
Repeated swim stress increases pain-induced expression of c-Fos in the rat lumbar cord
10
Brain Research 965 (2003) 259–268 www.elsevier.com / locate / brainres Research report Repeated swim stress increases pain-induced expression of c-Fos in the rat lumbar cord a a a a Luis Quintero , Maria Cecilia Cuesta , Jose Antonio Silva , Jose Luis Arcaya , b c a, * Lorena Pinerua-Suhaibar , William Maixner , Heberto Suarez-Roca a Section of Pharmacology, Institute of Clinical Investigation, School of Medicine, University of Zulia, Apartado Postal 1151, Maracaibo 4001- A, Venezuela b Fundacite-Zulia, Maracaibo, Venezuela c Dental Research Center, School of Dentistry, University of North Carolina, Chapel Hill, NC 27599-7455, USA Accepted 13 December 2002 Abstract We have previously demonstrated that repeated swim stress produces a long-lasting cutaneous hyperalgesia in rats.We have now looked at c-Fos expression in the spinal lumbar cord of male Sprague–Dawley rats subjected to 10–20 min daily sessions of forced swimming for 3 consecutive days. Control rats were subjected to sham swimming or were completely naive. Forty-eight hours later, nociception was assessed by recording for 90 min the nociceptive behavior evoked the injection of 1% formalin in the hind paw. Thirty min later, the rats’ spinal cords were removed for c-Fos immunocytochemistry. Total pain scores were 45% higher in swim stressed rats compared to control animals due an increased nociceptive behavior during last 70 min of the recording period. In addition, the number of c-Fos- immunoreactive nuclei was 40% higher in the lumbar ipsilateral dorsal horn (L4–L5) of swim stressed rats than in controls, being the highest relative increase, relative to the control groups, observed in laminae III–IV, followed by laminae V–VI, with the smallest ¨ difference in laminae I–II. c-Fos expression in the contralateral dorsal horn was higher in swim stressed rats than in sham and naıve rats. In the absence of a nociceptive stimulus, a low level of c-Fos expression was observed mainly in laminae I, II,V, and VI, being higher in swim stressed rats than in sham rats. These findings suggest that repeated inescapable and uncontrollable stress could induce a sensitization and activation of sensory neurons at the spinal level. 2002 Elsevier Science B.V. All rights reserved. Theme: Sensory systems Topic: Pain modulation: anatomy and physiology Keywords: Hyperalgesia; Stress; c-fos; Learned helplessness; Formalin test; Pain; Forced swimming 1. Introduction oscillations or exposure to ether vapors [11], inescapable holding or exposure to a novel environment [41], 15-min Under some experimental conditions, stressful environ- restraint / vibration [13], and partial restraint [15]. ments can elicit hyperalgesia instead of analgesia [17]. A more long-lasting hyperalgesia has been observed Frequently, this effect of stress on nociceptive behaviors following chronic stress. Repeated exposure to a cold has been misinterpreted as tolerance to the effect of a environment (4 8C for 30 min every hour for 1 day) repeated stressor [22] or even overlooked despite of its induces 3-day long mechanical hyperalgesia [34]. One- clinical significance. An immediate and transient (2-min hour restraint a day for 40 days produces thermal hy- long) thermal or mechanical hyperalgesia has been ob- peralgesia [9]. Finally, repeated non-noxious swim-stress served after brief stress situations, such as, horizontal (10–20 min a day for 3 days) elicits a delayed (after 24–48 h) and long-lasting (8–9 days) thermal and chemical cutaneous hyperalgesia [30]. *Corresponding author. Tel.: 158-261-793-1859; fax: 158-261-759- Although some of these studies have looked at the 8618. E-mail address: [email protected] (H. Suarez-Roca). mechanisms involved in this increased pain sensitivity, 0006-8993 / 02 / $ – see front matter 2002 Elsevier Science B.V. All rights reserved. doi:10.1016 / S0006-8993(02)04224-5
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Repeated swim stress increases pain-induced expression of c-Fos in the rat lumbar cord

Brain Research 965 (2003) 259–268www.elsevier.com/ locate/brainres
Research report
R epeated swim stress increases pain-induced expression of c-Fos inthe rat lumbar cord
a a a aLuis Quintero , Maria Cecilia Cuesta , Jose Antonio Silva , Jose Luis Arcaya ,b c a ,*Lorena Pinerua-Suhaibar , William Maixner , Heberto Suarez-Roca
aSection of Pharmacology, Institute of Clinical Investigation, School of Medicine, University of Zulia, Apartado Postal 1151, Maracaibo 4001-A,Venezuela
bFundacite-Zulia, Maracaibo, VenezuelacDental Research Center, School of Dentistry, University of North Carolina, Chapel Hill, NC 27599-7455,USA
Accepted 13 December 2002
Abstract
We have previously demonstrated that repeated swim stress produces a long-lasting cutaneous hyperalgesia in rats. We have now lookedat c-Fos expression in the spinal lumbar cord of male Sprague–Dawley rats subjected to 10–20 min daily sessions of forced swimming for3 consecutive days. Control rats were subjected to sham swimming or were completely naive. Forty-eight hours later, nociception wasassessed by recording for 90 min the nociceptive behavior evoked the injection of 1% formalin in the hind paw. Thirty min later, the rats’spinal cords were removed for c-Fos immunocytochemistry. Total pain scores were 45% higher in swim stressed rats compared to controlanimals due an increased nociceptive behavior during last 70 min of the recording period. In addition, the number of c-Fos-immunoreactive nuclei was 40% higher in the lumbar ipsilateral dorsal horn (L4–L5) of swim stressed rats than in controls, being thehighest relative increase, relative to the control groups, observed in laminae III–IV, followed by laminae V–VI, with the smallest
¨difference in laminae I–II. c-Fos expression in the contralateral dorsal horn was higher in swim stressed rats than in sham and naıve rats.In the absence of a nociceptive stimulus, a low level of c-Fos expression was observed mainly in laminae I, II, V, and VI, being higher inswim stressed rats than in sham rats. These findings suggest that repeated inescapable and uncontrollable stress could induce asensitization and activation of sensory neurons at the spinal level. 2002 Elsevier Science B.V. All rights reserved.
Theme: Sensory systems
Topic: Pain modulation: anatomy and physiology
Keywords: Hyperalgesia; Stress; c-fos; Learned helplessness; Formalin test; Pain; Forced swimming
1 . Introduction oscillations or exposure to ether vapors [11], inescapableholding or exposure to a novel environment [41], 15-min
Under some experimental conditions, stressful environ- restraint /vibration [13], and partial restraint [15].ments can elicit hyperalgesia instead of analgesia [17]. A more long-lasting hyperalgesia has been observedFrequently, this effect of stress on nociceptive behaviors following chronic stress. Repeated exposure to a coldhas been misinterpreted as tolerance to the effect of a environment (48C for 30 min every hour for 1 day)repeated stressor [22] or even overlooked despite of its induces 3-day long mechanical hyperalgesia [34]. One-clinical significance. An immediate and transient (2-min hour restraint a day for 40 days produces thermal hy-long) thermal or mechanical hyperalgesia has been ob- peralgesia [9]. Finally, repeated non-noxious swim-stressserved after brief stress situations, such as, horizontal (10–20 min a day for 3 days) elicits a delayed (after 24–48
h) and long-lasting (8–9 days) thermal and chemicalcutaneous hyperalgesia [30].*Corresponding author. Tel.:158-261-793-1859; fax:158-261-759-
Although some of these studies have looked at the8618.E-mail address: [email protected](H. Suarez-Roca). mechanisms involved in this increased pain sensitivity,
0006-8993/02/$ – see front matter 2002 Elsevier Science B.V. All rights reserved.doi:10.1016/S0006-8993(02)04224-5

260 L. Quintero et al. / Brain Research 965 (2003) 259–268
using anatomical lesions and pharmacological approaches, behaviors were immediately observed for the first 30 minnone have examined the spinal neuronal substrates that (intervals 1–10) and for two additional observation periodssupport the behavioral findings. In the present study, we of 15 min each (intervals 11–15 and 16–20) spaced bydetermined if the long-term cutaneous hyperalgesia in- two periods of 15 min without observation. Pain behaviorduced by repeated forced swim stress is associated with an was recorded at 180-s intervals, using a rating scale, asenhanced pain-induced activation of spinal cord neurons. described by Dubuisson and Dennis [8]. Briefly, 05weightWe used the expression of c-fos proto-oncogene, as is evenly distributed among all paws when the rat is still ormeasured by Fos protein expression, as a marker of in locomotion; 15little or no weight is placed upon thenociceptive-induced neuronal activity in the spinal cord injected paw; 25injected paw is elevated; 35injected paw[12]. is licked, bitten, or shaken. Numerical ratings were calcu-
lated from the following formula: pain rating5(T112T213T3) /180, where T1, T2, and T3 were the duration (in
2 . Methods seconds) spent in categories 1, 2 or 3, respectively, duringeach 180-s observation interval. An observer unaware of
2 .1. Animals the rat’s condition did a computer-assisted scoring of thenociceptive behavior. We also harvested spinal cords from
¨Forty male Sprague–Dawley rats weighing 150–300 g three additional groups; naıve rats, sham swim and forced(University of Los Andes Breeding Center, Merida, Ven- swim rats, which were not injected with formalin (withoutezuela) were used in this study. The rats were individually pain stimulation).caged 3 days prior to testing since this type of housingenhances the long-term disturbance produced by repeated2 .3. c-Fos immunocytochemistrystress [33]. The rats were kept in the same room where thebehavioral procedures were done 3 days before the begin- Two hours after the injection of formalin, the animalsning of the experimental protocol to reduce the effects of were anesthetized with CO and perfused intracardially2
environmental novelty [4]. Procedures were performed with cold PBS, followed by 4% paraformaldehyde. Thebetween 08:00 and 15:00 h. Animals were kept at a room lumbar spinal cords were removed, post-fixed in 4%temperature of 24–268C and food and water were avail- paraformaldehyde for 2 h and cryo-protected by soakingable ad libitum. The experimental protocol was approved them in 30% sucrose overnight. Cryostat-coronal sectionsby the Ethical Committee for Scientific Research on (40mm) of the lumbar enlargement of spinal cord wereAnimals at the University of Zulia. cut, being every third section collected and processed by a
free-floating slice immunocytochemistry procedure. Sec-2 .2. Experimental protocol tions were rinsed with Tris-buffered saline with Triton-X
(TBS/TX: 0.05 M Tris, 2.7% NaCl, 0.3% Triton-X, pHOn day 1, rats were subjected to a forced swim 7.6) two times for 5 min and two times for 10 min. Then,
procedure for 10 min by placing them in a plastic cylinder they were incubated in 0.3% hydrogen peroxide for 30 min(diameter 30 cm, height 50 cm) that contained 20 cm of followed by three 10-min washes with TBS/TX andwater at 24–268C. On days 2 and 3 of testing, rats were incubation with a blocking solution of 4% normal goatsubjected to a similar swim stress for 20 min. During the serum diluted in TBS/TX (NGS; Vector Labs, USA) for 60swimming sessions, the ‘immobility’ and ‘escape’ times min at room temperature. The sections were then incubatedwere recorded as previously described [28]. Immobility with polyclonal anti-c-Fos antiserum raised from rabbitoccurred when the rat made bodily movements just suffi- (Oncogene, USA) diluted 1:2000 in NGS. After a 48-hcient to keep its head above the water. Escape occurred incubation at 48C, sections are rinsed four times withwhen the rat was actively diving, jumping, or vigorously TBS/TX, and treated with NGS for 30 min as an addition-moving all four limbs to break the surface of the water or al blocking step. Sections were then incubated for 2 h withwhen it attempted to climb the wall of the container with a biotinylated goat-anti-rabbit IgG (Vector Labs) diluteits forelimbs. Control rats were subjected to a sham 1:500 in NGS. After additional rinses in TBS/TX, allswimming session by allowing them to wade in a similar sections are incubated in streptavidin-HRP (ABC Elite;cylinder that contained only 2–3 cm of water at 24–268C. Vector; diluted 1:100 in buffer) for 30 min at roomThe water was changed and the container thoroughly temperature. Sections were rinsed four times again andcleaned between each swimming session. Rats were al- treated with diaminobenzidine (0.02% in Tris buffer,lowed to dry in a warm environment (30–338C) after each 0.01% hydrogen peroxide) as a chromogen, intensifiedswimming session. The rats rested on day 4. On day 5, with nickel to visualize the c-Fos immunoreactive nuclei.chemical nociception was assessed with the formalin test Finally, sections were rinsed four times again and mounted[8]. Briefly, 0.1 ml of 1% formalin (37.5% formaldehyde on gelatin-subbed slides, air-dried, dehydrated with in-in 0.9% saline) was subcutaneously injected in the plantar creasing concentrations of ethanol (50, 75, 95, and 100%,pad of the right hind paw of the rats and their nociceptive 1 min each) and xylene (4 min), and protected with
L. Quintero et al. / Brain Research 965 (2003) 259–268 261
Permount-glued coverslips. A neuron was consideredlabeled only if the nucleus, with proper size and shape,showed an uniform, diffuse, and distinctly outlined brown-black staining distinguishable from background, and anunlabeled nucleolus, at magnifications of 403, 1003, and2003.
2 .4. Data reduction and statistical analysis
For the formalin test, we calculated the total pain scoreby summing all the pain scores recorded during each of the20 observation periods. We quantified the effect of swimstress on c-Fos expression by counting all the labelednuclei of 10–20 slices under bright-field illumination. The
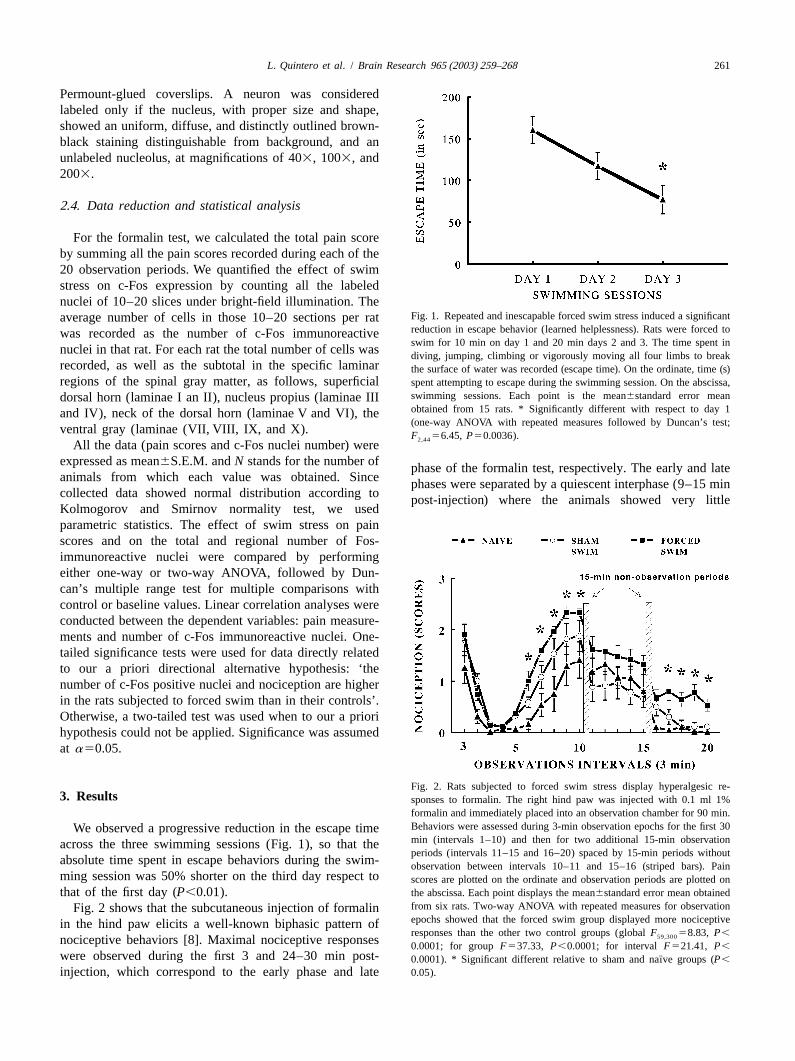
Fig. 1. Repeated and inescapable forced swim stress induced a significantaverage number of cells in those 10–20 sections per ratreduction in escape behavior (learned helplessness). Rats were forced towas recorded as the number of c-Fos immunoreactiveswim for 10 min on day 1 and 20 min days 2 and 3. The time spent in
nuclei in that rat. For each rat the total number of cells was diving, jumping, climbing or vigorously moving all four limbs to breakrecorded, as well as the subtotal in the specific laminar the surface of water was recorded (escape time). On the ordinate, time (s)regions of the spinal gray matter, as follows, superficial spent attempting to escape during the swimming session. On the abscissa,
swimming sessions. Each point is the mean6standard error meandorsal horn (laminae I an II), nucleus propius (laminae IIIobtained from 15 rats. * Significantly different with respect to day 1and IV), neck of the dorsal horn (laminae V and VI), the(one-way ANOVA with repeated measures followed by Duncan’s test;
ventral gray (laminae (VII, VIII, IX, and X). F 56.45, P50.0036).2,44All the data (pain scores and c-Fos nuclei number) were
expressed as mean6S.E.M. andN stands for the number of phase of the formalin test, respectively. The early and lateanimals from which each value was obtained. Since phases were separated by a quiescent interphase (9–15 mincollected data showed normal distribution according to post-injection) where the animals showed very littleKolmogorov and Smirnov normality test, we usedparametric statistics. The effect of swim stress on painscores and on the total and regional number of Fos-immunoreactive nuclei were compared by performingeither one-way or two-way ANOVA, followed by Dun-can’s multiple range test for multiple comparisons withcontrol or baseline values. Linear correlation analyses wereconducted between the dependent variables: pain measure-ments and number of c-Fos immunoreactive nuclei. One-tailed significance tests were used for data directly relatedto our a priori directional alternative hypothesis: ‘thenumber of c-Fos positive nuclei and nociception are higherin the rats subjected to forced swim than in their controls’.Otherwise, a two-tailed test was used when to our a priorihypothesis could not be applied. Significance was assumedat a50.05.
Fig. 2. Rats subjected to forced swim stress display hyperalgesic re-3 . Results sponses to formalin. The right hind paw was injected with 0.1 ml 1%
formalin and immediately placed into an observation chamber for 90 min.Behaviors were assessed during 3-min observation epochs for the first 30We observed a progressive reduction in the escape timemin (intervals 1–10) and then for two additional 15-min observationacross the three swimming sessions (Fig. 1), so that theperiods (intervals 11–15 and 16–20) spaced by 15-min periods without
absolute time spent in escape behaviors during the swim-observation between intervals 10–11 and 15–16 (striped bars). Painming session was 50% shorter on the third day respect toscores are plotted on the ordinate and observation periods are plotted onthat of the first day (P,0.01). the abscissa. Each point displays the mean6standard error mean obtained
from six rats. Two-way ANOVA with repeated measures for observationFig. 2 shows that the subcutaneous injection of formalinepochs showed that the forced swim group displayed more nociceptivein the hind paw elicits a well-known biphasic pattern ofresponses than the other two control groups (globalF 58.83, P,59,300nociceptive behaviors [8]. Maximal nociceptive responses 0.0001; for groupF537.33, P,0.0001; for intervalF521.41, P,
were observed during the first 3 and 24–30 min post- ¨0.0001). * Significant different relative to sham and naıve groups (P,
injection, which correspond to the early phase and late 0.05).

262 L. Quintero et al. / Brain Research 965 (2003) 259–268
nociceptive behavior. Two-way ANOVA showed that 48 h 1), which is consistent with the topographical projection offollowing the last forced swim session, pain behavior the hindpaw afferents to spinal cord [25,26].during the formalin test was greater in forced swim The mean total number of nuclei expressing c-Fos
¨compared to sham swim and naıve (P,0.0001; Fig. 2). across all ipsilateral dorsal horn laminae was significantlyThe total nociceptive behavior (sum of all pain scores greater in forced swim rats (86621 nuclei) than in sham
¨across the formalin test) was about two times greater in swim (5469 nuclei) and naıve rats (3168 nuclei) (Fig. 3),¨swim stressed rats than in sham swim and naıve rats while no significant difference was observed between sham
¨ ¨(naıve59.9961.35; sham swim512.8361.93; forced swim and naıve rats (one-way ANOVA;F 54.30; P52,15
swim520.6661.60,N56; one-way ANOVA,F 57.413, 0.033). There were significant group differences in the2,15
P50.006). This increased total nociceptive behavior in the number of c-Fos nuclei in the laminae of the ipsilateralforced swim rats was mainly due to a more robust late dorsal horns (P,0.0001; Fig. 4). Forced swim rats had aphase from 18 to 90 min post-injection (observation higher number of labeled nuclei in laminae I–VI than the
¨intervals 6–20). Specifically, the magnitude of the maxi- control rats (sham swim and naıve) (P,0.01). Althoughmal (peak) and submaximal nociceptive behavior during two-way ANOVA of the number of c-Fos nuclei bythe late phase was greater in forced swim rats. Also, the laminae did not demonstrate a significant overall difference
¨duration of the late phase was longer in forced swim rats between sham and naıve rats, one-way ANOVA showedsince nociceptive behavior was still present 87–90 min that the number of c-Fos nuclei in laminae I–II and III–IV
¨post-injection in forced swim while it had almost dis- was greater in sham swim rats than in naıve rats (P,0.05).appeared 75–81 min following the injection of formalin in We observed few c-Fos positive nuclei in the lateral areas
¨naıve and sham swim rats. There were not significant of the ipsilateral ventral horn (laminae VII and IX) of¨differences in the magnitude of the early phase and forced and sham swim rats but not in the naıve rats. The
interphase of the formalin test among the three experimen- number of c-Fos positive nuclei in the ipsilateral dorsaltal groups. horn was positively correlated with the total nociceptive
scores evoked by formalin administration (r50.529; P,0.0239;N518;) (FIG. 5). We also found a positive lineal
3 .1. Effect of swim stress on formalin-induced c-Fos correlation between total pain scores and the number ofexpression the spinal cord c-Fos positive nuclei in laminae I–II (r50.497; P5
0.0356; N518), III–IV (r50.496; P50.0361; N518),¨In the naıve rats injected with 1% formalin c-Fos and V–VI (r50.481; P50.0434;N518).
immunoreactive nuclei were almost exclusively localized Two-way ANOVA showed a significant interactionin the medial aspects of the superficial laminae (I and II) between stress and the laminar distribution of c-Fos nucleiand the neck of the dorsal horn (laminae V and VI) (FIG. in the ipsilateral dorsal horn (F52.09;P50.0233). Indeed,
¨Fig. 3. Photomicrographs (original magnification 1003) show formalin-induced expression of c-Fos protein in the ipsilateral dorsal horn of naıve rats (A)¨or rats subjected to sham (B) or forced (C) swimming rats. When compared to naıve and sham swim rats, the larger relative increase in forced swim rats
was observed in laminae III–IV, followed by laminae IV–V, while the smallest increase was seen laminae I–IIo. DC5dorsal columns.
L. Quintero et al. / Brain Research 965 (2003) 259–268 263
increase seen in laminae I–II (42%) and laminae III–IV(78%), respectively. In contrast to swim stress rats, theincrease in the number of c-Fos nuclei in the sham swim
¨rats relative to naıve rats was similar for each dorsallamina, by about 100%.
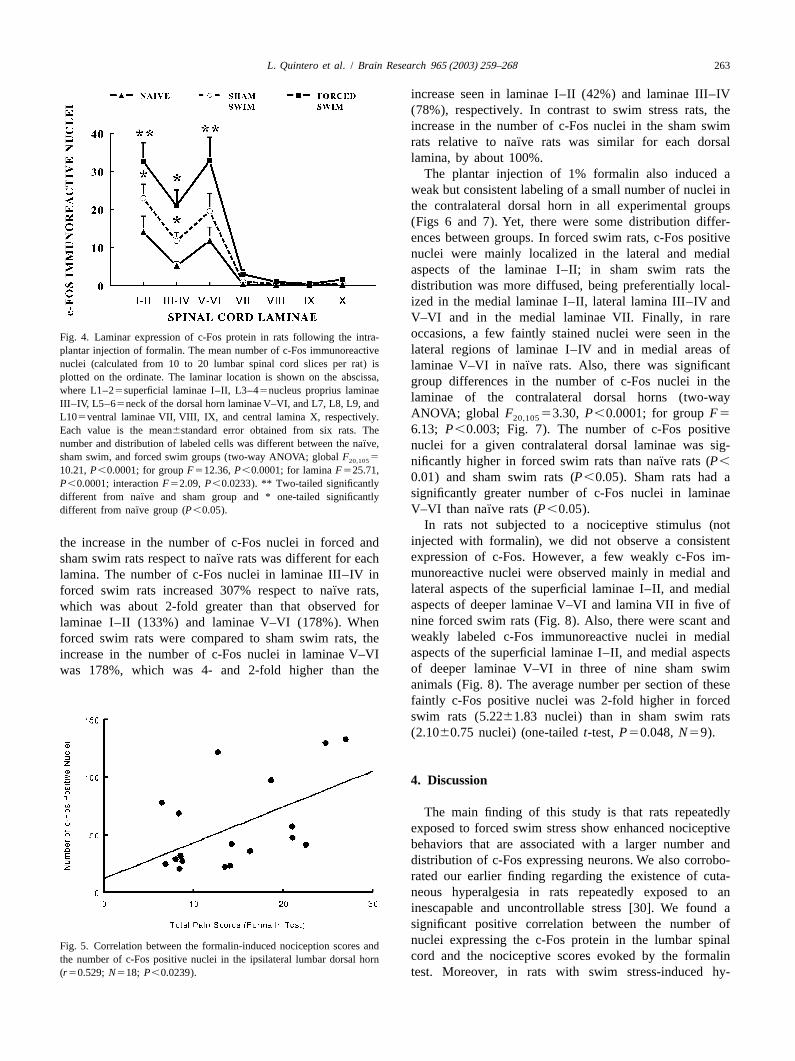
The plantar injection of 1% formalin also induced aweak but consistent labeling of a small number of nuclei inthe contralateral dorsal horn in all experimental groups(Figs 6 and 7). Yet, there were some distribution differ-ences between groups. In forced swim rats, c-Fos positivenuclei were mainly localized in the lateral and medialaspects of the laminae I–II; in sham swim rats thedistribution was more diffused, being preferentially local-ized in the medial laminae I–II, lateral lamina III–IV andV–VI and in the medial laminae VII. Finally, in rareoccasions, a few faintly stained nuclei were seen in theFig. 4. Laminar expression of c-Fos protein in rats following the intra-lateral regions of laminae I–IV and in medial areas ofplantar injection of formalin. The mean number of c-Fos immunoreactive
nuclei (calculated from 10 to 20 lumbar spinal cord slices per rat) is ¨laminae V–VI in naıve rats. Also, there was significantplotted on the ordinate. The laminar location is shown on the abscissa, group differences in the number of c-Fos nuclei in thewhere L1–25superficial laminae I–II, L3–45nucleus proprius laminae
laminae of the contralateral dorsal horns (two-wayIII–IV, L5–65neck of the dorsal horn laminae V–VI, and L7, L8, L9, andANOVA; global F 53.30, P,0.0001; for groupF5L105ventral laminae VII, VIII, IX, and central lamina X, respectively. 20,105
6.13; P,0.003; Fig. 7). The number of c-Fos positiveEach value is the mean6standard error obtained from six rats. The¨number and distribution of labeled cells was different between the naıve, nuclei for a given contralateral dorsal laminae was sig-
sham swim, and forced swim groups (two-way ANOVA; globalF 520,105 ¨nificantly higher in forced swim rats than naıve rats (P,10.21,P,0.0001; for groupF512.36,P,0.0001; for laminaF525.71,
0.01) and sham swim rats (P,0.05). Sham rats had aP,0.0001; interactionF52.09, P,0.0233). ** Two-tailed significantlysignificantly greater number of c-Fos nuclei in laminae¨different from naıve and sham group and * one-tailed significantly
¨¨ V–VI than naıve rats (P,0.05).different from naıve group (P,0.05).
In rats not subjected to a nociceptive stimulus (notinjected with formalin), we did not observe a consistentthe increase in the number of c-Fos nuclei in forced andexpression of c-Fos. However, a few weakly c-Fos im-¨sham swim rats respect to naıve rats was different for eachmunoreactive nuclei were observed mainly in medial andlamina. The number of c-Fos nuclei in laminae III–IV inlateral aspects of the superficial laminae I–II, and medial¨forced swim rats increased 307% respect to naıve rats,aspects of deeper laminae V–VI and lamina VII in five ofwhich was about 2-fold greater than that observed fornine forced swim rats (Fig. 8). Also, there were scant andlaminae I–II (133%) and laminae V–VI (178%). Whenweakly labeled c-Fos immunoreactive nuclei in medialforced swim rats were compared to sham swim rats, theaspects of the superficial laminae I–II, and medial aspectsincrease in the number of c-Fos nuclei in laminae V–VIof deeper laminae V–VI in three of nine sham swimwas 178%, which was 4- and 2-fold higher than theanimals (Fig. 8). The average number per section of thesefaintly c-Fos positive nuclei was 2-fold higher in forcedswim rats (5.2261.83 nuclei) than in sham swim rats(2.1060.75 nuclei) (one-tailedt-test, P50.048,N59).
4 . Discussion
The main finding of this study is that rats repeatedlyexposed to forced swim stress show enhanced nociceptivebehaviors that are associated with a larger number anddistribution of c-Fos expressing neurons. We also corrobo-rated our earlier finding regarding the existence of cuta-neous hyperalgesia in rats repeatedly exposed to aninescapable and uncontrollable stress [30]. We found asignificant positive correlation between the number ofnuclei expressing the c-Fos protein in the lumbar spinalFig. 5. Correlation between the formalin-induced nociception scores andcord and the nociceptive scores evoked by the formalinthe number of c-Fos positive nuclei in the ipsilateral lumbar dorsal horn
(r50.529; N518; P,0.0239). test. Moreover, in rats with swim stress-induced hy-
264 L. Quintero et al. / Brain Research 965 (2003) 259–268
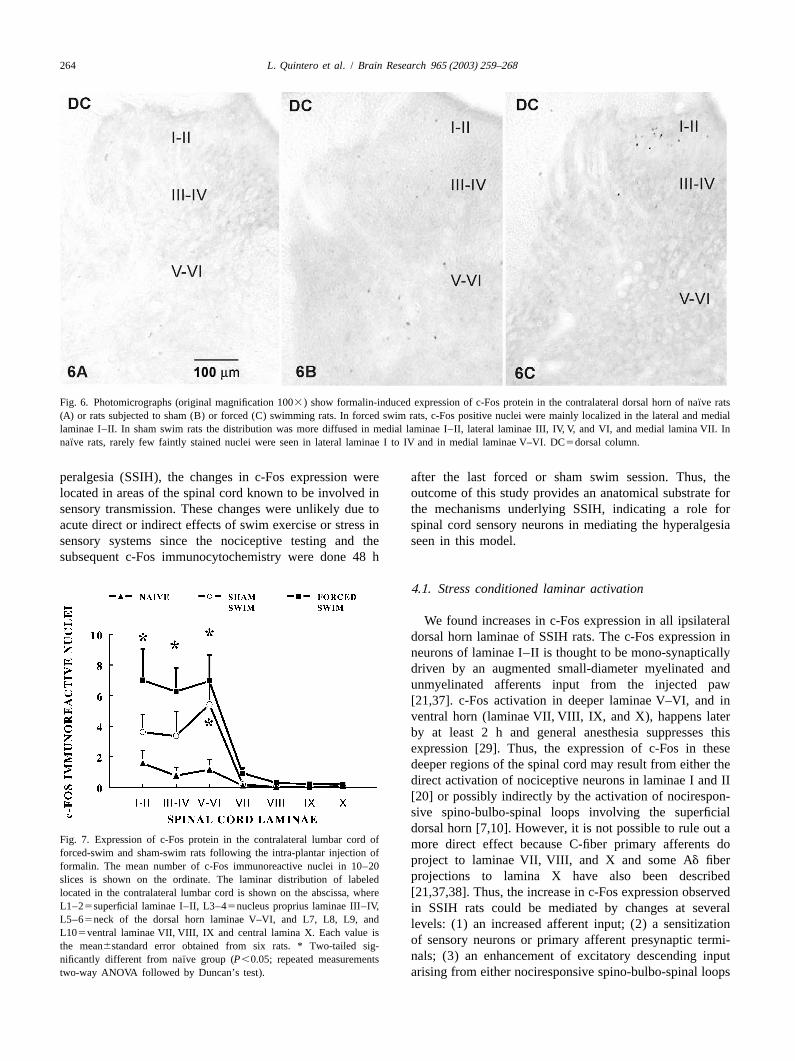
¨Fig. 6. Photomicrographs (original magnification 1003) show formalin-induced expression of c-Fos protein in the contralateral dorsal horn of naıve rats(A) or rats subjected to sham (B) or forced (C) swimming rats. In forced swim rats, c-Fos positive nuclei were mainly localized in the lateral and mediallaminae I–II. In sham swim rats the distribution was more diffused in medial laminae I–II, lateral laminae III, IV, V, and VI, and medial lamina VII. In
¨naıve rats, rarely few faintly stained nuclei were seen in lateral laminae I to IV and in medial laminae V–VI. DC5dorsal column.
peralgesia (SSIH), the changes in c-Fos expression were after the last forced or sham swim session. Thus, thelocated in areas of the spinal cord known to be involved in outcome of this study provides an anatomical substrate forsensory transmission. These changes were unlikely due to the mechanisms underlying SSIH, indicating a role foracute direct or indirect effects of swim exercise or stress in spinal cord sensory neurons in mediating the hyperalgesiasensory systems since the nociceptive testing and the seen in this model.subsequent c-Fos immunocytochemistry were done 48 h
4 .1. Stress conditioned laminar activation
We found increases in c-Fos expression in all ipsilateraldorsal horn laminae of SSIH rats. The c-Fos expression inneurons of laminae I–II is thought to be mono-synapticallydriven by an augmented small-diameter myelinated andunmyelinated afferents input from the injected paw[21,37]. c-Fos activation in deeper laminae V–VI, and inventral horn (laminae VII, VIII, IX, and X), happens laterby at least 2 h and general anesthesia suppresses thisexpression [29]. Thus, the expression of c-Fos in thesedeeper regions of the spinal cord may result from either thedirect activation of nociceptive neurons in laminae I and II[20] or possibly indirectly by the activation of nocirespon-sive spino-bulbo-spinal loops involving the superficialdorsal horn [7,10]. However, it is not possible to rule out a
Fig. 7. Expression of c-Fos protein in the contralateral lumbar cord of more direct effect because C-fiber primary afferents doforced-swim and sham-swim rats following the intra-plantar injection of
project to laminae VII, VIII, and X and some Ad fiberformalin. The mean number of c-Fos immunoreactive nuclei in 10–20projections to lamina X have also been describedslices is shown on the ordinate. The laminar distribution of labeled
located in the contralateral lumbar cord is shown on the abscissa, where[21,37,38]. Thus, the increase in c-Fos expression observedL1–25superficial laminae I–II, L3–45nucleus proprius laminae III–IV, in SSIH rats could be mediated by changes at severalL5–65neck of the dorsal horn laminae V–VI, and L7, L8, L9, and levels: (1) an increased afferent input; (2) a sensitizationL105ventral laminae VII, VIII, IX and central lamina X. Each value is
of sensory neurons or primary afferent presynaptic termi-the mean6standard error obtained from six rats. * Two-tailed sig-nals; (3) an enhancement of excitatory descending input¨nificantly different from naıve group (P,0.05; repeated measurements
two-way ANOVA followed by Duncan’s test). arising from either nociresponsive spino-bulbo-spinal loops
L. Quintero et al. / Brain Research 965 (2003) 259–268 265
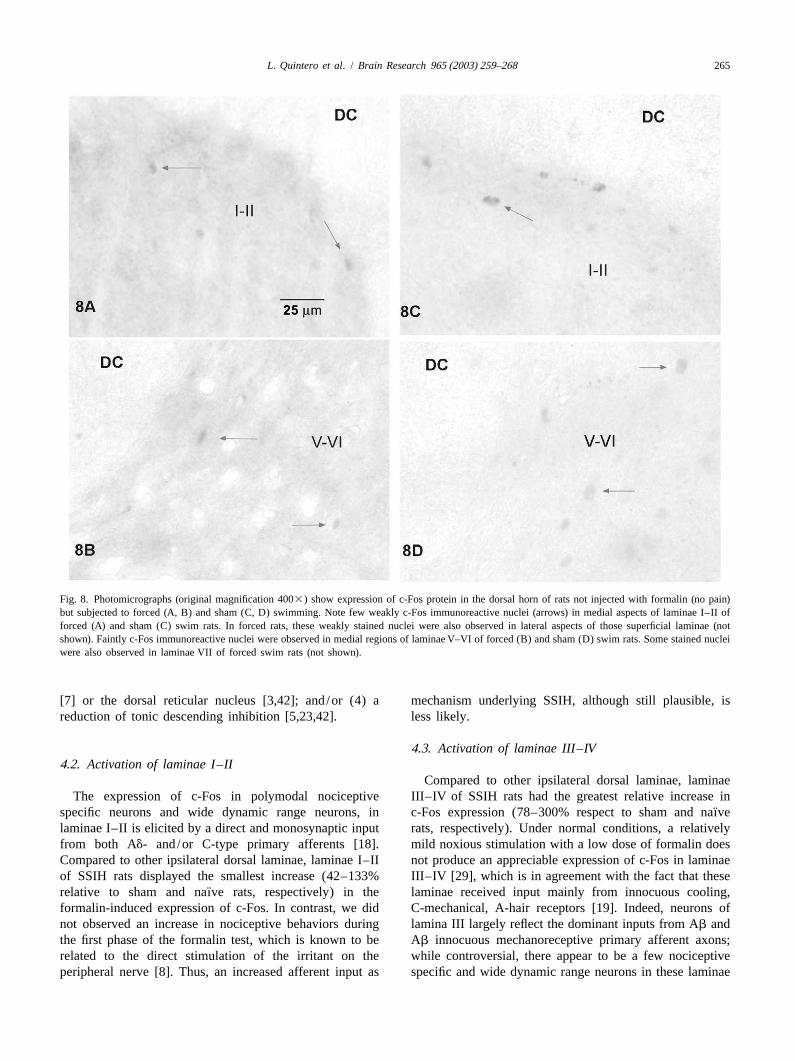
Fig. 8. Photomicrographs (original magnification 4003) show expression of c-Fos protein in the dorsal horn of rats not injected with formalin (no pain)but subjected to forced (A, B) and sham (C, D) swimming. Note few weakly c-Fos immunoreactive nuclei (arrows) in medial aspects of laminae I–II offorced (A) and sham (C) swim rats. In forced rats, these weakly stained nuclei were also observed in lateral aspects of those superficial laminae (notshown). Faintly c-Fos immunoreactive nuclei were observed in medial regions of laminae V–VI of forced (B) and sham (D) swim rats. Some stained nucleiwere also observed in laminae VII of forced swim rats (not shown).
[7] or the dorsal reticular nucleus [3,42]; and/or (4) a mechanism underlying SSIH, although still plausible, isreduction of tonic descending inhibition [5,23,42]. less likely.
4 .3. Activation of laminae III–IV4 .2. Activation of laminae I–II
Compared to other ipsilateral dorsal laminae, laminaeThe expression of c-Fos in polymodal nociceptive III–IV of SSIH rats had the greatest relative increase in
¨specific neurons and wide dynamic range neurons, in c-Fos expression (78–300% respect to sham and naıvelaminae I–II is elicited by a direct and monosynaptic input rats, respectively). Under normal conditions, a relativelyfrom both Ad- and/or C-type primary afferents [18]. mild noxious stimulation with a low dose of formalin doesCompared to other ipsilateral dorsal laminae, laminae I–II not produce an appreciable expression of c-Fos in laminaeof SSIH rats displayed the smallest increase (42–133% III–IV [29], which is in agreement with the fact that these
¨relative to sham and naıve rats, respectively) in the laminae received input mainly from innocuous cooling,formalin-induced expression of c-Fos. In contrast, we did C-mechanical, A-hair receptors [19]. Indeed, neurons ofnot observed an increase in nociceptive behaviors during lamina III largely reflect the dominant inputs from Ab andthe first phase of the formalin test, which is known to be Ab innocuous mechanoreceptive primary afferent axons;related to the direct stimulation of the irritant on the while controversial, there appear to be a few nociceptiveperipheral nerve [8]. Thus, an increased afferent input as specific and wide dynamic range neurons in these laminae

266 L. Quintero et al. / Brain Research 965 (2003) 259–268
[18]. Lamina IV contains neurons that received innocuous The effect of stress in nociception could be even moremechanoreceptive inputs [18]. Specific nocireceptive or complex and dynamic. Wei et al. [42] proposed that bothwide dynamic range neurons of lamina IIo are known to nucleus raphe magnus (serotoninergic) and locusproject to laminae III–IV [18]. Thus, the neuronal activa- coeruleus/subcoeruleus (noradrenergic) descending path-tion of laminae III–IV in SSIH animals, subjected to ways are major sources of enhanced inhibitory modulationnoxious stimulation (formalin injection), could be due to a in acute inflammatory pain. Instead, the persistent hy-stress-enhanced relay of nociceptive input from lamina IIo, peralgesia and neuronal hyperexcitability may be mediatedor a sensitization of laminae III–IV non-nociceptive and in part by a descending pain facilitatory system involvingnociceptive neurons. A facilitated activation of laminae nuclei reticularis gigantocellularis [3,42]. Thus, the intensi-III–IV could be associated with a cutaneous allodynic state ty of perceived pain and hyperalgesia is fine-tuned byin SSIH animals. descending pathways. Repeated inescapable and uncon-
trollable stress might produce an imbalance of these4 .4. Activation of laminae V–VI modulating systems to favor hyperalgesic conditions.
The second greatest increase (about 178% respect to4 .6. Enhanced contralateral activation of the dorsal¨sham or naıve rats) in the expression of c-Fos was seen inhorn
laminae V–VI of SSIH rats. Lamina V contains neuronsthat project mainly to the thalamus and receive both Under normal conditions, painful stimuli applied to theinnocuous and nociceptive primary afferent fibers [10]. foot induce c-Fos expression almost exclusively in theInterestingly, lamina V is known to contain the highest ipsilateral spinal cord. Yet, we observed that the injectionpopulation of wide dynamic range neurons, which receive of a relatively low dose of formalin induced a weak butinputs from innocuous mechanoreceptors and nociceptors. consistent c-Fos expression in the contralateral side theThe activity of these neurons is tightly coupled to nocicep- dorsal horn in both forced and sham swim groups. Thetive behaviors and the sensory discrimination of nocicep- respective increase in c-Fos labeling was, 6- and 3-fold
¨tive stimuli [18]. In addition, neurons in laminae V–VI are greater than that observed in naıve rats. Previous studiesstrongly modulated by supraspinal structures [42,43], have shown that procedures, which produce hyperalgesia,which suggest that neurons in these laminae contribute to enhance the contralateral expression of c-Fos. For exam-SSIH, as a result of changes in the activity of descending ple, morphine-tolerant rats, when compared to control rats,modulatory pathways. show significant and bilateral increases in Fos-like im-
munoreactivity in the L4/5 and L2 segments of the dorsal4 .5. Role of descending pain modulation in SSIH spinal cord in response to the unilateral injection of
formalin into a hindpaw [32]. The authors of this studySpinoparabrachial neurons located in the lateral reticu- suggested that lumbar spinal cord neurons are sensitized
lated neck of the dorsal horn (laminae V–VI) and the during the development of tolerance, and that the sensitiza-superficial dorsal horn (laminae I–II) are inhibited by tion can be unmasked by the administration of a noxiousserotoninergic and noradrenergic descending fibers, respec- stimulus.tively [42,43]. After acute exposure to different types of The anatomical bases for this contralateral activationadverse psychological or physical stimuli, the extracellular have been reported. The anterograde transport of tritiatedconcentrations of serotonin increase in several brain re- amino acids injected into dorsal ganglia has demonstratedgions, especially in the raphe magnus [1]. Conversely, projections to the contralateral marginal zone and lamina Vprolong stress diminishes the efflux of serotonin in some of spinal sacral and coccigeal segments [31]. Thus, re-brain structures known to be activated by stress, such as peated stressful conditions could produce a contralateralthe amygdala and lateral septum [2,14]. Interestingly, the sensitization of the spinal cord, manifested as increasedactivity of serotoninergic descending fibers from the raphe pain-induced expression of c-Fos protein. It is also pos-magnus tonically inhibits pain transmission within the sible that bilateral alterations in descending control sys-spinal cord [23] and the inhibition of serotonin synthesis at tems can also contribute to the bilateral expression ofthe spinal level produces hyperalgesia in rats [5]. The c-Fos in sensory pathways of the spinal cord. Thismagnitude of tonic inhibition of pain transmission within enhanced contralateral spinal activation induced by re-the spinal dorsal horn appears to depend on the behavioral peated swim stress might contribute to the referral ofstate of the organism (depressed mood, anxiety, fear) [23]. painful sensations to the contralateral limb.Finally, we have reported that serotoninergic-selectiveantidepressants, such as, clormipramine and fluoxetine as4 .7. Pre-activation of spinal sensory neurons bywell as tryptophan, prevents the development of SSIH repeated-swim stress:[30]. Thus, the increased expression of c-Fos in SSIH ratsmay be linked, at least in part, to a stress-induced decrease In the absence of a nociceptive stimulus (no formalinin serotoninergic tonic inhibition of nociception. injection), we found few faintly stained cells for c-Fos in

L. Quintero et al. / Brain Research 965 (2003) 259–268 267
lateral and medial aspects of laminae I–II, V–VI, and VII stimulus applied 2 weeks later [6]. Thus, the outcomes ofin 55% of forced swim rats and 33% of sham swim rats, the present study seem to be related to a generalizedwhile the number of the c-Fos labeled nuclei was about sensitization by an uncontrollable stress. Noteworthy,2-fold higher in forced swim rats than in sham swim rats. controllable challenges generally activate mechanisms thatThis pre-activation or sensitization of sensory neurons in consolidate and coordinate ongoing behavior, and repeatedthe absence of a nociceptive stimulus could be related to exposure to the same stressor will thus lead to habituationthe repeated swim stress procedure. In support of this of the neuro-endocrine stress response. In contrast, expo-view, repeated chronic (but not acute) social stress sure to an uncontrollable stress, such as forced swim, canproduces a persistent c-Fos expression in sensory relay induce an increase in the behavioral and neuro-endocrinenuclei in the brain, brain stem, and spinal cord of the responses following repeated exposure to the same stressormouse at least after 24 h the last stressor [24]. On the other [35].hand, persistent c-Fos expression has been seen with longsurvival times following formalin injection [44]. Thesuperficial pattern gives way to the appearance of a A cknowledgementspopulation of immunoreactive cells in the deeper laminaethat is still present after 24 h, having become even more This work was supported by FONACIT S-1 2000000787diffuse and symmetrical [44]. Thus, at early stages, stress(Ministry of Science and Technology, Venezuela) andor injury appears to trigger a complex pattern of molecular VAC-CONDES CC-0325-02 (University of Zulia, Ven-events within the spinal cord, which are initially mono- ezuela).synaptic but in the longer term are associated to an inducedpattern of synaptic activity set up within the spinal cord.
Since c-Fos expression, in the absence of a nociceptiveR eferencesstimulus, was located in laminae of the dorsal horninvolved in pain processing, this suggests that a pre- [1] A. Adell, J. Casanovas, F. Artigas, Comparative study in the rat ofactivation or sensitization of sensory neurons by repeated the actions different types of stress on the release of 5-HT in rapheswim stress underlies SSIH. The number and distribution nuclei and forebrain areas, Neuropharmacology 36 (1997) 735–741.
[2] H.A. Akil, M.I. Morano, Stress, in: F.E. Bloom, D.J. Kupter (Eds.),of sensory neurons pre-sensitized by stress might bePsychopharmacology: The Fourth Generation of Progress, Ravencrucial for the expression of hyperalgesia because thePress, New York, 1995, pp. 773–785.
hyperalgesic forced swim rats expressed 70% more c-Fos [3] A. Almeida, R. Storkson, D. Lima, K. Hole, A. Tjolsen, Thelabeled cells, which were also more widely distributed medullary dorsal reticular nucleus facilitates pain behaviour inducedcompared to sham swim controls. Although we did not by formalin in the rat, Eur. J. Neurosci. 11 (1999) 110–122.
[4] A. Badiani, M.M. Oates, H.E. Day, S.J. Watson, H. Akil, T.E.observed any gross behavioral difference between ourRobinson, Amphetamine-induced behavior, dopamine release, andexperimental groups, we cannot ruled out that this swim-c-fos mRNA expression: modulation by environmental novelty, J.
stress induced c-Fos expression in the dorsal horn is Neurosci. 18 (1998) 10579–10593.associated with some degree of sensory discomfort (subtle [5] R.J. Bodnar, J.H. Kordower, M.M. Wallace, H. Tamir, Stress andallodynia and/or spontaneous pain). morphine analgesia: alterations followingp-chlorophenylalanine,
Pharmacol. Biochem. Behav. 14 (1981) 645–651.[6] A.W. Bruijnzeel, R. Stam, J.C. Compaan, G. Croiset, L.M. Akker-4 .8. Long-lasting sensitization by uncontrollable stress
mans, B. Olivier, V.M. Wiegant, Long-term sensitization of Fos-responsivity in the rat central nervous system after a single stressful
A number of animal models has been developed in experience, Brain Res. 819 (1999) 15–22.which intense stressful experiences (shocks, social con- [7] F. Cervero, J.H. Wolstencroft, A positive feedback loop between
spinal cord nociceptive pathways and antinociceptive areas of thefrontations) result in long-term altered behavioural, au-cat’s brain stem, Pain 20 (1984) 125–138.tonomic and hormonal responsivity to aversive challenges
[8] D. Dubuisson, S.G. Dennis, The formalin test: A quantitative study(for a review, see Ref. [35]). Rats exposed to a single brief of the analgesic effects of morphine, meperidine, and brain stemsession of inescapable foot shock display long-lasting (up stimulation in rats and cats, Pain 4 (1977) 161–174.to 4 weeks) increased immobility, enhanced ACTH and [9] G.D. Gamaro, M.H. Xavier, J.D. Denardin, J.A. Pilger, D.R. Ely,
M.B. Ferreira, C. Dalmaz, The effects of acute and repeated restraintcorticosterone response in the open field test [16,39,40],stress on the nociceptive response in rats, Physiol. Behav. 63 (1998)increased colonic responses, and increased heart rate693–697.
responses to an electrified prod inserted in the home cage[10] G.J.J. Giesler, D. Menetrey, A.I. Basbaum, Differential origins of[27,36]. They also show increased release of norepineph- spinothalamic tract projections to medial and lateral thalamus in therine in different brain areas upon the exposure to a second rat, J. Comp. Neurol. 184 (1979) 107–126.
[11] R.L. Hayes, G.J. Bennett, P.G. Newlon, D.J. Mayer, Behavioral andnovel stimulus [40]. Moreover, a single stressful ex-physiological studies of non-narcotic analgesia in the rat elicited byperience can produce the acute expression of c-Fos in thecertain environmental stimuli, Brain Res. 155 (1978) 69–90.
brain that quickly disappears in few hours [24]. However, [12] S.P. Hunt, A. Pini, G. Evan, Induction of c-fos-like protein in spinala long-term activation of c-Fos occurs in several brain cord neurons following sensory stimulation, Nature 328 (1987)regions following a second exposure to an aversive 632–634.

268 L. Quintero et al. / Brain Research 965 (2003) 259–268
[13] E. Jorum, Noradrenergic mechanisms in mediation of stress-induced morphine suppresses noxious stimulus-evoked Fos protein-likehyperalgesia in rats [abstract], Pain 32 (1988) 349–355. immunoreactivity in the rat spinal cord, J. Neurosci. 10 (1990)
[14] L.G. Kirby, A.R. Allen, I. Lucki, Regional differences in the effects 323–335.of forced swimming on extracellular levels of 5-hydroxytryptamine [30] L. Quintero, M. Moreno, C. Avila, J.L. Arcaya, H. Suarez-Roca,and 5-hydroxyindoleacetic acid [abstract], Brain Res. 682 (1995) Long-lasting delayed hyperalgesia after subchronic swim stress189–196. [abstract], Pharmacol. Biochem. Behav. 67 (2000) 449–458.
[15] C. Lacheze, A.M. Coelho, J. Fioramonti, L. Bueno, Influence of [31] M. Rethelyi, J.J. Capowski, The terminal arborization pattern oftrimebutine on inflammation- and stress-induced hyperalgesia to primary afferent fibers in the substantia gelatinosa of the spinal cordrectal distension in rats, J. Pharm. Pharmacol. 50 (1998) 921–928. in the cat, J. Physiol. (Paris) 73 (1977) 269–277.
[16] S. Levine, J. Madden, R.L. Conner, J.R. Moskal, D.C. Anderson, [32] D.S. Rohde, D.J. Detweiler, A.I. Basbaum, Formalin-evoked FosPhysiological and behavioral effects of prior aversive stimulation expression in spinal cord is enhanced in morphine-tolerant rats,(preshock) in the rat, Physiol. Behav. 10 (1973) 467–471. Brain Res. 766 (1997) 93–100.
[17] J.W. Lewis, J.T. Cannon, J.C. Liebeskind, Opioid and nonopioid [33] M.A. Ruis, J.H. te Brake, B. Buwalda, S.F. De Boer, P. Meerlo,mechanisms of stress analgesia [abstract], Science 208 (1980) 623– S.M. Korte, H.J. Blokhuis, J.M. Koolhaas, Housing familiar male625. wildtype rats together reduces the long-term adverse behavioural
[18] A.R. Light, The organization of nociceptive neurons in the spinal and physiological effects of social defeat, Psychoneuroendocrinolo-grey matter, in: P.L. Gildenberg (Ed.), The Initial Processing of Pain gy 24 (1999) 285–300.and Its Descending Control: Spinal and Trigeminal Systems, Karger, [34] M. Satoh, Y. Kuraishi, M. Kawamura, Effects of intrathecal anti-Basel, 1992, pp. 109–177. bodies to substance P, calcitonin gene-related peptide and galanin on
[19] A.R. Light, Spinal terminations of nociceptive primary afferent repeated cold stress-induced hyperalgesia: comparison with car-axons, in: P.L. Gildenberg (Ed.), The Initial Processing of Pain and rageenan-induced hyperalgesia [abstract], Pain 49 (1992) 273–278.Its Descending Control: Spinal and Trigeminal Systems, Karger, [35] R. Stam, A.W. Bruijnzeel, V.M. Wiegant, Long-lasting stress sensiti-Basel, 1992, pp. 87–108. sation, Eur. J. Pharmacol. 405 (2000) 217–224.
[20] A.R. Light, A.M. Kavookjian, Morphology and ultrastructure of [36] R. Stam, G. Croiset, L.M. Akkermans, V.M. Wiegant, Sensitizationphysiologically identified substantia gelatinosa (lamina II) neurons of the colonic response to novel stress after previous stressfulwith axons that terminate in deeper dorsal horn laminae (III–V), J. experience, Am. J. Physiol. 271 (1996) R1270–R1273.Comp. Neurol. 267 (1988) 172–189. [37] Y. Sugiura, C.L. Lee, E.R. Perl, Central projections of identified,
[21] A.R. Light, E.R. Perl, Spinal termination of functionally identified unmyelinated (C) afferent fibers innervating mammalian skin,primary afferent neurons with slowly conducting myelinated fibers, Science 234 (1986) 358–361.J. Comp. Neurol. 186 (1979) 133–150. [38] J.E. Swett, C.J. Woolf, The somatotropic organization of primary
[22] J. Madden, H. Akil, R.L. Patrick, J.D. Barchas, Stress-induced afferent terminals in the superficial laminae of the dorsal horn of theparallel changes in central opioid levels and pain responsiveness in rat spinal cord, J. Comp. Neurol. 231 (1985) 66–77.the rat, Nature 265 (1977) 358–360. [39] H.H. van Dijken, D.C. de Goeij, W. Sutanto, J. Mos, E.R. de Kloet,
[23] P. Mason, K. Gao, Raphe magnus serotonergic neurons tonically F.J. Tilders, Short inescapable stress produces long-lasting changesmodulate nociceptive transmission, Pain Forum 7 (1998) 143–150. in the brain–pituitary–adrenal axis of adult male rats, Neuroen-
[24] S. Matsuda, H. Peng, H. Yoshimura, T.C. Wen, T. Fukuda, M. docrinology 58 (1993) 57–64.Sakanaka, Persistent c-fos expression in the brains of mice with [40] J.G. Veening, L.W. Swanson, P.E. Sawchenko, The organization ofchronic social stress, Neurosci. Res. 26 (1996) 157–170. projections from the central nucleus of the amygdala to brainstem
[25] C. Molander, G. Grant, Cutaneous projections from the rat hindlimb sites involved in central autonomic regulation: a combined retro-foot to the substantia gelatinosa of the spinal cord studied by grade transport–immunohistochemical study, Brain Res. 303 (1984)transganglionic transport of WGA-HRP conjugate, J. Comp. Neurol. 337–357.237 (1985) 476–484. [41] C. Vidal, J.J.C. Jacob, Stress hyperalgesia in rats: an experimental
[26] C. Molander, G. Grant, Laminar distribution and somatotropic animal model of anxiogenic hyperalgesia in human, Life Sci. 31organization of primary afferent fibers from hindlimb nerves in the (1982) 1241–1244.dorsal horn. A study by transganglionic transport of horseradish [42] F. Wei, R. Dubner, K. Ren, Nucleus reticularis gigantocellularis andperoxidase in the rat, Neuroscience 19 (1986) 297–312. nucleus raphe magnus in the brain stem exert opposite effects on
[27] M.J. Nijsen, G. Croiset, M. Diamant, R. Stam, P.J. Kamphuis, A. behavioral hyperalgesia and spinal Fos protein expression afterBruijnzeel, D. de Wied, V.M. Wiegant, Endogenous corticotropin- peripheral inflammation, Pain 80 (1999) 127–141.releasing hormone inhibits conditioned-fear-induced vagal activation [43] F. Wei, R. Dubner, K. Ren, Laminar-selective noradrenergic andin the rat, Eur. J. Pharmacol. 389 (2000) 89–98. serotoninergic modulation includes spinoparabrachial cells after
[28] R.D. Porsolt, G. Anton, N. Blavet, M. Jalfre, Behavioral despair in inflammation, Neuroreport 10 (1999) 1757–1761.rats: A new model sensitive to antidepressant treatments, Eur. J. [44] S. Williams, G.I. Evan, S.P. Hunt, Changing patterns of c-fosPharmacol. 47 (1978) 379–391. induction in spinal neurons following thermal cutaneous stimulation
[29] R.W. Presley, D. Menetrey, J.D. Levine, A.I. Basbaum, Systemic in the rat, Neuroscience 36 (1990) 73–81.




















