
Reissner’s fiber formation depends on developmentally regulated factors extrinsic to the...
13
Cell Tissue Res (2005) 321: 429–441 DOI 10.1007/s00441-004-1040-1 REGULAR ARTICLE C. Hoyo-Becerra . M. D. López-Avalos . M. Alcaide-Gavilán . M. C. Gómez-Roldán . J. Pérez . P. Fernández-Llebrez . J. M. Grondona Reissner’s fiber formation depends on developmentally regulated factors extrinsic to the subcommissural organ Received: 31 August 2004 / Accepted: 29 October 2004 / Published online: 7 July 2005 # Springer-Verlag 2005 Abstract Reissner ’ s fiber (RF) is a threadlike structure present in the third and fourth ventricles and in the central canal of the spinal cord. RF develops by the assembly of glycoproteins released into the cerebrospinal fluid (CSF) by the subcommissural organ (SCO). SCO cells differen- tiate early during embryonic development. In chick em- bryos, the release into the CSF starts at embryonic day 7 (E7). However, RF does not form until E11, suggesting that a factor other than release is required for RF formation. The aim of the present investigation was to establish whether the factor(s) triggering RF formation is (are) intrinsic or extrinsic to the SCO itself. For this purpose, SCO explants from E13 chick embryos (a stage at which RF has formed) were grafted at two different developmental stages. After grafting, host embryos were allowed to survive for 6– 7 days, reaching E9 (group 1) and E13 (group 2). In ex- perimental group 1, the secretion released by the grafted SCOs never formed a RF; instead, it aggregated as a floc- culent material. In experimental group 2, grafted SCO explants were able to develop an RF-like structure, similar to a control RF. These results suggest that the factor trig- gering RF formation is not present in the SCO itself, since E13 SCO secretion forms an RF in E13 brains but never develops RF-like structures when placed in earlier devel- opmental environments. Furthermore, the glycoproteins released by implanted SCOs bind specifically to sev- eral structures: the apical portion of the mesencephalic floor plate and the choroid plexus of the third and fourth ventricles. Keywords Subcommissural organ . Reissner ’ s fiber . Isograft . Cerebrospinal fluid . Chick embryo (White Leghorn) Introduction The subcommissural organ (SCO) is an ependymal differ- entiation located in the roof plate, midline of the caudal- most portion of the diencephalon (prosomere 1), under the posterior commissure. The SCO is an ancient and phylo- genetically conserved structure, present throughout the vertebrate phyla (Oksche 1961). SCO ependymal cells are specialized in the synthesis and secretion of glycoproteins of high molecular mass that are released into the cerebro- spinal fluid (CSF), where they aggregate on top of the cilia of the SCO cells. Subsequently, this thin film over the cilia becomes further packed in an highly ordered fashion to form a threadlike supramolecular structure known as Reissner ’ s fiber (RF; Oksche 1969; Rodriguez et al. 1992, 1998; Meiniel 2001; Nualart and Hein 2001). In the encephalon, glycoproteins forming RF can be found in two conforma- tions: (1) the material aggregated over the SCO cilia (the so-called pre-RF) and (2) the RF proper, which is a cylin- drical regular structure. RF grows caudally by the addition of SCO glycoproteins at its cephalic end and extends along the brain aqueduct, the fourth ventricle and the central canal of the spinal cord (Sargent 1900; Sterba et al. 1967; Leonhardt 1980; Rodriguez et al. 1992). The SCO develops early during ontogeny (Oksche 1956; Naumann 1986; Schoebitz et al. 1986; Naumann et al. 1987). The embryonic development of the SCO-RF com- plex has been studied in rat (Castaneyra-Perdomo et al. 1983; Marcinkiewicz and Bouchaud 1983, 1986; Naumann et al. 1987), rabbit (Kimble and Mollgard 1975), chicken (Wingstrand 1953; Schoebitz et al. 1986; Naumann et al. C. Hoyo-Becerra and M. D. López-Avalos contributed equally to this study and should be considered as joint first authors. C. Hoyo-Becerra was the recipient of a predoctoral fellowship (PFPI) from the Ministerio de Ciencia y Tecnología (Spain). This work was supported by grants from DGICYT (BFI2003-03348; Spain) and FIS (01/0948; Spain), FIS (01–0948, PI021517; Spain) and ISCIII (red CIEN, nodo Fundación Carlos Haya). C. Hoyo-Becerra . M. D. López-Avalos . M. Alcaide-Gavilán . M. C. Gómez-Roldán . J. Pérez . P. Fernández-Llebrez . J. M. Grondona (*) Departamento de Biología Celular, Genética y Fisiología, Facultad de Ciencias, Campus de Teatinos, Universidad de Málaga, 29071 Málaga, Spain e-mail: [email protected] Tel.: +34-952-134113 Fax: +34-952-0132000
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Reissner’s fiber formation depends on developmentally regulated factors extrinsic to the...

Cell Tissue Res (2005) 321: 429–441DOI 10.1007/s00441-004-1040-1
REGULAR ARTICLE
C. Hoyo-Becerra . M. D. López-Avalos .M. Alcaide-Gavilán . M. C. Gómez-Roldán . J. Pérez .P. Fernández-Llebrez . J. M. Grondona
Reissner’s fiber formation depends on developmentally regulatedfactors extrinsic to the subcommissural organ
Received: 31 August 2004 / Accepted: 29 October 2004 / Published online: 7 July 2005# Springer-Verlag 2005
Abstract Reissner’s fiber (RF) is a threadlike structurepresent in the third and fourth ventricles and in the centralcanal of the spinal cord. RF develops by the assembly ofglycoproteins released into the cerebrospinal fluid (CSF)by the subcommissural organ (SCO). SCO cells differen-tiate early during embryonic development. In chick em-bryos, the release into the CSF starts at embryonic day 7(E7). However, RF does not form until E11, suggesting thata factor other than release is required for RF formation. Theaim of the present investigation was to establish whetherthe factor(s) triggering RF formation is (are) intrinsic orextrinsic to the SCO itself. For this purpose, SCO explantsfrom E13 chick embryos (a stage at which RF has formed)were grafted at two different developmental stages. Aftergrafting, host embryos were allowed to survive for 6–7 days, reaching E9 (group 1) and E13 (group 2). In ex-perimental group 1, the secretion released by the graftedSCOs never formed a RF; instead, it aggregated as a floc-culent material. In experimental group 2, grafted SCOexplants were able to develop an RF-like structure, similarto a control RF. These results suggest that the factor trig-gering RF formation is not present in the SCO itself, sinceE13 SCO secretion forms an RF in E13 brains but neverdevelops RF-like structures when placed in earlier devel-opmental environments. Furthermore, the glycoproteins
released by implanted SCOs bind specifically to sev-eral structures: the apical portion of the mesencephalicfloor plate and the choroid plexus of the third and fourthventricles.
Keywords Subcommissural organ . Reissner’s fiber .Isograft . Cerebrospinal fluid . Chick embryo (WhiteLeghorn)
Introduction
The subcommissural organ (SCO) is an ependymal differ-entiation located in the roof plate, midline of the caudal-most portion of the diencephalon (prosomere 1), under theposterior commissure. The SCO is an ancient and phylo-genetically conserved structure, present throughout thevertebrate phyla (Oksche 1961). SCO ependymal cells arespecialized in the synthesis and secretion of glycoproteinsof high molecular mass that are released into the cerebro-spinal fluid (CSF), where they aggregate on top of the ciliaof the SCO cells. Subsequently, this thin film over the ciliabecomes further packed in an highly ordered fashion to forma threadlike supramolecular structure known as Reissner’sfiber (RF; Oksche 1969; Rodriguez et al. 1992, 1998;Meiniel 2001; Nualart and Hein 2001). In the encephalon,glycoproteins forming RF can be found in two conforma-tions: (1) the material aggregated over the SCO cilia (theso-called pre-RF) and (2) the RF proper, which is a cylin-drical regular structure. RF grows caudally by the additionof SCO glycoproteins at its cephalic end and extends alongthe brain aqueduct, the fourth ventricle and the centralcanal of the spinal cord (Sargent 1900; Sterba et al. 1967;Leonhardt 1980; Rodriguez et al. 1992).
The SCO develops early during ontogeny (Oksche 1956;Naumann 1986; Schoebitz et al. 1986; Naumann et al.1987). The embryonic development of the SCO-RF com-plex has been studied in rat (Castaneyra-Perdomo et al.1983; Marcinkiewicz and Bouchaud 1983, 1986; Naumannet al. 1987), rabbit (Kimble and Mollgard 1975), chicken(Wingstrand 1953; Schoebitz et al. 1986; Naumann et al.
C. Hoyo-Becerra and M. D. López-Avalos contributed equally tothis study and should be considered as joint first authors.C. Hoyo-Becerra was the recipient of a predoctoral fellowship(PFPI) from the Ministerio de Ciencia y Tecnología (Spain). Thiswork was supported by grants from DGICYT (BFI2003-03348;Spain) and FIS (01/0948; Spain), FIS (01–0948, PI021517; Spain)and ISCIII (red CIEN, nodo Fundación Carlos Haya).
C. Hoyo-Becerra . M. D. López-Avalos . M. Alcaide-Gavilán .M. C. Gómez-Roldán . J. Pérez . P. Fernández-Llebrez .J. M. Grondona (*)Departamento de Biología Celular, Genética y Fisiología,Facultad de Ciencias, Campus de Teatinos,Universidad de Málaga,29071 Málaga, Spaine-mail: [email protected].: +34-952-134113Fax: +34-952-0132000

1987; Karoumi et al. 1990; Meiniel et al. 1993) andhumans (Oksche 1956; Wislocki and Roth 1958; Oksche1961; Olsson 1961; Møllgard 1972; Rodriguez et al. 2001).By using antibodies raised against bovine RF (Rodriguezet al. 1984) and recognizing chick SCO secretion, Schoebitzet al. (1986, 1993) have performed a detailed immuno-cytochemical study of chick SCO development. With re-spect to secretory material synthesis, ventricular release,assembly and RF formation, these authors have establishedthat, in the chick embryo, the SCO secretory material isfirst synthesized at embryonic day 3 (E3) by morphologi-cally undifferentiated neuroepithelial cells in the dorsaldiencephalon. At E7, the SCO starts to release, into theCSF, glycoproteins that aggregate as pre-RF onto thesurface of the neuroepithelium. No RF proper has beendescribed until E11, at which time it appears in the cerebralaqueduct. At E12, the RF is present in the lumbar spinalcord. The elapsed time between the ventricular release ofthe secretion and RF formation has led Schoebitz et al.(1986) to conclude that factors other than ventricular re-lease must be required for the formation of the RF. Thisdelay in RF formation during fetal life has also beenreported in rat development (Schoebitz et al. 1993).
The mechanisms that trigger RF formation remainunknown. However, evidence has been obtained suggest-ing that both the hydrodynamics of the CSF (Olsson 1958;Oksche 1961; Rodriguez et al. 1989; Irigoin et al. 1990)and an unknown CSF factor (Schoebitz et al. 1986, 1993)are involved in such process. In order to improve ourunderstanding of the assembly of RF, transplants of SCOinto the lateral ventricle of adult rats have been performed(Rodriguez et al. 1999): isografts (rat SCO into rat brainventricles) are able to form RF even in the lateral ventricle,whereas xenografts (bovine SCO into rat brain ventricles)never produce an RF, despite releasing secretory materialinto the CSF. These results suggest that (1) the hydrody-namics of the CSF are not essential for RF formation, and(2) the putative CSF factor controlling this process is spe-cies-specific (Rodriguez et al. 1999).
The existence of a species-specific CSF factor involvedin RF formation in adult rat, as proposed by Rodriguezet al. (1999) and the presence of putative factor(s) induc-ing the RF formation at E11 in chick embryos (Schoebitzet al. 1986) have led us to question whether such a factortriggering the embryonic RF is intrinsic (i.e., is producedby the SCO or is dependent on the molecular nature of thesecretion) or extrinsic to the SCO itself. The present in-vestigation has been designed to gain new insights intoRF assembly during embryonic development and to de-termine whether intrinsic factor(s) is (are) involved in thisassmebly. For this purpose, we have grafted SCO explantsfrom E13 chick donor embryos, which are able to formRF, into younger host chick embryos subsequently al-lowed to survive until E9 or E13. These two groups ofembryos have been used to study the assembly of secre-tory material released by the grafted SCO explants in thedifferent developmental environments present in E9 andE13 brains.
Materials and methods
Animals
Fertilized eggs from White Leghorn chicken (Gallusdomesticus) were obtained from a local farm (GranjaSanta Isabel, Córdoba) and incubated in a forced-draftincubator at 38.5°C with high humidity. Embryos werestaged according to Hamburger and Hamilton (Hamburgerand Hamilton 1951). The animals used in this study werehandled in conformity with the current national legislation(decree 223/1988. BOE no. 67) and guidelines from theCouncil of the European Communities (86/609/CEE).
Dissection and grafting techniques
In order to obtain E13 SCO explants, chick brains from thisdevelopmental stage were dissected out under a dissectingmicroscope (MZ6; LeicaMicrosystems, Wetzlar, Germany)under sterile conditions. E13 SCOs were isolated fromsurrounding tissue by using fine tweezers and scissors;however, the posterior commissure could not be completelyremoved. The collected SCO explants were kept in Hanks’solution (Sigma, St. Louis, Mo., USA) for no longer than30 min before grafting them into host chick embryos. TheseE13 SCO explants were used for the two recipient groupsdescribed below.
In experimental group 1, E3 (H/H18) host embryos weregrafted with 2–3 E13 SCO explants introduced into theventricular cavity of the optic tectum through a small in-cision made with fine steel needles in the lateral tectum.Most explants remained free in the CSF of this ventricularcavity. After the surgery, the eggs were sealed with ad-hesive tape and kept in the incubator for 6 further days toobtain E9 embryos (H/H35–H/H36) (Fig. 1a). In experi-mental group 2, E6 (H/H28–H/H29) host embryos weregrafted with 3–4 E13 SCO explants in the optic tectumventricular cavity as described above. The eggs were sealedand incubated for 7 days until they reached E13 (H/H39)(Fig. 1a). In both experimental groups, about 70% of theembryos survived the manipulations. At the end of theincubation period, 12 E9 embryos (group 1) and 16 E13embryos (group 2) were fixed and processed for histolog-ical analysis.
Histology and immunocytochemistry
At the end of the incubation period, complete heads of E9chick embryos (experimental group 1) were skinned andfixed by immersion in Bouin’s fluid for 48 h at room tem-perature. Brains from E13 embryos (experimental group 2)were removed and fixed as for the E9 embryos. The tissueswere dehydrated and embedded in paraffin wax. Trans-verse sections (10 μm thick) were obtained on a microtome(RM2125; Leica Microsystems) and processed accordingto the immunoperoxidase method of Sternberger et al.(1970) with some modifications. Briefly, sections were first
430
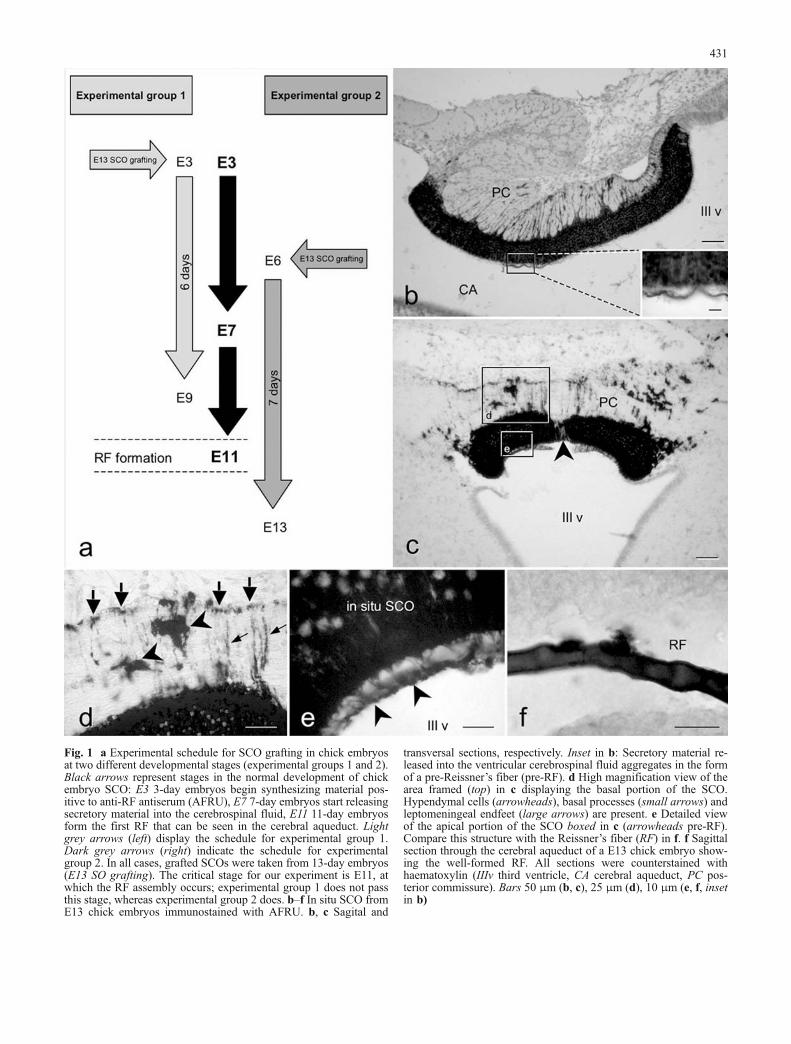
Fig. 1 a Experimental schedule for SCO grafting in chick embryosat two different developmental stages (experimental groups 1 and 2).Black arrows represent stages in the normal development of chickembryo SCO: E3 3-day embryos begin synthesizing material pos-itive to anti-RF antiserum (AFRU), E7 7-day embryos start releasingsecretory material into the cerebrospinal fluid, E11 11-day embryosform the first RF that can be seen in the cerebral aqueduct. Lightgrey arrows (left) display the schedule for experimental group 1.Dark grey arrows (right) indicate the schedule for experimentalgroup 2. In all cases, grafted SCOs were taken from 13-day embryos(E13 SO grafting). The critical stage for our experiment is E11, atwhich the RF assembly occurs; experimental group 1 does not passthis stage, whereas experimental group 2 does. b–f In situ SCO fromE13 chick embryos immunostained with AFRU. b, c Sagital and
transversal sections, respectively. Inset in b: Secretory material re-leased into the ventricular cerebrospinal fluid aggregates in the formof a pre-Reissner’s fiber (pre-RF). d High magnification view of thearea framed (top) in c displaying the basal portion of the SCO.Hypendymal cells (arrowheads), basal processes (small arrows) andleptomeningeal endfeet (large arrows) are present. e Detailed viewof the apical portion of the SCO boxed in c (arrowheads pre-RF).Compare this structure with the Reissner’s fiber (RF) in f. f Sagittalsection through the cerebral aqueduct of a E13 chick embryo show-ing the well-formed RF. All sections were counterstained withhaematoxylin (IIIv third ventricle, CA cerebral aqueduct, PC pos-terior commissure). Bars 50 μm (b, c), 25 μm (d), 10 μm (e, f, insetin b)
431
incubated in phosphate-buffered saline (PBS) containing20% methanol and 5% hydrogen peroxide (Merck,Darmstadt, Germany) to inactivate endogenous peroxidase.After being rinsed in PBS with 0.05% Tween-20 (Sigma),the sections were incubated in the primary antibody for18 h. The secondary antibody, anti-rabbit IgG, raised insheep in our laboratory, was used at a dilution of 1:20 for30 min, followed by rabbit PAP (Sigma), at a dilution of
1:400 for 30 min. Peroxidase activity was detected byincubating the tissue sections for 5 min in TRIS-bufferedsaline (50 mM TRIS-HCl, 137 mM NaCl), pH 7.8, con-taining 0.1% 3,3′ diaminobenzidine (DAB, Sigma) and0.03% hydrogen peroxide (Merck). The reaction wasstopped by several washes with distilled water. After im-munostaining, sections were counterstained with haemat-oxylin solution (Panreac, Barcelona, Spain). The primary
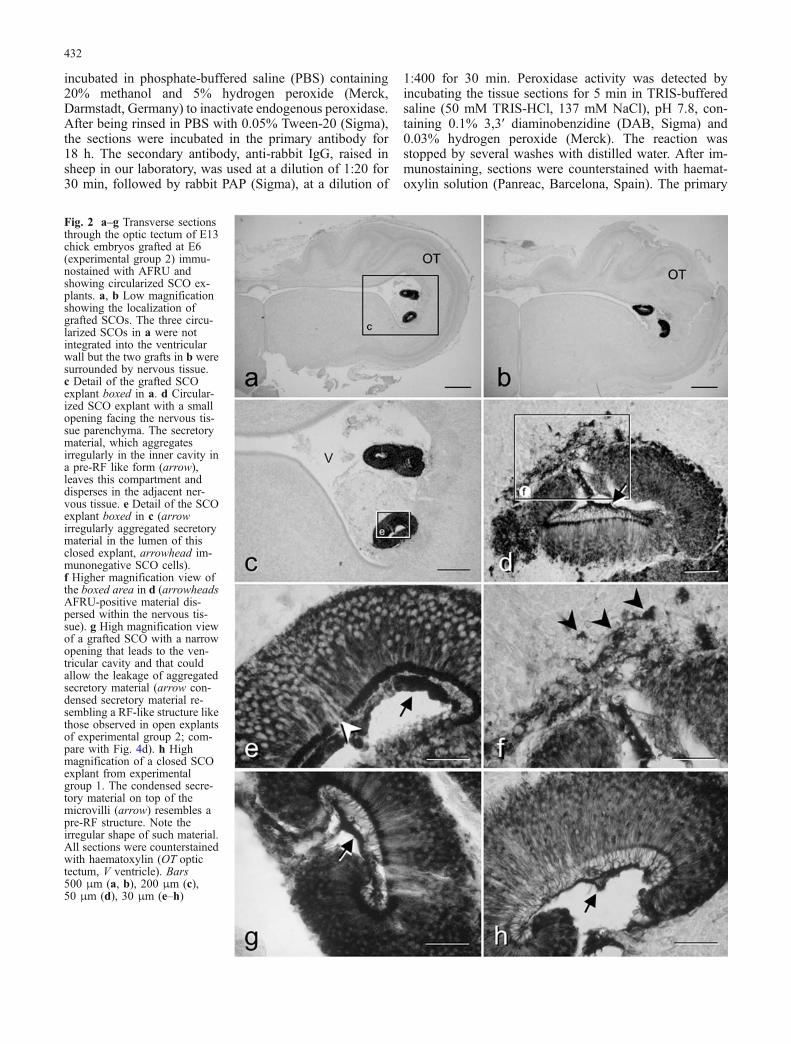
Fig. 2 a–g Transverse sectionsthrough the optic tectum of E13chick embryos grafted at E6(experimental group 2) immu-nostained with AFRU andshowing circularized SCO ex-plants. a, b Low magnificationshowing the localization ofgrafted SCOs. The three circu-larized SCOs in a were notintegrated into the ventricularwall but the two grafts in b weresurrounded by nervous tissue.c Detail of the grafted SCOexplant boxed in a. d Circular-ized SCO explant with a smallopening facing the nervous tis-sue parenchyma. The secretorymaterial, which aggregatesirregularly in the inner cavity ina pre-RF like form (arrow),leaves this compartment anddisperses in the adjacent ner-vous tissue. e Detail of the SCOexplant boxed in c (arrowirregularly aggregated secretorymaterial in the lumen of thisclosed explant, arrowhead im-munonegative SCO cells).f Higher magnification view ofthe boxed area in d (arrowheadsAFRU-positive material dis-persed within the nervous tis-sue). g High magnification viewof a grafted SCO with a narrowopening that leads to the ven-tricular cavity and that couldallow the leakage of aggregatedsecretory material (arrow con-densed secretory material re-sembling a RF-like structure likethose observed in open explantsof experimental group 2; com-pare with Fig. 4d). h Highmagnification of a closed SCOexplant from experimentalgroup 1. The condensed secre-tory material on top of themicrovilli (arrow) resembles apre-RF structure. Note theirregular shape of such material.All sections were counterstainedwith haematoxylin (OT optictectum, V ventricle). Bars500 μm (a, b), 200 μm (c),50 μm (d), 30 μm (e–h)
432

antibody used in this study was a rabbit polyclonal anti-serum against bovine RF (AFRU, 1:1,000; Rodriguezet al. 1984) kindly provided by Prof. Rodriguez (Valdivia,Chile). This antiserum recognizes chick SCO secretorymaterial and has been previously used to study the em-bryonic development of chick RF (Schoebitz et al. 1986).All antisera were diluted in PBS, pH 7.2, containing 10%normal goat serum and 0.3% Triton X-100 (Sigma). Allincubations with antisera (primary and secondary) and PAPwere performed in a moist chamber at room temperature.Controls for immunostaining included incubation of sec-tions with (1) non-immune rabbit serum or (2) diluentbuffer, instead of the primary antiserum. No immunostain-ing was observed in such controls. Images were taken witha Leica DMLB photomicroscope and a Nikon Coolpix 990digital camera. Colour images were processed with AdobePhotoshop 5.0 to obtain black and white images.
Results
Morphology of in situ SCO in E13 embryos
The structure of the SCO of E13 embryos has been thor-oughly described elsewhere (Schoebitz et al. 1986). Here,we highlight several morphological features of E13 in situSCO to provide a better understanding of the alterationsthat it undergoes after grafting.
At E13, SCO cells were organized into two cell layers:ependymal and hypendymal. Ependymal cells were ar-ranged as a tall-stratified epithelium with basal processesthat ended on the external basal membrane (large arrows inFig. 1d). Hypendymal cells were spread under the epen-dymal layer, some of them lying within the posteriorcommissure (arrowheads in Fig. 1d). Both cell types werestrongly immunoreactive to the antiserum against thebovine RF (AFRU), with the exception of a small group ofcells that lay in the midline of the ependymal layer and thatwas devoid of immunoreactive material as could be seen intransverse sections through E13 SCO (arrowhead in Fig. 1c).Such non-immunoreactive cells were observed in previousstages of development (not shown) and in the grafted SCOs(arrowheads in Figs. 2e, 3b). Another remarkable feature ofthe E13 in situ SCO was its prominent secretory activity, asrevealed by the presence of a thin layer of AFRU-immu-noreactive material lying on the surface of the ependymalcells (Fig. 1b, c, e) and, most importantly, the capacity ofthat material to aggregate into the form of an RF (Fig. 1f).
Morphology and secretory activity of grafted SCOs
All the grafted SCOs survived the surgical dissection, theimplantation in the optic tectum and the 6–7 days ofincubation period until the time of sacrifice. Most of thetransplanted SCOs appeared to be attached to the optictectum ventricular wall (Figs. 2c, 4a, e) or, less frequently,in the fourth ventricle (Fig. 3i–k). In some cases, theywere partially or totally integrated into such brain areas
(Figs. 2b, d, 3a, b). Almost 60% of the grafted SCOs tookon a circularized conformation with the apical surfacefacing a small internal cavity that usually was not incommunication with the ventricular space (Fig. 2a–c).The ependymal layer of the grafted SCO kept the sametall-stratified epithelium arrangement as the in situ SCO,although the basal processes became shorter or weremissing (Figs. 2e, 3j, 4c). However, the hypendymal layerbecame disorganized and disappeared in most cases. Themorphology of the ependymal cells in the grafted SCOswas similar to that of E13 in situ SCO. Such cellsdisplayed profuse immunoreactivity to the AFRU. ThisAFRU-positive material filled most of the cell cytoplasm,being more abundant in the apical and perinuclear regions(Figs. 2e, g, 3j, 4c).
The grafted SCOmaintained its capacity to release immu-noreactive material, as evidenced by the AFRU-positivematerial that could be found on the apical surface of thetransplanted SCO and in its surroundings (Figs. 2e, g, h, 3c,j, 4b, d, f).
The mode of aggregation of the glycoproteins releasedby the grafted SCOs depended on the structural configu-ration adopted by the graft. As mentioned above, two con-formations were observed: (1) circularized inwards, and(2) with the apical surface open to the ventricular cavity. Inthe two experimental groups, viz. incubation until devel-opmental stage E9 or until E13, the secretion released bycircularized SCOs appeared to be assembled similarly.However, when the apical surface of the grafted SCO facedthe ventricular cavity, the assembly of the released materialdiffered considerably between the experimental groups, asdescribed below.
Aggregation of secretory material in circularizedSCO explants
As mentioned above, the closed circular conformation wasfound in both experimental groups. In the inner cavity ofsuch explants, the released secretory material aggregatedon top of the microvilli in the form of AFRU-immunore-active material of variable thickness and irregular shape(arrows in Fig. 2e, g, h), sometimes resembling a pre-RF(Fig. 2h). In a few cases, the lumen of the circularized SCOwas partially open to the ventricular cavity (Fig. 2g). Inthese situations, small amounts of secretion left the innercavity, passing into the CSF of the optic tectum ventricularspace in the form of small irregular fibrillar aggregates (notshown). In other cases, in which the opening faced theventricular wall, these immunoreactive aggregates werefound immersed the nearby nervous tissue (arrowheads inFig. 2f). Even though no conspicuous differences wereobserved in the mode of aggregation in such circularizedexplants, we noticed that, in some partially opened explantsfrom experimental group 2 (embryos kept until E13), theassembled secretion appeared to be more regularly orga-nized than that in closed SCO explants, resembling the RF-like structures observed in the open explants (comparearrow in Fig. 2g with arrowheads in Fig. 4d). Apart from
433

434

this exception, we never observed structures similar to anRF in circularized explants, irrespective of the experi-mental group.
Aggregation of secretory material released by graftedSCOs with the apical portion open to the ventricularcavity
The assembly pattern of the secretory material releasedby the grafted SCOs that adopted an open conformationclearly differed in the E9 embryos of experimental group 1and in the E13 embryos of group 2.
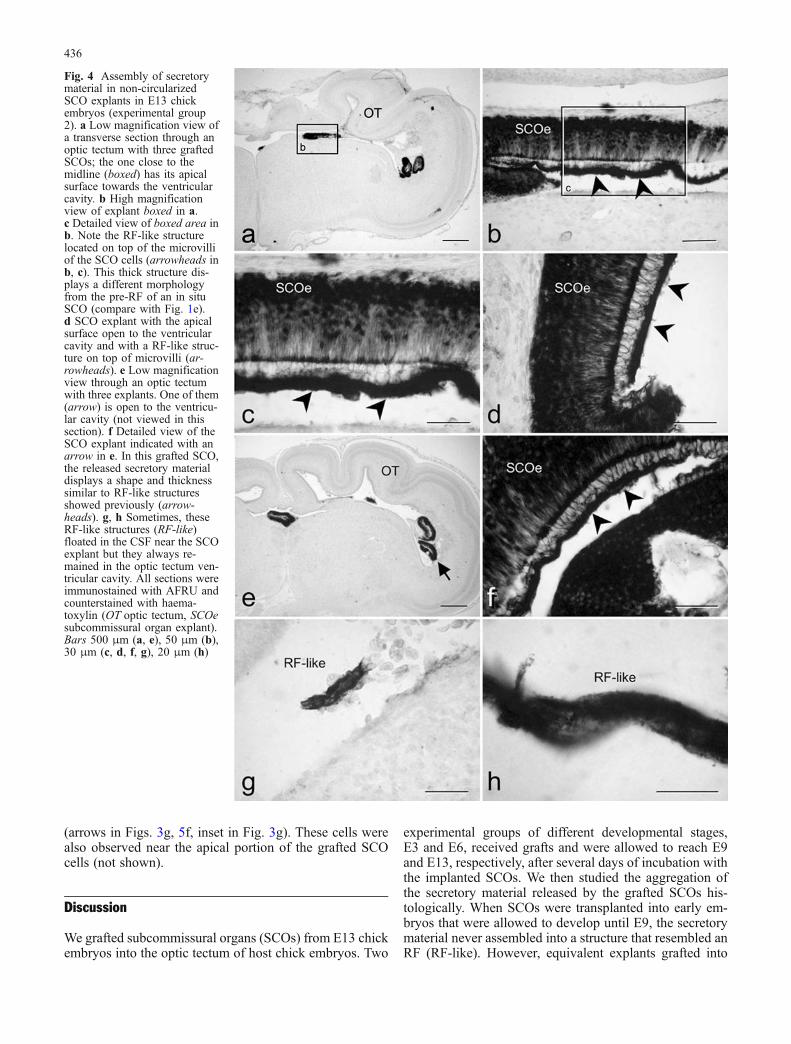
Assembly in experimental group 1
In this group, E3 embryos were grafted with E13 SCOs andmaintained until E9, a stage in which the RF is not yetformed in non-grafted control chick embryos. The secre-tion released into the CSF by the grafted SCOs aggregatedin the form of small immunoreactive particles that weredispersed in the CSF (Figs. 3b, c, inset in e) or attached tothe ventricular surface in the vicinity of the graft (arrow-heads in Fig. 3d, e). Sometimes, these particles clumpedtogether and appeared as large immunoreactive structuresdisseminated in the fourth ventricle (asterisks in Figs. 3f–h).In addition to this flocculent material, these embryos dis-played AFRU-positive aggregates in the form of smallfibrils similar to those forming the pre-RF. Such structureswere also produced by the in situ SCO at E9. We could notfind a structure with the morphological appearance of an
RF or an RF-like structure in any of the sections analysedfrom embryos in this experimental group, even at the apicalsurface of the grafted SCOs (compare Fig. 3i–k withFig. 4c, d, f).
Assembly in experimental group 2
Embryos in this group reached E13, a stage in which awell-organized RF can be observed in non-grafted controlembryos. In this E13 ventricular environment, the materialreleased by the grafted SCO aggregated to form an AFRU-positive cylindrical structure (8 μm of diameter) similar tothat of an RF proper (Fig. 4c, d, f–h). Nonetheless, suchstructures did not display the perfect cylindrical shape ofthe normal RF (compare Fig. 1f with Fig. 4g); in addition,its homogeneous AFRU-immunostaining pattern was dif-ferent from that seen in the RF proper, which showedstrong immunoreactivity at the periphery and weakerimmunostaining at the core (compare Fig. 1f with Fig. 4c,d, f). For these reasons, the AFRU-positive aggregateswere termed “RF-like” structures. They could be found ontop of the microvilli of the grafted SCOs (Fig. 4c, d, f) andin other optic tectum ventricular areas (Fig. 4g, h). Usually,the RF-like structures were present as small fragments ofabout 50 μm (Fig. 4b), although occasionally they reachedapproximately 200 μm (Fig. 4d). Moreover, in some em-bryos, irregular masses of immunoreactive material (50–100 μm in diameter) were seen in the fourth ventricle(Fig. 5g) suggesting that this could be the final fate of theRF-like material (see below).
In experimental group 2, the SCO proper was able toform its own RF, which appeared to be normal in structureand immunostaining pattern (data not shown). This RFreached the fourth ventricle but we could not verify itspresence in the central canal of the spinal cord, as we didnot study this area. In addition, the in situ SCO in bothexperimental groups had a histologically and immunocy-tochemically normal appearance compared with that ofnon-grafted embryos (data not shown).
AFRU-positive material on the mesencephalic floorplate and choroid plexus in grafted embryos
AFRU demonstrated the presence of immunoreactivematerial on the apical surface of the mesencephalic floorplate (Fig. 5c, d) and in the choroid plexus of the third(Fig. 5a, b) and fourth (Fig. 5e) ventricles. Although thisfeature was previously described in non-grafted embryos(Schoebitz et al. 1986), we found that it was more con-spicuous in transplanted embryos, particularly in thosewith the so-called “open” explants, irrespective of the ex-perimental group. These mesencephalic floor plate cellswith apical extracellular immunoreactive material also dis-played a weak dispersed cytoplasmic immunoreactivity(not shown).
In both experimental groups, numerous cells containingAFRU-positive material were present free in the CSF
3Fig. 3 Transverse sections through the optic tectum (a–e) andfourth ventricle (f–k) of E9 chick embryos grafted at E3 with E13SCOs (experimental group 1). All sections were immunostainedwith AFRU. Only embryos with non-circularized SCOs explants(open explants) are shown. a Low magnification view of the optictectum with an open explant partially integrated into the nervoustissue but with its apical surface towards the ventricular cavity.b Detail of the graft shown in a (arrowhead unstained region). Notethe AFRU-positive material dispersed in the optic tectum ventricularcavity. c Higher magnification of the optic tectum ventricular areaboxed in b. d Detailed view of boxed area in c. AFRU-positivesecretory material aggregates in the form of small irregular sizedspheres (arrowheads). e High magnification of boxed area inc. Similar aggregates stick to the apical neuroepithelium (arrow-heads) or bind to each other and remain free in the cerebrospinal fluid(CSF) as flocculent material (see inset, which corresponds to a highermagnification of the boxed area in the middle of c). f–h AFRU-positive structures (white asterisks) in the CSF of the fourth ventricle,composed of flocculent material probably released by the graftedSCOs. Note the cells containing AFRU-positive material (arrows ing). Inset in g: Higher magnification of these immunoreactive cells.i SCO explant located in the fourth ventricle and with the apicalportion towards the ventricular cavity. j High magnification of thearea boxed in i (arrowheads AFRU-positive material close to theapical portion of the grafted SCO). k Grafted SCO with its apicalportion towards the ventricular cavity. In this case, the secretorymaterial aggregates as a pre-RF that does not form a RF-likestructure. All sections were counterstained with haematoxylin (OToptic tectum, IV v fourth ventricle, SCOe subcommissural organexplant). Bars 500 μm (a), 100 μm (b, i), 50 μm (c), 20 μm (f, g,j, k), 10 μm (d, e, h), 5 μm (insets)
435
(arrows in Figs. 3g, 5f, inset in Fig. 3g). These cells werealso observed near the apical portion of the grafted SCOcells (not shown).
Discussion
We grafted subcommissural organs (SCOs) from E13 chickembryos into the optic tectum of host chick embryos. Two
experimental groups of different developmental stages,E3 and E6, received grafts and were allowed to reach E9and E13, respectively, after several days of incubation withthe implanted SCOs. We then studied the aggregation ofthe secretory material released by the grafted SCOs his-tologically. When SCOs were transplanted into early em-bryos that were allowed to develop until E9, the secretorymaterial never assembled into a structure that resembled anRF (RF-like). However, equivalent explants grafted into
Fig. 4 Assembly of secretorymaterial in non-circularizedSCO explants in E13 chickembryos (experimental group2). a Low magnification view ofa transverse section through anoptic tectum with three graftedSCOs; the one close to themidline (boxed) has its apicalsurface towards the ventricularcavity. b High magnificationview of explant boxed in a.c Detailed view of boxed area inb. Note the RF-like structurelocated on top of the microvilliof the SCO cells (arrowheads inb, c). This thick structure dis-plays a different morphologyfrom the pre-RF of an in situSCO (compare with Fig. 1e).d SCO explant with the apicalsurface open to the ventricularcavity and with a RF-like struc-ture on top of microvilli (ar-rowheads). e Low magnificationview through an optic tectumwith three explants. One of them(arrow) is open to the ventricu-lar cavity (not viewed in thissection). f Detailed view of theSCO explant indicated with anarrow in e. In this grafted SCO,the released secretory materialdisplays a shape and thicknesssimilar to RF-like structuresshowed previously (arrow-heads). g, h Sometimes, theseRF-like structures (RF-like)floated in the CSF near the SCOexplant but they always re-mained in the optic tectum ven-tricular cavity. All sections wereimmunostained with AFRU andcounterstained with haema-toxylin (OT optic tectum, SCOesubcommissural organ explant).Bars 500 μm (a, e), 50 μm (b),30 μm (c, d, f, g), 20 μm (h)
436
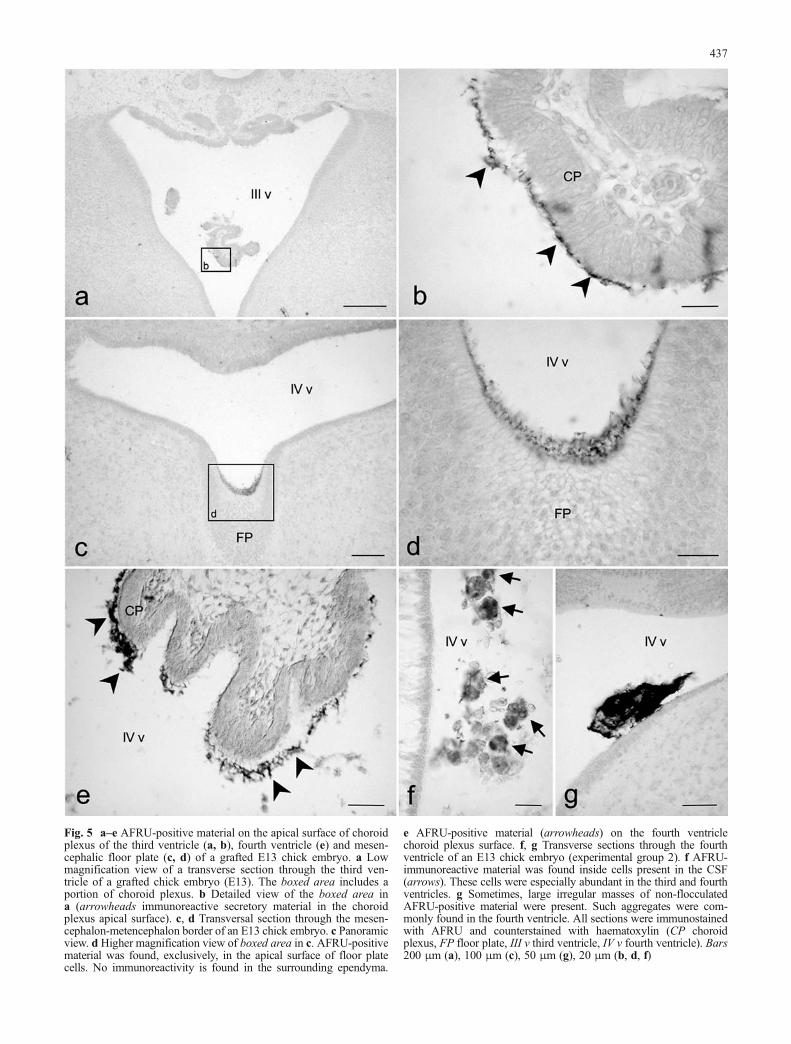
Fig. 5 a–e AFRU-positive material on the apical surface of choroidplexus of the third ventricle (a, b), fourth ventricle (e) and mesen-cephalic floor plate (c, d) of a grafted E13 chick embryo. a Lowmagnification view of a transverse section through the third ven-tricle of a grafted chick embryo (E13). The boxed area includes aportion of choroid plexus. b Detailed view of the boxed area ina (arrowheads immunoreactive secretory material in the choroidplexus apical surface). c, d Transversal section through the mesen-cephalon-metencephalon border of an E13 chick embryo. c Panoramicview. d Higher magnification view of boxed area in c. AFRU-positivematerial was found, exclusively, in the apical surface of floor platecells. No immunoreactivity is found in the surrounding ependyma.
e AFRU-positive material (arrowheads) on the fourth ventriclechoroid plexus surface. f, g Transverse sections through the fourthventricle of an E13 chick embryo (experimental group 2). f AFRU-immunoreactive material was found inside cells present in the CSF(arrows). These cells were especially abundant in the third and fourthventricles. g Sometimes, large irregular masses of non-flocculatedAFRU-positive material were present. Such aggregates were com-monly found in the fourth ventricle. All sections were immunostainedwith AFRU and counterstained with haematoxylin (CP choroidplexus, FP floor plate, III v third ventricle, IV v fourth ventricle). Bars200 μm (a), 100 μm (c), 50 μm (g), 20 μm (b, d, f)
437

older embryos and maintained until E13 gave rise to RF-like structures. These results suggest that the factor trig-gering RF formation is not present in the SCO itself but isfound in the embryonic ventricular CSF microenvironmentand might be regulated during development.
Secretory activity of the grafted SCOs
All transplanted SCOs survived the surgical procedure andgrafting period (6–7 days). The ependymal layer kept itsoriginal arrangement, although almost 60% of the explantscircularized. The ependymal cells of the grafted SCOsdisplayed the morphology and AFRU immunoreactivitysimilar to those of ependymal cells of E13 in situ SCOs.Various experimental strategies, such as in vitro SCOculture (Lehmann and Sterba 1989, 1993; Lehmann et al.1989, 1993; Schoebitz et al. 2001) and SCO grafting(Rodriguez et al. 1989, 1999; Chouaf et al. 1991), havebeen designed to gain new insights regarding the releaseand aggregation of secretory material of SCOs placed in aforeign environment. In our study, several data suggest thatthe grafted SCO releases its secretion into the CSF: (1) incircularized closed explants, the internal lumen containsabundant extracellular immunoreactive material, (2) inopen explants, the apical extracellular surface of the graftedSCO is covered with AFRU-positive material, (3) in theseopen explants, the secretory material could also be found inthe CSF and nearby the explant and, (4) the binding of SCOsecretory material to specific brain areas increases in thosecases of grafted embryos with open explants.
A remarkable feature of our grafted SCO was the processof circularization. Bovine SCO explants from adult ani-mals (Lehmann et al. 1993; Lehmann and Sterba 1993;Cifuentes et al. 1995; Schoebitz et al. 2001) acquire around configuration, but with the apical membrane ofependymal cells towards the external surface of the explant.However, our embryonic explants adopted an inwards cir-cular conformation. Schoebitz et al. (2001) suggests that thecircularization could be attributable to: (1) rearrangement ofthe ependymal layer and/or (2) growth involving mitoticactivity.
Assembly of secretory material releasedby grafted SCOs
Several studies with SCO grafts have been previouslyperformed and the assembly of secretory material hasbeen extensively described (Rodriguez et al. 1989, 1999;Chouaf et al. 1991). The first attempt to graft rat SCOsinvolved their transplantation under the rat kidney cap-sule (Rodriguez et al. 1989). In that experimental model,the SCO cells released AFRU-positive material into aninner cavity formed after grafting; however, such mate-rial did not form a RF. In a more recent work specifical-ly designed to study the factors affecting RF assembly,Rodriguez et al. (1999) carried out isografts and xeno-
grafts, implanting SCO explants from adult rat or adultbovine into adult rat brains. The results from these exper-iments led the authors to conclude that a species-specificCSF factor, and not only CSF hydrodynamics, triggered RFassembly (Rodriguez et al. 1999).
In our experimental design, the host animals were chickembryos at various developmental stages. During normalchick development, the SCO starts releasing secretorymaterial at E7 but the RF does not appear until E11(Schoebitz et al. 1986; Meiniel et al. 1993; Schoebitz et al.1993) suggesting that factors other than release are requiredfor RF polymerization. Based on these previous data, wehave grafted SCOs from E13 embryos, which have an RF,into the tectum of chick embryos at two different devel-opmental stages: (1) E3 embryos that, 6 days after im-plantation, reach E9, which represents an “immature”environment for RF formation, and (2) E6 embryos that,7 days later, reach E13, which can be considered a“mature” environment for RF formation.
We have observed remarkable differences between thetwo experimental groups with respect to the assembly ofthe material released by open SCO grafts, viz. those ex-plants that adopt a conformation with the apical surfacetowards the CSF of the tectum ventricular cavity. In E9embryos, the material released by the graft aggregates inthe form of small AFRU-positive particles or flocculentmaterial that is dispersed throughout the ventricles. An RF-like structure has never been seen under this experimentalcondition. Conversely, the material released by the graftedSCOs in E13 embryos forms an RF-like structure. Theseresults suggest that the assembly of secretory material intothe form of an RF-like structure depends on a factor that isnot present at E9 but that exists at E13. In addition, thisfactor is probably not produced by the SCO itself, sincegrafted E13 SCOs, which are “mature” enough to develop aRF, do not do so when placed in an “immature” environ-ment (E9 embryos).
The nature of the factor(s) triggering the RF assembly isunknown. Post-release processing of secretory material byextracellular proteases has been suggested because bovineRF contains components (electrophoretic bands) that arenot present in bovine SCO extracts (Nualart et al. 1991;Rodriguez et al. 1992). This idea is supported by the exis-tence of numerous proteases that are developmentallyregulated and that have implicated in the control of ex-tracellular matrix composition during brain ontogenesis(Sheffield and Graff 1991; Vaillant et al. 1999; Yong et al.2001). The recent finding, in chick CSF, of a protein frac-tion whose concentration changes during embryonic devel-opment (Gato et al. 2004) further supports this possibility.
The formation of an RF-like structure rather than anormal RF, as takes place in rat SCO isograft experiments(Rodriguez et al. 1999), indicates that, even in E13 em-bryos (“mature” environment), some factor or conditionthat normally occurs in SCO in situ is missing in graftedSCOs. The most obvious difference is the explant location,which would alter the CSF hydrodynamic forces that nor-mally act over the in situ SCO. Although such a factor does
438

not seem to affect RF assembly in isografted rat SCOs(Rodriguez et al. 1999), it might be an important factor inour grafted E13 chick embryos.
Despite the final fate of the newly generated RF-likestructures being unknown, the presence of irregular massesof AFRU–positive material in the fourth ventricle demon-strates that it reaches caudal areas of the ventricular system.Unlike the findings of other authors (Rodriguez et al.1990), we have not found immunoreactive material withinthe CNS neuropil.
On the other hand, the existence of unusual flocculentmaterial in our E9 embryos could be the result of alteredmolecular assembly. Similar aggregates occur in the ven-tricles of rats injected with neuraminidase (a glycosidasethat removes the terminal sialic acid of secretory glyco-proteins) into the CSF (Grondona et al. 1998). Anotherpossible explanation could be the different CSF hydrody-namic force operating on the SCO in situ (cerebral aque-duct) and on the grafted SCO (optic tectum). In accordance,the RF-like structures formed in E13 transplanted embryosare not as regular in shape as the RF proper. However,such flocculent material might correspond to CSF-solu-ble AFRU-positive glycoproteins (Irigoin et al. 1990;Lehmann and Sterba 1993; Rodriguez et al. 1993;Schoebitz et al. 1993) that are released by the graftedSCOs and that precipitate during fixation.
In circularized explants, the secretory material releasedinto the small inner lumen aggregates in a completelydifferent way from that observed in open explants. Thispeculiar assembly is similar in closed explants from bothexperimental groups and consists of irregular structuresof variable thickness that resemble a pre-RF on top of themicrovilli. The different behaviour of the secretion en-trapped within the closed explants could have severalnon-excluding explanations: (1) the putative CSF factorpromoting RF-like structures in group 2 does not reachthe inner cavity; (2) there is no CSF circulation; (3) themicroenvironment in the small lumen may avoid RF-likeassembly and (4) the limited physical space of the innercavity does not allow the RF to grow and achieve itscorrect assembly. The finding that a few partially openedexplants in experimental group 2 form an aggregate thatresembles an RF-like structure supports the above-sug-gested explanations.
Specific binding of AFRU-positive secretory materialto the mesencephalic floor plate and choroid plexus
Immunoreactive material was revealed by AFRU on theapical surface of the mesencephalic floor plate and thechoroid plexus of the third and fourth ventricles in graftedchick embryos. Several authors have reported a transientRF immunoreactivity during embryonic development infloor plate cells of the mesencephalon, metencephalon andspinal cord in various vertebrate species (Schoebitz et al.1993; Fernandez-Llebrez et al. 1996; Rodriguez et al.1996; Lopez-Avalos et al. 1997; Yulis et al. 1998;Lichtenfeld et al. 1999; del Brio et al. 2000, 2001; Richter
et al. 2001; Guinazu et al. 2002). Some of these authorshave shown that the RF immunostaining within floor platecells is confined to the supranuclear and apical regions. Thepossibility that this material is released into the CSF hasbeen proposed by various workers (Rodriguez et al. 1996;Yulis et al. 1998; Guinazu et al. 2002). Moreover,Schoebitz et al. (1986) and del Brio et al. (2000) havedescribed aggregates of AFRU-immunoreactive materialon the free surface of the floor plate at the mesencephalic-metencephalic boundary of chick embryos.
In the present study, several observations suggest that theimmunoreactive material found in the apical portion ofboth floor plate and choroidal cells in grafted embryos isnot released by these cells: (1) almost no immunoreactivityhas been detected inside the cells displaying apical AFRU-positive material, (2) even though this extracellular immu-noreactivity has been found in non-grafted embryos, it ismore conspicuous in transplanted embryos, mainly in thosewith “open” explants, indicating that some of this materialcomes from the grafted SCO. Hence, if floor plate andchoroid plexus cells do not synthesize and release theapical AFRU-positive material, a specific binding of suchglycoproteins to these cells might be suggested. Further-more, Miranda et al. (2001) have reported the binding ofradiolabelled RF glycoproteins perfused into CSF to themesencephalic floor plate cells. The same authors havefound AFRU-positive material attached to choroidal cellsin the following situations: (1) rat SCO grafted into lateralventricle of rat brain (this increases the secretion of ma-terial into the CSF) and (2) in vitro co-culture experimentsof SCO and choroid plexus explants. In addition, an anti-idiotypic antibody raised against anti-RF monoclonalantibodies binds to choroidal cells (Miranda et al. 2001).All of the above results and our own evidence stronglysuggest the existence of specific binding sites for RF gly-coproteins in the mesencephalic floor plate and in thechoroid plexus cells, although the functional meaning ofthese binding sites is unknown.
References
Castaneyra-Perdomo A, Meyer G, Ferres-Torres R (1983) De-velopment of the subcommissural organ in the albino mouse(a Golgi study). J Hirnforsch 24:363–370
Chouaf L, Didier-Bazes M, Hardin H, Aguera M, Fevre-MontangeM, Voutsinos B, Belin MF (1991) Developmental expression ofglial markers in ependymocytes of the rat subcommissuralorgan: role of the environment. Cell Tissue Res 266:553–561
Cifuentes M, Rodriguez EM, Hernandez S, Perez J, Peruzzo B,Perez-Figares JM, Fernandez-Llebrez P (1995) Antibody-me-diated lysis of the bovine subcommissural organ maintained inculture. Exp Brain Res 107:9–51
Brio MA del, Riera P, Munoz RI, Montecinos H, Rodriguez EM(2000) The metencephalic floor plate of chick embryos ex-presses two secretory glycoproteins homologous with the twoglycoproteins secreted by the subcommissural organ. Histo-chem Cell Biol 113:415–426
Brio MA del, Riera P, Peruzzo B, Rodriguez EM (2001) Hindbrainfloor plate of the rat: ultrastructural changes occurring duringdevelopment. Microsc Res Tech 52:615–626
439

Fernandez-Llebrez P, Lopez-Avalos MD, Mota MD, Cifuentes M,Andrades JA, Grondona JM, Perez J, Perez-Figares JM,Rodriguez EM (1996) Secretory glycoproteins of the roof andfloor plates and their rostral derivatives, the subcommissuraland flexural organs, in the developing central nervous system ofvertebrates. An immunocytochemical study. Int J Dev BiolSuppl 1:151S-152S
Gato A, Martin P, Alonso MI, Martin C, Pulgar MA, Moro JA(2004) Analysis of cerebro-spinal fluid protein composition inearly developmental stages in chick embryos. J Exp Zool AComp Exp Biol 301:280–289
Grondona JM, Perez-Martin M, Cifuentes M, Perez J, Estivill-Torrus G, Perez-Figares JM, Fernandez-Llebrez P, RodriguezEM (1998) Neuraminidase injected into the cerebrospinal fluidimpairs the assembly of the glycoproteins secreted by the sub-commissural organ preventing the formation of Reissner’s fiber.Histochem Cell Biol 109:391–398
Guinazu MF, Richter HG, Rodriguez EM (2002) Bovine floor plateexplants secrete SCO-spondin. Cell Tissue Res 308:177–191
Hamburger V, Hamilton H (1951) A series of normal stages in thedevelopment of the chick embryo. J Morphol 88:49–91
Irigoin C, Rodriguez EM, Heinrichs M, Frese K, Herzog S, OkscheA, Rott R (1990) Immunocytochemical study of the sub-commissural organ of rats with induced postnatal hydroceph-alus. Exp Brain Res 82:384–392
Karoumi A, Croisille Y, Croisille F, Meiniel R, Belin MF, Meiniel A(1990) Glycoprotein synthesis in the subcommissural organ ofthe chick embryo. II. An immunochemical study. J NeuralTransm Gen Sect 80:203–212
Kimble JE, Mollgard K (1975) Subcommissural organ-associatedneurons in fetal and neonatal rabbit. Cell Tissue Res 159:195–204
Lehmann W, Sterba G (1989) Tissue culture of rat subcommissuralorgan in vitro. Biomed Res 10(Suppl 30):11–18
Lehmann W, Sterba G (1993) The subcommissural organ in vitro.In: Oksche A, Rodriguez EM, Fernandez-Llebrez P (eds) Thesubcommissural organ. Springer, Berlin Heidelberg New York,pp 133–140
Lehmann W, Sterba G, Wobus AM (1989) Primary culture of themouse subcomissural organ in vitro. Acta Zool 70:199–203
Lehmann W, Naumann W, Wagner U (1993) Tissue culture ofbovine subcommissural organ. Anat Embryol 187:505–514
Leonhardt H (1980) Ependym und circumventrikuläre Organe. In:Oksche A, Vollrath L (eds) Neuroglia I (Handbuch der M-ikroskopischen Anatomie des Menschen, vol IV, part 10).Springer, Berlin Heidelberg New York, pp 176–665
Lichtenfeld J, Viehweg J, Schützenmeister J, Naumann WW (1999)Reissner’s substance expressed as a transient pattern in ver-tebrate floor plate. Anat Embryol 200:161–174
Lopez-Avalos MD, Cifuentes M, Grondona JM, Miranda E, Perez J,Fernandez-Llebrez P (1997) Rostral floor plate (flexural organ)secretes glycoproteins immunologically similar to subcommis-sural organ glycoproteins in dogfish (Scyliorhinus canicula)embryos. Brain Res Dev Brain Res 102:69–75
Marcinkiewicz M, Bouchaud C (1983) The ependymal secretion ofthe fetal and adult rat subcommissural organ. Morphologicalaspects linked to the synthesis, storage and release of the se-cretory products. Biol Cell 48:47–52
Marcinkiewicz M, Bouchaud C (1986) Formation and maturation ofaxo-glandular synapses and concomitant changes in the targetcells of the rat subcommissural organ. Biol Cell 56:57–65
Meiniel A (2001) SCO-spondin, a glycoprotein of the subcommis-sural organ/Reissner’s fiber complex: evidence of a potentactivity on neuronal development in primary cell cultures. Mi-crosc Res Tech 52:484–495
Meiniel R, Didier R, Molat JL, Meiniel A (1993) Developmentalaspects of the subcommissural organ: an approach using lectinsand monoclonal antibodies. In: Oksche A, Rodriguez EM,Fernandez-Llebrez P (eds) The subcommissural organ. Spring-er, Berlin Heidelberg New York, pp 51–59
Miranda E, Almonacid JA, Rodriguez S, Perez J, Hein S, CifuentesM, Fernandez-Llebrez P, Rodriguez EM (2001) Searching forspecific binding sites of the secretory glycoproteins of the sub-commissural organ. Microsc Res Tech 52:541–551
Møllgard K (1972) Histochemical investigations on the humanfoetal subcommissural organ. I. Carbohydrates and mucosub-stances, proteins and nucleoproteins, esterase, acid and alkalinephosphatase. Histochemie 32:31–48
Naumann W (1986) Immunhistochemische Untersuchungen zurOntogenese des Subcommissuralorgans. Acta Histochem Suppl33:265–272
Naumann W, Müller G, Kloss P (1987) Immunoreactive glycopro-teins of the subcommissural organ in the embryonic stages ofthe vertebrate brain. Wiss Z Karl-Marx-Univ Leipzig Math-Naturwiss R 36:17–20
Nualart F, Hein S (2001) Biosynthesis and molecular biology of thesecretory proteins of the subcommissural organ. Microsc ResTech 52:468–483
Nualart F, Hein S, Rodriguez EM, Oksche O (1991) Identificationand partial characterization of the secretory glycoproteins of thebovine subcommissural organ-Reissner’s fiber complex. Evi-dence for the existence of two precursor forms.Brain Res MolBrain Res 11:227–238
Oksche A (1956) Funktionelle histologische Untersuchungen überdie Organe des Zwischenhirndaches der Chordaten. Anat Anz102:404–419
Oksche A (1961) Vergleichende Untersuchungen über die sekretor-ische Aktivität der Subkommissuralorgans und den Gliachar-akter seiner Zellen. Z Zellforsch Mikrosk Anat 54:549–612
Oksche A (1969) The subcommissural organ. J Neurovisc RelatSuppl 9:111–139
Olsson R (1958) Studies on the subcommissural organ. Acta Zool39:71–102
Olsson R (1961) Subcommissural ependyma and pineal organ de-velopment in human fetuses. Gen Comp Endocrinol 1:117–123
Richter HG, Munoz RI, Millan CS, Guinazu MF, Yulis CR,Rodriguez EM (2001) The floor plate cells from bovine expressthe mRNA encoding for SCO-spondin and its translation pro-ducts. Brain Res Mol Brain Res 93:137–147
Rodriguez EM, Oksche A, Hein S, Rodriguez S, Yulis R (1984)Comparative immunocytochemical study of the subcommissur-al organ. Cell Tissue Res 237:427–441
Rodriguez EM, Rodriguez S, Schoebitz K, Yulis CR, Hoffmann P,Manns V, Oksche A (1989) Light- and electron-microscopicinvestigation of the rat subcommissural organ grafted under thekidney capsule, with particular reference to immunocytochem-istry and lectin histochemistry. Cell Tissue Res 258:499–514
Rodriguez S, Rodriguez EM, Jara P, Peruzzo B, Oksche A (1990)Single injection into the cerebrospinal fluid of antibodiesagainst the secretory material of the subcommissural organ re-versibly blocks formation of Reissner’s fiber: immunocyto-chemical investigations in the rat. Exp Brain Res 81:113–124
Rodriguez EM, Oksche A, Hein S, Yulis CR (1992) Cell biology ofthe subcommissural organ. Int Rev Cytol 135:39–121
Rodriguez EM, Jara P, Richter H, Montecinos H, Flandez B,Wiegand R, Oksche A (1993) Evidence for the release of CSF-soluble secretory material from the subcommissural organ, withparticular reference to the situation in the human. In: Oksche A,Rodriguez EM, Fernandez-Llebrez P (eds) The subcommissuralorgan. Springer, Berlin Heidelberg New York, pp 121–131
Rodriguez EM, Brio Leon MA del, Riera P, Menendez J, SchoebitzK (1996) The floor plate of the hindbrain is a highly specializedgland: immunocytochemical and ultrastructural characteristics.Brain Res Dev Brain Res 97:153–168
Rodriguez EM, Rodriguez S, Hein S (1998) The subcommissuralorgan. Microsc Res Tech 41:98–123
Rodriguez S, Navarrete EH, Vio K, Gonzalez C, Schobitz K,Rodriguez EM (1999) Isograft and xenograft of the subcom-missural organ into the lateral ventricle of the rat and the for-mation of Reissner’s fiber. Cell Tissue Res 296:457–469
440

Rodriguez EM, Oksche A, Montecinos H (2001) Human subcom-missural organ, with particular emphasis on its secretory activ-ity during the fetal life. Microsc Res Tech 52:573–590
Sargent PE (1900) Reissner‘s fibre in the canalis centralis ofvertebrates. Anat Anz 17:33–44
Schoebitz K, Garrido O, Heinrichs M, Speer L, Rodriguez EM(1986) Ontogenical development of the chick and duck sub-commissural organ: an immunocytochemical study. Histochem-istry 84:1–40
Schoebitz K, Rodriguez EM, Garrido O, Brio MA del (1993)Ontogenetic development of the subcommissural organ withreference to the flexural organ. In: Oksche A, Rodriguez EM,Fernandez-Llebrez P (eds) The subcommissural organ. Spring-er, Berlin Heidelberg New York, pp 41–49
Schoebitz K, Gonzalez C, Peruzzo B, Yulis CR, Rodriguez EM(2001) Organ culture of the bovine subcommissural organ:evidence for synthesis and release of the secretory material.Microsc Res Tech 52:496–509
Sheffield JB, Graff D (1991) Extracellular proteases in developingchick neural retina. Exp Eye Res 52:733–741
Sterba G, Ermisch A, Freyer K, Hartmann G (1967) Incorporation ofsulphur-35 into the subcommissural organ and Reissner’s fibre.Nature 216:504
Sternberger LA, Hardy PH Jr, Cuculis JJ, Meyer HG (1970) Theunlabeled antibody enzyme method of immunohistochemistry:preparation and properties of soluble antigen-antibody complex(horseradish peroxidase-antihorseradish peroxidase) and its usein identification of spirochetes. J Histochem Cytochem 18:315–333
Vaillant C, Didier-Bazes M, Hutter A, Belin MF, Thomasset N(1999) Spatiotemporal expression patterns of metalloprotei-nases and their inhibitors in the postnatal developing rat cere-bellum. J Neurosci 19:4994–5004
Wingstrand KG (1953) Neurosecretion and antidiuretic activity inchick embryos with remarks on the subcommissural organ. ArkZool 6:41–67
Wislocki GB, Roth WD (1958) Selective staining of the humansubcommissural organ. Anat Rec 130:125–133
Yong VW, Power C, Forsyth P, Edwards DR (2001) Metallopro-teinases in biology and pathology of the nervous system. NatRev Neurosci 2:502–511
Yulis CR, Mota MD, Andrades JA, Rodriguez S, Peruzzo B,Mancera JM, Ramirez P, Garrido M, Perez-Figarez JM,Fernandez-Llebrez P, Rodriguez EM (1998) Floor plate andthe subcommissural organ are the source of secretory compounds of related nature: comparative immunocytochemicalstudy. J Comp Neurol 392:19–34
441




















