
Regulation of Bacterial Communities Through Antimicrobial Activity by the Coral Holobiont
14
HOST MICROBE INTERACTIONS Regulation of Bacterial Communities Through Antimicrobial Activity by the Coral Holobiont E. Charlotte E. Kvennefors & Eugenia Sampayo & Caroline Kerr & Genyess Vieira & George Roff & Andrew C. Barnes Received: 18 June 2011 /Accepted: 15 September 2011 # Springer Science+Business Media, LLC 2011 Abstract Interactions between corals and associated bacteria and amongst these bacterial groups are likely to play a key role in coral health. However, the complexity of these interactions is poorly understood. We investigated the functional role of specific coral-associated bacteria in maintaining microbial communities on the coral Acropora millepora (Ehrenberg 1834) and the ability of coral mucus to support or inhibit bacterial growth. Culture-independent techniques were used to assess bacterial community structures whilst bacterial culture was employed to assess intra- and inter-specific antimicrobial activities of bacteria. Members of Pseudoalteromonas and ribotypes closely related to Vibrio coralliilyticus displayed potent antimicro- bial activity against a range of other cultured isolates and grew readily on detached coral mucus. Although such bacterial ribotypes would be expected to have a competitive advantage, they were rare or absent on intact and healthy coral colonies growing in situ (analysed using denaturing gradient gel electrophoresis and 16S rRNA gene sequenc- ing). The most abundant bacterial ribotypes found on healthy corals were Gammaproteobacteria, previously de- fined as type A coral associates. Our results indicate that this group of bacteria and specific members of the Alphaproteobacteria described here as ‘type B associates’ may be important functional groups for coral health. We suggest that bacterial communities on coral are kept in check by a combination of host-derived and microbial interactions and that the type A associates in particular may play a key role in maintaining stability of microbial communities on healthy coral colonies. Introduction Scleractinian corals are the building blocks of coral reefs worldwide and are well known for their biodiversity and productivity [12]. Increasingly, corals are considered as complex communities that involve not only the cnidarian coral host but also their dinoflagellate symbionts and a range of microbial associates [15, 38, 39]. The coral holobiont (the coral host plus all of its associates) includes diverse intracellular and extracellular bacterial communities that (a) differ from those of the overlying water column [22, 38, 39], (b) can be host species specific and (c) display some level of spatial variability from local to geographic scales [22, 39]. Despite this knowledge, the direct function of bacterial associates remains largely unknown. Potential important roles of some of the associated bacteria may be to protect against invasive pathogens or fouling agents by occupying entry niches and space (acting as competitors) or Electronic supplementary material The online version of this article (doi:10.1007/s00248-011-9946-0) contains supplementary material, which is available to authorized users. E. C. E. Kvennefors : E. Sampayo : C. Kerr : G. Vieira : A. C. Barnes Centre for Marine Science, The University of Queensland, St Lucia, Brisbane, Queensland 4072, Australia G. Roff : A. C. Barnes School of Biological Sciences, The University of Queensland, St Lucia, Brisbane, Queensland 4072, Australia E. C. E. Kvennefors (*) : G. Roff ARC Centre of Excellence for Coral Reef Studies, The University of Queensland, St Lucia, Brisbane, Queensland 4072, Australia e-mail: [email protected] Microb Ecol DOI 10.1007/s00248-011-9946-0
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Regulation of Bacterial Communities Through Antimicrobial Activity by the Coral Holobiont

HOST MICROBE INTERACTIONS
Regulation of Bacterial Communities Through AntimicrobialActivity by the Coral Holobiont
E. Charlotte E. Kvennefors & Eugenia Sampayo &
Caroline Kerr & Genyess Vieira & George Roff &Andrew C. Barnes
Received: 18 June 2011 /Accepted: 15 September 2011# Springer Science+Business Media, LLC 2011
Abstract Interactions between corals and associatedbacteria and amongst these bacterial groups are likely toplay a key role in coral health. However, the complexity ofthese interactions is poorly understood. We investigated thefunctional role of specific coral-associated bacteria inmaintaining microbial communities on the coral Acroporamillepora (Ehrenberg 1834) and the ability of coral mucusto support or inhibit bacterial growth. Culture-independenttechniques were used to assess bacterial communitystructures whilst bacterial culture was employed to assessintra- and inter-specific antimicrobial activities of bacteria.Members of Pseudoalteromonas and ribotypes closelyrelated to Vibrio coralliilyticus displayed potent antimicro-bial activity against a range of other cultured isolates andgrew readily on detached coral mucus. Although such
bacterial ribotypes would be expected to have a competitiveadvantage, they were rare or absent on intact and healthycoral colonies growing in situ (analysed using denaturinggradient gel electrophoresis and 16S rRNA gene sequenc-ing). The most abundant bacterial ribotypes found onhealthy corals were Gammaproteobacteria, previously de-fined as type A coral associates. Our results indicate thatthis group of bacteria and specific members of theAlphaproteobacteria described here as ‘type B associates’may be important functional groups for coral health. Wesuggest that bacterial communities on coral are kept incheck by a combination of host-derived and microbialinteractions and that the type A associates in particular mayplay a key role in maintaining stability of microbialcommunities on healthy coral colonies.
Introduction
Scleractinian corals are the building blocks of coral reefsworldwide and are well known for their biodiversity andproductivity [12]. Increasingly, corals are considered ascomplex communities that involve not only the cnidariancoral host but also their dinoflagellate symbionts and arange of microbial associates [15, 38, 39]. The coralholobiont (the coral host plus all of its associates) includesdiverse intracellular and extracellular bacterial communitiesthat (a) differ from those of the overlying water column [22,38, 39], (b) can be host species specific and (c) displaysome level of spatial variability from local to geographicscales [22, 39]. Despite this knowledge, the direct functionof bacterial associates remains largely unknown. Potentialimportant roles of some of the associated bacteria may be toprotect against invasive pathogens or fouling agents byoccupying entry niches and space (acting as competitors) or
Electronic supplementary material The online version of this article(doi:10.1007/s00248-011-9946-0) contains supplementary material,which is available to authorized users.
E. C. E. Kvennefors : E. Sampayo : C. Kerr :G. Vieira :A. C. BarnesCentre for Marine Science,The University of Queensland,St Lucia,Brisbane, Queensland 4072, Australia
G. Roff :A. C. BarnesSchool of Biological Sciences,The University of Queensland,St Lucia,Brisbane, Queensland 4072, Australia
E. C. E. Kvennefors (*) :G. RoffARC Centre of Excellence for Coral Reef Studies,The University of Queensland,St Lucia,Brisbane, Queensland 4072, Australiae-mail: [email protected]
Microb EcolDOI 10.1007/s00248-011-9946-0

by the production of antibiotic compounds [14, 16, 36, 40].Given the global deterioration of coral reef ecosystemsthrough climate change [6] and disease outbreaks [50],understanding the community ecology and function ofcoral-associated microbial communities has become in-creasingly important.
Shifts in microbial community structure have beenobserved following coral bleaching and disease [22, 35,45]. Such shifts have also been recorded in diseased coralsin the absence of the specified causative agent [1, 22, 25,32, 45], and it is therefore at times unclear as to whetherthese changes are a result from specific bacterial pathogens,or are merely symptomatic of unhealthy corals. It ispossible that the disappearance of key bacterial associates(by biotic or abiotic disturbances) amongst these commu-nities provide an entry niche for opportunistic species thatcan further interfere with the microbial community structureand health status of the coral holobiont.
Factors that could lead to an environment that allowsproliferation of potentially harmful bacteria include (a) theloss of bacterial ribotypes with specific antimicrobialactivity or competitive advantage or (b) changes in host-derived antimicrobial activity (e.g. immunity). Alternative-ly, growth of specific bacterial ribotypes may be promotedor less favoured through provision of secondary metabolitesby the coral holobiont, which directly or indirectly lead to afavourable environment for certain species. Evidence fromthe current literature suggests that both bacterial associatesand the coral itself produce compounds that have the abilityto affect associated bacteria [20, 31, 37, 43, 49]. Consid-ering the high diversity of coral-associated bacterialcommunities, it is likely that promotion and inhibition bya multiplicity of bacterial species occur simultaneously in acomplex manner within the coral holobiont.
Previously, a specific group of Gammaproteobacteria,termed type A associates, was identified as a keymicrobial group associated with two unrelated GreatBarrier Reef (GBR) coral species, Acropora hyacinthusand Stylophora pistillata [22]. These specific bacteriawere not only common across healthy coral colonies butalso represented the dominant component of the microbialcommunity [22]. Furthermore, these types of bacteria havebeen found in other coral species on the GBR andelsewhere (as outlined in [22]). During environmentalstress conditions causing bleaching or the manifestation ofcoral disease signs (such as ‘white syndrome’), type Aassociates are displaced by Vibrio and Serratia speciesamongst others [5, 22]. Upon recovery from bleaching, thecoral-associated microbial community returned to onedominated by type A Gammaproteobacteria (classified in[22]) [5], and these observations demonstrate the dynamicnature of the microbial community structure on healthy,diseased and bleached corals.
Several bacteria have been isolated from corals thatposses antimicrobial activity against putative coral patho-gens [31, 37, 43], and the antimicrobial activity of coralmucus is observed to decline in corals that display signs ofcoral bleaching or disease [37]. These observations hint atintriguing relationships between different coral-associatedbacteria and between bacterial associates and the coral host.It is currently unknown whether bacterial communities areselected by extrinsically mediated factors or whether theholobiont itself selects for beneficial associates. Analogousstudies of sponges suggest that the latter is possible, whereit is proposed that the species Mycale adhaerens mayselectively sequester bacterial epibionts with antimicrobialproperties [24]. Here, we use a multidisciplinary approachto investigate how coral-associated bacterial communitiesare influenced by specific members of the bacterialcommunity as well as by compounds secreted by the coralholobiont. Highly replicated culture-independent techni-ques were employed to identify the most common bacterialassociates on healthy individuals of the reef building coralAcropora millepora (Ehrenberg 1834), whilst bacterialinteractions and the effects of coral mucus on differentbacterial groups were investigated in culture.
Materials and Methods
Coral Sample Collection
Samples of visibly healthy A. millepora were collected at1–5-m depth from Heron reef (23°27′ S; 151°54′ E),southern Great Barrier Reef, Australia in March 2006.Replicate samples consisted of the outer 3–4 cm of a branchtip from separate colonies dispersed within ~1-km2 area ofthe reef, in order to account for small-scale spatial differ-ences in the coral bacterial community and avoid samplingof coral clone mates. For DNA-based analyses of microbialcommunity structure, 11 replicate samples from differentcolonies were prepared following previously establishedprotocols [22]. Briefly, the coral tissue and mucus wereremoved using an airbrush, and the slurry was preserved indimethyl sulfoxide (DMSO) conservation buffer [41] andstored at −20°C until further processing.
For mucus collection, six replicate coral samples ofdifferent colonies were each placed in separate steriledisposable 50-ml centrifuge tubes (Greiner) and allowedto produce mucus for 30 min. Branches were retained forsubsequent bacteriological analyses, and the mucus wascollected by centrifugation at 8×g for 5 min at 4°C. Analiquot of the recovered mucus was immediately used toculture bacteria as described below and the remainingmucus was frozen at −80°C for further analysis. Toinvestigate the effect of mucus obtained from wild versus
E.C.E. Kvennefors et al.

aquarium-reared corals on bacterial growth, six replicatelive coral pieces (7×5×2 cm) were transported back toindoor aquaria facilities at The University of Queenslandand maintained in artificial seawater prepared from RO-filtered water and a commercial ocean salts mixture (OceanNature, Aquasonic, Wauchope, Australia). After 5 weeks ofaquarium culture, mucus was collected from these corals asdescribed above.
Isolation of Bacteria from Coral Samples
Bacterial isolates were obtained from mucus and coraltissues independently to avoid potential bactericidal effectfrom coral tissues on mucus-associated bacteria. Freshlyextracted (immediately upon sampling) coral mucus andtissues (removed using sterile scalpel) was diluted 10-, 100-and 1,000-fold in sterile artificial seawater after whichdilutions and undiluted samples (100 μl) were immediatelyspread onto marine agar plates (Difco, BD Biosciences,North Ryde, Australia). All agar plates were incubated at25°C, and after 24 and 48 h, a total of 27 bacterial isolateswith differing colony morphologies from each isolationsource were selected for further study. Of these, 25 wereunique and were used for further analyses. Cross-inhibitionstudies were immediately performed with these freshcultures as described below. A sample from each bacterialisolate was also immediately stored at −80°C in marinebroth (Difco, BD Biosciences, North Ryde, Australia)containing glycerol (25% v/v) for subsequent replicatestudies and identification.
Cross-inhibition of Bacterial Growth
The potential of each bacterial isolate obtained in thepresent study to inhibit the growth of each other wasassessed by an agar plate cross-streaking method on marineagar following previously described methodologies [13].Inhibition was measured from the edge of the vertical streakwith the ‘test isolate’ to the first colony of ‘target isolate’cross-streak and divided into three distance-dependentcategories: 1–5, 5–20 and >20 mm (Fig. 1). The experimentwas repeated three times using three dilutions of the ‘testisolate’ each time and the inhibition zones recorded. Ifinhibition for an isolate was not consistently observed inreplicate assays, this was reported as no inhibition in thefinal results.
Effect of Coral Mucus on Bacterial Growth
To investigate the effect of coral mucus on the growth ofbacteria isolated from the coral, isolates were grown inmucus. Samples from both wild and aquarium-reared coralswere used in order to assess mucus properties in changed
environmental conditions as many studies on coral–bacte-rial interactions are performed in aquaria environments.Mucus samples were collected from six replicate wild andsix replicate aquarium-reared A. millepora nubbins (allfrom different colonies) and were pooled respectively.Pooled samples were centrifuged at 8,000×g for 2 min atroom temperature to remove any debris and associated
Figure 1 Representative culture plates showing the assay method fortesting inhibitory activity of cultured isolates from corals against eachother. The triple horizontal streak represents three dilutions of (top tobottom represents undiluted, 1:10 diluted and 1:100 diluted) isolatetested for inhibitory activity whilst the vertical streak represents theisolate against which inhibition was tested. Typical inhibitionproperties are shown in a for isolate T15, Pseudolateromonads rubra;b isolate T25, Pseudoalteromonads luteoviolaceae; c M13, V.coralliilyticus-related isolate. The distance from the ‘test isolate’ tothe ‘target isolate’ is measured and termed the ‘inhibition zone’ (iz)
Regulation of Bacterial Communities by the Coral Holobiont

bacteria. The pellet was removed and the supernatant wassterilised by UV irradiation in uncovered Petri dishes for30 min at 302 nm in a biosafety cabinet. The proteinconcentration of the mucus samples was determinedspectrophotometrically using a NanoDrop ND-1000 spec-trophotometer (Protein A280 module) (NanoDrop Technol-ogies, Rockland, DE, USA) and also using bicinchoninicacid protein assay (Pierce, Rockford, IL, USA) with bovineserum albumin as a standard. Differences in the proteinconcentrations of the mucus from wild versus aquarium-maintained corals were corrected by standardising theprotein concentration in the mucus to 0.25 mg ml−1 usingsterile artificial seawater.
Sterilised mucus samples were inoculated with bacterialisolates of interest. Inocula were prepared from freshovernight cultures of the bacterial isolates previously storedat −80°C in glycerol buffer. Bacteria were washed inphosphate buffered saline (PBS)+2% NaCl (dilutionbuffer), and standard suspensions prepared in dilutionbuffer to OD600=1.0. In assays with mucus only, 50-μlmucus was added to the wells of a sterile 96-well microplateand inoculated with 10 μl of a 100-fold dilution of the standardbacterial suspension described above. Plates were incubated ina thermomixer (Eppendorf, Hamburg, Germany) at 28°C at100 rpm. Growth of bacteria was measured by reading opticaldensity at 600 nm after 0 and 20 h of incubation in a BMGFluostar Optima plate reader (BMG Labtech, Melbourne,Australia). Non-inoculated controls (here used as blanks) wereincluded to account for any variations in OD resulting from themucus itself, or from any potential contaminating growth insterilised mucus. Additional controls contained dilution bufferonly (PBS+2% NaCl) to control for airborne contamination ordilution buffer with corresponding inoculate to control forgrowth without mucus. Following incubations, aliquots (10 μl)from all controls were spotted in triplicate onto marine agarplates to detect any viable bacteria in control samples and thiswas repeated three times (including all the control incuba-tions). Bacterial growth was determined by the change inoptical density at 600 nm after 20 h of incubation.
To determine differences between growth of bacterialisolates on mucus from wild versus aquarium-reared coralcolonies, a factorial analysis of variance was performedusing the software package Statistica (8.0, Statsoft, Tulsa,OH, USA), and post hoc tests were performed usingFisher’s LSD test. Normality and homogeneity of variancewas assessed using Levene’s test of equal variances andresidual plots.
Identification of Bacterial Isolates by Amplificationand Sequencing of 16S rRNA Gene
DNA from each of the isolates was extracted from freshlygrown cultures using a Plant Mini Kit (QIAGEN). The 16S
rRNA gene of the isolates was amplified by polymerase chainreaction (PCR) in 50-μl reaction volumes containing 0.2 mMof each deoxynucleotide triphosphate (Biotech, USA),0.5 μM of each primer 27f (5′-AGA GTT TGA TCM TGGCTC AG-3′) and 1492r (5′-TAC GGY TAC CTT GTT ACGACT T-3′) [23], 1.5-mM MgCl2, 1× reaction buffer, 1 U ofPlatinum Taq DNA polymerase (Invitrogen, Melbourne,Australia) and approximately 50 ng of total DNA and sterilewater to volume. Amplification was performed in a Master-cycler Gradient Ep S (Eppendorf, Hamburg, Germany) withan initial denaturation at 94°C for 5 min, followed by30 cycles of 94°C for 1 min, 48°C for 1 min, 72°C for 2 minand a final extension of 72°C for 10 min. The resulting PCRproduct was purified using a MEGA-spin PCR purificationkit (Intron Biotechnology, South Korea) according to themanufacturer’s instructions, and three replicate sequencingreactions were performed using the primers 27f, 1492r (asabove) and 530 F (5′-GTGCCAGCMGCCGCGG-3′)[23] at the Australian Genome Research Facility usingBigDye Terminator v3.1 chemistry (Applied Biosystems).Sequences were visually inspected and assembled usingDNASTAR (Laser gene) software and their identityestablished using a BLAST search on GenBank [2](www.ncbi.nlm.gov/BLAST/). Closest published relativesand percent homology were used to assign affiliations toknown ribotypes.
Analysis of Bacterial Community Compositionby Denaturing Gradient Gel Electrophoresis
Tissue and mucus samples conserved in DMSO preserva-tion buffer were used for microbial community analyses ofhealthy coral colonies. Samples were extracted, PCR-amplified and prepared for denaturing gradient gel electro-phoresis (DGGE) analyses following previously describedmethods [22]. Briefly, a Plant Mini Kit (Qiagen, Australia)was used to extract DNA and the universal primers 27f and1492r were used to amplify the bacterial 16S rRNA gene[23]. The resulting PCR product was purified using aQIAquick PCR purification kit (Qiagen) and a nested PCRwas performed using the internal primers GC358f (5′-CGCCCG CCG CGC CCC GCG CCC GTC CCG CCG CCCCCG CCC CCC TAC GGG AGG CAG CAG-3′) and 517r(5′-ATT ACC GCG GCT GCT GG-3′) [29]. For visual-isation of the bacterial community structure, DGGEsamples were run on a D-Code universal mutation detectionsystem (Bio-Rad, USA) using a 25–60% denaturinggradient run at 100 V for 10 h at 60°C [22]. Gels werestained with SYBR Green (Sigma-Aldrich, USA) andvisualised using a UVIDOC camera system fitted withSYBR Green filter.
All bands were scored for the presence or absence inreplicate samples with the aid of Quantity One®
E.C.E. Kvennefors et al.

fragment analysis software (BioRad), and the resultswere confirmed by sequencing. Sharply featured bandsoccurring more than once were excised using sterilescalpels from a minimum of two different samples.Dominant bands that occurred in one sample only werecut from two different gels to ensure repeatability ofresults [22]. Ribotypes with high band intensity werepresumed to be more abundant members of the overallbacterial community [30]. DNA from excised bands wasamplified using the primers 517r and 358f without the GCclamps (5′-CCT ACG GGA GGC AGC AG-3′), andsequencing was performed as described above for isolates,but using the primers 517r and 358 f.
Phylogenetic Analysis
Phylogenetic trees were based on analysis of partial 16SrRNA gene sequences (>1,350 bp) aligned to closepublished matches using the BLAST algorithm [2] andother common bacterial ribotypes previously identifiedfrom healthy and diseased corals. Evolutionary relation-ships with Aquifex pyrophilus as the out-group werecalculated using maximum likelihood methods with theARB software package [27]. Shorter partial sequences(<1,350 bp) were subsequently added to the trees withoutaltering tree topology using the parsimony-interactive toolin ARB. Bootstrap values (1,000 replicates) for a maximumlikelihood tree were calculated in PAUP 4.0b10 [47] usingthe sequences (>1,350 bp) initially used to construct thetree in ARB, and bootstrap values greater than 50% wereadded to the ARB tree.
Results
Identification and Characterisation of Bacterial Isolates
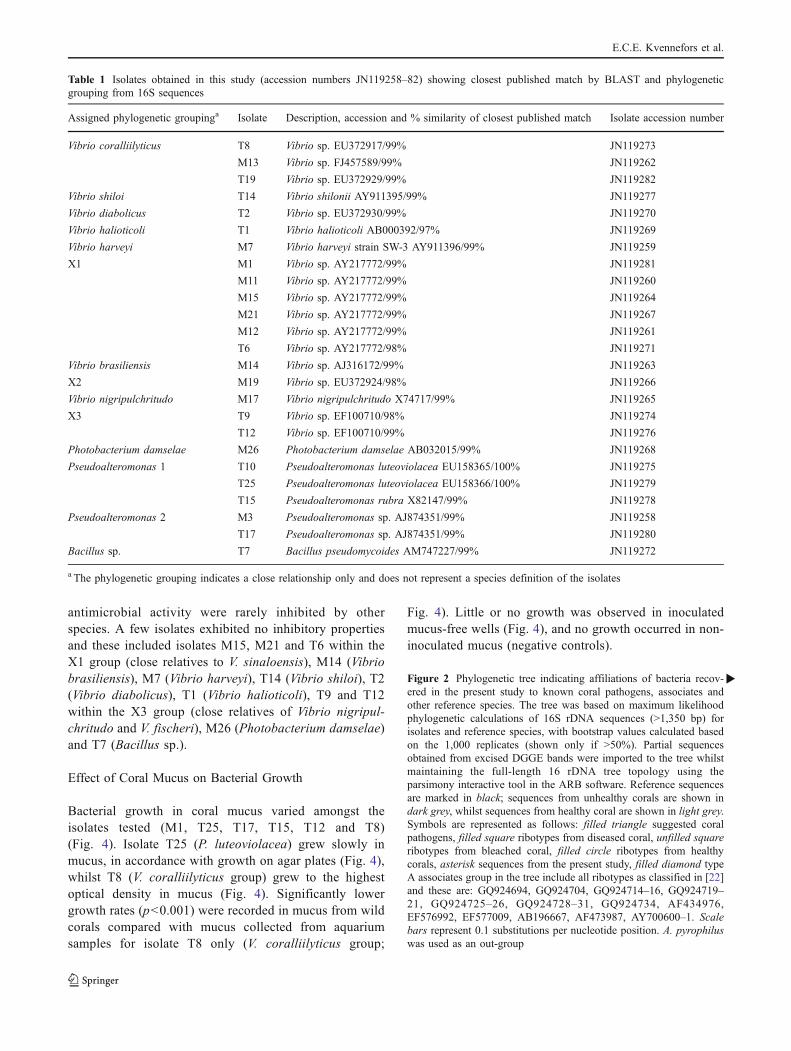
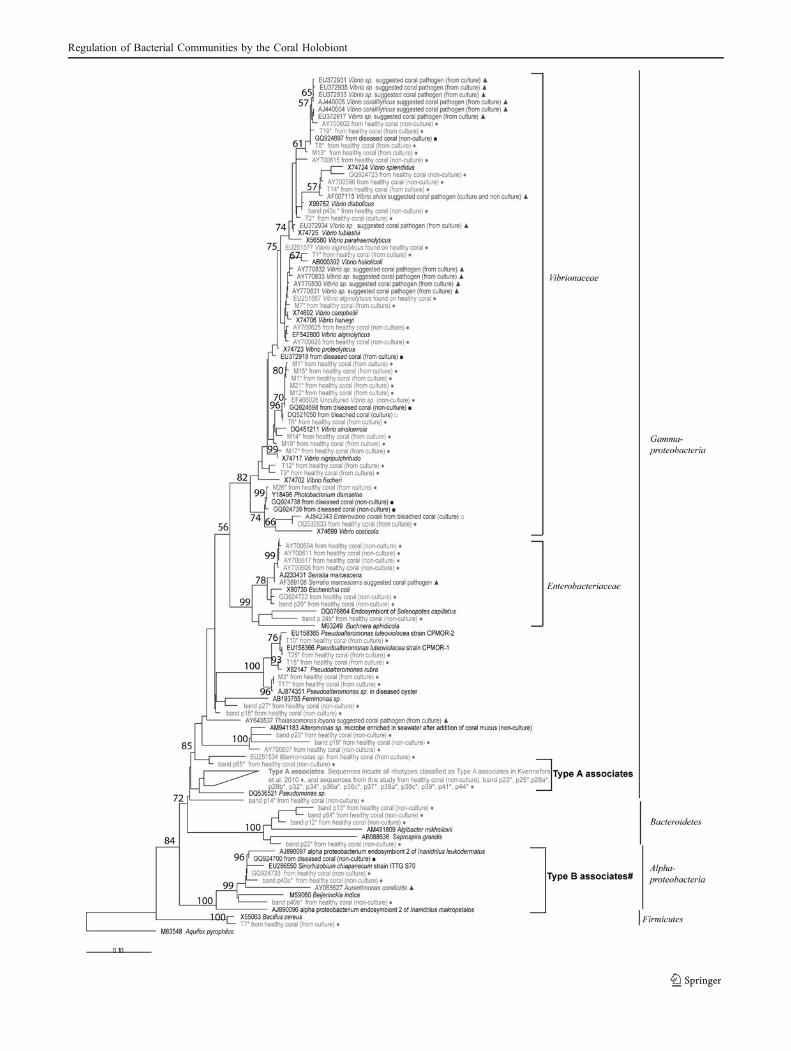
Identities of cultured bacterial isolates, determined byBLAST analysis of 16S rRNA gene sequence supportedby phylogenetic analyses, are presented in Table 1. Allisolates designated with the prefix M were derived frommucus only, whilst isolates with the prefix T wereobtained from tissue samples where bacteria originatefrom either the tissue or adherent mucus. Sequencing andsubsequent BLAST analysis of the 16S rDNA geneamplified from the isolates identified them as membersof the Vibrionaceae, Pseudoalteromonidae, and Bacillus(Table 1), with similarities to numerous bacterial ribotypespreviously found in association with healthy or diseasedcorals (Fig. 2).
In total, 12 isolates were obtained from mucus onlysamples and 13 from tissue/mucus samples (Table 1). Themajority of cultured isolates (76%) were members of the
Vibrionaceae (genera Vibrio and Photobacterium), with 11of the Vibrionaceae isolates associated with mucus only(M1, M7, M11, M12, M13, M14, M15, M17, M19, M21,M26) and 8 with tissue/mucus samples (T1, T2, T6, T8, T9,T12, T14, T19). Five remaining isolates were Pseudoalter-omonas spp., four of which were associated with tissue/mucus (T10, T15, T17, T25) and one with mucus only(M3), and one Bacillus sp. associated with tissue/mucus(T7) (Table 1).
Isolates T10 and T25 were identified as tworibotypes of Pseudoalteromonas luteoviolacea by phylo-genetic identification (100% match; Table 1, Fig. 2), aswell as colony appearance (characteristic dark purplecolouring) and isolate T15 was very closely related toPseudoalteromonas rubra (Figs. 1 and 2; Table 1) and hadthe characteristic red colouring. Interestingly, both strainsof P. luteoviolacea (T10 and T25) displayed lower growthrates compared to all other isolates in this study. All of theisolated cultured bacteria appeared to represent closerelatives of previously observed ribotypes and certainisolates were closely related to previously unidentifiedstrains or species of known coral-associated bacterial.Some bacteria, within a phylogenetic grouping hereassigned as X1, X2 and X3, did not appear to affiliatewith any one species within the Vibrionaceae, but weremost similar to Vibrio sinaloensis in the case of X1 andX2 and Vibrio fischeri in the case of X3 (Table 1). Furtheranalysis is required to assign these unknown groups asspecific species within the genus Vibrionaceae.
Inhibitory Properties of Bacterial Isolates
Inhibitory properties of the different isolates were, ingeneral, consistent for close relatives (Table 2, Figs. 2and 3), where most isolates within a phylogeneticgrouping showed either similar patterns of target inhibi-tion or were targeted as a group (Fig. 3). However, therewere some exceptions to this general trend, and someclose relatives had strikingly different patterns of inhibi-tion, most notably isolate T19 within the Vibrio corallii-lyticus group. Isolates T8 and M13 within the V.coralliilyticus group did not inhibit any members of theX1 group, and in turn, members of the X1 group did notinhibit isolates T8 and M13, indicating these were not incompetition with each other. Furthermore, members ofthese two groups showed widespread inhibition properties.In contrast, T19 did not share targets of inhibition with theother members of the V. coralliilyticus group but insteadtargeted members of the X1 group and these, in turn,inhibited T19. Overall, isolates M13 and T8 within the V.coralliilyticus group and T25 and T15 from the Pseudoal-teromonas group 2 were the most broadly inhibitory(Fig. 3). These isolates with potent and broad ranging
Regulation of Bacterial Communities by the Coral Holobiont
antimicrobial activity were rarely inhibited by otherspecies. A few isolates exhibited no inhibitory propertiesand these included isolates M15, M21 and T6 within theX1 group (close relatives to V. sinaloensis), M14 (Vibriobrasiliensis), M7 (Vibrio harveyi), T14 (Vibrio shiloi), T2(Vibrio diabolicus), T1 (Vibrio halioticoli), T9 and T12within the X3 group (close relatives of Vibrio nigripul-chritudo and V. fischeri), M26 (Photobacterium damselae)and T7 (Bacillus sp.).
Effect of Coral Mucus on Bacterial Growth
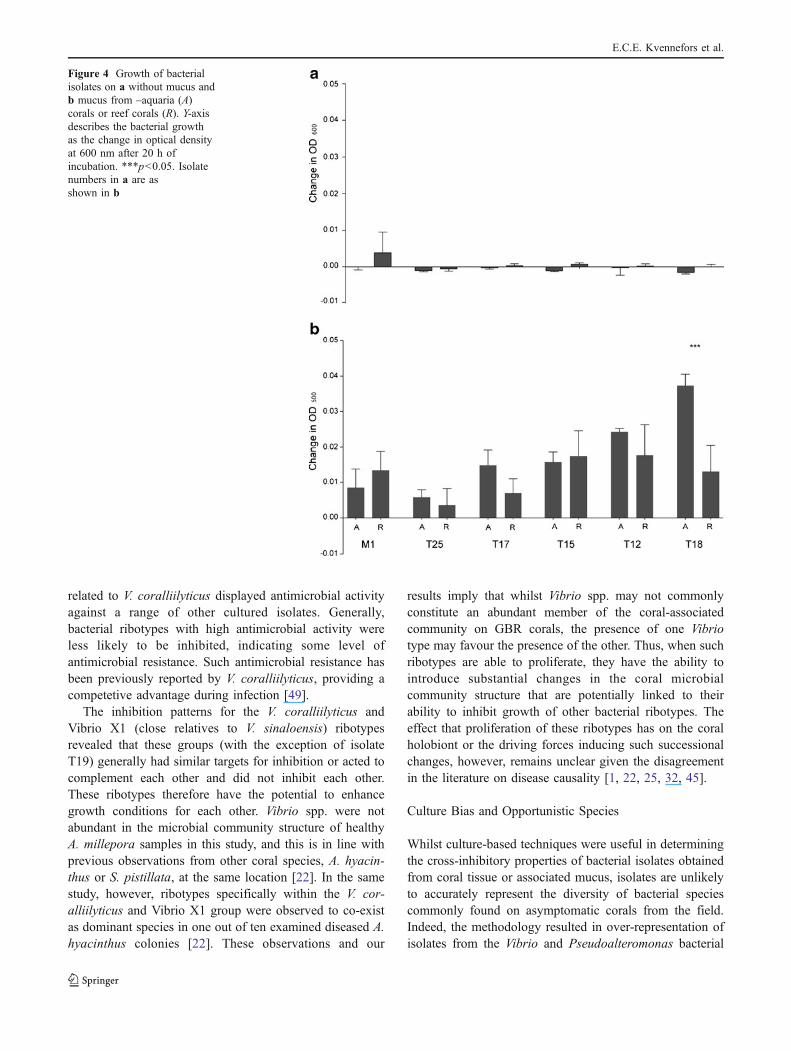
Bacterial growth in coral mucus varied amongst theisolates tested (M1, T25, T17, T15, T12 and T8)(Fig. 4). Isolate T25 (P. luteoviolacea) grew slowly inmucus, in accordance with growth on agar plates (Fig. 4),whilst T8 (V. coralliilyticus group) grew to the highestoptical density in mucus (Fig. 4). Significantly lowergrowth rates (p<0.001) were recorded in mucus from wildcorals compared with mucus collected from aquariumsamples for isolate T8 only (V. coralliilyticus group;
Fig. 4). Little or no growth was observed in inoculatedmucus-free wells (Fig. 4), and no growth occurred in non-inoculated mucus (negative controls).
Table 1 Isolates obtained in this study (accession numbers JN119258–82) showing closest published match by BLAST and phylogeneticgrouping from 16S sequences
Assigned phylogenetic groupinga Isolate Description, accession and % similarity of closest published match Isolate accession number
Vibrio coralliilyticus T8 Vibrio sp. EU372917/99% JN119273
M13 Vibrio sp. FJ457589/99% JN119262
T19 Vibrio sp. EU372929/99% JN119282
Vibrio shiloi T14 Vibrio shilonii AY911395/99% JN119277
Vibrio diabolicus T2 Vibrio sp. EU372930/99% JN119270
Vibrio halioticoli T1 Vibrio halioticoli AB000392/97% JN119269
Vibrio harveyi M7 Vibrio harveyi strain SW-3 AY911396/99% JN119259
X1 M1 Vibrio sp. AY217772/99% JN119281
M11 Vibrio sp. AY217772/99% JN119260
M15 Vibrio sp. AY217772/99% JN119264
M21 Vibrio sp. AY217772/99% JN119267
M12 Vibrio sp. AY217772/99% JN119261
T6 Vibrio sp. AY217772/98% JN119271
Vibrio brasiliensis M14 Vibrio sp. AJ316172/99% JN119263
X2 M19 Vibrio sp. EU372924/98% JN119266
Vibrio nigripulchritudo M17 Vibrio nigripulchritudo X74717/99% JN119265
X3 T9 Vibrio sp. EF100710/98% JN119274
T12 Vibrio sp. EF100710/99% JN119276
Photobacterium damselae M26 Photobacterium damselae AB032015/99% JN119268
Pseudoalteromonas 1 T10 Pseudoalteromonas luteoviolacea EU158365/100% JN119275
T25 Pseudoalteromonas luteoviolacea EU158366/100% JN119279
T15 Pseudoalteromonas rubra X82147/99% JN119278
Pseudoalteromonas 2 M3 Pseudoalteromonas sp. AJ874351/99% JN119258
T17 Pseudoalteromonas sp. AJ874351/99% JN119280
Bacillus sp. T7 Bacillus pseudomycoides AM747227/99% JN119272
a The phylogenetic grouping indicates a close relationship only and does not represent a species definition of the isolates
Figure 2 Phylogenetic tree indicating affiliations of bacteria recov-ered in the present study to known coral pathogens, associates andother reference species. The tree was based on maximum likelihoodphylogenetic calculations of 16S rDNA sequences (>1,350 bp) forisolates and reference species, with bootstrap values calculated basedon the 1,000 replicates (shown only if >50%). Partial sequencesobtained from excised DGGE bands were imported to the tree whilstmaintaining the full-length 16 rDNA tree topology using theparsimony interactive tool in the ARB software. Reference sequencesare marked in black; sequences from unhealthy corals are shown indark grey, whilst sequences from healthy coral are shown in light grey.Symbols are represented as follows: filled triangle suggested coralpathogens, filled square ribotypes from diseased coral, unfilled squareribotypes from bleached coral, filled circle ribotypes from healthycorals, asterisk sequences from the present study, filled diamond typeA associates group in the tree include all ribotypes as classified in [22]and these are: GQ924694, GQ924704, GQ924714–16, GQ924719–21, GQ924725–26, GQ924728–31, GQ924734, AF434976,EF576992, EF577009, AB196667, AF473987, AY700600–1. Scalebars represent 0.1 substitutions per nucleotide position. A. pyrophiluswas used as an out-group
�
E.C.E. Kvennefors et al.
Regulation of Bacterial Communities by the Coral Holobiont

Microbial Community Structure of A. milleporaDetermined by DGGE
DGGE analysis of the coral-associatedmicrobial communitiesof A. millepora around Heron Island was performed in orderto assess whether cultured bacterial isolates were amongstthose present as dominant members of the microbialcommunity in healthy coral samples. A total of 11 collectedsamples from individual A. millepora colonies at HeronIsland showed significant variation of the microbial commu-nity structure amongst colonies (Table 2; Fig. 5).
Members of Alphaproteobacteria and Gammaproteobacte-ria, Bacteroidetes and Firmicutes were identified in the DGGEprofiles (Fig. 5 and Table 2) but Gammaproteobacteria andAlphaproteobacteria represented the dominant constituents ofthe microbial community across different colonies. Five of
the 11 samples were dominated by Gammaproteobacterialribotypes belonging to a group termed type A associates(Fig. 5 and Table 2), which are also commonly found acrosscoral colonies of healthy A. hyacinthus and S. pistillata fromthe same reef [22]. One sample appeared to be completelydominated by a single ribotype (sample 3; Fig. 5) with highsimilarity to Escherichia. The remaining samples contained amixed community of Gammaproteobacteria and Alphapro-teobacteria, Alteromonadales and Bacteroidetes. None of thecultured isolates had a 100% match with those retrieved fromthe DGGE profiles of healthy colonies of A. millepora, buttwo Alteromonas ribotypes (bands p15 and p27) were veryclosely related to the cultured Pseudoalteromonas isolates(Fig. 2) as well as a single Vibrio spp. (band p40c) found thatwas closely related to isolate T2 and grouping with V.diabolicus (Fig. 2).
Table 2 DGGE band identities (sequence data; Supplemental material 1) showing closest published match by BLAST and phylogenetic groupingfrom 16S sequences
Grouping Band number Description, accession and % similarity of closest published match
Type A associate (Gammaproteobacteria) Band p23 Uncultured bacterium GQ924718/98%
Band p25 Uncultured bacterium FJ809618/100%
Band p28a Uncultured bacterium clone PDA-OTU3 AY700601/96%
Band p28b Uncultured bacterium clone PDA-OTU3 AY700601/97%
Band p32 Uncultured bacterium FJ809618/99%
Band p34 Uncultured bacterium FJ809618/99%
Band p36a Uncultured bacterium GQ924730/100%
Band p36c Uncultured bacterium GU184753/97%
Band p37 Uncultured bacterium GQ924730/97%
Band p38a Uncultured bacterium GQ924728/98%
Band p38c Uncultured bacterium GQ924728/97%
Band p39 Uncultured bacterium GQ924731/97%
Band p41 Uncultured Gammaproteobacterium EF630125/95%
Band p44 Uncultured bacterium GQ924734/96%
Gammaproteobacteria Band p14 Uncultured Gammaproteobacterium DQ234178/93%
Bacteroidetes Band p12 Mesoflavibacter zeaxanthinifaciens AB265181/98%
Band p13 Mesoflavibacter zeaxanthinifaciens AB265181/96%
Band p84 Mesoflavibacter zeaxanthinifaciens AB265181/97%
Band p22 Saprospira grandis AB088636/98%
Alteromonadales Band p15 Pseudoalteromonas luteoviolacea EU158366/98%
Band p27 Ferrimonas sp. AB193755/98%
Band p19 Alteromonas sp. EU251538/94%
Band p20 Alteromonas macleodii Y18232/97%
Oceanospirillales Band p85 Marinomonas sp. EU251534/99%
Enterobacteriaceae Band p24b Endosymbiont of Solenopotes capillatus DQ076664/96%
Band p29 Escherichia coli EU438991/98%
Type B associate (Alphaproteobacteria) Band p40a Ochrobactrum sp. EU438986/97%
Band p40b Uncultured Methylocella sp. EF625937/97%
Vibrionaceae Band p40c Vibrio diabolicus X99762/100%
The identity of bands were resolved by sequencing using both forward and reverse primers from a minimum of two different samples to assuretrue identity. Bands that occurred only once were sequenced from two separate DGGE gel extractions
E.C.E. Kvennefors et al.

Discussion
Inter-species Interactions of Cultured Coral-AssociatedBacteria
Our results indicate that interactions between coral-associated bacterial species are highly complex. Bacterialisolates were observed to inhibit a broad range of othercoral-associated bacteria, potentially affecting the overallmicrobial community structure in situ through direct
inhibition of competing species. Several bacterial isolatesinhibited bacteria that were closely related to putative coralpathogens, and these findings corroborate previous reportsthat many bacteria cultured from corals inhibit putativecoral pathogens in vitro [31, 37, 43]. However, bacterialspecies associated with, and often found on, diseased coralalso had potent and specific antimicrobial activity indicat-ing that these may also hold competitive advantage andinhibit ‘probiotic’ species if conditions are favourable.Members of Pseudoalteromonas and ribotypes closely
Figure 3 Inhibition properties of coral bacterial isolates. Isolatesexerting inhibition are shown on the left column (vertical), and isolatesthat are inhibited are shown in the top row (horizontal). Assignedphylogenetic groupings (see Table 2) are listed on the left. Inhibitionwas measured from the edge of the vertical streak on the agar plates(inhibitors) to the first colony of the cross-streak (inhibited isolates;
horizontal) and divided into three categories (1–5, 5–20, >20 mm).Indicated inhibition is the mean result of three independently repeatedtrials, showing the average inhibition observed on three differentdilutions of tester strains (horizontal). No designation indicates noobserved inhibition
Regulation of Bacterial Communities by the Coral Holobiont
related to V. coralliilyticus displayed antimicrobial activityagainst a range of other cultured isolates. Generally,bacterial ribotypes with high antimicrobial activity wereless likely to be inhibited, indicating some level ofantimicrobial resistance. Such antimicrobial resistance hasbeen previously reported by V. coralliilyticus, providing acompetetive advantage during infection [49].
The inhibition patterns for the V. coralliilyticus andVibrio X1 (close relatives to V. sinaloensis) ribotypesrevealed that these groups (with the exception of isolateT19) generally had similar targets for inhibition or acted tocomplement each other and did not inhibit each other.These ribotypes therefore have the potential to enhancegrowth conditions for each other. Vibrio spp. were notabundant in the microbial community structure of healthyA. millepora samples in this study, and this is in line withprevious observations from other coral species, A. hyacin-thus or S. pistillata, at the same location [22]. In the samestudy, however, ribotypes specifically within the V. cor-alliilyticus and Vibrio X1 group were observed to co-existas dominant species in one out of ten examined diseased A.hyacinthus colonies [22]. These observations and our
results imply that whilst Vibrio spp. may not commonlyconstitute an abundant member of the coral-associatedcommunity on GBR corals, the presence of one Vibriotype may favour the presence of the other. Thus, when suchribotypes are able to proliferate, they have the ability tointroduce substantial changes in the coral microbialcommunity structure that are potentially linked to theirability to inhibit growth of other bacterial ribotypes. Theeffect that proliferation of these ribotypes has on the coralholobiont or the driving forces inducing such successionalchanges, however, remains unclear given the disagreementin the literature on disease causality [1, 22, 25, 32, 45].
Culture Bias and Opportunistic Species
Whilst culture-based techniques were useful in determiningthe cross-inhibitory properties of bacterial isolates obtainedfrom coral tissue or associated mucus, isolates are unlikelyto accurately represent the diversity of bacterial speciescommonly found on asymptomatic corals from the field.Indeed, the methodology resulted in over-representation ofisolates from the Vibrio and Pseudoalteromonas bacterial
Figure 4 Growth of bacterialisolates on a without mucus andb mucus from –aquaria (A)corals or reef corals (R). Y-axisdescribes the bacterial growthas the change in optical densityat 600 nm after 20 h ofincubation. ***p<0.05. Isolatenumbers in a are asshown in b
E.C.E. Kvennefors et al.

groups. Culture bias towards these groups has previouslybeen noted in coral bacterial studies [38], yet studies usingculture techniques only are common in this field. Mostmarine bacteria remain unculturable [33], and culture of themost common bacterial associates of intact and healthy A.millepora around Heron Island was not possible. We werethus unable to ascertain the inhibitory/susceptibility profileof a potential keystone associate in coral microbialcommunities. This finding has significant ramificationssince a multitude of previous studies on antimicrobialactivity from the coral holobiont [37, 49] or on coralpathogens [3, 7, 46] rely exclusively on culture-basedtechniques and thus do not incorporate the effects of keybacterial associates on corals. Our results highlight thecontinuing importance of investigating microbial commu-nities through culture-independent techniques simulta-neously when assessing functional capabilities of bacterialisolates in vitro.
Although culture-independent methods such as DGGEmay be unable to detect bacterial ribotypes that are presentin very low abundances on the coral, a significantadvantage is that it allows for the rapid and inexpensivescreening of a high number of replicate samples. Given thatthe microbial community diversity associated with corals isvariable between colonies of the same species, with theadded complexity of spatial diversity within the same reefsystem, obtaining an estimate of community diversity isimportant to allow for a reliable interpretation of resultsbased on culture-based methods or those using highthroughput sequencing of few samples. Some of thecultured isolates were indeed found by DGGE but neverin high abundance in several replicates of healthy, intactcolonies of A. millepora sampled around Heron Island. It islikely that many of these isolates persist in low abundanceand their growth appears to be favoured in the nutrient-richcondition of laboratory media.
Figure 5 Denaturing gradientelectrophoresis gel showing themicrobial community structureof different colonies of A.millepora. Numerical codesindicate representative bandsthat were excised forsequencing; their identitiesare shown in Table 2 andphylogenetic grouping in Fig. 2.Each lane (1–11) represents areplicate sample froma different colony
Regulation of Bacterial Communities by the Coral Holobiont

Proliferation of Vibrio in coral mucus and on coralsduring stress or disease conditions has been observed [5,28], and several ribotypes, including those closely relatedto V. coralliilyticus, are increasingly being implicated ascoral pathogens for different syndromes [3, 7, 18, 46].Vibrio spp. are easily cultured from apparently healthycorals [8, 43], and many of the isolates obtained fromhealthy corals in the present study are indeed closelyrelated to Vibrio species implicated to be pathogenic tocorals (Fig. 2). Whilst this group of bacteria are oftensuggested as major coral disease agents when present,some such Vibrio ribotypes may equally constituteopportunistic secondary invaders [44, 48]. In the presentstudy, the ability of V. coralliilyticus to grow rapidly onrich media and in mucus from coral under alteredenvironmental conditions coupled with with its potentantibacterial properties and resistance to inhibition byother coral bacteria suggests that these specific ribotypesare opportunistic species, able to rapidly proliferate whensupression from one or several members of the coralholobiont fails.
In Situ Microbial Community Analysis of A. millepora
The microbial community analysis of the samples of A.millepora was consistent with previous findings from thesame region [22] and showed that a major constituentwithin and amongst A. millepora colonies was thepreviously identified ‘type A associates’, indicating thatthis group of Gammaproteobacteria is associated withhealthy corals of the GBR. Not only are these ribotypescommon on various coral species of unrelated genera(Pocilloporidae and Acroporidae, [22]) and when certainribotypes within this cluster disappear from the coralassociated community, subsequent increases are seen inthe presence of members of the Vibrionaceae, Enter-obacteriaceae and Alphaproteobacteria [22]. Similarly, ourstudy on A. millepora observed that in samples notdominated by type A associates, one sample was complete-ly dominated by a member of the Enterobacteriaceae and asecond sample contained a Vibrio ribotype, and higherdensity of bands representing Alphaproteobacteria was seenin additional samples. Whilst PCR bias cannot be com-pletely excluded as a confounding factor, the observedpatterns of potential species interactions in the present studyare supported not only by our previous findings [22], butalso by those of a study utilising differing amplificationmethods that observed simultaneous increases of Vibrio andloss of type A associates during bleaching [5].
In addition to type A associates, our DGGE resultshighlight a group of Alphaproteobacteria that alsofrequently associate with colonies of A. millepora (thisstudy) and A. hyacinthus [22] around Heron Island. These
Alphaproteobacteria ribotypes fall within a group thatcontains several nitrogen fixers (Fig. 2; [34]) and anumber of endosymbionts, including those from gutlessworms found on tropical and subtropical coral reefs(Fig. 2; [4, 9]). As a specific group of ribotypes ofAlphaproteobacteria are present in the microbial commu-nities of several apparently healthy colonies of twodifferent species at different times, we propose them hereas a second group of key associates of corals of the GBRtermed ‘type B associates’ until further phylogeneticgrouping and description can be provided. Given theirrelatedness to a group of nitrogen fixers and endo-symbionts of marine invertebrates, there is the potentialfor ‘type B associates’ to contribute nutrients to the coralhost. These are interesting topics for future investigationthat may further clarify the roles of the bacterial associatesof reef-building corals, building on recent work onnitrogen fixation of coral-associated microbes [21, 26].Future challenges lie in culturing both type A and Bassociates in order to assess their potential nutritional orprobiotic contributions to the coral holobiont, but it ispossible that culturing will be complex if these specificgroups of Gammaproteobacteria and Alphaproteobacteriaare dependent on live coral host or their obligatedinoflagellate symbionts (Symbiodinium) for growth.
Only a few of the cultured isolates were observed in theDGGE analyses. Whilst PCR bias again cannot completelybe excluded, the primers used for DGGE in the presentstudy have the ability to amplify ribotypes from thecultured isolates. Thus, a more likely scenario to the lackof overlap is a combination of culture bias (as describedpreviously in the text) and the inability of the DGGEmethod to detect ribotypes that are present in extremely lowabundance.
Growth Promotion or Inhibition of Bacterial Speciesby the Coral Holobiont
Whilst it is likely that the presence of some bacterialspecies on a coral can affect the abundance of others—atheory supported by a metagenomic analysis on the coralPorites astreoides—in which coral associated bacteria wereshown to carry genes for ‘waging chemical warfare’ [51], itis not known to what extent such interactions are governedby the bacteria themselves or mediated by the coral host.Extracts from scleractinian corals have been shown tocontain antimicrobial compounds [10] but antimicrobialcompounds have also been reported in coral-associatedbacteria [31, 37, 43]. Thus, antimicrobial activity is likelyto arise from both microbial and host sources. Recentstudies showing functional immunological molecules withbacterial binding capacity [19, 20] indicate that corals havethe potential means of controlling their microbial asso-
E.C.E. Kvennefors et al.

ciates. These interactions are further complicated as coral orbacteria-derived quorum-sensing molecules or inhibitorsmay exercise control over the expression of antimicrobialcompounds from coral-associated microbes [37].
In the present study, mucus removed from the coralsupported growth of most bacterial isolates tested. Similar-ly, Sharon and Rosenberg [42] found that isolates from themarine environment, which were not readily observed onthe coral, grew well on sterilised coral mucus and that thenumber of bacteria in the mucus increases dramaticallywhen the mucus was detached from the coral. In particular,Vibrio species proliferated in detached mucus after incuba-tion at 30°C [42]. These results imply that growthpreviously inhibited by the coral holobiont in situ ispermitted once the mucus is removed. Whilst some residualantimicrobial compounds from the coral holobiont arelikely to remain even in detached mucus, it is likely thatwithout replenishment from the coral host or associatedmicrobial community, these rapidly degrade in in vitromodels.
Coral mucus taken directly from corals on the reefwas less supportive of growth for V. coralliilyticus thanmucus from corals that had been kept in aquaria forextensive periods of time. It is not apparent whether thischange is due to altered microbial communities inaquarium corals [17] or a response from the coral host,but our results suggest that changing environmentalconditions affect the ability of mucus to support growthof V. coralliilyticus. These findings do not only imply thatchanges in the external environment may directly and/orindirectly affect bacterial growth on corals, but alsofurther indicate that caution should be exercised wheninterpreting results from studies on coral–bacterial inter-actions performed in aquaria environments. Other studieshave observed loss of antimicrobial activity in bleached ordiseased corals in comparison to apparently healthycolonies [11, 37], and thus a change in the environmentor the health status of the coral allows for changes to themicrobial community.
In conclusion, the present study reveals several impor-tant aspects and hints at an intriguingly complex web ofinteractions in the coral holobiont. Thus, simplified modelsof coral health and disease in assessing the function ofmembers of the associated coral microbial communityseparately may not always provide a basis for accurateinterpretation. Our study further identified a number ofpotential major players in the coral bacterial communityand highlighted the dynamic nature of these in relation toeach other and their surrounding environment.
Acknowledgements This project was supported in part by grantsfrom the PADI Foundation, the Australian Coral Reef Society and theWinifred Violet Scott Foundation.
References
1. Ainsworth T, Fine M, Roff G, Hoegh-Guldberg O (2008) Bacteriaare not the primary cause of bleaching in the Mediterranean coralOculina patagonica. Isme Journal 2:67–73
2. Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990)Basic local alignment search tool. J Mol Biol 215:403–410
3. Ben-Haim Y, Rosenberg E (2002) A novel Vibrio sp pathogen ofthe coral Pocillopora damicornis. Mar Biol 141:47–55
4. Blazejak A, Kuever J, Erseus C, Amann R, Dubilier N (2006)Phylogeny of 16S rRNA, ribulose 1,5-bisphosphate carboxylase/oxygenase, and adenosine 5′-phosphosulfate reductase genes fromgamma- and alphaproteobacterial symbionts in gutless marineworms (Oligochaeta) from Bermuda and the Bahamas. ApplEnviron Microbiol 72:5527–5536
5. Bourne D, Iida Y, Uthicke S, Smith-Keune C (2008) Changes incoral-associated microbial communities during a bleaching event.Isme Journal 2:350–363
6. Bruno JF, Selig ER (2007) Regional Decline of Coral Cover in theIndo-Pacific: Timing, Extent, and Subregional Comparisons.PLoS ONE 2(8): e711
7. Cervino J, Thompson FL, Gomez-Gil B, Lorence EA, Goreau TJ,Hayes RL, Winiarski-Cervino K, Smith GW, Hughen KA, BartelsE (2008) The Vibrio core group induces yellow band disease inCaribbean and Indo-Pacific reef-building corals. J Appl Microbiol105:1658–1671
8. Chimetto LA, Brocchi M, Thompson CC, Martins RCR, RamosHR, Thompson FL (2008) Vibrios dominate as culturablenitrogen-fixing bacteria of the Brazilian coral Mussismilia hispida.Syst Appl Microbiol 31:312–319
9. Dubilier N, Amann R, Erseus C, Muyzer G, Park SY, Giere O,Cavanaugh CM (1999) Phylogenetic diversity of bacterial endo-symbionts in the gutless marine oligochete Olavius loisae(Annelida). Mar Ecol Prog Ser 178:271–280
10. Geffen Y, Rosenberg E (2005) Stress-induced rapid release ofantibacterials by scleractinian corals. Mar Biol 146:931–935
11. Gochfeld D, Aeby G (2008) Antibacterial chemical defenses inHawaiian corals: possible protection from disease. Mar Ecol ProgSer 362:119–128
12. Harborne AR, Mumby PJ, Zychaluk K, Hedley JD, Blackwell PG(2006) Modeling the beta diversity of coral reefs. Ecology87:2871–2881
13. Hill JE, Baiano JCF, Barnes AC (2009) Isolation of a novel strainof Bacillus pumilus from penaeid shrimp that is inhibitory againstmarine pathogens. Journal of Fish Diseases 32:1007–1016
14. Kelman D, Kashman Y, Rosenberg E, Kushmaro A, Loya Y(2006) Antimicrobial activity of red sea corals. Mar Biol 149:357–363
15. Knowlton N, Rohwer F (2003) Multispecies microbial mutualismson coral reefs: the host as a habitat. Am Nat 162:S51–S62
16. Koh EGL (1997) Do scleractinian corals engage in chemicalwarfare against microbes? J Chem Ecol 23:379–398
17. Kooperman N, Ben-Dov E, Kramarsky-Winter E, Barak Z,Kushmaro A (2007) Coral mucus-associated bacterial communi-ties from natural and aquarium environments. FEMS MicrobiolLett 276:106–113
18. Kushmaro A, Loya Y, Fine M, Rosenberg E (1996) Bacterialinfection and coral bleaching. Nature 380:396–396
19. Kvennefors ECE, Leggat W, Kerr CC, Ainsworth TD, Hoegh-Guldberg O, Barnes AC (2010) Analysis of evolutionarilyconserved innate immune components in coral links immunityand symbiosis. Developmental and Comparative Immunology 34(11): 1219–1229
20. Kvennefors ECE, Leggat W, Hoegh-Guldberg O, Degnan BM,Barnes AC (2008) An ancient and variable mannose-binding
Regulation of Bacterial Communities by the Coral Holobiont

lectin from the coral Acropora millepora binds both pathogensand symbionts. Dev Comp Immunol 32:1582–1592
21. Kvennefors ECE, Roff G (2009) Evidence of cyanobacteria-likeendosymbionts in Acroporid corals from the Great Barrier Reef.Coral Reefs 28:547–547
22. Kvennefors ECE, Sampayo E, Ridgway T, Barnes AC, Hoegh-Guldberg O (2010) Bacterial communities of two ubiquitous GreatBarrier Reef corals reveals both site- and species-specificity ofcommon bacterial associates. PLoS ONE 5:e10401
23. Lane DJ (1991) 16S/23S rRNA sequencing. Wiley, New York24. Lee OO, Qian PY (2004) Potential control of bacterial epibiosis
on the surface of the sponge Mycale adhaerens. Aquat MicrobEcol 34:11–21
25. Lesser MP, Bythell JC, Gates RD, Johnstone RW, Hoegh-Guldberg O (2007) Are infectious diseases really killing corals?Alternative interpretations of the experimental and ecologicaldata. J Exp Mar Biol Ecol 346:36–44
26. Lesser MP, Falcon LI, Rodriguez-Roman A, Enriquez S, Hoegh-Guldberg O, Iglesias-Prieto R (2007) Nitrogen fixation bysymbiotic cyanobacteria provides a source of nitrogen for thescleractinian coral Montastraea cavernosa. Mar Ecol Prog Ser346:143–152
27. Ludwig W, Strunk O, Westram R, Richter L, Meier H, BuchnerYA, Lai T, Steppi S, Jobb G, Forester W, Brettske I, Gerber S,Ginhart AW, Gross O, Grumann S, Hermann S, Jost R, Koenig A,Liss T, Luemann R, May M, Nonhoff B, Reichel B, Strehlow R,Stamatakis A, Stuckmann N, Vilbig A, Lenke M, Ludwig T, BodeA, Schleifer K-H (2004) ARB: a software environment forsequence data. Nucleic Acids Research 32:1363–1371
28. Luna GM, Biavasco F, Danovaro R (2007) Bacteria associatedwith the rapid tissue necrosis of stony corals. Environ Microbiol9:1851–1857
29. Muyzer G, de Waal EC, Uitterlinden AG (1993) Profiling ofcomplex microbial populations by denaturing gradient gelelectrophoresis analysis of polymerase chain reaction-amplifiedgenes coding for 16S rRNA. Appl Environ Microbiol 59:695–700
30. Nicolaisen MH, Ramsing NB (2002) Denaturing gradient gelelectrophoresis (DGGE) approaches to study the diversity ofammonia-oxidizing bacteria. J Microbiol Methods 50:189–203
31. Nissimov JRE, Munn CB (2009) Antimicrobial properties ofresident coral mucus bacteria of Oculina patagonica. FEMSMicrobial Letters 292:210–215
32. Pantos O, Bythell JC (2006) Bacterial community structureassociated with white band disease in the elkhorn coral Acroporapalmata determined using culture-independent 16S rRNA techni-ques. Dis Aquat Org 69:79–88
33. Rappe MS, Giovannoni SJ (2003) The uncultured microbialmajority. Annu Rev Microbiol 57:369–394
34. Rincon-Rosales R, Lloret L, Ponce E, Martinez-Romero E (2009)Rhizobia with different symbiotic efficiencies nodulate Acaciellaangustissima in Mexico, including Sinorhizobium chiapanecumsp. nov. which has common symbiotic genes with Sinorhizobiummexicanum (vol 67, pg 103, 2009). FEMS Microbiol Ecol68:255–255
35. Ritchie KB, Dennis JH, McGrath T, Smith GW (1994) Bacteriaassociated with bleached and nonbleached areas of Monastreaannularis. Proc Symp Nat Hist Bahamas 5:75–80
36. Ritchie KB, Smith GW (2004) Microbial communities of coralsurface mucopolysaccharide layers. In: Rosenberg E, Loya Y(eds) Coral Health and Disease. Springer, Berlin, pp 259–264.
37. Ritchie KB (2006) Regulation of microbial populations by coralsurface mucus and mucus-associated bacteria. Mar Ecol Prog Ser322:1–14
38. Rohwer F, Breitbart M, Jara J, Azam F, Knowlton N (2001)Diversity of bacteria associated with the Caribbean coralMontastraea franksi. Coral Reefs 20:85–91
39. Rohwer F, Seguritan V, Azam F, Knowlton N (2002) Diversity anddistribution of coral-associated bacteria. Mar Ecol Prog Ser 243:1–10
40. Rohwer F, Kelley S (2004) Culture-independent analyses of coralassociated microbes. In: Rosenberg E, Loya Y (eds) Coral Healthand Disease. Springer, Berlin, pp 265–277
41. Seutin G, White BN, Boag PT (1991) Preservation of avian bloodand tissue samples for DNA analyses. Canadian Journal ofZoology-Revue Canadienne De Zoologie 69:82–90
42. Sharon G, Rosenberg E (2008) Bacterial growth on coral mucus.Curr Microbiol 56:481–488
43. Shnit-Orland M, Kushmaro A (2009) Coral mucus-associatedbacteria: a possible first line of defense. FEMS Microbiol Ecol67:371–380
44. Sudheesh PS, Xu HS (2001) Pathogenicity of Vibrio para-haemolyticus in tiger prawn Penaeus monodon Fabricius: possiblerole of extracellular proteases. Aquaculture 196:37–46
45. Sunagawa SDT, Piceno YM, Brodie EL, Desalvo MK,Voolstra CR, Weil E, Andersen GL, Medina M (2009)Bacterial diversity and white plague disease-associated com-munity changes in the Caribbean coral Montastraea faveolata.Isme Journal 3:512–521
46. Sussman M, Willis B, Victor S, Bourne DG (2008) Coralpathogens identified for white syndrome (WS) epizootics in theIndo-Pacific. PLoS ONE 3:e2393
47. Swofford LSD. PAUP*, phylogenetic analysis using parsimony(*and other methods). Version 4.0b10. Sinauer, Sunderland,2000.
48. Vandenberghe J, Thompson FL, Gomez-Gil B, Swings J (2003)Phenotypic diversity amongst Vibrio isolates from marine aqua-culture systems. Aquaculture 219:9–20
49. Vizcaino MI, Johnson WR, Kimes NE, Williams K, Torralba M,Nelson KE, Smith GW, Weil E, Moeller PD, Morris PJ (2009)Antimicrobial resistance of the coral pathogen Vibrio coralliilyti-cus and Caribbean sister phylotypes isolated from a diseasedoctocoral. Microb Ecol 59:646–657
50. Ward JR, Lafferty KD (2004) The elusive baseline of marinedisease: Are diseases in ocean ecosystems increasing? PLoS Biol2:542–547
51. Wegley L, Edwards R, Rodriguez-Brito B, Liu H, Rohwer F(2007) Metagenomic analysis of the microbial communityassociated with the coral Porites astreoides. Environ Microbiol9:2707–2719
E.C.E. Kvennefors et al.




















