
Rissian, Eemian and Würmian Coleoptera assemblages from La Grande Pile (Vosges, France)
41
ELSEVIER Palaeogeography, Palaeoclimatology,Palaeoecology114 (1995) 1-41 Rissian, Eemian and Wiirmian Coleoptera assemblages from La Grande Pile (Vosges, France) Philippe Ponel Laboratoire de Botanique historique et Palynologie (Bofte 451), UA CNRS 1152, Facultk des Sciences et Techniques de Saint Jbrdme, F-13397 Marseille Cedex 20, France Received 31 January 1994; revised and accepted 24 August 1994 Abstract The Grande Pile peat-bog sequence is one of the few west European sites that cover the entire time span of the last major climatic cycle (140,000 years). A recent program of coring has provided material for insect analysis. The aim of this palaeoentomological study is to interpret the environmental and climatic evolution from the end of the Rissian glaciation to the Holocene using subfossil Coleoptera. The studied samples yielded 394 taxa of Coleoptera, half of them identified to species level; 19 of which do not belong to the present-day French fauna. The large number of taxa suggests a wide variety of habitats and provides much detailed palaeoecological evidence for the period studied. The lowermost sediments of the sequence, corresponding to the end of the Rissian glaciation, were deposited under very cold conditions in a tundra environment. This is succeeded by a forest period in which two cool interludes of grassland environment occur. Although these periods are decidedly poor in tree-dependent Coleoptera they do not contain any really cold-adapted taxa. They divide the forest phase into three periods. The first one, corresponding to the Eemian Interglacial, shows an early stage in which the beetle fauna is characterized by species dependent on deciduous trees, a later stage in which this fauna is mixed with many conifer-dependent elements, some of which (e.g. Platypus oxyurus) suggest warmer and perhaps wetter climatic condition than today. The two later woodland periods yielded coleopteran assemblages rather similar to those recorded in the second part of the Eemian, i.e. with both deciduous- and conifer-dependent taxa. There is some evidence to suggest that these two periods were slightly cooler than the Eemian proper. Marked climatic deterioration becomes obvious in the upper half of the sedimentary sequence attributed to the last glacial period (Wiarm), with the reappearance of tundra beetle assemblages. Sediment and insect evidence suggest that the climate was extremely cold and continental at La Grande Pile at about 30,000 B.P. A comparison of the insect analysis with previous palynological works enables precise correlation between the results provided by these two independent approaches. However, large numbers of running-water Coleoptera in the forest periods, replaced by standing-water Coleoptera in cold periods, raise questions concerning the lacustrine origin of the sedimentation at La Grande Pile. 1. Introduction The Grande Pile site shows an almost con- tinuous sedimentary record, about 20 m thick, covering the last climatic cycle from the Riss glaciation through the whole of the Last Interglacial Complex, much of the Last Glaciation and the Holocene. It was the first west European 0031-0182/95/$9.50 © 1995 ElsevierScience B.V. All rights reserved SSDI 0031-0182(95)00083-2 site for the interpretation of the palaeoclimate of the last 140,000 years. Subsequent pollen analysis of two additional sites, the Echets mire near Lyon (Beaulieu and Reille, 1984) and the Massif Central maars (Reille and Beaulieu, 1990; Beaulieu and Reille, 1992b), have been shown to cover the same period. Since its discovery (Seret, 1967), La Grande Pile
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Rissian, Eemian and Würmian Coleoptera assemblages from La Grande Pile (Vosges, France)

E L S E V I E R Palaeogeography, Palaeoclimatology, Palaeoecology 114 (1995) 1-41
Rissian, Eemian and Wiirmian Coleoptera assemblages from La Grande Pile (Vosges, France)
Philippe Ponel Laboratoire de Botanique historique et Palynologie (Bofte 451), UA CNRS 1152, Facultk des Sciences et Techniques de
Saint Jbrdme, F-13397 Marseille Cedex 20, France
Received 31 January 1994; revised and accepted 24 August 1994
Abstract
The Grande Pile peat-bog sequence is one of the few west European sites that cover the entire time span of the last major climatic cycle (140,000 years). A recent program of coring has provided material for insect analysis. The aim of this palaeoentomological study is to interpret the environmental and climatic evolution from the end of the Rissian glaciation to the Holocene using subfossil Coleoptera. The studied samples yielded 394 taxa of Coleoptera, half of them identified to species level; 19 of which do not belong to the present-day French fauna. The large number of taxa suggests a wide variety of habitats and provides much detailed palaeoecological evidence for the period studied.
The lowermost sediments of the sequence, corresponding to the end of the Rissian glaciation, were deposited under very cold conditions in a tundra environment. This is succeeded by a forest period in which two cool interludes of grassland environment occur. Although these periods are decidedly poor in tree-dependent Coleoptera they do not contain any really cold-adapted taxa. They divide the forest phase into three periods. The first one, corresponding to the Eemian Interglacial, shows an early stage in which the beetle fauna is characterized by species dependent on deciduous trees, a later stage in which this fauna is mixed with many conifer-dependent elements, some of which (e.g. Platypus oxyurus) suggest warmer and perhaps wetter climatic condition than today. The two later woodland periods yielded coleopteran assemblages rather similar to those recorded in the second part of the Eemian, i.e. with both deciduous- and conifer-dependent taxa. There is some evidence to suggest that these two periods were slightly cooler than the Eemian proper. Marked climatic deterioration becomes obvious in the upper half of the sedimentary sequence attributed to the last glacial period (Wiarm), with the reappearance of tundra beetle assemblages. Sediment and insect evidence suggest that the climate was extremely cold and continental at La Grande Pile at about 30,000 B.P. A comparison of the insect analysis with previous palynological works enables precise correlation between the results provided by these two independent approaches. However, large numbers of running-water Coleoptera in the forest periods, replaced by standing-water Coleoptera in cold periods, raise questions concerning the lacustrine origin of the sedimentation at La Grande Pile.
1. Introduction
The Grande Pile site shows an almost con- tinuous sedimentary record, about 20 m thick, covering the last climatic cycle from the Riss glaciation through the whole of the Last Interglacial Complex, much of the Last Glaciation and the Holocene. It was the first west European
0031-0182/95/$9.50 © 1995 Elsevier Science B.V. All rights reserved SSDI 0031-0182(95)00083-2
site for the interpretation of the palaeoclimate of the last 140,000 years. Subsequent pollen analysis of two additional sites, the Echets mire near Lyon (Beaulieu and Reille, 1984) and the Massif Central maars (Reille and Beaulieu, 1990; Beaulieu and Reille, 1992b), have been shown to cover the same period.
Since its discovery (Seret, 1967), La Grande Pile

2 P. Ponel/Palaeogeography, Palaeoclimatology, Palaeoecology 114 (1995) 1-41
has been the subject of intense palaeoecological research: pollen analyses by Woillard (1973, 1974a,b, 1975, 1977a,b, 1978a,b, 1979)then by Beaulieu and Reille (1992a); algal (diatoms) analy- ses by Louis and Smeets (1981), Louis et al. (1981), Louis and Ermin (1983), Louis and Peters (1983), Louis et al. (1983), Cornet (1988); paly- nology and sedimentology (Seret et al., 1992); palaeoclimatology (Guiot et al., 1989, 1992, 1993). Up to now, no palaeoentomological investigation had been undertaken at La Grande Pile, although it is now well established (Buckland and Coope, 1991) that in Quaternary ecology and climatology, insects and particularly Coleoptera can yield pre- cise information and permit quantification of cli- matic reconstructions to be made (Atkinson et al., 1986, 1987).
It must be emphasized that the present study is the first palaeoentomological analysis tracing the environmental and climatic evolution from the end of the penultimate glaciation to the Holocene (this paper dealt with this period up to the phase of maximum cold of the Wt~rm Glaciation).
2. Study area and site
The location of the Grande Pile peat-bog is so well known that only a brief summary will be
given here (Figs. 1 and 2). It occupies a closed depression located on an interfluvial plateau about 20 m above the Ognon valley. On a geological point of view, this peat-bog is located on the southern vosgian piedmont which culminates at Ballon de Servance (1216 m). It lies in the Saint Germain basin made up of Visean coal-bearing formations covered with Triassic sandstone sedi- ments. These formations are buried under a thick fluvioglacial cover deposited during the Rissian and Wtirmian glaciations. The Triassic formations underlying the Grande Pile depression show an alternance of marl and sandstone sediments with local dolomitic facies (Th6obald et al., 1974).
The peat-bog itself has a surface area of 25 ha at an altitude of 325 m above sea level. It is, at the present day, isolated from the surface hydro- graphic network (Seret et al., 1990) and no tribu- tary feeds it (the Coleoptera provide evidence that, in the past, this isolation was not continuously present throughout the period studied). The mire has been recently drained by two channels, the first one northwestwards, the second one south- wards, dug for peat extraction.
The Ecromagny plateau (where La Grande Pile lies) was covered by the Linexert (or Rissian) ice sheet as evidenced by the presence of glaciolac- ustrine tills of the Linexert glaciation at the basis of the sedimentary sequence. The absence of
~ "~'-~ PARIS ~
EPlNAL I
FRANCE
Fig. 1. Location of the Grande Pile peat-bog (47°44'N, 6°30'14"E).
La Grande Pile
BESANCON

P. Ponel/Palaeogeography, Palaeoclimatology, Palaeoecology 114 (1995) 1-41 3
1 2 3
Fig. 2. Topographic map of the Grande Pile peat-bog and its surroundings. 1 = The Grande Pile peat-bog. 2 = Lakes. 3 = Contour levels every 10 m, altitude in m a.s.l.
Wtirmian till deposits suggests that it was not covered by the Wtirmian ice sheets. Thus, La Grande Pile is located between the westward exten- sion limits of the Rissian and Wtirmian moraines (Seret et al., 1990; Beaulieu et al., 1992).
The Grande Pile mire is today surrounded by a
forest that belongs to the phytosociological com- munity Quercion robori-petraeae; Carpinus betulus is also abundant and indicates the euro-siberian character of the local climate. Molinia coerulea occupies the peat-bog itself, concurrently with Sphagnum spp., Drosera rotundifolia, Oxycoccos

4 P. Ponel/Palaeogeography, Palaeoclimatology, Palaeoecology 114 (1995) 1-41
palustris, Eriophorum vaginatum, Menyanthes tri- foliata and several Carex: C. fusca, C. canescens, C. vesicaria, C. stellulata. Several tree species are also colonizing the mire itself, e.g. Betula pubescens, Populus tremula, Frangula alnus, Salix spp. and Quercus robur (Woillard, 1975). The mean temperatures for January and July are 0°C and 18.6°C, respectively (mean yearly temperature: 9.5°C). The yearly precipitation is 1040 mm.
3. Methods
3.1. Sampling
In order to get sufficient sample volume, two series of parallel cores were undertaken using a
stationary piston corer (O 8cm) (Aaby and Diggerfeldt, 1986). The first series of 8 corings about 1 m/1.5 m away from one another and about 20 m depth was made in the centre of the mire near to the site where Woillard's most com- plete diagram (1975, diagram X) was derived (Fig. 3, corings A, cores GP 90-1, GP 90-2, GP 90-3, GP 90-4, GP 90-5, GP 90-6, GP 90-7 and GP 90-8). The second series of cores (Fig. 3, corings B, cores GP 90-9, GP 90-10, GP 90-11, GP 90-12, GP 90-13 and GP 90-14) was bored nearer to the littoral area, 100 m southwards on the same line as the old GP XX coring site that yielded the diagram of Beaulieu and Reille (1992a). The cores were stored in PVC tubes. Only the core series from the central area (A) has been studied in detail so far.
~ drain LINEXERT " ~ \
,/ i'
" i "I\
N ~
o E }
$ i i
lOOm / / /"
,,,,, .,,.._.~f"~"~-?~ ~.
I I ,
i~.,,~, I ~ '"'" /7';' /I ......... ~.~ ................ ' I A l/ '~
'
o 5o ", % t','.
"' I~ J " , '< t I
', . . . . . - t/ , > , 7
Ili ditch/~', ..-~#
- - 4 .......... S';' ...// © location of the cores ana l ys~y - -W-~LARD ( 1 9 7 ~ - - - ~ % ] I ~ D > ~ . - ~ "~'~ " ,lir location ofthe core XX analysed by BEAULIEU & REILLE (1992a III ~ ' ,~ i ST GERMAIN
I palaeoentomological corings of June 1990 = ~-~ (A : cores 90-1 to 90-8 ; B : cores 90-9 to 90-14)
- - - peat-bog limit • boundary stones
forest tracks
I ' ' '¸' t t . . . . . .
ST GERMAIN
Fig. 3. Location of the cores on the Grande Pile peat-bog.

P. Ponel/Palaeogeography, Palaeoclimatology, Palaeoecology 114 (1995) 1-41 5
3.2. Lithostratigraphy
The correlation of the stratigraphy of the cores took into account the nature, texture, colour and consistency of the sediment, the latter classifying into 4 main types: sand, silt, clay, and gyttja (Fig. 4). This permits the equivalent layers to be recognized from one core to another and the definition of more or less homogeneous sedimento- logical zones. This operating procedure has already been used at La Taphanel peat-bog (Massif Central, France) (Ponel and Coope, 1990; Ponel et al., 1991), This method enabled the subdivision of nearly all the cores GP 90-1 to GP 90-8 into 44 sedimentary slices, 8-9 kg each, numbered from 0 to 41 (layers 3 and 4 are split in two: 3A-3B, 4A-4B). The location of the 44 samples is shown on Fig. 4; it is an "average" location since the equivalent levels collected within a sample do not necessarily correspond to the same depth in each core.
Good correlations can be established between our core stratigraphy and the stratigraphical logs published by Woillard (1975), but with some minor discrepancies. Of particular interest is a layer (sample 38 and 39) of an in situ intraclastic breccia which was not described in previous investigations at this site. This layer consists of fragments of bedded and disrupted sediments with no evidence of having been transported. It could indicate an episode of dessication or shallow water sedimenta- tion within the range of winter freezing that could similarly disrupt the bedding. It is clear however that the conditions that lead to the disruption did not occur either above or below this particular horizon. The role of the insect fauna in this interpretation will be discussed later (study of GP-A7 faunal unit).
3.3. Correlation with palynology and chronology
The chronology of this sequence (Fig. 5) is established by correlation with previous studies but is mainly derived from the interpretation pro- posed by Beaulieu and Reille (1992a); the latter is an attempt to correlate recent pollen diagrams and radiocarbon dates from La Grande Pile (Beaulieu and Reille, 1992a; Woillard, 1975, 1978a; Woillard
10.
11.
12-
13-
14-
15,
16.
17.
18.
19.
6 - ~ 4 1
7 - 3 8
" 3 7
35
3 4 9-
:~,~= 33
........... 2 9
...... 2 6
2 5
i;';'-'S' ¢ 2~
;;,; ...... 16
.... 15
, ,v 14 13:
. . . . 12 1'I
. . . . . 1 0
9 8 \M /
7
; ' , ; ( . . . . . . 6
v~ 4b . . . . . . . . . 4a:
~, 3a ~'-"-- - . 2 " - - - - - ' 1 : _- -_~
~ . ~ _ ~ c)
Fig. 4. Synthetic lithostratigraphy of the central core series at La Grande Pile and location of samples for insect analysis. I = Bedded clay, silt and sand, 2 = silt, 3 = organic silt, 4 = silty breccia, 5 = minerogenic gyttja, 6 = organic gyttja, 7--- Late Glacial gyttja. Numbers left: depth (m); numbers right: sampling levels.

,-? ,.c 1£ r~ m g g g o o o
m m m m m g g g g g g o o o o o o g g g ~ ~ g
I~ ~I 0 a'l" I N ~ ' I d i ' I 'O'S I I i ,
1 NlVMI~l~9 L S
p i ~ ~t
i I i i i i
,i
i ,
R a l l ~ l a x
-+++
g 8 ~ o "o®
o ~ +-
t . . . .
I
I! ........ i . . . . . . . ~+
+ I +++ + + ....... i
, , t r I i
I[ '
j,.,,LII , ,~ . . . . J . . . . . ill, + i I - j J r +~I.II~I. I h+l
I ' ' ' [+ , ,
+ + . ~ + ' , +, , I +
, . + -+ : ~ . . . . . , + + . ~ . . . - + ~ ~ + - + - - - J + ~ - - ~ + + - , . . . . . . +.+-4~+_..
g
, i
i
B 0
× ×
~ , .0
0
J j I [ [ J ! t ] ] .r ! ] I I [ ! I ! U I I i I+ ~ ! [ ! I + I J J f ! r itHitt ~ ' ~ .... ......../ . . - .............................+i...........'./.....~............i/ .........+...~:::i~..-.............../ ...- /. / ./-. . .- ..- / : . / ...- .... /..../~-.~..~>. + >
i i ii ++i iiiiiiii?.i.. !:.:.!!i!i))))) !! :: iiiiiiiiiiiiiii iiii~ iiiii .... / / / / ~:.:~:: • /::/." //' / : : / : / :

P. Ponel/Palaeogeography, Palaeoclimatology, Palaeoecology 114 (1995) 1-41 7
and Moock, 1982) on the basis of palynological events.
4. Insect analysis
4.1. Extraction procedure for insect remains
The sediment from each level was treated using the extraction method recommended by Coope (1986). The samples were first broken gently by hand in water in a bowl. Mild chemical treatment (sodium carbonate solution) was occasionally nec- essary. Water with macroremains mixed with dis- aggregated sediment was allowed to flow in a 300 #m sieve. The material collected in this way was largely made up of plant debris and in most cases an insect concentration had to be undertaken. The damp residue was mixed with kerosene, then excess oil poured off and water added to the bowl. After decanting the floating fraction, rich in insect remains, was then poured into the same sieve and washed, first with detergent then with water and with alcohol. The material obtained was sorted under a binocular microscope. The insect remains were eventually preserved in alcohol or glued on pieces of cardboard. Beetles macrofossils were identified by direct comparison with a modern reference collection.
A total of 41 samples were analysed in this study (the insect assemblages from levels 24, 25 and 28 have been left out because of possible contamina- tion during sampling). 394 taxa of Coleoptera were identified, more than half to species level (Table 1); 19 species present as fossils at La Grande Pile are not members of the present-day French fauna. Every sample yielded Coleoptera remains. Other insect remains such as chironomid larva heads and Trichoptera larval sclerites were observed but not identified. The sediment yielded also numerous remains of Crustacea such as Cladocera and Notostraca; the occurrences of the latter are briefly discussed in this paper in addition to beetle analysis.
4.2. The insect fauna
The Coleoptera may be classified into several categories according to their ecological require-
ments, i.e. aquatic species (both running-water and standing-water species), terrestrial species, riparian species (that live on the shores of standing-water or on fiver banks), coprophagous and coprophi- lous species (directly or indirectly dependent on mammal faeces), necrophagous species (that live on dead animals), tree-dependent species (insects linked to leaves, wood, bark, etc) and plant detritus feeders. The wide variety of habitat requirements suggests that the assemblages obtained from La Grande Pile shows that the insect fauna was derived from a broad area of diverse environments. The main part of the Coleopteran record probably represents a fauna that lived in the close vicinity around the site (Lemdahl, 1990). However, most beetles fly readily and some specimens may have originated some kilometres away from the site.
The list of taxa (Table 1) also indicates the occurrence of species with diverse climatic require- ments. Their modern distribution range from boreal, boreo-montane to mediterranean. The suc- cession of these species clearly reflects the large- scale climatic changes that took place in this part of Europe during the last climatic cycle (Pons et al., 1992).
4.3. Analysis of the ma& ecological groups at La Grande Pile
Total representation of Coleoptera (Fig. 6) The histogram of taxa shows some variations in
the number of taxa per sample and in the variations of the total number of individuals per sample. It should be noted that no sample is totally devoid of beetle remains. Great variation is found in the total number of individuals which ranges from as low as 15 individuals to as many as 260 individuals per sample. Two drastic decreases occurred around samplel 0 and around samples 17-18.
Tree-dependent Coleoptera (Fig. 7) In this category are gathered all conifer and
deciduous tree-dependent Coleoptera, the latter including willow and boreo-montane dwarf- willow-dependent beetles. The histograms of taxa and individuals (Fig. 7a) show clearly a dramatic change at the transition between sample 20 and

Tab
le 1
L
ist
of c
oleo
pter
a sh
owin
g th
e m
inim
um n
umbe
r of
ind
ivid
uals
rec
over
ed f
rom
eac
h sa
mpl
e. N
omen
clat
ure
and
taxo
nom
ic o
rder
fro
m L
ucht
(19
87).
Spe
cies
whi
ch
are
not
mem
bers
of
the
pres
ent-
day
Fre
nch
faun
a ar
e m
arke
d w
ith
an a
ster
isk
(*)
0 1
2 3A
3B
4A
4B
5
6 7
8 9
10
11
12
13
14
15
16
17
18
19 2
0 21
22
23
26 2
7 29
30
31
32
33
34
35
36
37
38
39 4
0 41
CA
RA
BID
AE
C
icin
dela
sp.
C
alos
oma
syco
phan
ta
(L.)
C
arab
us c
lath
ratu
s L
. C
. ca
ncel
latu
s C
. ni
tens
L.
C.
arve
nsis
Hbs
t.
1 C
arab
us s
p.
Neb
ria
gyll
enha
lli
(Sch
rnh.
) N
ebri
a sp
. N
otio
phil
us s
p.
1 1
*Ela
phru
s la
ppon
icus
G
yll.
E
laph
rus
ripa
rius
(L
.)
* Dia
chei
la a
rcti
ca
(Gyl
l.)
*D. p
olit
a (F
ald.
) 1
Lor
icer
a pi
lico
rnis
(F
.)
Dys
chir
ius
glob
osus
(H
bst.
) P
eril
eptu
s ar
eola
tus
1 (C
reut
z.)
Tre
chus
sec
alis
(P
ayk.
) T.
rub
ens
(F.)
1
T. o
btus
us E
r./
1 4-
stri
atus
(S
chrk
.)
Tre
chus
sp.
B
embi
dion
bi
punc
tatu
m (
L.)
B
. ni
tidu
lum
(M
arsh
.)
*B.
daur
icum
(M
ots.
) 1
1 B
. (T
este
diol
um)
sp.
1 B
. sc
happ
eli
Dej
. B
. ae
neum
Ger
m.
R
unic
olor
Cha
ud.
2
121
1 1
1 1
1 1
1 1
2 1
11
1 1
1 1
1 1
1
1
1
11
1 3
11
1
1 1 11
1
1 1
1
e~

B.
guttu
la (
F.)
Bem
bidi
on s
p.
* Pat
robu
s as
sim
ilis
Cha
ud.
Pat
robu
s sp
. B
rady
cellu
s ru
ficol
lis
(Ste
ph.)
Poe
cilu
s le
pidu
s (L
eske
) P
oeci
lus
sp.
Pte
rost
ichu
s pu
mili
o (D
ej.)
P.
dilig
ens
(Stu
rm)
P.
vern
alis
(Pa
nz.)
P
. ni
grita
(Pa
yk.)
P
. ni
ger
(Sch
all.)
P
. m
elan
ariu
s (I
ll.)
P
. m
adid
us (
F.)
P
tero
stic
hus
sp.
Aba
x sp
. Sy
nuch
us n
ival
is
(Pan
z.)
Cal
athu
s m
elan
ocep
h-
alus
(L
.)
~4go
num
eri
ceti
(Pan
z.)
A.
mue
lleri
(H
bst.)
A
. ful
igin
osum
(P
anz.
) A
gonu
m s
p.
Am
ara
luni
colli
s (S
ehdt
e)
A.
quen
seli
(Sch
~nh.
) A
mar
a sp
. A
. (C
yrto
notu
s) s
p.
HA
LIP
LID
AE
B
rych
ius
elev
atus
(V
anz.)
H
alip
lus
sp.
DY
TIS
CID
AE
H
yphy
drus
ova
tus
(L.)
C
oela
mbu
s im
pres
so-
punc
tatu
s (S
chal
l.)
Hyd
ropo
rus
palu
stri
s (L
.)
1 1
1 1
1 1 1
1 4
1 2 1 1
1
1
1 1 1 113
2 2
¢ 7"

Tab
lel(
cont
inue
d)
0 1
2 3A
3B
4A
4B
5
6 7
8 9
10
11
12
13
14
15
16
17
18
19 2
021
2223
26
2729
30
31
32
33
34
35
36
37
38
39
40
41
2 1
22
43
1 I
11
19
1
34
14
11
1
I1
12
511
Hyd
ropo
rus
sp.
Pot
amon
ecte
s gr
iseo
stri
atus
(G
eer)
P
. as
sim
ilis
(Pay
k.)
Pot
amon
ecte
s sp
. A
gabu
s bi
pust
ulat
us
(L.)
A
. st
urm
i (G
yll.
) *A
. ar
ctic
us (
Payk
.)
Aga
bus
sp.
llybi
us s
p.
Rha
ntus
sp.
C
olym
bete
s fu
scus
(L
.)
*C.
dola
brat
us
(Pay
k.)
Col
ymbe
tes
sp.
Gra
phod
erus
sp,
A
ciliu
s sp
. D
ytis
cus
sp.
1 1
I 1
1
1 1
1 1
GY
RIN
IDA
E
Gyr
inus
min
utus
F.
1 G
. aer
atus
Ste
ph.
?1
1 3
G. a
erat
us
2 1
1 1
Step
h./m
arin
us
Gyl
l.
Gyr
inus
sp.
1
1 1
1 1
1 1
RH
YS
OD
IDA
E
Rhy
sode
s su
lcat
us
(F.)
HY
DR
AE
NID
AE
H
ydra
ena
gr.
ripa
ria
Kug
. H
. gr
acili
s H
ydra
ena
sp.
Och
theb
ius
gr.
fove
olat
us
Ger
m.
Och
theb
ius
sp.
2 1
1 3
34
1
2 i
2
43
t
1
2
22
1
1 1 2
1
1
1
1 1
1 1
I 1
1 I
I
,2 4~
4~

Lim
nebi
us s
p.
Hyd
roch
us s
p.
Hel
opho
rus g
rand
is Il
L
* H.
sibi
ricu
s (M
ots.
) H
. aq
uatic
us (
L.)
*H
. ob
long
us L
eC.
type
11
. gla
cial
is V
illa
H
. br
evip
alpi
s Bed
el
H.
?fla
vipe
s F.
H.
gr. m
inut
us F
. H
elop
horu
s sp
. C
oelo
stom
a or
bicu
lare
(F
.) Sp
haer
idiu
m s
p.
Cer
cyon
mel
anoc
epha
- /u
s (L
.)
Cer
cyon
sp.
M
egas
tern
um
bole
to-
phag
um (
Mar
sh.)
C
rypt
ople
urum
m
inu-
tu
rn (
F.)
H
ydro
bius
fusc
ipes
(L
.)
Ana
caen
a sp
. La
ccob
ius
sp.
Eno
chru
s af
finis
( T
hunb
. )/c
oar c
tatu
s (G
r.)
Eno
chru
s sp
. C
haet
arth
ria
sem
inu-
lu
m (
Hbs
t.)
HIS
TE
RID
AE
P
lega
deru
s vu
lner
atus
(P
anz.
) P
arom
alus
fla
vico
rnis
(H
bst.
) H
iste
r sp
.
12
13
1 16
1
1 1
1 1
22
1 2 1
1
2
1
1
1
2 1
11
22
11
2
1 1
1 1
2 5
47
4
1 1 I
34
14
3
21
1
12
12
2
3 3
1
1 11
11
7 3
53
1
6 2
43
1 1
1
1 12
2
1 7
.¢
E"
xt~
SIL
PH
IDA
E
Than
atop
hilu
s sp
. P
tero
lom
a fo
rsst
roem
i (G
yll.
) N
ecro
phor
us s
p.
1
1 1
2 1
1

Tab
le 1
(co
ntin
ued)
0 l
2 3A
3B
4A
4B
5
6 7
8 9
l0
II
12
13
14
15
16
17
18
19
20
21
22
23
26
27
29
30
31
32
33
34
35
36
37
38
39
40
41
G
CA
TO
PID
AE
C
hole
ra s
p.
1 C
atop
s sp
. 1
1 I
1
LIO
DID
AE
G.
sp.
1 2
SC
YD
MA
EN
IDA
E
Neu
raph
es s
p.
1 1
I 3
2
PT
ILID
AE
A
crot
rich
is s
p.
I 1
1
ST
AP
HY
LIN
IDA
E
Mic
rope
plus
te
sser
ula
Cur
t.
Meg
arth
rus
sp. 1
M
egar
thru
s sp
.2
Pro
tein
us s
pp.
Eus
phal
erum
spp
. A
crul
ia i
nfla
ta (
Gyl
l.)
Pyc
nogl
ypta
lur
ida
(Gyl
l.)
Acr
oloc
ha c
f. s
ult'u
la
(Ste
ph.)
O
mal
ium
cae
sura
G
rav.
*O
loph
rum
?co
nsim
ile
(Gyl
l.)
O. f
useu
m (
Gra
v.)
*0.
bore
ale
(Pay
k.)
Olo
phru
m s
p.
Euc
neco
sum
br
achy
pter
um
(Gra
y.)
Aci
dota
cre
nata
(F
.)
A.
crue
ntat
a (M
ann
h.)
Le
stev
a sp
. G
eodr
omic
us n
igri
ta
(Mal
l.)
G. p
lagi
atus
(F
.)
G.
cf.
kunz
ei H
eer
Ant
hoph
agus
sp.
1 1
1
I 2 1
I 2
2 4
2 8
6
2 2
1 2
I 21
15
10
1
1
3 2
2 1
3 2
1 1
1 1
1
I~1
12
1
32
1
3
I
1 1
1 1
1
1 1
1 1
I 1
8 1
I 1
1 1
1 2 1
2 1
1 2
1 1
1 1
1 1 1
e~
e~
o~
e~
t~

* Bor
eaph
ilus h
enni
ngi-
an
us S
ahib
. *B
. nor
dens
kioe
ldi
Mhk
l.
Apl
oder
us c
aela
tus
(Gra
y.)
Oxy
telu
s in
seca
tus
Gra
v.
O. p
iceu
s (L
.)
O. l
aque
atus
(M
arsh
.)
O. s
culp
tura
tus
Gra
v.
*0.
?pol
itus
Er.
O
. niti
dulu
s G
rav.
1
*0.
gibb
ulus
Epp
. O
xyte
lus
sp.
Pla
tyst
hetu
s co
rnut
us
(Gra
v.)
Ble
dius
sp.
St
enus
sp.
1
1 E
uast
hetu
s b i
punc
tatu
s (L
jung
h)
Scop
aeus
sp.
La
thro
bium
1
term
inat
um G
ray.
X
anth
olin
us s
p. s
l. 1
Stap
hylin
us s
p.
Phi
lont
hus/
Que
dius
1
1 sp
. M
ycet
opor
us s
p.
1 B
olito
bius
sp.
1
Tach
ypor
us
chry
som
elin
us (L
.)
Tach
ypor
us s
p.
Tach
inus
rufip
es
(Cre
er)
T. l
atic
ollis
Gra
v.
T. c
ortic
inus
Gra
v.
T. e
long
atus
Gyl
l.
T. ?
fimet
ariu
s Gra
v.
Tach
inus
sp.
Myl
laen
a sp
. A
leoc
hari
nae
inde
t.
1011
1
PS
EL
AP
HID
AE
B
atri
sode
s sp
. B
ryax
is s
p.
1 1
1 2
1
1 1
1 1
1
1 1
1 1
I1
1 2
12
12
1
1
1
1 6
2 1
1 1
1 1
3 1
1 4
1 2
1 7
3 1
1 2
1 I
1 1
2 2
4
12
12
14
21
1
1
11
111
1 12
1 1
1 1
1 1
1 1
1 1
1 1
1 1
2 2
1 5
1 2
4 5
5 2
8 1
1 2
1 ?1
1
1 1
1 1
2 4
126
134
8 14
109
7 1
6 26
128
4 10
4
1 1
3 3
8 5
3 15
4
19
10
5 1
74
2
¢%
-&
U,

Tab
le 1
(co
ntin
ued)
.~
CA
NT
HA
RID
AE
R
hago
nych
a sp
, M
alth
odes
sp.
ME
LY
RID
AE
H
aplo
cnem
us s
p.
Das
ytes
sp.
EL
AT
ER
IDA
E
Am
pedu
s cf
. ni
gerr
imus
(La
c.)
Ade
loce
ra m
urin
a ( L
. I
Cte
nice
ra
pect
inie
orni
s (L
.)
C.
eupr
ea (
F.)
Cte
nice
ra s
p.
Pro
ster
num
te
ssel
latu
m
(L.)
Se
lato
som
us a
eneu
s (L
.)
Den
tico
llis
line
aris
(L
.)
Fle
utia
uxel
lus
mar
itim
us (
Cur
t.)
Ela
teri
dae
inde
t, p
l. sp
.
EU
CN
EM
IDA
E
?Hyl
ocha
res
dubi
us
(Pil
l. M
itt.
) G
. sp
.
TH
RO
SC
IDA
E
Thro
scus
car
inifr
ons
Bon
v.
BU
PR
ES
TID
AE
A
gril
us s
p.
G.
sp,
HE
LO
DID
AE
in
det.
0 1
2 3A
3B
4A
4B
5
6 7
8 9
1 I
1 1
I t
1 1
2 1
1 I
1 1
1 1
l 1
1 1
2 2
1
l0
II
12
13
14
15
16
17
18
19
20
21
22
23
26
27
29
30
31
32
33
34
35
36
37
38
39
40
41
1 I
3
1 1
3
2 1
1 1
1 2
1 1
1 5
t 2
1
2 1
1
1
18
42
1
1 1
o~ 7"

DR
YO
PID
AE
D
ryop
s sp
. E
lmis
t.a
enea
(M
Oll
.)
2 E
solu
s pa
ralle
lipip
edus
(M
oll.
) O
ulim
nius
tube
rcul
atus
(M
Oll
.)
O.
trog
lody
tes
(Gyl
l.)
Oul
imni
us s
p.
1 Li
mni
us
1 1
1 vo
lckm
ari
(Pan
z.)
L. o
pacu
s (M
Oll
.)
Lim
nius
sp.
N
orm
andi
a ni
tens
(M
011.
) R
iolu
s sp
. 1
BY
RR
HID
AE
C
ytilu
s sp
. B
yrrh
us s
p.
* Sim
ploc
aria
m
etal
lica
(Stu
rm)
OS
TO
MID
AE
N
emos
oma
elon
gatu
m
(L.)
NIT
IDU
LID
AE
C
ater
etes
sp.
M
elig
ethe
s sp
. E
pura
ea s
p.
Poc
adiu
s fe
rrug
ineu
s (F.)
RH
IZO
PH
AG
IDA
E
Rhi
zoph
agus
de
pres
sus
(F.)
R
hizo
phag
us s
pp.
2 1
2 3
1 1
1 1
1
1 2
7 4
8 5
5 1
?1
6 9
4 6
2 3
1 2
3 4
2 2
2 1
4 1
35
6
2 3
2 2
8 1
3 3
1 5
2 3
5 2
8 4
3 1
1 3
1 1
2 1
3 1
1 1
18
7
3 10
I1
1
12
71
44
8
1 1
01
01
83
21
71
71
3
8 8
7 1
1 6
2 1
1 3
7 2
3 7
15
21
10
4
8 10
7
6 9
6
1 1
1 1
11
1
11
1
45
12
1
1
1 1
1 2
1 2
1 1
1 1
1 1 1
3
2 1
1 2
1 2
1 1
4 1
41
21
48
21
26
2
4 2 1
1 2
1
1 1
1 1
1 1
CU
CU
JID
AE
M
onot
oma
brev
icol
lis
Aub
6 A
irap
hilu
s sp
. Si
lvan
opru
s fa
gi (
Gu6
r.)

Tab
le I
(c
onti
nued
)
0 1
2 3A
3B
4A
4B
5
6 7
8 9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
26
27
29
30
31
32
33
34
35
36
37
38
39 4
0 41
1 P
edia
cus
derm
esto
ides
(F
.)
Laem
ophl
oeus
bi
mac
ulat
us (
Pay
k.)
Laem
ophl
oeus
sp.
CR
YP
TO
PH
AG
IDA
E
Cry
ptop
hagu
s sp
. C
(M
icra
mbe
) sp
. 2
3 1
A to
mar
ia s
p.
3
PH
AL
AC
RID
AE
P
hala
crus
car
icis
1
2 S
turm
P
hala
crus
sp.
O
libru
s sp
. 1
1 I
G.
sp.
I
LA
TH
RID
IID
AE
E
nicm
us s
p.
I C
orti
cari
a/C
orti
cari
na
2 2
1 1
5 6
sp.
type
CO
LY
DII
DA
E
Pyc
nom
erus
ter
ebra
ns
(oi.)
D
itom
a cr
enat
a ( F
. )
Col
ydiu
m e
long
atum
(F
.)
C fi
lifo
rme
(F.)
EN
DO
MY
CH
IDA
E
?End
omyc
hus
sp.
1 12
11
4
e~
CO
CC
INE
LL
IDA
E
?Epi
lach
na s
p.
Scym
nus
sp.
Chi
loco
rus
bipu
stul
atus
(L.
) * H
ippo
dam
ia
7-m
acul
ata
( Gee
r)
1
1

H.
arct
ica
(Sch
neid
er)
Har
mon
ia 4
-pun
ctat
a (P
ont.)
CIS
IDA
E
Sulc
acis
?bi
dent
ulus
(R
osh.
) C
/s s
p.
AN
OB
IID
AE
D
ryop
hilu
s pus
illus
(G
yll.)
E
rnob
ius
cf.
nigr
inus
(S
turm
) E
rnob
ius
sp.
Ano
bium
pun
ctat
um
(Gee
r)
A. o
f. fu
lvic
orne
Stu
rm
A. c
f. pe
rtin
ax (
L.)
A
nobi
um s
p.
Dor
cato
ma
sp.
PT
INID
AE
P
tinus
fur
(L
.)
OE
DE
ME
RID
AE
O
edem
era
luri
da
( Mar
sh.)/
vire
scen
s (L
.)
AN
TH
ICID
AE
A
nthi
cus
sp.
MO
RD
EL
LID
AE
A
nasp
is h
umer
alis
(F
.)
Ana
spis
sp.
SE
RR
OP
AL
PID
AE
D
irca
ea 4
-gut
tata
(P
ayk.
)/au
stra
lis
Fair
m.
SC
AR
AB
AE
IDA
E
Geo
trup
es s
p.
Ont
hoph
agus
ve
rtic
icor
nis (
Lai
ch.)
O
ntho
phag
us s
p.
1 2 1
1
1 3 1
E-
.¢
E"
oa
4~

Tab
le 1
(co
ntin
ued)
o
c
Aph
odiu
s t.
foss
or
(L.)
*A.
hold
erer
i R
eitt
er
A.
rufip
es (
L.)
A
. t.
fim
etar
ius
(L.)
A
phod
ius
spp.
Se
rica
bru
nnea
( L
.)
?Tri
odon
ta s
p.
Ano
mal
a sp
. V
algu
s he
mip
teru
s (L
.)
LU
CA
NID
AE
C
eruc
hus
chry
som
elin
us
(Ho
ch.)
CE
RA
MB
YC
IDA
E
Rha
gium
bifa
scia
tum
iF
.)
dlos
tern
a ta
baci
colo
r (G
eer)
CH
RY
SO
ME
LID
AE
D
onac
ia c
lavi
pes
F.
D.
?ver
sieo
[ore
a (B
rah
m)
D,
thal
assi
na G
erm
, D
. ei
nere
a H
bst.
D
onae
ia s
p.
Don
aeia
/Pla
teum
aris
sp
. P
late
umar
is s
p.
Mae
ropl
ea
appe
ndic
ulat
a (P
anz.
) Le
ma
sp.
Cry
ptoc
epha
lus
?mor
aei
Cry
ptoc
epha
lus
sp.
Ado
xus
obsc
urus
( L
.)
0 1
2
2 2
11
2
!
3A
3B
4A
4B
5
1 1
6 5
2 4
67
89
2 1
2 1
1 1
1 1
1 1
1
1 1
1 1
1 l
10
11
12
13
14
15
16
17
18
19
20
21
22
23
26
27
29
30
31
32
33
34
35
36
37
38
39
40
41
2 1
06
1
1
1 2
1 3
1 1
2 1
1 I
1 1
1 1
1 1
2 7
4 1
5 4
2 1
1 1
1 !
1 1
l l
1 1
I
1
1 1
,2
-m
e,
4t~

Chr
ysom
ela
sp.
C. ?
cere
alis
L.
Chr
ysoc
hloa
sp.
P
haed
on s
p.
Pla
giod
era
vers
icol
ora
(Lai
ch.)
P
hyto
dect
a vi
min
alis
(L
.)
Phy
tode
cta
sp.
Phy
llode
cta
latic
ollis
Su
ffr.
P
hyllo
dect
a sp
. G
aler
uca
tana
ceti
(L.)
Lu
peru
s sp
. A
gela
stic
a al
ni (
L.)
P
hyllo
tret
a sp
. A
phth
ona
sp.
Hal
tica
sp.
Cre
pido
dera
sp.
C
halc
oide
s fu
lvic
orni
s (F
.)
Cha
etoc
nem
a sp
. C
assi
da s
p.
BR
UC
HID
AE
B
ruch
us/B
ruch
idiu
s sp
. B
ruch
idiu
s fas
ciat
us
(Ol.
) B
. de
bilis
(GyU
.)
B.
unic
olor
/deb
ilis
(Gyn
.)
AN
TH
RIB
IDA
E
Bra
chyt
arsu
s ne
bulo
sus
( For
st. )
G
. sp
.
SC
OL
YT
IDA
E
Scol
ytus
cf
. am
ygda
li (G
u&
.)
S. i
ntri
catu
s (R
atz.
) S.
cf.
mal
i (B
echs
t.)
S. c
f. e
arpi
ni (R
atz.
) S.
sco
lytu
s (F
.)
S. r
atze
burg
i Jan
son
6
4
2 8
1
5 4
6 3
2 4
3 5
2 3
6 1
1 1 4
2 1
2 2
2
2
2
1 1 1
2 1
4 1
1
1 I
1
1 1
E"

Tab
le 1
(co
ntin
ued)
0 l
2 3A
3B
4A
4B
5
6 7
8 9
10
11
12
13
14
15
16
17
18
19 2
0 21
22
23
26
27
29
30
31
32
33
34
35
36
37
38
39 4
0 41
S. m
ulti
stri
atus
1
(Mar
sh.)
H
ylas
tes
ater
(P
ayk.
) 1
?1
3 1
2 8
4 H
ylur
gops
pal
liat
us
1 6
3 2
5 4
6 1
1 2
(Gyl
l.)
Pol
ygra
phus
1
1 5
3 6
5 3
39
11 4
2
2 3
,~
poli
grap
hus
(L,)
P~
?su
bopa
cus
Tho
rns.
1
1 2
2 H
yles
inus
ole
iper
da
1 ~
- (F.)
~-
Lepe
risi
nus
vari
us
1 (F.)
Kis
soph
agus
hed
erae
I
""
(Sch
mit
t.)
Kis
soph
agus
/ ~
""
Xyl
echi
nus
sp.
~ P
ityo
phth
orus
2
1 !
! pi
tyog
raph
us (
Rat
z.)
~.
Pit
yoph
thor
us s
p.
I t,,
,,,
Pit
yoge
nes
I I
2 2
I 5
2 8
4 "~
ch
aleo
grap
hus
(L.)
P
. tr
epan
atus
2
I 2
5 2
(N6r
dl.)
~"
P
. bM
enta
tus
(Hbs
t.)
1 P
ityo
phth
orus
sp.
1
Pit
yokt
eine
s sp
inid
ens
5 8
2 ,~
(R
eitt
er)
P.
curv
iden
s (G
erm
.)
4 "~
P
. vo
ront
zow
i 10
12
8 (J
acob
s.)
Ort
hoto
mic
us
1 su
tura
lis
(GyU
.)
O.
lari
cis
(F.)
I
?Ort
hoto
mic
us s
p.
l lp
s se
xden
tatu
s 1
2 1
1 2
(Boe
rner
) X
yleb
orus
sax
esen
i I
1 (R
atz.
) X
. dr
yogr
aphu
s 1
(Rat
z.)
Xyl
oter
us d
omes
ticu
s 3
(L.)
X
ylot
erus
sp.
1

PL
AT
YP
OD
IDA
E
Pla
typu
s ox
yuru
s (D
uf.)
P.
cylin
drus
(F.
)
CU
RC
UL
ION
IDA
E
Pse
laph
orhy
nchi
tes
nanu
s (P
ayk.
) P
. to
men
tosu
s (G
yll.
) R
hync
hite
s ?a
urat
us
(Sco
p.)
Rhy
nchi
tes
sp.
Byc
tiscu
s be
tula
e (L
.)
B. p
opul
i (L
.)
Dep
orau
s tr
istis
(F
.)/se
min
iger
Rtt
. 1)
. bet
ulae
(L
.)
Api
on c
erdo
Ger
st.
Api
on s
pp.
Otio
rhyn
chus
gr.
cla
v-
ipes
(B
onsd
.)
O.
dubi
us (
StrU
m.)
O
. ruf
ifron
s (G
yU.)
O
tiorh
ynch
us s
p.
Phy
llobi
us/
Pol
ydru
sus
sp.
Pol
ydru
sus
sp.
Stro
phos
oma
capi
tatu
m (
Gee
r)
Bar
ynot
us o
bscu
rus
(F.)
Sito
na
of.
flave
scen
s (M
arsh
.)
Sito
na s
p.
Cle
onus
(sl.)
sp.
D
ryop
htho
rus
cort
ical
is (
Payk
.)
Rhy
ncol
us e
long
atus
(G
yU.)
Rhy
ncol
us s
p.
Sten
osce
lis s
ubm
uric
a-
tus
(Sch
finh
.)
Bag
ous
sp.
Not
aris
acr
idul
us (
L.)
N
. ae
thio
ps (
F.)
Ant
hono
mus
sp.
1 5 1
91
2 2
1 1
1
2112
11
1 1
1 1
1 1
11
1
22
23
21
1
1 1
2 1
322
1 1
1 211
5 5
1 1
3 1
1
231
1 1 1 14
1
1 1
7 3
2 1
1 1
1
1 11
11
11
21
1
1
1
11
31
35
31
14
21
1
.¢
7"

Tab
le 1
(co
ntin
ued)
0 I
2 3A
3B
4A
4B
5
6 7
8 9
l0
ll
12
13
14
15
16
17
18
19 2
0 2l
22
23
26 2
7 29
30
31
32
33
34
35
36 3
7 38
39
40
41
Bra
chon
yx p
inet
i 1
1 1
(Pay
k.)
Cur
culio
ven
osus
1
l 1
1 (G
ray.
) C
. gl
andi
um M
arsh
. 2
3 1
.~
C. p
yrrh
ocer
as
1 I
3 2
5 3
2 1
1 1
6 1
Mar
sh.
Pis
sode
s pi
ni (
L.)
1
P.
?har
cyni
ae (
Hbs
t.)
1 ~"
M
agda
lis
niti
da
2 2
3 4
1 1
(Gyl
l.)
Alo
phus
tri
gutt
atus
Lim
noba
ris
T-al
bum
I
l (L
.)/p
ilis
tria
ta
(Ste
ph.)
E
ubry
chiu
s ve
lutu
s I
Phy
tobi
us m
uric
atus
B
ris.
/gra
natu
s G
yll.
-
~"
Mic
relu
s er
icae
t
1 1
2 (G
yll.
) F
Gym
netr
on
sp.
l 1
Mia
rus
sp,
Ano
plus
pla
ntar
is
8 2
1 2
(Nae
zen)
R
hync
haen
us
quer
cus
2 2
2 2
1 2
2 2
-Q
(L.)
R_
ave
llan
ae
1 1
~-~
(Oon
ov.)
R
. ru
sci
(Hbs
t.)
3
R.
foB
orum
(M
fill
.)
2 1
~ 1
1 3
1 1
1 R
. (P
seud
orch
este
s)
1 sp
. R
hync
haen
us
sp.
1 1
1 R
ham
phus
pul
icar
ius
1 1
1 !
1 (H
bst.
) 1

P. Ponel/Palaeogeography, Palaeoclimatology, Palaeoecology 114 (1995) 1-41 23
41 40 39 38 37 36 35 34 33 32 31. 30 28 2;' 26 23 22 21 20 19 18 17 16 15 14 13 12 11 10
9 8 ?
6 5
4B 4A 3B 3A
2 1 0
0 260 0 lO I I I I I I I I I I l l I I I r i l l I I I I I l l l l l t l l " l I I g. .
l l
......... ~m~mmm _ _
m mmm m m m m m m
.-.I
x --e
~o 133
$ $ (--
--i
GP-A7b
GP-A7a
GP-A6
GP-A5
GP-A4
-GP-A3 -
GP-A2b
Fig, 6. Total number of taxa (T) and individuals (/) of Coleoptera per sample.
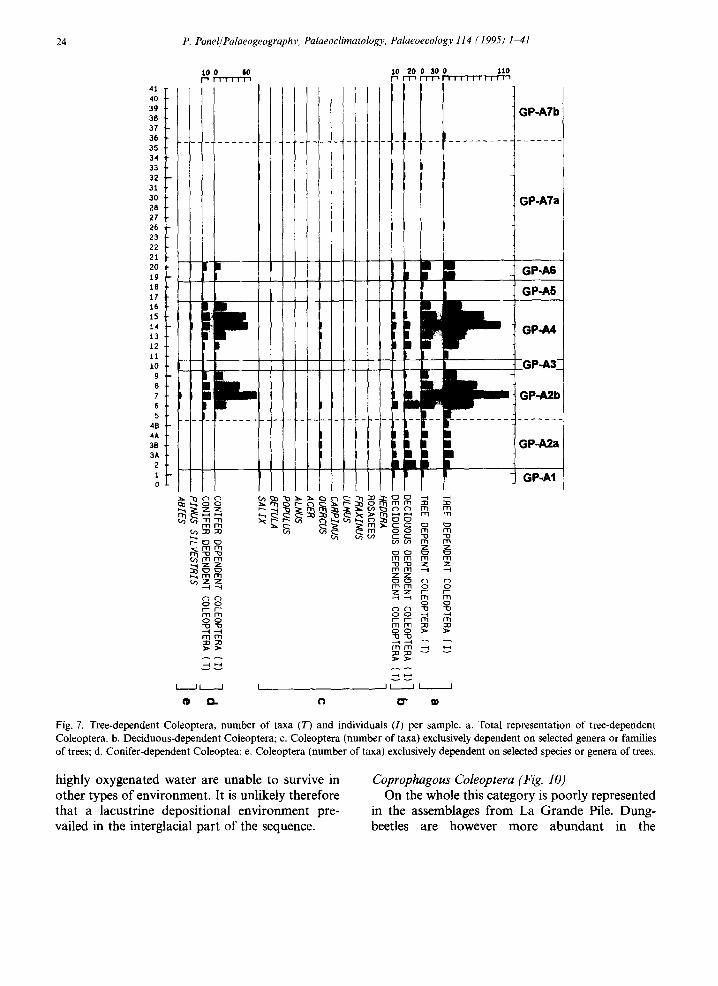
sample 21: the tree-dependent Coleoptera which are common or very common from sample 1 to sample 20 disappear almost totally from sample 21 upwards; only a few isolated individuals of willow-dependent taxa persist. The basal sample 0 does not contain any tree-dependent Coleoptera. As described above, both histograms show two
sudden decreases during the predominantly forest period: one in sample 10, the other in samples 17-18.
Many beetle species are dependent on deciduous trees. Fig. 7b shows the variations in numbers of taxa and individuals of such species. However, it is interesting to analyse the histograms relating to beetles specifically linked to particular host-plants (Fig. 7c). Comparisons with previously published pollen diagrams (Woillard, 1975, 1979; Beaulieu and Reille, 1992a) show an excellent agreement between pollen and beetle analysis, for example the agreement between oak pollen and oak- dependent Coleoptera; the beetle taxa linked to deciduous Quercus appear in samples that are exactly equivalent to those from which deciduous oak pollen has been recorded.
When compared with deciduous tree-dependent taxa, conifer-dependent taxa occur rather later in the sequence; the first individuals are not being recorded until sample 5 (Fig. 7d,e). Here again, the data obtained from palaeoentomological analysis agree perfectly with those obtained from pollen analysis, for example the bark-beetle Platypus oxyurus lives in the trunks of silver fir and its occurrence here matches the pollen curve for Abies (Fig. 8).
Aquatic Coleoptera (Fig. 9) This histogram clearly indicates that numbers
of aquatic taxa and individuals are remarkably stable throughout the sequence, implying that aquatic environments were continuously present on the site. It is much more informative however to split the frequency histogram into two, one for running-water Coleoptera and another one for standing-water Coleoptera. This reveals a striking contrast between the two categories. Running- water species are markedly predominant (up to 71 individuals in sample 14) in the forest domi- nated part of the sequence. From the top of the Saint Germain period, standing-water species become relatively more abundant. It is worth noting that sample 1 (the Riss glacial period) with its cold-adapted species of Coleoptera is also characterized by a predominance of standing- water Coleoptera.
Running-water Coleoptera being dependent on
24 P. Ponel/Palaeogeography, Palaeoclimatology, Palaeoecology 114 (1995) 1-41
41 40
39 38 37 36 35 34 33
32 31 30 28 27 26 23
22 21 20
19 18 17 16 15 ~4 13 12 11
10 9 8 7 6 5
4B 4A 3B 3A
2 1 0
10 0 60 10 20 0 30 0 110 i-~ i i [ i l ' l , ~'-~ I ' - ~ i 11 ,1"~ i i I I I [ ~ i 7 ,
~.~ I'rl I'rl 7- : 7
I '~ I '~ m 0 3 I'rl m "13 "X3 Z Z
O O O O
m r ~
I II , , ,i I ~ 1 I I I
Fig. 7. Tree-dependent Coleoptera, number of taxa (T) and individuals (I) per sample, a. Total representation of tree-dependent Coleoptera. b. Deciduous-dependent Coleoptera; c. Coleoptera (number of taxa) exclusively dependent on selected genera or families of trees; d. Conifer-dependent Coleoptea; e. Coleoptera (number of taxa) exclusively dependent on selected species or genera of trees.
highly oxygenated water are unable to survive in other types of environment. It is unlikely therefore that a lacustrine depositional environment pre- vailed in the interglacial part of the sequence.
Coprophagous Coleoptera (Fig. 10) On the whole this category is poorly represented
in the assemblages from La Grande Pile. Dung- beetles are however more abundant in the

P. Ponel/Palaeogeography, Palaeoclimatology, Palaeoecology 114 (1995) 1-41 25
41 40 39 38 3? 36 35 34 33 32 3t 30 28 21 26 23 Z2
21 20 19
17 16 15 14 13 12 11 10
9 B 7 6 5
4B 4A 3B 3A
2 1 0
20 10 0 700 40 10 0 80 m r- . I i i i ~ i i i ~1 i i 1 i 1--,i f i i i i ) i i I
-..t
¢o
F i g . 8. C o m p a r i s o n o f t h e o c c u r r e n c e s o f Abies p o l l e n a n d
Platypus oxyurus.
lowermost and the uppermost levels (where Aphodius holdereri appears) of the sequence (samples 1 and 2, samPles 33-37), that is those corresponding to the glacial episodes, suggesting that many of the herbivorous mammals lived on the open grassland but avoided the thick forest.
4.4. The sequence of coleopteran assemblages and their palaeoenvironmental interpretation
In order to make easier the description of the faunal and palaeoenvironmental changes through-
41 40 39 38 37 36 35 34 33 32 31 30 28 2;' 26 23 22 21 20 19 18 17 16 15 14 13 12 11 10
9 8 7 8 5
48 4A 3B 3A
2 1 0
I
m m m m
l,,
"n ¢.rl ~10
~--4 t:~ H E:J
g~g g ~To
Fig. 9. Aquatic Coleoptera, uals (I).
I I I GP-A6 I l l | | GP-A~
, .n.. l ' GP-A4 |
' l | | | • GP-A3
number of taxa (T) and individ-
out the sequence, the beetle assemblages are grouped into 7 main faunal units numbered from GP-A1 to GP-A7. They have been established on the basis of differences in the specific composition of the Coleoptera assemblage as a whole and not on the significance of certain indicator species. The term unit is preferred to zone because the latter

26 P. Ponel/Palaeogeography, Palaeoclimatology, Palaeoecology 114 (1995) 1-41
20 r r 7
41 40 39 38 37 36 35 34 33 32 31 30 28 2 l 26 23 22 21 20 19 18 1? 16 15 14 13 12 11 10
9 8 7 6 5
4B 4A 3B 3A
2 1 0
S " o
g 212
?, r -
Fig. 10. Coprophagous Coleoptera, number of individuals.
pearance of cold-adapted Coleoptera, the abun- dance of tree-dependent and running-water Coleoptera. This unit may be divided in two sub- units GP-A2a (rich in deciduous tree-dependent Coleoptera but totally devoid of conifer-dependent Coleoptera) and GP-A2b (many conifer-dependent taxa, mixed with deciduous tree-dependent Coleoptera).
--GP-A3: a small unit showing a significant decrease in tree-dependent Coleoptera and the dominance of standing-water Coleoptera over run- ning-water Coleoptera.
--GP-A4: this unit is very similar to GP-A2b, but with sporadic occurrence of isolated specimens of the relatively cold-adapted species Potamonectes assimilis.
--GP-A5: fairly similar to GP-A3, with a pro- nounced decrease of tree-dependent Coleoptera, a slight rise of standing-water beetles and a rare occurrence of cold-adapted taxa.
--GP-A6: this unit has similar beetle assem- blages to that recorded in units GP-A4 and GP-A2b. Tree-dependent Coleoptera reappear but are less abundant in GP-A6 than in GP-A4 and GP-A2b. There is a predominance of running- water Coleoptera. Cold-adapted Coleoptera are rare.
--GP-A7: This large unit is made up of 18 samples. The beetle assemblage shows a great change compared with the lower samples. The tree-dependent taxa disappear almost totally, cold- adapted and standing-water species increase in numbers and there is a corresponding decline in the numbers of running-water beetles. This unit may be divided into two subunits GP-A7a and GP-A7b, the latter is defined by an increase in cold-adapted species and an almost total loss of any running-water element.
has been used in a rather different manner to subdivide pollen diagrams. 4.5. Details o f faunal units
Main faunal units - -GP-AI: characterized by the occurrence of
cold-adapted Coleoptera, the scarcity of tree- dependent Coleoptera and the high number of standing-water Coleoptera.
--GP-A2: characterized by the complete disap-
GP-A 1 faunal unit This unit is characterized by the presence of a
number of cold-adapted beetle species that do not occur in the Interglacial samples above. Many of these have arctic distributions today (e.g. Diacheila polita, Bembidion dauricum, Amara quenseli,

P. Ponel/Palaeogeography, Palaeoclimatology, Palaeoecology 114 (1995) 1-41 27
Helophorus glacialis, Eucnecosum brachypterum, Hippodamia arctica).
Diacheila polita is today widespread in the tundra of northwest North America and Eurasia. In Fennoscandia it is restricted to the eastern part of the Kola Peninsula in northern Russia. It usu- ally lives on the peaty soil of open tundra, some- times on the margin of pools with Carex or in drier places where Betula nana occurs (Lindroth, 1985). Bembidion dauricum is almost circumpolar. In North America it is restricted to the west of the Hudson Bay and to the Rocky Mountains. In Scandinavia it is known from a very few localities only, where it is limited to the birch zone and to the lower alpine region of the mountains. It occurs mainly on rather dry and sandy soils with sparse vegetation. It is found under stones among dry grass (Lindroth, 1985). Amara quenseli is a circum- polar species present also in Iceland and in Scotland. This rather xerophilous species inhabits open environments such as sandy or gravelly soils with scarce vegetation; it is characteristic of grass- lands and heaths in alpine and subalpine regions (Lindroth, 1986). Eucnecosum brachypterum is more widely distributed: British Isles, northern Scandinavia, Central Europe from Germany to Russia, Alps (but not in the French Alps), Transylvania, Bulgaria, Caucasus, Siberia, north Mongolia, North America, mainly in subalpine and alpine regions (Zanetti, 1987). Helophorus glacialis is a boreo-alpine taxon present in Scandinavia and in the high mountains of southern and central Europe. It is the most stenotherm Helophorus species (Angus, pers. comm.). Restricted to glacial snow-melt water, usually in shallow ponds left on black soil behind the retreating ice, sometimes in rocky or clayey ponds (Hansen, 1987). In the southern Europe moun- tains, Helophorus glacialis is typical of glacier regions, at about 2700-2800 m (Mani, 1968). Hippodamia arctica is a very northern species that is not found south of latitude 65°N; in the south- ernmost part of its area it is restricted to moun- tains. According to Strand (1946), Hippodamia arctica was found on Salix scrub as well as on Betula, Empetrum and Arctostaphylos. Like many ladybirds, H. arctica probably feeds upon aphids
according to Strand (1946) and Coope and Sands (1966).
With this beetle assemblage that corresponds clearly to an arctic tundra fauna are associated some species whose modern distribution and ecol- ogy do not fit with such an hostile environment, like Plagiodera versicolor and Gonioctena viminalis that feed upon willows (but not upon dwarf wil- lows according to Koch, 1992), Galeruca tanaceti that feeds upon Compositae (Tanacetum vulgare, Achillea millefolium) or Perileptus areolatus and Bembidion iricolor whose geographical ranges are dominantly southern European today (Lindroth, 1985). Two hypotheses may be put forward to explain the anomalous presence of these two rela- tively southern species: ( 1 ) long-distance transport, or (2) the samples covered a climatic transition. The first hypothesis is improbable because La Grande Pile is located in a plain which does not favour long distance eolian transport, as it is the case with some mountain sites (Ponel et al., 1992; Tessier et al., 1993). The second hypothesis is more likely since the top of sample 1 certainly corres- ponds to a period of very sudden climatic improve- ment leading to the temperate conditions that prevail in the overlying faunal unit (GP-A2).
During most of the GP-A1 unit, the local envi- ronment of La Grande Pile may be described as open and extremely cold, similar to an arctic tundra, with a dominantly herbaceous vegetation and scattered shrubs on which Hippodamia arctica may have hunted aphids. The presence of open water is suggested by the dytiscid Potamonectes griseostriatus. However, the occurrence of several xerophilous ground-dependent Coleoptera (Notio- philus, Bembidion dauricum, Amara quenseli) sug- gests that in places the surroundings of the site may have been rather dry. The top of GP-A1 unit shows a diversification of the willow-dependent fauna, with the leaf-beetles Plagiodera versicolor and Gonioctena viminalis, and the weevil Rhynchaenus saliceti. Running water and small streams are suggested by Perileptus areolatus and Linmius volckmari, the latter occurring as early as sample 1. This climatic improvement mentionned above was not followed by the immediate appear- ance of trees and tree-dependent Coleoptera sug- gesting a lag in their response time, possibly

28 P. Ponel/Palaeogeography, Palaeoclimatology, Palaeoecology 114 (1995) 1-41
because of unsufffcient soil maturity or slower rates of spread of woody plants (Coope and Angus, 1975).
GP-A2a faunal unit This unit is characterized by the abundance of
deciduous tree-dependent Coleoptera and by the total absence of conifer-dependent and cold- adapted Coleoptera. Curculio venosus, C. glandium and Rhynchaenus quercus feed exclusively upon oaks, so does Curculio pyrrhoceras that has its early live history in Cynips quercusfolii galls, a parasitic Hymenoptera living only on oaks. The occurrence of ash is attested by Hylesinus oleiperda and Leperisinusfraxini, while the presence of Acer pseudoplatanus is indicated by Deporaus tristis/ seminiger. This unit contains a vast number of willow, birch and poplar-associated beetles, e.g. Phyllodecta laticollis, Plagiodera versicolor, Chal- coides fulvicornis, Byctiscus populi, Rhynchaenus rusci and Rhamphus pulicarius. Bark-beetles are represented by many species, including Scolytus multistriatus and S. scolytus that are mainly elm- dependent. Some click-beetle larvae such as Adelocera murina and Ctenicera pectinicornis feed underground on various roots, whereas others develop in hollow trees. This is certainly the case for Denticollis linearis and Prosternum tessellatum, however the latter can also be found in alpine grasslands (Leseigneur, 1972). A true forest envi- ronment is clearly suggested by a number of species such as Pycnomerus terebrans, Colydium elongatum and Dryophthorus corticalis that live in old trees, or the anthribid Brachytarsus nebulosus, a parasite of Lecaninae (tree-dependent coccids) according to Hoffmann (1945). Herba layer taxa are poorly represented, with Bruchidae (the larvae of which develop within the seeds of Fabaceae) and Apion, including Apion cerdo which feeds on the genus Vicia.
Although the riparian Coleoptera are repre- sented only by Stenus, Lathrobium and several species of Donaeia, truly aquatic taxa are very abundant. This group is dominated by running- water insects such as Dryops, Elmis, Esolus, Oulimnius, Limnius, Normandia and Riolus.
Lastly, it should be noted that present in this unit and nowhere else, there is a chafer not iden-
tiffed yet but probably belonging to the genus Triodonta. All the Triodonta species today are confined to southern Europe so the presence of this taxon in this unit suggests particularly mild climatic conditions that were more favourable to such thermophilous Coleoptera than those of the present. Thus the overall environment of the close surroundings during unit GP-A2a may be described as a forested landscape with various deciduous tree species and a poorly developed herbaceous stratum (probably due to the density of trees). The extraordinary abundance of running- water beetles and the rarity of species of standing water suggests that the sediment was carried into the area by running water or even directly depos- ited by it.
GP-A2b faunal unit Deciduous tree-dependent Coleoptera are still
abundant, with many of the taxa already recorded in the underlying unit, but also Agelastica alni (specific to alder), Platypus cylindrus (mainly on oak) and Anoplus plantaris (specific to birch). The most important event is the addition of a rich xylophagous fauna made up of many conifer- dependent species, mostly scolytids and weevils, such as Hylastes ater, Hylurgops palliatus, Ips sexdentatus, Polygraphus polygraphus, Pityo- phthorus pityographus, Pityogenes chalcographus, P. bidentatus, P. trepanatus, Pityokteines spinidens, P. eurvidens, P. vorontzovi, Xyleborus dryographus, X. saxeseni, Rhyncolus elongatus, Magdalis nitida. Other taxa are corticolous, for example Rhizophagus spp, Colydium filiforme, Paromalus flavicornis and Plegaderus vulneratus. According to Lindroth (1985) the large Carabid Calosoma sycophanta also inhabits conifer and deciduous forests, since it is a predator species that exclusively feeds upon tree-dependent moth caterpillars (Lymantriidae, Thaumatopoeidae). Among tree- dependent Coleoptera, three other species (Rhysodes sulcatus, Ceruehus ehrysomelinus, Platypus oxyurus) with relict modern distributions are extremely significant from a biogeographical and ecological point of view. Rhysodes sulcatus is today a very rare species recorded from very few localities between Europe and Asia Minor: Bohemia, Slovakia (Freude, 1971), Lombardy,

P. Ponel/Palaeogeography, Palaeoclimatology, Palaeoecology 114 (1995) 1-41 29
Tuscany, southern Sweden, Haute-Savoie (Ch~tenet, 1986), extreme south-west of France (western Pyr6n6es woodlands) (Tiberghien, 1969). According to Koch (1989), it lives under beech barks, but in the Pyr6n6es, Tiberghien (1969) reports that this species is exclusively found in decaying fir trunks (Abies alba) in which it seems to be localized in a wet and dense deep wood layer, underlying a rotten superficial layer. Ceruchus chrysomelinus is highly characteristic of wet forest in the European mountains, in France it is known from the Jura, the Alps and the Pyr6n6es. According to Paulian (1959), it lives within old decaying stumps of Abies and Picea. It was discovered by Ponel et al. (1992) above 2000 m in the alpine zone of the Taillefer Massif (Is6re, France), in a fossil assemblage of forest beetles in Holocene peat deposits. Platypus oxyurus is today much more restricted than the common species P. cylindrus. In France it is confined to the northern slopes of the Pyr6n6es (Pyr6n6es°Atlantiques, Corbi6res) where it lives at middle altitude. Its European distribution is discontinuous since it is also recorded from Corsica (For& d'Aitone), Calabria, Greece (island of Euboea) and Turkey (no precise locality). This insect is specific to Abies, in the trunks of which it digs deep ramified galleries (Sainte-Claire Deville, 1914; Balachowsky, 1949). The present-day restriction of its distribution sug- gests that some climatic factors may be involved, since Abies is today widespread in Europe. Furthermore, because this species is unable to develop in branches or in small-size trunks of young trees, it indicates that adult trees were involved. Platypus oxyurus was also discovered in deposits that have been dated from an earlier interglacial (Hoxnian Interglacial), in regions located as far away from its present-day area as Britain is today (Shotton and Osborne, 1965; Coope, 1990). From a purely climatic point of view the occurrence of Rhysodes sulcatus, Ceruchus chrysomelinus and Platypus oxyurus is significant: the known localities for these three species are characterized by a high humidity with heavy rain- fall; it is especially the case for Rhysodes sulcatus and Ceruchus chrysomelinus (Herv6, 1951) whose humidity requirements seem to be very high. Thus the presence of these insects provides evidence for
the establishment in unit GP-A2b of a mature forest environment, with rather mild and wet cli- matic conditions.
GP-A3 faunal unit This unit is limited to sample 10 and is charac-
terized by an almost complete disappearance of the tree-dependent insect fauna, which is here represented by only a single specimen of Platypus oxyurus and a single specimen of Rhyncolus elonga- tus. Bearing in mind the decidedly non-forest char- acter of the assemblage as a whole, it is likely that these specimens really belong either to the overly- ing or the underlying faunal unit. Platypus oxyurus certainly appears to belong to the underlying unit GP-A2b in which it shows a major but short expansion spanning two samples, with up to at least 33 specimens. This problem may be attributed to inevitable imprecision in the cutting of the cores.
This faunal unit shows a striking absence of any cold-adapted taxa in this unit in spite of the almost total disappearance of tree-dependent insects, and thus of the mature forest environment. Two hypotheses can be presented: first the climate dete- rioration was perhaps too short to allow the north- ern fauna to establish itself. Atkinson et al. (1987) and Coope (1987) demonstrated the exceptionally fast ability of beetle species to respond to climatic change, so this hypothesis is unlikely to be correct. Moreover the decline of tree-dependent taxa and of the total number of individuals clearly begins as early as the middle of unit GP-A2b, suggesting that, if declining temperatures were responsible for the diminution in the numbers of trees it was by no means a short sharp deterioration. Second, the climatic deterioration may not have been severe enough to permit the incoming of really cold- adapted Coleoptera. This second hypothesis is supported by the occurrence of other taxa recorded in this assemblage, which include the riparian species Bembidion guttula, B. aeneum, Stenus, Platysthetus cornutus. Many aquatic beetles are also present, with a dominance of standing-water taxa such as Acilius, Hydroporus, Colymbetes, Hyphydrus ovatus, Helophorus spp. over running- water taxa. One may conclude from these data that the community that lived around La Grande Pile throughout unit GP-A3 is consistent with an

30 P. Ponel/Palaeogeography, Palaeoclimatology, Palaeoecology 114 (1995) 1-41
open grassland environment, with open birch forest as suggested by pollen analysis. The beetles recorded in this unit do not provide any evidence for a real arctic tundra. The Grande Pile sediments at this time would appear to have been mainly deposited in standing water.
GP-A4 faunal unit This unit, which corresponds to 6 samples, is
marked by the re-appearance of a beetle assem- blage fairly similar to that described in GP-A2b unit. This assemblage is characterized by a rich fauna of tree-dependent species indicating the pres- ence of both conifer and deciduous trees. Compared with unit GP-A2 some species have disappeared, e.g. Platypus oxyurus. Other species are recorded for the first time, for instance Alosterna tabacicolor (whose larvae develop in the dead wood of various deciduous trees), the scolyt- ids Orthotomicus laricis and O. suturalis which are both parasite of many coniferous trees, Xyloterus domesticus that lives exclusively on various decidu- ous trees and Kissophagus hederae, another scolytid species that develops in large stems and dying twigs of ivy, Hedera helix (Balachowsky, 1949). Nemosoma elongatum is a today a very rare tree- dependent Ostomidae indirectly linked to trees. It is not a phytophagous insect but feeds upon larval exsuviae and excreta produced by scolytids. Pissodes pini has larvae that dig superficial galleries under the bark of Pinus sylvestris and Pinus unci- nata. Brachonyx pineti feeds exclusively on Pinus sylvestris and has larvae that develop at the basis of the needles and not within the wood (Hoffmann, 1954). Other bark-beetles include Ditoma crenata, a predator on many scolytid species, Pediacus dermestoides, dependent on deciduous trees such as Quercus, Fagus, Acer and Laemophloeus bimacu- latus, a rare Cucujid that hunts Dryocoetes villosus inside its own galleries under oak and beech barks (Lefkovitch, 1958). The last five species occurs for the first time in this unit.
Most of the phytophagous taxa were already present in the lower units, but this is not the case for some weevils such as Rhynchites nanus (chiefly feeding on Salix, Betula, Alnus), Rhynchites tomen- tosus (feeding on Salix and sometimes on Populus) and Rhynchaenus avellanae (mainly feeding on
Quercus). Taxa belonging to the genera Phyllotreta, Apion and Bruchidius, provide evidence for the persistence of an herbaceous stratum, and an open- ing up of the environment is indicated by the ground-beetles Bradycellus ruficollis and Amara lunicollis which are usually found in moors or in forest clearings. The presence of marshes at the site is revealed by the occurrence of hygrophilous species such as Lathrobium terminatum and Stenus, and by many phytophagous taxa such as Eubrychius velutus (which feeds chiefly on Myriophyllum), Lirnnobaris and Donacia. Phala- crus caricis is a small phytophagous Coleoptera feeding on smutted Carex (Thomson, 1958), it is another typical marsh species. A few taxa indica- tive of standing water are recorded (Agabus, Enochrus, Anacaena, Coelostoma, Helophorus). However, running-water taxa (Elmis, Esolus, Oulimnius, Limnius, Normandia) are again domi- nant and represented by large numbers of individuals.
The occurrence of the water beetle Potamonectes assimilis is rather more difficult to interpret. Widespread today throughout northern and central Europe, it reaches the northeast of France, such as the lakes in Hautes-Vosges and Alsace (Guignot, 1947), Strasbourg, from where several recent findings are reported by Callot (1990). This insect does not occur in the underlying unit GP-A3 but appears for the first time in GP-A4 unit (in the lowermost and uppermost samples), where the abundance of tree-dependent Coleoptera suggest a temperate climate. The presence or absence of Potamonectes assirnilis may not depend only on thermal conditions, but also on the quality of water, this species being usually found in mountain lakes and springs according to most of the pub- lished data. Nevertheless the presence of this insect in GP-A4 unit probably denotes climatic condi- tions less temperate than those in GP-A2 unit.
GP-A5 faunal unit This unit, which is composed of only two
samples, presents similarities to GP-A3 unit. The fall in the number of tree-dependent taxa obviously indicates a marked thinning out of the forest although pollen analysis suggests the continued presence of some birch. The tree-dependent beetle

P. Ponel/Palaeogeography, Palaeoclimatology, Palaeoecology 114 (1995) 1-41 31
taxa do not disappear totally since Dircaea australis/quadriguttatus, Polygraphus polygraphus (on conifers), Deporaus betulae and Anoplus plan- taris (on birch), Curculio pyrrhoceras (on oak) persist in very small numbers. The Melandryidae species belonging to the genus Dircaea may also reveal the presence of birch since several living specimens of this extremely rare insect were discov- ered at La Grande Pile during the coring opera- tions, on erect dead birches covered with Polyporus (tree-dependent fungus). The cold-adapted species here are represented by Potamonectes griseostriatus only, a boreoalpine species whose distribution area expands from northern Europe to the Moroccan Atlas across the mountains of central and southern Europe. It is also known from northern Asia and boreal North America. In the Alps and the Pyrrnres it lives in high-altitude lakes (1800-2400 m) (Guignot, 1947). Some phytophagous genera such as Chaetocnema, Bruchidius, Apion, indicate the presence of herbaceous vegetation. The preda- tor ground-beetle Synuchus nivalis that lives on dry and sandy soils (Koch, 1989) suggests an open environment also.
GP-A6 faunal unit This unit is characterized by a moderate increase
of tree-dependent Coleoptera assemblages but in rather less profusion than in GP-A2 and GP-A4. Typical forest species from this unit are Acrulia inflata, Paromalus flavicornis, Laemophloeus bima- culatus, Colydium filiforme, Ceruchus chrysomeli- nus, Polygraphus polygraphus, Hylurgops palliatus, Scolytus intricatus, S. ratzeburgi, Strophosoma capitatum, Curculio pyrrhoceras, Dryophthorus cor- ticalis, Brachonyx pineti and Magdalis nitida. These species indicate a return of a truly forested land- scape. However, the occurrence of a single indivi- dual of the northern swamp weevil Notaris aethiops is of interest here since this insect is only known in France from two localities in Puy-de-D6me (Massif Central) where it lives in peat-bogs and on the border of mountain-lakes and in cold marshes (Hoffmann, 1958), and probably develops on various Cyperaceae. This species possibly pro- vides a hint that the climate was cooler than in either unit GP-A2 or GP-A4. Running-water aquatic genera such as Normandia, Esolus,
Limnius, Oulimnius, Normandia and Elmis once more return in abundance, both by the number of taxa and by the number of individuals present, and the still-water species are relatively rare, sug- gesting again that the sediment was washed in or even deposited by a stream.
GP-A 7faunal unit This long and rather homogeneous unit extends
over 18 samples. It is characterized by the almost complete disappearance of tree-dependent taxa except for the weevils Rhynchaenus foliorum and Anoplus plantaris, whose larvae mine the leaves of many willows (including dwarf-willows) and of birch (including Betula nana), respectively.
Phytophagous taxa from the herb layer are extremely rare, they are represented, for example, by Micrelus ericae, exclusively feeding on Calluna vulgaris and Erica tetralix.
The most striking feature of this faunal unit is the disappearance of the entire tree-dependent fauna and the appearance of a number of very cold-adapted species whose modern distribution is mainly restricted to northern Europe, Fennoscandia and the arctic regions of Russia, most of which are able to live above the tree-line or even obliged to do so. This cold fauna consists of ground Coleoptera (Carabidae) and aquatic Coleoptera (Dytiscidae, Hydraenidae, Hydro- philidae). Elaphrus lapponicus is only known today from northern England (one locality), Scotland and Fennoscandia. According to Lindroth (1985), this species occurs in the birch and the upper conifer region, and also in the lower alpine region. It inhabits small marshes with Carex, Eriophorum, mosses, etc. Also according to Lindroth, it is sometimes found associated with Diacheila arctica, probably because these species have similar ecolog- ical requirements; both species occur together in sample 36. This shows the extent to which the ecological requirements of Coleoptera remained stable despite the great climatic and biogeographi- cal upheavals they have to withstood during the glacial/interglacial cycles. Unlike many alpine or subalpine species, E. lapponicus is occasionally found in relatively warmer places, which is consis- tent with its preference for sunny microhabitats (Lindroth, 1985). Diacheila arctica inhabits

32 P. Ponel/Palaeogeography, Palaeoclimatology, Palaeoecology 114 (1995) 1 41
northern Scandinavia, the Kola peninsula and the north of Russia. It is an hygrophilous species living on lake shores and muddy depressions from the upper conifer zone to the lower alpine zone. Its favourite habitat seems to be fens with Carex, Eriophorum, Scirpus, Juncus and mosses (Lindroth, 1985). Diacheila polita, last found in GP-A1, reap- pears in this unit.
The circumpolar water beetles are represented by many taxa. Agabus arcticus is a carnivorous water beetle that lives in Sphagnum pools on peat- bogs, and on mossy lake shores in the north of the British Isles and in Fennoscandia (Balfour- Browne, 1950). Colymbetes dolabratus is today known from the north of Fennoscandia, the north of Russia, Siberia, North America, Greenland and Iceland (Coope, 1968). It lives in standing water or in slowy moving streams. Helophorus sibiricus is an holarctic species, widespread in the palaearc- tic region extending from the north of Fennoscandia to Mongolia and China. In Fennoscandia it lives on river margins, but it can also be found in shallow grassy pools, especially in the eastern part of its area (Angus, 1973; Hansen, 1987). Helophorus oblongus is known today from North America and Siberia; according to Angus (1973) this species is probably wide- spread in the Siberian taiga and tundra. In the southern part of its area this species is apparently restricted to forest although it is common else- where in grassland pools (Angus, 1973). Helophorus glacialis, a highly stenothermic species whose ecological requirements were described above (GP-A1 unit), reappears in almost every sample of GP-A7 unit.
Among the terrestrial insects, the silphid Pterolomaforsstroemi, is found in Scandinavia and also on high mountains of Central Europe. It lives in mosses, under gravel and pebbles, and along small streams and torrents where it preys upon molluscs (Chgttenet, 1986). The Omaliinae are common, with Euenecosum brachypterum pre- viously found in GP-A1 unit, here accompanied by three other species characteristic of this type of assemblage (Coope, 1975; Taylor and Coope, 1985): Pycnoglypta lurida (northern Europe, in the south toward Denmark and northern Germany; Siberia, North America; in wetlands and marshes
according to Zanetti, 1987), Boreaphilus henningia- nus (Scandinavia, north Russia and the Harz mountains; under plant debris, in wet mosses and grassy marshes according to Coope, 1962) and Boreaphilus nordenskioeldi (isolated localities in the north of North America and Asia; its ecological requirements are similar to those of the previous species according to Coope, 1975). Another sta- phylinid, the coprophilous Oxytelus gibbulus, is found living today only in the Caucasus mountains and maybe also in eastern Siberia (Hammond et al., 1979). The oldest fossils for this species are dated from ca 600,000 yr B.P. (Waverley Wood, Warwickshire) (Shotton et al., 1993). It was spo- radic in its occurrence at several sites in England dating from the mid-WOrm (Devensian, Weichselian). O. gibbulus was the most common staphylinid in England about 200,000 yr B.P. and was regularly associated with mammoths. Simplocaria metallica is widely distributed in Scandinavia and in few isolated localities in Central Europe. It is exclusively a moss feeder (Coope, 1961). Aphodius holdereri is today an asiatic species (a Tibetan endemic, known from lake Ko Ko Nor region in the north to the northern slopes of Himalaya, according to Coope, 1973). It appears in three samples only: 33, 36 and 37. The rare occurrences of these exotic species might be of use as stratigraphical markers permitting corre- lation of the Grande Pile sedimentary levels with the British sediments where similar fossil assem- blages occur within a restricted time period (Coope et al., 1961; Coope, 1969). According to Coope (1979), Aphodius holdereri is "by far the most abundant large dung-beetle in fossil assemblages from the British Isles that date from the middle of the Last Glaciation". The occurrence of this and many other exclusively Asiatic species in western Europe at this time (Coope, 1994) indicates a climatic episode whose modern analogues may be found in Tibetan mountains, at altitudes from 3000 to 5000 m.
As regards aquatic Coleoptera, running-water species decline gradually in the GP-A7 unit and are eventually replaced by standing-water species in the five upper samples, suggesting that the Glacial sedimentary environment was quite different from that of the Interglacial.

P. Ponel/Palaeogeography, Palaeoclimatology, Palaeoecology 114 (1995) 1-41 33
It is convenient to divide GP-A7 unit into two subunits:
--GP-A7a (samples 21-35) is characterized by the almost complete disappearance of tree- dependent species, the first indications of the re-appearance of the most cold-adapted Coleoptera, the persistence of running-water bee- tles and of many taxa that are rather catholic in their ecological requirements.
--GP-A7b (samples 36-41) is characterized by a rise in arctic-alpine Coleoptera, the complete disappearance of running-water beetles and an impoverishment of ubiquitous species. There can be little doubt that this fauna indicates a further increase in coldness. It is thus interesting to note that the breccia level contained in sample 38 and 39, thought to be the result of frost-shattering phenomena in shallow water, occurs just above the marked rise of many cold-adapted beetles, including the Tibetan dung-beetle Aphodius holder- eri, in samples 36 and 37. This may well be the result of the progressive intensification of the cold in which the increasingly harsh conditions allow a rich cold-adapted fauna to become established. Later, the thermal conditions must have become so severe that even many of the cold-adapted species eventually succumbed. The environment was by now similar to a polar tundra with an impoverished fauna highly adapted to extremely cold conditions. This interpretation is also sup- ported by the occurrence of the crustacean Notostraca Lepidurus arcticus Pallas which appears in huge numbers (223 mandibles were
recorded) in samples 38 and 39. The ecology and climatic requirements of this tadpole shrimp is summarized by Taylor and Coope (1985). It is today mostly found north of the Arctic circle in very shallow water of impermanent water bodies and is able to stand very cold conditions.
The climatic interpretation of sedimentological and biological data does not correspond exactly to the timing of the early land-ice glacial maximum dated between 50,000 and 30,000 yr B.P. described by Seret et al. (1990). By comparison (Fig. 5) with the pollen zones and dates provided by Beaulieu and Reille (1992a: figs. 1 and 5) the cold maximum inferred from arthropod assemblages may be dated roughly just after ca 30,000 yr B.P. As a whole, the insects of the GP-A7b unit indicate extremely harsh climatic conditions, similar to those pre- vailing today in arctic tundra or in the highest mountains.
Summary of the correlation of the coleopteran succession with pollen and isotope stratigraphy
The faunal units established here upon Coleoptera and those based on pollen zonation (Beaulieu and Reille, 1992a) and isotope zones (Woillard and Moock, 1982) may be readily corre- lated (Table 2). The two very cold periods GP-A1 and GP-A7 correspond to the termination of the penultimate glacial period (Linexert) and to the last glacial period (Pleniwiirm+Late Warm of Beaulieu and ReiUe, 1992a), respectively. Isotope stage 4 does not appear in the palaeoentomologlcal records; it may correspond to the non-analyzed
Table 2 Correlations between insect, pollen and extrapolated marine isotope zones
Faunal units Pollen chronozones Isotope stages Beaulieu and Reille (1992a) Woillard and Moock (1982)
GP-A7b Wiirmian Pleniglacial 2 GP-A7a Wttrmian Pleniglacial 3 samples missing Lower Wiirmian Pleniglacial 4 GP-A6 Saint Germain II 5a GP-A5 Mrlisey II 5b GP-A4 Saint Germain I 5c GP-A3 Mrlisey I 5d GP-A2b Eemian Interglacial 5e GP-A2a Eemian Interglacial 5e GP-A1 Riss (Linexert) 6

34 P. Ponel/ Palaeogeography, Palaeoclimatology, Palaeoecology 114 (1995) 1-41
samples 24 and 25. The forested periods GP-A2, GP-A4 and GP-A6 correspond to the Eemian, to the Saint Germain I and to the Saint Germain II, respectively. They were interrupted by two treeless episodes that are equivalent to Mrlisey I and Mrlisey II and show that trees were greatly reduced. As observed from pollen analysis by Pons et al. (1989), there is no entomological evidence for an extremely cold phase at La Grande Pile during these two last episodes. These correlations are in broad agreement with the palaeoenviron- ment implications derived by Beaulieu and Reille (1992a) from the pollen analysis of core XX.
4.6. Discrepancy between water beetles occurrences and the possible lacustrine origin of La Grande Pile
Fig. 9 clearly shows the great abundance of running-water Coleoptera in the samples attrib- uted to the Eemian and to the Saint Germain I and II, with two declines in abundance in samples 10 and 17 corresponding to M~lisey I and M~lisey II, respectively, which are short treeless spells interpreted as reflecting moderate climatical deteri- orations according to both pollen and insect analy- sis. Though occurring in low numbers, running- water Coleoptera continue to be present from sample 22 to sample 36 and then disappear totally from sample 37 upwards (beginning of the Late WOrm). The lowermost samples 0 and 1 (Linexert, termination of the Rissian glaciation) also contain very few running-water beetles. The occurrence of standing-water beetles is virtually the reverse of this; large numbers in sample 1, small numbers in samples 2-23, quite large numbers in 26-37, then large numbers again in the uppermost sample 41. As regards Lepidurus arcticus, the abundance of this crustacean in samples 38 and 39 indicates that very shallow water was present during the Upper Wiirmian Pleniglacial.
It is noteworthy that the insect data are sup- ported by some pollen data. Thus Fig. 1 in Beaulieu and Reille (1992a) shows that shallow and calm water plants such as Isoetes are practi- cally absent until the end of the Eemian. The Cyperacaeae, mostly shallow-water plants, are well represented at the end of the penultimate glacial period, then disappear until the Upper Wtirmian
Pleniglacial except two short occurrences in Mrlisey I and Mrlisey II (samples 10 and 17). Furthermore the isolated occurrences of Sphagnum in several points of the sequence indicate that exposed places were available and favoured the development of mosses. Lastly, the Ranunculus batrachium group appears just at the beginning of the Pleniglacial and increases gradually until the end of the sequence, probably in association with the infilling in of the site.
With regards to the diatoms, Cornet (1988) states that sedimentological data suggesting a high water level (several metres) are not in agreement with algal data which indicated rather shallow water at the end of the Eemian.
It is tempting to interpret the variations of running- and standing-water beetles as reflecting the hydrological regime that prevailed at La Grande Pile during the two last climatic cycles. The large numbers of running-water beetles com- bined with the scarcity of lake or pond taxa certainly indicate the presence or even the predomi- nance of small streams during the Eemian and other forested periods on this site. These streams would have carried sediments sporadically into the depositional environment and the rates of accumu- lation are unlikely to have been constant. Estimations of rates of environmental changes based on the assumption of constant sedimentary input are thus unlikely to be reliable.
The presence of such streams during these periods has further significant implications since the mire is located today on a slightly elevated interfluve plateau as stated by Cornet (1988) and by Woillard (1975): "Le bassin d'alimentation de la tourbi~re est tr~s limitr: aucun cours d'eau ne l'alimente et elle ne poss6de que deux exutoires de vidange (...)". The hydrology of the site must have been rather different in the past. La Grande Pile may not always have been in such a raised position above the alluvial plains of Ognon and Lanterne. Today the depression is only 20 m higher than the plains mentioned above, some geomorphological and hydrodynamical changes must have occurred contemporaneously with the climatical changes, especially during the Wiarmian deglaciation. The replacement of running-water Coleoptera by standing-water Coleoptera during cold episodes

P. Ponel/ Palaeogeography, Palaeoclimatology, Palaeoecology 114 (1995) 1-41 35
(Riss, Mrlisey I, Mrlisey II, Pleniglacial) suggests a fundamental environmental change and the replacement of a running-water environment by a marsh or a very shallow lake environment. During Pleniglacial times the depth of the water must have been very slight (occurrence of Lepidurus arcticus) to permit the frost disturbance of the deposit. Maybe a decrease in temperature and precipitation caused the interruption in water flow. Analysis of group B core series, extracted close to the margin of the Grande Pile depression 150 m away from group A, might provide some new insight into this intriguing problem.
4. 7. Climatic reconstruction
One of the most important aspects of modern Quaternary entomological analysis lies in the fact that many fossil sclerites can be identified to the species level. Furthermore these species appear to be identical to living specimens. A mass of biogeo- graphical, ecological and climatical data can thus be obtained from the rich literature that has been devoted to Coleoptera during the two last centu- ries. Moreover a recent computerized palaeocli- mate reconstruction method based on Coleoptera has been recently developed: the Mutual Climatic Range Method, or M.C.R.M. (Atkinson et al., 1986), used to obtain the climatic reconstruction presented in this paper. This method enables quan- titative palaeoclimatic reconstructions to be made that are not matched by any other palaeoecological approaches to continental environments. It is based on the range of climates corresponding to the total geographical area occupied today by the species present in a fossil assemblage. The climatic estimate proposed for the whole assemblage is given by the overlap of the climatic ranges of the species identified in this assemblage. It is important to note that the precision of this reconstruction method relies in part on the identification of the taxa to species level. Moreover, the taxa included in the M.C.R.M. data-base only include Coleoptera that are thought to be not directly dependent of higher plants: these insects are especi- ally significant from a palaeoclimatic point of view because their distribution is not likely to be linked to the occurrence of particular host-plants but
imposed by climatic factors. Usually, the food chains of such species ultimately depend on micro- organisms (algae, unicellular fungi,...).
At La Grande Pile this category is mainly represented by aquatic, riparian, terrestrial and coprophilous beetles belonging to the families Dytiscidae, Hydrophilidae, Staphylinidae, Cara- bidae and Scarabaeidae families (e.g. Elaphrus lapponicus, Diacheila arctica, Diacheila polita, Bembidion dauricum, Amara quenseli, Potamonectes griseostriatus, Potamonectes assimilis, Agabus arcti- cus, Colymbetes dolabratus, Helophorus glacialis, Helophorus sibiricus, Helophorus oblongus, Ptero- loma forsstroemi, Pycnoglypta lurida, Eucnecosum brachypterum, Boreaphilus henningianus, Borea- philus nordenskioeldi, Simplocaria metallica, Hippodamia arctica, Aphodius holdereri, Notaris aethiops; Fig. 11). Many of these cold-adapted taxa are only able to complete their biological cycle under very harsh thermal conditions and are of great value for climatic reconstructions. For example, it is the case for the circumpolar ground- beetle Diacheila polita or the Tibetan dung-beetle Aphodius holderei which cannot tolerate average summer temperatures much higher than about 10°C; on the other hand they can withstand and even seem to prefer average winter temperatures as low as - 1 2 / - 2 4 ° C (Coope, pers. comm.).
The Mutual Climatic Range Method has been used for each coleopteran assemblage obtained from the 41 Grande Pile samples (Fig. 12). This diagram shows clearly an early cold period repre- sented by samples 1 and 2 during which the climate was also very continental. The Eemian warming took place from sample 2 upwards after which temperature fluctuates slightly but remains, on average, quite high until sample 22. After a warm maximum of summer temperature centred on samples 5, 6, 7 (broadly synchronized with the Taxus phase in Fig. 5 and overlapping the peak of Platypus oxyurus), the second half of the Eemian shows slowly declining temperature. At La Grande Pile therefore no indication is found for any cold periods "to levels more typical of the mid-glacial period" during the Eemian (as reported by the G.R.I.P. project, 1993); this result is in accord with Boulton (1993). The modest climatic deterio- ration of Mrlisey I is clearly visible in sample 10,

3 6 P. Ponel/Palaeogeography, Palaeoclimatology, Palaeoecology 114 (1995) 1-41
10 10 10 10 0 30 i TM i-~ i-'n F - ~ I I ,
4, !I"i' 'l,ilE" ,o I i l t I i ! G 39 P-A7b 3 8
36 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 35 a,t
33 , a2 31 30 2~ 21 26 ~3 ~2
~0 19 18
16 15
13 12 11 10
9
?
5 4B
4A 313 3A
2 1 O
~
~ ~ NN
2 m ~
Fig. 11. Cold-adapted Coleoptera, number of taxa (T) and individuals (I): and occurrences of selected cold-adapted Coleoptera (number of individuals).
it does not indicate a return to the same level of coldness as sample 0 and 1. In contrast, the M61isey II episode does not appear clearly. Samples 21-22 correspond to a fundamental climatic deterioration which can be attributed to the beginning of the Pleniglacial. Temperature remains low until sample 41 (probably final Pleniglacial), except in samples 23 and 34 that appears to represent short warming.
According to Fig. 11, these events would seem to be related to a reduction in the numbers of the cold-adapted elements rather than the reappear- ance of therrnophilous species. These events are not clearly recorded by pollen analysis (Fig. 5) and illustrate the rapidity of response of Coleoptera in comparison with the reaction time of the vegetation. Sample 34 corresponds to the

P. Ponel/Palaeogeography, Palaeoclimatology, Palaeoecology 114 (1995) 1-41 3 7
'C
?ii i ? i ? \illii i? :ii? illllli ??/?Lil li????/????/??i
• ° I ' 2 l,.[3bl,.14b 516 I ' 1~ 1 9 ,o , ,I ,2f ,~l , , l , s[ ,~ ,el,8,912o 2,12212~1~612,1291,o1,,1,21,31~,1,5 ~61~71,,1~91,o1,, b G,,-,~, ~P-~. G~AZ. I ~ GP-A4 Q,.-,. GP,.A6 GP,.A7a GP-A7b
c L E E M I SG I M II SG II PW LW
"C
+ 10'
- 1 0 ,
. 2 0 ¸
- 30
iii iif: iiliiii:i - i ii ::liii i - ii:: iii i i:i!l !!i/ii!ili!!!i!!!!
. . . . . . . . . . . . . . . . . . . . . . . . . . . . ,..2122-- ;21212; ,°o12222-2221h . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Fig. 12. "Mutual climatic range" climatic reconstruction for the Coleoptera from La Grand Pile (°C). The horizontal axis is approximately equivalent to time B.P.a. Insect samples, b, Entomological biostratigraphic units, c. Pollen chronozones of Beaulieu and Reille (1992a), L= Linexert (or Riss), E= Eemian, M I= M61isey I, M H= M61isey II, SG I= Saint Germain I, SG H= Saint Germain II, PW= Pleniwllrm, LW= Late WOrm.
upper half of palynozone 8f, the termination of which (sharp rise of Pinus curve) is dated ca 34,000 yr B.P. (Fig. 5). This short warming may be related to one of the episodes of relatively mild climate conditions described during the mid and late parts of the last glaciation by Coope and Angus (1975), Coope (1977) and more recently by Johnsen et al. (1992). The coldest part of the climatic reconstruc- tion for the mean temperature of the warmest month corresponds to sample 38 (dated just after 30,000 yr B.P. according to Fig. 5) and (together with sample 39) to sediments that shows signs of frost activity, with an impoverished and extremely cold-adapted Coleopteran fauna (e.g. Diacheila polita, Boreaphilus nordenskioeldi) and the occur- rence of the crustacean Lepidurus arcticus, just after the occurrence of the Tibetan element Aphodius holdereri in samples 36 and 37.
No other climatic episodes are clearly recorded, either because such episodes are too faint to be detected by the insect record, or because of possible sedimentary hiatuses. However, no such sedi- mentary gaps were recognised during pollen analy- ses (Woillard, 1975; Beaulieu and Reille, 1992a). An analysis of climatic reconstructions based on both Coleoptera and pollen evidence is proposed by Guiot et al. (1993); the climatic data obtained from La Grande Pile are discussed by these authors.
5. Conclusions
This unique continuous beetle record in conti- nental Western Europe gives a detailed picture of the drastic changes in the insect fauna that corre- late with changes that affected the flora and the

38 P. Ponel/Palaeogeography, Palaeoclimatology, Palaeoecology 114 (1995) 1 41
vegetation. These changes clearly reflect the major climatic changes that took place within the last glacial cycle, i.e. the last 140,000 years, in this part of the world.
The steep warming of the transition Riss- Eemian lead to the replacement of a cold-adapted fauna and of a tundra-type environment by a tree- dependent insect fauna made up dominantly of deciduous tree-dependent species in the first part of the Eemian interglacial. In the second part of the Eemian this tree-dependent fauna was largely dominated by conifer-dependent taxa. After a warm episode that overlaps partly the peak of the Mediterranean Platypus oxyurus, a progressive cooling ended in a change of the environment during the Mrlisey I event when the tree-dependent species disappeared but were not replaced by very cold-adapted taxa The landscape was then similar to an open grassland rather than a true tundra. The return of a forest environment is proved by the development of a rich tree-dependent fauna that thrived in the next period, namely Saint Germain I. This fauna showed some similarities with that of the second part of the Eemian (how- ever Platypus oxyurus is now absent) and must be likened to a ta~ga-type insect fauna. After another cool event, namely Mrlisey II (which appears as a kind of replica of Mrlisey I), the Saint Germain II corresponds with a brief re-appearance of the forest fauna; it was abruptly interrupted by the faunal and environmental upheavals that marked the transition with the Pleniglacial and the estab- lishment of a very cold-adapted insect fauna, con- currently with a tundra-like landscape. A period of severe cold occurred towards the top of the sequence (sample 38) just after 30,000 yr B.P., as indicated by biological and stratigraphical evidence.
Concurrently with these fundamental climatic changes, the hydrological regime at the site seems to have undergone some important variations: the mild forested periods correspond to large numbers of running-water-dependent Coleoptera (Dryopidae) that live in highly oxygenated water only. Thus, this faunal community seems to indi- cate an hydrological regime characterized by the presence of streams. In contrast, the cold periods are characterized by the replacement of this fauna
by standing-water Coleoptera that denote the pres- ence of a lake or pool environment. The exact mechanism of these hydrological variations is not yet fully understood but raises interesting questions about the steadiness of the sedimentation at La Grande Pile.
Acknowledgments
First of all, I would like to thank G.R. Coope who took an essential part in the realization of this work. I am grateful to the "Commissariat h l'l~nergie Atomique" who supported this study and to V. Andrieu, J.-L. de Beaulieu, M. Campy, F. David, C. Goeury, J. Guiot, P. Guenet, M. Reille, W. Safar, G. Seret and his team for help and advice. Figs. 6-11 were drawn with the pro- gram GPAL3 created by Goeury (1988). Numerous helpful comments on an early version of the manuscript were made by H.J.B. Birks, G.R. Coope, P. de Deckker and G. Lemdahl; Michrle Pellet helped improve the final form of the English text.
References
Aaby, B. and Diggerfeldt, G., 1986. Sampling techniques for lakes and bogs. In: B.E. Berglund (Editor), Handbook of Holocene Palaeoecology and Palaeohydrology. Wiley, Chichester, pp. 181-194.
Atkinson, T.C., Briffa, K.R. and Coope, G.R., 1987. Seasonal temperatures in Britain during the past 22,000 years, reconstructed using beetles remains. Nature, 325: 587-592.
Atkinson, T.C., Briffa, K.R., Coope, G.R., Joachim, M.J. and Perry, D.W., 1986. Climatic calibration of coleopteran data. In: B.E. Berglund (Editor), Handbook of Holocene Palaeoecology and Palaeohydrology. Wiley, Chichester, pp. 851-858.
Angus, R,B., 1973. Pleistocene Helophorus [Coleoptera Hydrophilidae] from Borislav and Starunia in the Western Ukraine with a reinterpretation of M. Lomnicki's species. Description of a new Siberian species, and a comparison with British Weichselian faunas. Philos. Trans. R. Soc. London, B 265: 299-326.
Balachowsky, A.S., 1949. Faune de France. 50, Colroptrres Scolytidae. Libr. Fac. Sci., Paris, 320 pp.
Balfour-Browne, F., 1950. British Water Beetles, 2. Ray Soc., London, 394 pp.
Beaulieu, J.-L. de and Reille, M., 1984. A long Upper

P. Ponel/Palaeogeography, Palaeoclimatology, Palaeoecology 114 (1995) 1-41 39
Pleistocene pollen record from Les Echets, near Lyon, France. Boreas, 13:111-132.
Beanlieu, J.-L. de and Reille, M., 1992a. The last climatic cycle at la Grande Pile (Vosges, France): a new pollen profile. Quat. SCI. Rev., 11: 431-438.
Beaulieu, J.-L. de and Reille, M., 1992b. Long Pleistocene pollen sequences from the Velay Plateau (Massif Central, France). I. Ribains maar. Veget. Hist. Archaeobot., 1: 233-242.
Beaulieu, J.-L. de, Monjuvent, G., Nicoud, G., Richard, H. and Seret, G. (with the collaboration of: Campy, M., Clerc, J., Dricot, E., Either, U., Mandier, P., Ponel, P., Reille, M., Rousseau, D.D., Ruffaldi, P. and Wansard, G.), 1992. Long pollen sequences and the last glaciations from the Southern Alps to the Vosges mountains. In: 8th Int. Palynol. Congr., Aix-en-Provence, 13-16th Sept. 1992, Excursion I. Cah. Micropaltontol., 7: 215-257.
Boulton, G.S., 1993. Two cores are better than one. Nature, 366: 507-508.
Buckland, P.C. and Coope, G.R., 1991. A Bibliography and Literature Review of Quaternary Entomology. Collis Publ., Dep. Archaeol. Prehist., Univ. Sheffield, 85 pp.
Callot, H.J., 1990. Hydradephaga. In: Catalogue et atlas des Coltopt~res d'Alsace. Soc. Alsac. Entomol. Mus. Zool. Univ. Strasbourg, 68 pp.
Ch~tenet, G. du, 1986. Guide des Coltoptbres d'Europe. Delachaux et Niestlt, Lausanne, 480 pp.
Coope, G.R., 1961. A Pleistocene coleopterous fauna with arctic affinities from Fladbury, Worcestershire. Q. J. Geol. Soc. London, 118: 103-123.
Coope, G.R., 1962. Coleoptera from a peat interbedded between two Boulder Clays at Burnhead near Airdrie. Trans. Geol. Soc. Glasgow, 24: 279-286.
Coope, G.R., 1968. An insect fauna from Mid Weichselian deposits at Brandon, Warwickshire. Philos. Trans. R. Soc. London, B 254: 425-456.
Coope, G.R., 1969. The response of coleoptera to gross thermal changes during the Mid Weichselian interstadial. Mitt. Int. Ver. Theor. Angew. Limnol., 17: 173-183.
Coope, G.R., 1973. Tibetan species of dung beetle from Late Pleistocene deposits in England. Nature, 245: 335-336.
Coope, G.R., 1975. Mid-Weichselian climatic changes in Western Europe re-interpreted from coleopteran assem- blages. In: R.P. Suggate and M.M. Cresswell (Editors), Quaternary Studies. R. Soc. N. Z., Wellington, pp. 101-108.
Coope, G.R., 1977. Fossil coleopteran assemblages as sensitive indicators of climatic changes during the Devensian (Last) cold stage. Philos. Trans. R. Soc. London, B 280: 313-340.
Coope, G.R., 1986. Coleoptera analysis. In: B.E. Berglund (Editor), Handbook of Hoiocene Palaeoecology and Palaeohydrology. Wiley, Chichester, pp. 703-713.
Coope, G.R., 1987. The response of late Quaternary insect communities to sudden climatic changes. In: J.H.R. Gee and P.S. CAller (Editors), Organization of Communities Past and Present. Blackwell, Oxford, pp. 421-438.
Coope, G.R., 1990. The invasion of Northern Europe during the Pleistocene by Mediterranean species of Coleoptera. In:
F. di Castri, A.J. Hansen and M. Debussche (Editors), Biological Invasions in Europe and the Mediterranean Basin. Kluwer, Dordrecht, pp. 203-215.
Coope, G.R., 1994. The response of insects faunas to glacial- interglacial climatic fluctuations. Philos. Trans. R. Soc. London, B 344: 19-26.
Coope, G.R. and Angus, R.B., 1975. An ecological study of a temperate interlude in the middle of the last glaciation, based on fossil Coleoptera from Isleworth, Middlesex. J. Animal Ecol., 44: 365-391.
Coope, G.R. and Sands, C.H.S., 1966. Insect faunas of the last glaciation from the Tame Valley, Warwickshire. Proc. R. Soc. London, B 165: 389-412.
Coope, G.R., Shotton, F.W. and Strachan, I., 1961. A Late Pleistocene fauna and flora from Upton Warren, Worcestershire. Philos. Trans. R. Soc. London, B 244: 379-421.
Comet, C., 1988. Interprttation paltotcologique des diatomtes /t la Grande Pile (France). MOrt. Soc. R. Bot. Belg., 10: 77-86.
Freude, H., 1971. Rhysodidae. In: Freude-Harde-Lohse, Die Irdifer Mitteleuropas. Goecke and Evers, Krefeld, pp. 93-94.
Goeury, C., 1988. Acquisition, gestion et reprtsentation des donntes de l'analyse pollinique sur micro-ordinateur. In: Actes 10~ne Symp. Assoc. Palynol. Langue Fr. Inst. Fr. Pondichtry, Tray. Sect. Sci. Tech., 25: 405-4-416.
G.R.I.P. members, 1993. Climate instability during the last interglacial period recorded in the GRIP core. Nature, 364: 203-207.
Guignot, F., 1947. Faune de France. 48. Coltopt~res Hydrocanthares. Lechevalier, Paris, 287 pp.
Guiot, J., Beaulieu, J.-L. de, Cheddadi, R., David, F., Ponel, P. and Reilte, M., 1993. The climate in Western Europe during the last Glacial/Interglacial cycle derived from pollen and insect remains. Palaeogeogr. Palaeoclimatol. Palaeoecol., 103: 73-93.
Guiot, J., Pons, A., Beaulieu, J.-L. de and Reille, M., 1989. A 140,000-year continental climate reconstruction from two European pollen records. Nature, 338: 309-313.
Guiot, J., Reille, M., Beaulieu, J.-L. de and Pons, A., 1992. Calibration of the climatic signal in a new pollen sequence from La Grande Pile. Clim. Dyn., 6: 259-264.
Hammond, P., Morgan, A. and Morgan, A.V., 1979. On the gibbulus Group of Anotylus, and fossil oeeurences of Anotylus gibbulus (Staphylinidae). Syst. Entomol., 4: 215-221.
Hansen, M., 1987. Fauna Entomologica Scandinavica. 18. The Hydrophiloidea (Coleoptera) of Fennoscandia and Denmark. Brill, Scan. Sci. Press, Leiden, Copenhagen, 254 pp.
Herv~, P., 1951. Apropos de captures de Ceruchus chrysomeli- nus Hoch. darts les Alpes-Maritimes. Conditions ~tholog, iques, climatiques et foresti~res. Entomologiste, 7: 30-35.
Hoffmann, A., 1945. Faune de France. 44. Coltopt~res Bruchides et Anthribides. Libr. Fac. Sci., Paris, 184 pp.
Hoffmann, A., 1954. Faune de France. 59. Col~opt~res Curculionides (2~me partie). Libr. Fac. Sci., Paris, pp. 487-1208.
Hoffmann, A., 1958. Faune de France. 62. Col~opt~res

40 P. Ponel/Palaeogeography, Palaeoclimatology, Palaeoecology 114 (1995) 1-41
Curculionides (36me partie). Libr. Fac. Sci., Paris, pp. 1209-1840.
Johnsen, S.J., Clausen, H.B., Dansgaard, W., Fuhrer, K.. Gundestrup, N., Hammer, C.U., Iversen, P., .Iouzel, J., Stauffer, B. and Steffensen, J.P., 1992. Irregular glacial interstadials recorded in a new Greenland ice core. Nature, 359: 311-313.
Koch, K., 1989. Die Kafer Mitteleuropas, Okologie 1. Goecke and Evers, Krefeld, 440 pp.
Koch, K., 1992. Die Kafer Mitteleuropas, Okologie 3. Goecke and Evers, Krefeld, 389 pp.
Lefkovitch, L.P., 1958. A revision of the European Laemophloeinae (Coleoptera: Cucujidae). Trans. R. Entomol. Soc. London, 111:95 -118.
Lemdahl, G., 1990. Water-surfaces as insect traps and some consequences for palaeoentomology. Quat. Newsl., 62: 26 29.
Leseigueur, L., 1972. Col6opt6res Elateridae de la faune de France continentale et de Corse. Bull. Soc. Linn. Lyon, 382 pp. (suppl.).
Lindroth, C.H., 1985. Fauna Entomologica Scandinavica. 15 (part 1). The Carabidae (Coleoptera) of Fennoscandia and Denmark. Brill, Scand. Sci. Press, Leiden, Copenhagen, 226 pp.
Lindroth, C.H., 1986. Fauna Entomologica Scandinavica. 15 (part 2). The Carabidae (Coleoptera) of Fennoscandia and Denmark. Brill, Scand. Sci. Press, Leiden, Copenhagen, pp. 227 598.
Louis, A. and Smeets, J., 1981. Etude d6taill6e des fluctuations algales comme param6tres des fluctuations thermales dans un refroidissement (M61isey II) de l'Interglaciaire Riss- Wtlrrn. Stud. Algol. Lov., 10, 241 pp.
Louis, A., De Cremer, D. et coll., 1981. Les fluctuations algales param6tres des fluctuations thermales au St Germain II et/i l'Eowfirm. Stud. Algol. Lov., 11,370 pp.
Louis, A. and Ermin, L., 1983. Les fluctuations climatiques au Pr~-E6mien et au d6but de l'E6mien d~duites des fluctuations algales. Stud. Algol. Lov., 12, 126 pp.
Louis, A. and Peters, J., 1983. Les fluctuations climatiques de l'E~mien, du M~lisey I e t du St Germain I d6duites des fluctuations algales fossiles. Stud. Algol. Lov., 13, 270 pp.
Louis, A. et al., 1983. Fluctuations climatiques au N6ow~rm et Pr6bor6al d6duites des fluctuations algales. Stud. Algol. Lov., 15, 190pp.
Lucht, W.H., 1987. Die K~fer Mitteleuropas, Katalog. Goecke and Evers, Krefeld, 342 pp.
Mani, M.S., 1968. Ecology and Biogeography of High Altitude Insects. Junk, The Hague, 527 pp.
Paulian, R., 1959. Faune de France. 63. Col~opt6res Scarab6ides. Libr. Fac. Sci., Paris, 298 pp.
Ponel, P., Beaulieu, J.-L. de and Tobolski, K., 1992. Holocene palaeoenvironment at the timberline in the Taillefer Massif, French Alps: a study of pollen, plant macrofossils and fossils insects. Holocene, 2:117-130.
Ponel, P. and Coope, G.R., 1990. Lateglacial and Early Flandrian Coleoptera from La Taphanel, Massif Central,
France: Climatic and Ecological Implications. J. Quat. Sci., 5:235 249.
Ponel, P., Etlicher, B., Beaulieu, J.-L. de, Debard, E., Thinon, M., Vasari, A. and Petiot, R., 1991. La fin de la derni6re glaciation dans le Cantal (France): la tourbi&e de la Taphanel et son environnement. Quaternaire, 2: 147-163.
Pons, A., Campy, M. and Guiot, J., 1989. The last climatic cycle in France: the diversity of records. Quat. Int., 3/4: 49-55.
Pons, A., Guiot, J., Beaulieu, J.-L. de and Reille, M., 1992. Recent contributions to the climatology of the last glacial interglacial cycle based on french pollen sequences. Quat. Sci. Rev., 11: 439-448.
Reille, M. and Beaulieu, J.-L. de, 1990. Pollen analysis of a long younger Pleistocene continental sequence in a Velay maar (Massif Central, France). Palaeogeogr. Palaeoclimatol. Palaeoecol., 80:35 48.
Sainte-Claire Deville, J., 1914. Catalogue critique des Col~opt6res de la Corse. Caen, 573 pp.
Seret, G., 1967. Les syst6mes glaciaires du bassin de la Moselle et leurs enseignements. Soc. R. Beige G6ogr., 77, 577 pp.
Seret, G., Dricot, E. and Wansard, G., 1990. Evidence for an early glacial maximum in the French Vosges during the last glacial cycle. Nature, 346:453 456.
Seret, G., Guiot, J., Wansard, G., Beaulieu, J.-L. de and Reille, M., 1992. Tentative palaeoclimatic reconstruction linking pollen and sedimentology in La Grande Pile (Vosges, France). Quat. Sci. Rev., 11: 425-430.
Shotton, F.W., Keen, D.H., Coope, G.R., Currant, A.P., Gibbard, P.L., Aalto, M., Peglar, S.M. and Robinson, .I.E., 1993. The Middle Pleistocene deposits of Waverley Wood Pit, Warwickshire, England. J. Quat. Sci., 8: 293-325.
Shotton, F.W. and Osborne, P.J., 1965. The fauna of the Hoxnian Interglacial deposits of Nechells, Birmingham. Philos. Trans. R. Soc. London, B 248: 353-378.
Strand, A., 1946. Nord-Norges Coleoptera. Tromso Mus. Arsh. Naturhist. Avd., 34, 67 ( 1 ), 629 pp.
Taylor, B.J. and Coope, G.R., 1985. Arthropods in the Quaternary of East Anglia--Their role as indices of local palaeoenvironments and regional palaeoclimates. Mod. Geol., 9: 159-185.
Tessier, L., Beaulieu, J.-L. de, Cof~teaux, M., Edouard, J.-L., Ponel, P., Rolando, C., Thinon, M., Thomas, A. and Tobolski, K., 1993. Holocene palaeoenvironments at the timberline in the French Alps--a multidisciplinary approach. Boreas, 22: 244-254.
Theobald, N., Thi6baut, J. and Bernatzky, M., 1974. Carte g6ologique /~ 1/50,000, Giromagny XXXV-20. BRGM, Orleans.
Thomson, R.T., 1958. Coleoptera Phalacridae. In: Handbooks for the Identification of British Insects, V, 5 (b). R. Entomol. Soc. London, London, 17 pp.
Tiberghien, G., 1969. Nouvelles observations sur Rhysodes sulcatus F. [Col. Rhysodidae]. Entomologiste, 25: 85-92.
Woillard, G., 1973. Mise en 6vidence de l'E~mien sur le plateau de Haute-Sa6ne. C. R. Acad. Sci., Paris, 276: 939-942.
Woillard, G., 1974a. Recherches sur le P16istoc+ne dans l'Est

P. Ponel/Palaeogeography, Palaeoclimatology, Palaeoecology 114 (1995) 1-41 41
de la Belgique et dans les Vosges lorraines. Thesis. Univ. Cathol. Louvain, 216 pp.
Woillard, G., 1974b. Expos6 des recherches palynologiques sur le Plgistoc~ne dans l'Est de la Belgique et des Vosges lorraines. Trav. Lab. Palynol. Phytosociol., Univ. Cathol. Louvain, 19 pp.
Woillard, G., 1975. Recherches palynologiques sur le P16istoc.bne dans l'est de la Belgique et dans les Vosges lorraines. Acta Geogr. Lov., 14, 118 pp.
Woillard, G., 1977a. Comparison between the chronology from the beginning of the classical Eemian to the beginning of the classical Wllrm in Grande Pile peat-bog, and other chronolo- gies in the world. In: V. Sibrava (Editor), Quaternary Glaciations in the Northern Hemisphere. Rep., 4, Praha, pp. 72-82.
Woillard, G., 1977b. Grande Pile: a continuous climatic record for the last 140,000 years. In: Abstr. 10th INQUA Congr., Birmingham, 1977, p. 505.
Woillard, G., 1978a. Grande Pile peat-bog: a continuous pollen record for the last 140,000 years. Quat. Res., 9: 1-21.
Woillard, G., 1978b. The last interglacial-glacial cycle at Grande Pile in northeastern France. Trav. Lab. Palynol. Phytosociol., Univ. Louvain, 21 pp.
Woillard, G., 1979. The last interglacial-glacial cycle at Grande Pile in Northeastern France. Bull. Soc. Beige G-6ol., 88: 51-69.
Woillard, G. and Moock, W.G., 1982. Carbon dates at Grande Pile: correlation of land and sea chronologies. Science, 215: 159-161.
Zanetti, A., 1987. Fauna d'Italia. 25. Coleoptera Omaliinae. Calderini, Bologna, 472 pp.




















