
Physical and chemical changes of polystyrene nanospheres irradiated with laser
Upload
independentCategory
view
0download
0
Cell Biology International 2000, Vol. 24, No. 9, 635–648doi:10.1006/cbir.2000.0558, available online at http://www.idealibrary.com on
RELEASE OF MITOTIC DESCENDANTS BY GIANT CELLS FROM IRRADIATEDBURKITT’S LYMPHOMA CELL LINES
JEKATERINA A. ERENPREISA1*, MARK S. CRAGG2, BIRGITTA FRINGES4, IGOR SHARAKHOV5
and TIMOTHY M. ILLIDGE2,3
1Laboratory Tumor Cell Biology, Latvian University Biomedicine Centre, Riga, Latvia; 2Tenovus ResearchLaboratory, Cancer Sciences Division, Southampton General Hospital, Southampton, U.K.; 3The CRC Laboratory,
Southampton General Hospital, Southampton, U.K.; 4Institute of Pathology, Justus-Liebig-University, Giessen,Germany; 5Institute of Biology and Biophysics, Tomsk State University, Tomsk, Russia
Received 11 January 2000; accepted 3 May 2000
Polyploid giant cells are produced as part of the response of p53 mutant Burkitt’s lymphoma celllines to high doses of irradiation. Polyploid giant cells arise by endo-reduplication in the firstweek after a single 10 Gray dose of irradiation. Within the giant cells a sub-nuclear structure isapparent and within this, sub-nuclear autonomy is evident, as displayed by independent nuclearstructure and DNA replication in different parts of the nucleus. The majority of these cells soondie as apoptotic polykaryons. However, approximately 10–20% of giant cells remain viable intothe second week after irradiation and begin vigorous extrusion of large degraded chromatinmasses. During the second week, the giant cells begin to reconstruct their nuclei into polyploid‘bouquets’, where chromosome double-loops are formed. Subsequently, the bouquets return toan interphase state and separate into several secondary nuclei. The individual sub-nuclei thenresume DNA synthesis with mitotic divisions and sequester cytoplasmic territories aroundthemselves, giving rise to the secondary cells, which continue mitotic propagation. This processof giant cell formation, reorganization and breakdown appears to provide an additionalmechanism for repairing double-strand DNA breaks within tumour cells. � 2000 Academic Press
K: lymphoma; irradiation; giant cells; mitotic survivors.
*Please address all correspondence to: Dr Jekaterina Erenpreisa,Latvian University Biomedicine Centre, Ratsupites str.1, Riga,LV-1067, Latvia. E-mail: [email protected]
INTRODUCTION
A tight co-ordination of DNA replication andmitosis is thought to be required in the eukaryoticcell cycle to preserve the genetic fidelity of daughtercells and is regulated, amongst other factors, by thetumour suppressor gene p53 (Levine, 1997). Muta-tions of this gene are found in 50–60% of primarytumours and the frequency increases with tumourprogression (Greenblatt et al., 1994). Uncouplingof DNA replication from mitosis leads to theformation of polyploid giant cells (Nagl, 1995). Assuch, the formation of giant cells occurs throughwhat has been termed the endocycle and appears tobe a characteristic feature of the genomic instability
1065–6995/00/090635+14 $35.00/0
of p53 mutant or null cells (Cross et al., 1995; Oliveet al., 1996; Merritt et al., 1997). Giant cell forma-tion is further enhanced by irradiation (Peled et al.,1996) and is only one of a set of responses dis-played by damage-resistant tumours after geno-toxic insult, which have been grouped under thecollective term ‘mitotic’ or ‘reproductive’ death.The mechanisms which define mitotic death arecurrently ill-defined (Bernhard et al., 1996; Grafi,1998).
Mitotic death was reported to be associated withabrogation of the G1/S DNA damage checkpointand delay in the G2/M checkpoint. After adapta-tion, a few rounds of aberrant mitoses ensue,frequently followed by delayed (post-mitotic)apoptosis, micronucleation, and mitotic catas-trophe, culminating in the death of the polyploidgiant cells (Lock and Ross, 1990). Importantly,
� 2000 Academic Press

636 Cell Biology International, Vol. 24, No. 9, 2000
tumours that display mitotic death after severegenotoxic insults are more likely to be more resist-ant to, and ultimately survive, these treatments(Bernhard et al., 1996). In accordance, it is notablethat p53 mutant Burkitt’s lymphoma (BL) cellsare more radioresistant than wild-type possessingcounterparts (O’Connor et al., 1993).
It is widely accepted that, once initiated, endocy-cles do not reverse and that giant cells are thereforereproductively dead (Weibezahn et al., 1985). Thisview was based upon observations of cells in exper-imental models of genotoxic damage that did notextend any longer than 1 week post-damage. Incontrast, we have followed cells for up to 3 weekspost-damage and found by various methods,including separation, labelling, and cultivation ofgiant cells produced after severe irradiation ofseveral p53 mutant Burkitt’s lymphoma cell lines,that although the vast majority of giant cells die inthe first week post-irradiation, a small but signifi-cant proportion survive and persist for another 1–2weeks and may be a source of clonogenic survivors(Illidge et al., 2000, in this issue). In the presentstudy, we have examined the fate of the survivingpolyploid cells and revealed that giant cells of thesecond week post-irradiation undergo intranuclearsorting and complex nuclear reconstruction, whichresult in their final disintegration into mitoticdescendants. We believe that this novel mechanismmay provide an additional pathway by which p53mutant lymphomas may survive severe genotoxicdamage.
MATERIALS AND METHODS
Materials
Unless otherwise stated, all materials werepurchased from Sigma Chemical Co.
Maintenance of cell lines
The Ramos and Namalwa Burkitt’s lymphoma celllines used in this study have established p53 mutantalleles and are highly radioresistant (O’Connoret al., 1993). The cell lines were maintained inRPMI 1640 medium (Gibco, Paisley, Scotland),supplemented with antibiotics, 50 U/ml amphoter-icin B (Fungizone; Squibb and Sons, Cramlington,U.K.), 2 m L-glutamine (Gibco), 1 m pyruvate(Gibco) and 10% Myoclone plus foetal calf serum(FCS; Gibco) at 37�C in a 5% CO2 humidifiedincubator. For experimental studies, cells were
maintained in log-phase of growth for at least 24 hprior to experiment.
Irradiation of cells
All cell lines were irradiated with 10 Gy, using aPhillips 5 Mv linear accelerator photons at a doserate of 3.0 Gy/min.
Flow cytometric analysis of DNA
The DNA content of cells was assessed essentiallyas described previously (Cragg et al., 1999). Briefly,aliquots of cells (5–10�105) were centrifuged for5 min at 500�g and then washed once in PBS. Thecells were then resuspended in hypotonic fluoro-chrome solution (50 �g/ml propidium iodide (PI),0.1% (w/v) sodium citrate, 0.1% (w/v) TritonX-100) and incubated in the dark at 4�C overnightor 4 h at room temperature. Intensity of PI stainingof DNA from 10,000 cells was detected on aFACScan flow cytometer (Becton Dickinson,Bedford, U.K.). Untreated samples were used ascontrols to allow gating of cells in G1, S, G2/Mphases of the cell cycle. The proportion of DNAgiving fluorescence below the G1 peak (sub-G1)was taken as a measure of apoptosis and cells withlevels of fluorescence higher than in G2/M cellswere defined as polyploid.
[3H]-thymidine autoradiography
DNA replicative activity was determined at variousperiods post-irradiation by adding fresh [3H]-thymidine (Amersham, Little Chalfont, U.K.) to afinal concentration of 5 �Ci/ml (for a 60 min pulse)or 1–2 �Ci/ml (for 24 h labelling) to the culturemedium. In some cases, the 24 h labelled cells werechased following further cultivation. Cell sampleswere fixed as smears or cytospins, washed withdiluted perchloric acid and tap water and the slideswere exposed with fresh emulsion (Ilford K5) for1–8 weeks. Subsequently, radioautographs weredeveloped using Kodak 4-Dektol developer andlightly counterstained with 0.005% methylene blue.
Cytological examination using light and electronmicroscopy
For standard light microscopy observation, cellswere fixed as cytospins with ethanol/acetone (1:1),dried, treated with 0.1 N HCl, 4�C for 10 min,washed in distilled water and stained with0.05% toluidine blue (TB) in 50% citrate–phosphate McIlvain buffer, pH 5 for 10 min.

Cell Biology International, Vol. 24, No. 9, 2000 637
These preparations were used to determinecounts of mitotic, apoptotic, and segmentating–micronucleating cells. 1000–2000 cells werecounted per sample. For DNA staining, conven-tional Feulgen reaction or its TB-variant wasused. In the latter case, after hydrolysis with 5NHCl at 21�C for 20 min and thorough rinsing,the cytospins were stained with 0.05% TB in50% McIlvain buffer, pH 4 for 10 min. For elec-tron microscopy (EM), conventional fixation andembedding in Spurr resin was used. Subsequently,ultra-thin sections were cut and contrasted withlead citrate.
TUNEL (terminal dUTP nucleotide end labelling)
This method labels the double strand DNA(dsDNA) breaks and is therefore a direct indicatorof apoptosis. TUNEL analysis was carried out oncytospin cells, which were fixed in methanol/acetone (1:1) at �20�C for 10 min and then air-dried. The subsequent reaction was carried outaccording to the method of Gavrieli et al. (1992),with the proteinase K pretreatment omitted asdescribed by Roach et al. (1995). Briefly, the reac-tion was performed on slides under a cover slip byaddition of 25 �l of the end-labeling buffer (30 mTris/HCl, pH 7.2, 1% BSA, 140 m Na-cacodylate,1 m CoCl2) containing 2.5 � dNTP with thebiotin-conjugated dUTP, and 7.5 U terminal trans-ferase (Boehringer Mannheim Corp., Lewes, U.K.)at 37�C, for 1 h. This was followed by treatmentwith avidin–peroxidase conjugate, which wasfurther stained by 3-amino-9-ethylcarbazole (AEC)for visualization of the product.
In situ DNA denaturation test with acridine orange(AO)
This is an indirect test for the detection of singleand double DNA strand breaks (Darzynkiewicz,1994). In this test, nicked DNA acquires a red ororange colour, while non-degraded cell nucleiexhibit green fluorescence. Although usuallyapplied in flow cytometric analysis, here themethod was adjusted for visual observation ofcytospins or cell smears essentially as describedby Erenpreisa et al. (1997), omitting RNAsetreatment.
Fluorescence in situ hybridization (FISH)
FISH of centromeric probes for chromosomes 1, 3,4 and 9 was performed essentially according toRooney and Czepulkowski (1992) and the protocol
of ONCOR (http://www.oncor.com/prod-gen.htm)using biotinylated probes. For detection of probehybridization, slides were incubated with avidin-FITC and counterstained with propidium iodide inAntifade.
RESULTS
Origins of polyploid giant cells
BL cells undergo nearly full G2 arrest on day 1post-irradiation, comprising 86.7�3.3% of cells inG2 arrest in six experiments for Namalwa and89.2�2.1% for Ramos cells (Fig. 1A). Octaploidcells begin to appear simultaneously with therelease from G2-arrest (Fig. 1B). Electronmicroscopy (EM) revealed that many of these giantcells possess two pairs of centrioles lying side byside, suggesting that giant cells originate fromG2-arrested cells. The kinetics of polyploidizationreveal that subsequently there follows a dailydoubling of the DNA content in the giant cells,reaching 64N at the first week after irradiation(Fig. 1B). During this time, intense incorporationof [3H]-thymidine pulse label is observed ingiant cells (Fig. 2A). Together, these data suggestthat a sequence of endo-reduplication cycles isoccurring and that this mechanism elicits themajority of highly polyploid giant cells. Thoughthe numbers of polyploid cells does not exceedthe number of diploid cells, their mass on thefirst week considerably dominates the depresseddiploid stem-line, as can be seen from Figure1B. The polyploidization stage in the firstweek is followed by de-polyploidization of giantcells during the second week post-irradiation(Fig. 1B).
From day 2 onwards, aberrant mitoses areevident, which are often seen in pyknosis andoccasionally in restitution (Fig. 2B). The mitoseswhich occur up to anaphase disappear after day 5up to day 11–12 for Namalwa cells and day 14–16for the Ramos cell line, coinciding with the returnof the diploid stem-line. Although the daily dou-bling of the DNA content and intense uptake of[3H]-thymidine in the giant cell nuclei suggest thatendoreduplication is the predominant mechanisminvolved in giant cell formation, we cannot excludecell fusion as a minor source of polyploid giantcells and have seen rare cytological picturesthat may indicate this. Therefore, the source ofgiant cells is potentially three-fold, namelythrough endo-reduplication, mitotic restitution orcell fusion.

638 Cell Biology International, Vol. 24, No. 9, 2000
0
140
Days post irradiation
B
DN
A c
onte
nt
per
cell
fra
ctio
n (
arbi
trar
y u
nit
s)
Control
0
100A
Cel
l nu
mbe
r (%
)
Control
20
40
60
80
100
120
2 4 6 8 10 12 14 16 18 20 22
10
20
30
40
50
60
70
80
90
51 9 18Days post irradiation
Fig. 1. A: Histograms of the DNA content showing response of Ramos cell line to a single 10 Gy dose of external beamirradiation (averaged from 6 experiments). Cells have been harvested at the time-point indicated, washed and stained with PI andthen DNA content analysed by flow cytometry (as described in Materials and Methods). The DNA histograms are showing: G2arrest at day 1, postponed apoptosis, depression of the diploid cell line and accumulation of polyploids in the first week (day 5),high level of apoptosis, even more depressed diploid cell line and decrease in % of giant cells during the second week (day 9), andrestoration of the cycling rapidly growing diploid stem-line in the third week (day 18). <2N, �; 2N, ; S-phase, �; 4N, ;>4N, . B. Changes in DNA content of each ploidy fraction (4N fraction is omitted) after irradiation over the 3-week timecourse. The changes are shown for the Namalwa cell line as averages of six experiments and demonstrate: depression of thediploid stem-line from day 1 to 14, step-wise rounds of DNA doubling and increasing ploidy in giant cells during the first week,de-polyploidization during the second week, and steep restoration of the diploid cell line from the end of the second week. DNAcontent (corresponding cellular mass) per cell fraction is represented in arbitrary units and was calculated by multiplying thepercentage of cells in each gated region by the DNA ploidy for that region. 2N, – –�– –; 8N ——; 16N —�—; 32N, —*—;64N, ——.
Sub-nuclear composition of giant cells andautonomy of sub-nuclei
From 48–72 h after irradiation and thereafter, asub-nuclear composition of many polyploid cell
nuclei is apparent and the autonomy of thesesub-nuclei becomes evident. As such, the giantnuclei are often seen to be composed of two ormore lobes or sub-nuclei, which are positionedin close contact. These sub-nuclei can possess
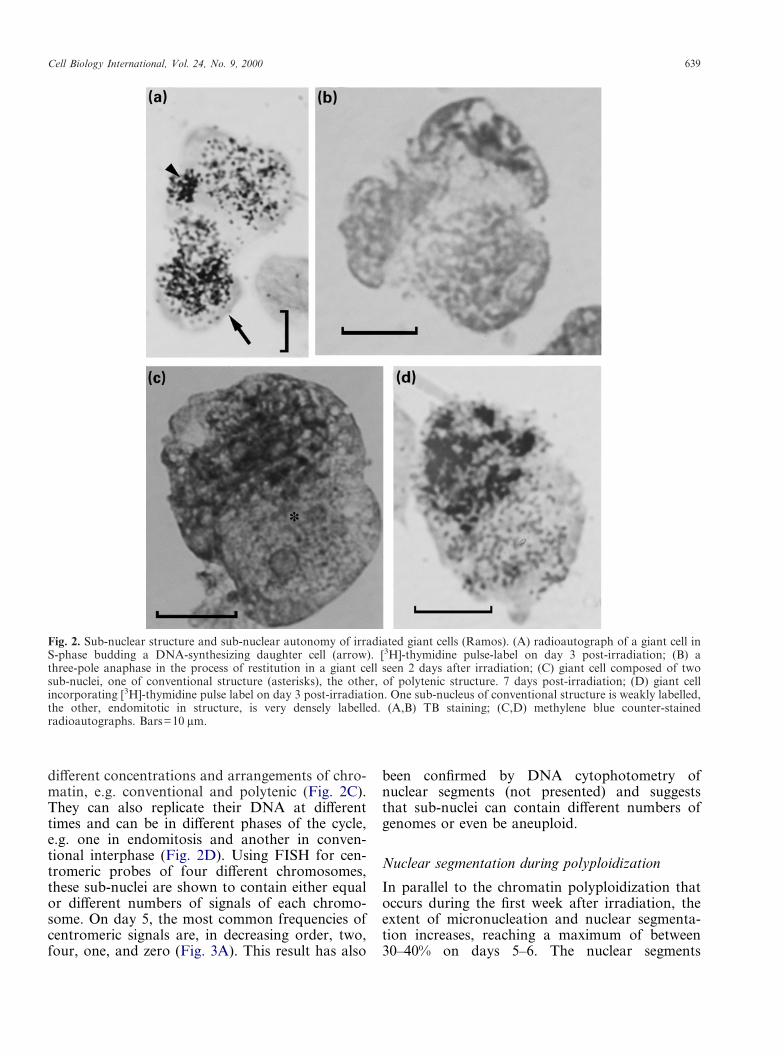
Cell Biology International, Vol. 24, No. 9, 2000 639
different concentrations and arrangements of chro-matin, e.g. conventional and polytenic (Fig. 2C).They can also replicate their DNA at differenttimes and can be in different phases of the cycle,e.g. one in endomitosis and another in conven-tional interphase (Fig. 2D). Using FISH for cen-tromeric probes of four different chromosomes,these sub-nuclei are shown to contain either equalor different numbers of signals of each chromo-some. On day 5, the most common frequencies ofcentromeric signals are, in decreasing order, two,four, one, and zero (Fig. 3A). This result has also
been confirmed by DNA cytophotometry ofnuclear segments (not presented) and suggeststhat sub-nuclei can contain different numbers ofgenomes or even be aneuploid.
Fig. 2. Sub-nuclear structure and sub-nuclear autonomy of irradiated giant cells (Ramos). (A) radioautograph of a giant cell inS-phase budding a DNA-synthesizing daughter cell (arrow). [3H]-thymidine pulse-label on day 3 post-irradiation; (B) athree-pole anaphase in the process of restitution in a giant cell seen 2 days after irradiation; (C) giant cell composed of twosub-nuclei, one of conventional structure (asterisks), the other, of polytenic structure. 7 days post-irradiation; (D) giant cellincorporating [3H]-thymidine pulse label on day 3 post-irradiation. One sub-nucleus of conventional structure is weakly labelled,the other, endomitotic in structure, is very densely labelled. (A,B) TB staining; (C,D) methylene blue counter-stainedradioautographs. Bars=10 �m.
Nuclear segmentation during polyploidization
In parallel to the chromatin polyploidization thatoccurs during the first week after irradiation, theextent of micronucleation and nuclear segmenta-tion increases, reaching a maximum of between30–40% on days 5–6. The nuclear segments
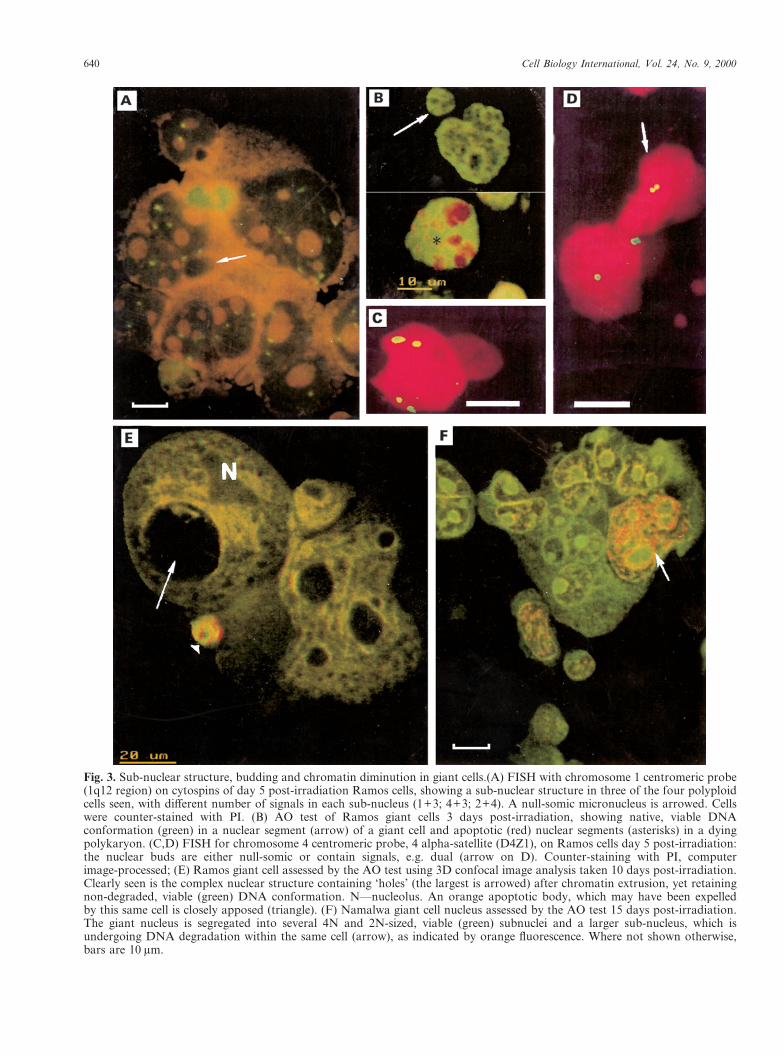
640 Cell Biology International, Vol. 24, No. 9, 2000
Fig. 3. Sub-nuclear structure, budding and chromatin diminution in giant cells.(A) FISH with chromosome 1 centromeric probe(1q12 region) on cytospins of day 5 post-irradiation Ramos cells, showing a sub-nuclear structure in three of the four polyploidcells seen, with different number of signals in each sub-nucleus (1+3; 4+3; 2+4). A null-somic micronucleus is arrowed. Cellswere counter-stained with PI. (B) AO test of Ramos giant cells 3 days post-irradiation, showing native, viable DNAconformation (green) in a nuclear segment (arrow) of a giant cell and apoptotic (red) nuclear segments (asterisks) in a dyingpolykaryon. (C,D) FISH for chromosome 4 centromeric probe, 4 alpha-satellite (D4Z1), on Ramos cells day 5 post-irradiation:the nuclear buds are either null-somic or contain signals, e.g. dual (arrow on D). Counter-staining with PI, computerimage-processed; (E) Ramos giant cell assessed by the AO test using 3D confocal image analysis taken 10 days post-irradiation.Clearly seen is the complex nuclear structure containing ‘holes’ (the largest is arrowed) after chromatin extrusion, yet retainingnon-degraded, viable (green) DNA conformation. N—nucleolus. An orange apoptotic body, which may have been expelledby this same cell is closely apposed (triangle). (F) Namalwa giant cell nucleus assessed by the AO test 15 days post-irradiation.The giant nucleus is segregated into several 4N and 2N-sized, viable (green) subnuclei and a larger sub-nucleus, which isundergoing DNA degradation within the same cell (arrow), as indicated by orange fluorescence. Where not shown otherwise,bars are 10 �m.

Cell Biology International, Vol. 24, No. 9, 2000 641
produced are often released as DNA synthesizingdaughter cells (Fig. 2A), which initially have nativeDNA conformation (Fig. 3B). These nuclear budscontain either one, two, three or often zero signalsof one of the chromosome probes (Fig. 3C,D).Thus, they often contain an incomplete chromo-some set. These cells are distinguishable by apeculiar (‘endocyclic’) chromatin ultrastructure(not shown) and appear to be reproductively dead,since they are never seen to undergo mitosis andinstead experience premature chromosome conden-sation, further segmentation and/or apoptosis.Other researchers (Dini et al., 1996; Shimizu et al.,1998) have arrived at the same conclusion as to thefate of these buds.
Death of giant cells by postponed apoptosis
The majority of giant cells that form die by the endof the first week after irradiation, becoming apop-totic polykaryons which display disconnected sub-nuclei (Fig. 3B). In our experiments, it appearsfrom flow cytometric analysis that after 10 Gy,only 10–20% of giant cells survive the first weekafter irradiation. Thus, we propose that thesecells have in some way been selected by lateapoptosis.
Chromatin diminution during de-polyploidization
The majority of giant cells that have been rescuedfrom death by the end of the first week then appearradically to change their behaviour. First, as judgedby DNA flow cytometry (Fig. 1B) and DNA pulseand pulse-chase radioautography, these cells stoptheir endo-reduplication cycles. Subsequently, aftera few days’ interval, they undergo only a singlefurther round of DNA replication. In addition,during this time these giant cells begin increasinglyto degrade and extrude large portions of geneticmaterial.
Although this process is seen rarely in giant cellsfrom days 5–6, the majority of the giant cells thatsurvive into the second week perform it. Prior toextrusion from the cell, a large number of activemitochondria (and in some cases lysosomes) aregathered near the relevant part of the nucleus (Fig.4A). Next, a portion of the chromatin starts to budfrom the nuclear wall (not shown), or is sequesteredby the nuclear envelope, and extruded into thecytoplasm (Fig. 4A). This transition from nucleusto cytoplasm appears to be very rapid, as we haveseen only rare cases of it, such as that shown inFigure 4A,B. The sequestered chromatin subse-quently undergoes degradation by nucleolysis and
autophagy, followed by active expulsion from thecell.
In many instances, the autophagic digestion isvery prompt and the chromatin is rapidly loosenedand cleared from autophagosomes, leaving behinda vacuole which is usually located in a nuclearinvagination (Figs 3E; 4B,C,E) and/or is opened tothe cell exterior (4B,C). In some cases, the seques-tered nuclear segment is shed out of the mother cellas a sub-cell with a pyknotizing nucleus, character-istically anchored to the mother nucleus (Figs 3E;4B,F), and the formation of an autophagosomemay follow (Fig. 4D). Unexpectedly, after selectivedegradation of its genetic material, the mothergiant cell retains viability (i.e. is capable of furthersynthesizing DNA) and the remaining chromatin isleft intact, as shown by the AO test (Fig. 3E).
On days 12–17 after irradiation, the nucleus andespecially the cytoplasm of giant cells become per-forated by multiple vacuoles. These vacuoles con-tain chromatin remnants, suggesting that theprocess of chromatin diminution by autophagy andextrusion occurs repeatedly and contributes signifi-cantly to the decrease in polyploidy of giant cellsseen by DNA cytophotometry.
The formation of polyploid ‘bouquets’
In the second week post-irradiation, the majority ofgiant cells are seen to undergo a complex nuclearreorganization. Initially, double chromatin loops,fixed by both ends to the nuclear envelope, areformed (Fig. 5A). Simultaneously the whole giantnucleus begins to extend in a polar fashion, form-ing a ‘bouquet’ or ‘parachute’ structure (Fig.5B,D), where both ends of the loop are closely fixedon a narrow pole of the nucleus within a smallcircular area (not shown). This bouquet has fea-tures similar to the specific bouquet structure ofmeiotic prophase. It proceeds through a stage ofthin loops and then thick loops. Our current studiesusing monoclonal antibodies against tubulins showthat initially the bouquet possesses a compositecentrosome at a foot pole and undergoes rotation(manuscipt in preparation). [3H]-thymidine pulse-labelling and chase experiments suggest that thesingle, postponed round of DNA replication in thegiant cells during the second week coincides withthe initial stage of bouquet formation (data notshown).
Final disintegration of giant cells
Simultaneously with the chromatin diminution, thebouquets undergo their final segregation. Initially
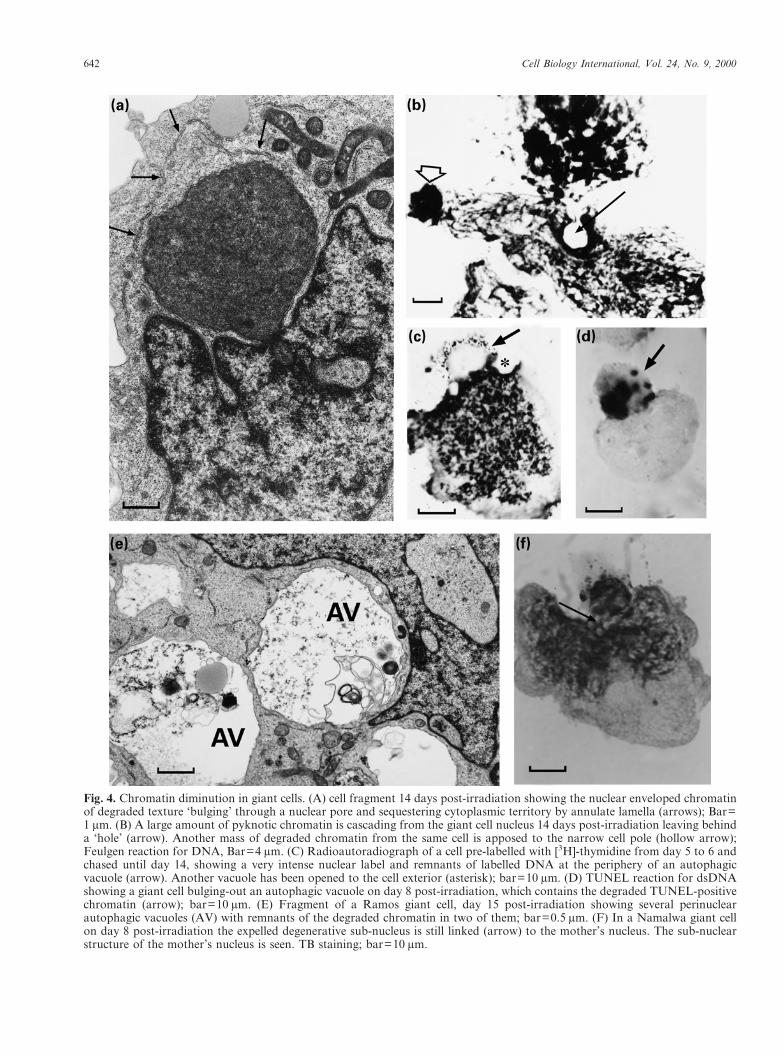
642 Cell Biology International, Vol. 24, No. 9, 2000
Fig. 4. Chromatin diminution in giant cells. (A) cell fragment 14 days post-irradiation showing the nuclear enveloped chromatinof degraded texture ‘bulging’ through a nuclear pore and sequestering cytoplasmic territory by annulate lamella (arrows); Bar=1 �m. (B) A large amount of pyknotic chromatin is cascading from the giant cell nucleus 14 days post-irradiation leaving behinda ‘hole’ (arrow). Another mass of degraded chromatin from the same cell is apposed to the narrow cell pole (hollow arrow);Feulgen reaction for DNA, Bar=4 �m. (C) Radioautoradiograph of a cell pre-labelled with [3H]-thymidine from day 5 to 6 andchased until day 14, showing a very intense nuclear label and remnants of labelled DNA at the periphery of an autophagicvacuole (arrow). Another vacuole has been opened to the cell exterior (asterisk); bar=10 �m. (D) TUNEL reaction for dsDNAshowing a giant cell bulging-out an autophagic vacuole on day 8 post-irradiation, which contains the degraded TUNEL-positivechromatin (arrow); bar=10 �m. (E) Fragment of a Ramos giant cell, day 15 post-irradiation showing several perinuclearautophagic vacuoles (AV) with remnants of the degraded chromatin in two of them; bar=0.5 �m. (F) In a Namalwa giant cellon day 8 post-irradiation the expelled degenerative sub-nucleus is still linked (arrow) to the mother’s nucleus. The sub-nuclearstructure of the mother’s nucleus is seen. TB staining; bar=10 �m.

Cell Biology International, Vol. 24, No. 9, 2000 643
Fig. 5. ‘Bouquet’ formation and segregation in BL cells. (A) EM picture showing chromatin double loops attaching by both endsto the nuclear envelope (arrowheads), characteristic of the ‘bouquet’ arrangement of early meiosis. Sample taken 12 dayspost-irradiation; bar=1 �m. (B–D) Giant ‘bouquets’, 14 days post-irradiation. TB staining; bars=10 �m; (C) An ‘opened’ giantbouquet with clearly visible radial double loops (arrow). (D) Giant ‘parachute’ in the process of return in interphase and splittingin secondary nuclei; the loop structures are arrowed. (E) EM picture of the ordered splitting of the Ramos giant cell nucleusdisplaying a centrosome in the centre of the split (enlarged on insertion); bar=1 �m.
the chromatin loops de-condense and the nucleusreturns to an interphase state (Fig. 5C,D). Thebouquet then splits from the centre of its wider
pole, like a water melon (Fig. 5D,E). Secondarynuclei are yielded, which are located in a commonarea of cytoplasm and are often seen to be

644 Cell Biology International, Vol. 24, No. 9, 2000
Fig. 6. Final disintegration of giant cells into mitosis-competent sub-cells, 14–18 days post-irradiation. (A) Ramos giant cellsegregating a number of sub-nuclei, constricted at a single point (arrow). (B) Namalwa giant cell segregating into severaloligoploid sub-cells; one of them contains one metaphasic and two anaphasic plates. (C) Radioautograph of the Namalwa giantcell segregating into two labelled oligoploid interphase sub-cells and one sub-cell in metaphase. The sample was obtained fromthe giant cell fraction separated on day 5 post-irradiation (as described by Illidge et al., 2000 in this issue), labelled by[3H]-thymidine from day 5 to 6, and chased upon cultivation up to day 17. The vacuoles remaining after chromatin diminution(asterisks) as well as the edge of the common cytoplasm (arrow) are indicative of the origin of these subcells from one giantmother. (D) Three large and one small chromosome plates in a single cell (cytoplasm not counterstained). The mitotic figuresappear to be connected by DNA-containing links (arrow); (A,B) TB staining; (D) Feulgen staining. Bars=10 �m.
constrained to a narrow pole of the formerbouquet (Fig. 6A). While still within the commoncytoplasm, the daughter nuclei asynchronouslybegin to undergo DNA synthesis (not shown) andthis is followed by mitosis. Giant cells separated bylight scattering and labelled by [3H]-thymidinefrom day 5 to 6 and further cultivated (see com-panion article in this issue by Illidge et al., 2000)
show these thymidine-labelled sub-cells includingmitoses (Fig. 6C). Simultaneously, the daughternuclei enclose individual cytoplasmic regionsaround themselves and leave their mother cell asindividual cells. In rarer cases, several chromosomeplates are found simultaneously within the body ofa single oligo-ploid sub-cell (Fig. 6B) or wholegiant cell (Fig. 6D). Furthermore, it seems that in

Cell Biology International, Vol. 24, No. 9, 2000 645
the latter case, the chromosome plates are alsoconnected by the DNA-containing links (Fig. 6D).Often, one or two of the sub-nuclei, usually thosethat are largest and of a coarser structure, appearto stay in place and undergo selective endonucleo-lytic degradation. These can be highlighted withinthe cytoplasm of the giant cell by virtue of theirorange fluorescence using the AO test (Fig. 3F).The other smaller sub-nuclei fluoresce green inthis test, indicative of intact DNA. [3H]-thymidinepulse and pulse-chase experiments show that,unlike the DNA of vital sub-nuclei, the degradingchromatin of the giant cell does not take part inthe DNA replication of the 2nd–3rd week (notshown). This is either extruded or the wholeremnant of the giant cell undergoes degradation.Soon after the beginning of the sub-cellularmitoses, the giant cells disintegrate completely.This coincides with very intense growth of thediploid cell fraction over the next few days asshown by flow cytometry (Fig. 1) and incorpor-ation of [3H]-thymidine DNA pulse label, leadingto an increase in the S-phase index (1 h label) up to50–60%. Together with the presence of largenumbers of mitoses, a high percentage of whichdisplay anaphases and telophases, this indicates therestoration of the stem-line.
DISCUSSION
We have demonstrated with observations over anextended period that after irradiation with a singledose of 10 Gy, giant cells are formed in Ramos andNamalwa BL cell lines that may produce mitoticdescendants. Our observation is contrary to thewidely accepted view that survivors can only arisefrom the mitotic stem-line. Indeed, this was thecase in our experiments with the same cells afterless severe irradiation at 2 and 5 Gy (Cragg et al., inpreparation). However, after genotoxic damage at10 Gy, the giant cells were capable of producing themitotic cells, while the mitotic pathway wasstrongly suppressed, as documented by our cyto-logical and flow cytometric study. Our observa-tions are largely compatible with those of Brodskyand Uryvaeva (1982) and Hall et al. (1996), whodemonstrated that following the initiation of endo-cycles mammalian cells are capable of undergoingmultipolar mitoses; the first authors also indicatedreconstitution of the diploid DNA content. How-ever, this mode of return to the mitotic cycle isnot likely to be the major mechanism at work inour model, where much more complex nuclearrearrangements of giant cells were preceding the
restoration of the mitotic cycle. In addition, wepresent here three novel morphological facets ofmammalian giant cells, which are sub-nuclearautonomy, chromatin diminution, and reconstruc-tion of giant nuclei into ‘bouquets’. Importantly,these bouquets were characteristic only for thosegiant cells which survived into the second weekafter irradiation. We believe that these phenomenamay represent important components of the resti-tution process and that they provide an additionalmechanism of repairing dsDNA breaks.
Prior to the final segregation, the giant cellsdisplayed a major nuclear sorting and selectionprocess. As such, only 10–20% of giant cells surviveinto the second week after irradiation. The selectedcells that survive then begin to sort and expelmasses of degraded chromatin. In contrast toapoptosis of the entire polyploid cell which occursin the first week, this second process represents aselective degradation of distinct parts of the giantcell nucleus. In its biological sense, this represents aprocess of chromatin diminution, as discovered byBovery in 1887 (see review of Wilson, 1925), anddefined as the partial degradation of the geneticmaterial of a living cell. Although selective autoph-agic chromatin degradation of this type hasbeen observed in certain metamorphosinginsects (Bataillon, 1891: cited from Clarke, 1990;Matsuura et al., 1968; Andries et al., 1975) and is away of eliminating the macronucleus in heter-okaryotic protists (reviewed by Raikov, 1982), ithas not previously been reported to our knowledgefor somatic mammalian cells.
Chromatin diminution and the sub-nuclearautonomy and differentiation observed in giantcells early in their development seem very strangeprocesses for a somatic metazoan cell to undertake.Rather, it bears analogy with the behaviour ofsome higher protists. The life cycle of someheterokaryotic protists involves sub-nuclear differ-entiation, endo-reduplication of the DNA in amacronucleus, budding of temporarily viablenuclear segments during the vegetative phase andelimination of this macronucleus in the generativephase of the life cycle (Raikov, 1982; Prescott,1994; Donhoff and Klein, 1996). Similarity inthe behaviour of tumour and Tetrahymenacells in the phenomenon of nuclear buddingwas also noticed by Shimizu et al. (1998). Alsosimilar to our observations with irradiated tumourcells, in Tetrahymena this selective degenerationevidently combines the features of autophagy(Levy and Elliott, 1968) and apoptosis (Mpokeand Wolfe, 1996, 1997) and is preceded bydisruption of DNA replication (Raikov, 1982).

646 Cell Biology International, Vol. 24, No. 9, 2000
Furthermore, cycling polyploidization (followed byde-polyploidization) of the primary nucleus ischaracteristic of the asexual life-cycle of Radiolaria(Raikov, 1982).
After leaving cell cycle arrest at G2, the irradi-ated tumour cells show aberrant mitoses and it isclear that the giant cells that form must harbourunrepaired dsDNA. In turn, the restoration ofmitotic potency within giant cells should mean thatthis broken DNA has been repaired. Non-homologous end joining represents the primarymechanism of dsDNA repair in mammals. How-ever, this process is prone to error. The pathwaycapable of repairing dsDNA with greatest effi-ciency is homologous recombination between adamaged DNA molecule and an undamagedhomologous partner (Friedberg et al., 1995).Intriguingly, recent data indicate that this processcan take place in mammalian cells (Liang et al.,1998) and is enhanced soon after irradiation (Haafet al., 1995). In addition, it has been shown thatp53 mutant tumours have an increased ability torepair in this manner (Xia et al., 1994).
In irradiated tumour cells, we see rapid initiationof endocycles, but simple multiplication of brokenchromatids during these endo-reduplication cycleswould not provide an undamaged template forhomologous recombination. Therefore, only twoprocesses involving polyploid giant cells could inprincipal be capable of providing repair: (1) cellfusion and exchange or sorting of parental chromo-somes, assembling an ‘assorted’ non-damagedgenome(s) or, (2) a process similar to meiosisinvolving pairing and exchange of homologouschromosomes, providing undamaged DNAtemplates for homologous recombination.
The first possibility is rather improbable as manydata, and indirectly our own, suggest that eachgenome persists within a giant cell as a physicallyassembled supra-chromosomal entity (Raikov,1982). Therefore the second possibility, somaticreduction, seems more likely. Somatic reductionhas also been termed ‘meiosis without karyogamy’(Raikov, 1982). In this context, a third phenom-enon should be considered, which is also novel andunusual for a somatic metazoan cell, that is rear-rangement of giant cell nuclei into ‘bouquets’. Thebouquet structure is a very specific chromosomearrangement of meiotic prophase, which is neededfor, and involved in, chromosome pairing andreciprocal genetic recombination (Shertan et al.,1996). In the asexual life cycle of Radiolaria,the pairing and recombination of homologouschromosomes was found to be involved inthe so called ‘segregation of genomes’ from
polygenomic cells (Grell, 1953), which is a kindof somatic reduction. In our polygenomic gianttumour cells this would correspond to de-polyploidization. A sub-nuclear theory explainingde-polyploidization of polygenomic nuclei inprotists was proposed by Sonneborn (1947) over50 years ago and was later confirmed (Raikov1982), who said that each sub-nucleus of a giantcell contains an individual genome(s), which isitself capable of independent reproduction. Thesub-nuclear structure and autonomy of DNA-replicative activity, as well as the asynchronousreturn to the mitotic cycle in separate sub-nuclei inour giant tumour cells, seems highly analogouswith this behaviour.
In the context of the final segregation of gianttumour cells into mitosis-competent survivors, thisgenetic re-organization process could potentiallyprovide repair of the damaged DNA and faithfulsegregation of genetic material into secondarynuclei. The process of somatic reduction was con-sidered by Nagl (1978) as a theoretically possibleway for an endopolyploid nucleus to give rise to amitotic diploid cell.
In conclusion, a working hypothesis can be putforward which suggests that the response of p53mutant tumour cells to genotoxic insult is that, ifrepair is not accomplished through the mitoticcycle, it can occur by an additional rescue mechan-ism via giant cells. This novel mechanism involvesformation and de-polyploidization of giant cells,which may provide a basis for true repair ofdsDNA and return to the mitotic cell cycle. Inmany aspects, it mirrors the life-cycle program ofprotozoa.
ACKNOWLEDGEMENTS
We would like to commemorate our discussionswith the late Prof. Igor Raikov and to thank DrThomas Haaf and Prof. Ivor Bowen for theircomments on the manuscript. We acknowledge thetechnical skills of Ms Sue Cox, Mr Nick Barnet,Mr Roger Alston (Southampton General Hospital)and Mr Vladislavs Zelchs (Latvian University).The assistance of Dr Galina Selivanova and MrVladimir Timoshevsky is acknowledged. Finally,Prof. Sergei A. Nazarenko is thanked for providingthe image analysis system for fluorescencemicroscopy of FISH. The work in Latvia waspartially funded by the Latvian State Program onMalignant Tumours. M. Cragg was funded by theTENOVUS charity and T. Illidge was supported bythe Cancer Research Campaign, U.K.

Cell Biology International, Vol. 24, No. 9, 2000 647
REFERENCES
A J-C, 1975. Differenciation et mort cellulaire au coursde la metamorphose mesenterique de la larve d’Aeshnacynea. J Microsc Biol Cell 4: 327–350.
B EJ, M RJ, B VJ, MKWG, 1996. Reducing the radiation-induced G2-delay causesHeLa cells to undergo apoptosis instead of mitotic death. IntJ Radiat Biol 69: 575–584.
B VY, U IV, 1981. Genome Multiplication inGrowth and Development. Cambridge, Cambridge UniversityPress.
C PGH, 1990. Developmental cell death: morphologicaldiversity and multiple mechanisms. Anat & Embryol 181:190–213.
C M-S, Z L, F R-R, G MJ, 1999.Analysis of the interaction of monoclonal antibodies withsurface IgM on neoplastic B cells. Brit J Cancer 79: 850–857.
C SHM, S C-A, M C-A, S M-K,R S, I R-L, R W-H, R BJ, 1995. Ap53-dependent mouse spindle checkpoint. Science 26: 1353–1356.
D Z, 1994. Acid-induced denaturation of DNAin situ as a probe of chromatin structure. Meth Cell Biol 41:527–541.
D L, C S, R MT, G L, 1996. Multiplepathways for apoptotic nuclear fragmentation. Exp Cell Res223: 340–347.
D T, K A, 1996. Timing of differential amplifica-tion of macronucleus destined sequences during macronu-clear development in the hypotrichous ciliate. Chromosoma105: 172–179.
E J, F T, R H, A R, 1997.Apoptotic cell nuclei favour aggregation and fluorescencequenching of DNA dyes. Histochem & Cell Biol 108: 67–75.
F EC, W GC, S W, 1995. Mutagenesis inEukaryotes. In: DNA Repair and Mutagenesis. AmericanSociety for Microbiology, Washington, DC, 662–669.
G Y, S Y, B-S SA, 1992. Identificationof programmed cell death in situ via specific labelling ofnuclear DNA fragmentation. J Cell Biol 119: 493–501.
G G, 1998. Cell cycle regulation of DNA replication. Theendoreduplication perspective. Mini-review. Exp Cell Res244: 372–378.
G DT, B WP, H M, H CC,1994. Mutations in the p53 tumour suppressor gene: Cluesto cancer etiology and molecular pathogenesis. Cancer Res54: 4855–4878.
G KG, 1953. Die Chromosomen von Aulacantha scoly-mantha Haeckel. Arch Protist 99: 1–54.
H T, G EI, R G, R CM, W DC,1995. Nuclear foci of mammalian Rad51 recombinationprotein in somatic cells after DNA damage and its localiza-tion in synaptonemal complexes. Proc Nat Acad Sci USA92: 2298–2302.
H LL, T’ JPH, C XW, T RL, B EM,1996. A brief Staurosporine treatment of mitotic cellstriggers premature exit from mitosis and polyploid cellformation. Cancer Res 56: 3551–3559.
I TM, C MS, F B, O P, E J,2000. Polyploid giant cells provide a survival mechanism forp53 mutant cells after DNA damage. Cell Biol Int 24:621–633.
L AL, 1997. p53, the cellular gatekeeper for growth anddivision. Cell 88: 323–331.
L MR, E AM, 1968. Biochemical and ultrastructuralchanges in Tetrahymena pyriformis during starvation. JProtozool 15: 208–222.
L FM, H M, R PJ, J M, 1998.Homology-directed repair is a major double-strand breakrepair pathway in mammalian cells. Proc Nat Acad Sci USA95: 5172–5177.
L RB, R WE, 1990. Possible role for p34cdc2kinase inEtoposide-induced cell death of Chinese hamster ovary cells.Cancer Res 50: 3767–377.
M S, M T, N SPJ, T Y, 1968.Studies on the posterior silk gland of the silkworm, Bombyxmori. II. Cytolytic processes in posterior silk gland cellsduring metamorphosis from larva to pupa. J Cell Biol 38:589–603.
M AJ, T DA, P CHS, H JA, 1997.Apoptosis in small intestinal epithelia from p53-null mice:evidence for a delayed, p53-independent G2/M-associatedcell death after irradiation. Oncogene 14: 2759–2766.
M S, W J, 1996. DNA digestion and chromatincondensation during nuclear death in Tetrahymena. Exper-imental Cell Research 225: 357–365.
M SS, W J, 1997. Differential staining of apoptoticnuclei in living cells: Application to Macronuclear Elimina-tion in Tetrahymena. J Histochem Cytochem 45: 675–684.
N W, 1995. Cdc2-kinases, cyclins, and the switch fromproliferation to polyploidization. Protoplasma 188: 143–150.
N W, 1978. Endopolyploidy and Polyteny in Differentiationand Evolution. Amsterdam, North-Holland.
O’C PM, J J, J D, B K, MJ, K KW, 1993. Role of the p53 tumour suppressor genein cell cycle arrest and radiosensitivity of Burkitt’slymphoma cell lines. Cancer Res 5: 4776–4780.
O PL, B JP, D RE, 1996. Development ofapoptosis and polyploidy in human lymphoblast cells as afunction of the cell cycle at the time of irradiation. RadiationRes 146: 595–602.
P A, S D, E NB, W R, L R,R V, 1996. The role of p53 in the induction ofpolyploidity of myelomonocytic leukemic M1/2 cells. Onco-gene 13: 1677–1685.
P DM, 1994. The DNA of ciliated protozoa. MicrobiolRev, 58: 233–267.
R IB, 1982. The Protozoan Nucleus. Morphology andEvolution. Wien–New York, Springer Verlag.
R HI, E JA, A T, 1995. Osteogenicdifferentiation of hypertrophic chondrocytes involves asym-metric cell divisions and apoptosis. J Cell Biol 131: 483–494.
R DE, C BH, 1992. In: Human cytogen-etics. A practical approach. V.1. Oxford, Oxford Univ. Press.91–117.
S HS, W H, S H, H C, H M,C T, 1996. Centromere and telomere movementsduring early meiotic prophase of mouse and man areassociated with the onset of chromosome pairing. J Cell Biol134: 1109–1125.
S N, I N, U H, W GM, 1998. Selectiveentrapment of extrachromosomally amplified DNA bynuclear budding and micronucleation during S-phase. J CellBiol 140: 1307–1320.
S T, 1947. Recent advances in the genetics ofParamecium and Euplotes. Advances in Genet 1: 263–358.
W KF, L H, H P, 1985. Double-strandbreak repair in G2 block in Chinese hamster ovary cells andtheir radiosensitive mutants. Mutat Res 145: 177–183.

648 Cell Biology International, Vol. 24, No. 9, 2000
W EB, 1925. The Cell in Development and Heredity.Russian edition (1936). Moscow-Leningrad, State Publ.Biol. & Med. Liter.
X F, A SA, N JA, L HL, 1994.Different capacities for recombination in closely related
human lymphoblastoid cell-lines with different muta-tional responses to X-irradiation. Mol Cell Biol 14: 5850–5857.
Copyright © 2022 FDOKUMEN




















