
Reproductive biology of turbot (Psetta maxima) and brill (Scophthalmus rhombus) (Teleostei,...
8
This article was downloaded by: [186.89.52.65] On: 24 March 2014, At: 16:08 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Italian Journal of Zoology Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/tizo20 Reproductive biology of turbot (Psetta maxima) and brill (Scophthalmus rhombus) (Teleostei, Pleuronectiformes) in the Adriatic Sea Vincenzo Caputo a , Giacomo Candi a , Sabrina Colella b & Enrico Arneri b a Istituto di Biologia e Genetica , Università di Ancona , via Brecce Bianche, I‐60100, Ancona, Italy b IRPEM ‐ CNR , Istituto di Ricerche sulla Pesca Marittima , largo Fiera della Pesca, I‐60125, Ancona, Italy Published online: 28 Jan 2009. To cite this article: Vincenzo Caputo , Giacomo Candi , Sabrina Colella & Enrico Arneri (2001) Reproductive biology of turbot (Psetta maxima) and brill (Scophthalmus rhombus) (Teleostei, Pleuronectiformes) in the Adriatic Sea, Italian Journal of Zoology, 68:2, 107-113, DOI: 10.1080/11250000109356393 To link to this article: http://dx.doi.org/10.1080/11250000109356393 PLEASE SCROLL DOWN FOR ARTICLE Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should be independently verified with primary sources of information. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use of the Content. This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http:// www.tandfonline.com/page/terms-and-conditions
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Reproductive biology of turbot (Psetta maxima) and brill (Scophthalmus rhombus) (Teleostei,...

This article was downloaded by: [186.89.52.65]On: 24 March 2014, At: 16:08Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: MortimerHouse, 37-41 Mortimer Street, London W1T 3JH, UK
Italian Journal of ZoologyPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/tizo20
Reproductive biology of turbot (Psetta maxima)and brill (Scophthalmus rhombus) (Teleostei,Pleuronectiformes) in the Adriatic SeaVincenzo Caputo a , Giacomo Candi a , Sabrina Colella b & Enrico Arneri ba Istituto di Biologia e Genetica , Università di Ancona , via Brecce Bianche, I‐60100,Ancona, Italyb IRPEM ‐ CNR , Istituto di Ricerche sulla Pesca Marittima , largo Fiera della Pesca,I‐60125, Ancona, ItalyPublished online: 28 Jan 2009.
To cite this article: Vincenzo Caputo , Giacomo Candi , Sabrina Colella & Enrico Arneri (2001) Reproductive biologyof turbot (Psetta maxima) and brill (Scophthalmus rhombus) (Teleostei, Pleuronectiformes) in the Adriatic Sea, ItalianJournal of Zoology, 68:2, 107-113, DOI: 10.1080/11250000109356393
To link to this article: http://dx.doi.org/10.1080/11250000109356393
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purposeof the Content. Any opinions and views expressed in this publication are the opinions and views of theauthors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content shouldnot be relied upon and should be independently verified with primary sources of information. Taylorand Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses,damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connectionwith, in relation to or arising out of the use of the Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Ital. J. Zool., 68: 107-113 (2001)
Reproductive biology of turbot(Psetta maxima) and brill(Scophthalmus rhombus) (Teleostei,Pleuronectiformes) in the Adriatic Sea
VINCENZO CAPUTOGIACOMO CANDIIstituto di Biologia e Genetica, Università di Ancona,via Brecce Bianche, I-60100 Ancona (Italy)
SABRINA COLELLAENRICO ARNERIIRPEM - CNR, Istituto di Ricerche sulla Pesca Marittima,largo Fiera della Pesca, I-60125 Ancona (Italy)
ABSTRACT
The reproductive biology of two commercially important flat-fish, turbot (Psetta maxima) and brill (Scophthalmus rhombus), inthe Adriatic Sea was investigated by histology and macroscopicanalysis of gonads. Different patterns of vitellogenesis and spawn-ing were observed in the two species. In turbot, gonad develop-ment culminates annually from March to May and the seasonalegg batch develops synchronously, the eggs becoming ready atabout the same time. In brill, the ovary contains several oocytebatches in different stages of maturity, indicating intermittent re-lease of ripe eggs in the course of a long spawning season (Janu-ary-July). These data are discussed and compared with observa-tions on reproduction of turbot and brill for other areas of theirdistribution ranges.
KEY WORDS: Reproduction - Gonad histology - Psetta maxima- Scophthalmus rhombus - Pleuronectiformes - Adriatic Sea.
ACKNOWLEDGEMENTS
This work is part of a study of the growth, feeding habits, andreproductive biology of turbot and brill funded by the EuropeanUnion (Contract MED/93/017). Dr Silvia Modena reviewed theEnglish version of the manuscript.
(Received 13 July 2000 - Accepted 3 January 2001)
INTRODUCTION
Turbot Psetta maxima (Linnaeus, 1758) and brillScophthalmus rhombus (Linnaeus, 1758) are flatfishes ofthe family Scophthalmidae that are widely distributedfrom Norway to the Mediterranean and the Black Sea(Nielsen, 1986). They inhabit sandy and muddy bottomsin the shallower part of the continental shelf down toabout 70-80 metres. In the Adriatic they have alwaysbeen highly valued commercial fishes (Faber, 1883) andare caught mainly by bottom and beam trawls but alsoby fixed gear such as trammel nets and bottom longlines. Official catch data for both species are unreliablebecause a considerable proportion of the catch is notsold on the market and because other flatfishes are fre-quently recorded as turbot (Froglia, 1993)- The annualcatch of turbot and brill in the upper and central Adriaticcan be tentatively put at around 400 tonnes.
Despite their commercial importance to Adriatic fish-eries, data on the biology of these two species in thisarea of the Mediterranean are scanty. Reproductive biol-ogy, providing information on species life history, iscritically important in fishery science, and useful to fish-eries management. Fish reproduction is generally inves-tigated by studying gonad maturity on the basis ofmacroscopic characteristics, gonado-somatic index, andhistology. The first two methods are gross indicators ofreproductive biology while histology allows the pro-cesses of gonadal development and maturation to bestudied in details (Hunter et al., 1992). The presentstudy was undertaken to describe these aspects in orderto increase current knowledge on the reproductivestrategies of turbot and brill in the Mediterranean Sea.
MATERIALS AND METHODS
From November 1994 to July 1996, 155 turbot (size range 13 to79 cm in total length, TL) and 149 brill (size range 15 to 49 cmTL) were sampled in the central Adriatic from the landings of thecommercial fishing fleet of Ancona (Italy). These two species arenot constantly present in the landings and the price for large indi-viduals can be very high; therefore the sampling was opportunis-tic, with sample size varying from few specimens (in months withnearly no landings) to about 30 individuals per species per monthin months with substantial landings. For each fish sampled, TL (tothe nearest 1 mm), total weight (to the nearest 1 g for fishes up to2500 g, and to 50 g for larger fishes), and sex were recorded; allmeasurements were taken after capture, on chilled individuals.The maturity stage of the gonads was determined at first macro-scopically following the eight-point scale for total spawners(Holden & Raitt, 1974). Weight of ovaries was recorded to an ac-curacy of 1 g and the gonado-somatic index (GSI = gonad weight/ body weight x 100) was calculated excluding immature individu-als (Kartas & Quignard, 1984), which do not show substantialvariation in the weight of gonads during the year.
Because females of these two species present a uniform struc-ture of the cystovary, a fragment of the gonad from 35 female brilland 22 female turbot, collected in different months, was fixed in10% neutral formalin or in Bouin's fluid and embedded in Para-plast, to investigate seasonal changes in gonad morphology; 6-umsections stained with Mayer's haematoxylin-eosin and Galgano'strichrome were prepared for histological examination (Beccari &
Dow
nloa
ded
by [
186.
89.5
2.65
] at
16:
08 2
4 M
arch
201
4

108 V. CAPUTO, G. CANDI, S. COLELLA, E. ARNERI
Mazzi, 1972). Ovarian follicles were classified into five stages (I =early perinucleolar stage; II = late perinucleolar stage; III = yolkvesicles stage; IV = vitellogenic stage; V = mature stage), accord-ing to Wallace & Selman (1981) and West (1990).
Ovarian activity was assessed by counting the ovarian folliclesat different stages on five different sections, 1 mm apart, of eachovary. The presence of atretic follicles was also recorded. In 20oocytes for each stage of development, the cellular and nucleardiameters were measured and the nucleo-plasmatic index wascalculated (NPI = nucleus diameter / cytoplasm diameter). The re-sults are expressed as means ± SE.
The lobular testes of 24 male brill and 18 male turbot were re-moved and fixed in 10% neutral formalin or in Bouin's fluid, andparaffin sections were prepared as described for the ovaries. Sper-matogenic activity was assessed by measuring the relative surfaceoccupied by different types of gametocytes (from primary spermato-cytes to spermatozoa) in 10 seminiferous lobules randomly selectedfrom three different sections, 50 um apart, of each testis. Presenceor absence of sperms in the sperm duct was also recorded. Sectionswere observed with a Leica Diaplan photomicroscope. Measure-ments were performed with the Optimas videoanalysis package.
In ripe females of turbot, the number of mature ova was count-ed in a 0.5-g fragment of the ovary to obtain an estimate of thetotal fecundity (gravimetric method; Holden & Raitt, 1974).
RESULTS
The morphometric features that characterize the stagesof ovarian development for the two species are summa-
rized in Table I; Figures 1, 4 show the seasonal varia-tions observed in ovarian and spermatogenic activities.
Scophthalmus rhombus
At the end of September and in October, some ovarieshave no cytoplasmic inclusions in oocytes; these arenevertheless developing, because the nucleoli have mul-tiplied and migrated to the periphery of the nucleus(stages I and II) (Figs 1, 2A; Table I). Other individualsalso display larger oocytes with yolk vesicles in the cyto-plasm (stage III) (Figs 1, 2B; Table I). At this stage, thezona radiata begins to form and lampbrush chromo-somes can be seen in the nucleus.
In December three oocyte batches are evident (Fig.1): one consists of large oocytes filled with yolk gran-ules (stage IV) for the winter spawning; one of oocytesin stage III which will mature for the spring spawning;and one is a batch of oocytes in late stage II, for thesummer spawning (Fig. 2C; Table I).
The first spawning takes place in January and Febru-ary: oocytes in late vitellogenesis grow to a diameter ofabout 900 mm and the yolk granules become indistinctso that the yolk melts into a homogeneous, transparentmass; due to resumption of meiosis, the nucleus is notvisible (stage V) (Figs 1, 2D; Table I).
TABLE I - Morphometric characteristics identifying oocyte stages (mean ± SE, range). NPI, nucleo-plasmatic index.
Stage Oocyte diameter (mm) Nucleus diameter (mm) NPI
Scophthalmus rhombus
Early stage II
Late stage II
Stage III
Early stage IV
Late stage IV
Stage V
Psetta maxima
Early stage II
Late stage II
Stage III
Late stage IV
Stage V
58.18 ± 2.33(47.07-68.27)
73.94 ± 4.14(52.04-96.85)
131.49 ± 6.41(103.14-156.81)
165.13 ± 7.29(128.74-221.23)
568.70 ± 28.66(425.94-693.58)
889-17 ± 12.53(814.75-989.3D
47.17 ± 2.75(37.72-65.21)
76.63 ± 3.39(60.32-90.30)
143.19 ± 5.04(124.51-168.53)
397.89 ± 13.84(315.53-460.14)
724.97 ± 12.05(664.89-794.71)
27.97 ± 1.07(22.98-33.40)
30.87 ± 1.97(22.93-40.35)
52.86 ± 3-14(36.80-67.55)
67.52 ± 3.41(49.80-87.99)
127.80 + 5.65(106.42-171.46)
23.08 + 1.41(16.90-31.46)
38.74 ± 2.59(27.27-49.43)
71.97 ± 3-30(60.31-86.86)
88.91 ± 3-88(74-114.56)
48.52 ± 2.15(40.62-59.57)
41.79 + 1.35(34.63-49.71)
40.10 ± 1.05(31.94-43.89)
41.26 ± 2.14(29.74-53.95)
2311 ± 1.66(15.48-31.02)
50.04 ± 1.86(41.40-59.81)
50.45 + 2.25(36.05-59.45)
48.12 ± 1.72(38.42-53.18)
22.39 ± 0.73(19-28-28.04)
Dow
nloa
ded
by [
186.
89.5
2.65
] at
16:
08 2
4 M
arch
201
4
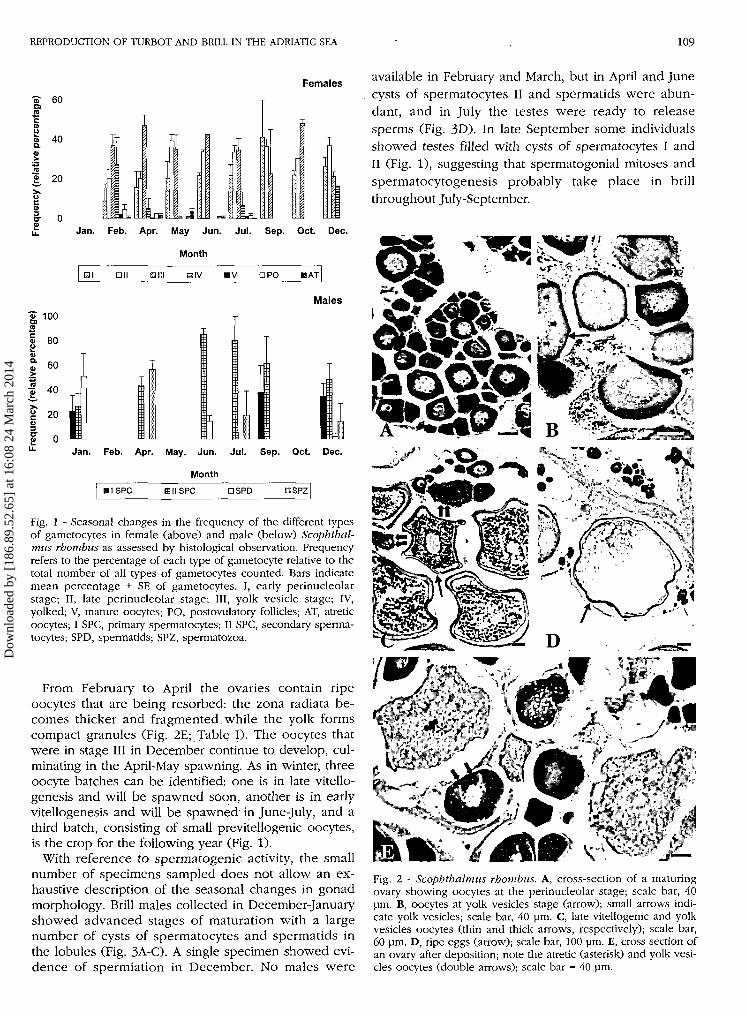
REPRODUCTION OF TURBOT AND BRILL IN THE ADRIATIC SEA 109
Females
a60
40
2 20
Jan. Feb. Apr. May Jun. Jul. Sep. Oct. Dec.
Month
01 Dll 0111 BIV «V QPO HAT
Males
Jan. Feb. Apr. May. Jun. Jul. Sep. Oct. Dec.
Month
• ISPC HII SPC DSPD ESSPZ
Fig. 1 - Seasonal changes in the frequency of the different typesof gametocytes in female (above) and male (below) Scophthal-mus rhombus as assessed by histological observation. Frequencyrefers to the percentage of each type of gametocyte relative to thetotal number of all types of gametocytes counted. Bars indicatemean percentage + SE of gametocytes. I, early perinucleolarstage; II, late perinucleolar stage; III, yolk vesicle stage; IV,yolked; V, mature oocytes; PO, postovulatory follicles; AT, atreticoocytes; I SPC, primary spermatocytes; II SPC, secondary sperma-tocytes; SPD, spermatids; SPZ, spermatozoa.
From February to April the ovaries contain ripeoocytes that are being resorbed: the zona radiata be-comes thicker and fragmented while the yolk formscompact granules (Fig. 2E; Table I). The oocytes thatwere in stage III in December continue to develop, cul-minating in the April-May spawning. As in winter, threeoocyte batches can be identified: one is in late vitello-genesis and will be spawned soon, another is in earlyvitellogenesis and will be spawned in June-July, and athird batch, consisting of small previtellogenic oocytes,is the crop for the following year (Fig. 1).
With reference to spermatogenic activity, the smallnumber of specimens sampled does not allow an ex-haustive description of the seasonal changes in gonadmorphology. Brill males collected in December-Januaryshowed advanced stages of maturation with a largenumber of cysts of spermatocytes and spermatids inthe lobules (Fig. 3A-C). A single specimen showed evi-dence of spermiation in December. No males were
available in February and March, but in April and Junecysts of spermatocytes II and spermatids were abun-dant, and in July the testes were ready to releasesperms (Fig. 3D). In late September some individualsshowed testes filled with cysts of spermatocytes I andII (Fig. 1), suggesting that spermatogonial mitoses andspermatocytogenesis probably take place in brillthroughout July-September.
Fig. 2 - Scophthalmus rhombus. A, cross-section of a maturingovary showing oocytes at the perinucleolar stage; scale bar, 40pm. B, oocytes at yolk vesicles stage (arrow); small arrows indi-cate yolk vesicles; scale bar, 40 pm. C, late vitellogenic and yolkvesicles oocytes (thin and thick arrows, respectively); scale bar,60 pm. D, ripe eggs (arrow); scale bar, 100 pm. E, cross section ofan ovary after deposition; note the atretic (asterisk) and yolk vesi-cles oocytes (double arrows); scale bar = 40 pm.
Dow
nloa
ded
by [
186.
89.5
2.65
] at
16:
08 2
4 M
arch
201
4

110 V. CAPUTO, G. CANDI, S. COLELLA, E. ARNERI
J»,
€• •
% •
"**!
-$ *
f f
*l*''
•&,'
.1 -,
.'N
m j.
< *,•*»
; * *•« * • - •#» . , -.•« *
wm^MFig. 3 - Scophthalmus rhombus. A, spermatocyst filled with primary spermatocytes; scale bar, 10 um. B, spermatocyst filled with sec-ondary spermatocytes; scale bar, 10 pm. C, spermatocyst full of spermatids (SPD) surrounded by spermatozoa (SPZ) free in the lobulelumen; scale bar, 30 pm. D, cross section of a sperm duct filled with sperms (arrows); scale bar, 100 pm.
Dow
nloa
ded
by [
186.
89.5
2.65
] at
16:
08 2
4 M
arch
201
4

REPRODUCTION OF TURBOT AND BRILL IN THE ADRIATIC SEA 111
The smallest mature female recorded by macroscopicgonad analysis was 37 cm TL. This finding was support-ed by histological data where females of 40 cm inlength and about 1000 g in weight showed oocytes inactive vitellogenesis whereas in females weighing up toabout 200 g and 25 cm long, only ovaries with oocytesin previtellogenic stage (stage I-III) were observed. Onthe contrary, small males (TL from 23 to 28 cm andweight from 160 to 300 g) showed active spermatogene-sis in December and late January: cysts prevalently filledwith spermatocytes I in December, and with spermato-cytes II, spermatids, and spermatozoa in late January. Inbrill females, due to the limited number of samples permonth, the GSI did not produce any significant result.
Psetta maxima
In turbot the ovaries rest from June to November andcontain small chromophilic oocytes devoid of cytoplas-mic inclusions (early stage II) (Fig. 4). Oocyte matura-tion starts in December (stages III and IV) and peaks(stage V) between March and May. Vitellogenesis seemsto take place in one batch of oocytes at a time, as in-
Females
8, 8°n
S 60
» 40 ^
1£. 20
s-o 0
£
Jan. Feb. Mar. Apr. May Jun. Sep. Nov. Dec.
Month
• i Dll BIV • V • PO BAT
Males
0)
c100
80 -
60 -
40
20 -\c<u3 0
Jan. Feb. Mar. Apr. May Jun. Sep. Nov. Dec.
Month
• ISPC HIISPC DSPD 0SPZ
Fig. 4 - Seasonal changes in the frequency of the different typesof gametocytes in female (above) and male (below) Psetta maxi-ma as assessed by histological observation. Frequency refers tothe percentage of each type of gametocyte relative to the totalnumber of all types of gametocytes counted. Bars indicate meanpercentage + SE of gametocytes. Legends as in Figure 1.
ferred from the presence in the ovary of two distinctgroups of oocytes before the spawning season, oneconsisting of small previtellogenic oocytes and the otherof large yolked oocytes: i.e. the current season's crop(Figs 4, 5A).
With reference to spermatogenic activity, the smallnumber of specimens of male turbot collected fromFebruary to May showed evident signs of spermiation(sperm duct filled with sperms) (Figs 4, 5B).
Gonado-somatic index values were obtained from 56female turbot (size range 45 to 79 cm, in order to ex-clude immature individuals). Although based on a limit-ed number of observations, GSI in turbot showed a peakin March and April with average values of about 11,compared to values from 1 to 3 for the rest of the year.This suggests that these months are the main spawningseason and thus it confirms the histological data.
The smallest macroscopically observed ripe femaleturbot was 50 cm in TL. Histological data supported thisfinding: in winter months (when the ovaries of largerfemales are in full vitellogenesis), female P. maxima of700-1000 g and 34-40 cm in length, showed only pre-vitellogenic oocytes (stage I). For males, macroscopicanalysis is rather uncertain and sexual maturity is appar-ently reached at 40-45 cm TL.
From 23 ripe females (ranging from 50 to 77 cm inTL) an estimate of total fecundity was obtained: itranged from 2xl06 to l6xl06 eggs per female. A linearregression between body weight and fecundity, fitted tothe experimental data, is shown in Figure 6.
DISCUSSION
The present investigation shows a different pattern ofvitellogenesis and spawning in turbot and brill. In theformer, gonad development culminates annually, fromMarch to May, in a relatively short spawning season:the whole seasonal batch of eggs develops synchro-nously and all the eggs to be spawned in a season areripe at about the same time. Towards the spawning sea-son, two clearly distinct groups of oocytes are recog-nized in the ovary by their stage of development: oneconsists of small previtellogenic oocytes, which may bethe recruitment stock for the following year, the other ismade up of large yolked oocytes for the current sea-son's crop. A different pattern was identified in brill,where the ovary contains several oocyte batches in dif-ferent stages of maturity, indicating many intermittentreleases of ripe eggs over a long spawning season (Jan-uary-July). Further histological evidence of prolongedspawning is provided by the presence of atretic ripeeggs from February to July in ovaries containing manydeveloping oocytes. This has been observed also inother multiple spawner pleuronectiforms, e.g. somesoleids, where batches of oocytes of different size arereleased in different waves (Deniel, 1984).
The spawning period of turbot in the Adriatic extendsfrom March to May and is therefore about two months
Dow
nloa
ded
by [
186.
89.5
2.65
] at
16:
08 2
4 M
arch
201
4

112 V. CAPUTO, G. CANDI, S. COLELLA, E. ARNERI
Fig. 5 - Psetta maxima. A, cross section of the ovary showing late vitellogenic and early perinucleolar oocytes (thin and thick arrows,respectively); scale bar, 80 yim. B, cross section of the sperm duct filled with sperms (arrows); scale bar, 100 pm.
earlier than spawning in the North Sea (Rae & Devlin,1972; Jones, 1974) and the coast of Brittany (Lahaye,1972). The spawning period of brill extends from Jan-uary to July according to our data, and from May to Ju-ly in Brittany according to Lahaye (1972). This is notsurprising, since geographic differences in timing of re-production have been observed in many fish species:they frequently occur in latitudinal clines (Leggett &Carscadden, 1978) and are apparently controlled bytemperature and photoperiod (Nash, 1999).
Our data indicate that in the Adriatic Sea, turbot is asingle spawner, in agreement with similar observationsfor the North Atlantic (Lahaye, 1972; Jones, 1974; De-niel, 1982; Mugnier, 1990). Also our fecundity data, al-though based on a limited number of specimens, aresubstantially in line with those of Jones (1974) for At-lantic turbot. However, the present findings on Mediter-ranean brill point to multiple spawning with a protract-ed deposition period, at variance with the histologicaldata reported by Lahaye (1972) and Deniel (1982) forAtlantic brill, which seem to indicate these populationsas single spawners.
Most fish species show geographic variations in batchdeposition and fecundity. This has led some Authors toattribute most of them to adaptation by individual spec-imens to such proximal factors as water temperatureand food supply (Angelini & Ghiara, 1984; Minami &Tanaka, 1992). At higher latitudes, for example, the lim-itation of productivity to short periods of the year can-not support a long season of repeated spawning. Infact, Fox (1978) found that Coitus gobio spawns once a
year in northern, low-productive streams, whereas inthe relatively warm, highly-productive, southern chalkstreams, it exhibits multiple spawning within a-season*Wootton (1977) observed that Gasterosteus aculeatuscan increase the number of egg batches produced inone season in response to increased food levels. Ac-cording to paleohistorical hypotheses (Borsa et ah,1987; Berrebi P., 1988, unpubl. Thesis, Univ. MontpellierII; Renaud et ah, 1990) and genetic data (Blanquer etah, 1992), the Mediterranean populations of both brilland turbot originated rather recently from parent At-lantic populations and therefore they probably under-went the same intensity of climatic events and environ-ment constraints. Multiple spawning as a fine adaptationto the Mediterranean environment might thus be, inbrill, a consequence of a higher degree of plasticity inreproductive traits rather than caused by selectiveprocesses.
With reference to the onset of first sexual maturity, theavailable data did not allow us to calculate a proper sizeat first maturity for the two species. Due to the lownumber of specimens, our data do not allow us to drawdefinitive conclusions; the results must be taken withcaution and considered as a first attempt to overcomethe absolute lack of data for these two important speciesin the Mediterranean. Males of both brill and turbotreach sexual maturity at a smaller size than females.During the spawning season, brill males mature earlierthan females, as has been observed in other teleosts(Brown-Peterson et ah, 1988; Burton, 1998). As regardsfemales, in the early winter season young brills weigh-
Dow
nloa
ded
by [
186.
89.5
2.65
] at
16:
08 2
4 M
arch
201
4

REPRODUCTION OF TURBOT AND BRILL IN THE ADRIATIC SEA 113
18
£ 14oe•2 10
6 -•aa
y= 1907.6 x - 2905216 ( n = 23; r = 0.818)
2000 4000 6000
Body weight (g)
8000
Fig. 6 - Relationships between body weight and total fecundity infemales of Psetta maxima.
ing about 200 g and of 25 cm TL (one year old, Arneri etal., 2001) show yolk vesicles in oocytes which remain inthis stage in the following months. Probably the en-docrine supply is still insufficient to allow the achieve-ment of sexual maturity during their first year of life.Lastly, there is evidence that turbot in the Adriatic ma-ture more or less at the same size as in the North Sea(Jones, 1974), but, according to different growth rates(Arneri et al., 2001), this size is reached on average oneyear earlier, thus suggesting that maturity in turbot maybe size-dependent rather than age-dependent.
REFERENCES
Angelini F., Ghiara G., 1984 - Reproductive modes and strategiesin vertebrate evolution. Boll. Zool., 51: 121-203.
Arneri E., Colella S. and Giannetti G., 2001 - Age determinationand growth of turbot and brill in the Adriatic Sea: reversal ofthe seasonal pattern of otolith zone formation. J. appl. Ichthy-ol., in press.
Beccari N., Mazzi V., 1972 - Manuale di tecnica microscopica. Val-lardi, Milano, 366 pp.
Blanquer A., Alayse J.-P., Berrada-Rkhami O., Berrebi P., 1992 -Allozyme variation in turbot (Psetta maxima) and brill (Scoph-thalmus rhombus) (Osteichthyes, Pleuronectiformes, Scoph-thalmidae) throughout their range in Europe. J. Fish Biol., 41:725-736.
Borsa P., Berrebi P., Blanquer A., 1987 - Mécanisme de la forma-tion en Méditerranée des sous-espèces du flet Platichthys flesusL. (poisson plat). In: Actes Coll. nat. CNRS "Biologie des popu-lations". Université Claude Bernard, Lyon, pp. 472-481.
Brown-Peterson N., Thomas P., Arnold C. R., 1988 - Reproductionbiology of the spotted seatrout, Cynoscion nebulosus, in southTexas. Fish. Bull., 86: 373-388.
Burton M., 1998 - Gametogenesis in north west Atlantic teleosts.Ital. J. Zool, 65 (suppl.): 199-202.
Deniel C, 1982 - Les poissons plats (Téléostéens-Pleuronecti-
formes) en baie de Douarnenez. Reproduction, croissance etmigration des Bothidae, Scophthalmidae Pleuronectidae etSoleidae. La Peche maritime, 1248: 155-160.
Deniel C, 1984 - La reproduction des poissons plats (Téléostéens-Pleuronectiformes) en baie de Douarnenez. II. Cycles sexuelset fécondité des Soleidae: Solea vulgaris vulgaris, Solea lascaris,Buglossidium luteum et Microchirus variegatus. Cah. Biol. mar.,25: 257-285.
Faber G. L., 1883 - The fisheries of the Adriatic and the fish there-of. Bernard Quaritch, London, 292 pp.
Fox P. J., 1978 - Preliminary observations on different reproduc-tion strategies in the bull-head (Cottus gobio L.) in northern andsouthern England. J. Fish Biol., 12: 5-11.
Froglia C, 1993 - Indagine biologica sulle variazioni dei quantita-tivi comercializzati presso alcuni Mercati Ittici all'ingrosso del-1'Alto Adriatico in relazione all'attuazione del fermo tempora-neo di pesca a strascico. Ministero della Marina Mercantile, Ro-ma, 137 pp.
Holden M. J., Raitt D. F. S., 1974 - Manual of fisheries science.Part 2 - Methods of resource investigation and their application.FAO Fish. tech. Pap., 115: 1-214.
Hunter J. R., Macewicz D. J., Chyan-huel Lo N., Kimbrel C. A.,1992 - Fecundity, spawning, and maturity of female Dover soleMicrostomus pacificus, with an evaluation of assumption andprecision. Fish. Bull., 90: 101-128.
Jones A., 1974 - Sexual maturity, fecundity and growth of the tur-bot Scophthalmus maximus L. J. mar. biol. Assoc. UK, 54: 109-205.
Kartas F., Quignard J.-P., 1984 - La fécondité des poissonstéléostéens. Masson, Paris, 121 pp.
Lahaye J., 1972 - Cycles sexuels de quelques poissons plats descôtes bretonnes. Rev. Trav. Inst. Peches marit., 36: 191-207.
Leggett W. C, Carscadden J. E., 1978 - Latitudinal variation in re-productive characteristics of American shad (Alosa sapidissimd):evidence for population specific life history strategies in fish. J.Fish. Res. Board. Can., 35: 1469-1478.
Minami T., Tanaka M., 1992 - Life history cycles in flatfish fromthe northwestern Pacific, with particular reference to their earlylife histories. Neth. J. Sea Res., 29: 35-48.
Mugnier C, 1990 - Etude des gametogeneses male et femelle duturbot Scopthalmus maximus L. en captivité et en photoperiodenaturelle. Mem. DEA (Oceanol. biol. et environ, mar.: Exploit,des Syst. aquat.), Brest, 27 pp.
Nash J. P., 1999 - Seasonal reproduction, fish. In: E. Knobil & J. P.Neill (eds), Encyclopedia of reproduction, vol 4. AcademicPress, San Diego, London, Boston, New York, Sydney, Tokyo,Toronto, pp. 329-340.
Nielsen J. G., 1986 - Scophtalmidae. In: P. J. P. Whithead, M. L.Bauchot, J.-C. Hureau & E. Tortonese (eds), Fishes of theNorth-eastern Atlantic and Mediterranean. Unesco, Paris, pp.1287-1293.
Rae B. B., Devlin B. E., 1972 - The turbot, its fishery and biologyin the Scottish area. Mar. Res., 1: 1-27.
Renaud F., Blanquer A., Gabrion C, 1990 - Genetic divergence inBothriocephalus gregarius: a hypothesis based on the paleo-geographic movements of their teleost (Psetta) host. Int. J. Par-asitol., 16: 553-558.
Wallace R. A., Selman K., 1981 - Cellular and dynamic aspects ofoocyte growth in teleosts. Am. Zool., 21: 325-343.
West G., 1990 - Methods of assessing ovarian development infishes: a review. Aust. J. mar. freshwater Res., 41: 199-222.
Wootton R. J., 1977 - Effect of food limitation during the breedingseason on the size, body components and egg production offemale sticklebacks (Gasterosteus aculeatus). J. anim. Ecol., 46:823-834.
Dow
nloa
ded
by [
186.
89.5
2.65
] at
16:
08 2
4 M
arch
201
4




















