390.444
Ss/Bl
Total 566312.222 8
HSD Tukey test
HSD [0.05]=184.14; HSD [0.01]=268.03
No ICP vs. Cry1Ac P \ 0.5
No ICP vs. Cry6 P \ 0.01
Cry1Ac vs. Cry6 P \ 0.01
Each leaf was coated with 250 ng/ll of Bt protein as indicated. Each replication contains data from 6 plates and the experiment was repeated
three times
Fig. 3 Effect of Cry6B protein
on leaf damage in Medicagosativa caused by lucerne weevil.
Five neonate larvae were
released on each trifoliate leaf
coated with 250 ng/ll of
insecticidal protein and the
damage was recorded after
2 days of release
A. Sharma et al.: Cry6B is Toxic to Hypera postica
123

nematocidal Bt toxins, but only sparse data were presented
[13]. Mark et al. [9] described activity of truncated Cry6
toxin against coleopterans. In the present study, full Cry6B
gene was recombinantly expressed in E.coli and was found
to be active against coleopteran lucerne weevil with nearly
5- and 9-fold mortality than that of Cry1Ac and negative
control, respectively. The Cry6B protein is found to have
LC50 and LC90 values *280 and 630 ng/ll, respectively.
These results clearly identified lucerne weevil to be sen-
sitive to Cry6B protein. Besides killing the pests, the
presence of Cry6B mitigates the leaf damage as evidenced
by nearly three times less damaged Cry6B coated leaves as
compared with the uncoated or Cry1Ac coated leaves.
Minor injury (feeding) on the Cry6B coated leaves
Table 3 Growth and mortality of larvae during insect bioassay using Bt protein
Treatment Replication Weight of 5 larvae (mg) Growth rate (%) Mortality (%) (N = 15 per replication)
Initial Wt. Wt. 2 days
after inoculation
No ICP I 62.0 80.5 18.5 (29.84) 21.8 ± 2.8 (32.83) 1 (06.60) 1.33 ± 0.58 (8.8)
II 67.8 91.5 23.7 (34.95) 2 (13.20)
III 68.5 91.6 23.1 (33.72) 1 (06.60)
Cry1Ac I 81.6 102.1 20.5 (25.12) 22.5 ± 3.6 (25.26) 4 (26.60) 2.33 ± 1.52 (15.46)
II 85.8 106.1 20.3 (23.66) 2 (13.20)
III 98.5 125.1 26.6 (27.00) 1 (06.60)
Cry6 I 61.0 67.1 6.1 (10.00) 7.2 ± 1.6 (10.59) 13 (93.20) 12.67 ± 1.52 (84.33)
II 65.3 71.8 6.5 (9.95) 11 (73.20)
III 77.0 86.1 9.1 (11.82) 14 (86.60)
ANOVA summary for mortality
Source Sum of squares Degree of freedom Mean of squares F value P value
Treatment (Between groups) 78740741 2 39.37037 70.87 \0.0001
Error 13.333 24 0.5556
Ss/Bl
Total 92.074 26
HSD Tukey test
HSD [0.05] = 3.24; HSD [0.01] = 4.71
No ICP vs. Cry1Ac Non-significant
No ICP vs. Cry6 P \ 0.01
Cry1Ac vs. Cry6 P \ 0.01
ANOVA summary for loss of larval weight
Source Sum of squares Degree of freedom Mean of squares F value P value
Treatment (Between groups) 443.7622 2 221.8811 28.24 \0.001
Error 47.14 6 7.8567
Ss/Bl
Total 490.9022 8
HSD Tukey test
HSD [0.05] = 7.03; HSD [0.01] = 10.23
No ICP vs. Cry1ac Non-significant
No ICP vs. Cry6 P \ 0.01
Cry1ac vs. Cry6 P \ 0.01
Each leaf was coated with 250 ng/ll of insecticidal protein. Difference in the body weight of surviving larvae before and after 2 days of
treatment was taken as indicative of their growth rate. Mortality in the larvae was recorded after 5 days of incubation
A. Sharma et al.: Cry6B is Toxic to Hypera postica
123
indicates that larvae do start feeding on the coated leaves
but subsequently fails to do so. Bt toxins are known to
induce the paralysis of larval midgut, followed by the death
of the larvae due to starvation and massive septiemia [3].
Feeding on Cry toxins is known to retard the growth of
larvae of various insects [8, 16]. Larvae with retarded
growth are more prone to other infections since, B. thur-
ingiensis and other bacteria are known to be more virulent
on weakened insect larvae [12].
Conclusion
The study describes identification of toxicity range of the
Cry6B protein. The E. coli-expressed Cry6B was found to be
toxic to lucerne weevil. High mortality in Cry6B fed insects,
their less growth, and less damage of leaves suggests a
possible bio-pesticidal control molecule to minimize the
damage caused by lucerne weevil. Transgenesis of M. sativa
with Cry6B gene, thus, seems to be an answer to the problem
imposed upon lucerne crop by the lucerne weevil.
Acknowledgments Financial assistance from the Department of
Biotechnology, Ministry of Science and Technology, Government of
India, New Delhi, is gratefully acknowledged.
References
1. Beegle CC, Yamamoto T (1992) History of Bacillus thuringiensisBerliner research and development. Can Entomol 124:587–616
2. Bradford M (1976) A rapid and sensitive method for the quan-
titation of microgram quantities of protein utilizing the principle
of protein-dye binding. Anal Biochem 72:248–254
3. Bravo A, Jansens S, Peferoen M (1992) Immunocytochemical
localization of Bacillus thuringiensis insecticidal crystal proteins
in intoxicated insects. J Invertebr Pathol 60:237–246
4. Chandra A, Pandey KC (2006) Resistance source for weevil
(Hypera postica Gyll.) in genus Medicago. IGFRI Newsl. 12:7
5. Feitelson JS (1993) The Bacillus thuringiensis family tree. In:
Kim L (ed) Advanced engineered pesticides. Marcel Dekker, Inc.,
New York, NY, pp 63–71
6. Feitelson JS, Payne J, Kim L (1992) Bacillus thuringiensis:
insects and beyond. BioTech 10:271–275
7. Guo S, Liu M, Peng D et al (2008) New strategy for isolating
novel nematicidal crystal protein genes from Bacillus thuringi-ensis strain YBT-1518. Appl Environ Microbiol 74:6997–7001
8. Jackson RE, Marcus MA, Gould F et al (2007) Cross-resistance
responses of Cry1Ac-selected Heliothis virescens (Lepidoptera:
Noctuidae) to the Bacillus thuringiensis protein Vip3A. J Econ
Entomol 100:180–186
9. Mark T, Mark K, Guy C (1999) US Patent 5874288—Bacillusthuringiensis toxins with improved activity
10. Pandey KC, Hasan N, Bhaskar RB et al (1995) Losses-problem
and present state of knowledge. In: Pests and diseases of major
forage crops. All India Co-ordinated Research Project of Forage
Crops, I.G.F.R.I., Jhansi, India, pp 2–3
11. Sambrook J, Fritsch EF, Maniatis T (1989) Molecular cloning: a
laboratory manual, 2nd edn. Cold Spring Harbor Laboratory
Press, New York, p 253
12. Schnepf E, Crickmore N, Van Rie J et al (1998) Bacillus thur-ingiensis and its pesticidal crystal proteins. Microbiol Mol Biol
Rev 62:775–806
Table 4 Bioassay for calculation of lethal concentration values
Bt Toxin Conc. (ng/ll) Mortality (N: 15 9 3 = 45)
I II III Total Percent
Cry6 0 0 0 0 0 0
125 2 1 1 4 9
250 7 7 8 22 49
500 10 12 11 33 75.5
750 14 14 13 41 91
1000 15 15 14 44 98
Cry1Ac 0 0 0 0 0 0
125 0 0 0 0 0
250 1 1 1 3 7
500 2 1 2 5 11
750 2 2 1 5 11
1000 3 2 2 6 13
The leaves were coated with purified Cry6B in the range 100 to
1,000 ng/ll. Five lucerne larvae were released per leaf in three such
replicates and mortlity of the larvae were calculated on fifth day of
release of larvae. Each experiment was independently carried out thrice
Lethal concentrations LC50 and LC90 values are 280 and 630 ng/ll,
respectively
Cry1Ac vs Cry6B
Call: survdiff(formula = Surv(time, censor) ~ group)
N Obs Exp (O-E)^2/E (O-E)^2/V Cry1Ac 300 65 235 123 425 Cry6B 300 300 130 220 425
Chisq= 425 on 1 degrees of freedom, p= 0
0
50
100
150
200
250
300
1 2 3 4 5 6 7 8 9
Days
Mo
rtal
ity
Cry1Ac Cry6
Fig. 4 Survivalship curve of lucerne larvae allowed to feed on
Cry1Ac and Cry6B toxins coated leaves. Bt toxins were applied on
the leaves at concentration slightly higher than the LC50 value (i.e.,
375 ng/ll). Mortality was monitored per day till ninth day of setting
up the bioassay. No mortality, but yellowing of the larvae, was
observed with in 2 days of inoculation. All the larvae fed upon Cry6B
coated leaves were found dead on ninth day, while the larvae on
Cry1Ac coated leaves started pupating
A. Sharma et al.: Cry6B is Toxic to Hypera postica
123

13. Schnepf EH, Schwab GE, Payne J et al. (1998) U.S. Patent
5,753,492—Genes encoding nematode-active toxins from Bacil-lus thuringiensis strains
14. Singh LN (1987) Fodder production strategies for temperate and
sub-temperate regions of India. In: Singh P (ed) Forage produc-
tion in India. Range Management Society of India. IGFRI, Jhansi,
pp 21–27
15. Wei J-Z, Hale K, Carta L et al (2003) Bacillus thuringiensiscrystal proteins that target nematodes. Proc Natl Acad Sci
100:2760–2765
16. Wu X, Leonard BR, Zhu YC et al (2009) Susceptibility of
Cry1Ab-resistant and -susceptible sugarcane borer (Lepidoptera:
Crambidae) to four Bacillus thuringiensis toxins. J Invertebr
Pathol 100:29–34
A. Sharma et al.: Cry6B is Toxic to Hypera postica
123
Copyright © 2022 FDOKUMEN




















