
Ethnobotany of the Iroquois with an Emphasis on the Seneca ...
Upload
khangminh22Category
view
0download
0
REVIEW
THE ETHNOBOTANY OF FERNS AND LYCOPHYTES
H. A. KELLER1 & G. T. PRANCE2
1Consejo Nacional de Investigaciones Científicas y Tecnológicas, Instituto de Botánicadel Nordeste, Sargento Cabral 2131, Corrientes, Argentina.
2Royal Botanic Gardens, Kew, Richmond, Surrey, TW9 3AB, UK, correspondingauthor: [email protected]
Keywords: Fern ethnobotany, ethnopteridology, archaeobotany, medicinal plants,ornamental plants
ABSTRACTA summary is presented of the most important ways in which ferns have beenimportant to humanity. Many of these categories are positive such as the use offerns for subsistence. On the negative side is their role as weeds and as bearers ofsubstances harmful to human health. Many of the traditional uses such as formedicines have been transferred to modern life as societies have modernized. Someuses have even become important in industrial society, for example in the assay ofnew medicines.
INTRODUCTIONFerns are distributed in all climate zones of the planet, but have a greater diversity in thetropics (Smith et al., 2006; Strasburger et al., 2003). The discipline that studies therelationship between the uses of ferns and humans has been termed ethnopteridology, andit was well explained and amplified by Boom (1985). These reciprocal interactions may ormay not be related to a particular use category in family or regional cultures, but the conceptof ethnobotany goes beyond the utilitarian spheres of economics and uses to includesymbolic values, nomenclature, religion and also the place that particular plants occupy inthe cosmology of peoples. Strictly economic aspects of uses are addressed by the disciplineof economic botany. It is not easy to separate economic botany and ethnobotany and indeedeconomic botany implies a type of relationship between human groups and plants.
The object of this work is to present a global panorama of the state of the art inethnobotany of pteridophytes. We will show the great diversity of relationships betweenthese plants and humans. Pteridophytes feature in some way in many papers aboutethnobotany and economic botany and here we will concentrate mainly on papers whoseobject was specifically about the study of ferns or lycophytes. A few previous papers haveaddressed similar topics at least locally. For example, May (1978) made a summary of theeconomic and folkloric uses of ferns. Díaz de León et al. (2007) reviewed the many usesof ferns and lycophytes in Mexico and to some extent the rest of the world. Murillo (1983)produced a major work on the uses of ferns in South America with a special emphasis onthe ferns of Colombia. Mannar Mannan et al. (2008) gave a short review of the potentialuses of ferns. We have used the names as cited in the various papers rather than trying toupdate the nomenclature in any way. The literature about the reciprocal relationshipsbetween humans and pteridophytes is so extensive that here we cannot possibly cover itall, but we hope to show the variety of possible relationships through presenting a numberof wide-ranging examples.
FERN GAZ. 20(1):1-13. 2015 1

ARCHAEOBOTANYArchaeobotany is the study of ancient plant remains found in archaeological contexts.The name is obviously a parallel to, and a derivative from, ethnobotany. In botharchaeobotany and ethnobotany the focus of attention is upon the uses of plants by, andtheir association with people (Wiley, 1995). Since remote times human populations inmany parts of the world knew and made use of ferns and lycophytes. The archaeologicalliterature offers much evidence of the close relationship between humans and these plantsover many generations. Anderson & White (2001) suggested that Cyathea was used forconsumption by humans at least 888 years ago in Norfolk Island. Dental evidence fromhuman remains in New Zealand show ancient use of the rhizomes of Pteridiumesculentum (G.Forst.) Cockayne (Houghton, 1978). In China there was use of edibleferns at least 3000 years ago (Zhang, 2007). There is evidence of the prehistoric use ofthe leaves of Marattia fraxinea Sm. in ritual involving the incipient use of iron in Africa(Schmidt & Avery, 1983).
FERNS IN PRE-LINNEAN LITERATUREOur study has shown that fern ethnobotany is nothing new and that there is much aboutferns and local cultures in ancient literature. We have mainly concentrated here on themore recent literature, but a good example of early studies of fern ethnobotany is that ofGeorgius Everhardus Rumphius (1627-1702) in Ambon, now in Indonesia (Rumphius,2011). Rumphius devoted 44 pages to descriptions of ferns, lycophytes and their uses.The pre-Linnean names are often hard to identify to species, but the good drawings andthe interpretation by E. D. Merrill (1917) make it possible to relate to current speciesnames. These pages are full of ethnobotanical information about fern uses by theAmbonese and natives of other islands in the seventeenth century. Many of these usescome under the subheadings we have used below. For example one chapter is entitled“The Edible Fern” and is about Athyrium esculentum (Retz.) Copel. Rumphius said thatthis fern and related species “are a renowned potherb of all of these islanders. One canmake a good salad from its leaves and shoots….it cools moderately and loosens thebowels, especially if one drinks some tree-wine after it.” Rumphius describes the culinaryuse of several different species of ferns together with great details about the effects ofeating them. The Balinese stick the tops of Tectaria crenata Cav. behind their ears whenthey go to war, because this plant will keep them from getting hurt by dart poison,rendering it powerless. They also rub the dry leaves over their bodies when bathing toget rid of sweat and odours. The lower stems of Lygodium circinnatum (Burm. f.) Sw.are split into four strips and used for seams around the edges of baskets by the Ambonese.An interesting use of the leaves of Drynaria sparsisora (Desv.) Moore is to tie them toa baited fishhook and use them as sails to carry the hooks out to sea until fish bites. Theleaves of the same species are suspended over little children to keep them safe from evilspirits. The pages of Rumphius contain much interesting fern ethnobotany and manysimilar uses are reported in the more recent literature on the subject.
ETHNOPTERIDOLOGYIn the literature surveyed we found a number of articles that were specifically aboutethnopteridology of human groups and also ethnobotany about individual species ofpteridophytes, and we highlight examples of recent works from different continents.From the Americas, Navarrete et al. (2006) presented information on the uses attributedto more than 200 species of pteridophytes of Ecuador, Peru and Bolivia. Boom (1985)
2 FERN GAZ. 20(1):1-13. 2015

treated the use of ferns by the Chácobo tribe of Amazonian Bolivia and Macía (2004)compared the ethnopteridology of the Tacana of Bolivia with that of the Huaorani ofEcuador. The only species in common was Cyathea pungens (Willd.) Domin. HernándezCibrián & Sutherland (2007) carried out an ethnobotanical study of the ferns of a nationalpark in Honduras and found only eight species that were used by the local population.For Argentina Keller et al. (2011) treated the various uses of 50 species of ferns andlycophytes by the Guaranies of Misiones Province (Figures 1-3) and Hurrell & De LaSota (1996) did the same for the villagers of Santa Victoria in Salta Province. In AsiaChristensen (1997) studied the ethnopteridology of ethnic groups in Malaysia. A studyof the ethnobotanical uses of ferns in the Indian States of Jammu and Kashmir (Kirn &Kapahi, 2001) listed 17 species of which 11 were medicinal, four used for thatchingroofs and three as foods. Joshi (1997) listed ethnobotanical uses of 44 species of fernsin Uttar Pradesh State of India.
For Africa the ethnopteridological study of ethnic groups by Nwosu (2002)mentioned 36 species in 23 families. In addition to fern uses for food and medicines,many of these studies mention uses in rituals of love, for magic ceremonies, as indicatorsof cardinal points and the presence of animals and also as material to make crafts andweapons. The demands of today’s markets have led various indigenous groups tocommercialize ornamental ferns and flowerpots made out of erect rhizomes and thetrunks of tree ferns.
FERNS IN TRADITIONAL MEDICINEStudies of uses of ferns in ethnomedicine are abundant on all inhabited continents. InCórdoba (Argentina) the study of ferns used in traditional medicine has developed to
KELLER & PRANCE: THE ETHNOBOTANY OF FERNS AND LYCOPHYTES 3
Figure 1. Microgramma squamulosa (Kaulf.) de la Sota, a multipurpose species for theGuarani, used for slimming, menstrual analgesic, post partum washing, and treatmentof lumbago.

such an extent that anatomical and morphological evidence is used to detect adulterantsof the products (Luján et al., 2007; 2011). However, the adulteration of medicinal fernsis nothing new. Hipólito Ruiz (1805) described the species Polypodium calguala Ruizwith the intention of clearly differentiating this medicinal ethnospecies of the indigenouspeoples of Peru from other fern species that were being imported into Spain from theNew World as adulterants of the legitimate “calaguala.”
In a comparison between the ethnopteridology of the Tacana of Bolivia and theHuaorani of Peru, Macía (2004) found that 76% of the recorded uses for ferns weremedicinal either for people or for animals to heal wounds or expel parasites. Most of theuses by the Tacana are external, whereas the Huaorani uses are mainly internal. Thisstudy cites uses for 24 species of ferns and lycophytes. There are many medicinal usesof ferns in India. For example, Sharma and Vyas (1985) described the use of six speciesin Rajastan. Srivastava (2007) emphasised the importance of ferns in tribal medicinefrom a study made in various places throughout India. Benniamin (2011) reported onthe use of 51 species of ferns in the east of India and Kumari et al. (2011) gaveinformation about the use of 66 species in ethnomedicine in India. Dixit (1982) is anexample of the ethnobotanical use of a single species, Selaginella bryopteris (L.) Bak.This species is much revered in local medicine and commands a high market price. Themedicinal use of Helminthostachys zeylanica (L.) Hook. has had serious effects on itsstate of conservation in Himalaya (Joshi, 2011). Osmunda regalis L. is used as a medicinein the north of Spain (Molina et al., 2009).
Of the 36 species of ferns reported in the ethnobotany study of ferns of SouthernNigeria by Nwosu (2002), 34 have medicinal uses. The paper reads like a complete
4 FERN GAZ. 20(1):1-13. 2015
Figure 2. Alsophylla setosa Kaulf. The base of the rhizomes are used as stands forornamental plants. The Guaranis and local farmers agree that the presence of this fernis an indicator that the soil is not suitable for agriculture. The Guarani use a soup of thepetioles for the treatment of herpes.

pharmacy to treat many different ailments and all from pteridophytes. Ferns supplytreatment for external injuries and wounds and many are taken internally to treat suchdiseases as malaria, ulcers, intestinal worms, liver disease etc. Many of the species listedhave multiple uses, for example, a decoction of the rhizome and leaves of Polypodiummicrorhizoma Clarke ex Bak. is used for the relief of gastrointestinal disorders, backacheand jaundice; a paste from the dried leaves (dried over an open fire) is applied externallyfor fissures on hands and wound healing; and a paste mixed with palm-kernel oil isapplied externally to domestic animals such as sheep and cattle. The whole plant ofOsmunda regalis is taken internally for psychosis as it is believed that the tonic can chaseaway evil spirits, and an infusion of the roots is used to treat malaria and jaundice.
The importance of medicinal ferns is evidenced by the growing interest in methodsfor their reproduction. An example is the achievement of in vitro propagation of theAsiatic fern Drynaria quercifolia (L.) J.Sm. that is much used in traditional phytotherapy(Mazumder et al., 2011).
VETERINARYThe literature has many examples of the use of ferns for treating animals. According toNwosu (1922), the leaves of Tectaria macrodonta (Fée) C.Chr. are powdered and mixedwith castor oil and given to goats and sheep to stop a running stomach; young fronds arechewed by cows after delivery of a calf to accelerate the expulsion of the afterbirth.
EDIBLE FERNSFern rhizomes were an important source of food for Native Americans in western North
KELLER & PRANCE: THE ETHNOBOTANY OF FERNS AND LYCOPHYTES 5
Figure 3. The rhizome of Dicksonia sellowiana Hook. in a Guarani basket offered forsale at a roadside stand.

America. Turner et al. (1992) produced a summary of this in a detailed paper that listedat least 15 species of ferns together with their native nomenclature. Ferns are much eatenin India. For example, Pandy and Pangtey (1987) list seven different species of fernsconsumed in Western Himalaya and Joshi (1997) lists 10 edible species used in UttarPradesh State. For China, Liu et al. (2012) listed a total of 42 edible pteridophytes, butthey estimated that the potential total could be as high as 144 species. Some ferns areeaten as though they were sweets, as in the case of Pecluma pectinatiformis (Lindm.)M.G.Price, where the sweet leaves are commonly chewed by Guaraní children inMisiones, Argentina (Keller et al., 2011).
Some species of Polypodium are known for their property of sweetness. A variety ofPolypodium vulgare L. was used to flavour tobacco for its liquorice taste and it containssmall amounts of ostadin, a steroid saponin 3000 times as sweet as sucrose. In formertimes the fronds of this species were used in Ireland to treat coughs, colds and asthma.Polypodium glycyrrhiza D.Eaton also has a liquorice flavour and was eaten by NativeAmerican peoples (Mabberley, 2008).
FERNS AS BUILDING MATERIALSThe Guaranies of southeastern Brazil use the stems of Dicksonia sellowiana Hook. tosupport the walls of their traditional houses (Prudente, 2007).
Joshi (1997) lists Cyathea spinulosaWall. ex Hook. and Dicranopteris liniaris (Burm.f.) Underw. as used for thatching roofs in India. Kirn and Kapahi (2001) mentionPteridium aquilinum (L.) Kuhn. var. wightianum (Ag.) Tryon, Pteris vittata L.,Thelypteris erubescens (Wall. ex Hook.) Ching and Woodwardia unigemmata (Makino)Nakai for the same purpose.
FERNS IN ORNAMENTATION AND ARTFerns are often used for body ornamentation. The stipes of Cheilanthes farinosa (L.)Brogn. and Adiantum lunulatum Burm. are used as nose and ear studs by children andpoor women in Uttar Pradesh (Pande & Pangey, 1987) and Adiantum venustum D. Donis used as ear studs by girls in Kashmir (Kirn & Kapahi, 2001). The tree ferns Cyatheadivergens var. tuerckenheimii R.M.Tryon and C. fulva (Martens & Galeotti) Fée areharvested to produce handicrafts for garden ornamentation by artisans of the mountainsof Cuetzalan, Mexico (Elutério, 2006). In the Philippines the petiole and leaf rachis ofLygodium japonicum (Thunb.) Sw. are used to decorate baskets (Novellino, 2006). InArgentina the Guaranies use the petioles of various ferns to make necklaces (Keller etal., 2011).
USES OF FERN SPORESFrye (1934) reported that the spores of Lycopodium clavatum L. were used for dustingon open raw wounds and chafed infants by natives of northwestern North America. Thespores are fine and light and so repel water and prevent stickiness. Lycopodium powderhas also been used as a lubricating dust on latex gloves and condoms, though the latteruse is not recommended (see Balick & Beitel, 1989), because these spores have beenknown to cause allergic reactions, ranging from hay-fever to more serious giant cellgranulomas. May (1978) reported that the easily flammable spores of species ofLycopodium have been used in theatre as a flash powder.
6 FERN GAZ. 20(1):1-13. 2015

FERNS AS PLACE NAMESThe importance of ferns to local communities has often led to the names of places,topographic formations, watercourses and political divisions. In Misiones Province(Argentina), the most tropical and wettest province of Argentina, there are various placesbased on plant names. The ‘Diccionario geográfico toponímico’ of Stefañuk (2009) givesseveral examples. “Los helechos” is the name of a stream and a municipality in thisprovince. In the town of Oberá there is a place and a stream named “Samambaya” thecommon name for ferns derived from the Guaraní word for them “amambái” whichtranslated into English is a generic name of ferns. Locally amambái refers to the largepopulations of Pteridium arachnoideum (Kaulf.) Maxon. In the Department of L.N. Alemthere is a stream called “Chachi” which is the Guaraní name for the tree fern Cyatheaatrovirens (Langsd. & Fisch.) Domin (Cyatheaceae). In the Department of San Martínthere is stream called “Culandrillo”, a term that refers to species of the genus Adiantum(Pteridaceae). In Paraguay the derivation of the name of the political division AmambayDepartment is derived from the Guaraní word for fern. There are many place names inthe United Kingdom associated with ferns, for example, Ferndown in Dorset, Fernileein Derbyshire and Ferness in the Highlands. Fern is a town in Tayside and Fernie a streamand a castle in Fife. Bracken (Pteridium aquilinum) features in Brackenfield inDerbyshire and Brackenthwaite in Cumbria.
FERNS IN PHARMACOLOGYSubstances with antioxidant activity are now used in medicine to reduce the effects ofoxidation stress. Antioxidant activity has been reported in Adiantum capillus-veneris L.,a widely distributed fern (Kumar, 2009). A recent study of the lateral branches ofEquisetum giganteum L. of South and Central America showed that they can be used asa source of antioxidant compounds (Ricco et al., 2011). In Malaysia studies have shownsimilar properties in various ferns: Blechnum orientale L., Cibotium barometz (L.) J.Sm.,Cyathea latebrosa (Wall. ex Hook.) Copel., Dicranopteris linearis Burm., Drynariaquercifolia (L.) J.Sm. and Stenochlaena palustris (Burm. f.) Bedd. (Chai et al., 2012;Lai & Lim, 2011, Lai et. al., 2010). In China several fern rhizomes have been shown tohave antioxidant properties: Drynaria fortunei (Kze.) J.Sm., Pseudodrynaria coronans(Wall. ex Mett.) Ching, Davallia divaricata Bl., D. mariesii Moore ex Bak., D. solida(Forst.) Sw., and Humata griffithiana (Hook.) C.Chr. (Chang et al., 2007).
Ferns also contain substances with antibacterial activity as was shown in the studyby Thomas (2011) of Osmunda regalis. India has made important advances in this areaand Kumarpal (2013) showed that there was good antimicrobial activity in three speciesof ferns from three different families used in traditional medicines in Darjeeling,Athyrium filix-femina (L.) Roth (Woodsiaceae), Dicranopteris linearis (Burm. f.)Underw. (Gleicheniaceae) and Pleopeltis macromarpa (Bory ex Willd.) Kaulf.(Polypodiaceae). Patric Raja et al. (2012) found antibacterial and antifungal activity inCyathea nilgiriensis Holttum, C. crinita (Hook.) Copel., Leptochilus lanceolatus Féeand in Osmunda hugeliana Presl. Studies made in Romania by Soare et al. (2012) showedthat the bladder fern Cystopteris fragilis (L.) Bernh. and Polypodium vulgare L. stronglyinhibited various bacteria, especially Escherichia coli. Most of the species that have beenstudied for their pharmaceutical properties have notable antecedents in traditionalmedicine. The development of pharmaceuticals from plants used in the pharmacopeiaof local peoples ought to ensure that the benefits are shared with the traditionalcommunities from where the original information came (Prance, 1991).
KELLER & PRANCE: THE ETHNOBOTANY OF FERNS AND LYCOPHYTES 7

FERNS IN TOXICOLOGYThe relationship between plants and people is not always a positive one This includesthose species that have strong substances that are damaging either to human health or tothat of domesticated animals. Pteridium aquilinum has been studied for the relationshipbetween the various uses and its toxicity (Alonso Amelot, 1999; Franca et al., 2002;Ortega, 1993; Vetter, 2011). Recent studies show that Blechnum orientale is able toabsorb heavy metals from the environment and it is therefore a potentially toxic sourceof food and medicine. The accumulation is mostly in the rhizomes and leaves, the veryparts of the plant that are eaten (Zhu et al., 2013). A positive side for the environment ofthis absorptive capacity of ferns is the accumulation of arsenic by Pteris vittata L., whichcould be used as a method of decontaminating toxic sites. Phytoremediation of arseniccontaminated environments will involve growing the arsenic hyperaccumulator ferns inthe contaminated environment, harvesting the arsenic-rich biomass and the safe disposalof the biomass (Gumaelius, 2004; Rathinasabapathi et al., 2006).
FERNS AS ORNAMENTAL PLANTSA large number of fern species are used as ornamentals in the gardens around the world,and this dates back to ancient times and often to domestication for other uses. Studies ofthe ornamental potential of pteridophytes of regional floras has led to an increase of theiruse as cultivated plants. Abraham et al. (2012) listed a total of 153 ferns and 18lycophytes from Nilgiris, India with ornamental potential. Macaya (2004) mentioned 20species of ferns native to Chile that are cultivated as ornamentals and in Macaya (2008)he expanded this to a list of 75 species. An example of the relevance of ethnobotany formaking strategies for use of populations of native vegetation is that of Baldauf et al.(2007), who made an ethnobotanical study of the management systems of Rumohraadiantiformis (G. Forst.) Ching. This species is used as an ornamental in southern Brazil.Another fern of the southern cone region of South America with high ornamentalpotential is Blechnum tabulare (Thunb.) Kuhn, a species that resembles the Cycadaceaein appearance. A population study of this species was made in southern Brazil(Rechenmacher et al., 2007). The stems of some tree ferns are used extensively as asubstrate for the cultivation of epiphytes. However, the indiscriminate use of this resourceis threatening populations of these plants in some places. An example of this is the useof Dicksonia sellowiana Hook., the conservation of which has led to studies of growth,phenology and germination of spores (Filippini et al., 1999; Schmitt et al., 2009).
FERNS AND SOIL QUALITYAzolla Lam. has long been used as a fertilizer in rice paddies (Jones, 1987). In Misiones,Argentina farmers identify compacted and degraded soils by means of observations onthe presence of Pteridium aquilinum. In the central Andean region of Peru indigenouspeoples of the Quechua language group use the fronds of Dennstaedtia glauca (Cav.)Looser to fertilize the soils where they cultivate potatoes (Camino & Jhons, 1988), whichis similar to the use of bracken peat as a fertilizer in some places in the United Kingdom.
CONCLUSIONSThis brief summary has shown the diversity of important uses to which humans haveput ferns and lycophytes, and some of these are illustrated in Figures 1-3. Many of theseuse categories originated in traditional societies (the white area of Figure 4) such as usein rituals, the construction of houses and to make weapons. Other uses are typical of
8 FERN GAZ. 20(1):1-13. 2015
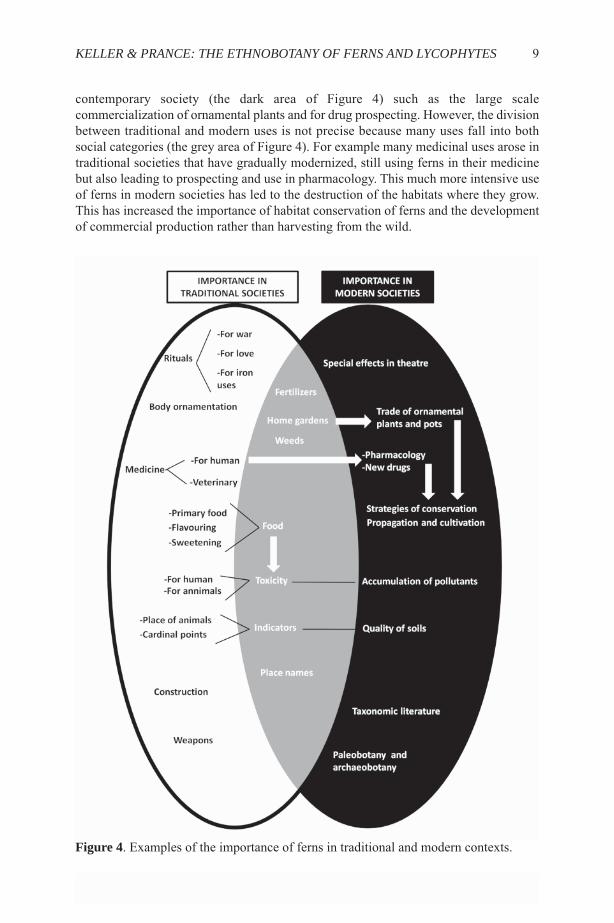
contemporary society (the dark area of Figure 4) such as the large scalecommercialization of ornamental plants and for drug prospecting. However, the divisionbetween traditional and modern uses is not precise because many uses fall into bothsocial categories (the grey area of Figure 4). For example many medicinal uses arose intraditional societies that have gradually modernized, still using ferns in their medicinebut also leading to prospecting and use in pharmacology. This much more intensive useof ferns in modern societies has led to the destruction of the habitats where they grow.This has increased the importance of habitat conservation of ferns and the developmentof commercial production rather than harvesting from the wild.
KELLER & PRANCE: THE ETHNOBOTANY OF FERNS AND LYCOPHYTES 9
Figure 4. Examples of the importance of ferns in traditional and modern contexts.

ACKNOWLEDGMENTSWe thank Blanca León for helpful suggestions when reviewing the manuscript.
REFERENCESABRAHAM, S., RAMACHANDRAN, V.S. & SOFIA, C. 2012. Potential ornamental
Ferns from Nilgiris, Tamil Nadu. Advances in Applied Science Research 3 (4):2388-239.
ALONSO AMELOT, M. E. 1999. Helecho macho, salud animal y salud humana.Rev.Fac. Agron. (LUZ) 16: 528-541.
ANDERSON, A. & WHITE, P. 2001. Approaching the Prehistory of Norfolk Island.Records of the Australian Museum, Supplement 27: 1–9.
BALDAUF, C., HANAZAKI, N. & SEDREZ DOS REIS, M. 2007. Caracterizaçãoetnobotânica dos sistemas de manejo de samambaia-preta (Rumohra adiantiformis(G. Forst) Ching - Dryopteridaceae) utilizados no sul do Brasil. Acta bot. bras. 21(4): 823-834.
BALICK, M. J. & BEITEL, J. M. 1989. Lycopodium spores used in condommanufacture: associated health hazards. Economic Botany 43: 373-377.
BENNIAMIN, A. 2011. Medicinal ferns of North Eastern India with special referenceto Arunachal Pradesh. Indian Journal of Traditional Knowledge 10(3): 516-522.
BOOM, B. M. 1985. Ethnopteridology of the Chácobo Indians in Amazonian Bolivia.Amer. Fern J. 75:19–21.
CAMINO, A. & JOHNS. T. 1988. Laki-Laki (Dennstaedtia glauca, Polypodiaceae): A Green Manure Used in Traditional Andean Agriculture. Economic Botany42(1):45-53.
CHRISTENSEN, H. 1997. Uses of ferns in two indigenous communities in Sarawak,Malaysia. In JOHNS, R. J. (Ed.) Holttum Memorial Volume, pp. 177-192. RoyalBotanic Gardens, Kew.
CHAI, T.-T., PANIRCHELLVUM, E., ONG, H.-C. & WONG, F.-C. 2012. Phenoliccontents and antioxidant properties of Stenochlaena palustris, an edible medicinalfern. Botanical Studies 53: 439-446.
CHANG, H.C., HUANG, G.J., AGRAWAL, D.C., KUO, C.L., WU, C.R. & TSAY, H.S.2007. Antioxidant activities and polyphenol contents of six folk medicinal ferns usedas “Gusuibu”. Bot. Stud. 48: 397-406.
CLINE, W. 1937. Mining and Metallurgy in Negro Africa. George Banta Pub. Co.: Menasha, Wisconsin.
DÍAZ DE LEÓN, M. E. M., MENDOZA-RUÍZ, A. & PÉREZ-GARCÍA, B. 2007. Usosde los helechos y plantas afines. Etnobiología 5: 117-125.
DIXIT, R.D. 1982. Selaginella bryopteris (L.) Bak. – An ethnobotanical study IV. J.Econ. Tax. Bot. 3: 309-312.
ELUTÉRIO, A.A. 2006. Management of tree fern (Cyathea spp.) for handicraftproduction in Cuetzalan, Mexico. Economic Botany 60: 182-191.
FILIPPINI, E.C.P., DUZ, S.R. & RANDI, Á.M. 1999. Light and storage on thegermination of spores of Dicksonia sellowiana (Presl) Hook., Dicksoniaceae. Revta.Bras. Bot. 22: 21-26.
FRANCA, T. N., TOKARNIA, C. H. & PEIXOTO, P. V. 2002. Enfermidadesdeterminadas pelo princípio radiomimético de Pteridium aquilinum (Polypodiaceae).Pesq. Vet. Brass. 22(3): 85-96.
FRYE, T. C. 1934. Ferns of the Northwest. Metropolitan Press, Portland.GUMAELIUS, L., LAHNER, B., SALT, D. E. & BANKS, J. A. 2004. Arsenic
10 FERN GAZ. 20(1):1-13. 2015

hyperaccumulation in gametophytes of Pteris vittata. A new model system foranalysis of arsenic hyperaccumulation. Plant Physiol. 136:3198–3208.
HERNÁNDEZ CIBRIÁN, R. K. & SUTHERLAND, C. H. N. 2007. Etnobotánica delos Helechos de Honduras. Ceiba 48(1-2): 1-10.
HOUGHTON, P. 1978. Dental evidence for dietary variation in prehistoric New Zealand.Journal of the Polynesian Society 87(3): 257-263.
HURREL, J. A. & DE LA SOTA, E. R. 1996. Etnobotánica de las Pteridofitas de losPastizales de altura de Santa Victoria (Salta, Argentina). Revista Mus La Plata (NS).Botánica 15(105): 353–364.
JONES, D. L. 1987. Encyclopaedia of Ferns. An Introduction to ferns. Their structure,biology, importance, cultivation and propagation. Lothian Publishing Company PTYLTD. Melbourne, Sydney, Auckland.
JOSHI, P. 1997. Ethnobotany of Pteridophytes of hilly districts of Uttar Pradesh, India.Indian Fern J. 14: 14-18.
JOSHI, P. 2011. Ecology and Medicinal Uses of Helminthostachys zeylanica (L.) Hook.“An endangered flora of India” reported at Foothills of Kumaun Himalaya(Kashipur), Uttarakhand. Researcher 3(4): 51-54.
KELLER, H.A., MEZA-TORRES, E.I. & PRANCE, G.T. 2011. Ethnopteridology of theGuaranis of Misiones Province, Argentina. Amer. Fern J. 101(3): 193-204.
KIRN, H.S. & KAPAHI, B.K. 2001. Ethnobotanical notes on some ferns and fern-alliesof Jammu and Kashmir State, India. Indian Fern J. 18: 35-38.
KUMAR A. 2009. Antioxidant effect of Adiantum capillus veneris Linn. on humanlymphocyte: an in vitro study. Journal of Cell and Tissue Research 9 (2): 1899-1902.
KUMARI, P., OTAGHVARI, A. M., GOVINDAPYARI, H., BAHUGUNA, Y. M. &UNIYAL, P. L. 2011. Some ethno-medicinally important Pteridophytes of India. Int.J. Med. Arom. Plants 1(1): 18-22.
KUMARPAL, S. 2013. Study of Activity of Some Medicinal Ferns of Darjeeling.International Journal of Scientific and Research Publications 3(8): 1-4.
LAI, H.Y. & LIM, Y.Y. 2011. Evaluation of Antioxidant Activities of the MethanolicExtracts of Selected Ferns in Malaysia. International Journal of EnvironmentalScience and Development 2(6): 442-447.
LAI, H.Y., LIM, Y. Y. & KIM, K.H. 2010. Blechnum Orientale Linn - a fern withpotential as antioxidant, anticancer and antibacterial agent. BMC Complementaryand. Alternative Medicine, vol.10: 15.
LIU, Y., WUJISGULENG, W. & LONG, C. 2012. Food uses of ferns in China: a review.Acta Soc. Bot. Poloniae 81: 263-270. (52 species used)
LOOSER, G. & RODRÍGUEZ, R. 2004. Los helechos medicinales de Chile y susnombres vulgares. Gayana Bot. 61(1): 1–5.
LUJÁN, M. C., MORERO, R., BONZANI, N. E. & BARBOZA, G. E. 2007. Sobre laidentidad de algunos helechos medicinales que se comercializan en Córdoba(Argentina). Boletín Latinoamericano y del Caribe de Plantas Medicinales yAromáticas 6(6): 376-377.
LUJÁN, M. C., MORERO, R. & BARBOZA, G. E. 2011. Estudios epidérmicos enhelechos y licófitas medicinales de la Provincia de Córdoba, Argentina. Hoehnea38(4): 609-659, 2 tab., 28 fig.
MABBERLEY, D.J. 2008. Mabberley’s Plant-book, ed. 3: 690. Cambridge Univ. PressMACAYA, J. 2004. Helechos nativos de Chile cultivados con fines ornamentales. Chloris
Chilensis 7(1) URL: http://www.chlorischile.cl.MACAYA, J. 2008. Helechos introducidos en Chile con fines ornamentales (parte I).
Chloris Chilensis Año 11, N°2. URL: www.chlorischile.cl.
KELLER & PRANCE: THE ETHNOBOTANY OF FERNS AND LYCOPHYTES 11

MACÍA, M.J. 2004. A comparison of useful Pteridophytes between two AmerindianGroups from amazonian Bolivia and Ecuador. Amer. Fern J. 94: 39-46.
MANNAR MANNAN, M., MARIDASS, M. & VICTOR, B. 2008. A Review on thePotential Uses of Ferns. Ethnobotanical Leaflets 12: 281-285.
MAY, L. W. 1978. The economic uses as associated folklore of ferns and fern allies. Bot. Rev. 44: 491-528.
MAZUMDER, P. B., MAZUMDER, B., CHOUDHURY, M D. & SHARMA, G.D. 2011.In Vitro Propagation of Drynaria quercifolia (L.) J. Sm., a Medicinal Fern. AssamUniversity Journal of Science & Technology, Biological and Environmental Sciences7(1): 79-83.
MERRILL, E. D. 1917. An interpretation of Rumphius’s “Herbarium Amboinense”.Manila Bureau of Printing.
MOLINA, M., REYES-GARCIA V. & PARDO-DE-SANTAYANA, M. 2009. Localknowledge and management of the Royal Fern (Osmunda regalis L.) in NorthernSpain: Implications for Biodiversity Conservation. Amer. Fern J. 99 (1): 45–55.
MURILLO, M.T. 1983. Usos de los helechos en Suramérica con especial referencia aColombia. Ed. Instituto de Ciencias Naturales, Biblioteca José Jerónimo Triana, Nº5,156 pp. Bogotá.
NAVARRETE, H., LEÓN, B., GONZALES, J., AVILES, D. K., SALAZAR LECARO,J., MELLADO, F., ALBAN, J. & ØLLGAARD, B. 2006. Helechos. In: MORAESR., M., ØLLGAARD, B., KVIST, L. P., BORCHSENIUS, F. & BALSLEV, H. (Eds.):Botánica Económica de los Andes Centrales. Universidad Mayor de San Andrés, La Paz, p: 385-411.
NOVELLINO, D. 2006. An introduction to basketry in Island Southeast Asia.Proceedings of the IVth International Congress of Ethnobotany (ICEB 2005), 2006:621-625.
NWOSU, M. O. 2002. Ethnobotanical studies on some Pteridophytes of southern Nigeria.Econ. Bot. 56: 255–259.
ORTEGA, F. 1993. La etnobotánica de Pteridium aquilinum (L.) Kuhn en Venezuela ysus posibles riesgos asociados a la carcinogénesis. Medula 2 (3-4): 51-56.
PANDE, P.C. & PANGTEY, Y.P.S. 1987. Studies on ethnobotany –I. On some less knownedible and economic ferns of Kumaun region of Western Himalaya. J. Econ. Tax.Bot. 11: 81-85.
PATRIC RAJA, D., JOHNSON, M., IRUDAYARAJ, V. & JANAKIRAMAN, N. 2012.Antimicrobial efficacy of selected ferns of Western Ghats, South India. InternationalJournal of Current Pharmaceutical Research 4 (2): 58-60.
PRANCE, G. T. 1991. What is ethnobotany today? J. Ethno-pharmacol. 32: 209-216.PRUDENTE, L. T. 2007. Arquitetura Mbyá-Guaraní na Mata Atlântica do Rio Grande
do Sul. Estudo do caso do Tekoá Nhüu Porã. Tese (Mestrado) em engenharia.Universidades Federal do Rio Grande do Sul. Escola de Engenharia. Porto Alegre,164 pp.
RATHINASABAPATHI, B., MA, L. Q. & SRIVASTAVA, M. 2006. ArsenicHyperaccumulating Ferns and their Application to Phytoremediation of ArsenicContaminated Sites. Floriculture, Ornamental and Plant Biotechnology 3: 304-311.
RECHENMACHER, C., SCHMITT, J. L. & BUDKE, J. C. 2007. Estrutura e distribuiçãoespacial de uma população de Blechnum tabulare (Thunb.) Kuhn (Pteridophyta,Blechnaceae) em um mosaico floresta-campo no sul do Brasil. Pesquisas, Botânica58: 177-186.
RICCO, R. A., AGUDELO, I., GARCÉS, M., EVELSON, P., WAGNER, M. L. &GURNI, A. A. 2011. Polifenoles y actividad antioxidante en Equisetum giganteum
12 FERN GAZ. 20(1):1-13. 2015

L. (Equisetaceae). Boletín Latinoamericano y del Caribe de Plantas Medicinales yAromáticas 10 (4): 325 – 332.
RUIZ, H. 1805. Memoria sobre la legítima calaguala y otras dos raíces que nos vienende la América meridional. Imprenta de D. José del Collado, Madrid, 60 pp.
RUMPHIUS, G. E. 2011. The Ambonese Herbal vol 5: 121-165. English translation byE.M. BEEKMAN of The Ambonese herbal first published in 1741, Yale UniversityPress.
SHARMA, B.D & VYAS, M.S. 1985. Ethnobotanical studies on the ferns and fern alliesof Rajastan. Bull. Bot. Survey India 27: 1-4.
SCHMITT, J. L., SCHNEIDER, P. H. & WINDISCH, P. G. 2009. Crescimento docáudice e fenologia de Dicksonia sellowianaHook. (Dicksoniaceae) no sul do Brasil.Acta bot. bras. 23(1): 282-291.
SCHMIDT, P. R. & AVERY, D. H. 1983. More Evidence for an Advanced PrehistoricIron Technology in Africa. Journal of Field Archaeology 10(4): 421-434.
SMITH, A. R., PRYER, K. M., SCHUETTPELZ, E., KORALL, P., SCHNEIDER, H.& WOLF, P. G. 2006. A classification for extant ferns. Taxon 55(3): 705-731.
SOARE, L. C., FERDEŞ, M., DELIU, I. & GIBEA, A. 2012. Studies regarding theantibacterial activity of some extracts of native pteridophytes. U.P.B. Sci. Bull., SeriesB 74(1): 21-26.
SRIVASTAVA, K. 2007. Importance of Ferns in Human Medicine. EthnobotanicalLeaflets 11: 231-234.
STEFAÑUK, M. Á. 2009. Diccionario geográfico toponímico de Misiones.Contratiempo Ediciones. Buenos Aires, 817 pp.
STRASBURGER, E., NOLL, F., SCHENCK, H. & SCHIMPER, A. F. W. 2003. Tratadode Botánica, 35ª edición, actualizada por SITTE, P., WEILER, E. W., KADEREIT,J. W., BRESINSKY, A. & KÖRNER, C. Ediciones Omega, Barcelona, 1134 pp.
THOMAS, T. 2011. Preliminary Antibacterial and Phytochemical Assessment ofOsmunda regalis L. International Journal of Pharmaceutical & Biological Archives;2(1): 559-562.
TURNER, N.J, JOHNSON GOTTESFELD, L.M., KUHNLEIN, H.V. & CESKA, A.1992. Edible wood fern rootstocks of western North America: solving anethnobotanical puzzle. J. Ethnobiol. 12: 1-34.
VETTER, J. 2011. Toxicological and medicinal aspects of the most frequent fern species,Pteridium aquilinum (L.) Kuhn. Chapter 25. In H. Fernández et al., (eds.): Workingwith ferns: Issues and applications. Springer Sciences & Business Applications. P.361.
WILEY, G. R. 2003. Archaeobotany: Scope and Significance . In: Richard EvansSchultes & Siri Von Reis (Eds.): Ethnobotany. Timber Press, Portland. P: 400-406.
ZHANG, Y. M. 2007. A preliminary investigation of the history of pteridophyte eatingin China. Journal of Southwest Agricultural University (Social Science Edition) 5(4):103–106.
ZHU, X., KUANG, Y., XI, D., LI, J. & WANG, F. 2013. Absorption of HazardousPollutants by a Medicinal Fern Blechnum orientale L., BioMed ResearchInternational Vol. 2013: 1-6.
KELLER & PRANCE: THE ETHNOBOTANY OF FERNS AND LYCOPHYTES 13

FERN GAZ. 20(1). 2015 14
A SHORT BIOGRAPHY OF THE AUTHORS
Héctor Alejandro Keller, holds a Forestry Engineer’s degree from the UniversidadNacional of Misiones, Argentina (2000), and a Ph.D. in Natural Resources from theUniversidad Nacional del Nordeste (2008). He is currently lecturer in botany andvegetation systematics at the National University of Misiones. Technical officerresponsible for the Multiple-use Guaraní Reserve, 2006-2007. Since 2010 ScientificResearcher of the National Council for Scientific and Technological Research ofArgentina. He has collected 12,300 herbarium specimens, mainly from MisionesProvince, Argentina including ten new species and one new beetle. Author of one bookchapter and 87 articles in scientific journals on ethnobotany, ethnography, ethnobiologyand floristic studies. Epononyms: : Lasiodactylus kelleri Cline (Nitidulidae); Gaya kelleriKrapov. (Malvaceae), Phaeostemma kelleri Morillo (Asclepiadaceae).
Professor Sir Ghillean Prance, University of Oxford BA in Botany (1960) M.A., D.Phil.(1963); research assistant The New York Botanical Garden 1963, B A Krukoff Curatorof Amazonian Botany (1966), Director and Vice-President of Research (1976-1982),Senior Vice President for Science (1982-1988). 39 expeditions to Amazonia, collected350 new species of plants. Director of the Royal Botanic Gardens, Kew 1988 -1999;McBryde Professor, National Tropical Botanical Garden Hawaii 2001-02, McBrydeSenior Fellow (2003-). Visiting Professor at Reading University; author of 20 books and529 scientific and general papers in taxonomy, ethnobotany, economic botany,conservation and ecology; International COSMOS Prize 1993; Fellow of the RoyalSociety; Knight Bachelor; 1995; Victoria Medal of Honour of Royal HorticulturalSociety, 1999; David Fairchild Medal for plant exploration, 2000; Allerton Award in2005; Commander of the Order of the Southern Cross Brazil, 2000; Order of the RisingSun from Japan, 2012.
The authors in a Guarani village in Misiones, Héctor Keller at left, Ghillean Prancesecond from left.

NOTE ON THE REDISCOVERED TYPE SPECIMEN OFANGIOPTERIS INDICA DESV. (MARATTIACEAE)
J. MAZUMDAR
Department of Biological Sciences, Burdwan Town School, Burdwan-713101, India Email: [email protected]
Key words: Angiopteris indica, Herb. Desvaux, India, Marattiaceae, type.
ABSTRACTThe type of the tree fern Angiopteris indicaDesv. (Marattiaceae) was rediscoveredin Herb. Desvaux at P and its status is discussed.
INTRODUCTIONThree species of the marattioid fern genus Angiopteris Hoffm. (Marattiaceae) aregenerally accepted to occur in India (Fraser-Jenkins, 2008; Fraser-Jenkins & Benniamin2010), namely Angiopteris indicaDesv., A. helferiana C.Presl, and A. palmiformis (Cav.)C.Chr. Fraser-Jenkins (2008) accepted A. indica as the oldest available name for plantscharacterized by the combination of the following characters: the soral lines are locatedclose or at the margin, the lamina segments possess prominent teeth near their tips, thelamina colour is darker than in other Indian species, and with the false (recurrent) veinsreaching the soral line or just passing beyond it. In contrast, A. helferiana isdistinguishable from A. indica by its inframarginal sori, whereas A. palmiformis has longfalse veins extended up to the pinnule-midrib.
Angiopteris indica was described by Desvaux in 1813 (Desvaux 1813: 267) and notin 1811 (Desvaux 1811: 207), as misquoted by Moore (1857: 75) and Christensen (1906:57), but see Hooker & Greville (1831) for the correct citation of the name. In theprotologue, Desvaux (1813: 267) described the plants as “frondibus pinnatis, pinnislanceolatis utrinque attenuates” and mentioned the area of origin as “Habitat in Indiaorientali”. He further considered the non-cordiform base on the segments as the mainfeature distinguishing this species from previously described taxa. Desvaux in hisprotologue mentioned that he saw a specimen of the species at Herb. de Jussieu. Laterauthors (Vriese & Harting, 1853; Fraser-Jenkins, 2008) concluded that no specimens ofthis species were found in the Herb. de Jussieu, which is located in the herbarium at the“Museum national d’Histoire naturelle” at Paris (P).
Subsequently, Fraser-Jenkins (2008) designated a neotype: Herb. Wight e Nilghiry[“S. India”], alt. 5000 ped. Wallich List No. 187.8. (K-W). This designation is based onthe crucial assumption that no specimen seen by Desvaux can be found.
RESULTSHere, I present the discovery of a single specimen located at “Herbier de A.N. DesvauxDonné par Mme Vve Lavallée en 1896” in the herbarium P (Fig. 1A) with the barcodeP01646217 (http://sonneratphoto.mnhn.fr/2010/08/09/4/P01646217.jpg). The specimencarries two labels on the left side that were interpreted as written by Desvaux showingthe name “Angiopteris indica Desv., journ. bot. appl, p. 267, Habitat in India orientali”(Fig. 1D). The second label mentions (in French) “was not described in my memoir givenat Berlin” (Fig. 1D). By this annotation Desvaux indicated that the name A. indica was
FERN GAZ. 20(1):15-18. 2015 15

not published in “Berlin Magazine” in 1811 along with many new taxa he described there.A third label below the pinna fragment shows the name “Angiopteris glauca Desv.” (Fig.1B). This name does not correspond to Angiopteris glaucaAlderw. Mal. Ferns, Suppl. I,Corr. 61 (1917) and it is interpreted here as an unpublished name considered by Desvaux.
16 FERN GAZ. 20(1):15-18. 2015
Figure 1. Specimens of Angiopteris indicaDesv. A. Herbarium sheet (P01646217) withtwo pinnule-segments, B. fragmented pinnule (Type), C. second pinnule with long falseveins, D. labels written by Desvaux. Images © Muséum national d’Histoire naturelle,Paris Herbarium (P), reproduced with permission.

The putative type is a fragment of pinnule showing prominent marginal teeth,inconspicuous false veins and sori inframarginal (Fig. 1B). A morphologically distinctsecond pinnule pasted above it (see Fig. 1A) has long false veins extended up to pinnule-midrib, rounded base and nearly median sori (Fig. 1C).
DISCUSSIONOne of these pinnules should be considered as type of A. indica as it was definitelystudied by Desvaux, supported by the evidence of his own handwritten labels, and whichcan be assumed to be the lost type of A. indica from Herb. de Jussieu.. The second pinnule(Fig. 1C) is not a separate portion of the same plant but was pasted from a differentspecimen (pinna): Java, Leschenault 642 (barcode P01646218,http://sonneratphoto.mnhn.fr/2010/08/09/4/P01646218.jpg); Jean-Baptiste Leschenaultwas a French botanist and his plant collections from Java were subsequently used byA.L. de Jussieu and others. But there is no evidence to support the view that Desvauxselected this second pinnule (Fig. 1C) as type and pasted it himself. Actually the secondpinnule was originally cut and pasted in an inverted position in P01646218. As a resultit was easily detached and later erroneously pasted in P01646217.
Desvaux coined the name A. glauca Desv. probably due to glaucous lamina of thepinnule (Fig. 1B), but later revised it as A. indica considering the distribution “IndiaOrientali,” which includes modern India, Sri Lanka, Indonesia, Philippines and Marianas(Cook 2009).
CONCLUSIONThe fragment of pinnule (Fig. 1B) is chosen here as the type of A. indica from the twopinnules in Figure 1A. This type, actually lectotype (designated here), refutes theapplication of the name A. indica to plants from India with marginal sori by Fraser-Jenkins (2008) in displaying inframarginal sori (Fig. 1B), similar to A. helferiana (havingsori about 1 mm distant from margin). But due to wide range of morphological variabilityof species of Angiopteris further additional data are required to confirm the identity andcorrect number of species of Angiopteris in India.
ACKNOWLEDGEMENTSI thank Mr C.R. Fraser-Jenkins, Nepal for discussing the present topic and translation ofDesvaux’s French annotation; Muséum national d’Histoire naturelle (MNHN), ParisHerbarium (P) and Dr G. Rouhan, Curator of Ferns & Lycophytes, MNHN for grantingpermission for publication of the image; and an anonymous reviewer for improving themanuscript.
REFERENCESCHRISTENSEN, C. 1906. Index Filicum. H. Hagerup, Copenhagen. 744 pp.
(http://www.biodiversitylibrary.org/item/9241)COOK, A. 2009. Linnaeus and Chinese plants: A test of the linguistic imperialism thesis.
Notes Rec. R. Soc. (doi:10.1098/rsnr.2009.0051)DESVAUX, N.A. 1811. Observations sur quelques nouveaux generes de fougères et sur
plusieures espéces nouvelles de la meme famille. Berl. Mag. 5: 297-330.(http://babel.hathitrust.org/cgi/pt?id=hvd.32044106317993;view=1up;seq=218)
DESVAUX, N.A. 1813. Description de Fougères nouvelles. J. Bot. Agric. 1: 266-273.(http://gallica.bnf.fr/ark:/12148/bpt6k96848g)
MAZUMDAR: THE REDISCOVERY OF ANGIOPTERIS INDICA 17

FRASER-JENKINS, C.R. & BENNIAMIN, A. 2010 [“2009”]. Fifty rarities andadditions to the pteridophytic flora of Arunachal Pradesh, N.E. India. PanjabUniversity Research Journal (Science) 59: 1-38.
FRASER-JENKINS, C.R. 2008. Taxonomic revision of three hundred Indiansubcontinental pteridophytes with a revised census-list. Bishen Singh Mahendra PalSingh, Dehra Dun. 685 pp.
HOOKER, W.J. & GREVILLE, R.K. 1831. Icones Filicum. Vol. 1, Prostant VenalesApud Treuttel et Wurtz, Treuttel Fil. et Richter etc., London.(http://dx.doi.org/10.5962/bhl.title.3867)
MOORE, T 1857. Index Filicum. W. Pamplin, London. 396 pp.(http://dx.doi.org/10.5962/bhl.title.19640)
VRIESE, W.H. DE & HARTING, P. 1853. Monographie des Marattiacées. Noothovenvan Goor, Leiden & Arnz, Düsseldorf. 60 pp.
18 FERN GAZ. 20(1):15-18. 2015

FIRST CHROMOSOME NUMBER REPORT FOR CYSTOPTERISFRAGILIS (CYSTOPTERIDACEAE: PTERIDOPHYTA) IN IRAN
G. BABAEE & *M. HAJI MONIRI
Department of Biology, Faculty of Sciences, Mashhad Branch,Islamic Azad University, Mashhad, Iran
*Author for correspondence: e-mail: [email protected]
Keywords: fern, polyploidy, tetraploid.
ABSTRACTThe chromosome number and ploidy level of Cystopteris fragilis (L.) Bernh. aredocumented for the first time from Iran. The count was tetraploid, with 2n= 4x=168. This agrees with previous reports of tetraploid C. fragilis from Europe, Asiaand the United States.
INTRODUCTIONCystopteris Bernh. (Cystopteridaceae) comprises about 27 species distributed throughoutthe temperate regions and tropical mountains (Rothfels et al., 2013). Three species occurin Iran: C. alpina (Lamark) Desvaux, C. dickieana Sim and C. fragilis (L.) Bernh. (Roux2001; Khoshravesh et al., 2009). The two latter species have also been reported fromhigh elevations in the Hindu Kush Mountains of neighboring Afghanistan (Breckle,1987). In general, these species of Cystopteris are found in shady, rocky, wet habits(Bhellum and Razdan, 2012). The taxonomy and distribution of its species in Iran havebeen reported by Khoshravesh et al. (2009). Cystopteris fragilis differs from C. dickieanain having spiny spores (vs. verrucate in C. dickieana).
The chromosome numbers of n=84 and n=126 (tetra- and hexaploid, respectively)of C. fragiliswere first reported from Europe (Manton, 1950). Vida (1976) later reportedoctaploids in Central Europe. In North America, tetraploids and hexaploids of C. fragilishave been found (Wagner, 1955; Haufler and Windham, 1991; Haufler et al.; 1995, Palerand Barrington, 1995). In Asia, tetraploids and hexaploids have been reported from Indiaand Japan (Bhellum and Razdan, 2012; Mitui, 1970). In 2010, a pentaploid with 2n=210was also recorded from Mongolia Altai by Kawakami et al. (2010). For Iran, however,chromosome numbers of C. fragilis have not been reported previously. This paperrepresents the first such report for the country.
MATERIAL AND METHODSFixed cytological material and voucher specimens were collected by the first author fromthree populations in natural habitats in Akhlamad waterfall valley, ca. 65 km NW ofMashhad, NE Iran at an elevation 1500 m in May and June 2013. The populationsoccurred on stony or rocky slopes exposed to waterfall, within shaded habitats, andoccasionally in the cement of the roadside. Vouchers of examined specimens aredeposited in IAUM and NY. Our initial identification was checked by comparison withdescriptions in the floristic literature (Khoshravesh et al., 2009) and confirmed by RobbinC. Moran at NY.
The chromosome squash method used followed that of Windham and Yatskievych(2003). Pinnae with whitish sporangia were collected every half hour from 9 am to 1pm. The pinnae were fixed in Farmer’s Solution (1:3 glacial acetic acid-ethyl alcohol)
FERN GAZ. 20(1):19-22. 2015 19

and Carnoy’s Solution (ethanol, glacial acetic acid and chloroform in the proportion of6: 1: 3). Large numbers of sporangia were squashed in acetocarmine. Photography weretaken with a digital camera Dino-Lite connected to Labomed LX 400 microscope.
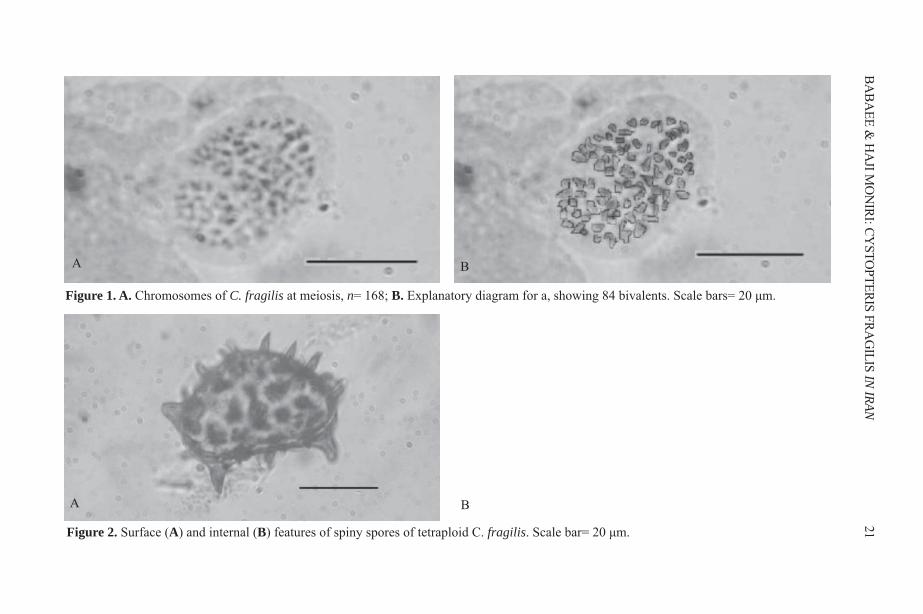
RESULTS AND DISCUSSION We found n=84 pairs (bivalents) at meiotic metaphase I (Figure 1). This is the firstchromosome count of C. fragilis from Iran. This number agrees with previous countsfrom Jammu and Kashmir ((Bhellum and Razdan, 2012), Mongolian Altai (Kawakamiet al., 2010) in Asia; Finland, (Sorsa, 1961), and Iceland (Löve & Löve, 1943) in Europe;Ontario, British Columbia, and Greenland in Canada (Britton, 1953; Taylor and Mulligan,1968; Dalgaard, 1989), and Michigan, USA (Wagner, 1955).Each sorus contained 6–19 sporangia. The most important factor in obtaining meioticfigures at the proper stage was the time of fixation. Descriptions and the figures in thepresent paper refer to fixation at 11am. Tetrad spores were produced by the secondmeiotic division and many well filled, spiny spores were observed (Figure 2). The sizeof 25 spores from one leaf was measured in distilled water as followed: 38.48–48.60 ×23.40–28.65 mm. The size straddles that reported for tetraploids and hexaploids withinCystopteris by Blasdell (1963). He found that tetraploids had spore lengths from 32–42mm and hexaploids from 38–48mm. It is not known what mounting media Blasdell usedto measure his spores—whether distilled water, Hoyer’s solution, or Canada balsam. Useof different mounting media might explain why our results of spore length measurementsdo not fall exactly within the tetraploid range reported by Blasdell (1963).
ACKNOWLEDGMENTSWe thank Dr. R. C. Moran (NY) for protocol advice, comments on the manuscript, andconfirmation of the species identification. We also thank Mr. M. Joharchi for the originalidentification the species and M. Basiri for his assistance in the field.
REFERENCESBHELLUM, B.L. & RAZDAN, B. 2012. Cystopteris fragilis f. kashmiriensis Bhellum
and Razdan- A new forma from India. International Journal of Scientific and ResearchPublication 2(4): 1– 4.
BLASDELL, R.F. 1963. A monographic study of the fern genus Cystopteris. Memoirsof the Torrey Botanical Club 21(4): 1–102.
BRAITHWAITE, A.F. 1964. A new type of apogamy in ferns. New Phytol. 63 (3): 293–305.
BRECKLE, S.-W. 1987. Distribution and ecology of Cystopteris (Athyriaceae) Withinthe Flora Iranica region. Plant Syst Evol. 155: 59–69.
BRITTON, D.M. 1953. Chromosome studies on ferns. Am J Bot. 40: 575–583.DALGAARD, V. 1989. Additional chromosome numbers in vascular plants from the
Disko Bugt area (west Greenland). Willdenowia 19: 199–213. HAUFLER, C.H., MORAN, R.C., & WINDHAM, M.D. 1995. Cystopteris. Pages 263–
270. In: Flora of North America Editorial Committee, editors. Flora of NorthAmerica, volume 2. Pteridophytes and Gymnosperms. Oxford University Press, NewYork.
HAUFLER, C.H. & WINDHAM, M.D. 1991. New species of North AmericanCystopteris and Polypodium, with comments on their reticulate relationship. Am FernJ. 81: 7–23.
20 FERN GAZ. 20(1):19-22. 2015
BABA
EE & H
AJI M
ON
IRI: CYSTO
PTERIS FRAG
ILISIN IRAN
21
Figure 1. A. Chromosomes of C. fragilis at meiosis, n= 168; B. Explanatory diagram for a, showing 84 bivalents. Scale bars= 20 μm.
A B
Figure 2. Surface (A) and internal (B) features of spiny spores of tetraploid C. fragilis. Scale bar= 20 μm.
BA

KAWAKAMI, S.M., KAWAKAMI, S., KATO, J., KONDO, K., SMIRNOV, S.V. &DAMDINSUREN, O. 2010. Cytological study of a fern Cystopteris fragilis inMongolian Altai. Chromosome Botany 5: 1–3.
KHOSHRAVESH, R., AKHANI, H. & GREUTER, W. 2009. Ferns and fern allies ofIran. Rostaniha (Botanical Journal of Iran) 10 (1): 1–129.
LÖVE, A., LÖVE, D. 1943. The significance of differences in the distribution of diploidsand polyploids. Hereditas 29: 145-163.
MANTON, I. 1950. Problems of cytology and evolution in the Pteridophyta. Cambridge:University Press. London. 316 Pp.
MITUI, K. 1970. Chromosome studies on Japanese ferns. Japanese Journal of Botany 45: 84–90.
PALER, M.H. & BARRINGTON, D.S. 1995. The hybrid Cystopteris fragilis × C. tenuis(Dryopteridaceae) and the relationship between its tetraploid progenitors. Syst Bot.20: 528–545.
ROTHFELS, C., WINDHAM, M.D. & PRYER, K.M. 2013. A plastid phylogeny of thecosmopolitan fern family Cystopteridaceae (Polypodiopsida). SystBot. 38: 259–306.
ROUX, J. P. 2001. Conspectus of southern African Pteridophyta. ABC Press, EppingInd., Cape Town. 223 Pp.
SORSA, V. 1961. Chromosome studies on Finish Pteridophyta II. Hereditas 47: 480–488.
TAYLOR, J.A., & MULLIGAN, G.C. 1968. Flora of the Queen Charlotte islands. Part2: Cytological aspects of the vascular plants. Canada Department of Agriculture Press,first edition. 659 Pp.
WAGNER, W.H. 1955. Cytotaxonomic observations on North American ferns. Rhodora57: 219–240.
WINDHAM, M.D., & YATSKIEVYCH, G. 2003. Chromosome studies of cheilanthoidferns (Pteridaceae: Cheilanthoideae) from the western United States and Mexico. AmJ of Bot. 90: 1788–1800.
22 FERN GAZ. 20(1):19-22. 2015

THE DUTCH RUSH: HISTORY AND MYTH OF THE EQUISETUM TRADE
W. DE WINTER
Alterra, P.O. box 47, 6700AA Wageningen, NetherlandsEmail: [email protected]
Key words: Equisetum hyemale, economic botany, ethnopteridology, trade history
ABSTRACTIn England in the early 19th century at least two products went by the commercialname Dutch Rush, viz. the Rough Horsetail Equisetum hyemale L. used in cabinetmaking and similar crafts, and the Common Club-rush/Bulrush Schoenoplectuslacustris (L.) Palla used in matting and chair manufacturing. Some authors didnot heed the scientific names and confused the properties and geo-culturalbackgrounds of both products. Thus the myth took hold that E. hyemale was inculture in the Netherlands and that is was deliberately planted and cared for toprotect that country from the sea. Scarce but widespread evidence of trade revealsthat this species was economically insignificant. The idea that it owes its commonname to imports from Holland could be correct; however, other parts of Northand Central Europe, especially the upper Rhine Valley, are more likely to be theoriginal sources from where the Dutch obtained the plants. North America canbe reasoned to be an alternative origin, but evidence for this hypothesis is stilllacking.
INTRODUCTIONEver since the sixteenth century, many authors in a number of Western Europeancountries have reported the use of Equisetum hyemale L. by various trades (e.g. Fuchs,1543; Gerarde, 1597; Bock & Sebisch, 1630; Pexenfelder, 1670; Ruppe & Haller, 1745;Anon., 1749; Pernety, 1771; Krünitz, 1785; J. E. Smith, 1802; Headrick, 1813; Stewart,1815; Phillips, 1818; Hooker, 1821; Gill, 1822). Material evidence of its use is found ininventories of workshops (e.g. Giskes, 1979) and characteristic scratches from itssiliceous skin on antique woodcraft (Esterly, 1998). In addition, several authors duringthe past two centuries showed that the vernacular name “Dutch Rush” used in Englandrelates to large-scale imports from Holland1 (Pratt, 1846; Francis, 1851; Johnson &Sowerby, 1856; Moore, 1861; Pratt, 1866). Up to the present day the statement is repeatedfrequently (Weeda et al., 1985; Øllgaard & Tind, 1993; Page, 1997). The apparent sourceto which this can be traced back is Edward Newman (Newman, 1842, 1844), who alsonoted that the species was cultured in Holland and played an important role in the defenceof the coast against the eroding action of the sea:
“… for this purpose it is imported, under the name of “Dutch Rush” in large quantities,from Holland, where it is grown on the banks of canals and on the sea ramparts, whichare often bound together and consolidated by its strong and matted roots.”
FERN GAZ. 20(1):23-45. 2015 23
1Wherever the toponym Holland is used, literal citations excepted, it is to be understoodin the strict sense, i.e. the western part of the Netherlands, or the present provincesNoord-Holland and Zuid-Holland.

This is at odds with the ecological preferences of the species, as well as with both itshistorical and present day distribution (de Winter, 2007). These problems can be summarisedunder three main points:1. the alleged abundance of E. hyemale in Holland finds no evidence in its current
distribution in that country, nor is it supported in historical Dutch reports; 2. the reported occurrence and even cultivation of E. hyemale along canals and the practice
of using it for the solidification of dykes, dunes, or any kind of coastal protectionconstructions finds no reference in Dutch literature, nor in the present distribution andhabitat preferences of European E. hyemale;
3. the plant detailed by Newman by its size and number of ridges suggests American E.hyemale subsp. affine (Engelm.) Calder & Roy L.Taylor rather than European E. hyemalesubsp. hyemale, which casts doubt on the continental origin of the merchandise.If these doubts are well-founded, then the question must be addressed as to whether E.
hyemale really was imported from Holland to England, and where the origin should be soughtof the plants sold on London markets.
HISTORY OF THE NAME “DUTCH RUSH”A number of vernacular names have been used for E. hyemale in English, but most of themnever became popular and have disappeared into oblivion (Table 1). The oldest historic oneis “shave-grass”, which was first recorded in the 14th century: “Cauda equina, caudaCaballina idem est. angl, schauegres” (Anon., 1350-1400; Lancaster, 1887; Murray, 1971).To shave is derived from the Anglo-Saxon scafan through the Old English shaven, and MiddleEnglish schaven/schafen (Flexner & Hauck, 1993), whence there is a direct relation with theOld High German scafthawi (Graff, 1838) and the Dutch stem schaaf - as in schaafstro (deWinter, 2012). Although the verb “to shave” at present is predominantly used to refer to theprocess of removing the beard, the general meaning is to make a surface smooth (Webster,1913). The name is continuous through Turner (1538) and Gerard (1597). “Shave-grass” hasbeen largely replaced now by the more official “rough horse-tail” (Bolton, 1790; J. E.Smith, 1802), and “Dutch Rush”.
24 FERN GAZ. 20(1):23-45. 2015
Table 1. Vernacular names used in British English
Name Source
Shave-grass (discussion in this paper)Dutch Rush (discussion in this paper)Rough horsetail From the French/Latin asprella (de Winter, 2012), introduction
unknownDutch shave-grass species list of Scarborough (Hinderwell, 1811)Holland rushes In advertisements (Glover, 1843; Measom, 1861)Dutch reed rarely (Bailey, 1756; Martin, 1813; Henslow, 1839; Trimmer,
1842)polishing rushes customs’ duties lists (Burn et al., 1831; Parnell, 1831; Ellis, 1837)shave weed (Aubrey, 1848)shauynge gyrs (Turner, 1538)dyßhewaßhynges (Turner, 1538)Pewterwort e.g. (Withering, 1776; Targioni Tozzetti, 1813; Wilkinson, 1858)scouring[-]rush American (Law Olmstedt et al., 1924)
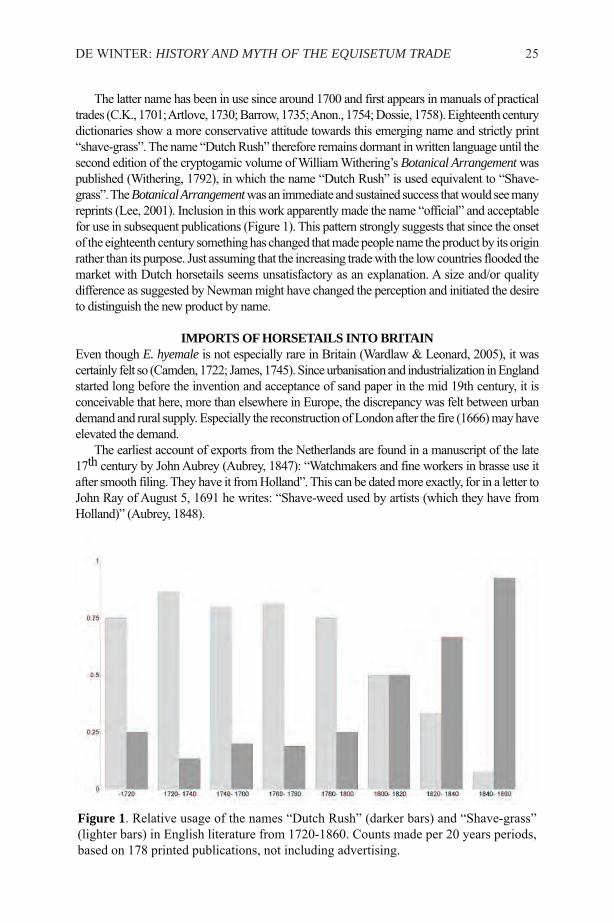
The latter name has been in use since around 1700 and first appears in manuals of practicaltrades (C.K., 1701; Artlove, 1730; Barrow, 1735; Anon., 1754; Dossie, 1758). Eighteenth centurydictionaries show a more conservative attitude towards this emerging name and strictly print“shave-grass”. The name “Dutch Rush” therefore remains dormant in written language until thesecond edition of the cryptogamic volume of William Withering’s Botanical Arrangementwaspublished (Withering, 1792), in which the name “Dutch Rush” is used equivalent to “Shave-grass”. The Botanical Arrangementwas an immediate and sustained success that would see manyreprints (Lee, 2001). Inclusion in this work apparently made the name “official” and acceptablefor use in subsequent publications (Figure 1). This pattern strongly suggests that since the onsetof the eighteenth century something has changed that made people name the product by its originrather than its purpose. Just assuming that the increasing trade with the low countries flooded themarket with Dutch horsetails seems unsatisfactory as an explanation. A size and/or qualitydifference as suggested by Newman might have changed the perception and initiated the desireto distinguish the new product by name.
IMPORTS OF HORSETAILS INTO BRITAINEven though E. hyemale is not especially rare in Britain (Wardlaw & Leonard, 2005), it wascertainly felt so (Camden, 1722; James, 1745). Since urbanisation and industrialization in Englandstarted long before the invention and acceptance of sand paper in the mid 19th century, it isconceivable that here, more than elsewhere in Europe, the discrepancy was felt between urbandemand and rural supply. Especially the reconstruction of London after the fire (1666) may haveelevated the demand.
The earliest account of exports from the Netherlands are found in a manuscript of the late17th century by John Aubrey (Aubrey, 1847): “Watchmakers and fine workers in brasse use itafter smooth filing. They have it from Holland”. This can be dated more exactly, for in a letter toJohn Ray of August 5, 1691 he writes: “Shave-weed used by artists (which they have fromHolland)” (Aubrey, 1848).
DE WINTER: HISTORY AND MYTH OF THE EQUISETUM TRADE 25
Figure 1. Relative usage of the names “Dutch Rush” (darker bars) and “Shave-grass”(lighter bars) in English literature from 1720-1860. Counts made per 20 years periods,based on 178 printed publications, not including advertising.

The demand declined when glass paper came into use, but it never completelydisappeared. In the USA it was still advertised in 1938 (Kaliban’s Grocery andMarket, 1938). Nowadays some still prefer it to scrape their clarinet reeds (Intravaia& Resnick, 1965; Taillard & Dalmont, 2012).
Testimony of imports into England, which lasted more than one and a halfcenturies, exists in lists of import duties of the English customs (Burn et al., 1831;Parnell, 1831; Ellis, 1837), but is absent in earlier legislation (e.g. Steel, 1796).Often the horsetails could be imported, transported, and exported tax-free (deMartens & Murhard, 1836; Anon., 1837; MacGregor, 1843; Anon., 1849a, 1853,1858) or they were not considered important enough to justify their own category(Koninkrijk der Nederlanden, 1816, 1822). As a consequence, no accounts of theirtrade were kept and they do not show in yearly statistical surveys (Departement vanFinanciën, 1848). England and Holland were the two countries with the worst keptstatistics on agriculture and trade (MacGregor, 1843). Yet, although far fromcomplete, a few statistics of the early period remain, covering the imports into theport of London and allowing sampling of the existing data of imports of E. hyemale.However, in all of them the horsetail trade is conspicuous by its absence, viz. inJanuary 1683 (Houghton, 1728), from May to October 1735 (Anon., 1735), andfrom January to June, 1776 (Whitworth, 1777b, 1777a).
In the nineteenth century Dutch newspapers meticulously reported freight loadedand unloaded per port, frequently even per ship. Any product of trade of anyimportance would be expected to be listed. Nonetheless horsetails are not mentionedby any of their known names. With one exception, however: in the second half of1855, 142 inland navigation barges with in total 6050 bundles of shave-grass arrivedin Brussels (Anon., 1856). Estimating the total volume of Equisetum transportedrequires acquaintance with the size of such a bundle, which we do not know. Arough estimate for a bundle of 80 cm perimeter (as is used at present for bulrushes:de Vries, 2008) would be 500 – 1000 stems. Given that dense stands have 200 –500 stems per m² (Rutz & Farrar, 1984), the annual production for Brussels onlywould have required the depletion of c. 0.6 – 3 ha. Such small amounts could easilyhave been found on Belgian territory, but apparently for reasons of quality or cost,long-distance transport per ship was more attractive.
In the mid-nineteenth century, exports of reeds and rushes to Britain exceededthose to Belgium by about 50% (Departement van Financiën, 1848) and if the exportof horsetail may be estimated proportionally to this, the economic value must havebeen insignificant. John Yeats, an English commercial-geographer, who lived inHolland in 1845/46, compares E. hyemale, which “is occasionally imported fromHolland” with Scirpus lacustris L. (now Schoenoplectus lacustris (L.) Palla), sayingthat “Many vessels laden with this rush arrive annually in England from Hollandand Belgium, bringing thirty or forty tons of rushes each voyage. This is a verylarge quantity considering the lightness of the material. More than 1,000 tons ofbulrushes are annually imported into the United Kingdom” (Yeats, 1870). It shouldbe noted, though, that these figures stem from the second half of the nineteenthcentury, when the use of E. hyemale was already on the decline in favour ofsandpaper, of which mass production in London had started by 1833. But also muchearlier, in 1827, international horsetail trade was but marginal. From an examplerecord of the net produce of customs duties it can be deduced that the total value ofpolishing rushes legally imported into Britain must have been ₤13.25 for that year
26 FERN GAZ. 20(1):23-45. 2015

(Parnell, 1830).CULTIVATION AND CARE IN HOLLAND: COAST PROTECTION
Newman’s statement, that the Dutch grew E. hyemale “on the sea ramparts, which are oftenbound together and consolidated by its strong and matted roots” (Newman, 1844), isrepeated with slight variations in later literature. The authors obviously have copied eachother in sequence, rephrasing the allegation without adding new observations:
“it is grown on the banks of canals and on the sea ramparts, which are often boundtogether and consolidated by its strong and matted roots” (Newman, 1844)
“The Dutch are well acquainted with the value of its long and matted roots in restrainingthe wasting effects of the ocean, which would soon undermine their dykes were it not forthe Equisetum hyemale which is planted upon them” (Francis, 1851)
“the plant is of immense value in its native country from the extraordinary length andinterlaced growth of its root-fibres, which mat together and consolidate the loose andswampy soil in which they grow, and thus form one of the most effectual water-dams of solevel a land.” (Wilkinson, 1858)
“this species is planted to support embankments, which it does by means of itsbranching underground stems” (Moore, 1861)
Along long stretches of the coast of Belgium and Holland, and the outward coasts ofthe islands in the North, belts of natural and semi-natural sand dunes protect the countryfrom the sea. The hybrid E. × moorei Newman (E. hyemale × E. ramosissimum Desf.) isfound in great numbers at several locations in the dunes of Zuid-Holland and southernNoord-Holland. It was first reported by Du Mortier in 1825 (as E. trachyodonA.Br.) nearBeverwijk (Du Mortier, 1869), where it still can be found today. It is locally abundantbetween Scheveningen and Hook of Holland, where it has been known since the late 18thcentury (Anon., 1796; van Hall et al., 1832). It is surprising that J.E. Smith (1793) did notnote it when visiting the place. Leiden (L) has a number of specimens collected in the Westof Holland during the 19th century; however, until 1870, when the first collection wasmade from the fore-dunes south of Scheveningen, all the collections originate from theinterior dunes around Haarlem.
In 1797, Jan Kops set out to report on the state of the dunes of Holland. He was aneager botanist, paid special attention to any plants that could be used to control shiftingsand and, as an agronomist working for the ministry, was also keen to find plants that couldbe made profitable in any way (Baert, 1943). If Newman with “the consolidation of searamparts” referred to the fixation of shifting sands in the dunes, it is hard to imagine thatthis would have escaped Kops’s attention. Yet Kops explicitly states that no other meansare known to him to fix the dunes than marram (Ammophila arenaria (L.) Link), straw,and reed mats (Kops, 1798; van Eys et al., 1799). Neither earlier (Montin, 1771; Le Francqvan Berkheij, 1780; van Geuns, 1789) nor later authors (Spengler, 1891; Vuyck, 1898) oftechnical treatments about sand fixation mention Equisetum for this purpose, even thoughLe Francq van Berkheij lists “Paardestaart, Equisetum” among the plants of the fore-dunes.Despite the present abundance of the hybrid in the dunes of southern Holland never, fromthe 18th century to the present, has its distribution been said to be deliberately promotedby man, and the remarks of Francis and Newman above cannot be attributed to it.
Where dunes are weak or absent, dykes have been built to protect the land, as in thetopographically complex estuarine area of Zeeland and Zuid-Holland, along the rivers,along the entire coast of the Wadden Sea, and, before 1932, along the Zuiderzee and itsinlets. Sea-dykes must be protected by a hard surface since no vascular plant can form matsfirm enough to protect the dyke’s surface from the action of the waves. At the onset of the
DE WINTER: HISTORY AND MYTH OF THE EQUISETUM TRADE 27

19th century such hard-shells were not yet customary.Reed and rushes were recognised as effective agents to break the action of the waves
and to prevent erosion of the banks of lakes (Meese, 1768), but they grow neither inseawater, nor at the high water limits of rivers. Sea-dykes were protected by staplingbundles of straw, eelgrass and rushes to the dyke-surface to form a soft shell that had tobe renewed every autumn (Bréval, 1726; Schraver, 1807).
Inland river dykes consist of a core of sand or clay, covered with a water resistantlayer of clay and finished with a grass-covered top layer (Fliervoet, 1992). Mats ofsuperficially rooting herbs protect the surface against erosion, but deep-rooting plantsweaken the construction (de Haan et al., 2001; 2003) and can perforate the water resistantclay mantle (Technische Adviescommissie voor de Waterkeringen, 1985), without addingmuch to the stability (Sykora & Liebrand, 1987). High growing dense stands of E.hyemale could outcompete lower herbs that offer better protection. Finally, frequentlygrazed or mown low vegetation is less than optimal for this woodland species and atpresent it is not found on dykes, nor are there collections in the National Herbarium fromsuch places. The hybrid E. × moorei has a number of stations along the river Rhine, butit is still rare on dykes. Altogether both technical and historical evidence for Newman’sstatement appear to be entirely lacking.
OVER-REPORTING OF DUTCH RUSH DUE TO DUTCH RUSHESThe likely source of the delusion is the 18th century bestseller, The Gardeners Dictionary(Miller, 1754, Ed. 4) that depicts how species of rush “grow on the Sea-shores, wherethey are frequently watered by the Salt-water. These two Sorts2 are planted with greatCare on the Banks of the Sea in Holland, in order to prevent the Water from washingaway the Earth; which, being very loose, would be in Danger of removing every Tide, ifit were not for the Roots of these Rushes; which fasten themselves very deep in theGround, and mat themselves near the Surface, so as to hold the Earth closely together.Therefore, whenever the Roots of these Rushes are destroyed, the Inhabitantsimmediately repair them to prevent farther Damage.” This paragraph is quoted from thesection on Juncus as “Juncus acutus”.
Miller must have meant the glaucous bulrush Schoenoplectus tabernaemontani (C.C.Gmelin) Palla that was cultured in great numbers in the brackish water of the Maasestuary, and possibly to some extent S. maritimus (L.) Lye, which was not cultured, butcommon in the more seaward parts of the estuary and known for its even greater capacityof promoting the sedimentation rate of silt than the former (Clevering & van Gulik, 1990;Weeda et al., 1994).
Miller’s handbook on wild and cultivated plants went into eight editions3, and wascopied (e.g. MacFarquhar & Gleig, 1797; Knight, 1833), and translated into French,German and Dutch. As Miller’s text was copied over and over during more than a century,gradually elements changed, and new ones slipped in (Table 2).
Noteworthy is Loudon’s addition that combines the name Dutch rush, and its usabilityfor scouring metals (Loudon, 1829). Rushes (for making chairs) were imported from the
28 FERN GAZ. 20(1):23-45. 2015
2i.e. Juncus acutus, capitulis sorghi C.B.P. & J. acutus maritimus Anglicus Park.
3Cited here is the fourth edition of 1754, but essentially the same text is found in theDutch translation of 1745 (Miller, 1745) and therefore in earlier English editions notseen by the author.

Netherlands into England in large quantities (Yeats, 1870). Inevitably, they weresometimes designated as “Dutch rushes”, an adjective combination that never made it toa taxonym.
Traders used the name indiscriminately for both E. hyemale and Schoenoplectus. Inadvertising in the first half of the century, it is rarely made explicit which kind of rush isoffered (e.g. “Dutch polishing rushes” or “Dutch bull rushes”). In less than half of suchadvertisements the context, such as chairmakers’ or cabinet-makers’, revealed the nature
DE WINTER: HISTORY AND MYTH OF THE EQUISETUM TRADE 29
Period Interpretation
1724to1754
The first and second Sorts grow on the Sea Shores, where they are frequentlywashed by the Salt Water. These two Sorts are planted with great Care on theBanks of the Sea in Holland, in order to prevent the Water from washing awaythe Earth; which, being very loose, would be in Danger of removing every Tide,if it were not for the Roots of these Rushes; which fasten themselves very deepin the Ground, and mat themselves near the Surface, so as to hold the Earthclosely together. Therefore, whenever the Roots of these Rushes are destroyed,the Inhabitants immediately repair them to prevent farther Damage. In theSummer-time, when the Rushes are fully grown, the Inhabitants cut them, andtie them up into Bundles, which are dried, and afterward carried into the largerTowns and Cities, where they are wrought into Baskets, and several other usefulThings, which are frequently sent into England. These Sorts do not grow sostrong in England, as they do on the Maese, and some other Places in Holland,where I have seen them upward of four Feet high (Miller, 1754).
1797 The conglomeratus, and acutus or marine rush, are planted with great care onthe banks of the sea in Holland (MacFarquhar & Gleig, 1797)
1829 J. acutus and maritimus are planted on the sea-embankments of Holland, andalso in some parts of our own coasts, and in America. The roots run deep into thesand, and form a matted body which holds it together. In Holland, when the plantsare fully grown and in flower, they are cut down, dried, and bound up like corn.The J. acutus, being very rough, is used for scouring copper and other vessels,and is one of the plants imported into this country for that purpose, under thename of the Dutch rush. The other species, and often both, are plaited into mats,baskets, chair-bottoms, ropes, etc. (Loudon, 1829)
1833 (…) are made of bulrushes; these grow in this country, naturally but not verycommonly, in deep slow streams. The demand for them is greater than the homesupply, and a considerable quantity is imported from Holland. (Society for theDiffusion of Useful Knowledge, 1833)
1842 Equisetum hyemale: for this purpose it is imported, under the name of “ DutchRush,” in large quantities, from Holland, where it is grown on the banks of canalsand on the sea ramparts, which are often bound together and consolidated by itsstrong and matted roots. Bundles of this imported Dutch Rush are exposed forsale by many London shopkeepers. (Newman, 1842)
Table 2. The evolution of Miller’s account on bulrush-culture to Newman’s account ofE. hyemale

of the product offered (Table 3). It must have been this ambiguous use of the name thatled Newman to his misunderstanding. Evidently, when writing about E. hyemale, he didnot make the distinction and copied unreliable sources without checking.
CULTIVATION AND CARE IN HOLLAND: HARVESTING FOR EXPORTWhereas in English literature it is often stated that Dutch Rushes were cultivated andexported by the Dutch (Newman, 1842, 1844; Francis, 1851; Moore, 1861; Pratt, 1866),the continental literature remains almost taciturn about any culturing activities. Theearliest mention of such a trade in Dutch literature is found in the Flora Batava (Kops& Van der Trappen, 1846), published shortly after Newman’s publication in ThePhytologist (Newman, 1842).
In the Netherlands, E. hyemale is not an abundant species. In the east of the countryseveral healthy populations exist, but it is hard to imagine that these could have supportedprolonged harvesting for both domestic and export markets. In Holland in the strict sense,i.e. the western part, E. hyemale is virtually absent, but the hybrid E. × moorei is quitecommon in the dunes and along railroads (de Winter & Lubienski, 2012). It would beconceivable that present day populations are not representative for the abundance in the18th and 19th century. Both habitat destruction and harvesting might have caused sucha decline. De Gorter (1781) had found the species in forests in the county Zutphen andKops & Van der Trappen (loc. cit.) say it is abundantly found near Zutphen, at a numberof locations in the West, the latter almost certainly to be attributed to the hybrid, and atsome riverine locations for many of which the same applies. In the 17th century, Comelin(1683) saw it in stagnant-water ditches near the river Vecht near “den Bergh”4, at theborder of the firm, sandy soil of Utrecht and the fen areas of Holland.
E. hyemale is a species of moist woodlands. The area of woodland in the Netherlandshas not declined since 1800, but has actually increased (Bijlsma, 2003), as before thattime much had been cleared. Older sources often associate the species with ditches rather
30 FERN GAZ. 20(1):23-45. 2015
Table 3. Advertisements with “Dutch Rushes” or “Holland Rushes” from before 1850.
Advertisements where product defined as: Number
Equisetum 4Schoenoplectus 8Unknown 13Total 25
Sources: Bury and Norwich Post: 29 Nov 1815; Carlisle Journal: 09 Mar 1833, 24 Aug1833; Carlisle Patriot: 28 Oct 1820; Chelmsford Chronicle: 03 Oct 1783; ChesterChronicle: 23 Mar 1849; Hampshire Advertiser: 15 Sep 1832; Hampshire Chronicle: 26Jul 1819 2x; Hampshire Telegraph and Sussex Chronicle: 23 Jul 1832; HampshireTelegraph: 02 Mar 1829, 25 Apr 1831; Hull Packet: 07 Jan 1801, 03 Mar 1801, 27 Feb1835; Kentish Gazette: 07 Jan 1845; Liverpool Mercury: 13 Jun 1823, 17 May 184 4;Norfolk Chronicle: 09 Dec 1815; Salisbury and Winchester Journal: 19 Sep 1803, 26Feb 1821; Stamford Mercury: 21 Apr 1848; The Ipswich Journal: 22 Jun 1805, 14 Sep1805; The Times: 08 Sep 1831
4presumably Nederhorst den Berg, about 25km SE. of Amsterdam

than woodland, which may be explained by its habit of persisting long after the treeshave been felled. Altogether it seems unlikely that the magnitude of the population wouldhave permitted large scale harvesting to support an export trade over a long time and itmust be questioned whether this country has ever been the source of such trade.
Comelin (loc. cit.) noted that at den Bergh it was collected for turners to polish withit5. But later, from 1806, a number of annual reports have been compiled about the Dutchagriculture, including products gathered from uncultivated lands, such as reed(Phragmites), rushes (Schoenoplectus) and herbs (Kops, 1807, 1808, 1809, 1816, 1819,1821, 1822c, 1822b, 1822a, 1829). No mention is made of any production or harvestingof Equisetums.
Nozeman (1783) expresses his amazement that E. hyemale is imported into theNetherlands from Spain (see below), rather than collected from local stocks. Subsequentbotanists agree: “This plant does grow in some places of our home country, in woodsand sandy lands, but it is, as far as I have been able to find out, not used” (van Hall,1854). “The shave-grass that the cabinet-makers use does not originate fromour land, but is imported from elsewhere. From [the Netherlands] it is further transportedto England, where it, completely undeserved, is designated by the name DutchRushes”(Oudemans, 1862; likewise: de Vries et al., 1870).
An interesting phenomenon is that, analogous to the English “Dutch Rush”, in theNetherlands it was sometimes called “Spanish rush” (Noel Chomel & De Chalmot, 1778;Van Meerten-Schilperoort, 1830; 1843; van Hall, 1843). The epithet “Spanish”intermittently used in the Dutch names of the species does not indicate a Spanish origin.Though it is found in Spain (Prada, 1986; Salvo Tierra, 1990), it occurs too infrequentlyto allow any significant yield (C. Fraser-Jenkins, pers. com.). Also, historically andregionally the predicate has been applied to at least thirty other species (Heukels, 1907),which seem to have little in common, though for quite a few of them it might beinterpreted as “conspicuous” or “foreign”.
A few authors hold the view that that the Netherlands does or could producecommercial horsetails: “This wood [Zalkerbos] also furnishes very good shave-grassesto polish wood” (van der Aa, 1851), without revealing his source nor the scale of theexploitation. In almanacs beggar boys were urged to collect shave-grass (among otherproducts of nature) to earn their own living (Heldring et al., 1837; Heldring, 1845). Thismay denote more about the low value and little enthusiasm to collect the stems in theNetherlands. Altogether, the conclusion seems justified that if Dutch-produced horsetailswere ever imported into England, this must have taken place longer ago than the memoryof the 19th century writers cited above, therefore no later than the first half of the 18thcentury.
CONTINENTAL TRADE AND ORIGIN OF THE SPECIESEven if the plants have never been cultivated in the Netherlands, nor commercially harvestedfrom the wild, it is still possible that Dutch traders imported them into London from othercontinental or overseas sources. The question then is where they would have originated.
DE WINTER: HISTORY AND MYTH OF THE EQUISETUM TRADE 31
5Comelin’s habitat description is more typical for E. fluviatile L. with which E. hyemalewas often confused, and so was his description: Equisetum foliis nudum, non ramosumsive junceum. B. pin. Hippuris foliis & nuda Tab. Equisetum majus aquaticum prim.icon J. Bauh. Belg. Groot Paarde-staart. However, his observation on collecting theplant shows that he was at least partially correct.

There is evidence of horsetail trade elsewhere in Europe. Tariffs of the Zollverein(customs union, 1818-1871) name Schachtelhalm in near complete lists of itemstransported through the German lands (Anon., 1824; de Martens & Murhard, 1836;Anon., 1849a, 1849b, 1853, 1858). Likewise, Russia levied a small duty on horsetails(MacGregor, 1850), as did Norway, (Arntzen, 1830), France (Dujardin-Sailly, 1813) andAustria (Klenner, 1822; Anon., 1824, 1838), whereas Illyria (Anon., 1836) let them passfor free.
A number of more incidental sources can be found too. According to an early 19thcentury encyclopedia, E. hyemalewas a lucrative product and exports from the southernFrench department Bouches-du-Rhone amounted to 10,000 francs annually (Leman,1826). The amount in francs suggests the figure applies to statistics postdating 1795,when this currency had been introduced. The 1802 overview of this department, however,makes no mention at all of such an export (Michel, 1802). Moreover, it is stated that thedepartment is very poor in woodland, a condition that has not changed to the present,and this explains the near absence of E. hyemale in those parts (J. le Paslier & C. Déliry,pers. com.). Duval-Jouve also expressed his doubt about the figures he had from deVilleneuve (de Villeneuve, 1821), but after investigating the matter he found that theyreferred to E. ramosissimum and E. palustre L., of which at least 30,000 bundles weresold each year for kitchen usage (Duval-Jouve, 1864). Local trade in Provence wasprovided by vendors crying “Leis fretadous de coussooudo”6 (Achard, 1785), who werenot likely to be selling the hard skinned E. hyemale, since their merchandise would haveruined the kettles.
Abundant sources of E. hyemale were found in the forests along the Rhine, wherespecialised collectors took it to sell it to carpenters and cabinetmakers, or to plait ringsof them (the so-called Schaftheukränze, Figure 2) to clean kitchen utensils and floors(Kirschleger, 1857). These rings, also called Schlutte (Acker, 1982), are identical to theone depicted by Bock (1630) and described by Bauhin: apud nos ancillae circulos (qualisfigurae adpositus,) ad suppellectilem expoliendam conficiunt7 (Bauhin, 1658). Longdistance commercial trade of E. hyemale for use by craftsmen to polish wood and metalhas been reported from south-west Germany, where they were imported intoWürttemberg in great masses from the Rhine valley in Baden (von Martens & Kemmler,1882). French collectors apparently drew a heavy toll on the population, since Hausser(1894) complained that “a few decades ago” at Neubreisach the traders took cartloadsof E. hyemale, but that at the time of his writing they would have come in vain. Plantscollected near Strasbourg (dept. Bas-Rhin) were sold in Nancy, 120 km to the west(Braconnot, 1828; Kirschleger, 1857) and to Paris (Duval-Jouve, 1864). Halfwaybetween Strasbourg and Paris, in the Orient Forest near Troyes, it was available forcollecting, but, to the amazement of the Troyans, traders did not know this, or preferredto pull it in other places (des Étangs, 1841). Plumier tells us, presumably from Lyon8,that it is “a plant brought to us from the mountains”. In southern France commercialtrade existed where Villeneuve functioned as the hub selling horsetails collected in theLot-valley to the market in Bordeaux (de Saint-Amans, 1821).
32 FERN GAZ. 20(1):23-45. 2015
6“scouring pads of horsetails”
7at ours the maids make wreaths (illustrated adjacent) to polish the furniture
8The printer as well as the religious authorities approving the book resided in Lyon.

Yet one other possible origin should be considered, viz. North America. For this routeI have found no historical data, unless it would account for the surprisingly large exportvolume of “reeds” from the USA (Wolcott, 1795). However, the species is abundant inthe USA to the extent of becoming a nuisance (Millhollon, 1987), and the largerAmerican subspecies would explain observations of Newman and contemporaries on thesize of the marketed horsetails.
Newman expressed uncertainty about the Dutch Rushes of the London markets beingconspecific with English E. hyemale, and was supported in that opinion by other, notexplicitly named, botanists. His doubts arose from the “much larger size than any Britishexamples of E. hyemale” and “the much greater number of striæ, amounting in someinstances to thirty-two” (Newman, 1844). Large size and high number of ridges (Figure3) better conform to the American E. hyemale subsp. affine which has 14 – 50 ridgesthan the European subspecies with 14 – 26 ridges (Hauke, 1963). Johnson (Johnson &Sowerby, 1856), however, disagrees, stating that “British plant from Gamlingay Bog,Cambridgeshire, cultivated in my garden for thirty years, frequently rivals the imported“Dutch Rushes” in these respects, the number of ridges and smaller tubes varying fromtwenty to twenty-eight in the larger stems”. In the Zalkerbos near Zwolle, the only Dutchpopulation which historically has been associated with usage of the plants as smoothingagents, currently has stems of 1.20m tall with 25 – 26 ribs. Although such thick specimensof E. hyemale subsp. hyemale are memorable in itself, there is still a gap betweenNewman’s 32 ridges, and the Dutch and Johnson’s 26-28 ridges.
In the Netherlands, Van Hall (1854) wrote: “This plant does grow in some places ofour home country, in woods and sandy lands, but it is, as far as I have been able to findout, not used, and it is by far not as heavy as the one that occurs in commerce and isbrought in from elsewhere”. Herman van Hall, a student of Kops, was co-author of earliervolumes of the Flora Batava (Kops & Van Hall, 1828, 1832, 1836, 1844) and thereforelikely to be very well aware of the note given there, that E. hyemale is exported toEngland. His 1854 remark reads as if he has double-checked the fact, while bearing inmind that a different, larger taxon might be involved. Oudemans (1862) agreed: “Thereason why, both at ours and in England, the foreign shave-grass is preferred, seems tobe the latter’s thicker stems”.
DISCUSSIONAt the onset of the eighteenth century, the name Dutch Rush for E. hyemale had becomeso established that it started to show up in print, at first with authors addressing theartificers and amateurs who would actually be using it and, almost a century later, alsoin the scientific botanical literature. Something must have happened with the product forpeople to dub it with that new name. Later authors ascribe the name to imports fromHolland, but they all might have been deluded by the Schoenoplectusus/Equisetum-misconception, with the possible exception of John Smith (1802), who published inadvance of the earliest date so far discovered that shows the entanglement. The bestargument that Dutch import does explain the name is given by Aubrey’s 1691 letter.While he writes that the shave-grass comes from Holland, he does not use the nameDutch Rush, and therefore this is not likely to be an ad hoc explanation of that name.
As for the scale of the imports during the 150 years between Aubrey and Newman,again we must sift through the mystery caused by the Schoenoplectusus/Equisetum-misconception. All the imports of Dutch Rush, in “great quantities” or similar wording,are likely to be ascribed to bulrushes. In the eighteenth century, the Netherlands counted
DE WINTER: HISTORY AND MYTH OF THE EQUISETUM TRADE 33

over 4000 ha of land with culture of bulrushes (Maas, 2000) and this was an importantexport product. In 1827, 1600 loads of 63 bundles of bulrushes were declared at theBritish customs, accounting for over 360 times more duty revenues than received onpolishing rushes (calculations based on Parnell, 1830). If, in spite of Yeats’s opinion, E.hyemale would have been of any observable economic significance, one would haveexpected it to have been seen in the exceedingly detailed port statistics, where it is not.Evidently, half a year could pass without any horsetails being legally imported. Inaccordance with these incidental observations, it is absent from English and Dutch tradestatistics. The possibility that significant amounts of rushes have deliberately been keptfrom import registration is not very likely. Smugglers concentrated on low-volumecontraband where the import duty exceeded the intrinsic value. It is not unreasonable toassume that anything that was taxed was also smuggled, but 20% is a low tariff comparedwith that levied on alcohol and tobacco, so the incentive for smugglers to bring in shave-grass must have been slight, given that at some points in British history, smugglingcarried the death penalty (Richard Platt, pers. com.). The conclusion must be that evenif Holland has been an important source in the horsetail trade, the trade as a whole servedbut a niche market.
The emerging pattern of several disjunct historic observations is that of trade routesfrom scattered source regions into the most accessible larger centres of population. Forunknown reasons source populations closer at hand were ignored. Like Oudemans, DeSt.-Amans (loc. cit.) thinks it a matter of quality: “Celle des bords du Lot est belle etplus estimée”. Sustained exploitation sometimes resulted in depletion of local stocks. Inthe upper Rhine Valley an infrastructure of collectors and transporters had developedwho transported E. hyemale overland to France and downstream by river, as suggestedby the toll-treaties of German rivers (Anon., 1837, 1853). The Dutch had virtuallymonopolised navigation on the river Rhine (MacGregor, 1843). When brought all theway down to the sea, goods would have ended up in the port of Rotterdam, ideallysituated for further transport to Belgium and England. Since the species was “rathercommon in whole Europe, even in Russia, and especially in Sweden” (de Chalmot &Chomel, 1792), it may have been that the collectors took the product from wherever itwas available in north-west Europe, including the eastern part of the Netherlands andRussia (which then included Poland). Tariffs categorising horsetails are available frommany north and central European countries.
During the Napoleonic wars around 1800, reciprocal trade embargos causedinterruptions in the supply of indispensable commodities. Since the French invasion ofthe Netherlands in 1794, bulrushes from Holland had become difficult to obtain (Anon.,1819) and consequently become expensive: “during the non-commercial intercoursebetween Holland and Great Britain, which lasted a number of years, a great scarcityexisted in (…) Dutch Rushes, which were not to be had during the war, except in veryprecarious chance lots, occasionally smuggled over, at any price” (E. Smith, 1841). Thesame will have happened to E. hyemale from the continent and it may be hypothesisedthat in circumvention a new supply route from the United States was opened. Theaberrant size of the plants reported by nineteenth century authors indicates the NorthAmerican subspecies rather than the slenderer European form. If the shipping cost hadallowed a profitable transport, the availability would have been virtually unlimited fromthe largely uncultivated American lands where the species is abundant. However, doubtexists whether the economic opportunities were recognised: “The rushes used for rubbingdown are a valuable article, and grow abundantly in various parts of the United States,
34 FERN GAZ. 20(1):23-45. 2015

they are found in our own neighbourhood in the State of New Jersey; though extensivelyused in Europe they are scarcely known to our workmen” (Gill, 1828).
REFERENCESACHARD, C. F. 1785. Dictionnaire de la Provence et du Comté-Venaissin. 2 vols.
Vol. 2, Marseille; Jean Mossy.ACKER, A. 1982. Prêles, Encyclopédie de l’Alsace, 6146. Editions Publitotal,
Strasbourg.ANONYMOUS. 1350-1400. MS. Arch. Selden. B. 35, Bodleian, Oxford.ANONYMOUS. 1735. Goods imported and exported to and from the port of London,
from the 23rd Day of May, to the 16th Day of October inclusive, 1735. The PoliticalState of Great Britain 50: 450-456.
ANONYMOUS. 1749. Arts Companion, Or A New Assistant for the Ingenious.London, Dublin; I. Jackson.
ANONYMOUS. 1754. The British legacy: or, fountain of knowledge. . London;Thomas Chandler.
ANONYMOUS. 1796. Kannenkraut. In: HÖPFNER, L. J. F. [ed.], DeutscheEncyclopädie oder Allgemeines Real-Wörterbuch aller Künste und Wissenschaftenvon einer Gesellschaft Gelehrten, 97-102. Varrentrapp und Wenner, Frankfurt amMayn.
ANONYMOUS. 1819. Transactions of the Society, instituted at London for theEncouragement of Arts, Manufactures, and Commerce; with Ihe premiums offeredin the year 1817. The Literary Panorama and National Register 8 (New Series): 754-764.
ANONYMOUS. 1824. Sr. K.k. Majestät Franz des Ersten Politische Gesetze undVerordnungen für die Oesterreichischen, Böhmischen und Galizischen Erbländer.Vol. 50, Wien; K.K. Hof- und Staats-Druckerey.
ANONYMOUS. 1836. Sammlung der politischen Gesetze und Verordnungen für dasLaibacher Gouvernement-Gebiet im Königreiche Illyrien. Vol. 18, Laibach(Lubljana); Eger’schen Gubernial-Buchdruckerei.
ANONYMOUS. 1837. Tarif de Douanes concerté entre la Prusse, I’Electorat de Hesseet le Grandduché de Hesse d’une part, et la Bavière et le Wurtemberg de l’autre part.En date du 31. Oct. 1833. In: DE MARTENS, G. F. & MURHARD, F. [eds.],Nouveau recueil de traités d’Alliance, de Paix, de Trève, de Neutralité, deCommerce, de Limites, d’Echange etc, et de plusieurs autres actes servant à laconnoisancce des relations étrangères des Puissances et d’Etats de l’Europe., 736-777. Librairie de Dieterich, Göttingen.
ANONYMOUS. 1838. Zoll-Tariff für die Ein- und Ausfuhr der Waaren in demösterreichischen Kaiserstaate. Wien; der kaiserliche und königliche Hof- undStaats-Arearial-Druckerey.
ANONYMOUS. 1843. Verhandeling der Stuco, of zoogenaamd Kunstmarmer.Bouwkundig magazijn of schetsen voor handwerkslieden 1: 11-12.
ANONYMOUS. 1849a. Traité entre l’Autriche, la Prusse, la Saxe, le Hannovre, leDanemarc, le Mecklenbourg-Schwerin , les duchés d’Anhalt et les villes libresanséatiques de Hambourg et de Lübeck pour regulariser les droits de douaneshanovriennes payables à Brunshaus (ci-devant à Stade) Sur l’Elbe. Conclu et signeä Drèsde, le 13 Avril 1844. In: DE MARTENS, M. & MURCHARD, F. [eds.],Nouveau recueil général de traités: conventions et autres transactionsremarquables, servant à la Connaissance des Relations étrangères des Puissances
DE WINTER: HISTORY AND MYTH OF THE EQUISETUM TRADE 35

et États dans leurs Rapports mutuels, 473-525. Libraire de Dieterich, Göttingen.ANONYMOUS. 1849b. Acte additionel au traité du 23 Juin 1821 sur la navigation de
l’Elbe, signé à Drèsde le 13 Avril 1844, entre l’Autriche, la Prusse, la Saxe-royale,le Hanovre, le Danemarc, le Grand-duché de Mecklenbourg-Schwerin, les Duchésd’Anhalt-Coethen, d’Anhalt-Dessau et d’Anhalt-Bernbourg et les villes libresanséatiques de Hambourg et de Lübeck. (Gesetzsamml. für die Königl. PreussischenStaaten. Jahrg. 1844. Nro 37 v. 1. Novemb.). In: DE MARTENS, G. F. &MURCHARD, F. [eds.], Nouveau recueil général de traités: conventions et autrestransactions remarquables, servant à la Connaissance des Relations étrangères desPuissances et États dans leurs Rapports mutuels, 386-462. Libraire de Dieterich,Göttingen.
ANONYMOUS. 1853. Traité de commerce et de douanes entre l’Autriche et la Prusse,signé à Berlin, le 9 Février 1853. In: DE MARTENS, C. & DE CUSSY, F. [eds.],Recueil manuel et pratique de traités, conventions et autres actes diplomatiques,134-173. F.A. Brockhaus, Leipzig.
ANONYMOUS. 1856. Binnenlandsche Berigten. Nederlandsche staatscourant, 28-03-1856.
ANONYMOUS. 1858. Traité de commerce et de douanes entre la Prusse et l’Autriche)signé à Berlin, le 19 fébrier 1853. In: MARTENS, G. F. & SAMWER, C. [eds.],Nouveau recueil général de traités: conventions et autres transactionsremarquables, servant à la Connaissance des Relations étrangères des Puissanceset Etats dans leurs Rapports mutuels, 382-429. Libraire de Dieterich, Göttingen.
ARNTZEN, K. A. 1830. Love, Anordninger, Kundgjørelser, aabne Breve, Resolutioner,m.m., der vedkomme Kongeriget Norges Lovgivning og offentlige Bestyrelse, foraaret 1829. Christiania; Chr. Grøndahl.
ARTLOVE, M. 1730. The art of japanning, Varnishing, Pollishing, and Gilding. Beinga collection of very plain directions and receipts. Written for the Use of those whohave a mind to follow those diverting and useful Amusements, and Published at theRequest of several Ladies of Distinction. London; T. Warner.
AUBREY, J. 1847. The Natural History of Wiltshire. Wiltshire Topographical Society.AUBREY, J. 1848. Letter to Mr. Ray. In: LANKESTER, E. [ed.], The Correspondence
of John Ray, 237-238. The Ray Society, London.BAERT, J. 1943. Jan Kops, pionier van Hollands landbouw. ‘s Gravenhage; MArtinus
Nijhoff.BAILEY, N. 1756. Universal Etymological English Dictionary. 4th ed. Vol. 2, London;
T. Waller.BARROW, J. 1735. Dictionarium polygraphicum: or, the Whole Body of Arts Regularly
Digested. Vol. II, London; C. Hithch & C. Davis.BAUHIN, G. 1658. Caspari Bauhini theatri botanici. Vol. 1, Basel; Joannes König.BIJLSMA, R. J. 2003. Bosrelicten op de Veluwe. Een historisch-ecologische
beschrijving. Alterra, Wageningen, report no. 647.BOCK, H. & SEBISCH, M. 1630. Kräutterbuch Weylandt des Weitberhüm[m]ten vndt
Hocherfharnen Herren Hieronymi Tragi genant Bock. Auff Vielfaltiges begeren vndfreundtliches ersuchen Mit vleiss übersehen, vnd mit der Kräutter zunamen, derNamen ursachen, qualitete[n], deren gradibus, gebrauch in den Apotecken, vndsehr vielen in[n]erlichen vnd eusserlichen experimentis gemheret und gebessert.Durch Melchiorem Sebizium, der Artzeney Doctorem, vnd Professorem, auch derStatt Strassburg bestellten Physicum. Sampt Angehenckten Speisskammer ermelten
36 FERN GAZ. 20(1):23-45. 2015

herrn H. Bocken, darin[n] aber für dissmhal nichts geändert worde[n]. 4th ed.Strassburg; Wilhelm Christian Glasern.
BOLTON, J. 1790. Filices Britannicae. 2 vols. Vol. 2, London.BRACONNOT, H. 1828. Recherches chimiques sur la nature des Prêles. Annales de
chimie et de physique 39: 5-24.BRÉVAL, J. D. 1726. Remarks on several parts of Europe: relating chiefly to the
History, Antiquities and Geography, of those countires through which the author hastraveled : as France, the Low Countries, Lorrain, Alsacia, Germany, Savoy, Tyrol,Switzerland, Italy and Spain. Vol. 1, London; Bernard Lintot.
BURN, R., CHITTY, J. & CHITTY, T. 1831. The Justice of the Peace and ParishOfficer. 6 vols. Vol. 2, London; S. Sweet & A. Maxwell.
C.K., 1701. Art’s master-piece: or, A companion for the ingenious of either sex. In twoparts. London; G. Gonyers.
CAMDEN, W. 1722. Britannia: Or a Chorographical Description of Great Britain andIreland (etc.) Written in Latin and Translated Into English. 2nd ed. Vol. 1, London;Awnsham Churchhill.
CLEVERING, O. A. & VAN GULIK, W. J. M. 1990. De aanleg vanbiezenbegroeiingen: mogelijkheden voor her-introduktie van biezen in hetHaringvliet - Hollandsch Diepgebied. Oostvoorne; Instituut voor OecologischOnderzoek.
COMELIN, J. 1683. Catalogus plantarum indigenarum Hollandiae, cui praemissaLamb. Bidloo dissertatio de re herbaria. Amsterdam; Henr. & Viduam Theod.Boom.
DE CHALMOT, J. A. & CHOMEL, N. 1792. Vervolg op M. Noël Chomel Algemeenhuishoudelyk-, natuur-, zedekundig- en konst-woordenboek. Vol. agtste deel, zijndehet XV. deel van het woordenboek, Campen; J.A. de Chalmot.
DE GORTER, D. 1781. Flora VII provinciarum Belgii foederati indigena. Haarlem;Bohn & Filius.
DE HAAN, W. A., VAN DE PAVERD, M., VAN RIJNSBERGEN, J., LEENDERS, P.H. J. A., WORMGOOR, J. A. & VAN GELDER, P. H. A. J. M. 2001. Dijken alsonderdeel van een systeem van bescherming tegen overstromingen. Delft; DelftUniversity.
DE MARTENS, G. F. & MURHARD, F. 1836. Publication officielle du Tarif deDouanes concerté entre la Hesse électorale d’une part et entre la Prusse et la Hessegrandducale de l’autre part, en exécution du Traité de douanes et de commerce,conclu le 25 Août 1831 entre les dits Etats. En date du 30 Décembre 1831. (Kurhess.Gesetz-Sammlung 1831. Dezemb. Nr.XXII.), Supplément au Recueil de principauxtraités d’Alliance, de Paix, de Trêve, de Neutralité, de Commerce, de Limites,d’Echange etc conclus par les puissance de l’Europe tant entre elles., 495-521.Librairie de Dieterich, Göttingen.
DE SAINT-AMANS, J. F. B. 1821. Flore Agenaise ou Description Methodique DesPlantes observées dans le département de Lot-et-Garonne et dans quelques partiesdes départements voisins. Agen; Prosper Noubel.
DE VILLENEUVE, C. 1821. Statistique du département des Bouches-du-Rhône: avecAtlas Vol. 1, Marseille; Antoine Ricard.
DE VRIES, H. 2008. De laatste biezentelers. Reformatorisch Dagblad, 10-10-2008.DE VRIES, H., VAN DER SANDE LACOSTE, C. M. & SURINGAR, W. F. R. 1870.
Flora, Algemeene statistiek van Nederland: beschrijving van den maatschappelijken
DE WINTER: HISTORY AND MYTH OF THE EQUISETUM TRADE 37

toestand van het Nederlandsche volk in het midden der negentiende eeuw, 172-241.Vereeniging voor de Statistiek in Nederland / A.W. Sijthoff, Leiden.
DE WINTER, W. P. 2007. What’s Dutch about the Dutch rush? The Pteridologist 4: 195-197.
DE WINTER, W. P. 2012. In Nomine Equiseti. Varen-Varia 25: 4-7.DE WINTER, W. P. & LUBIENSKI, M. 2012. Equisetum ×ascendens Lubienski &
Bennert: de eerste triploïde paardestaartbastaard in Nederland. Gorteria 36: 1-17.DEPARTEMENT VAN FINANCIËN. 1848. Statistiek vand den handel en de
scheepvaart van het Koningrijk der Nederlanden over het jaar 1846. ‘s-Gravenhage; Departement van Financiën.
DES ÉTANGS, S., ed. 1841. Notes sur quelques plantes observées dans le départementde l’Aube.Mémoires de la Société no. 77-78. Troyes; Société académiqued’agriculture, des sciences, arts et belles-lettres du département de l’Aube.
DOSSIE, R. 1758. The handmaid to the arts. London; J. Nourse.DU MORTIER, B. 1869. Bouquet du littoral belge. Bulletin de la Societe de Botanique
de Belgique 7: 318-371.DUJARDIN-SAILLY, E. 1813. Tarif chronologique des douanes de l’empire française:
avec des explications. 8th ed. Paris; Dujardin-Sailly.DUVAL-JOUVE, J. 1864. Histoire Naturelle des Equisetum de France. Paris; J.B.
Baillière et Fils, Libraires de ‘Académie Impériale de Médicine.ELLIS, R. 1837. The Laws and practical Regulations forming the water-side and
general Practice of the Customs. The Laws and practical Regulations of theCustoms. 4 vols. Vol. 3, London; O.H. Bailey & Co.
ESTERLY, D. 1998. Grinling Gibbons and the Art of Carving. New York; HarryAbrahams Inc.
FLEXNER, S. B. & HAUCK, L. C. 1993. Random House unabridged dictionary. 2nded. New York; Random House.
FLIERVOET, L. M. 1992. Aanleg en beheer van grasland op rivierdijken. Den Haag;Unie van Waterschappen etc.
FRANCIS, G. W. 1851. An Analysis of the British Ferns and their Allies. London;Simpkin, Marshall & Co.
FUCHS, L. 1543. Den nieuwen Herbarius, dat is dat boeck van den cruyden, int welckemet groote neersticheyt beschreven is niet alleen die gantse historie, dat is, dienamen, tfaetsoen, natuere, cracht ende operatie van meesten deel de cruyden, diehier ende in anderlanden wassende sijn, midtsgaders der plecken oft stede, ende dentijt waer ende wanneer sij wassen, maer oock allede wortelen, stele, bladeren,bloemen, saet, vruchten, ende insumma dat gantse wesen ende gelegenheyt van denselven, is hierinne so constelijcken ende natuerlicken na dleven gefigureert endegeconterfeyt, dat desghelijcks voormaels noyt ghesien en heeft gheweest, noch oytinde printe ghecomen en is. Basel; Michel Insingrin.
GERARDE, J. 1597. The Herball or Generall Historie of Plantes. London; JohnNorton.
GILL, T. 1822. The Technical repository. Vol. II, London; T. Cadell.GILL, T. 1828. Gill’s technological repository. 6 vols. Vol. 2, London; Thomas Hurst,
Edward Chance & Company.GISKES, J. H. 1979. Jaarboeken van het genootschap Amstelodanum 71: De
vioolmakersfamilies Boumeester en Menslage te Amsterdam. Debussy EllermanHarms bv., Amsterdam.
38 FERN GAZ. 20(1):23-45. 2015

GLOVER, S. 1843. The history and directory of the borough of Derby, a guide. Derby;S. Clover & Son.
GRAFF, E. G. 1838. Althochdeutscher Sprachschatz: oder Wörterbuch deralthodeutschen Sprach. Vol. 4, Berlin; Beim Verfasser und in Commission derNikolaischen Buchhandlung.
HAUKE, R. A. 1963. A taxonomic review of the genus Equisetum subgenusHippochaete. Beiheft Nova Hedwigia B: 1 - 123.
HAUSSER, E. 1894. Auf welche Weise fördern wir die Kenntnis unserer einheimischerFlora? Mitteilungen der Philomathischen Gesellschaft in Elsaß-Lothringen 2: 19-25.
HEADRICK, J. 1813. General View of the Agriculture of the County of Angus, orForfarshire; With Observations on the Means of Its Improvement. Edinburgh; Boardof Agriculture, Great Britain.
HELDRING, O. G. 1845. Eene vergelijking tusschen Geldersche en Noord hollandschezeden, Geldersche volksalmanak, 71-101. G. van Eldik Thieme, Arnhem.
HELDRING, O. G., VAN ELDIK THIEME, G. & NIJHOFF , I. A. 1837. Gedachtenvan Volksvrienden, Geldersche volksalmanak, (s.n.). C.A. Thieme, Arnhem.
HENSLOW, J. S. 1839. Description and Physiological Botany. new edition ed.London; Longman &c.
HEUKELS, H. 1907. Woordenboek der Nederlandsche volksnamen van planten: uit degegevens verzameld door de Commissie voor Nederlandsche plantennamen. ‘s-Gravenhage; Versluys.
HINDERWELL, T. 1811. The history and antiquities of Scarborough and the vicinity.2nd ed. (1st ed. 1798) ed. York; Longman et al.
HOOKER, W. J. 1821. Flora scotica, or, A description of Scottish plants. London;Archibald Constable etc.
HOUGHTON, J. 1728. A collection for the improvement of husbandry and trade:Consisting of many valuable materials relating to corn, cattle, coals, hops, wool,&c. With a compleat catalogue of the several sorts of earths, ... With many otheruseful particulars, communicated by several eminent members of the Royal Society,to the collector, John Houghton, F.R.S. Now revised, corrected, and published, witha preface and useful indexes, by Richard Bradley, ... In three volumes. London;Woodman and Lyon.
HULTÉN, E. 1962. The circumpolar plants. 1, Vascular cryptogams, conifers,monocotyledons. . Handlingar / Kungl. Svenska vetenskapsakademiens (Fjaerdeseries). Vol. 4:8:5, Stockholm; Almqvist & Wiksell.
INTRAVAIA, L. J. & RESNICK, R. S. 1965. A Research Study of a Technique forAdjusting Clarinet Reeds. Journal of Research in Music Education 16: 45-58.
JAMES, R. J. 1745. A Medicinal Dictionary, Including Physic, Surgery, Anatomy,Chymistry, and Botany, in All Their Branches Relative to Medicine; Together with aHistory of Drugs, an Account of Their Various Preparations, Combinations, andUses; and an Introductory Preface, Tracing the Progress of Physic and Explainingthe Theories Which Have Principally Prevail’d in All Ages of the World. 3 vols. Vol. 2, London; T. Osborne.
JOHNSON, C. & SOWERBY, J. E. 1856. The Fern Allies: a supplement to the Fernsof Great Britain. London; J.E. Sowerby.
KALIBAN’S GROCERY AND MARKET. 1938. Advertisement of Kaliban’s Groceryand Market. The mount Vernon Hawkeye-Record and the Lisbon Herald, 24 Nov.1938.
DE WINTER: HISTORY AND MYTH OF THE EQUISETUM TRADE 39

KIRSCHLEGER, F. 1857. Flore d’Alsace. 3 vols. Vol. 2, Strasbourg, Paris; VictorMasson etc.
KLENNER, F. W. 1822. Allgemeiner Zoll-Tariff für den österreichischen Kaiserstaat.147 vols Wien; der k.k. Hof- und Staats-Arearial-Druckerey.
KNIGHT, C. 1833. Vegetable Substances: materials of manufacturers. The Library ofEntertaining Knowledge. London; Society for the Diffusion of Useful Knowlledge.
KONINKRIJK DER NEDERLANDEN. 1816. Wet van den derden October 1816 overde heffing der regten van in- en uitvoer: benevens het tarief derzelve regten voor hetKoningrijk der Nederlanden. P. den Hengst & Zoon, ‘s-Gravenhage & Amsterdam.
KONINKRIJK DER NEDERLANDEN. 1822. Wet van den 26. Augustus 1822,houdende het Tarief van regten op den In-, Uit- en Doorvoer van alle goederen,waren en koopmanschappen. Staats-Secretarij, ‘s-Gravenhage.
KOPS, J. 1798. Algemeen rapport der Commissie van superintendentie over hetonderzoek der duinen; vol.1: Tegenwoordige staat der duinen van het voormaaliggewest Holland. 2 vols. Vol. 1, Leyden; Herdingh en Du Mortier.
KOPS, J., ed. 1807. Staat van den landbouw in het Koninkrijk van Holland gedurendeden jare 1806. Haarlem; Loosjes.
KOPS, J., ed. 1808. Staat van den landbouw in het Koninkrijk van Holland gedurendeden jare 1807. Haarlem; Loosjes.
KOPS, J., ed. 1809. Staat van den landbouw in het Koninkrijk van Holland gedurendeden jare 1808. Haarlem; Loosjes.
KOPS, J., ed. 1816. Staat van den landbouw in de Vereenigde Nederlanden gedurendeden jare 1814. Den Haag; Algemeende ‘s Lands Drukkerij.
KOPS, J., ed. 1819. Staat van den landbouw in het Koninkrijk der Nederlanden,(Noordelijke provincien) gedurende het jaar 1815. Brussel; Stapleaux.
KOPS, J., ed. 1821. Staat van den landbouw in het Koninkrijk der Nederlanden,(Noordelijke provincien) gedurende het jaar 1818. Brussel; Stapleaux.
KOPS, J., ed. 1822a. Staat van den landbouw in het Koninkrijk der Nederlanden,(Noordelijke provincien) gedurende het jaar 1821. Brussel; Stapleaux.
KOPS, J., ed. 1822b. Staat van den landbouw in het Koninkrijk der Nederlanden,(Noordelijke provincien) gedurende het jaar 1820. Brussel; Stapleaux.
KOPS, J., ed. 1822c. Staat van den landbouw in het Koninkrijk der Nederlanden,(Noordelijke provincien) gedurende het jaar 1819. Brussel; Stapleaux.
KOPS, J., ed. 1829. Staat van den landbouw in de Vereenigde Nederlanden gedurendehet jaar 1828. ‘s Gravenhage; Algemeende Lands Drukkerij.
KOPS, J. & VAN HALL, H. C. 1828. Flora Batava of Afbeelding en Beschrijving vanNederlandsche Gewassen. Vol. 5, Amsterdam; J.C. Sepp en Zoon.
KOPS, J. & VAN HALL, H. C. 1832. Flora Batava of Afbeelding en Beschrijving vanNederlandsche Gewassen. Vol. 6, Amsterdam; J.C. Sepp en Zoon.
KOPS, J. & VAN HALL, H. C. 1836. Flora Batava of Afbeelding en Beschrijving vanNederlandsche Gewassen. Vol. 7, Amsterdam; J.C. Sepp en Zoon.
KOPS, J. & VAN HALL, H. C. 1844. Flora Batava of Afbeelding en Beschrijving vanNederlandsche Gewassen. Vol. 8, Amsterdam; J.C. Sepp en Zoon.
KOPS, J. & VAN DER TRAPPEN, J. E. 1846. Flora Batava of Afbeelding enBeschrijving van Nederlandsche Gewassen. Vol. IX, Amsterdam; J.C. Sepp enZoon.
KRÜNITZ, J. G. 1785. Kannen-kraut, Kandelkraut, Kandelwisch. In: KRÜNITZ, J. G.,FLOERKEN, F. J., FLÖRKE, H. G., KORTH, J. W. D., HOFFMANN, C. O. &
40 FERN GAZ. 20(1):23-45. 2015

KOSSARSKI, L. [eds.], Oekonomisch-technologische Encyklopädie, oderallgemeines System der Stats- Stadt- Haus- und Land-Wirthschaft, und der Kunst-Geschichte, in alphabetischer Ordnung, 146-183. Joachim Pauli, Berlin.
LANCASTER, J. 1887. Alphita, a medico-botanical glossary from the Bodleianmanuscript, Selden B. 35. Oxford; Clarendon Press.
LAW OLMSTEDT, F., COVILLE, F. V. & KELSEY, H. P. 1924. Standardized plantnames : a catalogue of approved scientific and common names of plants inAmerican commerce. Salem; American Joint Committee on HorticulturalNomenclature.
LE FRANCQ VAN BERKHEIJ, J. 1780. Antwoord op de vraage, voorgesteld door deHollandsche Maatschappye der Weetenschappen, te Haarlem: Welke Boomen,Heesters en Planten zyn ‘er, behalve den Helm ( Arundo Arenaria) en den Sleedoorn(Prunus Sylvestris), dewelke op de Zandduinen ter weeringe der Zandverstuivingekunnen geplant worden ? Kan men ook eenige andere Planten aan onze Zee-stranden met voordeel gebruiken ? Zyn daar mede hier te Lande al eenigeproeven gedaan ? En welke is de uitkomst daar van geweest ? Verhandelingenuitgegeeven door de Hollandse Maatschappye der Weetenschappen te Haarlem.XIX: 1-80.
LEE, M. R. 2001. William Withering (1741-1799): A Birmingham Lunatic. Proceedingsof the Royal College of Physicians of Edinburgh 31: 77-83.
LEMAN, M. 1826. Prêle. In: CUVIER, F. [ed.], Dictionnaire des sciences naturelles.F.G. Levrault, Strasbourg, Paris.
LOUDON, J. C. ed. 1829. An encyclopædia of plants: comprising the description,specific character, culture, history, application in the arts, and every other desirableparticular respecting all the plants indigenous, cultivated in, or introduced toBritain, combining all the advantages of a Linnean and Jussieuean speciesplantarum, an historia plantarum, a grammar of botany, and a dictionary of botanyand vegetable culture. The whole in English; with the synonymes of the commonerplants in the different European and other languages. London; Longman, Rees,Orme, Brown & Green.
MAAS, G. J. 2000. Historische geomorfologie Maas en benedenrivieren : Oude Maas,Merwede-Hollandse Biesbosch, Afgedamde Maas en Maaskant. Alterra-rapport;075. Wageningen; Alterra, Research Instituut voor de GroeneRuimte.
MACFARQUHAR, C. & GLEIG, G. 1797. Encyclopædia Britannica: Or, A Dictionaryof Arts, Sciences, and Miscellaneous Literature. Edinburgh; A. Bell & C.Macfarquhar.
MACGREGOR, J. 1843. Commercial statistics: A digest of the productive resources,commercial legislation, customs tariffs, of all nations. Including all Britishcommercial treaties with foreign states. 1st. ed. 2 vols. Vol. 1, London; Whittaker &Co.
MACGREGOR, J. 1850. Commercial statistics: A digest of the productive resources,commercial legislation, customs tariffs, of all nations. Including all Britishcommercial treaties with foreign states. 5 vols. Vol. 2, London; Whittaker & Co.
MARTIN, T. 1813. The circle of the mechanical arts; containing practical Treatises onthe various manual Arts, Trades, and Manufactures. London; Richard Rees.
MEASOM, G. S. 1861. Official Illustrated Guide to the Great Northern, Manchester,Sheffield, and Lincolnshire, and Midland Railways. London; Griffin Bohn.
DE WINTER: HISTORY AND MYTH OF THE EQUISETUM TRADE 41

MEESE, D. 1768. Antwoord op de vraag, door de Hollandsche Maatschappy derWeetenschappen te Haarlem, in den jaare 1764. Voor de eerfte, en in het jaar 1766.Voor de tweedemaal opgegeeven , om voor het begin van het jaar 1767. Tebeantwoorden: welke zyn de beste en minst kostbaare middelen, om het afnemender oevers van het Haarlemmer Meer te beletten? Verhandelingen uitgegeeven doorde Hollandse Maatschappye der Weetenschappen te Haarlem. 10: 1-46.
MICHEL, J. E. 1802. Statistique du département des Bouches du Rhône. Paris;Hachette.
MILLER, P. 1745. Groot en algemeen kruidkundig, hoveniers, en bloemistenwoordenboek, behelzende de manier om moes-, bloem-, vrugt-, kruid-tuinen, wild-bossen, wijngaarden, Oranje-huizen, stook-kassen, enz. aanteleggen, enz. enallerlei gewassen te kweeken [...]. II vols. Vol. I, Leiden; Pieter vander Eyk, en deweduwe Jakob vander Kluis.
MILLER, P. 1754. The Gardeners Dictionary: Containing the Methods of Cultivatingand Improving All Sorts of Trees, Plants, and Flowers, for the Kitchen, Fruit, andPleasure Gardens; as Also Those which are Used in Medicine. With Directions forthe Culture of Vineyards, and Making of Wine in England. In which Likewise areIncluded the Practical Parts of Husbandry. 3 vols. Vol. II, London; John & JamesRivington.
MILLHOLLON, R. W. 1987. Control of Equisetum hyemale on drainage ditches insugarcane. Journal of the American Society Sugar Cane Technologists 7: 57-64.
MONTIN, L. 1771. Bemerkungen über den Anbau des Flugsandes. Der Königl.Schwedischen Akademie der Wissenschaften neue Abhandlungen aus derNaturlehre, Haushaltungskunst und Mechanik 30: 276-283.
MOORE, T. 1861. British Ferns and their Allies. London; George Routledge and Sons.MURRAY, J. A. H., ed. 1971. Compact edition of the Oxford English dictionary,
complete text reproduced, micrographically. 2 vols. Vol. 2; Oxford; OxfordUniversity Press.
NEWMAN, E. 1842. Art. LXXVI. - A History of the British Equiseta. The Phytologist2: 273-278.
NEWMAN, E. 1844. A History of British Ferns and Allied Plants. Second (revised)edition. London; John van Voorst.
NOEL CHOMEL, M. & DE CHALMOT, J. A. 1778. Algemeen huishoudelijk-, natuur-, zedekundig-, en konst- woordenboek. 2nd. ed. Vol. 4, Joh. le Mair.
NOZEMAN, C. 1783. Verhandeling over heermoes, unjer of kattestaart (Equisetum).Antwoord op de vraeg der Maatschappy ter Bevordering vav de Landbouw: Welkezyn de Eigenschappen van de verschillende soorten van het Equisetum, by onsbekend onder den naam van Heermoes, Unjer, of Kattestaart ? Waarin bestaat de aarten hoedanigheid van deszelfs Vruchtdeelen en Voortplanting? Welk nadeel wordtdoorhetzelve aan het Wei en Bouw- land toegebragt ? En welk is het door deondervinding beproefde beste middel om hetzelve op de minst kostbaare wyzeuitteroeijen ? Verhandelingen uitgegeeven door de Maatschappy ter Bevorderingvan de Landbouw 2: 1-66.
ØLLGAARD, B. & TIND, K. 1993. Scandinavian Ferns. Copenhagen; Rhodos,International Science & Art Publishers.
OUDEMANS, C. A. J. A. 1862. De natuurlijke historie van Nederland : de flora vanNederland. 1st ed. ed. 3 vols. Vol. 3, Haarlem; A.C. Kruseman.
PAGE, C. N. 1997. The Ferns of Britain and Ireland. Cambridge; Cambridge
42 FERN GAZ. 20(1):23-45. 2015

University Press.PARNELL, H. 1830. On Financial Reform. London; J. Murray.PARNELL, H. 1831. On Financial Reform. 3rd ed. London; John Murray.PERNETY, D. 1771. The history of a voyage to the Malouine, or Falkland, islands,
made in 1763. London; T. Jeffrys.PEXENFELDER, M. 1670. Apparatus eruditionis tam rerum quam verborum per
omnes artes et scientias. Nürnberg; Michael & Joh. Friedrich Endter.PHILLIPS, R. 1818. Book of english trades and library of the useful arts.New Enlarged
ed. London; J. Souter, at the Juvenile Library.PRADA, C. 1986. Equisetum. In: CASTROVIEJO, S., LAÍNZ, M.,
LÓPEZ-GONZÁLEZ, G., MONTSERRAT, P., MUÑOZ GARMENDIA, F.,PIAVA, J. & VILLAR, L. [eds], Flora Iberica, 21-29. Real Jardín Botánoco,C.S.I.C., Madrid.
PRATT, A. 1846. Wild flowers of the year. London; The Religious Tract Society.PRATT, A. 1866. The Flowering Plants, Grasses, Sedges, and Ferns of Great Britain.
Vol. VI: British grasses, sedges and ferns. London; Frederick Warne & Co.RIJNLAND, H. V. 2003. Waterkeringsbeheerplan 2003-2007 - Katern 3:
Beplantingsbeleid. Leiden, report no.RUPPE, H. B. & HALLER, A. 1745. Flora Jenensis Henrici Bernhardi Ruppii, ex
posthumis auctoris schedis et propriis observationibus aucta et emendata. Jena;C.H. Cuno.
RUTZ, L. M. & FARRAR, D. R. 1984. The habitat characteristics and abundance ofEquisetum × ferrissii and its parent species, Equisetum hyemale and Equisetumlaevigatum, in Iowa. American Fern Journal 74: 65-76.
SALVO TIERRA, E. 1990. Guía de helechos de la Península Ibérica y Baleares.Madrid; Pirámide,.
SCHRAVER, A. 1807. Antwoord op de vraag: Is ‘er eenig middel, om het Kram- enMat-werk aan de zeedijken, het zij met daartoe geschikte kruiden, heesters, ofandere gewassen, tusschen hetzelve te planten; het zij op eenige andere wijze,duurzamer, en tegen den slag van ‘t zeewater meer bestand ie maken ? NieuweVerhandelingen Van Het Zeeuwsch Genootschap der Wetenschappen 1: 67-131.
SMITH, E. 1841. Proposed cultivation of the Dutch species of rushes in England &c.The Mechanics’ Magazine, Museum, Register Journal, and Gazette 35: 34-35.
SMITH, J. E. 1793. A Sketch of a Tour on the Continent, in the Years 1786 and 1787.Vol. 1, London; James Edward Smith.
SMITH, J. E. 1802. English Botany; or, coloured figures of British Plants, with theiressenntial characters, synonyms, and places of growth, to which will be addedoccasional remarks. Vol. XIV, London; J. Sowerby.
SOCIETY FOR THE DIFFUSION OF USEFUL KNOWLEDGE. ed. 1833. Vegetablesubstances: materials of manufactures.The Library of Entertaining KnowledgeLondon; C. Knight.
SPENGLER, G. C. 1891. Het vastleggen en opbosschen van duinen enzandverstuivingen. Ede; Menger.
STEEL, D. 1796. New and complete tables of the net duties payable, and drawbacksallowed, on goods, wares, and merchandises, imported into, exported from, andcarried coastwise in, Great-Britain, agreeable to the act of the 27th Geo. III. calledthe Consolidation-Act, and corrected to the year 1796. London; D. Steel.
STEWART, J. A. 1815. The young woman’s companion; or, Female instructor. Oxford;
DE WINTER: HISTORY AND MYTH OF THE EQUISETUM TRADE 43

Bartlett & Newman.SYKORA, K. V. & LIEBRAND, C. I. J. M. 1987. Natuurtechnische en civieltechnische
aspecten van rivierdijkvegetaties. Wageningen; Landbouwuniversiteit.TAILLARD, P.-A. & DALMONT, J.-P. 2012. Scraping technique for clarinet reeds
derived from a static bending simulation. Proceedings of the Acoustics 2012 NantesConference, Nantes, France: 2735-2740.
TARGIONI TOZZETTI, O. 1813. Istituzioni botaniche del dottore Ottaviano TargioniTozzetti. 3rd ed. Vol. III, Firenze; Guglielmo Piatti.
TECHNISCHE ADVIESCOMMISSIE VOOR DE WATERKERINGEN. 1985.Leidraad voor het ontwerpen van rivierdijken. 2 vols. Vol. 1, Den Haag;Staatsuitgeverij.
TRIMMER, J. 1842. Practical chemistry for farmers and landowners. London; John W.Parker.
TURNER, W. 1538. Libellus de herbaria novus, in quo herbarum aliquot nominagreca, latina, & Anglica habes, una cum nominibus officinarum, in gratiam studioseinventutis nunc primum in lucem aeditus. William Turner : Libellus de re herbaria,1538. The names of herbes, 1548. London; Ray Society, facsimile 1965.
VAN DER AA, A. J., ed. 1851. Aardrijkskundig woordenboek der Nederlanden.13 vols.Vol. 13 (Z & app.). Gorinchem; J. Noorduyn.
VAN EYS, J. N., REINHART VAN MARLE, C. G., TEMMINCK, M., VAN DERGOES, F. & VAN STYRUM, J. 1799. Ontwerp tot vruchtbaarmaking der duinen:zijnde het tweede deel van het Algemeen Rapport der Commissie vanSuperintendentie over het Onderzoek der Duinen. 2 vols. Vol. 2, Leyden; Herdinghen Du Mortier.
VAN GEUNS, S. J. 1789. Antwoord op de vraag, voorgesteld door de HollandscheMaatschappye der Weetenschappen, te Haarlem: Welken zyn de onderwerpenbetreffende de Natuurlyke Historie onzes Vaderlands, waarvan men met gegrondereden te verwachten hebbe, dat eene verdere naspooring ten nutte van het Vaderlandverstrekken zal? Verhandelingen uitgegeeven door de Hollandse Maatschappye derWeetenschappen te Haarlem. XXVI: 231-316.
VAN HALL, H. C. 1843. De Heermoes of kwade aard en de middelen tot hareverdelging voorgeslagen. Tijdschrift ter bevordering van Nijverheid 8: 1.
VAN HALL, H. C. 1854. Neerlands plantenschat, of landhuishoudkundige flora,behelzende eene beschrijving der onkruiden, vergiftige en nuttige inlandscheplanten en der in onzen landbouw gekweekte gewassen. Leeuwarden; G.T.N.Suringar.
VAN HALL, H. C., MIQUEL, F. A. W. & DASSEN, M. 1832. Flora belgiiseptentrionalis, sive index plantarum indigenarum, quae hucusque in 7 provinciisfoederatis repertae sunt = Flora van Noord-Nederland, of korte aanwijzing der inhet wild wassende planten, welke tot nu toe in de voormalige 7 vereenigde gewestengevonden zijn. Pars 1: Equisetaceae, Filices, Marsiliaceae, Lycopodiaceae, Musciet Hepaticae = Bedektbloeijende planten, (paardestaartigen, varens, wortelzadigen.2 vols. Vol. 2, Amsterdam; J.C. Sepp.
VAN MEERTEN-SCHILPEROORT, A. B. 1830. Een theekistje à la milionade of à lachinoise. Penélopé, of Maandwerk aan het vrouwelijk geslacht toegewijd 6: 85-87.
VON MARTENS, G. M. & KEMMLER, C. A. 1882. Flora von Württemberg undHohenzollern. 3rd ed. 2 vols. Vol. 2, Heilbronn; Gebr. Henniger.
VUYCK, L. 1898. De plantengroei der duinen: overzicht van het voorkomen der
44 FERN GAZ. 20(1):23-45. 2015

planten in het duingebied en de veranderingen die in de flora zijn opgetreden in delaatste vijftig jaar in het bijzonder met het oog op den aanleg vanduinwaterleidingen. PhD, Leiden University, Leiden.
WARDLAW, A. C. & LEONARD, A., eds. 2005. New Atlas of Ferns & Allied Plantsof Britain & Ireland. BPS Special Publication no. 8. London; The BritishPteridological Society.
WEBSTER, N. 1913. Webster’s Revised Unabridged Dictionary of the EnglishLanguage. G. & C. Merriam Co.
WEEDA, E. J., WESTRA, R., WESTRA, C. & WESTRA, T. 1985. NederlandseEcologische Flora. 5 vols. Vol. 1, Amsterdam; IVN.
WEEDA, E. J., WESTRA, R., WESTRA, C. & WESTRA, T. 1994. NederlandseEcologische Flora. 5 vols. Vol. 5, Amsterdam; IVN.
WHITWORTH, C. 1777a. A register of the trade of the port of London; specifying thearticles imported and exported, arranged under the respective countries; with a listof the ships entered inwards and cleared outwards. Number I. January, February,and March, 1776. London; G. Kearsly, W. Flexney, G. Robinson, J. Robson, J.Walter, T. Cadell, J. Sewell.
WHITWORTH, C. 1777b. A register of the trade of the port of London; specifying thearticles imported and exported, arranged under the respective countries; with a listof the ships entered inwards and cleared outwards. Number II. April, May, and June,1776. London; G. Kearsly, W. Flexney, G. Robinson, J. Robson, J. Walter, T. Cadell,J. Sewell.
WILKINSON, C. C. 1858. Weeds and wild flowers: their uses, legends, and literature.London; John van Voorst.
WITHERING, W. 1776. A botanical arrangement of all the vegetables naturallygrowing in Great Britain. 1st ed. 2 vols. Vol. 2, London; T. Cadell etc.
WITHERING, W. 1792. A botanical arrangement of British Plants. 2nd ed. 3 vols. Vol. 3, Birmingham; G.G.J. & J. Robinson, etc.
WOLCOTT, O. 1795. Abstract of the Goods, Wares, and Merchandize, exported fromthe United States, from the 1st October 1793, to the 30th September 1794.Philadelphia; Francis Childs.
YEATS, J. 1870. The natural history of commerce. London; Cassell, Petter & Galpin.
DE WINTER: HISTORY AND MYTH OF THE EQUISETUM TRADE 45

SHORT NOTE
PLAGIOGYRIA MINUTA IS DISTINCT FROM P. EGENOLFIOIDESVAR. EGENOLFIOIDES
Two fern collections from Gunung Gaharu, Serian District, 1st Division, Sarawak, onthe border with Kalimantan in Borneo, have usually been filed with grammitid ferns inherbaria. They are Anderson SAR 15696 and Ilias & Azahari SAR 35673, both seen in Kand SAR, and said to be in L also. Both collections grew on sandstone cliffs, at 610 mand 790 m respectively, and are conspecific. They differ from grammitids in numerousmorphological characters, however, and actually belong to Plagiogyria.
The Flora Malesiana treatment of Plagiogyria (Zhang & Nooteboom, 1998) providesthe most up-to-date account of the genus in Borneo, but the specimens were difficult tokey to species because although they had the crenate sterile pinna margins of P.egenolfioides (Baker) Copel., the sterile pinnae were slightly narrowed at base, notauriculate which is a character of this species. The key to varieties of P. egenolfioidesindicates that the collections belong to var. egenolfioides, being dwarf and with obscureaerophores only at the stipe base, but the description of this variety also states that mostof the sterile pinnae are auriculate. Plagiogyria minuta Copel. is listed in the synonymyof P. egenolfioides var. egenolfioides and examination of the image of the type, MICH1190812, shows that not only is it the same taxon as the Gunung Gaharu plants, but alsothat it would not key to P. egenolfioides var egenolfioides either, because of the shape ofthe pinna bases. Copeland (1915) noted that P. minuta was “an evident and near relativeof P. egenolfioides, but much smaller throughout, and the sterile pinnae usually narrowedat the base instead of cordate or auricled”. The Flora Malesiana account needs to berevised to take into account the differences between the two taxa.
REFERENCESCOPELAND, E. B. 1915. Notes on Bornean Ferns. Philippines Journal of Science C
Botany 10(2): 145-149.ZHANG, X. C. & NOOTEBOOM, H. P. 1998. Plagiogyriaceae, in Flora Malesiana
Series II, Volume 3: 295-316, Flora Malesiana Foundation.
B. S. ParrisFern Research Foundation
KerikeriNew Zealand
Email: [email protected]
FERN GAZ. 20(1). 2015 46

SHORT NOTE
ACROSORUS NUDICARPUS TRANSFERRED TO XIPHOPTERELLA
Acrosorus nudicarpus P.M.Zamora & Co. is a grammitid fern (Polypodiaceae) knownonly from ultramafic substrates on Palawan Island in the Philippines (Zamora & Co,1981) and Mount Kinabalu in Sabah, Malaysia (Borneo) (Parris et. al., 1992 asXiphopteris nudicarpa (P.M.Zamora & Co) Parris). Although it was described inAcrosorus, it differs from all other member of the genus in having hydathodes at the veinendings on the adaxial surface of the lamina, a character shared with several species ofXiphopterella. The following new combination is proposed:
Xiphopterella nudicarpa (P.M.Zamora & Co) Parris, comb. nov., based on Acrosorusnudicarpus P.M.Zamora & Co, Nat. Appl. Sci. Bull. Univ. Philipp. 32: 47 (1981). Type:Philippines, Palawan, Mt Mantalingahan, Iloilo Ridge, c. 1700 m, March-April 1977,Co 1776 (holotype PNH 199345!; isotypes A, K, L, LWG, MICH barcode 1115915!,PUH).
REFERENCESPARRIS, B. S., BEAMAN, R. S. & BEAMAN, J. H. 1992. The Plants of Mount
Kinabalu. 1. Ferns and Fern Allies. Royal Botanic Gardens, Kew. 165 pp.ZAMORA, P. M. & CO, L. L. 1981. New species of ferns (Filicopsida) from Palawan,
Philippines. Nat. Appl. Sci. Bull. Univ. Philipp. 32: 43-51.
B. S. ParrisFern Research Foundation
KerikeriNew Zealand
Email: [email protected]
47 FERN GAZ. 20(1). 2015

INSTRUCTIONS FOR AUTHORS
PAPERS should not usually exceed 20 printed pages and are generally expected to beconsiderably shorter. As well as reports of original research, review articles and shortnotes, e.g. new records, are encouraged. The senior author should supply an email addressto facilitate correspondence. REVIEWS should cover recent developments in any aspect of research on ferns andlycopods. This can include unpublished research by the author, and/or the possibledirection of future research. They should be aimed at the general reader who may not bean expert in the chosen topic. Proposals for a review should be discussed first with thereview editors.MANUSCRIPTS should be submitted in English (British) in electronic format in10-point Times New Roman font and double spaced. Electronic versions of text andtables should be compatible with WORD, with figures as pdf or jpg files, and sent asemail attachments or CDroms. All manuscripts will be refereed.THE TITLE should reflect the content of the paper and be in BOLD CAPITALS(11-point) and centrally aligned. Generic and specific names should be in italics and anytitle containing a generic or specific name should be followed by the family name inbrackets e.g.
TRICHOMANES SPECIOSUM (HYMENOPHYLLACEAE)IN SOUTHERN SPAIN
AUTHOR ABBREVIATIONS should follow Pichi Sermolli’s (1996) Authors ofscientific names in Pteridophyta, Royal Botanic Gardens, Kew.MAIN HEADINGS: should be in BOLD CAPITALS (10-point) and centrally aligned.SUBSIDIARY HEADINGS: should be in bold, the first letter of each word in capitals,the rest in lower case and left-aligned.AUTHORS’ NAMES AND FULL ADDRESSES (including email address): followthe title and are centrally aligned.KEY WORDS: up to ten.ABSTRACT: should reflect the content of the paper.FIGURES: there is no distinction between photographs and line drawings in numbering.All should be presented in a form ready for reproduction, colour, TIF format and 300dpi (please contact editor with queries), with a scale bar where appropriate. Lettering ornumbers (Arabic) should be in the bottom left using uppercase Times Roman. Figurecaptions should be on a separate sheet.TABLES: can be printed in either portrait or landscape format. Authors should considerthis when preparing tables. Authors should ensure that tables fit the printed page size ina legible form.PAGESIZE: The Gazette has a useable page size of 123 × 204 mm. This can be used inportrait or landscape mode but any supplied artwork should not be larger than this.MEASUREMENTS: follow the metric system.CHECKLISTS: follow the format of Baksh-Comeau, Fern Gaz. 16(1, 2): 11- 122.REFERENCES: follow the style of a recent issue of The Fern Gazette, e.g.:-HOOKER, W.J. 1864. Species Filicum, 5. Dulau & Co., London.MORTON, C.V. 1947. The American species of Hymenophyllum, section
Sphaeroconium. Contr. U.S. Natl. Herb. 29(3): 139-201.STEVENSON, D.W. & LOCONTE, H. 1996. Ordinal and familial relationships of
pteridophyte genera. In: CAMUS, J.M., GIBBY, M. & JOHNS, R.J. (Eds)Pteridology in perspective, pp. 435-467. Royal Botanic Gardens, Kew.
JOURNAL ABBREVIATIONS: should follow Botanico Periodicum Huntianum &Supplements.Alterations from the original text at proof stage will be charged for unless they are minorpoints of detail. A pdf will be provided free to the senior author.
48 FERN GAZ. 20(1). 2015

TWO NEW DIPLAZIUM (WOODSIACEAE) SPECIES FROM EASTMALESIA
P. HOVENKAMP
Naturalis Biodiversity Center, PO box 9517, NL-2300 RA Leiden, The NetherlandsEmail: [email protected]
Keywords: Diplazium, taxonomy, rhizome scales, Sulawesi, Seram,
ABSTRACTTwo new species of Diplazium are described from Indonesia: D. walkeriHovenk. from Sulawesi, and D. murkele Hovenk. from Seram.
INTRODUCTIONDuring identification of ferns collected by A.C. Jermy and T. G. Walker in 1969 in theLatimojong Mountains, Sulawesi Selatan (Indonesia), a number of specimens ofDiplazium (Woodsiaceae) could not be identified with any known species. Comparisonwith the collections in BM, K and L revealed similarities with Diplazium mutabileHovenk. (Hovenkamp & De Joncheere, 1988), described from a different mountain areaon Sulawesi, with D. furculicolum Alderw. (Van Alderwerelt van Rosenburgh, 1918),described from Seram, and with a number of collections from Seram identified by Katoet al. (Kato, 1994) as D. vestitum C.Presl. With D. mutabile the unidentified specimensshare a characteristic combination of evanescent basal scales and pinnule rachisornamentation. The basal scales characteristically disintegrate starting with the outercell walls (Figure 1A), and the abaxial surfaces of the pinna and pinnule rachis areornamented with a combination of somewhat fleshy hair-like protrusions and appressedflat scales (Figure 1B). The same rachis indument is found in D. furculicolum. and inthe Seram collections identified as D. vestitum, but accompanied by different types ofscales. The scales of D. furculicolum have the same thin texture as those of D. mutabile,but retain their general outline more strongly; the scales of the Seram “D. vestitum”specimens are thick, brown and fleshy, but also appear disintegrated somewhatespecially at the margin and the surface of the scale, leaving the exact shape and outlineof the scales uncertain. Diplazium vestitum, as represented in L by collections from thePhilippines and Borneo, differs significantly in having basal scales that are translucentwith a dark dentate margin and smooth abaxial rachis surfaces and thereby is clearlydifferent from the Seram specimens.
Notwithstanding the general state of ignorance we are in about the exact speciesdelimitation in Malesian Diplazium, the four groups of specimens appear to besufficiently distinct to warrant the description of two new species. Table 1 provides asynopsis of the major morphological similarities and differences between the fourspecies discussed here.
METHODSPhotographs were taken with a Zeiss Discovery V20 microscope equipped with a ZeissAxiocam MRC5 camera, and processed with Axiovision software (Axiovision Vs40x64V. 4.9.10). For scanning electron microscopy (SEM) spores were sputter-coated withgold and examined in a JEOL JSM 5300 microscope (Figure 2A) or coated with c. 5 nmPt/Pd and observed with a Jeol JSM 7600F FEG-SEM (Figure. 2B, C, D).
FERN GAZ. 20(2):49-54. 2015 49

50 FERN GAZ. 20(2):49-54. 2015
Figure 1. Diplazium walkeri.A: Rhizome scale, showing the fibrous transverse cell walls remaining after thedisappearance of the superficial walls (rehydrated). Scale bar 2mm. B: Abaxial surface of penultimate midrib, showing combination of fleshy hairs andappressed flat scales (in sicco). Scale bar 1mm (Walker 12367, BM).
Figure 2. Spores:A: Diplazium mutabileB: Diplazium walkeriC: Diplazium murkele, inset: detail of scaly outer layerD: Diplazium furculicolumScale bars 10µm except inset in C 1µm. (Hennipman 5571 (L): A; Kato et al. C 5043(L): C; C 1124 (L): D, Walker T12373 (BM): B).
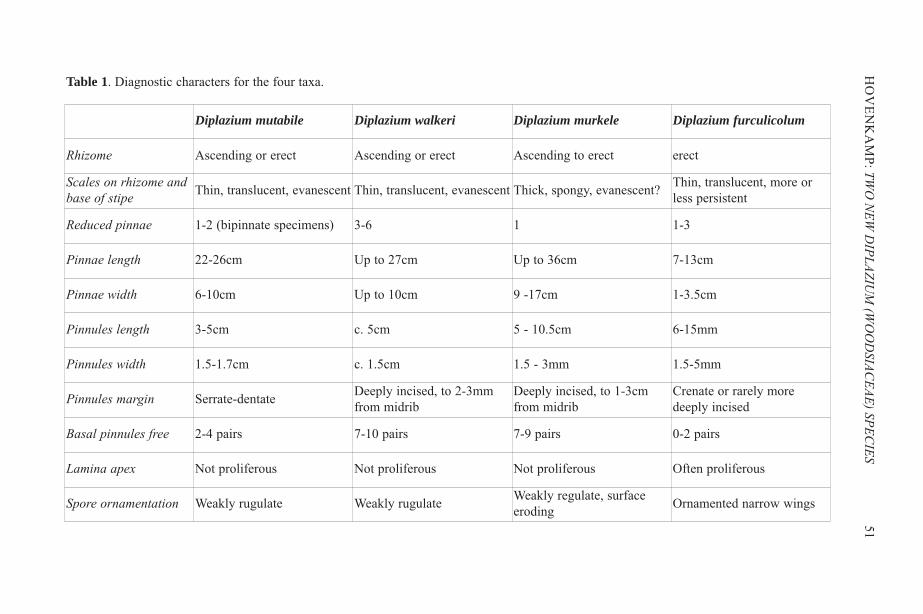
HO
VEN
KA
MP: TW
O N
EW D
IPLAZIUM
(WO
OD
SIACEAE) SPEC
IES 51
Table 1. Diagnostic characters for the four taxa.
Diplazium mutabile Diplazium walkeri Diplazium murkele Diplazium furculicolum
Rhizome Ascending or erect Ascending or erect Ascending to erect erect
Scales on rhizome andbase of stipe Thin, translucent, evanescent Thin, translucent, evanescent Thick, spongy, evanescent? Thin, translucent, more or
less persistent
Reduced pinnae 1-2 (bipinnate specimens) 3-6 1 1-3
Pinnae length 22-26cm Up to 27cm Up to 36cm 7-13cm
Pinnae width 6-10cm Up to 10cm 9 -17cm 1-3.5cm
Pinnules length 3-5cm c. 5cm 5 - 10.5cm 6-15mm
Pinnules width 1.5-1.7cm c. 1.5cm 1.5 - 3mm 1.5-5mm
Pinnules margin Serrate-dentate Deeply incised, to 2-3mmfrom midrib
Deeply incised, to 1-3cmfrom midrib
Crenate or rarely moredeeply incised
Basal pinnules free 2-4 pairs 7-10 pairs 7-9 pairs 0-2 pairs
Lamina apex Not proliferous Not proliferous Not proliferous Often proliferous
Spore ornamentation Weakly rugulate Weakly rugulate Weakly regulate, surfaceeroding Ornamented narrow wings

DESCRIPTION OF NEW TAXADiplazium walkeri Hovenk., spec. nov.Type: Trevor G. Walker 12373 (holo BM; iso L), Indonesia, Sulawesi Selatan,Enrekang, above the village of Rantelimo, 25km SE of Kalosi, on the south-westernslopes of Mt. Rantemario, 1950-2100m, on ridge above northern branch of R. Pasui, 24November 1969.Terrestrial fern, rhizome ascending or erect, incl. stipe-base remnants 2-3cm thick;scaly but glabrous in older parts; rhizome scales basifix, periclinal cell walls very thinand quickly disappearing, often leaving the scale disintegrating into a fibrous network,shape, margin, apex of the scales accordingly not observed. Fronds to c. 130cm long;stipes to c. 46cm, sparsely scaly at base with scales similar to those on the rhizome,upwards with papillose multicellular protrusions; lamina bipinnate, to c. 90 by 50cm,widest at c. 1/3-1/5, gradually narrowing from c. halfway up; free pinnae 12-15 pairs,lowest 1-6 somewhat reduced, alternate to subopposite, largest pinnae to 2 cm stalked,to 37 by 18cm; pinnules alternate to opposite, basal 7-10 pairs free, to 5 by 1.5cm,symmetric at base, incised to 1.5-3mm from midrib, apex obtuse to acute, lobestruncate; pinnae and pinnule rachis on the adaxial surface with two flanges interruptedat the vein junctions, on the abaxial surface with flat translucent scales and papillosemulticellular protrusions, veins free. Sori diplazioid, on all veins of a vein-group,running from costule to c. 1mm from the margin, spores c. 50 µm long, very lightlyrugulate. Distribution – Indonesia: Sulawesi Selatan (Enrekang District, Latimojong Mountains). Ecology – From lowland forest to upper montane forest, in deep shade, in ravines or bystreams, 1700-2150m.Etymology - The species is named after Trevor G. Walker (1927-2006), Britishpteridologist. Note - Diplazium walkeri differs from D. mutabile mainly in the more highly dissectedlamina. Diplazium furculicolum differs from both in a much smaller size, basal scalesthat retain more of their shape and in much more strongly ornamented spores (Figure2).
Diplazium murkele Hovenk., spec. nov.Type: M. Kato, K. Ueda & U.W. Mahjar C 1292 (holo L; iso TI, not seen, BO, not seen),Indonesia, Seram, Tehoru & Seram Utara: Manusela National Park: along a trailbetween Hatumete (seaside) and Maraina (810 m) in Manusela Valley via Hoale Pass(1770m) in Murkele Ridge, 03° 10-14' S, 129° 35-37' E, 18 Nov. 1983.Terrestrial fern, rhizome ascending or erect, incl. stipe-base remnants 2-3cm thick;scaly but glabrous in older parts; rhizome scales basifix, to c. 10 by 2mm wide, thick,fleshy, margin entire, in the basal part thin, flaccid; apex obtuse. Fronds to c. 120 orlonger; stipes to 75cm, naked or sparsely scaly at base with thick brown scales to 10 by6 mm, lamina bipinnate, to c. 70 by 50cm, widest at c. 1/4, free pinnae 8-12 pairs,lowest pair slightly reduced, alternate to opposite, largest pinnae to 1.5cm stalked, to 37by 17cm; pinnules alternate to opposite, basal 7-9 pairs free, to 8.5 (10.5) by 1.5-2(3)cm, symmetric at base, incised to 1—3 mm from midrib, apex obtuse to acute, lobestruncate; pinnae and pinnule rachis on the adaxial surface with two flanges interruptedat the vein junctions, on the abaxial surface with flat translucent scales and papillosemulticellular protrusions, veins free. Sori diplazioid, on all veins of a vein-group,running from costule to c. 1mm from the margin, spores c. 40µm long when well-
52 FERN GAZ. 20(2):49-54. 2015

formed, many collapsed or otherwise malformed, perispore with a thin almost smooth,eroded, scaly outer layer uncovering a finely irregularly porous inner layer.Distribution – Indonesia, Moluccas: Seram.Ecology – Terrestrial, in deep shade, near brooks, in ravines etc., in lowland to montaneforest, disturbed forest, often noted for calcareous areas, lowland to 1350m, possiblyhigher. Etymology - The epithet is to be treated as a word in apposition and derives fromMurkele Ridge, the location of the type specimen and a number of other specimens.Note – Diplazium murkele differs from D. walkeri mainly in the basal scales that arenot thin and translucent but thick and fleshy. However, they often also appear to showsigns of disintegration, which may explain the scarcity of intact scales in the collectedspecimens. The lamina is somewhat smaller, and has distinctly fewer reduced pinnaenear the basis, the pinnules on the other hand are a bit larger. The outer layer of thespores appears to erode away, leaving only a partial, scaly cover (Figure 2 C).
Additional Specimens Seen:
Diplazium mutabile Sulawesi. Palolo: Hennipman, E. 5003 (L) 21 April 1979; Track to Roroka Timbu,Sopu-valley: Hennipman, E. 5151 (L) 30 April 1979; Sopu-valley: Hennipman, E. 5571(L) 23 May 1979; 5587 (L) 1979, 5625 (L) 26 May 1979; 5640 (L); Gumbassa:Hennipman, E. 5711 (L) 30 May 1979; Rantemario: Walker, T.G. 11985 (BM) 1November 1969; 12369 (BM) 24 November 1969.
Diplazium murkele.Moluccas. Manusela National Park: Iwatsuki et al. C 1027 (L) 12 November 1983;Kato et al. C 801 (L) 11 November 1983, C 1292 (L) 18 November 1983; C 1475 (L)22 November 1983; C 1773 (L) 29 November 1983, C 2924 (L) 30 December 1984; C3223 (L) 2 January 1985; C 3553 (L) 3 January 1985, C 5043 (L) 22 January 1985, C12022 (L) 24 July 1986; C 13799 (L) 26 August 1986; C 14089 (L) 27 August 1986.
Diplazium walkeri.Sulawesi. Rantemario: Walker, T.G. 11958 (BM, L) 1 November 1969; 11959 (BM,L)11961 (BM,L) 12365 (BM,L) 24 November 1969; 12368 (BM,L) 12373 (BM,L)
Diplazium furculicolum.New Guinea. Sattelberg: Clemens 7059 (BM) 10 September 1937; Moluccas.Manusela National Park: Kato, M et al. C 949 (L) 16 November 1983, C 1602 (L) 25November 1983; C 3216 (L) 2 January 1985, C 6887 (L) 17 February 1985; C 7437 (L)20 February 1985, C 12191 (L) 27 July 1986; C 12431 (L) 28 July 1986; C 12582 (L)31 July 1986; C 14325 (L) 7 September 1986; C 14457 (L) 1 September 1986; C 1124(L) 17 November 1983; C 1207 (L) 16 November 1983; Parris, B.S. 11089 (L) 27August 1987.
ACKNOWLEDGMENTSA visit to the Natural History Museum, London to identify collections from Sulawesiwas supported by Synthesys grant GB-TAF-4095. I gratefully acknowledge thepatience of the Curator of Pteridophytes at BM for allowing me the loan of a numberof specimens for an extensive period.
HOVENKAMP: TWO NEW DIPLAZIUM (WOODSIACEAE) SPECIES 53

REFERENCESHOVENKAMP, P. & DE JONCHEERE, G.J. 1988. Additions to the fern flora of
Sulawesi. Blumea; 33(2): 395-409.KATO, M. 1994. Taxonomic studies of Pteridophytes of Ambon and Seram. J. Fac. Sc.
Tokyo, III. Botany 15: 135-152.VAN ALDERWERELT VAN ROSENBURGH, C. R. W. K. 1918. New or interesting
Malayan Ferns 10. Bull. Jard. Bot. Buitenzorg sér. 2 28: 1-56.
54 FERN GAZ. 20(2):49-54. 2015

EFFECT OF TEMPERATURE ON THE VIABILITY, LIFESPAN ANDVIGOUR, OF CHLOROPHYLLOUS SPORES OF OSMUNDASTRUM
CINNAMOMEUM (OSMUNDACEAE)
Y.L. CHANG1,2, M.H. TSENG2,3, W.L. CHIOU4 & Y.M. HUANG4,5
1Dr. Cecilia Koo Botanic Conservation Center, 31 Tongsing Rd., Gaoshu Township,Pingtung 90646, Taiwan.
2These authors contributed equally to this work.3Department of Applied Physics and Chemistry, University of Taipei, 1 Ai-Guo West
Rd., Taipei 10048, Taiwan.4Taiwan Forestry Research Institute, 53 Nanhai Rd., Taipei 10066, Taiwan.
5Corresponding author, e-mail: [email protected]
ABSTRACTShort-lived chlorophyllous spores need special treatment to extend theirlongevity and vigour for ex situ conservation. Germination rates, mean longevityand germination times of chlorophyllous spores of the rare fern Osmundastrumcinnamomeum were determined to assess their viability, lifespan and vigour at18–25°C (room temperature, RT), 4°C and -80°C during 96 weeks of storage.Results showed that at the low temperatures, the viability and vigour weresignificantly maintained, and the lifespan of O. cinnamomeum spores wasextended. Spores stored at RT had completely lost all ability to germinate within12 weeks. Some spores retained their viability for more than 96 weeks at 4°Cand -80°C. Mean longevities were 6.1, 41.8, and >96 weeks with storage at RT,4°C and -80°C respectively. The mean spore germination time (MGT), hereinregarded as an indicator of vigour, was affected by both storage temperature andtime. Spores stored at RT and 4°C began to lose their vigour after three weeks.However, it did not decline at -80°C until 24 weeks of storage. This studydemonstrates that low temperature (4°C) and cryopreservation (-80°C)significantly extended the longevity and vigour of chlorophyllous spores of O.cinnamomeum.
Keywords: cryopreservation, mean germination time.
INTRODUCTIONAs plants are threatened by habitat loss, pollution, invasive species, and other factors,in situ conservation is considered as the primary focus for conserving threatenedspecies. Ex situ germplasm storage, on the other hand, is an important complementaryaspect of plant conservation (Pence, 2004). A germplasm bank, storing livingpropagules in a small space, can efficiently maintain genetic variability (Dickie et al.,1990; Smith et al., 2003; Pence, 2008). Conservation plans for a number of threatenedfern species include propagation from spores (e.g., Estrelles et al., 2001; Lusby et al.,2002; Simabukuro et al., 1998). Spore banks have low financial costs and have beendeveloped for more than 20 years to conserve ferns ex situ (Page et al., 1992; Ibars &Estrelles, 2012).
Most fern spores can typically retain their viability for a long period but there isgreat disparity among species (Okada, 1929). One of the principal features causing such
FERN GAZ. 20(2):55-64. 2015 55

variation is the presence of chlorophyll pigments. Fern spores are primarily divided intochlorophyllous spores (also called green spores) and non-chlorophyllous spores (non-green spores), depending on whether or not chlorophyll pigments exist in them (Gabriely Galán & Prada, 2011). Compared with non-chlorophyllous spores, which retain theirviability for several months to years, chlorophyllous spores are relatively short-lived,surviving for only a few weeks or months at room temperature (Lloyd & Klekowski,1970).
Chlorophyllous spores quickly lose their viability because of a lack of desiccationtolerance, a high metabolic rate, and the absence of dormancy (Kato, 1976; Li & Shi,2014). Therefore they require specialised treatment for long-term conservation ingermplasm banks (Ballesteros et al., 2011). It is well documented that the viability ofchlorophyllous spores can usually be extended under low temperatures (Ballesteros,2011; Gabriel y Galán & Prada, 2011). The longevity of chlorophyllous and non-chlorophyllous spores has been studied for over 50 years, and some protocols have beenestablished to optimise spore preservation in fern spore banks (Ibars & Estrelles, 2012).For example, low temperature (-70°C) storage substantially prolonged chlorophyllousspore viability of Equisetum hyemale L. for over 16 months (Whittier, 1996), andat -20°C for more than 28 months for Osmunda regalis L. (Magrini & Scoppola, 2012).The “viability” of spores has been variously defined, such as the length of time thatspores retain their ability to germinate (Raghavan, 1989; Gabriel y Galán & Prada,2011), or the capacity of a spore to germinate (Qunitanilla et al., 2002). The former oneis mutually used with the term “longevity” (e.g., Aragón & Pangua, 2004, Pence, 2004).We herein define “spore longevity” (the lifespan of a spore) to be the period from sporeharvest to loss of germination ability as per Windham et al. (1986) and Ballesteros et al.(2011). “Spore viability” is defined as the spore germination ability as indicated by thegermination rate (%) (Gabriel y Galán & Prada, 2011).
The quality of spores (spore vigour) is related to the germination rate and ageingduring storage. Because spore germination time is prolonged with storage time (Smith& Robinson, 1975; Camloh, 1999), germination time was used to indicate spore vigourin this study.
Osmundastrum cinnamomeum (L.) C.Presl is a temperate fern, usually growing inswamps and widely distributed from Asia to the Americas (Metzgar et al., 2008).However, the population number is low (≤4) and relatively small (<200 matureindividuals in the largest population) in Taiwan (Chiu et al., 2013). It was evaluated tobe a vulnerable species based on IUCN red list categories and criteria (Wang et al.,2012). The goal of this study was to determine the effect of various storagetemperatures on the longevity and vigour of chlorophyllous spores of O. cinnamomeum.The results provide useful information for long-term ex situ conservation of this rarefern in spore banks.
MATERIALS AND METHODSFresh spores of O. cinnamomeum were obtained from 12 fertile fronds of six plantsgrowing in a marsh at Tsaopi (24°46'03"N, 121°36'26"E, elev. 809m), Yilan County,northeastern Taiwan, in March 2012. The fertile fronds released spores after beingair-dried for three days. Spores were isolated from sporangia and transferred tomicrotubes (Labcon Screw-Cap Microcentrifuge Tubes, 2.0ml) for dry storage.Microtubes of spores were stored at 18−25°C (room temperature, RT), 4°C or -80°C forup to 96 weeks.
56 FERN GAZ. 20(2):55-64. 2015

Spores stored at RT, 4°C and -80°C were sown separately onto four sheets ofmembrane filters (with a pore size of 0.45µm and diameter of 47mm; GelmanLaboratory) after 0 (fresh spores), 1, 2, 3, 4, 8, 12, 24, 48 and 96 weeks of storage. Eachsheet was laid on a 4cm thick layer of moist sphagnum moss in a plastic box(114×86×102mm, PHYTATRAY IITM, Sigma). All cultures were incubated underwhite fluorescent illumination at approximately 25−35µmol m-2 s-1 for 12h d-1 at 20± 1°C.
Germination refers to the breakage of the spore wall and the appearance of the firstrhizoid or gametophyte cell (Lloyd & Klekowski, 1970). To calculate the germinationrate (%), 200 spores (50 spores/filter × 4 filters) were randomly observed under amicroscope (Leica, Wild M8) every week after sowing until no more germination wasdetected.
The mean germination time (MGT) was calculated for each replication pertreatment according to the equation: MGT = Σ(tn)/(Σn), where t is the time in weeksstarting from spore sowing and n is the number of spores that had germinated in weekt (modified from De Brum & Randi, 2006). The spore lifespan was indicated by thespores’ mean longevity calculated by the equation: ML = Σti (pi − pi+1), where ti is thespore storage time (ith week), pi is the germination rate in the ith week, and pi+1 is thegermination rate for the next storage time (i+1th week).
Germination rates and mean germination times for each temperature were analysedby a one-way analysis of variance (ANOVA) and Fisher's least significant difference(LSD) test (p<0.05) to estimate significant differences of means among differentstorage times.
RESULTSAll (100%) fresh spores of O. cinnamomeum germinated within one week. With RTstorage, the spore germination rate remained at almost 100% for four weeks in storage.Then it decreased significantly to nearly 50% after eight weeks in storage, and sporeshad completely lost viability after 12 weeks in storage. When the storage temperaturewas lowered to 4°C, spores retained almost all of their viability for 24 weeks in storage.Then it decreased significantly to 69.5% and 3.0% after 48 and 96 weeks of storage,respectively. For spores stored at -80°C, the germination percentage was almost 100%during 48 weeks of storage and still remained at 91.5% even after 96 weeks of storage(Table 1).
The MGT of fresh spores was one week. Under RT storage, the MGT increasedsignificantly to 1.4 weeks after three weeks in storage, and up to 2.0 and 2.1 weeks afterfour and eight weeks in storage, respectively. The MGT of spores stored at 4°C did notincrease within two weeks of storage, then slightly (but significantly) increased to1.3−1.5 weeks during 3−12 weeks of storage, and again significantly increased to2.4−2.5 weeks after 24−48 weeks of storage. However MGT was found to havedecreased to 2.0 weeks after 96 weeks of storage. The MGT was ≤1.2 weeks whenspores were stored at -80°C after 12 weeks in storage, but significantly increased to 2.2weeks after 24 weeks of storage, then decreased to 1.9 and 1.7 weeks respectively whenspores were stored for 48 and 96 weeks (Table 2).
Spores had completely lost their viability after 12 weeks of storage at RT, whereassome viability was retained at 4°C and -80°C storage at the end of the observationperiod (96 weeks) (Table 2). Thus the lifespan of at least some proportion of spores at4°C and -80°C was more than 96 weeks. The mean longevity of spores significantly
CHANG et al.: EFFECT OF TEMPERATURE ON CHLOROPHYLLOUS SPORES 57
58 FERN GAZ. 20(2):55-64. 2015
Table 1. Germination rates of spores of Osmundastrum cinnamomeum at differentstorage times and temperatures
1) Data are the mean±standard deviation (n=4).2) Means with the same superscript letter do not significantly differ within atemperature (p<0.05; LSD test).3) Room temperature of 18−25°C.
Germination rate (%)1,2)
Storage time
(weeks)
Storage temperature
RT3) 4°C - 80°C
0 100a -- --
1 100a 100a 100a
2 100a 100a 100a
3 100a 100a 100a
4 99.5±1.0a 100a 100a
8 51.5±4.4b 100a 100a
12 0c 100a 100a
24 0c 97.0±3.5a 96.5±4.7a
48 0c 69.5±10b 98.5±1.9a
96 0 c 3.0±1.2c 91.5±5.3b
CHANG et al.: EFFECT OF TEMPERATURE ON CHLOROPHYLLOUS SPORES 59
Table 2. Mean germination times of spores of Osmundastrum cinnamomeum atdifferent storage times and temperatures
1) Data are the mean±standard deviation (n=4).2) Means with the same superscript letter do not significantly differ within atemperature (p<0.05; LSD test).3) Room temperature of 18−25°C.4) -, no data.5) --, spores did not germinate.
Mean germination time (weeks)1,2)
Storage time
(weeks)
Storage temperature
RT3) 4°C - 80°C
0 1.0a -4) -
1 1.1±0.0a 1.1±0.0a 1.1±0.0a
2 1.1±0.1a 1.1±0.0a 1.1±0.1a
3 1.4±0.2b 1.3±0.1b 1.1±0.0a
4 2.0±0.2c 1.3±0.0b 1.2±0.1a
8 2.1±0.2c 1.5±0.0c 1.1±0.0a
12 --5) 1.4±0.1bc 1.1±0.0a
24 -- 2.5±0.2e 2.2±0.3d
48 -- 2.4±0.2e 1.9±0.1c
96 -- 2.0± 0.0d 1.7±0.2b

differed among the three temperature treatments, with 6.1, 41.8, and >96 weeks at RT,4°C and -80°C storage, respectively.
DISCUSSIONThe viability of chlorophyllous spores under RT conditions is known to be lost rapidlybut there are great disparities among species. For example, longevity of chlorophyllousspores of Equisetum hyemale, Osmunda regalis, and Matteuccia sp. are two weeks, ca.one month, and more than one year, respectively (Stokey, 1951; Lloyd & Klekowski,1970; Lebkuecher, 1997). This study demonstrated that the lifespan of a proportion ofspores of O. cinnamomeum at RT reached eight weeks, slightly longer than previousrecords from Japan (1.4–1.8 months) (Okada, 1929). This difference may have resultedfrom plant genetic variations and/or spore maturity. Fully mature spores usually havegreater longevity than do immature ones (Huang et al., 2014). Some physiologicalmechanisms and compounds, including the respiration rate, water content, catalasecontent and granules of lipids and lipoproteins, were documented to affectchlorophyllous spore viability (Okada, 1929; Gullvåg, 1969; Olsen & Gullvåg, 1973).The decay of fern spore viability might also result from the loss of some housekeepingsubstances, such as sugars, amino acids and proteins (Beri & Bir, 1993; Ballesteros etal., 2004). Spore viability is also related to various kinds of sulphur metabolites, suchas glutathione (GSH), glutathione disulphide (GSSG), S-adenosylmethionine (SAM),and S-adenosylhomocysteine (SAH), which are involved in modulating proteinactivities or other metabolites under stress (Giovanelli et al., 1985; Dixon et al., 2005;Michelet et al., 2005; Loenen, 2006; Roje, 2006). Chang et al. (2013) furtherdocumented that GSH/GSSG and SAM/SAH ratios of spores of O. cinnamomeumsignificantly declined as they lost viability. Apparently those physiochemical changesaffect the germination rates of O. cinnamomea spores and probably are correlated withthe loss of viability in other chlorophyllous fern spores.
Chlorophyllous spores tend to germinate quickly because their photosyntheticallyactive chloroplasts can rapidly transform light energy to metabolic activity (Raghavan,1989). The MGT of chlorophyllous spores at RT is ca. 1.5 days for the Equisetaceae,Osmundaceae, Grammitidaceae, Hymenophyllaceae, Blechnum nudum (Labill.) Mett.ex Luerss., Lomariopsis sorbifolia (L.) Fée, and some species of Christiopteris,Marginariopsis, Matteuccia, Onoclea and Onocleopsis (Lloyd & Klekowski, 1970). Itis obvious that the germination rate of chlorophyllous spores should be measured atshort intervals. Daily observations would provide more accurate data than the weeklyobservations that were used in this study. Nevertheless, correlations between sporevigour and different storage temperatures were distinguishable.
The results of this study show that the MGT increased before the germination ratedeclined. However, the MGT of O. cinnamomeum spores increased significantly afterthree weeks of storage at 4°C, then decreased after 96 weeks of storage. A similarsituation also occurred with storage at -80°C. The increase and decrease in the MGTmay have resulted from the self-regulation of spores in response to stress during storage(Chang et al., 2013). More studies are needed to determine whether the MGT is a goodindicator for evaluating the germination vigour of fern spores.
The successful establishment of individuals and populations of a species in a newhabitat after spore dispersal depends on how long the spores retain their viability andvigour (Sheffield, 1996; Shorina, 2001; Gabriel y Galán & Prada, 2011). When thespore vigour decreases, subsequent gametophytes may grow abnormally and even
60 FERN GAZ. 20(2):55-64. 2015

affect subsequent sporophyte production (Smith & Robinson, 1975; Beri & Bir, 1993)and population establishment (Gabriel y Galán & Prada, 2011).
An increase in the storage time results in decline in spore viability and vigour aspreviously reported for O. cinnamomeum and a few other ferns (Smith & Robinson,1975; Ballesteros et al., 2011, 2012). Evidence was presented that suggests spore agemay be partly attributable to the declining ability of spores to synthesise metabolitesessential for germination (Gabriel y Galán & Prada, 2011). It is well known thatchlorophyllous spores have shorter lifespans compared with non-chlorophyllous spores(Lloyd & Klekowski, 1970). Using low temperature to prolong their lifespan and vigourwas documented (Pence, 2000, 2008; Ballesteros et al., 2006, 2011, 2012; Ballesteros,2011; Gabriel y Galán & Prada, 2011). It was reported that chlorophyllous spores ofOsmunda regalis and O. japonica Thunb. stored at low temperatures (-25 and 4°C) alsoretained viability for up to 24 and 12 months respectively (Ballesteros et al., 2011; Li& Shi, 2014). This study also showed that spores of O. cinnamomeum rapidly lostviability at RT storage, and the lowest temperature of -80°C provided the bestconditions to retain their spore viability and vigour. Cryopreservation is, therefore,recommended for the long-term storage of spores of this species, and this proceduremay be applicable to other fern spores in terms of ex situ conservation.
CONCLUSIONSChlorophyllous spores of ferns have shorter lifespans at room temperature storagecompared with non-chlorophyllous spores. However, cryogenic temperatures mightsuppress metabolic activities and effectively extend their longevity and retain the vigourof chlorophyllous spores as shown in this study. It is suggested that cryopreservation isa feasible method for establishing long-term germplasm of the chlorophyllous spores ofOsmundastrum cinnamomeum.
REFERENCESARAGÓN, C.F. & PANGUA, E. 2004. Spore viability under different storage
conditions in four rupicolous Asplenium L. taxa. Amer. Fern J. 94: 28-38.BALLESTEROS, D. 2011. Conservation of fern spores. In: FERNÁNDEZ, H.,
KUMAR, A. & REVILLA, M.A. (Eds) Working with Ferns: Issues andApplications, pp. 165-172. Springer press, New York.
BALLESTEROS, D., ESTRELLES, E. & IBARS A.M. 2006. Responses ofpteridophyte spores to ultrafreezing temperatures for long-term conservation ingermplasm banks. Fern Gaz. 17: 293-302.
BALLESTEROS, D., ESTRELLES, E., WALTERS, C. & IBARS, A.M. 2011. Effect ofstorage temperature on green spore longevity for the ferns Equisetum ramosissimumand Osmunda regalis. Cryoletters 32: 89-98.
BALLESTEROS, D., ESTRELLES, E., WALTERS, C, & IBARS, A.M. 2012. Effectof temperature and desiccation on ex situ conservation of nongreen fern spores.Amer. J. Bot. 99: 721-729.
BALLESTEROS, D., IBARS, A.M. & ESTRELLES, E. 2004. New data aboutpteridophytic spore conservation in germplasm banks. In: PLANTA EUROPACOMMITTEE (Eds) 4th European Conference on the Conservation of Wild Plants,Valencia.
BERI, A. & BIR, S.S. 1993. Germination of stored spores of Pteris vittata L. Amer.Fern J. 83:73-78.
CHANG et al.: EFFECT OF TEMPERATURE ON CHLOROPHYLLOUS SPORES 61

CAMLOH, M. 1999. Spore age and sterilization affects germination and earlygametophyte development of Platycerium bifurcatum. Amer. Fern J. 89: 124-132.
CHANG, Y.L., HSIEH, C.L., HUANG, Y.M., CHIOU, W.L., KUO, Y.H. & TSENG,M.H. 2013. Modified method for determination of sulfur metabolites in plant tissuesby stable isotope dilution-based liquid chromatography-electrospray ionization-tandem mass spectrometry. Anal. Biochem. 442: 24-33.
CHIU, T.Y., CHIOU, W.L. & HUANG, Y.M. 2013. Phenological differences betweensterile and fertile fronds in a dimorphic fern, Osmundastrum cinnamomeum (L.) C.Presl (Osmundaceae). Taiwan J. Biodivers. 15: 311-322.
DE BRUM, F.M.R. & RANDI, A.M. 2006. Germination of spores and growth ofgametophytes and sporophytes of Rumohra adiantiformis (Forst.) Ching(Dryopteridaceae) after spore cryogenic storage. Revista Brasil. Bot. 29: 489-495.
DICKIE, J.B., ELLIS, R.H., KRAAK, H.L., RYDER, K. & TOMPSETT, P.B. 1990.Temperature and seed storage longevity. Ann. Bot. 65: 197-204.
DIXON, D.P., SKIPSEY, M., GRUNDY, N.M. & EDWARDS, R. 2005. Stress-inducedprotein S-glutathionylation in Arabidopsis. Plant Physiol. 138: 2233-2244.
ESTRELLES, E, IBARS, A.M., IRANZO, J. & MORALES, F. 2001. Recuperacio yreintroduccio de Marsilea quadrifolia L. en los arrozales del delta del Ebro(Tarragona, Españ). Bot. Complut. 25: 251-259.
GABRIEL Y GALÁN, J.M. & PRADA, C. 2011. Pteridophyte spores viability. In:FERNÁNDEZ, H., KUMAR, A. & REVILLA, M.A. (Eds) Working with Ferns:Issues and Applications, pp. 193-205. Springer press, New York.
GIOVANELLI, J., MUDD, S.H. & DATKO, A.H. 1985. Quantitative analysis ofpathways of methionine metabolism and their regulation in Lemna. Plant Physiol.78: 555-560.
GULLVÅG, B.M. 1968. On the fine structure of the spores of Equisetum fluviatile var.verticillatum studied in the quiescent, germinated and non-viable state. GranaPalynol. 8: 23-69.
HUANG, Y.M., LEE, P.H., FU, C.H. & CHANG, Y.H. 2014. The relation betweenspore maturity and viability of tree ferns. J. Nat. Taiwan Museum 67(2): 1-14.
IBARS, A.M. & ESTRELLES, E. 2012. Recent developments in ex situ and in situconservation of ferns. Fern Gaz. 19(3): 67-86.
KATO, Y. 1976. The effect of freezing and organic solvents on viability ofchlorophyllous fern spores. Cytologia 41: 387-393.
LEBKUECHER, J.G. 1997. Desiccation-time limits of photosynthetic recovery inEquisetum hyemale (Equisetaceae) spores. Amer. J. Bot. 84: 792-797.
LI, Y. & SHI, L. 2014. Effect of desiccation level and storage temperature on greenspore viability of Osmunda japonica. Cryobiology 68: 446-450.
LLOYD, R.M. & KLEKOWSKI, E.J. 1970. Spore germination and viability inpteridophyta: evolutionary significance of chlorophyllous spores. Biotropica 2: 129-137.
LOENEN, W.A.M. 2006. S-adenosylmethionine: jack of all trades and master ofeverything? Biochem. Soc. Transact. 34: 330-333.
LUSBY, P., LINDSAY, S. & DYER, A.F. 2002. Principles, practice and problems ofconserving the rare British fern Woodsia ilvensis (L.) R. Br. Fern Gaz. 16: 350-355.
MAGRINI, S. & SCOPPOLA, A. 2012. First results from conservation studies ofchlorophyllous spores of the Royal fern (Osmunda regalis, Osmundaceae).Cryobiology 64: 65-69.
62 FERN GAZ. 20(2):55-64. 2015

METZGAR, J.S., SKOG, J.E., ZIMMER, E.A. & PRYER, K.M. 2008. The paraphylyof Osmunda is confirmed by phylogenetic analyses of seven plastid loci. System.Bot. 33: 31-36.
MICHELET, L, ZAFFAGNINI, M., MARCHAND, C., COLLIN, V.,DECOTTIGNIES, P., TSAN, P., LANCELIN, J.M., TROST, P., MIGINIAC-MASLOW, M., NOCTOR, G. & LEMAIRE, S.D. 2005. Glutathionylation ofchloroplast thioredoxin f is a redox signaling mechanism in plants. Proc. Natl. Acad.Sci. U.S.A 102: 16478-16483.
OKADA, Y. 1929. Notes on the germination of the spores of some Pteridophytes withspecial regard to their viability. Sci. Rep. Tdhoku Imp. Univ. Biol. 4: 127-182.
OLSEN, L.T. & GULLVÅG, B.M. 1973. A fine structural and cytochemical study ofmature and germinating spores of Equisetum arvense. Grana 13: 113-118.
PAGE, C.N., DYER, A.F., LINDSAY, S. & MANN, D.G. 1992. Conservation ofPteridophytes – the ex situ approach. In: IDE, J.M., JERMY, A.C. & PAUL, A.M.(Eds) Fern Horticulture; Past, Present and Future Perspectives, pp. 269-278.Intercept, Andover.
PENCE, V.C. 2000. Survival of chlorophyllous and nonchlorophyllous fern sporesthrough exposure to liquid nitrogen. Amer. Fern J. 90: 119-126.
PENCE, V.C. 2004. Ex situ conservation methods for bryophytes and pteridopytes. In:GUERRANT, E.O. Jr., HAVENS, K. & MAUNDER, M. (Eds) Ex Situ PlantConservation Supporting Species Survival in The Wild, pp. 206-227. Island Press,Covelo.
PENCE, V.C. 2008. Cryopreservation of bryophytes and ferns. In: REED, B.M. (Ed)Plant Cryopreservation: A Practical Guide, pp. 117-140. Springer, New York.
QUINTANILLA, L.G., AMIGO, J. & PAJARÓN, S. 2002. Effect of storage method onspore viability in five globally threatened fern species. Ann. Bot. 90: 461-467.
RAGHAVAN, V. 1989. Developmental Biology of Fern Gametophytes. CambridgeUniversity Press, New York.
ROJE, S. 2006. S-adenosyl-l-methionine: beyond the universal methyl group donor.Phytochemistry 67: 1686-1698.
SHEFFIELD, E. 1996. From pteridopyte spore to sporophyte in the naturalenvironment. In: CAMUS, M.G.J.M. & JOHNS, R. (Eds) Pteridology inPerspective, pp. 541-549. Royal Botanic Gardens, Kew.
SHORINA, N.I. 2001. Population biology of gametophytes in homosporouspolypodiophyta. Russ. J. Ecol. 32: 164-169.
SIMABUKURO, E.A., DYER, A.F. & FELIPPE, G.M. 1998. The effect of sterilizationand storage conditions on the viability of the spores of Cyathea delgadii. Amer. FernJ. 88: 72-80.
SMITH, D.L. & ROBINSON, P.M. 1975. The effects of spore age on germination andgametophyte development in Polypodium vulgare L. New Phytol. 74: 101-108.
SMITH, R.D., DICKIE, J.B., LININGTON, S.H., PRITCHARD, H.W. & PROBERT,R.J. 2003. Seed Conservation: Turning Science into Practice. Royal BotanicGardens, Kew.
STOKEY, A.G. 1951. Duration of viability of spores of the Osmundaceae. Amer. FernJ. 41: 111-115.
WANG, J.C., CHIOU, W.L. & CHANG, H.M. 2012. A Preliminary Red List ofTaiwanese Vascular Plants. Endemic Species Research Institute & Taiwan Societyof Plant Systematics, Nantou.
CHANG et al.: EFFECT OF TEMPERATURE ON CHLOROPHYLLOUS SPORES 63

WHITTIER, D.P. 1996. Extending the viability of Equisetum hyemale spores. Amer.Fern J. 86: 114-118.
WINDHAM, M.D., WOLF, P.G. & RANKER, T.A. 1986. Factors affecting prolongedspore viability in herbarium collections of three species of Pellaea. Amer. Fern J.76:141-148.
64 FERN GAZ. 20(2):55-64. 2015

PLEOPELTIS ×CERRO-ALTOENSIS (POLYPODIACEAE), A NEWFERN HYBRID FROM ROBINSON CRUSOE ISLAND (JUAN FERNANDEZ ARCHIPELAGO, CHILE)
P. DANTON1*, M. BOUDRIE2, A. BIZOT3 & R.L.L. VIANE4
15, rue Galilée, F-38000 Grenoble, France. E-Mail: [email protected]
216, rue des Arènes, F-87000 Limoges, France.E-mail: [email protected]
3 1, rue de la Faye, F-08160 Hannogne-Saint-Martin, France. E-mail: [email protected]
4 Universiteit Ghent, Vakgroep Biologie, Pteridologie, K.L. Ledeganckstraat35, B-9000 Ghent, Belgium.
E-mail: [email protected]* Author for correspondence
Keywords : Pleopeltis, hybrid, Polypodiaceae, Juan Fernández, Chile
ABSTRACTA fern hybrid of the genus Pleopeltis was discovered on Robinson Crusoe Islandin the Juan Fernández Archipelago, off the coast of Chile, and is described as P.×cerro-altoensis. Its putative parents are P. macrocarpa and P. masafuerae, twospecies present in the archipelago.
Mots-clés : Pleopeltis, hybride, Polypodiaceae, Juan Fernández, Chili
RÉSUMÉUn hybride de fougère appartenant au genre Pleopeltis a été découvert sur l’îleRobinson Crusoë, dans l’archipel Juan Fernández, au large du Chili, et est décritsous le nom de P. ×cerro-altoensis. Ses parents probables sont P. macrocarpa etP. masafuerae, deux espèces présentes dans l’archipel.
Palabras clavas : Pleopeltis, híbrido, Polypodiaceae, Juan Fernández, Chile
RESUMENUn híbrido de helecho que pertenece al género Pleopeltis ha sido descubierto enla isla Robinson Crusoe, en el archipiélago Juan Fernández, a la altura de Chile,y es descrito con el nombre de P. ×cerro-altoensis. Sus parientes probables sonP. macrocarpa y P. masafuerae, dos especies presentes en el archipiélago.
INTRODUCTIONSince 1997, the first author has undertaken, with the association ROBINSONIA1,regular expeditions in the Juan Fernández Archipelago, off the coast of Chile, to study
FERN GAZ. 20(2):65-78. 2015 65
1 French association registered under Law 1901, on 9th October 2001, involved in the inventoryand conservation of the Flora of Juan Fernández Archipelago, in relation with local communitiesand notably with the Corporación Nacional Forestal (CONAF) of Chile.

both the indigenous and the introduced flora of the three islands that constitute thisarchipelago (Figure 1): the main island, Robinson Crusoe (or Masatierra) and thenearby Santa Clara Island (also called Île aux Chèvres), and the third one, AlejandroSelkirk Island (or Masafuera), located 180 km further west.
During these expeditions, on 29th January 1999, a small isolated population of apolypodiaceous fern was discovered on a cliff of volcanic rocks, at Cerro Alto (Figures2 & 3) on Robinson Crusoe Island (Masatierra). The presence of peltate scales on bothsurfaces of the lamina and its axes showed that it belongs to the genus Pleopeltis Humb.& Bonpl. ex Willd. (Figures 4 & 7).
We compared this plant with the Pleopeltis species known from the area, but ourreview of botanical literature relating to Juan Fernández (Hemsley, 1884; Johow, 1892-93, 1896; Christensen & Skottsberg, 1920; Skottsberg, 1953; Barrera Moscoso, 1997;Marticorena et al., 1998; Boudrie, 2003; Danton & Perrier, 2006) and to continentalChile (Gay, 1853; Looser, 1961, 1962, 1965-66, 1968; de la Sota, 1967; Marticorena &Rodriguez, 1995; Zuloaga et al., 2008) has shown that it was not recorded in any of thefloras or floristic catalogues, nor any description applied to it. Therefore, whatever itsidentity or origin, we believe that it is a taxon new to the Juan Fernández Archipelago.
Two other species of Pleopeltis (previously placed in the genus Polypodium) occurin the Juan Fernández Archipelago (Figure 5):
- Pleopeltis macrocarpa (Bory ex Willd.) Kaulf., Berlin. Jahrb. Pharm.Verbundenen Wiss. 21: 41. 1820.Basionym: Polypodium macrocarpum Bory ex Willd., Sp. Pl. ed. 4, 5: 147. 1810.Syn. : Polypodium lanceolatum L., Sp. Pl. 2: 1802. 1753.Distribution: almost pantropical, America (from Central America to Chile andArgentina, Juan Fernández, Galapagos), tropical southern Africa, India.
- Pleopeltis masafuerae (Phil.) de la Sota, Darwiniana 45(2): 239. 2007.Basionym : Polypodium masafuerae Phil., Linnaea 29: 107. 1857.Distribution: Juan Fernández Archipelago (only on Alexander Selkirk Island =Masafuera), Chile (province of Antofagasta); mentioned from southern Peru(Marticorena & Rodriguez, 1995), but not confirmed to our knowledge.
Concerning the identity of P. masafuerae, its relationships with several South AmericanPleopeltis such as P. pinnatifida (Gillies) Hook. & Grev., P. pycnocarpa (C.Chr.)A.R.Sm. and Polypodium mollendense Maxon are unresolved (pers. comm. A.R. Smith,2015). Although the circumscription of Pleopeltis has recently been enlarged, basedboth on molecular and on morphological studies (Otto et al., 2009; Smith & Tejero-Díez, 2014), Neotropical Pleopeltis taxa are a taxonomically difficult group, withpolyploids and numerous hybrids, and a modern revision is needed to clarify thetaxonomy. Tryon & Stolze (1993), treating the Peruvian species (in Polypodium),alluded to these complexities.
Because neither of these two species matched the newly discovered plant, whichthus might represent a new taxon (Danton et al. 1999), a more careful study of it wasundertaken, as described below.
MATERIAL AND METHODSThe material used for this study comprises:
- Fronds collected in the field at Cerro Alto, now pressed and dried herbariumspecimens:
● Danton B(652)619 (P, SGO, private herb. Ph. Danton)
66 FERN GAZ. 20(2):65-78. 2015
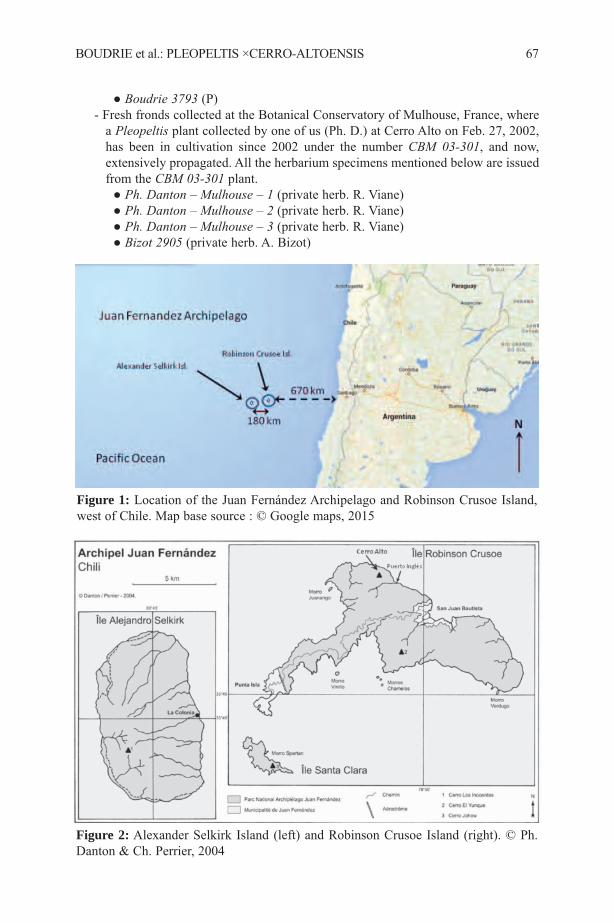
● Boudrie 3793 (P)- Fresh fronds collected at the Botanical Conservatory of Mulhouse, France, where
a Pleopeltis plant collected by one of us (Ph. D.) at Cerro Alto on Feb. 27, 2002,has been in cultivation since 2002 under the number CBM 03-301, and now,extensively propagated. All the herbarium specimens mentioned below are issuedfrom the CBM 03-301 plant.
● Ph. Danton – Mulhouse – 1 (private herb. R. Viane)● Ph. Danton – Mulhouse – 2 (private herb. R. Viane)● Ph. Danton – Mulhouse – 3 (private herb. R. Viane)● Bizot 2905 (private herb. A. Bizot)
BOUDRIE et al.: PLEOPELTIS ×CERRO-ALTOENSIS 67
Figure 1: Location of the Juan Fernández Archipelago and Robinson Crusoe Island,west of Chile. Map base source : © Google maps, 2015
Figure 2: Alexander Selkirk Island (left) and Robinson Crusoe Island (right). © Ph.Danton & Ch. Perrier, 2004

68 FERN GAZ. 20(2):65-78. 2015
Figure 3: Pleopeltis ×cerro-altoensis in situ at Cerro Alto. © Photo Ph. Danton,22/01/1999
Figure 4: Pleopeltis ×cerro-altoensis. © Ph. Danton, 25/01/1999. a - entire plant; b -portion of adaxial laminar surface; c - abaxial surface of fertile frond; d – peltate-basedlanceolate laminar scale; e – peltate-based lanceolate rhizome scale.

BOUDRIE et al.: PLEOPELTIS ×CERRO-ALTOENSIS 69
Figure 5 – Left: Pleopeltis masafuerae (Phil.) de la Sota, rocks at the beginning of theQuebrada Las Casas, Alejandro Selkirk Island, 19/12/2003, Danton G(1319)1083. –Right : Pleopeltis macrocarpa (Bory ex Willd.) Kaulf., epiphytic in forest under theMirador de Selkirk, Robinson Crusoe Island, 07/12/1998, Danton B(410)366. © PhotoPh. Danton, 26/05/2015.

70 FERN GAZ. 20(2):65-78. 2015
Figure 6: Pleopeltis ×cerro-altoensis Danton & Boudrie, Cerro Alto, Robinson CrusoeIsland, Juan Fernández Archipelago, holotype (SGO). Danton B(658)619. © Photo Ph.Danton, 17/11/2014.

BOUDRIE et al.: PLEOPELTIS ×CERRO-ALTOENSIS 71
Figure 7: Abaxial laminar surface of Pleopeltis ×cerro-altoensis, showing peltate-lanceolate scales and sori (from Bizot 2905). © Photo A. Bizot, 8/10/2014.
Figure 8: Peltate-based, lanceolate, fimbriate scale of abaxial laminar surface ofPleopeltis ×cerro-altoensis (from Bizot 2905). © Photo A. Bizot, 10/10/2014.

72 FERN GAZ. 20(2):65-78. 2015
Figure 9: Sporangium and malformed spores of Pleopeltis ×cerro-altoensis (fromBizot 2905). © Photos A. Bizot, 8/10/2014 & 18/10/2014.

- Dried herbarium specimens issued from the cultivated plant at Mulhouse undernumber CBM 03-301 :
● Boudrie 3966 (P)● Boudrie 4500 (P)● Danton Z(hs)1727 (private herb. Ph. Danton)
- Other Pleopeltis taxa from Mexico, deposited in the personal herbarium of one ofus (R. V.), but used, when freshly collected, for FCM analyses. Only one, P.guttata, is in cultivation in R.V.’s garden in Serskamp, near Ghent.
● Viane 11001: Pleopeltis guttata (Maxon) E.G. Andrews & Windham● Viane 11005: Pleopeltis polylepis (Roem. ex Kunze) T.Moore var. erythrolepis(Weath.) Wendt● Viane 11013: Pleopeltis polypodioides (L.) E.G.Andrews & Windham var.knoblochiana (Mickel) A.R.Sm. & Tejero
Herbarium specimens of the respective private herbaria of A.B., Ph. D. and R. V., citedabove, will be deposited in the future in official institutions.
To establish its characters and identity, several methods have been applied to thematerial mentioned above.
Micromorphological observations Micromorphological observations were carried out by A.B. using a microscope Bressertriocular TRM-301 microscope at 400x to 1000x enlargements, equipped with a BresserMicroCamLab-1 ocular camera. Fresh fronds from material in cultivation in theBotanical Conservatory of Mulhouse [Bizot 2905] were used.
- Laminar scales were observed (Figures 7 & 8) on fresh fronds. Rhizome scaleswere not microscopically observed. However, despite their more lanceolate shapethan the laminar scales, they show the same clathrate pattern of cells.
- Stomata: Using tweezers, three epidermial slices were excised from the abaxialside of the lamina in the middle of a median pinna. The epidermis was observedin glycerin at 1000x enlargement. Eighty (80) stomata were measured.
- Spores were observed (Figure 9) in glycerin liquid at 1000x enlargement. Theywere collected from several (3-4) sori of three different fresh fronds as well asfrom more than two sori of a frond collected in the field [Danton B(652)619].
Comparison of size of stomata of the Cerro Alto plant with stomata from plants of P.macrocarpa and P. masafuerae may reveal important information, but this investigationwas impossible owing to the lack of adequate fresh material.
Measurements of rhizomesThe width (rhizomes are slightly flattened) and the thickness of rhizomes of the 10specimens of P. macrocarpa, four specimens of P. masafuerae, and four specimens ofthe Cerro Alto plant were measured from dried herbarium specimens. Sectioned piecesof rhizomes were observed under a binocular and measured using a plastic ruler withthe smallest calibrated unit being 0.5mm.
DNA content using FCM A few fresh fronds (Botanical Conservatory of Mulhouse, France) of the Pleopeltisfrom Cerro Alto (in cultivation since 2002) were sent to the laboratory of Pteridologyof Ghent University (Belgium) on 6th October 2014 for flow cytometric measurements(FCM). For this analysis, fresh samples were first chopped, using a sharp razor blade,
BOUDRIE et al.: PLEOPELTIS ×CERRO-ALTOENSIS 73
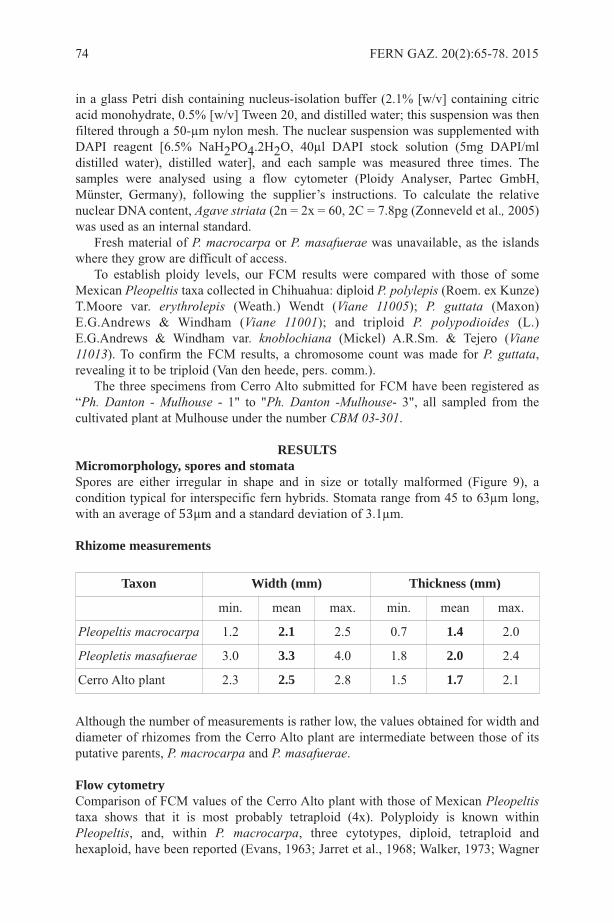
in a glass Petri dish containing nucleus-isolation buffer (2.1% [w/v] containing citricacid monohydrate, 0.5% [w/v] Tween 20, and distilled water; this suspension was thenfiltered through a 50-µm nylon mesh. The nuclear suspension was supplemented withDAPI reagent [6.5% NaH2PO4.2H2O, 40µl DAPI stock solution (5mg DAPI/mldistilled water), distilled water], and each sample was measured three times. Thesamples were analysed using a flow cytometer (Ploidy Analyser, Partec GmbH,Münster, Germany), following the supplier’s instructions. To calculate the relativenuclear DNA content, Agave striata (2n = 2x = 60, 2C = 7.8pg (Zonneveld et al., 2005)was used as an internal standard.
Fresh material of P. macrocarpa or P. masafuerae was unavailable, as the islandswhere they grow are difficult of access.
To establish ploidy levels, our FCM results were compared with those of someMexican Pleopeltis taxa collected in Chihuahua: diploid P. polylepis (Roem. ex Kunze)T.Moore var. erythrolepis (Weath.) Wendt (Viane 11005); P. guttata (Maxon)E.G.Andrews & Windham (Viane 11001); and triploid P. polypodioides (L.)E.G.Andrews & Windham var. knoblochiana (Mickel) A.R.Sm. & Tejero (Viane11013). To confirm the FCM results, a chromosome count was made for P. guttata,revealing it to be triploid (Van den heede, pers. comm.).
The three specimens from Cerro Alto submitted for FCM have been registered as“Ph. Danton - Mulhouse - 1" to "Ph. Danton -Mulhouse- 3", all sampled from thecultivated plant at Mulhouse under the number CBM 03-301.
RESULTSMicromorphology, spores and stomataSpores are either irregular in shape and in size or totally malformed (Figure 9), acondition typical for interspecific fern hybrids. Stomata range from 45 to 63µm long,with an average of 53µm and a standard deviation of 3.1µm.
Rhizome measurements
Although the number of measurements is rather low, the values obtained for width anddiameter of rhizomes from the Cerro Alto plant are intermediate between those of itsputative parents, P. macrocarpa and P. masafuerae.
Flow cytometryComparison of FCM values of the Cerro Alto plant with those of Mexican Pleopeltistaxa shows that it is most probably tetraploid (4x). Polyploidy is known withinPleopeltis, and, within P. macrocarpa, three cytotypes, diploid, tetraploid andhexaploid, have been reported (Evans, 1963; Jarret et al., 1968; Walker, 1973; Wagner
74 FERN GAZ. 20(2):65-78. 2015
Taxon Width (mm) Thickness (mm)
min. mean max. min. mean max.
Pleopeltis macrocarpa 1.2 2.1 2.5 0.7 1.4 2.0
Pleopletis masafuerae 3.0 3.3 4.0 1.8 2.0 2.4
Cerro Alto plant 2.3 2.5 2.8 1.5 1.7 2.1
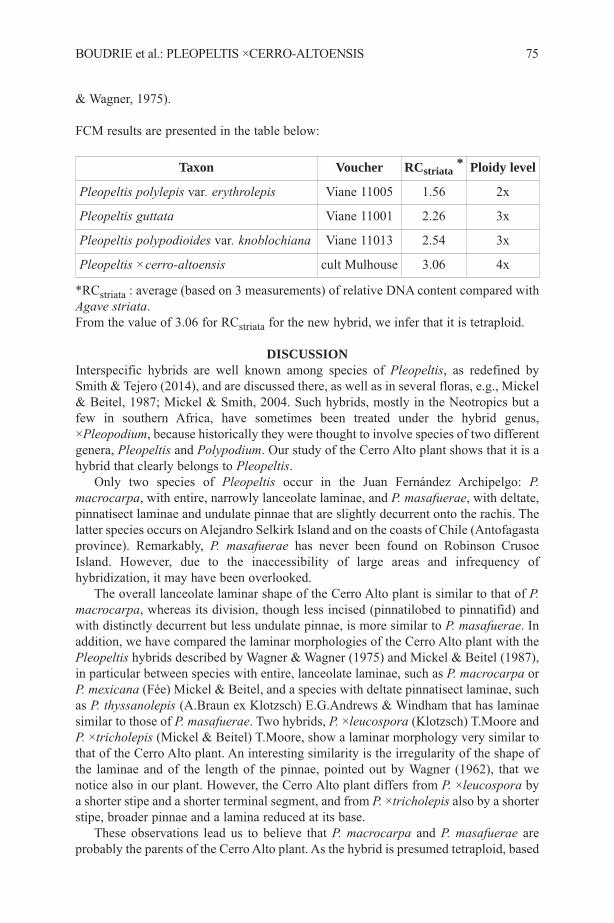
& Wagner, 1975).
FCM results are presented in the table below:
*RCstriata : average (based on 3 measurements) of relative DNA content compared withAgave striata. From the value of 3.06 for RCstriata for the new hybrid, we infer that it is tetraploid.
DISCUSSIONInterspecific hybrids are well known among species of Pleopeltis, as redefined bySmith & Tejero (2014), and are discussed there, as well as in several floras, e.g., Mickel& Beitel, 1987; Mickel & Smith, 2004. Such hybrids, mostly in the Neotropics but afew in southern Africa, have sometimes been treated under the hybrid genus,×Pleopodium, because historically they were thought to involve species of two differentgenera, Pleopeltis and Polypodium. Our study of the Cerro Alto plant shows that it is ahybrid that clearly belongs to Pleopeltis.
Only two species of Pleopeltis occur in the Juan Fernández Archipelgo: P.macrocarpa, with entire, narrowly lanceolate laminae, and P. masafuerae, with deltate,pinnatisect laminae and undulate pinnae that are slightly decurrent onto the rachis. Thelatter species occurs on Alejandro Selkirk Island and on the coasts of Chile (Antofagastaprovince). Remarkably, P. masafuerae has never been found on Robinson CrusoeIsland. However, due to the inaccessibility of large areas and infrequency ofhybridization, it may have been overlooked.
The overall lanceolate laminar shape of the Cerro Alto plant is similar to that of P.macrocarpa, whereas its division, though less incised (pinnatilobed to pinnatifid) andwith distinctly decurrent but less undulate pinnae, is more similar to P. masafuerae. Inaddition, we have compared the laminar morphologies of the Cerro Alto plant with thePleopeltis hybrids described by Wagner & Wagner (1975) and Mickel & Beitel (1987),in particular between species with entire, lanceolate laminae, such as P. macrocarpa orP. mexicana (Fée) Mickel & Beitel, and a species with deltate pinnatisect laminae, suchas P. thyssanolepis (A.Braun ex Klotzsch) E.G.Andrews & Windham that has laminaesimilar to those of P. masafuerae. Two hybrids, P. ×leucospora (Klotzsch) T.Moore andP. ×tricholepis (Mickel & Beitel) T.Moore, show a laminar morphology very similar tothat of the Cerro Alto plant. An interesting similarity is the irregularity of the shape ofthe laminae and of the length of the pinnae, pointed out by Wagner (1962), that wenotice also in our plant. However, the Cerro Alto plant differs from P. ×leucospora bya shorter stipe and a shorter terminal segment, and from P. ×tricholepis also by a shorterstipe, broader pinnae and a lamina reduced at its base.
These observations lead us to believe that P. macrocarpa and P. masafuerae areprobably the parents of the Cerro Alto plant. As the hybrid is presumed tetraploid, based
BOUDRIE et al.: PLEOPELTIS ×CERRO-ALTOENSIS 75
Taxon Voucher RCstriata * Ploidy level
Pleopeltis polylepis var. erythrolepis Viane 11005 1.56 2x
Pleopeltis guttata Viane 11001 2.26 3x
Pleopeltis polypodioides var. knoblochiana Viane 11013 2.54 3x
Pleopeltis ×cerro-altoensis cult Mulhouse 3.06 4x

on FCM values, we assume that tetraploid P. macrocarpa is involved in thecombination, and that P. masafuerae is also tetraploid.
We conclude that the Cerro Alto plant is a hybrid new to the Juan FernándezArchipelago.
Pleopeltis ×cerro-altoensis Danton & Boudrie, hyb. nov.Abortivis sporis, tetraploideum, hybridum filix est, repente rhizomate, 5-7 cm inlongitudine fronde, angustissime alato atrobrunneo petiolo, appressis, fimbriatispeltatisque paleis penitus pinnatifido lanceolato limbo atque infra atrobrunnea et superviridiluteola rhachidi. A P. macrocarpa pinno-pinnatifido limbo, et a P. masafuerae basireducta lanceolato limbo praecipue differt.
Types: CHILE. Archipel Juan Fernández, île Robinson Crusoe, Cerro Alto, QuebradaPuerto Inglés, rochers en falaise sur le versant est-sud-est du cerro, 300 m, 22 janvier1999, Danton B(652)619 [holotype: SGO (Figure 6); isotypes: P, private herb. P.Danton].
Description: Epilithic fern. Rhizomes creeping, 2.5-3 mm diam. (4-5 mm including thescales), branched, densely covered with oblanceolate, acute, clathrate, bicolorousscales, with dark centres and narrow, light brown, and finely dentate-fimbriate margins.Fronds monomorphic, 0.5-1 cm apart, 6-15 × 0.6-4 cm. Stipes 2-5 cm, about 1/3 - ¼ aslong as laminae, dark brown, with sparse, elongate scales. Laminae coriaceous, darkgreen, matt adaxially, light green to yellowish and slightly glaucous abaxially, narrowlyelliptic to lanceolate, deeply pinnatifid, tapering proximally to reduced lobes, and belowthat long-decurrent (young fronds entire to sinuately lobed), distally ending in a longpointed to obtuse segment; laminar surfaces covered with numerous, scattered, peltate-based, appressed, bicolourous lanceolate scales with dentate or fimbriate margins,scales more abundant abaxially than adaxially. Dark brown rachis colour ending at thebase of the apical segment abaxially, adaxially the colour ending at the first or secondproximal pinnae and becoming green-yellowish distally. Pinnae 5-7 per side, triangularto oblong, 24-26 × 5.5-6 mm (for the longest ones), of variable length, oblique to rachis,alternate to more opposite in the distal half of the laminae, apices obtuse to rounded;laminar margins cartilaginous and slightly recurved. Stomata (45)-53.5-(63) µm long.Sori round to slightly oblong, ca. 0.3 cm diam., exindusiate, mixed with peltate scales,median on both sides of the costae or (in less divided blades) of the rachis. Sporeswhitish to yellowish, malformed and irregular in shape and in size, (53)-73.5-(90) µmlong. Tetraploid (4x).
Etymology: The epithet of the taxon refers to the locality, Cerro Alto, where the plantwas found.
Distribution and habitat: To our knowledge, endemic to Robinson Crusoe Island(Masatierra) in Juan Fernández Archipelago (Chile), on southern flanks of Cerro Alto.Plants grow in a limited population of four small, relatively close patches, on smallrocky edges covered by thin soil, on subvertical unstable volcanic cliffs, exposed to thewinds, at 300 m elevation (Figure 3). Due to its ability to revive, it can tolerate periodsof drought.
76 FERN GAZ. 20(2):65-78. 2015

Specimens examined (paratypes): CHILE. Juan Fernández Archipelago, RobinsonCrusoe Island, Quebrada Puerto Inglés, rebords de rochers en falaise et balmes sur leversant est-sud-est du Cerro Alto, 300 m, M. Boudrie et session SBF, 20 février 2002,Boudrie 3793 (P).
Specimens ex horto examined (paratypes): CHILE. Juan Fernández Archipelago,Robinson Crusoe Island, Puerto Inglés, rebords de rochers en falaise et balmes sur leversant est-sud-est du Cerro Alto, 300 m, ex horto Conservatoire Botanique de la villede Mulhouse (Haut-Rhin, France), M. Boudrie et J.-P. Reduron, 22 août 2003, Boudrie3966 (P); idem, Ph. Danton et M. Hildebrand, 13 juin 2006, Z(hs)1727 (private herb.Ph. Danton); idem, M. Boudrie et J.-P. Reduron, 5 octobre 2009, Boudrie 4500 (P);idem, M. Hildebrand, 29 septembre 2014, Bizot 2905 (private herb. A. Bizot), and P.Danton - Mulhouse - 1, 2 & 3 (private herb. R. Viane).
IUCN Status: “CR”
Conservation ex situ: A plant was previously cultivated, from 1999 to 2004, in thegarden of the Administration of the National Park, on Robinson Crusoe Island, but isnow dead. Another specimen has been maintained and propagated in the BotanicalConservatory of the town of Mulhouse (France) since 2002.
ACKNOWLEDGEMENTSWe thank Mr. Christian Diaz (San Juan Bautista, Isla Robinson Crusoe, Chile), theGuards of the Parque Nacional Archipiélago Juan Fernández (San Juan Bautista, IslaRobinson Crusoe, Chile), MM. Hernan Gonzalez (San Juan Bautista, Isla RobinsonCrusoe, Chile) and Ivan Leiva (San Juan Bautista, Isla Robinson Crusoe, Chile). We arealso extremely grateful to Dr. Caroline Van den heede for the chromosome count ofPleopeltis guttata, Dr. Jacques Florence (IRD-MNHN, Paris) for the translation of thediagnosis into Latin, Dr. Alan R. Smith (University of Berkeley, USA) for usefulrecommendations, Mathieu Hildebrand (Mulhouse, France), Christophe Perrier (Saint-Crépin, France), Jean-Pierre Reduron (Mulhouse, France), as well as to the SociétéBotanique de France (Paris, France).
REFERENCESBARRERA MOSCOSO E., 1997. Helechos de Juan Fernández. Publ. Ocas. Mus. Nac. Hist.
Nat., 51: 5-104.BOUDRIE M., 2003. Les ptéridophytes de l’île Robinson Crusoe (archipel Juan
Fernández, Chili). J. Bot. Soc. Bot. France 24: 32-48.CHRISTENSEN C. & SKOTTSBERG C., 1920. The Pteridophyta of Juan Fernandez Islands.
In Skottsberg C., Natural History of Juan Fernandez and Easter Island. Vol. II.Botany. Almqvist & Wiksells Eds., Uppsala, 46 p.
DANTON P., BRETEAU E. & BAFFRAY M., 1999. Les îles de Robinson. Trésor vivant desmers du Sud – Entre légende et réalité. Yves Rocher, Nathan, Paris, 144 p.
DANTON P. & PERRIER C., 2006. Nouveau catalogue de la flore vasculaire de l’archipelJuan Fernández (Chili). Acta Bot. Gallica 153 : 399-587.
EVANS A.M., 1963. New chromosome observations on the Polypodiaceae andGrammitidaceae. Caryologia 16: 671-677.
GAY C., 1853. Historia fisica y politica de Chile, Botanica. Tome VI. C. Gay (Ed.),
BOUDRIE et al.: PLEOPELTIS ×CERRO-ALTOENSIS 77

Paris, Santiago, Chile, 531 p.HEMSLEY W.B., 1884. Report on the Botany of Juan Fernandez, the South-Eastern
Moluccas and the Admiralty Islands [Voyage of H.M.S. Challenger]. JuanFernandez and Masafuera. Botany, vol. 1, part 4, 100 pp. + Plates LIV-LWIII.
JARRETT F.M., MANTON I. & ROY S.K., 1968. Cytological and taxonomic notes on asmall collection of living ferns from Galapagos. Kew Bull. 22: 475-480.
JOHOW F., 1892-93. Los Helechos de Juan Fernández. Anales Univ. Chile 82: 741-757,977-1005.
JOHOW F., 1896. Estudios sobre la Flora de las Islas de Juan Fernández. ImprentaCervantes, Santiago de Chile, XI + 289 p. + XVIII láminas + 2 mapas.
LOOSER G., 1961. Los Pteridofitos o helechos de Chile I. Revista Univ. (Santiago) 46:213-262.
LOOSER G., 1962. Los Pteridofitos o helechos de Chile (Excepto Isla de Pascua) II.Revista Univ. (Santiago) 47: 17-31.
LOOSER G., 1965-66. Los Pteridofitos o helechos de Chile (Excepto Isla de Pascua) III.Revista Univ. (Santiago) 50-51: 75-93.
LOOSER G., 1968. Los Pteridofitos o helechos de Chile (Excepto Isla de Pascua) IV.Revista Univ. (Santiago) 53: 27-39.
MARTICORENA C. & RODRIGUEZ R. (Eds.), 1995. Flora de Chile. Vol. I, Pteridophyta –Gymnospermae. Universidad de Concepción, Concepción, Chile.
MARTICORENA C., STUESSY T.F. & BAEZA C. M., 1998. Catalogue of the vascular floraof the Robinson Crusoe or Juan Fernandez Islands, Chile. Gayana, Bot. 55: 187-211.
MICKEL J.T. & BEITEL J.M., 1987. Notes on ×Pleopodium and Pleopeltis in TropicalAmerica. Amer. Fern J. 77: 16-27.
OTTO E.M., JANSSEN T., KREIER H.-P. & SCHNEIDER H., 2009. New insights into thephylogeny of Pleopeltis and related Neotropical genera (Polypodiaceae,Polypodiopsida). Molec. Phylogen. Evol. 53: 190-201.
SKOTTSBERG C., 1953. A supplement to the Pteridophytes and Phanerogams of the JuanFernandez and Easter Island. The Natural History of Juan Fernandez and EasterIsland. 2: 763-792.
SMITH A.R. & TEJERO-DIéZ J.D., 2014. Pleopeltis (Polypodiaceae), a redefinition of thegenus and nomenclatural novelties. Bot. Sci. 92: 43-58.
SOTA E.R. DE LA, 1967. Sinopsis de las familias y géneros de Pteridófitas de Argentina,Uruguay y Chile (Incluyendo las Islas Juan Fernández y Pascua). Revista Mus.LaPlata, Secc. Bot. 10: 187-221.
TRYON R.M. & STOLZE R.G., 1993. Pteridophyta of Peru. Part V. 18. - Aspleniaceae. -21. Polypodiaceae. Fieldiana, Bot. 32: 1-190.
WAGNER W.H., 1962. Irregular morphological developments in hybrid ferns.Phytomorphology 12: 87-100.
WAGNER JR. W.H. & WAGNER F.S., 1975. A hybrid polypody from the New Worldtropics. Fern Gaz. 11: 125-135.
WALKER T.G., 1973. Additional cytotaxonomic notes on the pteridophytes of Jamaica.Trans. Roy. Soc. Edinburgh 69: 109-135.
ZULOAGA F., MORRONE O. & BELGRANO M., 2008. Catálogo de las Plantas Vascularesdel Cono Sur (Argentina, Sur de Brasil, Chile, Paraguay y Uruguay). Vol. 1:Pteridophyta, Gymnospermae, Monocotyledoneae. Missouri Botanical GardenPress, St. Louis, USA.
78 FERN GAZ. 20(2):65-78. 2015

A CONTRIBUTION TO THE PHYLOGENY OF DRYOPTERISREMOTA BY GENOTYPING OF A FRAGMENT OF THE NUCLEAR
PgiC GENE
W. BUJNOCH
Associated contributor of the Biogeographic Institute of Trier University / GermanyNeuwiese 13, D-54296 Trier, Germany
Email: [email protected]
Keywords: Dryopteris remota, PCR-direct, PgiC 15/16 fragment, genotyping,phylogeny
ABSTRACTSince its discovery by A. Braun in 1834, there has been speculation on theorigins of Dryopteris remota. The present contribution suggests that none of theprevious assumptions are correct. Dryopteris remota is triploid and of hybridderivation. Its parental species are the sexual allotetraploid D. carthusiana andan as yet unidentified sexual diploid species, most likely identical with theunknown parent of the apomictic diploid D. affinis. These assertions are basedon comparison of traits, including habit, apomixis, colour of insertions of pinnaeof the species under examination, and fragment length variation of the nuclearPgiC gene.
INTRODUCTIONThe phylogeny and taxonomic status of Dryopteris remota (A.Br.) Druce is probablyone of the most controversially discussed of any Central European fern species. WhenAlexander Braun discovered the plant near Baden-Baden in 1834 (Baden-Württemberg/S-Germany), he thought the new taxon to be a variation of D. villarii (Bellardii) Woynarex Schinz & Tell. As such it was listed in 1843 by J. C. Döll in “Rheinische Flora”. Inhis book “Betrachtungen über die Erscheinung der Verjüngung in der Natur” of 1851,Braun revised his view in a footnote and described the taxon as a hybrid of D. filix-mas(L.) Schott and D. carthusiana (Vill.) H.P.Fuchs or D. dilatata (Hoffm.) A.Gray. Henonetheless attributed species rank to this taxon by calling it “Aspidium remotum”, i.e.in current nomenclature D. remota. Since then a substantial number of renowned fernexperts have speculated on its origin and taxonomic classification. In 1973 Benl andEschelmüller presented an exhaustive description of this continuing dispute amongscientists.
Further confusion had arisen following the discovery of a fern hybrid near LakeWindermere in north-west England in the middle of the 19th century. The morphologyof this hybrid resembled very much that of D. remota, but it had abortive spores. In1859 A. Braun found a single plant of D. remota among a large population of D. filix-mas near Aachen (Nordrhein-Westfalen/ Germany), which led him to theconclusion that D. remota must be a variation of D. filix-mas. He did not realize thatthis plant was not D. remota but actually the same hybrid as that from LakeWindermere. He transplanted this hybrid to the Botanical Garden of Berlin and forapproximatively 15 years he sent incorrectly labelled exsiccata from there to herbariaand to individual scientists before this mistake was discovered (Benl & Eschelmüller,
FERN GAZ. 20(2):79-89. 2015 79

1973). Further examination of a frond sample in the herbarium at Bonn revealed thatBraun’s finding was the second record of the tetraploid hybrid D. carthusiana × D. filix-mas, as discovered earlier at Lake Windermere (Krause et al., 2001), and whichhas now been typified as D. × brathaica Fraser- Jenk. & Reichst. (1977). It is nowpossible to distinguish the two taxa through cytology (Manton, 1950), morphology(Fraser-Jenkins & Reichstein, 1984; Krause et al., 2001; Freigang et al., 2013),chemotaxonomy (Widén et al., 1976) and molecular genetics (Freigang et al., 2013).
Dryopteris remota is today recognised as a triploid species of hybrid derivation, butthe dispute concerning its origin continues. From fundamental works by W. Döpp(1932; 1933; 1935; 1939; 1955) and I. Manton (1932; 1938; 1939; 1950), Döppconsidered D. remota to be the hybrid of D. dilatata × D. affinis (Lowe) Fraser-Jenk.(Döpp, 1955). Reichstein, Fraser-Jenkins and others are of the opinion that the diploidapomictic D. affinis must be one of the parents due to the characteristics of D. remota(Fraser-Jenkins & Reichstein, 1984). In consequence the second parent would have tobe a diploid sexual taxon that contributes the characteristics known from the D. carthusiana group to the genotype of D. remota. Dryopteris expansa (C.B.Presl)Fraser-Jenk. & Jermy was proposed (Widén et al., 1971, Fraser-Jenkins & Reichstein,1984), even though chemo-taxonomic results did not match well (Widén et al., 1970;1971; 1996). Furthermore, in an on-line discussion of the British Fern Group,Fraser-Jenkins (30 December 2008) reiterated doubts as to D. expansa being involved:“Although I agree that D. expansa is the most likely second ancestor of D. remota, itought to have a ? along that line as it has never been proved in any way and thepossibility of some other, even perhaps Asiatic species being involved still remains.”
To shed light on the ongoing discussions, the nuclear PgiC-marker of many speciesof the genus Dryopteris, including D. remota, has been examined.
MATERIAL AND METHODSMaterialThe samples used in the investigation are listed in Table 1. Herbarium vouchers,currently in the collectors’ private herbaria will be deposited in JE.
MethodsSamples were analyzed at the nuclear PgiC gene (cytosolic 6-Phosphoglucose-isomerase), individual fragments were generated and their lengths compared. Theintron fragment between exons 15 and 16 proved especially suitable for comparison asit shows a species specific length for a number of species of the genus Dryopteris,especially in the D. carthusiana group (Juslén et al., 2011, Freigang et al., 2013). Forthis fragment Ishikawa et al., 2002 constructed an ‘EPIC’- primer pair (Exon PrimedIntron Crossing), which offers almost general applicability. Due to a high frequency ofpoor PCR yields or complete PCR failures, the method described in Freigang et al.(2013) was partly replaced by the PCR-direct method of ThermoScientific (Phire PlantDirect PCR Kit, F-130WH). This method worked without DNA isolation and PCRyields were often higher and, according to the producer instructions the workflowconsisted of only two steps. 1. A disc of 0.3-0.5 mm diameter was punched out from fresh or silica gel-dried leafmaterial and put into PCR-solution. 2. PCR was carried out according to the producerinstructions. The uncleaned amplicon was genotyped in a sequencer (ABI 3500,Applied Biosystems or MegaBace Instrument, Healthcare). Employment of different
80 FERN GAZ. 20(2):79-89. 2015

sequencers made transformation of results necessary (marked with an asterisk). Theanalysis of the generated data was first made with Fragment Profiler software(Healthcare), then later with GeneMapper software (Applied Biosystems).Sometimesthe sequencing device produced a double peak in consecutive basepairs (bp). This couldbe an artefact or may be the result of PCR-conditions.
The relative height of signals within an electropherogram depended firstly on thenumber of sets of chromosomes causing the signal and, secondly, on the relative PCRyield of the 15/16-fragment of the existing alleles.
RESULTSThe genotypes of samples 1 to 24 are summarised in Table 2. Signals ofelectropherograms which were not separated are noted in the same table-field.Electropherograms of a subset of samples are shown in Figures 1-3 (underlined inTables 1 and 2).
DISCUSSIONIn the case of apomictic reproduction in ferns the sporophyte grows directly from agametophyte with the unreduced number of chromosomes, and without gametes beinginvolved. The sporophyte produces genetically uniform diplospores through arestitution nucleus; in the case of apomicts of Dryopteris, restitution occurs in the lastmitotic cell division before the production of spore mother cells (Manton, 1950),resulting in the production of eight, rather than the usual 16 spore mother cells.Sporogenesis then results in the production of 32 (instead of the usual 64) spores, eachwith the same number of chromosomes as the sporophyte.
Fischer (1909; 1919) detected apomixis in D. remota and Manton demonstrated thatit is a triploid taxon, with three different (non-homologous) chromosome sets (i.e. aheterotriploid fern) as in the occasional production of a spore mother cell of the ‘16 celltype’, the chromosomes are unpaired (non-homologous), with 123 univalents (3x). Theheterotriploid character is also shown by the electropherogram of PgiC 15/16 fragmentsof the species (Figure 1), where three distinct peaks are observed.The electropherogramshows the following: The genome of D. remota contains at least three chromosome setspossessing fragments of different length and therefore derived from different alleles.This supports the results of cytology that the species is heterotriploid. The two shorterfragments of 488 bp and 500 bp correspond to the results seen in D. carthusiana (Figure2), an allotetraploid species; no other taxon of the European ferns of the genusDryopteris examined up to now shows this pattern.
Comparison with D. cristata (L.) A.Gray (sexual allotetraploid) and D. intermedia(Mühl. ex Willd.) A.Gray (sexual diploid) reveals that the peak at 488 bp differs fromthat of D. intermedia (500 bp), and is generated by the chromosome set derived fromthe common parental taxon of D. cristata and D. carthusiana, referred to as D. “semicristata” (Sc) by Walker (1955), a diploid species as yet unknown. The signalat 500 bp in D. remota is identical with that of D. intermedia (I). The chromosome setsof D. carthusiana can be symbolized by ScScII. The assumption that the parentalspecies of D. carthusiana are D. intermedia and D. “semicristata” is supported (Table2) and confirms the result of Sessa et al. (2012). The results are also in agreement withchloroplast sequence data by Juslén et al. (2011), which showed that D. carthusiana andD. remota share a maternally inherited set of chromosomes, referred to as“semicristata”. Table 2 shows that the 15/16-fragment length of D. expansa is the same
BUJNOCH: THE PHYLOGENY OF DRYOPTERIS REMOTA 81

82 FERN GAZ. 20(2):79-89. 2015
Sample no. &species Origin
Collector/Determination/
DateInternal Data
1. Dryopterisaemula
Portugal/ Azores/ São Miguel/ N-side and ridge at Pico da Vara,1100 m MSL
GZ 28.5.2013Dr 8
MPb1458,DE807,RUN32/2H
2. D. affinis Portugal/ Madeira/ N Funchal/ c.600 m S of restaurant Ribeiro Frioat the roadside EN 103
JF/GZ25.8.2011JFV1086
MPb1724,RUN45/7A
3. D. affinis
Germany/ RP/ Palatinate/ NEWilgartswiesen, rock atWiligartsburg, ca. 230 m MSL, RF6713/32
WB/GZ6.8.1996
DE032,RUN21/11B
4. D.carthusiana
Germany/ NRW/ MärkischesLand, SW Hagen/ near Hasperdam, RF 4603
HWB27.05.2013
MPb1442,DE791,RUN32/1B
5. D.carthusiana
Germany/ NRW/ niederrheinischesTiefland/ E Venlo/ Nettetal/Poelvenn–Kuhlen/ RF 4603/23
N.Neikes05.07.2009
MPb1482,DE812,RUN32/3E
6. D.carthusiana
Germany/ RP/ middle Mosel/ WTrier/ Sirzenicher Bachtal, RF6205/23
WB 17.7.2005 DE083,RUN22/10B
7. D.carthusiana
Germany/ BW/ Oberschwaben/NE Ravensburg/ SE Bergatreute/near Kiliansweiher, RF 8124/32
JF 22.10.2006JF 2006
DE385,RUN20/12A
8. D. cristataGermany/ RP/ Eifel/ SSE Daun/Schalkenmehrener Maar, RF5807/13
WB 28.8.1984GWBT2 sporecultured
DE024RUN20/G11
9. D. dilatata
France/ Haut–Rhin/ Vosges/ S Colde la Schlucht/ Sentier des Roches/NE “les Trois Four“, 1050 mMSL, RF 7908/31
GZ/P.Holveck27.8.2014 Dr 4
MPb1665,RUN40/11D
10. D. expansa
Germany/ BY/ upper Allgäu/West–Grünten/ N Burgberg/ N„Weinhalde“, northern slope, 780–795 m MSL, RF 8427/41
GZ 19.7.2004Dr 20
MPb1229,DE695,GWBT156RUN28/11E
11. D. expansaSwitzerland/ Wallis/ S Leukerbad/Dala Valley/ lower Lochwald,1260 m MSL, RF 9611/23
GZ 18.7.2000Dr 1
MPb1230DE696,GWBT182RUN28/11F
Table 1: Genotyped samples (Nomenclature according Fraser-Jenkins & Reichstein,1984 and Fraser-Jenkins, 2007). RF: numbers according to rasters (see Niklfeld, 1971).Abbreviations of the names of collectors: GZ (G. Zenner/ Kirn), HWB (H. W. Bennert/ Ennepetal), JF (J. Freigang/ Bergatreute), SJ (S. Jeßen/ Chemnitz), WB (W. Bujnoch, Trier).

BUJNOCH: THE PHYLOGENY OF DRYOPTERIS REMOTA 83
12. D.intermedia
USA/ Vermont/ Rutland/ LeicesterHollow Trail/ NNE Brandon
HWB15.8.2001 SJ-3476
DE368RUN25/A11
13. D. oreades
France/ Hérault/ HautLanguedoc,/W Bédarieux/ NColombières-s-Orb/ near “LaFage”, c. 760 m MSL
GZ/WB23.7.1994 Dr 1
DE067, GZ-58,RUN18/11C
14. D. oreades
Italy/ Toscana/ Appennino ToscoEmiliano/ NW Pistoia/ W Abetone/W M Gomito/Val di Luce, c. 1480m MSL
SJ/J.Müller12.8. 1992 SJ-2226
DE288,RUN19/11B
15. D. pallidasubsp. pallida
Italy/ Sicily/ Mount Etna aboveZafferana
JF 1.1.2007JFV542
MPb1562DE861RUN37/B12
16. D. remota
Germany/BW/ Northern–Schwarzwald/ SSE Bad Herrenalb/valley of Rothenbach, 440 mMSL, RF 7216/21
GZ/WB1.8.1990spore cultured
FZ-90/7,GWBT 32,DE26,RUN19/12G
17. D. remota
Germany/ BY/ W Berchtesgaden/Ramsau/ SW ofSchwarzbachwacht, c. 900 mMSL, RF 8343/31
GZ/JF 5.8.2013Dr 2,
MPb1516, DE 824,RUN35/11B
18. D. remotaSchwitzerland/ Berner Oberland/Innertkirchen/ Urbachtal inVordertalwald, RF 9315/12
SJ 21.7.1992SJ-2270
MPb1235, DE637, RUN28/11C
19. D. remotaAustria/ Salzburg/ Pinzgau/ ERauris/ valley of Geißbach, c.1060 m MSL, RF 8744/31
GZ 6.8.2014 Dr 10
MPb1650,RUN44/11H
20. D. remota
Romania/ S-Karpaten/ N- Fǎgǎraș/NW des Rifugio “Negoiu”/ SSEPorumbacu de Sus/ Şerbota Tal c.1000 m MSL
SJ 5.8.1985 SJ-2560-2
MPb1236,DE638,RUN28/ 11D
21. D. remotaGermany/ BY/ West-Allgäu/ NWScheidegg/ forest N “Lötz”, c. 700m MSL, RF 8425/11
JF 4.6.2011 JF-V1008
MPb1250,DE710,RUN28/12D
22. D. remota
North-Italy/ Prov. Varese /E Luino/W Ponte Tresa/ Tresa-Valley nearBiviglione, 250 m MSL, RF0018/21
GZ/JF/SJ15.6.2014 Dr33
MPb1580,RUN40/9D
23. D. remotaNortheast-Turkey/ Rize/ ETrabzon, S Ardeşen, Cat -Ҫamlihemşin, c. 400m MSL
SJ-3026, TR-4011, CRFJ4037,27.08.1973
MPb1738,RUN47/A1
24. D. villarii
Bulgaria / North-Pirin/ E desKameniski vrah/ NW des Vihren/W Bansko, Bela-reka-Valley, 2150m MSL
SJ 24.7.1981SJ-335
MPb1452DE801,RUN32/2C
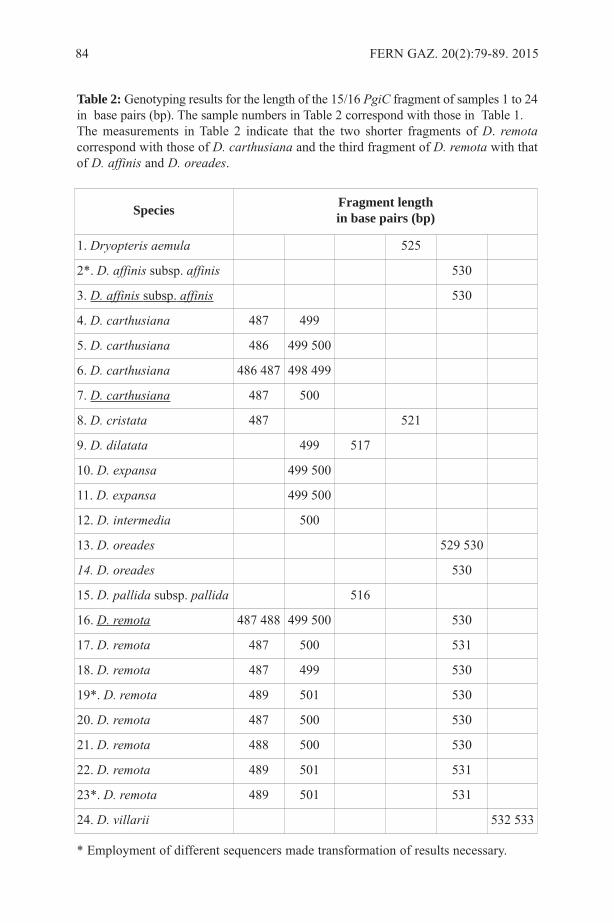
84 FERN GAZ. 20(2):79-89. 2015
Species Fragment lengthin base pairs (bp)
1. Dryopteris aemula 525
2*. D. affinis subsp. affinis 530
3. D. affinis subsp. affinis 530
4. D. carthusiana 487 499
5. D. carthusiana 486 499 500
6. D. carthusiana 486 487 498 499
7. D. carthusiana 487 500
8. D. cristata 487 521
9. D. dilatata 499 517
10. D. expansa 499 500
11. D. expansa 499 500
12. D. intermedia 500
13. D. oreades 529 530
14. D. oreades 530
15. D. pallida subsp. pallida 516
16. D. remota 487 488 499 500 530
17. D. remota 487 500 531
18. D. remota 487 499 530
19*. D. remota 489 501 530
20. D. remota 487 500 530
21. D. remota 488 500 530
22. D. remota 489 501 531
23*. D. remota 489 501 531
24. D. villarii 532 533
Table 2: Genotyping results for the length of the 15/16 PgiC fragment of samples 1 to 24in base pairs (bp). The sample numbers in Table 2 correspond with those in Table 1.The measurements in Table 2 indicate that the two shorter fragments of D. remotacorrespond with those of D. carthusiana and the third fragment of D. remota with thatof D. affinis and D. oreades.
* Employment of different sequencers made transformation of results necessary.

as that of D. intermedia. As mentioned in the introduction, the involvement of D. expansa in the parentage of D. remota has been the subject of much discussion. Butthis scenario would entail that the second parent of D. remota must be a diploidapomictic taxon of the D. affinis agg. with signals at 488 bp and 530 bp. Despite studyof many taxa of the D. affinis agg., this combination has never been found. The sameapplies to the combination of signals at 500 and 530 bp.
During the first half of the last century D. remota was thought to be a hybrid of D. carthusiana × D. filix-mas, and Döpp endeavoured to backcross D. remota with thesupposed parental species. He was successful in hybridizing D. remota only with D. carthusiana (Döpp, 1935). This provides some support for the conclusion that D. carthusiana is one parent of D. remota.
Already H. Fischer pointed to the fact that such a hybrid should also emerge innature and he mentioned that the pharmacist H.K. Woynar (Graz) allegedly foundintermediate forms in mixed stands of the two species at different sites in Tirol. Since2011 putative hybrids of D. remota × D. carthusiana have been found at variouslocations in the wild (Freigang et al., in prep.).
The remaining question relates to the unknown identity of the third signal in theelectropherogram of D. remota, at 530 bp. The chromosome set causing this third signalat 530 bp would have to contribute two qualities to D. remota that are characteristic ofit and cannot be derived from the genetic material of D. carthusiana - the obligatoryapomixis and the blue-black colour at the insertion of the pinnae (Fraser-Jenkins &Reichstein, 1984). Both features also exist in the D. affinis agg., but not in that of D. filix-mas. Examination of the diploid taxon D. affinis (Figure 3) shows a single peakat 530 bp.
According to current opinion D. affinis is a diploid apomict fern that showsnon-homology of the two chromosome sets in 16-celled sporangia (Fraser-Jenkins,2007) and it is presumed to have been derived from a cross between the sexual diploidD. oreades Fomin (OO) and a sexual diploid taxon, not yet identified but referred to asD. “semiaffinis” (SaSa) (Krause, 1998). The PgiC 15/16 fragments of both taxacorrespond to the third peak in D. remota. As D. remota is triploid and its genome
BUJNOCH: THE PHYLOGENY OF DRYOPTERIS REMOTA 85
Figure 1. Electropherogram of Dryopteris remota (Schwarzwald, Smp.No. 16)
Figure 2. Electropherogram of Dryopteris carthusiana (Bergatreute, Smp. No. 7)
Figure 3. Electropherogram of Dryopteris affinis (Wilgartswiesen, Smp. No. 3)

contains two sets of chromosomes of allotetraploid D. carthusiana, D. affinis as thesecond parent is out of question, because as an apomictic taxon it would contribute bothits chromosome sets to D. remota, resulting in a tetraploid taxon. This implies that thethird peak (530 bp) in D. remota must be derived from a diploid sexual taxon with the530 bp peak, and the only possible candidate from the results presented here are D. oreades or the missing diploid taxon D. “semiaffinis”.
Unfortunately the length of PgiC 15/16 fragments of D. oreades and D. “semiaffinis” cannot be distinguished with the method used here. Of these two, D. “semiaffinis” is the more likely parent; D. oreades does not exhibit blue-blackinsertions, nor apomixis. We are left to infer that both diploid D. affinis and triploid D. remota have inherited these characteristics from D. “semiaffinis”, and hence thephylogeny of D. remota could be analogous to that of D. affinis, as shown in Table 3.
All the evidence suggests that “semiaffinis” induces apomixis in its hybrids andspecies of hybrid derivation, in contrast to hybrids involving D. oreades. This is thecase in all heterodiploid as well as heterotriploid taxa of the D. affinis complex, whichare all fertile but to varying degrees. In hybrids of taxa of the D. affinis complex withD. filix-mas, apomixis is also expressed, despite that fact the “semiaffinis” chromosomeset is only one of four or five sets in the nucleus. The hybrid of D. carthusiana and D. “semiaffinis” (Table 3) has three non-homologous sets of chromosomes and wouldbe expected to be sterile, in contrast to D. remota which is fertile and can produce viableoffspring in the wild. This case of D. remota, gives further support to the hypothesis that
86 FERN GAZ. 20(2):79-89. 2015
Parental Taxa D. oreadeshomodiploid, sexual
D. “semiaffinis”homodiploid, sexual
D. carthusianaallotetraploid, sexual
Chromosome sets ofa somatic cell
Chromosome set(s)of a gamete
Hybridisation
Primary hybrid
Development ofapomixis
Progeny
chromosome sets of asomatic cell
OO SaSa IScISc
O Sa ISc
OSa SaIScheterodiploid, sterile heterotriploid, sterile
D. affinis D. remota
OSa SaIScheterodiploid, heterotriploid,
apomictic apomicticfertile fertile
Table 3. Diagram of the hypothetical genesis of Dryopteris remota and D. affinisAbbreviations of chromosome sets: O = oreades, Sa = “semiaffinis“, I = intermedia,Sc = “semicristata“

the set of chromosomes from “semiaffinis” has the ability to transfer the qualities ofdark insertion of pinnae, and apomixis coupled with fertility.
The distribution of D. remota lies between the Pyrenees and the Caucasus and thusfalls within the distribution of D. carthusiana and D. affinis (Fraser-Jenkins &Reichstein, 1984). The genetic variability of D. remota was examined by theisozyme-method (Schneller & Holderegger, 1994) and also by RAPDs (Schneller et al.,1998). The first method did not reveal any genetic variability within or betweenpopulations, the second only little. The authors suggested that the species developedrelatively recently and only once, presumably in Caucasus or the Mediterranean region,and spread over the now populated area by ‘long-distance spore dispersal’. Since sporesof apomicts produce sporophytes without the need for sexual fertilization, then anindividual spore has the potential to give rise to a new individual or eventually even anew population.
During the past 180 years of discussion about the origin of D. remota other taxahave been proposed as possible parents, for example D.villarii (Braun in Döll, 1843),D. dilatata (Döpp, 1935; 1955), D. aemula (Aiton) O.Kuntze (Widén et al., 1971) orD.pallida subsp. pallida (Bory) Fomin and subsp. raddeana Fraser-Jenk. (Peroni et al.,1991). The results in Table 2 rule out almost all of these possibilities, with the exceptionof D. pallida subsp. raddeana, as samples of this taxon were not included in the study.
ACKNOWLEDGEMENTSI would like to thank Prof. Dr. M. Veith, head of the Institute of Biogeography at TrierUniversity, for providing me with a working place and also PD. Dr. A. Hochkirch, headof laboratory and the technicians at the laboratory. In particular I would like thank mycolleagues, listed in Table 1, who made samples available for me. I would like to thankS. Jeßen and G. Zenner for their discussion and their valuable suggestions, also E.Erfurth and J. Beninde who helped with the translation of this paper into English.Finally I would like to thank the two reviewers and the editor for help and advice.
REFERENCESBENL,G. & ESCHELMüLLER, A. 1973. über „Dryopteris remota“ und ihre
Vorkommen in Bayern. Ber. Bayer. Bot. Ges. 44: 101-141.BRAUN, A. 1851. Betrachtungen über die Erscheinung der Verjüngung in der Natur,
ins- besondere in der Lebens- und Bildungsgeschichte der Pflanzen. Engelmann,Leipzig.
DöLL, J. C. 1843. Rheinische Flora. Brönner, Frankfurt a. M.DöPP, W. 1932. Die Apogamie bei Aspidium remotum A.Br. Planta 17: 86-152.DöPP, W. 1933. Weitere Untersuchungen an apogamen Farnen. Ber. Dtsch. Bot. Ges.
51: 341-347.DöPP, W. 1935. Versuche zur Herstellung von Artbastarden bei Farnen, I. Ber.Dtsch.
Bot. Ges. 53: 630-636.DöPP, W. 1939. Cytologische und genetische Untersuchungen innerhalb der Gattung
Dryopteris. Planta 29: 481-533.DöPP, W. 1955. Experimentell erzeugte Bastarde zwischen Dryopteris filix-mas (L.)
und D. paleacea (Sw.) C.Chr. Planta 46: 70-91.FISCHER, H. 1909. über Aspidium remotum A.Br.: Kreuzung oder Mutation? – Ein
neuer Fall von Apogamie. Ber. Dtsch. Bot. Ges. 27: 495-502.FISCHER, H. 1919. Apogamie bei Farnbastarden. Ber. Dtsch. Bot. Ges. 37: 286-292.
BUJNOCH: THE PHYLOGENY OF DRYOPTERIS REMOTA 87

FRASER-JENKINS, C. R. & REICHSTEIN, T. 1977. Dryopteris ×brathaicaFraser-Jenk. & Reichst. hybr. nov., the putative hybrid of D. carthusiana × D.filix-mas. Fern Gaz. 11: 337.
FRASER-JENKINS, C. R. & REICHSTEIN, T. 1984. Dryopteris. In: Kramer, K. U.(ed.) Hegi, Illustrierte Flora von Mitteleuropa, 1(1), ed. 3, pp. 136-169. Paul Parey,Berlin & Hamburg.
FRASER-JENKINS, C. R. 2007. The species and subspecies in the Dryopteris affinisgroup. Fern Gaz. 18: 1-26.
FRASER-JENKINS,C. R. 2008. http://tech.groups.yahoo.com/group/ukferns/message/863.FREIGANG, J., BUJNOCH, W. & ZENNER, G. 2013. Erstfunde von Dryopteris
×brathaica Fraser-Jenk. & Reichst. (Dryopteridaceae, Pteridophyta) inSüd-Deutschland – morphologische und molekulargenetische Untersuchungen.Kochia 7: 67-86.
ISHIKAWA, H., WATANO, Y., KANO, K., ITO, M. & KURITA, S. 2002.Development of primer sets for PCR amplification of the PgiC gene in ferns. J.Plant. Res. 115: 65-70.
JUSLéN, A.,VäRE, H., WIKSTRöM, N. 2011. Relationships and evolutionary originsof polyploid Dryopteris (Dryopteridaceae) from Europe inferred using nuclear PgiCand plastid trnL-F sequence data. Taxon 60: 1284-1294.
KRAUSE, S. 1998. Dryopteris Adans. (Dryopteridaceae). In: Wisskirch, R. &Haeupler, H. (Eds) Standardliste der Farn- und Blütenpflanzen Deutschlands, Vol.1, pp. 182-183. Ulmer, Stuttgart.
KRAUSE, S., HACHTEL, M. & BENNERT, H. W. 2001. Dryopteris ×brathaicaFraser-Jenk. & Reichst. (Englischer Wurmfarn; Drypoteridaceae, Pteridophyta) inDeutschland. Feddes Repert. 112: 247-260.
MANTON, I. 1932. Contributions to the cytology of apospory in ferns in Britain.Nature 144: 291.
MANTON, I. 1950. Problems of Cytology and Evolution in the Pteridophyta.University Press, Cambridge.
MILDE, J. 1865. Die höheren Sporenpflanzen Deutschland’s und der Schweiz. Felix,Leipzig.
NIKLFELD, H. 1971. Bericht über die Kartierung der Flora Mitteleuropas. Taxon 20:545 – 571.
PERONI, A., PERONI, G., RASBACH, H., RASBACH, K. & REICHSTEIN, T. 1991.Dryopteris remota (A.Braun) Druce in Italien. Farnblätter 23: 1-13.
SCHNELLER, J. J. & HOLDEREGGER, R. 1994. Lack of isozyme variation in theagamosporous fern Dryopteris remota (A.Braun) Druce. Amer. Fern J. 84: 94-98.
SCHNELLER, J. J., HOLDEREGGER, R., GUGERLI, F., EICHENBERGER, K. &LUTZ,E. 1998. Patterns of genetic variation detected by RAPDs suggest a singleorigin with subsequent mutations and long-distance dispersal in the apomictic fernDryopteris remota (Dryopteridaceae). Amer. Fern J. 85: 1038-1042.
SESSA, E. B., ZIMMER, E.A. & GIVNISH, T. J. 2012. Unraveling reticulate evolutionin North American Dryopteris (Dryopteridaceae). BMC Evol. Biol. 12: 104-127.
WALKER, S. 1955. Cytogenetic studies in the Dryopteris spinulosa complex I.Watsonia 3: 193-209.
WIDéN, C.-J., EUW, J. VON, & REICHSTEIN, T. 1970. Trispara-aspidin, ein neuesPhloroglucid aus dem Farn Dryopteris remota (A.Br.) Hayek. Helv. Chim. Acta 53:2176-2188.
88 FERN GAZ. 20(2):79-89. 2015

WIDéN, C.-J., VIDA, G., EUW, J. VON, REICHSTEIN, T. 1971. Die Phloroglucidevon Dryopteris villarii (Bell.) Woynar und anderer Farne der Gattung Dryopterissowie die mögliche Abstammung von D. filix-mas (L.) Schott. Helvetica ChimicaActa 54: 2824-2850.
WIDéN, C.-J., LOUNASMAA, M., JERMY, A.C., EUW, V. J., REICHSTEIN, T. 1976. Die Phloroglucide von zwei Farnhybriden aus England und Schottland, vonauthentischem “Aspidium remotum” A. Braun und von Dryopteris aemula (Aiton)O. Kuntze aus Irland. Helv. Chim. Acta 59: 1725 -1744.
WIDéN, C.-J., FRASER-JENKINS, C. R., REICHSTEIN, T., GIBBY, M. &SARVELA, J. 1996. Phloroglucinol derivatives in Dryopteris sect. Fibrillosae andrelated taxa (Pteridophyta, Dryopteridaceae). Ann. Bot. Fennici 33: 69-100.
BUJNOCH: THE PHYLOGENY OF DRYOPTERIS REMOTA 89

90 FERN GAZ. 20(2). 2015

SHORT NOTE
NOMENCLATURAL NOTE ON HEMIONITIS ARIFOLIA(PTERIDACEAE)
J. MAZUMDARDepartment of Biological Sciences, Burdwan Town School, Burdwan-713101, India
Email: [email protected]
The fern name Hemionitis arifolia (Burm.f.) T.Moore (Pteridaceae) is based onAsplenium arifolium Burm.f., first proposed by Burmann in 1768. Morton (1974: 316)found a sheet in the Geneva Herbarium (G, barcode G00360112, Figure 1) carrying twospecimens and labeled as “Typus”. Alston in August, 1952, had determined thesespecimens as juvenile Acrostichum aureum L., not Asplenium arifolium. As the typebelonged to another taxon Morton (1974) suggested the next available synonymHemionitis cordata Roxb. ex Hook. & Grev. as the correct name for this taxon. Thesheet carries Burmann’s annotation “Arifolium. Asplenium Arifolio Petiv Tab 50 Fig12” indicating that Burmann saw these specimens (Figure 1). But in diagnosis hementioned “simple ovate auriculate sinuous frond” and these specimens showing non-auriculate, non-sinuous frond (Figure 1), and not the basis of description or diagnosisvalidating the name (McNeill et al. 2012, Art. 9.3). Thus these specimens are not Typesand on that basis the name Asplenium arifolium cannot be reduced as synonym ofAcrostichum aureum. Hemionitis arifolia is the correct name for Hemionitis cordata.
In the protologue Burmann mentioned Petiver’s (1702, t. 50, f. 12) and van Rhedetot Drakestein’s (1692-1703, t. 10, Fig. 2zzz) figures showing auriculate sinuous frondsand his diagnosis was based on these figures. Although Morton (1974) used the term“Type” for the specimen in G (Figure 1), he did not accept it, as is clear from hiscomment “If this really is the type…,”; it is not acceptable as lectotype (McNeill et al.2012, Art. 7.10). Here I select van Rhede tot Drakestein’s figure from Hortus IndicusMalabaricus (1703) as Lectotype. But, as no collection data is available for thespecimen on which this drawing was based (Johnston 1970; Wagenitz 1978; McNeill etal. 2012, Rec. 8A.1 & 8A.2) an epitype is selected.
TYPIFICATIONAsplenium arifolium Burm. f., Fl. Ind. 231. 1768 — Hemionitis arifolia (Burm. f.) T.Moore, Index Fil. 114. 1859. — Parahemionitis arifolia (Burm. f.) Panigrahi, Amer.Fern J. 83(3): 90. 1993.Lectotype (designated here): van Rhede tot Drakestein, Hortus Indicus Malabaricus 12:21, t. 10, 1703 (Figure 2).Epitype (designated here): India. West Bengal: Burdwan, Kanchannagar, near bridgeabove canal, 30 m, 21 Nov 2007, J. Mazumdar 59 (CAL Figure 3).
ACKNOWLEDGEMENTSI sincerely thank Dr. I. Valette, Secrétaire Herbiers and Conservatoire et jardinbotaniques de la Ville de Genève, Switzerland (G) for providing permission forpublication of image of G specimens; and Dr. A.R. Smith, University of California,Berkeley, U.S.A. and Mr. N.J. Turland, Botanischer Garten und Botanisches MuseumBerlin, Germany for useful discussions.
FERN GAZ. 20(2):91-94. 2015 91

REFERENCESBURMANN, N.L. 1768. Flora Indica etc. Apud Cornelium Haek, Amsterdam.
(http://www.biodiversitylibrary.org/item/123275).JOHNSTON, M.C. 1970. Still No Herbarium Records for Hortus Malabaricus. Taxon
19(4):655.
92 FERN GAZ. 20(2):91-94. 2015
Figure 1. Type sheet of Asplenium arifolium Burm.f. from G (barcode G00360112).Image © Conservatoire et jardin botaniques de la Ville de Genève, Switzerland,reproduced with permission.

MCNEILL, J., BARRIE, F.R., BUCK, W.R., DEMOULIN, V., GREUTER, W.,HAWKSWORTH, D.L., HERENDEEN, P.S., KNAPP, S., MARHOLD, K.,PRADO, J., PRUD’HOMME VAN REINE, W.F., SMITH, G.F., WIERSEMA, J.H.& TURLAND, N.J. (eds) 2012. International Code of Nomenclature For algae,fungi, and plants (Melbourne Code) adopted by the Eighteenth InternationalBotanical Congress Melbourne, Australia, July 2011 . (http://www.iapt-taxon.org/nomen/main.php?page=title).
MORTON, C.V. 1974. William Roxburgh's fern types. Contr. U. S. Natl. Herb. 38(7):283-396. (http://www.biodiversitylibrary.org/item/13798).
PETIVER, J. 1702. Catalogus Classicus & Topicus Omnium Rerum Figuratum in V.Decadibus, Seu primo [-secundo] volumine Gazophylacii naturae & artis; Singulisad proprias Tabulas & Numeros relatis. Ex Officinâ Christ. Bateman, vico vulgòdicto Pater-noster-row, London. (http://bibdigital.rjb.csic.es/ing/Libro.php?Libro=4831).
VAN RHEDE TOT DRAKESTEIN, H.1692-1703. Hortus Indicus Malabaricus etc.Vols. 11 & 12. Sumptibus Johannis van Someren, et Joannis van Dyck, Amsterdam.(http://www.biodiversitylibrary.org/item/14380).
WAGENITZ, G. 1978. The “Plantae Malabaricae” of the Herbarium at GöttingenCollected near Tranquebar. Taxon 27(5/6): 493-494.
MAZUMDAR: NOMENCLATURAL NOTE ON HEMIONITIS ARIFOLIA 93
Figure 2. Lectotype of Asplenium arifolium Burm.f.: Tab. 10 from van Rhede totDrakestein’s Hortus Indicus Malabaricus.

94 FERN GAZ. 20(2):91-94. 2015
Figure 3. Epitype of Asplenium arifolium Burm.f. (J. Mazumdar 59) (CAL).

SHORT NOTE
CONFIRMATION OF LOXOGRAMME LANKOKIENSIS(POLYPODIACEAE) IN INDIA
J. MAZUMDARDepartment of Biological Sciences, Burdwan Town School, Burdwan-713101, India
Email: [email protected]
The small epiphytic fern Loxogramme lankokiensis (Rosenst.) C.Chr. was first reportedto occur in India from the Tirap District, Arunachal Pradesh, by Singh & Panigrahi(1987). Later Singh & Panigrahi (2005) suspected that an earlier record (Bhargavan &Joseph 1978) of L. grammitoides (Baker) C.Chr., from Arunachal Pradesh, LohitDistrict, may also represent L. lankokiensis and not L. grammitoides (Baker) C.Chr.However, they were unable to locate the specimen at CAL and could not make a definitedecision (Singh & Panigrahi 2005). During the taxonomic study of ferns of EasternIndia, I found this specimen of Loxogramme lankokiensis: INDIA. Arunachal Pradesh:Lohit District (N.E.F.A.), forest around Hayuliang along Dalai river, J. Joseph 48934(originally from CAL, now at BURD; Figures 1A–C).
In recent revisions Fraser-Jenkins (2008, 2012) followed the opinion of Dr. M.G.Price (University of Michigan, Michigan) and claimed that the report of L. lankokiensisfrom India was based on the misidentification of small plants of L. grammitoides. Closeexamination of J. Joseph 48934 showed features that matched with characters of L.lankokiensisas described in literature (Singh & Panigrahi 2005, Zhang et al. 2013).Loxogramme lankokiensis has dimorphic fronds (Figure 1C) and rhizome scales withentire margins whereas L. grammitoides has mostly monomorphic fronds and rhizomescales with slightly dentate margins, according to Zhang et al. 2013. This specimen (J.Joseph 48934) confirms the earlier report of L. lankokiensis in India, Lohit District,Arunachal Pradesh by Singh & Panigrahi (2005), in addition to China, Thailand andVietnam.
Selected digitized specimens examined— Loxogramme lankokiensis: VIETNAM.Lankok [“Roches moussues du Mont Bavi, prèt de la vallée de Lankok”], 12 Jun. 1888,B. Balansa 1934 (Isotypes B-20 0089150, BR-0000006979492, BM-001039924, BM-001039925, P-01356591); Tonkin: Cao-Bang, D. Billet s.n., Herb. H. Christ (P-01356590). THAILAND. Udawn: Phu (Mt.) Luang, about 900 m, 11 Jan. 1966, E.Hennipman 3631A (P-01353469).— Loxogramme grammitoides: CHINA. S. Wushan,West Hupeh, E.H. Wilson 620 (Isotypes B-20 0089149, US00067489, BM-001039922).
ACKNOWLEDGEMENTSI sincerely thank the Director, Botanical Survey of India and Prof. R. Mukhopadhyay,Pteridology Section, Department of Botany, The University of Burdwan for kindlyproviding opportunity to study the specimen and Dr M. Lehnert, University Bonn,Germany for constructive suggestions.
REFERENCESBHARGAVAN, P. & JOSEPH, J. 1978. Loxogramme grammitoides (Bak.) C.Chr.- a
fern record for India. New Botanist 5: 43-44.
FERN GAZ. 20(2):95-97. 2015 95

FRASER-JENKINS, C.R. 2008. Taxonomic revision of three hundred Indiansubcontinental Pteridophytes with a revised Census- List. Bishen Singh MahendraPal Singh, Dehra Dun, 685 pp.
FRASER-JENKINS, C.R. 2012. Rare and Threatened Pteridophytes of Asia 2.Endangered Species of India—the Higher IUCN Categories. Bull. Natl. Mus. Nat.Sci., Tokyo, B. 38: 153-181.
SINGH, S. & PANIGRAHI, G. 1987. Notes on two species of Loxogramme Presl(Pteridophyta) in Eastern Himalayas. Bull. Bot. Surv. India. 27 (1-4): 252-254.
96 FERN GAZ. 20(2):95-97. 2015
Figure 1. A.–C. Loxogramme lankokiensis (J. Joseph 48934 CAL): A. & B. Specimenfrom Arunachal Pradesh, India, C. Part of plant showing distinct fertile and sterilefronds.

SINGH, S. & PANIGRAHI, G. 2005. Ferns and Fern-allies of Arunachal Pradesh(Adiantaceae to Nephrolepidaceae). Vol. 1. Bishen Singh Mahendra Pal Singh,Dehra Dun, 426 pp.
ZHANG, X.C., LU, S.G., LIN, Y.X., QI, X.P., MOORE, S., XING, F.W., WANG, F.G.,HOVENKAMP, P.H., GILBERT, M.G., NOOTEBOOM, H.P., PARRIS, B.S.,HAUFLER, C., KATO, M. & SMITH, A.R. 2013. Polypodiaceae. 758-850 pp., inWU, Z.Y., RAVEN, P.H. & HONG, D.Y. (eds), Flora of China, Vol. 2-3 (Pteridophytes).(http://flora.huh.harvard.edu/china/mss/volume02/Flora_of_China_Volume_2_3_Polypodiaceae.pdf)
MAZUMDAR: LOXOGRAMME LANKOKIENSIS IN INDIA 97

SHORT NOTE
THREE NEW COMBINATIONS IN CTENOPTERELLA(POLYPODIACEAE)
Three new combinations are proposed for Ctenopterella (Polypodiaceae) in preparationfor a monograph of the genus.
Ctenopterella lasiostipes (Mett.) Parris comb. nov.Based on Polypodium lasiostipes Mett., Ann. Sci. Nat. 4, 15: 76-77 (1861). Type: NewCaledonia, Poila, Vieillard 1601 (lectotype P, chosen by Brownlie, Fl. Nouv.-Caléd. 3:274 (1969); isolectotype NSW!). Synonym: Ctenopteris lasiostipes (Mett.) Brownlie,Fl. Nouv.-Caléd. 3, Pteridophytes: 273 (1969).
Ctenopterella thwaitesii (Beddome) Parris comb. nov.Based on Polypodium thwaitesii Beddome, Ferns Brit. India: t. 188 (1866). Type:Ceylon, Thwaites C. P. 3900 (lectotype K!, chosen by Parris, Fern Gaz. 16(5): 243(2002); isolectotypes BM!, CAL 20454 images seen, CGE!, E 00195204!, GH!, K!, P02141982!, PDA!). Synonym: Ctenopteris thwaitesii (Beddome) Sledge, Bull. B.M.(Nat. Hist.) Bot. 2(5): 155 (1960).
Ctenopterella undosa (Baker) Parris comb. nov. Based on Polypodium undosum Baker, J. Bot. 28: 108 (1890). Type: New Guinea, MtMusgrave, 1889, MacGregor 61 (holotype K 000507064!; isotypes MEL!). Synonyms:Ctenopteris undosa (Baker) Copel., Philipp. J. Sci. 81(2): 103 (1953); Polypodiumpycnophyllum C.Chr., Brittonia 2(4): 308 (1937). Type: Papua, Central Division, MtAlbert Edward, 3680 m alt., 5 July 1933, Brass 4421 (holotype BM; isotypes BRI!,MICH 1115954!, NY) (synon. nov.), Ctenopteris pycnophylla (C.Chr.) Copel., Philipp.J. Sci. 81(2): 102 (1953).
B. S. ParrisFern Research Foundation, Kerikeri, New Zealand.
Email: [email protected]
98 FERN GAZ. 20(2). 2015

BOOK REVIEW
Farne als Lebensleidenschaft – Festschrift für H. Wilfried Bennert anlässlichseines 65.Geburtstages. Berichte des Instituts für Landschafts- undPflanzenökologie der Universität Hohenheim. Beiheft 22. 2013. 238pp. 21.0 x29.7cm (in German).Available from A. Kleinsteuber, Weiβdornweg 35, D-76149, Karlsruhe, Germany.email: [email protected]
This festschrift for Wilfried Bennert on the occasion of his 65th birthday is a collectionof 11 papers gathered together by Karsten Horn and illustrated with a series of etchingsof ferns by Reinhard Böcker. The subjects of the articles reflect Bennert’s wide interestsin pteridology and ecology. The volume starts with a description of his life and careerin science, the Bochum days, and how he fell under the spell of ferns, horsetails andclubmosses. He carried out fieldwork not only in Europe but also in California, Iceland,Southeast Asia and Russia, and Horn describes the work that culminated in one of hismajor fern projects, the publication of Die seltenen und gefährdeten FarnpflanzenDeutschlands in 1999. This is followed by a full list of his publications.
The subjects and authors of the other papers are summarised below.Reinhard Bornkamm describes the ferns, horsetails, Ephedra and gymnosperms ofEgypt and their role in plant communities. Herbert Sukopp looks at three ferns in C19thhorticulture around Berlin. Andreas Sarazin et al. describe new records of neophyte ferntaxa that have been discovered in Northwest Germany, including species of Adiantum,Cyrtomium and Pteris. Böcker describes the historical and present distributions offerns, lycophytes and horsetails from the Stuttgart area, and assesses the impact ofhuman activity. Marcus Lubienski provides photographs, detailed descriptions anddistributional data for all known hybrids of Equisetum in Europe, and theirmorphological variability is discussed. Caroline Van den heede and Ronnie Viane reportthe discovery of Asplenium lolegnamense and A.×chasmophilum and discuss theirorigins. Wolfgang Jäger reports the occurrence of ascomycete fungi on ferns in theBergisches Land, Northrhine-Westphalia. Karsten Horn has published an account of thelife and scientific output of K.A.J. Milde, a great pteridologist of C19th, withillustrations from some of his significant monographs. Henning Haeupler reviews andclarifies the work on life forms to describe vegetation, first introduced by Humboldt in1806 and extended by Raunkiaer in 1907. The volume concludes with plantcolonisation in shallow waters - plants of the littoral zone, by Klaus Kaplan. Wilfried Bennert has been a member of BPS since 1975, and is active in the Group ofEuropean Pteridologists.
M. Gibby
FERN GAZ. 20(2). 2015 99

INSTRUCTIONS FOR AUTHORS
PAPERS should not usually exceed 20 printed pages and are generally expected to beconsiderably shorter. As well as reports of original research, review articles and shortnotes, e.g. new records, are encouraged. The senior author should supply an email addressto facilitate correspondence. REVIEWS should cover recent developments in any aspect of research on ferns andlycopods. This can include unpublished research by the author, and/or the possibledirection of future research. They should be aimed at the general reader who may not bean expert in the chosen topic. Proposals for a review should be discussed first with thereview editors.MANUSCRIPTS should be submitted in English (British) in electronic format in10-point Times New Roman font and double spaced. Electronic versions of text andtables should be compatible with WORD, with figures as pdf or jpg files, and sent asemail attachments or CDroms. All manuscripts will be refereed.THE TITLE should reflect the content of the paper and be in BOLD CAPITALS(11-point) and centrally aligned. Generic and specific names should be in italics and anytitle containing a generic or specific name should be followed by the family name inbrackets e.g.
TRICHOMANES SPECIOSUM (HYMENOPHYLLACEAE)IN SOUTHERN SPAIN
AUTHOR ABBREVIATIONS should follow Pichi Sermolli’s (1996) Authors ofscientific names in Pteridophyta, Royal Botanic Gardens, Kew.MAIN HEADINGS: should be in BOLD CAPITALS (10-point) and centrally aligned.SUBSIDIARY HEADINGS: should be in bold, the first letter of each word in capitals,the rest in lower case and left-aligned.AUTHORS’ NAMES AND FULL ADDRESSES (including email address): followthe title and are centrally aligned.KEY WORDS: up to ten.ABSTRACT: should reflect the content of the paper.FIGURES: there is no distinction between photographs and line drawings in numbering.All should be presented in a form ready for reproduction, colour, TIF format and 300dpi (please contact editor with queries), with a scale bar where appropriate. Lettering ornumbers (Arabic) should be in the bottom left using uppercase Times Roman. Figurecaptions should be on a separate sheet.TABLES: can be printed in either portrait or landscape format. Authors should considerthis when preparing tables. Authors should ensure that tables fit the printed page size ina legible form.PAGESIZE: The Gazette has a useable page size of 123 × 204 mm. This can be used inportrait or landscape mode but any supplied artwork should not be larger than this.MEASUREMENTS: follow the metric system.CHECKLISTS: follow the format of Baksh-Comeau, Fern Gaz. 16(1, 2): 11- 122.REFERENCES: follow the style of a recent issue of The Fern Gazette, e.g.:-HOOKER, W.J. 1864. Species Filicum, 5. Dulau & Co., London.MORTON, C.V. 1947. The American species of Hymenophyllum, section
Sphaeroconium. Contr. U.S. Natl. Herb. 29(3): 139-201.STEVENSON, D.W. & LOCONTE, H. 1996. Ordinal and familial relationships of
pteridophyte genera. In: CAMUS, J.M., GIBBY, M. & JOHNS, R.J. (Eds)Pteridology in perspective, pp. 435-467. Royal Botanic Gardens, Kew.
JOURNAL ABBREVIATIONS: should follow Botanico Periodicum Huntianum &Supplements.Alterations from the original text at proof stage will be charged for unless they are minorpoints of detail. A pdf will be provided free to the senior author.
100 FERN GAZ. 20(2). 2015

REVIEW
MYCORRHIZAL RELATIONSHIPS IN LYCOPHYTES AND FERNS
MARCUS LEHNERT1,2* & MICHAEL KESSLER3
1 Nees-Institut für Biodiversität der Pflanzen, Rheinische FriedrichWilhelms-Universität Bonn, Meckenheimer Allee 170, D-53115 Bonn, Germany.
2 Staatliches Museum für Naturkunde, Am Löwentor, Rosenstein 1, D-70191 Stuttgart,Germany
3 Systematic and Evolutionary Botany, University of Zurich, Zollikerstrasse 107,CH-8008 Zurich, Switzerland.
* Author for correspondence, email: [email protected]
ABSTRACTMycorrhizae, i.e., symbiotic associations between fungi and plant roots, occur inabout 80% of land plant species and have been shown to benefit both the fungus(carbon uptake) and plant (nutrient and water uptake, protection againstpathogenic fungi). We here provide a brief overview of the state of knowledge ofmycorrhization in ferns. Only about 62% of species studied to date havemycorrhizae, with arbuscular mycorrhizal fungi (mainly Glomeromycta) beingthe dominant partners, while other associations are made with ascomycetes andthe so-called Dark Septate Endophytes. There is no clear phylogenetic signal inmycorrhization among ferns, with both basal (e.g., Anemiaceae, Gleicheniaceae,Cyatheaceae) and derived families (e.g., non-grammitid Polypodiaceae,Tectariaceae, Aspleniaceae) having <50% of species mycorrhizal. Ecologically,epiphytes have lower degrees of mycorrhization than terrestrial species, withaquatic taxa almost completely lacking mycorrhizae. This probably reflects therequirements of the fungi. There are no experimental studies on the benefits ofmycorrhizae for ferns, but field experiments suggest that there is a fine balancebetween positive and negative effects, so that many fern species have disposedof the fungi either generally or facultatively. Much remains to be learnt aboutmycorrhization in ferns, especially in an evolutionary context in comparison withbryophytes and seed plants.
INTRODUCTIONThe term mycorrhiza (pl. mycorrhizae) was introduced by Frank (1885) for structures inangiosperm roots caused by fungi and translates as “fungus root”. Since then it has beenshown that it is one of the most common and important symbioses in the land plants(Brundrett, 2002; 2004; 2009), even in plants like bryophytes and Psilotaceae that lackroots. It is estimated that ca. 80% of all land plants engage either in facultative or obligatemycorrhizal associations (Trappe, 1987; Wang & Qiu, 2006; Brundrett, 2009).
It is likely that the colonization of land by early land plants was facilitated bymycorrhizae (Pyrozinski & Malloch, 1975; Remy et al., 1994; Blackwell, 2000; Selosseet al. 2015), even before roots first arose (Taylor et al., 1995; Brundrett, 2002; Kenrick& Strullu-Derrien, 2014). This assumption is based on the presence of structures verysimilar to extant symbiotic Glomeromycota in the earliest tracheophyte fossils from theRhynie Chert (Remy et al., 1994; Strullu-Derrien & Strullu, 2007; Selosse et al., 2015)
FERN GAZ. 20(3):101-116. 2016 101

and supported by the conserved nature of the genes regulating mycorrhizal associationsacross all land plant lineages (Wang et al., 2010).
Mycorrhizal fungi are known to provide numerous benefits to their plant hosts(Selosse & Rousset, 2011). They enhance nutrient uptake, especially of phosphorous(Siddiqui & Pichtel, 2008), increase drought tolerance by facilitating water uptake (Faberet al., 1991; Warren et al., 2008), and compete with parasitic fungi thus protecting plantsfrom parasitic fungal infections (Newsham et al., 1995; Brundrett, 2002). In return, thefungal partners receive carbohydrates from the host plant (Selosse & Rousset, 2011).The intensity of this symbiosis may vary with external ecological factors (Allen et al.,2003; Antunes et al., 2011) and apparently can be regulated more actively by the planthost than by the fungal symbiont (Barker et al., 1998; Jones et al., 2004).
Most research on mycorrhizae has been conducted on seed plants because of theirdominance, diversity and economic importance (Brundrett, 2004; 2009). In recent years,the scientific focus has shifted more to the origin of this symbiosis and hence to itspresence in old land plant lineages like extant bryophytes and lycophytes (Read et al.,2000; Rimington et al., 2015). Mycorrhizae in ferns have been regularly screened forbut rarely been in the focus of physiological or synthetic studies combining ecology andphylogeny (Wang & Qiu, 2006; Brundrett, 2009).
In the present paper, we provide a brief overview of the still incomplete knowledgeof mycorrhization in lycophytes and ferns (for simplicity called ferns in the following).
TYPES OF MYCORRHIZAEFour major types of mycorrhiza have been distinguished (Barker et al., 1998; Smith &Read, 2008; Siddiqui & Pichtel, 2008) with some characteristic deviations andaberrations in some major clades of host plants (Figure 1; Brundrett, 2002; 2004).
Arbuscular mycorrhiza (AM) is an endosymbiosis, in which the fungi penetrate thecell walls of the root cortex and form characteristic organs in the living cells for nutrientexchange, the so-called arbuscules and vesicles (Figure 2a,b; Bonfante & Genre, 2008).It is the most common, and probably the ancestral form of mycorrhizae (Field et al.,2015a), occurring in ca. 80% of extant land plants (Wang & Qiu, 2006; Brundrett, 2009).The fungal partners are aseptate Eumycota, in the majority Glomeromycota (Schüßleret al., 2001), which are asexual and obligate symbionts (Brundrett, 2004; 2009). In somebasal lineages of landplants (hornworts, some liverworts and lycophytes, a few ferns),Mucoromycotina, which are either basal to or sister to Glomeromycota, may also forma similar endomycorrhiza but without the presence of arbuscules (Desirò et al., 2013;Strullu-Derrién et al., 2014; Field et al., 2015a, b).
Ectomycorrhizae (ECM) are mainly formed by the more derived septate Ascomycotaand Basidiomycota (Taylor & Alexander, 2005; Taylor et al., 2010) but in a few casesalso endogonalean fungi (Mucoromycotina, Hibbet et al., 2007). They build an internalnetwork of hyphae that surround the intact root cells (Hartig net) and an external hyphalsheath around the root tips (Smith & Read, 2008; Brundrett, 2004). ECM occur in2.0–4.7% of seed plants but dominate among tree species in temperate and boreal zonesfamilies (Smith & Read, 2008; Brundrett, 2009). Until now, ECM have not beenconfirmed for ferns. It has been suggested that ferns shift from AM to ECM in forestswhere ectomycorrhizal tree species are dominant (Cooper, 1976; Iqbal et al., 1981), butthe claim that the investigated roots really belonged to ferns has been strongly contested(Smith & Read, 2008; Brundrett, 2002). Fungal symbioses interpreted as derivations ofECM are the Arbutoid mycorrhiza and the Monotropid mycorrhiza (Brundrett, 2004),
102 FERN GAZ. 20(3):101-116. 2016
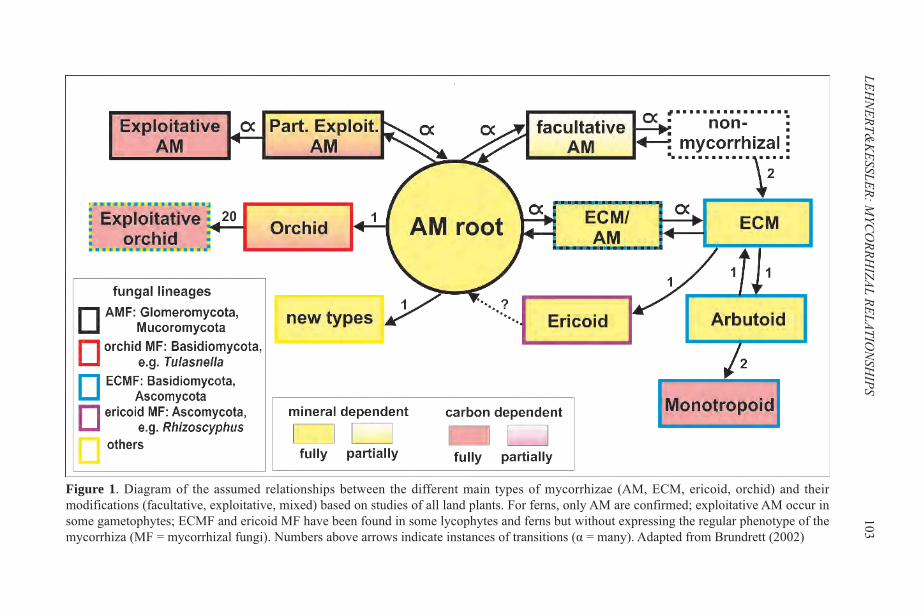
LEHN
ERT&K
ESSLER:MYC
ORRH
IZAL RELATION
SHIPS
103
Figure 1. Diagram of the assumed relationships between the different main types of mycorrhizae (AM, ECM, ericoid, orchid) and theirmodifications (facultative, exploitative, mixed) based on studies of all land plants. For ferns, only AM are confirmed; exploitative AM occur insome gametophytes; ECMF and ericoid MF have been found in some lycophytes and ferns but without expressing the regular phenotype of themycorrhiza (MF = mycorrhizal fungi). Numbers above arrows indicate instances of transitions (α = many). Adapted from Brundrett (2002)

but these are also not reported for ferns. Ericoid mycorrhizae are formed with septate Ascomycota, which all are primarily
decomposers and saprophytes (Hibbet et al., 2007). They are uncommon but typical ofEricaceae (Cullings, 1996; Brundrett, 2004) and some other plant groups occurring onnutrient-deficient substrates characterized by low primary production (Cairney &Meharg, 2003). Typical of this symbiosis is that the hyphae enter through root hairs andform hyphal coils in cortical cells (Read, 1996). ascomyceta infections of the Ericoidtype have been reported for tropical ferns, mainly epiphytes, such as Elaphoglossum,grammitid ferns, Hymenophyllum and Trichomanes (Schmid et al., 1995; Lehnert et al.,2009). Records of the so-called Dark Septate Endophytes (DSE) in ferns (Dhillion, 1993;Jumpponen & Trappe, 1998; Fernández et al., 2008) may include still unrecognizedEricoid mycorrhizae since the taxonomic identity of most of these fungi is unknown(Jumpponen, 2001; Rains et al., 2003). Like the Ericoid mycorrhizal Ascomycota, DSEare more common in epiphytic fern species (Kessler et al., 2010).
Orchid mycorrhizae involve a small group of Basidiomycota (Brundrett, 2002), andare characterized by the initial carbon dependency of the host plant seedling on thesaprophytic fungal partner; several orchid lineages have independently developed anexploitative life-style from this condition (Brundrett, 2002; 2004).
Such exploitative mycorrhizae are not known from the sporophytes of lycophytesand ferns, but gametophytes of Lycopodiaceae (Duckett & Ligrone, 1992; Winther &Friedmann, 2008), Ophioglossaceae (Schmid & Oberwinkler, 1994; Field et al., 2015c)and Psilotaceae (Duckett & Ligrone, 2005; Winther & Friedmann, 2009) are well knownfor being mycoheterotropic. These mostly involve AMF but recently Horn et al. (2013)reported the steady prevalence of basidiomycetes of the order Sebacinales (as found inorchid mycorrhiza and ECM) in gametophytes of Diphasiastrum alpinum (L.) Holub.This had been anticipated because basidiomycetes had already been documented asmycorrhizal endophytes for bryophytes (Pressel et al., 2010)
Mycorrhizae are species-specific regarding their phenotype (i.e., one fungus and oneplant always engage in a specific type of interaction) but not regarding the taxa involved:Ferns can host a mixture of up to five different mycorrhizal fungi at the same time butwith one usually dominating (Jansa et al., 2008), and one fungus can colonize different
104 FERN GAZ. 20(3):101-116. 2016
Figure 2a. Internal structures of arbuscular mycorrhiza (vesicles and arbuscules,lower ones partially released from cell due to preparation) in the root cortex of thetree fern Cyathea tortuosa R.C.Moran from Ecuador (Prov. Napo, 500 m) (20 x).Figure 2b. The roots of a typical non-mycorrhizal plant with pronounced root hairformation, represented by Elaphoglossum castaneum (Baker) Diels from Ecuador(Prov. Napo, 2000 m). (10 x) Photos Ramona Gudel.

plant species (Gemma et al., 1992; van der Heijden et al., 2015). The true symbiotic nature of a fungal association can only be confirmed by
physiological experiments aiming at the flow of nutrients (e.g., Jurkiewicz et al., 2010).As this has not been done for any fern yet, it is uncertain whether most observedassociations are really symbiotic or functionally neutral (Brundrett, 2004; 2009). AsGlomeromycota are fully dependent on the symbiosis and only known to occurindependently from a host plant when they are dormant spores (Brundrett, 2009), it seemslikely that a symbiotic relationship is involved in most cases but not necessarily with theroots in which they are found. Under high inoculum pressure in the substrate, typicalnon-mycorrhizal taxa may harbor AMF as neutral endophytes while the same myceliumprobably forms a true mycorrhiza with a typical host plant nearby (Lekberg et al., 2015).Similarly, some typical non-mycorrhizal taxa have been found to be able to host AMFunder nutrient deficiency (Brundrett, 2009). Furthermore, a low presence of fungi in theroots must not necessarily imply a low physiological activity between both partners. Thecategorization into obligate and facultative mycorrhizal plant species can only beevaluated in controlled experiments, e.g., by establishing sterile cultures of both partnersand later re-inoculation (Pearson & Read, 1973), and not by visual estimation alone.
TAXONOMIC DISTRIBUTION OF MYCORRHIZAEThe mycorrhizal status is regarded to be ancestral among ferns (Zhao, 2000; Wang &Qiu, 2006), as is likely for all land plants (Pirozynski & Malloch, 1975; Strullu-Derriénet al., 2014; Field et al., 2015a).
The scant reviews of fern mycorrhizae have focused on the sporophyte and havecome up with highly diverging results regarding the general presence of fungal symbiontsas well as the distribution among the different groups. Newman and Reddell (1987) found87% to be always mycorrhizal, 7% facultative, and 6% not mycorrhizal (based on 180fern species), which is within the range of the values found for angiosperms (Brundrett,2009). Others reported 75% of the studied fern species to have AM (Gemma et al., 1992).Zhao (2000) found only 33% of 256 species from China to be mycorrhizal (17% obligate+ 16% facultative), and similarly Moteetee et al. (1996) reported 36% of the species tobe mycorrhizal. Cooper (1976) found 100% of the investigated ferns to harbourmycorrhiza, but the sampling included only terrestrials and low epiphytes from humidsubtropical to southern temperate forest, i.e., conditions that foster fungal growth. Onthe other extreme, Lesica & Antibus (1990) focused on epiphytes and found nomycorrhiza in the 12 investigated species. Wang & Qiu (2006) found 52% of all speciesand 92% of all families to harbour AMF. Brundrett (2009) summarized only more recentreports on mycorrhizae that he could trust, but contrary to his detailed summary onspermatophytes, he could not give an exact evaluation for ferns.
Lehnert (2007) used all available data on fern mycorrhiza worldwide starting fromBoullard (1958, 1979), and evaluated simple presence/absence of mycorrhizalcolonisations in three classes (aseptate fungi = AMF; septate fungi = DSE/ascomycetes;unspecified mycorrhiza) in 675 taxa (ca. 6% of the global diversity; Smith et al., 2006).The observed average of 62% for fungal colonisation lies in the middle of the range ofprevious reports for ferns but is lower than the values for seed plants (Brundrett, 2009).AMF are dominant among the fungal symbionts with 66%, whereas the remaining 34%are made up of ascomycetes, DSE and unspecified mycorrhizations. Across thephylogeny at the family level (sensu Smith et al., 2006), below average mycorrhization(≤62%) was found among several families of derived Polypods (non-grammitid
LEHNERT&KESSLER: MYCORRHIZAL RELATIONSHIPS 105

Polypodiaceae 23%, Tectariaceae 25%, Aspleniaceae 45%) but other families of thePolypods such as Davalliaceae and Onocleaceae had 100% mycorrhization. Vice versa,among the other leptosporangiate ferns, several families had below averagemycorrhization rates (Anemiaceae 50%, Gleicheniaceae 48%, Cyatheaceae 53%).Among the eusporangiate lineages, only the Equisetaceae showed low mycorrhization(22%) whereas the remainder had average to high values (Psilotacae 67%, Marattiaceae91%). Families largely restricted to aquatic habitats showed low (Isoetaceae; one of twosamples) or no (Marsileaceae, Salviniaceae) fungal colonization. Mycorrhizal epiphyteswere found mainly among grammitid ferns (84% of epiphytes and 74% of all speciesmycorrhizal), Hymenophyllaceae (76% of epiphytes and 76% of all species mycorrhizal)and Dryopteridaceae (63% of epiphytes [mainly Elaphoglossum] and 62% of all speciesmycorrhizal). Interestingly, only 42% of the investigated sporophytes of Lycopodiaceaehad mycorrhizae despite their predominantly mycoheterotrophic gametophytes (Winther& Friedmann, 2008).
Later substantial additions to the survey (Kessler et al., 2009; 2010b; 2014a; Zubeket al., 2010; Ogura Tsujita et al., 2013; Muthukumar & Prabha, 2012; 2013; Lara-Pérezet al., 2015) have increased the species count but have not significantly changed theoriginally found total average and ratios between the substrates (Lehnert et al.,unpublished data).
A caveat to bear in mind when comparing many different reports is the often unevenapproaches regarding evaluation and quantification of the presence of fungi, as has beenpointed out for AMF by Brundrett (2009). However, considering that it is more likelythat fungi present have been over-interpreted as symbionts rather than that true,functioning mycorrhizas have been overlooked, these numbers are more likely to dropthan to rise after a thorough re-evaluation. Thus, the observed pattern that lycophytesand ferns have a below-average colonization rate compared with seed plants or landplants in general is expected to hold true.
A similar but even stronger uncertainty lies in the data supporting the mutualisticnature of these colonizations, which would allow them to be addressed undisputedly asmycorrhizae (Brundrett, 2002; 2009). Most reports used only light microscopicalpreparation techniques to investigate the presence of fungi, which render the sampleunsuitable for futher ultrastructural and molecular studies (pers. obs. for ferns, but seeIshii & Loynachan, 2004). Even if obligately mycorrrhizal fungi abound in the roots,this does not necessarily mean that the partners interact physiologically (Lekberg et al.,2015). Electron microscopy may allow the larger classification of the fungus and couldreveal whether the host cell responded more in a symbiotic way or if it just tried to rejectand contain the penetrating fungus (e.g. Schmid & Oberwinkler, 1994; Schmid et al.,1995). So far such ultrastructural and/or genetic investigations of mycorrhizae in thesporophyte generation covered only 12 species of lycophytes (Duckett & Ligrone, 1992;Winther & Friedmann, 2007a; Rimington et al., 2015) and 35 species of ferns (Schmidet al., 1995; Schmid & Oberwinkler, 1996; Pazmiño, 2006; Winther & Friedmann, 2007b;2009; Lehnert et al., 2009; Field et al., 2015c; Rimington et al., 2015), which representsonly a small fraction (3.7 %) of the 1,287 species of lycophytes and ferns for whichreports are available (ca. 11 % of the global diversity; Lehnert et al., unpubl. data)
LIFE-STAGE DISTRIBUTION OF MYCORRHIZAEMycorrhizal interactions with fern gametophytes are not as uniform as often stated(Mehltreter, 2010). With a few exceptions (Duckett & Ligrone, 1992), gametophytes of
106 FERN GAZ. 20(3):101-116. 2016

Lycopodiaceae, Psilotaceae (Duckett & Ligrone, 2005) and Ophioglossaceae areachlorophyllous and mycoheterotrophic, meaning not only do they receive water andminerals but also carbohydrates from the fungi (Schmid & Oberwinkler, 1994). By thismeans, the gametophytes may tap into the carbon source of another, photoautotrophichost of the fungus (Harley & Harley, 1987; Horn et al., 2013), possibly even the parentalsporophyte (Winther & Friedmann, 2008). The anatomy of these mycoheterotrophicgametophytes, which are tuberous and relatively thick textured (Winther & Friedmann,2007; 2008; 2009), allows an easy colonisation by fungi, whereas across fern lineageswith photoautotrophic gametophytes, we see a gradual reduction of size (especiallythickness) and longevity along the phyletic sequence (Smith et al., 2006). Mycorrhizationrates seem to follow this pattern.
The eusporangiate Marrattiaceae (Bower, 1923; Ogura-Tsuijita et al., 2013) and thebasal leptosporangiate Osmundaceae, Plagiogyriaceae, Gleicheniaceae and Cyatheaceae(Ogura-Tsuijita et al., 2015) show relatively constant and high presence of AM fungi(58–97%) in the thick centre of the gametophytes. However, most leptosporangiatefamilies have the common heart-shaped Polypodium-prothallium and offer only restrictedspace for fungal symbionts; accordingly fungal colonization is observed infrequently(e.g., Reyes-Jaramillo et al., 2005; Turnau et al., 2005; Muthukumar & Prabha, 2012)and most taxa are considered to be only facultatively mycorrhizal (Read et al., 2000). Inleptosporangiate lineages with strongly dissected to filiform prothallia (e.g., someHymenophyllaceae, grammitid Polypodiaceae), AM are not observed (Boullard, 1979).
Even if both generations in the life cycle are mycorrhizal, the sporophyte does notdirectly inherit the fungal partner from the gametophyte but has to acquire it anew fromthe substrate (Schmid & Oberwinkler, 1995), so that each generation can have a differentassortment of fungal partners, although usually with a strong overlap (Winther &Friedmann, 2008).
HABITAT DISTRIBUTION OF MYCORRHIZAEAmong ferns, most studies have only considered selected species; whole ferncommunities have been studied only in Canada (Berch & Kendrick, 1982), China (Zhao,2000), La Réunion (Kessler et al., 2010), and Ecuador (Kessler et al., 2014a). In general,the mycorrhization rate of soil-growing fern species is typically higher than that ofepiphytic species (Gemma et al., 1992; Kessler et al., 2010). Thus, Lehnert (2007)reported putative mycorrhizal fungi mainly in terrestrial and saxicolous species (67%and 59%, repectively) but only in 53% of epiphytic species and in 9% of aquatic species.The percentage in epiphytes included a high presence of ascomycetes + DSE (67% ofcolonisations), whereas the remaining 33% of epiphytic mycorrhiza represented only8% of the total of AMF colonisations in all lycophytes and ferns.
The low rate of mycorrhizal infections among epiphytes may be caused by either thefungi or the ferns. Glomeromycetes, in particular, are expected to have a lowercolonization potential than fungi with air-borne spores (e.g., ascomycetes,basidiomycetes) because they produce spores subterraneously and require stable substrateconditions for growth and development (Zubek et al., 2010), thus being hardly suited forthe dynamic canopy habitat (Janos, 1993). It has also been suggested that epiphytic fernsmay be less dependent on mycorrhizae because they have special adaptations (e.g.,drought resistance, leaf litter traps, thick storage rhizomes) that limit requirements formycorrhizae (Mehltreter, 2010).
Among the ferns growing on the ground, there are also slight differences between
LEHNERT&KESSLER: MYCORRHIZAL RELATIONSHIPS 107

substrates, with those on well-developed soil and rocks having high levels ofmycorrhization than those on sand – again a habitat that is unsuitable for fungal growth(Brundrett, 2009) – having fewer mycorrhizal fern species (Gemma et al., 1992).
A special case is provided by raw volcanic soils, especially on islands such asHawai’i. It has been suggested that such soils are more easily colonized bynonmycotrophic or facultatively mycorrhizal fern species due to the paucity of fungalinnoculum, perhaps facilitating later colonization of obligate mycotrophic ferns andangiosperms (Gemma & Koske, 1990; Gemma et al., 1992). This contrasts, however,with the regular presence of mycoheterotrophic gametophytes of Psilotum in volcanicsoils (Holloway, 1938; Duckett & Ligrone, 2005) and observations on La Réunion, wherethe percentage of terrestrial fern individuals with mycorrhizae was higher on shallow,raw soils with high pH values, high base-saturation and low nutrient availability (Kessleret al., 2010). These contrasting results may be due partly to the fact that the Hawai’istudies were based on the species level, whereas the results from La Réunion are basedon numbers of individuals.
Mycorrhization is absent to very rare in aquatic environments (Boullard, 1958; 1979;Lehnert et al., 2007; Brundrett, 2009), presumably reflecting the paucity of fungi in suchenvironments. Furthermore, the benefits mycorrhiza could bring are no longeradvantageous in a medium in which water and most nutrients are freely available andoften in surplus, but – at least in the case of fully submerged plants – assimilation ishampered by lower CO2 partial pressure and decreased light intensity. In some cases,amphibious species that grow both inundated and on dried-out soils are facultativelymycorrhizal, as found in some Isoetes (Beck-Nielsen & Madsen, 2001; Radhinka &Rodrigues, 2007). Likewise, species of Equisetum are non-mycorrhizal in wetlands, butcommonly mycorrhizal in dry environments (Harley & Harley, 1987; Marsh et al., 2000;Read et al., 2000; Brundrett, 2002).
Focussing on climatic gradients, the degree of mycorrhization has been documentedto decrease with elevation in Ecuador (Kessler et al., 2010). In addition to the commonlyproposed fungus-oriented explanations based on reduced fungal growth at lowtemperatures (Read & Haselwandter, 1981; Olsson et al., 2004; Schmidt et al., 2008),these authors also proposed a fern-oriented one in that mycorrhization might decreasebecause the relative costs of allocating carbon to the fungal partner increase withelevation (where photosynthesis decreases: Kessler et al., 2014b) and possibly exceedthe benefits of the fungal nutrient supply.
ECOLOGICAL EFFECTS OF MYCORRHIZATION IN FERNSBased on work on seed plants, it is known that fungi provide plants with soil nutrients(especially PO4), water, and other benefits (Blackwell, 2000; Wilkinson, 2001). Inexchange, the fungi obtain organic C from the photosynthetic host plants (Smith & Read,2008; Olsson et al., 2010), which allocate up to 20% of their net photosynthetic C totheir fungal partners (Allen et al., 2003). The relative costs of this fungal C use have tobe assessed in relation to the benefits derived from increased nutrient uptake, andpresumably can be offset partly by higher rates of photosynthesis, if the plants are notlight limited (Smith & Smith, 2011). In greenhouse experiments with potted plants,mycorrhizal fungi have been shown to increase leaf tissue nutrient concentrations, mostimportantly those of P (Stribley et al., 1980; Smith & Smith, 2011).
For ferns, very few data are available, and contrary to seed plants there are nogreenhouse experiments. In an early study, Cooper (1976) showed that mycorrhizal plants
108 FERN GAZ. 20(3):101-116. 2016

of the facultatively mycorrhizal Pteridium aquilinum (L.) Kuhn in New Zealand hadstronger growth than non-mycorrhizal individuals. More recently, it has been shown bothon La Réunion and in Ecuador that terrestrial mycorrhizal fern species are locally moreabundant than those lacking mycorrhizae (Kessler et al., 2010; 2014a). For example, onLa Réunion, the 78% mycorrhizal species made up 98% of all fern individuals. Thissuggests a clear ecological advantage provided by mycorrhizae to ferns.
On the other hand, a study in Ecuador found that fern species with mycorrhizae hadlower relative biomass increment per year than those without mycorrhizae (Kessler etal., 2010), suggesting a high C cost of mycorrhization. Even more surprisingly, non-mycorrhizal fern species had significantly higher concentrations of N, P, Mg, and Ca intheir leaves. Negative growth effects of mycorrhization have previously been documentedin a number of cases among seed plants (e.g., Johnson et al., 1992; Smith & Read, 2008;Veiga et al., 2011). These often reflect the parasitic nature of mycorrhizal fungi insituations in which the nutritional benefits for the plant do not outweigh the C loss to thefungi (Johnson et al., 1992; Karst et al., 2008).
The very sparse data available until now thus provide evidence for benefits ofmycorrhization in ferns as well as for negative effects. While this may initially besurprising, in fact it makes sense if we consider that not all plant species havemycorrhizae (Brundrett, 2009), even if they do not grow under special habitat conditionsthat limit fungal growth such as aquatic or epiphytic environments. If we further bear inmind that the ancestral state is most likely the presence of mycorrhization, then the 33%of terrestrial and 47% of epiphytic fern species lacking mycorrhizae (Lehnert, 2007)must have lost them over evolutionary times, suggesting that there must be a benefit oflosing mycorrhiza. It thus appears that there is a fine balance between positive andnegative effects of mycorrhization among land plants in general and ferns in particular.It is much too early to speculate under which conditions this balance switches either way,especially considering that the balance is likely to be not simply determined by a nutrient-carbon trade-off but also by water relations and the influence of symbiotic fungi onparasitic or pathogenic fungi and other biotic interactions.
OPEN QUESTIONSWe conclude this overview of what is known about mycorrhizae in ferns with a series ofquestions that readily develop from the above text. This list is certainly incomplete, butit may offer some guidelines as to what we would like to know to better understand therole of mycorrhization in ferns and lycophytes:
Are all visually detectable fungal associations of ferns truly mutualistic? Most of the•records are based on light microscopical screenings only and await furthermicrostructural and molecular conformation. Until now, true mycorrhizae with mutualbenefits have been confirmed only for lycophytes and basal fern lineages and involveonly Glomeromycota and Mucoromycotina.Why is the overall degree of mycorrhization lower among lycophytes and ferns (and•liverworts) than among seed plants? Does this reflect some basic physiologicaldifference between spore-bearing and seed-bearing vascular plants, e.g., in water useefficiency (Brodribb et al., 2009; Watkins et al., 2010) that tips the balance more inthe direction of non-mycorrhization in ferns?What is the role of mycorrhization in ferns that are very difficult to cultivate or•transplant, such as Gleicheniaceae, Lycopodiaceae and grammitids (Page, 2002)?For ferns with facultative or variable mycorrhization, does the degree of•
LEHNERT&KESSLER: MYCORRHIZAL RELATIONSHIPS 109

mycorrhization vary with ecological conditions? Are there implications of mycorrhizal associations in ferns for insect herbivory or•other ecological interactions?
ACKNOWLEDGEMENTSWe thank Bridget Laue and Adrian Dyer for the invitation to do this review, and MaryGibby and the reviewers for helping us improve the manuscript.
REFERENCESALLEN, M. F., SWENSON, W., QUEREJETA, J. I., EGERTON-WARBURTON, L. M.
& TRESEDER, K. K. 2003. Ecology of mycorrhizae: a conceptual framework forcomplex interactions among plants and fungi. Ann. Rev. Phytopath. 47: 271–303.
ANTUNES, P. M., KOCH, A. M., MORTON, J. B., RILLIG, M. C. & KLIRONOMOS,J. N. 2011. Evidence for functional divergence in arbuscular mycorrhizal fungi fromcontrasting climatic origins. New Phytol. 189(2): 507–514.
BARKER, S. J., TAGU, D. & DELP, G. 1998. Regulation of root and fungalmorphogenesis in mycorrhizal symbioses. Plant Physiol. 116(4): 1201–1207.
BECK-NIELSEN, D. & MADSEN, T.V. 2001. Occurrence of vesicular-arbuscularmycorrhiza in aquatic macrophytes from lakes and rivers. Aquatic Botany 71: 141–148.
BERCH, S. M. & KENDRICK, B. 1982. Vesicular-arbuscular mycorrhizae of SouthernOntario ferns and fern-allies. Mycologia 74: 769–776.
BLACKWELL, M. 2000. Terrestrial life - Fungal from the start? Science 289: 1884–1885.
BOULLARD, B. 1958. La mycotrophie chez les ptéridophytes. Sa fréquence, sescaractéres, sa signification. Doctor thesis, Université de Caen. E. Droulliard,Bordeaux.
BOULLARD, B. 1979. Consideration sur la symbiose fongique chez les Pteridophytes.Syllogeous 19: 1–59.
BONFANTE, P. & GENRE, A. 2008. Plants and arbuscular mycorrhizal fungi: anevolutionary-developmental perspective. Trends Plant Sci. 13(9): 492–498.
BOWER, F. O. 1923. The Ferns. Vol. I. University Press, Cambridge, 376 pp.BRODRIBB, T. J., MCADAM, S. A., JORDAN, G. J. & FEILD, T. S. 2009. Evolution
of stomatal responsiveness to CO2 and optimization of water‐use efficiency amongland plants. New Phytol. 183(3): 839–847.
BRUNDRETT, M. C. 2002. Coevolution of roots and mycorrhizas of land plants. NewPhytol. 154: 275–304.
BRUNDRETT, M. C. 2004. Diversity and classification of mycorrhizal associations.Bot. Rev. 79: 473–495.
BRUNDRETT, M. C. 2009. Mycorrhizal associations and other means of nutrition ofvascular plants: understanding the global diversity of host plants by resolvingconflicting information and developing reliable means of diagnosis. Plant Soil 320:37–77.
CAIRNEY, J. W. G. & MEHARG, A. A. 2003. Ericoid mycorrhiza: a partnership thatexploits harsh edaphic conditions. European Journal of Soil Science 54(4): 735–740.
CAVAGNARO, T. R., GAO, L. L., SMITH, F. A. & SMITH, S. E. 2001. Morphology ofarbuscular mycorrhizas is influenced by fungal identity. New Phytol. 151(2): 469–
110 FERN GAZ. 20(3):101-116. 2016

475.COOPER, K. M. 1976. A field survey of mycorrhizas in New Zealand ferns. N.Z. J. Bot.
14: 169–181.CULLINGS K. W. 1996. Single phylogenetic origin of ericoid mycorrhizae within the
Ericaceae. Can. J. Bot. 74: 1896–1909.DESIRÒ, A., DUCKETT, J. G., PRESSEL, S., VILLARREAL, J. C. & BIDARTONDO,
M. I. 2013. Fungal symbioses in hornworts: a chequered history. P. Roy. Soc. Lond.B Bio. 280(1759), p.20130207.
DHILLION, S. S. 1993. Vesicular-arbuscular mycorrhizas of Equisetum species inNorway and the USA: occurrence and mycotrophy. Mycol. Res. 97: 656–660.
DUCKETT, J. G. & LIGRONE, R. 1992. A light and electron microscope study of thefungal endophytes in the sporophyte and gametophyte of Lycopodium cernuum withobservations on gametophyte-sporophyte junction. Can. J. Bot. 70: 58–72.
DUCKETT, J. G. & LIGRONE, R. 2005. A comparative cytological analysis of fungalendophytes in the sporophyte rhizomes and vascularized gametophytes of Tmesipterisand Psilotum. Can. J. Bot. 83: 1443–1456.
FABER, B. A., ZASOSKI, R. J., MUNNS, D. N. & SHACKEL, K. 1991. A method formeasuring hyphal nutrient and water uptake in mycorrhizal plants. Can. J. Bot. 69(1),pp.87–94.
FERNANDEZ, N., MESSUTI, M. I. & FONTENLA, S. 2008. Arbuscular mycorrhizasand dark-septate fungi in Lycopodium paniculatum (Lycopodiaceae) and Equisetumbogotense (Equisetaceae) in a Valdivian temperate forest of Patagonia, Argentina.Amer. Fern J. 98: 117–127.
FIELD, K. J., PRESSEL, S., DUCKETT, J. G., RIMINGTON, W. R. & BIDARTONDO,M. I. 2015a. Symbiotic options for the conquest of land. Trends Ecol. Evol. 30(8):477–486.
FIELD, K. J., RIMINGTON, W. R., BIDARTONDO, M. I., ALLINSON, K. E.,BEERLING, D. J., CAMERON D. D., DUCKETT, J.G., LEAKE, J. R. & PRESSEL,S. 2015b. First evidence of mutualism between ancient plant lineages(Haplomitriopsida liverworts) and Mucoromycotina fungi and its response tosimulated Palaeozoic changes in atmospheric CO2. New Phytol. 205(2): 743–56.
FIELD, K. J., LEAKE, J. R., TILLE, S., ALLINSON, K. E., RIMINGTON, W. R.,BIDARTONDO, M. I., BEERLING, D. J. & CAMERON, D. D. 2015c. Frommycoheterotrophy to mutualism: mycorrhizal specificity and functioning inOphioglossum vulgatum sporophytes. New Phytol. 205(4): 1492–1502.
FRANK, B. 1885. Über die auf Wurzelsymbiose beruhende Ernährung gewisser Bäumedurch unterirdische Pilze. Ber. dt. Bot. Ges. (3): 128–145.
GEMMA, J. N. & KOSKE, R. E. 1990. Mycorrhizae in recent volcanic substrates inHawaii. Am. J. Bot. 77 (9): 1193–1200.
GEMMA J. N., KOSKE, R. E., & FLYNN, T. 1992. Mycorrhizae in Hawaiianpteridophytes: Occurrence and evolutionary significance. Am. J. Bot. 79: 843–852.
HARLEY, J. L. & HARLEY, E. L. 1987. A check‐list of mycorrhiza in the British flora.New Phytol. 105(s1): 1–102.
HIBBETT, D. S., BINDER, M., BISCHOFF, J. F., BLACKWELL, M., CANNON, P.F.,ERIKSSON, O.E., HUHNDORF, S., JAMES, T., KIRK, P. M., LÜCKING, R. &LUMBSCH, H. T. 2007. A higher-level phylogenetic classification of the Fungi.Mycol. Res. 111(5): 509–547.
HOLLOWAY, J.E. 1938. The embryo and gametophyte of Psilotum triquetrum. A
LEHNERT&KESSLER: MYCORRHIZAL RELATIONSHIPS 111

preliminary note. Ann. Bot. (Lond.) 2: 807–809. HORN, K., FRANKE, T., UNTERSEHER, M., SCHNITTLER, M. & BEENKEN, L.
2013. Morphological and molecular analyses of fungal endophytes of achlorophyllousgametophytes of Diphasiastrum alpinum (Lycopodiaceae). Am. J. Bot. 100(11):2158–2174.
IQBAL, S.H., YOUSAF, M. & YOUNUS, M. 1981. A field survey of mycorrhizalassociations in ferns of Pakistan. New Phytol. 87: 69–79.
ISHII, S. & LOYNACHAN, T. E. 2004. Rapid and reliable DNA extraction techniquesfrom trypan-blue-stained mycorrhizal roots: comparison of two methods. Mycorrhiza14(4): 271–275.
JANOS, D. P. 1993. Vesicular-arbuscular mycorrhizae of epiphytes. Mycorrhiza, 4(1):1–4.
JANSA, J., SMITH, F. A. & SMITH, S. E. 2008. Are there benefits of simultaneous rootcolonization by different arbuscular mycorrhizal fungi? New Phytol. 177(3): 779–789.
JOHNSON, N. C., COPELAND, P. J., CROOKSTON, R. K. & PFLEGER, F. L. 1992.Mycorrhizae: possible explanation for yield decline with continuous corn andsoybean. Agron. J. 84(3): 387–390.
JONES, D. L., HODGE, A. & KUZYAKOV, Y. 2004. Plant and mycorrhizal regulationof rhizodeposition. New Phytol. 163(3): 459–480.
JUMPPONEN, A. R. I. 2001. Dark septate endophytes–are they mycorrhizal?Mycorrhiza 11(4): 207–211.
JUMPPONEN, A. R. I. & TRAPPE, J. M. 1998. Dark septate endophytes: a review offacultative biotrophic root‐colonizing fungi. New Phytol. 140(2): 295–310.
JURKIEWICZ, A., RYSZKA, P., ANIELSKA, T., WALIGÓRSKI, P., BIAŁOŃSKA,D., GÓRALSKA, K., TSIMILLI-MICHAEL, M. & TURNAU, K. 2010.Optimization of culture conditions of Arnica montana L.: effects of mycorrhizal fungiand competing plants. Mycorrhiza 20(5): 293–306.
KARST, J., MARCZAK, L., JONES, M. D. & TURKINGTON, R. 2008. The mutualism-parasitism continuum in ectomycorrhizas: a quantitative assessment usingmeta-analysis. Ecology 89(4): 1032–1042.
KENRICK, P. & STRULLU-DERRIEN, C. 2014. The origin and early evolution of roots.Plant Physiol. 166(2): 570–580.
KESSLER, M., JONAS. R., STRASBERG, D. & LEHNERT, M. 2010. Mycorrhizalcolonizations of ferns and lycophytes on the island of La Réunion in relation tonutrient availability. Basic Appl. Ecol. 11: 329–336.
KESSLER, M., JONAS, R., CICUZZA, D., KLUGE, J., PIATEK, K., NAKS, P. &LEHNERT, M. 2009. A survey of the mycorrhization of Southeast Asian ferns andlycophytes. Plant Biol. 12: 788–793.
KESSLER, M., GÜDEL, R., SALAZAR L., HOMEIER J. & KLUGE J. 2014a. Impactof mycorrhization on the abundance, growth and leaf nutrient status of ferns along atropical elevational gradient. Oecologia 175: 887–900.
KESSLER, M., SALAZAR, L., HOMEIER J. & KLUGE, J. 2014b. Species richness-productivity relationships of tropical terrestrial ferns at regional and local scales. J.Ecol. 102: 1623–1633.
LARA-PÉREZ, L.A., VALDÉS-BAIZABAL, M.D., NOA-CARRAZANA, J.C.,ZULUETA-RODRÍGUEZ, R., LARA-CAPISTRÁN, L. & ANDRADE-TORRES,A. 2015. Mycorrhizal associations of ferns and lycopods of central Veracruz, Mexico.
112 FERN GAZ. 20(3):101-116. 2016

Symbiosis 65(2): 85–92.LEHNERT, M., KOTTKE, I., SETARO, S., PAZMINO, L. F., SUÁREZ, J. P. &
KESSLER, M. 2009 (2010). Mycorrhizal associations in ferns from SouthernEcuador. Amer. Fern J. 99: 292–306.
LEKBERG, Y., ROSENDAHL, S. & OLSSON, P.A. 2015. The fungal perspective ofarbuscular mycorrhizal colonization in ‘nonmycorrhizal’plants. New Phytol. 205(4):1399–1403.
LESICA, P. & ANTIBUS, R. K. 1990. The occurrence of mycorrhizae in vascularepiphytes of two Costa Rican rain forests. Biotropica 33: 250–258.
MARSH, A. S., ARNONE, J. A. A., III., BORMANN, B. T. & GORDON, J. C. 2000.The role of Equisetum in nutrient cycling in an Alaskan shrub wetland. J. Ecol. 88:999–1011.
MEHLTRETER, K. 2010. Interactions of ferns with fungi and animals. Pp. 220–254. In:Mehltreter, K., Walker, L. R. & Sharpe, J. M. (Eds) Fern Ecology. Cambridge Univ.Press, Cambridge.
MOTEETEE, A., DUCKETT, J. G. & RUSSELL, A. J. 1996. Mycorrhizas in the fernsof Lesotho. In: Camus, J. M., M. Gibby and R. J. Johns (Eds) Pteridology inperspective, pp. 621–631. Royal Botanic Gardens, Kew.
MUTHUKUMAR, T. & PRABHA, K. 2012. Fungal associations in gametophytes andyoung sporophytic roots of the fern Nephrolepis exaltata. Acta Bot. Croat., 71(1):139–146.
MUTHUKUMAR, T. & PRABHA, K. 2013. Arbuscular mycorrhizal and septateendophyte fungal associations in lycophytes and ferns of south India. Symbiosis59(1): 15–33.
NEWMAN, E. I. & REDDELL, P. 1987. The distribution of mycorrhizas among familiesof vascular plants. New Phytol. 106: 745–751.
NEWSHAM, K. K., FITTER, A. H. & WATKINSON, A. R. 1995. Arbuscularmycorrhiza protect an annual grass from root pathogenic fungi in the field. J. Ecol.83(6): 991–1000.
OGURA-TSUJITA, Y., HIRAYAMA, Y., SAKODA, A., SUZUKI, A., EBIHARA, A.,MORITA, N., & IMAICHI, R. 2016. Arbuscular mycorrhizal colonization in field-collected terrestrial cordate gametophytes of pre-polypod leptosporangiate ferns(Osmundaceae, Gleicheniaceae, Plagiogyriaceae, Cyatheaceae). Mycorrhiza 26(2):87–97.
OGURA-TSUJITA, Y., SAKODA, A., EBIHARA, A., TOMOHISA YUKAWA, T. &IMAICHI. R. 2013. Arbuscular mycorrhiza formation in cordate gametophytes oftwo ferns, Angiopteris lygodiifolia and Osmunda japonica. J. Plant Res. 126 (1): 41–50.
OLSSON, P. A., ERIKSEN, B. & DAHLBERG, A. 2004. Colonization by arbuscularmycorrhizal and fine endophytic fungi in herbaceous vegetation in the Canadian HighArctic. Can. J. Bot. 82: 1547–1556.
OLSSON, P. A., RAHM, J. & ALIASGHARZAD, N. 2010. Carbon dynamics inmycorrhizal symbioses is linked to carbon costs and phosphorus benefits. FemsMicrobiol. Ecol. 72: 125–131.
PAGE, C. N. 2002. Ecological strategies in fern evolution: a neopteridological overview.Rev. Palaeobot.Palyn. 119(1): 1–33.
PAZMINO, L. 2006. Estudio preliminar de hongos que forman micorrizas en helechosde las familias Grammitidaceae y Polypodiaceae en el bosque de la ECSF y El Tiro,
LEHNERT&KESSLER: MYCORRHIZAL RELATIONSHIPS 113

Loja, Ecuador. BSc thesis, Universidad Tecnica Particular de Loja, Loja, Ecuador.pdf available upon request from the author.
PEARSON, V. & READ, D.J. 1973. The biology of mycorrhiza in the Ericaceae: I. Theisolation of the endophyte and synthesis of mycorrhizas in aseptic culture. NewPhytol. 72(2): 371–379.
PIROZYNSKI, K. A. & MALLOCH, D. W. 1975. The origin of land plants: A matter ofmycotrophism. BioSystems 6: 153–164.
PRESSEL, S., BIDARTONDO, M. I., LIGRONE, R. O. & DUCKETT, J. G. 2010.Fungal symbioses in bryophytes: new insights in the twenty first century. Phytotaxa9(1): 238–53.
RADHINKA, K. P. & RODRIGUES, B. F. 2007. Arbuscular mycorrhizae in associationwith aquatic and marshy plants in Goa, India. Aquatic Bot. 86: 291–294.
READ, D. J. 1996. The structure and function of the ericoid mycorrhizal root. Ann. Bot.77(4): 365–374.
READ, D. J. & HASELWANDTER, K. 1981. Observations on the mycorrhizal status ofsome alpine plant communities. New Phytol. 88: 341–352.
READ, D.J., DUCKETT, J.G., FRANCIS, R., LIGRONE, R. & RUSSELL, A. 2000.Symbiotic fungal associations in “lower” land plants. P. Trans. Roy. Soc. B 355: 815–831.
REMY, W., TAYLOR, T.N., HASS, H. & KERP, H. 1994. Four hundred-million-year-old vesicular arbuscular mycorrhizae. PNAS 91(25): 11841–11843.
REYES–JARAMILLO, I., CAMARGO-RICALDE, S. L. & AQUIAHUATL-RAMOS,M.A. 2008. Mycorrhizal-like interaction between gametophytes and youngsporophytes of the fern Dryopteris muenchii (Filicales) and its fungal endophyte.Rev. Biol. Trop. 56(3): 1101–1107.
RIMINGTON, W.R., PRESSEL, S., DUCKETT, J.G. & BIDARTONDO, M.I. 2015.Fungal associations of basal vascular plants: reopening a closed book? New Phytol.205(4): 1394–1398.
SCHMID, E. & OBERWINKLER, F. 1994. Light and electron microscopy of the host–fungus interaction in the achlorophyllous gametophyte of Botrychium lunaria. Can.J. Bot. 72(2): 182–188.
SCHMID, E. & OBERWINKLER, F. 1995. A light‐and electron‐microscopic study ona vesicular–arbuscular host‐fungus interaction in gametophytes and youngsporophytes of the Gleicheniaceae (Filicales). New Phytol. 129(2): 317–324.
SCHMID, E. & OBERWINKLER, F. 1996 Light and electron microscopy of a distinctiveVA mycorrhiza in mature sporophytes of Ophioglossum reticulatum. Mycol. Res.100: 843–849.
SCHMID, E., OBERWINKLER, F. & GOMEZ L. D. 1995 Light and electronmicroscopy of a host-fungus interaction in the roots of some epiphytic ferns fromCosta Rica. Can. J. Bot. 73: 991–996.
SCHMIDT S. K., SOBIENIAK-WISEMAN, L.C., KAGEYAMA, S. A., HALLOY, S.R. P. & SCHADT, C. W. 2008. Mycorrhizal and dark-septate fungi in plant rootsabove 4270 meters elevation in the Andes and Rocky Mountains. Arct. Antarct. Alp.Res. 40: 576–583.
SCHUßLER, A., SCHWARZOTT, D. & WALKER, C. 2001. A new fungal phylum, theGlomeromycota: phylogeny and evolution. Mycological Research 105: 1413–1421.
SELOSSE, M. A. & ROUSSET, F. 2011. The plant-fungal marketplace. Science333(6044): 828–829.
114 FERN GAZ. 20(3):101-116. 2016

SELOSSE, M. A., STRULLU‐DERRIEN, C., MARTIN, F. M., KAMOUN, S. &KENRICK, P. 2015. Plants, fungi and oomycetes: a 400‐million year affair thatshapes the biosphere. New Phytol. 206(2): 501–506
SIDDIQUI, Z.A. & PICHTEL, J. 2008. Mycorrhizae: an overview. In: Siddiqui, Z. A.,Akhtar, M. S. & Futai, K. (Eds) Mycorrhizae: Sustainable Agriculture and Forestry,pp. 1–35. Springer Science and Business Media B.V.
SMITH, A. R., PRYER, K. M., SCHUETTPELZ, E., KORALL, P., SCHNEIDER, H.& WOLF, P.G. 2006. A classification for extant ferns. Taxon 55: 705–731.
SMITH, S. E. & READ, D. J. 2008. Mycorrhizal Symbiosis. Academic Press; 3rd edition.SMITH, S. E. & SMITH, F. A. 2011. Roles of arbuscular mycorrhizas in plant nutrition
and growth: New paradigms from cellular to ecosystem scales. Ann. Rev. Plant Biol.62: 227–50.
STRIBLEY, D.P., TINKER, P. B. & RAYNER, J. H. 1980. Relation of internalphosphorus concentration and plant weight in plants infected by vesicular-arbuscularmycorrhizas. New Phytol. 86: 261–266.
STRULLU-DERRIEN, C. & STRULLU, D. G. 2007. Mycorrhization of fossil and livingplants. Comptes Rendus Palevol, 6(6), pp.483–494.
STRULLU‐DERRIEN, C., KENRICK, P., PRESSEL, S., DUCKETT, J. G., RIOULT,J. P., & STRULLU, D. G. 2014. Fungal associations in Horneophyton ligneri fromthe Rhynie Chert (c. 407 million year old) closely resemble those in extant lowerland plants: novel insights into ancestral plant–fungus symbioses. New Phytologist203(3): 964–979.
TAYLOR, T. N., REMY, W., HASS, H. & KERP, H. 1995. Fossil arbuscular mycorrhizaefrom the Early Devonian. Mycologia 87(4): 560–573.
TAYLOR, A. F. & ALEXANDER, I. A. N. 2005. The ectomycorrhizal symbiosis: life inthe real world. Mycologist 19(03): 102–112.
TRAPPE, J. M. 1987. Phylogenetic and ecologic aspects of mycotrophy in theangiosperms from an evolutionary standpoint. In: Safir, G.R. (Ed.) Ecophysiologyof VA mycorrhizal plants, pp. 2–25. CRC Press, Boca Raton.
TURNAU, K., ANIELSKA, T. & JURKIEWICZ, A. 2005. Mycothallic/mycorrhizalsymbiosis of chlorophyllous gametophytes and sporophytes of a fern, Pellaea viridis(Forsk.) Prantl (Pellaeaceae, Pteridales). Mycorrhiza 15(2): 121–128.
VAN DER HEIJDEN, M. G., MARTIN, F. M., SELOSSE, M. A. & SANDERS, I. R.2015. Mycorrhizal ecology and evolution: the past, the present, and the future. NewPhytol. 205(4): 1406–1423.
VEIGA, R. S. L, JANSA, J., FROSSARD, E. & VAN DER HEIJDEN M. G. A. 2011.Can arbuscular mycorrhizal fungi reduce the growth of agricultural weeds? PloS One6: e27825.
WANG, B. & QIU, Y. L. 2006. Phylogenetic distribution and evolution of mycorrhizasin land plants. Mycorrhiza 16: 299–363.
WANG, B., YEUN, L., XUE, J. Y., LIU, Y., ANÉ, J. M. & QIU, Y. L. 2010. Presence ofthree mycorrhizal genes in the common ancestor of land plants suggests a key roleof mycorrhizae in the colonisation of land by plants. New Phytol. 186(2): 514–525.
WARREN, J. M., BROOKS, J. R., MEINZER, F. C. & EBERHART, J. L. 2008.Hydraulic redistribution of water from Pinus ponderosa trees to seedlings: evidencefor an ectomycorrhizal pathway. New Phytol. 178(2): 382–394.
WATKINS, J.E., HOLBROOK, N.M. & ZWIENIECKI, M.A. 2010. Hydraulic propertiesof fern sporophytes: consequences for ecological and evolutionary diversification.
LEHNERT&KESSLER: MYCORRHIZAL RELATIONSHIPS 115

Amer. J. Bot. 97(12): 2007–2019.WILKINSON, D. M. 2001. Mycorrhizal evolution. Trends Ecol. Evol. 16: 64–65.WINTHER, J. L. & FRIEDMAN, W. E. 2007a. Arbuscular mycorrhizal associations in
Lycopodiaceae. New Phytol. 177(3): 790–801.WINTHER, J. L. & FRIEDMAN, W. E. 2007b. Arbuscular mycorrhizal symbionts in
Botrychium (Ophioglossaceae). Amer. J. Bot. 94(7): 1248–1255.WINTHER, J. L. & FRIEDMAN, W. E. 2009. Phylogenetic affinity of arbuscular
mycorrhizal symbionts in Psilotum nudum. J. Plant Res. 122(5): 485 496.ZHAO, Z. W. 2000. The arbuscular mycorrhizas of pteridophytes in Yunnan, southwest
China: evolutionary interpretations. Mycorrhiza 10: 145–149.ZUBEK, S., PIĄTEK, K., NAKS, P., HEISE, W., WAYDA, M. & MLECZKO, P. 2010.
Fungal root endophyte colonization of fern and lycophyte species from CelaqueNational Park in Honduras. Amer. Fern. J 100: 126–136.
116 FERN GAZ. 20(3):101-116. 2016

A SHORT BIOGRAPHY OF THE AUTHORS
Michael Kessler was born and grew up in Peru where he spent much of his childhoodchasing birds across the forest and mountains of the country. He studied biology at theUniversity of Göttingen, Germany from 1986 onwards, culminating with his PhD thesison the taxonomy, ecology, and biogeography of the high-Andean tree genus Polylepis(Rosaceae) in 1995. He then shifted his focus to the plant diversity of tropical montaneforests, and, inspired by the work of Hanna Tuomisto in Amazonia, picked ferns as astudy group. Over the years, ferns have become the main focus of his work, with researchmainly on diversity patterns and ecology, but also numerous taxonomic studies,especially on Bolivian ferns. Since 2008, he has worked as scientific curator of thebotanical garden and researcher at the University of Zurich, Switzerland. Currently, hisfern work is mainly concerned with understanding the evolutionary and ecologicalprocesses that determine the richness and composition of fern assemblages, involvingfieldwork in, e.g., Mexico, the Andes, Uganda, Indonesia, and New Guinea. Otherprojects involve studies on the diversification of bromeliads and palms in the neotropicsand the flora of the canton of Zurich in Switzerland.
FERN GAZ. 20(3). 2016 117

FERN GAZ. 20(3). 2016 118
Marcus Lehnert was born on 1 October 1975 in Neutstadt/Ostholstein, Germany. Heattended Göttingen University, where he received his Diplom (M.S. equivalent) in 2002and his Ph.D. in 2007. His research was based first at the Albrecht-von-Haller Institutefor Plant Sciences, Göttingen (2000–2007), with interludes at the Instituto de Ecología,Universidad Mayor de San Andres, La Paz, Bolivia (2002–2003) and the University ofCalifornia, Berkeley (2006–2007). From 2008-2012, he was employed as a PostdoctoralResearcher in the Botany Department of the State Museum for Natural History inStuttgart. Since 2013 he has been a research assistant at the Nees Institute for PlantBiodiversity, University Bonn. His research interest focuses on the taxonomy, phylogeny,and floristics of ferns but also includes mycorrhiza, general biogeography, and thesystematics of several angiosperm families. For his fieldwork, he has visited amongothers Puerto Rico, Panama, Peru, Argentina, Brazil, and several times Ecuador,Colombia, Bolivia and New Guinea. Marcus Lehnert is currently working on a large-scale projects focusing on the use of DNA barcoding for the rapid assessment ofbiodiversity (Accelaration of Biodiversity Assessment—ABA Ecuador and GermanBarcode of Life-GBoL). He has contributed his taxonomical expertise on scaly tree ferns(Cyatheaceae) to floral projects like “Catálogo de las Plantas Vasculares del Cono Sur”and “Ferns of Bolivia.”

THE FERN FAMILY PTERIDACEAE IN TURKEY
M. BONA1 & M. GIBBY2
1Department of Botany, Faculty of Science, Istanbul University, Istanbul, Turkey. E-mail: [email protected]
2Royal Botanic Garden Edinburgh, Edinburgh, UK. E-mail: [email protected]
Keywords: Adiantum, Allosorus, Anogramma, Cheilanthes, Cryptogramma,Paragymnopteris, Pteridaceae, Pteris, Cosentinia, Turkey.
ABSTRACTThe last comprehensive study of Turkish Pteridaceae was made over 50 yearsago. In this study the family Pteridaceae in Turkey is reviewed according to recenttaxonomic and nomenclatural changes and a full description, synonyms anddistribution information are given for each taxon and a key provided.
INTRODUCTIONA generic classification of cheilanthoid ferns (Pteridaceae) has eluded taxonomists formore than 200 years (Grusz & Windham, 2013). Several taxonomic and nomenclaturalupdates have been made in recent decades, based on morphological and molecularmethods (Gastony & Rollo, 1995; Sanchez-Baracaldo, 2004; Zhang et al., 2005; Smithet al., 2006; Kamau, 2007; Schuettpelz et al., 2007; Windham et al., 2009; Grusz &Windham, 2013; Schneider et al., 2013; Abotsi et al., 2015). The current family diversityis about 1000 species distributed in 53 genera (Schuettpelz et al., 2007; Christenhusz etal., 2011).
The last comprehensive study of Turkish Pteridaceae was made over 50 years ago(Henderson, 1965) for the Flora of Turkey. A major project was recently initiated toillustrate and describe the flora of Turkey in the Turkish language and the first volumeof Resimli Türkiye Florası [Illustrated Flora of Turkey] was published by Güner andEkim (2014). The second volume, which will include the family Pteridaceae, is scheduledfor publication in 2016. The aim of the present study has been to review TurkishPteridaceae according to recent taxonomic and nomenclatural changes and to providedescriptions and distributions for each taxon. It is based on recent literature onPteridaceae (e.g. Nardi & Reichstein, 1985; Benlioğlu et al., 1997; Kaynak et al., 2008;Christenhusz et al., 2011; Greuter & Raab-Straube, 2012; Jessen et al., 2012; Güner etal., 2012), and on herbarium specimens from Ankara University Faculty of Pharmacy(AEF), Ankara University Faculty of Science (ANK), Natural History Museum, London(BM), Royal Botanic Garden Edinburgh (E), Geneva (G), Gazi University Faculty(GAZI), Hacettepe University (HUB), Istanbul University Faculty of Pharmacy (ISTE)and Royal Botanic Gardens, Kew (K). All specimens listed below are from E, exceptwhere indicated otherwise.
PTERIDACEAE E.D.M.Kirchn., Schul-Bot.: 109 (1831).Synonym: Adiantaceae Newman, Hist. Brit. Ferns: 5 (1840), Sinopteridaceae Koidz.,Acta Phytotax. Geobot. 3: 50 (1934). Cryptogrammaceae Pic.Serm., Webbia 17: 299(1963). Cheilanthaceae B.K.Nayar, Taxon 19: 233 (1970).
FERN GAZ. 20(3):119-132. 2016 119

Key for Turkish Pteridaceae taxa 1. Fertile and sterile leaves or pinnae conspicuously dimorphic ................................... 21. Fertile and sterile leaves or pinnae not, or weakly dimorphic ....................................42.Annual, sori on the lower surface of the leaf, indusium absent .......................................................................................................................................Anogramma leptophylla2. Perennial, sori marginal, covered by the recurved leaf margin ...................................33. Lamina of sterile leaves up to 16 cm long, ovate to deltoid-ovate, spores <60μm long.......................................................................................................Cryptogramma crispa3. Lamina of sterile leaves up to 7 cm long, broadly deltoid, spores >60μm long ............................................................................................................Cryptogramma bithynica4. Pinnules fan-shaped; petiole blackish, glabrous, fine and wiry ....................................................................................................................................Adiantum capillus-veneris4. Pinnules not fan-shaped; petiole not as above.............................................................55. Fronds pinnate to pinnatisect, weakly dimorphic........................................................65. Fronds 2-pinnate, not dimorphic .................................................................................7 6. Leaves 1-pinnate, lanceolate, >10 pinnae pairs ...................................... Pteris vittata6. Lower pinnae pinnatisect, frond ovate-triangular, with 7 or fewer pinnae pairs................................................................................................................................Pteris cretica7. Underside of pinnules glabrous or with glandular hairs..............................................87. Underside of pinnules densely covered with scales or eglandular hairs ...................108. Pseudoindusium entire, frond sparsely glandular below.................... Allosorus tinaei8. Pseudoindusium ciliate, frond eglandular below ........................................................99. Pseudoindusium narrow, with dense long marginal hairs, spores <45 μm long ........................................................................................................................Allosorus persicus9. Pseudoindusium broad, irregular, with short cilia, spores >45 μm long ................................................................................................................................Allosorus acrosticus10. Upper surface of pinnules glabrous ...............................Paragymnopteris marantae10. Upper surface of pinnules white, woolly ........................................Cosentinia vellea
Adiantum L., Sp. Pl. 1094 (1753).Short-creeping, scaly rhizome. Fronds dense or sparse; stipe dark brown or black shiny.Pinnae fan-shaped, without a midrib, sori borne on reflexed marginal lobes of lamina.A. capillus-veneris L., Sp. Pl. 2: 1096 (1753).Synonyms: Adiantum capillus Sw., J. Bot. (Schrader) (2): 83 (1801). A. formosum R.Br.,Prodr. Fl. Nov. Holland 155 (1810). A. schaffneri Fourn., Bull. Soc. Bot. France 27: 328(1880). A. modestum Underw., Bull. Torrey Bot. Club. 28: 46 (1901). A. michelii Christ,Bull. Geogr. Bot. Mans. 12 (1910).Lectotype: Herb. Linn. 1252.9 (LINN)!Rhizome densely covered with shiny, narrow lanceolate scales. Fronds bipinnate, (5–)15–30(–45) × (1.5–)4–6(–10) cm, glabrous; stipe (1.5–)8–12(–20) cm long, brown orblack, shiny, glabrous. Pinnae 2–7 × 1–4 cm with stalk 0.3–3 cm long. Pinnules fan-shaped, (0.7–)1–2(–3.5) × (0.7–)1–2.4(–3) cm, cuneate at base, without midrib, 2–8 lobedwith stalk (1–)2–5(–16) mm long. Sori borne on reflexed marginal lobes of lamina.Habitat: In crevices of limestone and serpentine or tufa and base-rich soil on shady banks,1–1000 m. Distribution in Turkey:Adana: Feke; Göksu gorge, 800 m, 09.vii.1952, Davis 19855!Antalya: Kemer, Sumakşehiri Deresi, kalkerli G. Deresi değil vadisi, P. nigra-C.
120 FERN GAZ. 20(3):119-132. 2016

sempervirens ormanı 500-700 m, 06.x.1977, H. Peşmen 3569 & Ş. Yıldırımlı HUB36734! Alanya, 120 m, 04.viii.1982, Ş. Yıldırımlı 4451 HUB 36732! Aksu, DüdenÇağlayanı, 06.ix.1988, Ş. Yıldırımlı 11596 HUB 36736!Artvin: Çoruh gorge, between Yusufeli and Artvin. 600 m, 01.viii.1966, Davis 47690!Demirkent nahiyesi, zwischen Tortum Gölü und Artvin, 620 m, 03.viii.1973, F. Holtz, P.Hanel & T. Kesercioğlu 991! Hopa, Kemalpaşa-Sarp arası, denize dik kayalıklar, 0-50m, 19.v.1985, Ş. Yıldırımlı 7898 HUB 36740!Bursa: Uludağ, 25.vi.1946, A. Yılmak!Denizli: 13.vii.1947, Davis 13263! Pamukkale, 16.x.1980, Ş. Yıldırımlı HUB 36728!Düzce: Akçakoca, Edilli Köyü, kıyı kumul, orman, 50 m, 29.ix.2001, A. Doğru Koca1489 (HUB)!Hatay: Dörtyol, Kuzuculu to Bülke on forest road, 1000 m, 04.iv.1965, M.J.E. Coode &B.M.G. Jones 416! Valley above Dörtyol, south bank, north facing, boulder clay andserpentine, 150 m, 12.viii.1969, J. Darrah 623! Valley above Yeşilkent, 305 m,11.viii.1969, J. Darrah 505! Samadağ, Çevlik, Vespasiyanus ve Titus Tüneli boyunca,Beşikli mağarası ve taşlık maki, 10-150 m, 04.ix.1990, Ş. Yıldırımlı 13926 (GAZI)!Antakya, Harbiye, Şelale, 180 m, 25.iv.1978, M. Coşkun 332, B. Şener & F. Ilisulu AEF6353! Kırıkhan, Delibekirli, nemli kayalıklar, 500 m, 21.iv.1977, M. Koyuncu AEF 5668!Musa Dağı, nemli yerlerde, 14.iv.1967, Y. Akman ANK 7943!İçel: Kanlıdivane, 23.iv.1974, H. Peşmen & S. Erik HUB 36733! Anamur, NasrettinKöyü üstü, 21.iv.1975, M. Koyuncu & M. Coşkun AEF 4854!İzmir: Kemalpaşa, Next to a stream in shady ravine in macchia in the foothills of NifDağ, 06.vi.1966, R. Alava & C. von Regel 5040!Karamanmaraş: Andırın; 8 miles South of Çatak, 800 m, 17.v.1965, M.J.E. Coode &B.M.G. Jones 1147! Süleymanlı, Akdağ, nemli dere içi, 1500 m, 24.vii.1977, B. Yıldız1555 HUB 36731!Konya: Ermenek, Kazancı kasabası, Yeşilköy, Göksu Nehri boyunca, 700 m, 12.x.1983,H. Sümbül 2536 HUB 36730! Ermenek, Göktepe Kasabası, Darcın Köyü, Ayaşlı mevkii,Göksu Nehri Boyunca, 600 m, 15.ix.1983, H. Sümbül 2477 HUB 36729!Malatya: Pütürge, Ferikhan Köyü, dere kenarı, ca. 650 m, 06.v.1981, Y Altan ANK 1252!Manisa: Manisa Dağı (Spil), Karaçay boyunca, 150-450 m, 23.iv.1984, Ş. Yıldırımlı 6014,HUB 36739!Mardin: Hessana at S foot of Cudi Dağ, 900 m, 10.v.1966, Davis 42776! Kasrık Boğazı,11 km from Cizre to Şırnak, shady river bank near bridge, 400 m, 7.v.1966, Davis 42641!Dargeçit, Çelik Köyü, Umutlu mezrası, kaya üzeri, 520 m, S. Aslan 3308 et al. (GAZI)!Muğla: 13 km South of Köyceğiz, by road to Dalyan, just east of the village of Tepearası,10–20 m, 19.v.1990, R. Lampinen 7492! Köyceğiz, Yangı Köyü Yangı Deresi, kalkerli,sarp derin vadi, kazmofit, 22.iii.1981, A. Güner 8421 et al. HUB 36725! Fethiye, KatrancıAdası, 0-20 m, 21.v.2011, Z. Aytaç 9830 (GAZI)! Boyobası Köyü, Saklayıcı mevkii,şistik arazi, nemli çakıllık, 120 m, 18.ii.1991, A. Güner 8103 et al. (GAZI)! Yatağan-Milas arası, su kenarı, 21.v.1970, N. Tanker AEF 4851!Rize: Rize-Çayeli arası, c. 10 m, yol kenarı, 19.viii.1985, A. Güner 6848 & M. VuralHUB 36726! Pazar-Hemşin, nemli kaya üzerinde, 150 m, 19.vi.1984, M. Vural 2913HUB 36735!Sakarya: Adapazarı; Karasu to Söğütlü, 30 m, 07.viii.1962, Davis & Coode, D. 39101!Samsun: Samsun’un 10 km doğusu, asfalt kenarı, 10 m, 12.ix.1964, H. Kayacık & G.Elçin 2211! Seaside cliffs, 3 miles east of Samsun, 10 m, 19.v.1963, Tobey 133!Sinop: On south wall of bath, 10 m, 23.iv.1966, Tobey 1624! Ayancık, Bakırlı Köyü yol
BONA & GIBBY: THE FERN FAMILY PTERIDACEAE IN TURKEY 121

ayrımından Akgöl’e 2 km, 15.ix.2009, A. Dönmez 16205 & Z. Uğurlu (HUB)!Şanlıurfa: Urfa-Gaziantep c. 1 km from Urfa, 600 m, 11.ix.1956, McNeill 794!Trabzon: Maçka, 24 km south of Trabzon, 250 m, 28.vi.1960, Stainton & Henderson5895!General Distribution: Widespread in temperate and subtropical regions of the New andOld World, absent from South America.
Anogramma Link, Fil. Spec. 137 (1841). Small, annual, delicate plants with short rhizome. Prothallus perennial and mostlypersistent. Fronds dimorphic, mature fertile fronds bi-tripinnate, pinnae simple. Sorilinear, indusium absent.A. leptophylla (L.) Link, Fil. Spec. 137 (1841).Synonyms: Polypodium leptophyllum L., Sp. Pl. 2: 1092 (1753); Acrostichumleptophyllum Lam. & DC., Fl. Franc. (de Candolle & Lamarck), ed. 3. 2: 565. (1805);Grammitis leptophylla (L.) Sw. Syn. Fil. (Swartz) 23, 218, t. 1. f. 6. (1806);Gymnogramma leptophylla (L.) Desv. in Berl. Mag. 5: 305 (1811); Hemionitisleptophylla Lag. Gen. Sp. Pl. (Lagasca) 33: (1816).Lectotype: Herb. Linn. 1251: 56 (LINN)!Plant delicate, up to 15 cm high, annual, with short rhizomes. Prothallus perennial,reniform, bilobed, 7–10 mm long. Fronds yellow-green, glabrous, young fronds reniformand 3–5 lobed, mature fronds bipinnate 5–15 ×1–1.5 (–2) cm, stipe glabrous 2.5–7 cmlong, dark brown-black below and yellow-green above. Pinnae glabrous, 0.5–2 × 0.5–1cm, dichotomously veined, pinnae stipe 1–2 mm. Pinnule mostly bilobed or lanceolate.Sori linear along veins; no indusium.Habitat: Dry metamorphic rocks or moist shady serpentine rocks, 30–1000 m.Distribution in Turkey:Adana: Feke; Senean dere between Gurumze and Suphandere, 1000 m, 01.vii.1952,Davis 1961, Dodds & Çetik!Antalya: Rocks near the sea, 12.iv.1955, H. Walter 3891! Side, c. 40 m, 19.v.1994, K.Karamanoğlu, M. Koyuncu & M. Coşkun AEF 16! Kemer, Kesmeboğaz, kalker kayalığı,200 m, 25.iii.1978, H. Peşmen 3649 & B. Yıldız HUB 36798! Kemer, Beycik-DelikinTepesi arası, 500-700 m, kalker kayalığı, P. brutia maki karışık ormanı, 28.iv.1980, H.Peşmen 4818 HUB 36796!Hatay: Dörtyol; Rabat, vertical serpentine flush, c. 100 m, 05.v.1965, M.J.E. Coode &B.M.G. Jones!Isparta: Ş.Karaağaç, Kızıldağ Milli Parkı, Kuzgu Tepe, kaya üzeri, 1350 m, 24.vii.1994,B. Mutlu 1084 HUB 36794!İçel: Anamur, Nasrettin Köyü üstü, pınar civarı nemli yerlerde, 250 m, 21.iv.1975, M.Koyuncu & M. Coşkun HUB 36797!Kastamonu: Taşköprü, Kayabaşı Köyü yukarısı, yangın kulesine tırmanış, dağ bozkırı,P. nigra-P. sylvestris açıklığı, 04.viii.2010, A. Dönmez 17533 & Z. Uğurlu (HUB)!Muğla: behind Marmaris, 50–100 m, 14.iv.1965, Davis 41407! Marmaris, On soilamongst rocks in ravine forest near Hotel Lidya, 06.v.1984, E.C. Wallace! Marmaris,Pinetum brutiae, on serpentine, 30 m, 24.iii.1956, Davis 25258 & O. Polunin! Ortaca,Dalyan, Gökbel Köyü, Kızıl Çam ormanı, kalkerli arazi, 100-150 m, 14.iv.1991, A. Güner8622 et al. HUB 36795! Köyceğiz, Ekincik Köyü, Sandallı tepe, makilik, kalkerli arazi,10-500 m, 19.v.1991, A. Güner 9097, M. Vural & H. Şağban HUB 36799! Ortaca,Dalyan, Marmarlı çevresi, makilik arazi, 30 m, 19.iv.1991, A. Güner 8918 et al. (GAZI)!
122 FERN GAZ. 20(3):119-132. 2016

General Distribution: Mediterranean area, Switzerland, W. France and Channel Islands,Crimea, N. America; scattered to S. Africa, India and Australasia.
Allosorus Bernh., Neues J. Bot. 1(2): 36 (1805).Rhizome short-creeping or tufted. Fronds persistent, bipinnate; stipe wiry. Sori terminalon veins, protected by a linear pseudoindusium, a recurved and modified leaf margin.A. acrosticus (Balbis) Christenh. Willdenowia 42(2):284. (2012) Synonyms: Pteris acrostica Balb. Elenco 98. (1801). Cheilanthes acrostica (Balbis) Tod.Giornale de Scienze Naturali ed Economiche de Palermo 1(10): 215. (1866). Lectotype: “Polypodium ilvense Bell. / puto esse potius novam plantam pteridis genus”[Balbis’s handwriting] / “est Polypodium fragrans L.” [not Balbis’s handwriting] (TO).Four plants. (see Nardi & Reichstein. 1985).Plant up to 20 cm, rhizome brown and covered with long linear, reddish brown scales.Fronds bipinnate, 3–20 × 1–2.5 cm, glabrous, green; stipe dark brown-black 1.5–11.5cm with a few scales. Pinnae 0.5–3 × 0.3–1 cm, stalk sessile, 1–3 mm long at end of thefronds. Pinnule 0.5–2 mm broad, without midrib. Sori many; pseudoindusium broad,irregularly lobed, ciliate.Habitat: Crevices in limestone rocks, sea level up to 500 m.Distribution in Turkey:Antalya: Antalya-Burdur yolu, 33. km, 12.iv.1965, A. Baytop & T. Baytop 7926! ĢarmaDağ, 700ft., 17.vi.1958, E.R.B.Little! Aspendos Amphitheatre near Serik, c.50m.02.viii.1975, C.R.Fraser Jenkins 4573. BM! Kemer Küçükburun, kasabanın doğusundakiküçük yarımada, P. brutia ormanı, 20 m, 21.iii.1949, H. Peşmen 4150 & A. Güner HUB36705! Kemer, Kesmeboğaz, kalker kayalığı, 300 m, 25.iii.1978, H Peşmen & B. YıldızHUB 36707! Kaş, Kekova, Alan-Kale Mahallesi, kalker kaya çatlağı, 10-20 m,14.ii.2009, Z. Aytaç 9286 & H. Duman (GAZI)! Alanya, Hacımemetli Köyü, Bağderesi,200 m, 21.iii.1993, M. Koyuncu 9595 & H. Duman (AEF)!Aydın: Söke Milet haraberleri, anfitiyatro duvarları üzerinde, 10.v.1965, Kayacık &Yaltırık ISTO 3545!Balıkesir: Ayvalık, Pınar Adası, 60 m, 10.v.1996, K. Alpınar ISTE 71797!Çorum: Alacahöyük Köyü, 01.vii.1995, U. Sönmez ISTE 68382!Denizli: Mt. above Pamukkale (Hierapolis) hot springs near Burdur, rock crevice, dryplace on vertical earth bank with C. persica and Selaginella denticulata, 1000-1150m,16.v.1973, Betty E.G. Molesworth Allen 8102. BM!Hatay: Castle of Payas, near Dörtyol, sea level, 03.iv.1965, M.J. Coode & B.M.G. Jones394! Kırıkhan to Hamam, c. 100 m, 06.v.1965, M.J. Coode & B.M.G. Jones 533!İçel: Işıktepe (Seydüşıh) Köyü üstü, maki, kalker anakaya, 150-500 m, 19.iv.1985, Ş.Yıldırımlı 7594 HUB 36723! Mersin, Anamur çevresi, 09.v.1972, M. Koyuncu HUB36697! Dibek Mağarası girişi, Cennet-Cehennem, 09.xi.1973, A. Baytop & E. TuzlacıISTE 27072!İzmir: Schuth hinter Friendhof von Ahmetbeli, ca. 100 m, 19.iii.1966, G. Bocquet 1523!Selçuk harabeleri, 09.iv.1971, A. Baytop & T. Baytop ISTE 19112!Konya: Bucakkışla, Dede Dağı, Kayalık, kurak yamaçlar, maki, ca.580 m, 25v.1979, M.Vural 1873 GAZI)!Muğla: ca. 4 km nw Muğla. Felsen in der Macchie, 03.iv.1974, K. Lewejohann (Tu-74-137) & F. Holtz! Dereköy, between Marmaris and Muğla, 10 m, 26.iii.1956, Davis 25395& O. Polunin! BM! Fethiye, 30 m, 27.iii.1956, Davis 25421 & O. Polunin! BM! Kişleto Kozağaç (Milas-Yatağan), 550 m, 13.iv.1965, Davis 41056! Marmaris, Söğüt to
BONA & GIBBY: THE FERN FAMILY PTERIDACEAE IN TURKEY 123

Bozburun, sea level–50 m, 15.iv.1965, Davis 41168! Dalyan-Sülüngür Gölü, Ortaca arası,100-200 m, 14.iv.1981, A Güner 8571 et al. (HUB)! Bodrum, Türkbükü-Bodrum, 200-500 m, 15.iii.1984, Ş. Yıldırımlı 5883 HUB 36696! Köyceğiz, Çandır Köyü, KaunosHarabeleri, duvar arası, 80 m, 08.xi.1991, A. Güner 10323 (GAZI)! Köyceğiz, YangıKöyü, Yangı Deresi, sarp ve derin vadi, kalkerli yamaçlar, 60-80 m, 24.v.1981, A. Güner8324, M. Vural & H. Şağban (GAZI)!General Distribution: From Mediterranean area to West Himalayas.Note: There has been confusion in the past between Allosorus acrosticus and A.pteridioides (Reichard) Christenh. (see Nardi & Reichstein, 1985). Allosorus acrosticusis a tetraploid taxon, whereas A. pteridioides is diploid. This latter has a more westernMediterranean distribution and, so far, has not been recorded for Turkey, although thereare records from Cyprus and Jordan in BM and from Syria and Palestine in E, and it mayhave been overlooked in Turkey. The two taxa have similar morphology but can bedistinguished; A. acrosticus has a pseudoindusium with ciliate margins, with scales onrachis and costae, whereas A. pteridioides has pseudoindusium entire, and rachis andcostae are ± glabrous.A. persicus (Bory) Christenh. Willdenowia 42(2): 284 (2012).Synonyms: Notholaena persica Bory in Bélanger & al., Voy. Indes Or.: 23 (1833);Cheilanthes szovitsii Fisch. & Mey. apud Hohen. in Bull. Soc. Nat. Mosc. 1838 No. 3:241 (1838); Cheilanthes persica (Bory) Kuhn in Bot. Zeitung (Berlin) 26: 234 (1868).Lectotype: [Iran] La Perse, parmi les rochers, 1821, C. Bélanger s.n. P00607931 (P)!Plant up to 20 cm, rhizomes covered densely in narrow dark brown-black scales. Frondsbipinnate, 3–16 × 1–4 cm, green and glabrous above and with brown scales on rachisand costae, eglandular; stipe 1–8 cm, covered with dark brown-black and narrowlanceolate or linear long brown hairs. Pinna (0.3–) 1–3 × 0.2–2 cm; pinna stalk very shortc. 1 mm, pinnules lobed, lobes lancolate. Sori many; pseudoindusium narrow, with manylong marginal hairs that cover the lower surface of the segment.Habitat: Shady crevices in limestone or metamorphic rocks, 200–2300 m.Distribution in Turkey:Adana: Feke, Sencan Dere between Belenköy and Suphandere, 1000 m, 02.vii.1952,Davis 19586A, Dodds, Çetik! Kozan, (Anti-taurus), 300 m, 12.iv.1957, Davis 26614,Hedge! Osmaniye-Yarpuz, 15 km e Osmaniye, 800 m, 29.iv.1986, M. Nydegger HUB36695!Ankara: mamak, Kıbrıs Köyü vadisi, Dipsiz gölü mevkii, kaya çatlağı, gölgeli yerler,1000-1100 m, 26.vi.2005, S. Arslan 2352 (GAZI)!Antalya: Kumluca, Sazak Koyu, kalker kayalığı, 40 m, 29.iv.1980, H. Peşmen 4876 HUB36704! Konyaaltı, 10 m, 23.v.1977, M. Koyuncu & M. Coşkun HUB 36703! Koslu Dere,n.of Bozburun Dağ, 1200 m, 27.vii.1949 Davis 15754.Aydın: 3 km westlich Ortaklar, Abangh am Ovacık Dag oberhalb der Strasse Selçuk-Ortaklar, 200 m, 22.v.1966, G. Bocquet 2472! Söke, piriyen, 400 m, 30.v.1972, N. Tanker,AEF 17!Denizli: Bozdağ above Geynam yaila, 16.vii.1947, Davis 13432!Erzincan: Kemaliye, Yaka Köyü, Sazak deresi, kazmofit, 850-1000 m, 18.xi.1980, Ş.Yıldırmlı 4186 HUB 36708!Isparta: Ş. Karaağaç, Kızıldağ Milli Parkı girişi, sedir ormanı, kalker, 1200-1300 m,17.xi.1993, B. Mutlu 163 HUB 36701! Eğridir, Yaka Köy, 2 km güneyi, derin kalkervadisi, P. nigra-J. excelsa ormanı, 1800 m, 20.v.1973, Peşmen 3401, Erik, PamukçuoğluHUB 36709! Eğridir, Anamas Köyü, Kapız Deresi, kalkerli sarp ve derin vadi, 1250-
124 FERN GAZ. 20(3):119-132. 2016

1450 m, 05.viii.1974, H. Peşmen & A. Güner 1836 HUB 36706! Eğridir, Aksu,Karacahisar Köyü, Başak mevkii, kalkerli vadi, 1000 m, 03.vii.1974, H. Peşmen & A.Güner 1653 HUB 36702!Konya: Ereğli, Aydos Dağı, Delimahmutlu, Kapız, kayalık, 1600 m, 21.vi.1977, S. Erik2179 HUB 36718! Beyşehir, Kurucuova Köy, Kuzgun tepe, P. ningra ormanı, kalkerliK:B. yamaç kayalık arazi, 1300-1500 m, 24.iv.1974, H. Peşmen & A. Güner 1019 HUB36710!Mardin: Cudi Dağ above Hessana (d. Silopi), 900–1150 m, 11.v.1966, Davis 42862A!Savur, 900 m, 24.v.1957, Davis 28568, Hedge! Dargeçit, Çelik Köyü, Umutlu mezrası,kaya üzeri, 520 m, S. Arslan 3297 (GAZI)!Muğla: Kara Dağ, 750 m, 18.v.1988, P. Ainsworth T27/88! Ortaca, Dalyan-SülüngürGölü arası, 10 m, kalkerli arazi, makilik dik yamaçlar, 12.ii.1991, A. Güner 7999 et al.(HUB)! Fethiye, Babadağı, batı etekleri, gıdırak mevkii, 100 m, 10.v.1984, E. TuzlacıISTE 53054!Niğde: Side valley W. of Maden, 30.vii.1969, J. Darrah 349!Samsun: Vezirköprü at Kızılırmak, 400 m, 06.iv.1969, Tobey 2446!Şanlıurfa: Yenice Köyü üstü, Nemrut Dağları, kalker kayalığı tabakalar, taşlık, bozkır,475-675 m, 26.vi.1988, Ş. Yıldırımlı 10791a HUB 36700!Şırnak: Beytüşebap, Dağdibi Köyünden anayola, kalker kayalığı, 25.v.2004, A. Dönmez11916 (HUB)!Siirt: Botan Çay, by the bridge 19 km from Siirt, 400 m, 17.v.1966, Davis 43220!Tunceli: Ovacık, Munzur Dağları, Karagöl vadisi, 1300-1500 m, 06.v.1979, Ş. Yıldırımlı1388 & Ş. Kaplan HUB 36711!Yozgat: Sorgun to Çekerek, 1200 m, 27.v.1965, M.J.E. Coode & B.M.G. Jones 1598!General Distribution: Distributed in Europe, Asia, and Africa.A. tinaei (Tod.) Christenh. Willdenowia 42(2): 284 (2012). Synonyms: Cheilanthes tinaei Tod. in Giorn. Sci. Nat. Econ. Palermo 1: 217 (1866); C.duriensis Mendonça & Vasc., Anais Inst. Vinho Porto 15(4): 47 (1956); C. pteridioidessubsp. tinaei (Tod.) O. Bolòs & al., Fl. Man. Països Catalans 1213 (1990).Holotype: [Sicily] “Messina, Cheilanthes odora var. stipites glabrato” Id. 9541 (PAL).Isotype: Id. 9542 (PAL) (see Nardi, Rasbach & Reichstein, 1979).Plant up to 25 cm, rhizome covered with brown long scales. Fronds bipinnate 5–25 × 1–2.5 cm, shortly and sparingly pilose when young, later ± glabrate above, scattered2-5-celled red glandular hairs beneath; stipe densely covered with long scales at baseand with a few scattered scales further up when young then almost glabrous, brown-black, 1–10 cm. Pinnae 0.5–1.5 × 0.3–0.5 cm with very short stalks c. 1 mm. Pinnule0.5–2 mm broad. Pseudoindusium with rounded, entire membranous margin.Habitat: Coastal, bare rocks, sea level to 1250 m.Distribution in Turkey:Istanbul: Büyükada, Eskibağ, kayalıklar, 17.08.1977, H. Demiriz, B. Tütel, A. Aydın(ISTE 31028!)General Distribution: N.W. Africa, Portugal, S. Europe.
Cryptogramma R.Br., Narr. Journey Polar Sea [Franklin] 767 (1823).Rhizome creeping to ascending covered with light brown to black long scales. Frondsdimorphic, tripinnate; sori terminal on veins protected by recurved membranous lateralmargins of segments.C. bithynica S.Jess., L.Lehm. & Bujnoch, Fern Gaz. 19(2): 52 (2012).
BONA & GIBBY: THE FERN FAMILY PTERIDACEAE IN TURKEY 125

Holotype: Northwestern Turkey: Uludağ southeastern of Bursa: silicate scree near theroadway, NNE of the summit of Uludağtepe, 1770 m, 09.ix.2009, leg. S. Jesen & L.Lehmann (SJ-3820), JE. Isotype: 09.ix.2009, leg. S. Jesen & L. Lehmann (SJ-3820),ANK!Plant 10(–22) cm, - covered with light brown long scales. Fronds tripinnate, green,reddish at the end of the fronds, glabrous above, veins visible at and of the lobes in white-yellow colour. Stipe straw-coloured to greenish, glabrous to sparsely covered with lightbrown scales. Fertile fronds 8–10 × 3–4 cm; stipe 6–7 cm. Pinnae 2 × 1–2 cm; pinnastalks 1–3 mm. Pinnules linear-lobed, convolute at the edge of lobes. Vegetative frondsslightly thick and relatively tough, 8–10(–22) × 3–4.5 cm, broadly deltate; stipe 5–6 cm.Pinnae ovate to orbicular, 1.5–2.5 × 1–2.5 cm; pinna stalks 1–3 mm. Pinnules lanceolate-lobed. Sori terminal on veins protected by recurved margins of lobes.Habitat: Silicate scree near the cycle roadway, 1700–2300 m.Distribution in Turkey:Northwestern Turkey: Uludağ southeastern of Bursa: silicate scree near the roadway,NNE of the summit of Uludağtepe, 1770 m, 09.ix.2009, leg. S. Jesen & L. Lehmann (SJ-3820), JE. Isotype: 09.ix.2009, leg. S. Jesen & L. Lehmann (SJ-3820)(ANK)!General Distribution: Endemic to Turkey.C. crispa (L.) R.Br. ex Hook., Gen. Fil. pl. 115B (1842).Synonyms: Osmunda crispa L., Sp. Pl. 1067 (1753); Allosorus crispus (L.) Bernh. inSchrad Jour. 1(2): 36 (1800).Type: Habitat in Anglia, Helvetia, Herb. Sloane 85: 128 (BM-SL).Plant to 30 cm, rhizome covered with brown-black long scales. Fronds thin, tripinnate,green, reddish at the end of the fronds, glabrous above, veins visible at and of the lobesin white-yellow colour. Stipe straw-coloured to greenish, glabrous. Fertile fronds 15–30× 2.5–5 cm; pinna stalks 8–20 cm. Pinnae 2–5 × 1–1.5 cm; stipe 1–6 mm. Pinnules linear-lobed, convolute at the edge of lobes. Vegetative fronds ovate to deltoid–ovate, 15–20 ×3–4 cm; pinna stalks 4–11 cm. Pinnae narrowly ovate, 1.5–3 × 0.7–1.8 cm; pinna stalks1–4 mm. Pinnules lanceolate-lobed. Sori terminal on veins protected by recurved marginsof lobes.Habitat: Screes, 200–2400 m.Distribution in Turkey:Artvin: Arhavi, above Dikyamaç village, 2200 m, 26.vii.1974, M. Çoşkun AEF 19!Çoruh, Şaval Tepe above Murgul (igneus), 2000 m, 13.viii.1957 Davis 32393 & Hedge!Şavşat, Kocabey Kışlası, Camız patlatan mevkii, kayalık alanlar, 1900-1950 m,14.v.2004, H. Altınözlü 3945 (HUB)! Kaçkal Dağı, Pırnallı Üst Yaylası, CehennemDeresi Mevkii, 1750-3100 m, 12.vii.1984, Demirkuş 2641b HUB 36758! Arhavi,Dikyamaç Köyü yaylası, Arpaçay civarı, 2200 m, M. Coşkun AEF!Rize: Kaçkar Mountains, east of Elevit, north of road beyond village, 2365 m, 27.ix.2008!Çamlıhemşin, Amlakıt Yaylası, çayırlık, 1350 m, 10.viii.1984, A. Güner 6117 & M. VuralHUB 36755! Çamlıhemşin, Amlakıt Yaylası, Kayguttan Çovinovit yoluyla yayla, kalkerliarazi, alpinik step, 2200-2720 m, 19.vii.1974, A. Güner 1087 HUB 36753! Çamlıhemşin,Yukarı Amlakıt Yaylası-Kışyatağı arası, Rhododendron caucasicum çalılığı, granitanakaya, 2200-2350 m, 20.ix.1979, A. Güner 2190 HUB 36754! General Distribution: Europe and Caucasus.
Cosentinia Tod. Giorn. Sci. Nat. Econ. Palermo 1: 219 (1866).Rhizomes short-creeping to erect with long linear brown scales. Fronds pinnate, both
126 FERN GAZ. 20(3):119-132. 2016

surfaces of the frond covered, densely below, with long whitish multicellular hairs.C. vellea (Aiton) Tod. Giorn. Sci. Nat. Econ. Palermo 1: 220 (1866).Synonyms: Acrostichum velleum Aiton Hort. Kew. 3: 457 (1789). Notholaena vellea(Aiton) Desv. in J. Bot. Appl., 1:92 (1813) non R. Br. (1810); N. lanuginosa (Desf.) Desv.ex Poir., Fragenta Phytographiae Australiae 5 (1865); Cheilanthes vellea (Aiton) F. Muell.Domin Bibl. Bot. 85. 133 nota. (1913); Acrostichum catanense Cosent., Atti Accad.Gioenia Sci. Nat. Catania 1/2: 207 (1827); Cheilanthes catanensis (Cosent.) H.P.Fuchs,Brit. Fern Gaz. 9(2): 45 (1961). Lectotype: Portugal, 1777, F. Masson s.n. BM000829683 (BM)!Plant 10–20 cm, rhizomes brown with long linear pale brown scales. Fronds pinnate, 8–16 × 1.5–4 cm, green above and brownish below, both surfaces of the frond covered,densely below, with tangled long whitish multicellular hairs; stipe brown, 1–4.5 cm,densely covered with hairs when young, becoming glabrous when mature, young frondsalmost sessile, lobed, 0.5–2 × 0.3–0.8 cm. Sori not discrete, sporangia spread along veinsfrom leaf margin, ± protected by recurved but not modified leaf-margin.Habitat: Dry limestone, calcareous rocks, sea level to 300 m.Distribution in Turkey:Ankara: Beynam, Karaçam ormanı, 1300-1500 m, 15.v.1986, Ş. Yıldırımlı 9221 HUB36713!Antalya: Kumluca, Adrasan Köyü kuzeyi, serpantin arazi, 300 m, 29.ii.1980, H. Peşmen4791 & A Güner HUB 36721!Aydın: Priene, 100 m, 09.iv.1965, Davis 40788! Milet harabeleri, R. Lampinen 7441ISTE 63662!Bolu: Eğerci’den Yedigöller’e 4. km, Yağmurca Köyü ilerisi, 193 m, 03.xi.2001, A.Dönmez 10387 & S. Işık (HUB)! Kıbrıscık, Kıbrıscık’tan Beypazarı’na 5 km, Paliurusspina-christi açıklığı, 1110 m, 08.viii.2010, A. Dönmez 17616 & Z. Uğurlu (HUB)!Çankırı: Eldivan, Seydiköy üstü, step, 1300-1350 m, 09.viii.1984, Ş. Yıldırımlı 7409HUB 36716!Hatay: İskenderun, Musa Dağı, Kale ilerisi, P. brutia ormanı, 200 m, 30.iii.1995, A.Dönmez 4423 (HUB)!Isparta: Ş.Karaağaç, Kızıldağ Milli Parkı girişi, Sedir ormanı açıklığı, kalker, 1200-1300m, 18.xi.1993, B. Mutlu 180!İçel: Anamur’un üstü, Kayalıklarda 200 m, 06.iii.1977, M. Koyuncu HUB 36722!Kastamonu: Hanönü, Karapınar Köyü’nden, Derencik Köyü’ne gidiş, köprü çevresi,çakıllı yamaçlar, 26.ii.2010, A. Dönmez 16603 & Z. Uğurlu (HUB)!Kayseri: Yılanlı Dağ, 1200 m, 12.vi.1975, M. Coşkun & N. Çelik HUB 36719!Muğla: Marmaris-Emecik, 5 m, 25.iii.1956, Davis 25349 & O. Polunin! Marmaris,Hisarönü to Turgut, 20 m, 15.iv.1965, Davis 41147! Ören, 50–100 m, 10.iv.1965, Davis40855! Köyceğiz, Yargı Köyü, Yargı Deresi, 60-80 m, sarp ve derin vadi, kalkerli kayalıkyamaçlar, kazmofit, 24.v.1981, A. Güner 9322, M. Vural & H. Şağban HUB 36720!Köyceğiz, Sultaniye, Kersele Deresi, dere yatağı, metamorfik arazi, 15-30 m, 21.v.1991,A. Güner 9195, M. Vural & H. Şağban HUB 36715!General distribution: Macaronesia, Mediterranean region including S. Europe and NorthAfrica and east to Turkey, Egypt, Yemen and Oman.Note: Cosentinia vellea has two subspecies, a tetraploid C. vellea subsp. vellea and adiploid C. vellea subsp. bivalens (Reichstein) Rivas Mart. & Salvo. Onmacromorphological characters the two are indistinguishable, but can be separated bycomparison of spore size and stomatal length (see Haou et al., 2011). Of four Turkish
BONA & GIBBY: THE FERN FAMILY PTERIDACEAE IN TURKEY 127

specimens for which spores have been measured, three correspond with subsp. vellea[64-(67)-71µm: Davis 25349,40855, 41147] and one with subsp. bivalens [55-(57)-62µm: Davis 40788], although cytological verification is still lacking.
Paragymnopteris K.H. Shing, Indian Fern J. 10(1–2): 227 (1994) [1993].Rhizome short-creeping, covered with long, linear scales. Fronds pinnate, linear-lanceolate scales along the midrib above and covered with lanceolate brown scales below.P. marantae (L.) K.H. Shing, Indian Fern J. 10(1–2): 229 (1994) [1993]. Synonyms: Acrostichum marantae L., Sp. Pl. 2: 1071 (1753). Ceterach marantae (L.)DC., Fl. Franc. ed. 3. 2: 567 (1805); Cheilanthes marantae (L.) R.Br., Prodr. Fl. Nov.Holland. 145 (1810); Notholaena marantae (L.) R.Br., Prod. Fl. N. Holl. 146 (1810);Cincinalis maranthae (L.) Desv., Berl. Mag. 5. 312 (1811); Gymnogramma marantae(L.) Mett., Fil. Hort. Bot. Lips. 43 (1856); Gymnopteris marantae (L.) Ching, ActaPhytotax. Sin. 10: 303 (1965); Paraceterach marantae (L.) R.M.Tryon, Amer. Fern J.76(4): 186 (1986).Lectotype: Pierre Magnol s.n. Herb. Linn. 1245.12 (LINN)!Plant 10–30 cm. Rhizome covered with long, linear light brown scales. Fronds pinnate-pinnatisect, 10–30 × 1.2–4.5 cm, linear-lanceolate scales only along the midrib above,lower frond surface densely covered with lanceolate brown scales; stipe 4–20 cm, withscattered brown scales. Pinnae 1–2.5 × 0.5–1.2 cm, 3–7 lanceolate or broad triangularlobed, main lobe almost lyre-shaped, edge of pinnae undulate or simple; pinna stalks 1–2 mm or sessile. Sori not discrete, sporangia spread along veins from leaf margin, ±protected by slightly recurved and modified leaf-margin.Habitat: Serpentine rocky slopes, sea level to 1400 m.Distribution in Turkey:Adana: Karsantı, Kumbükü, Hızar yolu, ca. 880 m, 05.v.1972, E Yurdakul ANK 1768!Feke: Sencan Dere between Belankey and Suphandere, metamorphic rocks, 100 m,02.vii.1952, Davis 19586B, Dodds, Cetik! Bahçe (N. Amanus), Dumanli Dağ aboveHaruniye, shady metamorphic rocks, 1200 m, 19.iv.1957, Davis 26845, Hedge!Ardahan: In Felsritzen etwa 10-15 km östlich von Kosor zw. Göle und Olta, 22.iv.1957,E. Sauer 306/57!Artvin: Yusufeli, Altıparmak Köyü üzeri, P. sylvestris-P. orientalis ormanı, 1250 m,20.v.1983, A. Güner 4829 & M. Vural (GAZI)!Erzincan: Erzincan’ın 30 km batısında serpantin boğazı, ca. 1500 m, 18.vii.1958, H.Birand ANK 10909! Keşiş dağ above Cimin, among igneous rocks, 2700 m, 28.vii.1957,Davis 31652, Hedge!Giresun: Şebinkarahisar, Arslanyurdu Deresi, ca. 1380-1460 m, 28.viii.1976, K.P. ButtlerANK 21819!Hatay: Hassa to Dörtyol, mountain road, slopes above Dörtyol, 1100 m, A.J. Byfield &D. Pearman B 2620 ISTE 73362!İçel: Mersin, Fındıkpınar, Demirışık, P. brutia altı, nemli yerler, 24.vi.1971, T. Uslu ANK526!Kahramanmaraş: Andırın, Elma Dağı, orman açıklığı, 1550 m, 22.vi.1979, E. Tuzlacı &M. Saraçoğlu ISTE 42386!Kastamonu: Tosya, Sekiler Köyü, Seki Deresi, kayaların arası, ca. 1100 m, 28.ix.1977,M. Kılınç ANK 6866!Konya: 2 km from Çamlık village, Akseki-Beyşehir area, 1360–1370 m, 03.vi.2003, R.D.Reeves 2693 & N. Adıgüzel!
128 FERN GAZ. 20(3):119-132. 2016

Muğla: Marmaris, 50 m, 13.iv.1965, Davis 41021! Marmaris, Pinetum brutiae, onserpentine, shady rocks, 30 m, 24.iii.1956. Davis 25,255, O.Polunin! NordwestlichKöyceğiz oberhalb Ağla, Felsspalte im Kiefernforst, 950 m, 30.vi.1973, F. Holtz 428, P.Hanel, T. Kesercioğlu! Köyceğiz, Sultaniye, Kersele Deresi, dere yatağı, metamorfikarazi, 5-30 m, 21.v.1991, A. Güner 9195, M. Vural, H. Şağban HUB 36715! SandrasDağı, Çiçekbaba Tepesinin güney yamaçları, 1700 m, 14.vii.1978, N. Özhatay & E.Özhatay ISTE40635!Samsun: Ladik Istasyon, Karadağ, 1050 m, 08.v.1965, Tobey 947! Vezirköprü atKızılırmak, 400 m, 20.iv.1969, Tobey 2451!Zonguldak: Dirgine to Devrek, cliff edges (serpentine), 180m, 18.vii.1962. Davis 37906,Coode, Yaltirik,!General Distribution: Mediterranean area, northern Austria and Caucasus. C. & S.Europe, Macaronesia, N. Africa.
Pteris L., Sp. Pl. 2: 1073 (1753).Fronds pinnate to bipinnate, green and glabrous, midrib and secondary veins distinct.P. cretica L., Mant. Pl. 130 (1767). Synonyms: none.Lectotype: Pietro Arduino s.n. Herb. Linn. 1246.7 (LINN)!Plant 30–70 cm. Rhizomes short-creeping dark brown with dark brown scales. Frondspinnate to bipinnate, green and glabrous, midrib and secondary veins distinct. Pinnae ≤7 pairs, often with a long basal segment. Fertile fronds serrate only at the end, convoluteat the edge of lobes, 40–70 × 15–20 cm; stipe 20–40 cm, pinnae 10–15 cm and sessileor with c. 1 mm stalk. Vegetative fronds serrate at margin, 30–70 × 10–20 cm; stipeglabrous, 20–35 cm. Pinnae 5–20 cm, sessile or with 1–3 mm stalk. Sori submarginal,in a continuous line protected by scarious recurved leaf-margin.Habitat: Shady banks and shady damp places in deciduous forests, sea level to 800 m.Distribution in Turkey:Artvin: 10 km south of Hopa on road to Artvin, 120 m, 25, 25.ix.2005, B. Güner et al.,TUDI 46! Arhavi, Dikyamaç village, 800 m, 15.vii.1994, M. Coşkun AEF 20! Çoruh, 1km südwestlich von Hopa neben Küstenstrabe steiler, 05.viii.1973, F. Holtz (01.056), P.Hanel & T. Kesercioğlu! Çoruh, Borçka, Hopa, 300 m, 21.vi.1957, Davis & Hedge29832! Çoruh, deciduous forest, 2 ft. 6 ins. high, 18.viii.1966, Tong 496! Çoruh, Hopa,near sea level, 22.vi.1957, Davis & Hedge 29876! Hopa, Sarp Köyü, 5 m, 11.iv.1982, Ş.Yıldırımlı 4414 HUB 36746! Hopa, Kemalpaşa-Sarp arası denize dik kayalıklar, 0-50m, 19.v.1985, Ş. Yıldırımlı 7895 HUB 36751! Hopa, orman altı, 0-50 m, 25.01.1992, D.Osmanoğlu & Z. Aytaç 4473 (GAZI)! Murgul, Meratit Köyü, 02.viii.1959, R. Çetik &Davis ANK 20500!Giresun: Denize yakın çalılık ve ıslak yerlerde, 07.vii.1958, H. Birand, A 10692a!Gengene Köyü, Fagus ormanı, 24.vi.1977, Y. Akman ANK 6977!Ordu: Fatsa, rocks above Kayalar Dere, 60 m, 23.iv.1963, C. Tobey 46!Rize: 2 km south east of Rize, 100 m, 04.viii.1974, J.R. Akeroyd 173! Shady banks, 100–200 m, 21.viii.1952, Davis 20785 & Dodds! Çamlıhemşin, Aşağı Vice Mahallesi üstü,karışık orman ve nemli çayırlık, 500-600 m, 13.vii.1979, A. Güner 2095 HUB 36750!Çamlıhemşin, Boğazıçik Köyü, Pogina çevresi, karışık orman, 650 m, 04.ix.1978, A.Güner 1820 HUB 36748! Alipaşa Köyü civarı, 29.vii.1956, K. Karaosmanoğlu HUB36749!Trabzon: Valley above Çaykara, 700 m, 05.viii.1957, Davis & Hedge 32107! Sürmene,
BONA & GIBBY: THE FERN FAMILY PTERIDACEAE IN TURKEY 129

Gökçesu Mahallesi, İdrum tepe çevresi, 350-550 m, 04.vii.1997, H. Duman 6464(GAZI)!General Distribution: S. Europe, W. Syria, Caucasus, subtropical Asia, E. and S. Africa,N. America, Hawaii.P. vittata L., Sp. Pl. 1074 (1753).Synonyms: P. ensifolia Poir. in Lam, Encycl. 5: 711 (1804); P. diversifolia Sw. Syn. Fil.96: 288 (1806); P. costata Bory, Sp. Pl. 5: 367 (1810); P. inaequilateralis Poir. Encycl.4: 601 (1816); Pycnodoria vittata (L.) Small, Ferns Florida 89 (1931).Lectotype: Herb. Linn. 1246.3 (LINN)!Plant 10–70 cm. Rhizomes short-creeping, brown with fine long scales. Fronds pinnate,10–70 × 5–20 cm, green, long thin scales along with midrib; stipe 1.5–10 cm, yellowishbrown with thin long scales. Pinnae (2–)6–10 × 0.3–1 cm, ≥10 pairs, longer at apex offrond, reducing in length towards frond base, narrowly lanceolate, undivided, serrate,almost sessile. Sori submarginal, in a continuous line protected by scarious recurvedleaf-margin.Habitat: Shady rocks or serpentine.Distribution in Turkey:Antalya: Kemer-Ulupınar arası, 200 m, 08.vii.1979, H. Peşmen 4380 & A. Güner HUB36744! Kemer, Götnük Deresi, Kalker vadisi, 50-200 m, 22.iii.1979, H. Peşmen 4199 &A. Güner HUB 36743! Alanya, P. brutia altı, c. 300 m, 04.viii.1982, Ş. Yıldırımlı 4452HUB 36741! Düden Şelalesi, 23.iv.1976, N. Kızılgül AEF 5714!Hatay: Valley above Dörtyol, S. bank, N. facing, 150 m, 12.viii.1969, J. Darrah 534,634, 620! Antakya, Harbiye, şelale civarı, 180 m, M. Coşkun 321, B. Şener & F. Ilisulu!Mersin: Silifke-Anamur arasında Değirmenönü, köy değirmeni yakını, 50 m, 09.ix.1962,H. Kayacık 2958!General Distribution: Europe and Asia.
CONCLUSIONIn the first volume of the Flora of Turkey (Henderson, 1965) the family currentlyrecognised as Pteridaceae was represented by nine species from five genera and classifiedin five different families. Since this publication, further new records of Cheilanthes tinaei(Davis, 1988) and Cryptogramma bithynica (Jessen et al., 2012), have been published.In this revision the single family Pteridaceae in Turkey includes 11 species from sevendifferent genera. Families Adiantaceae, Cryptogrammaceae, Gymnogrammaceae andSinopteridaceae have been combined in the family Pteridaceae (see Christenhusz et al.,2011). The names of the genera Anogramma, Adiantum, Cryptogramma and Pteris havenot changed since 1965. However, Turkish Cheilanthes species are now placed in threedifferent genera: Allosorus, Cosentinia and Paragymnopteris. Cheilanthes marantae isa synonym of Paragymnopteris marantae and Cheilanthes catanensis is a synonym ofCosentinia vellea, whilst Cheilanthes persica, Cheilanthes acrostica and Cheilanthestinaei are now placed within Allosorus (Christenhusz et al., 2011; Greuter & Raab-Straube, 2012).
ACKNOWLEDGEMENTSWe are grateful to Prof. Adil Güner for his help in accessing resources on Pteridaceae,to Prof. Gönül Kaynak who sent images of Allosorus tinaei and to Miss Alison Paul andDr Fred Rumsey for reviewing the manuscript. This study was supported financially bythe Edinburgh Botanic Garden (Sibbald) Trust to M.B.
130 FERN GAZ. 20(3):119-132. 2016

REFERENCESABOTSI, K. E., RADJI, A. R., ROUHAN, G., DUBUISSON, J. Y. & KOKOU, K. 2015.
The Pteridaceae family diversity in Togo. Biodivers. Data J. 3: e5078. BENLIOĞLU, O., KAYNAK, G. & TARIMCILAR, G. 1997. Ferns of the Black Sea
region of Turkey: chorological and ecological studies. Fern Gaz. 15(5): 169-192.CHRISTENHUSZ, M.M., ZHANG, X. & SCHNEIDER, H. 2011. A linear sequence of
extant families and genera of lycophytes and ferns. Phytotaxa 19: 7-54.DAVIS, P.H. 1988. Flora of Turkey and The East Aegean Islands, vol. 10. Edinburgh
University Press, Edinburgh.GASTONY, G. & ROLLO, D. 1995. Phylogeny and generic circumscriptions of
Cheilanthoid ferns (Pteridaceae: Cheilanthoideae) inferred from rbcL nucleotidesequences. Am. Fern J. 85(4): 341-360.
GREUTER, W. & RAAB-STRAUBE, E.V. 2012. Euro+ Med Notulae, 6. Willdenowia,42: 283-285.
GRUSZ, A.L. & WINDHAM, M.D. 2013. Toward a monophyletic Cheilanthes: Theresurrection and recircumscription of Myriopteris (Pteridaceae). PhytoKeys 32:49-64.
GÜNER, A., ASLAN, S., EKIM, T., VURAL, M. & BABAÇ, M.T. 2012. TürkiyeBitkileri Listesi (Damarlı Bitkiler). Nezahat Gökyiğit Botanik Bahçesi ve FloraAraştırmaları Derneği Yayını.
GÜNER, A. & EKIM, T. 2014. Resimli Türkiye Florası, Vol. 1. Ali Nihat Gökyiğit Vakfı,Flora Araştırmaları Derneği and Türkiye İş Bankası Kültür Yayınları, İstanbul.
HAOU, S., DE BÉLAIR, G. & VIANE, R.L.L. 2011. Inventory of the ferns (filicopsida)of Numidia’s (North-Eastern Algeria). Int. J. Biodiv. Cons. 3(6): pp. 206-223.
HENDERSON, M.D. 1965. Filicales. In: Davis, P.H. (Ed.), Flora of Turkey and the EastAegean Islands, Vol. 1, pp. 38-63. Edinburgh: Edinburgh University Press.
JESSEN, S., LEHMANN, L. & BUJNOCH, W. 2012. Cryptogramma bithynica spec.nov. (Pteridaceae, Pteridophyta) - A new fern species from NorthwesternAnatolia/Turkey. Fern Gaz. 19(2) 47-54.
KAMAU, P.W. 2007. Systematic Revision of Pteris L. in Tropical Africa and Ecology ofFerns and Lycophytes in Lowland Tropical Rainforests. PhD Thesis. UniversitätKoblenz-Landau, Germany.
KAYNAK, G., TARIMCILAR, G., DAŞKIN, R. & YILMAZ, Ö. 2008. Chorologicaland Ecological investigations on Cheilanthoid ferns in Turkey. J. Env. Biol. 2(6)83-88.
NARDI, E., RASBACH, H. & REICHSTEIN, T. 1979. Cheilanthes tinaei Tod., an earliername for C. corsica Reichstein et Vida and related species in Sicily. Webbia, 33:449-456.
NARDI, E. & REICHSTEIN, T. 1985. Nomenclatural notes on Cheilanthes pteriodioides(Reichard) C. Chr. (Sinopteridaceae, Pteridophyta). Webbia 39(1): 135-139.
SANCHEZ-BARACALDO, P. 2004. Phylogenetics and biogeography of the neotropicalfern genera Jamesonia and Eriosorus (Pteridaceae). Am. J. Bot. 91(2): 274-284.
SCHNEIDER, H., HE, L., HENNEQUIN, S. & ZHANG, X. 2013. Towards a naturalclassification of Pteridaceae: inferring the relationships of enigmatic pteridoid fernspecies occurring in the Sino-Himalaya and Afro-Madagascar. Phytotaxa 77(4):49-60.
SCHUETTPELZ, E., SCHNEIDER, H., HUIET, L., WINDHAM, M. & PRYER, K.2007. A molecular phylogeny of the fern family Pteridaceae: Assessing overall
BONA & GIBBY: THE FERN FAMILY PTERIDACEAE IN TURKEY 131

relationships and the affinities of previously unsampled genera. Mol. Phyl. Evol.44(3): 1172-1185.
SMITH, A.R., PRYER, K.M., SCHUETTPELZ, E., KORALL, P., SCHNEIDER, H. &WOLF, P.G. 2006. A classification for extant ferns. Taxon 55(3) 705-731.
WINDHAM, M.D., HUIET, L., SCHUETTPELZ, E., GRUSZ, A.L., ROTHFELS, C.,BECK, J., YATSKIEVYCH, G. & PRYER, K.M. 2009. Using plastid and nuclearDNA sequences to redraw generic boundaries and demystify species complexes incheilanthoid ferns. Am. Fern J. 99: 128-132.
ZHANG, G., ZHANG, X. & CHEN, Z. 2005. Phylogeny of cryptogrammoid ferns andrelated taxa based on rbcL sequences. Nord. J. Bot. 23: 485-493.
132 FERN GAZ. 20(3):119-132. 2016

THE PUBLICATIONS OF E. “BERT” HENNIPMAN1
HENNIPMAN, E. 1966. Pteridoblechnum, a new genus of blechnoid ferns fromAustralia. Blumea Supplement, Leiden 13: 397-403.
HENNIPMAN, E. & TOUW, A. 1966. Report on the Thai-Dutch Botanical Expedition1965/1966. Nat. Hist. Bull. Siam Soc., 21: 269-281.
HENNIPMAN, E. 1967. Some notes on Dutch Cladoniae (Lichenes). Persoonia 4: 427-428.
HENNIPMAN, E. & WIEFFERING, J. 1967. Ophioglossaceae. Dansk Botanisk Arkiv25: 68-70.
HENNIPMAN, E. 1968. Cladonia incrassata Floerke, een nieuw licheen voor ons land.Gorteria 4: 28-29 (in Dutch).
HENNIPMAN, E. 1968. The mucilage secreting hairs on the young fronds of someleptosporangiate ferns. Blumea 16: 97–103.
HENNIPMAN, E. 1968. A new Todea from New Guinea with remarks on the genericlimitation in recent Osmundaceae (Filices). Blumea 16: 105–108.
HENNIPMAN, E. 1969. De Nederlandse Cladonia's. KNNV (in Dutch).HENNIPMAN, E. 1970. Electron & light microscopical observations on the perine of
the spores of some Bolbitis species (Filices). Acta Bot. Neerl. 19: 671-680.HENNIPMAN, E. 1970. Some novelties in Bolbitis from Asia & the Pacific (Filices).
Blumea 18: 147-149.JONCHEERE, G.J.DE & HENNIPMAN, E. 1970. Two new species of Platycerium &
the identification of P. grande (Fée) Presl. Br. Fern Gaz. 10: 113–118.HENNIPMAN, E. 1974. The lectotypification of Acrostichum acuminatum Willd. & the
reinstatement of the name Photinopteris speciosa (Bl.) Presl. Blumea 22: 51-52.HENNIPMAN, E. 1975. New combinations in American Bolbitis species. Am. Fern J.
65: 30.HENNIPMAN, E. 1975. A re-definition of the gymnogrammoid genus Austrogramme
Fournier. Fern Gaz. 11: 61–72.HENNIPMAN, E. 1976. A second species of Pteridoblechnum (Blechnaceae) formerly
referred to Leptochilus acuminatus C.T.White & Goy. Proc. Roy. Soc. Queensl. 87:95–98.
HENNIPMAN, E. 1977. A monograph of the fern genus Bolbitis (Lomariopsidaceae).Leiden Bot. Ser. 2: 1–331.
HENNIPMAN, E. 1978. Bolbitis. In: HOLTTUM, R.E. Lomariopsis-group. FloraMalesiana II,1 4. The Hague Boston, London, M. Nijhoff / W. Junk.
HENNIPMAN, E., SIPMAN, H. J. M. & VIJSMA, E. 1978. De Nederlandse Cladonia'sLichenes. 2nd Ed., KNNV in Dutch.
SEN, U. & HENNIPMAN, E. 1981. Structure & ontogeny of stomata in Polypodiaceae.Blumea 27: 175-201.
HENNIPMAN, E. & ROOS, M.C. 1982. A monograph of the fern genus Platycerium(Polypodiaceae). Amsterdam, Oxford, New York, North-Holland PublishingCompany.
HENNIPMAN, E. & ROOS, M.C. 1983. Phylogenetic systematics of thePolypodiaceae (Filicales). Verh. Naturwiss. Ver. Hamburg 26: 321–342.
1 An obituary for Bert Hennipman by Peter Hovenkamp was published in the BritishPteridological Society Bulletin, Volume 8(1), 2014.
FERN GAZ. 20(3). 2016 133

HENNIPMAN, E. & HETTERSCHEID, W.L.A. 1984. Emendation of the fern genusChristiopteris, including the transference of two taxa to the microsorioidPolypodiaceae. Bot. Jahrb. Syst. 105: 1-10.
HETTERSCHEID, W.L.A. & HENNIPMAN, E. 1985. Venation patterns, soralcharacteristics, and shape of the fronds of the Microsoroid Polypodiaceae.Bot.Jahrb.Syst. 105: 11-47.
UFFELEN, G.A.VAN & HENNIPMAN, E. 1985. The spores of Pyrrosia Mirbel(Polypodiaceae), a SEM study. Pollen et spores 27: 155–198.
HENNIPMAN, E. 1986. Notes on the ant-ferns of Lecanopteris sensu stricto inSulawesi, with the description of two new species. Kew Bull. 41: 781–788.
RAVENSBERG, W.J. & HENNIPMAN, E. 1986. The Pyrrosia species formerlyreferred to Drymoglossum and Saxiglossum. Leiden Bot. Ser.. Leiden, Brill. 9:281-310.
HOVENKAMP, P., GITTENBERGER, E., HENNIPMAN, E., DE JONG, R., ROOS,M.C., SLUYS, R. & ZANDEE, M. (Eds). 1987. Systematics and evolution: a matterof diversity. Utrecht, Utrecht University, 341 pp.
HENNIPMAN, E. & VERDUYN, G.P. 1987. A taxonomic revision of the genusLecanopteris Polypodiaceae in Sulawesi, Indonesia. Blumea 32: 313–319.
BAAYEN, R.P. & HENNIPMAN, E. 1987. The paraphyses of the PolypodiaceaeFilicales. I. General part. Beitr. Biol. Pfl. 62: 251–315.
BAAYEN, R. & HENNIPMAN, E. 1988. The paraphyses of the PolypodiaceaeFilicales. II. Taxonomic part. Beitr. Biol. Pfl. 62: 317–347.
HENNIPMAN, E. 1990. The significance of the SEM for character analysis of sporesof Polypodiaceae (Filicales). In: CLAUGHER, D. (Ed.). Scanning ElectronMicroscopy in Taxonomy and Functional Morphology. Syst. Ass. Spec. Oxford,Clarendon Press. 41: 23-44.
HENNIPMAN, E., VELDHOEN, P., KRAMER, K.U. & PRICE M.G. 1990.Polypodiaceae. In: KRAMER, K. U. & GREEN, P.S. (Eds). The Families andGenera of Vascular plants. 1. Pteridophytes and Gymnosperms. Berlin etc.,Springer: 203-214.
KRAMER, K.U., CHAMBERS, T.C. & HENNIPMAN, E. 1990. Blechnaceae. In:KRAMER, K.U. & GREEN, P.S. (Eds). The Families and Genera of Vascularplants. 1. Pteridophytes and Gymnosperms. Berlin etc., Springer: 60–68.
HENNIPMAN, E. 1996. Scientific consensus classification of Pteridophyta. In:CAMUS, J.M., GIBBY, M. & JOHNS, R.J. (Ed.) Pteridology in perspective.Proceedings of the Holttum memorial pteridophyte symposium. Kew, 1995. RoyalBotanic Gardens, Kew. 730pp.
HENNIPMAN, E. & HOVENKAMP, P.H. 1998. Lecanopteris. In: HOVENKAMP, P.H. (Ed.) Polypodiaceae. Flora Malesiana II 3: 59-72. Leiden,Rijksherbarium/Hortus Botanicus.
HENNIPMAN, E. 2000. Sustainable exploitation in ornamental horticulture, anexample: Hippeastrum (Amaryllidaceae). Acta Hortic. 541, 67-73.
HAUFLER, C.H., GRAMMER, W.A. HENNIPMAN, E., RANKER, T.A., SMITH,A.R. & SCHNEIDER, H. 2003. Systematics of the ant-fern genus Lecanopteris(Polypodiaceae): Testing phylogenetic hypotheses with DNA sequences. Syst. Bot.282: 217-227.
134 FERN GAZ. 20(3). 2016

PhD Theses supervised
ROOS, M.C. 1985. Phylogenetic systematics of the Drynarioideae (Polypodiaceae).Utrecht, Universiteit Utrecht.
HOVENKAMP, P.H. 1986. A monograph of the fern genus Pyrrosia. Leiden BotanicalSeries 9. Brill, Leiden.
RÖDL-LINDER, G. 1990. A monograph of the fern genus Goniophlebium. Blumea342: 277-423.
BOSMAN, M.T.M. 1991. A monograph of the fern genus Microsorum. LeidenBotanical Series 14, Rijksherbarium/Hortus Botanicus.
UFFELEN, G.A. VAN 1993. Morphogenesis & evolution of the spore wall ofPolypodiaceae (Filicales). Leiden, Rijksherbarium/Hortus Botanicus.
COMPLIED BY P.H. HOVENKAMP
FERN GAZ. 20(3). 2016 135

THE PUBLICATIONS OF A.C. JERMY1
BARRY, D.H. & JERMY, A.C. 1953. Observations on Najas marina. Trans. Norf.Norw. Nat. Soc. 17: 294-297.
JERMY, A.C. 1959. A preliminary census list of British pteridophytes. Br. Fern Gaz.9(1): 4-15.
JERMY, A.C. 1960. A revised preliminary census list of British Pteridophytes Br. FernGaz. 9(1) (Suppl.): 1-12.
EVANS, G.B. & JERMY, A.C. 1962. Notes on the history of British Fern species I:confusion in the filmy ferns. Br. Fern Gaz. 9(3): 81-84.
JERMY, A.C. 1963. Name changes in the British fern list. Br. Fern Gaz. 9(4): 119. CRABBE, J.A., JERMY, A.C. & LOVIS, J.D. 1964. Asplenium L. In: TUTIN, T.G. et
al. (Eds) Flora Europaea 1: 14-17. Cambridge University Press.JERMY, A.C. 1964. Athyrium Roth, Cyclosorus Link, Diplazium Swartz,
Gymnocarpium Newman, Isoetes L., Thelypteris Schmidel. In: TUTIN, T.G. et al.(Eds) Flora Europaea 1: 5-6, 13-14, 18, 22. Cambridge University Press.
JERMY, A.C. & FUCHS, H.P. 1964. Cheilanthes Swartz. In: TUTIN, T.G. et al. (Eds)Flora Europaea 1: 10. Cambridge University Press.
WALKER, S. & JERMY, A.C. 1964. Dryopteris assimilis S.Walker in Britain. Br. FernGaz. 9(5): 137-140.
JERMY, A.C. 1964. Two species of Diplazium in Europe. Br. Fern Gaz. 9(5): 160-162. JERMY, A.C. & LOVIS, J.D. 1964. Asplenium majoricum Litardière. Br. Fern Gaz.
9(5): 163-167.PICHI-SERMOLLI, R.E.G. with BALLARD, F., HOLTTUM, R.E., ITÔ, H.,
JARRETT, F.M., JERMY, A.C., SCHELPE, E.A.C.L.E., TARDIEU-BLOT, M.-L.& TRYON, R.M. 1965. Index filicum: Supplementum quartum (pro annis1934-1960), xiv, 370 pp. Firenze.
CRABBE, J.A., JERMY, A.C. & MATTHEWS, G.A. 1965. Two forms of Equisetumvariegatum at Braunton Burrows. Proc. Bot. Soc. Br. Isl. 6: 40-42.
JERMY, A.C. 1965. Isoetes in DAVIS, P.H. (Ed.) Flora of Turkey and the East AegeanIslands, Vol. 1, pp. 37-38. Edinburgh University Press, Edinburgh.
JERMY, A.C. 1966. British Museum (Natural History) and University ofNewcastle-upon-Tyne Expedition to New Guinea, 1964-1965. London. 52 pp.
JERMY, A.C. 1967. [Fern] Abstracts . Br. Fern Gaz. 9(8): 362. JERMY, A.C. 1967. Carex section Carex (= Acutae Fr.). Proc. Bot. Soc. Br. Isl. 6(4):
375-379.JERMY, A.C., JONES, K. & COLDEN, C. 1967. Cytomorphological variation in
Selaginella. J. Linn. Soc., Bot. 60: 147-158. JERMY, A.C. & SAYERS, C.D. 1967. An expedition to the Finisterre mountains of
Eastern New Guinea. J. R. Hort. Soc. 92(3): 117-128. JERMY, A.C. 1968. Two new hybrids involving Dryopteris aemula. Br. Fern Gaz.
10(1): 9-12. JERMY, A.C. 1968. A new Selaginella from Zambia. Br. Fern Gaz. 10(1): 30-32. JERMY, A.C. & TUTIN, T.G. 1968. British sedges. Botanical Society of the British
Isles, London. 199 pp.
1 An obituary for Clive Jermy by Alison Paul was published in the BritishPteridological Society Bulletin, Volume 8(1), 2014.
136 FERN GAZ. 20(3). 2016

JERMY, A.C. 1970. Selaginellaceae; Isoetaceae. In: EXELL, A.W. & LAUNERT, E.(Eds) Flora Zambesiaca: Pteridophyta. 22-29; 29-32. Crown Agents for OverseasGovernments and Administrations. London.
CRABBE, J.A., JERMY, A.C. & WALKER, S. 1970. The distribution of Dryopterisassimilis S.Walker in Britain. Watsonia 8: 3-15.
CHAMBERS, T.C., JERMY, A.C. & CRABBE, J.A. 1971. A collection of ferns fromEspiritu Santo, New Hebrides. Br. Fern Gaz. 10(4): 175-182.
JERMY, A.C. & HARPER, L. 1971. Spore morphology of the Cystopteris fragiliscomplex. Br. Fern Gaz. 10(4): 211-213.
JERMY, A.C. 1972. Distribution of British pteridophytes. Br. Pterid. Soc. Newsl. 10:32.
JERMY, A.C. & WORLAND, A.J. 1972. The code of conduct: A list of rare plants. Br.Pterid. Soc. Newsl. 10: 33-34.
BROWNSEY, P.J. & JERMY, A.C. 1973. A fern collecting expedition to Crete. Br. FernGaz. 10(6): 331-348.
CRABBE, J.A. & JERMY, A.C. 1973. Seven new species of Selaginella from tropicalSouth America. Am. Fern J. 63(3): 135-144.
JERMY, A.C., CRABBE, J.A. & THOMAS, B.A. (Eds). 1973. The phylogeny andclassification of the ferns. Academic Press for The Linnean Society of London,London. xiii, 284 pp.
HOLMAN, J.H. & JERMY, A.C. 1973. An international directory of Pteridologists. TheBritish Pteridological Society. London. 31 pp.
JERMY, A.C. & STOTT, P.A. (Eds). 1973. Chalk grassland - studies on itsconservation and management in South-East England. Kent Trust for NatureConservation. Maidstone. pp. 59.
JERMY, A.C. & STOTT, P.A. 1973. Appendix 1:Select bibliography; Appendix 2:chalk grassland reserves and SSSI’s of S.E. England. pp. 48-50; 51-59 in JERMY,A.C. & STOTT, P.A. (Eds). Chalk grassland - studies on its conservation andmanagement in South-East England. Kent Trust for Nature Conservation.Maidstone. pp. 59.
BRITTON, D.M. & JERMY, A.C. 1974. The spores of Dryopteris filix-mas and relatedtaxa in North America. Can. J. Bot. 52(8): 1923-1926.
PETTITT, J.M. & JERMY, A.C. 1974. The surface coats on spores. Biol. J. Linn. Soc.6(3): 245-257.
BROWNSEY, P.J. & JERMY, A.C. 1975. Asplenium × khaniense, a new wild hybridfrom Crete. Candollea 30: 21-28.
CRABBE, J.A., JERMY, A.C. & MICKEL, J.T. 1975. A new generic sequence for thepteridophyte herbarium. Fern Gaz. 11(2&3): 141-162.
JERMY, A.C. 1975. Cystopteris, Dryopteris, Woodsia, Gymnocarpium. pp. 112-118,120. In STACE, C.A. (Ed.) Hybridization and the flora of the British Isles.Academic Press & Botanical Society of the British Isles. London. 626 pp.
JERMY, A.C. 1975. The tropical rain forest: an ecosystem on resource. Proc. 13thPacific Sci. Congr. (Abstract of paper read).
JERMY, A.C. & WALKER, T.G. 1975. Lecanopteris spinosa - a new ant-fern fromIndonesia. Fern Gaz. 11(2&3): 165-176.
MOORE, J.A., JERMY, A.C. & MULLIN, J.M. 1975. Lamprothamnium papulosum, anew record for Ireland. Irish Nat. J. 18(8): 233-237.
PETTITT, J.M. & JERMY, A.C. 1975. Pollen in hydrophilous angiosperms. Micron 5:
FERN GAZ. 20(3). 2016 137

377-405.CRABBE, J.A. & JERMY, A.C. 1976. Six new species of Selaginella from tropical
South America. Fern Gaz. 11(4): 255-262. FRASER-JENKINS, C.R. & JERMY, A.C. 1976. Nomenclatural notes on Dryopteris
Adans. Taxon 25(5&6): 659-676. JERMY, A.C. & WILSON, R.A.I. 1976. Guide to reserves and other sites managed by
the Trust. Kent Trust for Nature Conservation, Maidstone. pp. 68.WIDÉN, C.-J., LOUNASMAA, M., JERMY, A.C., JOSEF, V.E. & REICHSTEIN, T.
1976. Die Phloroglucide von zwei Farnhybriden aus England und Schottland, vonauthentischen “Aspidium remotum” A.Braun und von Dryopteris aemula (Aiton)O.Kuntze aus Irland. Helv. Chim. Acta 59(5): 1725-1744.
FRASER-JENKINS, C.R. & JERMY, A.C. 1977. Nomenclatural notes on Dryopteris:2. Fern Gaz. 11(5): 338-340.
GIBBY, M., JERMY, A.C., RASBACH, H, RASBACH, K., REICHSTEIN, T. &VIDA, G. 1977. The fern genus Dryopteris in the Canary Isles and Azores and thedescription of two new tetraploid species. Bot. J. Linn. Soc.: 74: 251-277.
HOLLIS, D., JERMY, A.C. & LINCOLN, R.J. 1977. Biological collecting for the smallexpedition. Geogrl J. 143(2): 249-265.
JERMY, A.C. & WALKER, T.G. 1977. A note on the cytology of Botrychiumlanuginosum and the occurrence of the genus in Malesia. Gard. Bull. StraitsSettlem. 30: 293-298.
SCHELPE, E.A.C.L.E. & JERMY, A.C. 1977. Selaginellaceae; Isoetaceae. pp. 23-31,31-32. In SCHELPE, E.A.C.L.E.. Conspectus Florae Angolensis, vol.Pteridophyta. Junta de Investigações Cientificas do Ultramar, Lisbon. pp. vi,142.
BANGERTER, E.B., CANNON, J.F.M. & JERMY, A.C. 1978. Ferns and their allies(chap. 12). In JERMY, A.C. & CRABBE, J.A. (Eds). The island of Mull, a surveyof its flora and environment. British Museum (Natural History), London. 631pp.
JERMY, A.C. 1978. General description and topography (chap. 3); Climate (chap. 6).In JERMY, A.C. & CRABBE, J.A. (Eds). The island of Mull, a survey of its floraand environment. British Museum (Natural History), London. 631pp.
JERMY, A.C. & CRABBE, J.A. (Eds) 1978. The island of Mull, a survey of its floraand environment. British Museum (Natural History), London. 631pp.
JERMY, A.C., HIBBERD, D.J. & SIMS, P.A. 1978. Brackish and freshwaterecosystems (chap. 9). In JERMY, A.C. & CRABBE, J.A. (Eds). The island of Mull,a survey of its flora and environment. British Museum (Natural History), London.631pp.
JERMY, A.C., JAMES, P.W. & EDDY, A. 1978. Terrestrial ecosystems (chap. 10). InJERMY, A.C. & CRABBE, J.A. (Eds). The island of Mull, a survey of its flora andenvironment. British Museum (Natural History), London. 631pp.
WOOLLEY, A.R. & JERMY, A.C. 1978. Geology (chap. 4). In JERMY, A.C. &CRABBE, J.A. (Eds). The island of Mull, a survey of its flora and environment.British Museum (Natural History), London. 631pp.
JERMY, A.C., ARNOLD, H.R., FARRELL, L. & PERRING, F.H. (Eds). 1978. Atlas offerns of the British Isles. Botanical Society of the British Isles & BritishPteridological Society. London. 101 pp.
FRASER-JENKINS, C.R. & JERMY, A.C. 1978. Nomenclatural Notes on DryopterisAdans. [3] Taxon 27(1): 129-131.
138 FERN GAZ. 20(3). 2016

FRASER-JENKINS, C.R. & JERMY, A.C. 1979. Dryopteris hybrids: a correction. FernGaz. 12(1): 56.
HANBURY-TENISON, A.R. & JERMY, A.C. 1979. The RGS Expedition to GunongMulu, Sarawak 1977-78. Geogrl J. 145: 175-191.
JERMY, A.C. & PAGE, C.N. 1980. Additional field characters separating thesubspecies of Asplenium trichomanes in Britain. Fern Gaz. 12(2): 112-113.
JERMY, A.C. 1980. Notulae et novitates Muluenses: No. 1. Bot. J. Linn. Soc. 81(1):1-46.
JERMY A.C. 1980. Biosystematic studies of Dryopteris. Acta Phytotax. Sin.18(1):37-44.
JERMY, A.C. 1980. Eriophorum. pp. 280-281. In TUTIN, T.G. et al. (Eds) FloraEuropaea Vol. 5. Cambridge University Press. xxxvi, 452 pp.
JERMY, A.C. 1980. Diphasiastrum (Lycopodium) issleri (Rouy) Holub inGloucestershire. Gloucs. Nats Soc. 31(11&12): 438-439.
ALSTON, A.H.G., JERMY, A.C. & RANKIN, J.M. 1981. The genus Selaginella intropical South America. Bull. Brit. Mus. (Nat. Hist.) Bot. 9(4): 233-330.
JERMY, A.C. 1981. Asplenium cuneifolium Viv. erroneously recorded in the BritishIsles. Watsonia 13(4): 322-323.
JERMY, A.C. 1981. Youth and Exploration: Tropical rain forests. BP InternationalLimited and Royal Geographical Society. London. 27 pp.
ANDERSON, J.A.R., JERMY, A.C. & CRANBROOK, Earl of. 1982. Gunung MuluNational Park: A management and development plan. (1979 Report to the SarawakGovernment) Royal Geographical Society. London.
JERMY, A.C. 1982. Gunung Mulu National Park: the 1977-78 survey. Sarawak Mus.J. 30 (51 new ser.): 1-15.
JERMY, A.C. & KAVANAGH, K.P. (Eds). 1982. Gunung Mulu National Park,Sarawak. An account of its environmemt and biota being the results of theRGS/Sarawak Government expedition and survey 1977-78. Sarawak Mus. J. 30 (51new ser.) Special Issue No. 2, Pt 1: 1-279.
JERMY, A.C. PARRIS, B.S. & RANKIN, J.M. 1982. A preliminary list ofPteridophytes recorded in Gunung Mulu National Park. App. IV in ANDERSON,J.A.R., JERMY, A.C. & CRANBROOK, Earl of. Gunung Mulu National Park: Amanagement and development plan. (1979 Report to the Sarawak Government)Royal Geographical Society. London.
JERMY, A.C. & SCHELPE, E.A.C.L.E. 1982. Isoetaceae. In New species and newcombinations in African Pteridophyta and Orchidaceae. Contr. Bolus Herb.10:150-151.
WALKER, T.G. & JERMY, A.C. 1982. The ecology and cytology of Phanerosorus(Matoniaceae). Fern. Gaz. 12(4): 209-213.
JERMY, A.C., CHATER, A.O. & DAVID, R.W. 1982. Sedges of the British Isles. BSBIHandbook No l, 2nd edition. Botanical Society of the British Isles, London. 268 pp.
JERMY, A.C. 1983. Gunung Mulu National Park, Sarawak. Oryx. 17: 6-14.JERMY, A.C. 1984. Origin and distribution of Pteridophytes in the Mediterranean area.
Webbia 38(1): 397-416.JERMY, A.C. 1984. The scaly male fern – problem fern number one. Pteridologist 1(1):
10-12.JERMY, A.C. (Ed.) 1984. Studies on the flora of Gunung Mulu National Park,
Sarawak. Forest Department, Sarawak. Kuching. xv, 233 pp.
FERN GAZ. 20(3). 2016 139

JERMY, A.C. 1984. Introduction. pp. v-xv. In JERMY, A.C. (Ed.). Studies on the floraof Gunung Mulu National Park, Sarawak. Forest Department, Sarawak. Kuching.xv, 233 pp.
JERMY, A.C., PARRIS, B.S., CAMUS, J.M. & PAUL, A.M. 1984. The Pteridophyta ofGunung Mulu National Park. pp. 145-233. In JERMY, A.C. (Ed.). Studies on theflora of Gunung Mulu National Park, Sarawak. Forest Department., Sarawak.Kuching. xv, 233 pp.
JERMY A.C. 1985. Cytotaxonomic studies of the ferns of Trinidad 1. The climate,geology, and vegetation of Trinidad with particular reference to the ecology offerns. Bull. Br. Mus. Nat. Hist. (Bot.) 13(2): 133-147.
JERMY A.C. & WALKER, T.G. 1985. Cytotaxonomic studies of the ferns of Trinidad3. Descriptions of new species and hybrids and a new combination. Bull. Br. Mus.Nat. Hist. (Bot.) 13(2): 251-147.
GIVEN, D.R. & JERMY, A.C. 1985. Conservation of pteridophytes: a postscript. Proc.Roy. Soc. Edinburgh. Section B. Biol. Sci. 86: 435-437.
JERMY, A.C. 1986. Subgeneric names in Selaginella. Fern Gaz. 13(2): 117-118. JERMY, A.C. 1986. Two new Selaginella species from Gunung Mulu National Park,
Sarawak. Kew Bull. 41: 547-559. DERRICK, L.N., JERMY, A.C. & PAUL, A.M. 1987. Checklist of European
pteridophytes. Sommerfeltia 6: xx, 94 pp. FITZGERALD, R. & JERMY, A.C. 1987. Equisetum ramosissimum in Somerset.
Pteridologist 1(4): 178-181.JERMY, A.C. & WALKER, T.G. 1987. Adiantum × spurium, a new name for A. ×
villosolucidum. Fern Gaz. 13(3): 188.JERMY, A.C. & WOODHAMS, J.R. 1987. Ferns at Kew Garden - A review.
Pteridologist 1(4): 159-161.VIANE, R., MAYOR LÓPEZ, M. & JERMY, A.C. 1987. The systematics and ecology
of the pteridophytes of northern Spain. Excursion guide. XIV InternationalBotanical Congress. Berlin. 54pp.
JERMY, A.C. 1988. Responsibilities for plant conservation: past, present and future.Plants Today. November-December 1988: 175-177.
JERMY, A.C. 1989. The history of Diphasiastrum issleri (Lycopodiaceae) in Britainand a review of its taxonomic status. Fern Gaz. 13(5): 257-265.
SHEFFIELD, E., WOLF, P.G., HAUFLER, C.H., RANKER, T. & JERMY, A.C. 1989.A re‐evaluation of plants referred to as Pteridium herediae (Colmeiro) Löve &Kjellqvist. Bot. J. Linn. Soc. 99(4): 377-386.
CAMUS, J.M. & JERMY, A.C. 1990. Ferns of woods and moorland. Br. Wildl. 1(6):350-353.
JERMY, A.C. 1990. Conservation of Pteridophytes, p. 14, In K.U. Kramer & P.S. Green(Eds.) The families and genera of vascular plants. Vol.1. Pteridophytes andGymnosperms. Springer-Verlag, Berlin, 1.
JERMY, A.C. 1990. Isoetaceae. pp. 26-31, In K.U. Kramer & P.S. Green (Eds.) Thefamilies and genera of vascular plants. Vol.1. Pteridophytes and Gymnosperms.Springer-Verlag, Berlin, 1.
JERMY, A.C. 1990. Selaginellaceae. pp. 39-45, In K.U. Kramer & P.S. Green (Eds.)The families and genera of vascular plants. Vol.1. Pteridophytes andGymnosperms. Springer-Verlag, Berlin, 1.
JERMY, A.C. & CAMUS, J.M. 1991. The illustrated field guide to ferns and allied
140 FERN GAZ. 20(3). 2016

plants of the British Isles. Natural History Museum Publications, London. xiv, 194pp.
CAMUS, J.M., JERMY, A.C. & THOMAS, B.A. 1991. A world of ferns. NaturalHistory Museum Publications, London. 112 pp.
JERMY, A.C. & JARVIS, C.E. 1992. The lectotypification of Aspleniumtrichomanes-ramosum L., the correct name for A. viride Hudson. pp. 319-321. InNewton, M.E. Flora Europaea: notulae systematicae ad floram Europaeamspectantes. Ser. 2 No. 5. Bot. J. Linn. Soc. 109(3): 319-328.
JERMY, A.C. & VIANE, R. 1992. New subgenera within Asplenium L.. pp. 321-322.In Newton, M.E. Flora Europaea: notulae systematicae ad floram Europaeamspectantes. Ser. 2 No. 5. Bot. J. Linn. Soc. 109(3): 319-328.
IDE, J.M., JERMY, A.C. & PAUL, A.M. (Eds). 1992. Fern horticulture: past, presentand future perspectives. Intercept, Andover. xii, 308 pp.
JERMY, A.C. 1992. What’s in a name?: Nomenclature, of fern ‘varieties’. pp. 87-96.In: IDE, J.M., JERMY, A.C. & PAUL, A.M. (Eds) Fern horticulture: past, presentand future perspectives. Intercept, Andover. xii, 308 pp.
CRABBE, J. A. & JERMY, A.C. 1993. Cystopteris Bernh. In: TUTIN, T.G.,BURGESS, N.A., CHATER, A.O. et al. Flora Europaea, vol. 1: Psilotaceae toPlatanaceae, pp. 24-24.
JERMY, A.C., TITTLEY, I. & GEORGE, J.D. 1992. A brief biological survey of theproposed pipeline route across Walney Island. A report for British Gas Explorationand Production Limited. Natural History Museum. London. 13pp.
JERMY, A.C. 1993. British clubmosses. Br. Wildl. 4(4): 216-220.JERMY, A.C. 1993. Athyrium Roth, Deparia Hooker & Grev., Holub, Diplazium
Swartz, Gymnocarpium Newman. In: TUTIN, T.G., BURGESS, N.A., CHATER,A.O. et al. Flora Europaea, vol. 1: Psilotaceae to Platanaceae, pp. 24, 26.
JERMY, A.C. & AKEROYD, J. R. 1993. Isoetes L. In TUTIN, T.G., BURGESS, N.A.,CHATER, A.O. et al. Flora Europaea, vol. 1: Psilotaceae to Platanaceae, pp. 6-7.
JERMY, A.C. & CHAPMAN, R. 1993. Tropical forest expeditions. 4th edition. RoyalGeographical Society Expedition Advisory Centre. London. 104 pp.
JERMY, A.C., FUCHS, H.P. & PAUL, A.M. 1993. Cheilanthes Swartz. In TUTIN, T.G.,BURGESS, N.A., CHATER, A.O. et al. Flora Europaea, vol. 1: Psilotaceae toPlatanaceae, p. 12.
JERMY, A.C. & PAUL, A.M. 1993. Christella Léveillé, Cosentinia Tod., NotholaenaR.Br., Oreopteris J. Holub, Phegopteris (C.Presl) Fée, Stegnogramma Blume,Thelypteris Schmidel. In TUTIN, T.G., BURGESS, N.A., CHATER, A.O. et al.Flora Europaea, vol. 1: Psilotaceae to Platanaceae, pp. 12-13, 17-18.
ROTHMALER, W. & JERMY, A.C. 1993. Diphasiastrum J. Holub, Lycopodiella J.Holub, Lycopodium L. In TUTIN, T.G., BURGESS, N.A., CHATER, A.O. et al.Flora Europaea, vol. 1: Psilotaceae to Platanaceae, pp. 3-5.
VALENTINE, D.H., JERMY, A.C. & AKEROYD, J.R. Polypodium L. In TUTIN, T.G.,BURGESS, N.A., CHATER, A.O. et al. Flora Europaea vol. 1: Psilotaceae toPlatanaceae, pp. 15-16.
VIANE, R., JERMY, A.C. & LOVIS, J.D. 1993. Asplenium. In TUTIN, T.G.,BURGESS, N.A., CHATER, A.O. et al. Flora Europaea, vol. 1: Psilotaceae toPlatanaceae, pp. 18-23.
VOGEL, J.C., JESSEN, S., GIBBY, M., JERMY, A.C. & ELLIS, L. 1993.Gametophytes of Trichomanes speciosum Willd. (Hymenophyllaceae,
FERN GAZ. 20(3). 2016 141

Pteridophyta) in Central Europe. Fern Gaz. 14: 227-232.TITTLEY, I., HUXLEY, R. & JERMY, A.C. 1993. Investigation of the effects of
sewage spills on the vegetation of Walthamstow Marshes. A report to ThamesWater Utilities. Natural History Museum. London. 72pp.
MORAN R.C., ZIMMER, B. & JERMY A.C. 1995. Adiantum. In: Moran, R.C. & Riba,R. (Eds). Flora Mesoamericana. Volumen 1. Psilotaceae a Salviniaceae. pp.106-117. Univ. Nac. Autónoma de México. México D. F., México.
WORKMAN, C., GIMINGHAM, A. & JERMY, A.C. (Eds) 1995. Environmentalresponsibility for expeditions – A guide to good practice. British EcologicalSociety and Young Explorers’ Trust.
JERMY, A.C., LONG, D., SANDS, M.J.S., STORK, N.E. & WINSER, S. 1996.Biodiversity assessment review: a guide to good practice. HMSO PublicationsCentre.
JERMY, A.C. 1996. British Equisetum - horsetails. Br. Wildl. 8(1): 37-41.JERMY, A.C. 1998. Fern recording and monitoring: a programme to take the BPS into
the next century. Pteridologist 3(3): 5-12.RICH, T.C.G. & JERMY, A.C. 1998. Plant crib. Bot. Soc. Br. Isl., London. 392 pp.RUMSEY, F.J., JERMY, A.C. & SHEFFIELD, E. 1998. The independent gametophytic
stage of Trichomanes speciosum Willd. (Hymenophyllaceae), the Killarney Fernand its distribution in the British Isles. Watsonia 22(1): 1-19.
STEWART, N.F., FITZGERALD, R., JERMY, A.C. & LANSDOWN, R.V. 1999. Theconservation of pillwort Pilularia globulifera in England. Interim Report No. 1.Plantlife Report No. 130. Plantlife, London.
JERMY, A.C. & CHAPMAN, R. 2002. Tropical forest expeditions. 5th edition.RGS-IBG Expedition Advisory Centre. London. 197 pp.
DEAN, M., HUTCHEON, K., CAYOUETTE, J., JERMY, A.C. & ASHTON, P.A.2005. Carex salina - a new species of sedge for Britain. BSBI News (99): 17-19.
JERMY, A.C., SIMPSON, D.A., FOLEY, M.J.Y. & PORTER, M.S. 2007. Sedges of theBritish Isles. BSBI Handbook No. 1, 3rd edition. Bot. Soc. Br. Isl., London. 554pp.
SIMPSON, D.A. & JERMY, A.C. 2007. The status of Trichophorumcespitosum nothosubsp. foersteri (Cyperaceae). Kew Bull. 62(1): 165.
Also abstracts of presentations, obituaries and c. 50 reviews of books in a number ofbiological and geographical journals.
COMPILED BY M. GIBBY & A.M.PAUL
142 FERN GAZ. 20(3). 2016

SHORT NOTE
RETYPIFICATIONS OF ADIANTUM INCISUM (PTERIDACEAE) ANDPTERIS INTERRUPTA (THELYPTERIDACEAE)
Species-rich eastern India includes the states of Sikkim, West Bengal, Arunachal Pradesh,Meghalaya, Assam, Tripura, Manipur, Nagaland and Mizoram (Ghosh et al. 2004).Rejection of records of a number of new species and new distributional records fromthis region by Fraser-Jenkins (2008, 2012) necessitate a revision to the treatment byGhosh et al. As a part of ongoing revision of the ferns of Eastern India, Adiantum incisumForssk. (Pteridaceae) and Pteris interrupta Willd. (current name: Cyclosorus interruptus(Willd.) H.Ito, Thelypteridaceae) are retypified.
Adiantum incisum Forssk., Fl. Aegypt.-Arab. 187. 1775.Lectotype (designated here): Yemen. Al Hadiyah [‘Hadîe’], 1763, P. Forsskål s.n.(C10001579, Isolectotype C10001580 images!).
The name Adiantum incisum was proposed by Linnaeus’s disciple Peter Forsskål(1775) based on his collection from Yemen. The Botanical Garden, University ofCopenhagen, Denmark (C) stores two specimens: C10001579(http://www.daim.snm.ku.dk/digitized-type-collection-details-simple?catno=C10001579) and C10001580(http://www.daim.snm.ku.dk/digitized-type-collection-details-simple?catno=C10001580).C10001579 is a complete plant with Forsskål’s field label “Adianthum *incisum Hadîe”,thus undoubtedly original material and type of Adiantum incisum. C10001580 is singleisolated frond. It does not have Forsskål’s annotation but resembles C10001579 and ismost probably from same gathering.
During a revision of the ferns of tropical East Africa the renowned botanist the lateDr. Bernard Verdcourt selected the specimen in C with the original field label (i.e.C10001579) as lectotype (see Verdcourt 2002: 55). Although his lectotype selection istaxonomically correct, it unfortunately did not fulfill requirements specified in theMelbourne Code (McNeill et al. 2012). Lectotypification statements published on orafter 1 Jan 2001 must include the phrase “designated here” (hic designatus) or anequivalent (McNeill et al. 2012 Art. 7.10). Verdcourt’s lectotypification was publishedin 2002 without such statement. Here it is republished to fix the application of the nameA. incisum.
Pteris interruptaWilld., Phytographia 13 t. 10 f. 1. 1794. [Sep 1794].Lectotype (designated here): Willdenow (1794), Phytographia t. 10, f. 1. [Icon].Epitype (designated here): India. Tamil Nadu [“Madras”], Aug 1794, Rottler ex Kleins.n. (BW19770011, Isoepitype BW19770012).
Pteris interrupta (Cyclosorus interruptus) is the type of the name Cyclosorus Link.Fosberg & Sachet (1972) considered a specimen (BW19770011;http://herbarium.bgbm.org/object/BW19770011) in Herb. Willdenow from HerbariumBerolinense, Berlin (B) as the type of Pteris interrupta and this has been considered tobe a lectotype. Holttum (1973) also treated it as such.
The name Pteris interrupta was published in September 1794 by Willdenow, but the
FERN GAZ. 20(3). 2016 143

supposed type sheet (BW19770011) was annotated by Willdenow as “Klein Ind. 1797.W.” This indicates that Willdenow received this specimen in 1797 from Johann GottfriedKlein, a missionary and botanist from South India. Thus this specimen is not originalmaterial, rather a later collection.
The putative type (BW19770011) also carries the annotation “Pteris sp. n.? Aug.1794. Auf den Reise nach Madras” written by Johan Peter Rottler, a missionary andbotanist from South India (Dr. Frits Adema pers. comm. 2015). However the annotation“(Klein)” was written by Willdenow. So this specimen was actually collected by Rottlerin August 1794 during a trip through Madras (now Tamil Nadu, India) and reachedWilldenow through Klein. It is unlikely that Rottler’s collection (in August 1794) wassubsequently studied and published by Willdenow within short period of one month (inSeptember 1794).
This sheet does not carry Willdenow’s name Pteris interrupta, but Willdenow’sannotation “Asp. obtusatum 1a.” The name Aspidium obtusatum Sw. was published in1801 by Swartz. As explained by Fosberg & Sachet (1972: 8) Willdenow sent a pinnafrom BW19770011 to Swartz. The pinna, now in the Natural History Museum, London(BM), was identified by Swartz as Aspidium obtusatum and he informed Willdenow ofthis.
Another sheet (BW19770012; http://herbarium.bgbm.org/object/BW19770012)exists in Herb. Willdenow in B carrying two isolated fronds and Willdenow’s annotation“Asp. obtusatum 1b.” These two specimens resemble BW19770011 and were mostprobably collected by Rottler from same individual or population.
Thus no type specimen for Pteris interrupta is known to exist. The illustration “t. 10f. 1.” of Pteris interrupta in the protologue (Willdenow 1794) is the only original materialand I select it as lectotype and BW19770011 as epitype.
ACKNOWLEDGEMENTSI sincerely thank Dr. J. McNeill, Royal Botanic Garden Edinburgh for constructiverevision and Dr. F. Adema, Naturalis Biodiversity Center, Leiden for helpful advice.
REFERENCESFORSSKÅL, P. 1775. Flora Aegyptiaco-Arabica etc. Post mortem auctoris edidit Carsten
Niebuhr. ex officina Molleri, Copenhagen.FOSBERG, F.R. & SACHET, M.H. 1972. Three Indo-Pacific Thelypteroid species
reinterpreted and a new African species described. Smiths. Contr. Bot. 8: 1–10.FRASER-JENKINS, C.R. 2008. Taxonomic revision of three hundred Indian
subcontinental Pteridophytes with a revised Census- List. Bishen Singh MahendraPal Singh, Dehra Dun.
FRASER-JENKINS, C.R. 2012. Rare and Threatened Pteridophytes of Asia 2.Endangered Species of India—the Higher IUCN Categories. Bull. Natl. Mus. Nat.Sci., Ser. B, 38: 153–181.
GHOSH, S.R., GHOSH, B., BISWAS, A. & GHOSH, R.K. 2004. Flora of India, Series4: The Pteridophytic flora of eastern India. Vol. 1, Botanical Survey of India, Kolkata.
HOLTTUM, R.E. 1973. The identity of three type specimens in the WilldenowHerbarium. Amer. Fern J. 63: 81–84.
MCNEILL, J., BARRIE, F.R., BUCK, W.R., DEMOULIN, V., GREUTER, W.,HAWKSWORTH, D.L., HERENDEEN, P.S., KNAPP, S., MARHOLD, K., PRADO,J., PRUD’HOMME VAN REINE, W.F., SMITH, G.F., WIERSEMA, J.H.,
144 FERN GAZ. 20(3). 2016

TURLAND, N.J. (Eds.) 2012. International Code of Nomenclature for algae, fungi,and plants (Melbourne Code): adopted by the Eighteenth International BotanicalCongress Melbourne, Australia, July 2011. Regnum Vegetabile 154, 1–274.
SWARTZ, O. 1801. Genera et species filicum ordine systematico redactorum. J. Bot.(Schrader) 1800: 1–136.
VERDCOURT, B. 2002. Adiantaceae. In: Flora of Tropical East Africa. Beentje, H.J.(Ed.), A.A. Balkema Publishers, Rotterdam.
WILLDENOW, C.L. VON 1794. Phytographia seu descriptio rariorum minus rognitarumplantarum etc. Sumptibus Wolfgangi Walther, Alemania.(http://bibdigital.rjb.csic.es/ing/Libro.php?Libro=6074).
J. MAZUMDARDepartment of Biological Sciences, Burdwan Town School, Burdwan-71301, West
Bengal, India. Email: [email protected]
FERN GAZ. 20(3). 2016 145

BOOK REVIEW
Ferns and Fern-allies of Nepal, Volume 1. Fraser-Jenkins C.R., Kandel D.R. &Pariyar S. 2015. pp 508. ISBN: 978-9937-2-9496-6 : National Herbarium and PlantLaboratories, Department of Plant Resources, Ministry of Forests and Soil Conservation,Kathmandu, Nepal.
Christopher Fraser-Jenkins has been studying Himalayan pteridophytes for over 30 yearsand for much of this time also living in Nepal. He is the pre-eminent expert of ferns inHimalayan region and also possesses vast expertise on the ferns of wider Asia andEurope. In this volume (the first of 2) the results of his studies of Nepalese ferns andlycophytes are summarised. The richness of the flora is highlighted by the fact that 580species and subspecies will be included once the second volume has been completed.By comparison, England and Wales occupying an area of similar size if not topography,has about 80 taxa including naturalised introductions.
The introduction to this volume includes interesting and useful summaries onphytogeography, collectors, collections and publications pertaining to Nepal. Thenfollows a section outlining the classification utilised (basically that of Smith et al. (2006)),and a longer section on the apparent deficiencies of recent molecular-based phylogenies.Fraser-Jenkins is well known for his scepticism of the utilisation of small sections ofDNA as a basis for classification. His frequent strongly stated views and objections ofstudies that don’t coincide with his ideas based on traditional morphological taxonomiesare presented again here. Next are short sections with notes on the history and progressof the current volume and the format of the species accounts. This is followed with a listof accepted species and subspecies included in both Volume 1 and 2. The list of taxa tobe published in Volume 2 includes taxonomic and nomenclatural notes on various speciesas well as new combinations, new species and even a new genus, Katoella (Davalliaceae).This latter example highlights one of the inconsistencies of his morphological approach.Davalliaceae is split into a number of small genera whereas the much larger andmorphologically diverse family Thelypteridaceae contains a single genus, Thelypteris.Recent molecular studies are starting to unravel the phylogeny of this family, but willthey be acceptable to Fraser-Jenkins? Maybe we will find out when volume 2 ispublished. Intriguingly, in this list, Dryopsis is lumped back in to Ctenitis withoutexplanation, which seems to be at odds with both morphological and molecular evidence.Hopefully this decision will also be explained in more detail in the second volume.
The main bulk of the book is taken up with the taxonomic accounts of 20 familiesincluding those of the Lycophyta and the early branching fern orders through to thePteridaceae in the Polypodiales (Vittariaceae remains separated and is the concludingfamily to Volume 1). The amount of information and knowledge collated here isastounding. For example, a page opened at random revealed the treatment ofAleuritopteris bicolor. It starts with a list of synonyms and misapplied names. Thenfollows a short overview of its confused history, cytology, ethnobotany and a briefcomparative description. Next is a list of dozens of examined specimens listed under thedifferent Nepalese regions and finally a note on its total distribution and status in Nepal.It should be emphasised that this is not an identification manual as such and no keys areprovided. To confidently identify one of the many different Pteris, Onychium orAleuritopteris species without prior experience would be difficult. However for smallergenera, such as the difficult Coniogramme, accurate identification should be possible
146 FERN GAZ. 20(3). 2016

from the brief descriptions. Incidentally under Coniogramme and several other genera,Fraser-Jenkins, with some justification, draws attention to the many errors of taxonomyand nomenclature contained within the recent Flora of China Pteridophyta treatment. Aswe have come to expect, his opinion is strongly presented, finishing with a radicalproposal to resolve the problems. In regard to Coniogramme I have spent some timetrying to sort out the species in cultivation and in my view his ideas make far more sensethan those found in the Flora of China account.
As is often the case in Fraser-Jenkins’ works, many interesting comments andobservations are concealed under various species accounts, including unexpected noteson species far away from Nepal. For example, both Adiantum viridimontanum and Pterislongifolia of the New World are reduced to subspecies of Adiantum pedatum and Pterisvittata respectively. Several important accounts clarifying the taxonomy and distributionof difficult genera are included. Onychium is comprehensively revised for the region,with descriptions of two new species and a new hybrid. Selaginella and Pteris are thetwo largest genera in Volume 1 with 23 and 27 species respectively. I noticed that thename Pteris longipes is considered to have been misapplied and is actually an earliername for the well-known P. wallichiana. Fraser-Jenkins favours rejection of the formername and I suspect a formal proposal is in the pipeline. Other complex genera such asHuperzia, Adiantum, Aleuritopteris and Microlepia are very fully treated and the latter,in particular, includes much new and useful information. One important name changeaffects the widespread and common fern, Dennstaedtia scabra. The rules of priority willmean D. zeylanica now takes precedence.
At the end of the book are 138 colour plates that show many of useful discriminatingcharacters for a range of species. Finally there is an appendix explaining in detail theconfused identity and nomenclature of Athyrium setiferum and the index.
I noted very few printing or spelling mistakes. Only one error stood out, Sambucusedulis (page 270), presumably meaning S. nigra (Elder). This is an example thatdemonstrates the author’s intimate knowledge of Asian ferns; he notes that living frondsof Coniogramme procera smell strongly of Elder when crushed. I doubt that this smallbut useful piece of information has ever been published previously.
Rather frustratingly there is no bibliography in the present volume and presumablythis will be included in Volume 2. Also a separate and complete list of new taxa andcombinations would be most useful. Although they are included in bold type in the indexof this volume I noticed at least one had been overlooked. The book is reasonably wellproduced although the binding leaves much to be desired and my copy is already showingsigns of distress.
In summary, Chris Fraser-Jenkins and his co-authors are to be warmly congratulatedon producing this landmark in Asian pteridology. It will provide a detailed baseline ofdata that will be used widely throughout the Himalayan region and beyond. I and manyother pteridologists, both professional and amateur, will eagerly await the arrival of thesecond volume.
REFERENCESSMITH, A.R., PRYER, K.M., SCHUETTPELZ, E., KORALL, P., SCHNEIDER, H &
WOLF, P.G. 2006. A classification for extant ferns. Taxon 55(3): 705-731.
T. PYNER
FERN GAZ. 20(3). 2016 147

INSTRUCTIONS FOR AUTHORS
PAPERS should not usually exceed 20 printed pages and are generally expected to beconsiderably shorter. As well as reports of original research, review articles and shortnotes, e.g. new records, are encouraged. The senior author should supply an email addressto facilitate correspondence. REVIEWS should cover recent developments in any aspect of research on ferns andlycopods. This can include unpublished research by the author, and/or the possibledirection of future research. They should be aimed at the general reader who may not bean expert in the chosen topic. Proposals for a review should be discussed first with thereview editors.MANUSCRIPTS should be submitted in English (British) in electronic format in10-point Times New Roman font and double spaced. Electronic versions of text andtables should be compatible with WORD, with figures as pdf or jpg files, and sent asemail attachments or CDroms. All manuscripts will be refereed.THE TITLE should reflect the content of the paper and be in BOLD CAPITALS(11-point) and centrally aligned. Generic and specific names should be in italics and anytitle containing a generic or specific name should be followed by the family name inbrackets e.g.
TRICHOMANES SPECIOSUM (HYMENOPHYLLACEAE)IN SOUTHERN SPAIN
AUTHOR ABBREVIATIONS should follow Pichi Sermolli’s (1996) Authors ofscientific names in Pteridophyta, Royal Botanic Gardens, Kew.MAIN HEADINGS: should be in BOLD CAPITALS (10-point) and centrally aligned.SUBSIDIARY HEADINGS: should be in bold, the first letter of each word in capitals,the rest in lower case and left-aligned.AUTHORS’ NAMES AND FULL ADDRESSES (including email address): followthe title and are centrally aligned.KEY WORDS: up to ten.ABSTRACT: should reflect the content of the paper.FIGURES: there is no distinction between photographs and line drawings in numbering.All should be presented in a form ready for reproduction, colour, TIF format and 300dpi (please contact editor with queries), with a scale bar where appropriate. Lettering ornumbers (Arabic) should be in the bottom left using uppercase Times Roman. Figurecaptions should be on a separate sheet.TABLES: can be printed in either portrait or landscape format. Authors should considerthis when preparing tables. Authors should ensure that tables fit the printed page size ina legible form.PAGESIZE: The Gazette has a useable page size of 123 × 204 mm. This can be used inportrait or landscape mode but any supplied artwork should not be larger than this.MEASUREMENTS: follow the metric system.CHECKLISTS: follow the format of Baksh-Comeau, Fern Gaz. 16(1, 2): 11- 122.REFERENCES: follow the style of a recent issue of The Fern Gazette, e.g.:-HOOKER, W.J. 1864. Species Filicum, 5. Dulau & Co., London.MORTON, C.V. 1947. The American species of Hymenophyllum, section
Sphaeroconium. Contr. U.S. Natl. Herb. 29(3): 139-201.STEVENSON, D.W. & LOCONTE, H. 1996. Ordinal and familial relationships of
pteridophyte genera. In: CAMUS, J.M., GIBBY, M. & JOHNS, R.J. (Eds)Pteridology in perspective, pp. 435-467. Royal Botanic Gardens, Kew.
JOURNAL ABBREVIATIONS: should follow Botanico Periodicum Huntianum &Supplements.Alterations from the original text at proof stage will be charged for unless they are minorpoints of detail. A pdf will be provided free to the senior author.
148 FERN GAZ. 20(3). 2016

DECAPLOID GAMETOPHYTE FORMATION FROM SPORES OF APENTAPLOID CYSTOPTERIS FRAGILIS (CYSTOPTERIDACEAE)
COLLECTED IN MONGOLIAN ALTAI
SUZUE M. KAWAKAMI1*, SHOGO KAWAKAMI2, OYUNCHIMEGDAMDINSUREN3, JUNTARO KATO4, SERGEY V. SMIRNOV5 &
KATSUHIKO KONDO
1Nishio Nursing College, Togasaki, Nishio City, Aichi Prefecture 445-0074, Japan. email:[email protected]
2Aichi University of Education, Hirosawa, Kariya, Aichi Prefecture, 448-8542, Japan.email:[email protected]
3Biology Division, Hovd State University, Hovd, Hovd Province 213500, Mongolia. 4Department of Biology, Aichi University of Education, Kariya City, Aichi Prefecture
448-8542, Japan. email: [email protected] of Plant Systematics and South-Siberian Botanical Garden, Altai State
University, Barnaul 656099, Russia. email: [email protected] Research Institute of Evolutionary Biology, Tokyo University of Agriculture,
2-4-28, Kamiyoga, Setagaya, Tokyo 158-0098, Japan. email: [email protected]*Author for correspondence, email:[email protected]
Key words: Cystopteris fragilis, decaploid, meiosis, pentaploid, sporogenesis
ABSTRACTGerminating spores were obtained from a pentaploid Cystopteris fragilis collectedin Mongolian Altai. DNA content of nuclei in gametophytes showed for the firsttime that gametophytes that developed from spores of the pentaploid sporophytehad a larger number of genomes than those of the mother sporophyte, and someappeared to be decaploid. Fertile decaploid (10n) spores may be produced in thepentaploid C. fragilis and derived from monad spores observed in sporogenesis.Antheridia were produced on the 10n gametophytes, although antherozooids withswimming ability were not observed. Archegonia were not observed. Apogamoussporophytes with 10n genome content were induced from decaploidgametophytes. The monad spores produced may contribute to the formation ofhigher polyploid sporophyte series in ferns.
INTRODUCTIONCystopteris fragilis (L.) Bernh. collected in Mongolian Altai showed three cytotypes,tetraploid, pentaploid and hexaploid, and although numerous univalent and bivalentchromosomes were observed in the spore mother cells of the pentaploid C. fragilis (2n= 5x = ca. 210), some germinating spores were obtained from the sporophyte (Kawakamiet al., 2010). In the present paper, therefore, we aimed to investigate the genome contentsof germinating spores produced and also to determine how these viable spores areproduced in the pentaploid sporophyte. From the results of DNA contents of nuclei ingametophytes raised from spores of the pentaploid sporophyte, it was shown for the firsttime that the gametophytes produced had not the same but a larger number of genomesthan those of the mother sporophyte, and surprisingly, some gametophytes had decaploid
FERN GAZ. 20(4):149-155. 2016 149
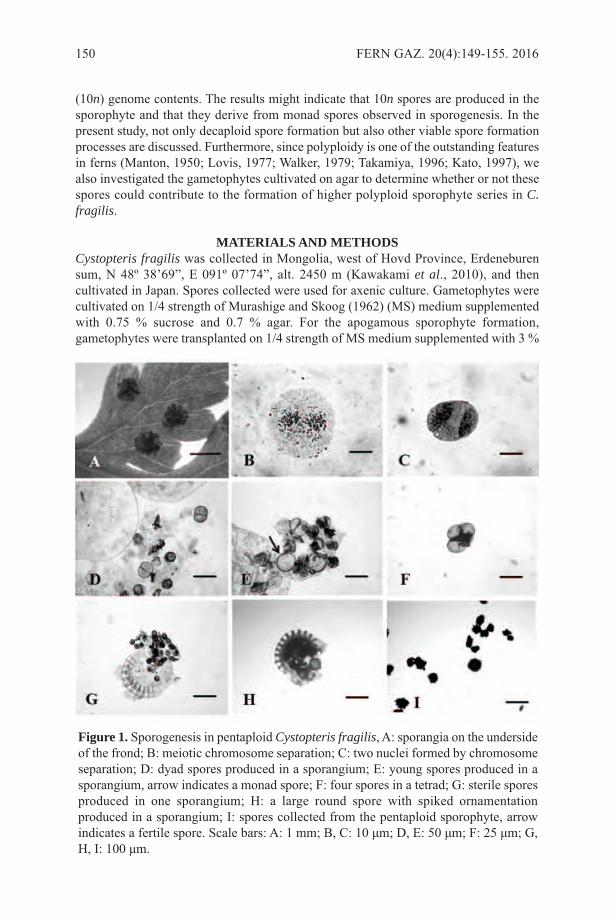
(10n) genome contents. The results might indicate that 10n spores are produced in thesporophyte and that they derive from monad spores observed in sporogenesis. In thepresent study, not only decaploid spore formation but also other viable spore formationprocesses are discussed. Furthermore, since polyploidy is one of the outstanding featuresin ferns (Manton, 1950; Lovis, 1977; Walker, 1979; Takamiya, 1996; Kato, 1997), wealso investigated the gametophytes cultivated on agar to determine whether or not thesespores could contribute to the formation of higher polyploid sporophyte series in C.fragilis.
MATERIALS AND METHODSCystopteris fragilis was collected in Mongolia, west of Hovd Province, Erdeneburensum, N 48º 38’69”, E 091º 07’74”, alt. 2450 m (Kawakami et al., 2010), and thencultivated in Japan. Spores collected were used for axenic culture. Gametophytes werecultivated on 1/4 strength of Murashige and Skoog (1962) (MS) medium supplementedwith 0.75 % sucrose and 0.7 % agar. For the apogamous sporophyte formation,gametophytes were transplanted on 1/4 strength of MS medium supplemented with 3 %
150 FERN GAZ. 20(4):149-155. 2016
Figure 1. Sporogenesis in pentaploid Cystopteris fragilis, A: sporangia on the undersideof the frond; B: meiotic chromosome separation; C: two nuclei formed by chromosomeseparation; D: dyad spores produced in a sporangium; E: young spores produced in asporangium, arrow indicates a monad spore; F: four spores in a tetrad; G: sterile sporesproduced in one sporangium; H: a large round spore with spiked ornamentationproduced in a sporangium; I: spores collected from the pentaploid sporophyte, arrowindicates a fertile spore. Scale bars: A: 1 mm; B, C: 10 μm; D, E: 50 μm; F: 25 μm; G,H, I: 100 μm.

sucrose, 0.1 % casamino acid and 0.8 % agar. Cultures were maintained at 25 ℃ andilluminated by two fluorescent lamps (NEC FL 15BR) to keep 800 lux at the surface.Meiotic chromosomes were observed by fixing sporangia with 3:1 ethanol-acetic acidfor 30 min at 5 ℃ and squashing them in 2 % aceto-orcein solution. The DNA contentsof nuclei in fronds were estimated by flow cytometry using a Partec Ploidy Analyzer PA(Partec, Münster, Germany) (Kawakami et al., 2003).
RESULTSThe pentaploid Cystopteris fragilis produced sporangia on the underside of the frond(Figure 1A). In spore mother cells, numerous univalent and bivalent chromosomes wereobserved at meiotic metaphaseⅠ. By meiotic chromosome separation (Figure 1B) twonuclei were formed (Figure 1C), and dyad spores were observed frequently in sporangia(Figure 1D). Some of these consisted of both large and small spores. Monad spores(Figure 1E) and spores in tetrads (Figure 1F) were more rarely observed. Mostly, youngspores produced in sporangia did not mature and the sporangia did not develop (Figure1G). In a few expanded sporangia, a few large spores with spiked ornamentation wereobserved (Figure 1H). Although spores obtained were mostly abortive, some were ableto germinate (Figure 1I).
DNA contents of nuclei in 13 gametophytes derived from spores of the pentaploidsporophyte were investigated. Their genome contents were greater than those of the donorpentaploid sporophyte (Figure 2A). Two gametophytes had approximately 8n genomecontent (Figure 2B), another two had approximately 9n and nine gametophytes hadapproximately 10n genomes (Figure 2C).
Eight out of nine 10n gametophytes produced antheridia (Figure 3A), however,swimming antherozooids were not observed. Archegonia were not observed on anygametophytes during three years of culture. Apogamous sporophytes were induced fromtwo 10n gametophytes after one year of culture (Figure 3B). They grew to approximately10 mm in height but then died.
DISCUSSIONManton (1950) made a cytological study of the pteridophyta, and following her, similarinvestigations on pteridophyta were carried out by many researchers (e.g. Wagner, 1954;Lovis, 1964; Sleep, 1966, 2014*; Reichstein, 1981; Pinter, 1995; Ekrt & Koutecky,2016). In these studies, various suggestions have been proposed about differentiationand development in ferns through observations of meiosis in hybrid species.
Generally speaking, meiosis of sporophytes with many univalent chromosomes isirregular and spores produced are mostly abortive. If fertile spores could be obtainedfrom those sporophytes, one might consider whether they are reduced spores withaneuploid chromosome numbers, as reported in triploid Osmunda regalis L. (Manton,1950) or unreduced spores with the same chromosome number as the mother plant,produced by the pathway of Döpp-Manton or Braithwaite (Manton, 1950; Braithwaite,1964; Walker, 1979; Kato, 1997; Kawakami et al., 2003). In the present study, however,the genome contents of gametophytes developed from spores of the pentaploid C. fragiliswere greater than those of the donor pentaploid sporophyte and some gametophytes had
*Sleep examined Polystichum and proposed her ideas on the development of this genus. Her studieswere not limited to Polystichum but also extended to other ferns. Because of her early death, thisstudy (Sleep, 2014) was published posthumously.
KAWAKAMI ET AL: CYSTOPTERIS FRAGILIS 151

10n contents that double the genome contents of the donor sporophyte. Sincegametophytes develop directly from spores, their genome content must be consideredto be the same as that of the spore from which an individual is derived. The results,therefore, suggest that spores with a higher ploidy level than the mother sporophyte are,surprisingly, produced in the pentaploid C. fragilis. The formation of gametophytes withgenomes doubled that of the donor sporophyte is reported here for the first time.
From the observation of meiosis, it might be considered that spores with 10n genome
152 FERN GAZ. 20(4):149-155. 2016
Figure 2. Relative DNA content of gametophytes derived from spores of the pentaploidCystopteris fragilis, A: distribution of relative DNA content of 13 gametophytes; B:sample of gametophytes with approximately 8n genome contents; C: Sample ofgametophytes with approximately 10n genome contents.

contents are from monad spores produced in sporangia. They might be produced withoutmeiotic cell division from the spore mother cells with doubled genome contents. Whenmonad spores are produced, restitution nuclei might occur; and if some chromosomesare lost when they occur, monad spores with genomes smaller than 10n, for example, 9nor 8n, might be produced. However, in the formation of spores with 9n or 8n genomecontents, especially in the case of 8n spore formation, another pathway might beconsidered. Since in sporogenesis dyad spores consisting of large and small spores wereobserved in the present study, the larger spore with 8n might be fertile, though the smallerspore with 2n might be sterile. Although dyad spores are well known to be produced inmany ferns, they are unreduced spores with the same genome contents as the donorsporophyte (Walker, 1979; Kato, 1997; Kawakami et.al., 2003). Why fertile 5n sporeswere not obtained in this study is unknown. Further studies of fertile spores may berequired to understand sporogenesis of the pentaploid sporophyte. The present studyrevealed the monad spore formation in the pentaploid C. fragilis.
It is well known that polyploidy is one of the outstanding cytological features in ferns(Manton, 1950; Wagner, 1954; Lovis, 1968; Walker, 1979; Kato, 1997). The highestpolyploid fern in Japan is 10x (Nakato, 1987; Takamiya, 1996) and the highest polyploidin the world is reported to be 16x (Walker, 1979). The evolutionary process of howpolyploid ferns such as 10x or 16x are produced has not been investigated in detail. Sinceunreduced spores are well known to play a very important role for polyploid formation(Gastony, 1986; Kato, 1997), the pathway of the decaploid sporophyte (10x) formationcould well be that they are produced by fertilization of pentaploid gametes producedfrom pentaploid gametophytes derived from unreduced spores produced in pentaploidplants. The present study, however, may well indicate another method of decaploidsporophyte formation: the pentaploid sporophytes produce decaploid spores and fromdecaploid gametophytes developed from spores, decaploid sporophytes could be inducedapogamously. Furthermore, if antherozooids with an ability of fertilization could beproduced from the decaploid gametophytes, though these were not observed in thepresent study, it might be possible to produce plants with higher polyploid levels thandecaploid, such as 12x plants, by fertilization between decaploid male gametes andfemale diploid gametes derived from tetraploid sporophytes. Whether the fertile sporesproduced in pentaploid sporophytes can play a role in the formation of higher polyploid
KAWAKAMI ET AL: CYSTOPTERIS FRAGILIS 153
Figure 3. An antheridium with antherozooids (A) and apogamous sporophytes (B)produced on gametophytes with approximately 10n genome content in Cystopterisfragilis. Scale bars: A: 25 μm; B: 1 mm.

C. fragilis in nature or not is quite intriguing. If decaploid C. fragilis were discovered inMongolian Altai, it might have been produced from decaploid gametophytesapogamously. To understand the evolution of polyploidy in these ferns, further studieson C. fragilis in Mongolian Altai may be necessary. .
ACKNOWLEDGEMENTThe study was supported by a Grant-in-Aid for Scientific Research Program (A)19255004 (the representative: Katsuhiko Kondo) from the Japan Society for thePromotion of Science.
REFRENCESBRAITHWAITE, A.F. 1964. A new type of apogamy in ferns. New Phytol. 63: 293-305.EKRT, L. & KOUTECKY, P. 2016. Between sexual apomictic: unexpectedly variable
sporogenesis and production of viable polyploids in the pentaploid fern of theDryopteris affinis agg. (Dryopteridaceae). Annals of Botany 117:97-106.
GASTONY, G.J. 1986. Electrophoretic evidence for the origin of a fern species byunreduced spores. Am. J. Bot. 73:1563-1569.
KATO, M. 1997. Diversity and evolution of land plants. Shokabo Press, Tokyo, pp. 75-104.
KAWAKAMI, S. M., KAWAKAMI, S., KONDO, K., KATO, J. & ITO, M. 2003.Sporogenesis in haploid sporophytes of Osmunda japonica (Osmundaceae). J PlantSci. 164:527-534.
KAWAKAMI, S.M., KAWAKAMI, S., KATO, J., KONDO, K., SMIRNOV, S.V. &DAMDINSUREN, O. 2010. Cytological study of a fern Cystopteris fragilis inMongolian Altai. Chromo. Bot. 5:1-3.
LOVIS, J.D. 1964. Autoplyploidy in Asplenium. Nature 203: 324-325.LOVIS, J.D. 1968. Artificial reconstruction of a Species of fern, Asplenium adulterinum.
Nature 217: 1163-1165.LOVIS, J.D. 1977. Evolutionary patterns and processes in ferns. Adv Bot Res 4: 229-
415.MANTON, I. 1950. Problems of cytology and evolution in the Pteridophyta. Cambridge
University Press, Cambridge.MURASHIGE, T. & SKOOG, F. 1962. A revised medium for rapid growth and bioassays
with tobacco tissue culture. Physiol Plant 15: 473-497.NAKATO, N. 1987. Chromosome numbers of three endemic species of the fern genus
Blechnum in Japan. J. Jap. Bot. 62: 261-267.PINTER, I. 1995. Progeny studies of the fern hybrid Polystichum ×bicknellii
(Dryopteridaceae: Pteridophyta). Fern Gaz. 15(1): 25-40.REICHSTEIN, T. 1981. Hybrids in European Aspleniaceae (Pteridophyta). Bot. Helv.
91: 89-139.SLEEP, A. 1966. Some cytotaxonomic problems in the fern genera Asplenium and
Polystichum. Unpublished Ph.D. thesis, University of Leeds.SLEEP, A. 2014. Hybridization in Polystichum (Dryopteridaceae: Pteridophyta). Fern
Gaz. 19(8): 319-341.TAKAMIYA, M. 1996. Index to chromosomes of Japanese pteridophyta (1910-1966).
Japan Pteridological Society.WAGNER, W.H. 1954. Reticulation evolution in the Appalachian Asplenium. Evolution
8: 103-118.
154 FERN GAZ. 20(4):149-155. 2016

WALKER, T.G. 1979. Experimental biology of ferns. In: DYER, A.F. (ed.) Thecytogenetics of ferns. Academic Press, London, pp. 87-132.
WALKER, T.G. 1984. Chromosomes and evolution in Pteridophytes. In: SHARMA, A.F.& SHARMA, A. (eds) Chromosomes in evolution of eukaryotic groups. CRC, BocaRaton, pp. 103-141.
KAWAKAMI ET AL: CYSTOPTERIS FRAGILIS 155

SHORT NOTE
TWO UPDATES FOR GRAMMITID FERNS (POLYPODIACEAE) ONTHE ‘FERNS OF THAILAND’ WEBSITE
The Ferns of Thailand website (http//rbg-web2.rbge.org.uk/thaiferns/Pages/thai-fern-list.htm), updated 23 June 2016, has one wrongly named set of images and one speciesthat has been re-identified amongst the grammitid ferns (Polypodiaceae).
The on-line image of Prosaptia celebica is not of that species, but belongs toCtenopterella blechnoides: the voucher specimen is Middleton et al. 5332 (E00700990!).The red-brown rhizome scales characteristic of most species of Ctenopterella can beseen in one of the images; in detail they are concolorous and glabrous. The rhizomescales of Prosaptia celebica are subclathrate to clathrate, medium to dark red-brown andciliate.
Thailand material formerly identified as Prosaptia alata (Middleton 4400E00700991!, Middleton E00690558!) has been re-identified as P. serriformis. The formerhas the rachis prominent and concolorous on the adaxial surface and sori in cylindricalpouches, while the latter has the rachis slightly prominent to slightly sunken andconcolorous to darker on the adaxial surface and sori in hemispherical or conical pouches.Peninsular Malaysian material of P alata has also been re-identified as P. serriformis.
ACKNOWLEDGEMENTSI wish to thank the Curator of the herbarium, Royal Botanic Garden Edinburgh forpermission to examine specimens of Grammitids in July 2016.
Barbara S. ParrisFern Research Foundation, 21 James Kemp Place, Kerikeri, Bay of Islands,
New Zealand 0230Email: [email protected]
156 FERN GAZ. 20(4). 2016

TWO NEW RECORDS FOR COLOMBIA OF PSILOTUM NUDUM(PSILOTACEAE) FROM THE MAGDALENA VALLEY AND IN THE
ANDEAN AMAZONIAN FOOTHILLS
D. SANÍN1, 2 & V. TORREZ3
1 Universidad de la Amazonia, Programa de Biología, Calle 17 diagonal 17 con carrera3F, Barrio el Porvenir, Florencia-Caquetá, Colombia;
2 Current address: Instituto de Ciências Biológicas, Departamento de Botânica,Universidade Federal de Minas Gerais, Caixa Postal 486, 30123-970, BeloHorizonte, Minas Gerais, Brazil. Email: [email protected];
3 Herbario Nacional de Bolivia, Instituto de Ecología, Universidad Mayor de SanAndrés, Cotacota, calle 27, Campus universitario, La Paz, Bolivia. Email:[email protected].
Key words: Inconspicuous species, Caldas, Caquetá, ferns, distribution.
ABSTRACTDespite its wide distribution, Psilotum nudum has been recorded previously onlyfrom the western slopes of the Occidental Andean Cordillera of Colombia,particularly in the Chocó Region, and in the Caribbean Golf of Urabá. As a resultof recent exploration, we here expand the distribution of P. nudum in Colombiaby reporting two new records from the Central and Oriental Andean Cordilleras.
INTRODUCTIONPsilotum nudum (L.) Beauv. is a remarkable fern taxon and the simple aerial axis waswidely regarded as a possible ancestral shoot design for sporophyte land plants (Figure1) (Bierhorst, 1977; Gensel, 1977; Schulte et al., 1987). The relationships amongOphioglossales, Marattiales, Psilotales and extinct taxa (Cladoxylales, Zigopteridales,Iridopteridales, Stauropteridales) remain highly ambiguous (Gensel, 1977; Bateman etal., 1998). However, based on more recent molecular analyses, it is currently classifiedin the family Psilotaceae as a sister group of Ophioglossaceae, with both included in theclass Psilotopsida (Smith et al., 2006). These molecular results are supported by evidencefrom morphology, for example, both families have cylindrical subterranean, non-green,mycorrhizal gametophytes and possess sporangia that are born on the adaxial surface ofthe leaf (Moran, 2004).
Psilotum nudum has a wide distribution, which is principally pantropical (Tryon &Tryon, 1982; Mickel & Smith, 2004), but it can be distributed in the subtropics ofAustralia and New Zealand (AVH, 2016) and in the more temperate zones of Spain(Molesworth-Allen, 1966; Galan et al., 1996). It can be found from sea level up to 1840m(Idárraga-Piedrahita et al., 2014; Murillo et al., 2016; TROPICOS, 2016). In America,it has been recorded from the south of the United States of America, Mexico, CentralAmerica, Colombia, Venezuela, Guayanas, Ecuador, Peru, Bolivia, Brazil, Paraguay,Argentina and in the Antilles (Mickel & Smith, 2004; Murillo et al., 2008; Moran 2011).It can be found either in pristine forest or urban zones (Moran, 2011).
The taxon may have few records in Colombia due to the fact that it does not looklike a fern (Moran, 2004) and it prefers to inhabit places in the shade (Galan et al., 1996).Both decrease the probability of its discovery, especially for the non-trained botanist. In
FERN GAZ. 20(4):157-162. 2016 157

Colombia, until now, it was recorded only from four localities in the OccidentalCordillera and in the Caribbean Golf of Urabá, principally in the Chocó Region (Table1, Figure 2).
As a result of a botanic exploration in the Magdalena Medio region and in the AndeanAmazonian foothills, it was discovered growing as an epiphytic fern in two new locations(Table 1, Figure 2). However, the habit of the plant can be epiphytic, terrestrial or
158 FERN GAZ. 20(4):157-162. 2016
Figure 1. Psilotum nudum. A: Habit of the plant, scale bar (2 cm). B: Stem and leaves,scale bar (5 mm). C: Sporangia, scale bar (1 mm). D: Ramification of the rhizome, scalebar (5 mm). All from D. Sanín & J. L. Peña 6046 (COL). Photos by J. L. Peña.
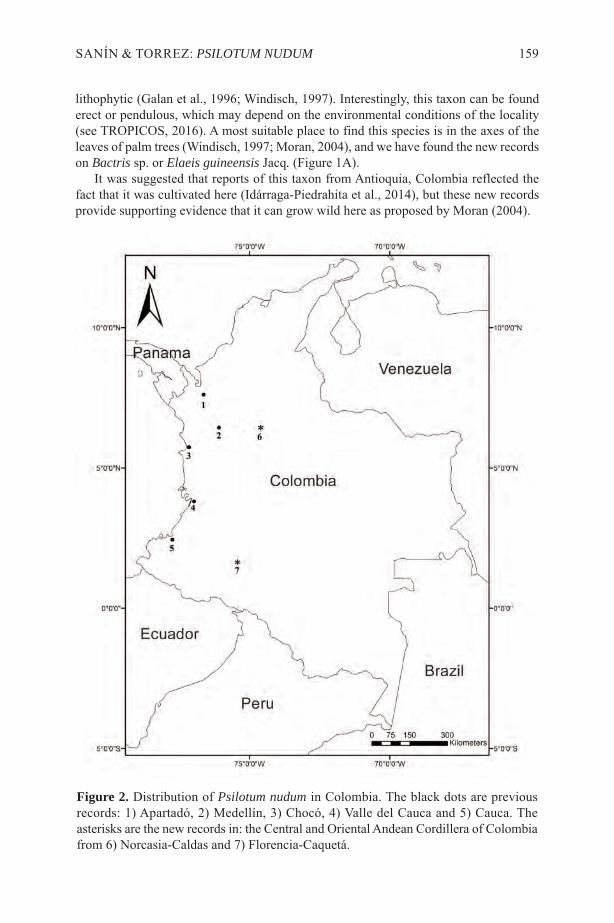
lithophytic (Galan et al., 1996; Windisch, 1997). Interestingly, this taxon can be founderect or pendulous, which may depend on the environmental conditions of the locality(see TROPICOS, 2016). A most suitable place to find this species is in the axes of theleaves of palm trees (Windisch, 1997; Moran, 2004), and we have found the new recordson Bactris sp. or Elaeis guineensis Jacq. (Figure 1A).
It was suggested that reports of this taxon from Antioquia, Colombia reflected thefact that it was cultivated here (Idárraga-Piedrahita et al., 2014), but these new recordsprovide supporting evidence that it can grow wild here as proposed by Moran (2004).
SANÍN & TORREZ: PSILOTUM NUDUM 159
Figure 2. Distribution of Psilotum nudum in Colombia. The black dots are previousrecords: 1) Apartadó, 2) Medellín, 3) Chocó, 4) Valle del Cauca and 5) Cauca. Theasterisks are the new records in: the Central and Oriental Andean Cordillera of Colombiafrom 6) Norcasia-Caldas and 7) Florencia-Caquetá.

160 FERN G
AZ. 20(4):157-162. 2016
Localities Altitude (m) Collection Collection date Bibliography
Antioquia: Apartadó yMedellín. 0-1500 J. Santa 378 (HUA) Not provided Idárraga et al. (2014).
Chocó: Nuqui-Pangui.Playa La Olimpica. 0 A. Gómez et al. 500 (MO) 14 Jan. 1992 TROPICOS (2016).
Cauca: Guapi, Orillas delrío Guapi: Población deGuapí.
5 B. R. Ramírez P. 17461(CAUP) 16 Nov. 2003 Ramírez & Macias-Pinto
(2007).
Valle del Cauca:Buenaventura, Bahía deBuenaventura.
0 E. P. Killip 35510 (COL) 05 Apr. 1939 Murillo et al. (2016).
Caldas: Caldas, Norcasia,Vereda Quiebra de Roque,Reserva Riomanso,desviación a manoizquierda de la Quebrada laTostada.
224
D. Sanín, A. F. Bohórquez-O., J. Ramírez-G., V.Ramírez-V. & J. P. Betancur5539 (HUA)
2 Dic. 2011 Current record
Caquetá: Florencia, JardínBotánico Uniamazonia,Sendero de la Anaconda.
269 D. Sanín & J.L. Peña 6046(COL) 14 Jul. 2015 Current record
Table 1. Records of Psilotum nudum in Colombia.

The discovery of these populations of Psilotum nudum in Caldas and Caquetádepartment expand its distribution to the Central and Oriental Andean Cordilleras ofColombia (Figure 2), and suggest that the plant may have a wider distribution inColombia. However, due the difficulties of recording it, its real distribution is still notwell known.
ACKNOWLEDGEMENTSDS would like to thank the University of the Amazonia for the funding support. DS isindebted to the Biology program, especially to Professor Alexander Velasquez-Valenciaand the staff of the Natural History Museum of the University of the Amazonia forproviding infrastructure to develop the research. The photographs were kindly providedby J.L. Peña. We thank Professor E. Trujillo Trujillo, for generously providing literatureand support in the initial phases of this research. We are grateful to Professor Mary Gibbyfor her kind comments that improved our manuscript.
REFERENCESAVH. 2016. Australia’s Virtual Herbarium, Council of Heads of Australasian Herbaria,
<http://avh.chah.org.au>, accessed <11 May. 2016>.BATEMAN, R.M., CRANE, P.R., DIMICHELE, W.A., KENRICK, P.R., ROWE, N.P.,
SPECK T. & STEIN, W.E. 1998. Early evolution of land plants: Phylogeny,physiology and ecology of the primary terrestrial radiation. Annual Rev. Ecol. Syst.29: 263–92.
BIERHOST, D.W. 1977. The systematic position of Psilotum and Tmesipteris. Brittonia29 (1): 3–13.
GALAN DE MERA, A., VICENTE ORELLANA, J.A., GONZALEZ, J.L. &FERNANDEZ LUNA J.C. 1996. New populations of Psilotum nudum in SW Europe(Psilotaceae: Pteridophyta). Fern Gaz. 15(3): 109–112.
GENSEL, P.G. 1977. Morphologic and taxonomic relationships of the Psilotaceaerelative to evolutionary lines in early land vascular plants. Brittonia 29 (1): 14–29.
IDÁRRAGA-PIEDRAHITA, A., ORTIZ, R.D.C., CALLEJAS POSADA, R. &MERELLO, M. 2014. Listado de las plantas vasculares del departamento deAntioquia. Pp. 127-939. In: IDÁRRAGA-PIEDRAHITA, A., ORTIZ, R.D.C.,CALLEJAS POSADA, R. & MERELLO, M. (Eds.) Flora de Antioquia: Catálogode las plantas vasculares. Vol II. Universidad de Antioquia y Jardín Botánico deMisouri & Gobernación de Antioquia. Editorial D’Vinni, Bogotá, Colombia.
MICKEL, J.T. & SMITH, A.R. 2004. The pteridophytes of Mexico. Mem. New YorkBot. Gard. 88: 1–1054.
MOLESWORTH-ALLEN, B. 1966. Psilotum nudum in Europe. Brit. Fern Gaz. 9: 249-251.
MORAN, R.C. 2004. Natural History of Ferns. Timber Press. 301 Pp.MURILLO-P., M. T., MURILLO-A., J.C. & LEÓN-P, A. 2016-07-29. Psilotum nudum
(L.) P.Beauv. En: BERNAL, R., GRADSTEIN, S.R. & CELIS, M. (Eds.). 2015.Catálogo de plantas y líquenes de Colombia. Instituto de Ciencias Naturales,Universidad Nacional de Colombia, Bogotá.http://catalogoplantasdecolombia.unal.edu.co
MURILLO-P., M.T., MURILLO-A., J.C. & LEÓN-P., A. 2008. Los Pteridofitos deColombia. Biblioteca José Jerónimo Triana No.18. Universidad Nacional deColombia. Arfo Editores e Impresos LTDA.
SANÍN & TORREZ: PSILOTUM NUDUM 161

RAMÍREZ PADILLA, B.R. & MACÍAS PINTO, D.J. 2007. Catálogo de helechos yplantas afines del Departamento del Cauca. Universidad del Cauca, Popayán, 215 p.
SCHULTE, P.J., GIBSON, A.C. & NOBEL, P.S. 1987. Xylem anatomy and hydraulicconductance of Psilotum nudum. Amer. J. Bot. 74 (9): 1438–1445.
SMITH, A. R., K. M. PRYER, E. SCHUETTPELZ, P. KORALL, H. SCHNEIDER & P.G. WOLF. 2006. A classification for extant ferns. Taxon 55 (3): 705–731.
TROPICOS. 2016. Missouri Botanical Garden. Accessed: 29 Jul 2016<http://www.tropicos.org/Name/26602814>
TRYON, R.M. & TRYON, A.F. 1982. Ferns and Allied Plants. Springer-Verlag. NewYork, Heidelberg, Berlin, p. 783–786.
WINDISCH, P.G. 1997. Pteridófitas do Estado de Mato Grosso: Psilotaceae. Bradea 8(10): 57–60.
162 FERN GAZ. 20(4):157-162. 2016

SHORT NOTE
TWO NEW COMBINATIONS IN GRAMMITID FERNS(POLYPODIACEAE): PROSAPTIA HORNEIAND RADIOGRAMMITIS
SETULIFERA
Two new combinations in Asian-Pacific Grammitid ferns are proposed: Polypodiumhornei (Baker) Parris and Radiogrammitis setulifera (Alderw.) Parris. A lectotype ischosen for Polypodium hornei Baker. Herbarium abbreviations follow Thiers (2016).
Prosaptia hornei (Baker) Parris, comb. nov., based on Polypodium hornei Baker, J.Bot. 17: 298 (1879). Type: Fiji, Viti Levu between Nadrau & Babuca, c. 1000 ft alt.,Horne 369 (lectotype K! barcode 001044319, chosen here). Syn. Ctenopteris hornei(Baker) Brownlie, Pacific Sci. 14: 402 (1960), Tomophyllum hornei (Baker) Parris, KewBull. 65(1): 124 (2010).
Baker’s description mentions two localities for the species, “Nadrau to Babuca, VitiLevu”, and “mountains of Ovalau”. The type sheet at Kew bears two plants; the left handone (K001044319) is well pressed and bears a label beneath it reading “on trees Nadrauto Babuca”. This plant is chosen as the lectotype. The right hand plant (K000604741)has rolled up fronds with the pinnae pressed together to obscure their abaxial surfacesand it has no label indicating its provenance.
Baker (1879) and Brownlie (1977: 359) both describe the rhizome scales of thespecies as ciliate, a character common to Prosaptia and some species of Tomophyllum.The type material of Polypodium hornei has a dorsiventral rhizome with stipes in tworows and articulated to prominent phyllopodia, and it also lacks hydathodes on the veinendings on the adaxial surface of the lamina, all of which are characters of Prosaptia.Brownlie’s illustration of part of the frond (1977: 355, pl. XLI, 3, as Ctenopteris hornei)shows the crenulate pinna margin often found in Tomophyllum, but very rarely inProsaptia, and material collected by him in CHR has the radial stipe arrangement ofTomophyllum. Nakamura & Kokubo (2008: 256, pl. 99, 1) illustrate (as Ctenopterishornei) a plant that is obviously a species of Tomophyllum; it has the evident hydathodeson the vein endings on the adaxial surface of the lamina characteristic of Tomophyllum,the same slightly crenulate lamina margin that Brownlie illustrated, and rhizome scaleswith a single apical hair that are found in numerous species of Tomophyllum. Clearly,two species have been confused under the name Ctenopteris hornei; Polypodium hornei= Prosaptia hornei, and an un-named species of Tomophyllum. The confusion is partlydue to the fact that Prosaptia hornei is one of a small group of Prosaptia species thathave superficial sori (like Tomophyllum), as opposed to sori sunken in steep-walleddepressions on the abaxial surface of the lamina or in marginal to submarginal pouchesthat are typical of Prosaptia.
Radiogrammitis setulifera (Alderw.) Parris, comb. nov., based on PolypodiumsetuliferumAlderw., Bull. Jard. Bot. Buitenzorg, sér. 2, 16: 32 (1914). Syntypes: Malacca, Matthew 408 (BO); Sumatra, Barisan Mts, Matthew 651 [657 in K](BO, K000501091!), Jan. 1913, Matthew s. n. (E00194034! p. p.). Syn. P. pumilumBrause. Bot. Jahrb. Syst. 49: 38 (1912) non Cockayne (1909). Xiphopteris setulifera(Alderw.) Parris, Kew Bull. 41(1): 69 (1986); Oreogrammitis setulifera (Alderw.) Parris,Gardens’ Bull. 58(2): 268 (2007).
FERN GAZ. 20(4). 2016 163

Detailed examination of the rhizomes of Matthew s. n in E and Matthew 657 in Kshows that they are radial rather than dorsiventral, thus the species belongs toRadiogrammitis, rather than Oreogrammitis. The species is very similar in frondindumentum and dissection to R. subpinnatifida (Blume) Parris, but differs in havingfewer and much shorter stipes arranged in a series of looser spirals or whorls on therhizome.
ACKNOWLEDGEMENTSI wish to thank the Keeper of E for loan of material, the Keeper of K for permission toexamine the holdings of Asian and Pacific Grammitids during July 2016, and EwenCameron (AK) for expediting the loan from E.
REFERENCESALDERWERELT VAN ROSENBURGH, C. R. W. K. 1914. New or interesting Malayan
ferns 6. Bull. Jard. Bot. Buitenzorg, sér. 2, 16: 1-60.BAKER, J. G. 1879. On a collection of ferns gathered in the Fiji Islands by Mr. John
Horne, F.L.S. J. Bot. 17: 292-300.BROWNLIE, G. 1977. The Pteridophyte Flora of Fiji. Nova Hedwigia, Beih. 55. J.
Cramer, Vaduz.NAKAMURA, T. & KOKUBA, K. 2008. Polypodiaceae. In KATO, M., NAKAMURA,
T., MATSUMOTO, S. & EBIHARA, A. (Eds) Illustrated Flora of Ferns & Fern Alliesof South Pacific Islands, pp. 297. National Museum of Nature and Science, Tsukuba.
THIERS, B. (2016). Index Herbariorum. New York Botanical Gardens, on-line.
Barbara S. ParrisFern Research Foundation, 21 James Kemp Place, Kerikeri, Bay of Islands,
New Zealand 0230Email: [email protected]
164 FERN GAZ. 20(4). 2016

BOOK REVIEW
FERN NAMES AND THEIR MEANINGS - 2nd EDITION. J.W. Dyce, revisedand expanded by J. Edgington. 2016. 59 pp. ISBN O-9509806-1-7. Published by TheBritish Pteridological Society. Price £5.
The scientific names of plants are often puzzling and their meanings frequently obscure.Many users of these names find their translation and origin fascinating. Findinginformation regarding the names of ferns and particularly the myriad of cultivarapplications is especially difficult. J.W (Jimmy) Dyce remedied this situation with thepublication of this small book in 1988. Proving to be very popular it was reprinted in1994 but has been difficult to obtain for several years. It is therefore a great pleasure toreceive this new edition, greatly expanded and enhanced by Professor John Edgington.
Compared with the first edition, Dyce’s original text is mostly retained unaltered,however there are significant additions that considerably improve the utility of the book.Starting with the introduction Dyce gives a brief explanation of history of using Latinfor the naming of plants and why it is still used in the modern era. He also alludes to theparticular problem of fern cultivar nomenclature, which even today does not seem to beresolved to the satisfaction of all. (The problem arises because many fern cultivar namesare actually brief descriptions or polynominals, and were abandoned in other areas ofbotanical nomenclature centuries ago). The next section, on the etymology of Britishfern generic names, has been fully revised and expanded by Professor Edgington. It nowincludes a brief statement regarding the origin of the name as well as the meaning.(Lycophytes were ignored by Dyce in the first edition but are happily now includeddespite making the book’s title slightly imprecise).
FERN GAZ. 20(4). 2016 165

The next chapter is completely new, an etymology of non-British genera includingall those found in Europe and many that are found in cultivation. I found this partparticularly interesting. As in the previous section, both the meaning and origin of thename is given. The next two sections, both helpful and informative, listing the wordelements at the beginning and end of fern names, are left unrevised.
The next chapter, again new, comprehensively covers most of the specific names ofEuropean ferns including hybrids. This is a most useful addition and many obscure namesare included, the exceptions being a large number of Asplenium hybrids, many onlyhaving been found once.
In 1984 Dyce published an updated list of British ferns and their current names andthis has been included here, suitably revised taking into account recent research. Underthe same heading, Fern nomenclature – old and new, are two new lists. The first coversnon-native species that have recorded growing in the wild in the British Isles and thesecond is a comprehensive list of synonyms of British ferns and lycophytes. Both theselists are of great interest and will prove to be very useful.
There then follows the largest section of the book, a glossary of fern varietal names.This formed the major part of the first edition and is included here unchanged. Many ofthe terms are also widely used in species names so it is not restricted to horticulturalvarieties. Finally there is a new, brief glossary of technical terms and an updated list ofreferences.
As I hope will be appreciated from this review this is a much improved publicationwhere John Edgington has managed to retain the content and spirit of Jimmy Dyce’soriginal but also sought to include much additional relevant and interesting information.He has achieved this most successfully and with the book priced at just £5 it must rankas one of the bargains of the year.
T. Pyner
166 FERN GAZ. 20(4). 2016

FERN GAZ. 20(4). 2016 167

INSTRUCTIONS FOR AUTHORS
PAPERS should not usually exceed 20 printed pages and are generally expected to beconsiderably shorter. As well as reports of original research, review articles and shortnotes, e.g. new records, are encouraged. The senior author should supply an email addressto facilitate correspondence. REVIEWS should cover recent developments in any aspect of research on ferns andlycopods. This can include unpublished research by the author, and/or the possibledirection of future research. They should be aimed at the general reader who may not bean expert in the chosen topic. Proposals for a review should be discussed first with thereview editors.MANUSCRIPTS should be submitted in English (British) in electronic format in10-point Times New Roman font and double spaced. Electronic versions of text andtables should be compatible with WORD, with figures as pdf or jpg files, and sent asemail attachments or CDroms. All manuscripts will be refereed.THE TITLE should reflect the content of the paper and be in BOLD CAPITALS(11-point) and centrally aligned. Generic and specific names should be in italics and anytitle containing a generic or specific name should be followed by the family name inbrackets e.g.
TRICHOMANES SPECIOSUM (HYMENOPHYLLACEAE)IN SOUTHERN SPAIN
AUTHOR ABBREVIATIONS should follow Pichi Sermolli’s (1996) Authors ofscientific names in Pteridophyta, Royal Botanic Gardens, Kew.MAIN HEADINGS: should be in BOLD CAPITALS (10-point) and centrally aligned.SUBSIDIARY HEADINGS: should be in bold, the first letter of each word in capitals,the rest in lower case and left-aligned.AUTHORS’ NAMES AND FULL ADDRESSES (including email address): followthe title and are centrally aligned.KEY WORDS: up to ten.ABSTRACT: should reflect the content of the paper.FIGURES: there is no distinction between photographs and line drawings in numbering.All should be presented in a form ready for reproduction, colour, TIF format and 300dpi (please contact editor with queries), with a scale bar where appropriate. Lettering ornumbers (Arabic) should be in the bottom left using uppercase Times Roman. Figurecaptions should be on a separate sheet.TABLES: can be printed in either portrait or landscape format. Authors should considerthis when preparing tables. Authors should ensure that tables fit the printed page size ina legible form.PAGESIZE: The Gazette has a useable page size of 123 × 204 mm. This can be used inportrait or landscape mode but any supplied artwork should not be larger than this.MEASUREMENTS: follow the metric system.CHECKLISTS: follow the format of Baksh-Comeau, Fern Gaz. 16(1, 2): 11- 122.REFERENCES: follow the style of a recent issue of The Fern Gazette, e.g.:-HOOKER, W.J. 1864. Species Filicum, 5. Dulau & Co., London.MORTON, C.V. 1947. The American species of Hymenophyllum, section
Sphaeroconium. Contr. U.S. Natl. Herb. 29(3): 139-201.STEVENSON, D.W. & LOCONTE, H. 1996. Ordinal and familial relationships of
pteridophyte genera. In: CAMUS, J.M., GIBBY, M. & JOHNS, R.J. (Eds)Pteridology in perspective, pp. 435-467. Royal Botanic Gardens, Kew.
JOURNAL ABBREVIATIONS: should follow Botanico Periodicum Huntianum &Supplements.Alterations from the original text at proof stage will be charged for unless they are minorpoints of detail. A pdf will be provided free to the senior author.
168 FERN GAZ. 20(4). 2016

REVIEW
DIVERSITY AND ADAPTATIONS OF RHEOPHYTIC FERNS
MASAHIRO KATODepartment of Botany, National Museum of Nature and Science, Tsukuba 305-0005,
Japan([email protected])
Key words: adaptive strategy, flood zone, heterochrony, Osmunda, recurrentadaptations, gametophyte, streamlined leaf
ABSTRACTRheophytes grow in the zone on river beds and banks that are regularly floodedafter heavy rains. In the tropics, this zone is of sufficient height (from the lowestto the highest water level) that different rheophytes can occur in the lower andupper subzones. Rheophytic species are obligate or facultative and some woodyplants, rarely climbers, are paedorheophytic. I enumerate 141 rheophytes andpossible rheophytes in 18 families and subfamilies of rheophytic ferns andlycophytes assigned to a variety of families, suggesting recurrent adaptations. Therheophyte morphology is adapted to the habitat and shared among the lineages,and especially the leaves or leaflets are streamlined with cuneate bases andglabrous, entire, tough blades. In leaf development the mesophyll cells andintercellular spaces, together with the epidermal cells, are smaller in size duringa shortened period of ontogeny. The gametophyte of rheophytes, which isindependent of the sporophyte, has a different adaptive strategy from thesporophyte: a shortened duration of generation and subtle morphologicalmodification.
ECOLOGYPlants include 300,000‒330,000 or more species of bryophytes, lycophytes, ferns,gymnosperms and angiosperms (Evert and Eichhon, 2013; Mauseth, 2014). They growin a wide range of habitats other than terrestrial, including alpine, arid, water, mangroveswamp, epiphytic, lithophytic (e.g., on limestone or ultramafic rocks), and other habitats(Crawford, 2008). The habitats are characterized by abiotic factors (temperature,precipitation, light, etc.) and associated organisms. Remarkable organismal associationsare seen in, for example, symbiotic plants (Paracer and Ahmadjian, 2000; Evert andEichhorn, 2013). This paper reviews our understanding of the diversity and adaptationsof ferns that grow in rivers.
A special habitat for plants is provided by rivers. Rivers are often subject to flashfloods after heavy rains, even as frequently as once a week in the tropics, and plantscalled rheophytes grow exclusively on banks and beds of rivers and streams (van Steenis,1981, 1987; Mehtreter, 2008). Van Steenis (1981) defined rheophytes in ecological termsas follows: rheophytes are “confined to the bank and bed and growing there up to floodlevel, but not beyond the reach of regularly occurring flash floods” (Figure 1).Rheophytes are largely tropical, but a few extend into temperate zones, e.g., Hokkaido,in northern Japan (43°N) (Iwatsuki, 1992). Most rheophytes live in the air much longerthan under relatively brief flooded conditions; this is because water levels return to the
FERN GAZ. 20(5):169-179. 2017 169

more normal levels shortly after rains stop or abate. A few other rheophytes arehydrophytic and always grow submerged, as in e.g., Hymenasplenium obliquissimum(Hayata) Sugim. & Sa. Kurata.
Similar habitats are present in seasonally inundated pools, where semiaquatic ferns,such as Platyzoma microphylla R.Br. (=Pteris platyzomopsis Christenh. & H.Schneid.)(Tryon, 1964) and Marsilea species (Kornas, 1988) grow, and in inundated riparian andmarshland forests, which are one of the habitats preferred by, e.g., Osmunda regalis L.(Lellinger, 1985; Page, 1997). However, the lands are covered by still or slow-runningwater.The height of the flood zone, from the normal low level of river water to the highestflood level reached, differs in different climates -- up to 3 metres in humid tropicalBorneo (Figure 1; M. Kato, unpubl. data). There are different preferences of rheophytesfor the tropical flood zone. Some rheophytes, e.g., Ctenitis vilis (Kunze) Ching, grow inthe higher zone, which is subject to rarer and shorter floods, while others, e.g.,Microsorum pteropus (Blume) Copel., prefer the lower zone, which is subject to morefrequent and longer floods. Microsorum paucijugum (Alderw.) K.Iwats. & M.Katoprefers the further lower zone.
Rheophytes are herbaceous or woody. In herbaceous rheophytes, whole plants aresubmerged when flooded, while in tall woody rheophytes only the basal part of the plantis submerged while flooded. In tall paedorheophytes, seedlings and low juveniles arerheophytic and the adults are emergent and thus typically not rheophytic. Alsophilaesmeraldensis R.C.Moran of the tree-fern family Cyatheaceae is regarded as a rheophyte(Moran, 1995), but it may not be paedorheophytic because of the short (up to 80 cm)trunk. Van Steenis (1987) considered Lomagramma species as paedorheophytic, because
170 FERN GAZ. 20(5):169-179. 2017
Figure 1. Habitat segregation of rheophytes and land species on river bank in WestKalimantan, Indonesia. Rheophytic Dipteris lobbiana occupies the flood zone, and landspecies such as D. conjugata Reinw. and Blechnum orientale L. grow above the zone

the juvenile creeping plants are rheophytic and only later climb trees. Further study isnecessary to reveal paedorheophily of ferns.
Rheophytes may be obligate or facultative. Usually, obligate-rheophytic speciescomprise only rheophytic populations, while facultative rheophytes comprise land-dwelling populations and rheophytic populations. Examples of facultative rheophyticferns are Bolbitis heteroclita (C.Presl) Ching in C.Chr. and B. sinuata (C.Presl)Hennipman. Ampelopteris prolifera (Retz.) Copel. and Diplazium esculentum (Retz.)Sw. may also belong to this category.
ADAPTIVE MORPHOLOGYThe impact of flash floods to plants is considerable because non-rheophytic land plantscannot invade the flood zone (Figure 1). Rheophytes have gained particular morphologiesto resist or reduce pressure of flooding, although their leaves may be damaged byflooding (Sharpe, 1997). The shoot-branches and leaf-petioles are flexible or tough withdeveloped bark in, e.g., Salix or with dense networks of cell walls in Osmunda (Imaichiand Kato, 1992). The roots adhere tightly to the river bank and bed. The most remarkablefeature of rheophytes is leaf morphology (van Steenis, 1981). The leaves or leaflets arestreamlined (usually narrow-lanceolate) with cuneate bases (Figure 2), entire or onlyminutely toothed at the margins, and hairless or sparsely hairy. Cuneate leaf-bases areparticularly important because the bases are under pressure from currents.Thelypteridaceae, a large family generally characterized by having hairy leaves, has thelargest number of rheophytes in ferns (Appendix), but the hairs of rheophytic species inthis family are usually sparse, short, or antrorse (ascending or appressed toward the distalend of the lamina or pinnae). In paedorheophytes, only the leaves of juvenile plants arestreamlined.
Sharpe (1997) found that in the Puerto Rican rheophyte Meniscium angustifoliumWilld. the leaf growth rate is faster than in other tropical land ferns, and the leaf life-spanis shorter. The faster growth rate would reduce exposure of the fragile uncoiling leaves
KATO: DIVERSITY AND ADAPTATIONS OF RHEOPHYTIC FERNS 171
Figure 2. Pinnules. A. Land-living Osmunda japonica. B. Rheophytic O. lancea.

to flood, and the shorter life span would allow escape from stresses of flooding. The leafphenology of rheophytic ferns should be further investigated to clarify the variousadaptations (see also section “Adaptations”).
DIVERSITYRheophytic ferns are most abundant in tropical areas with heavy rainfall, and van Steenis(1981, 1987) enumerated 67 rheophytic ferns and lycophytes. Kato et al. (1991) foundabout 40 species of rheophytic ferns from Borneo. It indicates that the ever-wet Borneois remarkably species-rich for rheophytes. At present 141 rheophytic fern taxa are knownin the world, and these are assigned to 18 families and subfamilies (Appendix 1). Thereare other species that require further study with respect to whether they are rheophyticor not.
In many fern genera, only one or a few rheophytic species are known, while thereare multiple rheophytes in other genera. This trend is obvious also in flowering-plantrheophytes, believed to be about 800 (0.3% of the total number) species belonging to 68(17%) families (van Steenis, 1981, 1987). The scattered distribution of rheophytes overmany lineages suggests that derivation of rheophytes from land-growing ancestors ismarkedly recurrent. Some rheophytes are secondarily derived from other rheophyticspecies in the same genus. I suggest that Microsorum paucijugum (Alderw.) K.Iwats. &M.Kato growing in the lower flood zone is derived from another rheophytic M. pteropus.Kato (1984) considered that two rheophytes, Deparia biserialis (Baker) M.Kato and D.confluens (Kunze) M.Kato, are derived from D. petersenii (Kunze) M.Kato, althoughmolecular evidence to support these relationships is not available (Kuo et al., 2016).Rheophytic fern species are distributed in a wide range of ferns (PPG I, 2016) from theprimitive Osmundaceae through the derived Polypodiaceae, and even in theeusporangiate Ophioglossaceae and the lycophytic family Isoëtaceae. Most fern familieswith rheophytic members are large (> 200 spp.) and tropical (for example,Thelypteridaceae). The most species-rich genus is Bolbitis (14 spp.), which prefers rocksurfaces in wet valleys. I suggest that the evolution of rheophytes happened by chancein the tropics where the rheophytic habitats are abundant. However, the basalleptosporangiate families Osmundaceae and Dipteridaceae are small and comprise fewerthan 20 species.
ADAPTATIONSNarrow streamlined leaves are characteristic of rheophytes (van Steenis, 1981; Vasco etal., 2013). Kato and Imaichi (1992a) examined 32 tropical rheophytic ferns and relatedland species, and found a strong correlation between leaf morphology and anatomy. Themesophyll cells and intercellular spaces are smaller in the narrow leaves than in the broadleaves of related species. Correlated with this, the epidermal cells are also smaller (Katoand Imaichi 1992a). In a comparative developmental study, Imaichi and Kato (1992,1997) showed that the mesophyll cells at the pre-expansion stage are cubical andequal-sized and then expand in both Osmunda japonica Thunb. and O. lancea Thunb.(Figure 3A, E). However, the expansion time-span is less and the mesophyll cell armsare shorter, surrounding the smaller intercellular spaces, in O. lancea (Figure 3). In thisstudy, we interpreted the evolution of the narrow leaves as heterochronic. Kato andImaichi (1992b) found a broad-leaved variant of the rheophytic Tectaria lobbii (Hook.)Copel.in East Kalimantan. This variant grew together with the normal form and differedonly in the broad pinnae with irregular lobes (Figure 4A, C). The broad-leaved variant
172 FERN GAZ. 20(5):169-179. 2017
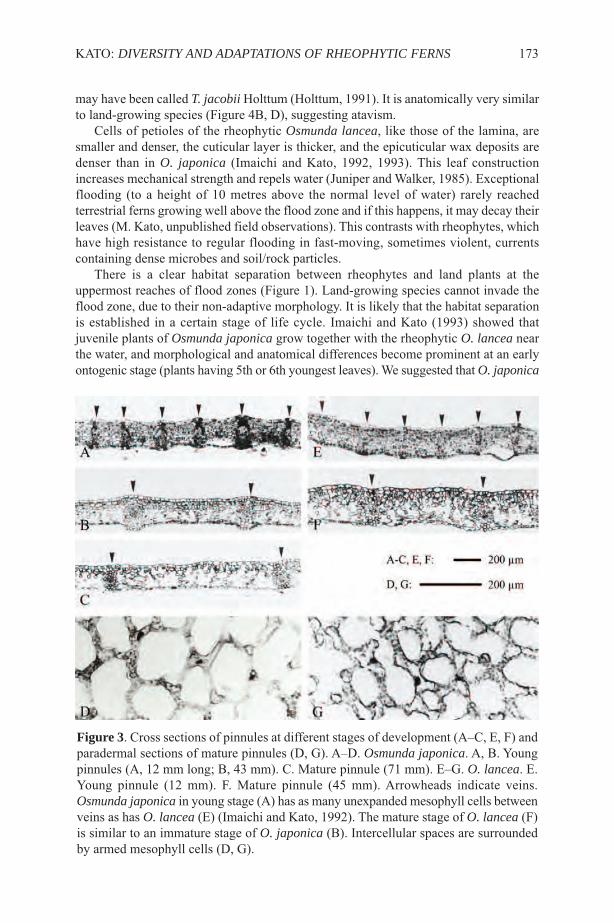
may have been called T. jacobii Holttum (Holttum, 1991). It is anatomically very similarto land-growing species (Figure 4B, D), suggesting atavism.
Cells of petioles of the rheophytic Osmunda lancea, like those of the lamina, aresmaller and denser, the cuticular layer is thicker, and the epicuticular wax deposits aredenser than in O. japonica (Imaichi and Kato, 1992, 1993). This leaf constructionincreases mechanical strength and repels water (Juniper and Walker, 1985). Exceptionalflooding (to a height of 10 metres above the normal level of water) rarely reachedterrestrial ferns growing well above the flood zone and if this happens, it may decay theirleaves (M. Kato, unpublished field observations). This contrasts with rheophytes, whichhave high resistance to regular flooding in fast-moving, sometimes violent, currentscontaining dense microbes and soil/rock particles.
There is a clear habitat separation between rheophytes and land plants at theuppermost reaches of flood zones (Figure 1). Land-growing species cannot invade theflood zone, due to their non-adaptive morphology. It is likely that the habitat separationis established in a certain stage of life cycle. Imaichi and Kato (1993) showed thatjuvenile plants of Osmunda japonica grow together with the rheophytic O. lancea nearthe water, and morphological and anatomical differences become prominent at an earlyontogenic stage (plants having 5th or 6th youngest leaves). We suggested that O. japonica
KATO: DIVERSITY AND ADAPTATIONS OF RHEOPHYTIC FERNS 173
Figure 3. Cross sections of pinnules at different stages of development (A‒C, E, F) andparadermal sections of mature pinnules (D, G). A‒D. Osmunda japonica. A, B. Youngpinnules (A, 12 mm long; B, 43 mm). C. Mature pinnule (71 mm). E‒G. O. lancea. E.Young pinnule (12 mm). F. Mature pinnule (45 mm). Arrowheads indicate veins.Osmunda japonica in young stage (A) has as many unexpanded mesophyll cells betweenveins as has O. lancea (E) (Imaichi and Kato, 1992). The mature stage of O. lancea (F)is similar to an immature stage of O. japonica (B). Intercellular spaces are surroundedby armed mesophyll cells (D, G).

was selected and excluded early from the flood zone and the two species eventuallybecome separated in space and habitat.
In contrast, O. lancea cannot extend its habitat above the flood zone. This restrictionmay be caused by the narrowed leaf area and reduced intercellular space in the mesophylllayer, which may reduce photosynthetic power. This anatomical and the suggestedphysiological character may be a trade-off. A reduced photosynthetic rate was found inthe rheophyte Farfugium japonicum var. luchuense (Masam.) Kitam. (Asteraceae)(Nomura et al., 2006), although it has a different mesophyll anatomy from fernrheophytes (see below). The isolation of the flood zone from the land zone appears strong,but this is not always strong. The diploid hybrid of O. japonica and O. lancea [= O.×intermedia (Honda) Sugimoto] occurs in the interface between both habitats and canreproduce F2 or later offspring (Yatabe-Kakugawa et al., 2013).
Ferns and lycophytes are characterized by the independence of the sporophyte andgametophyte. It is likely that the two generations have different adaptive strategies. Thegametophyte generation is much smaller, simpler, and shorter-lived. There is littlemorphological difference between gametophytes of O. japonica and O. lancea (Hiyamaet al., 1992). Hiyama et al. (1992) discovered that in the rheophytic O. lancea the lifespan of gametophytes is shorter and they interpreted that its gametophyte generation isshortened, relative to land ferns, showing a different strategy from the sporophyte.
Compared with the ferns investigated, the leaves of flowering plant rheophytes showdifferent anatomy. Narrow leaves of the facultative-rheophyte Farfugium japonicum var.
174 FERN GAZ. 20(5):169-179. 2017
Figure 4. Rheophytic Tectaria lobbii. A, B. Normal plants. C, D. Broad-leaved plants.In C, arrowhead indicates normal plants. B, D. Cross sections of pinnules.

luchuense have fewer mesophyll cells than the broad-leaved form (Usukura et al., 1994).Similar anatomy was found in other flowering plant rheophytes (Tsukaya, 2002;Setoguchi and Kajimaru, 2004; Okada and Tsukaya, 2010, 2013; Vermeulen and Tsukaya,2011; Ohga et al., 2012; Matsui et al., 2013). However, a rheophytic orchid ecotypeshows fern-type anatomy (Yorifuji et al., 2015).
EVOLUTIONThe pattern in which rheophytic ferns are scattered in various genera and familiesindicates that similar modes of adaptations produced rheophytes in a number ofindependent lineages (Kato et al., 1991). However, the phylogenetic relationships ofrheophytic ferns have been clarified in only a few cases (e.g., Osmunda, Tsutsumi et al.,2012; Deparia, Kuo et al., 2016). Most rheophytes are obligate-rheophytic species, whilesome others are distinguished at varietal rank, e.g., Deparia lancea var. rheophilaM.Kato, Darnaedi & K.Iwats., Lindsaea ensifolia var. rheophila K.Iwats. Still others,e.g., Bolbitis heteroclita and B. sinuata (C.Presl) Hennipman, are facultative and growin both habitats. Little is known about the evolutionary process of rheophytes. Onequestion to solve is whether facultative rheophytes or rheophytic varieties are transitionaltoward obligate rheophytes. Another is whether speciation of rheophytes is gradual orabrupt.
ACKNOWLEDGEMENTSI thank A. R. Smith for reviewing the manuscript and useful information, and A. Salino,N. Murakami, R. C. Moran and B. Parris for providing data of rheophytes.
REFERENCESCRAWFORD, R. M. M. 2008. Plants at the margin: ecological limits and climatic
change. Cambridge University Press, Cambridge.EVERT, R. G. & EICHHORN, S. E. 2013. Raven Biology of Plants, 8th ed. W. H.
Freeman and Company Publisher, New York.HIYAMA, T., IMAICHI, R. & KATO, M. 1992. Comparative development of
gametophytes of Osmunda lancea and O. japonica (Osmundaceae): adaptation ofrheophilous fern gametophyte. Bot. Mag. Tokyo 105: 215–225.
HOLTTUM, R. E. 1991. Tectaria group. Fl. Males. Ser. II, 2(1): 1–132.IMAICHI, R. & KATO, M. 1992. Comparative leaf development of Osmunda lancea
and O. japonica (Osmundaceae): heterochronic origin of rheophytic stenophylly. Bot.Mag. Tokyo 105: 199–213.
IMAICHI, R. & KATO, M. 1993. Comparative leaf morphology of young sporophytesof rheophytic Osmunda lancea and dryland O. japonica. J. Plant Res. 106: 37–45.
IMAICHI, R. & KATO, M. 1997. Speciation and morphological evolution in rheophytes.In: IWATSUKI, K. & RAVEN, P. H. (Eds) Evolution and diversification of landplants, pp. 309–318, Springer-Verlag, Tokyo.
IWATSUKI, K. 1992. Ferns and Fern Allies of Japan. Heibonsha, Tokyo. (In Japanese)JUNIPER, B. E. & JEFFREE, C. E. 1983. Plant surfaces. Edward Arnold, London.KATO, M. 1984. A taxonomic study of the athyrioid fern genus Deparia with main
reference to the Pacific species. J. Fac. Sci. Univ. Tokyo, Sect. III, 13: 371–430.KATO, M., DARNAEDI, D. & IWATSUKI, K. 1991. Fern rheophytes of Borneo. J. Fac.
Sci. Univ. Tokyo, Sect. III, 15: 91–110.KATO, M. & IMAICHI, R. 1992a. Leaf anatomy of tropical fern rheophytes with its
KATO: DIVERSITY AND ADAPTATIONS OF RHEOPHYTIC FERNS 175

evolutionary and ecological implications. Canad. J. Bot. 70: 165–174.KATO, M. & IMAICHI, R. 1992b. A broad-leaved variant of the fern rheophyte, Tectaria
lobbii. Int. J. Pl. Sci. 153: 212–216.KUO, L.-Y., EBIHARA, A., SHINOHARA, W., ROUHAN, G., WOOD, K. R., WANG,
C.-N. & CHIOU, W.-L. 2016. Historical biogeography of the fern genus Deparia(Athyriaceae) and its relation with polyploidy. Molec. Phylogenet. Evol. 104: 123–134.
LELLINGER, D. B. 1985. A field manual of the ferns & fern-allies of the United States& Canada. Smithsonian Institution Press, Washington, DC.
MATSUI, M., TAKEI, S., OHGA, K., HAYAKAWA, H., YOSHIDA M., ITO, K.,ARAKAWA, R., MASUMOTO, T. & FUKUDA, T. 2013. Morphological andanatomical variations in rheophytic ecotype of violet, Viola mandshurica var.ikedaeana (Violaceae). Amer. J. Pl. Sci. 4(4): 859–865.
MAUSETH, J. D. 2014. Botany; an introduction to plant biology, 5th ed. Jones andBartlett Learning. Burlington, Massachusetts.
MEHLTRETER, K. 2008. Phenology and habitat specificity. In: RANKER, T. A. &HAUFLER, C. H. (Eds) Biology and evolution of ferns and lycophytes, pp. 201–221. Cambridge University Press, Cambridge.
MORAN, R. C. 1995. Five new species and two new combinations of ferns from Ecuador(Polypodiopsida). Nord. J. Bot. 15: 49–58.
NOMURA, N., SETOGUCHI, H. & TAKASO, T. 2006. Functional consequences ofstenophylly for leaf productivity: comparison of the anatomy and physiology of arheophyte, Farfugium japonicum var. luchuense, and a related non-rheophyte, F.japonicum (Asteraceae). J. Pl. Res. 119: 645–656.
OHGA, K., MUROI, M., HAYAKAWA, H., YOKOYAMA, J., ITO, K., TEBAYASHI,S., ARAKAWA, R. & FUKUDA, T. 2012. Comparative morphology and anatomyof non-rheophytic and rheophytic types of Adenophora triphylla var. japonica(Campanulaceae). Amer. J. Pl. Sci. 3: 805–809.
OKADA, H. & TSUKAYA. H. 2010. A new species of Piptospatha (Araceae:Schismatoglottideae) from West Kalimantan, Indonesian Borneo. Acta Phytotax.Geobot. 61: 87–92.
OKADA, H. & TSUKAYA. H. 2013. A new species of Aridaram, Schimatoglottideae,Araceae, from West Kalimantan, Indonesian Borneo. Acta Phytotax. Geobot. 63:71–75.
PAGE, C. N. 1997. The ferns of Britain and Ireland. 2nd ed. Cambridge University Press,Cambridge.
PARACER, S. & AHMADJIAN, V. 2000. Symbiosis: an introduction to biologicalassociations. 2nd ed. Oxford University Press, Oxford.
PPG I. 2016. A community-derived classification for extant lycophytes and ferns. J. Syst.Evol. 54: 563–604.
SETOGUCHI, H. & KAJIMARU, G. 2004. Leaf morphology of the rheophyte,Rhododendron indicum f. otakumi (Ericaceae). Acta Phytotax. Geobot. 55: 45–54.
SHARPE, J. M. 1997. Leaf growth and demography of the rheophytic fern Thelypterisangustifolia (Willdenow) Proctor in a Puerto Rican rainforest. Pl. Ecol. 130: 203–212.
TSUKAYA, H. 2002. The leaf index: heteroblasty, natural variation, and the geneticcontrol of polar processes of leaf expansion. Pl. Cell Physiol. 43:372–378.
USUKURA, M., IMAICHI, R. & KATO, M. 1994. Leaf morphology of a facultative
176 FERN GAZ. 20(5):169-179. 2017

rheophyte, Farfugium japonicum var. luchuense (Compositae). J. Pl. Res. 107: 263–267.
VAN STEENIS, C. G. G. J. 1981. Rheophytes of the world. Sijthoff & Noordhoff, Alphenaan den Rijn.
VAN STEENIS, C. G. G. J. 1987. Rheophytes of the world: supplement. Allertonia 4(5):267–330.
VASCO, A., MORAN, R. C. & AMBROSE, B. A. 2013. The evolution, morphology,and development of fern leaves. In: BARTLETT, M. E. (Ed.) Frontiers in PlantScience 4: Article 4, pp. 1–16.
VERMEULEN, J. & TSUKAYA, H. 2011. An assumedly rheophytic orchid:Bulbophyllum rheophyton n. sp., from Borneo. Pl. Syst. Evol. 293:71–73.
WASER, N. M. & OLLERTON, J. (Eds) 2006. Plant-pollinator interactions: fromspecialization to generalization. University of Chicago Press, Chicago.
YATABE-KAKUGAWA, Y., TSUTSUMI, C., HIRAYAMA, Y., TSUNEKI, S.,MURAKAMI, N. & KATO, M. 2013. Transmission ratio distortion of molecularmarkers in a doubled haploid population originated from a natural hybrid betweenOsmunda japonica and O. lancea. J. Pl. Res. 126: 469–482.
YORIFUJI, E., ISHIKAWA, N., OKADA, H. & TSUKAYA, H. 2015. Arundinagraminifolia var. revoluta (Arethuseae, Orchidaceae) has fern-type rheophytecharacteristics in the leaves. J. Pl. Res. 128: 239–247
APPENDIXRheophytic ferns and suggested rheophytes. Family or subfamily names arefollowed by numbers of taxa.
Aspleniaceae (9) --- Asplenium delicatulum C.Presl, Asplenium dichotomum Hook.[facultative], Asplenium subaquatile Ces., Asplenium triculum A.R.Sm.;Hymenasplenium murakami-hatanakae Nakaike, Hymenasplenium obliquissimum(Hayata) Sugim. & Sa. Kurata, Hymenasplenium obtusifolium (L.) L.Regalado & Prada,Hymenasplenium riparium (Liebm.) L.Regalado & Prada [facultative],Hymenasplenium triquetrum (N. Murak. & R.C. Moran) L.Regalado & Prada
Athyriaceae (7) --- Athyrium haleakalae K.R.Wood & W.L.Wagner; Depariabiserialis (Baker) M.Kato, Deparia confluens (Kunze) M.Kato, Deparia lancea (Thunb.)Fraser-Jenk. var. rheophila (M.Kato, Darnaedi & K.Iwats.) M.Kato; Diplaziumaequibasale (Baker) C.Chr., Diplazium esculentum (Retz.) Sw. [facultative], Diplaziumwahauense M.Kato, Darnaedi & K.Iwats.
Blechnaceae (6) --- Austroblechnum difforme (Copel.) Gaspe & V.A.O.Dittrich;Cranfillia fluviatilis (R.Br.) Gaspe & V.A.O.Dittrich; Oceaniopteris francii (Rosenst.)Gaspe & Salino, Oceaniopteris gibba (Lab.) Gaspe & Salino; Parablechnum glaziovii(Christ) Gaspe & Salino, Parablechnum rheophyticum (R.C.Moran) Gaspe & Salino
Cyatheaceae (1) --- Alsophila esmeraldensis R.C.MoranDennstaedtiaceae (1) --- Microlepia rheophila K.Iwats. & M.KatoDipteridaceae (1) --- Dipteris lobbiana (Hook.) T.MooreDryopteridaceae subfamily Dryopteroideae (3) --- Ctenitis vilis (Kunze) Ching
[facultative]; Dryopteris rheophila Mitsuta ex Darnaedi, M.Kato & K.Iwats.;Polystichum yaeyamense (Makino) MakinoDryopteridaceae subfamily Elaphoglossoideae (17) --- Bolbitis fluviatilis (Hook.) Chingin C.Chr., Bolbitis heteroclita (C.Presl) Ching in C.Chr. [facultative], Bolbitis heudelotii
KATO: DIVERSITY AND ADAPTATIONS OF RHEOPHYTIC FERNS 177

(Bory) Alston, Bolbitis interlineata (Copel.) Ching in C.Chr. [facultative], Bolbitisinterpreta (C.Presl) Hennipman [rheophyte?], Bolbitis presliana (Fée) Ching in C.Chr.,Bolbitis quoyana (Gaud.) Ching in C.Chr., Bolbitis repanda Schott, Bolbitis rivularis(Brack.) Ching in C.Chr., Bolbitis salicina (Hook.) Ching in C.Chr., Bolbitis×singaporensis Holttum [rheophyte?], Bolbitis sinuata (C.Presl) Hennipman[facultative], Bolbitis taylorii (F.M.Bailey) Ching in C.Chr.; Elaphoglossum dumrongiiTagawa & K.Iwats., Elaphoglossum resiniferum Holttum, Elaphoglossum rheophilumM.Kato, Darnaedi & K.Iwats.; Mickelia furcata R.C.Moran, Labiak & Sundue
Hymenophyllaceae (2) --- Cephalomanes javanicum C.Presl var. asplenioides(C.Presl) K.Iwats. and var. sumatranum (Alderw.) K.Iwats. [facultative]
Isoëtaceae (2) --- Isoëtes hawaiiensisW.C.Taylor & W.H.Wagner [facultative], IsoëtesmartiiA.Br. ex Kuhn
Lindsaeaceae (13) --- Lindsaea decaryana (C.Chr.) K.U.Kramer [rheophyte?],Lindsaea ensifolia Sw. var. rheophila K.Iwats., Lindsaea integra Holttum, Lindsaealucida Blume [facultative?], Lindsaea plicata Baker; Odontosoria chinensis (L.) J.Sm.var. rheophila (K.U.Kramer) Parris, Odontosoria gracilis (Tagawa) Ralf Knapp,Odontosoria minutula (Sa.Kurata) Ebihara; Osmolindsaea odorata (Roxb.) Lehtonen &Christenh. [facultative], Osmolindsaea japonica (Baker) Lehtonen & Christenh.;Tapeinidium pinnatum (Cav.) C.Chr. [facultative], Tapeinidium prionoides K.U.Kramer[facultative?]; Xyropteris stortii (Alderw.) K.U.Kramer
Ophioglossaceae (2) --- Ophioglossum gramineum Willd. var. majus (Alderw.)Wieferring, Ophioglossum reticulatum L. var. dilatatum (Miq.) Wieferring [facultative]
Osmundaceae (4) --- Osmunda (Plenasium) angustifolia Ching, Osmunda(Plenasium) bromeliifolia (C.Presl) Copel. [facultative?], Osmunda hybrida Tsutsumi etal. [rheophyte?], Osmunda lancea Thunb.
Polypodiaceae (14) --- Calymmodon asiaticus Copel.; Leptochilus macrophyllus(Blume) Noot. var. fluviatilis (Lauterb.) Noot., Leptochilus minor Fée [facultative?],Leptochilus ×trifidusAlderw.; Loxogramme antrophyoides (Baker) C.Chr. var. rheophilaM.Kato, Darnaedi & K.Iwats.; Microsorum krayanense M.Kato, Darnaedi & K.Iwats.;Microsorum paucijugum (Alderw.) K.Iwats. & M.Kato, Microsorum pteropus (Blume)Copel., Microsorum submarginale M.Kato, Darnaedi & K.Iwats.; Oreogrammitisuniversa (Baker) Parris [facultative]; Selliguea oodes (Kunze) Hovenkamp, Selligueayakushimensis (Makino) Fraser-Jenk.; Xiphopterella alternidens (Ces.) Parris;Xiphopterella gracilis Parris.
Pteridaceae subfamily Pteridoideae (2) --- Pteris ensiformis Burm.f. var. rheophilaM.Kato, Darnaedi & K.Iwats.; Taenitis interrupta Hook. & Grev.
Pteridaceae subfamily Vittarioideae (3) --- Antrophyum vittarioides Baker[facultative?]; Austrogramme asplenioides (Holttum) Hennipman, Austrogrammedecipiens (Mett.) Hennipman [facultative]
Tectariaceae (10) --- Tectaria acutifolia (Hieron.) Maxon, Tectaria ×chaconianaA.Rajas, Tectaria fauriei Tagawa [facultative?], Tectaria jacobsii Holttum, Tectaria lobbii(Hook.) Copel., Tectaria menyanthidis (C.Presl) Copel., Tectaria microlepis Holttum,Tectaria rheophytica Holttum, Tectaria subdigitata (Baker) Copel., Tectaria trifida (Fée)Price
Thelypteridaceae (44) --- Amauropelta aspidioides (Willd.) Pic.Serm., Amauropeltaconsanguinea (Fée) Salino & T.E.Almeida [facultative], Amauropelta fluminalis(A.R.Sm.) Salino & T.E.Almeida, Amauropelta ptarmica (Kunze ex Mett.) Pic.Serm.,Amauropelta rheophyta (Proctor) Salino & T.E.Almeida, Amauropelta sancta (L.)
178 FERN GAZ. 20(5):169-179. 2017

Pic.Serm., Amauropelta saxicola (Sw.) Salino & T.E.Almeida, Amauropeltasoridepressa (Salino & V.A.O.Dittrich) Salino & T.E.Almeida, Amauropelta villana(L.D.Gómez) Salino & T.E.Almeida; Ampelopteris prolifera (Retz.) Copel.[facultative]; Christella subpubescens (Blume) Holttum [facultative]; Goniopteriscumingiana (Kunze) de Jonch. & S.Sen, Goniopteris francoana (E.Fourn.) A.Löve &D.Löve, Goniopteris riograndensis (Lindm.) Ching, Goniopteris skinneri (Hook.)Salino & T.E.Almeida; Meniscium angustifolium Willd., Meniscium arborescensHumb. & Bonpl. ex Willd. [facultative]; Meniscium chrysodioides Fée [facultative],Meniscium lanceum (A.R.Sm.) R.S. Fernandes & Salino [facultative], Menisciumnesioticum (Maxon & C.V.Morton) Pic.Serm. [facultative]; Menisorus pauciflorus(Hook.) Alston [facultative?]; Mesophlebion oligodictyon (Baker) Holttum; Plesioneuronprenticei (Carr.) Holttum; Pronephrium aquatiloides (Copel.) Holttum, Pronephriumfirmulum (Baker) Holttum, Pronephrium heterophyllum (C.Presl) Holttum [rheophyte?],Pronephrium hosei (Baker) Holttum, Pronephrium menisciicarpon (Blume) Holttum,Pronephrium peltatum (Alderw.) Holttum var. persetiferum Holttum [rheophyte?],Pronephrium rubidium (Hook.) Holttum [rheophyte?], Pronephrium salicifolium (Wall.ex Hook.) Holttum; Pseudocyclosorus falcilobus (Hook.) Ching, Pseudocyclosorusesquirolii (Christ) Ching [facultative]; Sphaerostephanos acrostichoides (Desv.) Holttum(=Thelypteris riparia (Copel.) C.F.Reed), Sphaerostephanos aquatilis (Copel.) Holttum,Sphaerostephanos cataractorum (W.H.Wagner & Grether) Holttum, Sphaerostephanoshispidifolius (Alderw.) Holttum, Sphaerostephanos mutabilis (Brause) Holttum,Sphaerostephanos perglanduliferus (Alderw.) Holttum, Sphaerostephanos warburgii(Kuhn & Christ) B.C.Stone; Steiropteris hottensis (C.Chr.) Salino & T.E.Almeida,Steiropteris wrightii (Mett. ex D.C.Eaton) Pic.Serm.; Trigonospora calcarata (Blume)Holttum in B.K.Nayar & S.Kaur, Trigonospora ciliata (Benth.) Holttum
KATO: DIVERSITY AND ADAPTATIONS OF RHEOPHYTIC FERNS 179

A SHORT BIOGRAPHY OF THE AUTHOR
Masahiro Kato was born in Japan in 1946. He graduated from Kyoto University andreceived Dr. Sci. in 1976 with a thesis entitled “Morphological and systematic studies ofAthyriaceae.” After being employed as research assistant in the Department of Botany,Kyoto University, he moved to the University of Tokyo in 1982 and became professorin 1997. Then he moved to the National Museum of Nature and Science as director ofthe Department of Botany and director of Tsukuba Botanical Garden. Now he is professoremeritus of the University of Tokyo and researcher emeritus of the National Museum ofNature and Science. He has undertaken a number of fieldwork projects over tropicalAsia. His research interest has focused on the fern flora of tropical Asia, classificationand systematics of pteridophytes, such as Athyriaceae, Davalliaceae, Ophioglossaceaeand Osmundaceae. He has also investigated the evolution of epiphytic Davalliaceae, andexplored rheophytes in Borneo and adjacent areas. In extended research of rheophytes,he has studied the flora, morphology and evolution of the torrenticolous rheophytesPodostemaceae (rosid angiosperms). This research extended into the genetic backgroundof their unique morphology. Additionally, he researched the evolution of primitiveangiosperms and single-leaf plants (Monophyllaea, Gesneriaceae), and the phylogeneticrelationships of bryophytes and pteridophytes.
180 FERN GAZ. 20(5). 2017

TWO HYBRIDS OF EQUISETUM SYLVATICUM (EQUISETACEAE)ON THE ISLAND OF SENJA, TROMS, NORWAY
MARCUS LUBIENSKI1 & VEIT MARTIN DÖRKEN2
1 Am Quambusch 25, 58135 Hagen, Germany; [email protected] Department of Biology, University of Konstanz, Universitätsstraße 10, 78457
Konstanz, Germany; [email protected]
Key Words: Equisetum sylvaticum, hybrids, macromorphology, micromorphology,SEM, Senja, Norway, distribution
ABSTRACTEquisetum ×mildeanum (E. pratense × E. sylvaticum) and E. ×lofotense (E.arvense × E. sylvaticum) have been discovered on the island of Senja (Troms,Norway). Their appearance in the field and macromorphology are extensivelycompared and imaged. Additionally micromorphological features of the shootand stomata of both rare hybrid taxa and the involved parental species have beenexamined by SEM technology. Distribution maps illustrate the known occurrencefor the European Nordic countries. A third taxon, E. ×bowmanii (E. sylvaticum ×E. telmateia), is included in the comparative study to include all known E.sylvaticum hybrids in a single SEM investigation for the first time.
INTRODUCTIONWood Horsetail, Equisetum sylvaticum L., belongs to the smooth horsetails (subgenusEquisetum) within the family of Equisetaceae (in comparison to the rough texturedmembers of subgenus Hippochaete frequently called “scouring rushes”) and is a delicateplant occurring over a wide range of wet woodland communities in the nemoral to boreo-arctic zone of Eurasia and North America (Hultén, 1950; Hultén & Fries, 1986; Jalas &Suominen, 1972).
Of the 11 hybrids that have been described within the subgenus (see overview inLubienski, 2010; Page & Gureyeva, 2009; 2013) three have E. sylvaticum as one parent:E. ×mildeanum Rothm. (E. pratense Ehrh. × E. sylvaticum), E. ×bowmanii C.N. Page(E. sylvaticum × E. telmateia Ehrh.), and E. ×lofotense Lubienski (E. arvense L. × E.sylvaticum).
E. ×mildeanum was originally described from the Baltic Isle of Rügen of NorthEastern Germany by Rothmaler (1944) and has been known to occur there since the1950s (Lubienski, 2013; Lubienski & Dörken, 2015). More recently the hybrid has beenrecorded from Scotland (Page, 1988; Acock, 2015), Norway (Lubienski, 2003, 2009),and Sweden (Lubienski & Dörken, 2015). Unfortunately it is often confused with theparents, and its real distribution in the British Isles may be overestimated (see Acock,2015).
E. ×bowmanii is recorded from a few localities within the New Forest area ofSouthern England (Page, 1988). Additionally a second site was reported from Scotlandmore recently (Acock, 2015), but this likely to be an error and confused with E.sylvaticum (H. McHaffie & P. Acock, pers. comm.). Consequently the New Forest sitesremain exclusive for this unique plant.
FERN GAZ. 20(5):181-196. 2017 181

In 2009 E. ×lofotense, a similarly remarkable hybrid incorporating E. sylvaticum, wasdetected at a single site in the far north of Norway on the Lofoten island of Austvågøya(Lubienski, 2010).
During a trip to the island of Senja (Troms, Norway) four colonies of E. sylvaticumhybrids were found. Two were identified as E. ×mildeanum and seem to fit into a seriesof E. ×mildeanum sites lying on the islands of North Western Norway, whereas the lattertwo represent the second record for E. ×lofotense. At first glance the two hybrid taxa arequite similar in habit and certain macromorphological features. Their adjacent sitesallowed for study and direct comparison of their macromorphology in situ. Additionalmicromorphological investigation using SEM focused on the silica spikes on the ridgesof the shoots and the structure of the stomata. As part of the morphological comparison,the hybrids were compared with E. sylvaticum and other taxa to confirm the identity ofthe second parents.
MATERIAL AND METHODSMaterialPlant material was collected on the the island of Senja (Troms province) that is locatednortheast of the Lofoten and Vesterålen islands. It is situated in the transition betweenthe Northern and Middle boreal vegetation zone (Sjörs et al., 2004). The islands andcoastal mainland of Northwestern Norway display an oceanic climate with highprecipitation values (Sjörs et al., 2004).There are two colonies of each hybrid alongsidea single fjord (Mefjorden) in the northern part of the island (Figure 1).
Equisetum ×mildeanum (E. pratense × E. sylvaticum)
1. Mefjorden near Svarthola, Berg kommune, Senja, Troms fylkeN 69°27’53.3’’ / E 17°34’29.2’’, c. 9 m a.s.l.
Growing over a length of approximately 90 metres along the wet western roadside ditchbeneath the northeastern slope of Litlehesten mountain, south of Bratthesten tunnel(Figure 2a, b).
182 FERN GAZ. 20(5):181-196. 2017
Figure 1.Mefjorden in the northern part of the island of Senja (Troms, Norway) showingthe four sites of the Equisetum sylvaticum hybrids (E. ×mildeanum 1 & 2, E. ×lofotense3 & 4) (based on Google Earth ©, Google Inc., 26.12.2016).

2. Mefjorden east of Mefjordvær, Berg kommune, Senja, Troms fylkeN 69°30’54.4’’ / E 17°28’08.2’’, c. 16 m a.s.l.
Occupying both ditches and shoulders of the road east of the town of Mefjordvær andwest of Mykjeneset beneath the northern slope of Kyle mountain. The colony stretchesover a length of approximately 100 metres (Figure 3a, b).
Equisetum ×lofotense (E. arvense × E. sylvaticum)
3. Mefjorden near Svarthola, Berg kommune, Senja, Troms fylkeN 69°27’42.2’’ / E 17°34’51.1’’, c. 5-13 m a.s.l.
Growing along a small creek that rises between the northeastern slopes of Store Hestenand Litlehesten mountain, crosses the road south of Bratthesten tunnel and flows intoMefjorden just a few metres beneath. The hybrid occupies the wet slope and the roadsideditch west of the road over an area of approximately 40 × 40 metres. The eastern side ofthe road descends to the sea and is colonized by E. ×lofotense over an area ofapproximately 40 × 5 metres (Figure 4a, b).
LUBIENSKI & DÖRKEN: TWO HYBRIDS OF EQUISETUM SYLVATICUM 183
Figure 2. a, b. Equisetum ×mildeanum at its site near Svarthola (25.07.2016, M.Lubienski).
Figure 3. a, b. Equisetum ×mildeanum at its site near Mefjordvær (30.07.2016, M.Lubienski).

4. Mefjorden north of Kvalvika, Berg kommune, Senja, Troms fylkeN 69°28’54.8’’ / E 17°33’15.9’’, c. 5-13 m a.s.l.
The site is situated by the same road two kilometres to the north, north of Bratthestentunnel. The hybrid grows over an area 10 × 30 metres on the wet slope between Burstindand Bringtinden mountain and in the western roadside ditch; it is also found on the other(eastern) side of the road, where it accompanies a small rivulet to the sea (Figure 5 a, b).
MethodsFreshly collected shoots were fixed in FAA (100 ml FAA = 90 ml 70% ethanol + 5 mlacetic acid 100% + 5 ml formaldehyde solution 37%) before being stored in 70% ethanol.For SEM with AURIGA ZEISS TM the fixed material was dehydrated in FDA(formaldehyde-dimethyl-acetal) and critical-point dried (CPD 030, BALZERS). CP-dried material was sputtered with gold-palladium (thickness 5 nm) with a BALTECSputterCoater SCD 030.
RESULTSComparison of morphology and field observations of Equisetum ×mildeanum andE. ×lofotense on SenjaIn the field both hybrids appear similar to E. sylvaticum at first glance and look like odd
184 FERN GAZ. 20(5):181-196. 2017
Figure 4. a, b. Equisetum ×lofotense at its site near Svarthola (25.07.2016, M.Lubienski).
Figure 5. a, b. Equisetum ×lofotense at its site near Kvalvika (25.07.2016, M.Lubienski).

LUBIENSKI & DÖRKEN: TWO HYBRIDS OF EQUISETUM SYLVATICUM 185
Figure 6. SEM images of the micromorphology of the shoot ridges and stomata ofEquisetum arvense (a-b), E. sylvaticum (c-d), E. pratense (e-f), and E. telmateia (g-h).Arrows and letters indicate ridges (r), furrows (f), mamillae (m) and pilulae (p).
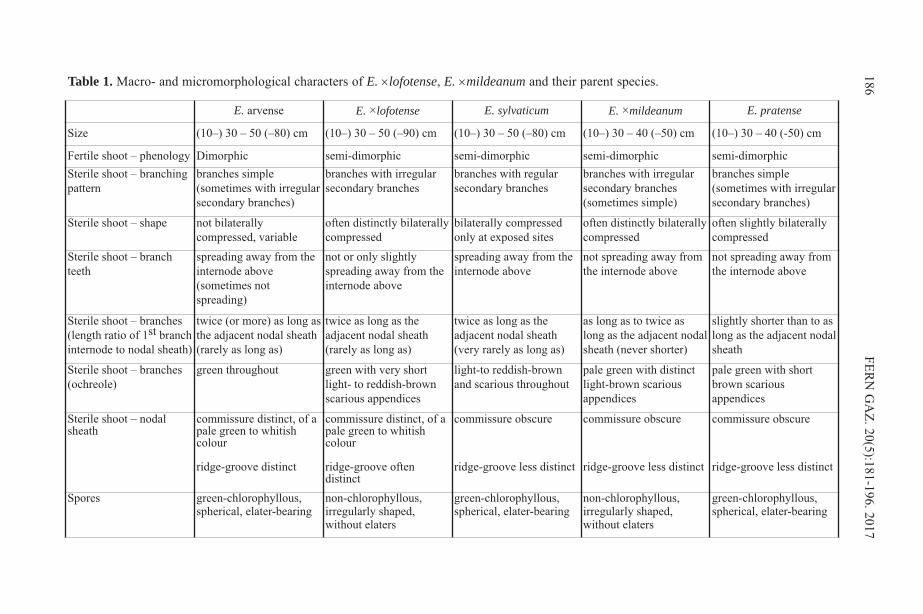
186 FERN G
AZ. 20(5):181-196. 2017
E. arvense E. ×lofotense E. sylvaticum E. ×mildeanum E. pratense
Size (10–) 30 – 50 (–80) cm (10–) 30 – 50 (–90) cm (10–) 30 – 50 (–80) cm (10–) 30 – 40 (–50) cm (10–) 30 – 40 (-50) cm
Fertile shoot – phenology Dimorphic semi-dimorphic semi-dimorphic semi-dimorphic semi-dimorphicSterile shoot – branchingpattern
branches simple(sometimes with irregularsecondary branches)
branches with irregularsecondary branches
branches with regularsecondary branches
branches with irregularsecondary branches(sometimes simple)
branches simple(sometimes with irregularsecondary branches)
Sterile shoot – shape not bilaterallycompressed, variable
often distinctly bilaterallycompressed
bilaterally compressedonly at exposed sites
often distinctly bilaterallycompressed
often slightly bilaterallycompressed
Sterile shoot – branchteeth
spreading away from theinternode above(sometimes notspreading)
not or only slightlyspreading away from theinternode above
spreading away from theinternode above
not spreading away fromthe internode above
not spreading away fromthe internode above
Sterile shoot – branches(length ratio of 1st branchinternode to nodal sheath)
twice (or more) as long asthe adjacent nodal sheath(rarely as long as)
twice as long as theadjacent nodal sheath(rarely as long as)
twice as long as theadjacent nodal sheath(very rarely as long as)
as long as to twice aslong as the adjacent nodalsheath (never shorter)
slightly shorter than to aslong as the adjacent nodalsheath
Sterile shoot – branches(ochreole)
green throughout green with very shortlight- to reddish-brownscarious appendices
light-to reddish-brownand scarious throughout
pale green with distinctlight-brown scariousappendices
pale green with shortbrown scariousappendices
Sterile shoot – nodalsheath
commissure distinct, of apale green to whitishcolour
ridge-groove distinct
commissure distinct, of apale green to whitishcolour
ridge-groove oftendistinct
commissure obscure
ridge-groove less distinct
commissure obscure
ridge-groove less distinct
commissure obscure
ridge-groove less distinct
Spores green-chlorophyllous,spherical, elater-bearing
non-chlorophyllous,irregularly shaped,without elaters
green-chlorophyllous,spherical, elater-bearing
non-chlorophyllous,irregularly shaped,without elaters
green-chlorophyllous,spherical, elater-bearing
Table 1. Macro- and micromorphological characters of E. lofotense, E. mildeanum and their parent species.
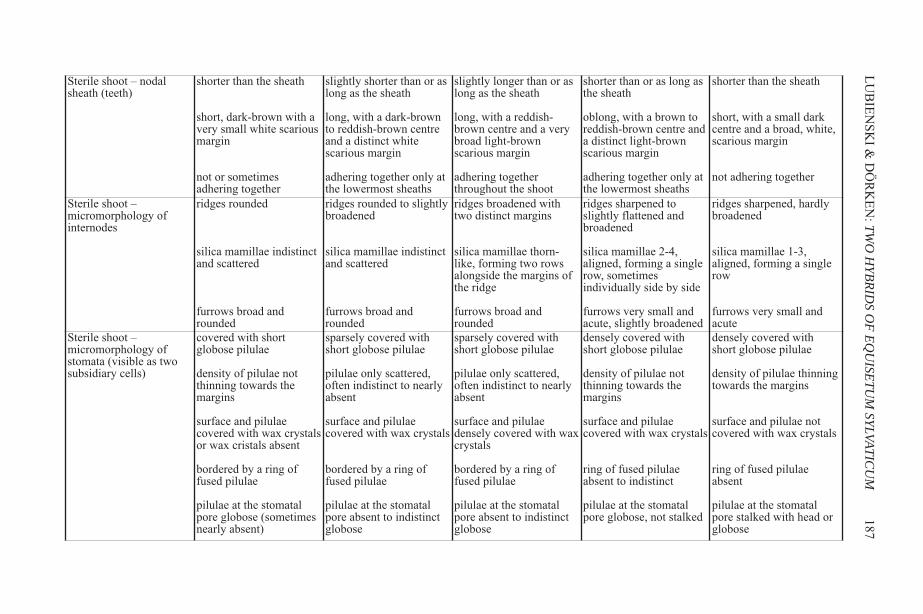
LUBIEN
SKI &
DÖ
RKEN
:TWO HYBRID
S OF EQ
UISETU
M SYLVATIC
UM
187
Sterile shoot – nodalsheath (teeth)
shorter than the sheath
short, dark-brown with avery small white scariousmargin
not or sometimesadhering together
slightly shorter than or aslong as the sheath
long, with a dark-brownto reddish-brown centreand a distinct whitescarious margin
adhering together only atthe lowermost sheaths
slightly longer than or aslong as the sheath
long, with a reddish-brown centre and a verybroad light-brownscarious margin
adhering togetherthroughout the shoot
shorter than or as long asthe sheath
oblong, with a brown toreddish-brown centre anda distinct light-brownscarious margin
adhering together only atthe lowermost sheaths
shorter than the sheath
short, with a small darkcentre and a broad, white,scarious margin
not adhering together
Sterile shoot –micromorphology ofinternodes
ridges rounded
silica mamillae indistinctand scattered
furrows broad androunded
ridges rounded to slightlybroadened
silica mamillae indistinctand scattered
furrows broad androunded
ridges broadened withtwo distinct margins
silica mamillae thorn-like, forming two rowsalongside the margins ofthe ridge
furrows broad androunded
ridges sharpened toslightly flattened andbroadened
silica mamillae 2-4,aligned, forming a singlerow, sometimesindividually side by side
furrows very small andacute, slightly broadened
ridges sharpened, hardlybroadened
silica mamillae 1-3,aligned, forming a singlerow
furrows very small andacute
Sterile shoot –micromorphology ofstomata (visible as twosubsidiary cells)
covered with shortglobose pilulae
density of pilulae notthinning towards themargins
surface and pilulaecovered with wax crystalsor wax cristals absent
bordered by a ring offused pilulae
pilulae at the stomatalpore globose (sometimesnearly absent)
sparsely covered withshort globose pilulae
pilulae only scattered,often indistinct to nearlyabsent
surface and pilulaecovered with wax crystals
bordered by a ring offused pilulae
pilulae at the stomatalpore absent to indistinctglobose
sparsely covered withshort globose pilulae
pilulae only scattered,often indistinct to nearlyabsent
surface and pilulaedensely covered with waxcrystals
bordered by a ring offused pilulae
pilulae at the stomatalpore absent to indistinctglobose
densely covered withshort globose pilulae
density of pilulae notthinning towards themargins
surface and pilulaecovered with wax crystals
ring of fused pilulaeabsent to indistinct
pilulae at the stomatalpore globose, not stalked
densely covered withshort globose pilulae
density of pilulae thinningtowards the margins
surface and pilulae notcovered with wax crystals
ring of fused pilulaeabsent
pilulae at the stomatalpore stalked with head orglobose

variants of that species. However detailed differences from the parents in macro- andmicromorphological aspects have been listed earlier: Page (1988; 1997), Lubienski(2003; 2009; 2013), Lubienski & Dörken (2015) (for E. ×mildeanum) and Lubienski(2010; 2013) (for E. ×lofotense). Additionally both hybrids show irregular meiotic celldivision resulting in aborted spores (for images see Lubienski, 2009; 2010; Lubienski &Dörken, 2015). In both taxa a remarkable discrepancy between their macro- andmicromorphological characters is expressed. Traits of the macromorphological dominantparent E. sylvaticum are much more obscure in the micromorphology of the main shootof both hybrids (especially regarding the highly distinctive ridge architecture, Figures 6,7). Macro- and micromorphological features of both hybrids and their parental species
188 FERN GAZ. 20(5):181-196. 2017
Figure 7. SEM images of the micromorphology of the shoot ridges and stomata ofEquisetum ×lofotense (a-b), E. ×mildeanum (c-d), and E. ×bowmanii (e-f). Arrows andletters indicate ridges (r), furrows (f), mamillae (m) and pilulae (p).

are listed in Table 1.The characteristic and highly distinct flattened and two-edged ridges bordered by
two rows of silica spikes of E. sylvaticum are not found in E. ×mildeanum and E.×lofotense. Both show a micromorphology more similar to their second parent, E.pratense and E. arvense respectively (Lubienski, 2010; Lubienski & Dörken, 2015).Interestingly this recessive behaviour of E. sylvaticum micromorphology is expressed toa similar degree in the third known E. sylvaticum hybrid, E. ×bowmanii, also a hybridclearly indicating Wood Horsetail as one parent in its macromorphological appearance.It therefore is added in Figures 6 and 7, so that the micromorphological characters of allthree known E. sylvaticum hybrids and their parental species can be compared directly.Because of the rarity of E. ×lofotense, in comparison with E. ×mildeanum, some furtherfield notes of the hybrid are given, thus supplementing the data given in Lubienski (2010;2013).
Plants on Senja were detected within the mixed stands with E. sylvaticum, becausethey displayed a characteristic asymmetric overhanging growth habit especially in theexposed roadside ditches and slopes (Figure 8a, b). In this character the hybrid is similarto E. ×mildeanum (Figure 8c, d).
Nevertheless in all dimensions it is a more robust and taller plant, reaching heightsup to 90 cm (the tallest shoot found on Senja was 78 cm long, Figure 9a), which is neverfound in E. ×mildeanum (max. height approx. 40 cm, Figure 9b). Additionally both E.sylvaticum hybrids show distinctly the influence of their second parent in their nodalsheath and branch characters (Figure 10) and can be differentiated by these. An additionaltask in the field is to distinguish E. ×lofotense from its E. sylvaticum parent with whichit frequently co-occurs. Differences between both are seen in the branches, the branchteeth, the sheath teeth, the shoot surface, and the fertile shoots in general.
The distinct branching pattern of all E. sylvaticum hybrids (Lubienski, 2010) seemsto be a very stable and reliable character. The side branches are long, spreading, andsecondarily branched only in their proximal part, so that a distal unbranched distinctlyoverhanging part results, giving the whole shoot a somehow irregular habit (Figure 11,14b; see also image in Lubienski 2010). This differs from the very symmetric and storiedbranching pattern of E. sylvaticum resulting in brush-like side branch clusters (Figure14a). In comparison with E. sylvaticum the teeth of the side branches of E. ×lofotenseare not, or to a much lesser degree, spreading from the internode above (Figure 12). Thesheath teeth are less reddish-brown, shorter or as long as the sheath itself (longer thanthe sheath in E. sylvaticum), only irregular adhering together, becoming darker brown,shorter and more free in the upper parts of the shoot, i.e. E. arvense-like. The main shootsurface is not as rough as in E. sylvaticum, a direct consequence of the micromorphology,where silica spikes are lacking on the less prominent ridges (Figure 7a). This differenceis also clearly seen at the surface of the nodal sheath, which shows shallow ridges witha smooth surface like E. arvense, in contrast to prominent ridges with rough silicaprojections in E. sylvaticum (Figure 13).
The fertile shoots of E. ×lofotense have a very striking appearance at the Senja sites.These are very sturdy in general habit and bigger in all dimensions than the sterile shoots.Although this is a typical feature of the fertile shoots of all semidimorphic species andhybrids (e.g. E. sylvaticum, E. pratense, E. ×mildeanum, E. ×bowmanii), the fertile E.×lofotense shoots from Senja are remarkable in their sturdy conifer-like habit. Whenviewed from above these shoots appear star-like and distinctly different from the fertileshoots of E. sylvaticum with which they grow intermixed. This difference results from
LUBIENSKI & DÖRKEN: TWO HYBRIDS OF EQUISETUM SYLVATICUM 189

190 FERN GAZ. 20(5):181-196. 2017
Figure 8. Equisetum ×lofotense (a & b) and E. ×mildeanum (c & d), showingcharacteristic overhanging growth habit.
Figure. 9. Shoots of Equisetum ×lofotense (a) and E. ×mildeanum (b), showingdifferences in size (bar = approx. 10 cm).

LUBIENSKI & DÖRKEN: TWO HYBRIDS OF EQUISETUM SYLVATICUM 191
Figure 10. Sterile shoots of Equisetum ×lofotense (a) and E. ×mildeanum (b), showinginfluence of the second parent species E. arvense in E. ×lofotense (a) (oblong nodalsheaths and sheath teeth, low number of teeth and side branches) and E. pratense in E.×mildeanum (b) (short nodal sheaths and sheath teeth, high number of densely spacedteeth and side branches).
Figure 11. Sterile and fertile shoots of Equisetum ×lofotense, showing irregularbranching pattern.
Figure 12. Sterile shoots of Equisetum sylvaticum (a) and E. ×lofotense (b), showingdifferences in branch teeth, spreading (a) and not spreading (b) from the internode above.

192 FERN GAZ. 20(5):181-196. 2017
Figure 13. Nodal sheaths of sterile shoots of Equisetum sylvaticum (a) and E. ×lofotense(b), showing differences in sheath surface and length and sheath teeth colour and length.
Figure 14. Fertile shoots of Equisetum sylvaticum (a) and E. ×lofotense (b), showingdifferences in branching pattern.
Figure 15 Equisetum ×lofotense, nodal sheaths, sterile (a) and fertile (b) shoot, showingdifferent influence of the parent species E. arvense and E. sylvaticum in themacromorphological appearance of the hybrid.

the fact that the 1st order side branches of the fertile shoots of E. ×lofotense are in generallonger than in E. sylvaticum, while having 2nd order side branches shorter than in E.sylvaticum (Figure 14).
Concerning the sterile and fertile shoots of E. ×lofotense, a different influence of theparent species is also expressed in the macromorphological appearance of the hybrid,making the sterile shoots more E. arvense-like and the fertile shoots more E. sylvaticum-like (Figure 15).
Distribution of Equisetum ×mildeanum and E. ×lofotenseFigure 16a shows the known distribution of E. ×mildeanum for whole Norden, and itcan be presumed that other populations may be found within this area.
Figure 16b shows the distribution of E. ×lofotense, at present an endemic ofnorthwestern Norway. Further field work in this area should explore whether it is actuallymore frequent than currently known. The vigour of the hybrid populations on Austvågøyaand Senja makes this suggestion likely.
DISCUSSIONAll four sites of E. ×mildeanum and E. ×lofotense are very similar from ageomorphological and ecological point of view, being situated within a single fjord inthe north of Senja over a distance of approximately 7.25 kilometres. The habitat isexclusively a wet, north- to east-facing mountain slope characterised by huge bouldersand sparse woodland, interrupted at its base by a road only a few metres above and nearto the sea. All sites and the whole area in general harbour extensive stands of the parentspecies E. sylvaticum, E. arvense and E. pratense.
Horsetail hybrid colonies are often found in disturbed sites such as roadside ditchesor railway embankments. This can be caused either by vegetative spreading alongside
LUBIENSKI & DÖRKEN: TWO HYBRIDS OF EQUISETUM SYLVATICUM 193
Figure 16. Distribution of Equisetum ×mildeanum (a) and E. ×lofotense (b) for Norden(arrows indicate two localities, the semi-filled dot indicates the historic occurrence ofE. ×mildeanum at its type locality on the Isle of Rügen, Germany

the traffic system by rhizome or shoot cuttings or through an initial hybridization eventdue to ecological conditions that favour cross fertilization between Equisetumgametophytes. Wet roadside ditches like those on Senja display perfect conditions forgermination of the chlorophyllous and short-lived Equisetum spores and theestablishment of gametophytes. These are open raw soils without higher plant orbryophyte competition, especially after roads are periodically cleared and straightened(Hauke, 1967; Page, 1967; Duckett & Duckett, 1974; Mesler & Lu, 1977; Duckett &Duckett, 1980; Duckett, 1985). Such pioneer habitats fit with the rather narrow ecologicalrequirements of Equisetum gametophytes and for this reason can harbour mixedgametophyte populations, that increase the probability of crossing events betweendifferent species (Hauke, 1978; Page & Barker, 1985; Husby, 2013).
Intergametophytic fertilization and therefore hybridization in Equisetum isadditionally favoured by the reproduction biology of the genus in general. Equisetumgametophytes are initially unisexual, potentially becoming bisexual under changinggrowing conditions (e. g. accumulation of metabolites) (Duckett, 1972; Duckett &Duckett, 1980). However such an accumulation rarely takes place under natural growingconditions (Duckett & Duckett, 1980) and therefore bisexuality is obviously rare amongwild gametophyte populations (Duckett & Duckett, 1974, 1980; Duckett, 1979).Consequently, in Equisetum, intergametophytic fertilization is highly effective in nature,thus outbreeding processes are facilitated. This effect is further emphasized wheregametophytes of different species grow together, which in addition favours hybridizationevents (Duckett & Duckett, 1974; Duckett, 1979).
It seems likely that E. ×mildeanum and E. ×lofotense both originated in the roadsideditches of Mefjorden and succeeded in colonising the adjacent slopes. Each of the fourcolonies may have started from an individual crossing event or there may have been justtwo events with subsequent vegetative spread. As the two sites of E. ×lofotense are onlytwo kilometres apart, a single de novo hybridization event between the parent species toform this rare hybrid seems to be most likely, followed by vegetative spread as a resultof human disturbance.
Nevertheless, the fact that all four colonies stretch over at least several hundred squaremetres means they are unlikely to be of very recent origin.
ACKNOWLEDGEMENTSWe would like to thank Torbjørn Alm (Tromsø, Norway) for help with the map of Nordenand Michael Laumann and Lauretta Nejedli (Electron Microscopy Center, Departmentof Biology, University of Konstanz, Germany) for technical support (SEM). PatrickAcock (St. Mary Cray, Great Britain) and Heather McHaffie (Edinburgh, Great Britain)provided valuable advice concerning E. ×bowmanii in the British Isles.
REFERENCESACOCK, P. 2015. Equisetum L. In: STACE, C. A., PRESTON, C. D. & PEARMAN, D.
A. (Eds) Hybrid Flora of the British Isles, pp. 7-13, Botanical Society of Britain andIreland, Bristol.
DUCKETT, J. G. 1972. Sexual behaviour of the genus Equisetum subgenus Hippochaete.Bot. J. Linn. Soc. 65: 87-108.
DUCKETT, J. G. 1979. An experimental study of the reproductive biology andhybridization in the European and North American species of Equisetum. Bot. J. Linn.Soc. 79: 205-229.
194 FERN GAZ. 20(5):181-196. 2017

DUCKETT, J. G. 1985. Wild gametophytes of Equisetum sylvaticum. Amer. Fern J. 75:120-127.
DUCKETT, J. G. & DUCKETT, A. R. 1974. The ecology of Equisetum gametophytes.Amer. J. Bot. 61 Suppl.: 36.
DUCKETT, J. G. & DUCKETT, A. R. 1980. Reproductive biology and populationdynamics of wild gametophytes of Equisetum. Bot. J. Linn. Soc. 80: 1-40.
HAUKE, R. L. 1967. Sexuality in a wild population of Equisetum arvense gametophytes.Amer. Fern J. 57: 59-66.
HAUKE, R. L. 1978. A taxonomic monograph of Equisetum subgenus Equisetum. NovaHedwigia 30: 385-455.
HULTÉN, E. 1950. Atlas över växternas utbredning i Norden. Fanerogamer ochormbunksväxter. Generalstabens Litografiska Anstalts Förlag, Stockholm.
HULTÉN, E. & FRIES, M. 1986. Atlas of North European Vascular Plants. North of thetropic of cancer. I. Introduction. Taxonomic Index to the maps 1-996. Maps 1-996.Koeltz Scientific Books, Königsstein.
HUSBY, C. 2013. Biology and functional ecology of Equisetum with emphasis on theGiant Horsetails. Bot. Rev. 79: 147-177.
JALAS, J & SUOMINEN, J. (Eds) 1972. Atlas Florae Europaeae. 1. Pteridophyta(Psilotaceae to Azollaceae). The Committee for Mapping the Flora of Europe andSocietas Biological Fennica Vanamo. Helsinki.
LUBIENSKI, M. 2003. Equisetum ×mildeanum Rothm. (E. pratense Ehrh. × E.sylvaticum L.), en snellehybrid ny for Norge. Blyttia 61: 171-178.
LUBIENSKI, M. 2009. Nye funn av Equisetum ×mildeanum (E. pratense × E.sylvaticum) i Norge. Blyttia 67: 270-275.
LUBIENSKI, M. 2010. A new hybrid horsetail Equisetum ×lofotense (E. arvense × E.sylvaticum, Equisetaceae) from Norway. Nord. J. Bot. 28: 530-540.
LUBIENSKI, M. 2013. Hybriden der Gattung Equisetum (Equisetaceae, Equisetopsida,Monilophyta) in Europa. Ber. Inst. Landschafts- Pflanzenökologie Univ. Hohenheim,Beih. 22: 91-124.
LUBIENSKI, M. & DÖRKEN, V. M. 2015. Ängsfräken × skogsfräken funnen i Sverige– samt en översikt över de nordiska fräkenhybriderna. [Equisetum ×mildeanumRothm. found in Sweden.] Svensk Bot. Tidskr. 109: 228-239.
MESLER, M. R. & LU, K. L. 1977. Large gametophytes of Equisetum hyemale innorthern California. Amer. Fern J. 67: 97-98.
PAGE, C. N. 1967. Sporelings of Equisetum arvense in the wild. Br. Fern Gaz. 9: 335-338.
PAGE, C. N. 1988. Two hybrids of Equisetum sylvaticum L. new to the British flora.Watsonia 17: 273-277.
PAGE, C. N. 1997. The ferns of Britain and Ireland. 2nd ed. Cambridge University Press,Cambridge.
PAGE, C. N. & BARKER, M. 1985. Ecology and geography of hybridization in Britishand Irish horsetails. Proc. Roy. Soc. Edinb. 86b: 265-272.
PAGE, C. N. & GUREYEVA, I. I. 2009. A new horsetail hybrid from Western Siberia.Animadversiones Systematicae ex Herbario Kryloviano Universitatis Tomskensis101: 15-21.
PAGE, C. N. & GUREYEVA, I. I. 2013. Equisetum × sergijevskianum, a hybrid horsetailfrom Siberia. Fern Gaz. 19: 181-190.
ROTHMALER, W. 1944. Pteridophyten-Studien I. Feddes Repert. 54: 55-82.
LUBIENSKI & DÖRKEN: TWO HYBRIDS OF EQUISETUM SYLVATICUM 195

SJÖRS, H., JONSELL, B. & ELVEN, R. 2004. Features of Nordic environment andvegetation. In: Jonsell, B. (Ed) Flora Nordica. General Volume, pp. 37–100, TheBergius Foundation, The Royal Swedish Academy of Sciences, Stockholm.
196 FERN GAZ. 20(5):181-196. 2017

NEW RECORD OF THE RARE SERPOCAULON OBSCURINERVIUMD. SANÍN (POLYPODIACEAE) IN THE EASTERN CORDILLERA OF
COLOMBIA
DAVID SANÍN1, 2*, VANIA TORREZ3, JORGE L. PEÑA-NÚÑEZ4 &EDWIN TRUJILLO-TRUJILLO5
1Universidad de la Amazonia, Programa de Biología, Calle 17, diagonal 17 concarrera 3F, Barrio el Porvenir, Florencia-Caquetá, Colombia;
2Current address: Instituto de Ciências Biológicas, Departamento de Botânica,Universidade Federal de Minas Gerais, Caixa Postal 486, 30123-970, Belo Horizonte,
Minas Gerais, Brazil; 3Herbario Nacional de Bolivia, Instituto de Ecología, Universidad Mayor de San
Andrés, Cota cota, calle 27, Campus Universitario, La Paz, Bolivia; 4Semillero de Investigación en Biogeografía Evolutiva (SIBE), Universidad de la
Amazonia, Programa de Biología, Florencia-Caquetá, Colombia; 5Universidad de la Amazonia, Programa de Ingenieria Agroecológica,
Florencia-Caquetá, Colombia.*Email: [email protected]
Key words:Andean-Amazonian foothills; Caquetá; Huila; ferns; Florencia;distribution range; epipetric habit.
ABSTRACTSerpocaulon obscurinervium is considered as a rare and threatened species, withonly four exemplars registered including the type. It was recorded from Colombiaand Ecuador, but most commonly reported for central southern Colombia (centralwest of Chocó and south of Nariño departments). Here, we expand its distributionby reporting new records for this species on both sides of the Eastern Cordilleraof the Colombian Andes, specifically from the high Andean forest of Florencia,in the Caquetá department and Suaza in the Huila department. Furthermore, thisis the first record of the species as an epipetric plant. Pictures of the plant and anupdated distribution map are provided.
INTRODUCTIONSerpocaulon A. R. Sm. (Polypodiaceae) is a neotropical fern genus, which contains 52taxa (Sanín 2015). These species occur mainly in humid forests (Smith et al. 2006).Colombia harbours 27 species that are distributed in the inter Andean valleys, the ChocóRegion, the Sierra Nevada de Santa Marta, Oriental valley and Amazon, but most speciesoccur in the humid forests of the Andes and the Chocó Region (Sanín in press).
Serpocaulon obscurinervium D. Sanín (Figure 1) is an Andean species that wasdescribed in 2014 from herbarium specimens deposited in collections of Colombia(CHOCO, COL, CUVC and PSO), France (P) and United States (MO and UC) (Sanín2014). The first specimen was collected in Ecuador by R. Spruce in 1909. In 1973, otherspecimens were collected in Nariño at 2500m by O. Hagemann & L. Leist. The mostrecent collections were recorded in 1988 in the Andean zone of the Department of Chocóby P. Silverstone–Sopkin, and in La Planada Reserve, Nariño Department, by H. Herrera
FERN GAZ. 20(5):197-201. 2017 197

(Sanín 2014). Because the distribution of the species within Colombia was restricted tohigh montane forests outside of any protected area, its conservation status was consideredas Endangered (EN) following the IUCN criteria (Sanín 2014). Since its first description,the species had never been found again or reported alive in nature.
During the ongoing botanic exploration of the Caquetá and Huila departments,Serpocaulon obscurinervium was recorded growing in sites on both sides of the EasternCordillera. The specimens were found along the roadside of Puente Cerro Negro in ElPortico, at the western slope in a forest at 2190m and in Gabinete military station at2050m on the border of Caquetá and Huila Departments on the Eastern slope of theEastern Andean Cordillera. In Huila Department, it was also found on the road to Neiva,in the municipality of Suaza at 2190m (Figure 2). Two to five vouchers were collectedper site and were deposited at the following herbaria: CAUP, COAH, COL, HUA, FAUC& PSO.
The exemplars from Caquetá (Puente Cerro Negro and Gabinete) are the first recordsof Serpocaulon obscurinervium as an epipetric fern. In addition, it is the first time thatthe species has been collected in the Caquetá and Huila departments (Murillo-P. et al.2016), the easternmost distribution of the species with its nearest known population at
198 FERN GAZ. 20(5):197-201. 2017
Figure 1. Serpocaulon obscurinervium. A: Habit of the plant, scale bar (5 cm). B:Rhizome, scale bar (1 cm). C: Rhizome bundles, scale bar (3.5 cm). D: Petiole bundles,scale bar (2 cm). E: Fiddlehead, scale bar (1 cm). F: Lamina base, scale bar (3 cm). G:Lamina apex, scale bar (3 cm). H: Immature sori, scale bar (8 cm). I: Mature sori, scalebar (8 cm). J: Pinna veins, scale bar (8 cm). All from Sanín & Peña 6371 (COL, HUA).Photos by J.L. Peña.

250 km distance on the road of La Victoria−Monopamba, Dept. Nariño, in central southColombia.
Serpocaulon obscurinervium is closely related to S. eleutherophlebium (Fée) A. R.Sm., a species that is distributed from Costa Rica to Ecuador and Venezuela (Moran1995, Smith et al. 2006), and in Colombia is widely recorded in high Andean forest and
SANÍN ET AL: SERPOCAULON OBSCURINERVIUM 199
Figure 2. Distribution of Serpocaulon obscurinervium. The black dots are previousregisters of the species: 1) Cerro del Torrá, Valle del Cauca, Colombia; 2) La PlanadaReserve, Nariño, Colombia; 3) road La Victoria−Monopamba, Nariño, Colombia; 4)Tungurahua, Mt. Abitagua, Ecuador. The asterisks are the new registers on the OrientalAndean Cordillera of Colombia: 5) El Portico and 6) El Gabinete from Florencia,Caquetá department and 7) From Suaza, Huila department.

subpáramos at 2430-3950m, epiphytic, terrestrial and rupicolus (Sanín in press).Theidentification of these two taxa has been confused. Both species have similarlong-creeping rhizomes, scales with patent apices, concolorous and pinnatisect laminae.However, S. obscurinervium differs from S. eleutherophlebium in the scales characters,being linear-lanceolate (versus subulate), dark orange (versus dark brown), larger sized5.2-(6.3)-8.5 × 1.6-(1.7)-2.3 mm (versus 3.5-(5.1)-7.1 × 0.5-(1.1)-1.7 mm), and withlonger petioles 24-(2.6)-28 cm (versus 3.4-(12)-25.5 cm), more areoles along the middlesegment 19-(20)-28 rows (versus 6-(12)-21 rows), inconspicuous fertile veinlets (versusconspicuous fertile veinlets), and the presence of 2-4 celled, dispersed, catenate, tereteand dark brown trichomes on the blades and rachises (versus absent trichomes) (for acomplete comparison see Sanín 2014).
Even though three more populations of Serpocaulon obscurinervium were discoveredin the Caquetá and Huila departments, it is still categorized as Endangered [EN B1ab(i,ii, iii) + B2ab(i, ii, iii)] following the IUCN criteria. Here we present more support forthis, and include Criterion D to its IUCN categorization: [EN B1ab(i, ii, iii) + B2ab(i, ii,iii) + D], the latter given that the population size of S. obscurinervium is estimated to befewer than 250 mature individuals.
The sparse and vicariant distribution of the species on the Andean Cordillera ofEcuador and Colombia (Sanín 2014), and the reduction of high montane forest vegetationcaused by human settlements and agricultural expansion (Murcia et al. 2010), impose apermanent and increasing danger to the populations of this species, especially as all therecords of this species are outwith any protected area. In addition, from five to 10individuals were recorded in these new localities, including both mature and juvenileindividuals (pers. obs.).
MATERIAL EXAMINEDSerpocaulon obscurinervium—Caquetá: Florencia, Via Neiva, vereda El Condor, amano derecha de la vía, a 100m del Puente Cerro Negro [1°45'19.62" N , 75°44'59.98"W], 2050m, 27 Jul 2015, Sanín & Peña 6371 (CAUP, PSO, COL, HUA); Florencia, ViaGuadalupe, El Gabinete, a mano izquierda de la vía, a 100m de la estación militar[75º40’58.4” O, 1º52’44” N], 2700–2800m, 20 Feb 2016, Sanín, García & Camargo-García 6396 (COAH, COL, FAUC, HUA); Huila: Suaza, via Neiva, vereda CampoHermoso, costado izquierdo de la via, km 40 [1°46'11.71" N, 75°46'37.52" O], 2190m,1 Apr 2016, Peña 542 (COL) Chocó: San José del Palmar, Cerro Torrá, Filo de Cumbre,[04º48’ N, 76º29’ W], 2700–2800m, 21 Aug 1988, Silverstone-Sopkin 4633 (Paratypes:CHOCO, CUVC, MO, UC n.v.). Nariño: Reserva Natural La Planada, sendero entre ElHondón y Los Horquetas, Bosque primario, 01º10’ N, 78º00’ W, 1820m, 31 Jan [no yeargiven], Herrera 9302, 9193 (Paratypes: PSO, CUVC, UC n.v.); Carretera LaVictoria−Monopamba, [01º10’ N, 78º00’ W], 2500m, 10 Jan 1973, Hagemann & Leist1898 (holotype: PSO; isotype: COL).
ACKNOWLEDGEMENTSDS would like to thank the primatologist Javier Garcia for promoting the biologicalresearch in Caquetá department. DS is in debt to the Biology program, especially toProfessor Alexander Velasquez-Valencia and the staff of the Natural History Museum ofthe University of Amazonia for providing infrastructure. Special thanks to Drs MaryGibby and Klaus Melhtreter for comments that substantially improved the manuscript.The University of the Amazonia funded this research.
200 FERN GAZ. 20(5):197-201. 2017

REFERENCESIUCN, 2001. IUCN Red List Categories and Criteria: Version 3.1. [online]. Available
from http://www.iucnredlist.org/info/catego- ries_criteria2001 [accessed 24 Oct.2016]
MORAN, R.C. 1995. Polypodiaceae. Pp. 333–366. in: G. DAVIDSE, M. SOUSA, ANDS. KNAPP (eds). Flora Mesoamericana, Psilotaceaea a Salviniceae. México:Universidad Nacional Autónoma de México.
MURCIA, G.U.G., HUERTAS, M.C., RODRÍGUEZ, J.M., & CASTELLANOS, H.O.2010. Cambios multitemporales de los bosques y otras coberturas de la Amazoniacolombiana, a escala 1:100.000, en el periodo 2002 al 2007. Instituto Amazónicode Investigaciones Científicas SINCHI. Bogotá, D. C.
MURILLO-P., M.T., J. MURILLO-A. & A. LEÓN-P. 2016. Serpocaulon obscurinerviumD. Sanín. In: BERNAL, R. GRADSTEIN, S.R. & CELIS, M. (Eds.). 2015. Catálogode plantas y líquenes de Colombia. Instituto de Ciencias Naturales, UniversidadNacional de Colombia, Bogotá. http://catalogoplantascolombia.unal.edu.co
SANÍN, D. 2011. Revisión taxonómica de Serpocaulon (Polypodiaceae), para laCordillera Central y el Macizo Colombiano. MSc thesis, 96 pp., Universidad deCaldas, Manizales, Colombia.
SANÍN, D. 2014. Serpocaulon obscurinervium (Polypodiaceae), a new fern species fromColombia and Ecuador. Plant Ecol. Evol. 147: 127–133.
SANÍN, D. 2015. Serpocaulon tayronae (Polypodiaceae), a new species from the SierraNevada de Santa Marta, Colombia. Phytotaxa 213: 243–252.
SMITH, A.R., KREIER, H.-P., HAUFLER, C.H., RANKER, T.A. & SCHNEIDER, H.2006. Serpocaulon (Polypodiaceae), a new genus segregated from Polypodium.Taxon 55 (4): 919-930.
SANÍN ET AL: SERPOCAULON OBSCURINERVIUM 201

202 FERN GAZ. 20(5). 2017

POLYSTICHUM LONCHITIS (DRYOPTERIDACEAE:PTERIDOPHYTA) A MONTANE-ALPINE FERN SPECIES,MONITORED SINCE 1978 IN THE NETHERLANDS
P. BREMER & A. SMIT
Roelingsbeek 1, 8033 BM Zwolle, The Netherlands([email protected])
Key words: Polystichum lonchitis, Kuinderbos, monitoring, mortality
ABSTRACTPolystichum lonchitis was found in 1978 for the first time in The Netherlands ona dry ditch bank, on calcareous sand within a planted woodland (Kuinderbos). Insubsequent years six other specimens were found at three other sites, in awoodland, on a scree in a chalk stone quarry and on a wall, and four of theseplants became fertile. Mortality in five out of seven plants resulted from a smalllandslide, habitat destruction, browsing and frost damage. Long term monitoringof two plants showed the impact of rainfall and frost on vitality. No recruitmentwas observed in the spore shadow of adult plants.
INTRODUCTIONPolystichum lonchitis (L.) Roth. is a montane-alpine species, with a Holarcticdistribution. In North America it is a species found mostly in boreal and subalpine regionsbetween 0 and 3200 m (Morin, 1993). In Europe the species is found between 900 and2100 m and rarely at lower altitude up to 300 m, except for the coastal area of Norway(Mutter et al., 1998). It is also found in Greenland, Iceland and in isolated parts of Russiabut is rare in Japan (Dostál & Reichstein, 1984). Within the UK it is more widespread inthe central Scottish Highlands (c. 610 – 915 m). Occasionally it descends to loweraltitudes in steep upland river valleys (c. 150 m) and lower levels near the west coast,with its cool summers (Page, 1982). In 1978, Polystichum lonchitis was recorded for thefirst time in the Netherlands during a survey of 120 km of dry ditches in the Kuinderbos.Two plants were found growing at the sides of two adjacent dry ditches on the bottom ofthe former Zuiderzee at ca. 1 m below sea level (Bremer, 1980). Since then this smallpopulation has been monitored annually for the first 15 years and less intensely insubsequent years. After 1979 the species was found at three other sites in the Netherlands,and monitored annually near Lelystad (Smit, 1989). This paper analyses these recordsto try to determine whether a montane-alpine plant species is able to build up a viablelowland population.
METHODSTwo plants (plant 1 and 3, both in Flevoland, Table 1) were monitored for 24 years(Kuinderbos) and 27 years (Lelystad) with annual counting of the number of fronds andthe maximum frond length and fertility at the end of the summer season or start ofautumn, when frond formation had stopped and sori produced. Change in maximumfrond length, change in number of fronds and the log transformed product of bothparameters (always year t to year t + 1) were used as response variables to rainfall andthe number of frost days. The average values in Flevoland were used for annual rainfall
FERN GAZ. 20(5):203-213. 2017 203
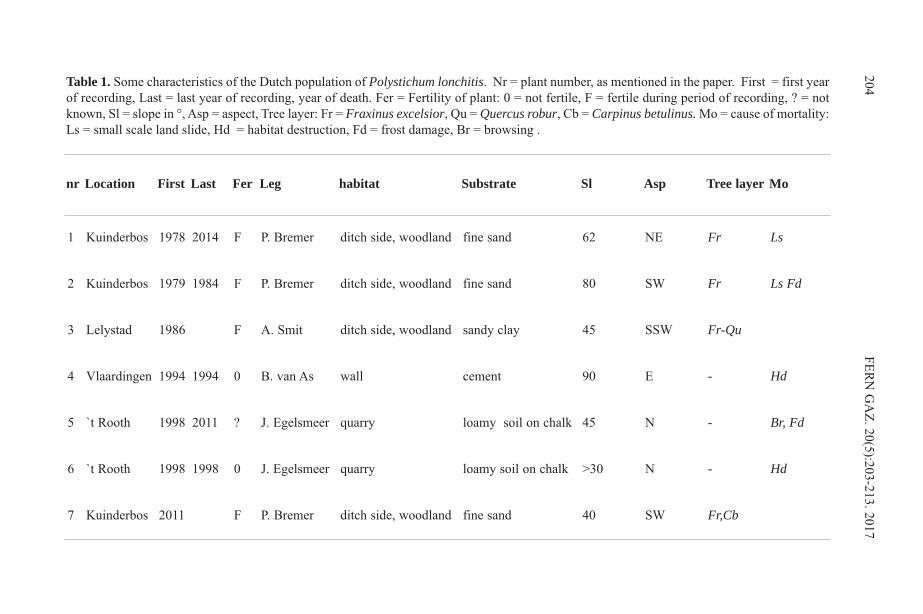
204 FERN G
AZ. 20(5):203-213. 2017
nr Location First Last Fer Leg habitat Substrate Sl Asp Tree layer Mo
1 Kuinderbos 1978 2014 F P. Bremer ditch side, woodland fine sand 62 NE Fr Ls
2 Kuinderbos 1979 1984 F P. Bremer ditch side, woodland fine sand 80 SW Fr Ls Fd
3 Lelystad 1986 F A. Smit ditch side, woodland sandy clay 45 SSW Fr-Qu
4 Vlaardingen 1994 1994 0 B. van As wall cement 90 E - Hd
5 `t Rooth 1998 2011 ? J. Egelsmeer quarry loamy soil on chalk 45 N - Br, Fd
6 `t Rooth 1998 1998 0 J. Egelsmeer quarry loamy soil on chalk >30 N - Hd
7 Kuinderbos 2011 F P. Bremer ditch side, woodland fine sand 40 SW Fr,Cb
Table 1. Some characteristics of the Dutch population of Polystichum lonchitis. Nr = plant number, as mentioned in the paper. First = first yearof recording, Last = last year of recording, year of death. Fer = Fertility of plant: 0 = not fertile, F = fertile during period of recording, ? = notknown, Sl = slope in °, Asp = aspect, Tree layer: Fr = Fraxinus excelsior, Qu = Quercus robur, Cb = Carpinus betulinus. Mo = cause of mortality:Ls = small scale land slide, Hd = habitat destruction, Fd = frost damage, Br = browsing .
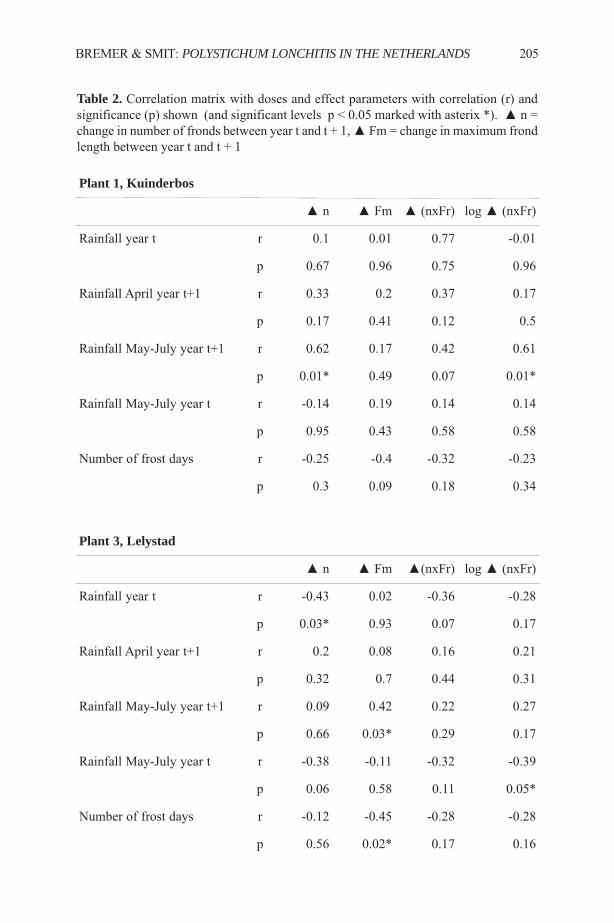
Plant 1, Kuinderbos
▲ n ▲ Fm ▲ (nxFr) log ▲ (nxFr)
Rainfall year t r 0.1 0.01 0.77 -0.01
p 0.67 0.96 0.75 0.96
Rainfall April year t+1 r 0.33 0.2 0.37 0.17
p 0.17 0.41 0.12 0.5
Rainfall May-July year t+1 r 0.62 0.17 0.42 0.61
p 0.01* 0.49 0.07 0.01*
Rainfall May-July year t r -0.14 0.19 0.14 0.14
p 0.95 0.43 0.58 0.58
Number of frost days r -0.25 -0.4 -0.32 -0.23
p 0.3 0.09 0.18 0.34
Plant 3, Lelystad
▲ n ▲ Fm ▲(nxFr) log ▲ (nxFr)
Rainfall year t r -0.43 0.02 -0.36 -0.28
p 0.03* 0.93 0.07 0.17
Rainfall April year t+1 r 0.2 0.08 0.16 0.21
p 0.32 0.7 0.44 0.31
Rainfall May-July year t+1 r 0.09 0.42 0.22 0.27
p 0.66 0.03* 0.29 0.17
Rainfall May-July year t r -0.38 -0.11 -0.32 -0.39
p 0.06 0.58 0.11 0.05*
Number of frost days r -0.12 -0.45 -0.28 -0.28
p 0.56 0.02* 0.17 0.16
BREMER & SMIT: POLYSTICHUM LONCHITIS IN THE NETHERLANDS 205
Table 2. Correlation matrix with doses and effect parameters with correlation (r) andsignificance (p) shown (and significant levels p < 0.05 marked with asterix *). ▲ n =change in number of fronds between year t and t + 1, ▲ Fm = change in maximum frondlength between year t and t + 1

in year t, rainfall in April in year t + 1, rainfall in May-July in year t and t + 1, as weexpected drought in spring or early summer to play a role. The number of frost days perwinter (maximum day temperature below 0°C) was used as indicator for the lowtemperature impact in winters regional data used (www.knmi.nl). These were the sameparameters as used by Bremer & Jongejans (2010) in their study on the demography ofAsplenium scolopendrium L. SPSS 10.1 was used to calculate correlations. Per locationdata about habitat, substrate, inclination, aspect and composition of the tree layer andherb layer were collected in order to classify the sites and vegetation.
RESULTSHabitat and vegetationOver a period of 39 years seven plants were found at the former sea floor in Flevoland,in the city of Vlaardingen and in a chalk quarry in the southern part of the Netherlands(Figure 1). The Kuinderbos plants grew in dry ditch sides within plantations of Fraxinusexcelsior L., accompanied by mosses of the Fissidentietum taxifolii bryophytecommunity (Bremer, 1999) and fern species such as Asplenium scolopendrium andPolystichum aculeatum (L.) Roth. Near Lelystad the plant grew under a canopy ofFraxinus excelsior L., with Oxyrhynchium hians (Hedw.) Loeske, Eurhynchium striatum(Hedw.) Schimp. and Geum urbanum L. The non-native Potentilla indica (Andrews)Wolf had invaded the site. This vegetation is related to the Fraxino-Ulmetum, a woodlandcommunity confined to clayey soils in the Netherlands. Both `t Rooth plants were foundgrowing among flint stones deposited at a chalk quarry. The vegetation at this site wascharacterized by mass growth of Asplenium trichomanes L., Gymnocarpium robertianum(Hoffm.) Newman and Asplenium scolopendrium. In the bryophyte layer Encalyptastreptocarpa Hedw. was a prominent species. Even Asplenium ruta-muraria L. wasgrowing among flint stones, one of the few examples where it shows terrestrial growthin the Netherlands. The vegetation belongs to the Asplenietum ruto-murario-trichomanes,a community strongly linked to wall habitats (Segal, 1969) (OV39, British NationalVegetation Classification, Rodwell, 1992). The Vlaardingen site was by a wall, whichwas destroyed two years after the discovery of Polystichum lonchitis. The vegetationbelonged to the Filici-Saginetum, a wall community with Dryopteris filix-mas (L.) Schottand Asplenium scolopendrium. All sites have a high pH in common and are related togradients. There was no difference in aspect comparing N-NE with S-SW (χ2 = 0.14 ns)(Table 1).
Distribution and trendThe number of plants varied from one to two plants per location. The small populationin the Netherland showed a positive trend until 2010 with decline in recent years (Figure2). Most plants were found as adults; the Vlaardingen plant was a juvenile/subadult plant,rather small in size, as is often the case with wall dwelling fern species. Five plants diedbecause of landslip, habitat destruction, frost damage and browsing or because of acombination of these factors (Table 1). At Vlaardingen, prior to wall destruction, theplant was conserved and planted in a garden but was subsequently lost through theftwithin two years. At `t Rooth quarry one of the sites was destroyed in favour of ponddigging because of a threatened toad species. The other plant died as a result of browsingin combination with frost damage (Bremer & Egelsmeer, 2015). In the Kuinderbos, oneplant slipped to the bottom of the dry ditch after a small scale land slip (initiated by heavyrain fall) and drowned as the normally dry ditches contain water (0.1 – 0.3 m depth) in
206 FERN GAZ. 20(5):203-213. 2017
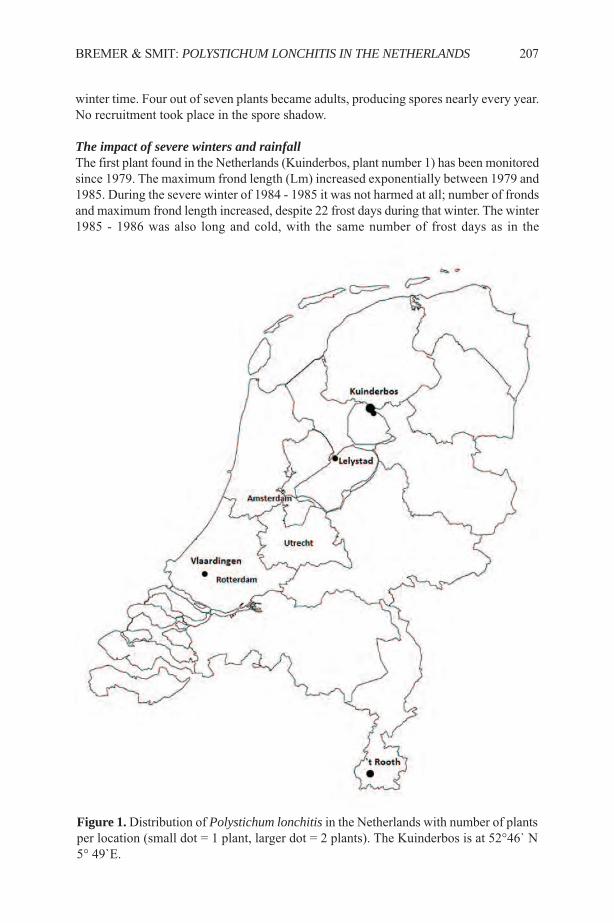
winter time. Four out of seven plants became adults, producing spores nearly every year.No recruitment took place in the spore shadow.
The impact of severe winters and rainfallThe first plant found in the Netherlands (Kuinderbos, plant number 1) has been monitoredsince 1979. The maximum frond length (Lm) increased exponentially between 1979 and1985. During the severe winter of 1984 - 1985 it was not harmed at all; number of frondsand maximum frond length increased, despite 22 frost days during that winter. The winter1985 - 1986 was also long and cold, with the same number of frost days as in the
BREMER & SMIT: POLYSTICHUM LONCHITIS IN THE NETHERLANDS 207
Figure 1.Distribution of Polystichum lonchitis in the Netherlands with number of plantsper location (small dot = 1 plant, larger dot = 2 plants). The Kuinderbos is at 52°46` N5° 49`E.
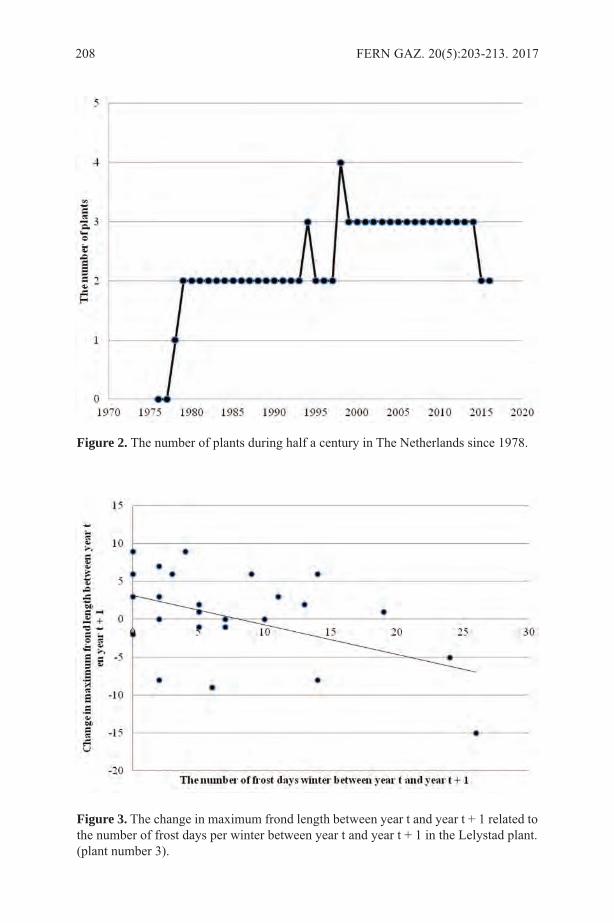
208 FERN GAZ. 20(5):203-213. 2017
Figure 2. The number of plants during half a century in The Netherlands since 1978.
Figure 3. The change in maximum frond length between year t and year t + 1 related to the number of frost days per winter between year t and year t + 1 in the Lelystad plant. (plant number 3).
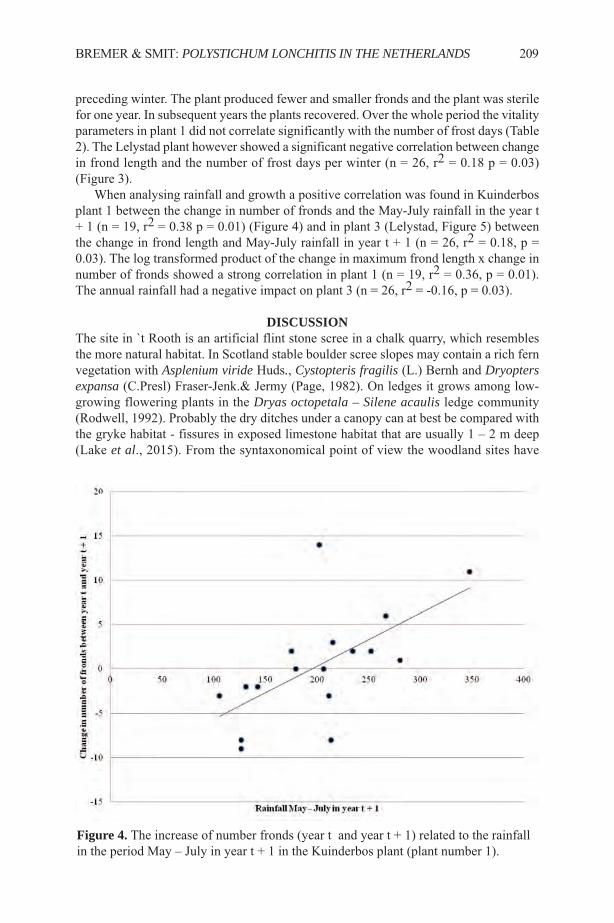
preceding winter. The plant produced fewer and smaller fronds and the plant was sterilefor one year. In subsequent years the plants recovered. Over the whole period the vitalityparameters in plant 1 did not correlate significantly with the number of frost days (Table2). The Lelystad plant however showed a significant negative correlation between changein frond length and the number of frost days per winter (n = 26, r2 = 0.18 p = 0.03)(Figure 3).
When analysing rainfall and growth a positive correlation was found in Kuinderbosplant 1 between the change in number of fronds and the May-July rainfall in the year t+ 1 (n = 19, r2 = 0.38 p = 0.01) (Figure 4) and in plant 3 (Lelystad, Figure 5) betweenthe change in frond length and May-July rainfall in year t + 1 (n = 26, r2 = 0.18, p =0.03). The log transformed product of the change in maximum frond length x change innumber of fronds showed a strong correlation in plant 1 (n = 19, r2 = 0.36, p = 0.01).The annual rainfall had a negative impact on plant 3 (n = 26, r2 = -0.16, p = 0.03).
DISCUSSIONThe site in `t Rooth is an artificial flint stone scree in a chalk quarry, which resemblesthe more natural habitat. In Scotland stable boulder scree slopes may contain a rich fernvegetation with Asplenium viride Huds., Cystopteris fragilis (L.) Bernh and Dryoptersexpansa (C.Presl) Fraser-Jenk.& Jermy (Page, 1982). On ledges it grows among low-growing flowering plants in the Dryas octopetala – Silene acaulis ledge community(Rodwell, 1992). Probably the dry ditches under a canopy can at best be compared withthe gryke habitat - fissures in exposed limestone habitat that are usually 1 – 2 m deep(Lake et al., 2015). From the syntaxonomical point of view the woodland sites have
BREMER & SMIT: POLYSTICHUM LONCHITIS IN THE NETHERLANDS 209
Figure 4. The increase of number fronds (year t and year t + 1) related to the rainfall in the period May – July in year t + 1 in the Kuinderbos plant (plant number 1).

210 FERN GAZ. 20(5):203-213. 2017
Figure 5. The Lelystad plant (plant number 3) accompanied by Potentilla indica in2007, 21 years after the first record (photo P. Bremer)
Figure 6. The plant recorded in the Kuinderbos in 2015 (plant number 7), firstdiscovered four years after the first record (photo P. Bremer).

much in common with, the Fraxinus excelsior-Acer campestre-Mercurialis perenniswoodland and in particular the Geranium robertianum-subcommunity, where sometimesit is found under a miniaturised version of this woodland in the Carboniferous Limestonegrykes in the Yorkshire Dales (Rodwell, 1998).
The montane-alpine species was not expected to colonise a former sea floor. Inneighbouring countries lowland sites for this species are not known (e.g.www.floraweb.de). In the year Polystichum lonchitis was recorded for the first time,Asplenium viride Huds. was also found in the Kuinderbos (Bremer, 1981), and somemontane bryophytes had been found on dry ditch sides on the former seafloor [e.g.Distichium inclinatum (Hedw.) Bruch & Schimp. (Bremer & Ott, 1979) and fungi (e.g.Suillus tridentinus (Bres.) Singer]. The Kuinderbos is the only known site within theNetherlands with a small viable population of the montane Knautia dipsacifolia Kreutzer(Meijer & Bremer, 1986). All these species came by long distance dispersal by wind,with Knautia dipsacifolia being the exception (probably with humans as vector).
Page (1982) states that the species occasionally descends to lower altitudes near theextreme west coasts of Scotland because of its cool summers. At open sites in theNetherlands the preferred N-E aspect was confirmed. In the Dutch woodlands it thriveswith a SW preference, although in April, without the protection of a sprouted canopy, itcan be warm in these ditches. A high temperature is probably compensated byevaporation of water, as dry ditches always contain water in this time of the year.
There is a continuous flow of diaspores from mountainous areas spreading over theNetherlands, implicating that under optimal conditions plants can establish from spores.But the chance of establishment is dependent on the extent of an area which is suitablefor colonisation, spore density, spore longevity (sporebank) and the ability ofintragametophytic selfing (Klekowski 1979). In the Kuinderbos 300 ha of calcareousdeposits on peat drained by more than 100 km of dry ditches provided a large area, withcolonization by numerous rare fern species (Bremer 2007, de Groot 2012). De Groot(2012) showed in four species [Asplenium scolopendrium, A. trichomanes, Polystichumaculeatum, P. setiferum (Forssk.) Woynar] a very high selfing capacity in both diploidand polyploid species. Single-spore establishment following long-distance dispersal mayhave resulted in selection for selfing genotypes. There was no recruitment from the adultplants of diploid Polystichum lonchitis, at woodland sites. This might indicate that selfingis less likely in a second generation and that the species depends on intergametophyticcrossing. There is a very low chance that spores from other genotypes are able toestablish within the spore shadow of an adult plant because of the large distance to Britishand Central European populations. Polystichum lonchitis does have ArbuscularMycorrhiza (Harley & Harley, 1987), but it is unknown whether fungi play a crucial rolein establishment. With only two plants left, stochastic extinction might occur. In theNetherlands Atlantic species such as Asplenium scolopendrium, Dryopteris borreriNewman, D. affinis (Lowe) Fraser-Jenk. and Polystichum setiferum are increasing innumber and becoming more common. Global warming with more precipitation insummer and higher temperatures in winter plays a role. With its fluctuating smallpopulation Polystichum lonchitis remains a threatened species on the Dutch Red List(Sparrius et al., 2012). Landslip within dry ditches, frost damage, habitat destruction andbrowsing played a role in the mortality of five out of seven plants. One plant lived forca. 45 years. Northridge writes that one of five Fermanagh plants (N. Ireland) was foundas a mature plant in 1979 and was still in excellent condition 32 years later(www.habitas.org.uk/priority/species). Although plants in the UK seem to be long-lived,
BREMER & SMIT: POLYSTICHUM LONCHITIS IN THE NETHERLANDS 211

no demographic studies are known for this species. In the Netherlands frost damageplayed a role in vitality and survival. One would expect a montane-alpine species to moreadapted to low temperatures, yet effects of severe winters were observed in four of sevenplants, with plants becoming smaller, with lower number of fronds per plant, smallerfronds per plant or lower fertility. The same was observed in Asplenium scolopendrium,but this species has an Atlantic distribution, and is susceptible to frost. In this speciessnow cover can protect plants from heavy damage (Bremer & Jongejans, 2010). Themore leathery fronds of Polystichum lonchitis (see Figure 6) seem to be more protectivethan those of Asplenium scolopendrium. However, our data suggest that snow covermight be crucial, a suggestion also made by Dostál & Reichstein (1984), indicatinggrowth in snow-rich habitats. Mutter et al. (1998) did not find a role of extensive snowcover. For many subalpine species snow cover is a prerequisite for survival duringwinters (Körner & Larcher, 1988) as snow protects from low temperatures. Buriedbeneath a layer of snow plants are exposed to milder temperatures close to 0°C. Withoutsnow cover desiccation plays a role or excessive natural radiation may result in frostdamage. As old fronds might serve as nutrient storage organs frost damage impacts thenew generation of fronds (Landi et al., 2014).
Within the Netherlands long term monitoring of a complete plant population isexceptional. While in other groups, e.g. breeding birds, annual monitoring iscommonplace, it is much more difficult in plants. Individuals in many species are moredifficult to identify [e.g. Pteridium aquilinum (L.) Kuhn] and a complete survey of alarge area time consuming. Plant species involved mostly have a restricted number ofpopulations, as is the case in Woodsia ilvensis (L.) R.Br. (McHaffie, 2010) and more longterm monitoring in ferns is needed (Sharpe & Mehlteter, 2010).
ACKNOWLEDGEMENTSWe thank Ben van As, Olaf Opdenkamp and Jan Egelsmeer for their data and KlausMehltreter for reviewing the paper.
REFERENCESBREMER, P. 1980. The ferns of the Kuinderbos (The Netherlands), the establishment
op 23 species in a planted forest. Acta Botanica Neerlandica 29(5/6): 351 – 357.BREMER, P. 1981. Polystichum lonchitis (L.) Roth. en Asplenium viride Huds. nieuw
voor Nederland. Gorteria 10(7): 113 - 120.BREMER, P. 1999. Mosgemeenschappen op greppelkanten in de Flevolandse bossen.
Stratiotes 19: 56 - 65.BREMER, P. 2007. The colonisation of a former sea floor by ferns. PhD thesis,
Wageningen University.BREMER, P. &. OTT, E.C.J. 1990. The establishment and distribution of bryophytes
in the woods of the IJsselmeerpolders, The Nether lands. Lind bergia 16:3 - 18.BREMER, P. & JONGEJANS, E. 2010. Frost and Forest Stand Effects on the Population
Dynamics of Asplenium scolopendrium. Population Ecology 52: 211 - 222.BREMER, P. & EGELMEERS, J. 2016. De Lansvaren in Nederland. Varenvaria 29(1):
8 - 13.GROOT, A. DE. 2012. The fate of a colonizer: successful but lonely? The establishment
of inter- and intraspecific diversity in ferns by means of long-distance dispersal. PhDthesis, Utrecht University.
DOSTáL, J. & REICHSTEIN, T. 1984. Polystichum. In: G. Hegi. Illustrierte Flora von
212 FERN GAZ. 20(5):203-213. 2017

Mitteleuropa. Band I. Teil 1: 169 - 187. HARLEY, J.L & HARLEY, E.O. 1987. A check-list of mycorrhiza in the British flora.
New Phytologist 105: 1 - 102. KLEKOWSKI, E.J., 1979. The genetics and reproductive biology of ferns. In: A.F. Dyer
(red.). The experimental biology of ferns. Pp. 133 – 170. London, Academic Press.KÖRNER, C & W. LARCHER, 1988. Plant life in cold climates. In: Plants and
Temperature. Eds. S. Long & F.I. Woodward. Cambridge: pp. 25 – 57. LAKE, S., D. LILEY, R.STILL & SWASH, A., 2015. Britain`s Habitats. A guide to the
Wildlife Habitats of Britain and Ireland. Princeton University Press.LANDI, M., ZOCCOLA, A., BACARO, G & ANGIOLINE, C., 2014. Phenology of
Dryopteris affinis ssp. affinis and Polystichum aculeatum: modelling relationshipsto the climatic variables in a Mediterranean area. Plant Species Biology 29(2): 129 –137.
MCHAFFIE, H., 2010. Recent monitoring of wild populations of Woodsia ilvensis.Pteridologist 5(3): 191 - 192.
MEIJER, K. & BREMER, P. 1986. Knautia dipsacifolia Kreutzer nieuw voor Nederland.Gorteria 13: 74 - 77.
MORIN, N.R. 1993. Flora of North America. North of Mexico. Volume 2. Pteridophytesand Gymnosperms. North America Editorial Committee. Oxford University Press.
MUTTER, H., H.J.B. BIRKSAND & ODLAND, A. 1998. The comparative ecology ofPolystichum aculeatum, P. braunii and P. lonchitis in Hordaland, western Norway.Nordic Journal of Botany 18(3): 267 - 288.
PAGE, C.N., 1982. The Ferns of Britain and Ireland. Cambridge University PressRODWELL. J.S. 1992. British Plant Communities Volume 3 - Grasslands and montane
communities. Cambridge University Press. RODWELL, J.S. 1998. British Plant Communities. Volume 1. Woodlands and scrub.
Cambridge University Press. SEGAL, S. 1969. Ecological notes on wall vegetation. PhD thesis, Universiteit of
Amsterdam.SHARPE , J.M. & K. MEHLTRETER, K. 2010. Ecological insight form fern population
dynamics. In: K. MEHLTRETER, L.R. WALKER & J.M. SHARPE. Fern Ecology:61 – 110. Cambridge University Press.
SMIT, A. 1989. De naaldvarens (Polystichum-soorten) van het Overijssels Hout bijLelystad. Gorteria 15: 90 – 94.
SPARRIUS, L., B.ODE & R.BERINGEN, 2012. Basisrapport voor de Rode Lijstvaatplanten 2012. FLORON. http://www.floron.nl/publicaties/rode-lijst-2012
BREMER & SMIT: POLYSTICHUM LONCHITIS IN THE NETHERLANDS 213

BOOK REVIEW
An Annotated Checklist of Indian Pteridophytes Part – 1 (Lycopodiaceae toThelypteridaceae. Fraser-Jenkins, C.R., Gandhi, K.N., Kholia, B.S. & Benniamin,A. 2017. pp 562. Bishen Singh Mahendra Pal Singh, Dehra Dun, India.
This new checklist of Indian pteridophytes is a welcome addition to our knowledge ofthe diversity of Asian ferns and lycophytes, and adds to the recent publications onpteridophytes from China (Wu et al. 2013) and Nepal (Fraser-Jenkins et al. 2015). Thisvolume covers Indian Lycopodiaceae, Selaginellaceae, Isoetaceae, Equisetaceae and 19fern families (Psilotaceae to Thelypteridaceae).
In addition to the detailed checklist that forms the bulk of the volume, there is a shortintroduction that sets the work in context and a summary of the numbers of families,genera and species that are to be included in both Parts 1 and 2. The checklist itself isfollowed by a series of 37 colour images, that range from some significant herbariumspecimens to many field photographs of living plants, either in close-up or as habitatshots, and all of these providing a beautiful visual introduction to the vast range ofdiversity among Indian pteridophytes.
In generating the information for this detailed checklist, the authors have combinedan extensive literature review with detailed studies of herbarium specimens anddistributions from both public and private herbaria from India and elsewhere, and soundfield knowledge. Critical comparison has been made with records from adjacent areas,and this has led to the inclusion of 34 families, 130 genera, 1107 species and 50 additionalsubspecies for the whole of the checklist, of which 23 families, 59 genera, 572 speciesand 32 additional subspecies are covered in Part 1. In addition, for each of the genera,the authors provide lists of excluded taxa; this is very valuable in clarifying thepresence/absence of taxa for which past records have proved to be erroneous, often theresult of confusion of nomenclature.
The new checklist has a traditional taxonomic framework; the authors haveconsidered recent taxonomic approaches built on phylogenetic analysis of DNA sequencedata but have given greater weight to morphological classification. One of the strengthsof this checklist is in the detailed citations for families, genera and species, includingcitation of types and synonyms, and information on distribution within and outwith India.For many of the species there are extensive notes to clarify previous reports, to point outmis-identified material or instances where there has been mis-application of names,sometimes in attempted lectotypification. This detailed clarification of the names used(and those excluded) in the checklist is invaluable for other researchers, and comparisoncan be made with, for example, fern taxa from elsewhere in Asia.
In a taxonomic work of this detail it is not surprising to find that some 55nomenclatural changes have been made, for example, Mickelopteris Fraser-Jenk., gen.nov. in the family Pteridaceae, and these changes can be identified readily as they arelisted in bold in the taxonomically arranged index of accepted and excluded taxa at theend of Part 1.
The authors are to be congratulated on a very welcome addition to current knowledgeof the rich diversity of pteridophytes in Asia.
214 FERN GAZ. 20(5). 2017

REFERENCESFraser-Jenkins C.R., Kandel D.R. & Pariyar S. 2015. Ferns and Fern-allies of Nepal,
Volume 1. pp 508. Kathmandu, Nepal. Wu, Z., Raven, P.H. & Hong, D. 2013. Flora of China, vols. 2-3, Lycopodiaceae through
Polypodiaceae. Missouri Botanical Garden Press.
M. Gibby
FERN GAZ. 20(5). 2017 215

INSTRUCTIONS FOR AUTHORS
PAPERS should not usually exceed 20 printed pages and are generally expected to beconsiderably shorter. As well as reports of original research, review articles and shortnotes, e.g. new records, are encouraged. The senior author should supply an email addressto facilitate correspondence. REVIEWS should cover recent developments in any aspect of research on ferns andlycopods. This can include unpublished research by the author, and/or the possibledirection of future research. They should be aimed at the general reader who may not bean expert in the chosen topic. Proposals for a review should be discussed first with thereview editors.MANUSCRIPTS should be submitted in English (British) in electronic format in10-point Times New Roman font and double spaced. Electronic versions of text andtables should be compatible with WORD, with figures as pdf or jpg files, and sent asemail attachments or CDroms. All manuscripts will be refereed.THE TITLE should reflect the content of the paper and be in BOLD CAPITALS(11-point) and centrally aligned. Generic and specific names should be in italics and anytitle containing a generic or specific name should be followed by the family name inbrackets e.g.
TRICHOMANES SPECIOSUM (HYMENOPHYLLACEAE)IN SOUTHERN SPAIN
AUTHOR ABBREVIATIONS should follow Pichi Sermolli’s (1996) Authors ofscientific names in Pteridophyta, Royal Botanic Gardens, Kew.MAIN HEADINGS: should be in BOLD CAPITALS (10-point) and centrally aligned.SUBSIDIARY HEADINGS: should be in bold, the first letter of each word in capitals,the rest in lower case and left-aligned.AUTHORS’ NAMES AND FULL ADDRESSES (including email address): followthe title and are centrally aligned.KEY WORDS: up to ten.ABSTRACT: should reflect the content of the paper.FIGURES: there is no distinction between photographs and line drawings in numbering.All should be presented in a form ready for reproduction, colour, TIF format and 300dpi (please contact editor with queries), with a scale bar where appropriate. Lettering ornumbers (Arabic) should be in the bottom left using uppercase Times Roman. Figurecaptions should be on a separate sheet.TABLES: can be printed in either portrait or landscape format. Authors should considerthis when preparing tables. Authors should ensure that tables fit the printed page size ina legible form.PAGESIZE: The Gazette has a useable page size of 123 × 204 mm. This can be used inportrait or landscape mode but any supplied artwork should not be larger than this.MEASUREMENTS: follow the metric system.CHECKLISTS: follow the format of Baksh-Comeau, Fern Gaz. 16(1, 2): 11- 122.REFERENCES: follow the style of a recent issue of The Fern Gazette, e.g.:-HOOKER, W.J. 1864. Species Filicum, 5. Dulau & Co., London.MORTON, C.V. 1947. The American species of Hymenophyllum, section
Sphaeroconium. Contr. U.S. Natl. Herb. 29(3): 139-201.STEVENSON, D.W. & LOCONTE, H. 1996. Ordinal and familial relationships of
pteridophyte genera. In: CAMUS, J.M., GIBBY, M. & JOHNS, R.J. (Eds)Pteridology in perspective, pp. 435-467. Royal Botanic Gardens, Kew.
JOURNAL ABBREVIATIONS: should follow Botanico Periodicum Huntianum &Supplements.Alterations from the original text at proof stage will be charged for unless they are minorpoints of detail. A pdf will be provided free to the senior author.
216 FERN GAZ. 20(5). 2017

REVIEWSIluRIan-DEVonIan oRIgInS of fERnS anD lycophytES -
What WE knoW, What WE nEED to fInD out
P.G. GENsElDepartment of Biology, University of North Carolina
Chapel Hill, NC 27599-3280, UsAEmail: [email protected]
Key words: silurian-Devonian, lycophytes, basal euphyllophytes, ferns evolution,phylogenetic relationships.
aBStRactThis represents a synopsis of current knowledge of the siluro-Devonian fossilrecord concerning evolution of lycophytes and ferns. This is the time period whenseveral taxa or lineages at different grades of organisation existed that may beinformative about the origins of these groups or structures typical of these groups.Considerable new data, including earlier first appearances of lineages and plantstructures, new data about siluro-Devonian lycopsids or basal euphyllophytes,and new whole plant reconstructions of small to tree-size plants in both lineages,have been published in recent years. It is not possible to be completelycomprehensive, but the taxa discussed are either central to established ideas, orprovide new information in relation to phylogenetic relationships and evolutionarytrends. It remains difficult to trace the phylogenetic relationships of early plantsrelative to extant lineages. New data are reviewed which may be important inreassessing homology of characters and/or hypotheses of such relationships or indetermining which taxa to exclude. Including fossils in estimates of relationshipsof these major lineages of plants will provide a more accurate and comprehensiveunderstanding of the past history of seedless vascular plants.
IntRoDuctIonA consensus classification of extant seedless vascular plants by PPG1 (2016), reflectingmolecular and some morphological phylogenies (smith et al., 2006b; Kenrick &Crane, 1997; Pryer et al., 2001; 2009; schneider et al., 2009; and others) recogniseslycopodiopsida (with three families and collectively referred to as lycophytes) and agrade “euphyllophytes” which consists of two clades - seed plants and Polypodiopsida(Figure 1). The latter includes four subclasses, Equisetidae, Ophioglossidae, Marattiidaeand Polypodiidae- all referred to as ferns.
lycopodiopsida appears to represent a monophyletic group defined by both molecularand morphological characters that diverged at least in the mid-silurian if not earlier(Kenrick & Crane, 1997; Nickrent et al., 2000; Renzaglia et al. 2000; Pryer et al., 2001;2004; Qiu et al., 2007; li et al., 2014) but see Hao and Xue (2013) for a differentinterpretation. Relationships of several putative stem lycophytes such as some Cooksoniapertoni, Aberlemnia (formerly Cooksonia) caledonica, Renalia and several others, somewithin more derived extinct clades where critical characters are missing, are poorlyresolved (Kenrick & Crane, 1997; Gensel & Berry, 2001; Hao & Xue, 2013). Additionaldata and more whole plant reconstructions of several Upper silurian or lower Devonianpolysporangiophytes are needed to address these issues.
FERN GAZ. 20(6):217-242. 2017 217

The term “fern” or “fern-like” now encompasses the majority of extant and extinctnon-lycophytic seed-free vascular plants and several extinct “stem” euphyllophytes(trimerophytes), those with “peripheral permanent protoxylems” such as cladoxylopsids,iridopterids, and in some cases, rhacophytes and stauropterids (Kenrick & Crane, 1997;Rothwell & stockey, 2008; Rothwell & Nixon, 2006; Berry & stein 2000; Berry & Wang2006). This results in a broad view of what represents a “fern”. This is tied up inconsiderations of evolution of the so-called megaphyllous leaf via modification of branchsystems. However it is clear such leaves evolved many times and possibly from differentparts of lateral branch systems. Furthermore, using criteria such as presence/absence ofa leaf gap and simple vs branched venation to distinguish microphyll vs megaphyll isnot clear-cut. For a more complete discussion of these issues, see Tomescu (2009) andlater sections of this paper. The term megaphyll is intentionally not used in this paper.
Fossils were intentionally excluded from the PPG1 classification, because “thephylogenetic affinities of most extinct plants are rather unclear” (p.565). While a practicalstarting point for names, it, along with molecular- derived phylogenies of only extantforms, implies misleading relationships by ignoring some of the major discoveries in thefossil record of the past several decades. For example, stating that ferns are the closestliving relative to seed plants is correct when only living plants are considered, but ignoresthe existence of extinct plants with unique combinations of characters representing oneor more now-extinct lineages, notably progymnosperms, but also others, indicating amore distant relationship between seed plants and ferns. While molecular-basedphylogenies may suggest a close relationship among the extant subclasses, incorporationof fossil and morphological data (Figure 2) suggest they are not monophyletic (Rothwell& Nixon, 2006).
The fossil record, while still incomplete, presently suggests each Polypodiopsidasubclass (PPG1 2016) differs in time of first appearance and possibly in origin. Firstappearances of undoubted extinct members of each clade are as follows: equisetophytes-the late Devonian, filicaleans (extinct Polypodiidae) - Early Carboniferous, Marattiidae-late Carboniferous, and Ophioglossidae -Cenozoic. Thus time gaps exist between thelater appearing taxa/lineages and putative early representatives of those clades. Findingmorphologically based synapomorphies to unite extant and extinct putative progenitor
218 FERN GAZ. 20(6):217-242. 2017
figure 1. A simplified phylogeny of ferns and lycophytes, showing only the majorlineages, modified from Figure 1 of the Pteridophyte Phylogeny Group 1 (PPG1) study.

or sister lineages is difficult. Characters that have been commonly employed among theDevonian age euphyllophytes are 1) branching systems, from which leaves are believedto have evolved and part or all of which are often interpreted as “leaf-equivalents”, 2)stelar anatomy and 3) protoxylem location. sometimes interpretations are retrospective,with extant leaf morphologies influencing ideas of what constitutes a leaf-equivalent.Not everyone agrees on which part or parts of a branching system constitutes a leaf-equivalent in each lineage, and lastly, for some, all leaves are considered homologous(ignoring the fossil record) rather than evaluating the fossil plants within the context oftheir time and environment (Berry & stein, 2000) and taking a more “bottom-up outlook”(Rothwell & stockey, 2008) of extinct plants relative to extant ones. Recently acquiredfossil data and whole- plant reconstructions, coupled with new developmentalinformation or tests of certain character distributions, may improve assessment ofhomology of structures and character state transformations and ultimately phylogeneticrelationships (Tomescu, 2009; Corvez et al., 2012 and authors cited therein). Additionalwork on understanding variation in anatomy also is needed. As our understanding ofcertain plant types improve, the fossil record may also indicate which possible progenitorcandidates to preclude.
What DoES thE foSSIl REcoRD cuRREntly tEll uS?
GENsEl: SILURIAN-DEVONIAN ORIGINS OF FERNS AND LYCOPHYTES 219
figure 2. From Rothwell and Nixon, 2006 showing a tree based on a combination offossil and extant plant morphological and molecular characters. Inclusion of fossilscauses changes in postulated relationships of so-called “fern-like plants” relative tofilicalean (=Polypodiidae) ferns. Terminology is that used by Rothwell and Nixon(2006). lignophytes refer to progymnosperms and seed plants. OP= ophioglossid fernsand psilotophytes. Palaeozoic leptosporangiate ferns include taxa such asMetaclepsydropsis, Ankyropteris, Psalixochlaena, Botryopteris, and others.
220 FERN GAZ. 20(6):217-242. 2017
figure 3. A stratigraphic chart showing major events in plant evolution or firstappearances of certain plant groups or plant structures, spanning Ordovician toTournaisian.

gEnERalItIES1) Everything happened earlier (Figure 3): Dispersed cryptospores and trilete sporesprovide evidence for Kingdom Plantae in the early Middle Ordovician to late MiddleOrdovician respectively (Rubenstein et al., 2014; steemans et al., 2009; 2010; Wellmanet al., 2008; 2015; Kenrick et al., 2012; Edwards et al., 2014, Edwards & Kenrick, 2015,and references cited therein). Cryptospore types and diversity increase until the earliestDevonian, trilete spores diversify from silurian on. Earliest forms occur in Gondwananregions, suggesting radiation of cryptospore producers and basal polysporangiates fromGondwanan microcontinents towards laurussia (Wellman et al., 2015; steemans et al.,2010). The absence of megafossils during this time span may partly reflect the lack ofknown terrestrial sediments in the Ordovician and silurian (Kenrick et al., 2012; Wellmanet al., 2015). some tiny Upper silurian to lower Devonian plants produce cryptospores,termed cryptophytes (Edwards et al., 2014), and others produce trilete spores, somepossibly representing precursors of a bryophyte or polysporangiophyte grade (Edwardset al., 2014). Unfortunately, only some dispersed spores can be tied to parent plants,which limits establishing evolutionary/phylogenetic links between cryptophyte,bryophyte or polysporangiate lineages, and thus to either lycophytes or fern-likeprecursors (Edwards et al., 2014).
2) Innovations: The silurian to Middle Devonian vascular plant diversificationdemonstrates a greater increase in plant diversity, size and complexity than previouslyknown. silurian to Early Carboniferous taxa exhibit a trend from considerablemorphological/anatomical plasticity to more predictable body plans or architecture inyounger, more derived forms (Bateman et al., 1998). Recent research suggests a transitionfrom apparently rhizomatous forms to upright trees by the Middle to late Devonianamong lycopsids and euphyllophytes (Figure 3), tree size ranging from 3 to at least 8mtall and of varying trunk diameters (4 to ~20 cm wide). some formed new vegetationassociations, most notably early forests. In euphyllophytes, no laminate leaves and no,or limited, secondary tissues exist (thus having alternative means of mechanical support)(stein et al., 2007; 2012; Giesen & Berry, 2013; Berry & Marshall, 2015).Archaeopteridalean forests of large trees with abundant secondary tissues dominatedsome regions in the late Devonian. Evidence of early evolution of leaves and rootsoccurs from late silurian on, as discussed in greater detail later.
Another major innovation, aligned xylem meeting criteria for secondary xylem andinterpreted as an early type of vascular cambium, is demonstrated by the PragianAmoricaphyton (Gerrienne et al., 2011; strullu-Derrien et al., 2013; 2014; Gerrienne &Gensel, 2015) and Emsian plants from eastern Canada (Gerrienne et al., 2011; Hoffman& Tomescu, 2013; Gensel in prep.). It is unknown if these possessed uni- or bifacialcambia. That, and lack of fertile remains, renders affinities and potential relevance as tohow vascular cambia evolved or to fern and lycophyte vs. lignophyte evolution unclear.Uni- and bifacial vascular cambia originated more than once, as they are known in severalextinct lineages from Middle Devonian on, including lycopsids (uni), progymnosperms(bi), early seed plants (bi), and possibly cladoxyls (unclear if secondary xylem or primary,aligned metaxylem).
Heterospory is established by the Middle Devonian in lycopsids and someeuphyllophytes (the incipiently heterosporous lower Devonian Chaleuria; the UpperDevonian archaeopterids and some lower Carboniferous stauropterids). The MiddleDevonian putative ovule precursor, Runcaria (Gerrienne et al., 2004) and numerous pre-
GENsEl: SILURIAN-DEVONIAN ORIGINS OF FERNS AND LYCOPHYTES 221

ovules of late Devonian (Famennian) age indicate an initial radiation of early seed plantsat nearly the same time as heterosporous euphyllophytes occurred (Prestianni &Gerrienne, 2010), but prior to appearance of all fern clades except Equisitidae.
lycophytE EVolutIonThe presence of late Wenlock Cooksonia sp. from Avalonia (Edwards & Feehan, 1980),and the Upper silurian (ludlow) occurrences of several zosterophylls, putative stemlycophytes, and the lycopsid Baragwanathia from Bathurst Island, Canada and/orAustralia (Kotyk et al., 2004; Rickards, 2000) indicate several grades and clades ofrhizomatous, homosporous lycophytes at that time and an early origin for lycophytes.Zosterophylls, stem lycopsids, pre-lycopsids and lycopsids occur widely in the EarlyDevonian. strobilate probable zosterophylls such as Distichophytum and plants similarto Bathurstia first appear in the late silurian and similar forms are diverse in the EarlyDevonian.
Zosterophyllopsids are leafless but exhibit similar anatomy and sporangialmorphology and dehiscence as lycopsids. Kaulangiophyton, lacking vascularized leaves,Asteroxylon with transfusion tissue in “leaves” (Hueber, 1992), and fertileDrepanophycus spp. all lack association of sporangia with leaves, and sometimes areconsidered pre-lycopsids (Gensel & Andrews, 1984; Gensel & Berry, 2001). leaves ofdiffering morphology occur in the siluro-Devonian Baragwanathia, in the pre-lycopsidDrepanophycus, the putative lycopsid Zhenglia (Hao et al., 2006) and in some Pragiantaxa of uncertain affinity from China such as Adoketophyton (li & Edwards, 1992; Haoet al., 2003). Mid-Upper Devonian and younger lycopsids exhibit mostly laminateuniveined lycophylls, some of which are much-divided (Protolepidodendrales) or havelaciniate to fimbriate margins (e.g. Longostachys, Wuxia). Ideas about how lycophyllsmay have evolved vary but are unresolved, ranging from change via progressivevascularization of enations, sterilization of sporangia (Crane & Kenrick, 1997), or byreduction of branch systems (reviewed in Tomescu, 2009).
Growth habit among these basal groups (stem taxa, zosterophylls, pre-lycophytes,early lycopsids) is either rhizomatous or tufted (Gensel et al., 2001; Raven & Edwards,2001; Gensel & Berry, 2001; Hao et al., 2010). Rooting structures appear to have arisenmore than once, ranging from downward trending axes (some Zosterophyllum spp. ofearliest to early Devonian age), to part of a bifurcation of a rhizome (H or K branching),with one half being stem-like and bearing enations or leaves, the other portion lackingthose structures and appearing root-like (Bathurstia, Drepanophycus - Gensel et al.,2001). An un-named lycopsid from the lower Devonian of Wyoming differs in that theinitial K-branch produces a “root-bearing axis”, from which dichotomizing root-likestructures depart (Matsunaga & Tomescu, 2016). These authors suggest rooting structuresin lycophytes are de novo structures rather than modified stems. Hetherington and Dolan(2017) note commonalities among lycopsid root morphology as well as disparities,recognizing multiple types of structures that bear the branched roots. None of the earlyforms are anatomically preserved, thus features that distinguish roots from stems (rootcap, root hairs, etc) in younger plants are lacking (Raven & Edwards, 2001; Gensel etal., 2001; Kenrick & strullu-Derrien, 2014).
The order Protolepidodendrales, plants characterized by much divided or hastateleaves, epiphyllous oval to possibly round sporangia, and faintly to deeply lobedprotosteles, is established by the Emsian (Leclercqia spp.) and taxa are much morediverse and widespread in the Middle and early late Devonian (Gensel & Berry, 2001).
222 FERN GAZ. 20(6):217-242. 2017

Most specimens may represent more distal or aerial branches and presumably wererhizomatous or in one case, climbing (L. uncinata Xu et al., 2011). Oval to elongatesporangia attach to the upper surface of sporophylls identical to vegetative leaves.Leclercqia complexa from the Middle Devonian Gilboa, NY locality is known to beligulate; L. complexa, L. cf. complexa and L. uncinata yield spores showing them to behomosporous (Grierson & Bonamo, 1979; Richardson et al., 1993; Gensel & Kasper,2005; Gensel & Albright, 2006; Wellman et al., 2009; Xu et al., 2011). spores and ligulesare unknown for other genera. Rooting structures, which might aid in betterunderstanding of relationships, are unknown in all taxa. These plants are often consideredto be putative precursors to the ligulate heterosporous clades selaginellales and Isoetales,but better understanding of all plant parts would allow a more precise assessment of theirrelationship to other extinct and extant clades. As noted below, additional putativelytransitional forms exist, although many are incompletely preserved.
A major change in growth habit and reproduction occurred in the late Mid Devonian,as shown by the bipolar/pseudobipolar, approximately half metre tall tree-like taxaLongostachys and Chamaedendron (Cai & Chen, 1996; schweitzer & li, 1996), both ofuncertain affinity, in China. In situ (and some isolated) lycopsid stems from the upperMiddle/lower Upper Devonian of svalbard, consisting of approximately 1m tall,incomplete trunk pieces, and identified as Protolepidodendropsis pulchra, are interpretedas a paleotropical forest (present in a low paleolatitude) by Berry and Marshall (2015).Sublepidodendron, especially S. songziense Wang et al. 2003, from the late Devonianof China, was at least a metre tall. A smaller, very slender, upright and unbranched lateDevonian lycopsid, Clevelandodendron ohioensis (Chitaly & Pigg, 1996), exhibits alobed/branched base and a bisporangiate strobilus reminiscent of some lepidodendrids.Wider stemmed lycopsids such as Cyclostigma, Lepidosigillaria, and the unidentifiedtree lycopsid from Gilboa, NY (Haughton, 1859; Chaloner, 1968; Kräusel & Weyland,1949; stein et al., 2012) also occur in late Mid to early late Devonian in laurussia.These taxa not only differ in size (length, width), but also in branching (presence/absence,type), leaf base patterns, leaves, rooting and reproductive structures. They also clearlydemonstrate the existence of upright lycopsids of varying heights, diameters, andbranching architecture by Middle to late Devonian time, coexisting with the presumablyrhizomatous protolepidodendrids and presaging growth habits and reproductive strategiespresent among the abundant Carboniferous and younger lycopsid lineages. Variousisolated anatomically preserved stem remains, when viewed in comparison to anatomyin more extensively preserved plants, also show differences in some features, not yetwell understood. secondary tissues occur in lycopsids from the Middle Devonian.Tracheid wall pitting among different taxa varies from one or more of the followingpitting types: scalariform, pitted, circular bordered pitted, scalariform with fibrilsconnecting the bars or pitlet sheets. Much remains to be learned about its possibletaxonomic implications.
Different types of stem bases with rooting structures occur among these taxa.Longostachys and Chamaedendron stems (and possibly Clevelandodendron) terminatein a series of downward trending, bifurcating root-like structures, but details are poorlyunderstood. The two major rooting structures of arborescent isoetalean lycopsidsdominant in the Carboniferous occur by the Middle Devonian. The earliest records ofcormose lycopsid bases may be those recorded by Xu and Wang (2016) from the lateMiddle-late Devonian Hujiersite Fm, China and the bases of the more extensivelypreserved Protolepidodendropsis pulchra, of similar age from svalbard (Berry &
GENsEl: SILURIAN-DEVONIAN ORIGINS OF FERNS AND LYCOPHYTES 223

Marshall, 2015). late Devonian Leptophloem and Otzinachsonia also exhibit cormosebases. A more branched, stigmarian type rooting system (Wang et al., 2003) occurs inSublepidodendron spp from China. laurussian forms are less easy to interpret. Pigg(2001) suggests that cormose and stigmarian forms represent variations of a singlerooting type, perhaps being influenced by ecological factors.
some of these Devonian lycopsid stems or branches bear sporangia aggregated intostrobili, ranging from mono- to bisporangiate (Longostachys, Chamaedendron,Sublepidodendron, Cyclostigma), while others apparently have alternating fertile andvegetative zones (Monilostrobus Wang & Berry, 2003; Wuxia Berry et al., 2003), as alsooccurs in younger tree lycopsids. The Upper Devonian Wuxia is interesting in thatmegasporangiate “cones/fertile zones” occur just below branching points of axes butpresumed microsporangiate ones are terminal (Berry et al., 2003). Hoxtolgaya robustaXu et al. (2012), Middle Devonian of China, is interpreted as arborescent based on 9cmwide stems of unknown length, and exhibits sporangia yielding spores of one size,interpreted as homospory, but preservaton is too incomplete to determine with certainty.In strobili or fertile zones, sporophylls are either identical to vegetative leaves, ormodified with a broad fimbriate to spiny base and distal lamina, as in the Middle to UpperDevonian small tree-like lycopsids such as Longostachys (Cai & Chen, 1996),Chamaedendron (schweitzer & li, 1996) or Wuxia (and others) (Berry et al., 2003).
Berry et al. (2003) note that variation in fertile regions/structures and in stemanatomical features occurs earlier and more broadly in China than laurussia, suggestingthat perhaps China may have been an important area of early evolutionary innovation inclade(s) from which the major lineages of arborescent, heterosporous lycopsids of theCarboniferous and later arose. More whole plant reconstructions of several of theselycopsids are needed, and particularly, clarification of the organisation of basal regions,presence/absence of branching, anatomical features, and type of reproductive structuresamong individual plants, in order to better understand their relationships to both earlierand later occurring lineages to resolve relationships of these several plants and other taxaall considered as part of the isoetalean clade.
The record of herbaceous, homosporous lycopsids (e.g. lycopodiaceae) is far lesswell known in the Devonian and Carboniferous. Lycopodites oosensis from the lateDevonian of Germany (Kräusel & Weyland, 1937) may represent one, but should berestudied. The lower Carboniferous herbaceous form Hestia possesses too fewcharacters to assess affinities (Bateman et al., 2007) but might represent an eligulateform. Younger fossils have been assigned to the genus Lycopodites but it is difficult todetermine the affinities of these leafy stems; one has been found to represent conifertwigs, others exhibit a mix of lycopodiacean and sellaginellacean characters but areusually allied to selaginellaceae (skog & Hill, 1992; Thomas, 1992). Rowe (1988)identified a rhizomatous plant from the late Early Carboniferous Drybrook sandstone asan early selaginallalean, Selaginellites resimus, but little is known about early evolutionof selaginellales.
several questions concerning early lycophyte evolution remain or are only partiallyanswered by recent data. The paraphyly of some Zosterophyllum species and stemlycophytes remains unresolved despite new data. Much remains to be clarified aboutoverall construction and relationships of the numerous Middle-late Devonianprotolepidodendraleans and small tree lycopsids, as discussed above, including howmany times have structures such as strobili, ligules, rooting structures, and leaf abscissionarisen and in what combinations? How do they relate to later-occurring lineages? And
224 FERN GAZ. 20(6):217-242. 2017

again, questions remain about the early history of herbaceous, eligulate, homosporouslycopodiaceae and ligulate, heterosporous selaginellaceae through much of theDevonian and Carboniferous and even early Mesozoic.
EuphyllophytE EVolutIonExtinct Devonian plants included in the Euphyllophyta mostly exhibit much branchedaerial axes, some terminated by mostly fusiform sporangia. Anatomy consists of variousprotosteles (haplo-, actino-, dissected). Pertinent to fern evolution are stemeuphyllophytes, such as Psilophyton and Pertica, the predominantly Mid-UpperDevonian Cladoxylopsida (=Pseudosporochnales [or Cladoxylales] + Iridopteridales),the enigmatic, but putative iridopterid Rhacophyton, and possibly stauropteridaleans.The latter groups have been considered by many authors to be derived from atrimerophyte ancestor, and possible fern-like or fern precursors. Reasons vary; somederive from cladistic studies, others from older comparative studies. While lackinglaminate leaves, either the whole or part of their lateral branch systems (lBs) areinterpreted as showing possible stage(s) in the evolution of a leaf, as mentioned earlier(Kenrick & Crane, 1997; Gensel et al., 2001; Boyce & Knoll, 2002; sanders et al., 2007;Tomescu, 2008; Galtier, 2010; Corvez et al., 2012). These occurred simultaneously butat different rates in different lineages (Galtier, 2010), and are not homologous withinferns or between “ferns” and seed plants. secondly, some of the above taxa exhibitperipherally located protoxylem in various types of actino- or dissected protosteles(where anatomy is known) that differs from that present in aneurophytaleanprogymnosperms and early seed plants. The latter, and some basal euphyllophytes,exhibit radiate protoxylem (Beck & stein, 1993), in which actinosteles or variants thereofhave a central protoxylem, which appears to extend out each arm of the stele to producelateral traces. These are considered mesarch but this depends on whether the protoxylemat the tips of arms persist, a feature presently not well understood. Anatomy of this typeis typical of aneurophytalean progymnosperms, possibly some of the larger (Pertica–grade) trimerophytes, stenokolealeans, and other plants of uncertain affinity. However,it remains unclear if these two protoxylem patterns represent a good indicator ofphylogenetic relationships. For example, scheckler et al. (2006), in a re-study ofLangoxylon stockmans, a taxon of uncertain affinity, point out that Langoxylon stemspossess a mixture of these two characters. Pseudosporochnus hueberi lacks a centralprotoxylem, but exhibits peripheral protoxylems plus several separate protoxylems alongthe midplane of an arm (stein & Hueber, 1989), thus not all early euphyllophytes can beso easily categorized.
The term “peripheral loop”, initially referring to an anatomical feature ofCarboniferous zygopteridalean petioles, has been used to describe protoxylem featuresin many Devonian plants. As originally defined, a peripheral loop consists of a centralrod of parenchyma surrounded by protoxylem (leclercq, 1970) located near the marginof an elongated vascular bundle. similar structures are termed the same in stems ofDevonian taxa. several studies show that most of the latter represent protoxylem lacunae,resulting from break-down of thinner walled protoxylem during development or representa different organisation of protoxylem with associated parenchyma (scheckler, 1974;scheckler et al., 2006; stein, 1982). This suggests that the presence of a so-calledperipheral loop should be carefully evaluated in any Devonian plant, and is notphylogenetically useful. Furthermore, care should be taken in use of the term“clepsydroid” and in any phylogenetic implications that might follow, because strap-
GENsEl: SILURIAN-DEVONIAN ORIGINS OF FERNS AND LYCOPHYTES 225

shaped strands in main axes or lateral branch traces in Devonian plants expand at theirends in preparation for producing the next higher order traces, thus appearing similar tothe dumbbell-shaped petiole trace so termed among some early ferns.
The earliest putative euphyllophyte laminate leaf may date from the PragianEophyllophyton (Hao, 1988; Hao & Beck, 1993), considered basal and sister to othereuphyllophytes by Hao and Xue (2013) but considered of uncertain affinity by this author.While laminate leaves are absent in most other coeval taxa, they occur in the Early tolate Devonian Platyphyllum (=Flabellifolium?) and the late Devonian Ellesmeris (Hillet al., 1997), enigmatic Shougangia bella (Wang DM et al., 2015), archaeopteridaleansand early seed plants, appearing earlier in time in lignophytes than in filicales (Galtier,2010). Clearly, resolution of what constitutes homologous structures in lBss amongthese different much- branched plants, and more documentation of the first appearanceof laminae, would address remaining questions concerning number of times and modesof leaf evolution in these groups and non-homology of so-called megaphylls in thesegroups.
claDoXylS, IRIDoptERIDS anD RhacophytES- callED “fERn-lIkE” But?
starting in the Mid-Eifelian and extending through the late Devonian, several taxareferable to Cladoxylopsids s.l. (e.g. including iridopterids, Berry & stein, 2000) andrhacophytes are part of a more complex vegetation, some of the former representingsmall to large trees recently described as major components of in situ Middle to earlyUpper Devonian forests in the UsA and Germany (stein et al., 2007; 2012).Concomitantly, major revision of taxon concepts for Pseudosporochnus (Berry & Fairon-Demaret, 1997; 2002), Calamophyton (now includes Duisbergia, Cladoxylon, and mostspecies of Hyenia - Fairon-Demaret & Berry, 2000; Berry, 2005; Giesen & Berry, 2013)and Eospermatopteris/Wattieza (stein et al., 2007; 2012) has resulted in critical newinformation about the architecture of these leafless trees.
pseudosporochnaleansPseudosporochnus trees (Figure 4) produce rows of branches that abscise at their base;the branches are digitately divided and produce higher-order, divided laterals that arethree-dimensional to subopposite and alternate, final divisions being vegetative and non-laminate or fertile (Berry & Fairon-Demaret, 2002). Calamophyton is a smaller tree, witha swollen base, stems enlarging near apex and bearing vertical rows of lateral branchsystems that then divide digitately (Figure 5), and produce higher order helically arrangeddichotomizing vegetative or fertile non-laminate units. Older branch systems break offabove their base, leaving the Duisbergia type morphology (Giesen & Berry, 2013).Eospermatopteris is larger, with a swollen base, a wide stem, and crown of digitatelydivided branch systems of the Wattieza type (stein et al., 2007; 2012), abscising at theirbase. Terminal fusiform sporangia are known for all of these, being either recurved(Calamophyton, Wattieza) or erect (Pseudosporochnus). These apparently represent theearliest tree architectural types among euphyllophytes, being similar to some palms,cycads and tree ferns (stein et al., 2007) in stem and root structure.
Where anatomy is known in the pseudosporochnaleans, it varies from being amultistranded, anastomosing stele, producing similar but smaller multi-stranded stelesin lateral branches, as in Calamophyton or Pseudosporochnus (Figure 6), to beingcomprised of individual radially oriented strands near the periphery of the stem adjacent
226 FERN GAZ. 20(6):217-242. 2017
GENsEl: SILURIAN-DEVONIAN ORIGINS OF FERNS AND LYCOPHYTES 227
figure 4. A recent reconstruction of the cladoxylopsid Pseudosporochnus, showing itscrown of relatively large branch systems, each of which is closely dichotomous so as toappear digitately divided. Reproduced from Berry and Fairon-Demaret, 2002, Figure 9,Int. J. Plant sci. 163(5) 699-713. © 2002 by The University of Chicago.

228 FERN GAZ. 20(6):217-242. 2017
figure 5. Calamophyton primaevum. Drawing of a stem showing two features- theremnant bases of branches along the stem and parts of the attached, digitately arrangedbranches at the crown. B. suggested anatomy of Calamophyton (Duisburgia) based onMustafa (1978) showing primary xylem in black, surrounded by aligned, possiblysecondary xylem (lines). Redrawn from Giesen and Berry, 2013; stem is from Figure5a (in part), anatomy redrawn from Figure 17d.

to a large pith in main trunk (Figure 5) as in the presumed base of Calamophyton stem(Giesen & Berry, 2013) and Eospermatopteris/Wattieza (Boyer & Matten, 1996). Opiniondiffers as to whether some of these plants exhibit a limited amount of aligned metaxylemor secondary tissues. Coeval progymnosperms possess a eustele and bifacial vascularcambium. Habit, branching, aspects of rooting and internal construction of the largepseudosporochnalean cladoxylopsids might mean they already had diverged to the extentthat they have little relationship with extant polypodiid or equisetid lineages, but insteadrepresent a distinct lineage that flourished globally, then went extinct without anysurvivors (see Rothwell & stewart, 1993).
IridopteridaleansNumerous Middle Devonian plants from laurussia, Gondwana, and China compriseanother lineage of Middle to Upper Devonian euphyllophytes, the Iridopteridales (stein,1982; Fu et al., 2011) based on anatomy, morphology, or a combination of thesepreservation forms (Figure 7). These plants as presently known are smaller thancladoxylopsids, consisting of stems with a deeply ribbed mesarch actinostele comprisedof two to three centrally united arms, each arm dividing one or more times to form sixto twenty arms/ribs depending on the taxon. A single protoxylem region exists near theend of each arm (Figure 7 A-C). Branching pattern is essentially whorled, eachprotoxylem pole emitting a trace that supplies either a lateral branch or a smallerdichotomous appendage, the two types occurring in the same whorl (Figure 7D).sometimes branching occurs in only part of a whorl, with successive partial whorls offset(stein, 1982). sporangia terminate ultimate dichotomizing appendages borne fromsecond order branches- some upright and some recurved (Berry &stein, 2000,Compsocradus). Other, often less extensively preserved, taxa attributed to theCladoxylopsida are not treated here. Relationship to Polypodiidae is unclear; depending
GENsEl: SILURIAN-DEVONIAN ORIGINS OF FERNS AND LYCOPHYTES 229
figure 6. Anatomy of a lateral branch system of Pseudosporochnus, which ischaracteristic of many cladoxylopsids; the vascular bundles are arrayed in a mannerinterpreted as representing the beginning of a dichotomy. Redrawn from stein andHueber, 1989.

on what combination of characters are selected, they may be closer to Equisetidae; ifothers, of uncertain affinity (stein et al., 1984; Berry & stein, 2000).
Rhacophyton and putative relativesRhacophyton (several spp) exhibits some characters that presage a fern type oforganisation, but also possesses secondary tissues and some morphology reminiscent ofprogymnosperms or iridopteridaleans. somewhat rhizomatous to upright axes (?stems)bearing adventitious roots in R. ceratangium (Cornet et al., 1976) produce single or pairedmuch divided lateral branch systems which overall appear three dimensional, with moretwo-dimensional second order axes. The secondary axes depart oppositely, and rangefrom dichotomous to slightly pseudomonopodial. Fertile lateral branch systems are morecomplex, each pair with one vegetative and one fertile unit. The vegetative segments areelongate and bear divided higher order non-laminate ultimates. The fertile units are muchdivided and recurved, forming rounded masses of dividing axes terminating in fusiformsporangia with elongate tips. Anatomy of less extensively preserved, more uprightappearing stems of R. zygopteroides was described by leclercq (1951) from poorlypreserved specimens, and interpreted as an actinostele with unequally sized arms,although these sections have deterioriated so this cannot be confirmed. similarly arrangedlateral branches produce clepsydroid shaped steles. Both were described as surroundedby secondary xylem. In R. ceratangium, both main axis and lateral branches areconsidered to have produced an elongated clepsydroid-shaped primary xylem,surrounded by secondary xylem with rays (Cornet et al., 1976; Dittrich et al., 1983).Traces to higher order axes also are known. R. condrusorum schultka (1978) is similar.Ellesmeris Hill et al. (1997) exhibits a simpler, but similar overall architecture, althoughthe ultimate units are laminate, and fertile structures unknown. Anatomy is not intact,but the authors suggest main axis and lateral branch produce an elongate haplostele, withprotoxylems near the tips of the arms. Knowing if all orders of branching possessed thesame type of stele, or alternatively that stelar configuration changed from radial to
230 FERN GAZ. 20(6):217-242. 2017
figure 7. line drawings of anatomy and stem portion of taxa included inIridopteridalean cladoxylopsids. A. Ibyka amphikoma; B. Iridopteris eriensis; C.Arachnoxylon kopfii; D. Portion of stem and branching structure of Anapaulia moodyi;whorls of branches alternate with dichotomizing structures similar to ultimate branchlets;each lateral branch bears second and ultimate order branches. A-C redrawn from stein,1982. D redrawn from Berry and Edwards, 1996.

bilateral from main axis to lateral branch is of some significance in consideringrelationships and what to interpret as stem vs. leaf in these plants. For the present theyusually are considered a separate lineage, Rhacophytaceae, although some authorspreviously placed them as a basal member of the zygopteridaleans and others considerthem more allied to progymnosperms. Berry and Wang (2006) suggest, intriguingly, thatRhacophyton shares some characteristics of iridopteridaleans.
StEm EuphyllophytES anD RElEVancE to EVolutIon of latERfoRmS
Trimerophyte grade euphyllophytes exhibit either dichotomous, anisotomous or stronglypseudomonopodial branching of varying degrees of predictable cladotaxy. some of theseterminate in fusiform sporangia, ranging from loosely (Psilophyton-grade plants) todensely (Pertica) clustered. Anatomy is known from parts of aerial systems of somePsilophyton species, while anatomy is unknown in published Pertica species. However,Gothanophyton and some undescribed plants at the Pertica-grade provide data aboutpossible anatomical variation in these entities, showing some differentiation betweenribbed main axes and lateral branches, and possibly indicating evolutionary trajectoriesdifferent from Pertica, although affinities of most of these are poorly understood.
Psilophyton-grade basal euphyllophytesPsilophyton Dawson (Banks et al.) Doran (1980), initially based on dichotomous toslightly pseudomonopodially branched axes, where anatomically preserved, exhibits ahaplostele with centrally located protoxylem (centrarch) with little or no differentiationbetween main axis and lateral branch traces (Figure 8A). It thus appears the leastspecialized of stem euphyllophytes, but that may be deceptive. At the same time, plantsof this type may be more closely related to polypodiids, or lineages leading to polypodiidsand equisetids, than many later-appearing taxa.
Over 14 species of Psilophyton are now recognized, some based on morphology, onefrom anatomy only (P. coniculum) and others with both, although anatomy so far is notextensively known in most species. Based on current knowledge, preservation rangesfrom nearly complete plants (P. crenulatum) to more fragmentary remains. Phylogeneticanalyses involving more than one species of Psilophyton indicate the genus is notmonophyletic (Kenrick & Crane, 1997; Hao & Xue, 2013) and comparative evaluationof known species further suggests the possibility that these species might represent morethan one taxon.
Psilophyton crenulatum Doran (1980), an apparently rhizomatous plant with uprightbranch systems, exhibits dense regions of mostly vegetative branching, some of whichappear immature (apices present) while more distally, spacing of vegetative and fertilelateral branches varies. Psilophyton dawsonii, based mainly on anatomy of a 2.5 cm longspecimen (Banks et al., 1975), possesses regions of closely spaced vegetative lateralbranches. The more distally located fertile lateral branches exhibit a trace suggested todiffer in shape from vegetative ones (Banks et al., 1975). some short dichotomouslydivided lateral appendages are considered possible aphlebiae or roots. These features arenot known for any other Psilophyton species, including P. coniculum (Trant & Gensel1985), known only from anatomy (no sporangia attached). In this species, more closelyspaced branching occurs mainly in lateral branches.
Comparison of the many other Psilophyton species indicates that relative distributionof vegetative vs fertile regions is unknown, partly because the genus is best recognized
GENsEl: SILURIAN-DEVONIAN ORIGINS OF FERNS AND LYCOPHYTES 231

morphologically from fertile specimens. In many instances the number of specimensavailable or mode of preservation limits information. Where numerous specimens areavailable, study shows branching is variously spaced. Frequently, lateral branches areincomplete and finding completely vegetative branch systems is difficult.
It was previously questioned whether certain previously described species ofPsilophyton conform sufficiently to the concept of the genus. P. kräuselii, from theMiddle Devonian of Bohemia (Obrhel, 1959) is too poorly preserved to determineaffinity, being based on fragmentary fertile and sterile remains (Hao & Xue, 2013; thispaper). Hao and Xue (2008) also question the generic identity of P. szaferi, P. striatum(Wang & Berry, 2001) and P. dapsile (Kasper et al. 1974) because they are either toopoorly known or lack twisted sporangia. Examination of P. dapsile sporangia in theauthor’s collection suggest that while small, they do show some torsion. Psilophytonszaferi, described by Zdebska (1986) from the lower Devonian of Poland, consists of aportion of a lateral branch system with upright sporangia borne in loose clusters moresimilar to those in Pertica and should be restudied. Psilophyton microspinosum from theEarly Devonian of Maine (Kasper et al., 1974) exhibits a more rigid pattern of lateralbranches and erect, untwisted sporangia, suggesting its affinity should be re-evaluated.
The few morphologically preserved Pragian Armoricaphyton and the Emsianunnamed plant from Canada (Gerrienne et al., 2011; strullu-Derrien et al., 2014;Gerrienne & Gensel, 2015) with an early type of secondary tissue are similar in stem
232 FERN GAZ. 20(6):217-242. 2017
figure 8. Diagrammatic drawings of small basal euphyllophytes; predominant mode ofbranching shown above, anatomy below. Axes approx. 2-5 mm. A. a generalized drawingof branch system of Psilopyton. Anatomy a centrarch haplostele, producing circular (orpossibly square, if fertile) lateral branches. B. Unnamed plant from Emsian, NewBrunswick (Gensel in prep). Ribbed, twice branched axes known. some stems withelongate haplostele producing lateral trace, others indicate same type of haplostelesurrounded by secondary xylem. C. Armoricaphyton chateaupannense, Pragian, France(Gerrienne et al., 2011; Gerrienne and Gensel, 2016). Ribbed, twice branched axes.Primary xylem a round haplostele, surrounded by secondary xylem. Basal part of lateralbranch with secondary xylem, more distal region without. Dark circles or lines in centerof steles indicate protoxylem, white area primary xylem, lines = secondary xylem.

diameter and branching, to Psilophyton (Figure 8 B, C). No fertile remains are known.All of these taxa, along with Pertica type trimerophytes showing preserved anatomy,exhibit P-type tracheid wall patterning sensu Kenrick and Crane, 1997 (Gensel,unpublished data). This character may be less useful at lower taxonomic levels, as thecombination of characters evident in these plants suggest broader disparity among theseearly simpler plants than previously realised.
Morphologically preserved Middle Devonian plants such as Planatophyton(Gerrienne et al., 2014), Pauthecophyton (Xue et al., 2012), and Tsaia (Wang & Berry,2001) are similar to Psilophyton in size and some aspects of branching but differ eitherin specifics of branching, sporangial orientation and/or dehiscence mode. These mayrepresent distal regions of much larger plants or, alternatively, fragments of plants similarto Psilophyton in size. Additional study of these plants may reveal features useful indetermining if they represent one, or several, lower to Middle Devonian genera and/orlineages and clarify their postulated phylogenetic relationships.
larger basal euphyllophytes similar to PerticaPertica and similar-appearing plants (Figure 9A-C) are taller and more robust in width,with a dominant main axis and variously arranged lateral branches. Pertica spp. exhibitmostly isotomously divided lateral branches with terminally borne, closely spaced, erectfusiform sporangia. Anatomy is unknown in published Pertica species.
A new genus, initially considered Pertica-like but definitely an unrelated taxon(Figure 9A), has a dominant main axis, helically arranged lateral branches that are singleor paired, and, in contrast to Pertica, dichotomously- divided ultimate units located alongtheir length, some sterile and some terminating in fusiform sporangia. Main axis anatomyis an actinostele (usually) with a single central protoxylem and a protoxylem near theend of each arm (Gensel, 1984). The lateral branch trace(s) are ovoid. There is noevidence of secondary xylem.
An undescribed plant from the lower Devonian of New Brunswick is similar toPertica spp. in morphology (Figure 9B), with a dominant main axis, helical lateralbranches, and dichotomous laterals terminating in tight clusters of fusiform sporangia(Gensel, in prep.). Anatomy is unusual, consisting of a “winged” stele in which fourmajor arms emanate from an elongate central region, but with one arm dividing further(Figure 9B). At least two types of lateral branch traces are formed, differing from mainaxis stelar configuration, but preservation is incomplete. Do other perticas have similaranatomy or might there be variation?
Gothanophyton Remy et Hass (Figure 9 C), with a 1cm wide main axis and single orpaired lateral branches, exhibits a “winged” stele, consisting of an elongated centralregion with two major arms extending from either end (Remy & Hass, 1986a). One ormore of the arms divide again, so that 4-7 armed steles are known. Protoxylems arelocated at either end of the central region near departure of major arms and near the endof each arm (Figure 9C). shape and/or lobing of lateral branch traces differ from themain axis configuration. Morphology is poorly preserved, but the authors suggest lateralbranches departed singly, in pairs or in threes. Much more needs to be learned about thiscomplex lower Devonian plant.
In all of these plant types, protoxylems emanate from one or more of the main axisarms to supply lateral branches and only primary xylem is present. Architecture of theseplants is morphologically and anatomically plastic, although with some common patterns,but evidence is building that these taxa exhibit enough differences to not be closely
GENsEl: SILURIAN-DEVONIAN ORIGINS OF FERNS AND LYCOPHYTES 233

234 FERN GAZ. 20(6):217-242. 2017
figure 9. Diagrammatic drawings of larger basal euphyllophytes, similar in size toPertica (main axes approx. 1 cm) - note that none show evidence of secondary xylem.A. Plant described by Gensel (1984), with large main axis, single or paired lateralbranches, dichotomously divided ultimates arranged along lateral branches. Anatomy athree-lobed stele with central protoxylem and protoxylem at end of each arm. First orderlateral trace (or pair of traces) oval, second order trace circular. (Gensel, in prep) B. APertica-like plant with large main axis, helically arranged predominantly dichotomouslydivided lateral branch, terminating in bunched sporangia (not shown). Anatomy consistsof slightly elongate central region from which four arms emanate, one of these dividesagain to produce five arms total. Protoxylem located at each end of central area, andends of arms. First order traces are elongate to v-shaped or oval and in pairs as shown.Gensel (in prep). C. Gothanophyton zimmermanni Remy and Hass. large main axis withsingle or paired lateral branches (poorly preserved), bearing at least second order ones.Main axis stele consists of an elongate central region from which 4-7 arms emanate.Protoxylem located at each end of central region, along and at ends of arms. lateraltraces change shape after departure to become 4-lobed. Protoxylem in black. Clear areain stele is primary xylem. C. redrawn from Remy and Hass, 1986.

related. They are similar in showing a change in shape between main axis and first-orderlateral branches which conforms more closely to that seen in many ferns or extinct fern-like plants than anything known in progymnosperms, despite apparently comparing moreclosely to the radiate protoxylem group.
Basal regions or rooting structures for Psilophyton and the taller euphyllophytesmentioned above are unknown. The fossils appear to represent aerial shoots extendingfrom a rhizome, but it also is possible that some represent the main part of a non-rhizomatous plant that may have had a basal rooting system. Fairly wide, branched butdetached rooting structures running perpendicular to bedding surfaces occur in sedimentsnear some of the larger taxa in northern New Brunswick, while one long, downwardextending axis was discovered by Elick (1998) in Gaspé which may represent a rootingstructure.
Basal euphyllophytes incertae sedisseveral upper lower Devonian plants (late Emsian) have been excluded fromphylogenetic studies, largely because their anatomy is unknown and branching patternand other features are unique. These include Oocampsa catheta, interpreted as atrimerophyte-progymnosperm intermediate (Andrews et al., 1975), Chaleuria cirrosa(Andrews et al., 1974) and an undescribed plant with similar branching architecture fromnorthern Maine (Gensel, in prep). They all exhibit a robust, fairly long, greater than 1cm wide dominant main axis with densely spiraled lateral branches. In Oocampsa, higherorder branches are several times irregularly dichotomous, bearing clusters of ovoidsporangia. Chaleuria and the new plant from Maine exhibit isotomous second-orderbranches, some terminating in fusiform to ovoid sporangia. Basal regions and thus growthhabit(s) are unknown. One might expect either a multilobed or cladoxyl- like stelarconfiguration based on the major branching patterns. Oocampsa differs from the othertwo in higher order branching and sporangial morphology, while the other two representone or more as yet unrecognized lineages, or may be early members of one of the later-occurring lineages such as cladoxyls or iridopterids.
stenokolealeans are still poorly known, but recent studies indicate a possible affinityto early seed plants (Moment et al., 2016).
SummaRysignificant new data now exist concerning the time of appearance of plants andmorphology and anatomy of some taxa. Exceptionally preserved plant remains haveindicated an earlier advent of plants, plant structures and habit, including transitions toupright, even arboreal types by the Middle Devonian. limited evidence indicates a widerarray of growth strategies among these. Although the euphyllophyte and lycophytelineages separate very early in time, with each demonstrating acquisition of major plantstructures, gaps in preservation preclude establishing evolutionary trends or clearphylogenetic relationships within each lineage, especially early vs later Devonian planttypes, and between Devonian and Carboniferous or younger lineages that appear moredirectly related to extant ferns or lycophytes (similar situation for seed plants). As wholeplant reconstructions are made and new approaches employed, with new discoveries orre-investigation of earlier material, assessment of character homology will be morepossible.
several putative candidate lineages leading to equisetids or polypodiids remain, butit is clear that some, such as cladoxyls, are already highly specialized in development
GENsEl: SILURIAN-DEVONIAN ORIGINS OF FERNS AND LYCOPHYTES 235

and construction by the Middle Devonian. Were more basal taxa similar, as suggestedby Lorophyton Fairon-Demaret et li (1993), Foozia minuta Gerrienne (1992) or perhapsas yet unrecognized taxa? Basal euphyllophytes at the Psilophyton, Pertica, andGothanophyton grade are increasingly interesting in that it is among plants of this typethat early differentiation of anatomical configurations of main axis- lateral branch isestablished. Within this group one might possibly be able to discern early divergence ofthe fern sensu lato vs seed plant lineages. Further examination of anatomy andorganisation of lateral branch architecture, incorporating signals of developmentaldifferences, may aid in recognizing early stages in evolution of leaves- these clearly willbe different in various lineages. similarly, the questions of transitions betweenpredominantly rhizomatous and upright habit, and between types of rooting and oforganisation of reproductive structures in both major lineages may become clearer withfuture discoveries or new approaches.
REfEREncESANDREWs, H.N., GENsEl, P.G., & FORBEs, W.H. 1974. An apparently
heterosporous plant from the Middle Devonian of New Brunswick. Palaeontol. 17:387-408.
ANDREWs, H.N., GENsEl, P.G. & KAsPER, A.E. 1975. A new fossil plant ofprobable intermediate affinities (Trimerophyte-Progymnosperm). Can. J. Bot. 53:11719-1728.
BANKs, H.P., lEClERCQ s., & HUEBER, F.M. 1975. Anatomy and morphology ofPsilophyton dawsoni, sp. n. from the late lower Devonian of Quebec (Gaspé) andOntario, Canada. Paleontographica Americana 8 (48): 77-127.
BATEMAN, R.M., CRANE, P.R., DIMICHElE, W.A., KENRICK, P., ROWE, N.P.,sPECK, T., sTEIN, W.E. 1998. Early Evolution of land plants: Phylogeny,physiology, and ecology of the primary terrestrial radiation. Ann. Rev. Ecol. syst.29: 263-92.
BATEMAN, R. M., KENRICK, P. & ROTHWEll, G.W. 2007. Do eligulate herbaceouslycopsids occur in Carboniferous strata? Hestia eremosa gen. et sp. nov. from theMississippian of Oxroad Bay, East lothian, scotland. Rev. Palaeobot. Palynol. 144(3): 323-335.
BECK, C.B. & sTEIN, W.E. 1993. Crossia virginiana gen. et sp. nov., a new memberof the stenokoleales from the Middle Devonian of southwestern Virginia.Palaeontographica B229: 115-134.
BERRY, C.M. 2005. ‘Hyenia’ vogtii Hoeg from the Middle Devonian of spitsbergen-its morphology and systematic position. Rev. Palaeobot. Palynol. 135(1-2), pp. 109-116.
BERRY, C. M. & FAIRON-DEMARET, M. 1997. Reinvestigation of the cladoxylopsidPseudosporochnus nodosus leclercq et Banks from the Middle Devonian of Goe,Belgium. Int. J. Plant sci. 158(3), pp. 350-372
BERRY, C. M. & sTEIN, W.E. 2000. A new iridopteridalean from the Devonian ofVenezuela. Int. J. Plant sci. 161(5), pp. 807-827.
BERRY, C. M. & FAIRON-DEMARET, M. 2002. The architecture of Pseudosporochnusnodosus leclercq et Banks: a Middle Devonian cladoxylopsid from Belgium. Int. J.Plant sci. 163(5), pp. 699-713.
BERRY, C. M. & WANG, Y. 2006. Eocladoxylon (Protopteridium) minutum (Halle)Koidzumi from the Middle Devonian of Yunnan, China: an early Rhacophyton-like
236 FERN GAZ. 20(6):217-242. 2017

plant? Int. J. Plant sci. 167(3), pp. 551-566. BERRY, C. M. & MARsHAll, J.E.A. 2015. lycopsid forests in the early late Devonian
paleoequatorial zone of svalbard. Geology 43(12): 1043-1046.BERRY, C. M., WANG, Y. & CHONGYANG, C. 2003. A lycopsid with novel
reproductive structures from the Upper Devonian of Jiangsu, China Int. J. Plantsci. 164(2), pp. 263-273.
BOYCE, C.K. & KNOll, A.H. 2002. Evolution of developmental potential and themultiple independent origins of leaves in Paleozoic vascular plants. Paleobiology28(1): 70-100.
BOYER, J. s. & MATTEN, l.C. 1996. Anatomy of Eospermatopteris eriana from theUpper Middle Devonian (Givetian) of New York. Int. Org. Palaeobot. 5, 13 (1996).
CAI C-Y & CHEN, l-Z. 1996. On a Chinese Givetian lycopod, Longostachyslatisporophyllus Zhu, Hu and Feng, emend.: its morphology, anatomy andreconstruction. Palaeontogr. Abt B. 1996; 238: 1–43.
CHAlONER, W. G. 1968. The cone of Cyclostigma kiltorkense Haughton, from theUpper Devonian or Ireland. J. linn. soc. london (Botany) 61: 25-36.
CHITAlY, s. & PIGG, K.B. 1996. Clevelandodendron ohioensis, Gen. et sp. Nov., Aslender upright lycopsid from the late Devonian Cleveland shale of Ohio. Am. J.Bot. 83(6): 781-789.
CORNET, B., PHIllIPs, T.l. & ANDREWs, H.N.1976. The morphology and variationin Rhacophyton ceratangium from the Upper Devonian and its bearing on frondevolution. Palaeotographica B 158: 105-129.
CORVEZ, A., BARIEl, B. & DUBUIsssON, J-Y. 2012. Diversity and evolution of themegaphyll in Euphyllophytes: phylogenetic hypotheses and the problem of foliarorgan definition. Comptes Rend. Palevol. 11: 403-418.
CRANE, P.R. AND KENRICK, P. 1997. Diverted development of reproductive organs:a source of morphological innovation in land plants. Plant syst. & Evol.206 (1997): 161-174.
DITTRICH, H.s., MATTEN, l.C. & PHIllIPs, T.l. 1983. Anatomy of Rhacophytonceratangium from the Upper Devonian (Famennian) of West Virginia. Rev. Palaeobot.Palynol. 40: 127-147.
DORAN, J.B. 1980. A new species of Psilophyton from the lower Devonian of northernNew Brunswick, Canada. Can. J. Bot. 58: 2241-2262.
EDWARDs, D. & FEEHAN, J. 1980. Records of Cooksonia-type sporangia from thelate Wenlock strata of Ireland. Nature 287: 41-42.
EDWARDs, D. & KENRICK, P. 2015. The early evolution of land plants, from fossilsto genomics: a commentary on lang (1937) ‘On plant remains from the Downtonianof England and Wales’. Phil. Trans. R. soc. B 370: 20140343.
EDWARDs, D., MORRIs, J. l., RICHARDsON, J.B. & KENRICK, P. 2014.Cryptospores and cryptophytes reveal hidden diversity in early land floras. NewPhytol. 202: 50-78.
ElICK, J. M., DRIEsE, s.G. & MORA, C.I. 1998. Very large plant and root traces fromEarly to Middle Devonian: Implications for early terrestrial ecosystems andatmospheric p (CO2) estimations. Geology 26: 143-146.
FAIRON-DEMARET, M. & lI, C-s. 1993. Lorophyton goense gen. et sp. nov. from thelower Givetian of Belgium and a discussion of the Middle Devonian Cladoxylopsida.Rev. Palaeobot. Palynol. 77(1-2): 1-22.
FAIRON-DEMARET, M. & BERRY, C.M. 2000. A reconsideration of Hyenia elegans
GENsEl: SILURIAN-DEVONIAN ORIGINS OF FERNS AND LYCOPHYTES 237

Krausel et Weyland and Hyenia ‘complexa’ leclercq: Two Middle DevonianCladoxylopsida from western Europe. Int. J. Pl. sci. 161(3): 473-494.
FU, Q., WANG, Y., BERRY, C.M. & XU, H-H.. 2011. Complex branching patterns in anewly recognized species of Compsocradus Berry et stein (Iridopteridales) from theMiddle Devonian of North Xinjiang, China. Int. J. Pl. sci. 172(5): 707-724.
GAlTIER, J. 2010. The origins and early evolution of the megaphyllous leaf. Int. J. Plantsci. 171(6):641-661.
GENsEl, P.G. & ANDREWs, H.N. 1984. Plant life in the Devonian. New York: PraegerPress, 379 pp.
GENsEl, P.G. & KAsPER, A.E. 2005. A new species of the Devonian lycopod genus,Leclercqia, from New Brunswick, Canada. Rev. Palaeobot. Palynol. 136, 105–123.
GENsEl, P.G. & AlBRIGHT, V.M. 2006. Leclercqia complexa from the EarlyDevonian (Emsian) of northern New Brunswick, Canada. Rev. Palaeobot. Palynol.142: 103-121.
GENsEl, P.G. & BERRY, C.M. 2001. Early lycophyte evolution. Am. Fern J. 91: 74-98.GENsEl P.G., KOTYK, M.E. & BAsINGER, J.F. 2001. Morphology of above- and
below-ground structures in Early Devonian (Pragian-Emsian) plants In GENsEl P.G.& D. EDWARDs, eds, Plants Invade the land: Evolutionary and EnvironmentalPerspectives. Columbia University Press, New York, pp. 83–102.
GERRIENNE, P. 1992. The Emsian plants from Fooz-Wépion (Belgium). III. Fooziaminuta gen. et spec. nov., a new taxon with probable cladoxylalean affinities. Rev.Palaeobot. Palynol. 74: 139-157.
GERRIENNE, P.M. & GENsEl, P.G. 2016. New data about anatomy, branching, andinferred growth patterns in the Early Devonian plant Armoricaphytonchateaupannense, Montjean-sur-loire, France. Rev. Palaeobot. Palynol. 224: 38-53.
GERRIENNE, P., MEYER-BERTHAUD, B., FAIRON-DEMARET, M., sTREEl, M.& sTEEMANs, P. 2004. Runcaria, a Middle Devonian seed plant precursor. science306: 856-858.
GERRIENNE, P., MEYER-BERTHAUD, B., YANG, N., sTEEMANs, P., & li, C-s.2014. Planatophyton gen. nov., a late Early or Middle Devonian euphyllophyte fromXinjiang, North-west China. Rev. Palaeobot. Palynol. 208:55-64.
GERRIENNE, P., GENsEl, P.G., sTRUllU-DERRIEN, C. lARDEUX, H.,sTEEMANs, P., & PREsTIANNI, C. 2011. A simple type of wood in two EarlyDevonian plants. 2011. science 333(6044):837.
GIEsEN, P. & BERRY, C.M. 2013. Reconstruction and growth of the early treeCalamophyton (Pseudosporochnales, Cladoxylopsida) based on exceptionallycomplete specimens from lindlar, Germany (Mid-Devonian): Organic connection ofCalamophyton branches and Duisbergia trunks. Int. J. Plant sci. 174 (4): 665-686.
GRIERsON, J.D. & BONAMO, P.M. 1979. Leclercqia complexa: Earliest ligulatelycopod (Middle Devonian). Am. J. Bot. 474-476.
HAO, s-G. 1988. A new lower Devonian genus from Yunnan, with notes on the originof leaves (in Chinese). Acta Bot. sin. 30: 441-448.
HAO, s G. & BECK, C. B. 1993. Further observations on Eophyllophyton bellum fromthe lower Devonian (siegenian of Yunnan, China. Palaeontogr. B 230: 27-41
HAO, s-G, & XUE, J. 2013. The Early Devonian Posongchong flora of Yunnan- acontribution to an understanding of the evolution and early diversification of vascularplants. science Press, 366 pp.
HAO s, XUE, J., GUO, D, & WANG, D-M.. (2010) Earliest rooting system and
238 FERN GAZ. 20(6):217-242. 2017

root:shoot ratio from a new Zosterophyllum plant. New Phytol 185: 217–225HAO, s., WANG, D., WANG, Q. & XUE, J. 2006. A New lycopsid, Zhenglia
radiata gen. et sp. nov., from the lower Devonian Posongchong Formation ofsoutheastern Yunnan, China, and Its Evolutionary significance. Acta Geol. sin. 80(1):11-19.
HAUGHTON, s. 1859. On Cyclostigma, a new genus of fossil plants from the Old Redsandstone of Kiltorcan Co Kilkenny. J. Roy. soc. Dublin 2:407-420.
HETHERINGTON, A.J. & DOlAN, l. 2017. The evolution of lycopsid rootingstructures: conservatism and disparity. New Phytologist 215 (2): 538-544.
HIll, s.A., sCHECKlER, s.E. & BAsINGER, J.F. 1997. Ellesmeris sphenopteroides,gen. et sp. nov., a new zygopterid fern from the Upper Devonian (Frasnian) ofEllesmere, N.W.T., Arctic Canada. Am. J. Bot. 84: 85-93.
HOFFMAN, l.A. & TOMEsCU, A.M. F. 2013. An early origin of secondary growth:Franhueberia gerriennei gen. et sp. nov. from the lower Devonian of Gaspé(Quebec, Canada). Amer. J. Bot. 100, 754-763.
HUEBER, F.M. 1983. A new species of Baragwanathia from the sextant Formation(Emsian) Northern Ontario, Canada. Bot. J. linn. soc. 86:57-79.
HUEBER, F.M. 1992. Thoughts on the early lycopsids and zosterophylls. Ann. Mo. Bot.Gdn. 79:474-499.
JOHNsON, T. 1913. On Bothrodendron (Cyclostigma) kiltorkense Haughton, sp. Roy.soc. Dublin, sci. Proceed. 13 (n.s.): 500-528.
KAsPER, A.E, ANDREWs, H.N. & FORBEs, W.H. 1974. New fertile species ofPsilophyton from the Devonian of Maine. Am. J. Bot. 61: 339-359.
KENRICK P. 2013. The origin of roots. In: Eshel A, Beeckamn T, eds. Plant roots: thehidden half. london, UK: Taylor & Francis, 1–14.
KENRICK P. & CRANE, P. R. 1997. The origin and early diversification of land plants:A cladistic study. Washington: smithsonian.
KENRICK P. & sTRUllU-DERRIEN, C. 2014. The Origin and early evolution of roots.Plant Phys. 166: 570-580.
KENRICK, P., WEllMAN, C.H., sCHNEIDER, H. & EDGECOMBE, G.D. 2012. Atimeline for terrestrialization: consequences for the carbon cycle in the Palaeozoic.Phil. Trans. Roy. soc. B 367: 519-536.
KOTYK, M. E., BAsINGER, J.F., GENsEl, P. G. & DE FREITAs, T. A. 2004.Morphologically complex plant macrofossils from the late silurian of Arctic Canada.Am. J. Bot. 89: 1004-1013.
KRÄUsEl, R. & WEYlAND, H. 1937. Pflanzenreste aus dem Devon. X. ZweiPflanzenfunde im Oberdevon der Eifel. senckenbergiana 19: 338-355.
KRÄUsEl, R. & WEYlAND, H. 1949. Gilboaphyton und die Protolepidophytales.senckenbergiana 30: 129-152.
lEClERCQ, s. 1951. Étude morphologique et anatomique d’une Fougère du Dévoniensuperieur. Ann. soc. Geol. Belg..Mem. Vol. XIV (3): 39 pp.
lEClERCQ, s. 1970. Classe des Cladoxylopsida Pichi-sermolli, 1959. In: Boureau, E>(Ed.). Traité de Paléobotanique. IV. Fasc. I. Filicophyta. Masson, Paris, pp. 119-165.
lI, F-W., VIllARREAl, J.C., KEllY, s., ROTHFEls, C.J., MElKONIAN, M.,FRANGEDAKIs, E., RUHsAM, M., sIGEl, E.M., DER, J.P., PITTERMANN, J.,BURGE, D.O., POKORNY, l., lARssON, A., CHEN, T., WEsTsTRAND, s.,THOMAs, P., CARPENTER, E., ZHANG, Y., TIAN, Z., CHEN, l., YAN, Z., ZHU,Y., sUN, X., WANG, J., sTEVENsON, D.W., CRANDAll-sTOTlER, B.J.,
GENsEl: SILURIAN-DEVONIAN ORIGINS OF FERNS AND LYCOPHYTES 239

sHAW, A. J., DEYHOlOs, M.K. sOlTIs, D.E. ,GRAHAM, s.W., WINDHAM,M.D., lANGDAlE, J.A., WONG, G.K-s. MATHEWs, s. & PRYER, K.M. 2014.Horizontal transfer of an adaptive chimeric photoreceptor from bryophytes to ferns.PNAs, 111:6672-6677
MATsUNAGA, K.K.s. & TOMEsCU, A.M.F. 2016. Root evolution at the base of thelycophyte clade: insights from an Early Devonian lycophyte. Ann. Bot117(4):585-598.
MOMENT, N., GERRIENNE, P. & PREsTIANNI, C. 2015. Brabantophyton, a newgenus of stenokolealean affinities from the Middle to earliest Upper Devonian localityfrom Belgium. Rev. Palaeobot. Palynol. 227: 77-96.
MUsTAFA, H. 1978. Beiträge zur Devonflora III. Argum. Palaeobot. 5: 91-132.NICKRENT, D. l., PARKINsON, C. l., PAlMER, J. D. & DUFF, R. J. 2000. Multigene
phylogeny of land plants with special reference to bryophytes and the earliest landplants. Mol. Biol. Evol. 17:1885–1895.
OBRHEl, J. 1959. Neue pflanzenfunde in den sbrsko-schichten (Mitteldevon). Vestn.Ustred. Ustavu Geol. 34: 384-388.
PPG1. 2016. A community-derived classification for extant lycophytes and ferns. J. syst.Evol. 54 (6): 563–603.
PREsTIANNI, C. & GERRIENNE, P.M. 2010. Early seed plant radiation: an ecologicalhypothesis pp. 71-80 in Vecoli, M., Clement, G. & Meyer-Berthaud, B. (eds). TheTerrestrialization Process: Modelling Complex Interactions at theBiosphere-Geosphere Interface. Geol. soc. lond. spec. Pub., 339.
PRYER, K.M., sMITH, A.R., & ROTHFEls, C. 2009. Polypodiopsida (14 January2009) [online]. In: Tree of life Web Project: http://tolweb.org/Polypodiopsida/20615/2009.01.14 [accessed 7 October 2016].
PRYER K.M., sCHNEIDER H., sMITH A.R., CRANFIll R., WOlF P.G., HUNT J.s.2001. Horsetails and ferns are a monophyletic group and the closest living relativesto seed plants. Nature 409: 618–621.
PRYER, K.M., sCHUETTPElZ, E., WOlF, P.G., sCHNEIDER H., sMITH A.R., &CRANFIll, R. 2004. Phylogeny and evolution of ferns (monilophytes) with a focuson the early leptosporangiate divergences. Amer. J. Bot. 91: 1582–1598
QIU Y-l, lI, l., WANG, B., CHEN, Z., DOMBROVsKA, O., lEE, J., KENT, l., lI,R., JOBsON, R.W., HENDRY, T.A., TAYlOR, D.W. 2007. A nonflowering landplant phylogeny inferred from nucleotide sequences of seven chloroplast,mitochondrial, and nuclear genes. Int. J. Plant sci. 168: 691–708.
RAVEN J.A. & EDWARDs, D. (2001) Roots: evolutionary origins and biogeochemicalsignificance. J. Exp. Bot. 52: 381–401
REMY, W. & HAss, H. 1986. Gothanophyton zimmermanni nov.gen., nov. spec., einePflanze mit komplexem stelar-Korper aus dem Emsian. Arg. Palaeobotanica 7 (9):9-69.
RENZAGlIA, K. s., DUFF, R. J. T, NICKRENT, D. l. & GARBARY, D. J. 2000.Vegetative and reproductive innovations of early land plants: implications for aunified phylogeny. Phil. Trans. R. soc. lond. B 355:769–793
RICHARDsON, J.B., BONAMO, P. M. & McGREGOR, D.C. 1993. The spores ofLeclercqia and the dispersed spore morphon Acinosporites lindlarensis Riegel: a caseof gradualistic evolution. Bull. Nat. Hist. Mus., lond. (Geol.) 49, 121–155.
RICKARDs, R. B. 2000. The age of the earliest club mosses: the silurian Baragwanathiaflora in Victoria, Australia. Geol. Mag. 137(2): 207-209.
240 FERN GAZ. 20(6):217-242. 2017

ROTHWEll, G.W. & NIXON, K. C. 2006. How does inclusion of fossil data changeour conclusions about the phylogenetic history of euphyllophytes? Int. J. Plant sci.167(3): 737-749.
ROTHWEll, G. W. & sTOCKEY, R. A. 2008. Phylogeny and evolution of ferns: apaleontological perspective. Pp. 332-364, Ranker, T.A. & Haufler, C. H. (eds)Biology and Evolution of Ferns and lycophytes. Cambridge Uiversity Press.
ROWE, N. 1988. A herbaceous lycophyte from the lower Carboniferous Drybrooksandstone of the Forest of Dean, Gloucestershire. Palaeontol. 31 (1): 69-83.
RUBINsTEIN, C.V.; GERRIENNE, P. DE lA PUENTE, G.s., AsTINI, R.A. &sTEEMANs, P. (2010). Early Middle Ordovician evidence for land plants inArgentina (eastern Gondwana). New Phytologist. 188 (2): 365–369
sANDERs, H., ROTHWEll, G.W. & WYATT, s. 2007. Paleontological context forthe developmental mechanisms of evolution. Int. J. Plant sci. 168: 719-728.
sCHECKlER, s.E., 1974. systematic characters of Devonian ferns. An. Mo. Bot. Gard.61:462-473.
sCHECKlER, s.E., sKOG, J.E. & BANKs, H.P.. 2006 Langoxylon asterochlaenoideumstockmans: Anatomy and relationships of a fern-like plant from the Middle Devonianof Belgium. Rev. Palaeobot Palynol. 142: 193-217.
sCHNEIDER H, sMITH, A. R. & PRYER, K. M. 2009. Is morphology really at oddswith molecules in estimating fern phylogeny? sys. Bot. 34: 455–475.
sCHUlTKA, s. 1978. Beiträge zur Anatomie von Rhacophyton condrusorum Crepin.Arg. Palaeobot. 5: 11-22.
sCHWEITZER H-J & lI, C-s. 1996 Chamaeodendron nov. gen., eine multisporangiatelycopsid aus dem Frasnium sudchinas. Palaeontogr B 238:45–69.
sKOG, J. & HIll, C. 1992. The Mesozoic herbaceous lycopsids. Ann. Mo. Bot. Gard.79: 648-675.
sMITH A.R., PRYER, K.M., sCHUETTPElZ, E. KORAll, P., sCHNEIDER, H.& WOlF. P.G. 2006b. A classification for extant ferns. Taxon 55: 705–731.
sTEEMANs, P., WEllMAN, C.H. & GERRIENNE, P. 2010. "Paleogeographic andpaleoclimatic considerations based on Ordovician to lochkovianvegetation". Geological Society, London, Special Publications. 339 (1): 49–58. doi:10.1144/sP339.5
sTEEMANs, P., lE HÉRIssÉ, A., MElVIN, J., MIllER, M.A., PARIs, F.,VERNIERs, J., WEllMAN, C.H.. 2009. Origin and radiation of the earliest vascularplants. science 324: 353.
sTEIN, W. 1982. Iridopteris eriensis from the Middle Devonian of North America, withsystematics of apparently related taxa. Bot. Gaz. 143(3): 401-416.
sTEIN, W. & HUEBER, F. M. 1989. The anatomy of Pseudosporochnus: P. hueberifrom the Devonian of New York. Rev. Palaeobot. Palynol. 60: 311-359.
sTEIN, W., Wight, D. C. & Beck, C. B. 1984. Possible alternatives for the origin ofsphenopsida. syst. Bot. 9: 102-118.
sTEIN, W., MANNOlINI, F., HERNICK, l V-A., lANDING, E., & BERRY, C.M..2007. Giant cladoxylopsid trees resolve the enigma of the Earth’s earliest foreststumps at Gilboa. Nature 446: 904-907.
sTEIN, W., BERRY, C.M., HERNICK, l V-A. & MANNOlINI, F. 2012. surprisinglycomplex community discovered in the mid-Devonian fossil forest at Gilboa.Nature 483(7387), pp. 78-81.
sTRUllU-DERRIEN, C., KENRICK, P., TAFFOREAU, P., COCHARD, H., JEAN-
GENsEl: SILURIAN-DEVONIAN ORIGINS OF FERNS AND LYCOPHYTES 241

lOUIs BONNEMAIN, J-l, lE HÉRIssÉ, A., lARDEUX, H. & BADEl, E. 2014.The earliest wood and its hydraulic properties documented in c. 407-million-year-old fossils using synchrotron microtomography. Bot. J. linn. soc. 175: 423-437.
THOMAs, B. 1992. Paleozoic herbaceous lycopsids and the beginnings of extantLycopodium sens. lat. and Selaginella sens. lat. Ann. Mo. Bot. Gdn 79: 623-631.
TOMEsCU, A.M.F. 2009. Megaphylls, microphylls and the evolution of leafdevelopment. Trends Plant sci.14:5-12.
TRANT, C. & GENsEl, P.G. 1985. Branching in Psilophyton: a new species from thelower Devonian of New Brunswick, Canada. Am. J. Bot. 72(8): 1256-1273.
WANG, DM, XU, H-H., XUE, J-Z., WANG,Q., lIU, l. 2015. leaf evolution in early-diverging ferns: insights from a new fern-like plant from the late Devonian of China.Ann. Bot. 115: 1133-1148.
WANG, QI, HAO, s-G., WANG, D-M., WANG, Y. & DENK, T. 2003. A late Devonianarborescent lycopsid Sublepidodendron songziense Chen emend.(sublepidodendraceae Krausel et Weyland 1949) from China, with a revision of thegenus Sublepidodendron (Nathorst) Hirmer 1927. Rev. Palaeobot. Palynol. 127: 269-305.
WANG, Y. & BERRY, C.M. 2001. A new plant from the Xichong Formation (MiddleDevonian) south China. Rev. Palaeobot. Palynol. 116:63-85.
WANG, Y. & BERRY, C.M. 2003. A novel lycopsid from the Upper Devonian of Jiangsu,China. Palaeontology 46(6): 1297-1311.
WEllMAN, C.H., sTEEMANs, P., MIllER, M.A. 2008. Trilete spores from theOrdovician of saudi Arabia: earliest evidence for vascular plants and their immediatepredecessors (“protracheophytes”). Terra Nostra 2, 304.
WEllMAN, C.H., sTEEMANs, P., MIllER, M.A. 2015. spore assemblages fromUpper Ordovician and lowermost silurian sediments recovered from the Qusaiba-1shallow core hole, Qasim region, central saudi Arabia. Rev. Palaeobot. Palynol. 212:111-126.
XU, H.H. & WANG, Y. 2016. The earliest cormose rhizomorph of putative lycopsidaffinity from the Middle Devonian of West Junggar, Xinjiang, China. Rev. Palaeobot.Palynol. 226: 54-57.
XU, H-H, BERRY, C.M., WANG, Y., MARsHAll, J.E.A. 2011. A new species ofLeclercqia Banks, Bonamo et Grierson (lycopsida) from the Middle Devonian ofNorth Xinjiang, China, with a possible climbing habit. Int. J. Plant sci. 172 (6): 836-846.
XUE, J.Z., HAO, s-G, ZHU, X., WANG, D-M. 2012. A new basal euphyllophyte,Pauthecophyton gen. nov., from the lower Devonian (Pragian) of Yunnan, China.Rev. Palaeobot. Palynol. 183(1): 9-20.
ZDEBsKA, D. 1986. Psilophyton szaferi sp. nov., from the lower Devonian of the Holycross Mountains, Poland. Acta soc. Bot. Poland 55: 315-324.
242 FERN GAZ. 20(6):217-242. 2017

a ShoRt BIogRaphy of thE authoRPatricia G. Gensel is a professor in the Biology Department, University of North Carolinaat Chapel HiIl, having started there in 1975. she received her B.A. at Hope College,Holland, Michigan, then she worked as a research assistant in the Geology Department,King's College, Univ. of london, England, for Dr. John Richardson in the area ofDevonian palynology. Patricia returned to the U.s.A. and embarked on graduate studiesof fossil plants and their spores with Professor Henry Andrews at the University ofConnecticut, storrs, CT., where she earned a M.s. and then Ph.D. After a few years ofpost-doctoral work, again with H. N. Andrews, she joined the faculty of (initially) theBotany Department, and later, Biology Department, at UNC.
Patricia continued work on both Devonian (from U.s. and Canada) and EarlyCarboniferous plants (from Us), studying both megafossils and where possible, in situspores. she also had students who worked on Cretaceous fossils from North Carolinaand prepared a field guide to Triassic Plants of the Deep River Basin, NC. she co-authored the book Plant Life in the Devonian with H.N. Andrews (1984), and co-editeda book of contributed chapters entitled Plants Invade the Land with Dianne Edwards(2001).
FERN GAZ. 20(6). 2017 243

she has authored or co-authored at least 68 peer-reviewed publications, the majorityon Devonian plants, but also some about in situ spores, spore ultrastructure, and EarlyCarboniferous plants, several book chapters, reviews or field guides. Patricia hasorganized many symposia, mostly about early land plant evolution, trained several Msstudents and a few PhD students. Her major interests center on producing a betterunderstanding of the morphology, structure, evolutionary relationships or patterns ofevolutionary change, and where possible ecology, of early land plants, mainly ofDevonian and Early Carboniferous age. she continues her interest in palynology,especially in situ spores or spore ultrastructure, in terms of what they may contributerelative to systematics or phylogenies of early plants. Recent publications include areport of early wood from the very Early to Early Devonian of France and NewBrunswick (Gerrienne et al., 2011, Gerrienne & Gensel, 2016, work in prep), a revisionof sporangial morphology and attachment in the iconic zosterophyll Sawdonia ornata(Gensel & Berry, 2016) and paleoenvironmental interpretations of some of the majorplant-bearing deposits in New Brunswick or early coal (Kennedy et al., 2012; Kennedyet al., 2013). Her on-going work focuses on elucidating anatomy and morphology ofbasal euphyllophytes (formerly trimerophytes), additional work on Sawdonia spp., andother Devonian or Carboniferous plants.
REFERENCEsGENsEl, P. & BERRY, C.M. 2016. sporangial Morphology of the Early Devonian
Zosterophyll Sawdonia ornata from the type locality (Gaspé). Int. J. Pl. sci. 177:618-632.
GERRIENNE, P.M. & GENsEl, P.G. 2016. New data about anatomy, branching, andinferred growth patterns in the Early Devonian plant Armoricaphytonchateaupannense, Montjean-sur-loire, France. Rev. Palaeobot. Palynol. 224: 38-53.
GERRIENNE, P., GENsEl, P.G., sTRUllU-DERRIEN, C. lARDEUX, H.,sTEEMANs, P., & PREsTIANNI, C. 2011. A simple type of wood in two EarlyDevonian plants. science 333(6044):837.
KENNEDY, K.l, GENsEl, P.G. & GIBlING, M.R., 2012. Paleoenvironmentalinferences from the classic Early Devonian plant-bearing locality of the CampbelltonFormation, New Brunswick, Canada. Palaios 27: 424–438.
KENNEDY, K.l., GIBlING, M.R, EBlE, C.F., GAsTAlDO, R.A., GENsEl, P.G.,WERNER-ZWANZIGER, U. & WIlsON, R.A. 2013. lower Devonian coaly shalesof northern New Brunswick, Canada: plant accumulations in the early stages ofterrestrial colonization. J. sediment. Res. 83:1202–1215
244 FERN GAZ. 20(6). 2017

a nEW REcoRD of BRoWn Rot DISEaSE In WatER fERn AZOLLAMICROPHYLLA (aZollacEaE): loSS of ImpoRtant BIo-RESouRcE
s. DEY, M. HORE, J. BIsWAs, M. BIsWAs, B.K. MANDAl, P. DAs & s. GUPTA
Pteridology and Palaeobotany section, Department of Botany, University of Kalyani, Kalyani- 741235, West Bengal, India
e-mail id: [email protected]
Key words: cottony white mycelium; yellow pigmentation; macroconidia;microconidia; chlamydospores; pathogenic fungus; Fusarium thapsinum
aBStRactseverely infected water fern, Azolla microphylla Kaulf., has been observed innatural habitats in different districts in the lower Gangetic plain of West Bengal,India. The infected plants have turned dark brown and been rotted entirely within5-7 days, causing loss of natural resource as the plant species has immensecommercial value. specific growing colonies with cottony white mycelium,yellow pigmentation on the cultured medium, macro- and micro- conidialfeatures, and absence of chlamydospore indicate the causal pathogen to beFusarium thapsinum Klittich., lesile, Nelson, Marases. The identification of thecausal pathogen has been authenticated by standard pathogenicity test (Koch’sPostulates). Extensive literature survey reveals that this is a new record of brownrot disease in A. microphylla.
IntRoDuctIonAzolla lam. is a free-floating, fast growing water fern. The plant is widely distributed intropical, subtropical, and warm temperate zones in different countries of the Worldincluding India (Wagner, 1997). Here Azolla spp. are naturally available, mostly on moistsoils, freshwater ditches, ponds, lakes, sluggish rivers and marshy lands. Azolla is treatedas a ‘green gold mine’, because of its high nutritive value and it is considered as ‘superplant’ due to its fast-growing capacity (Wagner, 1997). They can even grow in nitrogendeficit areas and can double the biomass in 3-5 days because of its unique symbioticrelationship with the nitrogen fixing endophytic blue-green cyanobacterium, Anabaenaazollae sterberger (Wagner, 1997).
High nutritive values (Buckingham et al., 1978; Alcantara & Querubin, 1985; Paolettiet al., 1987; lejeune et al., 2000; Alalade & lyayi, 2006) and huge productivity througheasy cultivation methods (Dao & Tran, 1979; lumpkin & Plucknett, 1982; liu et al.,2008) are the key factors for utilizing Azolla as one of the most promising aquatic plantsfor livestock feed (Becerra et al., 1995; Alalade & lyayi, 2006; Cagauan & Pullin, 1991;shiomi & Kitoh, 2001; Fiogbe et al., 2004), and as an effective natural fertilizer toincrease rice yield successfully (Peters, 1978; Tung & shen, 1985; Watanabe & liu,1992; Yadav et al., 2014). The plant has been employed in experiments during space andplanetary travel (liu et al., 2008; Carrapico, 2002; Katayama et al., 2008), and used forthe production of non-polluting high energy biofuel (Peters et al., 1976; Newton, 1976;Das et al., 1994; Hall et al., 1995), especially biodiesel (salehzadeh et al., 2014). Theplant can act as reducer of various greenhouse gases by ammonia volatilization
FERN GAZ. 20(6):245-254. 2017 245

(Watanabe & liu, 1992), by inhibiting methane production from rice field (Prasanna etal., 2002), and noticeably by sequestering atmospheric CO2 (Brinkhuis et al., 2006). Inaddition, weed control (Krock et al., 1991), mosquito repellence (Ansari & sharma,1991), phyto-remediation from waste water (Jain et al., 1989; saxena, 1995; Costa etal., 1999; Antunes et al., 2001; Khosravi et al., 2005; Umali et al., 2006; Rai, 2008; Costaet al., 2009; Rai & Tripathi, 2009; Elmachliy et al., 2010; Rai, 2010a, b; sood et al.,2012) and soil (Umali et al., 2006; Cohen et al., 2002 and Mashkani & Ghazvini, 2009)are added approaches for utilizing this vital bio-resource.
With view to the many valuable uses of Azolla, the present authors have attemptedto explore the species under various abiotic stresses through experimental set-up byculture of collected specimens from natural habitats of the lower Gangetic plains of WestBengal, India. During field survey, it has been observed that among the available Azollaspecies, A. microphylla Kaulf. grows dominantly over others and it can resist directsunlight in summer. However, the species has been found to be severely infected by afungal pathogen at the end of winter season (January-March) in most of the collectionsites, which disrupts normal growth of the plant and ultimately causes death within 5-7days.
From the available literature, it seems that pathogenic reports on Azolla spp. are rathermeagre (sasi et al., 1982; Kannaiyan, 1985; Kannaiyan & Kumar, 2005; lee et al., 2011;Banihashemi, 2014). The present communication describes a new causal organism forA. microphylla, which may help in identifying suitable measure(s) to protect thisimportant natural resource.
matERIal anD mEthoDSFresh and infected samples of Azolla microphylla Kaulf. were collected from differentsites of three districts, namely, North 24 Parganas (22.936 N, 88.392 E; 22.825 N, 88.786E), south 24 Parganas (22.021 N, 88.615 E), and Nadia (22.983 N, 88.445 E) in the lowerGangetic plain of West Bengal.
The morphological studies of infected and non-infected plants were made by handlens and stereo zoom microscope (model lEICA s8APO). The anatomical details ofinfected plants and that of the isolated pathogen were thoroughly studied under lightmicroscope (model leitz laborlux s). Macro and micro photographic documentationswere made using Nikon D3200 DslR and EC3 scientific cameras. To identify thepathogen from morphological characteristics, first the isolations were made from thefrond of the naturally infected A. microphylla plant using a sterile inoculating loop andplated onto 2.25% Potato Dextrose Agar (PDA). A total of five replicas of culture andthree replicas of control plates were incubated at 25˚C temperature in complete darkness.After 2–3 days of inoculation, the fungal colony was observed to grow in each of thefive replications of cultured plates but no changes were observed in control plates. Thecolony morphology, growth pattern, size and shape of macro- and micro- conidia werecharacterized thoroughly.
To confirm the identity of the pathogen, the standard pathogenicity test (Koch’sPostulates) was performed stepwise. student’s t-test was done to compare the variationsin two sets of data of pathogens detected from inoculated and re-inoculated pure culturedplates prepared through the pathogenicity test. The tests were performed by inoculatingnaturally-collected, healthy A. microphylla plants of a single genotype, because thedisease was observed in A. microphylla only in natural habitats. A total of 45 experimentalplants were surface sterilized through rinsing in 70% ethanol for 30 sec. followed by 1%
246 FERN GAZ. 20(6):245-254. 2017

sodium hypochlorite solution for 1 min., and sterile distilled water for three times. Outof 45 surface sterilized plants, 30 healthy plants were wounded on the fronds by sterileneedle. A small mycelial plug taken from a month-old colony of the pathogen wasinserted into the wounds using a sterile toothpick. Both 30 inoculated and 15non-inoculated plants were then placed onto nutrient medium, distributing five plants ineach plate so that six replicas were made for inoculated plants and three fornon-inoculated ones. The nutrient media was prepared using HIMEDIA PTO10Murashige and skoog medium with 17.04 gm./lit. and 10 gm. of agar. The plates werekept in a growth chamber at 25˚±2˚C temperature and 1800 lux intensity of light in 16:8hr day and night cycle.
RESultS anD DIScuSSIonsevere infection was observed in A. microphylla during the months of January to Marchin the field (Figure. 1A, B), as well as in a tank placed in the experimental garden of theBotany Department, Kalyani University (Figure 1C). In a fresh individual plant (Figure1D), at first the infection was observed in roots (Figure 1E), followed by darkbrown-coloured spots on the leaf (Figure 1F). later, the infected areas sequentiallyenlarged (Figure 1G), and eventually the whole leaf turned into brown, showing blochnotch symptoms (Figure 1H). Finally, the plant rotted within 5-7 days (Figure 1I).Outwardly protruding, cottony, white mycelial growth covering the entire leaf surfacewas observed with disease severity (Figure 1J). The transverse section of infected leavesat the initial stage showed loosening of leaf tissues with distinct entrance of hyphae(Figure 1K). Anatomical section of leaf was not possible at the final stage of infectiondue to total disintegration of leaf tissues. The browning and rotting of leaves and rootsare characteristic symptoms produced by the pathogen in A. microphylla. The dark browncoloration of the infected parts of the plant is because of the degradation ofchlorophyllous pigments and probably production of phenolic compounds as resistanceto pathogens (lattanzio et al., 2006). By comparing the symptoms of rot diseases (Agrios,2005), the studied infection in present investigation has been designated as brown rotdisease of A. microphylla.
Each replica of cultured PDA media produced fungal colonies with very thick,abundant, cottony white mycelium, finally attaining a diameter of 48.71±3.0 mm (Figure1l) after seven days of incubation. The growth pattern of the mycelium was radial. Acharacteristic yellow pigmentation was found to appear on the agar plate (Figure 1l)after 3-4 days of incubation. After 5-6 days, a slight purplish pigmentation was observedto ooze out and persisted for 1-2 days only. No sporodochia was observed in any of thecultured plates.
Under the microscope, the fungal hyphae appeared hyaline and septate (Figure 1M).Conidiogenous cells were mostly monophialidic (Figure 1N) and rarely polyphialidic.Monophilalidic conidiogenous cells were long (range 10.53-33.94 µm), with an averagelength of 21.43±7.41 µm (Table 1). The microscopic examination of the fungi taken fromthe pure culture plates revealed hyphae with characteristic macro- and microconidia(Table 1). Microconidia were produced in relatively short chains (Figure 1O) or in a falsehead; conidiophores were branched or unbranched, bearing hyaline, monophialidic,conidiogenous cells. Three different forms of microconidia [oval (Figure 1P), fusiform(Figure 1Q), and napiform (Figure 1R)] were found, which were hyaline, thin-walled,and yellowish in colour. Fusiform shaped microconidia had a single septum. The averagesizes of the oval, fusiform, and napiform shaped microconidia were
DEY et al.: BROWN ROT DISEASE IN AZOLLA MICROPHYLLA 247

248 FERN GAZ. 20(6):245-254. 2017
figure. 1(a.-W.): Caption opposite page

10.73±3.31x2.48±0.33 µm, 13.27±1.98 x 4.85±0.48 µm, and 7.17±0.77 x 3.37±0.18 µmrespectively (Table 1).
The macroconidia were found abundantly in aerial parts of the hyphae of the infectedleaf; 3-5 septate, but rarely 6; straight to falcate, thin walled, hyaline; slightly curved andtapering in a banana shape (Figure 1s). The average size of macroconidia is of33.10±5.92 x 3.62±0.28 µm (Table 1). The apical cell of each macroconidia was conical,whereas the basal cell was relatively ill-developed and not the usual foot-shaped. Theaverage length of the apical cell (6.47±0.77 µm) was longer than the penultimate cell(5.29±0.83 µm) (Table 1). It is significant to note that no chlamydospore was found inany one of the culture plates.
The growth pattern of the cultured pathogen onto PDA media and the occurrence ofdistinctive banana-shaped macroconidia and specific types of microconidia indicate thatthe causal pathogen is Fusarium link (Moretti, 2009). Fusarium is a well-known plantpathogenic fungus, which is soil borne in nature and causes severe plant diseases aroundthe world (Agrios, 2005). The Fusarium strain that produces diffusing yellow pigmentwas separated as a new species, F. thapsinum Klittich., lesile, Nelson, Marases(Teleomorph, Gibberella thapsina) (Klittich et al., 1997, summerell et al., 2003; leslie& summerell, 2006).
The characteristic features of cottony white mycelium, yellow pigmentation ontopure culture PDA medium, absence of chlamydospores, shape and size of macro- andmicroconidia corroborate with the diagnostic features of the holotype BPI 737885(Klittich et al., 1997) and hence identify the species epithet of the present causal pathogenas F. thapsinum.
After seven days incubation in the pathogenicity tests, identical disease symptomswith brown-coloured lesions and protruding cottony white mycelium were observed onlyon the inoculated plants (Figure 1V) and the fungus was consistently re-isolated fromlesions. The identity of the re-isolated pathogen was confirmed as F. thapsinum againby pure culturing onto PDA medium (Figure 1W) maintained at similar conditionsmentioned before and subsequent microscopic examinations of the hyphae with
DEY et al.: BROWN ROT DISEASE IN AZOLLA MICROPHYLLA 249
figure 1(a.-W.): Non-infected, and infected Azolla microphylla plant showingsymptoms of brown rot disease, cultured colony of causal pathogen, and microscopicobservation of different features of causal organism (bar= 0.1 mm except otherwisementioned). A. & B. Naturally infected plant on pond (A) and in rice field (B); C.Infected plant grown in tank at experimental garden of Botany Department, KalyaniUniversity; D. Non-infected plant; E. Infected root showing brown coloration; F. Browncoloured spots on leaf (arrow marked) at initial stage of infection; G. Gradualenlargement of infected area on leaf; H. leaf showing bloch notch symptom; I. Entirelyrotten plant at final stage of infection; J. Cottony white mycelial growth on infected leaf;K. Transverse section of infected leaf showing hyphal infiltration; l. Growth pattern ofpure cultured colony showing white mycelium and diffuse yellow pigmentation (dyp);M. septate fungal hyphae; N. Monophialidic conidiogenous cells; O. Occurrence ofmicroconidia in short chain; P.-R. Microconidia of different shapes- P. oval, Q. fusiform,R. napiform; s. Banana-shaped macroconidia; T. Experimentally diseased plants inpathogenicity test; U. Control plants in pathogenicity test; V. Magnified view ofexperimentally diseased plants showing same disease symptoms; W. Growth pattern ofpure cultured colony prepared from re-isolated pathogens from experimental diseaseplants showing white mycelium and diffuse yellow pigmentation (dyp).
250 FERN GAZ. 20(6):245-254. 2017
attributes
measurement (in µm) of macro- and micro- conidia*t value at
18 dfNDP EDP
Range Mean ± sD Range Mean ± sD
macRoconIDIa
No. ofseptation 3 - 6 5.00± 0.79 3 - 6 5.00± 0.94 ---
length ofapical cell 4.82 – 7.58 6.47 ± 0.77 4.78 – 7.83 6.27 ± 1.14 0.4485
length ofpenultimatecell
3.89 – 6.51 5.29 ± 0.83 4.12 – 6.62 5.00 ± 0.78 0.7648
Ratio of apicalto penultimatecell
0.95 – 1.58 1.25 ± 0.22 0.98 – 1.65 1.26 ± 0.18 0.1004
size [length(l) x width(w)]
19.43 – 39.23 (l) 3.06 – 3.98 (w)
33.10 ± 5.92 x3.62 ± 0.28
21.26 – 40.79 (l)2.80 – 4.00 (w)
32.53 ± 4.66 x 3.61 ± 0.35
0.2319 (l)0.0341 (w)
length widthratio 7.63 – 11.50 9.18 ± 1.70 5.84 – 11.39 9.09 ± 1.57 0.1253
mIcRoconIDIa
length ofconidiogenouscell
10.53 – 33.94 21.43 ± 7.41 10.96 – 34.08 22.42 ± 6.87 0.3012
size of ovalshapedmicroconidia[l x w]
7.22 - 16.08 (l)2.05 – 2.92 (w)
10.73 ±3.31 x2.48 ± 0.33
6.99 - 17.09 (l) 1.98 – 3.03 (w)
12.45 ±3.11 x 2.58 ± 0.35
1.1672 (l)0.6400 (w)
size ofnapiformshapedmicroconidia[l x w]
6.18 – 8.26 (l)3.14 – 3.68 (w)
7.17 ± 0.77 x3.37 ± 0.18
6.10 – 8.63 (l) 3.23 – 3.71 (w)
7.47 ± 0.80 x 3.42 ± 0.16
0.8199 (l)0.5843 (w)
size offusiformshapedmicroconidia[l x w]
10.08 – 16.86 (l)3.97 – 5.41 (w)
13.27 ± 1.98 x4.85 ± 0.48
9.77 – 16.23 (l)4.06 – 5.41 (w)
12.86 ± 2.20 x 4.76 ± 0.49
0.4284 (l)0.3894 (w)
* p>0.01 in all cases
table 1: Characterization of macro- and micro- conidia grown in pure cultures prepared from theisolated inoculum of naturally diseased plant (NDP) and the re-isolated inoculum ofexperimentally diseased plant (EDP) of Azolla microphylla

characteristics macro and micro conidia (Table 1). The control plants, however, did notshow any disease symptoms and no traces of F. thapsinum were recovered from them.
Thorough literature survey reveals that previously reported diseases in Azolla areblack rots in A. pinnata from India (Kannaiyan, 1985), Rhizoctonia blight in A. japonicafrom Korea (lee et al., 2011), and A. filiculoides from Iran (Banihashemi, 2014).Kanniyan and Kumar (2005) identified a fungal complex as a causal organism for blackrot disease in A. pinnata, with Rhizoctonia solani as the primary infecting organism.
concluSIonThe present investigation reports a new record of brown rot disease in Azolla microphyllacaused by the fungal pathogen Fusarium thapsinum. Because of the immense value ofthis plant, it will be essential to explore preventive measure(s) to protect this importantbio-resource from destructive fungal attack.
acknoWlEDgmEntSThe financial grant is aided by UGC-MRP (F. No. 41-489/2012 (sR) dated 16th July,2012) and DsT-PURsE programme of University of Kalyani. The first authoracknowledges the financial support received from University of Kalyani as a UniversityResearch scholar. The authors gratefully acknowledge the kind help provided by Prof.Animesh Kumar Datta of Botany Department, University of Kalyani. The authorsgratefully acknowledge the valuable comments and suggestions given by anonymousreviewers which helped a lot to improve quality of the manuscript.
REfEREncESAGRIOs, G.N. 2005. Plant pathology. In: DREIBElBIs, D. (Ed) Plant disease caused
by Fungi, pp 534-540. Elsevier academic press, UsA.AlAlADE, O.A. & lYAYI, E.A. 2006. Chemical composition and the feeding value of
Azolla (Azolla pinnata) meal for egg-type chicks. Int. J. Poult. sci. 5(2):137-141.AlCANTARA, P.F. & QUERUBIN, l.J. 1985. Feeding value of Azolla meal for broilers.
Philipp. J. Vet. Anim. sci. 11:1-8.ANsARI, M.A. & sHARMA, V.P. 1991. Role of Azolla in controlling mosquito breeding
in Ghaziabad district villages (U P). Indian J. Malariol. 28(1):51-54.ANTUNEs, A.P.M., WATKINs, G.M. & DUNCAN, J.R. 2001. Batch studies on the
removal of gold(III) from aqueous solution by Azolla filiculoides. Biotechnol. lett.23:249-251.
BANIHAsHEMI, Z. 2014. Rhizoctonia blight of Azolla in rice paddy fields in NorthernIran. Iran J. Plant Path. 50(4):189-190.
BECERRA, M., PREsTON, T.R. & OGlE, B. 1995. Effect of replacing whole boiledsoya beans with Azolla in the diets of growing ducks. livest. Res. Rural Devt.7(3):1-10.
BRINKHUIs, H., sCHOUTEN, s., COllINsON, M.E., slUIJs, A., DAMsTÃ, J.s.,DICKENs, G.R., HUBER, M., CRONIN, T.M., ONODERA, J. & TAKAHAsHI,K. 2006. Episodic fresh surface waters in the Eocene Arctic Ocean. Nature441:606-609
BUCKINGHAM, K.W., ElA, s.W., MORRIs, J.G. & GOlDMAN, C.R. 1978. Nutritivevalue of the nitrogen-fixing aquatic fern Azolla filiculoides. J. Agri. Food Chem.26(5):1230-1234.
CAGAUAN, A.G. & PUllIN, R.s.V. 1991. Azolla in aquaculture: Past present and
DEY et al.: BROWN ROT DISEASE IN AZOLLA MICROPHYLLA 251

future. In: MUIR, J., ROBERTs, R.J. (Eds) Recent Advances in Aquaculture, pp104-130. Blackwell science, Oxford.
CARRAPICO, F. 2002. The Azolla-Anabaena-Bacterial system as a natural microcosm.In: sPIE (Ed) Proceedings of sPIE Astrobiology Conference on Instruments Methodsand Missions for Astrobiology, IV. pp 261-265. sPIE, san Diego.
COHEN, M.F., WIllIAMs, J. & YAMAsAKI, H. 2002. Biodegradation of diesel fuelby an Azolla-derived bacterial consortium. J. Environ. sci. Health A Tox. Hazard.subst. Environ. Engg. A37(9):1593-1606.
COsTA, M. l., sANTOs, M.C. & CARRAPICO, F. 1999. Biomass characterization ofAzolla filiculoides grown in natural ecosystems and wastewater. Hydrobiologia415:323–327.
COsTA, M.l., sANTOs, M.C.R., CARRAPICO, F. & PEREIRA, A.l. 2009.Azolla-Anabaena’s behaviour in urban wastewater and artificial media – Influenceof combined nitrogen. Water Res. 43(15):3743-3750.
DAO, T.T. & TRAN, Q.T. 1979. Use of Azolla in rice production in Vietnam. In: IRRI(Ed) Nitrogen and Rice, pp 395-405. Int. Rice Res. Inst., Philippines.
DAs, D., sIKDAR, K. & CHETTERJEE, A.K. 1994. Potential of Azolla pinnata asbiogas generator and as a fish-feed. Indian J. Environm. Health 36(3):186-191.
ElMACHlIY, s., CHEFETZ, B., TEl-OR, E., VIDAl, l., CANAls, A. &GEDANKEN, A. 2010. Removal of silver and lead ions from water wastes usingAzolla filiculoides an aquatic plant which adsorbs and reduces the ions into thecorresponding metallic nanoparticles under microwave radiation in 5 min. Water Airsoil Pollut. 218:365-370.
FIOGBE, E. D., MICHA, J.C. & VAN, H.C. 2004. Use of a natural aquatic fern, Azollamicrophylla as a main component in food for the omnivorous-phytoplanktonophagoustilapia, Oreochromis niloticus l. J. Appl. Ichthyol. 20(6):517–520.
HAll, D.O., MARKOV, s.A., WATANABE, Y., RAO, K.K. 1995. The potentialapplications of cyanobacterial photosynthesis for clean technologies. Photosyn. Res.46(1):159-167.
JAIN, s.K., VAsUDEVAN, P.K. & JHA, N.K. 1989. Removal of some heavy metalsfrom polluted water by aquatic plants: studies on duckweed and water velvet. Biol.Wastes 28(2):115-126.
KANNAIYAN, s. & KUMAR, K. 2005. Black rot disease interactions and nitrogenfixation in Azolla. In: Daya Publishing House (Ed) Azolla biofertilizer for sustainablerice production, pp 404-421. Daya Publishing House, Delhi.
KANNIAIYAN, s. 1985. Use of Azolla in India. In: IRRI (Ed) Proceedings of theWorkshop on Azolla Use, pp 109-118. Fuzhou Fujian, China.
KATAYAMA, N., YAMAsHITA, M., KIsHIDA, Y., lIU, C.C., WATANABE, I.,WADA, H. & sPACE AGRICUlTURE TAsK FORCE. 2008. Azolla as a componentof the space diet during habitation on Mars. Acta Astronautica. 63(7-10):1093-1099.
KHOsRAVI, M., RAKHsHAEE, R. & GANJI M.T. 2005. Pre-treatment processes ofAzolla filiculoides to remove Pb(II), Cd(II), Ni(II) and ZN(II) from aqueous solutionin the batch and fixed-bed reactors. J. Hazard. Mater 127(1-3):228-237.
KlITTICH. C.J.R., lEslIE, J.F., NElsON, P.E. & MARAsAs, W.F.O. 1997. Fusariumthapsinum (Gibberella thapsina): A new species in section Liseola from sorghum.Mycologia 89(4):643-652.
KROCK, T., AlKAMPER, J. & WATANABE, I. 1991. Azolla’s contribution to weedcontrol in rice cultivation. Plant Res. Developm. 34:117-125.
252 FERN GAZ. 20(6):245-254. 2017

lATTANZIO, V., lATTANZIO, V. M.T. & CARDINAlI, A. 2006. Phytochemistry:Advances in Research. In: IMPERATO F. (Ed) Role of phenolics in the resistancemechanisms of plants against fungal pathogens and insects, pp 23-67. IsBN:81-308-0034-9. Research signpost, India
lEE, J., CHA, J., NOH, G., HAN, K., BAE, D., KWON, Y., lIM, C, JEONG, s, KWON,J., PARK, C. & KWAK, Y. 2011. Rhizoctonia blight of Azolla japonica caused byRhizoctonia solani. Res. Plant Dis. 17(3):405-409.
lEJEUNE, A., PENGA, J., BOUlENGE, E., lARONDEllE, Y. & VAN, H.C. 2000.Carotene content of Azolla and its variations during drying and storage treatments.Anim. Feed sci. Tech. 84:295-301.
lEslIE J.F. & sUMMEREll, B.A. 2006. species descriptions. In: The Fusariumlaboratory manual, pp 264-265. Blackwell Publishing, Iowa.
lIU, X., MIN, C., XIA-sHI, l. & CHUNGCHU, l. 2008. Research on some functionsof Azolla in CElss system. Acta Astronaut. 63:1061-1066.
lUMPKIN, T.A. & PlUCKNETT D.l. 1982. Azolla as a green manure: Use andmanagement in crop production, Westview Press, Boulder Colorado.
MAsHKANI, s.G. & GHAZVINI, P.T.M. 2009. Biotechnological potential of Azollafiliculoides for biosorption of Cs and sr: Application of micro-PIXE for measurementof Biosorption. Bioresource Technology 100(6):1915-1921.
MORETTI, A.N. 2009. Taxonomy of Fusarium genus a continuous flight betweenlumpers and splitters. Proc. Nat. sci. Matica. srpska. Novi. sad. 117:7-13.
NEWTON, J.W. 1976. Photoproduction of molecular hydrogen by a plant-algalsymbiotic system. sci. 191(4227):559-561.
PAOlETTI, C., BOCCI, F., lERCKER, G., CAPEllA, P. & MATERAssI, R. 1987.lipid composition of Azolla caroliniana biomass and its seasonal variation.Phytochem. 26(4):1045–1047.
PETERs, G.A. 1978. Blue-green algae and algal associations. Bioscience 28(9):580-585.PETERs, G.A., EVANs, W.R., TOIA, R.E. 1976 Azolla-Anabaena azollae relationship.
Plant Physiol. 58:119-126.PRAsANNA, R., KUMAR, V., KUMAR, s., YADAV, A.K., TRIPATHI, U., sINGH,
A.K., JAIN, M.C., GUPTA, P., sINGH, P.K. & sETHUNATHAN, N. 2002. Methaneproduction in rice soil is inhibited by cyanobacteria. Microbiol. Res. 157(1):1-6.
RAI, P.K. & TRIPATHI, B.D. 2009. Comparative assessment of Azolla pinnata andVallisneria spiralis in Hg removal from G.B Pant sagar of singrauli Industrial regionIndia. Environ. Monit. Assess. 148(1):75-84.
RAI, P.K. 2008. Phytoremediation of Hg and Cd from industrial effluents using anaquatic free floating macrophyte Azolla pinnata. Int. J. Phytoremediation. 10(5):430-439.
RAI, P.K. 2010a. Microcosm investigation on phytoremediation of Cr using Azollapinnata. Int. J. Phytoremediation 12(1):96-104.
RAI, P.K. 2010b. Heavy metal pollution in lentic ecosystem of subtropical industrialregion and its phytoremediation. Int. J. Phytoremediation 12(3):226-242.
sAlEHZADEH, A., NAEEMI, A.s. & ARAsTEH, A. 2014. Biodiesel production fromAzolla filiculoides (water fern). Trop. J. Pharm. Res. 13(6):957-960.
sAsI, P.s., WIlsON, K.I. & sAsIDHARAN, N.K. 1982. Incidence of Rhizoctoniasolani on Azolla pinnata. Curr. sci. 48(5):216.
sAXENA, D.K. 1995. Purification efficiency of Lemna and Azolla in WIMCO effluents.Proc. Natl. Acad. sci. India sect. B Biol. sci. 65:61-65.
DEY et al.: BROWN ROT DISEASE IN AZOLLA MICROPHYLLA 253

sHIOMI, N., KITOH, s. 2001. Culture of Azolla in a pond nutrient composition and useas fish feed. soil sci. Plant Nutr. 47(1):27–34.
sOOD, A., UNIYAl, P.l., PRAsANNA, R., AHlUWAlIA, A.s. 2012.Phytoremediation potential of aquatic macrophyte Azolla. Ambio. 41:122-137
sUMMEREll, B.A., sAllEH, B. & lEslIE J.F. 2003. A Utilitarian Approach toFusarium Identification. Plant Dis. 87(2):117-128.
TUNG, H.F. & sHEN, T.C. 1985. studies of the Azolla pinnata-Anabaena azollaesymbiosis: Concurrent growth of Azolla with rice. Aquatic Bot. 22(2):145-152.
UMAlI, l.J., DUNCAN, J.R. & BURGEss, J.E. 2006. Performance of dead Azollafiliculoides biomass in bioabsorption of Au from wastewater. Biotechnol. lett.28(1):45-49.
WAGNER, G.M. 1997. Azolla: A review of its biology and utilization. Bot. Rev.63(1):1-26.
WATANABE, I. & lIU, C.C. 1992. Improving nitrogen-fixing systems and integratingthem in to sustainable rice farming. Plant soil 141:57-67.
YADAV, R.K., ABRAHAM, G., sINGH, Y.V. & sINGH P.K. 2014. Advancements inthe utilization of Azolla-Anabaena system in relation to sustainable agriculturalpractices. Proc. Indian Natn. Acad. sci. sect. B Biol. sci. 80(2):301-316.
254 FERN GAZ. 20(6):245-254. 2017

CYSTOPTERIS FRAGILIS SuBSp. HUTERI, a taXon nEW to thEBRItISh ISlES anD RElatED taXa In thE CYSTOPTERIS
FRAGILIS complEX: CYSTOPTERIDACEAE (polypoDIopSIDa)
D.J. TENNANTlow Missise Farm, laverton, Ripon, HG4 3sY, UK.
Email: [email protected]
Keywords: Bladder ferns, Cystopteris, C. alpina, C. regia, Cyathea incisa, taxonomy,new taxa, Wales, scotland, distribution
aBStRactA taxon in the Cystopteris fragilis (l.) Bernh. complex that occurs in Britainappears up to now to have been overlooked there. It is proposed that this fern,which was recently rediscovered in snowdonia, North Wales, should be referredto a subspecies of C. fragilis, namely subspecies huteri (Hausm. ex Milde) C.Prada & salvo, that occurs in mainland Europe, but had not previously beenrecognised in the British Isles. The snowdonia fern is distinct in its morphologyfrom other British taxa in the C. fragilis complex and is described here togetherwith its historic records and present distribution, updated from information givenin an earlier paper in Watsonia (Tennant, 2010), and its relationship to affiliatedtaxa is discussed.
IntRoDuctIonThe Cystopteris fragilis complex is perhaps the most difficult one to resolve of all ferngroups. In the British Isles it presently includes the species C. fragilis (l.) Bernh., C.alpina (lam.) Desv., C. diaphana (Bory) Blasdell, and C. dickieana R. sim. It isgenerally accepted that more than one original diploid parent was probably involved inthe ancestry of the present species recognised in the complex. One result of which is thelarge variation between and within the species, making it especially difficult to recognisethe different cytotypes based solely on their gross morphology. Over many decades, astudy was made of the species within the complex, and especially their variants, both inthe field and in herbarium collections. This led to earlier papers being published on C.alpina and C. dickieana in Britain (Tennant, 1995; 1996; 2010), but two especiallydistinctive taxa were noted in Britain, which in their morphology did not readily conformto the appearance or descriptions of any of the known presently extant British taxa withinthe complex.
The first of these was found in 2008 in snowdonia, V.C. 49, North Wales, where itwas extremely local and scarce. A search in herbarium collections revealed that amorphologically identical taxon had been collected previously there. A paper publishedin Watsonia (Tennant, 2010) gave most of the records of this fern, which are updatedand relisted in this paper, together with its historical references, description, illustrations(Figures 1 & 2), and habitat, and evidence for the presently proposed identity of thissnowdonia fern is given.
The second distinctive Cystopteris taxon discovered during the course of these studiesoccurs in northern scotland, and a similar taxon also in North Wales. These ferns appearto be affiliated to certain variants currently included under C. alpina (lam.) Desv.,previously as “C. regia” auct., non (l.) Desv. Morphologically these scottish ferns
FERN GAZ. 20(6):255-266. 2017 255

closely match certain Cystopteris forms found by the author recently in alpine regionsof mainland Europe in three different countries (Figures 3 & 4). living material anddetails of these scottish ferns, which were grown from spores, have been donated to theNatural History Museum in london for further study and will be the subject of a laterpaper.
256 FERN GAZ. 20(6):255-266. 2017
figure 1. Cystopteris fragilis subsp. huteri. Cultivated examples fromsnowdonia, V.C.49, North Wales illustrating the degree of variation. (Furtherillustrations were given previously in Tennant (2010)).
0 1 2 4 cms

TENNANT: CYSTOPTERIS FRAGILIS SUBSP. HUTERI 257
figure 2. Cystopteris fragilis subsp. huteri. snowdonia, V.C. 49, North Wales.Cultivated specimen.
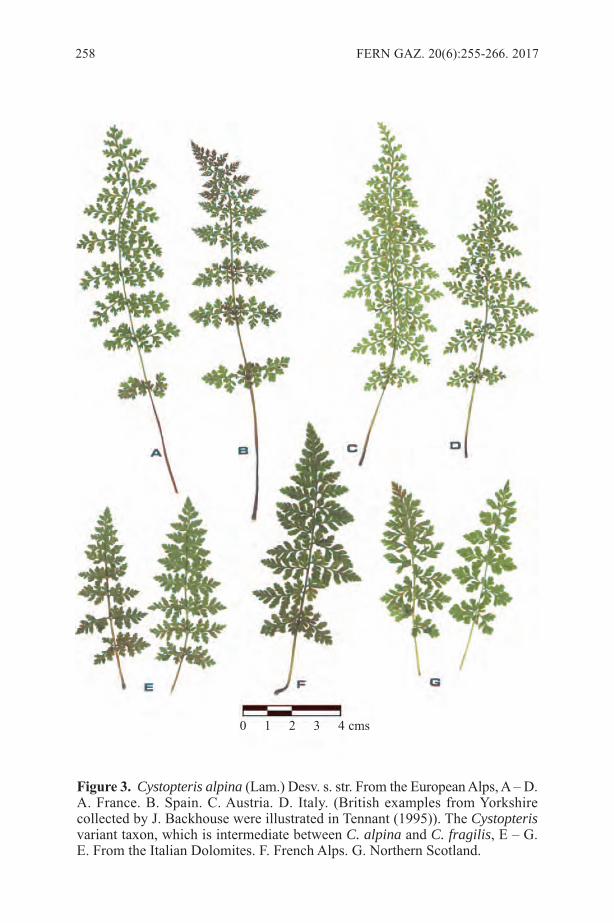
258 FERN GAZ. 20(6):255-266. 2017
figure 3. Cystopteris alpina (lam.) Desv. s. str. From the European Alps, A – D.A. France. B. spain. C. Austria. D. Italy. (British examples from Yorkshirecollected by J. Backhouse were illustrated in Tennant (1995)). The Cystopterisvariant taxon, which is intermediate between C. alpina and C. fragilis, E – G.E. From the Italian Dolomites. F. French Alps. G. Northern scotland.
0 1 2 3 4 cms

TENNANT: CYSTOPTERIS FRAGILIS SUBSP. HUTERI 259
figure 4. Cystopteris variant from Northern scotland, V.C. 105, which isintermediate between C. alpina and C. fragilis.

matERIalS anD mEthoDSThe fieldwork described below on Cystopteris in Britain dates back over many decadesthrough an early interest of the author, particularly in the British records for C. alpina.More recently, following several examinations of herbarium specimens at the NaturalHistory Museum (NHM) london (Bm) and at the Royal Botanic Garden, Edinburgh (E)between 2004 and 2006, details of all relevant specimens were abstracted and comparedwith recently found examples of Cystopteris. This was followed by further fieldwork inNorth Wales and in scotland. Permission was obtained from the Countryside Councilfor Wales to collect a small amount of material on snowdon, V.C. 49, which wasundertaken in 2009, following which this snowdonia fern was cultivated in Yorkshire.living examples of the snowdonia Cystopteris were then donated to NHM in londonfor further studies. This work is being undertaken by Dr F.J. Rumsey at NHM, and sofar has included a detailed examination of spores and some plastid DNA studies.
Meanwhile, historic publications were examined and information dating back to the17th century was abstracted, after it was discovered that the snowdonia Cystopteris hadfirst been collected at that time. Field studies on Cystopteris were more recently extendedin alpine regions of mainland Europe, namely Norway (Tromsø 2009), France (VanoiseAlps 2010), spain (Central Pyrenees 2012), Italy (Dolomites 2014, 2015), and AustrianTyrol (2016). Material found there was compared with examples from Britain.
RESultSDescription of the Snowdonia CystopterisThis fern was largely described and illustrated in an earlier paper (Tennant, 2010) fromsamples that had been found in snowdonia in 2008, and cultivated from 2009, althoughone crucial character was then omitted. During examination of the cultivated material itwas noted that the fronds were distinctly glandular, having scattered, capitate glandularhairs on the pinnae, a character unique in British Cystopteris. A further more detaileddescription is as follows, with illustrations given in Figures 1 and 2.
stipe slender, 1.5 – 6 cm. brownish, with scattered pale brown linear scales, frond(lamina) lanceolate, 4 – 10 x 2 – 5 cm, pinnae mainly narrowly ovate, 1.5 – 2.5 x 0.7 – 1.3cm, with scattered, capitate glandular hairs, which are most visible on the margins,pinnules stalked, the lower oblong-obdeltoid, subtruncate at the apex, often with short,sharply acute points, the upper narrower, linear-obdeltoid, more acute at the apex, theveins mainly terminating in the acute apical points. sori scattered, situated adjacent tothe veins, spores echinate, very large, mean c. 50 microns (F.J. Rumsey, pers.comm.).
The juvenile sporophytes have well-separated (divaricate), linear-oblong, bluntpinnules, unlike those of typical C. fragilis, but very similar to those of C. alpina. Theimmature, intermediate fronds usually have either obdeltoid (wedge-shaped) or linearpinnule lobes, again similar to those seen in C. alpina. The mature fronds are more finelydissected, sometimes with pinnae obliquely inclined to the plane of the lamina. Thesnowdonia ferns, although sharing several characters with C. alpina are readilydistinguished from it by the acute points at the apex of the pinnule lobes into which themajority of the veins terminate, and the unique presence of glandular hairs on the pinnae.Plastid DNA examination has also confirmed that the ferns are not C. alpina (F.J.Rumsey, pers. comm.). The relationship of these snowdonia ferns to a described,glandular subspecies of C. fragilis, which occurs in southern, central and westernmontane regions of mainland Europe is discussed below.
260 FERN GAZ. 20(6):255-266. 2017

habitatThe snowdonia fern is a plant of the alpine zone in Britain, probably not occurring ataltitudes below 700m. In the two localities in which it was rediscovered in 2008 it wasextremely local and scarce. It occurred on basic rocks in a north-facing vertical creviceand in a recess on the cliff face. No other vascular plants were present, but Cochleariapyrenaica subsp. alpina (Bab.) Dalby, Minuartia verna (l.) Hiern and Silene acaulis (l.)Jacq. occurred nearby, and typical C. fragilis was not far distant.Records of the Snowdonia Cystopterisspecimens in British herbarium collections and references in historical literature showedthat the fern that had been rediscovered in snowdonia in 2008 had been collected verymuch earlier. Identical material was located by the author in 2004-2006 at the NaturalHistory Museum (NHM) in london (Bm) and other relevant Cystopteris specimens werefound in the herbarium of the Royal Botanic Garden (RBGE) in Edinburgh (E), and thesnowdonia records are given below. This fern was first collected in the late 1680s byEdward lhwyd in snowdonia, and two of lhwyd’s original specimens were recentlylocated by F.J. Rumsey in the sloane herbarium at Bm; these specimens were identicalto those found on snowdon by the author in 2008. Further specimens collected bylhwyd, which are probably the same, are present in the herbarium of Oxford University(oXf) (Edgington, 2013). lhwyd’s specimens were also sent to John Ray and detailswere included in Ray’s Synopsis (Ray, 1724). lhwyd’s record and others fromsnowdonia were later included by sir J.E. smith (1828; 1834) in his English Flora, butunder Cystea regia smith, possibly the first, but illegitimate, binomial name given tothis fern, which is discussed later in this paper.
The snowdonia Cystopteris would meet the I.U.C.N. Threat Category of ‘criticallyendangered’, and is especially vulnerable to collecting, therefore precise details of itslocations are withheld, but otherwise its presently known records are as follows:snowdonia, sH65, Caernarvonshire, V.C.49 circa 1688, Edward lhwyd (sloane herb.,Bm; oXf) snowdon, sH65, V.C.49, 1824, collector unknown, herb.Rev. William Newbould as C.fragilis (Bm)snowdon, 1826, William Wilson (?herb. J.T.I. Boswell-syme), as Cyathea regia (in theC. fragilis folders) (Bm)snowdon, 1847, collector uncertain, herb. F.J. Hanbury, as C. fragilis var. (Bm)Near Glyder Fawr (Cwm Idwal), sH65, V.C.49, 1853, collector’s name illegible, in C.alpina folder (Bm)Cwm Glas, snowdon, 1874, collector uncertain (same sheet gives H. & J. Groves alsoW.R. linton), as C. fragilis (Bm)snowdon, 1890. J. lloyd Williams, in herb. Arthur Bennett, as C. alpina (later renamedC. fragilis) (Bm)snowdon, 2008, 2009, D.J. Tennant; Cwm Glas area, snowdon, 2008, D.J.T. (herb. DJt;Bm)In several cases not all of the specimens on the sheets for the above records were thesame taxon; as few as a single specimen conformed to the snowdonia Cystopteris inquestion in at least one case. A single specimen on the sheet which represents the 1853record given above, and those representing the 1853 record are morphologically identicalto the others, but require to be re-examined for the presence of glandular hairs. Glandularhairs were present on specimens representing the remaining seven records given above.
A specimen marked ‘W.J. Hooker’ in Bm, but undated, was collected at Castell Dinas
TENNANT: CYSTOPTERIS FRAGILIS SUBSP. HUTERI 261

Bran, Denbighshire, V.C.50, and named Cyathea regia by Hooker, but later renamed asa variety of C. fragilis. This record was listed by Tennant (2010), but is not the snowdoniaCystopteris for which records are given above. This interesting specimen bearsresemblance to those found in northern scotland, which were mentioned in theIntroduction, and require further investigation. A careful search by the author at CastellDinas Bran and adjacent rocks failed to locate Hooker’s fern, which is probably extinctthere. similarly, a recent but lesser search in Cwm Idwal did not reveal the snowdoniaCystopteris taxon.
Following the discovery that the snowdonia taxon was glandular, it was drawn tothe attention of the author by another botanist, Brian Burrow, that Flora Iberica (Prada,1986) describes an infra-specific form of C. fragilis that is distinguished by the presenceof glandular hairs, namely subspecies huteri (Hausm. ex Milde) C. Prada & salvo. Thedescription, illustration and spore size given (44-55 (60) microns) for this subspeciesmatched those of the snowdonia Cystopteris very well. subsp. huteri is a taxon foundmainly in montane regions of southern and western mainland Europe, with a generaldistribution similar to that of C. alpina (lam.) Desv., and has been linked to the latter inscandinavia (F.J. Rumsey, pers. comm.), although the extent of its further geographiclimits is not certain.
In 2014 and 2015 fieldwork was carried out by the author in the central ItalianDolomites, which is the centre of distribution of subsp. huteri (F.J. Rumsey, pers. comm.).subsp. huteri was found to be frequent there, enabling a comparison to be made withthe snowdonia Cystopteris, and material was donated to the NHM for further study byF.J. Rumsey. In the Dolomites C. fragilis subsp. huteri occurred mainly between 1600and 2400m on dolomitic limestone in open, rocky, alpine woodland, extending toboulders and scree at the base of cliffs well above the tree-line, both habitats with a veryrich alpine vascular flora. The other ferns often present were Cystopteris fragilis subsp.fragilis, C. alpina, C. montana (lam.) Desv., Gymnocarpium robertianum (Hoffm.)Newman, Asplenium trichomanes l. subsp. quadrivalens D.E. Meyer emend. lovis, A.trichomanes-ramosum l., A. ruta-muraria l. (including subsp. dolomiticum) andoccasionally Woodsia glabella R. Br. ex Richards. Cystopteris fragilis subsp. huteri inthe Dolomites was more variable than the snowdonia fern, but close matches to the latterwere found there, the majority being intermediate between C. fragilis and C. alpina ingeneral appearance. F.J. Rumsey has agreed that these snowdonia ferns should bereferred to C. fragilis subsp. huteri.
A very small population of another morphologically distinctive Cystopteris was alsofound by the author in the Dolomites, very locally at 2300m, and this matched the fernmentioned in the Introduction that occurred in scotland (Figures 3 & 4). The herbariumstudies at RBGE (E) showed that this scottish fern had been collected earlier there, andwas re-found in Orkney, V.C. 111, in 2010, and later in 2014 by the author in West Ross,V.C. 105, then cultivated from spores. Morphologically similar ferns had also been seenby the author in the French Vanoise and Norwegian Alps. All of these ferns are somewhatintermediate in their morphology between C. fragilis and C. alpina. Those from mainlandEurope occur there with C. alpina s. str., although often at lower altitudes, and at leastsome of these are currently referred to C. alpina. living examples from both recentlyfound localities in scotland that resemble the ferns from mainland Europe have beendonated to NHM for biosystematic analyses in order to study their phylogeneticrelationships and to establish ploidy levels.
262 FERN GAZ. 20(6):255-266. 2017

DIScuSSIonVarious authors have commented on the difficulties of diagnosing taxa in the Cystopterisfragilis complex by use of morphological characters alone, for example Vida (1974) andRothfels et al. (2013). Blasdell (1963), however, had successfully developed anevolutionary structure, which in most cases showed a good correlation with recentphylogenetic analyses (Rothfels et al., 2013). The main problems arise because ofmultiple ancestral origins, the existence of many polyploids, and in Europe, the absenceof the ancestral diploid parent. In Europe C. fragilis is tetraploid, hexaploid, or octoploid,but in the British Isles the last is so far unknown. Intermediate ploidy levels have arisenthrough intraspecific hybridisation, and pentaploid examples are said to be frequent(Vida, 1974). It should therefore be emphasised that the main conclusion in this paper,namely the recognition of C. fragilis subsp. huteri in the British Isles, is very largelybased on morphological characters. In this case, however, the morphology of thissnowdonia fern is quite distinct from other British variants of C. fragilis; the spores areexceptionally large, and the glandular condition is presently a unique character inEuropean Cystopteris. These characters provide the confidence to refer the snowdoniaCystopteris to this European taxon.
As stated above, diploid Cystopteris is not recorded in Europe, but in North Americaseveral glandular species of Cystopteris are known and their glandular condition hasresulted from the donation of a maternal genome from the diploid plant, C. bulbifera(l.) Bernh. (Rothfels et al., 2003). One glandular North American allohexaploid species,C. laurentiana (Weatherby) Blasdell, which possibly arose as a cross between C. fragilisand C. bulbifera, has been suggested to be synonymous with C. fragilis subsp. huteri(F.J. Rumsey, pers. comm.), but that would imply that C. bulbifera at one time hadoccurred in Europe, which at present is not proven.
The snowdonia fern here referred to C. fragilis subsp. huteri is clearly responsiblefor all of the previously published erroneous records in Wales for C. alpina s. str. (butsee comments above in Results, and below regarding a Welsh fern named by W.J.Hooker). The snowdonia fern does share some characters found in C. alpina, which mayhave led to the confusion, namely the close similarity of its juvenile sporophytes and themorphology of its immature pinnules (see under Results above and Tennant (2010)).These characters probably arose from an ancestral genome donated from C. alpina, whichmay therefore have once occurred in Wales. The very large spore size given for C. fragilissubsp. huteri in Flora Iberica was suggested by the authors of this publication to indicatethat this taxon is probably octoploid. spores of snowdonia examples were found to besimilar in size (F.J. Rumsey, pers. comm.), suggesting that these ferns were likely to beat least hexaploid, if not octoploid, a ploidy level not recorded for C. fragilis in the BritishIsles. Blasdell (1963) had suggested that spore size in Cystopteris could be used in thedetermination of the different cytotypes, but Vida (1974) found that, contrary to thatopinion, spore size was not always reliable in this respect. R.H. Roberts measured thespores of two different examples of C. alpina for the author. These both had almostidentical finely dissected fronds with linear pinnule segments that are associated withclassic C. alpina s. str. (Figure 3), but were found to differ widely in spore size (Tennant,1995), supporting Vida’s findings, which require further investigation using the variouscytotypes. Vida (1974) also found that a deliberately induced apogamous diploid of C.fragilis had a much more finely dissected frond than the natural tetraploid plants fromwhich it had been derived, and that can clearly be seen in his Plate 2, fig. 9, which mightsuggest that degree of frond dissection may not necessarily increase with an increasing
TENNANT: CYSTOPTERIS FRAGILIS SUBSP. HUTERI 263

ploidy level.Historic Cystopteris nomenclature used by British authors is very relevant to C.
fragilis subsp. huteri from snowdonia. This particularly applies to the names ‘incisa’,‘regia’ and ‘alpina’, and is also relevant to the related taxa from scotland and other taxafrom Wales, which are discussed earlier in this paper. smith (1794) in Volume 3, p.163published the name Cyathea incisa sm., ‘laciniated Cup-fern’ when describing andillustrating a fern collected pre 1790 by E. Forster at a single locality near Walthamstowin south Essex, V.C. 18. smith stated that this fern was not the Polypodium regium oflinnaeus and was also different from P. alpinum Jacquin. smith’s description of this ferngives ‘frond bipinnate’, (the pinnule lobes) ‘obtuse with no bristly point’, the lastcharacter clearly seeming to distinguish it from the snowdonia subsp. huteri. smith(1828; 1834) later published the name Cystea regia sm., and here he cited lhwyd’s late17th century snowdon record, two further records from snowdon, namely one by theRev. Hugh Davies in the early 19th century and one by W. Wilson in 1826, and a furtherrecord by Mr. Griffith from Cwm Idwell (Idwal), which all almost certainly referred tothe snowdonia subsp. huteri. smith, however, surprisingly also added here the Essexfern (Cyathea incisa and C. regia Forster), thus seeming to suggest that these weresynonymous with the snowdonia ferns, even though their descriptions differed. smithalso stated that Cystea regia sm. was unquestionably distinct from every other Britishfern, and from the ‘regia’ of alpine stations (in Europe), presumably meaning the fernnow referred to Cystopteris alpina (lam.) Desv., although confusingly he includedAspidium regium sw. (1802) in the synonymy.
W.J. Hooker (1842) then grouped together all of smith’s taxa under C. alpina Desv.,and added ‘tripinnate’ to smith’s earlier ‘bipinnate’ description of these taxa. Hookerhad earlier (before 1842) confirmed the name of one of the specimens from Essexcollected by Forster as Cyathea regia (stansfield, 1930), and in the same period had alsogiven the same name to an undated specimen in Bm that had been collected inDenbighshire. V.C. 50. These Essex and Denbighshire records, in the sense of Hooker(1842), therefore represent the first and second British records respectively for C. alpina.J. Backhouse’s record from Upper Teesdale in Yorkshire, V.C. 65 in 1872 (Tennant, 2010)would therefore be the third British record. In spite of smith (1828; 1834) and Hooker(1842) including the snowdonia ferns under the species which is now referred to C.alpina (lam.) Desv., C. fragilis subsp. huteri from snowdonia is shown here to bedifferent, and its first name, Cystea regia sensu sm. (parte) should be retained only insynonymy.
The use of the name ‘regia’ for some of these ferns is, however, incorrect on theassumption that the name ‘regia’ was entirely derived from P. regium l. The type sheetof P. regium in lInn contains more than one species and P. regium was therefore a‘nomen ambiguum’. None of the species on this sheet represent the taxa discussed inthis paper, including C. alpina (lam.) Desv.. This has long been known (stansfield,1929), and much earlier, for example P. regium l. was included by J.T. syme in thesynonymy under C. fragilis in the third edition of English Botany, Vol. 12 (1886), whichwould be correct, although an individual specimen still requires to be selected from thetype sheet and formally lectotypified as such.
Other authors also considered ‘regia’ to be distinct. Milde made ‘regia’ a variety ofC. alpina (Kestner, 1930), and stansfield (1929) proposed the name C. alpina var. obtusafor Forster’s Cystopteris from Essex. After examining the Essex specimen, stansfieldpointed out that it was bipinnate, with only shallowly notched pinnule lobes, many of
264 FERN GAZ. 20(6):255-266. 2017

the veins terminating in an apical point, unlike C. alpina s.str., which is at least tripinnatewith virtually all the veins ending in notches. Kestner (1930) had stated that the positionof the vein-endings was the only reliable character in the determination of C. alpina,additionally the Cystopteris key given in Murphy and Rumsey (2005) similarly relies onthis character for the determination of C. alpina.
The first name given to the Essex fern, Cyathea incisa sm. (1794), would be alegitimate name at specific level, and for the Denbighshire Cystopteris as this appears tobelong to the same taxon. It is doubtful, however, whether these ferns could be justifiedas a species distinct from C. alpina, in which case the name would be reduced tosynonymy under the latter. The Essex and Denbighshire ferns are both thought to beextinct at their original localities. A similar Welsh taxon from Merionethshire, V.C. 48,has not been collected since 1928, so none of these ferns are presently available forbiosystematic studies that might elucidate their correct status. The Cystopteris taxon fromscotland discussed briefly above also seems to have affinities with C. alpina. Twoexamples of this and a morphologically identical fern from Italy are in cultivation by theauthor, and material is now at NHM for further studies to establish their status.
several recently confirmed specimens of C. alpina seen by the author in the TromsøMuseum herbarium had exceptionally broad pinnule lobes. Following the fieldwork bythe author, uniformly similar Cystopteris forms were also encountered in alpine regionsof France, Italy, and in scotland, some of which are presently determined as C. alpina(lam.) Desv. This might suggest that the concept of C. alpina s.str. as a fern restrictedto one with a finely dissected frond and linear pinnule lobes, and with the veins virtuallyall ending in notches, unlike the ‘regia’ of some authors, may be one that is too narrow.An alternative suggestion may be that these ferns with broader pinnules truly representa distinct taxon, which is affiliated to C. alpina through the presence of a genome fromthis species. Hopefully biosystematics will resolve this.
Cystopteris fragilis subsp. huteri is presently known in Britain only from twolocalities in North Wales, where it is very local and scarce. It meets the I.U.C.N. ThreatCategory of Critically Endangered (CR) and should be considered for inclusion in theBritish Red Data list, its main vulnerability being one of collecting.
acknoWlEDgEmEntSI would especially like to thank Alison Evans for assistance in the preparation of themanuscript, Clive stace for general advice, Brian Burrow for information on the spanishpublication, Clive Jermy for details of previous papers, Eric Meek for organisingcollection of specimens from one of the scottish sites, Fred Rumsey for information,comments, opinions, and undertaking the preliminary studies at NHM, Arve Elvebakk,Torstein Engelskjøn and Douglas McKean for access to herbarium material, and JohnRatcliffe at the Countryside Council for Wales for permission to collect material.
REfEREncESBlAsDEll, R.F. 1963. A monographic study of the fern genus Cystopteris. Mem.
Torrey. Bot. Club 21:1-102EDGINGTON, J. 2013. Who found our ferns? British Pteridological society, special
Publication 12.HOOKER, W.J. 1842. The British Flora, Vol.1. longman, londonJERMY, A.C. & HARPER, l. 1971. spore morphology of the Cystopteris fragilis
TENNANT: CYSTOPTERIS FRAGILIS SUBSP. HUTERI 265

complex. British Fern Gazette 10: 211-213.KEsTNER, P. 1930. Cystopteris alpina Desv. British Fern Gazette 6 (2): 33-37.MURPHY, R.J. & RUMsEY, F.J. 2005. Cystopteris diaphana (Bory) Blasdell
(Woodsiaceae) – an overlooked native new to the British Isles? Watsonia 25:255-263.PRADA, C. 1986. Cystopteris pp. 115-121 in Flora Iberica, ed. s. Castroviejo. Madrid,
C.s.I.C.RAY, J. 1724. synopsis Methodica stirpium Britannicarum, edn. 3 1973 facsimile, Ray
society, london.ROTHFEls, C., WINDHAM, M.D., PRYER, K.M. 2013. A plastid phylogeny of the
cosmopolitan fern family Cystopteridaceae (Polypodiopsida). systematic Botany 38(2): 295-306. The American society of Plant Taxonomists.
sCHMAKOV, A. 2001. synopsis of the ferns of Russia. Turczaninowia 4: 36-72.sMITH, J.E. 1794 . English Botany, edn. 1, Vol. 3:163. london.sMITH, J.E. 1828. The English Flora, edn. 1, Vol. 4:302. london.sMITH, J.E. 1834. The English Flora, edn. 2, Vol. 4:289. longman, london.sTANsFIElD, F.W. 1929. Cystopteris alpina Desv. British Fern Gazette 6 (1): 15-19.sTANsFIElD, F.W. 1930. Cystopteris alpina Desv. British Fern Gazette 6 (3): 71-72.TENNANT, D.J. 1995. Cystopteris fragilis var. alpina Hook. in Britain. The Naturalist
120: 45-50.TENNANT, D.J. 1996. Cystopteris dickieana R. sim in the central and eastern scottish
Highlands. Watsonia 21: 135-139.TENNANT, D.J. 2010. The British records of Cystopteris alpina (lam.) Desv. Watsonia
28: 57-63.VIDA, G. 1974. Genome analysis of the European Cystopteris fragilis complex. 1
Tetraploid taxa. Acta Biologica Academiae scientiarum Hungaricae. Tomus 20 (1-2):181-192.
266 FERN GAZ. 20(6):255-266. 2017

FERN GAZ. 20(6). 2017 267

InStRuctIonS foR authoRSpapERS should not usually exceed 20 printed pages and are generally expected to beconsiderably shorter. As well as reports of original research, review articles and shortnotes, e.g. new records, are encouraged. The senior author should supply an email addressto facilitate correspondence. REVIEWS should cover recent developments in any aspect of research on ferns andlycopods. This can include unpublished research by the author, and/or the possibledirection of future research. They should be aimed at the general reader who may not bean expert in the chosen topic. Proposals for a review should be discussed first with thereview editors.manuScRIptS should be submitted in English (British) in electronic format in10-point Times New Roman font and single spaced. Electronic versions of text and tablesshould be compatible with WORD, with figures as TIFF files, and sent as emailattachments. All manuscripts will be refereed.thE tItlE should reflect the content of the paper and be in BOlD CAPITAls(11-point) and centrally aligned. Generic and specific names should be in italics and anytitle containing a generic or specific name should be followed by the family name inbrackets e.g.
TRICHOMANES SPECIOSUM (hymEnophyllacEaE)In SouthERn SpaIn
authoR aBBREVIatIonS should follow Pichi sermolli’s (1996) Authors ofscientific names in Pteridophyta, Royal Botanic Gardens, Kew.maIn hEaDIngS: should be in BolD capItalS (10-point) and centrally aligned.SuBSIDIaRy hEaDIngS: should be in bold, the first letter of each word in capitals,the rest in lower case and left-aligned.authoRS’ namES anD full aDDRESSES (including email address): followthe title and are centrally aligned.kEy WoRDS: up to ten.aBStRact: should reflect the content of the paper.fIguRES: there is no distinction between photographs and line drawings in numbering.All should be supplied in mono or colour, TIFF format, 300 dpi and CMYK. (pleasecontact the Production Editor with queries), with a scale bar where appropriate. letteringor numbers (Arabic) should be in the bottom left using uppercase Times Roman. Figurecaptions should be on a separate sheet.taBlES: can be printed in either portrait or landscape format. Authors should considerthis when preparing tables. Authors should ensure that tables fit the printed page size ina legible form.pagESIZE: The Gazette has a useable page size of 123 × 204 mm. This can be used inportrait or landscape mode but any supplied artwork should not be larger than this.mEaSuREmEntS: follow the metric system.chEcklIStS: follow the format of Baksh-Comeau, Fern Gaz. 16(1, 2): 11- 122.REfEREncES: follow the style of a recent issue of The Fern Gazette, e.g.:-HOOKER, W.J. 1864. species Filicum, 5. Dulau & Co., london.MORTON, C.V. 1947. The American species of Hymenophyllum, section
Sphaeroconium. Contr. U.s. Natl. Herb. 29(3): 139-201.sTEVENsON, D.W. & lOCONTE, H. 1996. Ordinal and familial relationships of
pteridophyte genera. In: CAMUs, J.M., GIBBY, M. & JOHNs, R.J. (Eds)Pteridology in perspective, pp. 435-467. Royal Botanic Gardens, Kew.
JouRnal aBBREVIatIonS: should follow Botanico Periodicum Huntianum &supplements.Alterations from the original text at proof stage will be charged for unless they are minorpoints of detail. A pdf will be provided free to the senior author.
268 FERN GAZ. 20(6). 2017

TAXONOMIC SURVEY OF OCCURRENCE, DIVERSITY ANDETHNOBOTANY OF PTERIDOPHYTES IN SOME PARTS OF
NASARAWA STATE, NIGERIA
G.F. AKOMOLAFE* & A. SULAIMON
Department of Botany, Federal University Lafia, PMB 146, Lafia, Nasarawa State,Nigeria
*Correspondence: [email protected], +2348068997606
Key words: Pneumatopteris afra, Nasarawa, Pteridophytes, Taxonomic survey
ABSTRACTThe study was carried out to investigate the taxonomic occurrence ofPteridophytes in Lafia and Doma Metropolis of Nasarawa State, Nigeria. A totalof 16 locations were sampled in Lafia and Doma respectively. These locationsinclude wetlands, swamps, forest shade and trees. From these locations, fernswere collected, pressed and identified. Their relative abundances in each locationwere calculated and their ethnobotanical uses by the local dwellers weredocumented using structured questionnaires. Ferns were observed only in 12locations in Lafia and nine locations in Doma. Seven species of ferns wereidentified namely, Pneumatopteris afra, Nephrolepis biserrata, N. undulata, N.cordifolia, Adiantum capillus-veneris, Adiantum sp. and Pityrogrammacalomelanos. Pneumatopteris afra was found to be the most widely distributedfern. Of the ferns observed, terrestrial ferns had the highest occurrence (85.7%)in Lafia with only 50% in Doma. Pneumatopteris afra was found to have thehighest relative frequency while Nephrolepis biserrata and N. undulata had thelowest relative frequency in Lafia and Doma. All these species have been reportedby the local dwellers to have ethnobotanical uses such as in the treatment of ulcer,fever, typhoid, chest pain, stomach pain and diabetes. Also, Pityrogrammacalomelanos had the highest percentage awareness by the local dwellers. It isconcluded that Lafia is more rich and diverse in fern species than Doma.Locations where ferns were absent are due to the long-term alterations in themicroclimatic conditions by various human activities in those places.
INTRODUCTIONIn most parts of Africa ferns are more diverse in the eastern than in the westernmountains (Moran & Smith, 2001). Ecological and physiological factors such as habitat,seasonality, elevation, soils, rainfall, cloud cover and temperature have been identifiedas the determinants for the diversity, richness and distribution of ferns in most parts ofthe world (Kornas, 1993).
Nigerian’s first biodiversity report stated that ferns and fern allies have been observedto grow in most ecological zones but different habitats (Akinsoji et al., 2016). Although,ferns have wide coverage in Nigeria, due to habitat change by factors such as unplannedurbanization, over-exploitation of minerals and farming, many species have beenreduced, endangered or are on the verge of extinction (Dixit, 2000).
Pteridophytes are often restricted to specific micro-habitats. However, as a result ofserious threats faced by these plants coupled with the fact that they are underutilized
FERN GAZ. 20(7): 269-279. 2018 269

and overlooked in this part of the world because of their relatively small size andcrytogamic nature, there is need for adequate survey and documentation of theiroccurrences, locations and economic values. Therefore, this research aimed to investigatethe occurrence, diversity and ethnobotany of ferns in Lafia and Doma metropolis,Nasarawa State, Nigeria.
MATERIALS AND METHODSThis study was carried out at Lafia and Doma Local Government areas of NasarawaState, Nigeria. Habitats assessed for the various diversities of pteridophytes include riverbanks, streams, forest (under shade), arboreal (top of trees), swamps and some other wetlands. The study was conducted towards the end of rainy season in 2016 and early dryseason of 2017. Plants specimens were collected using Random sampling method.Various fern species parts were collected which included leaflet, fronds and roots. Thesamples collected were all recorded in the field. Representative of all collected fernspecies or specimens, were also photographed. These specimens were later used foridentification and to provide permanent records for future use. The geographicalcoordinates of each sampling location were taken by using a GPS device.
Mature fern species were collected in each location and pressed, dried, poisoned thenmounted on standard herbarium sheets and classified according to standard procedures.Identifications were made with the aid of taxonomic literature (Oloyede & Odu, 2011).In the field proper care was taken in selecting the plant materials to enable correctidentification. Specimens were collected in a good condition, free of insect damage ordiseases. Voucher specimens are preserved in the Department of Botany Herbarium,Federal University, Lafia.
Ethnobotanical UsesA total of ninety (90) structured questionnaires were administered to local dwellers ofall the locations visited to gather information on the knowledge of the plants by thepeople, the uses of the plants and the part of the plant that had been used. The relativefrequency of occurrence of each fern species was determined to assess the distributionof the species.
Sørensen Similarity Coefficient was calculated to indicate their floristic similaritiesbetween the two towns (Lafia and Doma) as follows:
where Sc is the similarity coefficient, W is the species occurring in both communitiesunder consideration, a is the number of species occurred in Lafia and b is the number ofspecies occurred in Doma.
The relative abundance of the ferns at each site was determined according to Bongerset al. (1988) and Kayode (1999): less than five individuals as Rare; 5 to 10 as Occasional;11 to 30 as Frequent; 31 to 100 as Abundant and over 100 individuals as Very Abundant.
RESULTSFerns occurrence and distribution in Lafia A total of 16 locations were sampled in Lafia, of which ferns were observed in only 12
270 FERN GAZ. 20(7): 269-279. 2018
No of occurences of individual fern speciesNo of occurences of all fernsR F = x 100
2Wa + bSc = x 100
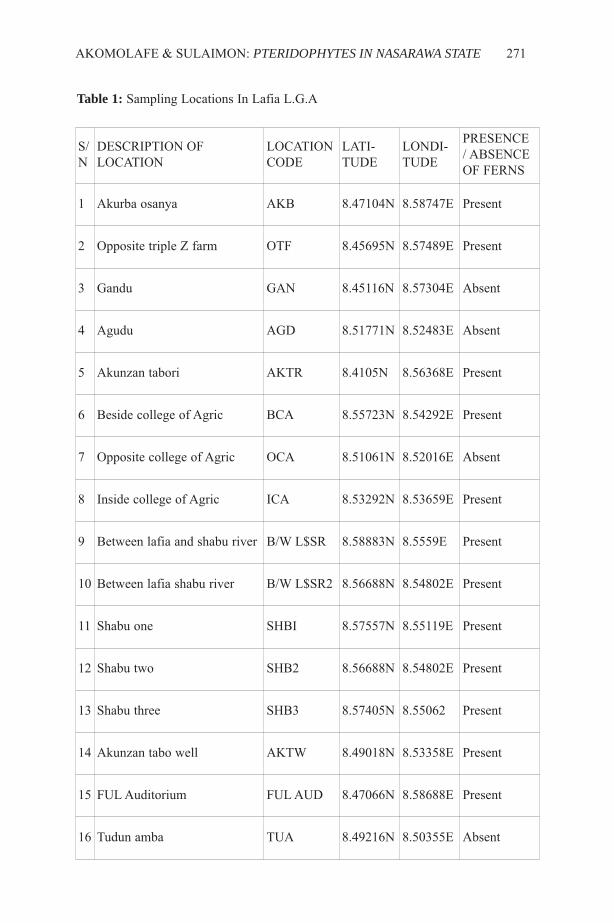
AKOMOLAFE & SULAIMON: PTERIDOPHYTES IN NASARAWA STATE 271
S/N
DESCRIPTION OFLOCATION
LOCATIONCODE
LATI-TUDE
LONDI-TUDE
PRESENCE/ ABSENCEOF FERNS
1 Akurba osanya AKB 8.47104N 8.58747E Present
2 Opposite triple Z farm OTF 8.45695N 8.57489E Present
3 Gandu GAN 8.45116N 8.57304E Absent
4 Agudu AGD 8.51771N 8.52483E Absent
5 Akunzan tabori AKTR 8.4105N 8.56368E Present
6 Beside college of Agric BCA 8.55723N 8.54292E Present
7 Opposite college of Agric OCA 8.51061N 8.52016E Absent
8 Inside college of Agric ICA 8.53292N 8.53659E Present
9 Between lafia and shabu river B/W L$SR 8.58883N 8.5559E Present
10 Between lafia shabu river B/W L$SR2 8.56688N 8.54802E Present
11 Shabu one SHBI 8.57557N 8.55119E Present
12 Shabu two SHB2 8.56688N 8.54802E Present
13 Shabu three SHB3 8.57405N 8.55062 Present
14 Akunzan tabo well AKTW 8.49018N 8.53358E Present
15 FUL Auditorium FUL AUD 8.47066N 8.58688E Present
16 Tudun amba TUA 8.49216N 8.50355E Absent
Table 1: Sampling Locations In Lafia L.G.A

Table 2: The locations occupied by the ferns and the respective abundance status in Lafia
272 FERN G
AZ. 20(7): 269-279. 2018
S/N
Species Location / Abundance status
AKB OTF GA AGD AKTR BCA OCA ICA B/WL&SR B/WL&SR2 SHB1 SHB2 SHB3 AKTW FULAUD TUA
1 Adiantum sp. X X X X X X X X X X X X X √ A X X
2 Adiantumcapillus-veneris X X X X X X X X X X X X X √ A X X
3 Pneumatopterisafra √ VA √ F X X X √ VA X X √ VA √ F X √ A √ A X X X
4 Nephrolepisbisserata X X X X X X X X X X X X √ R X X X
5 Nephrolepiscordifolia X X X X X X X X X X X X X X √ F X
6 Nephrolepisundulata X X X X √ F X X √ VA X √ A √ O X √ O X X X
7 Pityrogrammacalomelanos X X X X √ VA X X X X X X X X √ A X X
KEY: √ means present, X means absentA – abundant, VA – very abundant, F – frequent, O – occasional, R – rare

AKOMOLAFE & SULAIMON: PTERIDOPHYTES IN NASARAWA STATE 273
Figure 1: Relative frequency of Fern species in Lafia town
Figure 2: Relative frequency of Fern species in Doma town
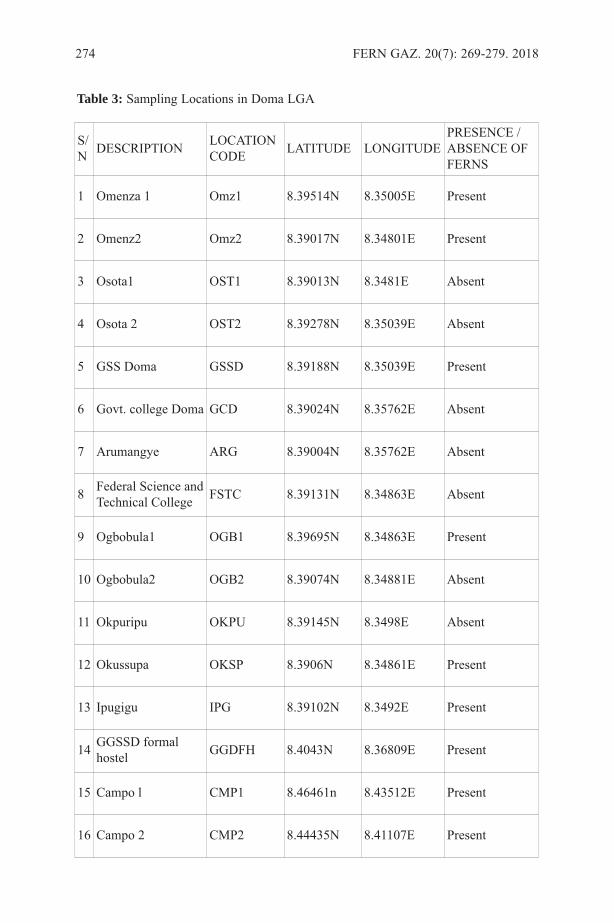
274 FERN GAZ. 20(7): 269-279. 2018
S/N DESCRIPTION LOCATION
CODE LATITUDE LONGITUDEPRESENCE /ABSENCE OFFERNS
1 Omenza 1 Omz1 8.39514N 8.35005E Present
2 Omenz2 Omz2 8.39017N 8.34801E Present
3 Osota1 OST1 8.39013N 8.3481E Absent
4 Osota 2 OST2 8.39278N 8.35039E Absent
5 GSS Doma GSSD 8.39188N 8.35039E Present
6 Govt. college Doma GCD 8.39024N 8.35762E Absent
7 Arumangye ARG 8.39004N 8.35762E Absent
8 Federal Science andTechnical College FSTC 8.39131N 8.34863E Absent
9 Ogbobula1 OGB1 8.39695N 8.34863E Present
10 Ogbobula2 OGB2 8.39074N 8.34881E Absent
11 Okpuripu OKPU 8.39145N 8.3498E Absent
12 Okussupa OKSP 8.3906N 8.34861E Present
13 Ipugigu IPG 8.39102N 8.3492E Present
14 GGSSD formalhostel GGDFH 8.4043N 8.36809E Present
15 Campo l CMP1 8.46461n 8.43512E Present
16 Campo 2 CMP2 8.44435N 8.41107E Present
Table 3: Sampling Locations in Doma LGA

AK
OM
OLA
FE & SU
LAIM
ON
:PTERIDOPH
YTES IN NASARAW
A STATE275
S/N Species Location / Abundance status
OMZ1 OMZ2 OST1 OST2 GSSD GCD ARG FSTC OGB1 OGB2 OKPU OKSP IPG GGFH CMP1 CMP2
1 Pneumatopterisafra √ VA √ A X X √ F X X X √ A X X X X X √ VA √ A
2 Nephrolepisundulata X X X X X X X X X X X √ VA √ VA √ O X X
Table 4: The locations occupied by the ferns and the respective abundance status in Doma
KEY: √ means present, X means absentA – abundant, VA – very abundant, F – frequent, O – occasional, R - rare

276 FERN G
AZ. 20(7): 269-279. 2018
S/N SPECIES % AWARENESS ETHNOBOTANICAL USES PARTS USED MODE OF PREPARATION
1 Pneumatopterisafra
90
Typhoid, fever, headache, ulcer Leaflets, fronds boiling, piles, stomach pain roots & fronds Drying
strengthening of bones whole plants drying & boilinghealing of wounds caused by guinea worm root Drying
2 Pityrogrammacalomelanos
100
enhancement of male sexual cells root boilingtreatment of chest pain, leaflets air dryinghealing of stomach pain, fever fronds boilingtreating shortage of blood leaflets boilingreducing prolonged labour whole plants boilingbreast cancer root boiling
3 Nephrolepisundulata
30Snake bite, convulsion, high blood pressure,poisoning, goitre, bone fracture, sickle cell anemia Whole plants drying, freshly pounded, boiling
diabetes roots boiling
4 Adiantumcapillus-veneris
40Convulsion, diabetes, headache, pile, Gonorrhea,scorpion bite, dysentery whole parts, roots &fronds air drying, freshly pounded, boiling
Measles leaflets only Boiling
5 Adiantum sp 33.3fibroid, shortage of blood, diabetes, abnormalmenstruation, stomach pain whole parts Boiling &air drying
piles, back pain, waist pain & joint cases leaf Boiling &air drying
6 Nephrolepiscordifolia
20sickler cases, high blood pressure, bone fracture,goiter, snake/scorpion bite, skin rashes &diabetes whole parts &roots Boiling, freshly pounded, air drying
headache &rheumatism leaf air drying
7 Nephrolepisbiserrata
30 Eyesight issues, bone fracture, goitre, high bloodpressure, snake / scorpion bite &diabetes Whole parts &roots Boiling, freshly pounded &drying
Table 5: Ethhnobotanical uses of the fern species in Lafia

AK
OM
OLA
FE & SU
LAIM
ON
:PTERIDOPH
YTES IN NASARAW
A STATE277
S/N SPECIES % AWARENESS ETHNOBOTANICAL USES PARTS USED MODE OF PREPARATION
1 Pneumatopteris afra 50
increases male sexual stamina Leaf, stem &whole parts Boiling
rheumatism, stmach pains,tuberculosis, fever Leaf, stem &whole parts Boiling
2 Nephrolepis undulata 50 Healing of wounds, asthma,pneumonia, cough &ulcer Leaf, whole parts Drying, boiling &freshly
pounded
Table 6: Ethhnobotanical uses of the fern species in Doma

locations (Table 1). Seven species of ferns were identified from the sampled locations inLafia: Pneumatopteris afra (Christ) Holttum, Nephrolepis undulata (Afzel.) J.Sm.,Nephrolepis cordifolia (L.) K.Presl, Nephrolepis biserrata (Sw.) Schott, Adiantum sp,Adiantum capillus-veneris L. and Pityrograma calomelanos (L.) Link (Table 2).Pneumatopteris afra was found to be the most widely distributed fern in Lafia where itwas found in seven locations. Of the ferns observed, terrestrial ferns were most frequentwith 85.7% occurrence while epiphytes were least frequent at 14.3% . Pneumatopterisafra was found to have the highest relative frequency at 65% while Nephrolepis biserrataand N. undulata had the lowest (5%) (Figure 1). The similarity coefficient between thetwo towns is 22.2%.
Ferns occurrence and distribution in DomaIn Doma, from a total of 16 locations sampled ferns were observed in only nine locations(Table 3). However, in Doma, only two species of ferns were recorded, one terrestrialand the other epiphytic, namely Pneumatopteris afra and Nephrolepis undulata (Table4). Pneumatopteris afra was found to be the most widespread fern in Doma where it wasfound in six locations. The relative frequency of Pneumatopteris afra was greater(66.67%) than that of N. undulata (33.34%) (Figure 2).
Ethnobotanical Uses of Ferns in LafiaThe results of the survey of ethnobotanical uses and awareness of the ferns in Lafia arepresented in Table 5. Every species was discovered to have ethnobotanical uses amongthe local population. Pneumatopteris afra had 90% percentage awareness and it was usedfor treating ulcer, fever, typhoid and piles. Pityrogramma calomelanos had the highestpercentage awareness (100%) and was used for enhancement of male sexual performanceand treating chest pain. The lowest percentage awareness (20%) was recorded forNephrolepis cordifolia which was used for treating high blood pressure and skin rashes.
Ethnobotanical Uses of Ferns in DomaIn Doma, Pneumatopteris afra had 50% percentage awareness and was used for treatingrheumatism, stomach pains and tuberculosis (Table 6). Nephrolepis undulata also had50% percentage awareness and was used for treating wounds, asthma and pneumonia.
DISCUSSSION AND CONCLUSIONThe occurrence of ferns in the two towns studied had only two species in common. Asreported by Jones et al., (2011), some fern species are highly flexible and are found innearly all habitats and also have various life forms. High species occurrence in areaswith suitable environmental conditions showed that such species have adapted to theenvironment (Richard et al., 2000). Consequently, the relatively high number of fernsspecies found in Lafia compared with Doma might be the result from more favourablemicroclimatic conditions than in Doma, where conditions have been affected by humanactivities such as deforestation and farming. This is supported by Sumesh et al. (2014)who stated that human activities usually increase pressures on plants.
The occurrence of some species in Lafia such as Pneumatopteris afra, Nephrolepisundulata and Adiantum capillus-veneris with high relative frequencies suggests that thesespecies will persist in this area if the environmental conditions remain favourable. Butspecies like Adiantum sp., Nephrolepis cordifolia, Nephrolepis biserrata andPityrogramma calomelanos with low relative frequencies are more likely to become
278 FERN GAZ. 20(7): 269-279. 2018

threatened in the future. Also in Doma town, only two ferns were recorded, andPneumatopteris afra was more frequent than Nephrolepis undulata. The paucity andoften total absence of ferns in some locations sampled in Lafia (four sites) and Doma(seven sites) is a clear indication of the accumulated long-term anthropogenic activitiesin the areas resulting in loss of ferns species at these sites.
It is concluded that, in comparison, Lafia has a more rich and diverse ferns flora thanDoma, and that despite the presence of relatively few species, all have been used by thelocal dwellers for various economic purposes. Therefore, conservative measures shouldbe put in place by the Government to avoid future extinction of these ferns.
REFERENCESAKINSOJI, A., AGBOOLA, O.O., ADEONIPEKUN, P.A., OYEBANJI, O.O.,
ADENIYI, T.A & AJIBODE, M.O. 2016. Distribution of pteridophytes in parts ofLagos and Osun State. Ife J. of Sci. 18(2): 447-453
BONGERS, F., POPMA, J., MEAVE DEL CASTILLO, J. & CARABIAS, J. 1988.Structure and floristic composition of the lowland rainforest of Los Tuxtlos, Mexico.Vegetatio. 74: 55-80.
DIXIT, R.D. 2000. Conspectus of pteridophytic diversity in India. Fern J. 17: 77-91.JONES, N.A., ROSS H., LYNAM, T. PEREZ P. & LEITCH, A. 2011. Mental models:
an interdisciplinary synthesis of theory and methods. Eco. & Soc. 16(1): 46KAYODE, J. 1999. Phytosociological investigation of compositae weeds in abandoned
farmlands in Ekiti State, Nigeria. Compositae News. 34: 62-68.KORNAS, J. 1993. The significance of historical factors and ecological preferences in
the distribution of bibliography of African Pteridophytes. J. of Biogeo. 20: 281-286. MORAN, A.C & SMITH, A.R. 2001. Phytogeographical relationship between
Neotropical and African –Madagascar pteridophytes. Brittonia 53: 304-351. OLOYEDE, F.A. & ODU, E.A. 2011. Taxonomic Evaluation of Homosporous
Leptosporangiate ferns in Southwestern Nigeria. J. of Curr. Res. 2(11): 009-017RICHARD, A. F., DEWAR, R. E., SCHWARTZ, M. & RATSIRARSON, J. 2000. Mass
change, environmental variability and female fertility in wild Propithecus verreauxi.J. of Human Evol. 39:381-391.
SUMESH, N.D., SUBASH, M.D.C. & RAMACHANDRA, T.V. 2014. Conservation ofPteridophytes to maintain vital link between lower and higher group of plants. Energyand Wetlands research group, center for Ecological Sciences, Indian Institute ofScience, Bangalore, pp. 560012
AKOMOLAFE & SULAIMON: PTERIDOPHYTES IN NASARAWA STATE 279

ERRATUM
The correct title and citation of this article should be:Gensel, P.G. 2017. Silurian-Devonian origins of ferns and lycophytes - what we know,what we need to find out. Fern Gaz. 20(6): 217-242.
280 FERN GAZ. 20(7). 2018

PHYTOSOCIOLOGICAL NOTES ON THE FERN-MEADOWVEGETATION OF MID-WEST SCOTLAND AND THE
NETHERLANDS
Rense HavemanCentral Government Estate Agency, Ministry of the Interior and Kingdom Relations,
Expert Centre Technique/Nature, P.O. Box 47, 6700 AA Wageningen, the Netherlands.Email: [email protected]
Keywords:Argyll, Melampyro-Holcetea mollis, Dryopteris, Oreopteris, climate,Luzulo luzuloides-Thelypteridetum limbospermae, phytosociology
ABSTRACTOn the basis of five phytosociological relevés, the fern-meadow vegetation withOreopteris limbosperma and Dryopteris cambrensis from the region around Oban,Argyll, West Scotland is described and compared with similar communities in theNetherlands and West Germany. It is concluded that these fern-meadows can beassigned to the Luzulo luzuloides-Thelypteridetum limbospermae, which inCentral and continental West Europe is restricted to forest edges. In West Scotlandthis community is also common in open landscapes, due to the hyperatlanticclimate. In comparison with the fern communities in the Netherlands andGermany, the Scottish examples are very rich in fern species, illustrating theoptimal development of the community in West Argyll. Possible futuredevelopment of this community in the Netherlands are discussed in the light ofthe increase in records of Oreopteris limbosperma and the ecological requirementsof Dryopteris cambrensis.
INTRODUCTIONFor a continental phytosociologist, one of the striking features of the vegetation of thecoastal lowlands of West-Argyll is the omnipresence of ferns in open landscapes (Figure1). Well-known is the invasive character of Pteridium aquilinum (L.) Kuhn in extensivelymanaged grasslands all over the British Isles (Rodwell et al., 1992), but many other fernspecies are found in the dense tall-forb vegetation on slopes in the hyperatlantic lowlandaround Oban. After Pteridium, probably Blechnum spicant (L.) Sm. is the most frequentspecies, but also Dryopteris cambrensis (Fraser-Jenk.) Beitel & W.R.Buck can be foundon virtually every slope and this species is much more abundant than Blechnum. Otherspecies which can be found often are, in no particular order, Dryopteris filix-mas (L.)Schott, D. borreri (Newm.) Newm. ex Oberh. & Tavel, D. dilatata (Hoffm.) A.Gray,D.carthusiana (Vill.) H.P.Fuchs, Athyrium filix-femina (L.) Roth, Phegopteris connectilis(Michx.) Watt, and Oreopteris limbosperma Holub.
In West-Europe, fern-dominated forest edge communities from lowland, hills andlower mountain areas were recently placed in a newly described alliance, the Holcomollis-Athyrion felicis-feminae (Royer et al.) Haveman & Weeda (class Melampyro-Holcetea mollis Passarge ex Weeda & Haveman; Weeda & Haveman, 2017). The firstauthor who drew the attention to these fern-rich forest edge communities was Wittig(2000), who described Oreopteris-rich forest-edges from the German lower mountainranges in the Rothhaargebirge, Taunus, and Schwarzwald as Luzulo luzuloides-Thelypteridetum limbospermae Wittig. Apart from Oreopteris limbosperma (syn.:
FERN GAZ. 20(7): 281-291. 2018 281

Thelypteris limbosperma (All.) H.P.Fuchs) also Athyrium filix-femina, Phegopterisconnectilis, Blechnum spicant, Gynmocarpium dryopteris (L.) Newman, Dryopteriscarthusiana, D. dilatata, and D. filix-mas are fern species mentioned for this vegetationtype. This Oreopteris-community is not strictly oreophytic (growing in the mountainbelt): Bremer (2016) and Weeda & Haveman (2017) recorded the Luzulo-Thelypteridetum also from the Netherlands, i.e. from the lowlands of Northwest-Europe.Rodwell et al. (1992) described a similar community as Thelyptris limbosperma-Blechnum spicant community (U19) from moist base-poor peaty soils on steep, shelteredbanks at low to moderate altitudes through the wetter west and north of Britain.Remarkably enough, other fern species are rare in this community: only Polypodiumvulgare L. and Athyrium distentifolium Tausch ex Opiz are mentioned in the tablepublished by these authors. Weeda & Haveman (2017) included this British communityin the Luzulo-Thelypteridetum.
During a three-weeks summer holiday in July 2017, I was able to study some of thefern-meadow communities in the wider surroundings of Oban, West-Argyll, Scotland. Iwas especially interested in the communities with the alleged mountain-dwellingOreopteris limbosperma and Dryopteris cambrensis, because the latter is very rare inthe Netherlands. From the species composition, it was immediately clear that the Scottishfern-meadow community bears a very close resemblance to the Luzulo-Thelypteridetum,but also that it had characteristic features of its own. In this paper, I will describe thiscommunity on the basis of a few phytosociological relevés. It will be compared with thecontinental fern communities of the aforementioned Holco-Athyrion and more inparticular the Luzulo-Thelypteridetum, in order to gain a better understanding of thesecommunities, both in hyperatlantic Scotland and the subatlantic Netherlands.
282 FERN GAZ. 20(7): 281-291. 2018
Figure 1. Fern-meadow on a lowland hillside in West-Argyll near Loch Fyne (photo:R. Haveman)

METHODSOn July 6th and 7th 2017, I made five phytosociological relevés of fern-rich forest-edgesin Ardcastle Forest at the shore of Loch Fyne and the area of Glencruitten House justoutside Oban, according to the methods of the French-Swiss school (Mueller-Dombois& Ellenberg, 1974; Westhoff & Van Der Maarel, 1978). In floristically homogeneousparts of the vegetation of 8-20 m2, all species were recorded, and their abundance wasestimated using the 9-level extended scale of Braun-Blanquet (Westhoff & Van DerMaarel, 1978) in every single layer (tree, shrub, field, and moss layer). The relevés weredigitised and stored in TurboVeg (Hennekens & Schaminée, 2001) in the personaldatabase of the author. After export from TurboVeg, the relevés were imported in JUICE(Tichý 2002) for further analysis. In JUICE, all species were ordered according to theirphytosociological position according to Weeda & Haveman (2017). Both a full and a(heavily abbreviated) synoptic table (in frequency percentages) of constant species(frequency > 40%) were exported, and the synoptic table compared with the synoptictables published by Weeda & Haveman (2017) and Wittig (2000) by arranging them ina longitudinal order (Scotland-the Netherlands-Germany). In the full table, allpteridophytes are included, as well as all species occurring in more than one relevé, orin one relevé with a cover ≥ 5%. In the synoptic tables, only the species with a frequency> 10% are included in at least one column.
RESULTSIn the Scottish relevés (Table 1), 11 fern species were recorded in total, and one fernhybrid. The most frequent fern species in the table is Blechnum spicant (in all fiverelevés), followed by Oreopteris limbosperma, and Dryopteris cambrensis (both in fourrelevés). In all relevés, Dryopteris species have an abundance prevalence over the otherfern species. Other species recorded in at least four out of five relevés are Luzula sylvatica(Huds.) Gaudin, Holcus mollis L., Hylocomium splendens (Hedw.) Schimp.,Rhytidiadelphus squarrosus (Hedw.) Warnst. and Polytrichum formosum Hedw.. In allrelevés, more than one fern species was found, and relevé 1 in Table 1 is especially richin ferns: in the vegetation sample of 20 m2 9 pteridophytes were recorded.
There are clear differences between the Scottish fern-communities with Oreopterislimbosperma and Dryopteris cambrensis (Table 2, column 1), and the Luzulo-Thelypteridetum in the Netherlands (Table 2, column 2) and Germany (Table 2, column3). Especially striking is the richness in ferns in the Scottish community: on average, 6.2fern species are recorded in the Scottish relevés, a number that declines strongly whengoing to the suboceanic regions in the Netherlands and Germany (viz. 3.9 and 3.4respectively). Also their absolute number in the Scottish dataset is higher than in theother two (12, vs. 7 and 9 respectively), although the number of Scottish relevés isevidently lower than in the other two groups. Dryopteris cambrensis, D. borreri andPteridium aquilinum are exclusively or, in case of the latter, mainly recorded in theScottish relevés, although it cannot be ruled out that Dryopteris affinis Fraser-Jenk.,mentioned in one relevé from Germany, is in fact D. borreri. Also Blechnum spicant andDryopteris filix-mas have a clear optimum in the Scottish relevés, and the same holdstrue for Holcus mollis, Luzula sylvatica, Epilobium montanum L., Betula pendula Roth(juveniles), and the mosses Hylocomium splendens and Rhytidiadelphus squarrosus.Typical for the Dutch relevés are Molinia caerulea (L.) Moench and Quercus robur L.(juveniles), whereas the German relevés are differentiated by Luzula luzuloides (Lam.)Dandy & Wilmott. However, the three regional groups of relevés have many taxa in
HAVEMAN: FERN-MEADOW VEGETATION 283
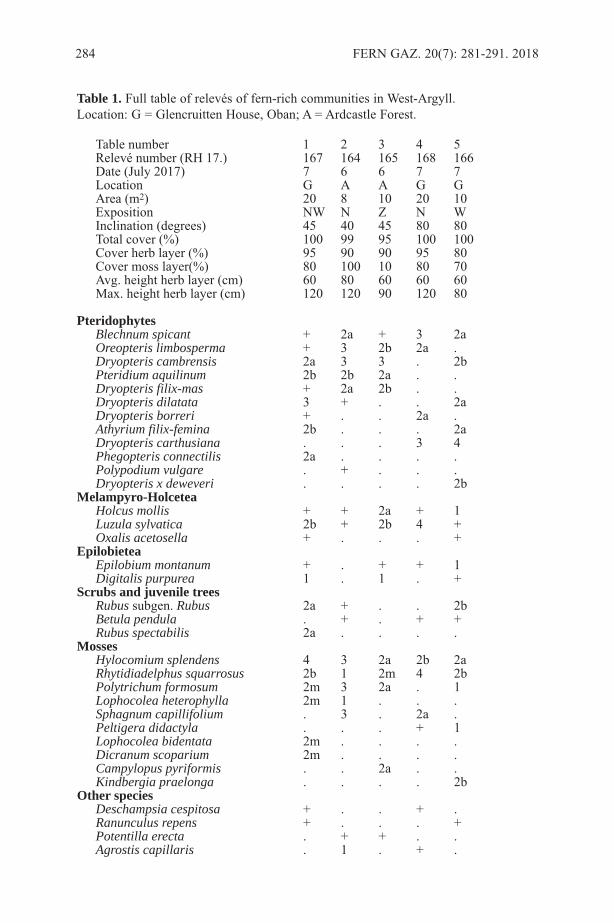
284 FERN GAZ. 20(7): 281-291. 2018
Table number 1 2 3 4 5Relevé number (RH 17.) 167 164 165 168 166Date (July 2017) 7 6 6 7 7Location G A A G GArea (m2) 20 8 10 20 10Exposition NW N Z N WInclination (degrees) 45 40 45 80 80Total cover (%) 100 99 95 100 100Cover herb layer (%) 95 90 90 95 80Cover moss layer(%) 80 100 10 80 70Avg. height herb layer (cm) 60 80 60 60 60Max. height herb layer (cm) 120 120 90 120 80
PteridophytesBlechnum spicant + 2a + 3 2aOreopteris limbosperma + 3 2b 2a .Dryopteris cambrensis 2a 3 3 . 2bPteridium aquilinum 2b 2b 2a . .Dryopteris filix-mas + 2a 2b . .Dryopteris dilatata 3 + . . 2aDryopteris borreri + . . 2a .Athyrium filix-femina 2b . . . 2aDryopteris carthusiana . . . 3 4Phegopteris connectilis 2a . . . .Polypodium vulgare . + . . .Dryopteris x deweveri . . . . 2b
Melampyro-HolceteaHolcus mollis + + 2a + 1Luzula sylvatica 2b + 2b 4 +Oxalis acetosella + . . . +
EpilobieteaEpilobium montanum + . + + 1Digitalis purpurea 1 . 1 . +
Scrubs and juvenile treesRubus subgen. Rubus 2a + . . 2bBetula pendula . + . + +Rubus spectabilis 2a . . . .
MossesHylocomium splendens 4 3 2a 2b 2aRhytidiadelphus squarrosus 2b 1 2m 4 2bPolytrichum formosum 2m 3 2a . 1Lophocolea heterophylla 2m 1 . . .Sphagnum capillifolium . 3 . 2a .Peltigera didactyla . . . + 1Lophocolea bidentata 2m . . . .Dicranum scoparium 2m . . . .Campylopus pyriformis . . 2a . .Kindbergia praelonga . . . . 2b
Other speciesDeschampsia cespitosa + . . + .Ranunculus repens + . . . +Potentilla erecta . + + . .Agrostis capillaris . 1 . + .
Table 1. Full table of relevés of fern-rich communities in West-Argyll.Location: G = Glencruitten House, Oban; A = Ardcastle Forest.

HAVEMAN: FERN-MEADOW VEGETATION 285
Column 1 2 3Country Sc NL GeNumber of relevés 5 11 21
PteridophytesDryopteris cambrensis 80 . .Dryopteris borreri 40 . .Polypodium vulgare 20 . .Dryopteris x deweveri 20 . .Pteridium aquilinum 60 9 .Blechnum spicant 100 64 24Oreopteris limbosperma 80 100 100Dryopteris dilatata 60 55 57Athyrium filix-femina 40 64 76Dryopteris carthusiana 40 64 14Phegopteris connectilis 20 36 33Dryopteris filix-mas 60 . 14Gymnocarpium dryopteris . . 14Dryopteris affinis . . 5
Hylocomium splendens 100 . .Pseudoscleropodium purum 20 55 .Potentilla erecta 40 45 .Holcus mollis 100 27 43Atrichum undulatum 20 45 10Oxalis acetosella 40 9 43Digitalis purpurea 60 9 43Rhytidiadelphus squarrosus 100 18 14Polytrichum formosum 80 64 43Rubus species 60 73 52Agrostis capillaris 40 45 67Juncus effusus 20 45 33Luzula sylvatica 100 . 52Epilobium montanum 80 . 24Betula pendula 60 . 5Rubus idaeus 20 . 71Molinia caerulea . 73 .Quercus robur . 55 .Deschampsia flexuosa . 27 51Dicranella heteromalla . 45 5Galium saxatile . 18 48Pellia epiphylla . 55 14Luzula luzuloides . . 81
Table 2. Synoptic table of Oreopteris limbosperma communities in Scotland (column1, source: Table 1), the Netherlands (column 2, source: Weeda & Haveman 2017), andGermany (column 3, source: Wittig 2000).

common, like Dryopteris dilatata, Athyrium filix-femina, Phegopteris connectilis,Atrichum undulatum (Hedw.) P.Beauv., Polytrichum formosum, Rubus L. subgen. Rubus,Agrostis capillaris L., and Juncus effusus L. Several other species are common to tworegions, whereas they are lacking from the third. Potentilla erecta (L.) Raeusch. is anexample, which was found in both the Scottish and the Dutch communities, and not inthe German.
DISCUSSIONPhytosociological AssignmentThe here described Scottish fern-meadows are at least for the greater part to be assignedto the Luzulo luzuloides-Thelypteridetum limbospermae, on the basis of the presence ofthe characteristic species Oreopteris limbosperma as well as the constant species, withmany pteridophytes and other species which are found in both the Scottish relevés aswell as in the German and/or the Dutch plots. The last relevé, without Oreopteris,probably represents a fragmentary example of this association. As was shown though,the Scottish fern-meadows have their own characteristics, with several species not foundin the suboceanic forms of the association. Especially typical is Dryopteris cambrensis.According to Fraser-Jenkins (2007), this species has its distribution centre in oceanicEurope. In the British Isles it is common in Scotland, contrary to its name less so inWales, and rather rare elsewhere (Page 1997; Trewren 2014). Considering this, theScottish fern-meadows could probably be described as a separate subassociation (Luzuloluzuloides-Thelypteridetum dryopteridetosum cambrensis prov.) but, at this stage, aformal description of such a subassociation is rejected, since the available dataset is toosmall for a proper assessment.
286 FERN GAZ. 20(7): 281-291. 2018
Figure 2. Abundant fern growth in a forest edge, Oban, Argyll. Apart from Pteridiumaquilinum, the vegetation is formed mainly by Dryopteris species and Oreopterislimbosperma (photo: R. Haveman)

EcologyIn the German lower mountain belt, the Luzulo-Thelypteridetum is almost strictlyconfined to forest edges (Wittig, 2000), and the same applies to the situation in theNetherlands (Weeda & Haveman, 2017). In Scotland, the community has a much broaderecological amplitude. Although it is frequently found in forest edges (relevés 3-5 in Table1, Figure 2), it also develops frequently at forest clearings on suitable soils (relevés 1and 2 in Table 1, Figure 3). Typical species of woodland clearings (grouped under theheading of Epilobietea and Scrubs and juvenile trees in Table 1) are not confined to thelatter environment: they are equally present in the relevés from forest edges, and also inthe relevés from Germany (Table 2 column 3, less so in the Dutch relevés).Unfortunately, no relevés were made in the stands on the third environment where thecommunity is found, viz. abandoned and extensively managed grasslands (Figure 1).However, several short checks of the stands in this situation made clear that the speciescomposition of the vegetation here is very similar to the community which develops inforest edges or at forest clearings.
The abundant occurrence of the combination of Oreopteris limbosperma, Blechnumspicant, Athyrium filix-femina, and several Dryopteris species outside the buffered forestmicro-climate indicate the high air-humidity and annual rainfall in this part of the UK,where water availability is never a problem (see also Knaben 1950, who drew the sameconclusion after study of the fern-communities of West-Norway). According to theclimate maps of the Met Office (www.metoffice.gov.uk), the region directly around Obanhas an average annual rainfall of 1,700-2,200 mm, which is 2.5 to 3 times the amount inthe Netherlands. The German lower mountains receive more rain than the Netherlands,but not as much as the Oban region: the higher regions of the Rothaargebirge e.g., havean average annual rainfall of 1,000 mm (www.wetter.de). Where the Luzulo-
HAVEMAN: FERN-MEADOW VEGETATION 287
Figure 3.A species rich fern community in a forest clearing in Glencruitten House forest,Oban, Argyll (photo R. Haveman)

Thelypteridetum grows in zonation with Pteridium-dominated communities, the first isconfined to the steep banks, or the shelter of woodlands (Figure 4). Probably this iscaused by difference in water demand, which is higher in Oreopteris than in Pteridium(Weeda & Haveman, 2017). In this respect, a striking description of the edaphic demandsof Oreopteris limbosperma is given by Page (1997): "... niches where trickling acidicpeaty water provides the most constant surface run-off or subterranean seepage, creatingcool and constantly moist but well-aerated edaphic conditions ..."
In West-Argyll, the Luzulo-Thelypteridetum can form complexes with different typesof grasslands. On the least fertile situations, the grasslands can be assigned to the classNardetea Rivas Goday et Borja Carbonell. On soils with a somewhat better availabilityof nutrients, the main grassland type that is found in complex with the fern-meadowcommunity is the Lolio-Cynosuretum Tüxen (alliance Cynosurion cristati Tüxen, classMolinio-Arrhenatheretea Tüxen). Usually, both Nardion and Cynosurion grasslands aregrazed by cattle or sheep. Where the grassland is mown, especially at the bottom ofvalleys, the grassland vegetation resembles the Crepido-Juncetum acutiflori Oberdorfer,a grassland community characteristic for peaty soils with laterally water movement andseepage. It is remarkable that this last grassland type is also mentioned by Bremer (2016)when he discusses the landscape in which the Luzulo-Thelypteridetum is found.
In The vegetation of Scotland, the classic book edited by Burnett (1964), Oreopterislimbosperma is mentioned only in the context of sub-alpine scrubs and fern-meadows(p. 162-163, and p. 516). Only one remark in the chapter on the history and regionalpatterns in Scottish vegetation (p. 574) associates the species with burning on north-exposed slopes in the Northern Highlands. The occurrence of fern-dominated forest edges
288 FERN GAZ. 20(7): 281-291. 2018
Figure 4.Abandoned hillside meadow with fern encroachment. Large areas are coveredby Pteridium aquilinum (in the background and in the lower right corner). Steep banksare invaded by Oreopteris limbosperma and Dryopteris species (lower left corner, andthe linear structure in the centre of the photo). Oban, Argyll (photo: R. Haveman)
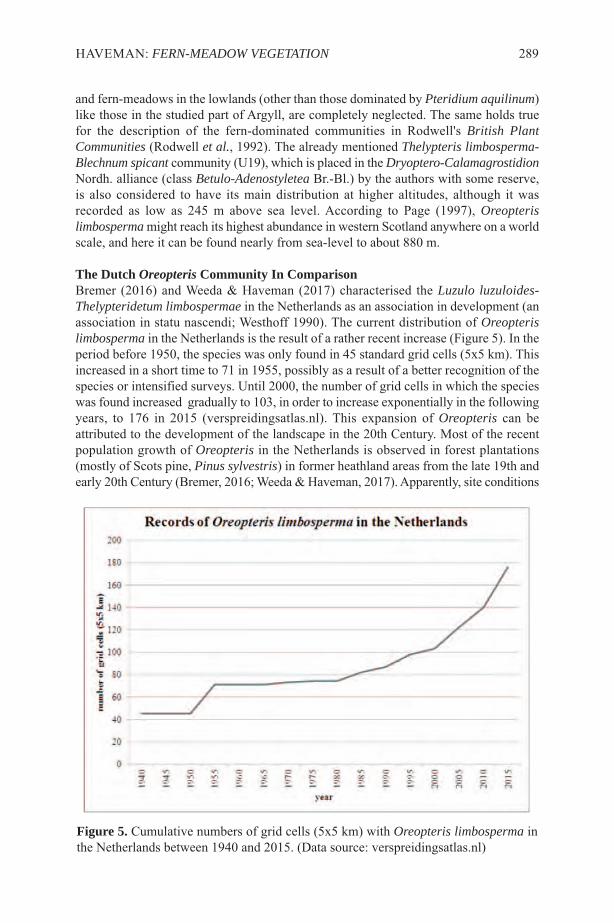
and fern-meadows in the lowlands (other than those dominated by Pteridium aquilinum)like those in the studied part of Argyll, are completely neglected. The same holds truefor the description of the fern-dominated communities in Rodwell's British PlantCommunities (Rodwell et al., 1992). The already mentioned Thelypteris limbosperma-Blechnum spicant community (U19), which is placed in the Dryoptero-CalamagrostidionNordh. alliance (class Betulo-Adenostyletea Br.-Bl.) by the authors with some reserve,is also considered to have its main distribution at higher altitudes, although it wasrecorded as low as 245 m above sea level. According to Page (1997), Oreopterislimbosperma might reach its highest abundance in western Scotland anywhere on a worldscale, and here it can be found nearly from sea-level to about 880 m.
The Dutch Oreopteris Community In ComparisonBremer (2016) and Weeda & Haveman (2017) characterised the Luzulo luzuloides-Thelypteridetum limbospermae in the Netherlands as an association in development (anassociation in statu nascendi; Westhoff 1990). The current distribution of Oreopterislimbosperma in the Netherlands is the result of a rather recent increase (Figure 5). In theperiod before 1950, the species was only found in 45 standard grid cells (5x5 km). Thisincreased in a short time to 71 in 1955, possibly as a result of a better recognition of thespecies or intensified surveys. Until 2000, the number of grid cells in which the specieswas found increased gradually to 103, in order to increase exponentially in the followingyears, to 176 in 2015 (verspreidingsatlas.nl). This expansion of Oreopteris can beattributed to the development of the landscape in the 20th Century. Most of the recentpopulation growth of Oreopteris in the Netherlands is observed in forest plantations(mostly of Scots pine, Pinus sylvestris) in former heathland areas from the late 19th andearly 20th Century (Bremer, 2016; Weeda & Haveman, 2017). Apparently, site conditions
HAVEMAN: FERN-MEADOW VEGETATION 289
Figure 5. Cumulative numbers of grid cells (5x5 km) with Oreopteris limbosperma inthe Netherlands between 1940 and 2015. (Data source: verspreidingsatlas.nl)

are developing into a favourable state for the establishment of Oreopteris in theselandscapes after about 100 years, especially in ditch sides in these plantations. In mostsites where the species occurs it is only present in very small populations, sometimesonly consisting of one single plant. This is in sharp contrast with the situation in WestScotland, where the species usually forms dense stands, and where a mature ferncommunity is developed.
The difference between the mature Scottish fern-meadows and the juvenile DutchOreopteris community is expressed in particular in the average number of fern species:6.2 in the Scottish relevés, and only 3.9 in the Dutch ones. Remarkably, in the only relevéthat was published by Bremer (2016) which can compete with the Scottish one in thisrespect (relevé 2003.20 in the table published by Bremer), Dryopteris borreri wasrecorded, the only observation of a species of the Dryopteris affinis complex in theassociation in the Netherlands.
Considering all available data, including vegetation structure, fern species numbers,as well as the ominipresence of the described fern-meadow vegetation, the Luzulo-Thelypteridetum may be optimally developed in the lowlands and low mountain areasof West Scotland. In the Central European lower mountain regions, O. limbosperma canform dense stands locally, but the number of fern species drops notably in comparisonwith the fern species numbers in the Scottish fern-meadows. In the Netherlands, thedistribution area of Oreopteris is still developing, and it is only rarely found in a densefern vegetation rich in pteridophytes.
The major absent in the Dutch relevés published by Bremer (2016) and Weeda &Haveman (2017) is Dryopteris cambrensis, which is so prominently present in theScottish relevés. This species was discovered only recently in the Netherlands, viz. in2010 by Koos Ballintijn in the Purmerbos just north of Amsterdam. The species isrecorded in eight grid cells now, although their identity is not yet verified in all cases(oral comm. Sipke Gongrijp). Whether the species is really as rare as the online map(verspreidingsatlas.nl) suggests is not clear. The apomicts in the Dryopteris affinisaggregate are difficult to separate, although it is possible after some experience.Considering the distribution pattern in the UK, where the species seems to be much morerestricted in the east than in the west, D. cambrensis may be and remain a rare species inthe Netherlands. It remains an open question whether this species will be able to establishin the Luzulo-Thelypteridetum. Its dispersion might be adequate to reach the siteseventually, but whether the conditions for the establishment of the species are appropriateis not certain. The current occurrence of the species in dense (young) forest plantationssuggest that D. cambrensis is confined to rather high levels of air humidity. In this respect,the environmental conditions in which the Luzulo-Thelypteridetum develops might befavourable also for the growth of D. cambrensis too. The future will show whether thisand other fern species can become established in this association in the Netherlands. Iexpect however, that the Luzulo-Thelypteridetum never will become as species rich andluxuriant as in the coastal areas of Argyll.
REFERENCESBREMER, P. 2016. Een nieuwe plantengemeenschap voor Nederland: het Luzulo
luzuloides-Thelypteridetum limbospermae Wittig 2000. Stratiotes 49: 11-22.BURNETT, J.H. (ed.) 1964. The vegetation of Scotland. Oliver & Boyd, Edinburgh &
London.FRASER-JENKINS, C. 2007. The species and subspecies in the Dryopteris affinis group.
290 FERN GAZ. 20(7): 281-291. 2018

Fern Gazette 18: 1-26.HENNEKENS, S.M. & SCHAMINÉE, J.H.J. 2001. TURBOVEG, a comprehensive data
base management system for vegetation data. J.Veg.Sci. 12: 589-591.KNABEN, G. 1950. Botanical investigations in the middle districts of western Norway.
Årbok for Universitetsmuseet 8: 1-177.MUELLER-DOMBOIS, D. & ELLENBERG, H. 1974. Aims and methods of vegetation
ecology. Wiley, London.PAGE, C.N. 1997. The ferns of Britain and Ireland. Cambridge University Press
Cambridge.RODWELL, J.S., PIGOTT, C.D., RATCLIFFE, D.A., MALLOCH, A.J.C., BIRKS,
H.J.B., PROCTOR, M.C.F., SHIMWELL, D.W., HUNTLEY, J.P., RADFORD, E.,WIGGINTON, M.J. & WILKINS, P. 1992. British Plant Communities. Volume 3.Grasslands and montane communities. Cambridge University Press, Cambridge.
TICHÝ, L. 2002. JUICE, software for vegetation classification. J.Veg.Sci. 13: 451-453.TREWREN, K. 2014. Some taxa within the Dryopteris affinis complex. A field guide.
British Pteridological Society, London.WEEDA, E.J. & HAVEMAN, R. 2017. Melampyro pratensis-Holcetea mollis. Stratiotes
50/51: 46-71.WESTHOFF, V. 1990. Neuentwicklung von Vegetationstypen (Assoziationstypen in statu
nascendi) an naturnahen neuen Standorten, erläutert am Beispiel der WestfriesischenInseln. Ber. Reinhold-Tüxen-Ges. 2: 11-23.
WESTHOFF, V. & VAN DER MAAREL, E. 1978. The Braun-Blanquet approach. In:WHITTAKER, R.H. (red.), Ordination and Classification of Communities, Springer,The Hague: 287-373.
WITTIG, R. 2000. Das Luzulo luzuloidis-Thelypteridetum limbospermae, eine azidoklineSaumgesellschaft der hoheren Mittelgebirge. Tuexenia 20: 131-142.
HAVEMAN: FERN-MEADOW VEGETATION 291

292 FERN GAZ. 20(7). 2018

MARSILEA AEGYPTIACA (MARSILEACEAE) ON THEMEDITERRANEAN ISLAND OF ELAFONISOS (LACONIA,
PELOPONNESE, GREECE)
Armin Jagel1 & Marcus Lubienski2
1 Danziger Str. 2, 44789 Bochum, Germany; [email protected] Am Quambusch 25, 58135 Hagen, Germany; [email protected]
Key Words: Marsilea aegyptiaca, Elafonisos, Greece, European species
ABSTRACTThe water-clover species Marsilea aegyptiaca was first detected on theMediterranean island of Elafonisos (Peloponnese, Greece) nearly 25 years ago.This was the first record for the species as part of the European flora. Recent workhas shown that M. aegyptiaca still occurs at the site, and data are presentedconcerning its identification, habitat and distribution. Morphological charactersof all known European species within the genus are compared.
INTRODUCTIONWater-clovers (Marsilea L.) are heterosporous ferns and the most species rich groupwithin the Marsileaceae. The family additionally comprises the pillworts (Pilularia L.)and the monotypic genus Regnellidium Lindm. All three genera are aquatic to semi-aquatic rhizomatous plants with roots and leaves born at nodes and sori arranged insporocarps (Kramer, 1990; Nagalingum et al., 2006). Recent phylogenetic studies haverevealed that the Clover ferns (Marsileaceae) together with the Floating ferns(Salviniaceae; Salvinia Seg. and Azolla Lam.) represent a monophyletic group ofheterosporous ferns within the fern clade, which evolved in the Mesozoic (Pryer, 1999;Smith et al., 2006; Nagalingum et al., 2006). Twentieth century treatments of the genusmainly focussing on morphology have been published for Africa (Launert, 1968, 1970,1971, 1984), Australia (Jones, 1998), India (Gupta, 1962), and the Americas (Johnson,1986). Recent molecular studies have thrown some light on the phylogeneticrelationships within Marsilea (Nagalingum et al., 2007, 2008; Whitten et al., 2012).
During extensive field work on the island of Elafonisos in 1991 (Laconia,Peloponnese, Greece; see Jagel, 1992) the semi-aquatic Water-clover species Marsileaaegyptiaca Willd. was found growing abundantly in a seasonal pool. This was not onlythe first record for Greece but also the first true European occurrence of the speciesknown at that time; its only known station at the time being from the Lower Volga inRussia (Jalas & Suominen, 1972; Tutin et al., 1993; Tzvelev, 2012) and an erroneousrecord for Spain (Alcober et al., 1980). The site was reinvestigated in 2016 and M.aegyptiaca confirmed to be still thriving at its obviously undisturbed habitat.
In regard to the biogeographical importance of the record, more detailed andcomprehensive data are given below.
BIOLOGY AND IDENTIFICATIONWater-clover species (Marsilea L.) are in general a variable and frequently confusinggroup of plants and depending on the life phase, are nearly impossible to separate fromeach other. Most members of the genus inhabit seasonally dry areas inundated only
FERN GAZ. 20(7): 293-300. 2018 293

during the rainy season and like most aquatic plants are characterized by a phenotypicplasticity, which renders identification on a species level quite difficult. This isemphazised when plants are in their sterile, vegetative growth stage with submergedrhizomes and floating leaves on the water surface. Sporocarps are formed only in theterrestrial life stage, but are essential for species determination. Sterile, aquatic plantslacking sporocarps are nearly impossible to identify, because the different species in thatlife stage are morphologically very similar.
Marsilea aegyptiaca (Figure 1) belongs to a group of xerophytic members within thegenus and is adapted to extreme dry conditions. Crouch et al. (2011) report that thespecies in Southern Africa inhabits arid semideserts and is obviously able to exist forseveral years without being submerged. Consequently M. aegyptiaca is quite small inits overall appearance (stipes in the dry land form max. 5 cm long with pinnae max. 1cm long) and shows hairy leaves (Figure 2 a-d), a feature often displayed by xerophyticMarsilea species. Nevertheless this is a very unstable and variable character, becausehairs on the leaves and petioles readily occur in hydrophytic species too, when being intheir terrestrial life stage. The pinnae of the land form are narrowly cuneate and pilosewith the distal margin being slightly truncate and shallowly bilobed. The sporocarp ofM. aegyptiaca is quite distinct (Figure 3): usually appearing in clusters of two or moreon short, free pedicels (2-8 mm), almost squarish in lateral view, with a distinct lateralfurrow, densely pilose at first, becoming subglabrous later, lacking an inferior but mostlyhaving a prominent superior tooth. In its aquatic life stage M. aegyptiaca is less distinct(see above). The floating leaves have stipes up to 30 cm long, broadly obovate, glabrousand larger pinnae (max. 2.5 cm) with rounded and shallowly sinuate distal margins(Figure 4 a, b).
294 FERN GAZ. 20(7): 293-300. 2018
Figure 1.Marsilea aegyptiaca at its growing site on the island of Elafonisos.

Table 1 shows the morphological characters of all known European species withinthe genus Marsilea. The former listed Azorean endemic species M. azorica Launert &Paiva has recently been shown to be a misidentification and synonymous with theinvasive M. hirsuta R. Br. from Australia and Asia (Schaefer et al., 2011) and thereforeis excluded here.
GEOGRAPHY, HABITAT, AND ECOLOGYElafonisos is located in the region of Laconia, an administration unit of the Peloponnese.The island lies just off the Southwest coast of Malea Peninsula and is separated from itby a 600 m wide strait. The history of Elafonisos as an island is short, being part of thepeninsula in former times. Therefore, it is not surprising that the largest part of the florais typical for the South Peloponnese. However, some species show relations to thefloristic region of the South Aegean (Jagel, 1992). In the Northwest part of Elafonisos at Leptos Kavos a seasonally moist habitat of about1000 m² exists (Figure 5 a, b), at a distance of not more than 100 m from theMediterranean sea. From late autumn until very early spring the shallow depression is
JAGEL & LUBIENSKI: MARSILEA AEGYPTIACA 295
Figure 2 a, b, c, d. Marsilea aegyptiaca, fertile dry land plant in terrestrial life stagewith hairy leaves.

296 FERN GAZ. 20(7): 293-300. 2018
M. quadrifolia M. strigosa M. batardae M. aegyptiaca
Sporocarp
Arrangement
clustered, 2-3,rarely solitary
stalked
mostly solitarycrowded in tworegularimbricating rowsalong the rhizome
sessile
solitary or inloose groups of 2-3, not crowdedalong the rhizome
stalked
clustered, usually2, rarely solitary
stalked
Shape
oblong-ellipsoid,beanshaped
longer than high
not laterallyflattened
oblique-elliptic,obliquelysquarish orrectangular,obovoid
longer than high
laterally usuallyflattened
irregularlysquarish tosubcircular
as long as high
laterally usuallyflattened
squarish
as long as high
laterally flattened
Size 3-6 mm 3-5 mm 3-4.5 mm 2-3 mm
Teeth
superior tooth 0-1(short toindistinguishable)
inferior tooth 0
superior tooth 1(short toindistinguishable)
inferior tooth 0-1
superior tooth 1(prominent)
inferior tooth 0-1
superior tooth 1(prominent, bluntto acute)
inferior tooth 0
Surface
glabrous (hairyonly whenyoung)
without a furrowon the lateralsides
strigose or silkypubescent, latersubglabrous
without a furrowon the lateralsides
strigose or silkypubescent, latersubglabrous
without a furrowon the lateralsides
densely piloselater subglabrous
with a distinctcurved verticalfurrow on thelateral sides
Pedicel (stalkof sporocarp)
2-20 mm
2- to 4-branched,rarely unbranched
1-2 mm
unbranched
2-8 mm
unbranched
2-8 mm
unbranched
Rhizome
long, creeping,profuselybranched
internodes morethan 10 mm
short, caespitose,sometimesbranched
nodes crowded
long, creeping,profuselybranched
internodes 5-20mm
long, creeping tosubcaespitose
internodes 5-10mm (sometimeslonger)
Pinnae Shape
obovate-cuneate
apex entire toundulate, rounded
obdeltate-cuneate
apex entire tocrenulate,rounded
obdeltate-cuneate
apex entire tocrenate
narrowly cuneateto obdeltate
apex deeplycrenate to sinuateor entire, rounded
Pubescence glabrous glabrous tosparsely hairy
glabrescent hairy or glabrous
Table 1: Macromorphological characters of the European species of Marsilea basedon Gupta (1962), Launert (1968, 1983, 1984), Castroviejo et al. (1986), Johnson(1986), Prelli (2001), Tutin et al. (1993), Delgado Vázquez & Plaza Arregui (2010),Crouch et al. (2011), Schaefer et al. (2011).

filled with fresh water not influenced by salt water. Marsilea aegyptiaca is very frequent in this small area, with hundreds or thousands
of fronds being found. A smaller population exists more or less 50 m east of the pond atthe border of a scrub. The sub-surface of the site is formed by limestone rocks(Symeonidis, 1969), covered by a shallow layer of sandy fine soil. After the beginningof the rainy season in autumn e.g. when Narcissus serotinus L. is flowering and whenthe shallow depression is filled up with rainwater, Ranunculus peltatus Schrank subsp.fucoides (Freyn) Munoz Garmendia (= Ranunculus saniculifolius Viv.) can be found asa water plant in the pond. At this time of year the Marsilea grows in its aquatic life stage(Figure 4). The pond is framed by tufts of Juncus heldreichianus Parl. and Schoenusnigricans L. At the end of winter or in early spring, usually at the end of March, the pondrapidly dries out within a few weeks. Species found growing together with M. aegyptiacaare Anagallis foemina Miller, Briza minor L., Isoetes phrygia (Boiss.) Hausskn., Isolepiscernua (Vahl) Roem. & Schult., Juncus capitatusWeigel, Juncus hybridus Brot., Lythrumborysthenicum (Schrank) Litv., Lythrum hyssopifolia L., Mentha pulegium L., Plantagoweldenii Rchb., Polypogon maritimus Willd., Ranunculus muricatus L., Romulearamiflora Ten. Sagina maritima G.Don, Solenopsis laurentia (L.) C.Presl, Trifoliumlappaceum L. Trifolium resupinatum L., and Triglochin barrelieri Loisel. Most of theseare highly specialized and characteristic for such sites in the Peloponnese.
DISTRIBUTIONMarsilea aegyptiaca is mainly an African species, recorded for Ethiopia, Botswana,
JAGEL & LUBIENSKI: MARSILEA AEGYPTIACA 297
Figure 3 a, b, c, d. Marsilea aegyptiaca showing characteristic clustered, squarish,pilose sporocarps with lateral furrow and blunt superior tooth on short pedicels.

Namibia, South Africa, Sudan, Egypt, Tunisia, Algeria, Libya, and Madagascar (Launert,1984; Crouch et al., 2011). The species also extends its range to the east with occurrenceson the Indian subcontinent (Gupta, 1962; Sharma, 2005).
Until the 1990s M. aegyptiaca was known from Europe only from the river Volgaand its delta near the Caspian Sea in Russia (Jalas & Suominen, 1972; Tutin et al., 1993;Tzvelev, 2012). A record for Spain (Alcober et al., 1980) turned out to be amisidentification and the plants are now known to belong to a different species, Marsileabatardae Launert, occurring in Southwest Spain and Southern Portugal (Launert, 1983;Castroviejo et al., 1986; Rosselló-Graell et al., 2000; Prelli, 2001; Delgado Vázquez &Plaza Arregui, 2010).
The discovery of M. aegyptiaca on the Peloponnesian island of Elafonisos in 1991therefore was a remarkable addition to the knowledge concerning the range of the speciesnow extending into the Eastern Mediterranean (Jagel, 1992) and representing the firstrecord for Greece and the European Union. Later a second occurrence for the region hasbeen reported from Cyprus (Hand, 2011), validating the species as an element of theEuropean mediterranean flora.
HABITAT AND THREATSIn former times sites of M. aegyptiaca on Elafonisos were used for agricultural purposes
298 FERN GAZ. 20(7): 293-300. 2018
Figure 5 a, b. Habitat of Marsilea aegyptiaca on Elafonisos, dried up in spring (a)and flooded in winter (b).
Figure 4 a, b. Marsilea aegyptiaca, sterile plant with floating leaves in aquatic lifestage.

perhaps because of the good water supply, with evidence of several low walls of collectedfield stones. Nowadays there is no noticeable human influence, neither huts, houses norstables are in the proximity so no current threat for the fern is perceived. The fact thatthe species has been known there for nearly 25 years now with no obvious change to thesite supports this assessment.
A large area of similar seasonally wet ponds is situated nearby on the mainland inViglafia north of the beach of Punta. The area seems to be comparable with the pond ofElafonisos but is quite overgrazed and in parts more brackish. Although it has beeninvestigated intensively over several years no plants of the Water-clover could be foundalthough most accompanying plants of the Elafonisos site were present. However thesite is nevertheless remarkable for hosting some very rare plant species which could notbe found on Elafonisos: Callitriche brutia Petagna, Cicendia filiformis (L.) Del.,Crassula vaillantii (Willd.) Roth, Isoetes gymnocarpa (Gennari) A.Braun, Isoetes histrixBory, and Myosurus heldreichii Heldr. ex H.Lév. Additionally a third Quillwort species,Isoetes phrygia (Boiss.) Hausskn., is present here (as it is at the Marsilea site onElafonisos), together with a hybrid probably involving this taxon.
ACKNOWLEDGEMENTSThe authors would like to thank Dr. H. W. Bennert (Ennepetal [formerly of Ruhr-University of Bochum], Germany) for the initial confirmation of Marsilea aegyptiacain 1992. He also contributed with literature and helpful discussions. Dr. V. M. Dörken(University of Konstanz, Germany) also supplied us with literature. Dr. A. Troia(University of Palermo, Italy) is gratefully acknowledged for his help with thedetermination of the different Isoetes species at the sites.
REFERENCESALCOBER, J.A., IBARS, A., MANSANET, J., MATEU, I. & PUCHE, M.F. 1980.
Marsilea aegyptiaca Willd. nueva cita para España. Anales Jard. Bot. Madrid 36: 55-59.
CASTROVIEJO, S., LAíNZ, M., LóPEZ GONZALES, C., MONTSERRAT, P.,MUñOZ GARMENDIA, F., PAIVA, J. & VILLAR L. 1986. Flora Iberica: Plantasvasculares de la Peninsula Iberica e Islas Baleares. Vol. 1. Lycopodiaceae-Papaveraceae. Real Jardín Botánico, C.S.I.C., Madrid.
CROUCH, N.R., KLOPPER, R.R., BURROWS, J.E. & BURROWS, S.M. 2011. Fernsof Southern Africa. A comprehensive guide. Struik Nature, Cape Town.
DELGADO VáZQUEZ, A.J. & PLAZA ARREGUI, L. 2010. Helechos Amenazados deAndalucía: Avances en Conservación. Consejería de Medio Ambiente. Junta deAndalucía, Sevilla.
GUPTA, K.M. 1962. Botanical Monograph No. 2. Marsilea. Council of Scientific &Industrial Research, New Delhi.
HAND, R. 2011. Supplementary notes to the flora of Cyprus VII. Willdenowia 41: 341-355.
JAGEL, A. 1992. Zur Flora und Vegetation der Insel Elafonisos (Lakonien,Griechenland). Diploma thesis. Spezielle Botanik, Ruhr-University, Bochum.
JALAS, J & SUOMINEN, J. (eds.) 1972. Atlas Florae Europaeae. 1. Pteridophyta(Psilotaceae to Azollaceae). The Committee for Mapping the Flora of Europe andSocietas Biological Fennica Vanamo, Helsinki.
JOHNSON, D.M. 1986. Systematics of the New World Species of Marsilea(Marsileaceae). Syst. Bot. Monographs 11. Ann Arbor.
JONES, D.L. 1998. Marsileaceae. In: Orchard, A. E. & McCarthy, P. M. (eds.) Flora of
JAGEL & LUBIENSKI: MARSILEA AEGYPTIACA 299

Australia, Vol. 48, Ferns, gymnosperms and allied groups, pp. 166-173,ABRS/CSIRO Australia, Melbourne.
KRAMER, K.U. 1990. Marsileaceae. In: KUBITZKI, K. (Ed) The Families and Generaof Vascular Plants. Volume I. Pteridophytes and Gymnosperms, pp. 180-183,Springer, Berlin/Heidelberg/New York/London/Paris/Tokyo/Hong Kong/Barcelona.
LAUNERT, E. 1968. A monographic survey of the genus Marsilea Linnaeus.Senckenbergiana biol. 49: 273-315.
LAUNERT, E. 1970. Marsilea Linnaeus: Variation on a theme. Senckenbergiana biol.51: 433-435.
LAUNERT, E. 1971. A remarkable new African species of Marsilea Linnaeus.Senckenbergiana biol. 52: 449-452.
LAUNERT, E. 1983. A new species of Marsilea from Portugal. Bol. Soc. Brot., Sér. 2,56: 99-104.
LAUNERT, E. 1984. A revised key to and new records of African species of the genusMarsilea. Garcia de Orta, Sér. Bot., Lisboa 6: 119-140.
NAGALINGUM, N.S., NOWAK, M.D. & PRYER, K.M. 2008. Assessing phylogeneticrelationships in extant heterosporous ferns (Salviniales), with a focus on Pilulariaand Salvinia. Bot. J. Linn. Soc. 157: 673-685.
NAGALINGUM, N.S., SCHNEIDER, H. & PRYER, K.M. 2006. Comparativemorphology of reproductive structures in heterosporous water ferns and a re-evaluation of the sporocarp. Int. J. Plant Sci. 167: 805-815.
NAGALINGUM, N.S. SCHNEIDER, H. & PRYER, K.M. 2007. Molecular phylogeneticrelationships and morphological evolution in the heterosporous fern genus Marsilea.Syst. Bot. 32: 16-25.
PRELLI, R. 2001. Les fougères et plantes alliées de France et d’Europe occidentale.Belin, Paris.
PRYER, K.M. 1999. Phylogeny of marsileaceous ferns and relationships of the fossilHydropteris pinnata reconsidered. Int. J. Plant Sci. 160: 931-954.
ROSSELLO-GRAELL, A., DRAPER, D., TAULEIGNE GOMES, C. & CORREIA,A.I.D. 2000. Distribuição de Marsilea batardae Launert em Portugal e determinaçãodo seu estatuto de ameaça. Portugaliae Acta Biol. 19: 219-224.
SCHAEFER, H., CARINE, M.A. & RUMSEY, F.J. 2011. From European priorityspecies to invasive weed: Marsilea azorica (Marsileaceae) is a misidentified alien.Syst. Bot. 36: 845-853.
SHARMA, O.P. 2005. Pteridophytic flora of Bundi district, Southeastern Rajasthan.Zoos’ Print Journal 20: 1836-1837.
SMITH, A.R., PRYER, K.M., SCHUETTPELZ, E., KORALL, P., SCHNEIDER, H. &WOLF, P.G. 2006. A classification for extant ferns. Taxon 55: 705-731.
SYMEONIDIS, N. 1969: Stratigraphisch-Paläontologische Untersuchungen über dieNeogenablagerungen von Lakonien, insbesondere der Gebiete von Neapolis(Eparchia Epidaurus-Limira) und gegenüber liegenden Insel Elaphonisos. Ann. Geol.Pays Helleniques 21: 531-553.
TUTIN, T.G., BURGES, N.A., CHATER, A.O., EDMONDSON, J.R., HEYWOOD,V.H., MOORE, D.M. VALENTINE, D.H., WALTERS, S.M. & WEBB, D.A. (eds.)1993. Flora Europaea. Volume 1. Psilotaceae to Platanaceae. 2nd ed. CambridgeUniversity Press, Cambridge.
TZVELEV, N.N. (ed.) 2012. Conspectus Florae Europae Orientalis. Tomus 1.,Consociatio editionum scientificarum KMK, Petropoli/Mosqua.
WHITTEN, W.M., JACONO, C.C. & NAGALINGUM, N.S. 2012. An expanded plastidphylogeny of Marsilea with emphasis on North American species. Amer. Fern J. 102:114-135.
300 FERN GAZ. 20(7): 293-300. 2018

SHORT NOTE
HYPODEMATIUM CRENATUM SUBSP. CRENATUM(HYPODEMATIACEAE): A NEW DISTRIBUTIONAL RECORD FOR
GUJARAT STATE
The state of Gujarat is divided into five zones viz., Kutch, Saurastra, North, Central andSouth Gujarat. These zones are based on climate and geographic location. The districtJunagadh is located on the Kathiawar peninsula in western Gujarat (Saurastra zone). Itcontains the Gir National Park and Girnar wildlife sanctuary, which is the only home forAsiatic lions. Girnar hills are also declared as a forest reserve for bamboos (Nakar &Jadeja, 2015), and ‘Girnari Giddh’, the long-billed vulture, is found only in the Girnarhills. The Girnar region alone accounts for about 25 per cent of the species in Gujaratstate (Patel et al., 1961). However, lycophyte and fern diversity of the whole of theGujarat state including Saurashtra regions (particularly Junagadh) is less understood,although recently there have been a few reports on the pteridophyte diversity of Central,North and South Gujarat (Kachhiyapatel et al., 2015, 2016; Rajput et al., 2016). Thepresent investigation was carried out to study pteridophytes from Girnar hills. Duringthis survey an interesting pteridophyte was collected which, after critical observation ofmorphological characters, was identified as Hypodematium crenatum (Forssk.) Kuhn.subsp. crenatum and it is reported here as new distributional record for Gujarat state.The identification was confirmed by comparing generated nucleotide sequences with theGenBank database, and the sequence data were submitted to BOLD.
The genus Hypodematium in the family Hypodematiaceae is represented by 20species in the world (PPG-I, 2016). A sole species of Hypodematium (divided into threesubspecies viz., H. crenatum subsp. crenatum, H. crenatum subsp. loyalii Fraser-Jenk.& Khullar and H. crenatum subsp. mehrae Fraser-Jenk.) has been reported from India(Fraser-Jenkins, 2008). We have found a single subspecies, H. crenatum subsp. crenatum,for Gujarat state. Its description, range of distribution, ecology, conservation status andspecimen examined is given.
Systematic TreatmentHypodematium crenatum (Forssk.) Kuhn, subsp. crenatum, Fraser-Jenkins, Taxon. Revis.Indian Sub-continental Pteridophytes., 204 - 206, 612. 2008; Patil, Systematic Studieson Pteridophytes of Satara District, 111-112. 2014. Hypodematium crenatum (Forssk) Kuhn. Manickam and Irudayaraj, Pterid. FI. W.Hypodematium hirsutum (Don) Ching. Indian Fern J. 1(1-2): 49.1985.Plant lithophytic, perennial herb; rhizome long creeping, densely red-brown scaly, givingwoolly look to rhizome; scales 1-2 cm × 1-3 mm, red-brown, oblong lanceolate-lanceolate, glossy, apex long acuminate, margin entire; frond 20-30 cm × 15-20 cm,green-dark green, bipinnatified-tripinnatifid; stipe 10-18 cm long, brown, densely scalyat base, grooved, glabrous above; lamina 5-12×15-20 cm, bi-pinnatifid-tri-pinnatifid,ovate, apex acute, base cuneate, basal half tri-pinnatifid, distal half base bi-pinnatifid,distal most part simply pinnatifid; pinnae 6-10 pairs, subopposite, lanceolate, apex acute,base decurrent, pale brown, stiff, hairs sparsely-densely distributed; texture herbaceous;vein forked 3-4 times, indistinct above and below, free, reaching up to margin; sorimedian on the vein lets in two rows; indusia reniform, entire, densely covered with shortstiff hair; spores 45-55 µm in dia., reniform to ellipsoid, exine with prominent
FERN GAZ. 20(7): 301-303. 2018 301

protuberances (and see Figure 1).Distribution: World: Africa, China, India, Japan, Malaysia, Myanmar, Philippines andThailandIndia: Himachal Pradesh, North East India, Punjab, Gujarat, Maharashtra, Kerala.Gujarat: Junagadh
302 FERN GAZ. 20(7): 301-303. 2018
Figure 1. A: Habit of Hypodematium crenatum subsp. crenatum. B: Frond C: RhizomeD: Stipe base showing dense scales E: Stipe base scale F: Pinna G: Pinnules showingarrangement of sori H: Enlarged view showing hairs on costules I: Sorus J: IndusiumK: Sporangium L: Spore. Scale bar:A-3 cm, B-3 cm, C- 1cm, D-0.5 cm, E-2 mm, F-2 cm, G-5 mm, H-1mm, I-0.5 mm, J-0.5 mm, K-100 µM, L-25 µM.

BOLD ID: MIPDG013-15Phenology: Vegetative phase: June-September; Reproductive phase: October-DecemberEcology: Lithophytic, collected from rock crevices at Girnar hill slopes (on the way toSitavan), between 650-700 m.Specimens Examined: INDIA-Maharashtra: Kolhapur Dt., Pankala, alt 1000 m.,26/10/1962, Bhandare, BLAT; 02/01/1952, V. D. Vartak (BLAT205), BLAT; Satara Dt.,Wai Tehasil; Panchagani, alt. 1100 m., Oct. 1918, Blatter (BLAT1211), BLAT; SataraDt., Panchgani, alt. 1200 m., Sept. 2014, (SMP10071) SUK; Gujarat: Junagadh dt, Girnarhills, on the way to Sitavan, 680 m, (21°32’03.50” N, 70°31’28.10” E, at 668 m), R.N.Kachhiyapatel & K.S. Rajput 129, 26/12/2014, BARO.
ACKNOWLEDGMENTSThe authors are grateful to the Gujarat Biodiversity Board (GBB) and Forest Department,Government of Gujarat for providing necessary permission to visit the area and permitcollection of the specimens, and to an anonymous reviewer and Dr Mary Gibby forvaluable suggestions on the previous version of the manuscript.
REFERENCESFRASER-JENKINS, C.R. 2008. Taxonomic Revision of Three Hundred Indian
Subcontinental Pteridophytes with a Revised Census-List. Bishen Sing MahendraPal Sing Dehara Dun.
KACHHIYAPATEL, R.N., SINGH, A.P., RAOLE, V.M. & RAJPUT, K.S. 2015.Distribution and occurrence of some pteridophytes in Gujarat: a new record for thestate. J. of Indian Bot. Soc. 94 (3&4): 236-244.
KACHHIYAPATEL, R.N., PATEL, S.K. & RAJPUT, K.S. 2016. Reporting Adiantumcapillus-veneris L. and Thelypteris prolifera (Retz.) C.f. Reed as new record forGujarat state together with their molecular identity. Indian Fern J. 33: 69-80.
NAKAR, R.N. & JADEJA, B.A. 2015. Flowering and fruiting phenology of some herbs,shrubs and under-shrubs from Girnar Reserve Forest, Gujarat, India. Curr. Sci. 108(1):111-118.
PATEL, G.D., RAJYAGOR, S.B., TRIPATHY, S. & CHOKSHI, U.M. 1961. Gujaratstate gazetteers. Directorate of Govt. Print., Stationery and Publications.
PATIL, S. 2014. Systematic studies on pteridophytes of Satara districts (Maharashtra).A Ph. D. Thesis submitted to Shivaji University, Kolhapur.
RAJPUT, K.S., KACHHIYAPATEL, K.N., PATEL, S.K. & RAOLE, V.M. 2016.Assessment of Pteridophyte Diversity and their status in Gujarat state, Western India.Plant Sci. Today’s. 3(4):337-348.
THE PTERIDOPHYTE PHYLOGENY GROUP, 2016. A community-derivedclassification for extant lycophytes and ferns. J. Syst. Evol. 54(6):563-603.
R. N. KACHHIYAPATEL, S. M. PATIL & K. S. RAJPUTLaboratory of Plant Anatomy & Pteridology, Department of Botany, The M. S.
University of Baroda, Vadodara – 390 002. *Address for correspondence: Email:[email protected]
KACHHIYAPATEL ET AL: HYPODEMATIUM CRENATUM 303

304 FERN GAZ. 20(7). 2018

SHORT NOTE
ELEVEN NEW COMBINATIONS FOR MALESIAN FERNS
The preparation of a checklist of pteridophytes from Gunung Mulu National Park,Sarawak, necessitates eleven new combinations to bring the generic treatment into linewith PPG 1 [Pteridophyte Phylogeny Group] (2016).
NEW COMBINATIONS
DAVALLIACEAE1. Davallia grammatosora (Copel.) Parris comb. nov., based on Davallodesgrammatosora Copel., Philipp. J. Sci. Bot. 3: 34, t. 6 (1908), syn. Davallodes hirsuta(C.Presl) Copel. non Davallia hirsuta Sw. (1801). Type: Philippines, Mindanao,Copeland 1724 (BM, P).
2. Davallia pusilloides (Copel.) Parris comb. nov., based on Humata pusilloides Copel.,Sarawak Mus. J. 2: 338 (1917). Type: Philippines, Mindanao, Zamboanga, San Ramon,May 1910, E B Copeland 153 (BM, K, MU, P, PNH, SING, UC).
3. Davallia subvestita (C.Chr.) Parris comb. nov., based on Humata kinabaluensis var.subvestita C.Chr., Gardens’ Bulletin Straits Settlement 7: 232 (1934). Type: Borneo,Sabah, Mt Kinabalu, near Lobang, 18 Nov. 1931, R E Holttum SFN 25549 (BM, BO,K,SING).
HYMENOPHYLLACEAE1. Abrodictyum saxatile (T.Moore) Parris comb. nov., based on Trichomanes saxatileT.Moore, Gard. Chron. 1862: 45 (1862). Type: Borneo, collector not specified (K?).
2. Abrodictyum setigerum (Backh. ex T.Moore) Parris comb. nov., based onTrichomanes setigerum Backh. ex T.Moore, Gard. Chron. 1862: 45 (1862). Type, Borneo,collector not specified (K?).
3. Crepidomanes novoguineense (Blume) Parris comb. nov., based on Trichomanesnovoguineense Brause, Bot. Jahrb. Syst. 49: 7 (1912). Type: Papua New Guinea, Lager“Hochmoos” 65 km südwärts der Tami-Mündung, 1600 m, Juli 1910, L Schulze 33/35(B).
4. Crepidomanes teysmannii (Bosch) Parris comb. nov., based on Trichomanesteysmannii Bosch, Ned. Kruid. Arch. 5: 142 (1861). Type: Sumatra, Teysmann s. n. (L).
5. Hymenophyllum album (Blume) Parris comb. nov., based on Trichomanes albumBlume, Enum. Pl. Javae: 226 (1828). Type: Java, Blume s. n. (L).
POLYPODIACEAE1. Leptochilus bolsteri (Copel.) Parris comb. nov., based on Polypodium bolsteriCopel., Philipp. J. Sci. 1, Suppl. 4: 257, pl. 4a (1906). Type: Philippines, Mindanao,Surigao, April 1906, Bolster s. n. (MICH).
FERN GAZ. 20(7). 2018 305

THELYPTERIDACEAE1. Amblovenatum immersum (Blume) Parris comb. nov., based on Aspidiumimmersum Blume, Enum. Pl. Javae: 156 (1828). Type: Java, Blume s. n. (L).
2. Amblovenatum paraphysophorum (Alderw.) Parris comb. nov., based onDryopteris paraphysophoraAlderw., Bull. Bot. Buitenzorg, ser. 3, 2: 143 (1920). Type:Sumatra, Lebong Tandai, C J Brooks 250S (BM, BO).
BARBARA S. PARRISFern Research Foundation, 21 James Kemp Place,
Kerikeri, Bay of Islands, New Zealand 0230Email: [email protected]
306 FERN GAZ. 20(7). 2018

FERN GAZ. 20(7). 2018 307

INSTRUCTIONS FOR AUTHORSPAPERS should not usually exceed 20 printed pages and are generally expected to beconsiderably shorter. As well as reports of original research, review articles and shortnotes, e.g. new records, are encouraged. The senior author should supply an email addressto facilitate correspondence. REVIEWS should cover recent developments in any aspect of research on ferns andlycopods. This can include unpublished research by the author, and/or the possibledirection of future research. They should be aimed at the general reader who may not bean expert in the chosen topic. Proposals for a review should be discussed first with thereview editors.MANUSCRIPTS should be submitted in English (British) in electronic format in10-point Times New Roman font and single spaced. Electronic versions of text and tablesshould be compatible with WORD, with figures as TIFF files, and sent as emailattachments. All manuscripts will be refereed.THE TITLE should reflect the content of the paper and be in BOLD CAPITALS(11-point) and centrally aligned. Generic and specific names should be in italics and anytitle containing a generic or specific name should be followed by the family name inbrackets e.g.
TRICHOMANES SPECIOSUM (HYMENOPHYLLACEAE)IN SOUTHERN SPAIN
AUTHOR ABBREVIATIONS should follow Pichi Sermolli’s (1996) Authors ofscientific names in Pteridophyta, Royal Botanic Gardens, Kew.MAIN HEADINGS: should be in BOLD CAPITALS (10-point) and centrally aligned.SUBSIDIARY HEADINGS: should be in bold, the first letter of each word in capitals,the rest in lower case and left-aligned.AUTHORS’ NAMES AND FULL ADDRESSES (including email address): followthe title and are centrally aligned.KEY WORDS: up to ten.ABSTRACT: should reflect the content of the paper.FIGURES: there is no distinction between photographs and line drawings in numbering.All should be supplied in mono or colour, TIFF format, 300 dpi and CMYK. (pleasecontact the Production Editor with queries), with a scale bar where appropriate. Letteringor numbers (Arabic) should be in the bottom left using uppercase Times Roman. Figurecaptions should be on a separate sheet.TABLES: can be printed in either portrait or landscape format. Authors should considerthis when preparing tables. Authors should ensure that tables fit the printed page size ina legible form.PAGESIZE: The Gazette has a useable page size of 123 × 204 mm. This can be used inportrait or landscape mode but any supplied artwork should not be larger than this.MEASUREMENTS: follow the metric system.CHECKLISTS: follow the format of Baksh-Comeau, Fern Gaz. 16(1, 2): 11- 122.REFERENCES: follow the style of a recent issue of The Fern Gazette, e.g.:-HOOKER, W.J. 1864. Species Filicum, 5. Dulau & Co., London.MORTON, C.V. 1947. The American species of Hymenophyllum, section
Sphaeroconium. Contr. U.S. Natl. Herb. 29(3): 139-201.STEVENSON, D.W. & LOCONTE, H. 1996. Ordinal and familial relationships of
pteridophyte genera. In: CAMUS, J.M., GIBBY, M. & JOHNS, R.J. (Eds)Pteridology in perspective, pp. 435-467. Royal Botanic Gardens, Kew.
JOURNAL ABBREVIATIONS: should follow Botanico Periodicum Huntianum &Supplements.Alterations from the original text at proof stage will be charged for unless they are minorpoints of detail. A pdf will be provided free to the senior author.
308 FERN GAZ. 20(7). 2018

GLOBAL REVIEW OF RECENT TAXONOMIC RESEARCH INTOISOETES (ISOETACEAE), WITH IMPLICATIONS FOR
BIOGEOGRAPHY AND CONSERVATION1
D.F. BRUNTON* & A. TROIA**
*216 Lincoln Heights Road, Ottawa, Ontario Canada K2B A8A,[email protected]
**Dipartimento di Scienze e Tecnologie Biologiche Chimiche e Farmaceutiche,Sezione di Botanica ed Ecologia Vegetale, Università degli Studi di Palermo, Via
Archirafi 38, I-90123, Palermo Italy, [email protected]
Key Words: Isoetes, lycophytes, taxonomy, global research, biogeography,conservation
ABSTRACTWith the advent of cytological analysis, Scanning Electron Microscopy, therecognition of sterile hybrids, improved access to wild populations andincreasingly comprehensive molecular investigations, there has been a revolutionin Isoetes taxonomy in recent times. The last 40 years have seen an almost 100%increase in the diversity documented during the previous two centuries. Fourgeographic areas account for over 90% of this diversity: northern-central SouthAmerica, eastern North America, western Eurasia (including the northernMediterranean) and South to Tropical Africa. There is a significant globalconservation concern for Isoetes, with many taxa being rare (known from one toa handful of populations) and some known only from herbarium specimens. Thetaxa of the extraordinary rock-outcrop pools and the ephemeral wetlands foundin disjunct areas around the world appear to be particularly vulnerable to habitatdestruction. We expect 100 or more new taxa to be described in coming years.Such taxonomic productivity will require more systematic cytological andmolecular investigations to be undertaken. It also requires the assembly of a largerglobal inventory of high-quality voucher specimens to provide the material forsuch investigations.
INTRODUCTIONSpeaking of quillworts (Isoetes, Isoetaceae), the late geneticist and pteridologist, DonaldBritton, said only half-jokingly “…. this is a crazy genus! Just add water and stir” (D.M. Britton in lit, 1989). In the following, we undertake a review of the dramatic changesthat have occurred in our global understanding of the taxonomy of this ‘crazy genus’ inrecent decades as a result of the work of Britton and many others.
Isoetes (Isoetaceae) is an ancient heterosporous lycophyte dating back to the JurassicPeriod (Pigg, 2001). With few exceptions, species exhibit a similar simple form andstructure (Figure 1). A spirally-arranged, typically small (3 - 30 cm tall), set of
1 The authors dedicate this paper to the memory of Donald M. Britton (1923-2012)and Enrico Bellini (1940-2002) who in addition to being fine gentlemen and scholars,inspired and developed our interest in Isoetes.
FERN GAZ. 20(8): 309-333. 2018 309

unbranched quill-like leaves with usually partially velum-covered sporangia embeddedinto their basal abaxial faces, is developed from a shallowly buried two or three-lobedcorm. At a gross scale there is not much to separate one taxon from another. Just thesame, a high degree of taxonomic diversity as well as a high degree of endemism (Jermy,1990) exists with approximately 200 species, subspecies and varieties currentlyrecognized (Troia et al., 2016).
Spore morphology is the primary character employed for distinguishing individualIsoetes taxa. The megaspores demonstrate a remarkable diversity of perispore (surface)ornamentation expressions (Figure 2), ranging from laevigate (virtually featureless) totuberculate (covered in broad, low, symmetrical projections) to echinate (spiny) orcristate-reticulate (intricate patterns of muri (walls), tubercles and/ or spines) (Hickey,1986b; Punt et al., 2007). These ornamentation patterns are typically expressedconsistently throughout the range of each taxon.
The notable exception to consistent morphological representation is with sterilehybrids. The megaspores of such taxa have been the source of much confusion for overa century (Engelmann, 1882; Motelay, 1892; Eaton, 1900), exhibiting a range of sizesand morphological features intermediate between their putative parents. Deformed (oftenflattened) individual megaspores with congested ornamentation patterns likened to braincoral and/ or with irregular, even ‘dumb-bell’ shapes (Figure 3) are frequent and arestrongly indicative of sterile hybrid status.
Microspores are typically less boldly ornamented, varying from plain to tuberculateto echinate (Figure 4). While consistent in size in relation to the cytology of the particulartaxon (Kott & Britton, 1983; Taylor et al., 1993; Musselman, 2002; Macluf et al., 2006),they often vary widely in appearance, even within individual populations. Microsporesof hybrids are variable in size. Typically they are amphibious and misshaped and, as withmegaspores, exhibit ornamentation patterns intermediate between those of the putativeparents.
All Isoetes occupy sites which are at least seasonally wet and grow in fresh (rarelybrackish) water, typically being amphibious or purely aquatic, most growing in sterileor at least nutrient poor, typically acidic to circumneutral substrates. These habitats varywidely from deep-water oligotrophic lakes (growing as deeply as 4-5 metres),permanently flowing rivers and high elevation bog pools, to open woodland seepages,shallow ephemeral swales and briefly inundated pools in open bedrock outcrops.
310 FERN GAZ. 20(8): 309-333. 2018
Figure 1: A typical Isoetes plant: A) in situ, and B) excavated, also showing corm androots (Isoetes minimaA.A.Eaton, Salmo, British Columbia, Canada, 11 June 2017 and28 June 2008, respectively) (Photos: D. F. Brunton).

Through all of this, be they like the more than half metre long, flexulose aquaticspecies Isoetes japonica A.Braun of eastern Asia, the more typically less than 30 cmlong tall, reflected, emergent I. drummondii A.Braun of southern Australia, or thestiffly-erect, 5-15 cm tall rock pool ephemeral I. piedmontana (N.Pfeiffer) C.Reed of thesoutheastern United States (USA), virtually all Isoetes retain this simple gross-scale
BRUNTON & TROIA: TAXONOMIC RESEARCH INTO ISOETES 311
Figure 2: Variations in Isoetesmegaspore ornamentation: A) diploid I. nuttalliiA.Braun,B) diploid I. cubana Engelm., C) decaploid I. lacustris L.(s. str.). (SEM images A & C- P. Sokoloff; B - D. M. Britton).
Figure 3: Aborted, sterile Isoetes hybrid megaspores: A) triploid I. engelmannii A.Braunx hyemalis D.F.Brunton, b) heptaploid I. ×harveyi A.A.Eaton, pro sp. (I. echinosporassp. muricata (Durieu) Löve & Löve x lacustris) (SEM image A - P. Sokoloff; B - D.M. Britton).
Figure 4: Variation in Isoetesmicrospore ornamentation: A) tetraploid I. septentrionalisD.F.Brunton., B) I. longissima Bory (s. str.) (SEM image A - P. Sokoloff; B - CarmelaDi Liberto, University of Palermo, Italy (with Angelo Troia). (Scale bar = 10 µm)

appearance.There has been a substantial change in our global understanding of the taxonomic
diversity of Isoetes, however, since it was last enumerated almost 100 years ago (Pfeiffer,1922). Change has been particularly dramatic in the latter half of that period (Troia etal., 2016), which is the focus of this review. For our purpose, ‘recent’ commences ca.1980 when Scanning Electron Microscope (SEM) and cytology were first applied incomprehensive reviews of Isoetes in North America (Boom, 1979; Kott, 1980; Kott &Britton, 1983), Europe (Berthet & Lecoq, 1977) and Australia (Marsden, 1979).
METHODSThe following assessments are based upon a comprehensive reading of the Isoetesliterature as well as the authors’ experience from investigations of the genus since the1980s. This includes on-site ecological and population investigations in southern Europe(A.T.) and across North America (D.F.B.). In concert with various research associatesand supplemented by extensive investigations of herbarium material, SEM imagery andthe application of cytological data, these studies have clarified or described a substantialnumber of taxonomic issues (see References, below). We pay special attention to theliterature concerning systematics, phylogeny, karyology and biogeography. We alsoconsidered the literature in other areas of investigation such as ecology, vegetationscience, physiology and conservation biology. For the nomenclatural purposes of thepresent study we follow Troia et al. (2016).
DISCUSSIONHistorical ContextPfeiffer (1922) conducted the most comprehensive global review of the genus, producinga mostly herbarium-based enumeration of the traditional 19th Century - early 20thCentury understanding of Isoetes diversity. Her study advanced the more modest reviewsof Baker (1880) and Motelay (1883) and relied especially upon the work of Braun (1846),Durieu (1864), Engelmann (1867; 1888) and Eaton (1900). Working independently, shedescribed 77 species and varieties globally. The publication of her remarkably insightfulreview coincided with an independent description of 15 new South American species(Weber, 1922), resulting in a total of 92 taxa being accepted at a species, subspecies orvarietal level by the end of that year. That fewer than 25 species were describedworldwide in the subsequent 50 years indicates how the technology and raw dataavailable to researchers in that earlier era had largely been exploited by Pfeiffer’s time.
After a long period of relative nomenclatural inactivity following Pfeiffer (1922),Isoetes has experienced a surge in taxonomic innovations. In effect, there has been a late20th Century - early 21st Century Isoetes taxonomic ‘renaissance’. This results in partfrom geographically expanded investigations of previously poorly explored regions andhabitats. More substantially, however, it reflects the application of new investigativetools for the reconsideration of both long-standing interpretations and new data.
Cytology and SEMFollowing the pioneering work of Manton (1950) on various pteridophyte genera, theconnection between chromosome number and Isoetes spore size was investigated in detailin northern North America (Kott, 1980; Taylor et al., 1985; Taylor & Hickey, 1992).These break-through investigations provided confident determination of both theconsistency and taxonomic significance of physical distinctions that had previously been
312 FERN GAZ. 20(8): 309-333. 2018

thought to represent only insignificant morphological variations within broadlycircumscribed species. Cytological data quickly became a major line of evidence insubsequent global taxonomic investigations.
Cytological investigations revealed that genetically distinct polyploid taxa arecommon in Isoetes, derived from basic diploid species (2x=2n=22) through hybridizationand subsequent chromosome doubling (Taylor et al., 1985; Taylor & Hickey, 1992; Tayloret al., 1993). Significant progress was made in documenting Isoetes cytology in thisperiod, with chromosome counts for over 80 taxa reported in Takamiya (1999) alone.The chromosome numbers for over 46% of all named taxa are presently known (Troiaet al., 2016).
The production of detailed, clear illustrations of seemingly subtle variations of variousIsoetes morphological features through the systematic application of SEM becamecommon in the 1980s and 1990s (Kott & Britton, 1983; Prada, 1983; Ferrarini et al.,1986; Schelpe & Anthony, 1986; Srivastava et al., 1993; Musselman & Knepper, 1994).For the first time this permitted commonly occurring but minute features to beconfidently and reliably related to larger-scale characteristics such as velum size,sporangium shape and colouring, sporangium-topping ligule characteristics and siteecology.
In the late 1980s the application of cytological data and SEM imagery in concert withcomprehensive reviews of preserved herbarium material and significantly expanded fieldinvestigations, lead to the detection and description of sterile hybrids in North America.The existence of natural Isoetes hybrids had been identified in Canada 50 years earlier(Jeffrey, 1937) in a publication that was overlooked by researchers for some decadesthereafter (Britton & Brunton, 1992). Independently, Matthews and Murdy (1969)reported probable naturally occurring hybrids in the southeastern USA. Other possiblehybrids were subsequently reported in the eastern USA (Boom, 1979, Taylor et al., 1985).The first sterile hybrid Isoetes to be formally named was I. ×hickeyi W.C.Taylor &N.Luebke (Taylor & Luebke, 1988).
The certain identification of sterile hybrids is taxonomically important. Theirexistence confirms that genetic barriers are in place between previously described andmorphologically similar species. They also indicate the existence of previouslyunsuspected species. Although research has not been undertaken evenly throughout theglobe, over a dozen hybrids have now been formally described in North America (Tayloret al., 1985; Taylor & Luebke, 1988; Britton & Brunton, 1989; Musselman & Knepper,1994; Brunton, 2015), with others named in Japan (Takamiya et al., 1997), Russia(Mochalova et al., 2015) and India (Singh et al., 2018). Undescribed sterile hybrids havebeen detected in western Europe (A. C. Jermy, pers. comm., Prada & Rolleri, 2003;D.F.B., pers. obs.) and northern South America (J. R. Hickey, pers. comm.) and aresuspected in South Africa (Musselman & Roux, 2002), Australia (Tasmania) (pers. obs.)and New Zealand (Hofstra & de Winton, 2016).
A Case Study of Recent DiversityThe combination of cytological data, SEM imagery and hybrid detection has confirmedthe existence of distinct taxa previously thought only to represent clusters of populationsdemonstrating insignificant morphological variation within broadly circumscribedspecies. The widespread North American Isoetes engelmannii s.l provides an excellentexample of how dramatic (and complex) this recent clarification has been. The specificdistinction of diploid I. engelmannii s.str. (Figure 5A and C) and I. valida (Engelm.)
BRUNTON & TROIA: TAXONOMIC RESEARCH INTO ISOETES 313
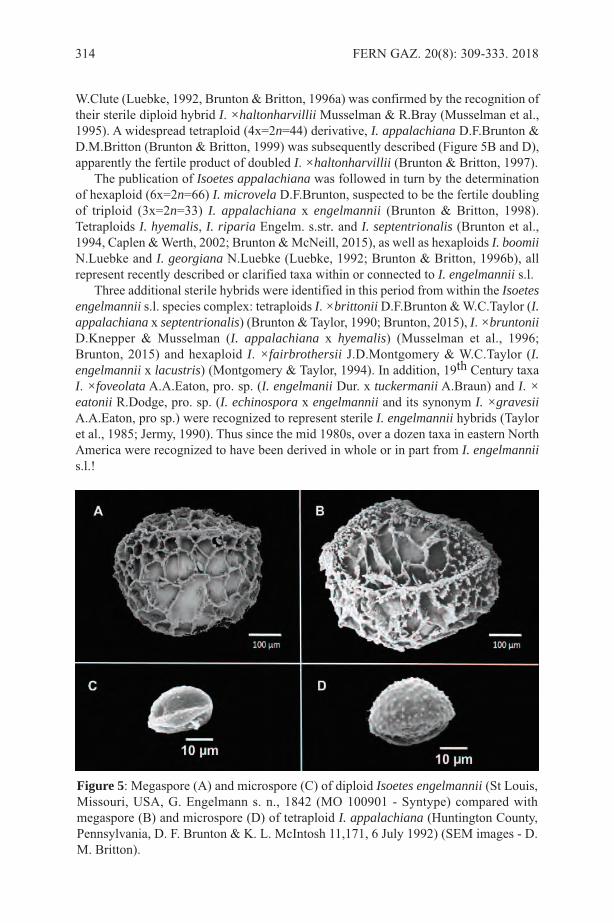
W.Clute (Luebke, 1992, Brunton & Britton, 1996a) was confirmed by the recognition oftheir sterile diploid hybrid I. ×haltonharvillii Musselman & R.Bray (Musselman et al.,1995). A widespread tetraploid (4x=2n=44) derivative, I. appalachiana D.F.Brunton &D.M.Britton (Brunton & Britton, 1999) was subsequently described (Figure 5B and D),apparently the fertile product of doubled I. ×haltonharvillii (Brunton & Britton, 1997).
The publication of Isoetes appalachiana was followed in turn by the determinationof hexaploid (6x=2n=66) I. microvela D.F.Brunton, suspected to be the fertile doublingof triploid (3x=2n=33) I. appalachiana x engelmannii (Brunton & Britton, 1998).Tetraploids I. hyemalis, I. riparia Engelm. s.str. and I. septentrionalis (Brunton et al.,1994, Caplen & Werth, 2002; Brunton & McNeill, 2015), as well as hexaploids I. boomiiN.Luebke and I. georgiana N.Luebke (Luebke, 1992; Brunton & Britton, 1996b), allrepresent recently described or clarified taxa within or connected to I. engelmannii s.l.
Three additional sterile hybrids were identified in this period from within the Isoetesengelmannii s.l. species complex: tetraploids I. ×brittoniiD.F.Brunton & W.C.Taylor (I.appalachiana x septentrionalis) (Brunton & Taylor, 1990; Brunton, 2015), I. ×bruntoniiD.Knepper & Musselman (I. appalachiana x hyemalis) (Musselman et al., 1996;Brunton, 2015) and hexaploid I. ×fairbrothersii J.D.Montgomery & W.C.Taylor (I.engelmannii x lacustris) (Montgomery & Taylor, 1994). In addition, 19th Century taxaI. ×foveolata A.A.Eaton, pro. sp. (I. engelmanii Dur. x tuckermanii A.Braun) and I. ×eatonii R.Dodge, pro. sp. (I. echinospora x engelmannii and its synonym I. ×gravesiiA.A.Eaton, pro sp.) were recognized to represent sterile I. engelmannii hybrids (Tayloret al., 1985; Jermy, 1990). Thus since the mid 1980s, over a dozen taxa in eastern NorthAmerica were recognized to have been derived in whole or in part from I. engelmanniis.l.!
314 FERN GAZ. 20(8): 309-333. 2018
Figure 5: Megaspore (A) and microspore (C) of diploid Isoetes engelmannii (St Louis,Missouri, USA, G. Engelmann s. n., 1842 (MO 100901 - Syntype) compared withmegaspore (B) and microspore (D) of tetraploid I. appalachiana (Huntington County,Pennsylvania, D. F. Brunton & K. L. McIntosh 11,171, 6 July 1992) (SEM images - D.M. Britton).
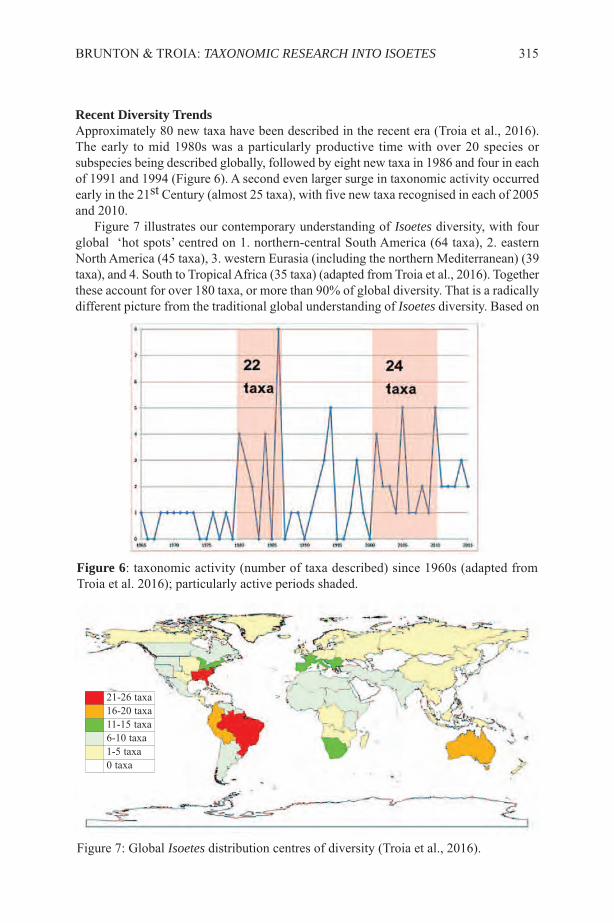
Recent Diversity TrendsApproximately 80 new taxa have been described in the recent era (Troia et al., 2016).The early to mid 1980s was a particularly productive time with over 20 species orsubspecies being described globally, followed by eight new taxa in 1986 and four in eachof 1991 and 1994 (Figure 6). A second even larger surge in taxonomic activity occurredearly in the 21st Century (almost 25 taxa), with five new taxa recognised in each of 2005and 2010.
Figure 7 illustrates our contemporary understanding of Isoetes diversity, with fourglobal ‘hot spots’ centred on 1. northern-central South America (64 taxa), 2. easternNorth America (45 taxa), 3. western Eurasia (including the northern Mediterranean) (39taxa), and 4. South to Tropical Africa (35 taxa) (adapted from Troia et al., 2016). Togetherthese account for over 180 taxa, or more than 90% of global diversity. That is a radicallydifferent picture from the traditional global understanding of Isoetes diversity. Based on
BRUNTON & TROIA: TAXONOMIC RESEARCH INTO ISOETES 315
Figure 6: taxonomic activity (number of taxa described) since 1960s (adapted fromTroia et al. 2016); particularly active periods shaded.
Figure 7: Global Isoetes distribution centres of diversity (Troia et al., 2016).
21-26 taxa16-20 taxa11-15 taxa6-10 taxa1-5 taxa0 taxa

Pfeiffer (1922) and Weber (1922), the traditional five primary centres of diversity wouldhave been identified as western Eurasia (including the northern Mediterranean) (19 taxa- 21% of global diversity), western North America (16 taxa - 17% of global diversity),eastern North America (12 taxa - 13% of global diversity), northern South America (12taxa - 13% of global diversity), and Australia-New Zealand (9 taxa - 10% of globaldiversity).
While the prominence of these contemporary ‘hot spots’ is undoubtedly biasedtowards regions where ‘Isoetologists’ have lived and/ or could most easily access, webelieve the overall pattern at least broadly represents global reality. Recent developmentswithin the major Isoetes diversity areas identified in Figure 7 are summarized below.
Northern-central South America (31% of global diversity)The expansion of known documented Isoetes diversity in northern South America inrecent years is remarkable, with more than a doubling of the number of taxa that wereknown to exist prior to 1930. Most of these are from Brazil, with a particular focus inthe Atlantic Rainforest region where almost 40% of pteridophytes are considered endemic(Prado, Sylvestre & Labiak et al., 2015; Prado, Sylvestre & Pereira, 2015). This totalincludes at least seven Isoetes taxa (Pereira, Mittelbach & Labiak, 2015).
A number of new species represent clarifications of taxa first described (thoughfrequently not legitimately so) in the works of Fuchs-Ekert (1982; 1992). Most new taxaare rare, with several being known from only a single (or handful of) site(s) (Pereira etal., 2016). At least one is known only from 19th Century herbarium specimens (Pereira& Stutzel et al., 2017).
This recent neotropical work was substantially built upon the foundation of taxonomicresearch established by James Hickey and his associates. Their work focused particularlyon species diversity in the northwest of the continent (Hickey, 1984; 1985; 1986a; 1989).The taxonomy of Isoetes in the northwestern portion of the continent remainssubstantially unresolved (Pinto-Zarata et al., 2017), being hampered by the limitedavailability of herbarium material and a confused nomenclatural history. A number ofnew species have been described in this region (e.g. five of nine species reported fromPeru (Hickey, 1994) have been described since 1979 as well as others in the highelevation I. karstenii A.Braun complex (Small & Hickey, 2001)). Undescribed speciesand hybrids remain to be discriminated and documented, however (R. J. Hickey, pers.comm.).
Eastern North America (22% of global diversity) The recent expansion of our knowledge of Isoetes diversity on the eastern side of theNorth American continent commencing with the investigation of species diversity in thesoutheastern USA by Boom (1979; 1982) has been simply explosive. The systematicreview of herbarium collections in the 1980s and 1990s and subsequent fieldinvestigations supported by cytological analysis of morphologically, ecologically and/or distributionally distinctive elements detected within long-established species, has ledto the recognition of numerous new taxa. While 32 taxa (24 species or subspecies andeight named hybrids) were enumerated in the definitive Flora of North America (Tayloret al., 1993), the total now stands at 51 taxa (35 species and subspecies and 16 describedhybrids). A number of additional candidate species, subspecies and hybrids are underactive investigation. This growth represents an almost 60% increase in total NorthAmerican Isoetes diversity over the last 25 years.
316 FERN GAZ. 20(8): 309-333. 2018

Somewhat akin to the geographic concentration witnessed in the Atlantic CoastalRainforest region of South America (see Northern-central South America, above), theAtlantic Coastal Plain of the southeastern USA is particularly species rich and supportsa number of rare and endemic taxa (Reed, 1965; Musselman et al., 2001; Brunton &Britton, 2006; Brunton, 2016).
Considerable attention has been focussed in recent years on species of shallow,ephemeral pools occurring on open, acidic, granitic or sandstone bedrock outcrops (flat-rocks) in the southeastern USA (Matthews & Murdy, 1969; Heafner & Bray, 2005;Brunton, 2016). Comparable species have been described from central and southernAfrica (Hall, 1971; Schlepe & Anthony, 1986; Roux et al., 2009), the Mediterraneanregion (Troia & Raimondo, 2009; Troia & Greuter, 2015b), southern and central Australia(Johnson, 1984; Chinnock, 1993; 1998) and Mexico (Mora-Oivo, 2013) from similaroutcrop pools and ephemeral swales. Such pools are otherwise referred to as erosion pits,!gua, gnammas and vleis. These outcrop taxa include the only known distichous-leavedIsoetes, the diminutive and remarkably similar Isoetes tegetiformans Rury of Georgia,USA (Rury, 1985; Allison, 1993) and I. australis Williams of western Australia(Williams, 1943).
Outcrop taxa are frequently rare and are confined to widely disjunct sites. Manyspecies, such as the aforementioned Isoetes tegetiformans and I. australis or I.melanospora Engelm. of the southeastern USA and I. pusilla C.R.Marsden &R.J.Chinnock of eastern Australia (Entwhisle, 1994), bear remarkable ecological andmorphological similarities (Figure 8) despite being hugely separated geographically.
Western Eurasia (including northern Mediterranean) (19% of global diversity) Pfeiffer (1922) initially enumerated 13 taxa in this region. Although Berthet & Lecocq(1977) made an important contribution to European Isoetes research with their SEM-based review of French taxa, taxonomic reconsiderations in the period began with thework of Carmen Prada (Prada, 1979; 1980; 1983) focussed on the Iberian peninsula.Beyond that, only occasional taxonomic investigations (including karyology) wereundertaken (Schneller, 1982; Britton & Brunton, 1996). With the start of the newMillennium, however, dedicated Isoetes research was initiated in Italy (Troia & Bellini,2001; Cesca & Peruzzi, 2001; Peruzzi et al., 2003), including the review andreconsideration of known diversity and the description of new species (Troia &Raimondo, 2009; Troia & Azzella, 2013).
In this period Prada and associates expanded their studies beyond Spain (Prada &Rolleri, 2005; Rolleri & Prada, 2007). Romero and associates focused on critical Spanishtaxa (Romero et al., 2004; Romero & Real, 2005) while Musselman and associates(Musselman, 2002; Bolin et al., 2008; 2011) studied Isoetes in the eastern part of theMediterranean basin. Troia and associates concentrated their studies on expanding andclarifying previously documented Isoetes taxonomy in the central and easternMediterranean region. This included national syntheses for Italy (Troia & Greuter, 2014,2015a) and Greece (Troia & Greuter, 2015b). In addition, a nomenclatural clarificationof the taxonomically important but perplexing I. longissima-velata complex of theMediterranean has been prepared (Troia & Rouhan, 2018).
The treatment of Isoetes in Flora Europaea (Jermy & Akeroyd, 1993) lists 14 taxa.Jermy also studied aquatic taxa in northwestern Europe, providing the first confirmationof a European sterile hybrid (I. echinospora s.str. x lacustris - A. C. Jermy, pers. comm.).Documentation was also provided at that time for triploid plants of I. echinospora (s.
BRUNTON & TROIA: TAXONOMIC RESEARCH INTO ISOETES 317

318 FERN GAZ. 20(8): 309-333. 2018
Figure 8: Morphologically similar bedrock outcrop pool species Isoetes pusilla ofeastern Australia, LEFT (top to bottom): full plant; megaspore and microspore from thisspecimen (SEM image P. Sokoloff); habitat at Ingram’s Rock, Victoria (Photo: N.Walsh), and I melanospora of the southeastern USA, RIGHT (top to bottom): full plant;megaspore from this specimen (SEM image D. M. Britton); habitat at Mount Arabia,DeKalb County, Georgia (Photo: D. F. Brunton). Scale bar = 100 µm.

str.) in England that were morphologically indistinguishable from the otherwise diploidplants of this species but showed no evidence of hybridization (Rumsey et al., 1993);this unique circumstance for at least a Northern Hemisphere Isoetes species remainsunresolved (F.J Rumsey, pers. comm.).
Vegetation inventories of subalpine Isoetes communities in the Pyrenees Mountainsof France, Spain and Andorra have led to the unexpected discovery and description of I.creussensis J.J.Lazare & S.Riba (Lazare & Riba, 2010; Aymerich & Saez, 2013),probably the duodecaploid (12x=2n=132) detected earlier in this region by Taylor &Hickey (1992). A hexaploid (6x=2n=66) or heptaploid (7x=2n=77) I. ×hickeyi-like sterilehybrid has also has been reported (Taylor & Hickey, 1992) here. A reconsideration ofthe rare and ambiguous (possibly hybrid) taxon I. brochonii M.L.Motelay, and itsrelationship to other Pyrenean taxa, is on-going (D.F.B., pers. obs.).
South to Tropical Africa (17% of global diversity) Pfeiffer (1922) enumerated only four taxa in this region, probably because she waslimited by the availability of voucher material and the restricted nature of botanicalexplorations in the region up to that time. That number of documented taxa has increasedsteadily over the last century, with at least 10 taxa now known. The increase to date isentirely from southern Africa (Wanntorp, 1970; Schelpe & Anthony, 1980; Musselman& Roux, 2002). Most new taxa are found in briefly-flooded ephemeral ponds (vleis) orbedrock outcrop solution pits (Roux et al., 2009).
Tropical African species remain less well known, presumably due to continuinglimited field exploration opportunities and relatively limited herbarium collections. Avariety of amphibious species in ephemeral grassy wetlands and perhaps also in flatrock-like habitats over sandstone bedrock, are known here (Halls, 1971).
Areas of Lesser DiversityA number of areas of lesser diversity have also received particular attention in recentyears. These are described below in descending order of the documented diversity thatis apparent.Australia - New Zealand (16 - 23 species; 8 to 11% of global diversity): Isoetes of thatregion, along with those in a major area of eastern Asia, were enumerated in acomprehensive investigation by Marsden (1979). In that foundational study a number ofnew taxa from Australia were described, several of which are confined to ephemeralopen bedrock pools (Chinnock, 1993). These share remarkably close morphological andecological affinities with flat rock species in the southeastern USA (see Eastern NorthAmerica, above). Approximately double the traditional number of Australian Isoetesknown to Pfeiffer (1922) are now recognized (Chinnock, 1998) - a total of 16 species,with at least one Tasmanian species remaining undescribed (Garrett, 1996). Muchremains to be resolved here, however, with possible hybrids and cryptic species suggestedby abnormal morphological characteristics in several species (referred to by Marsden(1979) as Type I, Type II and Type III megaspores) and multiple ploidy levels reportedfrom other taxa such as apparently apomictic I. muelleri A.Braun s.l. (Chinnock, 1998).
The taxonomy of New Zealand taxa was unresolved in Marsden’s (1979) study andremains so today. There appear to be up to five aquatic taxa represented here, probablyincluding hybrids (Hofstra & de Winton, 2016).
Eastern Asia (Japan/ China/ Korea) (12 taxa; 6% of global diversity): Researchover the last three decades has doubled the known diversity of Isoetes taxa in this region.
BRUNTON & TROIA: TAXONOMIC RESEARCH INTO ISOETES 319

All the newly described taxa are aquatics and most are very rare (Liu et al., 2005), beingknown from one to a handful of sites. Two (including I. ×michinokuana M.Takamiya,M.Watanabe & K.Ono, the first Asian hybrid to be described) result from the especiallycomprehensive morphological and cytological investigations of Japanese Isoetes by M.Takamiya and associates (Takamiya et al., 1997; Takamiya, 2001). Another two newspecies are found on a single island off the coast of Korea (Choi et al., 2008).
Central America - Caribbean (10 taxa; 5% of global diversity): Known diversity inthis floristically transitional area between North and South America is moderate in extentbut demonstrates wide ecological variation and significant endemism (e.g. four speciesconfined exclusively to Mexico). Known diversity has increased in recent years with thedescription of three new endemic species, including I. jamaicensis J.R.Hickey from itsnamesake island (Hickey, 1981), the lowland I. pallida J.R.Hickey of the South AmericanI. triangulaWeber s.l. complex found in southern Mexico (Hickey, 1988), and a bedrockoutcrop pool taxon I. tamaulipana A.Mora-Olivio, A.Menoza-Ruiz &J.G.Martinez-Ávalos, known from a single site in north-eastern Mexico (Mora-Olivio etal., 2013).
India (6-23 taxa; 3 to 11% of global diversity): Ironically, while the endemic Isoetescoromandalina L.f. s.str. was one of the first Isoetes to be described (Linnaeus 1781), itwas the only known member of the genus from India known to Pfeiffer (1922). The 18species now reported from the subcontinent (Shukla et al., 2002, Pantil & Rajput, 2017)were almost all described after 1960 (Troia et al., 2016).
The disposition of a number of recently described taxa and the various subspeciesand varieties within several species complexes (Pant & Shrivastava, 1962; Gena &Bhardwaja, 1984; Shrivastava et al., 1993; Srivastava, 1998; Shukla et al., 2002) iscontroversial and remains unresolved. This is due to uncertainties from the reports ofmultiple and irregular chromosome counts and possibly apomictic reproduction amongstwidely scattered, rarely occurring taxa with distinctions dependent on variablemorphological characteristics. At least six of the 23 described taxa (Pantil & Rajput,2017), however, seem solidly established at a species level (Fraser-Jenkins et al., 2016;Troia et al., 2016). The first Indian sterile hybrid, I. xgopalkrishnae S.K.Singh,P.K.Shukla & N.K.Dubey, has also recently been described (Singh et al. 2018). North Pacific Beringia (ca. 7 taxa; 3% of global diversity) - The Isoetes taxa of the areabetween Far East Asia (Siberia, Russia) and northwest North America (Alaska, USA andYukon, Canada) have received attention in recent years due to their location within thephytogeographically significant Beringian region (Brunton et al., 2015), and one sterilehybrid, I. ×gopalkrishnae S.K.Singh, P.K.Shukla & N.K.Dubey (Singh et al., 2018) hasrecently been described. Britton and Brunton (1999), Mochalova (2006) and Mochalovaet al. (2015) document at least seven taxa (three hybrids) intermixing across this NorthPacific transition zone. One of these, Isoetes ×paratunica D.F.Brunton, O.Mochalova &A.A.Bobrov (I. asiatica (Makino) Makino x I. maritima Underw.) (Mochalova et al.,2015), represents the first hybrid reported between putative parents, that occurpredominantly on separate continents (Asia and North America, respectively). Researchcontinues on the intercontinental relationships of these taxa and their phytogeographicimplications (A. Bobrov, pers. comm.). Kim et al. (2009) and Kim and Choi (2016) haveconducted DNA analyses that further refine the origins and taxonomic distinctions ofthis Beringian complex.
320 FERN GAZ. 20(8): 309-333. 2018

Molecular Investigations Molecular investigations of Isoetes commenced with the Hoot & Taylor (2001)assessment of the value of these data for species determination and the clarification ofIsoetes phylogeny. Although employed in support of some morphologically andcytologically determined specific taxonomic decisions (e.g. Bolin et al., 2011) and toexamine the genetic diversity of particular taxa (Chen et al., 2005), molecular researchto date has largely been applied at a higher level. Molecular data have also been employedin many areas of the world over the last 15 years to investigate genetic diversity withinand between populations of rare and endangered Isoetes. This research has particularimportance for the development of conservation management strategies aimed atestablishing sustainable populations with such species as I. hypsophilaH.vonHandel-Mazzetti (Chen et al., 2005; Li et al., 2012), I. sinensis T.C.Parker (Kanget al., 2005; Li et al., 2012) and I. yunguiensis Q.F.Wang & W.C.Taylor (Chen et al.,2007; Dong et al., 2018) in China, I. coreana Y.H.Chung & H.K.Choi (Kim et al., 2008)in Korea and I. malinverniana V.deCesati & G.DeNotaris (Gentili et al., 2010; Abeli etal., 2017) in Italy. It has been similarly applied for clarifying diversity in undeterminedrare taxa in New Zealand (Hofstra & de Winton, 2016) and with both I. butleriG.Engelmann in the United States (Vander Stelt et al., 2017) and I. durieui J.Bory inItaly (Troia, 2003). Carrying on from fossil and morphological interpretation-basedstudies such as Retallack (1997) and Taylor and Hickey (1992), molecular research hasfocussed on broader questions of phylogeny and the evidence for extinct lineages (Rydin& Wikstrom, 2002; Hoot et al., 2004; Taylor et al., 2004; Larsen & Rydin, 2016; Pereiraet al., 2017). This permitted Hoot et al. (2006), for example, to identify six major well-supported Isoetes clades globally. In recent years Xiang Lui and associates have applieda significant focus of Isoetes molecular research in China on regional origins andprehistoric dispersal patterns (X. Liu, pers. comm.).
Bolin at al. (2018) employed DNA flow cytometry to consider the utility of C Valuesfor the identification of particular species and Freund et al. (2018) combined molecularphylogeny data with morphological documentation to clarify the evolution of particularcharacter states (e.g. the number of corm lobes).
Molecular investigations, both at a high level and in regards to particular taxarepresent a new and powerful tool that will be fundamental to future taxonomic studiesof Isoetes, as they now are to virtually all groups of vascular plants.
Other Areas of InvestigationWhile the limited scope of our review directs focus onto recent taxonomicaccomplishments, it is important to note that significant strides in our understanding ofother aspects of Isoetes were made during this period. The following briefly notes someof the major achievements of other areas of biological investigation.
Vegetation classification involving Isoetes communities was conducted extensivelyin Eurasia, with considerable work being undertaken recently in the Mediterraneanregion. A preliminary synthesis of the Isoetes - related vegetation classification in thisregion was made by Quézel (1998). Studies on the ecology and vegetation science ofIsoetes communities are now available for most (though not all) species around theMediterranean: northwest Africa (Boutin et al., 1982), Azores (Pietsch, 1994), Spain(Romero & Amigo, 1995; Molina, 2005; Molina et al., 2011), France (Médail, 2003; M.Rhazi et al., 2004; L. Rhazi et al., 2009), Italy (Abeli et al., 2014), Sardinia (Bagella &Caria, 2013) and Turkey (Kürschner & Parolly, 1999). These studies are clearly
BRUNTON & TROIA: TAXONOMIC RESEARCH INTO ISOETES 321

conservation-oriented in some cases (e.g. Médail, 2003; M. Rhazi et al., 2004; L. Rhaziet al., 2009; Gentili et al., 2010).
Troia (2016) investigated the dispersal and possible migration routes of some of thecurrent Mediterranean taxa, especially those with African affinities and particularlyconsidering faunal and climatic vectors. Amongst climatic issues, severe wind stormswere speculated to be a distributional vehicle for bedrock outcrop Isoetes taxa in thesouthern USA (Brunton, 2001) as they are for other types of terrestrial and wetlandvascular plants (Matthews et al., 1991; Brunton et al., 2018).
Presumably discouraged by long-standing taxonomic uncertainty in regards to manyspecies, other contemporary investigations of Isoetes biogeography have been relativelyfew. Studies in East Asia employing geological, cytological and molecular data, however,have examined the origins and original dispersal patterns of the genus in that region (Liuet al., 2004; Taylor et al., 2004). Other investigations on a more local scale consideredpossible recent Trans-Pacific dispersal between North America and Asia (Brunton et al.,2015).
Ecological and physiological investigations of Isoetes have been relatively few,despite being initiated early in the recent era in the pioneering studies of Matthews &Murdy (1969) regarding rock outcrop communities in the southern USA. Indeed, thebenefit of considering ecological characteristics as an aid to taxonomic clarification wasdemonstrated earlier with recognition of the importance of substrate chemistry fordistinguishing the morphologically similar ephemerals I. melanopoda Gay & Durieus.str. and I. butleri Engelm. in the southern USA (Taylor et al., 1975). Substrateconsiderations have subsequently been applied to investigations of many aquatic andamphibious species in North America and elsewhere.
Karrfalt and Hunter (1980) examined how the uniquely mucilage-covered sporophyllsof South American Isoetes andicola (Amstutz) L.D.Gomez (Stylites andicolaAmstutz)were literally squeezed out of the alpine vegetation in which they developed by theexpansion of new plant growth, thus aiding the dispersal of sporangia. Otherphysiological and morphometric investigations in this period relate to leaf structure(ligule) functions (Kristen et al., 1982; Sharma & Singh, 1984; Singh, 1984), the earlydevelopment of sporophytes (Hilger et al., 2002) leaf (intrastelar canal) anatomy (Romeoet al., 2000), and the early development of sporophytes (Hilger et al., 2002).
The only extensive in situ ecological investigations of purely aquatic Isoetes to datewere undertaken in central and northern Europe. Margrit Voge conducted underwaterstudies of numerous deep-water populations of I. lacustris at this time (Voge, 1997;2003). She focussed especially on the importance of water quality parameters forcontrolling the health and size of particular populations. The importance of water qualityto population sustainability was also investigated in China by Wen et al. (2003). Morerecently, Chappuis et al. (2015) examined the relative importance of light and organicelements in the stability (or instability) of southern European I. lacustris populations.Sharma (1998) and Sudova et al. (2011) examined the apparently symbiotic role ofmycorrhizal fungi connected with the roots of Isoetes taxa in India and central Europe,respectively.
The most dramatic physiological Isoetes investigations in recent decades resultedfrom the remarkable work of Jon Keeley and associates who determined that the CAMphotosynthetic process typically employed by desert plants was also widely employedby Isoetes (Keeley, 2014). Their field and laboratory studies determined that CO2 uptakeof Isoetes plants was at least substantially through their roots, enabling these plants to
322 FERN GAZ. 20(8): 309-333. 2018

prosper in sterile, otherwise prohibitively challenging habitats (Keeley & Busch, 1984;Keeley & Sandquist, 1991). Those investigations focussed mostly on ephemeral pooltaxa in semi-arid habitats in southern California, USA but also included subalpineAmerican taxa and South American tropical alpine species (Keeley et al., 1994).
Conservation Biology and ManagementConservation concerns are increasingly significant motivators of Isoetes investigations,with many global conservation initiatives focussing on rare species of precarious statusthat are in danger of extinction. Taxa known from only a single population are knownaround the world, including I. pallida in southern Mexico (Hickey, 1988), I.viridimontana M.A.Rosenthal & W.C.Taylor in the northeastern USA (Rosenthal et al.,2014), I. sabatina A.Troia & M.M.Azzella (Troia & Azzella, 2013) in central Italy, I.hallasanensis H-K.Choi, Ch.Kim & J.Jung in Korea (Choi et al., 2008), I. libanoticaMusselman, J.F.Bolin & R.D.Bray in Lebanon (Bolin et al., 2011) and I. junciformisD.F.Brunton & D.M.Britton in the southeastern USA (Brunton & Britton, 1999). Liu etal. (2005) note that all Isoetes taxa in China are in danger of extinction and several easternMediterranean taxa are similarly at risk (Bolin et al., 2011; Troia & Greuter, 2015a).Amphibious bedrock outcrop and ephemeral pool taxa may be the most threatened classof Isoetes globally, these being considered to be of even greater conservation value bysharing habitat with other specialized (often endemic) flora and fauna (Allison, 1993;Bauder & McMillian, 1998; Bagella & Caria, 2013; Ernandes & Marchiori, 2013;Bagella et al., 2016; Brunton, 2016).
FUTURE POSSIBILITIESBased on the rate of nomenclatural innovation in recent years (Figure 6) and the incentiveprovided by new research techniques supplementing traditional databases, we speculatethat 100 or more additional Isoetes taxa will be identified in future years. The richest,perhaps globally most important areas for investigation would appear to be the bedrockoutcrop and ephemeral pool habitats of the USA, Australia and South Africa and the lesswell investigated areas of Tropical Africa and interior South America. In addition topotentially providing taxonomic insights, the remarkably strong ecological andmorphological similarities of geographically widely dispersed bedrock outcrop taxa(Figure 8) suggest an exceptional opportunity for the study of convergent evolution.
Despite no Isoetes being known from the area, a strong case can be made forincluding the Caucasus region of central Asia amongst those having a high potential forfuture taxonomic contributions. The extraordinarily rich biodiversity of this region isknown to support the highest level of Temperate Zone endemism in the NorthernHemisphere (Batsatsachvii et al., 2013).
Molecular clarification of the subtle and even cryptic relationships within Isoetesspecies complexes could have major conservation management implications in manyparts of the world. Again, this is especially true in regards to bedrock outcrop andephemeral wetland taxa.
Particular regions such as India, central and northern South America, South toTropical Africa, central Asia and the eastern Mediterranean appear to be particularly inneed of the initiation or continuation of systematic cytological and molecular taxonomicinvestigations and clarification. That applies as well to a number of unusually challengingspecies complexes including Isoetes muelleri s.l. in Australia, I. coromandelina s.l. inIndia, I. longissima s.l. in the Mediterranean region of Europe, I. karstenii s.l. in South
BRUNTON & TROIA: TAXONOMIC RESEARCH INTO ISOETES 323

324 FERN GAZ. 20(8): 309-333. 2018
America, and both I. riparia s.l. and I. melanopoda s.l. in North America.Although considerable improvements have been seen in recent decades in the growth
of Isoetes herbarium collections globally, more systematic gathering of high qualitymaterial is needed to facilitate future physiological and molecular investigations,particularly with morphologically challenging species groups. This is especiallyimportant in geographic regions (identified above) which traditionally have beendeficient in preserved plant material. Similarly, cytological determinations remainoutstanding for over half of the described taxa (Troia et al., 2017). These determinationsare fundamental to understanding polyploid taxa and interspecfic hybrid relationships.And of course, the application of rapidly evolving and increasingly sophisticatedmolecular investigative tools is required in a systematic basis, not just in regards toindividual taxa or species complexes.
Despite the laudable achievements of pioneering 19th and 20th Century investigatorsand the remarkable advances of recent decades, the potential for the identification ofnew taxa remains high. Indeed, one could accurately state that such potential exitsbecause of those pioneering efforts. Regardless, it is clear that previously undetectedIsoetes taxa can be expected just about anywhere in the world where these curious - and‘crazy’ - plants are to be found.
ACKNOWLEDGMENTSOur thanks to W. Carl Taylor, Botany, Smithsonian Institution, Washington, DC, USAfor providing background information and encouragement for this project. We appreciatepermission for use of the SEM images produced by Paul Sokoloff, Botany, CanadianMuseum of Nature, Ottawa, Ontario, Canada (with Daniel Brunton), the late Donald M.Britton, University of Guelph, Guelph, Ontario, Canada and Carmela Di Liberto,University of Palermo, Italy (with Angelo Troia). We are grateful as well to AlexanderBobrov, I. D. Papanin Institute for Biology of Inland Waters, Borok, Russia, R. JamesHickey, University of Miami, Ohio, USA and Xiang Lui, Laboratory of Plant Systematicsand Evolutionary Biology, Wuhan University, Wuhan, China for their contributions ofregional information. Neville Walsh, Royal Botanic Gardens, Melbourne, Australia alsoprovided regional information and photographic material and offered helpful commentson the treatment of Australian taxa. We are particularly appreciative of the invaluableobservations that James R. Hickey, Miami University, Oxford Ohio USA. Jim possiblyhas the longest continuous record of and perspective on Isoetes research in the world.And finally our sincere thanks to Editor Bridget Laue, without whose patience andpersistence this paper would never have seen the light of day.
REFERENCESABELI, T., ORSENIGO, S., ARDENGHI, N.M.G., LUCASSEN, E.C.H.E.T. &
SMOLDERS, A.J.P. 2014. Hydrochemical characterization of a stand of thethreatened endemic Isoetes malinverniana. Amer. Fern J. 103: 241-244.
ALLISON, J.R. 1993. Recovery plan for three granite outcrop plant species: black-spored quillwort (Isoetes melanospora), mat-forming quillwort (Isoetestegetiformans), and little amphianthus (Amphianthus pusillus). United States Fishand Wildlife Service, Jackson, Mississippi.
AYMERICH, P. & SAEZ, L. 2013. Noves dades pteridològiques dels Pirineus catalans.Orsis 27: 175-194.
BAGELLA, S. & CARIA, M.C. 2013. Sensitivity of ephemeral wetland swards withIsoetes histrix Bory to environmental variables: implications for the conservation of

Mediterranean temporary ponds. Aquatic Conservation 23: 277-290. BAGELLA, S., GASCON, S., FILIGHEDDU, R., COGONI, A. & BOIX, D. 2016.
Mediterranean temporary pools: new challenges from a neglected habitat.Hydrobiologia 782: 1-10.
BAKER, J.G. 1880. A Synopsis of the species of Isoetes. J. Bot. Br. Foreign 9 65-70,105-110.
BATSATSACHVII, K., SCHATZ, G.E. & SCHULKINA, T. (Eds) 2013. Caucasus PlantInitiative: A Regional Plant Conservation Strategy.https://www.bgci.org/files/Worldwide/News/2013/caucasus_strategy.pdf
BAUDER, E.T. & MCMILLAN, S. 1998. Current distribution and historical extent ofvernal pools in southern California and northern Baja California, Mexico. In:WITHAM, C.W., BAUDER, E.T., BELK, D., FERREN, W.R. & ORNOFF, R. (Eds)Ecology, Conservation and Management of Vernal Pools, pp. 56-70. Proceedingsfrom a 1996 Conference. California Native Plant Society, Sacramento.
BERTHET, P. & LECOCQ, M. 1977. Morphologie sporale des espèces françaises dugenre Isoetes L. Pollen & Spores 19: 329-359.
BOLIN, J.F., BRAY, R.D., KESKIN, K. & MUSSELMAN, L.J. 2008. The genus IsoetesL. (Isoetaceae, Lycophyta) in South-Western Asia. Turk. J. Bot. 32: 447-457.
BOLIN, J.F., BRAY, R.D. & MUSSELMAN, L.J. 2011 A new species of diploidquillwort (Isoetes, Isoetaceae, Lycophyta) from Lebanon. Novon 21: 295-298.
BOLIN, J.F., HARTWIG, C.L., SCHAFRAN, P. & KOMARNYTSKY, S. 2018.Application of DNA flow cytometry to aid species delimitation in Isoetes. Castanea83: 38- 47.
BOOM, B.M. 1979. Systematic Studies of the Genus Isoetes in the Southeastern UnitedStates. Master’s Thesis, University of Tennessee, Knoxville.
BOOM, B.M. 1982. Synopsis of Isoetes in the southeastern United States. Castanea 47:38-59.
BOUTIN, C., LESNE, L. & THIÉRY, A. 1982. Ecologie et typologie de quelques marestemporaires à isoètes d’une région aride du Maroc occidental. Ecol. Medit. 8: 31–56.
BRAUN, A. 1846. Uber die nordamericanischen Isoetes Arten. Flora oder Bot. Zeit.(Regensburg) 12: 177-180.
BRITTON, D.M. & BRUNTON, D.F. 1989. A new Isoetes hybrid (Isoetes echinosporax riparia) for Canada. Canad. J. Bot. 67: 2995-3002.
BRITTON, D.M. & BRUNTON, D.F. 1992. Isoetes ×jeffreyi, hyb. nov.: a new Isoetes(I. macrospora x riparia) from Quebec, Canada. Canad. J. Bot. 70: 447-452.
BRITTON, D.M. & BRUNTON, D.F. 1996. Isoetes ×pseudotruncata, a new triploidhybrid from western Canada and Alaska. Canad. J. Bot. 74: 51-59.
BRITTON, D.M. & BRUNTON, D.F. 1999. Isoetes in Alaska and the Aleutians. Amer.Fern J. 79: 133-141.
BRUNTON, D.F. 2001. Quillwort dispersal: which way is the wind blowing? Chinquapin9: 20; 24-26.
BRUNTON, D.F. 2015. Key to the quillworts (Isoetes: Isoetaceae) of the southeasternUnited States. Amer. Fern J. 105: 86-100.
BRUNTON, D.F. 2016. Flat Rock Quillwort, Isoetes graniticola sp. nov. (Isoetaceae), anew lycophyte from the southeastern United States. Rhodora 118: 261-275.
BRUNTON, D.F. & BRITTON, D.M. 1996a. The status, distribution and identificationof Georgia Quillwort (Isoetes georgiana; Isoetaceae). Amer. Fern J. 86: 105-113.
BRUNTON & TROIA: TAXONOMIC RESEARCH INTO ISOETES 325

BRUNTON, D.F. & BRITTON, D.M. 1996b. Taxonomy and distribution of Isoetesvalida (Isoetaceae). Amer. Fern J. 86: 16-25.
BRUNTON, D.F. & BRITTON, D.M. 1997. Appalachian Quillwort, (Isoetesappalachiana, sp. nov.; Isoetaceae), a new pteridophyte from the eastern UnitedStates. Rhodora 99: 118-133.
BRUNTON, D.F. & BRITTON, D.M. 1998. Isoetes microvela (Isoetaceae), a newquillwort from the southeastern coastal plain of the United States. Rhodora 100: 261-275.
BRUNTON, D.F. & BRITTON, D.M. 1999. Rush Quillwort (Isoetes junciformis, sp.nov.), a new pteridophyte from southern Georgia. Amer. Fern J. 79: 187-197.
BRUNTON, D.F. & BRITTON, D.M. 2006. Isoetes melanopoda spp. silvatica (subsp.nov.), a new quillwort (Isoetaceae) from eastern North America. Castanea 71: 15-30.
BRUNTON, D.F., BRITTON, D.M. & TAYLOR, W.C. 1994. Isoetes hyemalis, sp. nov.(Isoetaceae); a new quillwort from the southeastern United States. Castanea 59 (1):12-21.
BRUNTON, D.F. & MCNEILL, J. 2015. Status, distribution and nomenclature ofNorthern Quillwort, Isoetes septentrionalis (Isoetaceae), in Canada. Canad. Field-Nat. 129: 174-180.
BRUNTON, D.F., MOCHALOVA, O.A. & BOBROV, A.A. 2015. Isoëtes (Isoëtaceae)in the northern Pacific: diversity, taxonomy and distribution. In: Problems oftaxonomy and geography of aquatic plants, pp. 90-91. Proceedings of theInternational Conference (Borok, Russia, 21-24 October, 2015),Yaroslavl, Filigran.
BRUNTON, D.F. & TAYLOR, W.C. 1990. Isoëtes ×brittonii hyb. nov. (Isoëtaceae): Anaturally occurring hybrid (I. engelmannii ´ I. riparia) in the eastern United States.Amer. Fern J. 80: 82-89.
CAPLEN, C.A. & WERTH, C.R. 2000. Isozymes of the Isoetes riparia complex, II.Ancestry and relationships of polyploids. Syst. Bot. 25: 260-280.
CESCA, G. & PERUZZI, L. 2001. Isoetes (Lycophytina, Isoetaceae) with terrestrialhabitat in Calabria (Italy). New karyological and taxonomical data. Flora Medit. 11:303-309
CHAPPUIS, E., LUMBRERAS, L., BALLESTEROS, E. & GACIA, E. 2015.Deleterious interaction of light impairment and organic matter enrichment on Isoeteslacustris (Lycopodiophyta, Isoetales). Hydrobiologia 760: 145-158.
CHEN, J.-M., LUI, X., WANG, J.-Y., GITURU, W.R. & WANG, Q.F. 2005. Geneticvariation within the endangered quillwort Isoetes hypsophila (Isoetacae) in China asevidenced by ISSR analysis. Aquatic Bot. 82: 89-98.
CHINNOCK, R.J. 1993. Notes on Isoetes and Tmesipteris in Victoria. Muelleria 8: 57-60.
CHINNOCK, R.J. 1998. Isoetaceae. In: Flora of Australia, Volume 4. AustralianBiological Resources Study, Department of the Environment and Energy, Canberra.
CHOI, H, JUNG, J. & KIM, C. 2008. Two new species of Isoetes (Isoetaceae) from JejuIsland, South Korea. J. Pl. Biol. 51: 354-358.
DURIEU DE MAISONNEUVE. 1864. Notes sur quelques espèces nouvelles d’Isoetesde l’Amérique du Nord et de l’Australie. Bull. Soc. Bot. France 11(4): 100-105.
EATON, A.A. 1900. The genus Isoetes in New England, In: Fernwort Papers, pp. 1-16.W. N. Clute & Co., Binghamton.
ENGELMANN, G. 1867. Isoetaceae. In: GRAY, A. Manual of the Botany of the
326 FERN GAZ. 20(8): 309-333. 2018

Northern United States, 5th Edition, pp. 698-700. Iverson, Blakeman & Company,New York.
ENGELMANN, G. 1882. The Genus Isoetes in North America. Trans. Acad. Sci. St.Louis 4: 358-390.
ENTWISLE, T.J. 1994. Ferns and allied plants (Psilophyta, Lycopodiophyta,Polypodiophyta). In: WALSH, N.G. & ENTWISLE, T.J. (Eds), Flora of Victoria Vol.2, Ferns and Allied Plants, Conifers and Monocotyledons. Inkata Press, Melbourne.
ERNANDES, P. & MARCHIORI, S. 2013. Mediterranean temporary ponds in Puglia(South Italy): a “joyau floristique” to protect. Acta Bot. Gallica 160: 53-64.
FERRARINI, E., CIAMPOLINI, F., PICHI SERMOLLI, R.E.G. & MARCHETTI, D.1986. Iconographia Palynologica Pteridophytorum Italiae. Webbia 40: 1-202.
FRASER-JENKINS, C.R., GANDHI, K.N., KHOLIA, B.S. & BENNIAMIN, A. 2016.Isoeteaceae, In: An annotated checklist of Indian pteridophytes Part I (Lycopodiaceaeto Thelypteridaceae), pp. 54–60. Bishen Singh Mahendra Pal Singh, Dehradun, India.
FREUND, F.D., FREYMAN, W. & ROTHFELS, C. 2018. Inferring the evolutionaryreduction of corm lobation in Isoetes using Bayesian model-averaged ancestral statereconstruction. Amer. J. Bot. 105: 275-286.
FUCHS-ECKERT, H.P. 1982. Zur heutigen Kenntnis von Vorkommen und Verbreitungder sudamerianischen Isoetes-Arten. Proceedings of the Koninklijke NederlandseAkademie va Weetenschappen, Series C 85: 2015-260.
FUCHS-ECKERT, H.P. 1992. Supplementum ad indicem Isoetalium. Bionature 12: 99-159.
GARRETT, M. 1996. The Ferns of Tasmania, their ecology and distribution. TasmanianForest Research Council, Hobart.
GENA, C.B. & BHARDWAJA, T.N. 1984. Three new species of genus Isoëtes fromRajasthan. J. Bombay Nat. Hist. Soc. 81: 165-168.
GENTILI, R., ABELI, T., ROSSI, G., LI, M., VAROTTO, C. & SGORBATI, S. 2010.Population structure and genetic diversity of the threatened quillwort Isoetesmalinverniana and implication for conservation. Aquatic Bot. 93: 147-152.
HALLS, J.B. 1971. Observations on Isoetes in Ghana. Bot. J. Linn. Soc. 64: 117-139.HEAFNER, K.D. & BRAY, R.D. 2005. Taxonomic reassessment of North American
granite outcrop Isoetes species with emphasis on vegetative morphology and I.piedmontana (Pfeiffer) Reed sensu lato. Castanea 70: 204-221.
HICKEY, R.J. 1981. A new Isoetes from Jamaica. Amer. Fern J. 71: 69 - 74.HICKEY, R.J. 1984. Chromosome numbers of Neotropical Isoetes. Amer. Fern J. 74: 9-
13.HICKEY, R.J. 1985. Revisionary Studies of Neotropical Isoetes. University of
Connecticut, Storrs University of Adelaide [Phd Thesis].HICKEY, R.J. 1986a. The early evolutionary and morphological diversity of Isoetes,
with descriptions of two new Neotropical species. Syst. Bot. 11: 304-321.HICKEY, R.J. 1986b. Isoetes megaspore surface morphology: nomenclature, variation
and systematic importance. Amer. Fern J. 76: 1-16. HICKEY, R.J. 1988. Isoetes pallida, a new species from Mexico. Amer. Fern J. 78: 35-
36.HICKEY, R.J. 1989. A new species of Isoetes from Territorio Federal Amazonas,
Venezuela. Ann. Missouri Bot. Gard. 76: 1160-1162.HICKEY, R.J. 1994. Pteridophyta of Peru, Part VI - Family 28. Isoetaceae. Fieldiana 34:
88-97.
BRUNTON & TROIA: TAXONOMIC RESEARCH INTO ISOETES 327

HILGER, H.H., WEIGEND, W. & FREY, W. 2002. The gametophyte-sporophytejunction in Isoetes boliviensisWeber (Isoetaceae, Lycopodiophyta). Phyton 42: 149-157.
HOFSTRA, D. & DE WINTON, M. 2016. Geographic scales of genetic variationamongst Isoëtes in New Zealand. Aquatic Bot. 131: 28-37.
HOOT, S.B., NAPIER, N.S. & TAYLOR, W.C. 2004. Revealing unknown or extinctlineages with Isoetes (Isoetaceae) using DNA sequences from hybrids. Amer. J. Bot.91: 899-904.
HOOT, S.B. & TAYLOR, W.C. 2001. The Utility of nuclear ITS, a LEAFY homologintron, and chloroplast atpB-rncL spacer region data in phylogenetic analyses andspecies delimitation in Isoetes. Amer. Fern J. 91: 166-177.
HOOT, S.B., TAYLOR, W.C. & NAPIER, N.S. 2006. Phylogeny and biogeography ofIsoetes (Isoetaceae) based on nuclear and chloroplast DNA sequence data. Syst. Bot.31: 449-460.
JEFFREY, E.C. 1937. The cytology of a heterosporous Isoetes. Bot. Mag. (Tokyo) 521:203-209.
JERMY, A.C. 1990. Isoetaceae, In: KRAMER, K.U. & GREEN, P.S. (Eds) The Familiesand Genera of Vascular Plants Volume 1: Pteridophyes and Gymnosperms, pp. 24-31. Springer-Verlag, Berlin.
JERMY, A. C. & AKEROYD, J. R. 1993. Isoetes L. In: TUTIN, T.G., BURGES, N.A.,CHATER, O., EDMONDSON, J.R., HEYWOOD, V. MOOR, D., VALENTINE,D.H., WALTERS, S.M. & WEBB, D.A. Flora Europaea, Volume 1 Psilotaceae toPlatanaceae, 2nd ed., pp. 6-7. Cambridge University Press, Cambridge.
JOHNSON, E.R.L. 1984. Taxonomic revision of Isoetes L. in Western Australia. J. Roy.Soc. Western Australia 66, Part 4: 28-43.
KARRFALT, E.E. & HUNTER, D.M. 1980. Notes on the natural history of Stylitesgemmifera. Amer. Fern J. 70: 69-72.
KEELEY, J.E. 2014. Aquatic CAM Photosynthesis: a brief history of its discovery.Aquatic Bot. 118: 38-44.
KEELEY, J.E. & BUSCH, G. 1984. Carbon assimilation characteristics of the aquaticCAM plant Isoetes howellii. Pl. Physiol. 76: 525-530.
KEELEY, J.E. & SANDQUIST, D.R. 1991. Diurnal photosynthesis Cycle in CAM andnon-CAM seasonal-pool aquatic macrophytes. Ecology 72: 716-727.
KEELEY, J.E., DEMASON, D.A., GONZALEZ, R. & MARKHAM, K.R. 1994.Sediment-based nutrition in tropical alpine Isoetes. In: RUNDEL, P.W., SMITH, A.P.& MIENZER, F.C. (Eds) Tropical Alpine Environments, pp.167-194. CambridgeUniversity Press, Cambridge.
KIM, C. & CHOI, H.-K. 2016. Biogeography of North Pacific Isoetes (Isoetaceae)inferred from nuclear and chloroplast DNA sequence data. J. Pl. Biol. 59: 386-396.
KIM, C., NA, N.R., SHIN, H. & CHOI, H.-K. 2009. Systematic evaluation of IsoetesasiaticaMakino (Isoetaceae) based on FLAP, nrITS and chloroplast DNA sequences.J. Pl. Biol. 52: 501-510.
KOTT, L.1980. Chromosome numbers for Isoetes in northeastern North America. Canad.J. Bot. 58: 980-984.
KOTT, L. & BRITTON, D.M. 1983. Spore morphology and taxonomy of Isoetes innortheastern North America. Canad. J. Bot. 61: 3140-3164.
KRISTEN, U., LIEBEZEIT, G. & BIEDERMANN, M. 1982. The ligule of Isoeteslacustris: ultrastructure, mucilage composition. and a possible pathway of secretion.
328 FERN GAZ. 20(8): 309-333. 2018

Ann. Bot. 49: 569-584.KÜRSCHNER, H. & PAROLLY, G. 1999. On the occurrence of Isoëtes histrix in the
Menderes Massif of western Turkey - a synecological study and the first record ofan Isoëtion community for Turkey. Bot. Jahrb. Syst. 121: 423-451.
LARSÉN, E. & C. RYDIN. 2016. Disentangling the phylogeny of Isoetes (Isoetales),using Nuclear and Plastid Data. Int. J. Pl. Sci. 177: 157–174.
LAZARE, J.J. & RIBA, S. 2010. Les Isoètes de l’Estany de Creussans (Andorre). J. Bot.Soc. France 30: 19-26.
LINNAEUS, C. 1781. Supplementum Plantarum Systematis Vegetabilium EditionisDecimae Tertiae, Generum Plantarum Editiones Sextae, et Specierum PlantarumEditionis Secundae. Brunsvigne.
LIU, X., GITURU, W.R. & WANG, Q.-F. 2004. Distribution of basic diploid andpolyploid species of Isoetes in East Asia. J. Biogeogr. 31: 1239-1250.
LIU, X., WANG, J.-Y. & WANG, Q.-F. 2005. Current status and conservation strategiesfor Isoetes in China: a case study for the conservation of threatened aquatic plants.Oryx 39: 335-338.
LUEBKE, N.T. 1992. Three new species of Isöetes for the southeastern United States.Amer. Fern J. 82: 23-26.
MACLUF, C.C., MORBILLI, M.A. & GIUDICE, G.E. 2006. Microspore morphologyof Isoetes Species (Lycophyta) from southern South America. Bot. Review 72: 121-124.
MANTON, I. 1950. Problems of cytology and evolution in the Pteridophyta. CambridgeUniversity Press, New York.
MARSDEN, C.R. 1979. Morphology and Taxonomy of Isoetes in Australia, India, North-East and South-East Asia, China and Japan. Department of Botany, University ofAdelaide [Phd Thesis].
MATTHEWS, J.F., FAIRCLOTH, W.R. & ALLISON, J.R. 1991. Portulaca bilobaUrban(Portulacaeae), a species new to the United States. Syst. Bot. 130: 53-61.
MATTHEWS, J.F. & MURDY, W.M. 1969. A study of Isoetes common to the graniteoutcrops of the southeastern Piedmont, United States. Bot. Gaz. 130: 53-61.
MÉDAIL, F. 2003. La colonisation des mares temporaires méditerranéennes par lesligneux menace la survie à court terme d’une ptéridophyte très rare et menacée,l’isoète sétacé (Isoetes setacea Lam.). Ecol. Medit. 29: 251-252.
MOCHALOVA, O.A. 2006. The genus Isoetes (Isoetaceae) in the northern Far East ofAsia. Bot. Zhurn. (Moscow & Leningrad) 91: 94-98 [in Russian].
MOCHALOVA, O.A., BOBROV, A.A. & BRUNTON, D.F. 2015. Isoetes in Kamchatka(northern Russian Far East) with description of a new hybrid I. ×paratunica (I.asiatica x I. maritima). Amer. Fern J. 105: 101-112.
MOLINA, J.A. 2005. The vegetation of temporary ponds with Isoetes in the IberianPeninsula. Phytocoenologia 35: 219-230.
MOLINA, J.A., LUMBRERAS, A., GALLARDO, T., AGOSTINELLI, E.,CASERMEIRO, M.A. & PRADA, C. 2011. Isoetes distribution pattern in aMediterranean vernal pool system. Acta Bot. Gallica 58: 27-36.
MONTGOMERY, J.D. & TAYLOR, W.C. 1994. Confirmation of a hybrid Isoetes fromNew Jersey. Amer. Fern J. 84: 115-120.
MORA-OLIVO, A., MENDOZA-RUIZ, A. & MARTÍNEZ-ÁVALOS, J.-G. 2013.Isoetes tamaulipana (Isoetaceae), a new species from Mexico. Phytotaxa 267: 113-120.
BRUNTON & TROIA: TAXONOMIC RESEARCH INTO ISOETES 329

MOTELAY, M.L. 1883. Monographie des Isoëteae. Actes Soc. Linn. Bordeaux 36:309‐404.
MOTELAY, M.L. 1892. Isoetes brochoni Motelay, spec. nov. Actes Soc. Linn. Bordeaux45: 45-48 + Plate II.
MUSSELMAN, L.J. 2002. Ornamentation of Isoetes (Isoetaceae, Lycophyta)microspores. Bot. Review 68: 474-487.
MUSSELMAN, L.J., BRAY, R.D. & KNEPPER, D.A. 1996. Isoetes ×bruntonii (Isoetesengelmannii ´ I. hyemalis), A new hybrid quillwort from Virginia. Amer. Fern J. 86:8-15.
MUSSELMAN, L.J. & KNEPPER, D.A. 1994. Quillworts of Virginia. Amer. Fern J. 84:48-68.
MUSSELMAN, L.J., KNEPPER, D.A., BRAY, R.D., CAPLEN, C.A. & BALLOU, C.1995. A new Isoetes hybrid from Virginia. Castanea 60: 245-254.
MUSSELMAN, L.J. & ROUX, J.P. 2002. Isoetes toximontana (Isoetaceae), a newquillwort with green megaspores from Northern Cape of South Africa. Novon 12:504-507.
MUSSELMAN, L.J., TAYLOR, W.C. & BRAY, R.D. 2001. Isoetes mattaponica(Isoetaceae), a new diploid quillwort from freshwater tidal marshes of Virginia.Novon 11: 200-2004.
PANT, D.D. & SRIVASTAVA, G.K. 1962. The genus Isoetes in India. Proc. Natl. Inst.Sci. India 28 (Part B): 242-280.
PATIL, S.M. & RAJPUT, K.S. 2017. The genus Isoetes from India: An Overview. PlantScience Today 4: 213-226.
PEREIRA, J.B.S., LABIAK, P.H., STUTZEL, T. & SHULTZ, C. 2017. Origin andbiogeography of the ancient genus Isoetes with focus on the neotropics. Bot. J. Linn.Soc. 185: 253-271.
PEREIRA, J.B.S., MITTELBACH, M. & LABIAK, P. 2015. Studies on chromosomenumbers and spore size in Brazilian Isoetes. Amer. Fern J. 105: 226-237.
PEREIRA, J.B.S., SALINO, A., ARRUDA, A. & STUTZEL, T. 2016. Two new speciesof Isoetes (Isoetaceae) from northern Brazil. Phytotaxa 272: 141-148.
PEREIRA, J.B.S., STUTZEL, T. & SCHULZ, C. 2017. Isoetes nana, a new species fromthe coastal mountains of southeastern Brazil. PhytoKeys 89: 91-107.
PERUZZI, L., CESCA, G. & PUNTILLO, D. 2003. Isoetes (Isoetaceae), Ophioglossumand Botrychium (Ophioglossaceae) in Calabria (S. Italy): more karyological andtaxonomical data. Caryologia 56: 359-363.
PFEIFFER, N.E. 1922. Monograph on the Isoetaceae. Trans. Acad. Sci. St. Louis 9: 79-233.
PIETSCH, W. 1994. Isöetes azorica Durieu ex Milde - ein Endemit der Azoren -Vegetations-und Standortsverhältnisse, Gefährdung und Schutzmaßnahmen.Phytocoenologia 24: 649-665.
PIGG, K.B. 2001. Isoetalean lycopsid evolution: from the Devonian to the present. Amer.Fern J. 91: 99-114.
PINTO-ZÁRATE, J.H., OROZCO-RINCÓN, E. & MARQUÍNEZ-CASAS, X. 2017.Panorama de la investigación sobre los Isoëtes de Colombia (Isoëtaceae,Lycopodiopsida). In: MORENO-GAONA, D.A. (Ed) Memorias IX CongresoColombiano de Botánica, 30 de julio al 3 agosto de 2017, Tunja-Boyacá. Ciencia enDesarrollo 8 (1, Supl. Esp.): 304-305, Asociación Colombiana de Botánica yUniversidad Pedagógica y Tecnológica de Colombia.
330 FERN GAZ. 20(8): 309-333. 2018

PRADA, C. 1979. Estudio de la anatomia foliar de las eespecies Espanolas del generoIsoetes L. Lagascalia 9: 107-113.
PRADA, C. 1980. Estudios palinologicos y cromosomicos en las especies Espanolas delgenero Isoetes (Isoetaceae). Palinologia 1: 211-225.
PRADA, C. 1983. El genero Isoetes L. en la Peninsula Iberica. Acta Bot. Malac. 8: 73-100.
PRADA, C. & ROLLERI, C.H. 2003. Caracteres diagnosticos foliares en taxonesIbericos de Isoetes L. (Isoetaceae, Pteridophyta). Anales Jard. Bot. Madrid 60: 371-386.
PRADA, C. & ROLLERI, C.H. 2005. A new species of Isoetes (Isoetaceae) from Turkey,with a study of microphyll intercellular pectic protuberances and their potentialtaxonomic value. Bot. J. Linn. Soc. 147: 213-228.
PRADO, J., SYLVESTRE, L., LABIAK, S. H., WINDISCH, P.G., SALINO, A.,BARROS, I.C.L., HIRAI, R.Y., ALMERIA, T.E., SANTIAGO, A.C.P., KLIELING-RUBIO, M.A., N. PEREIRA, A., OLLGAARD, B., RAMOS, C.G.V., MICKEL, J.T.,DIETRICH, V.A.O., MYNSSEN, C.M., SCHWARTSBURD, P., CONDACK, J.P.S.,PEREIRA, J.B.S. & MATOS, F.B. 2015. Diversity of ferns and lycophytes in Brazil.Rodriguesia 66: 1073-1083.
PRADO, J., SYLVESTRE, L. & PEREIRA, J.B.S. 2015. Isoetaceae, In: Lista de Espéciesda Flora do Brasil. Jardim Botânico do Rio de Janeiro.<http://floradobrasil.jbrj.gov.br/jabot/floradobrasil/FB119852 [accessed 22 March2018]
PUNT, W., HOEN, P.P, BLACKMORE, S., NILSSON, S. & LE THOMAS, A. 2007.Glossary of pollen and spore terminology. Review of Palaeobotany and Palynology143: 1–81.
QUEZEL, P. 1998. La végétation des mares transitoires à Isoetes en régionméditerranéenne, intérêt patrimonial et conservation. Ecol. Medit. 24: 111-117.
REED, C.F. 1965. Isoetes in the southeastern United States. Phytologia 12: 369-400.RETALLACK, G.J. 1997. Earliest Triassic origin of Isoetes and quillwort evolutionary
radiation. J. Paleontology 71: 500-521.RHAZI, L., FRILLAS, P., RHAZI, M. & AZNAR, J.-C. 2009. Ten-year dynamics of
vegetation in a Mediterranean temporary pool in western Morocco. Hydrobiologia634: 185-194. https://doi.org/10.1007/s10750-009-9893
RHAZI, M., GRILLAS, P., CHARPENTIER, A. & MEDAIL, F. 2004. Experimentalmanagement of Mediterranean temporary pools for conservation of the rare quillwortIsoetes setacea. Biol. Conservation 118: 675-684.
ROLLERI, C. & PRADA, C. 2007. Caracteres diagnosticos foliares en Isoetes(Pteridophyta, Isoetaceae). Ann. Missouri Bot. Gard. 94: 202-235.
ROMEO, D., TROIA, A., BURGARELLA, C. & BELLINI, E. 2000. Casparian stripesin the leaf intrastelar canals of Isoetes durieui Bory, a Mediterranean terrestrialspecies. Ann. Bot. (Oxford) 86: 1051-1054.
ROMERO, M.I. & AMIGO, J. 1995. Autecology and distribution of Isoetes longissimumin Europe. Nordic J. Bot. 15: 563-566.
ROMERO, M.I., AMIGO, J. & RAMIL, P. 2004. Isoetes fluitans sp. nov.: the identityof Spanish plants of ‘I. longissimum’. Bot. J. Linn. Soc 146: 231-236.
ROMERO, M.I. & REAL, C. 2005. A morphometric study of three closely related taxain the European Isoetes velata complex. Bot. J. Linn. Soc. 148: 459-464.
ROSENTHAL, M.A., ROSENTHAL, S.E., JOHNSON, G., TAYLOR, W.C. &
BRUNTON & TROIA: TAXONOMIC RESEARCH INTO ISOETES 331

ZIMMER, E. 2014. Isoetes viridimontana: a previously unrecognized quillwort fromVermont, USA. Amer. Fern J. 104: 7-15.
ROUX, J.P., HOPPER, S.D. & SMITH, R.J. 2009. Isoetes eludens (Isoetaceae), a newendemic species from Kamiesburg, Northern Cape, South Africa. Kew Bull. 64: 123-126.
RUMSEY, F.J., THOMPSON, P. & SHEFFIELD, E. 1993. Triploid Isoetes echinospora(Isoetaceae: Pteridophyta) in northern England. Fern Gaz. (UK) 14: 215-221.
RURY, P.M. 1985. A new and unique, mat-forming Merlin’s-grass (Isoetes) fromGeorgia. Amer. Fern J. 68: 102-104.
RYDIN, C. & WIKSTROM, N. 2002. Phylogeny of Isoëtes (Lycopsida): resolving basalrelationships using rbcL sequences. Taxon 51: 83-89.
SCHELPE, E.A.C.L.E. & ANTHONY, N.C. 1986. Pteridophyta. In: LEISTER, O.A.(Ed) Flora of Southern Africa. Botanical Research Institute, Pretoria.
SCHNELLER, J.J. 1982. Cytological investigations on Isoetes malinverniana. Webbia35: 307-309.
SHARMA, B.D. 1998. Fungal associations in the roots of three species of Isoetes L.Aquatic Bot. 61: 33-37.
SHARMA, B.D. & SINGH, R. 1984. The ligule In Isoetes. Amer. Fern J. 74: 22-28.SHRIVASTVA, G.K. 1998. Isoetaceae in India: morphology and taxonomy. Indian Fern
J. 15: 165-177.SHRIVASTVA, G.K., PANT, D.D. & SHUKLA, P.D. 1993. The genus Isoetes L. in India.
Amer. Fern J. 83: 105-119. SHUKLA, P.K, SHRIVASTAVA, G.K. & SHUKLA, S.K. 2002. The quillworts (Isoetes)
of India: distribution, endemism and species radiation. Biodivers. & Conservation11: 959-973.
SINGH, S.K., SHUKLA, S.K., DUBY, N.K. & SHUKLA, P.K. 2018. Isoetes×gopalkrishnae, a new interspecific hybrid from central India. Phytotaxa 362: 68-76.
SMALL, R.L. & HICKEY, R.J. 2001. Systematics of the North Andean Isoetes karsteniiComplex. Amer. Fern J. 91: 41-69.
SUDOVA, R., RYDLOVA, J., CTVRTILKOVA, M., HAVRANEK, P. & ADAMEC, L.2011. The incidence of arbuscular mycorrhiza in two submerged Isoetes species.Aquatic Bot. 94: 183-187.
TAKAMIYA, M. 1999. Natural history and taxonomy of Isoetes (Isoetaceae). ActaPhytotax. Geobot. 50: 101-138.
TAKAMIYA, M. 2001. Isoetes sinensis var. sinensis in Korea (Isoetes: Pteridophyta).Fern. Gaz. (UK) 16: 169-176.
TAKAMIYA, M., WATANABE, M. & ONO, K. 1997. Biosystematic studies on thegenus Isoetes in Japan IV. Morphology and anatomy of sporophytes, phytogeographyand taxonomy. Acta Phytotax. Geobot. 48: 89-122.
TAYLOR, W.C. & HICKEY, R.J. 1992. Habitat, evolution and speciation in Isoetes. Ann.Missouri Bot. Gard. 79: 613-622.
TAYLOR, W.C., LEKSCHAS A.R., WANG, Q.-F., LIU, X., NAPIER, N.S. & HOOT,S.R. 2004. Phylogenetic relationships of Isoetes (Isoetaceae) in China as revealed bynucleotide sequences of nuclear ribosomal ITS region and second intron of a LEAFYhomolog. Amer. Fern J. 94: 196-205.
TAYLOR, W.C. & LUEBKE, N.T. 1988. Isoetes ×hickeyi: A naturally occurring hybridbetween I. echinospora and I. macrospora. Amer. Fern J. 78: 6-13.
332 FERN GAZ. 20(8): 309-333. 2018

TAYLOR, W.C., LUEBKE, N.T., BRITTON, D.M., HICKEY, R.J. & BRUNTON, D.F.1993 Isoetaceae - Quillwort family. In: FLORA OF NORTH AMERICAEDITORIAL COMMITTEE (Eds) Flora of North America, volume 2: Pteridophytesand Gymnosperms, pp. 64-75. Oxford University Press, New York.
TAYLOR, W.C., LUEBKE, N.T. & SMITH, M.B. 1985. Speciation and hybridizationin North American Quillworts. Proc. Royal Soc. Edinburgh 86B: 259-263.
TAYLOR, W.C., MOHLENBROCK, R.H. & MURPHY, J.A. 1975. The spores andtaxonomy of Isoetes butleri and I. melanopoda. Amer. Fern J. 65: 33-38.
TROIA, A. 2016. Dispersal and colonization in heterosporous lycophytes: palynologicaland biogeographical notes on the genus Isoetes in the Mediterranean region. Webbia71: 277-281.
TROIA, A. & AZZELLA, M.M. 2013. Isoetes sabatina (Isoetaceae, Lycopodiophyta), anew aquatic species from central Italy. Pl. Biosyst. 147: 1052-1058.
TROIA, A. & BELLINI, E. 2001. Karyological observations on Isoetes durieui Bory(Lycophyta, Isoetaceae) in Sicily. Bocconea 13: 397-400.
TROIA, A. & GREUTER, W. 2014. A critical conspectus of Italian Isoetes (Isoetaceae).Pl. Biosyst. 148: 13-20.
TROIA, A. & GREUTER, W. 2015a. A conspectus of and key to Greek Isoetes(Isoetaceae) based on a reassessment of Haussknecht’s gatherings of 1885.Willdenowia 45: 391-403.
TROIA, A. & GREUTER, W. 2015b. Flora Critica d’’Italia: Isoetaceae. Fondazione perla Flora Italiana, Firenze. http://www.floraditalia.it/pdf/Isoetaceae.pdf [accessed 29April 2018].
TROIA, A., PEREIRA, J.B.S., KIM, C. & TAYLOR, W.C. 2016. The genus Isoetes(Isoetaceae): a provisional checklist of the accepted and unresolved taxa. Phytotaxa277: 101-145.
TROIA, A. & RAIMONDO, F.M. 2009. [published 2010]. Isoetes todaroana (Isoetes,Isoetaceae), a new species from Sicily (Italy). Amer. Fern J. 99: 238-243.
TROIA, A. & ROUHAN, G. 2018. Clarifying the nomenclature of some Euro-Mediterranean quillworts (Isoetes, Isoetaceae): Indicator species and species ofconservation concern. Taxon, https://doi.org/10.12705/675.10.
VOGE, M. 1997. Plant size and fertility of Isoetes lacustris L. in 20 lakes of Scandinavia.Arch. Hydrobiologia 139: 171-185.
VOGE, M. 2003. Environmentally related demography: field studies on Isoetes lacustrisL. (Lycophyta, Isoetaceae) in Europe. In: CHANDRA, S. & SHRIVASTAVA, M.(Eds) Pteridology in the New Millennium, pp. 233-260, Kluwer Academic Publishers,Dordrecht.
WAGNER, W.H., WAGNER, F.S. & TAYLOR, W.C. 1986. Detecting abortive spores inherbarium specimens of sterile hybrids. Amer. Fern J. 76: 129-140.
WANNTORP, H-E. 1970. The genus Isoetes in South West Africa. Svenska BotaniskTidskrift 64: 141-157.
WEBER, U. 1922. Zur anatomie und systematik der gattung Isoetes L. Hedwigia 63:219-262.
WEN, M.C., PANG, X.-A., WANG, Q.-F. & TAYLOR, W.C. 2003. Relationship betweenwater chemistry and the distribution of the endangered aquatic quillwort Isoetessinensis Palmer in China. J. Freshw. Ecol. 18: 361-367.
WILLIAMS, S. 1943. On Isoetes australis S. Williams, a new species from WesternAustralia. Proc. Roy. Soc. Edinburgh, Sec. B, 62: 1-11.
BRUNTON & TROIA: TAXONOMIC RESEARCH INTO ISOETES 333

SHORT BIOGRAPHIES OF THE AUTHORS
Daniel Brunton was born in Ottawa, Canada on 17 May 1948. He grew up in that areaand graduated with a Geography degree from Carleton University, Ottawa, in 1973. Akeen naturalist from an early age, he translated that interest into summer employmentand then permanent work in provincial parks in Ontario and Alberta (1967-1979),involved in the areas of environmental interpretation, ecological inventories and systemsplanning.
The lessons learned in parks prepared him for a career as an independent ecologicalconsultant from which he retired (mostly) in 2017. Encouragement from federalgovernment scientists in Ottawa and insights gained from expert associates in those fieldendeavors encouraged his interest in natural history generally and botany in particular.
334 FERN GAZ. 20(8). 2018
Isoetologists (left to right) Carl Taylor, Angelo Troia and Daniel Brunton visiting bedrockoutcrop Isoetes sites (I. piedmontana is abundant in the flooded erosion pits visiblebehind Taylor) at Clover, South Carolina, USA, 2 April 2016 (Photo: Gerry Taylor).

An extensive series of field based studies across Canada resulted in numerouspublications on rare flora and fauna, especially pteridophytes, from the 1970s throughthe 1990s and the development of a very large (20,000 specimen) private herbarium.
Association with the late Donald M. Britton, foremost Canadian pteridologist, drewBrunton into the challenging - perplexing! - world of ‘Isoetology’ in the late 1980s.Beginning with their discovery of an undescribed hybrid, the Britton and Brunton teamexplored Isoetes throughout North American and beyond. They generated numerouspapers between 1989 and 2006, describing over a dozen new species, hybrids andsubspecies. To date, Brunton has produced 49 publications dealing with the taxonomy,diversity, distribution, ecology and conservation of Isoetes in North America, Asia andEurope.
Angelo Troia was born in Palermo, Italy on 3 September 1967 and grew up in Sicily.After obtaining a degree in Natural Sciences at the University of Palermo in 1991 anddoing his (obligatory) service in the Italian Navy, he undertook PhD studies in PlantBiosystematics and Ecology between Florence and Bologna (Italy). His doctoral thesiswas on the systematics and biogeography of the rare insular endemic Cytisus aeolicusGuss. (Fabaceae). Returning to Sicily in 1997, he was introduced to the fantastic worldof Mediterranean quillworts by the late Prof. Enrico Bellini, his studies focussing oncaryological, taxonomic and ecological aspects of various species.
At the IX OPTIMA Meeting in Paris, in May 1998, Troia and Bellini presented aposter about the (previously unknown) chromosome number of the Mediterranean Isoetesdurieui. After a period as Director of the Trapani Saltworks Nature Reserve in westernSicily, he returned to academic research in 2007, working as a botanical researcher atthe University of Palermo. There he prepared the treatments for the three lycophytefamilies for the project “Flora Critica d’Italia”.
Troia’s ongoing research is focused on the diversity, ecology and biogeography ofplants, with a particular focus on the genus Isoetes (and other lycophytes and ferns),freshwater plants and their habitats, the role of islands in the ecology of theMediterranean, and a wide variety of conservation issues. In the course of his studies,he has produced 27 publications involving the taxonomy, biogeography, cytology,ecology and conservation of Isoetes. His most recent area of research is the study ofCharaceae (green algae).
FERN GAZ. 20(8). 2018 335

SHORT NOTE
TWO NEW COMBINATIONS IN GRAMMITID FERNS(POLYPODIACEAE) FROM NEW GUINEA: PROSAPTIALEDERMANNII AND RADIOGRAMMITIS HABBEMENSIS
Two new combinations in New Guinea Grammitid ferns (Polypodiaceae) are proposed:Prosaptia ledermannii (Brause) Parris and Radiogrammitis habbemensis (Copel.) Parris.Herbarium abbreviations follow Thiers (2018).
Prosaptia ledermannii (Brause) Parris, comb. nov., based on Davallia ledermanniiBrause, Bot. Jahrb. Syst. 56: 123 (1920). Type: Nordöstl. Neu-Guinea, Kaiserin-Augusta-Fluss-(Sepik-)Gebiet, Schraderberg, 2070 m alt., 31 May 1913, Ledermann 11842 (BM!image). Syn. Polypodium subhamatopilosum Alderw., Bull. Jard. Bot. Buitenzorg, sér.3, 5: 220 (1922), nomen novum pro Davallia ledermannii Brause, non Polypodiumledermannii Brause (1920); Prosaptia subhamatopilosa (Alderw.) Ching, Bull. Fan Mem.Inst. Biol. 10: 242 (1941); Ctenopteris ledermannii (Brause) Copel., Univ. Calif. Publ.Bot. 18: 225 (1942); Grammitis subhamatopilosa (Alderw.) Christenh., Global FloraGLOVAP Nomenclature Part 1, Vol. 4: 55 (2018).
Radiogrammitis habbemensis (Copel.) Parris, comb. nov., based on Grammitishabbemensis Copel., Univ. Calif. Publ. Bot. 18: 223 (1942). Type: Netherlands NewGuinea, 9 km east of Lake Habbema, 2860 m, Oct. 1938, Brass 10520 (holotype MICH!;isotypes A!, BO!, BRI, K! L! Syn. Oreogrammitis habbemensis (Copel.) Parris, Gard.Bull. Singapore 58(2): 260 (2007).
A detailed examination of the second collection of this species (Kessler 14049 &Abrahamczyk, Indonesia, Papua Barat, road from Wamena to Lake Habbema, 6 kmbefore summit, 4°08’S 138°47’E, 3000 m, 20 March 2011, UC) shows that the rhizomeis radial with stipes arranged in whorls, characteristic of Radiogrammitis Parris ratherthan dorsiventral, with stipes in two rows, characteristic of Oreogrammitis Copel., sothe species needs to be transferred to the former from the latter.
ACKNOWLEDGEMENTSI wish to thank the Keepers of A, K, L, MICH and UC for loan of material, the Keeperof BO for permission to examine Grammitid holdings in 2013 and Ewen Cameron (AK)for expediting the loans.
REFERENCESALDERWERELT VAN ROSENBURGH, C. R. W. K. 1922. New or interesting Malayan
ferns 12. Bull. Jard. Bot. Buitenzorg, sér. 3, 5: 179-240.BRAUSE, G. 1920. Bearbeitung der von C, Ledermann von der
Sepik-(Kaiserin-Augusta-) Fluss-Expedition 1912-bis 1913 und von anderenSammlern aus dem Papuagebiete früher mitgebrachten Pteridophyten, nebstÜbersicht über alle bis jetzt aus dem Papuagebiet bekannt gewordenen Artenderselben. Bot. Jahrb. Syst. 56: 31-250.
CHING, R. C. 1941. New family and combinations of Ferns. Bull. Fan. Mem. Inst. Biol.10: 235-256.
Continued on page 347
336 FERN GAZ. 20(8). 2018

GROWTH OF FERN GAMETOPHYTES AFTER 20 YEARS OF STORAGE IN LIQUID NITROGEN
V. C. PenceCenter for Conservation and Research of Endangered Wildlife (CREW)
Cincinnati Zoo & Botanical Garden, 3400 Vine Street, Cincinnati, OH 45220, USA email: [email protected]
Key words: cryopreservation, ex situ conservation, gametophyte; in vitro; long-termstorage
ABSTRACTIn vitro grown gametophytes of six species of ferns, which had beencryopreserved using the encapsulation dehydration procedure, were evaluated forsurvival after 20 yrs of storage in liquid nitrogen. Tissues were rewarmed andtransferred to a recovery medium with the same methods originally used to testpre-storage viability. All six species resumed growth. Post-storage viability wasnot consistently higher or lower than pre-storage viability of LN exposed tissues,likely reflecting the small sample sizes. However, these results demonstrate thatlong-term storage in liquid nitrogen is a viable option for preserving gametophytesof at least some fern species and could be utilized as an additional tool forpreserving valuable gametophyte collections and for the ex situ conservation offern biodiversity.
INTRODUCTIONFor many species of ferns, gametophyte tissues have proven to be highly adaptable togrowth in vitro (Table 1). Most of these have been initiated through the asepticgermination of spores, although the aseptic germination of gemmae has also beendemonstrated (Raine & Sheffield, 1997). As in vitro cultures, gametophytes can providetissues for research and for propagation, both for ornamental ferns as well as for ferns ofconservation concern. The ex situ conservation of ferns has traditionally relied on livingcollections and spore banks (Ballesteros, 2011). However, there are situations where itmay be desirable to conserve other types of fern tissues, particularly if spores are few orare difficult to access. If only a few spores are available, they can be germinated in vitroand multiplied to provide a theoretically unlimited supply of gametophytic tissue, whichcan be used to generate sporophytes.
Tissues of such in vitro-grown gametophytes have also been shown to be adaptableto storage using cryopreservation, or storage in liquid nitrogen (LN) (Barnicoat et al.,2011; Makowski et al., 2016, 2015, Mikula et al., 2010; Mikuła, Jata & Rybczyski, 2009;Pence, 2015; 2000; Wilkinson, 2002). This can serve as an alternative to maintainingactive cultures, which requires a constant input of labor and resources. Gametophytesappear to be particularly adaptable to cryostorage, largely due to their simplicity of formand high regenerability (Maeda & Ito, 1981). However, results thus far havedemonstrated the ability of gametophytic tissues to survive only short-term exposure toLN, generally 1 hr-3 days (Mikula et al., 2010; Pence, 2000).
In this lab in the 1990s, tissues from in vitro cultures of gametophytes of several fernspecies were shown to survive LN exposure (Pence, 2000). These common species werechosen because they provided readily accessible material for experimentation, but three
FERN GAZ. 20(8): 337-346. 2018 337
338 FERN G
AZ. 20(8): 337-346. 2018
Species Family Methoda ifLN Exposed Reference
Adiantum capillus-veneris L. Adiantaceae Somer et al., 2009Adiantum tenerum Sw. Adiantaceae ED Pence, 2000Adiantum trapeziforme L. Adiantaceae ED Pence, 2000Anemia phillitidis (L.) Sw. Schizaeaceae Kaźmierczak, 2003Asplenium adiantum-nigrum L. Aspleniaceae Somer et al., 2009Asplenium scolopendrium L. Aspleniaceae ED Pence, 2015Ceratopteris richardii Brongn. Parkeriaceae Kamachi & Noguchi, 2012Ceratopteris thalictroides (L.) Brongn. Parkeriacae ED Makowski et al., 2015Ceterach officinarum DC. Aspleniaceae Somer et al., 2009Cibotium glaucum (Sm.) Hook. & Arn. Dicksoniaceae ED, EV, V Mikula et al., 2010; Pence, 2000Cibotium schiedei Schltdl. & Cham. Dicksoniaceae ED, EV, V Mikula et al., 2010Ctenitis pauciflora (Kaulf.) Holttum Dryopteridaceae ED Barnicoat et al., 2011Cyathea australis Domin. Cyatheaceae ED, EV, V Mikula et al., 2010; Mikuła, Jata & Rybczyski, 2009Cyathea dealbata (G.Forst.) Sw. Cyatheaceae ED, EV, V Mikula et al., 2010Cyathea delgadii Sternb. Cyatheaceae ED, EV, V Mikula et al., 2010Cyathea smithii Hook.f. Cyatheaceae ED, EV, V Mikula et al., 2010Davallia canariensis (L.) Sm. Davalliaceae Somer et al., 2009Davallia fejeensis Hook. Davalliaceae ED Pence, 2000Dicksonia fibrosa Colenso Dicksoniaceae ED, EV, V Mikula et al., 2010
TABLE 1.Species of ferns that have been grown as gametophytes in vitro, some of which have also survived exposure to LN.

PENC
E: FERN GAM
ETOPH
YTES AFTER 20 YEARS IN LIQ
UID NITRO
GEN
339
Dicksonia sellowiana (Pr.) Hook. Dicksoniaceae Fiori, Santos & Randi, 2009Drynaria quercifolia (L.) J.Sm. Polypodiaceae ED Pence, 2000Dryopteris dilatata (Hoffm.) A.Gray Dryopteridaceae Somer et al., 2009Dryopteris filix-mas (L.) Schott Dryopteridaceae Somer et al., 2009Hymenophyllum tunbrigense (L.) Sm. Hymenophyllaceae ED Wilkinson, 2002Lepisorus longifolius (Blume) Holttum Polypodiaceae ED Barnicoat et al., 2011Macroglossum smithii (Rac.) Campbell Marattiaceae ED Barnicoat et al., 2011Notholaena R.Br. ‘Sun Tuff’ Adiantaceae Rogers & Banister, 1992Osmunda regalis L. Osmundaceae ED, EV, V Makowski et al., 2016 Mikula et al., 2010Osmundastrum cinnamomeum (L.) C.Presl Osmundaceae Hollingsworth et al., 2012Phlebodium aureum (L.) J.Sm. Polypodiaceae ED Pence, 2000Phyllitis (Asplenium) scolopendrium (L.) Newman Aspleniaceae ED, EV, V Mikula et al., 2010Polypodium cambricum L. Polypodiaceae Bertrand et al., 1999; Somer et al., 2009)Pteridium aquilinum (L.) Kuhn Dennstaedtiaceae Whittier & Steeves, 1960Pteris adscensionis Sw. Pteridaceae ED Barnicoat et al., 2011Tectaria devexa (Kunze) Copel. Dryopteridaceae Ashmore et al., 2011Thelypteris confluens (Thunb.) C.V.Morton Thelypteridaceae Sara & Manickam, 2007Pronephrium triphyllum (Sw.) Holttum Thelypteridaceae Marimuthu & Manickam, 2011Sphaerostephanos unitus (L.) Holttum Thelypteridaceae Marimuthu & Manickam, 2011Trichomanes speciosum Willd. Hymenophyllaceae Raine & Sheffield, 1997Woodwardia radicans (L.) Sm. Blechnaceae DeSoto et al., 2008
aMethods: ED = encapsulation dehydration; EV = encapsulation vitrification; V = vitrification

of the species are also congeneric with rare species (Kang et al., 2008; Kishida, 2015;Yu et al., 2017). In those studies, in addition to testing the initial survival of thegametophytes (Pence, 2000), samples were banked in the Frozen Garden™ of CREW’sCryoBioBank™ for long-term storage. In a recent study to evaluate the viability ofnumerous samples from the Frozen Garden after years of storage, several samples ofgametophytes of these six species were removed and evaluated for their viability after20 yrs in LN. The results of that study are presented here.
METHODSIn vitro gametophyte cultures of Davallia fejeensis Hook., Drynaria quercifolia (L.) J.Sm., Cibotium glaucum (Sm.) Hook & Arn., Adiantum trapeziforme L., Adiantumtenerum Sw., and Phlebodium aureum L. were originally initiated by the asepticgermination of spores in vitro, as previously described (Pence, 2000), and maintainedon a medium of half-strength Murashige and Skoog salts and minimal organics(Linsmaier and Skoog, 1965) with 1.5% sucrose and 0.22% Phytagel (gellan gum, SigmaChemical Co., St. Louis, MO) (= ½ MS medium), in 60 x 15 mm Petri plates (Corning™Falcon™, Durham, NC), approximately 15 ml of medium/plate. Cultures weremaintained at 26oC under cool white fluorescent lights at 10 – 20 µmol m-2 sec-1 PPFD,with a 16:8 hr light:dark cycle.
Tissue banking took place between 6/21/1995 and 7/26/1995. Fragments ofgametophyte tissue were isolated by cutting the gametophyte with a scalpel into smallpieces from throughout the gametophyte thallus. Pieces ranged in size from 1-3 mm inlength and width. Gametophyte fragments were prepared for cryopreservation byculturing for seven days on ½ MS medium with the addition of filter-sterilized 10 µMabscisic acid (ABA). They were then cryopreserved using the encapsulation dehydrationprocedure (Fabre & Dereuddre, 1990), in which the fragments were encapsulated inalginate beads before an18 hr incubation in a 0.75 M sucrose solution, with more thanone fragment being encapsulated per bead. The encapsulated tissues were then dried for3 or 5 hrs under the airflow of a laminar flow hood. The dried beads were stored in 2 mlpolypropylene cryovials (Corning®, Corning, NY), 6 to 19 beads per vial, the numberof beads varying depending on the amount of tissue available. The samples were storedin a MVE XC 47/11, 900 vial capacity LN storage tank (Chart MVE, Ball Ground, GA),submerged in LN (-196oC). As a control at the time of banking, one vial of each speciesfrom each time of banking was rewarmed after a one-day exposure to LN and culturedon ½ MS medium for recovery. Recovery growth was scored as the number of beadsshowing green growing tissues, and data for the samples were taken between nine and28 days after rewarming. In the case of three species, survival was also measured after4.5 months of growth, with data taken after four weeks of growth. Samples from eachspecies were also removed after 3.5 yrs of storage in LN, and recovery data was takenafter two and seven weeks of growth. The data taken seven weeks after rewarming werereported previously (Pence, 2000). Samples were removed from LN between 7/16/2015 and 8/14/2015 to evaluate 20-yrsurvival. Recovery conditions in the present study replicated as much as possible theoriginal recovery conditions, using ½ MS medium, only with modification in the sourceof the prepared MS salt mixture (Phytotechnology Laboratories, Overland Park, KS) andthe source and amount of the gellan gum (Gelzan, Caisson Labs, Smithfield, UT), addedat 0.33%. One vial representing each species and each banking time for that species wasremoved (9 vials total) and rewarmed at ambient temperature (22oC) for 15 min. The
340 FERN GAZ. 20(8): 337-346. 2018

beads were then plated on 1 – 4 plates of ½ MS medium, depending on the number ofbeads, 4-6 beads/plate. Cultures were maintained under the same temperature and lightingconditions originally used for growth. At the time of banking and at 3.5 yrs, survival was scored as the number of beads showingsome recovery growth of encapsulated tissues, regardless of the number of fragmentsper bead. When tissues were evaluated after 20 years, survival was scored both as thepercent of fragments and the percent of beads showing green, growing tissue at 3 – 4weeks.
RESULTSGrowth from cryopreserved gametophyte tissue occurred from all of the species of fernstested in this study (Table 2). The values for the original survival data reported here arethose recorded for the specific samples that were banked and are different from thosereported previously (Pence, 2000), which were collected from experiments done inpreparation for the long-term banking of samples. In all but two species, the percent oftissues showing growth after storage was greater than that observed before long-termstorage. In Drynaria quercifolia and both samples of A. trapeziforme, post-storage growthwas less than that observed originally, and in one sample of A. trapeziforme, there wasno post-storage growth. However, that sample produced fungal growth on a few beadswithin the first few days of culture, and the remaining beads were transferred to freshplates. The transferred beads did not show any contamination, but they also did not showany growth. Specific moisture levels of the banked samples used in this study are notavailable, except for two of the three samples, D. quercifolia and A. trapeziforme,whichwere dried for 5 hrs to 17.5% and 18.9% moisture, respectively.
Samples recovered at 20 yrs were scored at 3 – 4 weeks. When these were comparedwith samples recovered at 3.5 yrs, the percent survivals were, in most cases, more similarto those recorded at two weeks from the 3.5 yr samples than with survivals recorded atseven weeks for those samples, which in all cases were higher than two week survivalvalues (Table 2).
Since pre-storage viability was based on growth per bead, post-storage measurementswere calculated both on growth per bead and on growth per fragment for seven of thenine samples. The percent survival was very similar between the two measurements(Table 2). Most fragments showed some damaged areas, but ultimately, those with anysurviving areas went on to regenerate gametophyte tissues (Figure 1).
DISCUSSIONThese results demonstrate that in vitro-grown gametophytes of at least some fern speciescan survive up to 20 yrs in LN storage without appreciable deterioration using theencapsulation dehydration procedure. This method has been applied by others in theintervening yrs with good success (Table 1). There also have been reports of somesurvival of fern gametophytes using the encapsulation vitrification method (Makowskiet al., 2016; Mikula et al., 2010), although when it was compared with encapsulationdehydration, the latter resulted in higher survival. Studies using vitrification without theprotection of encapsulation have uniformly reported severe plasmolysis of thegametophyte tissues and no survival after exposure to LN (Makowski et al., 2016; Mikulaet al., 2010).
When the successful cryopreservation of fern gametophytes was reported originally,3 – 4 hrs of drying brought samples to 19-27% moisture (wet weight basis) (Pence, 2000).
PENCE: FERN GAMETOPHYTES AFTER 20 YEARS IN LIQUID NITROGEN 341
342 FERN G
AZ. 20(8): 337-346. 2018
SpeciesHrs ofDrying
Pre-storage 4.5 Mos 3.5 Yrs 20 Yrs
Day SurvivalaSurvival(at 4 wks)
Survival(at 2 wks)
bSurvival(at 7 wks) Day
Survivalper Bead
Survivalper Piece
Adiantum tenerum3 12 41.7 63.0 88.0 28 66.7 66.7
3 16 60.0 73.3 28 80.0 79.5
Adiantum trapeziforme3 12 25.0 33.0 50.0 28 0.0 0.0
5 4 84.6 20 39.3 nd
Cibotium glaucum 3 16 46.2 80 60.0 75.0 28 90.0 91.7
Davallia fejeensis5 8 46.2 80.0 100.0 28 82.3 82.3
3 9 6.7 40 20 50.0 nd
Drynaria quercifolia 5 6 91.7 50.0 67.0 28 41.3 40.5
Phlebodium aureum 3 27 19.3 67.0 90.0 28 83.5 83.5
TABLE 2Percent survival of six species of fern gametophytes, dried for 3 or 5 hrs, through LN exposure before storage and after 4.5 mos, 3.5 yrs, and 20yrs of LN storage. Day of observation is given in adjacent column, if different for different samples, or in the heading, if the same for all samples.
aData available for three species only.bSeven week survival data from Pence, 2000.

The moisture contents available for two of the samples in this study that had been driedfor five hrs were 18 – 19%. With seeds, a number of studies have been directed atdefining optimum storage moisture levels, which are related to the seed oil content (e.g.Pritchard, 1995; Hor et al., 2005). Whether the moisture levels used is this study areoptimum for fern gametophytes will require further research, but they were sufficient toallow regeneration of at least some of the tissues after cryostorage.
The fact that survival after long-term storage was greater in most cases than pre-storage survival may have resulted from data being taken much earlier in the case ofsome of the pre-storage samples, as these were done by different operators, at differenttimes, for various goals. The results of the 3.5 yr samples indicate that the initiation ofgrowth continued for several weeks, with more fragments growing at seven weeks thanat two weeks. Alternatively, the sample sizes in this study were small and may not belarge enough to clearly track any small changes in viability over the time of storage.Similar increases in viability after LN exposure have been noted with small samples ofother species, by others, as well as in this laboratory (Mix-Wagner, Schumacher & Cross,2003; Vanhove, Philpott & Pence, 2016).
The highly regenerative nature of fern gametophytes has been noted for severalspecies (Maeda & Ito, 1981; Ong & Ng, 1998), and this ability makes these tissues welladapted to cryopreservation. Even if some tissues are damaged in the process of eithercryoprotection or LN exposure, only a small amount of tissue is required (possibly onlyone cell) to regenerate the gametophyte tissue (Maeda & Ito, 1981). However, this abilitymakes it more difficult to evaluate survival quantitatively, because fragments are ofdifferent sizes and only portions of them may survive. Having more than one fragmentencapsulated in an alginate bead can increase the number of fragments available forregrowth and minimize the number of beads needed, and survival per bead appears to
PENCE: FERN GAMETOPHYTES AFTER 20 YEARS IN LIQUID NITROGEN 343
Figure 1. Encapsulated gametophtyes from two fern species resuming growth after 20yrs of storage in liquid nitrogen. A. Adiantum tenerum gametophytes, as well asfragments that did not grow (arrow); B. Phlebodium aureum gametophytes. Scale bars= 0.5 cm.

correlate well with fragment survival. When survival of the 20-yr old samples wasmeasured by both methods, the results were very similar.
Cryopreservation of fern gametophytes has thus far been applied to in vitro culturesof gametophytes. This requires the initiation of gametophyte cultures by the asepticgermination of spores or gemmae, and in vitro fern gametophyte cultures initiated in thisway have been established and reported for a number of species (Table 1). However, forspecies known only from the gametophyte in nature, in vitro cultures would need to beestablished directly from gametophytic tissue. Methods for this have been reported forgametophytes of bryophytes (Rowntree & Ramsay, 2005; Segreto et al., 2010) but notyet for fern gametophytes. Gametophytes of bryophytes are more complex structuresthan those of ferns, and further research is needed to determine whether ferngametophytes can survive the surface sterilization required for initiating in vitro cultures.
The results of this study indicate gametophyte cryopreservation is an option for long-term conservation of in vitro collections of fern gametophytes that have been developedfor horticultural or experimental purposes, as well as an option for the ex situconservation of fern biodiversity. Thus, it is a tool that can contribute to the ex situconservation goals (Target 8) of the Global Strategy for Plant Conservation (Conventionon Biological Diversity, 2010). Further research is needed to test the applicability ofencapsulation dehydration to more species with different habits and adaptations in orderto more clearly define the breadth of its potential use. In addition, more long-term studiesshould be initiated to evaluate the effectiveness of cryopreservation in preserving tissuesover the course of decades. However, these results with six species clearly demonstratethe potential of this method for fern conservation in the future.
ACKNOWLEDGEMENTSThe helpful technical assistance of Kirsten Stewart in banking the original samples isacknowledged. Removal and evaluation of samples after 20 yrs of storage was supportedby Institute of Museum and Library Services grant no. LG-25-12-0595-12.
REFERENCES ASHMORE, S.E., HAMILTON, K.N. & OFFORD, C.A. 2011. Conservation
technologies for safeguarding and restoring threatened flora: Case studies fromEastern Australia. In Vitro Cell. Develop. Biol.- Plant 47: 99-109.
BALLESTEROS, D. 2011. Conservation of Fern Spores. In: FERNÁNDEZ, H. & AL,E. (Eds) Working with ferns: Issues and applications, pp. 165-172. SpringerScience+Business Media, LLC, New York.
BARNICOAT, H., CRIPPS, R., KENDON, J. & SARASAN, V. 2011. Conservation invitro of rare and threatened ferns—case studies of biodiversity hotspot and islandspecies. In Vitro Cell. Develop. Biol.-Plant 47: 37-45.
BERTRAND, A.M., ALBUERNE, M.A., FERNÁNDEZ, H., GONZÁLEZ, A. &SÁNCHEZ-TAMÉS, R. 1999. In vitro organogenesis of Polypodium cambricum.Plant Cell, Tiss. Org. Cult. 57: 65-69.
CONVENTION ON BIOLOGICAL DIVERSITY (CBD). 2010. Consolidated update ofthe Global Strategy for Plant Conservation 2011-2020. Available online athttp://www.cbd.int/gspc/strategy.shtml. (Accessed September 15, 2017)
DESOTO, L., QUINTANILLA, L.G. & MÉNDEZ, M. 2008. Environmental sexdetermination in ferns: Effects of nutrient availability and individual density inWoodwardia radicans. J. Ecol. 96: 1319-1327.
344 FERN GAZ. 20(8): 337-346. 2018

FABRE, J., & DEREUDDRE, J. 1990. Encapsulation-dehydration: A new approach tocryopreservation of Solanum shoot-tips. CryoLett. 11: 413-426.
FIORI, C.C.L., SANTOS, M. & RANDI, Á.M. 2009. Aspects of gametophytedevelopment of Dicksonia sellowianaHook (Dicksoniaceae): an endangered tree fernindigenous to South and Central America. Amer. Fern J. 99: 207-216.
HOLLINGSWORTH, S.N., ANDRES, E.A. & GREER, G.K. 2012. Pheromonalinteractions among gametophytes of Osmundastrum cinnamomeum and the originsof antheridiogen systems in leptosporangiate ferns. Intern. J. Plant Sci. 173: 382-390.
HOR, Y.L., KIM, Y.J., UGAP, A., CHABRILLANGE, N., SINNIAH, U.R.,ENGELMANN, F. & DUSSERT, S. 2005. Optimal hydration status forcryopreservation of intermediate oily seeds: Citrus as a case study. Annals of Botany95: 1153-1161.
KAMACHI, H. & NOGUCHI. M. 2012. Negative gravitropism in dark-growngametophytes of the fern Ceratopteris richardii. Amer. Fern J. 102: 147-153.
KANG, M., HUANG, H., JIANG, M. & LOWE, A.J. 2008. Understanding populationstructure and historical demography in a conservation context: Population geneticsof an endangered fern. Divers. Distrib. 14: 799-807.
KAźMIERCZAK, A. 2003. Induction of cell division and cell expansion at the beginningof gibberellin A3-induced precocious antheridia formation in Anemia phyllitidisgametophytes. Plant Sci. 165: 933-939.
KISHIDA, W. 2015. Asplenium dielmannii. The IUCN Red List of Threatened Species2015: e.T80229074A80229092. http://dx.doi.org/10.2305/IUCN.UK.2015-4.RLTS.T80229074A80229092.en. Downloaded on 16 April 2018.
LINSMAIER, E.M. & SKOOG, F. 1965. Organic growth factor requirements of tobaccotissue cultures. Physiol. Plant. 18: 100-127.
MAEDA, M. & ITO, M. 1981. Isolation of protoplasts from fern prothallia and theirregeneration to gametophytes. Bot. Mag. Tokyo 94: 35-40.
MAKOWSKI, D., JRYBCZYńSKI, .J., MIKULA, A. & KLIMASZEWSKA, K. 2015.A simple way to overcome the recalcitrance of the water fern Ceratopteristhalictroides (L.) Brongn. to cryopreservation. Acta Soc. Bot. Pol. 84: 385-388.
MAKOWSKI, D., TOMICZAK, K., RYBCZYńSKI, J.J. & MIKUłA, A. 2016.Integration of tissue culture and cryopreservation methods for propagation andconservation of the fern Osmunida regalis L. Acta Physiol. Plant. 38: 1-12.
MARIMUTHU, J., & MANICKAM, V.S. 2011. Ex situ conservation of two threatenedferns of the Western Ghats through in vitro spore culture. J. Threat. Taxa 3: 1919-1928.
MIKUłA, A., JATA, K., & RYBCZYSKI, J.J. 2009. Cryopreservation strategies forCyathea australis (R. Br.) Domin. CryoLett. 30: 429-439.
MIKULA, A., MAKOWSKI, D., WALTERS, C. & RYBCZYNSKI, J.J. 2010.Exploration of cryo-methods to preserve tree and herbaceous fern gametophytes. In:FERNÁNDEZ, H., KUMAR, A. & REVILLA, M. A. (Eds), Working with ferns:Issues and applications, pp. 173-192. Springer Science+Business Media, LLC, NewYork.
MIX-WAGNER, G., SCHUMACHER, H.M. & CROSS, R.J. 2003. Recovery of potatoapices after several years of storage in liquid nitrogen. CryoLett. 24: 33-41.
ONG, B.L., & NG, M.L. 1998. Regeneration of drought-stressed gametophytes of theepiphytic fern, Pyrrosia pilosellodes (L.) Price. Plant Cell Rep. 18: 225-228.
PENCE, V.C. 2000. Cryopreservation of in vitro grown fern gametophytes. Amer. Fern
PENCE: FERN GAMETOPHYTES AFTER 20 YEARS IN LIQUID NITROGEN 345

J. 90: 16-23.PENCE, V.C. 2015. Propagation and cryopreservation of Asplenium scolopendrium var.
americanum, the American hart’s-tongue fern. Amer. Fern J. 105: 211-225.PRITCHARD, H.W. 1995. Cryopreservation of seeds. In: DAY, J.G. & MCLELLAN,
M.R. (Eds) Methods in molecular biology. Vol. 38: Cryopreservation and freeze-drying protocols, pp. 133-144. Humana Press Inc., Totowa, NJ.
RAINE, A.C.A. & SHEFFIELD, E. 1997. Establishment and maintenance of asepticculture of Trichomanes speciosum gametophytes from gemmae. Amer. Fern J. 87:87-92.
ROGERS, S.M.D. & BANISTER, S. 1992. Micropropagation of Notholaena. HortSci.27: 1224-1225.
ROWNTREE, J.K. & RAMSAY, M.M. 2005. Ex situ conservation of bryophytes :Progress and potential of a pilot project. Bol. Soc. Esp. Briol. 26-27: 17-22.
SARA, S.C. & MANICKAM, V.S. 2007. In vitro developmental ontogeny and life cycleof a rare fern species - Thelypteris confluens (Thunb.) Morton. Ind. J. Biotech. 6:372-380.
SEGRETO, R., HASSEL, K., BARDAL, R. & STENøIEN, H.K. 2010. Desiccationtolerance and natural cold acclimation allow cryopreservation of bryophytes withoutpretreatment or use of cryoprotectants. The Bryol. 113: 760-769.
SOMER, M., ARBESú, R., MENÉNDEZ, V., REVILLA, M.A. & FERNÁNDEZ, H.2009. Sporophyte induction studies in ferns in vitro. Euphytica 171: 203-210.
VANHOVE, A.-C., PHILPOTT, M. & PENCE, V.C. 2016. Evaluating survival in shoottips of several species stored in liquid nitrogen for 4 to 16 years. In Vitro Cell.Develop. Biol.-Plant 52: S33.
WHITTIER D.P. & STEEVES, T. 1960. The induction of apogamy in the bracken fern.Can. J. Bot. 38: 925-930.
WILKINSON, T. 2002. In vitro techniques for the conservation of Hymenophyllumtunbrigense (L.) Sm. Fern Gaz. 16: 458.
YU, R., ZHANG, G., LI, H., CAO, H., MO, X., GUI, M., ZHOU, X., JIANG, Y., LI, S.& WANG, J. (2017) In vitro propagation of the endangered tree fern Cibotiumbarometz through formation of green globular bodies. Plant Cell Tiss. Org. Cult. 128:369-379.
346 FERN GAZ. 20(8): 337-346. 2018

Continued from page 336
CHRISTENHUSZ, M. 2018. Plant Gateway’s The Global Flora A practical flora tovascular plant species of the world special edition GLOVAP Nomenclature Part 1,Vol. 4: 1-155. Plant Gateway, Bradford.
COPELAND, E. B. 1942. Filicum Novarum CXXXVIII Diagnoses. Univ. Calif. Publ.Bot. 18: 217-226.
PARRIS, B. S. 2007. Five new genera and three new species of Grammitidaceae(Filicales) and the re-establishment of Oreogrammitis. Gard. Bull. Singapore 58(2):233-274.
THIERS, B. 2018. Index Herbariorum. New York Botanical Gardens, on-line.
Barbara S. ParrisFern Research Foundation, 21 James Kemp Place, Kerikeri, Bay of Islands,
New Zealand 0230Email: [email protected]
FERN GAZ. 20(8). 2018 347

INSTRUCTIONS FOR AUTHORSPAPERS should not usually exceed 20 printed pages and are generally expected to beconsiderably shorter. As well as reports of original research, review articles and shortnotes, e.g. new records, are encouraged. The senior author should supply an email addressto facilitate correspondence. REVIEWS should cover recent developments in any aspect of research on ferns andlycopods. This can include unpublished research by the author, and/or the possibledirection of future research. They should be aimed at the general reader who may not bean expert in the chosen topic. Proposals for a review should be discussed first with thereview editors.MANUSCRIPTS should be submitted in English (British) in electronic format in10-point Times New Roman font and single spaced. Electronic versions of text and tablesshould be compatible with WORD, with figures as TIFF files, and sent as emailattachments. All manuscripts will be refereed.THE TITLE should reflect the content of the paper and be in BOLD CAPITALS(11-point) and centrally aligned. Generic and specific names should be in italics and anytitle containing a generic or specific name should be followed by the family name inbrackets e.g.
TRICHOMANES SPECIOSUM (HYMENOPHYLLACEAE)IN SOUTHERN SPAIN
AUTHOR ABBREVIATIONS should follow Pichi Sermolli’s (1996) Authors ofscientific names in Pteridophyta, Royal Botanic Gardens, Kew.MAIN HEADINGS: should be in BOLD CAPITALS (10-point) and centrally aligned.SUBSIDIARY HEADINGS: should be in bold, the first letter of each word in capitals,the rest in lower case and left-aligned.AUTHORS’ NAMES AND FULL ADDRESSES (including email address): followthe title and are centrally aligned.KEY WORDS: up to ten.ABSTRACT: should reflect the content of the paper.FIGURES: there is no distinction between photographs and line drawings in numbering.All should be supplied in mono or colour, TIFF format, 300 dpi and CMYK. (pleasecontact the Production Editor with queries), with a scale bar where appropriate. Letteringor numbers (Arabic) should be in the bottom left using uppercase Times Roman. Figurecaptions should be on a separate sheet.TABLES: can be printed in either portrait or landscape format. Authors should considerthis when preparing tables. Authors should ensure that tables fit the printed page size ina legible form.PAGESIZE: The Gazette has a useable page size of 123 × 204 mm. This can be used inportrait or landscape mode but any supplied artwork should not be larger than this.MEASUREMENTS: follow the metric system.CHECKLISTS: follow the format of Baksh-Comeau, Fern Gaz. 16(1, 2): 11- 122.REFERENCES: follow the style of a recent issue of The Fern Gazette, e.g.:-HOOKER, W.J. 1864. Species Filicum, 5. Dulau & Co., London.MORTON, C.V. 1947. The American species of Hymenophyllum, section
Sphaeroconium. Contr. U.S. Natl. Herb. 29(3): 139-201.STEVENSON, D.W. & LOCONTE, H. 1996. Ordinal and familial relationships of
pteridophyte genera. In: CAMUS, J.M., GIBBY, M. & JOHNS, R.J. (Eds)Pteridology in perspective, pp. 435-467. Royal Botanic Gardens, Kew.
JOURNAL ABBREVIATIONS: should follow Botanico Periodicum Huntianum &Supplements.Alterations from the original text at proof stage will be charged for unless they are minorpoints of detail. A pdf will be provided free to the senior author.
348 FERN GAZ. 20(8). 2018
Copyright © 2022 FDOKUMEN




















