
Review Phenotypic and genotypic analysis of drought-resistance traits for development of rice...
23
Review Phenotypic and genotypic analysis of drought-resistance traits for development of rice cultivars adapted to rainfed environments Akihiko Kamoshita a, *, R. Chandra Babu b , N. Manikanda Boopathi b , Shu Fukai c a Asian Natural Environmental Science Center, University of Tokyo, 1-1-1 Midoricho, Nishitokyo 188-0002, Japan b Department of Plant Molecular Biology and Biotechnology, Center for Plant Molecular Biology, Tamil Nadu Agricultural University, Coimbatore 641003, India c University of Queensland, School of Land, Crop and Food Sciences, Queensland, Australia 4072 Contents 1. Introduction ....................................................................................................... 2 2. Types of drought and drought-resistance traits ........................................................................... 2 2.1. Types of drought ............................................................................................. 2 2.2. Drought-resistance traits ....................................................................................... 3 2.2.1. Terminal drought ...................................................................................... 4 2.2.2. Vegetative stage drought................................................................................ 5 2.2.3. Intermittent drought ................................................................................... 5 3. Selection of drought-resistant genotypes ................................................................................ 5 3.1. Screening environments........................................................................................ 5 3.2. Selection criteria.............................................................................................. 6 Field Crops Research 109 (2008) 1–23 ARTICLE INFO Article history: Received 15 December 2007 Received in revised form 11 June 2008 Accepted 14 June 2008 Keywords: Drought avoidance Drought screening Drought type QTL MAS Rainfed lowland rice Upland rice ABSTRACT Many of the world’s rice-growing regions lack adequate irrigation facilities, and drought frequently reduces yield. This paper reviews drought-resistance traits in rice and their quantitative trait loci (QTLs), with emphasis on CT9993/IR62266, one of the most widely studied mapping populations, and suggests ways to develop cultivars that will perform well in drought-prone environments. Information about the type of drought faced in the target region – particularly the timing of the drought (late season terminal drought, early stage vegetative drought, and intermittent drought) and the intensity of the drought – are important in determining the specific plant traits required to improve drought resistance in rice. Most of these traits are related to drought avoidance strategy, so that the drought-resistant genotypes are able to maintain better internal water status, either by taking up more water through a better root system or by reducing the rate of plant water use. We identified and listed a number of QTLs for many drought- resistance traits, such as deep roots. We identified four key genomic regions on chromosomes 1, 4, 8, and 9 on which are co-located a number of QTLs for traits considered to be directly or indirectly responsible for grain yield under stress. These regions, once they have been more finely mapped, appear promising for eventual use in marker-assisted selection for development of drought-resistant rice varieties. In addition to selecting for specific traits or specific genomic regions, screening under managed drought conditions on the basis of yield itself or on spikelet fertility adjusted for flowering time appears useful, because of the relatively high degrees of heritability of these characters, for the development of drought-resistant rice cultivars, and it is currently practiced in some breeding programs. ß 2008 Elsevier B.V. All rights reserved. * Corresponding author. E-mail address: [email protected] (A. Kamoshita). Abbreviations: ABA, abscisic acid; DH, doubled haploid; DRI, drought response index; EW, epicuticular wax; GA, gibberellic acid; G E, genotype-by-environment interaction; LEA, late embryogenesis abundant; LWP, leaf water potential; MAS, marker-assisted selection; QTL, quantitative trait locus; RPR, root pulling resistance; RWC, relative water content; SSR, simple sequence repeat. Contents lists available at ScienceDirect Field Crops Research journal homepage: www.elsevier.com/locate/fcr 0378-4290/$ – see front matter ß 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.fcr.2008.06.010
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Review Phenotypic and genotypic analysis of drought-resistance traits for development of rice...

Field Crops Research 109 (2008) 1–23
Review
Phenotypic and genotypic analysis of drought-resistance traits for developmentof rice cultivars adapted to rainfed environments
Akihiko Kamoshita a,*, R. Chandra Babu b, N. Manikanda Boopathi b, Shu Fukai c
a Asian Natural Environmental Science Center, University of Tokyo, 1-1-1 Midoricho, Nishitokyo 188-0002, Japanb Department of Plant Molecular Biology and Biotechnology, Center for Plant Molecular Biology, Tamil Nadu Agricultural University, Coimbatore 641003, Indiac University of Queensland, School of Land, Crop and Food Sciences, Queensland, Australia 4072
A R T I C L E I N F O
Article history:
Received 15 December 2007
Received in revised form 11 June 2008
Accepted 14 June 2008
Keywords:
Drought avoidance
Drought screening
Drought type
QTL
MAS
Rainfed lowland rice
Upland rice
A B S T R A C T
Many of the world’s rice-growing regions lack adequate irrigation facilities, and drought frequently
reduces yield. This paper reviews drought-resistance traits in rice and their quantitative trait loci (QTLs),
with emphasis on CT9993/IR62266, one of the most widely studied mapping populations, and suggests
ways to develop cultivars that will perform well in drought-prone environments. Information about the
type of drought faced in the target region – particularly the timing of the drought (late season terminal
drought, early stage vegetative drought, and intermittent drought) and the intensity of the drought – are
important in determining the specific plant traits required to improve drought resistance in rice. Most of
these traits are related to drought avoidance strategy, so that the drought-resistant genotypes are able to
maintain better internal water status, either by taking up more water through a better root system or by
reducing the rate of plant water use. We identified and listed a number of QTLs for many drought-
resistance traits, such as deep roots. We identified four key genomic regions on chromosomes 1, 4, 8, and
9 on which are co-located a number of QTLs for traits considered to be directly or indirectly responsible
for grain yield under stress. These regions, once they have been more finely mapped, appear promising for
eventual use in marker-assisted selection for development of drought-resistant rice varieties. In addition
to selecting for specific traits or specific genomic regions, screening under managed drought conditions
on the basis of yield itself or on spikelet fertility adjusted for flowering time appears useful, because of the
relatively high degrees of heritability of these characters, for the development of drought-resistant rice
cultivars, and it is currently practiced in some breeding programs.
� 2008 Elsevier B.V. All rights reserved.
Contents lists available at ScienceDirect
Field Crops Research
journa l homepage: www.e lsev ier .com/ locate / f c r
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
2. Types of drought and drought-resistance traits . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
2.1. Types of drought . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
2.2. Drought-resistance traits . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3
2.2.1. Terminal drought. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4
2.2.2. Vegetative stage drought. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
2.2.3. Intermittent drought . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
3. Selection of drought-resistant genotypes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
3.1. Screening environments. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
3.2. Selection criteria. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6
* Corresponding author.
E-mail address: [email protected] (A. Kamoshita).
Abbreviations: ABA, abscisic acid; DH, doubled haploid; DRI, drought response index; EW, epicuticular wax; GA, gibberellic acid; G � E, genotype-by-environment
interaction; LEA, late embryogenesis abundant; LWP, leaf water potential; MAS, marker-assisted selection; QTL, quantitative trait locus; RPR, root pulling resistance; RWC,
relative water content; SSR, simple sequence repeat.
0378-4290/$ – see front matter � 2008 Elsevier B.V. All rights reserved.
doi:10.1016/j.fcr.2008.06.010

A. Kamoshita et al. / Field Crops Research 109 (2008) 1–232
4. Molecular approaches for development of drought-resistant varieties . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
4.1. QTL mapping . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
4.1.1. Co-location of QTLs for plant-type traits, and integrative, primary, and secondary drought-resistance traits . . . . . . . . . . . . 12
4.1.2. Fine mapping of QTLs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 17
4.2. Marker-assisted selection for drought resistance . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18
4.3. Transgenics . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18
5. Improvement of general adaptability and drought resistance . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18
6. Conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19
Acknowledgements. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19
1. Introduction
Large areas of rice are grown under lowland and upland rainfedconditions. These areas respectively occupy 31% and 11% of theglobal rice-growing area (IRRI, 2001). Drought is a majorlimitation for rice production in rainfed ecosystems. Evensonet al. (1996) estimated global rice yield lost to drought to be 18million tonnes annually or 4% of total rice production, which wasvalued conservatively at US$ 3.6 billion at that time. On average,the estimated yield lost to drought is 144 kg/ha annually ineastern India (Dey and Upadhyaya, 1996). Simulation modelingindicates that reduction in yield of rainfed lowland rice owing todrought averages from 13% to 35% for different locations innortheast Thailand (Jongdee et al., 1997b). Regardless of themonetary value placed on these losses, the ramifications for poorfarm families are often incalculable (O’Toole, 2000) since rice istheir staple food.
The ‘‘green revolution’’ in rice improvement has benefited manyfarmers in irrigated rice production but has had limited impact onrainfed production (Evenson and Gollin, 2003). Indeed, the gap inyield between irrigated and rainfed rice has increased from1.7 t ha�1 in the late 1960s to 3.6 t ha�1 in the late 1990s in Asia(Maclean et al., 2002).
Development of drought-resistant cultivars has been slow forrainfed lowland and upland rice. Fukai et al. (1999) indicated sixdifficulties inherent to selecting drought-resistant rice for rainfedlowlands: (1) coupling of photosynthesis and transpirationprocesses when water is limited, hence difficulty in increasingdry matter production and yield; (2) interaction between nutrientavailability and water stress, with even mild soil water deficitsreducing availability; (3) incomplete understanding of themechanisms of drought resistance; (4) the importance ofphenology and potential yield as components of yield underlimited water that override the effectiveness of drought-resistancecharacteristics; (5) large genotype-by-environment interaction(G � E) for yield, causing inconsistency in yield performance indifferent environments, and hence necessitating more testing indifferent locations and years; and (6) the different types ofdrought, and the different traits that may therefore be required foreach drought type.
The slow progress in developing drought-resistant rice alsoreflects the lack of a specific method for screening the largenumbers of genotypes required in breeding for drought (Zeiglerand Puckridge, 1995). Using natural field conditions in the wetseason is difficult because rainfall can eliminate water deficits.However, new drought-screening facilities and methods haverecently been developed (O’Toole, 2004), and they are facilitatingprogress in our understanding of drought-resistance traits and inour selection of drought-resistant genotypes.
Since McCouch et al. (1988) constructed the first linkage map inrice, several well-saturated maps have been developed (Causseet al., 1994; McCouch et al., 2002; Wu et al., 2002) using a widearray of molecular markers. Such maps have the potential to
facilitate the genetic analysis of complex traits (quantitative traitloci, QTLs) such as drought resistance.
Efforts to improve the drought resistance of rice and thusincrease yield in drought-prone rainfed areas led to the publicationof a manual for rice drought-resistance breeding (Fischer et al.,2004b). This book was designed primarily for inform plantbreeders about strategies for development of drought-resistantcultivars, including direct selection for yield under stress andindirect selection for physio-morphological traits. However, thebook does not fully deal with recent progress using moleculartechniques.
In this review, we emphasize results from a population derivedfrom CT9993 and IR62266. This population is unique in that both ofthe parents have different drought-resistance traits (e.g., Zhanget al., 2001a; Kamoshita et al., 2002a) and that the population hasbeen tested under rainfed field conditions. However, comprehen-sive meta-analysis of QTL collocation has not been conducted.Earlier research on drought-resistance traits in rice was reviewedby O’Toole (1982), Fukai and Cooper (1995), Nguyen et al. (1997),and Fukai et al. (1999) and will not be reviewed in detail here.
2. Types of drought and drought-resistance traits
2.1. Types of drought
Different types of drought develop in rainfed rice fields, andcharacterization of the prevalent drought regimes for the regionconcerned is essential for identifying appropriate drought-resistance traits. Types of drought can be classified first accordingto the rice ecosystem—rainfed lowland or upland. Upland ricefields seldom experience standing water, and the rice crop is byand large subjected to mild stress conditions continuously withmore severe stress developing between major rainfall events,particularly under conditions with high evapotranspiration. On theother hand, rainfed lowland rice commonly has standing water inthe paddies. However, the water may disappear during rainlessperiods, and drought may develop rapidly at any time during cropgrowth, suddenly affecting the crop and reducing crop yield.
Types of drought can be identified secondly by the nature of thedrought, such as the severity and timing of the drought in relationto the stage of crop development. Three common types of droughtin terms of timing may be recognized for rainfed lowland rice(Chang et al., 1979). Fig. 1 illustrates these types in relation to freewater level in paddies (Fischer et al., 2004a).
Terminal drought mainly develops towards the end of thegrowing season, but it sometimes begins well before flowering.Terminal drought occurs in northeastern Thailand and southernIndia in years when monsoon rains cease before rice maturity.Pandey et al. (2005) showed that in eastern India terminal droughtis the most frequent type of drought and severely affects yield. Freewater level from 3 weeks before anthesis until maturity is closelyrelated to grain yield in rainfed lowland rice (Fukai et al., 2001;
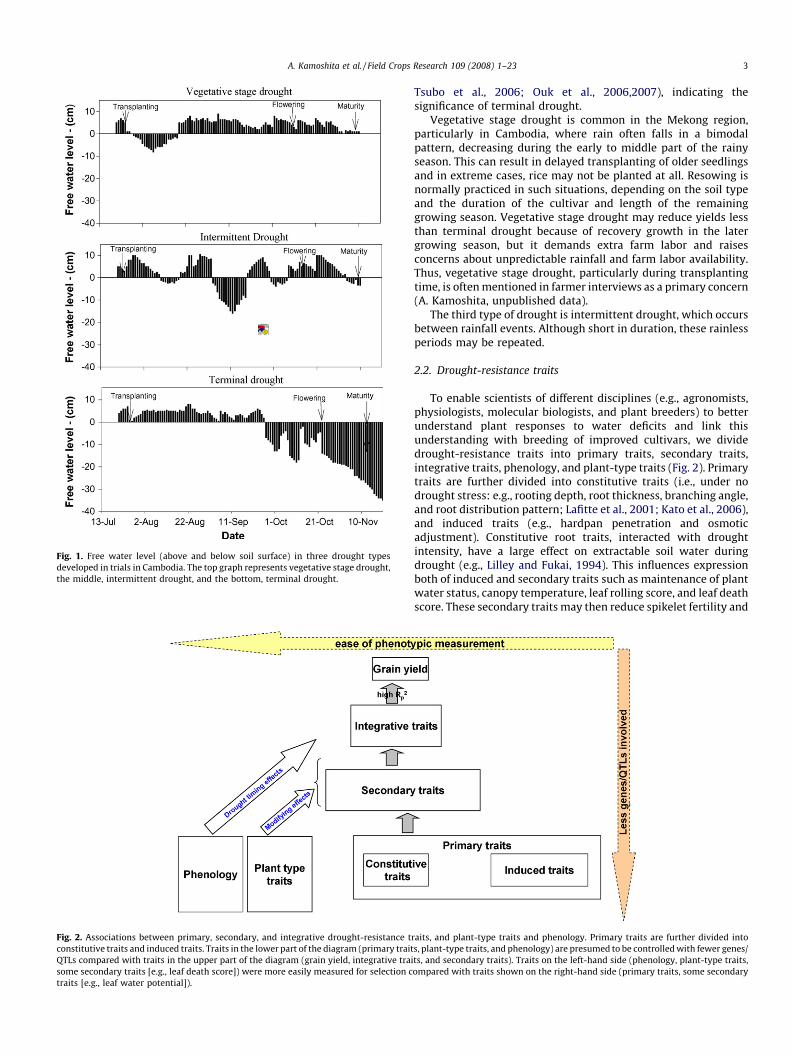
Fig. 1. Free water level (above and below soil surface) in three drought types
developed in trials in Cambodia. The top graph represents vegetative stage drought,
the middle, intermittent drought, and the bottom, terminal drought.
Fig. 2. Associations between primary, secondary, and integrative drought-resistance t
constitutive traits and induced traits. Traits in the lower part of the diagram (primary trait
QTLs compared with traits in the upper part of the diagram (grain yield, integrative trai
some secondary traits [e.g., leaf death score]) were more easily measured for selection c
traits [e.g., leaf water potential]).
A. Kamoshita et al. / Field Crops Research 109 (2008) 1–23 3
Tsubo et al., 2006; Ouk et al., 2006,2007), indicating thesignificance of terminal drought.
Vegetative stage drought is common in the Mekong region,particularly in Cambodia, where rain often falls in a bimodalpattern, decreasing during the early to middle part of the rainyseason. This can result in delayed transplanting of older seedlingsand in extreme cases, rice may not be planted at all. Resowing isnormally practiced in such situations, depending on the soil typeand the duration of the cultivar and length of the remaininggrowing season. Vegetative stage drought may reduce yields lessthan terminal drought because of recovery growth in the latergrowing season, but it demands extra farm labor and raisesconcerns about unpredictable rainfall and farm labor availability.Thus, vegetative stage drought, particularly during transplantingtime, is often mentioned in farmer interviews as a primary concern(A. Kamoshita, unpublished data).
The third type of drought is intermittent drought, which occursbetween rainfall events. Although short in duration, these rainlessperiods may be repeated.
2.2. Drought-resistance traits
To enable scientists of different disciplines (e.g., agronomists,physiologists, molecular biologists, and plant breeders) to betterunderstand plant responses to water deficits and link thisunderstanding with breeding of improved cultivars, we dividedrought-resistance traits into primary traits, secondary traits,integrative traits, phenology, and plant-type traits (Fig. 2). Primarytraits are further divided into constitutive traits (i.e., under nodrought stress: e.g., rooting depth, root thickness, branching angle,and root distribution pattern; Lafitte et al., 2001; Kato et al., 2006),and induced traits (e.g., hardpan penetration and osmoticadjustment). Constitutive root traits, interacted with droughtintensity, have a large effect on extractable soil water duringdrought (e.g., Lilley and Fukai, 1994). This influences expressionboth of induced and secondary traits such as maintenance of plantwater status, canopy temperature, leaf rolling score, and leaf deathscore. These secondary traits may then reduce spikelet fertility and
raits, and plant-type traits and phenology. Primary traits are further divided into
s, plant-type traits, and phenology) are presumed to be controlled with fewer genes/
ts, and secondary traits). Traits on the left-hand side (phenology, plant-type traits,
ompared with traits shown on the right-hand side (primary traits, some secondary

A. Kamoshita et al. / Field Crops Research 109 (2008) 1–234
yield components (i.e., integrative traits), and ultimately, yield(Kobata et al., 1994). Plant-type traits such as tiller number andplant height modify the expression of secondary and integrativetraits by affecting transpirational demand. Genotypes with greaterplant height are often larger in overall plant size, intercept morelight and use water faster by transpiration, leading to lower plantwater status (Kamoshita et al., 2004), higher leaf death scores, andmore spikelet sterility (Pantuwan et al., 2002c; Kato et al., 2007b).Phenology, interacted with timing of drought, has a large effect onyield through integrative traits.
Plant-type traits (e.g., plant height) and phenology (e.g.,flowering time) usually are highly heritable (discussed in Section3.1) and are extensively used in traditional plant breeding (Cooperet al., 1999a,b). Leaf rolling and canopy temperature (i.e.,secondary traits) are also useful (Lafitte et al., 2004a; Hirayamaet al., 2006) for quickly screening hundreds of lines. Althoughyields under stress generally have a higher phenotypic correlationwith some of the yield components (e.g., grain number and percentfertile spikelets; integrative traits) than they do with primarytraits, primary traits are likely to be controlled by fewer underlyinggenes or QTLs. Therefore, molecular characterization of primarytraits (e.g., QTL analysis or candidate gene approaches) willpresumably be a more promising avenue than the study of yieldcomponents. Recent transgenic studies have examined onlyprimary traits of which the expression is induced by water deficit,as is discussed in Section 4.3.
Trait type classifications are to some extent arbitrary. Thus, weclassify flowering delay as an integrative trait. The plant traits thatconfer drought resistance depend on the type of drought. Thefollowing section describes the physiological and morphologicaltraits likely to be useful against the three common types ofdrought.
2.2.1. Terminal drought
Early flowering genotypes can escape from late season drought,and this is a simple, but often the most effective, way of increasingyield under terminal drought. Replacing late maturing cultivarswith medium maturing cultivars that have good yield potential inrainfed lowlands, as has occurred in Cambodia, provides a betterchance of escaping late season drought (Ouk et al., 2007). Earlierflowering time may be especially useful for upper positions in atoposequence, because standing water often disappears earlierthere than in lower positions (Homma et al., 2003). Sets of uplandexperiments with diverse germplasm (i.e., indica, aus, and japonica
subspecies) also show the advantages of earlier flowering overlater flowering in terms of higher spikelet fertility, higher harvestindex, and higher yield (Lafitte and Courtois, 2002).
Among the genotypes with similar flowering times, main-tenance of high leaf water potential (LWP) was often related withhigher yields under terminal drought (Jongdee et al., 1997a, 2002;Pantuwan et al., 2002c; Jearakongman, 2005). Maintenance ofhigher LWP under drought is empirically related to better stemextension and panicle exsertion (Jearakongman, 2005), as well asto reduced delay in flowering (Pantuwan et al., 2002b). Jearakong-man (2005) found a positive relationship between panicleexsertion rate and LWP under drought among 55 near-isogeniclines of the cultivar IR64 into which root trait QTLs from thecultivar Azucena had been introduced (Shen et al., 2001).
Root signals such as ABA play important roles for stomatalcontrol under mild drought and hydraulic force becomesimportant under severe drought (Tardieu and Davies, 1993; Aliet al., 1999; Comstock, 2002), but evidences are few in rice thatgenotypic differences in LWP are related with root signals such asABA (Siopongco et al., 2008). It was suggested that some genotypesmay have stronger stomatal sensitivity to root signals in response
to soil water deficit to save water loss from stomata (Tardieu andDavies, 1993) and that others may continue higher assimilation ofCO2 in spite of high proportion of closed stomata (Hu et al., 2006).
Sibounheuang et al. (2001) showed consistent ranking of LWPas an important characteristic of drought resistance among sixgenotypes under different transpirational demands (by defoliationtreatment) and under similar soil water availability (by mixedplanting treatment) under upland drought conditions. Thissuggests an association between shoot water potential (e.g., inleaves and panicles) and internal plant water conductivity (Tsunoet al., 2004; Sibounheuang et al., 2006). Sibounheuang et al. (2006)found xylem diameter likely to be related to maintenance of xylemwater conductivity; however, this needs to be tested in a widerrange of germplasm together with vascular constrictions betweenroots and shoots. Umayal et al. (2001) observed that drought-tolerant indica landraces in southern India had thicker roots withwider xylem vessels (similar dimensions to the diameters of roots,steles, and xylem vessels in traditional upland japonica cultivars)than susceptible cultivars. Wider xylem vessels may decreasechances of cavitation, which would reduce xylem water con-ductivity (Hsiao, 1973; Hacke et al., 2001; Stiller et al., 2003).
Under terminal drought, slow use of water (conservativestrategy) is beneficial if the effective root depth is limited. Inrainfed lowland experiments in Thailand, where a severe droughtdeveloped suddenly, soil water from 0 to 45 cm soil depth tendedto be extracted more slowly during 7 days of no water supply atbooting stage by genotypes with smaller shoot systems that havereduced water demand (Pantuwan et al., 2002c). These genotypesproduced higher yields under the severe drought. This strategyappears particularly useful when drought develops rapidly late inthe growing season and the crop has high biomass and high waterrequirements. Genotypic variation in the amounts of epicuticularwax (EW) has been reported, production of EW increases underwater stress, which can reduce non-stomatal water loss fromleaves, but its effect on yield has not yet been demonstrated(Srinivasan, 2005).
Deeper and thicker roots also may occur under uplandconditions and some lowland conditions, resulting in the genotypeextracting larger amounts of soil water and maintaining higherplant water status (e.g., Yoshida and Hasegawa, 1982). Kumar et al.(2004) reported an association between higher root pullingresistance (RPR) and maintenance of higher LWP under severedrought stress, as well as a positive correlation between grain yieldand relative water content (r = 0.72, P = 0.01) and RPR (r = 0.59,P = 0.05). By using a system that restricted root growth to no morethan 25 cm below the soil surface by means of a water-permeablesheet, Kato et al. (2007b) showed that genotypes with deeper rootdevelopment (and with larger plant size) could maintain higherLWP when there was no root restriction, but suffered from agreater reduction in LWP and increased leaf death score with thepresence of root restriction. This shows that a well-developed rootsystem can help maintain plant water status, ultimately stabilizingyield under drought.
The ability of rice to function well at reduced plant waterpotentials has not been demonstrated to be useful under fielddrought regimes. In pot experiments, Capell et al. (2004) showedpositive effects of polyamine accumulation under drought condi-tions induced by replacing water with 20% polyethylene glycol for6 days. They identified several genotypes with high sperminecontent, a polyamine involved in cellular metabolism, which mayassist plant cells to maintain normal metabolisms under stress, andthey showed that transgenic plants overproducing polyaminesmaintained growth under water stress. Osmotic adjustment allowsmaintenance of growth when water uptake from the root system isinsufficient due to reduced availability of soil moisture, but

A. Kamoshita et al. / Field Crops Research 109 (2008) 1–23 5
possibly, it also could increase extraction of water by the roots(Serraj and Sinclair, 2002). Genotypic variation in osmoticadjustment has been demonstrated in rice (Lilley et al., 1996;Babu et al., 2001). However, there has been no report demonstrat-ing any positive phenotypic or genetic correlation between grainyield and osmotic adjustment in rice under stress. Genotypicvariation in cell membrane stability is also reported in rice(Tripathy et al., 2000). Babu et al. (2004b) showed that transgenicrice with HVA1, a LEA gene from barley, had less membrane leakagethan non-transgenic rice cultivars under water stress. Carbonisotope discrimination, often associated with transpiration effi-ciency, also varies with rice genotypes (Dingkuhn et al., 1991; Priceet al., 2002c), but its association with grain yield under drought hasnot been demonstrated (Laza et al., 2006).
Several studies have attempted to clarify physiologicalmechanisms underlying the genotypic variation in spikeletsterility under identical conditions of internal plant water deficit.Maintenance of open stomata, and hence continuous supply ofcarbohydrates to spikelets and higher enzyme activities (Sheoranand Saini, 1996), might assist fertilization and reduce spikeletsterility (Saini, 1997; Saini and Lalonde, 1998). The movement ofcarbohydrates to the anthers and peduncles is controlled notsimply by supply from source tissues but also by sink strength andactivity. The sink strength and activity is presumed to involve theGA versus ABA balance in inflorescences, ABA synthesis duringstress and its degradation during recovery (Zhang et al., 2006),expression of glycoprotein in pollens, and cell-wall acid invertaseactivity in anthers and pollens (Bennett et al., 2005). The drought-resistant cultivar N22 had higher spikelet fertility than thedrought-sensitive cultivar N118 as a result of increased antiox-idant enzyme activities in the panicles under water stress (Seloteand Khanna-Chopra, 2004). Liu et al. (2006) showed a smallerreduction in spikelet fertility in Moroberekan (16%) than in IR64(80%) after 6 days of withholding water, starting 3 days beforeheading. This smaller reduction in spikelet fertility was attributedto better anther dehiscence and higher stigma pollen density inMoroberekan.
2.2.2. Vegetative stage drought
Little research appears to have been conducted on genotypicvariation in rice in terms of germination (e.g., the lowest limits ofsoil water content for germination and emergence) or transplant-ing under water limiting conditions in rainfed lowlands. Ikeda et al.(2007) found a cultivar-by-water-regime interaction for rootingability after transplanting: a lowland cultivar produced new rootsmore quickly and allocated greater biomass to roots than anupland cultivar in the presence of standing water, but this was notthe case without standing water. Seedling vigor may allow fasterdevelopment of deep roots before or during the early stages ofdrought, thus accelerating water extraction and maintaininggrowth during drought (Kamoshita et al., 2000,2004). Landracesfrom southern India typically have higher seedling vigor, withrapid root and shoot growth and earlier ground coverage thanimproved cultivars. Price et al. (2002a,b) showed large interactiveeffects of initial soil moisture content on genotypic deep rootdevelopment and partitioning of assimilates during the seedlingstage when plants were grown in small containers.
Because of the long time from the time of stress to harvest,drought-resistance traits during vegetative stage drought may notbe related to grain yield (Lafitte et al., 2002a). Plant growthresumes after vegetative stage drought, and this recovery growththen affects the development of sink size as well as source supplyto meet the demand of the grain. Field studies (Lilley and Fukai,1994; Mitchell et al., 1998) and pot studies (Wade et al., 2000;Kamoshita et al., 2004) both show genotypic variation in short-
term recovery growth (e.g., 1 week to a few weeks) after vegetativestage drought, and these authors have reported the relationshipsbetween this genotypic variation and the amount of leaf remainingat the end of drought and the ability to tiller after drought.Although the benefits of short-term drought recovery traits onyield are difficult to demonstrate, a number of studies have shownthat later maturing and longer growth duration cultivars show lessgrowth stagnation and drought damage and have a higher yieldwhen they encounter mild water shortages during the vegetativeto panicle initiation stages (e.g., Fukai and Cooper, 1995; Hayashiet al., 2006; Ikeda et al., 2008).
2.2.3. Intermittent drought
A deep root system with higher root density is likely to be usefulunder intermittent drought if growing conditions permit rootdevelopment at depth. Under upland conditions, the associationbetween root length density and the amount of water extractedhas been well demonstrated (Lilley and Fukai, 1994), including forgenotypic variations (Nemoto et al., 1998; Kato et al., 2007a), anddeep and thick root traits contribute to better growth and higheryield under drought stress (Lafitte and Courtois, 2002; Babu et al.,2003). In CT9993/IR62266 doubled haploid (DH) lines, three roottraits measured in glasshouse experiments – root thickness, deeproot weight, and root penetration index – were correlatedpositively with yield and yield components under severe pre-flowering drought conditions in an upland field experiment (Babuet al., 2003).
On the other hand, deep rooting is often poorly expressed underrainfed lowland conditions (Pantuwan et al., 1997), and there ismuch less evidence of genotypic variation in root traits with regardto water extraction during drought (Kamoshita et al., 2000, 2004).The extent of deep root development measured by partitioningratio (deep root ratio) is less under anaerobic flooded conditions(ca. 0.3–1.2%) than under aerobic drought conditions (ca. 3–17%) inpot experiments (Azhiri-Sigari et al., 2000; Kamoshita et al., 2004).Genotypic rankings for deep root traits expressed under anaerobicconditions were generally similar to those expressed under aerobicconditions, but in some cases genotype by environment interac-tions were detected (Champoux et al., 1995; Azhiri-Sigari et al.,2000). Genotypic differences in the ability to penetrate compactedsoil layers have been reported (Babu et al., 2001; Samson et al.,2002); these differences are associated with the amount of waterextracted from below the compacted layer (Hoque and Kobata,2000; Kobata et al., 2000).
3. Selection of drought-resistant genotypes
3.1. Screening environments
Phenotyping facilities for screening rice genotypes for droughtresistance are increasingly available. Lafitte (2004) proposed theuse of a managed drought environment (i.e., a drought conditioncontrolled or intentionally manipulated to some degree byscientists) in order to provide the desired severity and timing ofthe drought. There are a number of advantages to such a system:(1) natural drought occurrence in rainfed lowland and upland ricein the wet season is unpredictable, thus limiting screening underthe desired drought types; (2) managed drought uses resources(e.g., the budget of a drought project of only a few years) moreefficiently than waiting for the occurrence of natural drought in thewet season; and (3) there is a high genetic correlation betweenyield under stress in a managed selection environment and that inthe target environment (Atlin, 2004). However, there are alsoweaknesses in screening genotypes from managed droughtenvironments (Kamoshita, personal communication). We will

A. Kamoshita et al. / Field Crops Research 109 (2008) 1–236
briefly discuss the strengths and weaknesses of several screeningenvironments.
A controlled drought environment can be established morereadily in the dry season (e.g., Lafitte et al., 2006), but theenvironment may not represent that of the farmer’s fields in thewet season and will confound the effects of photoperiod insensitive genotypes. Pantuwan et al. (2004) reported thatresponses to dry conditions among genotypes in the dry seasondiffered from responses under wet season drought. Nevertheless,the dry season may be useful for preliminary vegetative stagescreening of large numbers of genotypes (Pantuwan et al., 2004).
To reduce the chance of rainfall interfering with droughtdevelopment in the wet season, trials may be planted later andwater may then be drained from the field so that the crop has abetter chance of being exposed to drought; this is particularly sofor terminal drought trials (Pantuwan et al., 2002a; Jongdee, 2004).However, this method also causes bias against photoperiodsensitive cultivars. Late planting may not be readily achieved insome locations, as in southern India where pre-monsoon directseeding is practiced. Drainage techniques can create severe stressunder lowland conditions (Kumar et al., 2007), and the timing ofdrainage can be adjusted to induce different types of drought(Pantuwan et al., 2002a; Ouk et al., 2006). Another practice forestablishing a managed drought environment is to use a highposition in the toposequence (Kumar et al., 2007; Hayashi et al.,2007). Standing water disappears naturally often a few weeksearlier than it does in lower positions. A Thai breeding programcurrently utilizes different toposequence positions, and cultivarsare selected for different toposequence positions after considera-tion of the large differences in the water and soil nutrientenvironments (Homma et al., 2003; Tsubo et al., 2006,2007).
Rainout shelters can reduce damage from untimely rainfall andin China are used with drip irrigation and drainage systems(O’Toole, 2004; Li et al., 2005; Yue et al., 2005; Liu et al., 2008).However, the high cost of such facilities may limit their widespreaduse.
A Japanese upland rice breeding program uses raised beds (ca.30 cm above ground level) to screen drought-resistant genotypes(Hirayama and Suga, 1996; Kato et al., 2007b). These beds areprepared over a 5-cm-thick layer of gravel laid on the soil surface toprevent capillary rise of water into the raised bed. Kato et al.(2007b) showed that, compared with the natural rainfed uplandconditions, rice crops grown in the raised bed system were exposedto less soil moisture in the surface layers during rain-free periods,increasing the differences in yield between shallow- and deep-rooting cultivars.
3.2. Selection criteria
For efficient screening, Blum (2002) suggested two points forassessing the utility of traits. The first is that important drought-resistance traits are normally constitutive and not stress adaptive.
Table 1Broad sense heritability measured either greenhouse (pot) or field experiments for drou
and grain yield) under different drought stress types (non-stress, vegetative, intermitt
Measured systems
Drought-resistance traits drought stress types Pot
Primary traits
Deep and thick rootsa Non-stress 0.60–0.84b, 0.40–0
0.55–0.84e, 0.61–0
Terminal –
Intermittent 0.62–0.92f (0.50–0
Hard soils 0.37–0.58h
Constitutive traits, such as flowering time, the stay-green trait(delayed onset of leaf senescence), and root depth, can be routinelyscreened (without a drought challenge); generally, their roletowards drought resistance may be considered greater than stress-responsive traits. Stress adaptive traits (responsive, induced) arethose that are expressed only under drought. Such traits includeactive cellular accumulation of compatible solutes (osmoprotec-tants), antioxidant agents, heat shock proteins, and molecularchaperones, as well as osmotic adjustment and membranestability. Blum’s second point is that plant water status, ratherthan plant function, controls crop performance under drought.Therefore, those genotypes that can maintain higher LWP, andRWC are drought resistant simply because of their superiorinternal water status. He argues that cases of sustained functionunder low water status – a possible alternative for droughtresistance – are rare. His argument supports the classification ofrice as a drought avoider, and maintenance of plant water status,rather than tolerance of drought with low plant water potential, isthe key to drought resistance. For screening, traits, which are morelikely to be constitutive but may also include induced traits, needto be highly heritable and easily measured.
Calculation and interpretation of broad sense heritability (i.e.,approximately 0.9 as high, 0.6 as moderate, and 0.2 as low) wereexplained by Lafitte et al. (2004a). Numerous studies have foundmoderate heritability of yield under drought conditions (Babuet al., 2003; Atlin et al., 2004; Yue et al., 2005; Kumar et al., 2007),supporting direct selection for yield, rather than for drought-resistance traits (Table 1). Kumar et al. (2007) reported higherbroad-sense heritability of grain yield under severe terminaldrought stress in 2 years (0.37–0.54) in the CT9993/IR62266population than for secondary and integrative drought-resistancetraits such as harvest index (0.22–0.36), spikelet sterility (0.12–0.23), flowering delay (0.12–0.21), relative water content (0.07–0.13), root pulling force (0.32–0.49), or root dry weight (0.27–0.43),all under field conditions (Table 1). Heritability of yield understress is sometimes comparable with heritability under non-stressconditions (e.g., Kumar et al., 2007; Lafitte et al., 2004b) but alsocan be lower (e.g., 0.43 under terminal drought stress, 0.53–0.62under intermittent stress, 0.81 under non-stress conditions inJearakongman, 2005). In Brazil, direct selection for yield using amanaged drought environment that matches the environment inthe breeding domain has resulted in the identification of tolerantgenotypes and development of higher yielding cultivars for uplandrice (Pinheiro, 2004; Pinheiro et al., 2006). This approach has alsobeen used for rainfed lowland rice in Thailand and Cambodia(Jongdee, 2004; Ouk et al., 2006,2007).
On the other hand, because primary drought-resistance traitsare associated with fewer genes and are under simpler geneticcontrol than yield under stress, the heritability of primary traitsshould be higher than for secondary and integrative traits andyield, assuming that trait expression is accurately measured. In theCT9993/IR62266 population, primary traits such as deep and thick
ght-resistance trait groups (primary, secondary, integrative, plant-type, phenology,
ent, terminal)
Field
.81c, 0.44–0.61d,
.90f, 0.26–0.83g
0.80h, 0.32–0.49i
0.27–0.43i
.63)f, 0.50–0.92g (0.40–0.67)g –
–

Table 1 (Continued )
Measured systems
Drought-resistance traits drought stress types Pot Field
Cell membrane stability Vegetative (severe) 0.34j –
Osmotic adjustment Vegetative (gradual lengthy) 0.62h –
Secondary traits
Relative water content Intermittent – 0.39k
Terminal – 0.07–0.13i, 0.49–0.73l
Leaf water potential Non-stress – 0.41–0.53m
Intermittent – 0.43–0.46m
Terminal – 0.50–0.63m
Canopy temperature Terminal – 0.86l, 0.5–0.68n
Leaf death and rolling Intermittent 0.79f 0.33–0.63k, 0.78l, 0.69–0.83m, 0.58–0.79n
Terminal – 0.65–0.71l, 0.86–0.92m
Panicle exsertion Non-stress – 0.25m
Intermittent – 0.58–0.69m
Terminal – 0.55m
Integrative traits
Hearvest index Non-stress – 0.24l
Terminal – 0.22–0.36i, 0.60l
Spikelet sterility Non-stress 0.29–0.45i, 0.59k, 0.65–0.69l, 0.61–0.67m
Intermittent – 0.38k, 0.65–0.76m
Terminal – 0.12–0.23i, 0.43l, 0.63
Grain number per panicle Non-stress 0.89k, 0.51m
Intermittent – 0.67k, 0.62–0.70m
Terminal – 0.50l, 0.65m
1000-Grain weight Non-stress – 0.84k
Intermittent – 0.91k, 0.40m
Terminal – 0.37l, 0.28m
Biomass Non-stress 0.26–0.42i, 0.60–0.78l
Intermittent – 0.84l
Terminal – 0.40–0.58i, 0.37–0.60l
Drought response index Intermittent – 0.36–0.42m
Terminal – 0.46m
Flowering delay Terminal – 0.12–0.21i, 0.17k, 0.44–0.69n
Phenology
Flowering Non-stress – 0.36–0.53i, 0.94k, 0.75
Intermittent – 0.93k, 0.96l
Terminal – 0.32–0.48i, 0.70l
Plant-type traits
Plant height Non-stress – 0.78k, 0.94m
Intermittent – 0.66k, 0.93–0.95m
Terminal – 0.90m
Grain yield
Non-stress – 0.45–0.62i, 0.45k, 0.61–0.84l, 0.81m, 0.72–0.92n
Intermittent (0.86)f 0.45k, 0.81l, 0.53–0.62m, 0.73–0.84n
Terminal – 0.37–0.54i, 0.59l, 0.43m
Terminal and intermittent stress types were distinguished from each other by examining the reduction percentages of yield under stress relative to yield under non-stress
conditions in each reference. Values in ( ) indicate traits induced in response to water stress; these are either relative values (stress/non-stress) or increment under stress
(stress–non-stress).a Deep root weight below 30 cm depth, deep root ratio, deep root weight per tiller number, maximum root length, rooting depth, total root weight, root volume, root
thickness at depth.b Yadav et al. (1997).c Kamoshita et al. (2002a).d Ekanayake et al. (1985).e Courtois et al. (2003).f Yue et al. (2006).g Zheng et al. (2003).h Zhang et al. (2001a).i Kumar et al. (2007).j Tripathy et al. (2000).k Lafitte et al. (2004a,b).l Babu et al. (2003).m Jearakongman (2005).n Yue et al. (2005).
A. Kamoshita et al. / Field Crops Research 109 (2008) 1–23 7

Table 2aLiterature review of studies to identify QTL clusters correlating with plant-type traits, phenology, primary traits, secondary traits, and yield and integrative traits for drought
resistance across 15 mapping populations
Populationa Phenotyping
environmentb
Measured or mapped traitsc Reference
Plant-type Phenology Primary Secondary Yield and
integrative
1 CT/IR 100,154 Upland (Si),
lowland (W, Si)
– – – CT, DRS, DS,
LD, LR
GY, BY, GW, PSS Zhang et al. (1999)
CT/IR 104 Uplandp (S) – – CMS – – Tripathy et al. (2000)
CT/IR 154 Uplandp (S, hardpan) – – OA, PRL, PRT, PRW,
RPI, RT, RPF, TRW
– – Zhang et al. (2001a)
CT/IR 154 Lowlandp
(W, seedling stage)
– – DR%, DRW, DRW-T,
RD, RT
– SDW Kamoshita et al. (2002a)
CT/IR n.a. Uplandp (W) PH, TN, TPL – RN, RSLR, TRW – – Kanbar et al. (2002)
CT/IR 154 Upland (W, Si, St;
2 wet & 1 dry
seasons)
– – – LR, LD, RWC GY, BY, HI, SN Babu et al. (2003)
CT/IR 154 Lowland (W, Si, St;
wet season)
PN – – – GY, BY, SN,
PSS, GW
Lanceras et al. (2004)
CT/IR n.a. Upland (W, S) PH, PL, PTN DH EW CT, LD, LR, RWC GY, BY, HI, SY Srinivasan (2005)
CT/IR 105 Lowland (W, St;
2 years)
– DH RPF, TRW GY, BY, DFT,
HI, PSS, SDW
Kumar et al. (2007)
2 Co/Mo 203 Uplandp (W),
Upland (S; 3)
– – DRW-T, MRL, RSDR,
RT, TRW,
LR – Champoux et al. (1995)
Co/Mo 202 Uplandp (W,
hardpan)
– – PRN, RN, RPI – – Ray et al. (1996)
Co/Mo 42 Uplandp (S) – – DT, OA RWC – Lilley et al. (1996)
3 IR/Az 105 Uplandp (W) – – DRSR, DRW, DRW-T,
MRL, RT, TRW
– – Yadav et al. (1997)
IR/Az 85-105 Upland (S; 3
sites/seasons)
– – – LD, LR, RWC RGR Courtois et al. (2000)
IR/Az 56 Uplandp (W, S (mild
vegetative))
PH, TN – MRL, RL, RN, RSDR,
RT, TRW, TRW-T
– SDW Hemamalini et al. (2000)
IR/Az 109 Uplandp (W,
hardpan)
– – PRN, PRT, RN, RPI – – Zheng et al. (2000)
IR/Az 90 Lowland (W),
upland (Si),
uplandp (W, S)
PL, PTN, TN DH, DM MRL, RN, RSDR,
RT, RV, TRW
– GY, SDW, SY Venuprasad et al. (2002)
IR/Az 85 Upland (W, S
(flowering); 3
dry seasons)
PH, PN DH – – GY, RGY, GP,
GW, PSS, SN
Lafitte et al. (2002b)
4 Ba/Az 178# Lowlandh – – MRL, RCL, RT – – Price and Tomos (1997)
Ba/Az 178# Uplandp DH – MLRS, RSC,
SR, TFSC
– Price et al. (1997)
Ba/Az n.a. Upland – – RL – – Price and Courtois (1999)
Ba/Az 104 Uplandp – – PRN, RN, RPI – – Price et al. (2000)
Ba/Az 110–176 Upland (S; 2
sites in 3 dry
seasons)
– – – LD, LR, RWC – Price et al. (2002d)
Ba/Az 140 Uplandp (S; 2
seedling drought)
– – DRN, DRW, MRL,
RSDR, RT, TRW
– – Price et al. (2002b)
Ba/Az n.a. Uplandp (W; 2 dry
seasons)
– – d13C, SLA – – Price et al. (2002c)
Ba/Az 96 Upland (W, Si
(flowering);
2 dry seasons)
PH, PN DH – LD, LR, RWC GY, BY, FSP, GP,
GW, HI, SF
Lafitte et al. (2004b)
Ba/Az 177 Upland PH, PL DH – LD, LR GY, SY Gomez et al. (2005)
Ba/Az 168 Uplandp (W, S) PH – MRL, RT, DRW SDW MacMillan et al. (2006)
5 IR/IR 166 Uplandp (W, hardpan) – – PRL, PRN, RPI, PRT, RN – – Ali et al. (2000)
IR/IR 166 Lowlandp (W;
seedling stage)
– – DR%, DRW,
DRW-T, RD, RT
– SDW Kamoshita et al. (2002b)
6 IA/Co 125 Uplandp (W) PH, TN – DRSR, DRW, DRW-T,
MRL, RT, TRW
– SDW Courtois et al. (2003)
7 I/I 150 Uplandp (S) – – OA – – Robin et al. (2003)
8 I/A 150 Lowlandh (W),
uplandpa (W)
– – SRL – – Zhang et al. (2001b)
I/A 96 Lowlandp (W),
uplandp (S; mild)
– – ARN, LRL,
LRN, SRL
– – Zheng et al. (2003)
9 I/Yu 116 Lowland (W),
upland (Sv),
uplandp (Sv)
– – MRL, RN, RSDR,
RSFR, RT,
TRFW, TRW
– GY, IDR Li et al. (2005)
A. Kamoshita et al. / Field Crops Research 109 (2008) 1–238

Table 2a (Continued )
Populationa Phenotyping
environmentb
Measured or mapped traitsc Reference
Plant-type Phenology Primary Secondary Yield and
integrative
10 Z/I 180 Lowland (W, Si
(reproductive);
2 soils), uplandp
(W, St)
– – DRV, MRD,
RGV, RT
CT, LD, LR DFT, DRI, RGY, RSF Yue et al. (2005)
Z/I 150 Uplandp (W, Si
(flowering))
– – DIRD, DIDRV,
DRV, MRD, RGD,
RGV, RV
DLR, LD RGY, RBY, RFP,
RGW, RHI, RSF, RSN
Yue et al. (2006)
Z/I 187 Upland (W, Si) PN – – – GY, GP, GW, SF Zou et al. (2005)
Z/I 187 Upland (W, Si
(reproductive
stage))
PL, PN – – – GP, PBN, PND, SBN, SN Liu et al. (2008)
11 T/L 254 Lowland (W, St;
dry season)
PH, PN DH – – GY, GW Xu et al. (2005)
12 I/N n.a. Upland (W, S) PH, TN DH RT CT, DRS, LD,
LR, RWC
BY Boopathi et al. (2005)
I/N n.a. Upland (W, S) PH, PL, TN DH – CT, DRS, LD,
LR, RWC
GY, BY, HI, SY Beena (2005)
13 A/I 106 Lowlandh (W, S) – – – – RGR, WUE Kato et al. (2008)
A/I 106 Lowlandh (W) – – BI, RAL, TRW – – Horii et al. (2006)
14 O/Y 98 Lowland (W, S) PH, PN – MNRL, NRN, RA,
RSDR, TRW
– SDW, SPDW Ikeda et al. (2007)
15 V/W 436 Upland (W, S in 2
dry seasons)
PH, PN DH – – GY, DFT, DRI, HI, BY Bernier et al. (2007)
a The names of the populations are listed in Table 2b. The values under the population of each study signify the number of lines used to identify QTLs.b Subscript suffixes ‘‘p’’, ‘‘h’’, and ‘‘pa’’ stand for ‘‘pot experiments’’, ‘‘hydroponics’’, and ‘‘paper culture’’, respectively. W, S, Si, St, and Sv signify ‘‘well-watered’’, ‘‘stress’’,
‘‘intermittent stress’’, ‘‘terminal stress’’, and ‘‘vegetative stress’’, respectively.c Measured and mapped traits are grouped into (1) plant-type traits, (2) phenology, (3) primary traits, (4) secondary traits, and (5) yield and integrative traits.
(1) Plant-type traits contain plant height (PH), panicle length (PL), panicle number (PN), productive tiller number (PTN), tiller number (TN), and total plant length
(TPL).
(2) Phenology contains days to heading/flowering (DH), and days to maturity (DM).
(3) Primary traits contain (3a) constitutive root traits and (3b) other induced traits.
(3a) Constitutive root traits are total root weight (TRW), total root fresh weight (TRFW), root to shoot dry weight ratio (RSDR), root to shoot fresh weight ratio (RSFR),
deep root weight (DRW), deep root to total root dry weight ratio (DR%), deep root weight per tiller (DRW-T), deep root number (DRN), deep root to shoot dry
weight ratio (DRSR), rooting depth (RD), root length (RL), maximum root length (MRL), total root number (RN), root shoot length ratio (RSLR), adventitious root
number (ARN), lateral root number (LRN), lateral root length (LRL), seminal root length (SRL), root thickness at different depth (RT), root cell length (RCL),
maximum root depth (cm) (MRD), root volume (RV), deep root rate (%) in volume (DRV), root growth rate in depth (cm/day) (RGD), root growth rate in volume
(ml/day) (RGV), root pulling force (RPF), root axis length (RAL), branching index (BI), new root number (NRN), maximum new root length (MNRL), rooting ability
after transplanting (RA).
(3b) Other primary induced traits are drought induced root growth in depth (cm) (DIRD), deep root rate in volume (%) induced by drought (DIDRV), penetrated root
number (PRN), penetrated root thickness (PRT), root penetration index (RPI), penetrated root weight (PRW), penetrated root length (PRL), osmotic adjustment (OA),
cell membrane stability (CMS), carbon isotope discrimination as water use efficiency (d13C), specific leaf area (SLA), dehydration tolerance (DT).
(4) Secondary traits contain modified leaf rolling score (MLRS), leaf rolling (LR), number of days to leaf rolling (DLR), leaf drying (LD), drought score (DS), drought
recovery score (DRS), relative water content (RWC), canopy temperature (CT), stomatal resistance (SR), rate of stomatal closure (RSC), time of fastest stomatal
closure (TFSC).
(5) Grain yield (GY) and integrative traits contain straw yield (SY), biomass yield (BY), delay in flowering time by drought (DFT), primary and secondary branch
number in panicles (PBN, SBN), panicle neck diameter (PND), spikelet number (SN), percent spikelet sterility (PSS), fraction sterile panicle (FSP), grains per panicle
(GP), spikelet fertility (SF), 1000-grain weight (GW), harvest index (HI), shoot dry weight (SDW), single panicle dry weight (SPDW), water use efficiency (WUE),
relative grain yield (RGY), relative growth rate (RGR), relative spikelet fertility (RSF), relative biomass (RBY), relative rate of fertile panicle (RFP), relative harvest
index (RHI), relative grain weight (RGW), relative number of spikelets per panicle (RSN), panicle harvest index (PHI), drought response index (DRI), index of
drought resistance (IDR).
A. Kamoshita et al. / Field Crops Research 109 (2008) 1–23 9
roots (0.40–0.81 in Kamoshita et al., 2002a) and osmoticadjustment (0.62 in Zhang et al., 2001a) measured in greenhouseexperiments had heritabilities that were comparable to, or higherthan, that of yield under severe stress (0.59 in Babu et al., 2003;0.37–0.54 in Kumar et al., 2007) (Table 1). However, measurementsof root traits under drought conditions in the field usually havelarge errors (Pantuwan et al., 2002c; Samson et al., 2002), and thebroad-sense heritabilities of root traits measured in the field (e.g.,Kumar et al., 2007) are in general lower than those measured underhydroponic systems (Ekanayake et al., 1985) and pot systems(Kamoshita et al., 2002a; Yue et al., 2006). Measurement of inducedtraits such as osmotic adjustment or cell membrane stability couldinvolve greater errors than measurement of constitutive traitsbecause of differences in the degree of water stress if experimentalconditions are not precisely controlled. Even under managed
drought conditions (under both glasshouse and field conditions),expression of osmotic adjustment increases as drought intensifiesand LWP declines, resulting in the tendency toward a genotype-by-drought-intensity interaction for osmotic adjustment (Jongdeeet al., 2002; Kamoshita et al., 2000, 2004).
Heritabilities of secondary traits (such as plant water status,leaf death score, and leaf rolling) and integrated traits (such asspikelet sterility) are often comparable to or higher than theheritability of yield under stress. Jearakongman (2005) showedhigher broad-sense heritability for pre-dawn LWP (0.63), leafdeath and leaf rolling scores (0.86–0.92), panicle exsertion rate(0.55), plant height (0.90), total number of grains per panicle(0.66), and number of unfilled grains per panicle (0.75) than broad-sense heritability of yield under terminal stress (0.43) among 55IR64 near-isogenic lines (Table 1). In the CT9993/IR62266

A. Kamoshita et al. / Field Crops Research 109 (2008) 1–2310
population, Babu et al. (2003) measured secondary and integratedtraits in drought-affected fields and reported high broad-senseheritabilities for relative water content (0.49–0.73), canopytemperature (0.86), leaf rolling (0.65–0.78), leaf drying (0.70–0.71), harvest index (0.60), and spikelet fertility (0.43) comparedwith the broad-sense heritability of yield under stress (0.59–0.81).Integrative traits or secondary traits are likely to be more closelyassociated with grain yield under various drought conditions thanare primary traits, and they may have wider applicability inbreeding provided they add efficiency to selecting for yield per se.In the upland program in Brazil, integrative and secondary traits oflow leaf rolling, good panicle exsertion, and low level of spikeletsterility are used as selection criteria (Pinheiro et al., 2006).
Grain yield under drought is affected not only by drought-resistance traits but also by flowering time and by the potentialyield. The variation in potential yield and phenology can becorrected for by calculating a drought response index (DRI)(Pantuwan et al., 2002b; Lafitte, 2004; Ouk et al., 2006), anddrought-tolerant genotypes may be selected by using DRI.
4. Molecular approaches for development of drought-resistantvarieties
4.1. QTL mapping
QTLs linked to drought resistance have been mapped in at least15 different populations (Tables 2a and 2b). Most of the mappingpopulations were derived from indica � japonica parents, and it isoften the case that favorable alleles for drought-resistance traitsare contributed by japonica lines. Considering that indica andjaponica ecotypes are grown in different environments and thatmost breeding programs involve locally adapted rice accessions,the results for traits such as yield in indica � japonica populationshave to be interpreted with care. Hence, it is desirable to look forgenetic variation among indica ecotypes (e.g., IR58821/IR52561;Ali et al., 2000) as well as among japonica ecotypes (e.g., Akihikari/IRAT109, Otomemochi/Yumenohatamochi; Horii et al., 2006; Ikedaet al., 2007) (Ingram et al., 1994) and to map QTLs usingpopulations derived from lines adapted to target environments.
Although there are a number of untapped donors for drought-resistance traits available from wild species and landraces, nopopulation that uses a wild species as one of the parents has beendeveloped for drought studies. The possibility of selectivelyintrogressing useful genes from Oryza rufipogon to elite ricecultivars suggests a way of improving the performance of Oryza
sativa while broadening the genetic base (Xiao et al., 1998). Other
Table 2bThe 15 populations reviewed in this study; types of population and their parents
Population
CT/IR CT9993/IR62266 (DH)
Co/Mo Co39/Moroberekan (RI)
IR/Az IR64/Azucena (DH)
Ba/Az Bala/Azucena (F2, RI)
IR/IR IR58821/IR52561 (RI)
IA/Co IAC165/Co39 (RI)
I/I IR62266/IR60080 (BC)
I/A IR1552/Azucena (RI)
I/Yu IRAT109/Yuefu (DH)
Z/I Zhenshan97/IRAT109 (RI)
T/L Teqing/Lemont (BC)
I/N IR20/Nootripathu (RI)
A/I Akihikari/IRAT109 (BC)
O/Y Otomemochi/Yumenohatamochi (RI)
V/W Vandana/Way Rarem (F3–F5)
DH: doubled haploid lines; RI: recombinant inbred lines; F2–F5: F2–F5 populations; BC
wild species of interest include Oryza australiensis, Oryza glaber-
rima, Oryza officinalis, and Oryza nivara.A number of field studies provide information on QTLs linked to
grain yield and yield components under managed stress condi-tions, often during the dry season (Babu et al., 2003; Lanceras et al.,2004; Lafitte et al., 2004b; Xu et al., 2005; Yue et al., 2005;Jearakongman, 2005). It is important to summarize the currentstatus of QTLs for drought resistance and to evaluate the degree towhich they affect rice production under drought. By comparing thecoincidence of QTLs for specific drought-resistance traits and QTLsfor plant production under drought, it is possible to test whether aparticular constitutive or adaptive trait is likely to be useful forimproving drought tolerance in the field (Lebreton et al., 1995).
Here, we compare QTL analyses (i.e., a semi-meta analysis) toidentify the QTL segments that are expected to have an impact onrice yield under drought conditions. Several studies have beenconducted under both well-watered and drought-stressed condi-tions (e.g., Zou et al., 2005; Kumar et al., 2007), and two studies hadmultiple stress regimes (Lanceras et al., 2004; Jearakongman,2005), allowing preliminary assessment of the interaction betweenQTLs and drought stress types. We classified phenotypingenvironments into either well-watered control (C) or stress (S)conditions, and whenever possible, we further characterizeddrought stress environments as either intermittent (Si) or terminal(St). When the correlation coefficient between yield under stressand under well-watered conditions was relatively high (e.g.,r = 0.56**), only a few cases of QTL-by-environment interactionwere detected (Zou et al., 2005).
In such an analysis, we acknowledge several limitations:
(1) T
: bac
he sizes of the mapping populations are generally small (i.e.,about 100–200 lines, except for the 436 lines in Bernier et al.(2007)). For example, the progeny of 100 lines could cause alarge over-estimation of the phenotypic variance associatedwith detected QTLs (the Beavis effect) (Beavis, 1994; Xu, 2003).For a test with a significance level of 0.05, to achieve 90% powerfor detecting a QTL that accounts for 10% of the total traitvariance in a DH population, simulations show that samplesizes of n = 158 and 281 are required for recombinationfrequencies of 0.1 and 0.2, respectively (Hackett, 2002).
(2) P
henotyping protocols differ among studies. For example, thetraits measured for ‘‘deep and thick root system’’ varied.‘‘Thickness of roots’’ was measured at basal or deeper soil layers(e.g., Kamoshita et al., 2002b), and ‘‘deepness of roots’’ wasmeasured as maximum rooting length (e.g., Hemamalini et al.,2000), as seminal or adventitious root length (Zheng et al.,Parents
Upland tropical japonica/lowland indica
Irrigated lowland indica/upland japonica
Irrigated lowland indica with sd-1/upland tropical japonica
Upland indica with sd-1/upland tropical japonica
Lowland indica/lowland indica
Improved upland japonica/irrigated lowland indica
Lowland indica/elite lowland tropical japonica
Lowland indica/upland tropical japonica
Tropical upland japonica/lowland indica
Lowland indica/upland tropical japonica
Lowland indica with sd-1/lowland japonica with sd-1
Irrigated lowland indica/upland indica
Lowland japonica/upland tropical japonica
Lowland japonica/upland temperate japonica
Upland indica/upland tropical japonica
kcross population.

Fig. 3. Consistent QTLs mapped for drought resistance and plant production traits in rice chromosome 1 in different populations but using a common set of markers
(underlined). Parts of rice chromosome 1 are shown as vertical bars with mapped markers indicated to the right and a cM ruler to the left; the population used in genetic map
construction appears above the linkage group. Because of the differences in size and structure of the mapping populations, the different donors used for developing mapping
populations, the different DNA marker platforms, and the limited number of common DNA markers for comparison, placement of the common drought resistance QTLs on the
rice genetic linkage map in this review is tentative at best. The full names of the traits are indicated in Table 3.
A. Kamoshita et al. / Field Crops Research 109 (2008) 1–23 11

Fig.(und
A. Kamoshita et al. / Field Crops Research 109 (2008) 1–2312
2003), or as total or deep root dry weight (e.g., Yadav et al.,1997).
(3) T
here are differences in DNA marker platforms, limitednumbers of common DNA markers available for comparison,and inconsistent map distances across studies (e.g., Causseet al., 1994; McCouch et al., 2002).(4) P
henotyping errors are sometimes large. (5) R aw phenotypic and genotypic data were not available for allpopulations, as noted by Ahmadi et al. (2007), and we had torely on published data for our semi-meta analysis.
We first focused on previous studies using CT9993/IR62266,probably the most widely studied mapping population includingfield phenotyping, in which both of the parents are well adapted torainfed rice-growing environments. We identified 34 genomicregions with multiple QTLs for putative drought-resistance traits(i.e., primary, secondary, integrative, phenology, and plant-typetraits). Next, using maps developed by Causse et al. (1994),McCouch et al. (2002), and the International Rice GenomeSequencing Project (2005), we were able to compare with relativeprecision QTLs for drought-resistance traits that were mappedacross populations. We considered ‘‘common QTLs’’ to be thosethat were identified in the same marker intervals in bothpopulations when common markers were available in the area(i.e., the markers underlined in Figs. 3–6) (e.g., Kamoshita et al.,
4. Consistent QTLs mapped for drought resistance and plant production traits in
erlined). For full legend see Fig. 3.
2002b), as well as those found in both populations in the adjacentchromosomal segments judged by the locations of simplesequence repeats (SSRs) relative to genetic marker maps (i.e.,Supplementary Table 17 in International Rice Genome SequencingProject, 2005) when no common markers were available in thearea. Among the 34 regions, four key regions were identified whereputative QTLs for yield or yield components under stress anddrought-resistance component traits were identified acrosspopulations with their interval lengths of 35–64 cM (Figs. 3–6).We did not focus on common QTLs found in other populations ifthese were not also in CT9993/IR62266. There may be still otherQTLs that are equally important but are not highlighted in thisstudy, mainly because there is either limited or no information ontheir association with rice performance under drought conditionsin the field.
4.1.1. Co-location of QTLs for plant-type traits, and integrative,
primary, and secondary drought-resistance traits
The marker interval R2417–RZ909 (64 cM) on chromosome 1 inrice (with reference to the genetic map derived from CT9993/IR62266 DH lines; Zhang et al., 2001a) has frequently beenassociated with grain yield, various drought-resistance traits, andplant-type traits (Fig. 3, Table 3). Many QTLs for primary drought-resistance traits were identified under stress conditions in thisregion, including cell membrane stability (Tripathy et al., 2000),
rice chromosome 4 in different populations but using a common set of markers

Fig. 5. Consistent QTLs mapped for drought resistance and plant production traits in rice chromosome 8 in different populations but using a common set of markers
(underlined). For full legend see Fig. 3.
A. Kamoshita et al. / Field Crops Research 109 (2008) 1–23 13
osmotic adjustment (Lilley et al., 1996; Robin et al., 2003), andvarious root traits (e.g., maximum depth and penetrated rootthickness; Hemamalini et al., 2000; Price et al., 2002a; Nguyenet al., 2004; Zheng et al., 2003; Yue et al., 2006). Under well-watered control conditions, QTLs for various deep and thick roottraits were identified across different genetic backgrounds in thisregion (Ray et al., 1996; Yadav et al., 1997; Ali et al., 2000;Kamoshita et al., 2002b; Courtois et al., 2003; Li et al., 2005). Thisregion also contains QTLs for secondary traits such as plant waterstatus under stress (e.g., LWP and RWC; Courtois et al., 2000; Priceet al., 2002b; Babu et al., 2003; Srinivasan, 2005; Jearakongman,2005) (Table 3), as well as leaf rolling (Courtois et al., 2000; Priceet al., 2002b; Babu et al., 2003) and leaf drying (Courtois et al.,2000; Price et al., 2002b; Srinivasan, 2005; Boopathi et al., 2005;Yue et al., 2006). The positive effects of maintaining higher plantwater status under stress came from the deeper rooting parents(e.g., Azucena, CT9993) in some cases (Courtois et al., 2000; Babuet al., 2003; Srinivasan, 2005), but not in others (Jearakongman,2005). QTLs for integrated traits such as delay in flowering time(Yue et al., 2005), panicle exsertion rate, DRI (Jearakongman, 2005),and yield components (e.g., percentages of number of sterilepanicles, spikelet fertility, weight per grain) (Lafitte et al., 2004b;Jearakongman, 2005; Yue et al., 2006), as well as for grain yield(Kumar et al., 2007), all under drought stress in the field, have beenmapped in this region. This region is linked to plant-type QTLs with
maximum of 46% variability in the data set, such as plant height,panicle length, and tiller and panicle number, under both controland stress conditions across several mapping populations (Yu et al.,1995; Huang et al., 1996; Lanceras et al., 2004; Boopathi et al.,2005; Jearakongman, 2005). This segment contains the position ofsd-1, a major gene that controls semi-dwarfism that is widely usedby IRRI because of its strong association with harvest index andresponsiveness to fertilizer under well-watered conditions (Cour-tois et al., 1995). In populations in which a semi-dwarf cultivar(e.g., Bala or IR64) is used as one of the parents, alleles from thesemi-dwarf parents consistently reduce plant height (Hemamaliniet al., 2000; Lafitte et al., 2004b) and increase grain yield(Jearakongman, 2005) not only under control conditions but alsounder intermittent and terminal stress conditions. QTLs for grainyield under control or stress (Xu et al., 2005; Srinivasan, 2005), forrelative yield (Yue et al., 2005; Jearakongman, 2005), and fordifferences in yield between control and stress conditions (Xuet al., 2005) were also identified in this region. Thus, the sd-1 locusmay improve yield under both well-watered and droughtconditions, and it will be meaningful to estimate the effects ofthis QTL cluster, R2417-RZ909, after adjusting for the effects of sd-1
in future studies. In silico analyses of the QTL region, RM212–RM319 on chromosome 1 (Xu-Sheng et al., 2005; Zeng et al., 2006)have identified 175 annotated genes, 16 of which may be involvedin drought response (Xu-Sheng et al., 2005).

Fig. 6. Consistent QTLs mapped for drought resistance and plant production traits in rice chromosome 9 in different populations but using a common set of markers (bold and
underlined). For full legend see Fig. 3.
A. Kamoshita et al. / Field Crops Research 109 (2008) 1–2314
Another region consistently associated with drought tolerancein different populations is the interval RG939–RG620 (36 cM) onchromosome 4 (Fig. 4). A unique feature of this region is that bothQTL co-location and positive phenotypic correlation have beenobtained between root traits and yield under drought stress inCT9993/IR62266 (Babu et al., 2003), indicating the importance ofdrought avoidance through deep and thick roots. Many QTLs forroot traits have been identified in this region in populations grownunder well-watered control conditions (Champoux et al., 1995;Hemamalini et al., 2000; Zheng et al., 2000; Zhang et al., 2001b;Price et al., 2002a; Kamoshita et al., 2002b; Nguyen et al., 2004;Boopathi et al., 2005), as well as under simulated hardpanconditions (Ray et al., 1996; Zhang et al., 2001a; Nguyen et al.,2004) and drought (Yue et al., 2006) (Table 3). Secondary traitssuch as leaf rolling (Champoux et al., 1995; Courtois et al., 2000),leaf drying (Courtois et al., 2000; Boopathi et al., 2005), canopytemperature under stress, and stress recovery (Boopathi et al.,2005) have been mapped in this region, as well as QTLs forintegrative traits such as flowering delay (Lafitte et al., 2004b),yield components (Babu et al., 2003; Lanceras et al., 2004;Srinivasan, 2005; Lafitte et al., 2004b), and grain yield (Babu et al.,2003). QTLs for plant-type traits such as tiller number or plantheight are observed in some populations in this region (Ray et al.,1996; Babu et al., 2003; Lafitte et al., 2004b; Lanceras et al., 2004;
Srinivasan, 2005; Gomez et al., 2005; Xu et al., 2005; Boopathiet al., 2005), but their contributions (R2), unlike those in theinterval on chromosome 1, are not large compared with those ofother QTLs identified in the same region. Therefore, this region mayincrease yield under stress through drought avoidance byincreasing soil water uptake. Co-location of QTLs for root traitsand yield under stress was observed on this region only in theCT9993/IR62266 population (not in the IR64/Azucena or Bala/Azucena populations), in which both parent plants are of mediumto tall height and are well adapted to rainfed rice ecosystems.
Similarly, the marker interval RG978–RG598 (54 cM) onchromosome 8 has repeatedly shown associations with severalprimary drought-resistance traits in rice, as well as with plantwater status and grain yield, across a variety of genetic back-grounds and environments (Fig. 5). QTLs for primary traits such ascell membrane stability (Tripathy et al., 2000), osmotic adjustment(Lilley et al., 1996; Zhang et al., 2001a; Robin et al., 2003), leaf EW,rate of non-stomatal water loss (Srinivasan, 2005), and variousdeep and thick root traits (Price et al., 2002b; Yue et al., 2006;Zhang et al., 2001a; Nguyen et al., 2004) under stress were alsomapped in this region. A number of constitutive deep and thickroot traits under non-stressed conditions were also found indifferent populations (Champoux et al., 1995; Ray et al., 1996;Yadav et al., 1997; Hemamalini et al., 2000; Kamoshita et al.,

Table 3Four chromosome regions with clusters of QTLs detected for plant-type, phenology, primary, secondary, and integrated traits of drought resistance in CT9993/IR62266
population in previous studies across various environments
Trait Marker interval R2 Allele for the positive effect Reference
Chromosome 1
Plant-type traits
PH, plant height (S) RG109–ME10_14 27.8 CT9993 Babu et al. (2003)
PH, plant height (W) EM11_11–RG109 46.8, 46.5 CT9993 Babu et al. (2003)
PN-H, panicle number per hill (W) RG109–CDO345 21.7 IR62266 Lanceras et al. (2004)
PN-H, panicle number per hill (S) ME4_18–CDO345 20.7, 20.5, 20.8 IR62266 Lanceras et al. (2004)
PN-H, panicle number per hill (S) RG109–CDO345 18.6 IR62266 Lanceras et al. (2004)
PH, plant height (W) RM102–RG909 46.1 CT9993 Lanceras et al. (2004)
PH, plant height (S) RM102–RG909 41.1 CT9993 Lanceras et al. (2004)
PH, plant height (S) RM102–CDO345 36.2, 23.1 CT9993 Lanceras et al. (2004)
PH, plant height (S) C813–CDO345 32.6 CT9993 Lanceras et al. (2004)
PH, plant height (W) RG109–EM11_11 21.6 CT9993 Srinivasan (2005)
PH, plant height (S) RG109–EM11_11 22.7 CT9993 Srinivasan (2005)
PL, panicle length (S) RG109–EM11_11 13.5 CT9993 Srinivasan (2005)
PL, panicle length (W) RG109–EM11_11 11.7 CT9993 Srinivasan (2005)
Phenology traits
D50F, days to 50% flowering (S) CDO345–ME10_14 10.0 IR62266 Srinivasan (2005)
Primary traits
CMS, cell membrane stability (S) CDO345–ME10_14 20.1 CT9993 Tripathy et al. (2000)
PRT, penetrated root thickness (S) RG345–RG957 9.2 CT9993 Zhang et al. (2001a)
PRT, penetrated root thickness (S) RG345–RG957 12.0 CT9993 Nguyen et al. (2004)
TRDW, total root dry weight (S) RG109–EM11_11 9.3 CT9993 Zhang et al. (2001a)
DRW, deep root weight (W) R2417–RM212 9.4 CT9993 Kamoshita et al. (2002a)
DRR, deep root ratio (W) C813–RG957 7.8, 8.1 CT9993 Kamoshita et al. (2002a)
DR-T, deep root per tiller (W) R2417–RM212 9.8 CT9993 Kamoshita et al. (2002a)
RD, rooting depth (W) R2417–RM212 7.1 CT9993 Kamoshita et al. (2002a)
BRT, basal root thickness (W) CDO345–ME10_14 8.8 CT9993 Kamoshita et al. (2002a)
RSLR, root-shoot length ratio (W) CDO345–RZ909 7.8 CT9993 Kanbar et al. (2002)
TRDW, total root dry weight (S) RG727–RG109 9.3 CT9993 Nguyen et al. (2004)
PRT, penetrated root thickness (S) RG957–RZ19 12.0 CT9993 Nguyen et al. (2004)
Secondary traits
RWC, relative water content (S) RM212–C813 12.1 CT9993 Babu et al. (2003)
LR, leaf rolling (S) RG109–ME10_14 15.2 CT9993 Babu et al. (2003)
LD, leaf drying (S) RG109–ME10_14 20.8 CT9993 Babu et al. (2003)
LD, leaf drying (S) RZ909–CDO345 18.3 CT9993 Srinivasan (2005)
RWC, relative water content (S) RM212–RZ417 8.9 CT9993 Srinivasan (2005)
RWC50W+, time to reach 50% relative
water content (with wax)
RG109–EM11_11 8.7 IR62266 Srinivasan (2005)
Predawn LWP (W, Si, St) RM315 27.1, 47.6, – *IR64 Jearakongman (2005)
Midday LWP (W) RZ19 13.6 *Azucena Jearakongman (2005)
Midday LWP (Si) RM315 – *IR64 Jearakongman (2005)
Midday LWP (St) RG690 – *IR64 Jearakongman (2005)
RWC, relative water content (S) RZ730–RZ801 14.9, 12.0 *Azucena Courtois et al. (2000)
RWC, relative water content (S) RG810–RG331 6.1 *Azucena Courtois et al. (2000)
RWC, relative water content (S) C949–RZ14 13.9, 23.9 *Bala Price et al. (2002b)
Integrated traits
GP, grains per panicle (W) EM11_11–RG109 15.2 CT9993 Babu et al. (2003)
HI, harvest index (S) CDO345–RZ909 7.8 CT9993 Lanceras et al. (2004)
SDW, shoot dry weight at flowering (St) EM11_11–RG109 6.5 CT9993 Kumar et al. (2007)
GY, grain yield (St) EM11_11–RG109 14.5 CT9993 Kumar et al. (2007)
GY, grain yield (W) ME418–C813 17.0 CT9993 Srinivasan (2005)
HI, harvest index (W) ME418–C813 13.0 CT9993 Srinivasan (2005)
GY, grain yield (Si) RG690–RG730 – *IR64 Jearakongman (2005)
GY, grain yield (St) RZ810 12.6 *IR64 Jearakongman (2005)
GY, grain yield (W) OSR27–RM212 11.7, 5.0 *Teqing Xu et al. (2005)
GY, grain yield (d) OSR27–RM212 9.0 *Lemont Xu et al. (2005)
GY, grain yield (W) RM212 5.1 *– Beena (2005)
GY, grain yield (W) RM302 3.7 *– Beena (2005)
GY, grain yield (W) RM3825 6.7 *– Beena (2005)
Chromosome 4
Plant-type traits
PH, plant height (S) RG476–RG214 14.4 CT9993 Babu et al. (2003)
PH, plant height (W) RG476–RG214 16.0 CT9993 Babu et al. (2003)
PN, panicle number (W) ME4_9–RG214 21.3 IR62266 Lanceras et al. (2004)
PN, panicle number (S) ME4_9–RG214 16.3 IR62266 Lanceras et al. (2004)
PN, panicle number (S) RG939–RG214 18.9 IR62266 Lanceras et al. (2004)
PH, plant height (S) RG939–RG476 10.9 CT9993 Srinivasan (2005)
PTN, productive tiller number (S) RG214–RM280 16.5 CT9993 Srinivasan (2005)
PL, panicle length (S) C335–EM18_18 12.2 CT9993 Srinivasan (2005)
A. Kamoshita et al. / Field Crops Research 109 (2008) 1–23 15
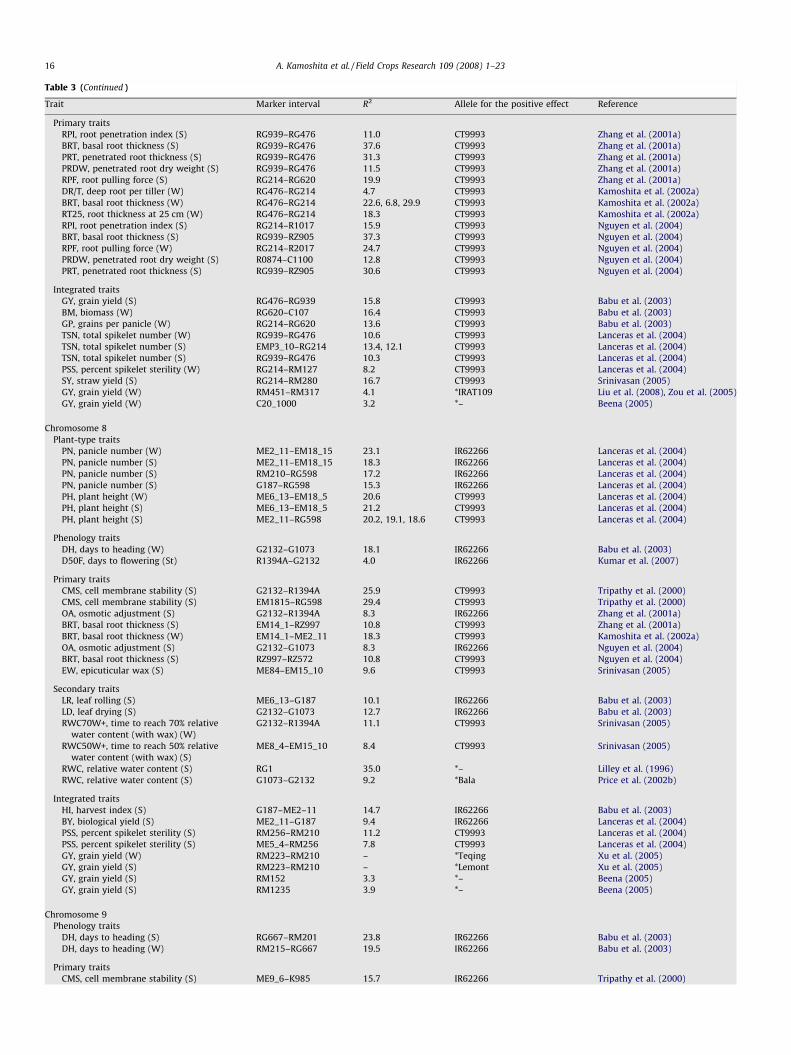
Table 3 (Continued )
Trait Marker interval R2 Allele for the positive effect Reference
Primary traits
RPI, root penetration index (S) RG939–RG476 11.0 CT9993 Zhang et al. (2001a)
BRT, basal root thickness (S) RG939–RG476 37.6 CT9993 Zhang et al. (2001a)
PRT, penetrated root thickness (S) RG939–RG476 31.3 CT9993 Zhang et al. (2001a)
PRDW, penetrated root dry weight (S) RG939–RG476 11.5 CT9993 Zhang et al. (2001a)
RPF, root pulling force (S) RG214–RG620 19.9 CT9993 Zhang et al. (2001a)
DR/T, deep root per tiller (W) RG476–RG214 4.7 CT9993 Kamoshita et al. (2002a)
BRT, basal root thickness (W) RG476–RG214 22.6, 6.8, 29.9 CT9993 Kamoshita et al. (2002a)
RT25, root thickness at 25 cm (W) RG476–RG214 18.3 CT9993 Kamoshita et al. (2002a)
RPI, root penetration index (S) RG214–R1017 15.9 CT9993 Nguyen et al. (2004)
BRT, basal root thickness (S) RG939–RZ905 37.3 CT9993 Nguyen et al. (2004)
RPF, root pulling force (W) RG214–R2017 24.7 CT9993 Nguyen et al. (2004)
PRDW, penetrated root dry weight (S) R0874–C1100 12.8 CT9993 Nguyen et al. (2004)
PRT, penetrated root thickness (S) RG939–RZ905 30.6 CT9993 Nguyen et al. (2004)
Integrated traits
GY, grain yield (S) RG476–RG939 15.8 CT9993 Babu et al. (2003)
BM, biomass (W) RG620–C107 16.4 CT9993 Babu et al. (2003)
GP, grains per panicle (W) RG214–RG620 13.6 CT9993 Babu et al. (2003)
TSN, total spikelet number (W) RG939–RG476 10.6 CT9993 Lanceras et al. (2004)
TSN, total spikelet number (S) EMP3_10–RG214 13.4, 12.1 CT9993 Lanceras et al. (2004)
TSN, total spikelet number (S) RG939–RG476 10.3 CT9993 Lanceras et al. (2004)
PSS, percent spikelet sterility (W) RG214–RM127 8.2 CT9993 Lanceras et al. (2004)
SY, straw yield (S) RG214–RM280 16.7 CT9993 Srinivasan (2005)
GY, grain yield (W) RM451–RM317 4.1 *IRAT109 Liu et al. (2008), Zou et al. (2005)
GY, grain yield (W) C20_1000 3.2 *– Beena (2005)
Chromosome 8
Plant-type traits
PN, panicle number (W) ME2_11–EM18_15 23.1 IR62266 Lanceras et al. (2004)
PN, panicle number (S) ME2_11–EM18_15 18.3 IR62266 Lanceras et al. (2004)
PN, panicle number (S) RM210–RG598 17.2 IR62266 Lanceras et al. (2004)
PN, panicle number (S) G187–RG598 15.3 IR62266 Lanceras et al. (2004)
PH, plant height (W) ME6_13–EM18_5 20.6 CT9993 Lanceras et al. (2004)
PH, plant height (S) ME6_13–EM18_5 21.2 CT9993 Lanceras et al. (2004)
PH, plant height (S) ME2_11–RG598 20.2, 19.1, 18.6 CT9993 Lanceras et al. (2004)
Phenology traits
DH, days to heading (W) G2132–G1073 18.1 IR62266 Babu et al. (2003)
D50F, days to flowering (St) R1394A–G2132 4.0 IR62266 Kumar et al. (2007)
Primary traits
CMS, cell membrane stability (S) G2132–R1394A 25.9 CT9993 Tripathy et al. (2000)
CMS, cell membrane stability (S) EM1815–RG598 29.4 CT9993 Tripathy et al. (2000)
OA, osmotic adjustment (S) G2132–R1394A 8.3 IR62266 Zhang et al. (2001a)
BRT, basal root thickness (S) EM14_1–RZ997 10.8 CT9993 Zhang et al. (2001a)
BRT, basal root thickness (W) EM14_1–ME2_11 18.3 CT9993 Kamoshita et al. (2002a)
OA, osmotic adjustment (S) G2132–G1073 8.3 IR62266 Nguyen et al. (2004)
BRT, basal root thickness (S) RZ997–RZ572 10.8 CT9993 Nguyen et al. (2004)
EW, epicuticular wax (S) ME84–EM15_10 9.6 CT9993 Srinivasan (2005)
Secondary traits
LR, leaf rolling (S) ME6_13–G187 10.1 IR62266 Babu et al. (2003)
LD, leaf drying (S) G2132–G1073 12.7 IR62266 Babu et al. (2003)
RWC70W+, time to reach 70% relative
water content (with wax) (W)
G2132–R1394A 11.1 CT9993 Srinivasan (2005)
RWC50W+, time to reach 50% relative
water content (with wax) (S)
ME8_4–EM15_10 8.4 CT9993 Srinivasan (2005)
RWC, relative water content (S) RG1 35.0 *– Lilley et al. (1996)
RWC, relative water content (S) G1073–G2132 9.2 *Bala Price et al. (2002b)
Integrated traits
HI, harvest index (S) G187–ME2–11 14.7 IR62266 Babu et al. (2003)
BY, biological yield (S) ME2_11–G187 9.4 IR62266 Lanceras et al. (2004)
PSS, percent spikelet sterility (S) RM256–RM210 11.2 CT9993 Lanceras et al. (2004)
PSS, percent spikelet sterility (S) ME5_4–RM256 7.8 CT9993 Lanceras et al. (2004)
GY, grain yield (W) RM223–RM210 – *Teqing Xu et al. (2005)
GY, grain yield (S) RM223–RM210 – *Lemont Xu et al. (2005)
GY, grain yield (S) RM152 3.3 *– Beena (2005)
GY, grain yield (S) RM1235 3.9 *– Beena (2005)
Chromosome 9
Phenology traits
DH, days to heading (S) RG667–RM201 23.8 IR62266 Babu et al. (2003)
DH, days to heading (W) RM215–RG667 19.5 IR62266 Babu et al. (2003)
Primary traits
CMS, cell membrane stability (S) ME9_6–K985 15.7 IR62266 Tripathy et al. (2000)
A. Kamoshita et al. / Field Crops Research 109 (2008) 1–2316

Table 3 (Continued )
Trait Marker interval R2 Allele for the positive effect Reference
PRT, penetrated root thickness (S) ME9_6–K985 18.5 CT9993 Zhang et al. (2001a)
PRDW, penetrated root dry weight (S) R41–ME2_10 16.8 CT9993 Zhang et al. (2001a)
PRDW, penetrated root dry weight (S) R41–ME2_10 16.8 CT9993 Nguyen et al. (2004)
Secondary traits
RWC, relative water content (S) RM215–RG667 58.8 CT9993 Babu et al. (2003)
Predawn LWP (St) RM107 – *Azucena Jearakongman (2005)
Midday LWP (St) RM107 – *Azucena Jearakongman (2005)
RWC, relative water content (S) RG451–RZ404 7.9 *IR64 Courtois et al. (2000)
Integrated traits
BM, biomass (W) ME9_3–ME9_6 14.3 CT9993 Babu et al. (2003)
SDW, shoot dry weight at
panicle initiation (W)
RM201–RG667 4.2 IR62266 Kumar et al. (2007)
BY, biological yield (S) R41–ME9_3 11.3 IR62266 Lanceras et al. (2004)
BY, biological yield (S) RM201–RM215 11.6 IR62266 Lanceras et al. (2004)
GY, grain yield (W, Si) OSR28 – *Azucena Jearakongman (2005)
GY, grain yield (Si) RM242 – *IR64 Jearakongman (2005)
GY, grain yield (Si) RM107 – *Azucena Jearakongman (2005)
GY, grain yield (S) PO463D04–RM242 8.7 *Azucena Gomez et al. (2005)
GY, grain yield (W) RM242–RM278 24.5 *Teqing Xu et al. (2005)
GY, grain yield (d) RM242–RM278 20.9 *Lemont Xu et al. (2005)
QTLs for plant water status and grain yield in the other populations are also added with asterisks (*). The letter in parenthesis after the trait name indicates the conditions
under which the trait value was measured. (W) Measured under well-watered conditions; (S) measured under water stress; (Si) and (St) intermittent and terminal types of
stress, respectively; (d) difference between stress and control. A dash (–) indicates that data are not available.
A. Kamoshita et al. / Field Crops Research 109 (2008) 1–23 17
2002b; Li et al., 2005). Interestingly, under irrigated conditions, aQTL for stable carbon isotope ratio (d13C) in grain, an indicator ofwater use efficiency, has been mapped in this region (Laza et al.,2006). QTLs for RWC under stress have been repeatedly foundacross different genetic backgrounds (Lilley et al., 1996; Price et al.,2002b; Srinivasan, 2005). Other secondary traits such as leafrolling (Champoux et al., 1995; Babu et al., 2003), leaf drying (Priceet al., 2002b; Babu et al., 2003), and canopy temperature (Yue et al.,2005) under drought stress have been mapped in this region(Table 3). QTLs for integrative traits such as harvest index (Babuet al., 2003), biomass, percent spikelet sterility, panicle number(Lanceras et al., 2004), delay in flowering time, relative yield,relative spikelet fertility, DRI (Yue et al., 2005), and grain yield (Xuet al., 2005) under stress have been mapped in this region. QTLs forplant-type traits have also been found in several populations (Rayet al., 1996; Hemamalini et al., 2000; Lafitte et al., 2004b; Xu et al.,2005; Boopathi et al., 2005).
The marker interval R41–RM215 (37 cM) on chromosome 9 islinked to drought resistance mostly through root traits, but alsoshows association with plant water status and grain yield (Fig. 6).QTLs for deep and thick root traits under control or stressconditions have been mapped in this region (Price et al., 2002b;Champoux et al., 1995; Yadav et al., 1997; Zhang et al., 2001b;Kamoshita et al., 2002b; Courtois et al., 2003; Zheng et al., 2003;Nguyen et al., 2004; Yue et al., 2006). A QTL for cell membranestability has also been identified (Tripathy et al., 2000). QTLs forplant water status – i.e., RWC (Babu et al., 2003; Courtois et al.,2000) and LWP (Jearakongman, 2005) – under stress have beenfound, with their positive effects coming from the deeper rootingparents (Babu et al., 2003; Jearakongman, 2005) and from theshort-statured parent (Courtois et al., 2000). QTLs for leaf rolling(Champoux et al., 1995; Courtois et al., 2000; Gomez et al., 2005)and leaf drying (Boopathi et al., 2005; Yue et al., 2006) under stresshave also been mapped in this segment. This region harbors QTLslinked to integrative traits such as biomass (Lanceras et al., 2004;Jearakongman, 2005), number of grains per panicle (Lafitte et al.,2004b), relative spikelet fertility, and delay in flowering time (Yueet al., 2005) under stress conditions. QTLs for grain yield have alsobeen identified in this region, under both control and stressconditions (Gomez et al., 2005; Xu et al., 2005; Jearakongman,2005).
These studies consistently identify regions on chromosomes 1,4, 8, and 9 that influence a range of drought-resistance traits, yield,and yield components under stress. Therefore, to increase droughttolerance, it may be more effective to use molecular methods toselect for specific regions rather than individual traits. Genomicregions mapped for several drought-resistance traits such as roottraits, phenology, yield, and yield components under water stressat the same locus are expected to be important, since such regionsnot only govern drought tolerance but are responsible for grainyield under stress (O’Toole, 2004). These alleles with effects acrossdifferent drought situations and genetic backgrounds are valuableand might be candidates for MAS. QTLs in these regions had R2
values ranging from 1.2% to 58.8% (Table 3), and although some areconsidered to be major (R2 value of greater than 10%; Yano andSasaki, 1997), many have only small effects on drought resistance.The contribution to genetic gain as a result of selecting these smalland large QTLs is likely to differ in each case, and quantitativeassessment of QTLs by simulation models is required (Cooper et al.,1999c; Yin et al., 2003), as well as verification of the agronomicvalues of the QTLs by the development of near-isogenic lines.
Interestingly, grain yield QTLs are contributed by both parents,as evidenced by transgressive segregation for grain yield underdrought stress (Babu et al., 2003; Lanceras et al., 2004; Xu et al.,2005). This indicates that neither parent carries all of the positiveor negative alleles for yield under stress.
4.1.2. Fine mapping of QTLs
A QTL may cover a region containing several hundred to severalthousand genes; this means that other factors are present in agiven region in addition to the loci of primary interest. Thus, theQTL regions on chromosomes 1, 4, 8, and 9 highlighted in thisreview (with their interval lengths of 35–64 cM) need to be finemapped to sequence candidate genes for drought-resistance. Insilico identification of candidate genes in the targeted QTL regionsassociated with drought tolerance traits has been describedrecently (Zeng et al., 2006; Fu et al., 2007). Fine mapping ofspecific QTL regions can also be achieved via strategies such asadding more markers in the regions (provided the population islarge enough to allow identification of recombinants around theQTLs), substitution mapping, and developing near-isogenic lines(Zhang et al., 1999). Nguyen et al. (2004) and Ganesh et al. (2003)

A. Kamoshita et al. / Field Crops Research 109 (2008) 1–2318
have added more markers to the linkage map of CT9993/IR62266DH lines and have fine-mapped certain already located QTLs to adistance of less than 1 cM.
4.2. Marker-assisted selection for drought resistance
Once stable QTLs for drought resistance are identified, theyshould be introduced into a near-isogenic background to assesstheir agronomic value and contributions to yield in targetenvironments (Price et al., 2002c). Shen et al. (2001) reportedimprovement of the rice root system by marker-assisted intro-gression of several root trait QTLs from Azucena into IR64. Theyalso studied effects of these introgressed segments on otheragronomic traits through pleiotropy or linkage drag. The potentialimpact of these QTLs on yield under drought was studied byevaluating the IR64 root introgression lines under rainfedconditions. The introgression lines had higher grain yield understress than IR64 (Babu et al., 2004a). Similarly, five QTLs associatedwith root morphological traits from Azucena were incorporatedinto a popular Indian cultivar, Kalinga III, by using marker-assistedbackcross breeding (Steel et al., 2006). The target segment onchromosome 9 (RM242–RM201) increased root length underirrigated and drought stress treatments, indicating that this QTLfunctions in a novel genetic background. Work is in progress inintrogressing the above ‘potential QTLs’ on chromosomes 4 and 9from CT9993 into IR20, IR64, and other locally adapted elitevarieties in southern India (Babu, personal communication).
Notwithstanding the potential suggested by QTL studies, MASfor drought resistance is not practiced widely. Reasons for thisinclude:
(1) R
esolution in mapping the target QTLs is low. QTL positions areimprecise, with the associated confidence interval coveringseveral megabases (Dupuis and Siegmund, 1999). This cancause crossovers and inefficient MAS (Zhang et al., 1999). Aresolution of 15–20 cM in the QTL location may be acceptableas a first step before dissection of the introgressed fragmentsinto smaller pieces to recover recombinants within thesesegments (Paterson, 1995). Ideally, the markers linked to theQTLs for a trait be within 1 cM of each other.(2) M
arkers flanking the QTLs often lack polymorphism in the elitelines into which the QTLs need to be introgressed, and large-scale genotyping is very costly (Morris et al., 2003; Tuberosaet al., 2003). Development of simple PCR-based markers such asSSRs (McCouch et al., 2002) may help in addressing theseproblems. The cost of MAS will further decrease as newgenotyping platforms, such as single nucleotide polymorphism(SNP) markers, become available (Collard et al., 2005).(3) A
ccurate phenotyping in the initial molecular mapping is themost important pre-requisite for success in selecting fordrought resistance. However, phenotyping is often inadequateowing to poor measurement techniques, irrelevant manipula-tion of experimental drought conditions, lack of reliable plantand soil data, and lack of field experts to interpret the results ofexperiments on plant function under drought stress (Blumet al., 2005).(4) M
ost putative drought-resistance traits considered for ricehave low heritability and are not consistently correlated withgrain yield under drought conditions in the target environ-ments because of the large G � E in rainfed rice as well as thelarge error variance under dry conditions (Atlin and Lafitte,2002).(5) T
he effects of individual QTLs on phenotypes are relativelysmall. O’Toole (2004) acknowledged the difficulty of develop-ing MAS when large numbers of small QTLs are often detectedfor putative drought-resistance traits. The task is to select threeto five QTLs in a breeding program that account for a highproportion of the variance.
4.3. Transgenics
Considerable progress has been made in developing transgenicrice lines tolerant to drought stress (Su et al., 1998; Datta, 2002;Toenniessen et al., 2003). Transgenic rice plants have beenproduced to over-express several candidate genes, such as
� a
dc, encoding arginine decarboxylase, which modulates the plantpolyamine content (Capell et al., 2004); � P 5CS, encoding pyrroline-5-carboxylate synthetase involved inproline biosynthesis (Zhu et al., 1998);
� H VA1, encoding late embryogenesis abundant (LEA) proteins (Xuet al., 1996; Cheng et al., 2002; Rohila et al., 2002);
� T PS and TP, encoding trehalose-6-phosphate synthase andtrehalose-6-phosphatase, involved in trehalose biosynthesis(Garg et al., 2002; Jang et al., 2003; Lee et al., 2003);
� R WC3, encoding the water channel protein aquaporin (Lian et al.,2004);
� O CPI1 (Oryza sativa chymotrypsin inhibitor-like 1), a stress-responsive proteinase inhibitor gene (Huang et al., 2007).
Transgenic rice plants have also been produced to over-expresstranscription factors involved in the regulation of stress-induciblegenes such as CBF/DREB1, DREB2, RD29B, RD22, and ICE1 (Shinozakiet al., 2003), CDPK encoding Ca2+-dependant protein kinase (Saijoet al., 2000), ABF3 and CBF3 (Oh et al., 2005), and SNAC1 encodingstress-responsive NAC 1 (Hu et al., 2006).
Although considerable progress has been made in developingtransgenic rice lines, their effectiveness in enhancing droughttolerance and yield under field conditions still needs to beassessed. The desiccation stress applied by most researchersevaluating transgenic plants for drought tolerance represents a‘shock’ treatment. The plants are grown in small containers thatprovide unrealistically small soil volumes, and they are subjectedto rapid stress cycles, mostly at the seedling stage, ranging froman hour to a few days (Rabbani et al., 2003; Babu et al., 2004b).When stress is imposed rapidly, there will be a greater number ofinjury-induced or survival-related responses than will be seenunder the slower, progressive water deficit found under fieldconditions. It is important that experiments be conducted underconditions that more closely approximate stress development inthe field in the target environments in terms of timing, duration,and intensity (e.g., Hu et al., 2006). In addition, because thecurrent transgenic approach can deal only with inducedresponses to water deficit, the importance of induced traitsrelative to that of constitutive traits needs to be quantified, asmentioned in Section 3.1.
Transgenic plants also can help us understand and manipulatethe responses of plants to stress. The use of transgenics, coupledwith recent developments in the areas of transcript mapping,functional genomics, and microarrays, may facilitate betterunderstanding of drought-resistance mechanisms in rice (Sale-kdeh et al., 2002a,b; Bennett et al., 2005; Gorantla et al., 2005;Kathiresan et al., 2006).
5. Improvement of general adaptability and drought resistance
When contrasting cultivars were compared over differentlocations and years under rainfed lowland conditions in southernIndia, Thailand, and Laos, genotypes with general adaptation andhigh yield potential performed better in most cases, particularly
Fig. 7. Grain yield of three groups of genotypes, high yielding (IR57514-PMI-5-B-1-
2, open circles), intermediate yielding (KDML105 and RD6, solid squares), and low
yielding (Chaing Seng, open triangles) plotted against environmental yield in
Thailand and Laos and analyzed by using Finlay–Wilkinson tests. Two linear
regression lines were drawn for the high and low yielding genotypes, respectively.
Thirty-five genotypes were tested in multi-location trials.
A. Kamoshita et al. / Field Crops Research 109 (2008) 1–23 19
when mean yields were high (Fukai and Babu, personal commu-nication, Fig. 7). Although we did not quantify the water deficits forall the data, this suggests the importance of improving thegermplasm base for general adaptability (i.e., mean yield overmulti-environment trails) and potential yield. Genotypic rankingof yield under mild drought conditions (i.e., with small yieldreduction) is in general similar to irrigated conditions (Pantuwanet al., 2002a). Traditional sources of parents for crossing in rainfedlowland rice breeding programs may not be the most appropriatefor improving potential yield level. The challenge is to combine thetraits of high potential yield and drought tolerance for genotypesthat flower at the most appropriate time for the target domain ofthe breeding program (Fukai et al., 1999). Recent IRRI genotypeevaluation trials in the Philippines under five widely differentenvironments, including irrigated lowlands and severely waterstressed uplands, showed low but positive correlations betweenyield under non-stress and yield under mild-stress conditions,confirming that some cultivars produced high yields under eithercondition, and supporting the potential for combining high yieldpotential and moderate water stress tolerance (Atlin et al., 2006).For improving rice yield in low-input upland environments, Lafitteet al. (2002b) listed the importance of potential yield together witha number of traits for specific adaptation (e.g., disease tolerance,tolerance to low pH, low-radiation tolerance before flowering, andweed competitiveness).
6. Conclusions
Managed drought environments in the field, such as dry seasontrials, delayed planting in the wet season, use of high toposequencelocations, drainage, raised beds, and large-scale rainout shelters,have been developed to simulate the target environments forbreeding. Selection for higher grain yield under managed stress,partly assisted by selection for secondary or integrative traits suchas low leaf rolling score, low spikelet sterility, and high DRI, withtheir moderate to high degrees of heritability, shows promise.
Understanding of genotypic responses to drought is increasing.Resistance traits differ under different types of drought (e.g.,terminal drought, vegetative stage drought, and intermittentdrought), but genotypic responses that contribute to droughtavoidance (e.g., deep and thick roots and conservative water use bymoderate plant size) and maintain higher plant water status areoften found to be more important for higher yield under stress than
are tolerance mechanisms. Transgenic rice, engineered forenhanced expression of primary induced traits for droughttolerance, has been studied under laboratory conditions, but theusefulness of these lines under field drought conditions remains tobe tested.
In our analysis of 15 mapping populations that have beenevaluated in drought-resistance studies, with emphasis on theCT9993/IR62266 population, four QTL clusters on chromosomes 1,4, 8, and 9 contained primary or secondary drought-resistancetraits or integrated traits. QTLs for constitutive primary traits suchas deep roots and plant-type traits such as plant height had higherR2 values than QTLs for induced traits, and were identified acrossdifferent populations under both well-watered and stress condi-tions. The QTLs for root traits and plant-type traits, together withQTLs for plant water status, were more often co-located withintegrated traits such as grain yield under stress. Although it isunlikely that a single primary or secondary trait will improve riceresistance to different types of drought, selection of some of theQTL clusters containing multiple drought-resistance traits ispromising.
In spite of the large amount of information on QTLs linked tovarious drought-resistance traits, routine use of these QTLs in MASis not widely practiced. The accuracy of phenotyping in these QTLmapping studies is one concern. Further, use of molecularapproaches may be limited because of the need to consider largenumber of QTLs with individually small effects. The effects thatMAS for such QTLs will have on improvement of plant breeding canbe estimated by the use of simulation models. Development ofnear-isogenic lines for these QTLs will allow testing of their trueagronomic value. Several labs are currently working on marker-assisted introgression of these QTLs into locally adapted elite ricelines.
Acknowledgments
We thank Dr Dave Mackill of IRRI and Professor Ken Fischer ofthe University of Queensland for their valuable comments on anearly draft. This review paper was written through our involve-ment in rice drought-resistance projects supported by the Rock-efeller Foundation and the Australian Centre for InternationalAgricultural Research. We acknowledge their generous support.We also acknowledge Grants-in-Aid for Scientific Research (No.20405019) from the Japan Society for the Promotion of Science,and Agreement on Academic Exchange Between University ofQueensland and University of Tokyo, which enabled involvementof the first author.
References
Ahmadi, N., Courtois, B., Khowaja, F., Price, A., Frouin, J., Hamelin, C., Ruiz, M., 2007.Meta-analysis of QTLs involved in rice root development using a QTL database.In: Proceedings of the International Symposium on Root Biology and MASStrategies for Drought Resistance Improvement in Rice, UAS-Bangalore, India,September 26–29, 2007. International Rice Research Institute, Los Banos,Philippines, pp. 29–30.
Ali, M., Jensen, C.R., Mogensen, V.O., Andersen, M.N., Henson, I.E., 1999. Rootsignaling and osmotic adjustment during intermittent soil drying sustain grainyield of field grown wheat. Field Crops Research 62, 35–52.
Ali, M.L., Pathan, M.S., Zhang, J., Bai, G., Sarkarung, S., Nguyen, H.T., 2000. MappingQTLs for root traits in a recombinant inbred population from two indicaecotypes in rice. Theoretical Applied Genetics 101, 756–766.
Atlin, G.N., 2004. Improving drought tolerance by selecting for yield. In: Fischer,K.S., Lafitte, R., Fukai, S., Atlin, G., Hardy, B. (Eds.), Breeding Rice forDrought-prone Environments. International Rice Research Institute, LosBanos, Philippines, pp. 14–22.
Atlin, G.N., Lafitte, H.R., 2002. Marker-assisted breeding versus direct selection fordrought tolerance in rice. In: Saxena, N.P., O’Toole, J.C. (Eds.), Field Screening forDrought Tolerance in Crop Plants with Special Emphasis on Rice: Proceedings ofthe International Workshop on Field Screening for Drought Tolerance in Rice,December 11–14, 2000, International Crop Research Institute for Semi-arid

A. Kamoshita et al. / Field Crops Research 109 (2008) 1–2320
Tropics, Patancheru, India. Patancheru (India); International Crop ResearchInstitute for Semi-arid Tropics, Rockefeller Foundation, New York, USA, pp.71–81.
Atlin, G.N., Lafitte, R., Venuprasad, R., Kumar, R., Jongdee, B., 2004. Heritability of riceyield under reproductive-stage drought stress, correlations across stress levelsand effects of selection: implications for drought tolerance breeding. In: Poland,D., Sawkins, J.M., Ribaut, M., Hoisington, D. (Eds.), Proceedings of a WorkshopHeld at Cuernavaca on Resilient Crops for Water Limited Environments, Mexico,May 24–28. International Maize and Wheat Improvement Center, Mexico, pp.85–87.
Atlin, G.N., Lafitte, H.R., Tao, D., Laza, M., Amante, M., Courtois, B., 2006. Developingrice cultivars for high-fertility upland systems in the Asian tropics. Field CropsResearch 97, 43–52.
Azhiri-Sigari, T., Yamauchi, A., Kamoshita, A., Wade, L.J., 2000. Genotypic variationin response of rainfed lowland rice to drought and rewatering. II. Root growth.Plant Production Science 3, 180–188.
Babu, R.C., Shashidhar, H.E., Lilley, J.M., Thanh, N.D., Ray, J.D., Sadasivam, S.,Sarkarung, S., O’Toole, J.C., Nguyen, H.T., 2001. Variation in root penetrationability, osmotic adjustment and dehydration tolerance among accessions of riceadapted to rainfed lowland and upland ecosystems. Plant Breeding 120, 233–238.
Babu, R.C., Nguyen, B.D., Chamarerk, V., Shanmugasundaram, P., Chezhian, P.,Jeyaprakash, P., Ganesh, S.K., Palchamy, A., Sadasivam, S., Sarkarung, S., Wade,L.J., Nguyen, H.T., 2003. Genetic analysis of drought resistance in rice bymolecular markers: association between secondary traits and field perfor-mance. Crop Science 43, 1457–1469.
Babu, R.C., Shanmugasundaram, P., Chezhian, P., Jeyaprakash, P., Balasubramanian, P.,Chararerk, V., Pathan, M.S., Babu, V., Nguyen, H.T., 2004a. Mapping quantitativetrait loci for drought tolerance in rice: comparison across environments, geneticbackgrounds and validation. In: Poland, D., Sawkins, J.M., Ribaut, M., Hoisington,D. (Eds.), Proceedings of a Workshop Held at Cuernavaca on Resilient Crops forWater Limited Environments, Mexico, May 24–28. International Maize andWheat Improvement Center, Mexico, pp. 204–205.
Babu, R.C., Zhang, J., Blum, A., Ho, T.H.D., Wu, R., Nguyen, H.T., 2004b. HVA1, a LEAgene from barley confers dehydration tolerance in transgenic rice (Oryza sativaL.) via cell membrane protection. Plant Science 166, 855–862.
Beavis, W.D., 1994. The power and deceit of QTL experiments: lessons fromcomparative QTL studies. In: Proceedings of the 49th Annual Corn & SorghumIndustry Research Conference. American Seed Trade Association, Washington,DC, pp. 250–266.
Beena, 2005. Studies on physio-morphological traits and genetic markers asso-ciated with drought response in rice (O. sativa). PhD thesis. Tamil NaduAgricultural University, Coimbatore, India.
Bennett, J., Liu, J.X., Ji, X.M., Raveendran, M., Oane, R., Liao, D.Q., Shobbar, Z., Chen,J.X., Tuberosa, R., Satoh, K.J., Kikuchi, S.S., Bruskiewich, R., Lafitte, R., Leung, H.,2005. Flowering stage drought stress in rice: proteome, metabolome andtranscriptome of anthers and peduncles. In: The 2nd International Conferenceon Integrated Approaches to Sustain and Improve Plant Production underDrought Stress. University of Rome, ‘‘La Sapienza’’, Rome, Italy, September24–28 L4.05.
Bernier, J., Kumar, A., Ramaiah, V., Spaner, D., Atlin, G., 2007. A large-effect QTL forgrain yield under reproductive-stage drought stress in upland rice. Crop Science47, 507–518.
Blum, A., 2002. Drought tolerance—is it a complex trait? In: Saxena, N.P., O’Toole,J.C. (Eds.), Field Screening for Drought Tolerance in Crop Plants with SpecialEmphasis on Rice: Proceedings of an International Workshop on Field Screeningfor Drought Tolerance in Rice, 11–14 December, 2000, International CropResearch Institute for Semi-arid Tropics, Patancheru, India. Patancheru, India;International Crop Research Institute for Semi-arid Tropics, Rockefeller Foun-dation, New York, USA, pp. 17–24.
Blum, A., Tuberosa, R., Nguyen, H.T., Araus-Ortega, J.L., 2005. Conference conclu-sions and recommendations. In: INTERDROUGHT-II. Proceedings of the 2ndInternational Conference on Integrated Approach to Sustain and Improve PlantProduction Under Drought Stress., Rome, Italy, September 24–28. www.plant-stress.com/id2/ID2-Report.pdf.
Boopathi, N.M., Chezhain, P., Jeyaprakash, P., Satheesh Kumar, S., Gomez, M., Suresh,R., Atlin, G., Subudhi, P.K., Shanmugasundaram, P., Babu, R.C., 2005. QTL map-ping of drought-resistance traits using indica rice (Oryza sativa L.) lines adaptedto target population of environment. In: Proceedings of the 2nd InternationalConference on Integrated Approaches to Sustain and Improve Plant Productionunder Drought Stress. University of Rome, ‘‘La Sapienza’’, Rome, Italy, Sep-tember 24–28 P6.05.
Capell, T., Bassie, L., Christou, P., 2004. Modulation of the polyamine biosyntheticpathway in transgenic rice confers tolerance to drought stress. PNAS 101, 9909–9914.
Causse, M.A., Fulton, T.M., Cho, Y.G., Ahn, S.N., Chunwongse, J., Wu, K., Xiao, J., Yu, Z.,Ronald, P.C., Harrington, S.E., Second, G., McCouch, S.R., Tanksley, S.D., 1994.Saturated molecular map of the rice genome based on an interspecific backcrosspopulation. Genetics 138, 1251–1274.
Champoux, M.C., Wang, G., Sarkarung, S., Mackill, D.J., O’Toole, J.C., Huang, N.,McCouch, S.R., 1995. Locating genes associated with root morphology anddrought avoidance in rice via linkage to molecular markers. Theoretical AppliedGenetics 90, 969–981.
Chang, T.T., Somrith, B., O’Toole, J.C., 1979. Potential of improving drought resis-tance in rainfed lowland rice. In: Rainfed Lowland Rice: Selected Papers from
the 1978 International Rice Research Conference. International Rice ResearchInstitute, Los Banos, Philippines, pp. 149–164.
Cheng, Z., Targoli, J., Huang, X., Wu, R., 2002. Wheat LEA genes, PMA80 andPMA1959, enhance dehydration tolerance of transgenic rice Oryza sativa L.Molecular Breeding 10, 71–82.
Collard, B.C.Y., Jahufer, M.Z.Z., Brouwer, J.B., Pang, E.C.K., 2005. An introduction tomarkers, quantitative trait loci (QTL) mapping and marker assisted selection forcrop improvement: the basic concepts. Euphytica 142, 169–196.
Comstock, J.P., 2002. Hydraulic and chemical signaling in the control of stomatalconductance and transpiration. Journal of Experimental Botany 53, 195–200.
Cooper, M., Rajatasereekul, S., Immark, S., Fukai, S., Basnayake, J., 1999a. Rainfedlowland rice breeding strategies for northeast Thailand. 1. Genotypic variationand genotype � environment interactions for grain yield. Field Crops Research64, 131–152.
Cooper, M., Rajatasereekul, S., Somrith, B., Sriwisut, S., Immark, S., Boonwite, C.,Suwanwongse, A., Ruangsook, S., Hanviriyapant, P., Romyen, P., Porn-uraisanit,P., Skulkhu, E., Fukai, S., Basnayake, L., Podlich, D.W., 1999b. Rainfed lowlandrice breeding strategies for northeast Thailand. 2. Comparison of intrastationand interstation selection. Field Crops Research 64, 153–176.
Cooper, M., Podlich, D.W., Fukai, S., 1999c. Combining information from multi-environment trials and molecular markers to select adaptive traits for yield ofrice in water-limited environments. In: Ito, O., O’Toole, J., Hartly, B. (Eds.), Ge-netic Improvement of Rice for Water-limited Environments. Proceedings of theWorkshop on Genetic Improvement of Rice for Water-limited Environments,IRRI, Los Banos, Philippines, December 1–3, 1998, pp. 13–34.
Courtois, B., Huang, N., Guiderdoni, E., 1995. RFLP mapping of genes controllingyield components and plant height in an indica � japonica DH population of rice.In: Fragile Lives in Fragile Ecosystems. Proceedings of the International RiceResearch Conference, International Rice Research Institute, Los Banos, Philip-pines, February 13–17, pp. 962–976.
Courtois, B., McLaren, G., Sinha, P.K., Prasad, K., Yadav, R., Shen, L., 2000. MappingQTLs associated with drought avoidance in upland rice. Molecular Breeding 6,55–66.
Courtois, B., Shen, L., Petalcorin, W., Carandang, S., Mauleon, R., Li, Z., 2003. LocatingQTLs controlling constitutive root traits in the rice population IAC165 � Co39.Euphytica 134, 335–345.
Datta, S.K., 2002. Recent developments in transgenics for abiotic stress tolerance inrice. JIRCAS Working Report 2002.
Dey, M.M., Upadhyaya, H.K., 1996. Yield loss due to drought, cold and submergencein Asia. In: Evenson, R.E., Herdt, R.W., Hossain, M. (Eds.), Rice Research in Asia:Progress and Priorities. Oxford University Press, Cary, NC, pp. 231–242.
Dingkuhn, M., Farquhar, G.D., De Datta, S.K., O’Toole, J.C., 1991. Discrimination of 13Camong upland rices having different water use efficiencies. Australian Journal ofAgricultural Research 42, 1123–1131.
Dupuis, J., Siegmund, D., 1999. Statistical methods for mapping quantitative traitloci from a dense set of markers. Genetics 151, 373–386.
Ekanayake, J., O’Toole, J.C., Garrity, D.P., Masajo, T.M., 1985. Inheritance of rootcharacters and their relations to drought resistance in rice. Crop Science 25,927–933.
Evenson, R.E., Gollin, D., 2003. Assessing the impact of the green revolution, 1960 to2000. Science 300, 758–762.
Evenson, R., Herdt, R.W., Hossain, M., 1996. Rice Research in Asia: Progress andPriorities. CAB International, Wallingford, UK.
Fischer, K.S., Fukai, S., Lafitte, R., McLaren, G., 2004a. Know your target environment.In: Fischer, K.S., Lafitte, R., Fukai, S., Atlin, G., Hardy, B. (Eds.), Breeding Rice forDrought-prone Environments. International Rice Research Institute, Los Banos,Philippines, pp. 5–11.
Fischer, K.S., Lafitte, R., Fukai, S., Atlin, G., Hardy, B., 2004b. Breeding Rice forDrought-prone Environments. International Rice Research Institute, Los Banos,Philippines, 98 pp.
Fu, B.Y., Xiong, J.H., Zhu, L.H., Zhao, X.Q., Xu, H.X., Gao, Y.M., Li, Y.S., Xu, J.L., Li, Z.K.,2007. Identification of functional candidate genes for drought tolerance in rice.Molecular Genetics and Genomics 278, 599–609.
Fukai, S., Cooper, M., 1995. Development of drought-resistant cultivars usingphysio-morphological traits in rice. Field Crops Research 40, 67–86.
Fukai, S., Pantuwan, G., Jongdee, B., Cooper, M., 1999. Screening for droughtresistance in rainfed lowland rice. Field Crops Research 64, 61–74.
Fukai, S., Basnayake, J., Cooper, M., 2001. Modelling water availability, crop growth,and yield of rainfed lowland rice genotypes in northeast Thailand. In: Tuong,T.P., Kam, S.P., Wade, L., Pandey, S., Bouman, B.A.M., Hardey, B. (Eds.), Pro-ceedings of the International Workshop on Characterizing and UnderstandingRainfed Environments, Bali, Indonesia, December 5–9, 1999. International RiceResearch Institute, Los Banos, Philippines, pp. 111–130.
Ganesh, S.K., Nadarajan, N., Yogameenakshi, P., Gomez, M., Manickavelu, A., 2003.Breeding for drought tolerance in rice. In: Proceedings of the 3rd Workshop onDrought Tolerance in Rice, National Network Program, University of Agricul-tural Sciences, Bangalore, India, July 22–25.
Garg, A.K., Kim, J.K., Owens, T.G., Ranwala, A.P., Choi, Y.D., Kochain, R.J., Wu, R.,2002. Trehalose accumulation in rice plants confers high tolerance to differ-ent abiotic stresses. Proceedings of the National Academy of Sciences 99,15898–15903.
Gomez, M., Babu, R.C., Shanmugasundaram, P., Satheesh Kumar, R., Suresh, R., Biji,K.R., Boopathi, N.M., Jeyaprakash, P., Gurumurthy, S., Price, A.H., 2005. QTLmapping and marker assisted selection for drought tolerance in rice (Oryzasativa L.). In: Proceedings of the 2nd International Conference on Integrated

A. Kamoshita et al. / Field Crops Research 109 (2008) 1–23 21
Approaches to Sustain and Improve Plant Production under Drought Stress.University of Rome, ‘‘La Sapienza’’, Rome, Italy, September 24–28 P6.19.
Gorantla, M., Babu, P.R., Reddy, V.B.L., Feltus, F.A., Paterson, A.H., Reddy, A.R., 2005.Functional genomics of drought stress response in rice: transcript mapping ofannotated unigenes of an indica rice (Oryza sativa L. cv. Nagina 22). CurrentScience 89, 496–514.
Hacke, U.G., Stiller, V., Sperry, J.S., Pittermann, J., McCulloh, K.A., 2001. Cavitationfatigue. Embolism and refilling cycles can weaken the cavitation resistance ofxylem. Plant Physiology 125, 779–786.
Hackett, C.A., 2002. Statistical methods for QTL mapping in cereals. Plant MolecularBiology 48, 585–599.
Hayashi, S., Kamoshita, A., Yamagishi, J., 2006. Effect of planting density on grainyield and water productivity of rice (Oryza sativa L.) grown in flooded and non-flooded fields in Japan. Plant Production Science 9, 298–311.
Hayashi, S., Kamoshita, A., Yamagishi, J., Kotchasatit, A., Jongdee, B., 2007. Genotypicdifference in grain yield of transplanted and direct seeded rainfed lowland rice(Oryza sativa L.) in northeast Thailand. Field Crops Research 102, 9–21.
Hemamalini, G.S., Sashidhar, H.E., Hittalmani, S., 2000. Molecular marker assistedtagging of morphological and physiological traits under two contrastingmoisture regimes at peak vegetative stage in rice (Oryza sativa L.). Euphytica112, 1–10.
Hirayama, M., Suga, R., 1996. Drought resistance. In: Yamamoto, R., Horisue, N., I-keda, R. (Eds.), Manual for Rice Breeding. Yokendo, Tokyo, (in Japanese; titletranslated by Dr A. Kamoshita), pp. 152–155.
Hirayama, M., Wada, Y., Nemoto, H., 2006. Estimation of drought tolerance based onleaf temperature in upland rice breeding. Breeding Science 56, 47–54.
Homma, K., Horie, T., Shiraiwa, T., Supapoj, N., Matsumoto, N., Kabaki, N., 2003.Toposequential variation in soil fertility and rice productivity of rainfed lowlandpaddy fields in mini-watershed (Nong) in northeast Thailand. Plant ProductionScience 6, 147–153.
Hoque, M.M., Kobata, T., 2000. Effect of soil compaction on the grain yield of rice(Oryza sativa L.) under water-deficit stress during the reproductive stage. PlantProduction Science 3, 316–322.
Horii, H., Nemoto, K., Miyamoto, N., Harada, J., 2006. Quantitative trait loci foradventitious and lateral roots in rice (Oryza sativa L.). Plant Breeding 125, 198–200.
Hsiao, T.C., 1973. Plant responses to water stress. Annual Review of Plant Physiology24, 519–570.
Hu, H., Dai, M., Yao, J., Xiao, B., Li, X., Zhang, Q., Xiong, L., 2006. Overexpressing aNAM, ATAF, and CUC (NAC) transcription factor enhances drought resistanceand salt tolerance in rice. Proceedings of the National Academy of Sciences 103,12987–12992.
Huang, N., Courtois, B., Khush, G.S., Lin, H., Wang, G., Wu, P., Zheng, K., 1996.Association of quantitative trait loci for plant height with major dwarfism genesin rice. Heredity 77, 130–137.
Huang, Y., Xiao, B., Xiong, L., 2007. Characterization of a stress responsive proteinaseinhibitor gene with positive effect in improving drought resistance in rice.Planta 226, 73–85.
Ikeda, H., Kamoshita, A., Manabe, T., 2007. Genetic analysis of rooting ability oftransplanted rice (Oryza sativa L.) under different water conditions. Journal ofExperimental Botany 58, 309–318.
Ikeda, H., Kamoshita, A., Yamagishi, J., Ouk, M., Bunna, L., 2008. Assessment ofmanagement of direct seeded rice production under different water conditionsin Cambodia. Paddy & Water Environment 6, 91–103.
Ingram, K., Bueno, T.F.D., Namuca, O.S., Yambao, E.B., Beyrouty, C.A., 1994. Rice roottraits for drought resistance and their genetic variation. In: Kirk, G.D. (Ed.), RiceRoots: Nutrient and Water Use. IRRI, Los Banos, Philippines, pp. 67–77.
International Rice Genome Sequencing Project, 2005. The map-based sequence ofthe rice genome. Nature 436, 793–800.
International Rice Research Institute, 2001. Rice ecosystem. Table 30. Distributionof rice crop area (000 ha), by environment, 2001. http://www.irri.org/science/ricestat/pdfs/Table%2030.pdf.
Jang, I.C., Oh, S.J., Seo, J.S., Choi, W.B., Song, S.I., Kim, C.H., Kim, Y.S., Seo, H.S., Choi,Y.D., Nahm, B.H., Kim, J.K., 2003. Expression of a bifunctional fusion of theEscherichia coli genes for trehalose-6-phosphate synthase and trehalose-6-phosphatase in transgenic rice plants increases trehalose accumulation andabiotic stress tolerance without stunting growth. Plant Physiology 131, 516–524.
Jearakongman, S., 2005. Validation and discovery of quantitative trait loci fordrought tolerance in backcross introgression lines in Rice (Oryza sativa L.)Cultivar IR64. PhD Thesis. Kasetsart University, p. 95.
Jongdee, B., 2004. Designing a national breeding program for developing drought-tolerant rainfed lowland varieties: the Thailand experiences. In: Fischer, K.S.,Lafitte, R., Fukai, S., Atlin, G., Hardy, B. (Eds.), Breeding Rice for Drought-proneEnvironments. International Rice Research Institute, Los Banos, Philippines, pp.64–69.
Jongdee, B., Fukai, S., Cooper, M., 1997a. Genotypic variation for water relationsand growth during the vegetative stage among six rice lines contrasting inmaintenance of high leaf water potential. In: Fukai, S., Cooper, M., Salisbury,J. (Eds.), Proceedings of the International Workshop on Breeding Strategiesfor Rainfed Lowland Rice in Drought-prone Environments, Ubon Ratchathani,Thailand, November 5–8, 1996. ACIAR, Canberra, pp. 180–191.
Jongdee, S., Mitchell, J.H., Fukai, S., 1997b. Modeling approach for estimation of riceyield reduction due to drought in Thailand. In: Fukai, S., Cooper, M., Salisbury, J.(Eds.), Proceedings of the International Workshop on Breeding Strategies for
Rainfed Lowland Rice in Drought-prone Environments, Ubon Ratchathani,Thailand, November 5–8, 1996. ACIAR, Canberra, pp. 65–73.
Jongdee, B., Fukai, S., Cooper, M., 2002. Leaf water potential and osmotic adjustmentas physiological traits to improve drought tolerance in rice. Field Crops Research76, 153–163.
Kamoshita, A., Wade, L.J., Yamauchi, A., 2000. Genotypic variation in response ofrainfed lowland rice to drought and rewatering. III. Water extraction duringdrought period. Plant Production Science 3, 189–196.
Kamoshita, A., Zhang, J., Siopongco, J., Sarkarung, S., Nguyen, H.T., Wade, L.J., 2002a.Effects of phenotyping environment on identification of QTL for rice rootmorphology under anaerobic conditions. Crop Science 42, 255–265.
Kamoshita, A., Wade, L.J., Ali, M.L., Pathan, M.S., Zhang, J., Sarkarung, S., Nguyen,H.T., 2002b. Mapping QTLs for root morphology of a rice populationadapted to rainfed lowland conditions. Theoretical Applied Genetics 104,880–893.
Kamoshita, A., Rodriguez, R., Yamauchi, A., Wade, L.J., 2004. Genotypic variation inresponse of rainfed lowland rice to prolonged drought and rewatering. PlantProduction Science 7, 406–420.
Kanbar, A., Shashidhar, H.E., Hittalmani, S., 2002. Mapping QTL associated withroot and related traits in DH population of rice. Indian Journal of Genetics 62,287–290.
Kathiresan, A., Lafitte, H.R., Chen, J., Manseuto, L., Bruskeiwich, R., Bennet, J., 2006.Gene expression microarrays and their application in drought research in rice.Field Crops Research 97, 101–110.
Kato, Y., Abe, J., Kamoshita, A., Yamagishi, J., 2006. Genotypic variation in rootgrowth angle in rice (Oryza sativa L.) and its association with deep rootdevelopment in upland fields with different water regimes. Plant and Soil287, 117–129.
Kato, Y., Kamoshita, A., Yamagishi, J., Imoto, H., Abe, J., 2007a. Growth of rice (Oryzasativa L.) cultivars under upland conditions with different levels of watersupply. 3. Root system development, water extraction and plant water status.Plant Production Science 10, 3–13.
Kato, Y., Kamoshita, A., Yamagishi, J., 2007b. Evaluating the resistance of six ricecultivars to drought: root restriction and the use of raised beds. Plant and Soil300, 149–161.
Kato, Y., Hirotsu, S., Nemoto, K., Yamagishi, J., 2008. Identification of QTLs control-ling rice drought tolerance at seedling stage in hydroponic culture. Euphytica160, 423–430.
Kobata, T., Tanaka, S., Utumi, M., Hara, S., Imaki, T., 1994. Sterility in rice (Oryzasativa L.) subject to drought during the booting stage occurs not because of lackof assimilate or of water deficit in the shoot but because of dehydration of theroot zone. Japanese Journal of Crop Science 63, 510–517.
Kobata, T., Hoque, M.M., Adachi, F., 2000. Effect of soil compaction on dry matterproduction and water use of rice (Oryza sativa L.) under water deficit stressduring reproductive stage. Plant Production Science 3, 306–315.
Kumar, R., Malaiya, S., Srivastava, M.N., 2004. Evaluation of morphophysiologicaltraits associated with drought tolerance in rice. Indian Journal of Plant Physiol-ogy 9, 305–307.
Kumar, R., Venuprasad, R., Atlin, G.N., 2007. Genetic analysis of rainfed lowland ricedrought tolerance under naturally-occurring stress in eastern India: heritabilityand QTL effects. Field Crops Research 103, 42–52.
Lafitte, R., 2004. Managing water for controlled drought in breeding plots. In:Fischer, K.S., Lafitte, R., Fukai, S., Atlin, G., Hardy, B. (Eds.), Breeding Rice forDrought-prone Environments. International Rice Research Institute, Los Banos,Philippines, pp. 23–26.
Lafitte, R., Courtois, B., 2002. Interpreting cultivar � environment interactions foryield in upland rice: assigning value to drought-adaptive traits. Crop Science 42,1409–1420.
Lafitte, H.R., Champoux, M.C., McLaren, G., O’Toole, J.C., 2001. Rice root morpho-logical traits are related to isozyme group and adaptation. Field Crops Research71, 57–70.
Lafitte, H.R., Courtois, B., Atlin, G., 2002a. The International Rice Research Institute’sexperience in field screening for drought tolerance and implications for breed-ing. In: Saxena, N.P., O’Toole, J.C. (Eds.), Field Screening for Drought Tolerance inCrop Plants with Special Emphasis on Rice: Proceedings of an InternationalWorkshop on Field Screening for Drought Tolerance in Rice, December 11–14,2000, International Crop Research Institute for Semi-arid Tropics, Patancheru,India; International Crop Research Institute for Semi-arid Tropics and Rock-efeller Foundation, New York, USA, pp. 25–40.
Lafitte, R., Courtois, B., Arraudeau, M., 2002b. Genetic improvement of rice in aerobicsystems: progress from yield to genes. Field Crops Research 75, 171–190.
Lafitte, R., Blum, A., Atlin, G., 2004a. Using secondary traits to help identify drought-tolerant genotypes. In: Fischer, K.S., Lafitte, R., Fukai, S., Atlin, G., Hardy, B.(Eds.), Breeding Rice for Drought-prone Environments. International RiceResearch Institute, Los Banos, Philippines, pp. 37–48.
Lafitte, H.R., Price, A.H., Courtois, B., 2004b. Yield response to water deficit in anupland rice mapping population: associations among traits and genetic mar-kers. Theoretical Applied Genetics 109, 1237–1246.
Lafitte, H.R., Li, Z.K., Vijayakumar, C.H.M., Gao, Y.M., Shi, Y., Xu, J.L., Fu, B.Y., Yu, S.B.,Ali, A.J., Domingo, J., Maghirang, R., Torres, R., Mackill, D., 2006. Improvement ofrice drought tolerance through backcross breeding: evaluation of donors andselection in drought nurseries. Field Crops Research 97, 77–86.
Lanceras, J.C., Pantuwan, G., Jongdee, B., Toojinda, T., 2004. Quantitative trait lociassociated with drought tolerance at reproductive stage in rice. Plant Physiol-ogy 135, 384–399.

A. Kamoshita et al. / Field Crops Research 109 (2008) 1–2322
Laza, Ma. R., Kondo, M., Ideta, O., Barlaan, E., Imbe, T., 2006. Identification ofquantitative trait loci for d13C and productivity in irrigated lowland rice. CropScience 46, 763–773.
Lebreton, C., Lazic-Jancic, V., Steel, A., Pekic, S., Quarrie, S.A., 1995. Identification ofQTL for drought responses in maize and their use in testing causal relationshipsbetween traits. Journal of Experimental Botany 46, 853–865.
Lee, S.B., Kwon, H.B., Kwon, S.J., Park, S.C., Jeong, M.J., Han, S.E., Byun, M.O., Daniell,H., 2003. Accumulation of trehalose within transgenic chloroplasts confersdrought tolerance. Molecular Breeding 11, 1–13.
Li, Z., Mu, P., Li, C., Zhang, H., Li, Z., Gao, Y., Wang, X., 2005. QTL mapping of roottraits in a doubled haploid population from a cross between upland andlowland japonica rice in three environments. Theoretical Applied Genetics110, 1244–1252.
Lian, H.L., Yu, X., Ye, Q., Ding, X.S., Kitagawa, Y., Kwak, S.S., Su, W.A., Tang, Z.C., 2004.The role of aquaporin Hong-Li Lian, Xin Yu, Qin Ye, Xiao-Song Ding, YoshichikaKitagawa, Sang-Soo Kwak, Wei-Ai Su, and Zhang-Cheng Tang RWC3 in droughtavoidance in rice. Plant Cell Physiology 45, 481–489.
Lilley, J.M., Fukai, S., 1994. Effect of timing and severity of water deficit on fourdiverse rice cultivars. I. Rooting pattern and soil water extraction. Field CropsResearch 37, 205–213.
Lilley, J., Ludlow, M., McCouch, S., O’Toole, J.C., 1996. Locating QTL for osmoticadjustment and dehydration tolerance in rice. Journal of Experimental Botany47, 1427–1436.
Liu, J.X., Liao, D.Q., Oane, R., Estenor, L., Yang, X.E., Li, Z.C., Bennett, J., 2006. Geneticvariation in the sensitivity of anther dehiscence to drought stress in rice. FieldCrops Research 97, 87–100.
Liu, G., Mei, H.W., Yu, X.Q., Zou, G.H., Liu, H.Y., Hu, S.P., Li, M.S., Wu, J.H., Chen, L., Luo,L.J., 2008. QTL analysis of panicle neck diameter, a trait highly correlated withpanicle size, under well-watered and drought conditions in rice (Oryza sativa L.).Plant Science 174, 71–77.
Maclean, J.L., Dawe, D.C., Hardy, B., Hettel, G.P., 2002. Rice Almanac. Los Banos,Philippines; International Rice Research Institute, Bouake, Cote d’Ivoire; WestAfrica Rice Department Association, Cali, Columbia; International Center forTropical Agriculture, Rome, Italy; Food and Agriculture Organization, 253 pp.
MacMillan, K., Emrich, K., Piepho, H.P., Mullins, C.E., Price, A.H., 2006. Assessing theimportance of genotype � environment interaction for root traits in rice using amapping population. II. Conventional QTL analysis. Theoretical Applied Genet-ics 113, 953–964.
McCouch, S.R., Kochert, G., Yu, H., Wang, Z.Y., Khush, G.S., 1988. Molecular mappingof rice chromosomes. Theoretical Applied Genetics 76, 815–829.
McCouch, S.R., Teytelman, L., Xu, Y., Lobos, K.B., Clare, K., Walton, M., Fu, B.,Maghirang, R., Li, Z., Zing, Y., Zhang, Q., Kono, I., Yano, M., Fjellstrom, R.,DeClerck, G., Chneider, D., Cartinhour, S., Ware, D., Stein, L., 2002. Developmentand mapping of 2240 new SSR markers for rice (Oryza sativa L.). DNA Research 9,199–207.
Mitchell, J.H., Siamhan, D., Wamala, M.H., Risimeri, J.B., Chinyamakobvu, E., Hen-derson, S.A., Fukai, S., 1998. The use of seedling leaf death score for evaluation ofdrought resistance of rice. Field Crops Research 55, 129–139.
Morris, M., Dreher, K., Ribaut, J.-M., Khairallah, 2003. Money matters (II): costs ofmaize inbred line conversion schemes at CIMMYT using conventional andmarker-assisted selection. Molecular Breeding 11, 235–247.
Nemoto, H., Suga, R., Ishihara, M., Okutsu, Y., 1998. Deep rooted rice varietiesdetected through the observation of root characteristics using the trenchmethod. Breeding Science 48, 321–324.
Nguyen, H.T., Babu, R.C., Blum, A., 1997. Breeding for drought tolerance in rice:physiology and molecular genetics considerations. Crop Science 37, 1426–1434.
Nguyen, T.T.T., Klueva, N., Chamareck, V., Aarti, A., Magpantay, G., Millena, A.C.M.,Pathan, M.S., Nguyen, H.T., 2004. Saturation mapping of QTL regions andidentification of putative candidate genes for drought tolerance in rice. Mole-cular Genetics Genomics 272, 35–46.
O’Toole, J.C., 1982. Adaptation of rice to drought prone environments. In: DroughtResistance in Crops with Emphasis on Rice. International Rice Research Insti-tute, Los Banos, Philippines, pp. 195–213.
O’Toole, J.C., 2000. Forward. In: Ribaut, J.M., Poland, D. (Eds.), Proceedings of aStrategic Workshop on Molecular Approaches for the Genetic Improvement ofCereals for Stable Production in Water-limited Environments. InternationalMaize and Wheat Improvement Center, El Batan, Mexico, June 21–25, 1999.International Maize and Wheat Improvement Center, Mexico, pp. iv.
O’Toole, J.C., 2004. Rice and water: the final frontier. In: Vanavichit A. (Ed.),Proceedings of the 1st International Conference on Rice for the Future,Kasetsert University, Bangkok. Bangkok, Thailand, 31 August to 2 September:Kasetsart University; National Center for Genetic Engineering and Biotech-nology, and Rice Research Institute, Ministry of Agriculture and Cooperatives,pp. 12–29.
Oh, S.L., Song, S.I., Kim, Y.S., Jang, H.J., Kim, S.Y., Kim, M., Kim, Y.K., Nahm, B.H.,Kim, J.K., 2005. Arabidopsis CBF3/DREB1A and ABF3 in transgenic riceincreased tolerance to abiotic stress without stunting growth. Plant Physiol-ogy 138, 341–351.
Ouk, M., Basnayake, J., Tsubo, M., Fukai, S., Fischer, K.S., Cooper, M., Nesbitt, H., 2006.Use of drought response index for identification of drought tolerant genotypesin rainfed lowland rice. Field Crops Research 99, 48–58.
Ouk, M., Basnayake, J., Tsubo, M., Fukai, S., Fischer, K.S., Kang, S., Men, S., Thun, V.,Cooper, M., 2007. Genotype-by-environment interactions for grain yield asso-ciated with water availability at flowering in rainfed lowland rice. Field CropsResearch 101, 145–154.
Pandey, S., Bhandari, H., Sharan, R., Ding, S., Prapertchob, P., Naik, D., Taunk, K.S.,2005. Coping with drought in agriculture of developing countries: insightsfrom rice farming in Asia. In: Proceedings of the 2nd International Conferenceon Integrated Approaches to Sustain and Improve Plant Production underDrought Stress. University of Rome, ‘‘La Sapienza’’, Rome, Italy, September24–28 L3.05.
Pantuwan, G., Fukai, S., Cooper, M., O’Toole, J.C., Sarkarung, S., 1997. Root traits toincrease drought resistance in rainfed lowland rice. In: Fukai, S., Cooper, M.Salisbury, J. (Eds.), Proceedings of an International Workshop, Ubon Ratch-athani, Thailand, 1996. Australian Centre for International AgriculturalResearch, Canberra, ACT, pp. 170–179.
Pantuwan, G., Fukai, S., Cooper, M., Rajatasereekul, S., O’Toole, J.C., 2002a. Yieldresponse of rice (Oryza sativa L.) genotypes to different types of drought underrainfed lowlands. Part 1. Grain yield and yield components. Field CropsResearch 73, 153–168.
Pantuwan, G., Fukai, S., Cooper, M., Rajatasereekul, S., O’Toole, J.C., 2002b. Yieldresponse of rice (Oryza sativa L.) genotypes to different types of drought underrainfed lowlands. 2. Selection of drought resistant genotypes. Field CropsResearch 73, 169–180.
Pantuwan, G., Fukai, S., Cooper, M., Rajatasereekul, S., O’Toole, J.C., 2002c. Yieldresponse of rice (Oryza sativa L.) genotypes to different types of drought underrainfed lowlands. 3. Plant factors contributing to drought resistance. Field CropsResearch 73, 181–200.
Pantuwan, G., Fukai, S., Cooper, M., Rajatasereekul, S., O’Toole, J.C., Basnayake, J.,2004. Drought resistance among diverse rainfed lowland rice (Oryza sativa L.)genotypes screened at the vegetative stage in dry season and its associationwith grain yield obtained under drought conditions in wet season. Field CropsResearch 89, 281–297.
Paterson, A.H., 1995. Molecular dissection of quantitative traits: progress andprospects. Genome Research 5, 321–333.
Pinheiro, B. da S., 2004. Integrating selection for drought tolerance into a breedingprogram: the Brazilian experience. In: Fischer, K.S., Lafitte, R., Fukai, S., A-tlin, G., Hardy, B. (Eds.), Breeding Rice for Drought-prone Environments.International Rice Research Institute, Los Banos, Philippines, pp. 75–83.
Pinheiro, B., da, S., de Castro, E., da, M., Guimaraes, C.M., 2006. Sustainability andprofitability of aerobic rice production in Brazil. Field Crops Research 97, 34–42.
Price, A.H., Tomos, A.D., 1997. Genetic dissection of root growth in rice (Oryza sativaL.). II. Mapping quantitative trait loci using molecular markers. TheoreticalApplied Genetics 95, 143–152.
Price, A.H., Courtois, B., 1999. Mapping QTLs associated with drought resistancein rice: progress, problems and prospects. Plant Growth Regulation 29, 123–133.
Price, A.H., Young, E.M., Tomos, A.D., 1997. Quantitative trait loci associated withstomatal conductance, leaf rolling and heading date mapped in upland rice(Oryza sativa). New Phytologist 137, 83–91.
Price, A.H., Steele, K.A., Moore, B.J., Barraclough, P.B., Clark, L.J., 2000. A combinedRFLP and AFLP linkage map of upland rice (Oryza sativa L.) used to identify QTLsfor root-penetration ability. Theoretical Applied Genetics 100, 49–56.
Price, A.H., Steele, K.A., Gorham, J., Bridges, J.M., Moore, B.J., Evans, J.L., Richardson,P., Jones, R.G.W., 2002a. Upland rice grown in soil-filled chambers and exposedto contrasting water-deficit regimes. I. Root distribution, water use and plantwater status. Field Crops Research 76, 11–24.
Price, A.H., Steele, K.A., Moore, B.J., Jones, R.G.W., 2002b. Upland rice grown in soil-filled chambers and exposed to contrasting water-deficit regimes. II. Mappingquantitative trait loci for root morphology and distribution. Field CropsResearch 76, 25–43.
Price, A.H., Cairns, J.E., Horton, P., Jones, H.G., Griffiths, H., 2002c. Linking drought-resistance mechanisms to drought avoidance in upland rice using a QTLapproach: progress and new opportunities to integrate stomatal and mesophyllresponses. Journal of Experimental Botany 53, 989–1004.
Price, A.H., Townend, J., Jones, M.P., Audebert, A., Courtois, B., 2002d. Mapping QTLsassociated with drought avoidance in upland rice grown in the Philippines andWest Africa. Plant Molecular Biology 48, 683–695.
Rabbani, M.A., Maruyama, K., Abe, H., Khan, A.A., Katsura, K., Ito, Y., Yoshiwara, K.,Seki, M., Shinozaki, K., Yamaguchi-Shinozaki, K., 2003. Monitoring expressionprofiles of rice genes under cold, drought, and high-salinity stresses and abscisicacid application using cDNA microarray and RNA gel-blot analyses. PlantPhysiology 133, 1755–1767.
Ray, J.D., Yu, L., McCouch, S.R., Champoux, M.C., Wang, G., Nguyen, H.T., 1996.Mapping quantitative trait loci associated with root penetration ability in rice.Theoretical Applied Genetics 92, 627–636.
Robin, S., Pathan, M.S., Courtois, B., Lafitte, R., Carandang, S., Lanceras, S., Amante, M.,Nguyen, H.T., Li, Z., 2003. Mapping osmotic adjustment in an advanced back-cross inbred population of rice. Theoretical Applied Genetics 107, 1288–1296.
Rohila, J., Jain, R.K., Wu, R., 2002. Genetic improvement of Basmati rice for salt anddrought tolerance by regulated expression of a barley Hva1 cDNA. Plant Science163, 525–532.
Saijo, Y., Hata, S., Kyozuka, J., Shimamot, K., Izhi, K., 2000. Overexpression of a singleCa2+ dependant protein kinase confers cold and salt/drought tolerance on riceplants. Plant Journal 23, 319–327.
Saini, H.S., 1997. Effects of water stress on male gametophyte development inplants. Sex Plant Reproduction 10, 67–73.
Saini, H.S., Lalonde, S., 1998. Injuries to reproductive development under waterstress, and their consequences for crop productivity. Journal of Crop Production1, 223–248.

A. Kamoshita et al. / Field Crops Research 109 (2008) 1–23 23
Salekdeh, Gh.H., Siopongoco, J., Wade, L.J., Ghareyazie, B., Bennet, J., 2002a. Pro-teomic analysis of rice leaves during drought stress and recovery. Proteomics 2,1131–1145.
Salekdeh, Gh.H., Siopongoco, J., Wade, L.J., Ghareyazie, B., Bennet, J., 2002b. Aproteomic approach to analyzing drought and salt responsiveness in rice. FieldCrops Research 76, 199–219.
Samson, B.K., Hasan, M., Wade, L.J., 2002. Penetration of hardpans by rice lines in therainfed lowlands. Field Crops Research 76, 175–188.
Selote, D.S., Khanna-Chopra, R., 2004. Drought-induced spikelet sterility is asso-ciated with an inefficient antioxidant defense in rice panicles. Plant Physiology121, 462–471.
Serraj, R., Sinclair, T.R., 2002. Osmolyte accumulation: can it really help increasecrop yield under drought conditions? Plant Cell Environment 25, 333–341.
Shen, L., Courtois, B., McNally, K.L., Robin, S., Li, Z., 2001. Evaluation of near-isogeniclines of rice introgressed with QTLs for root depth through marker-aidedselection. Theoretical Applied Genetics 103, 75–83.
Sheoran, I.S., Saini, H.S., 1996. Drought-induced male sterility in rice: changes incarbohydrate levels and enzyme activities associated with the inhibition ofstarch accumulation in pollen. Sex Plant Reproduction 9, 161–169.
Shinozaki, K., Shinozaki, K.Y., Seki, M., 2003. Regulatory network of gene expressionin the drought and cold stress responses. Current Opinion in Plant Biology 6,410–417.
Sibounheuang, V., Basnayake, J., Fukai, S., Cooper, M., 2001. Leaf water potential asdrought resistance character in rice. In: Fukai, S., Basnayake, J. (Eds.), ACIARProceedings 101; Increased Lowland Rice Production in the Mekong Region.Australian Centre for International Agricultural Research, Canberra, pp. 86–95.
Sibounheuang, V., Basnayake, J., Fukai, S., 2006. Genotypic consistency in theexpression of leaf water potential in rice (Oryza sativa L.). Field Crops Research97, 142–154.
Siopongco, J.D.L.C., Sekiya, K., Yamauchi, A., Egdane, J., Ismail, A.M., Wade, L.J., 2008.Stomatal responses in rainfed lowland rice to partial soil drying: evidence forroot signals. Plant Production Science 11, 28–41.
Srinivasan, S., 2005. Studies on mapping QTLs linked to leaf epicuticular wax,physio-morphological and plant production traits under water stress andnon-stress conditions in rice (Oryza sativa L.). PhD Thesis. Submitted to TamilNadu Agricultural University, Coimbatore, India.
Steel, K.A., Price, A.H., Shashidhar, H.E., Witcombe, J.R., 2006. Marker-assistedselection to introgress rice QTLs controlling root traits into an Indian uplandrice variety. Theoretical Applied Genetics 112, 208–221.
Stiller, V., Lafitte, H.R., Sperry, J.S., 2003. Hydraulic properties of rice (Oryza sativa L.)and the response of gas exchange to water stress. Plant Physiology 132, 1698–1706.
Su, J., Shen, Q., Ho, T.H.D., Wu, R., 1998. Dehydration stress regulated transgeneexpression in stably transformed rice plants. Plant Physiology 117, 913–922.
Tardieu, F., Davies, W.J., 1993. Integration of hydraulic and chemical signaling in thecontrol of stomatal conductance and water status of droughted plants. Plant,Cell & Environment 16, 341–349.
Toenniessen, G.H., O’Toole, J.C., DeVries, J., 2003. Advances in plant biotechnologyand its adoption in developing countries. Current Opinion in Plant Biology 6,191–198.
Tripathy, J.N., Zhang, J., Robin, S., Nguyen, T.T., Nguyen, H.T., 2000. QTLs for cell-membrane stability mapped in rice (Oryza sativa L.) under drought stress.Theoretical Applied Genetics 100, 1197–1202.
Tsubo, M., Basnayake, J., Fukai, S., Sihathep, V., Siyavong, P., Sipaseuth, Chan-phengsay, M., 2006. Toposequential effects on water balance and productivityin rainfed lowland rice ecosystem in Southern Laos. Field Crops Research 97,209–220.
Tsubo, M., Fukai, S., Tuong, T.P., Ouk, M., 2007. A water balance model for rainfedlowland rice fields emphasising lateral water movement within a topose-quence. Ecological Modelling 204, 503–515.
Tsuno, M., Takahashi, Y., Hirai, Y., 2004. Relationships between xylem hydraulicproperties and panicle characters in paddy and upland rice (Oryza sativa L.)genotypes. Japanese Journal of Tropical Agriculture 48, 131–136.
Tuberosa, R., Salvi, S., Sanguineti, M.C., Maccaferri, M., Giuliani, S., Landi, P., 2003.Searching for quantitative trait loci controlling root traits in maize: a criticalappraisal. Plant and Soil 255, 35–54.
Umayal, L., Babu, R.C., Sadasivam, S., 2001. Water stress induced histological andenzymatic changes in roots of rice cultivars. Plant Archives 1, 31–34.
Venuprasad, R., Shashidhar, H.E., Hittalmani, S., Hemamalini, G.S., 2002. Taggingquantitative trait loci associated with grain yield and root morphological traitsin rice (Oryza sativa L.) under contrasting moisture regimes. Euphytica 128,293–300.
Wade, L.J., Kamoshita, A., Yamauchi, A., Azhiri-Sigari, T., 2000. Genotypic variationin response of rainfed lowland rice to drought and rewatering. I. Growth andwater use. Plant Production Science 3, 173–179.
Wu, J., Maehara, T., Shimokawa, T., Yamamoto, S., Harada, C., Takazaki, Y., Ono, N.,Mukai, Y., Koike, K., Yazaki, J., Fujii, F., Shomura, A., Ando, T., Kono, I., Waki, K.,Yamamoto, K., Yano, M., Matsumoto, T., Sasaki, T., 2002. A comprehensive ricetranscript map containing 6591 expressed sequence tag sites. The Plant Cell 14,525–535.
Xiao, J., Li, J., Grandillo, S., Sang-Nag, A., Yuan, L., Tanksley, S.D., McCouch, S.R., 1998.Identification of trait-improving quantitative trait loci alleles from a wild ricerelative Oryza rufipogon. Genetics 150, 899–909.
Xu, D., Duan, B., Wang, B., Hong, B., Hong, Ho, T.H.D., Wu, R., 1996. Expression of lateembryogenesis abundant protein gene, HVA1, from barley confers tolerance towater deficit and salt stress in transgenic rice. Plant Physiology 110, 249–257.
Xu, S., 2003. Theoretical basis of the Beavis effect. Genetics 165, 2259–2268.Xu, J.L., Lafitte, H.R., Gao, Y.M., Fu, B.Y., Torres, R., Li, Z.K., 2005. QTLs for drought
escape and tolerance identified in a set of random introgression lines of rice.Theoretical Applied Genetics 111, 1642–1650.
Xu-Sheng, W., Jun, Z., Locedie, M., Bruskiewich, R., 2005. Identification of candidategenes for drought stress tolerance in rice by integration of a genetic (QTL) mapwith the rice genome physical map. Journal of Zhejiang University Science 6B(5), 382–388.
Yadav, R., Courtois, B., Huang, N., McLaren, G., 1997. Mapping genes controlling rootmorphology and root distribution in a doubled-haploid population of rice.Theoretical Applied Genetics 94, 619–632.
Yano, M., Sasaki, T., 1997. Genetic and molecular dissection of quantitative traits inrice. Plant Molecular Biology 35, 145–153.
Yin, X., Stam, P., Kropff, M.J., Schapendonk, A.H.C.M., 2003. Crop modeling, QTLmapping, and their complementary role in plant breeding. Agronomy Journal95, 90–98.
Yoshida, S., Hasegawa, S., 1982. The rice root system: its development and function.In: O’Toole, J.C. (Ed.), Drought Resistance in Crops with the Emphasis on Rice.International Rice Research Institute, Los Banos, Philippines, pp. 97–114.
Yu, Z.H., McCouch, S.R., Kinoshita, L., Tanksley, S.D., 1995. Association of morpho-logical and RFLP markers in rice. Genome 38, 566–574.
Yue, B., Xiong, L., Xue, W., Xing, Y., Luo, L., Xu, C., 2005. Genetic analysis for droughtresistance of rice at reproductive stage in field with different types of soil.Theoretical Applied Genetics 111, 1127–1136.
Yue, B., Xue, W., Xiong, L., Yu, Z., Luo, L., Cui, K., Jin, D., Xing, Y., Zhang, Q., 2006.Genetic basis of drought resistance at reproductive stage in rice: separation ofdrought resistance from drought avoidance. Genetics 172, 1213–1228.
Zeigler, R.S., Puckridge, D.W., 1995. Improving sustainable productivity in ricebased rainfed lowland systems of south and southeast Asia. Feeding fourbillion people. The challenge for rice research in the 21st century. Geo Journal35, 307–324.
Zeng, H., Zhong, Y., Luo, L., 2006. Drought tolerance genes in rice. Functional &Integrative Genomics 6, 338–341.
Zhang, J., Babu, R.C., Pantuwan, G., Kamoshita, A., Blum, A., Wade, L.J., Sarkarung, S.,O’Toole, J.C., Nguyen, H.T., 1999. Molecular dissection of drought tolerance inrice: from physio-morphological traits to field performance. In: Ito, O., O’Toole,J., Hardy, B. (Eds.), Genetic Improvement of Rice for Water-limited Environ-ments. Proceedings of the Workshop on Genetic Improvement of Rice forWater-limited Environments, International Rice Research Institute, Los Banos,Philippines, December 1–3, 1998, pp. 331–343.
Zhang, J., Zheng, H.G., Aarti, A., Pantuwan, G., Nguyen, T.T., Tripathy, J.N., Sarial, A.K.,Robin, S., Babu, R.C., Nguyen, B.D., Sarkarung, S., Blum, A., Nguyen, H.T., 2001a.Locating genomic regions associated with components of drought resistance inrice: comparative mapping within and across species. Theoretical AppliedGenetics 103, 19–29.
Zhang, W.P., Shen, X.Y., Wu, P., Hu, B., Liao, C.Y., 2001b. QTLs and epistasis forseminal root length under a different water supply in rice (Oryza sativa L.).Theoretical Applied Genetics 103, 118–123.
Zhang, J., Jia, W., Yang, J., Ismail, A.M., 2006. Role of ABA in integrating plantresponses to drought and salt stresses. Field Crops Research 97, 111–119.
Zheng, H.G., Babu, R.C., Pathan, Md. M.S, Ali, L., Huang, N., Courtois, B., Nguyen, H.T.,2000. Quantitative trait loci for root-penetration ability and root thickness inrice: comparison of genetic backgrounds. Genome 43, 53–61.
Zheng, B.S., Yang, L., Zhang, W.P., Mao, C.Z., Wu, Y.R., Yi, K.K., Liu, F.Y., Wu, P., 2003.Mapping QTLs and candidate genes for rice root traits under different water-supply conditions and comparative analysis across three populations. Theore-tical Applied Genetics 107, 1505–1515.
Zhu, B., Su, J., Chang, M.C., Verma, D.P.S., Fan, Y.L., Wu, R., 1998. Overexpression of aD1-pyrroline-5-carboxylate synthetase gene and analysis of tolerance to waterand salt stress in transgenic rice. Plant Science 139, 41–48.
Zou, G.H., Mei, H.W., Liu, H.Y., Liu, G.L., Hu, S.P., Yu, X.Q., Li, M.S., Wu, J.H., Luo, L.J.,2005. Grain yield responses to moisture regimes in a rice population: asso-ciation among traits and genetic markers. Theoretical Applied Genetics 112,106–113.




















