
High Strength Silicone-Urethane Copolymers: Synthesis and Properties

Available online at www.sciencedirect.com
Biomaterials 29 (2008) 1610e1619www.elsevier.com/locate/biomaterials
Response of mesenchymal stem cells to the biomechanical environmentof the endothelium on a flexible tubular silicone substrate
Eoin D. O’Cearbhaill a,b, Marie A. Punchard a, Mary Murphy c, Frank P. Barry c,Peter E. McHugh a,b, Valerie Barron a,*
a National Centre for Biomedical Engineering Science, National University of Ireland, Galway, Galway, Irelandb Department of Biomedical and Mechanical Engineering, National University of Ireland, Galway, Galway, Ireland
c Regenerative Medicine Institute, National Centre for Biomedical Engineering Science, National University of Ireland, Galway, Galway, Ireland
Received 30 July 2007; accepted 28 November 2007
Available online 14 January 2008
Abstract
Understanding the response of mesenchymal stem cells (MSCs) to forces in the vasculature is very important in the field of cardiovascularintervention for a number of reasons. These include the development of MSC seeded tissue engineered vascular grafts, targeted or systemicdelivery of MSCs in the dynamic environment of the coronary artery and understanding the potential pathological calcifying role of mechan-ically conditioned multipotent cells already present in the vessel wall. In vivo, cells present in the coronary artery are exposed to the primarybiomechanical forces of shear stress, radial stress and hoop stress. To date, many studies have examined the effect of these stresses in isolation,thereby not presenting the complete picture. Therefore, the main aim of this study is to examine the combined role of these stresses on MSCbehaviour. To this end, a bioreactor was configured to expose MSCs seeded on flexible silicone substrates to physiological forces e namely,a pulsatile pressure between 40 and 120 mmHg (5.33e1.6� 104 Pa), radial distention of 5% and a shear stress of 10 dyn/cm2 (1 Pa) at frequencyof 1 Hz for up to 24 h. Thereafter, the ‘pseudovessel’ was assessed for changes in morphology, orientation and expression of endothelial andsmooth muscle cell (SMC) specific markers. Hematoxylin and eosin (H&E) staining revealed that MSCs exhibit a similar mechanosensitiveresponse to that of endothelial cells (ECs); they reorientate parallel with direction of flow and have adapted their morphology to be similarto that of ECs. However, gene expression results show the cells exhibit greater levels of SMC-associated markers a-smooth muscle actinand calponin ( p< 0.05).� 2007 Elsevier Ltd. All rights reserved.
Keywords: Mesenchymal stem cell; Silicone; Bioreactor; Endothelial cell; Smooth muscle cell; Soft tissue biomechanics
1. Introduction
The studies of vascular disease, intervention and post-operative care are evolving as the role of undifferentiated cellsin the vasculature is being investigated more intensely. Themultipotentiality of these cells can be harnessed if properly di-rected and understood. However, their plasticity must be con-sistently controlled, while guarding against differentiation intoinappropriate cell types. The study of the effect of vascular
* Corresponding author. Tel.: þ353 91 492805; fax: þ353 91 494596.
E-mail address: [email protected] (V. Barron).
0142-9612/$ - see front matter � 2007 Elsevier Ltd. All rights reserved.
doi:10.1016/j.biomaterials.2007.11.042
physiological forces on mesenchymal stem cells (MSCs) isof particular interest. MSCs are well characterised and canbe successfully propagated in large numbers making themsuitable for clinical applications [1]. While the multipotential-ity of these cells has been widely explored under biochemicalstimulation [2], the role of biomechanical stimulation is lesswell understood.
Several aspects of cardiovascular research would benefitfrom a greater understanding of cell response to a vascularmechanical environment. The potential of being able to pro-vide autologous terminally differentiated vascular phenotypecell populations would be a major clinical breakthrough invascular tissue engineering [3,4].

1611E.D. O’Cearbhaill et al. / Biomaterials 29 (2008) 1610e1619
Similarly, many novel therapies currently being researchedto treat a broad range of diseases (including but not exclusiveto vascular diseases) involve the localised or systemic deliveryof MSCs as a direct therapeutic tool or with/on some form ofcarrier [5e8]. In many of these pilot studies, MSCs taggedwith a fluorescent protein have been seen in other non-targeted organs [9e11]. If these cells are circulating, thereis a chance they would adhere to vessel walls and differentiateunder biomechanical stimulation and lead to ectopic tissueformation.
Furthermore, subpopulations of cells found in the vesselmedial layer have been found to exhibit characteristics sim-ilar to those of MSCs isolated from the bone marrow. Thesecells have shown the capacity to undergo osteogenesis andchondrogenesis in vitro [12e14]. In addition to lipid accumu-lation at sites of vascular disease, more unusually, cartilageand bone-like tissues are also sometimes found at sites ofvascular disease [12e17]. This leads to the inference thatthese undifferentiated cells in the vessel wall under unfavour-able mechanical conditioning could lead to ectopic tissueformation. Therefore, a system which can apply a range ofloading conditions on similarly undifferentiated cells couldlead to a greater understanding of the influence of mechani-cal factors on pathology of certain types of arteriosclerosis[18,19].
In embryonic stem cells, FLK-1þ cells have shown thecapacity to differentiate into endothelial cells (ECs) throughbiomechanical [20] and biochemical signalling [21] in isola-tion and in combination [22]. In adult cells, attempts havebeen made to differentiate MSCs derived from bone marrow[23] and from the umbilical cord [24] into ECs, however,the effect of mechanical signalling along this differentiationpathway is largely unstudied. Cyclic tensile strain has beenshown to promote differentiation towards a smooth musclecell (SMC) phenotype [8,25]. Shear stress has been shownto promote EC differentiation from endothelial progenitorcells [26], while also playing a crucial role in vasculogenesisand re-endothelialisation [27].
Combinations of flow and pressure stimulation have beenshown to enhance expression of SMC markers in rat stromalcells [28]. The combined effects of laminar flow and cyclicstretching have recently been shown to enhance tissue growthin tissue engineered heart valves [29], however, the magnitudeof these forces is not as great as those found along the vesselsendothelium. It is clear from these studies that mechanical sig-nalling plays a crucial role in vascular cell development anddifferentiation.
It is the objective of this study to examine the combinato-rial effect of the three major forces of pulsatile flow, cyclicpressure and stretch experienced by cells lining the endothe-lium. The capacity of MSCs to differentiate into vascularphenotypes by these biomechanical stimuli is investigated.MSCs and human umbilical vein endothelial cells (HUVECs)are seeded onto silicone ‘pseudovessels’ and subjected tomechanical conditioning for 24 h. Cellular responses arecharacterised by changes in orientation, morphology andgene expression.
2. Materials and methods
2.1. Cell source
Bone marrow aspirates were obtained from the iliac crest of normal do-
nors. All procedures were performed with informed consent and approved
by the Clinical Research Ethical Committee at University College Hospital,
Galway. Donors are selected from what are generally considered ethically
approved guidelines: healthy males and non-pregnant females between the
ages of 18 and 45 years old, tested negative for HIV, hepatitis B and hepatitis
C. MSCs were isolated and expanded in culture as described previously by di-
rect plating [30]. Briefly, aspirates were washed with medium (DMEMelow
glucose containing penicillinestreptomycin solution (both SigmaeAldrich,
Ireland) at 100 U penicillin-G and 0.1 mg streptomycin/ml) and centrifuged;
the precipitated cells were suspended in medium with 10% selected foetal bo-
vine serum and plated at a final density of approximately 3.0� 105 cells/cm2.
Serum was selected based on maintenance of MSC proliferation and multipo-
tency in culture. Cells were seeded on T-175 flasks (Sarstedt, Ireland) and
maintained at 37 �C with 95% humidity and 5% CO2 in the same medium.
After 5 days, red blood cells were washed off with phosphate-buffered saline
and fresh medium added. Colonies of adherent cells formed within 9 days. At
the end of primary culture, adherent colonies were detached by treatment with
0.25% trypsin and 0.53 mM EDTA (SigmaeAldrich, Ireland). Cells were
plated in hMSC medium (DMEMeLG; 10% FBS; 1% antibiotic) at
5.7� 103 cells/cm2. All MSC preparations were characterised for surface ex-
pression of CD14, CD34, CD45, CD73 (BD Pharmingen, UK) and CD105
(Serotec, UK) using a FACs ARIA sorter (Becton Dickinson, UK). FACS anal-
ysis indicated the presence of a uniform population of cells negative for CD14,
CD34, CD45 (<2%) and positive for CD73 and CD105 (>95%) [31]. Cell
multipotency was confirmed through adipogenic, osteogenic and chondrogenic
assays. Cultures were passaged at 4e6 day intervals and expanded to passage
4 for experimentation.
2.2. Bioreactor design
This system is an enhanced version of that described in a previous study
[32] which now allows the incorporation of radial distention of tubular con-
structs under pressurisation. It is designed to fit inside a standard incubator
in an environment of 100% humidity, 5% CO2 at 37 �C. The bioreactor system
consists of a peristaltic pump (520U, Watson Marlow, UK) which operates at
a speed of 30 rpm. The pump generates a cyclic flow pattern of culture me-
dium through the system and the average flow rate is controlled by varying
the tubing diameter through which the medium is pulsed. The basal pressure
in the system is controlled by varying the height of the reservoir. Both the
mean and amplitude levels of the cyclic pressure waveform in the pseudovessel
are controlled by pinching the flow loop distal of the pseudovessel chamber.
Flow rate is monitored independently by a non-contact ultrasonic flowmeter
and recorded by a Labview� (National Instruments, TX, USA) program
[33]. Mean wall shear stress, tmean, is calculated by the HagenePoiseuille
equation [34]:
tmean ¼4mQ
pR3;
where m is the dynamic viscosity of the medium, Q is the total flow volume
and R is the vessel’s internal radius. Similarly, pressure is monitored by a pres-
sure probe and recorded in Labview�. Tube distention is examined by observ-
ing the expansion of the pseudovessel using a video-extensometer. Typical
waveforms for flow rate, pressure and radial distention are shown in Fig. 1.
Radial distention is quantified as the ratio of the instantaneous outer diameter
to the original unpressurised outer diameter of the pseudovessel.
2.3. Pseudovessel formation
Medical-grade silicone (4600, Elastosil, Wacker) was used to produce
pseudovessels. It exhibits similar levels of radial distention to those of a native
vessel when subjected to physiological levels of pressure [35]. When cured
this material has a tensile strength of 6 MPa and an elongation at break of

0
50
100
150
200
250
0 5 10 15 20 25 30Time (sec)
Flo
w (m
l/m
in
)
0
2
4
6
8
10
12
0 5 10 15 20 25 30Time (sec)
Strain
(%
)
0
20
40
60
80
100
120
140
0 5 10 15 20 25 30Time (sec)
Pressu
re (m
mH
g)
A
B
C
Fig. 1. Validation of vascular simulator outputs. Waveforms of typical (A)
pressure (40e120 mmHg), (B) radial distention (5% amplitude) and (C)
flow (mean w10 dyn/cm2) bioreactor output parameters when the peristaltic
pump is set to 30 rpm (corresponding to a pulse frequency of 1 Hz) and the
pinch is correctly positioned.
1612 E.D. O’Cearbhaill et al. / Biomaterials 29 (2008) 1610e1619
700%, compared to a tensile strength of 1.4e11.14 MPa and ultimate strain of
45e99% in the coronary artery [36]. For these experiments thin-walled tubes
were produced by dip coating a non-adherent mandrel (ID¼ 3.2� 0.01 mm).
Tube compliance was varied by either adding silicone oil to the liquid silicone
or by varying the number of dips. The dip coating was performed on ice to
delay the curing process at a speed of 100 mm/min. Curing was completed
by transferring the tubes to an oven at 100 �C for 3 h. This process generated
tubes with wall thicknesses in the range of 120e150 mm. Tubes were
individually inspected, measured and compliance tested in our system, to en-
sure all tubes selected for these experiments produced the same pressure, flow
and radial distention waveforms (as shown in Fig. 1) within a 5% tolerance.
2.4. Cell seeding
The silicone tubes were ultrasonically cleaned and ethanol sterilised prior
to cell seeding. Once seeded, these pseudovessels were attached to a specifi-
cally designed modular support structure which can be clipped into and out
of the bioreactor loop system. Pseudovessels were coated with human fibro-
nectin (8 mg/ml, SigmaeAldrich, Ireland) for 90 min on a bottle roller at
15 rev/h. They were then rinsed in Hank’s buffered saline solution (HBSS,
SigmaeAldrich, Ireland) and seeded with MSCs (P3eP4) at a concentration
of 1� 106 cells/cm2 or HUVECs (Clonetics, Cambrex Bio Science, UK)
(P4eP8) at a concentration of 1.75� 106 cells/cm2. The pseudovessels were
rotated for 24 h to allow cell adhesion. The presence of a confluent monolayer
is confirmed by H&E staining of a segment of the pseudovessel.
2.5. Cell conditioning
MSCs were subjected to 24 h of biomechanical conditioning. Pseudovessels
were transported into the bioreactor chamber and attached to the grips under
5% longitudinal tension. The chamber was then transferred to the incubator
and incorporated into the flow loop. The pump speed and pinch were gradually
increased to the desired levels (Fig. 1). Static controls were transferred into an
aliquot of the fresh hMSC medium with 1% fungizone used in the bioreactor
flow loop.
2.6. Cell visualisation and orientation analysis
In order to study gross cell morphology and orientation, pseudovessels
were stained with hematoxylin and eosin (SigmaeAldrich, Ireland) after being
fixed in methanol and mounted in a glycerol solution. Images were taken under
light microscopy (Fig. 2).
In order to quantify the orientation of cells, three random images were
taken from three biological replicate experiments (nine images in total, with
>1000 cells) and analysed using ImageJ� software (National Institutes of
Health, USA) [37]. Images were thresholded and in some cases manually
edited to highlight cell boundaries (Fig. 3A). The particle analysis tool was
used to give a best-fit ellipse for each cell. From this the cell orientation to
the pseudovessel’s longitudinal axis was determined (Fig. 3B for hMSCs
and Fig. 3C for HUVECs).
Cytoskeletal structure was analysis through F-actin staining (Fig. 4).
Cells were fixed in 4%-paraformaldehyde/2%-sucrose for 5 min prior to per-
meabilisation with 1% Triton X-100 (SigmaeAldrich, Ireland) for 5 min.
Following washes in a bovine serum albumin phosphate buffer saline solu-
tion (BSA/PBS), cells were placed in a 1 U/ml FITC-conjugated phalloidin
solution (Molecular Probes, UK) for 30 min at room temperature to visualise
actin fibres and then mounted with Vectashield� mounting medium with
DAPI nuclear stain (Vector Laboratories, UK). The samples were analysed
under a fluorescent microscope (Olympus 1�71, Olympus Biosystems
GMBH, Germany).
2.7. Immunostaining for vascular cell associated proteins
Sections of the pseudovessels were fixed as described previously. After
rinsing in PBS, samples were blocked for 1 h at room temperature with 1%
BSA/5% donkey serum. Samples were again washed in PBS and incubated
in 1:200 dilution of rabbit anti-human-von Willebrand factor (Dako) or
1:200 dilution of mouse anti-human-a-smooth muscle actin (Dako) for 1 h.
After another series of washes in PBS, cells were incubated for 1 h in 1:200
dilutions of FITC-conjugated anti-rabbit or TRITC-conjugated anti-mouse
IgG, respectively, in the dark (both Dako). Samples were then mounted with
Vectashield� mounting medium with DAPI nuclear stain and examined under
a fluorescent microscope.
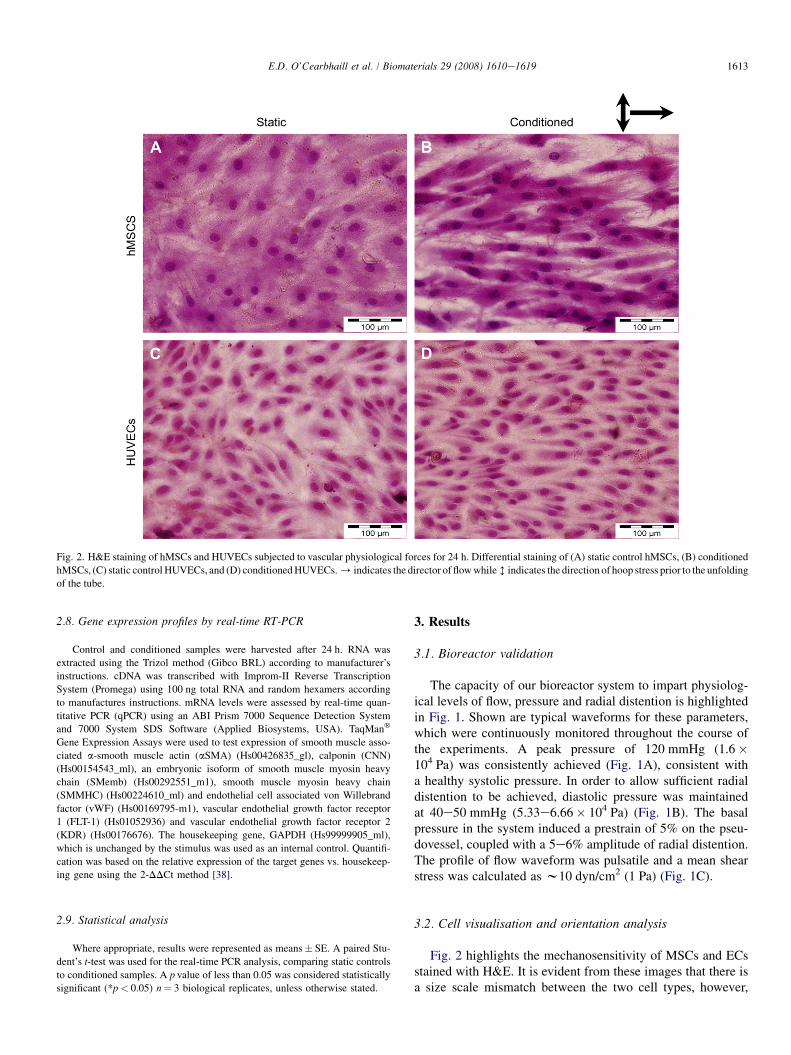
Fig. 2. H&E staining of hMSCs and HUVECs subjected to vascular physiological forces for 24 h. Differential staining of (A) static control hMSCs, (B) conditioned
hMSCs, (C) static control HUVECs, and (D) conditioned HUVECs. / indicates the director of flow while h indicates the direction of hoop stress prior to the unfolding
of the tube.
1613E.D. O’Cearbhaill et al. / Biomaterials 29 (2008) 1610e1619
2.8. Gene expression profiles by real-time RT-PCR
Control and conditioned samples were harvested after 24 h. RNA was
extracted using the Trizol method (Gibco BRL) according to manufacturer’s
instructions. cDNA was transcribed with Improm-II Reverse Transcription
System (Promega) using 100 ng total RNA and random hexamers according
to manufactures instructions. mRNA levels were assessed by real-time quan-
titative PCR (qPCR) using an ABI Prism 7000 Sequence Detection System
and 7000 System SDS Software (Applied Biosystems, USA). TaqMan�
Gene Expression Assays were used to test expression of smooth muscle asso-
ciated a-smooth muscle actin (aSMA) (Hs00426835_gl), calponin (CNN)
(Hs00154543_ml), an embryonic isoform of smooth muscle myosin heavy
chain (SMemb) (Hs00292551_m1), smooth muscle myosin heavy chain
(SMMHC) (Hs00224610_ml) and endothelial cell associated von Willebrand
factor (vWF) (Hs00169795-m1), vascular endothelial growth factor receptor
1 (FLT-1) (Hs01052936) and vascular endothelial growth factor receptor 2
(KDR) (Hs00176676). The housekeeping gene, GAPDH (Hs99999905_ml),
which is unchanged by the stimulus was used as an internal control. Quantifi-
cation was based on the relative expression of the target genes vs. housekeep-
ing gene using the 2-DDCt method [38].
2.9. Statistical analysis
Where appropriate, results were represented as means� SE. A paired Stu-
dent’s t-test was used for the real-time PCR analysis, comparing static controls
to conditioned samples. A p value of less than 0.05 was considered statistically
significant (*p< 0.05) n¼ 3 biological replicates, unless otherwise stated.
3. Results
3.1. Bioreactor validation
The capacity of our bioreactor system to impart physiolog-ical levels of flow, pressure and radial distention is highlightedin Fig. 1. Shown are typical waveforms for these parameters,which were continuously monitored throughout the course ofthe experiments. A peak pressure of 120 mmHg (1.6�104 Pa) was consistently achieved (Fig. 1A), consistent witha healthy systolic pressure. In order to allow sufficient radialdistention to be achieved, diastolic pressure was maintainedat 40e50 mmHg (5.33e6.66� 104 Pa) (Fig. 1B). The basalpressure in the system induced a prestrain of 5% on the pseu-dovessel, coupled with a 5e6% amplitude of radial distention.The profile of flow waveform was pulsatile and a mean shearstress was calculated as w10 dyn/cm2 (1 Pa) (Fig. 1C).
3.2. Cell visualisation and orientation analysis
Fig. 2 highlights the mechanosensitivity of MSCs and ECsstained with H&E. It is evident from these images that there isa size scale mismatch between the two cell types, however,
0
10
20
30
40
50
60
70
0-10 10-20 20-30 30-40 40-50 50-60 60-70 70-80 80-90Angle (°)
Freq
uen
cy (%
)
B
0
10
20
30
40
50
60
70
80
90
0-10 10-20 20-30 30-40 40-50 50-60 60-70 70-80 80-90Angle (°)
Freq
uen
cy (%
)
C
A
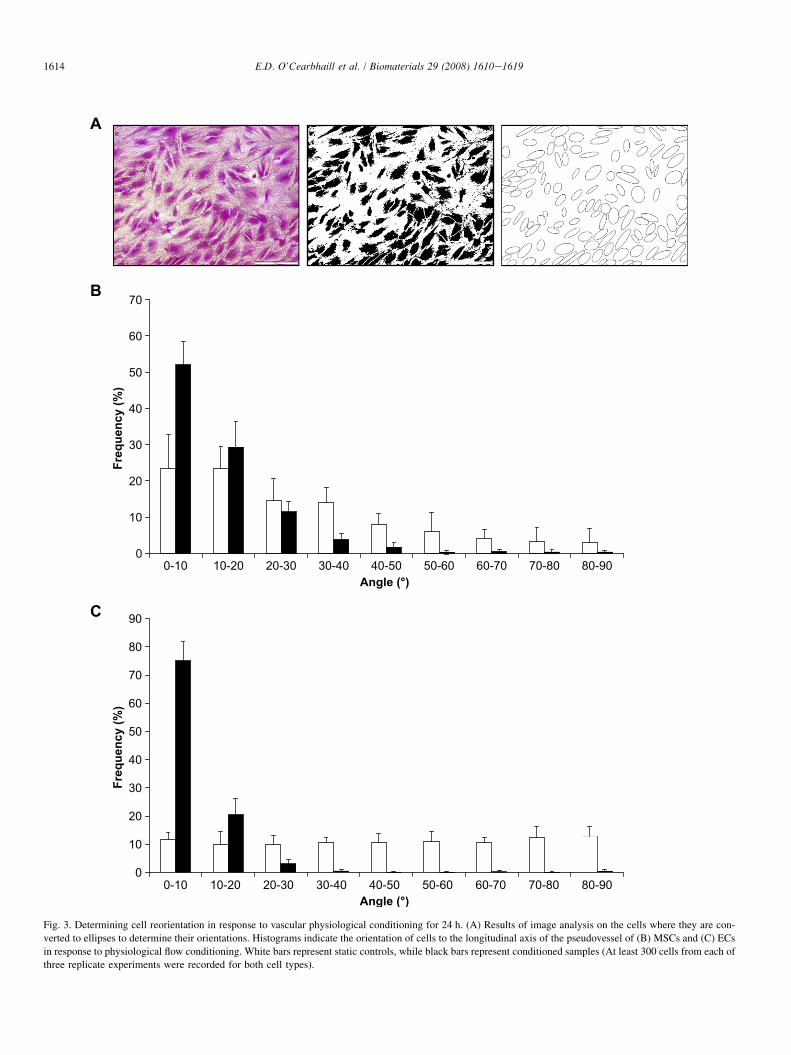
Fig. 3. Determining cell reorientation in response to vascular physiological conditioning for 24 h. (A) Results of image analysis on the cells where they are con-
verted to ellipses to determine their orientations. Histograms indicate the orientation of cells to the longitudinal axis of the pseudovessel of (B) MSCs and (C) ECs
in response to physiological flow conditioning. White bars represent static controls, while black bars represent conditioned samples (At least 300 cells from each of
three replicate experiments were recorded for both cell types).
1614 E.D. O’Cearbhaill et al. / Biomaterials 29 (2008) 1610e1619

Fig. 4. Cytoskeleton reorganisation in response to vascular physiological conditioning for 24 h. Comparison of cytoskeletal structure of static (A) hMSCs and (C)
HUVECs and mechanically conditioned (B) hMSCs and (D) HUVECs. Nuclei stained with DAPI in blue, F-actin in red shows orientation of cytoskeleton parallel
to the direction of flow (note that hMSCs and HUVECs are shown at different magnifications).
1615E.D. O’Cearbhaill et al. / Biomaterials 29 (2008) 1610e1619
MSCs reorientate perpendicular to the direction of hoop strainand parallel to the direction of flow along the longitudinal axisof the tube. MSCs in static culture tend to assemble in a spiralfashion, however, under physiological conditioning (Fig. 2B)their morphology more closely matches the length to widthratio of ECs, without specifically bearing a resemblance tothe ‘cobblestone’ morphology of ECs (Fig. 2D).
Fig. 3A shows a typical H&E stained section of the pseudo-vessel prior to ImageJ� image analysis. Shown also area thresholded image and a best-fit ellipses image from whichcell orientations are calculated.
Over 80% of MSCs (Fig. 3B) and over 90% of ECs(Fig. 3B) are seen to be aligned within 20� of the longitudinalaxis of the pseudovessel. It is noted that under static conditionsMSCs show a slight preference to align along the longitudinalaxis of the tube, compared to static ECs which show a uni-formly random distribution.
Fig. 4 illustrates that there is significant restructuring in thecytoskeleton of conditioned MSCs (Fig. 4B) compared tostatic controls (Fig. 4A). The mechanical stimulation de-creases cell length and width, resulting in a more compact cy-toskeleton, more closely resembling that of ECs (Fig. 4D).
3.3. Immunostaining for vascular cell associatedproteins
MSCs were found not to exhibit endothelial cell associatedvWF on a protein level under either static or mechanical stim-ulated conditions (Fig. 5A and B). MSCs are found to expressaSMA transiently with or without mechanical conditioningafter 24 h (Fig. 5C and D).
3.4. Gene expression profiles by real-time RT-PCR
qPCR results reveal statistically significant 6.5 and 33.8fold increases in aSMA and CNN expression, respectively,after 24 h. Neither SMemb nor SMMHC showed significantchanges. vWF did not show a significant change, while FLT-1 and KDR were not present at sufficient levels to be detected.
4. Discussion
MSCs clearly show the capacity to differentiate towardscells of mesodermal origin [2,6,39]. It is therefore not surpris-ing that they have a distinct potential to be developed into

Fig. 5. Presence of EC and SMC-associated proteins after 24 h of vascular physiological conditioning. (A) and (C) Static control hMSCs and (B) and (D) con-
ditioned hMSCs. In (A) and (B) nuclei are stained with DAPI in blue, while EC-associated vWF is shown in green. In (C) and (D) nuclei are stained with DAPI in
blue, while SMC-associated aSMA is shown in red.
1616 E.D. O’Cearbhaill et al. / Biomaterials 29 (2008) 1610e1619
SMCs phenotypes [8,25,40]. However, differentiation ofMSCs into functional ECs would appear to present a greaterchallenge. MSCs have been reported to differentiate into cellsexpressing endothelial cell markers in low serum conditions inthe presence of VEGF [23]. However, the percentage of cellswhich undergo this upregulation remains unclear and thismethod may be donor dependent (data not shown). Ganget al. [24] have performed similar studies showing more spe-cific differentiation towards ECs. However, the cells used inthese experiments were from the umbilical cord, which havebeen questioned as a suitable source for ‘stem’ cells [41].
It is clear that biochemical factors are a mainstay in promot-ing EC differentiation; however, the role of biomechanical con-ditioning in producing functional ECs cannot be discounted.There is a general lack of studies which have focussed onthe mechanical stimulation of MSCs, particularly with a syner-gistic approach [8,25,29].
The substrate onto which MSCs are seeded greatly influ-ences their differentiation pathway [42,43]. There are a limitednumber of biomaterials with properties suitable for mimickingthat of a native blood vessel. Polyurethane offers desirableelastic properties, but cell adhesion results can be variable[44]. Silicone has previously been shown to be an appropriate
substrate for studying the behaviour of cells under mechanicalstimulation [8,45e48]. Its suitability is governed by its com-pliance, biological inertness, manipulability, and translucency.When coated with fibronectin, silicone has been shown toallow good cell adhesion and ability to proliferate comparedto other biomaterials [49]. Furthermore, fibronectin has beenshown to promote endothelial cell differentiation in CD34þ
cells [50].This bioreactor system under a different configuration was
used to examine the effect of pathophysiological conditions onECs [32]. Indeed other groups have used similar bioreactors tomodel the effect of combined physiological forces on ECs inprevious studies [51,52]. However, to our knowledge, this isthe first study which has specifically examined the responseof MSCs to combined physiological levels of flow, pressureand radial distention.
Previous studies have shown that ECs align parallel to thedirection of flow in response to physiological levels of shearstress [53,54]. Similarly, ECs have been shown to reorientatethemselves perpendicular to the direction of stretch in systemswhich simulate hoop stretch [55,56]. While both forces playa role in determining cell orientation, it is there combined ef-fect which is of greatest physiological relevance. Here, MSCs

* p<0.05 Control vs Conditioned
*
*
0
10
20
30
40
50
60
aSMA Calponin SMemb SMMHC vWF
Re
la
tiv
e E
xp
re
ss
io
n
Static MSCs
Conditioned MSCs
Fig. 6. Changes in gene expression in response to vascular physiological con-
ditioning. Changes in gene expression of SMC-associated aSMA, calponin
and SMemb and SMMHC and endothelial cell associated vWF in response
to 24 h of mechanical stimulation.
1617E.D. O’Cearbhaill et al. / Biomaterials 29 (2008) 1610e1619
are identified as having similar mechanosensitivity to that ofECs. They adopt an analogous angle of orientation distributionprofile (Fig. 3) and a morphology similar to that of ECs undervascular physiological conditioning (Figs. 2e4). The conflu-ency, morphology and orientation of cells are consistent withthat of the in vivo endothelium [57]. Interestingly, it wasalso observed that MSCs show a tendency to orientate them-selves towards the longitudinal axis of the tube in static culture(Fig. 3B). Previous studies have shown the preferential orien-tation of cells towards the longitudinal axis of convex cylindri-cal substrates, where a high degree of surface curvature exists[58,59]. However, this also appears to be the case for concavesurfaces [60]. Here, it is thought that the degree of surfacecurvature is only sufficient to influence the morphology ofthe larger MSCs.
It is apparent that without biochemical stimulation, MSCsshow a greater capacity to differentiate towards a SMC pheno-type rather than an EC phenotype after a 24 h time period underthis biomechanical stimulation. It is acknowledged that vascu-lar cell differentiation may require longer time periods in whichto occur; nevertheless, interesting data can be gathered in termsof initial cellular response. Within this time point, MSCs do notappear to show signs of increases in endothelial cell markers ona protein (Fig. 5A and B) or mRNA level (Fig. 6). VEGF recep-tors, FLT-1 and KDR, were not present in sufficiently highlevels to be recorded. Conversely, SMC-associated gene ex-pression is upregulated, while aSMA expression is still tran-sient on a protein level (Fig. 5C and D). 6.5 and 33.8 foldincreases in contractile phenotype associated aSMA andCNN, respectively, were seen after 24 h of conditioning. Thesewere approximately one third of the levels expressed in SMCs(data not shown). This upregulation is larger than that reportedby Park et al. [8], where the effect of uniaxial stretch was ex-amined in isolation. This suggests a synergistic effect of themechanical forces may promote a more significant response.No upregulation was measured in synthetic SMC-associated
SMemb or the mature marker SMMHC. It is possible that thelatter would be observed over longer time points. Future studieswill seek to analysis MSC response over longer time periods ata protein synthesis level, as well as observing the combinedeffects of biochemical and biomechanical signalling on celldifferentiation.
The upregulation of SMC-associated genes in response tocyclic stretch has been widely reported in SMCs [61e65],but also in ECs [66]. Cevallos et al. [66] suggest that whereflow is distorted, ECs have the ability to transdifferentiateinto SMCs. This further highlights the synergistic effects ofphysiological levels of flow and stretch on cell behaviour,established by Zhao et al. [47]. Interestingly, when ECs aresubjected to the same physiological conditioning for 24 h,no significant upregulation in aSMA is observed (data notshown). While levels of CNN could not amplified sufficientlyin ECs to be successfully analysed, this highlights the key roleof shear stress in regulating cell phenotype.
5. Conclusions
It is clear that the biomechanical forces of the endotheliumplay a crucial role in cell differentiation at this milieu. Thepossible role of stem cells in restenosis has been identifiedelsewhere [67]. The tendency observed here for MSCs todifferentiate towards a SMC phenotype in a vascular physio-logical environment could have implications in their potentialas pathophysiological agents. MSCs do appear to offer an ap-propriate cell source for SMCs for vascular tissue engineeringapplications, through biochemical and biomechanical cellsignalling. MSCs, however, do not show an immediate poten-tial to differentiate into functional ECs under biomechanicalstimulation alone. While their morphology, orientation andcytoskeletal structure can be manipulated to resemble thatof ECs, they do not exhibit their functional markers, at leastwithin the time period observed here.
Acknowledgements
This work is supported by research funding from theProgramme for Research in Third Level Institutions (PRTLI)administered by the Higher Education Authority (HEA). Theauthors would like to thank Georgina Shaw (REMEDI, Na-tional University of Ireland, Galway) for her assistance andprovision of mesenchymal stem cells.
References
[1] Deans RJ, Moseley AB. Mesenchymal stem cells: biology and potential
clinical uses. Exp Hematol 2000;28(8):875e84.
[2] Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R, Mosca JD,
et al. Multilineage potential of adult human mesenchymal stem cells.
Science 1999;284(5411):143e7.
[3] Sales KM, Salacinski HJ, Alobaid N, Mikhail M, Balakrishnan V,
Seifalian AM. Advancing vascular tissue engineering: the role of stem
cell technology. Trends Biotechnol 2005;23(9):461e7.

1618 E.D. O’Cearbhaill et al. / Biomaterials 29 (2008) 1610e1619
[4] Riha GM, Lin PH, Lumsden AB, Yao Q, Chen C. Review: application of
stem cells for vascular tissue engineering. Tissue Eng 2005;11(9e10):
1535e52.
[5] Koc ON, Gerson SL, Cooper BW, Dyhouse SM, Haynesworth SE,
Caplan AI, et al. Rapid hematopoietic recovery after coinfusion of autol-
ogous-blood stem cells and culture-expanded marrow mesenchymal stem
cells in advanced breast cancer patients receiving high-dose chemother-
apy. J Clin Oncol 2000;18(2):307e16.
[6] Bruder SP, Fink DJ, Caplan AI. Mesenchymal stem cells in bone devel-
opment, bone repair, and skeletal regeneration therapy. J Cell Biochem
1994;56(3):283e94.
[7] Barbash IM, Chouraqui P, Baron J, Feinberg MS, Etzion S, Tessone A,
et al. Systemic delivery of bone marrow-derived mesenchymal stem cells
to the infarcted myocardium: feasibility, cell migration, and body distri-
bution. Circulation 2003;108(7):863e8.
[8] Park JS, Chu JS, Cheng C, Chen F, Chen D, Li S. Differential effects of
equiaxial and uniaxial strain on mesenchymal stem cells. Biotechnol
Bioeng 2004;88(3):359e68.
[9] Kraitchman DL, Tatsumi M, Gilson WD, Ishimori T, Kedziorek D,
Walczak P, et al. Dynamic imaging of allogeneic mesenchymal stem
cells trafficking to myocardial infarction. Circulation 2005;112(10):
1451e61.
[10] Kraitchman DL, Heldman AW, Atalar E, Amado LC, Martin BJ,
Pittenger MF, et al. In vivo magnetic resonance imaging of mesenchymal
stem cells in myocardial infarction. Circulation 2003;107(18):2290e3.
[11] Gao J, Dennis JE, Muzic RF, Lundberg M, Caplan AI. The dynamic in
vivo distribution of bone marrow-derived mesenchymal stem cells after
infusion. Cells Tissues Organs 2001;169(1):12e20.
[12] Tintut Y, Alfonso Z, Saini T, Radcliff K, Watson K, Bostrom K, et al.
Multilineage potential of cells from the artery wall. Circulation 2003;
108(20):2505e10.
[13] Watson KE, Bostrom K, Ravindranath R, Lam T, Norton B, Demer LL.
TGF-beta 1 and 25-hydroxycholesterol stimulate osteoblast-like vascular
cells to calcify. J Clin Invest 1994;93(5):2106e13.
[14] Tintut Y, Parhami F, Bostrom K, Jackson SM, Demer LL. cAMP stimu-
lates osteoblast-like differentiation of calcifying vascular cells. Potential
signaling pathway for vascular calcification. J Biol Chem
1998;273(13):7547e53.
[15] Demer LL, Tintut Y. Return to ectopia: stem cells in the artery wall.
Arterioscler Thromb Vasc Biol 2005;25(7):1307e8.
[16] Giachelli CM, Speer MY, Li X, Rajachar RM, Yang H. Regulation of
vascular calcification: roles of phosphate and osteopontin. Circ Res
2005;96(7):717e22.
[17] Rattazzi M, Bennett BJ, Bea F, Kirk EA, Ricks JL, Speer M, et al. Cal-
cification of advanced atherosclerotic lesions in the innominate arteries
of ApoE-deficient mice: potential role of chondrocyte-like cells. Arte-
rioscler Thromb Vasc Biol 2005;25(7):1420e5.
[18] O’Rourke M. Mechanical principles in arterial disease. Hypertension
1995;26(1):2e9.
[19] Owens GK, Kumar MS, Wamhoff BR. Molecular regulation of vascular
smooth muscle cell differentiation in development and disease. Physiol
Rev 2004;84(3):767e801.
[20] Yamamoto K, Sokabe T, Watabe T, Miyazono K, Yamashita JK, Obi S,
et al. Fluid shear stress induces differentiation of Flk-1-positive embry-
onic stem cells into vascular endothelial cells in vitro. Am J Physiol
Heart Circ Physiol 2005;288(4):H1915e24.
[21] Yamashita J, Itoh H, Hirashima M, Ogawa M, Nishikawa S, Yurugi T,
et al. Flk1-positive cells derived from embryonic stem cells serve as
vascular progenitors. Nature 2000;408(6808):92e6.
[22] Huang H, Nakayama Y, Qin K, Yamamoto K, Ando J, Yamashita J, et al.
Differentiation from embryonic stem cells to vascular wall cells under in
vitro pulsatile flow loading. J Artif Organs 2005;8(2):110e8.
[23] Oswald J, Boxberger S, Jorgensen B, Feldmann S, Ehninger G,
Bornhauser M, et al. Mesenchymal stem cells can be differentiated
into endothelial cells in vitro. Stem Cells 2004;22(3):377e84.
[24] Gang EJ, Jeong JA, Han S, Yan Q, Jeon CJ, Kim H. In vitro endothelial
potential of human UC blood-derived mesenchymal stem cells. Cytother-
apy 2006;8(3):215e27.
[25] Hamilton DW, Maul TM, Vorp DA. Characterization of the response of
bone marrow-derived progenitor cells to cyclic strain: implications for
vascular tissue-engineering applications. Tissue Eng 2004;10(3e4):
361e9.
[26] Tao J, Yang Z, Wang JM, Tu C, Pan SR. Effects of fluid shear stress on
eNOS mRNA expression and NO production in human endothelial
progenitor cells. Cardiology 2006;106(2):82e8.
[27] Zeng L, Xiao Q, Margariti A, Zhang Z, Zampetaki A, Patel S, et al.
HDAC3 is crucial in shear- and VEGF-induced stem cell differentiation
toward endothelial cells. J Cell Biol 2006;174(7):1059e69.
[28] Kobayashi N, Yasu T, Ueba H, Sata M, Hashimoto S, Kuroki M, et al.
Mechanical stress promotes the expression of smooth muscle-like prop-
erties in marrow stromal cells. Exp Hematol 2004;32(12):1238e45.
[29] Engelmayr Jr GC, Sales VL, Mayer Jr JE, Sacks MS. Cyclic flexure and
laminar flow synergistically accelerate mesenchymal stem cell-mediated
engineered tissue formation: implications for engineered heart valve
tissues. Biomaterials 2006;27(36):6083e95.
[30] Murphy JM, Fink DJ, Hunziker EB, Barry FP. Stem cell therapy in a cap-
rine model of osteoarthritis. Arthritis Rheum 2003;48(12):3464e74.
[31] Dominici M, Le Blanc K, Mueller I, Slaper-Cortenbach I, Marini F,
Krause D, et al. Minimal criteria for defining multipotent mesenchymal
stromal cells. The International Society for Cellular Therapy position
statement. Cytotherapy 2006;8(4):315e7.
[32] Punchard MA, Stenson-Cox C, O’Cearbhaill ED, Lyons E, Gundy S,
Murphy L, et al. Endothelial cell response to biomechanical forces under
simulated vascular loading conditions. J Biomech 2007;40(14):3146e54.
[33] Johnson GW, Jennings R. LabVIEW graphical programming. McGraw-
Hill Professional; 2001.
[34] Cheng CP, Parker D, Taylor CA. Quantification of wall shear stress in large
blood vessels using Lagrangian interpolation functions with cine phase-con-
trast magnetic resonance imaging. Ann Biomed Eng 2002;30(8):1020e32.
[35] White RA, Klein SR, Shors EC. Preservation of compliance in a small
diameter microporous, silicone rubber vascular prosthesis. J Cardiovasc
Surg (Torino) 1987;28(5):485e90.
[36] He W, Ma Z, Yong T, Teo WE, Ramakrishna S. Fabrication of collagen-
coated biodegradable polymer nanofiber mesh and its potential for endo-
thelial cells growth. Biomaterials 2005;26(36):7606e15.
[37] Abramoff MD, Magelhaes PJ, Ram SJ. Image processing with ImageJ.
Biophotonics Int 2004;11(7):36e42.
[38] Livak KJ, Schmittgen TD. Analysis of relative gene expression data
using real-time quantitative PCR and the 2(-Delta Delta C(T)) method.
Methods 2001;25(4):402e8.
[39] Mackay AM, Beck SC, Murphy JM, Barry FP, Chichester CO,
Pittenger MF. Chondrogenic differentiation of cultured human mesen-
chymal stem cells from marrow. Tissue Eng 1998;4(4):415e28.
[40] Seruya M, Shah A, Pedrotty D, du Laney T, Melgiri R, McKee JA, et al.
Clonal population of adult stem cells: life span and differentiation
potential. Cell Transplant 2004;13(2):93e101.
[41] Wexler SA, Donaldson C, Denning-Kendall P, Rice C, Bradley B,
Hows JM. Adult bone marrow is a rich source of human mesenchymal
‘stem’ cells but umbilical cord and mobilized adult blood are not. Br J
Haematol 2003;121(2):368e74.
[42] Curran JM, Chen R, Hunt JA. The guidance of human mesenchymal stem
cell differentiation in vitro by controlled modifications to the cell sub-
strate. Biomaterials 2006;27(27):4783e93.
[43] Curran JM, Chen R, Hunt JA. Controlling the phenotype and function of
mesenchymal stem cells in vitro by adhesion to silane-modified clean
glass surfaces. Biomaterials 2005;26(34):7057e67.
[44] Lin DT, Young TH, Fang Y. Studies on the effect of surface properties on
the biocompatibility of polyurethane membranes. Biomaterials 2001;
22(12):1521e9.
[45] Butcher JT, Barrett BC, Nerem RM. Equibiaxial strain stimulates fibro-
blastic phenotype shift in smooth muscle cells in an engineered tissue
model of the aortic wall. Biomaterials 2006;27(30):5252e8.
[46] Ziegler T, Bouzourene K, Harrison VJ, Brunner HR, Hayoz D. Influence
of oscillatory and unidirectional flow environments on the expression of
endothelin and nitric oxide synthase in cultured endothelial cells.
Arterioscler Thromb Vasc Biol 1998;18(5):686e92.

1619E.D. O’Cearbhaill et al. / Biomaterials 29 (2008) 1610e1619
[47] Zhao S, Suciu A, Ziegler T, Moore Jr JE, Burki E, Meister JJ, et al.
Synergistic effects of fluid shear stress and cyclic circumferential stretch
on vascular endothelial cell morphology and cytoskeleton. Arterioscler
Thromb Vasc Biol 1995;15(10):1781e6.
[48] Benbrahim A, L’Italien GJ, Kwolek CJ, Petersen MJ, Milinazzo B,
Gertler JP, et al. Characteristics of vascular wall cells subjected to
dynamic cyclic strain and fluid shear conditions in vitro. J Surg Res
1996;65(2):119e27.
[49] Van Kooten TG, Klein CL, Kohler H, Kirkpatrick CJ, Williams DF,
Eloy R. From cytotoxicity to biocompatibility testing in vitro: cell adhe-
sion molecule expression defines a new set of parameters. J Mater Sci
Mater Med 1997;8(12):835e41.
[50] Wijelath ES, Rahman S, Murray J, Patel Y, Savidge G, Sobel M. Fibro-
nectin promotes VEGF-induced CD34 cell differentiation into endothe-
lial cells. J Vasc Surg 2004;39(3):655e60.
[51] Moore Jr JE, Burki E, Suciu A, Zhao S, Burnier M, Brunner HR, et al. A
device for subjecting vascular endothelial cells to both fluid shear stress
and circumferential cyclic stretch. Ann Biomed Eng 1994;22(4):416e22.
[52] Benbrahim A, L’Italien GJ, Milinazzo BB, Warnock DF, Dhara S,
Gertler JP, et al. A compliant tubular device to study the influences of
wall strain and fluid shear stress on cells of the vascular wall. J Vasc
Surg 1994;20(2):184e94.
[53] Nerem RM, Girard PR. Hemodynamic influences on vascular endothelial
biology. Toxicol Pathol 1990;18(4 Pt 1):572e82.
[54] Galbraith CG, Skalak R, Chien S. Shear stress induces spatial reorgani-
zation of the endothelial cell cytoskeleton. Cell Motil Cytoskeleton
1998;40(4):317e30.
[55] Ives CL, Eskin SG, McIntire LV. Mechanical effects on endothelial cell
morphology: in vitro assessment. In Vitro Cell Dev Biol 1986;22(9):
500e7.
[56] Wang JH, Goldschmidt-Clermont P, Wille J, Yin FC. Specificity of
endothelial cell reorientation in response to cyclic mechanical stretching.
J Biomech 2001;34(12):1563e72.
[57] Rogers KA, McKee NH, Kalnins VI. The preferential orientation of
centrioles towards the heart in endothelial cells of major blood vessels
is reestablished following reversal of a segment. Proc Natl Acad Sci U
S A 1985;82:3272e6.
[58] Dunn GA, Heath JP. A new hypothesis of contact guidance in tissue cells.
Exp Cell Res 1976;101(1):1e14.
[59] Rovensky Yu A, Samoilov VI. Morphogenetic response of cultured
normal and transformed fibroblasts, and epitheliocytes, to a cylindrical
substratum surface. Possible role for the actin filament bundle pattern.
J Cell Sci 1994;107(Pt 5):1255e63.
[60] Frame MD, Sarelius IH. Flow-induced cytoskeletal changes in endothelial
cells growing on curved surfaces. Microcirculation 2000;7(6 Pt 1):419e27.
[61] Albinsson S, Nordstrom I, Hellstrand P. Stretch of the vascular wall in-
duces smooth muscle differentiation by promoting actin polymerization.
J Biol Chem 2004;279(33):34849e55.
[62] Birukov KG, Shirinsky VP, Stepanova OV, Tkachuk VA, Hahn AW,
Resink TJ, et al. Stretch affects phenotype and proliferation of vascular
smooth muscle cells. Mol Cell Biochem 1995;144(2):131e9.
[63] Hellstrand P, Albinsson S. Stretch-dependent growth and differentiation
in vascular smooth muscle: role of the actin cytoskeleton. Can J Physiol
Pharmacol 2005;83(10):869e75.
[64] Reusch P, Wagdy H, Reusch R, Wilson E, Ives HE. Mechanical strain
increases smooth muscle and decreases nonmuscle myosin expression
in rat vascular smooth muscle cells. Circ Res 1996;79(5):1046e53.
[65] Zeidan A, Nordstrom I, Albinsson S, Malmqvist U, Sward K,
Hellstrand P. Stretch-induced contractile differentiation of vascular
smooth muscle: sensitivity to actin polymerization inhibitors. Am J
Physiol Cell Physiol 2003;284(6):C1387e96.
[66] Cevallos M, Riha GM, Wang X, Yang H, Yan S, Li M, et al. Cyclic strain
induces expression of specific smooth muscle cell markers in human
endothelial cells. Differentiation 2006;74(9e10):552e61.
[67] Forte A, Cipollaro M, Cascino A, Galderisi U. Pathophysiology of stem
cells in restenosis. Histol Histopathol 2007;22(5):547e57.
Copyright © 2022 FDOKUMEN



















