
Response of bacterial community structure to seasonal fluctuation and anthropogenic pollution on...
9
Response of bacterial community structure to seasonal fluctuation and anthropogenic pollution on coastal water of Alang–Sosiya ship breaking yard, Bhavnagar, India Vilas Patel a , Hitendra Munot b , Yogesh S. Shouche b , Datta Madamwar a,⇑ a Environmental Genomics and Proteomics Lab, BRD School of Biosciences, Vadtal Road, Satellite Campus, Post Box No. 39, Sardar Patel University, Vallabh Vidyanagar 388 120, Gujarat, India b Molecular Biology Unit, National Center for Cell Science, Ganeshkhind, Pune 411007, Maharashtra, India highlights Total 2324 clones and 550 bacterial strains were retrieved from 7 samples. All clones were grouped in 525 distinct operational taxonomic units (OTUs). Anthropogenic activity exerts significant effect on bacterial diversity. Gammaproteobacteria shifted to Betaproteobacteria and Epsilonproteobacteria. Acidovorax and Arcobacter were found to be dominant in polluted samples. article info Article history: Received 6 November 2013 Received in revised form 26 February 2014 Accepted 4 March 2014 Available online 25 March 2014 Keywords: Alang–Sosiya ship breaking yard Bacterial community structure Seasonal variation 16S r-RNA gene Betaproteobacteria abstract Bacterial community structure was analyzed from coastal water of Alang–Sosiya ship breaking yard (ASS- BY), world’s largest ship breaking yard, near Bhavnagar, using 16S rRNA gene sequencing (cultured dependent and culture independent). In clone libraries, total 2324 clones were retrieved from seven sam- ples (coastal water of ASSBY for three seasons along with one pristine coastal water) which were grouped in 525 operational taxonomic units. Proteobacteria was found to be dominant in all samples. In pristine samples, Gammaproteobacteria was found to be dominant, whereas in polluted samples dominancy of Gammaproteobacteria has shifted to Betaproteobacteria and Epsilonproteobacteria. Richness and diversity indices also indicated that bacterial community in pristine sample was the most diverse followed by summer, monsoon and winter samples. To the best of knowledge, this is the first study describing bac- terial community structure from coastal water of ASSBY, and it suggests that seasonal fluctuation and anthropogenic pollutions alters the bacterial community structure. Ó 2014 Elsevier Ltd. All rights reserved. 1. Introduction Coastal and estuarine systems are amongst the most productive and diverse ecosystems in the world. The marine ecosystem harbors complex microbial communities that afford ecological sus- tenance by maintaining biogeochemical cycles. Advances in molec- ular techniques and ecological genomics have greatly improved understanding of bacterial diversity in marine ecosystem. These studies have proved that marine microbial community is complex, widespread and consists of unidentified or uncultivated members. However, there are still large gaps in basic ecological information about bacterial community due to their seasonal and biogeograph- ical distributions in marine ecosystem (Treusch et al., 2009). Effect of seasonal fluctuation on bacterial community structure from marine ecosystem such as the Sargasso, Baltic Seas and the English Channel and others have been studied thoroughly and revealed about tight coupling of environmental parameters with the bacte- rial community structure (Andersson et al., 2010; Carlson et al., 2009; Gilbert et al., 2009; Morris et al., 2005). In addition to these, the intertidal zone of marine ecosystem, which occupies the upper edge of the world’s coasts extending over 1,600,000 km, is considered as one of the most variable coast- al habitat due to higher biological productivity and economic va- lue. Coastal area mainly comprises of rocky platforms, sandy http://dx.doi.org/10.1016/j.biortech.2014.03.033 0960-8524/Ó 2014 Elsevier Ltd. All rights reserved. ⇑ Corresponding author. Tel.: +91 2692 229380; fax: +91 2692 236475. E-mail addresses: [email protected] (V. Patel), datta_madamwar@yahoo. com (D. Madamwar). Bioresource Technology 161 (2014) 362–370 Contents lists available at ScienceDirect Bioresource Technology journal homepage: www.elsevier.com/locate/biortech
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Response of bacterial community structure to seasonal fluctuation and anthropogenic pollution on...

Bioresource Technology 161 (2014) 362–370
Contents lists available at ScienceDirect
Bioresource Technology
journal homepage: www.elsevier .com/locate /bior tech
Response of bacterial community structure to seasonal fluctuation andanthropogenic pollution on coastal water of Alang–Sosiya ship breakingyard, Bhavnagar, India
http://dx.doi.org/10.1016/j.biortech.2014.03.0330960-8524/� 2014 Elsevier Ltd. All rights reserved.
⇑ Corresponding author. Tel.: +91 2692 229380; fax: +91 2692 236475.E-mail addresses: [email protected] (V. Patel), datta_madamwar@yahoo.
com (D. Madamwar).
Vilas Patel a, Hitendra Munot b, Yogesh S. Shouche b, Datta Madamwar a,⇑a Environmental Genomics and Proteomics Lab, BRD School of Biosciences, Vadtal Road, Satellite Campus, Post Box No. 39, Sardar Patel University, Vallabh Vidyanagar 388 120,Gujarat, Indiab Molecular Biology Unit, National Center for Cell Science, Ganeshkhind, Pune 411007, Maharashtra, India
h i g h l i g h t s
� Total 2324 clones and 550 bacterial strains were retrieved from 7 samples.� All clones were grouped in 525 distinct operational taxonomic units (OTUs).� Anthropogenic activity exerts significant effect on bacterial diversity.� Gammaproteobacteria shifted to Betaproteobacteria and Epsilonproteobacteria.� Acidovorax and Arcobacter were found to be dominant in polluted samples.
a r t i c l e i n f o
Article history:Received 6 November 2013Received in revised form 26 February 2014Accepted 4 March 2014Available online 25 March 2014
Keywords:Alang–Sosiya ship breaking yardBacterial community structureSeasonal variation16S r-RNA geneBetaproteobacteria
a b s t r a c t
Bacterial community structure was analyzed from coastal water of Alang–Sosiya ship breaking yard (ASS-BY), world’s largest ship breaking yard, near Bhavnagar, using 16S rRNA gene sequencing (cultureddependent and culture independent). In clone libraries, total 2324 clones were retrieved from seven sam-ples (coastal water of ASSBY for three seasons along with one pristine coastal water) which were groupedin 525 operational taxonomic units. Proteobacteria was found to be dominant in all samples. In pristinesamples, Gammaproteobacteria was found to be dominant, whereas in polluted samples dominancy ofGammaproteobacteria has shifted to Betaproteobacteria and Epsilonproteobacteria. Richness and diversityindices also indicated that bacterial community in pristine sample was the most diverse followed bysummer, monsoon and winter samples. To the best of knowledge, this is the first study describing bac-terial community structure from coastal water of ASSBY, and it suggests that seasonal fluctuation andanthropogenic pollutions alters the bacterial community structure.
� 2014 Elsevier Ltd. All rights reserved.
1. Introduction
Coastal and estuarine systems are amongst the most productiveand diverse ecosystems in the world. The marine ecosystemharbors complex microbial communities that afford ecological sus-tenance by maintaining biogeochemical cycles. Advances in molec-ular techniques and ecological genomics have greatly improvedunderstanding of bacterial diversity in marine ecosystem. Thesestudies have proved that marine microbial community is complex,widespread and consists of unidentified or uncultivated members.
However, there are still large gaps in basic ecological informationabout bacterial community due to their seasonal and biogeograph-ical distributions in marine ecosystem (Treusch et al., 2009). Effectof seasonal fluctuation on bacterial community structure frommarine ecosystem such as the Sargasso, Baltic Seas and the EnglishChannel and others have been studied thoroughly and revealedabout tight coupling of environmental parameters with the bacte-rial community structure (Andersson et al., 2010; Carlson et al.,2009; Gilbert et al., 2009; Morris et al., 2005).
In addition to these, the intertidal zone of marine ecosystem,which occupies the upper edge of the world’s coasts extendingover 1,600,000 km, is considered as one of the most variable coast-al habitat due to higher biological productivity and economic va-lue. Coastal area mainly comprises of rocky platforms, sandy

V. Patel et al. / Bioresource Technology 161 (2014) 362–370 363
beaches, mudflats, estuaries, salt marshes, mangrove forests, cer-tain coral reefs, and human-made infrastructures (Bertness et al.,2001). However, coastal zones of the world’s oceans are increas-ingly subjected to the discharge of anthropogenic pollutants, rang-ing from domestic to industrial effluents. These organic pollutantsare finally subjected to marine ecosystem. These input and accu-mulation of pollutant compounds is seen as a major threat forthe sea coast ecosystem, resulting in eutrophication. Eutrophica-tion is known to cause deleterious effects on bacterial communitystructure which lead resistant, resilient, functional redundancy orperturbation in bacterial community structure which can alterthe functionality of the community and implies a negative effecton ecosystem health, on its economic and social value (Nogaleset al., 2011).
One of such eutrophic coastal zone in India is Alang–Sosiya shipbreaking yard near Bhavnagar, Gujarat. The ASSBY is one of thelargest ship breaking yard in the world having over 41% of the totalworld’s ship breaking industries. With the rapid development ofthe international trade business and the sudden increase in popu-lation in the past 10 years, the coastal regions of the ASSBY are fac-ing many ecological problems. To date, previous studies hasreported fate of pollution due to ship breaking industries (Reddyet al., 2003, 2005; Basha et al., 2007). Preliminary study was to de-velop bacterial consortium for polycyclic aromatic hydrocarbon(PAH) degradation from coastal sediment of ASSBY (Patel et al.,2013). The present study describes analysis of bacterial communitystructure from coastal waters of the ASSBY during different seasonsusing 16S rRNA gene sequences. In present study, one more coastalwater sample (pristine) was chosen from the coastal ecosystem inthe outer part (10 km away) of the ASSBY to determine the effect ofanthropogenic activities on bacterial community structure.
2. Methods
2.1. Sample collection
The Alang–Sosiya ship scrapping yard is geographically situated2105� 21029� towards the north and 7205� 72015� towards the easton the Western Coast of the Gulf of Cambay (Fig. 1). The yardstretches to about 14 km along the north–south, encompassing atotal area of approximately 67 km2 with a small creek bifurcatinginto nearly two equal parts. The southern part is designated asAlang while the other is known as Sosiya, popularly called theAlang–Sosiya ship scrapping yard (Fig. 1). In present study, Thesediment samples were collected from two sampling sites DVASW(from Alang) and DVBSW (from Sosiya) at the Alang–Sosiya shipbreaking yard, Gujarat, India during three different seasons viz.winter (DVASW_D and DVBSW_D dated on 29-12-2010), monsoon(DVASW_J and DVBSW_J, D dated on 17-8-2011) and summer(DVASW_M and DVBSW_M, dated on 28-4-2012). Further, to eval-uate the effect of pollution on bacterial community structure atAlang–Sosiya, pristine sea water samples designated as DVPSWwas taken 10 km away from the Alang–Sosiya coast, wherein theno anthropogenic activity takes place. Total concentrations ofpotentially toxic metals and organic carbon in the coastal waterwere determined using Perkin-Elmer ICP Optima-3300RL (PerkinElmer, Norwalk, CT) and a TOC-V analyzer (Shimadzu, Japan)respectively.
2.2. Isolation of bacterial strains
Coastal water samples were serially diluted and spreaded on (1)high-nutrient media (Nutrient agar, Luria agar and Plate countagar) and incubated for 2–5 days at 20 �C, 30 �C and 37 �C and on(2) low-nutrient media (Zobell agar, Sea water agar, Soil extract
agar and Reasonar’s agar) and incubated for 15 days at 20 �C,30 �C and 37 �C. Bacterial strains were also isolated on differentmedia (as listed above) and were supplemented with petroleumhydrocarbons such as benzene, phenanthrene, diesel fuel (0.1%,w/v) and salt (3%, w/v). Since this coastal environment is contam-inated with anthropogenic pollutants, attempts were made to iso-late organisms on minimal media such as Bushnell-Hass Mediumagar (BHM agar) containing petroleum hydrocarbons like benzeneand phenanthrene as carbon source. Before use of BHM agar,0.025% (w/v) of phenanthrene was prepared in acetone and wasspreaded on the surface of the medium as the sole carbon source.A total of 60–70 bacterial strains were isolated from all sevencoastal water samples based on the distinct morphological charac-teristics of the colonies. Further, all bacterial strains were main-tained at 4 �C.
2.3. Extraction of genomic DNA from bacterial isolates and communityDNA extraction from coastal water samples
The DNA was extracted from freshly grown cultures using stan-dard Phenol, Chloroform method (Sambrook and Russell, 2000).For metagenomic DNA, 2.5 L of the coastal water sample was fil-tered through 0.22 lm filters. The filters obtained were used forthe isolation of metagenomic DNA as describe by Desai et al.(2009) with some modifications. In the present study extractedDNA was again purified by polyvinylpolypyrrolidone (PVPP) col-umn. DNA concentration was measured by means of a NanoDropspectrophotometer (NanoDrop Technologies Inc., Delaware, USA)and analyzed by gel electrophoresis.
2.4. Identification of isolates by 16S rRNA gene sequence analysis
All isolated bacterial strains were processed for Amplified r-DNA restriction analysis (ARDRA) as described by Desai et al.(2009) to remove similar bacterial species from all seven samplesand bacterial strain based on their unique banding pattern wereselected for sequences. For identification of unique bacterialstrains. 16S rRNA gene of all unique bacterial strains were ampli-fied using universal primer set 8F (50-CCAGAGTTTGATCGTGGCT-CAG-30) and 1492R (50-TTC TGC AGT CTA GAA GGA GGT GWTCCA GCC-30) (Desai et al., 2009) and sequenced using the Big DyesTerminator v3.1 cycle sequencing kit (Applied Biosystems).
2.5. Phylogenetic analysis
To characterize the bacterial community structure from all se-ven samples, 16S rRNA genes from metagenomic DNA were ampli-fied using 121F and 1492R primers and cloned into Escherichia coliDH5a competent cells as described by Desai et al. (2009). Plasmidsof positive clones were isolated using Alkali-lysis method (Sam-brook and Russell, 2000). A total of 2324 16S rRNA gene clones(approximately 320–350 from each site) were randomly selectedand subjected to 16S rRNA gene sequencing from the 7 clone li-braries using the Big Dyes Terminator v3.1 cycle sequencing kit(Applied Biosystems). Sequences from each of the clone librariesas well as from culture isolates were compared to the current data-base of 16S r-RNA gene sequences at Ribosomal Database Project II.The sequences were assembled and contig’s were obtained usingChromasPro software, alignment was done using CLUSTAL X2and the sequences were edited manually using DAMBE to getunambiguous sequence alignment. All sequences were checkedfor chimeric artifacts by Mallard program, reference sequence usedfor this purpose was E. coli U000096. Appropriate subsets of16S r-RNA gene sequences were selected and subjected to furtherphylogenetic analysis using DNADIST of Phylip (version 3.61). Thenumber of Operational Taxonomic Units (OTU) (clone sequences

Fig. 1. Geographical location of the study area of Alang–Sosiya and Mahuva on the Gulf of Cambay.
364 V. Patel et al. / Bioresource Technology 161 (2014) 362–370
with >97% similarity were grouped together and regarded as oneOTU) was obtained by DOTUR program (version 1.53) using fur-thest neighbor algorithm (Grote et al., 2008). Representative se-quences from each of the OTUs were retrieved and checkedagainst the previously determined 16S rRNA gene from the RDPIIrelease 10 version of the database and these sequences weredownloaded in FASTA format. Phylogenies were constructed withthe Molecular Evolutionary Genetics Analysis v3.0 program usingthe Kimura two-parameter model and the Neighbour-joining algo-rithm. The Significance of branching order was determined usingbootstrap analysis with 1000 re-sampled data sets. In order todetermine the significance of differences between the clone li-braries, the Principal co-ordination analysis plot was constructedusing online Unifrac program. Diversity indices and richness weredetermined as described previously by Grote et al. (2008).
The sequences are submitted to NCBI with accession numbersfrom KF722317 to KF722621 (DVASW_J), KF722010 to KF722316(DVBSW_J), KF721413 to KF721699 (DVASW_D), KF721700 toKF722009 (DVBSW_D), KF755695 to KF756010 (DVASW_M),KF755386 to KF755694 (DVBSW_M), KF756160 to KF75646(DVPSW) and KF756011 to KF756159 (Culturable isolates of all se-ven samples) on the basis of sequence similarities as obtained fromRibosomal Database Project II (RDP II).
3. Results
3.1. Physico-chemical parameters of sea coastal water of Alang–Sosiyaship breaking yard
The physicochemical characteristics of all seven coastal watersamples are given in Table 1. As expected, ASSBY showed clear sea-sonal fluctuation and effect of ship breaking activities on tempera-ture, COD, BOD and anthropogenic pollutants specifically on iron.As expected, water temperature of summer samples was highand lower in winter.COD and BOD concentration was high in pol-luted samples compared to pristine samples. Metal concentrationwas also found to be higher in polluted samples compared to pris-tine samples. Similarly, sulfate concentration varied from 0.98% to3.67%, in all samples whereas maximum concentration was foundin winter samples (DVBSW_D), 3.67% of ASSBY. Total organic car-bon was found to be 0.57–0.84% and 0.65% in polluted samplesand pristine sample, respectively.
3.2. Bacterial community structure using culture dependent approach
Total bacterial counts varied from 2.3 � 103 to 21.3 � 105 CFU/ml in all seven samples on high as well as low nutrient media.

Table 1Physico-chemical analysis of polluted coastal water from ASSBY as well as from pristine sea water.
Monsoon sea water Winter sea water Summer sea water Pristine
DVASW_J DVBSW_J DVASW_D DVBSW_D DVASW_M DVBSW_M DVPSW
Temperature (�C) 30 32 22 22 40 40 34COD (mg/l) 3400 5300 7000 4500 1690 5500 250BOD (mg/l) 1380 2460 3160 2300 870 2780 120Organic carbons 0.57 0.75 0.78 0.84 0.66 0.79 0.65Water soluble sulfur (%) 2.5 1.9 3.52 3.67 2.88 2.33 0.23Total solid (mg/l) 56,000 40,000 44,000 46,000 44,000 38,000 33,350Total dissolved solid (mg/l) 42,000 38,000 38,000 39,000 35,700 34,000 32,000Total suspended solid (mg/l) 14,000 12,000 6000 70,000 7300 4,000 1500Cadmium (Cd) (ppm) BDL BDL BDL BDL BDL BDL BDLChromium (Cr) 2.2051 2.5051 2.6 1.6051 1.1890 2.6051 BDLLead (Pb) (ppm) 0.8 1.0 1.1 BDL BDL 1.1 0.08Zinc (Zn) (ppm) 0.4 0.6 0.5 0.3208 BDL 0.520 0.18Iron (Fe) (ppm) 22 2.4 59 164.99 3.8 0.2 0.69
V. Patel et al. / Bioresource Technology 161 (2014) 362–370 365
Since this coastal environment is contaminated with anthropo-genic pollutants, attempts were made to isolate organisms onBHM agar containing petroleum hydrocarbons like benzene andphenanthrene as carbon source. Bacterial counts on BHM agarshowed clear difference between polluted and pristine sampleswith high abundance in polluted samples. Total bacterial countsranging from 1.9 � 102 to 4 � 104 CFU/ml were obtained from sam-ples of polluted ecosystems on BHM agar containing petroleumhydrocarbon, whereas only 35 bacterial counts were obtained onBHM agar containing petroleum hydrocarbon from pristine ecosys-tem. This could be an adaptation of the microorganisms in the pol-luted coastal water of Alang–Sosiya ship breaking yard to toleratehigh concentration of PAHs as well as to utilize them as the carbonsource. All isolated strains from all seven coastal water sampleswere processed for ARDRA analysis. Bacterial isolates showing un-ique banding pattern were selected and sequenced to analyze cul-turable bacterial community structure. Bacterial communitystructure using culture dependent approach is shown in Fig. 2.
Proteobacteria and Firmicutes were recovered from all samples.Different species of Planococcus and Bacillus were isolated fromall samples, whereas Staphylococcus, Planomicrobium and Exiguo-bacterium were isolated only from polluted samples. In Proteobac-teria phylum, Gammaproteobacteria was observed in polluted and
0
20
40
60
80
100
Rel
ativ
e ab
udan
ce o
f gen
us (%
)
Sea water samples
Alteromonas
Psychrobacter
Pusillimonas
Planomicrobium
Stenotrophomonas
Marinobacter
Halomonas
Exiguobacterium
Rheinheimera
Idiomarina
Planococcus
Brevibacillus
Alcaligenes
Staphylococcus
Pseudomonas
Bacillus
Fig. 2. Genus level comparison of culturable bacterial community structure fromASSBY. [Monsoon Season (Alang, DVASW_J and Sosiya, DVBSW_J), winter Season(Alang, DVASW_D and Sosiya, DVBSW_D), Summer Season of ASSBY (Alang,DVASW_M and Sosiya, DVBSW_M) and pristine coastal sea water (DVPSW)].
pristine samples, while Betaproteobacteria was acquired only frompolluted samples (Fig. 2). At genus level, Pseudomonas was ob-served in all samples, whereas Psychrobacter and Cobetia werefound to be present in monsoon and pristine sample of ASSBY. Inaddition to these, different species of Marinobacter, Alcaligenes,Rheinheimera, Pseudomonas, Halomonas, Stenotrophomonas, Psych-robacter, Pusillimonas and Alteromonas were also isolated from pol-luted samples.
3.3. Bacterial community structure using culture independentapproach
In culture independent approach, Total 2324 clone sequenceswere retrieved which were grouped in 525 OTUs based on DOTURanalysis, of which some sequences were archea and were thereforeexcluded in the subsequent analyses. These sequences (2324)/OTUs (525) were assigned to six different phyla or groups on thebasis of sequence similarities as obtained from Ribosomal Data-base Project II (RDP II). Each of the seven clone libraries containedbetween 300 and 350 clones, with OTUs ranging from 56 to 106.Rarefaction analysis for all seven cloned libraries also showed thatcoastal water samples from summer as well as from pristine coast-al sea water had higher diversity followed by monsoon and winter.The curve (Fig. 3) indicates that the diversity was sampled withgood level of confidence and majority of OTUs in the sample weredetected but still there is a need of more comprehensive samplingto cover rest of the undiscovered diversity from summer coastalwater samples of Alang–Sosiya ship breaking yard and pristinecoastal water. Statistical analyses of biodiversity provide interest-ing insights as shown in Table 2. The Good’s coverage index revealsthat 93% to 95% of the species were obtained in winter season sam-ples (DVASW_D and DVBSW_D), 86–88% for samples of summerseason (DVASW_M, DVBSW_M) and monsoon samples (DVASW_Jand DVBSW_J), while good coverage in the pristine sample(DVPSW) was found to be 85%. The maximum amount of OTUs ob-served was also reflected in the Chao Jcckknife and ACE values pre-dicted for all the samples (Table 2). The lowest Shannon diversityindices in the winter sample libraries (DVASW_D and DVBSW_J)indicated that its microbial community was composed of a fewphylotypes, while in the remaining samples, the diversity indicesshowed a higher level of species richness. Simpson index alsoshowed that dominance was more observed in DVASW_D andDVBSW_J libraries as compared to other samples.
3.3.1. Taxonomic compositionAll sequences were classified from phylum to genus according
to the (RDP II); four different phyla or groups were identified fromthese seven samples. The seven libraries showed similar 16S rRNA

0
20
40
60
80
100
120
1 31 61 91 121 151 181 211 241 271 301 331
Exp
ecte
d nu
ber
of O
TU
s per
sa
mpl
es
16S r-RNA gene sequences per sample
DVBSW_M DVASW_M DVPSW DVASW_D
DVBSW_D DVASW_J DVBSW_J
Fig. 3. Rarefaction curves of clone libraries from all seven samples.
Table 2Statistical indices calculated for seven libraries.
Monsoon coastal sea water Winter coastal sea water Summer coastal sea water Pristine
DVASW_D DVBSW_D DVASW_J DVBSW_J DVASW_M DVBSW_M DVPSW
Clones 338 338 329 319 319 316 315OTU 59 80 89 66 95 80 106Singleton 19 27 49 32 41 37 50Shannon Index 3.397 3.507 3.81 3.374 3.87 3.677 4.14Simpson Index 0.0553 0.0557 0.034 0.0513 0.039 0.042 0.023Good’s coverage 95 93 86 90 89 89 85Chaol 71.76 113.053 179.462 144.143 130.652 121.353 166.273Jackknife 78 116.218 192.164 124.716 136.782 123.074 180.946ACE 77.38 142.41 183.772 141.713 142.08 139.961 188.85
366 V. Patel et al. / Bioresource Technology 161 (2014) 362–370
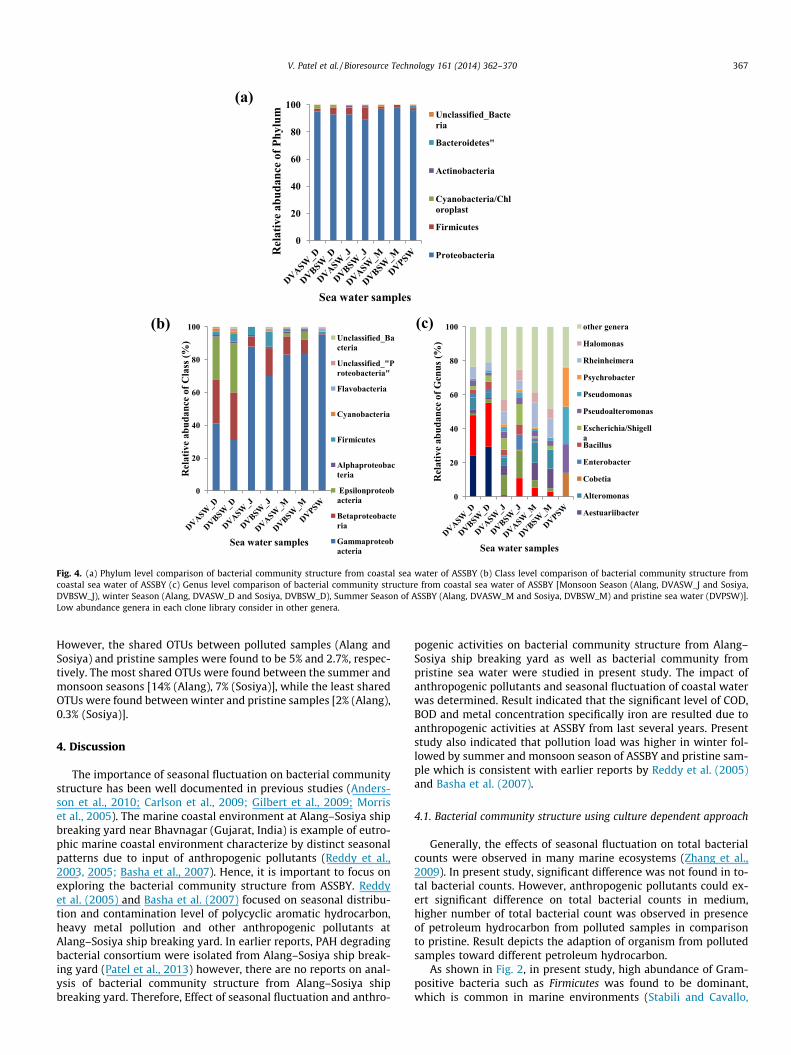
profiles at phylum level distributions but it showed different ten-dencies at class level and genus level as depicted in Fig. 4. The Phy-logenetic tree of the OTU’s obtained from all seven libraries isrepresented in Supplementary file, Figs. S1–S7.
As shown in Fig. 4, The DVPSW (pristine) clone library includedthree different phyla, where Proteobacteria (97%, with 80 OTU) wasthe most abundant phylum followed by Firmicutes (2%, 9 OTU) andBacteroidetes (2%, 5 OTU), representing 99% of the total sequences.The DVASW_J (Alang sample in monsoon) library was mainlynumerically dominated by Proteobacteria (93%, with 78 OTU), Fir-micutes (5%, 7 OTU), Actinobacteria (1%, 1 OTU) and Cyanobacteria(0.28%, with 1OTU), while DVBSW_J (Sosiya sample in monsoon) li-brary was dominated by Proteobacteria (90%, with 52 OTU), Firmi-cutes (9%, 8 OTU) and Actinobacteria (1%, 1 OTU). The summerseason samples, DVASW_M comprises (97%, with 75 OTU) and(1.5%, with 2OTU), while DVBSW_M comprised only one phylumProteobacteria (98%, with 89OTU). The winter seasons samples(DVASW_D and DVBSW_D) library was also comprised of three dif-ferent phyla Proteobacteria (95%, with 51 OUT; 93%, 62 OTU), Firmi-cutes (2%, 3 OTU; 5% 8OTU) and Cyanobacteria (2% with 1 OTU; 1%with 1OTU), respectively which accounted for 99% of totalsequences.
The most abundant OTUs within the different samples weredetermined to understand further the important bacteria and max-imum-parsimony was conducted to compare distribution of OTUsbetween all samples (Fig. 5). The most abundant genera retrievedfrom DVPSW clone library were related to Cobetia (14%, 24 OTU),Psychrobacter (23%, 24 OTU), Pseudoalteromonas (17%, 8 OTU) and
Pseudomonas (22%, 12 OTU). The DVASW_J library was dominatedby sequences related to Aceinetobacter (12%, 3 OTU) followed byRheinheimera (7.62%, 4OTU) and Escherichia/Shigella (6.7%, 3 OTU),whereas the DVBSW_J libraries were also dominated by Aceinetob-acter (16.3%, 4 OTU) followed by Escherichia/Shigella (12%, 3 OTU),Acidovorax (11%, 2 OTU) and Rheinheimera (5.34%, 1 OTU). In summer(DVASW_M) sample libraries, Rheinheimera (15%, 7 OTU) was themost abundant followed by Alteromonas (12%, 6 OTU), Aestuariibact-er (10.3%, 1 OTU), Halomonas (6%, 4 OTU) and Acidovorax (5.3%, 3OTU) whereas in DVBSW_M, Aestuariibacter (11.6%, 5 OTU) werethe most abundant followed by Alteromonas (11%, 6 OTU), Rheinhei-mera (11%, 7 OTU) and Halomonas (6%, 4 OTU). In winter samples(DVASW_D), Acidovorax (24%, 12 OTU) was the most abundant genusfollowed by Arcobacter (24%, 7 OTU), Alteromonas (7%, 3 OTU) andRheinheimera (7%, 4 OTU), whereas in DVBSW_D, Acidovorax (26%,13 OTU) and Arcobacter (30%, 11 OTU) were the most abundant fol-lowed by Alteromonas (3%, 1 OTU), Rheinheimera (4.7%, 5 OTU).
3.3.2. Comparison of all samples at OTUs levelThe Venn diagram demonstrates the distribution of OTUs at the
3% level of dissimilarity. Venn diagrams were constructed to eval-uate similarities of OTUs between all samples of Alang and pristineis depicted in Fig. 6(a), whereas similarities of OTUs between allsamples of Sosiya and pristine is depicted in Fig. 6(b). All three pol-luted samples of Alang and Sosiya shared 16% and 24% of thesetotal OTUs, respectively. These OTUs were affiliated to theAcidovorax, Enterobacter, Alteromonas, Aceinetobacter andPseudomonas indicated important role at ecosystem of ASSBY.
0
20
40
60
80
100
Rel
ativ
e ab
udan
ce o
f Phy
lum
Sea water samples
Unclassified_Bacteria
Bacteroidetes"
Actinobacteria
Cyanobacteria/Chloroplast
Firmicutes
Proteobacteria
0
20
40
60
80
100
Rel
ativ
e ab
udan
ce o
f Cla
ss (%
)
Sea water samples
Unclassified_Bacteria
Unclassified_"Proteobacteria"
Flavobacteria
Cyanobacteria
Firmicutes
Alphaproteobacteria
Epsilonproteobacteria
Betaproteobacteria
Gammaproteobacteria
0
20
40
60
80
100
Rel
ativ
e ab
udan
ce o
f Gen
us (%
)
Sea water samples
other genera
Halomonas
Rheinheimera
Psychrobacter
Pseudomonas
Pseudoalteromonas
Escherichia/ShigellaBacillus
Enterobacter
Cobetia
Alteromonas
Aestuariibacter
(a)
(b) (c)
Fig. 4. (a) Phylum level comparison of bacterial community structure from coastal sea water of ASSBY (b) Class level comparison of bacterial community structure fromcoastal sea water of ASSBY (c) Genus level comparison of bacterial community structure from coastal sea water of ASSBY [Monsoon Season (Alang, DVASW_J and Sosiya,DVBSW_J), winter Season (Alang, DVASW_D and Sosiya, DVBSW_D), Summer Season of ASSBY (Alang, DVASW_M and Sosiya, DVBSW_M) and pristine sea water (DVPSW)].Low abundance genera in each clone library consider in other genera.
V. Patel et al. / Bioresource Technology 161 (2014) 362–370 367
However, the shared OTUs between polluted samples (Alang andSosiya) and pristine samples were found to be 5% and 2.7%, respec-tively. The most shared OTUs were found between the summer andmonsoon seasons [14% (Alang), 7% (Sosiya)], while the least sharedOTUs were found between winter and pristine samples [2% (Alang),0.3% (Sosiya)].
4. Discussion
The importance of seasonal fluctuation on bacterial communitystructure has been well documented in previous studies (Anders-son et al., 2010; Carlson et al., 2009; Gilbert et al., 2009; Morriset al., 2005). The marine coastal environment at Alang–Sosiya shipbreaking yard near Bhavnagar (Gujarat, India) is example of eutro-phic marine coastal environment characterize by distinct seasonalpatterns due to input of anthropogenic pollutants (Reddy et al.,2003, 2005; Basha et al., 2007). Hence, it is important to focus onexploring the bacterial community structure from ASSBY. Reddyet al. (2005) and Basha et al. (2007) focused on seasonal distribu-tion and contamination level of polycyclic aromatic hydrocarbon,heavy metal pollution and other anthropogenic pollutants atAlang–Sosiya ship breaking yard. In earlier reports, PAH degradingbacterial consortium were isolated from Alang–Sosiya ship break-ing yard (Patel et al., 2013) however, there are no reports on anal-ysis of bacterial community structure from Alang–Sosiya shipbreaking yard. Therefore, Effect of seasonal fluctuation and anthro-
pogenic activities on bacterial community structure from Alang–Sosiya ship breaking yard as well as bacterial community frompristine sea water were studied in present study. The impact ofanthropogenic pollutants and seasonal fluctuation of coastal waterwas determined. Result indicated that the significant level of COD,BOD and metal concentration specifically iron are resulted due toanthropogenic activities at ASSBY from last several years. Presentstudy also indicated that pollution load was higher in winter fol-lowed by summer and monsoon season of ASSBY and pristine sam-ple which is consistent with earlier reports by Reddy et al. (2005)and Basha et al. (2007).
4.1. Bacterial community structure using culture dependent approach
Generally, the effects of seasonal fluctuation on total bacterialcounts were observed in many marine ecosystems (Zhang et al.,2009). In present study, significant difference was not found in to-tal bacterial counts. However, anthropogenic pollutants could ex-ert significant difference on total bacterial counts in medium,higher number of total bacterial count was observed in presenceof petroleum hydrocarbon from polluted samples in comparisonto pristine. Result depicts the adaption of organism from pollutedsamples toward different petroleum hydrocarbon.
As shown in Fig. 2, in present study, high abundance of Gram-positive bacteria such as Firmicutes was found to be dominant,which is common in marine environments (Stabili and Cavallo,
%62%50
1 2 3 4 5 6 7
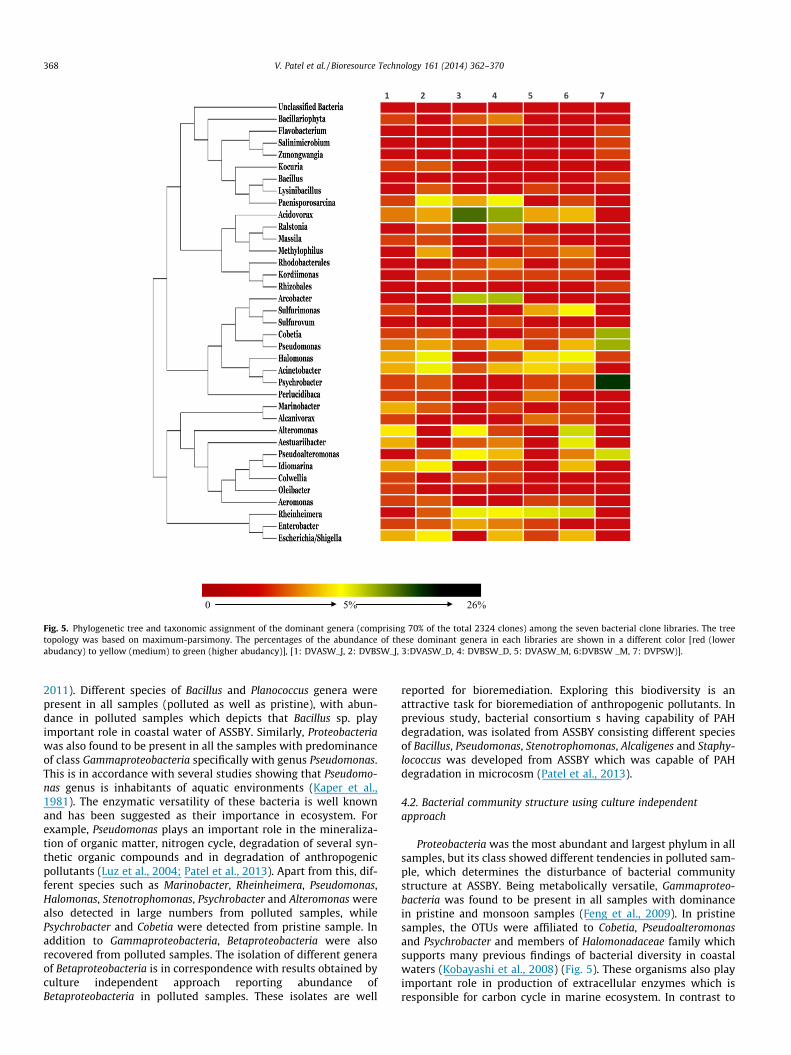
Fig. 5. Phylogenetic tree and taxonomic assignment of the dominant genera (comprising 70% of the total 2324 clones) among the seven bacterial clone libraries. The treetopology was based on maximum-parsimony. The percentages of the abundance of these dominant genera in each libraries are shown in a different color [red (lowerabudancy) to yellow (medium) to green (higher abudancy)], [1: DVASW_J, 2: DVBSW_J, 3:DVASW_D, 4: DVBSW_D, 5: DVASW_M, 6:DVBSW _M, 7: DVPSW)].
368 V. Patel et al. / Bioresource Technology 161 (2014) 362–370
2011). Different species of Bacillus and Planococcus genera werepresent in all samples (polluted as well as pristine), with abun-dance in polluted samples which depicts that Bacillus sp. playimportant role in coastal water of ASSBY. Similarly, Proteobacteriawas also found to be present in all the samples with predominanceof class Gammaproteobacteria specifically with genus Pseudomonas.This is in accordance with several studies showing that Pseudomo-nas genus is inhabitants of aquatic environments (Kaper et al.,1981). The enzymatic versatility of these bacteria is well knownand has been suggested as their importance in ecosystem. Forexample, Pseudomonas plays an important role in the mineraliza-tion of organic matter, nitrogen cycle, degradation of several syn-thetic organic compounds and in degradation of anthropogenicpollutants (Luz et al., 2004; Patel et al., 2013). Apart from this, dif-ferent species such as Marinobacter, Rheinheimera, Pseudomonas,Halomonas, Stenotrophomonas, Psychrobacter and Alteromonas werealso detected in large numbers from polluted samples, whilePsychrobacter and Cobetia were detected from pristine sample. Inaddition to Gammaproteobacteria, Betaproteobacteria were alsorecovered from polluted samples. The isolation of different generaof Betaproteobacteria is in correspondence with results obtained byculture independent approach reporting abundance ofBetaproteobacteria in polluted samples. These isolates are well
reported for bioremediation. Exploring this biodiversity is anattractive task for bioremediation of anthropogenic pollutants. Inprevious study, bacterial consortium s having capability of PAHdegradation, was isolated from ASSBY consisting different speciesof Bacillus, Pseudomonas, Stenotrophomonas, Alcaligenes and Staphy-lococcus was developed from ASSBY which was capable of PAHdegradation in microcosm (Patel et al., 2013).
4.2. Bacterial community structure using culture independentapproach
Proteobacteria was the most abundant and largest phylum in allsamples, but its class showed different tendencies in polluted sam-ple, which determines the disturbance of bacterial communitystructure at ASSBY. Being metabolically versatile, Gammaproteo-bacteria was found to be present in all samples with dominancein pristine and monsoon samples (Feng et al., 2009). In pristinesamples, the OTUs were affiliated to Cobetia, Pseudoalteromonasand Psychrobacter and members of Halomonadaceae family whichsupports many previous findings of bacterial diversity in coastalwaters (Kobayashi et al., 2008) (Fig. 5). These organisms also playimportant role in production of extracellular enzymes which isresponsible for carbon cycle in marine ecosystem. In contrast to

Fig. 6. (a) Venn diagram showing the comparison OTUs from all libraries of Alang[(monsoon (DVASW_J), winter (DVASW_D) and summer (DVASW_M)] and pristinesea water (b) Venn diagram showing the comparisons of OTUs from all libraries ofSosiya Alang [(monsoon (DVBSW_J), winter (DVBSW_D) and summer (DVBSW_M)]and pristine sea water, at 97% similarity. The size of the spheres is not consistentwith the amount of phylotypes present. The numbers of OTU are presentedseparately in each sphere represent unique OTU of each library.
V. Patel et al. / Bioresource Technology 161 (2014) 362–370 369
these, in polluted samples different genera of Alteromonadaceaeand Chromatiaceae family were found to be dominant which de-picts the effect of anthropogenic pollutants on bacterial diversityof ASSBY. The high abundance of Alteromonadaceae in water sam-ples of ASSBY correspondence with earlier report by Feng et al.(2009). Alteromonadaceae (such as Alteromonas and Aeustribacter)and is common copiotroph in marine ecosystem generally foundin nutrient rich coastal environment. Moreover, Alteromonas arealso reported for crude oil degradation. Thus it may be anticipatedthat members of the genus Alteromonas have potential role incrude oil degradation. Abundance of chemoautotrophic bacteriasuch as Rheinheimera can be explained by heavy metal contamina-tions such iron at ASSBY. Aceinetobacter commonly lives in soil andwater (Warskow and Juni, 1972) and have an ability to degradewide range of anthropogenic pollutants. The abundance of thesegenera has been recently reported in other eutrophic marine eco-system (Newton et al., 2013). Other hydrocarbon degrading organ-isms such as Marinobacter commonly identified as petroleumhydrocarbon degraders and Alcanivorax, known as oil hydrocarbondegrader in the marine environment (Yakimov et al., 2007) werealso recovered from coastal environment of ASSBY. The increasein relative abundance of these known hydrocarbon degradinggenera lends further evidence to the idea that community shiftsat genus level within Gammaproteobacteria class in ASSBY are
related to the presence of anthropogenic pollutants. Beside these,present study also revealed prevalence of Enterobacteriales in allpolluted samples specifically in winter samples and monsoon sug-gesting seasonal fluctuation on bacterial community structure. Fe-cal bacteria was negatively correlated to temperature suggestingthat these bacteria can persist longer at lower temperatures inmarine ecosystem (VandeWalle et al., 2012). Present study indi-cated that ASSBY is highly exposed to domestic sewage which isconsidered as one of the most significant threat for coastal environ-ments (Stabili and Cavallo, 2011).
Previous studies showed that Epsilonproteobacteria was one ofthe major marine bacterial lineages in polluted coastal ecosystemand played very important roles in the sulfur cycle (Lin et al.,2008). Grote et al. (2008) reported that Epsilonproteobacteria wasresponsible for �70% of the chemoautotrophic production in theBlack Sea, which is one of the world’s largest brackish water envi-ronments. These genera have been shown to use hydrogen sulfide,elemental sulfur and/or thiosulfate as electron donors for CO2 fixa-tion in anoxic condition. Result indicated that different genera ofEpsilonproteobacteria were also abundant in summer and winterseason of ASSBY, with high dominance in winter season. However,in monsoon samples Epsilonproteobacteria occur in high abundanceat oxic–anoxic in sediment (Wakeham et al., 2012; Gilbert et al.,2009). Epsilonproteobacteria was not recovered which indicates thatinflux of terrestrial input in sea reduces anoxic condition in mon-soon season of ASSBY. Furthermore, the present study showed thatArcobacter sp. was only present in winter sample of ASSBY whereconcentration of iron was high. Therefore, Arcobacter sp. might playimportant roles in iron metabolism. In addition to these, Arcobacteralso reported as ‘sewage indicator’ and pathogenic organisms(Newton et al., 2013), implies that eutrophication in coastal watersof ASSBY lead to loss of important bacterial phyla and build up somepathogenic bacteria which affects both the ecosystem of ASSBY andconsequently human health (White and Pickett, 1985).
Betaproteobacteria are not typically abundant in marine ecosys-tem (Zinger et al., 2011). However, present study revealed preva-lence of Betaproteobacteria in polluted samples specifically wintersamples suggested alteration in bacterial community structuredue to anthropogenic activities. Acidovorax was found to be mostdominant genus in all three seasons which suggests that Acidovo-rax has very important role in biogeochemical cycle and degrada-tion of anthropogenic pollutants. Recently, these genera werealso reported as iron oxidizing bacteria and for PAHs degradation(Singleton et al., 2009; Hedrich et al., 2011). In addition to this,Many OTUs from monsoon samples were affiliated to the Ralstonia,Alcaligenes Janthinobacterium, Masilla, Bordtella, Acidovorax, Vari-ovorax and Methylophilus, many of these were generally found interrestrial environment. In monsoon season, influx of fresh waterfrom terrestrial environment could be reason for the presence ofBetaproteobacteria in coastal water (Newton et al., 2013). Resultsalso indicated that ship breaking activities at ASSBY cause shiftof bacterial community structure toward Betaproteobacteria whichcan resist in such polluted environment.
Representatives of Alphaproteobacteria and Deltaproteobacteriawere the least diverse class of Proteobacteria found in ASSBY. OTUswere obtained from pristine library which was affiliated to Rhizob-ales which plays important role in symbiotic and asymbiotic nitro-gen fixation, production of DMS, methylhalides and EPS. FromPolluted samples OTUs were affiliated to Kordiimonas and Rhodo-bacterales. As reported earlier Kordiimonas was isolated from sedi-ment sample collected from a ship’s ballast water tank. In additionto these, OTUs from polluted samples were also affiliated to uncul-tured Deltaproteobacteria (Xu et al., 2011).
Besides Proteobacteria, Significant difference was not observeddifference in Firmicutes phylum. OTUs of Firmicutes displayeddistinct affiliation which belonged to two orders, Bacillales and

370 V. Patel et al. / Bioresource Technology 161 (2014) 362–370
Clostridiales. Most OTUs of Bacillales belonged to the Bacillus sp.from all the samples. However, One OTU was found similar withPanisarcina from only pristine sample. In other gram positivebacteria, one OTU was acquired from Actinobacteria from monsoonlibraries which was similar to Kocuria.
Cyanobacteria are also an important feature of the marine eco-system. In present study, Cyanobacteria were recovered from win-ters and monsoon season indicating the deterioration of coastwater in ASSBY due to ship breaking industries. Du et al. (2013)also reported abundance of Cyanobacteria in winter. The ecologicalsignificance of Bacteroidetes in marine ecosystem has been broughtto light because of their proficiency in degrading various biopoly-mers such as cellulose, chitin and pectin (Kirchman, 2002). TheBacteroidetes were well represented in both saline and freshwaterenvironments (Silveira et al., 2011). In this study, three OTUsclustered with Salibacter sp. and Salinimicrobium sp. were alsorecovered from other marine ecosystem.
5. Conclusion
To the best of knowledge, this is the first comparison of the bac-terial communities among different seasons in the coastal watersof the ASSBY by means of 16S rDNA sequence analysis. This com-prehensive study will enhance the overall understanding of themicrobial ecology of the ASSBY affected by ship breaking indus-tries. Strong relationships were observed between level of anthro-pogenic pollutants and bacterial community structure. Clonelibraries analysis for polluted and pristine stations have demon-strated significant differences in bacterial community composi-tions and also showed bacterial community has shifted fromGammaproteobacteria to Betaproteobacteria class due to effect ofship breaking activities.
Acknowledgements
Authors acknowledge the financial support by Department ofScience and Technology (DST) and Department of Biotechnology(DBT), New Delhi. Research fellowship awarded by Council ofScientific and Industrial Research (CSIR), New Delhi, India, to VilasPatel, is gratefully acknowledged. We are acknowledged to Dr.Varun shah (Institute of life sciences, Ahmadabad University,Ahmadabad) for valuable suggestion in manuscript preparation.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.biortech.2014.03.033.
References
Andersson, A.F., Riemann, L., Bertilsson, S., 2010. Pyrosequencing revealscontrasting seasonal dynamics of taxa within Baltic Sea bacterioplanktoncommunities. ISME J. 4, 171–181.
Basha, S., Gaur, P.M., Thorat, R.B., Trivedi, R.H., Mukhopadhyay, S.K., Anand, N.,Desai, S.H., Mody, K.H., Jha, B., 2007. Heavy metal content of suspendedparticulate matter at world’s largest ship-breaking yard, Alang–Sosiya, India.Water Air Soil Pollut. 178, 373–384.
Bertness, M.D., Gaines, S., Hay, M. (Eds.), 2001. Marine Community Ecology. SinauerAssociates Inc.
Carlson, C.A., Morris, R., Parsons, R., Treusch, A.H., Giovannoni, S.J., Vergin, K., 2009.Seasonal dynamics of SAR11populations in the euphotic and mesopelagic zonesof the northwestern Sargasso Sea. ISME J. 3, 283–295.
Desai, C., Parikh, R.Y., Vaishnav, T., Shouche, Y.S., Madamwar, D., 2009. Tracking theinfluence of long-term chromium pollution on soil bacterial communitystructures by comparative analyses of 16S rRNA gene phylotypes. Res.Microbiol. 160, 1–9.
Du, J., Xiao, K., Li, L., Ding, X., Liu, H., Lu, Y., Zhou, S., 2013. Temporal and spatialdiversity of bacterial communities in coastal waters of the South China Sea.PLoS One 8 (6), 1–13.
Feng, B.W., Li, X.R., Wang, J.H., Hu, Z.Y., Meng, H., Xiang, L.Y., 2009. Bacterialdiversity of water and sediment in the Changjiang estuary and coastal area ofthe East China Sea. FEMS Microbiol. Ecol. 70, 80–92.
Gilbert, J.A., Field, D., Swift, P., Newbold, L., Oliver, A., Smyth, T., 2009. The seasonalstructure of microbial communities in the Western English Channel. Environ.Microbiol. 11, 3132–3139.
Grote, J., Jost, G., Labrenz, M., Herdndl, G.J., Jurgens, K., 2008. Epsilonproteobacteriarepresent the major portion of chemoautotrophic bacteria in sulfidic waters ofpelagic redoxi clines of the Baltic and Black Seas. Appl. Environ. Microbiol. 74,7546–7551.
Hedrich, S., Schlomann, M., Johnson, D.B., 2011. The iron-oxidizing Proteobacteria.Microbiology 157, 1551–1564.
Kaper, J.B., Lockman, H., Colwell, R.R., 1981. Aeromonas hydrophila, ecology andtoxigenicity of isolates from estuary. J. Appl. Bacteriol. 50, 359–377.
Kirchman, D.L., 2002. The ecology of Cytophaga-Flavobacteria in aquaticenvironments. FEMS Microbiol. Ecol. 39, 91–100.
Kobayashi, T., Koide, O., Mori, K., Shimamura, S., Matsuura, T., Miura, T., Takaki, Y.,Morono, Y., Nunoura, T., Imachi, H., Inagaki, F., Takai, K., Horikoshi, K., 2008.Phylogenetic and enzymatic diversity of deep subseafloor aerobicmicroorganisms in organics- and methane-rich sediments off ShimokitaPeninsula. Extremophiles 12, 519–527.
Lin, X., Scranton, M.I., Chistoserdov, A.Y., Varela, R., Taylor, G.T., 2008.Spatiotemporal dynamics of bacterial populations in the anoxic Cariaco Basin.Limnol. Oceanogr. 53, 37–51.
Luz, A.P., Pellizari, V.H., Whyte, L.G., Greer, C.W., 2004. A survey of indigenousmicrobial hydrocarbon degradation genes insoils from Antarctica and Brazil.Can. J. Microbiol. 50 (5), 323–333.
Morris, R.M., Vergin, K.L., Cho, J.C., Rappe, M.S., Carlson, C.A., Giovannoni, S.J., 2005.Temporal and spatial response of bacterioplankton lineages to annualconvective overturn at the Bermuda Atlantic Time-series Study site. Limnol.Oceanogr. 50, 1687–1696.
Newton, R.J., Bootsma, M.J., Morrison, H.G., Sogin, M.L., McLellan, S.L., 2013. Amicrobial signature approach to identify fecal pollution in the waters off anurbanized coast of Lake Michigan. Microb. Ecol. 65, 1011–1023.
Nogales, B., Lanfranconi, M.P., Piña-Villalonga, J.M., Bosch, R., 2011. Anthropogenicperturbations in marine microbial communities. FEMS Microbiol. Rev. 35, 275–298.
Patel, V., Patel, J., Madamwar, D., 2013. Biodegradation of phenanthrene instimulated microcosm by consortium ASP isolated from Alang–Sosiya shipbreaking yard. Gujarat Mar. Pollut. Bull. 74 (1), 199–207.
Reddy, M.S., Basha, S., Joshi, H.V., Ramachandraiah, G., 2005. Seasonal distributionand contaminations levels of total PHCs, PAHs and heavy metals in coastalwaters of the Alang–Sosiya ship scrapping yard, Gulf of Cambay, India.Chemosphere 61, 1587–1593.
Reddy, S., Basha, M., Shaik, S., Kumar, V.G., Joshi, H.V., Ghosh, P.K., 2003.Quantification and classification of ship scrapping waste at Alang–Sosiya,India. Mar. Pollut. Bull. 46, 609–1614.
Sambrook, J., Russell, D.W., 2000. Molecular Cloning – A Laboratory Manual, Volume1, third ed. CSHL press, pp. 1.32–1.37.
Silveira, C.B., Vieira, R.P., Cardoso, A.M., Paranhos, R., Albano, R.M., 2011. Influence ofsalinity on bacterioplankton communities from the Brazilian rain forest to thecoastal Atlantic Ocean. PLoS One 6, 17789.
Singleton, D.R., Ramirez, L.G., Aitken, M.D., 2009. Characterization of a polycyclicaromatic hydrocarbon degradation gene cluster in a phenanthrene-degradingAcidovorax strain. Appl. Environ. Microbiol. 75 (9), 2613.
Stabili, L., Cavallo, R.A., 2011. Microbial pollution indicators and culturableheterotrophic bacteria in a Mediterranean area (Southern Adriatic Sea Italiancoasts). J. Sea Res. 65, 461–469.
Treusch, A.H., Vergin, K.L., Finlay, L.A., Donatz, M.G., Burton, R.M., 2009. Seasonalityand vertical structure of microbial communities in an ocean gyre. ISME J. 3,1148–1163.
VandeWalle, J.L., Goetz, G.W., Huse, S.M., Morrison, H.G., Sogin, M.L., Hoffmann, R.G.,Yan, K., McLellan, S.L., 2012. Aceinetobacter, Aeromonas and Trichococcuspopulations dominate the microbial community within urban sewerinfrastructure. Environ. Microbiol. 14 (9), 2538–2552.
Wakeham, S.G., Turich, C., Schubotz, F., 2012. Biomarkers, chemistry andmicrobiology show chemoautotrophy in a multilayer chemocline in theCariaco Basin. Deep Sea Res. I. 63, 133–156.
Warskow, A.L., Juni, E., 1972. Nutritional requirements of Acinetobacter strainsisolated from soil, water, and sewage. J. Bacteriol. 112, 1014–1016.
White, P.S., Pickett, S.T.A., 1985. The Ecology of Natural Disturbance and PatchDynamics. Academic Press, Orlando, FL, USA, pp. 3–13.
Xu, X.W., Huo, Y.Y., Bai, X.D., Wang, C.S., Oren, A., Li, S.Y., Wu, M., 2011. Kordiimonaslacus sp. nov., isolated from a ballast water tank, and emended description ofthe genus Kordiimonas. ISME J. 61, 422–426.
Yakimov, M.M., Timmis, K.N., Golyshin, P.N., 2007. Obligate oil-degrading marinebacteria. Curr. Opin. Biotechnol. 18, 257–266.
Zhang, R., Lau, S.C.K., Ki, J.S., Thiyagarajan, V., Qian, P.Y., 2009. Response ofbacterioplankton community structures to hydrological conditions andanthropogenic pollution in contrasting subtropical environments. FEMSMicrobiol. Ecol. 69, 449–460.
Zinger, L., Amaral-Zettler, L.A., Fuhrman, J.A., Horner-Devine, M.C., Huse, S.M., 2011.Global patterns of bacterial beta-diversity in seafloor and seawater ecosystems.PLoS One 6 (9), 24570.












