
Reevaluating What We Know About the Histories of Maize in Northeastern North America: A Review of...
43
Reevaluating What We Know About the Histories of Maize in Northeastern North America: A Review of Current Evidence John P. Hart • William A. Lovis Published online: 14 December 2012 Springer Science+Business Media New York 2012 Abstract The adoption of maize in northeastern North America is often seen as a catalyst for the development of settled village life. In this review we develop a theoretical framework centered on shifting-balance theory (SBT) and domesticated landscapes through which to understand the context for the adoption of maize agriculture in the Northeast. We review micro- and macrobotanical evidence and stable carbon isotope data from various sources to reevaluate maize histories and adoption trajectories. These data are coupled with contributions of subregionally
Transcript of Reevaluating What We Know About the Histories of Maize in Northeastern North America: A Review of...

Reevaluating What We Know About the Historiesof Maize in Northeastern North America: A Reviewof Current Evidence
John P. Hart • William A. Lovis
Published online: 14 December 2012� Springer Science+Business Media New York 2012
Abstract The adoption of maize in northeastern North America is often seen as acatalyst for the development of settled village life. In this review we develop atheoretical framework centered on shifting-balance theory (SBT) and domesticatedlandscapes through which to understand the context for the adoption of maizeagriculture in the Northeast. We review micro- and macrobotanical evidence andstable carbon isotope data from various sources to reevaluate maize histories andadoption trajectories. These data are coupled with contributions of subregionally

Introduction
In temperate northeastern North America (hereafter, Northeast), maize (Zea maysssp. mays) was the primary grain of Native American agricultural systems at thetime of initial European incursions. The advent of maize-based agriculture generallyseveral centuries earlier was long thought by many archaeologists to have been arevolutionary event (e.g., Hyde 1962, p. 17). The adoption of maize, specifically,was often seen as the cause of rapid transitions from mobile, logistical hunting andgathering systems to sedentary agricultural village life. Historically, maizeagriculture was thought to have been a necessary condition for the developmentof cultural elaborations, including the traditional culture-historic taxa of Hopewelland Mississippian (e.g., Hall 1980) and specific historical ethnic groups such as theIroquois of New York (Ritchie 1969; Snow 1995). Several decades ago across muchof the Northeast this transition was thought to have occurred within a century or twoof AD 1000 uncal (e.g., Ritchie 1969; Wright 1966; see Fritz 1990; unless we noteotherwise, dates in this article are calibrated with CALIB 6.01 using the IntCal09dataset [Stuiver and Reimer 1993]).
By the mid-1990s, there was substantiated evidence that maize was present inportions of eastern North America well before AD 1000 (e.g., Crawford et al.1997a; Riley et al. 1994). Various lines of evidence, however, continued to suggestthat there were major intensifications of maize production and shifts to village lifeacross much of the Northeast one or two centuries on either side of AD 1000 (Smith1992). These data include a pronounced increase in the frequency of maizemacrofossils recovered from archaeological sites and the number of archaeologicalsites from which they were recovered. Stable carbon isotope analyses of humanbone collagen showed distinct levels of d13C enrichment, indicating greater amountsof maize consumption across the region (e.g., Schwarcz et al. 1985; Stothers andBechtel 1987). The first occurrences of villages across much of the Northeastalso were linked to this date and the intensification of maize agriculture or thein-migration of maize agriculturists (e.g., Snow 1995).
In reviewing the evidence for maize and villages in portions of the Northeast 11years ago, Hart and Means (2002) found little evidence for a correlation between theappearance of maize macrofossils, enriched d13C values, and villages across theregion. Since then there has been an upsurge in research on the histories of maize inthe Northeast, incorporating new analytical techniques (e.g., Chilton 2006; Greenlee2006; Harrison and Katzenberg 2003; Hart et al. 2007a, 2012; Messner 2011a;Raviele 2010; Watts et al. 2011). This research has changed our understandings ofthe histories of maize in a number of subregions of the Northeast, sometimesradically so (Fig. 1).
Here we provide an overview of current evidence for the histories of maize,complementary plant foods, and settlement patterns in the Northeast. Rather thanbeing a radically transformative event, the adoption of maize involved theincorporation of a new grain resource into existing domesticated landscapes (Terrellet al. 2003) in which other grains were already exploited. The intensification ofmaize-based agriculture at different times by some populations in some areas, manycenturies later, was an extension of long traditions of plant exploitation and, in some
176 J Archaeol Res (2013) 21:175–216
123

cases, agricultural behaviors. What this review unintentionally reveals is support forthe conclusions reached by Hart and Means (2002) of no link between the adoptionand establishment of maize as an agricultural crop and settled village life. Rather, asChilton (1999) has hypothesized for post-AD 1000 interior New England, much ofthe sometimes now long history of maize in the Northeast is associated with at leastseasonally mobile local human populations. Our ability to build robust, subregionalhistories of maize, however, is hindered by the inconsistent availability of multiple,complementary lines of evidence.
The Northeast
The region of interest in this discussion is, broadly speaking, the northeastern part ofthe United States and parts of southeastern Canada, an area that incorporatesnorthern and central New York and adjacent parts of New England, including theupper St. Lawrence drainage, Pennsylvania, and northern Ohio (the Ohio Riverabove the Big Miami confluence), and southern Ontario adjacent to the lower GreatLakes of Erie, Ontario, and Huron to the eastern margins of Lake Michigan. This
Fig. 1 General location of sites mentioned in the text and Table 1: 1, Little Ossipee; 2, 19BN811;3, RI250; 4, Burnham-Shepard; 5, Ingalls; 6, Headquarters; 7, 211-1-1; 8, Bowmans Brook; 9, ParslowField, Westheimer; 10, Deposit Airport 1; 11, Fortin 2, Street; 12, Simmons, Vinette, Wickham;13, Hunter’s Home, Kipp Island; 14, Felix; 15, Broome Tech; 16, Catawissa Bridge; 17, Memorial park;18, Fisher Farm; 19, Backstrum #1; 20, Thorp; 21, Meadowcroft Rockshelter; 22, Grand Banks, Lone Pine;23, Cootes Paradise; 24, Forester; 25, Moatfield Ossuary; 26, Dunn; 27, Schultz, Fletcher, 20SA1276;28, Frazer-Tyra; 29, Schwerdt; 30, Edwin Harness; 31, Childers, Woods; 32, Cahokia; 33, Holding
J Archaeol Res (2013) 21:175–216 177
123

region is highly variable topographically, hydrologically, and climatically, as wellas in terms of precontact population densities, the relative reliance on maize as aprimary food source by contact-period indigenous groups, and archaeologicalknowledge of precontact subsistence regimes. Thus it provides a fruitful spacewithin which to explore the different impacts of maize on local populations at themacroregional, regional, and subregional scales, and at potentially differenttemporal rates.
The contact-period distribution of indigenous ethnic groups across this regionvaries somewhat depending on the author. The circum-Lake Erie and northern LakeOntario basin was home to the Huron, Tobacco, Neutral, and Erie. The area on thewest coast of Lake Huron has been variably attributed to the Sauk, Fox, Mascouten,Chippewa, and Potawatomi. The several components of what are known as Iroquois(Mohawk, Seneca, Cayuga, Onondaga, and Oneida) were centered in the FingerLakes region, the Mohawk River Valley, and north to the St. Lawrence River. Tothe south in the Susquehanna River Valley were the Iroquoian-speaking Susque-hannock. Societies along the coastal and lower riverine areas of New Englandinclude the Massachusetts, Wampanoag, Narragansett, Pequot, and the Penobscot(to the north). This catalog of named regional ethnic groups encompassestremendous variation in population densities and demography, organizational/institutional structure, investment in physical infrastructure, and relative degree ofreliance on maize and/or other cultigens, as well as the seasonal use of indigenousresources in more or less sedentary systems (Trigger 1976). Recognition of thisvariation conditions our assessment of their precontact precursors and thetrajectories and impacts of maize adoption subregionally and locally over differentperiods of time.
At a general scale, the area encompasses biotic zones of temperate continentalforest and temperate mountain ecosystems (United Nations 2001, fig. 33, table 18).This can be refined both latitudinally and altitudinally, with broad-leaf deciduousforests to the south and mixed deciduous/coniferous forest to the north and at higheraltitudes. With the exception of the higher elevations of northern Pennsylvania, NewYork, and New England, the majority of the region sustains an average of 120–140frost-free days, sufficient for local maize landraces; some areas in southern Ohiohave substantially longer average growing seasons (Environmental ProtectionAgency 2008). So-called lake-effect zones can be present locally due to theproximity of the large, temperature-ameliorating Great Lakes. These zones usuallylie to the east or southeast of the water masses and generally have greater seasonalprecipitation and longer frost-free day averages than adjacent inland areas. This is,however, only one of several microclimates present across the region. Currently,mean annual precipitation ranges from as little as 700 mm in southern lowerMichigan to over 1,200 mm in parts of the extreme Northeast, with average Julytemperatures ranging from 22 �C in the west and south to as little as 17–17.5 �C tothe east and at higher elevations (Environmental Protection Agency 2008). Arablesoils dominate most of the margins of the Great Lakes basins and the major rivervalleys of the Northeast, whereas higher elevations tend to have agriculturally pooror rocky soils. A combination of the above factors interacted with local-level socialphenomenon to affect indigenous settlement systems and economies.
178 J Archaeol Res (2013) 21:175–216
123

The degree to which the modern climatic regimes accurately reflect those ofvarious times spanning the millennial history of maize adoption in the region isdebatable. Major past climate events that may have had an impact on maizeproduction include the Medieval Warm Period, c. AD 900–1100, and the subsequentLittle Ice Age, c. AD 1500–1800. The former could well have had region-wide andlatitudinal affects on growing seasons, thereby potentially contributing to increasesin the spatial distribution of maize and local crop abundances. These climatevariations also might have resulted in regional or subregional microclimatic zonesmore favorable to sustainable maize production. That said, there is continued debateabout how much temperature may have increased or decreased during these periods;temperatures may have been as little as only 1 �C warmer or cooler than modernaverages. These factors achieve increasing salience as one applies specifictheoretical perspectives to understanding maize adoption and impact across theregion.
Maize agriculture adoption and intensification
Plants are sessile organisms. Animals, via the movement of propagules, mediatemany plant dispersals (e.g., Kays et al. 2011). Humans are the ultimate plantdispersers, with the ability to move propagules and entire plants extreme distancesto establish new populations. Maize, of course, is an excellent example. From itsbeginnings as a crop some 9,000 years ago in the central Balsas River basin ofMexico (Matsuoka et al. 2002; Piperno et al. 2009; Ranere et al. 2009), over thecourse of millennia humans dispersed maize to the northern and southern limits ofproduction in the Western Hemisphere (e.g., Boyd and Surette 2010; Staller et al.2006; Vigouroux et al. 2008). Through natural and artificial selection andhybridization, this dispersion resulted in the evolution of the hundreds of racesand thousands of varieties collected and described historically (Hallauer andMiranda 1988). After the arrival of Europeans, maize was dispersed around theglobe, which resulted in the evolution of new races and varieties (e.g., Camus-Kulandaivelu et al. 2006; McCann 2004; Rebourg et al. 2004).
Maize in the Northeast historically was dominated by Northern Flint (Brown1947) and prehistorically by its probable archaeological expression, Eastern 8-rowmaize (Crawford et al. 2006; Wagner 1987). Northern Flint shows geneticdivergence from other maize races to the extent it could be considered a separatespecies (Camus-Kulandaivelu et al. 2006; Doebley et al. 1986). This implies thatmaize was present in at least portions of the region for a long period of time,sufficient for natural and artificial selection to result in the evolution of NorthernFlint and its many landraces (e.g., Azar et al. 1997). This time requirement is borneout by recent advances in our knowledge of the history of maize across the region(e.g., Crawford et al. 1997a; Hart et al. 2007a; Raviele 2010; Riley et al. 1994).What processes were involved in the establishment, perpetuation, and evolution ofmaize in the Northeast?
Each time maize was introduced to a new local human population in theNortheast, the initial planting represented a founder event (Hart 1999a; King 1987).
J Archaeol Res (2013) 21:175–216 179
123

The amount of genetic variation present in such founder populations depends on theextent of genetic variation in the parent population, the sample of that population’sgenetic variation represented in the introduced seed, and any subsequent samplingof that seed done by the adopting local human population (Hart 1999a, 2008a; seeGillespie 1998; Rousset 2004). The low genetic diversity of Northern Flint relativeto Mexican highland and tropical lowland races is most likely explained as being aresult of serial founder effects as maize spread into the Northeast from the AmericanSouthwest (Vigouroux et al. 2008, p. 1250). The survival of any given maizefounder population depended on whether the local environmental conditions andmanagement of the crop by the interacting local human population were favorablefor its genotypes and the consequent plasticity of its phenotypes, i.e., genetic–environmental interactions (Mercer and Perales 2010).
The degree of human management in any given local context into which it wasintroduced depended on the perception of maize’s value relative to otherresources—how expensive maize was to grow in terms of time and energy perunit return relative to the procurement of other food resources, or marginal cost(Gremillion 1996a; Hart 1999a; Winterhalder and Goland 1997). If maize was notof sufficiently significant value and little effort was consequently applied to orexpended on crop management, the founder population was likely to remain small.Small, spatially isolated populations were likely to have gone extinct because ofstochastic climatic events (e.g., hail storms, late or early frosts, drought, floods),animal predation, and other conditions not controlled by the adopting local humanpopulation (Johannessen et al. 1970). If a small maize founder population survivedin the absence of gene flow, inbreeding depression would have had deleteriouseffects on productivity over time (e.g., Hallauer and Miranda 1988; Jugenheimer1976), causing its value relative to other resources to diminish and perhaps itsabandonment as a crop. Establishment of a replacement maize population dependedon the ability and desire of the local human population to obtain a new supply ofseed stock.
The only way for a maize population to increase its size was for its interactinglocal human population to plant larger numbers of kernels. Where maize was not afocal resource, the most likely means for population expansion and the perpetuationof maize populations was to be subdivided among components of dispersed localhuman populations (Hart 1999a). The establishment of numerous demes (subpop-ulations partially isolated from gene flow from one another) in a range ofenvironmental conditions increased the size of the population as well as thepopulation’s genetic variation through genetic drift, sampling of seed stock, andlocalized natural and artificial selection. Such metapopulations (subdividedpopulations) comprising numerous demes (see van Heerwaarden et al. 2010), arean important component of Wright’s (1932, 1978) shifting-balance theory (SBT) ofevolution that he developed on the basis of his knowledge of and experience withagricultural breeding (Provine 1986).
A key conceptual component of SBT is the fitness landscape. Fitness landscapesare n-dimensional but most easily conceived of in three dimensions, with the x andy axes representing alleles of two genes or the states of two phenotypic traits and thez axis representing fitness (see Niklas 1997, pp. 47–52). Conceptualized as a
180 J Archaeol Res (2013) 21:175–216
123

topographic map, peaks represent high-fitness states and valleys low-fitness states.Given enough genetic variation, selection will always act to increase fitness and thepopulation will climb the nearest fitness peak (local optimum/optima), even if that isnot the highest peak (global optimum) available. The issue then becomes howpopulations can access higher fitness peaks if selection acts to maintain thepopulation on its existing peak. Wright’s (1932) solution was SBT.
SBT consists of three phases. In phase I, demes of a population move across thefitness landscape, so-called adaptive walks (Niklas 1997), as a result of genetic drift,which is more likely to have an effect on genetic structure in smaller than in largerpopulations/demes (Gillespie 1998). In phase II, one or more demes ascend higherfitness peaks as a result of selection. During phase III, the fitter demes pass on theirfavorable gene complexes to other demes as a result of out-migration, thusincreasing the fitness of the entire metapopulation. Although problematic as ageneral model of adaptive evolution in natural populations (e.g., Rousset 2004),SBT does serve as a useful heuristic for adaptive evolution in agricultural settingsgiven the agricultural foundations of Wright’s theory; human–crop interactionsprovide analogs for each phase (e.g., van Heerwaarden et al. 2010; see Hall 1980 fora similar interpretation without reference to Wright).
Genetic drift and sampling within demes increased the amount of geneticvariation within the maize metapopulation (SBT phase I). Environmental variationincreased for the population as a whole through dispersal of demes across thephysical landscape (also diminishing the probability of population extinction bystochastic environmental events) and through exposure to varying managementpractices by units of the dispersed local human population. The interactions ofgenetic, environmental, and management variation would situate the demes acrossthe fitness landscape, with some climbing higher fitness peaks than others (SBTphase II) (Hart 2008a). While some demes may have gone extinct, new demes couldbe established with seed stock from a surviving deme or demes (Dyer and Taylor2008; van Heerwaarden et al. 2010). The perpetuation of any given deme and themetapopulation as a whole depended on the value of maize relative to otherresources—whether or not the interacting local human population was willing toexpend time and energy on its production.
As maize became more productive and its costs diminished relative to other foodsources, coevolutionary relationships were established with local human popula-tions (Hart 1999a; Rindos 1984). Increased productivity would include more plantssurviving to produce grain and/or increased production per ear, including more and/or larger kernels (see Crawford et al. 2006). Increased human attention andmanagement effort altered the fitness landscape, potentially resulting in higherfitness peaks. Higher fitness resulted in lowered costs relative to other resources,with humans increasing the amount of maize sown. Any deme that was able toascend these higher peaks had the potential to spread as humans exchanged seed(SBT phase III) (e.g., Louette et al. 1997). The productivity of the resulting newdemes relative to the originating deme depended on new microenvironmentalconditions, including such factors as local soil conditions and crop managementpractices. Technological innovations favorable to maize productivity also wouldhave shifted fitness landscapes, potentially resulting in maize demes ascending
J Archaeol Res (2013) 21:175–216 181
123

higher fitness peaks and, in turn, greater maize productivity and lowered costsrelative to other resources. The perpetuation of these more productive varieties ofmaize depended on the successful human intra- and intergenerational transmissionof seed stock, management practices, and tools. It also was necessary for successfulintra- and interhuman population maize dispersal (Hart 1999a, 2001).
The replacement of a deme with seed from a more productive deme would differfrom initial founder events because the adopting farmers would obtain enough seedto sow the desired, presumably large numbers of plants (van Heerwaarden et al.2010). If there was a major commitment to maize, the planting was extensive.Depending on the degree to which the agroecological conditions varied from that ofthe originating deme, productivity of the new deme might not have been as great. Insuch circumstances, however, farmers generally obtain seed from nearby sources(e.g., kin; Badstrue et al. 2007), so the extent of agroecological variation wouldlikely be low and productivity would not suffer greatly. The gradual dispersal ofhighly productive landraces of Northern Flint, adapted to local conditions, likelyoccurred under such a scenario. Demes and consequently metapopulations of suchlocally adapted landraces were expandable according to the needs of the interactinglocal human populations and the extent of available arable land, the nature ofagricultural management practices, and available labor. The spread of NorthernFlint across the Northeast would have been a gradual ‘‘diffusion wave’’ (Eswaran2002) that resulted in the evolution of different landraces as a consequence ofnatural and artificial selection and hybridization. Selection would have favoredadvantageous genotypes as Northern Flint spread from its location of evolution viaseed flow and farmers quickly assessed the productivity of new landraces obtainedfrom varying distances (Dyer and Taylor 2008). As Dyer and Taylor (2008, p. 474)suggest, maize evolution included complex social processes of seed flow and farmerassessments in addition to natural selection (e.g., Louette et al. 1997; Pressoir andBerthaud 2004).
Maize and domesticated landscapes
The processes described above reflect both opportunistic and continual evolution ofmaize and human–maize interactions. Variation in the exploitation of maize isexpected at any given temporal and spatial resolution—the mosaic that Smith (1992,p. 293) described as ‘‘different regions exhibiting variation in the types of maizebeing grown and its dietary importance relative to indigenous crops and wild animaland plant resources.’’ Smith (1992, p. 293) further suggested that while the exacttiming and routes of maize diffusion throughout the east were important issues, theprimary object of inquiry should remain ‘‘the role of food production in Woodlandperiod cultural development.’’ Such a focus requires a shift in conceptualization,from one of dichotomies (forager vs. farmer) to one of gradations (e.g., Smith 2001).One such conceptualization is the domesticated landscape (Terrell 2008; Terrell andHart 2008; Terrell et al. 2003).
Any group of humans in any landscape setting exploits many different species ofplants and animals. Depending on the nature, intensity, and duration of that
182 J Archaeol Res (2013) 21:175–216
123

exploitation, there may be genetic and resultant phenotypic changes in a givenspecies that constitute the traditional hallmarks of domestication. Maize is anarchetypical example of this kind of evolution, from a regionally restricted grasswith inedible kernels (Smalley and Blake 2003) to a grain crop that does notperpetuate without the aid of humans. But such extreme cases distract us from theoverall impacts of human behaviors on plant and animal populations. From thisstandpoint it makes more sense to gauge domestication on the basis of itsperformance than by its occasional genetic and phenotypic consequences. To thatend, the local population of a species can be considered domesticated when a localpopulation of another species knows how to exploit it (Terrell and Hart 2008,p. 330). By extension, entire landscapes can be considered domesticated as, in thiscontext, humans understand how to exploit the range of species on that landscape toobtain food, fuel, building materials, and other resources necessary for survival. Theadoption of maize or any other crop, then, is simply the addition of another speciesto the domesticated landscape.
Each of the species in a domesticated landscape occupies a distinct fitnesslandscape that is shaped, at least in part, by human interactions with it. The fitnesslandscapes of domesticated species are linked, given that changes in human patternsof exploitation of one species was likely to impact human exploitations of otherspecies (Hart 1999a). Maize adopted into any domesticated landscape became partof the suite of domesticated (as defined here) plants exploited by humans bothwithin and beyond any established agroecology. The nature of maize’s fitnesslandscape was determined in part by the extent to which human local populationsdevoted time and energy on its production relative to other species in thedomesticated landscape. Any changes in the marginal cost of maize productionrelative to those of other species shifted the fitness landscapes of all domesticatedspecies. Similarly, any change in the marginal cost of other domesticated specieschanged the fitness landscape of maize.
During late prehistoric times in many areas of the Northeast, the fitnesslandscapes of maize, bean, and squash became highly interwoven in intensivepolycultural agricultural systems (Mt Pleasant 2006). Early in its history of adoptioninto domesticated landscapes, however, maize was just one of a number of exploitedgrains in any given domesticated landscape. These other grains include such speciesas little barley (Hordeum pusillum), maygrass (Phalaris caroliniana), canary grass(Phalaris canariensis), panic grass (Panicum spp.), barnyard grass (Echinochloaspp.), and wild rice (Zizania spp.), among others (Crawford and Smith 2003). Theexploitation of these species varied throughout the Northeast (e.g., Asch Sidell2008; Simon 2000; Simon and Parker 2006). It is likely that the adopting localhuman populations recognized maize as just another starchy grain (e.g., Fritz 1993,p. 56) rather than a thoroughly exotic and intrinsically unusual plant/food source orsacred plant (e.g., Scarry 1993, p. 90).
The fact of prehistoric domesticated landscapes in the Northeast has becomeincreasingly clear over the past several decades. It is well documented that NativeAmericans persistently exploited large numbers of plant and animal species in theNortheast (e.g., Asch and Asch 1985; Asch Sidell 2008; Crawford 2011; Crawfordand Smith 2003; Simon 2000; Smith 1992; Styles 2011). The intensive exploitation
J Archaeol Res (2013) 21:175–216 183
123

of a particular series of seed-bearing annual plants in the riverine interior led to thedevelopment of what is commonly referred to as the Eastern Agricultural Complex(EAC) (Ford 1985; Smith 1992). The seeds of several plants, such as goosefoot(Chenopodium berlandieri), exhibit morphological characteristics indicative ofgenetic changes that resulted from selection in human-created/maintained settingsand are thus referred to in the literature as ‘‘domesticates.’’ The seeds of otherplants, such as little barley, commonly found in conjunction with these ‘‘domes-ticates,’’ lack such characteristics and are less frequently referred to as ‘‘domes-ticates,’’ although human-mediated range extensions of such plants indicate thelikelihood of cultivation (Asch and Asch 1985; Asch Sidell 2008).
Seeds of other widely and intensively exploited annual plants that lackmorphological changes and that were probably not cultivated in the traditionalsense have not been considered ‘‘domesticated,’’ even though they may have spreadacross the Northeast through purposeful human intervention (e.g., wild rice: Zizaniaspp.; Arzigian 2000; Crawford and Smith 2003). This is the case even when thosespecies (or other species in the same genus) were ‘‘domesticated’’ elsewhere (e.g.,Chenopodium in New England; George and Dewar 1999). Also increasinglyrecognized is the impact of human behavior on tree species diversity anddistributions across landscapes in eastern North America, including nut and fruit-bearing trees (e.g., Abrams and Nowacki 2008; Black et al. 2006; Moore and Dekle2010). The exploitation of fruits from shrubs and vines also is consistentlyrepresented in the archaeological record (e.g., Asch Sidell 2008; Crawford andSmith 2003; Simon 2000), although such plants are not considered ‘‘domesticated’’in the classic sense. The regular association of early maize agricultural societies infloodplain or wetland settings in much of the Northeast (e.g., Crawford et al. 1997b;Lovis et al. 2001) is an example of a category of domesticated landscapes intowhich maize was incorporated.
Maize macrofossil evidence
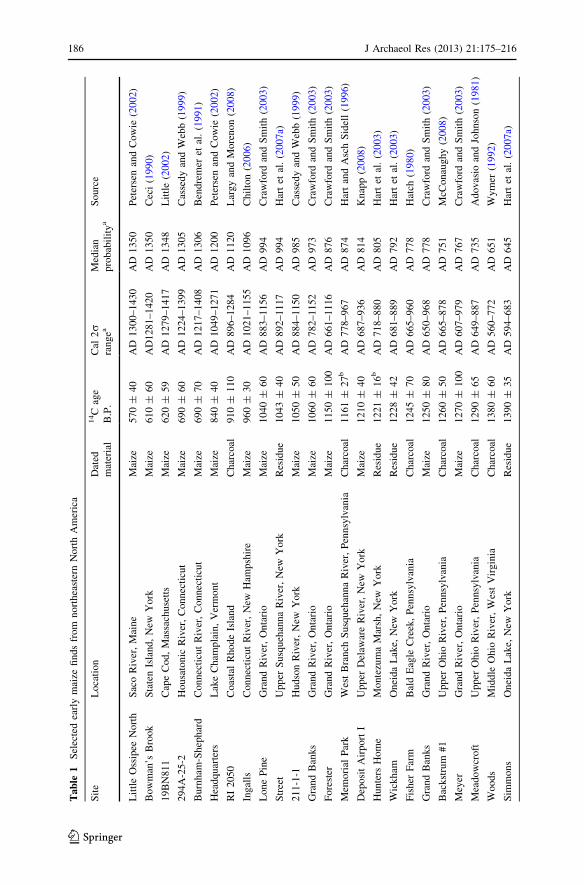
The recovery of maize macrofossils is a commonplace occurrence on post-AD900–1000 sites and components across much of the Northeast (Hart 1999b, 2008b;Hart and Rieth 2002). This was true even before the advent and common use offlotation processing in the 1970s and 1980s (e.g., Ritchie and Funk 1973). Althoughmaize had been found in earlier site components prior to the advent of flotation,particularly in the riverine interior, later direct accelerator mass spectrometry(AMS) dating proved many examples to be contaminants from late prehistoricoccupations (e.g., Conard et al. 1984). Even after the widespread use of flotation, therecovery of maize macrofossils in an early context was a rare and noteworthy event(e.g., Riley et al. 1994). Over the last few decades such finds have been doubted inthe absence of confirmatory direct AMS dating (Crawford et al. 1997a; Riley et al.1994), because there is no necessary chronological connection between an object ofchronological interest (e.g., a maize kernel) and spatially associated wood charcoal.
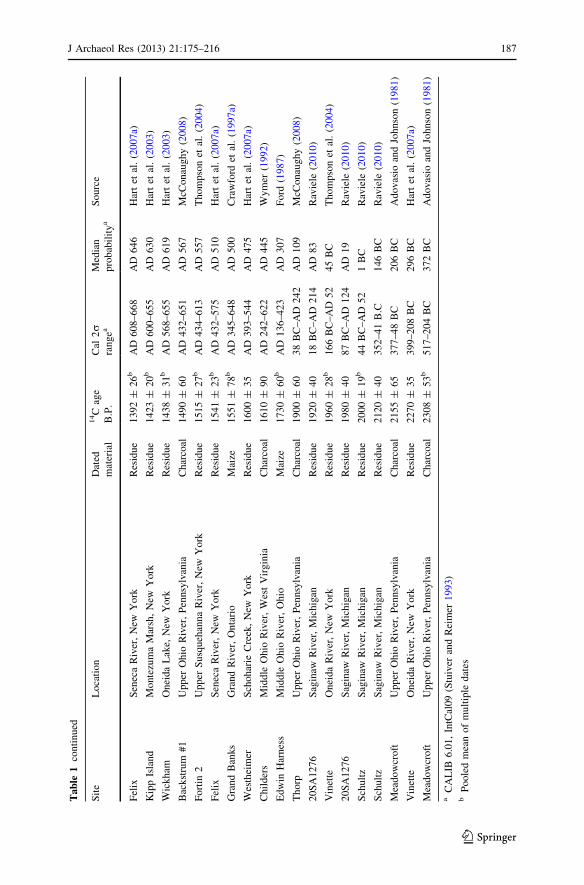
Despite the widespread adoption of flotation, intensive examination of thearchaeological record as a result of cultural resource management, and the now
184 J Archaeol Res (2013) 21:175–216
123

common practice of the direct AMS dating of potentially early crop macrofossils,little has changed in the macrofossil evidence for early maize in the Northeast overthe past decade. Late 20th century reports from southern Ontario (Crawford et al.1997a) and Ohio (Ford 1987) remain the earliest direct-dated macrofossil evidence(Table 1). Direct AMS dates on maize macrofossils in the Delaware Valley of NewYork (Knapp 2008) and across New England (Asch Sidell 2002; Chilton 2006;Little 2002; see Table 1), though centuries later, have added to our knowledge ofthe history of maize use in these areas. Other potentially early macrofossils, such asthose from very early contexts at Meadowcroft Rockshelter in southwesternPennsylvania (Adovasio and Johnson 1981), have not been directly dated (Table 1).Despite the apparent resolution of the dating controversy at Meadowcroft (Goldbergand Arpin 1999) and the undoubtedly early age of the maize macrofossils at thatsite, as with all potentially early macrofossils in the Northeast direct AMS dating isneeded to fully assess their chronological status. Given that AMS laboratoriesgenerally accept less than 5 mg of material for assay, it is not necessary tocompletely destroy individual macrofossils to obtain direct AMS dates (e.g.,Grobman et al. 2012).
Understanding the factors behind patterns of rare/nonexistent early and common/ubiquitous late macrofossils is important to substantive conceptualizations of thehistory of maize in the Northeast. How meaningful is the recovery of a smallamount of maize macrofossils at a short-term occupation site dating 2000 years agorelative to a large amount recovered from a village site occupied over the course ofdecades 700 years ago? One hypothesis is that the scarcity of early maizemacrofossils in the Northeast is simply a function of the crop’s limited use in theregion prior to AD 900–1000. A consideration of archaeological site formationprocesses and archaeological sampling methods, however, suggests that the presentrecord of maize macrofossils is incomplete.
The preservation of maize macrofossils at open-air archaeological sites is afunction of the length and intensity of site use and how maize was used at that site(Hart 1999a, 2008a; also see Lopinot 1992, pp. 55–59; Miksicek 1987). Maize entryinto the archaeological record is affected by the intensity, length, and frequency ofsite use. The effects of maize use include how it was prepared and cooked and theintensity and frequency of its use during any given occupation. The interaction ofsite use and maize use determine the probability that maize would be charredand lost or purposefully disposed of in a context favorable for preservation (Hart2008a). The probability of maize entering and preserving in the archaeologicalrecord at a small, short-term occupation site that had limited use of maize is verylow relative to a larger, longer-term occupation site where maize was used moreintensively.
King (1987) and Wright (2003) found that dried flint kernels are more likely tosurvive during charring than are fresh, moist kernels, which tend to pop and distort.Wright (2003, p. 582) and Goette et al. (1994, p. 12) found that moist kernels aremost likely preserved with short exposure at low temperatures in reducingatmospheres. Other experiments have shown that the most likely form of maize tobe preserved in the archaeological record is hominy (Goette et al. 1994; King 1987;Wright 2003). The crescent-shaped kernels missing pericarps, points of attachment,
J Archaeol Res (2013) 21:175–216 185
123
Tab
le1
Sel
ecte
dea
rly
mai
zefi
nds
fro
mn
ort
hea
ster
nN
ort
hA
mer
ica
Sit
eL
oca
tion
Dat
edm
ater
ial
14C
age
B.P
.C
al2r
ran
gea
Med
ian
pro
bab
ilit
yaS
our
ce
Lit
tle
Oss
ipee
No
rth
Sac
oR
iver
,M
ain
eM
aize
57
0±
40
AD
13
00
–1
430
AD
13
50
Pet
erse
nan
dC
owie
(20
02)
Bow
man
’sB
rook
Sta
ten
Isla
nd,
New
Yo
rkM
aize
61
0±
60
AD
12
81
–1
420
AD
13
50
Cec
i(1
99
0)
19B
N811
Cap
eC
od,
Mas
sach
use
tts
Mai
ze620
±5
9A
D1
27
9–
141
7A
D1
34
8L
ittl
e(2
00
2)
29
4A
-25
-2H
ou
sato
nic
Riv
er,
Con
nec
ticu
tM
aize
69
0±
60
AD
12
24
–1
399
AD
13
05
Cas
sed
yan
dW
ebb
(19
99)
Bur
nh
am-S
hep
har
dC
on
nec
ticu
tR
iver
,C
onn
ecti
cut
Mai
ze6
90
±7
0A
D1
21
7–
140
8A
D1
30
6B
endr
emer
etal
.(1
99
1)
Hea
dq
uar
ters
Lak
eC
ham
pla
in,
Ver
mo
nt
Mai
ze8
40
±4
0A
D1
04
9–
127
1A
D1
20
0P
eter
sen
and
Cow
ie(2
00
2)
RI
20
50
Co
asta
lR
hod
eIs
lan
dC
harc
oal
91
0±
11
0A
D8
96
–1
284
AD
11
20
Lar
gy
and
Mo
ren
on
(20
08)
Ing
alls
Co
nn
ecti
cut
Riv
er,
New
Ham
psh
ire
Mai
ze9
60
±3
0A
D1
02
1–
115
5A
D1
09
6C
hil
ton
(20
06)
Lo
ne
Pin
eG
rand
Riv
er,
On
tari
oM
aize
10
40
±6
0A
D8
83
–1
156
AD
99
4C
raw
ford
and
Sm
ith
(20
03)
Str
eet
Up
per
Su
squ
ehan
na
Riv
er,
New
Yo
rkR
esid
ue
10
43
±4
0A
D8
92
–1
117
AD
99
4H
art
etal
.(2
00
7a)
21
1-1
-1H
ud
son
Riv
er,
New
Yo
rkM
aize
10
50
±5
0A
D8
84
–1
150
AD
98
5C
asse
dy
and
Web
b(1
99
9)
Gra
nd
Ban
ksG
rand
Riv
er,
On
tari
oM
aize
10
60
±6
0A
D7
82
–1
152
AD
97
3C
raw
ford
and
Sm
ith
(20
03)
Fo
rest
erG
rand
Riv
er,
On
tari
oM
aize
11
50
±1
00
AD
66
1–
11
16A
D8
76
Cra
wfo
rdan
dS
mit
h(2
00
3)
Mem
ori
alP
ark
Wes
tB
ran
chS
usq
ueh
ann
aR
iver
,P
enn
sylv
ania
Cha
rcoa
l1
16
1±
27
bA
D7
78
–9
67
AD
87
4H
art
and
Asc
hS
idel
l(1
99
6)
Dep
osi
tA
irp
ort
IU
pp
erD
elaw
are
Riv
er,
New
Yo
rkM
aize
12
10
±4
0A
D6
87
–9
36
AD
81
4K
nap
p(2
00
8)
Hu
nte
rsH
om
eM
ont
ezu
ma
Mar
sh,
New
Yo
rkR
esid
ue
12
21
±1
6b
AD
71
8–
88
0A
D8
05
Har
tet
al.
(20
03)
Wic
kh
amO
nei
daL
ake,
New
Yo
rkR
esid
ue
12
28
±4
2A
D6
81
–8
89
AD
79
2H
art
etal
.(2
00
3)
Fis
her
Far
mB
ald
Eag
leC
reek
,P
enn
sylv
ania
Cha
rcoa
l1
24
5±
70
AD
66
5–
96
0A
D7
78
Hat
ch(1
98
0)
Gra
nd
Ban
ksG
rand
Riv
er,
On
tari
oM
aize
12
50
±8
0A
D6
50
–9
68
AD
77
8C
raw
ford
and
Sm
ith
(20
03)
Bac
kst
rum
#1
Up
per
Oh
ioR
iver
,P
enns
ylv
ania
Cha
rcoa
l1
26
0±
50
AD
66
5–
87
8A
D7
51
McC
on
aug
hy
(20
08)
Mey
erG
rand
Riv
er,
On
tari
oM
aize
12
70
±1
00
AD
60
7–
97
9A
D7
67
Cra
wfo
rdan
dS
mit
h(2
00
3)
Mea
do
wcr
oft
Up
per
Oh
ioR
iver
,P
enns
ylv
ania
Cha
rcoa
l1
29
0±
65
AD
64
9–
88
7A
D7
35
Ad
ov
asio
and
Joh
nso
n(1
98
1)
Wo
od
sM
idd
leO
hio
Riv
er,
Wes
tV
irg
inia
Cha
rcoa
l1
38
0±
60
AD
56
0–
77
2A
D6
51
Wy
mer
(19
92)
Sim
mo
ns
On
eida
Lak
e,N
ewY
ork
Res
idu
e1
39
0±
35
AD
59
4–
68
3A
D6
45
Har
tet
al.
(20
07a
)
186 J Archaeol Res (2013) 21:175–216
123
Tab
le1
con
tin
ued
Sit
eL
oca
tion
Dat
edm
ater
ial
14C
age
B.P
.C
al2r
ran
gea
Med
ian
pro
bab
ilit
yaS
our
ce
Fel
ixS
enec
aR
iver
,N
ewY
ork
Res
idu
e1
39
2±
26
bA
D6
08
–6
68
AD
64
6H
art
etal
.(2
00
7a)
Kip
pIs
lan
dM
ont
ezu
ma
Mar
sh,
New
Yo
rkR
esid
ue
14
23
±2
0b
AD
60
0–
65
5A
D6
30
Har
tet
al.
(20
03)
Wic
kh
amO
nei
daL
ake,
New
Yo
rkR
esid
ue
14
38
±3
1b
AD
56
8–
65
5A
D6
19
Har
tet
al.
(20
03)
Bac
kst
rum
#1
Up
per
Oh
ioR
iver
,P
enns
ylv
ania
Cha
rcoa
l1
49
0±
60
AD
43
2–
65
1A
D5
67
McC
on
aug
hy
(20
08)
Fo
rtin
2U
pp
erS
usq
ueh
ann
aR
iver
,N
ewY
ork
Res
idu
e1
51
5±
27
bA
D4
34
–6
13
AD
55
7T
ho
mps
on
etal
.(2
00
4)
Fel
ixS
enec
aR
iver
,N
ewY
ork
Res
idu
e1
54
1±
23
bA
D4
32
–5
75
AD
51
0H
art
etal
.(2
00
7a)
Gra
nd
Ban
ksG
rand
Riv
er,
On
tari
oM
aize
15
51
±7
8b
AD
34
5–
64
8A
D5
00
Cra
wfo
rdet
al.
(19
97
a)
Wes
thei
mer
Sch
ohar
ieC
reek
,N
ewY
ork
Res
idu
e1
60
0±
35
AD
39
3–
54
4A
D4
75
Har
tet
al.
(20
07a
)
Chi
lder
sM
idd
leO
hio
Riv
er,
Wes
tV
irg
inia
Cha
rcoa
l1
61
0±
90
AD
24
2–
62
2A
D4
45
Wy
mer
(19
92)
Ed
win
Har
nes
sM
idd
leO
hio
Riv
er,
Oh
ioM
aize
17
30
±6
0b
AD
13
6–
42
3A
D3
07
Fo
rd(1
98
7)
Th
orp
Up
per
Oh
ioR
iver
,P
enns
ylv
ania
Cha
rcoa
l1
90
0±
60
38
BC
–A
D2
42
AD
10
9M
cCo
nau
gh
y(2
00
8)
20
SA
12
76
Sag
inaw
Riv
er,
Mic
hig
anR
esid
ue
19
20
±40
18
BC
–A
D214
AD
83
Rav
iele
(20
10)
Vin
ette
On
eida
Riv
er,
New
Yo
rkR
esid
ue
19
60
±2
8b
16
6B
C–
AD
52
45
BC
Th
om
pso
net
al.
(20
04)
20
SA
12
76
Sag
inaw
Riv
er,
Mic
hig
anR
esid
ue
19
80
±40
87
BC
–A
D124
AD
19
Rav
iele
(20
10)
Sch
ultz
Sag
inaw
Riv
er,
Mic
hig
anR
esid
ue
20
00
±1
9b
44
BC
–A
D52
1B
CR
avie
le(2
01
0)
Sch
ultz
Sag
inaw
Riv
er,
Mic
hig
anR
esid
ue
21
20
±40
352–4
1B
.C146
BC
Rav
iele
(20
10)
Mea
do
wcr
oft
Up
per
Oh
ioR
iver
,P
enns
ylv
ania
Cha
rcoa
l2
15
5±
65
37
7–4
8B
C2
06
BC
Ad
ov
asio
and
Joh
nso
n(1
98
1)
Vin
ette
On
eida
Riv
er,
New
Yo
rkR
esid
ue
22
70
±3
53
99
–20
8B
C2
96
BC
Har
tet
al.
(20
07a
)
Mea
do
wcr
oft
Up
per
Oh
ioR
iver
,P
enns
ylv
ania
Cha
rcoa
l2
30
8±
53
b5
17
–20
4B
C3
72
BC
Ad
ov
asio
and
Joh
nso
n(1
98
1)
aC
AL
IB6.0
1,In
tCal
09
(Stu
iver
and
Rei
mer
19
93)
bP
ool
edm
ean
of
mu
ltip
led
ates
J Archaeol Res (2013) 21:175–216 187
123

and sometimes embryos found in many post-AD 900–1000 archaeological contextsin the Northeast are most likely hominy (Goette et al. 1994, p. 8). Myers (2006)suggested that hominy-processing technology arrived late in the Northeast. Despitethis, even at post-AD 1000 nucleated village sites where maize presumably wasused year-round by large, sedentary populations, maize macrofossils tend to befragmentary except when maize cobs were used as fuel, as in smudge pits, or whensubterranean granaries were burned and the contents charred. Macrofossil remains,however, do tend to be more ubiquitous on later rather than on earlier sitesregardless of the absolute amounts recovered.
The probability of maize macrofossils being recovered from an archaeologicalsite is a function of sampling relative to their frequency and distribution on a site,the recovery methods used (e.g., flotation), and subsequent sampling intensity andidentification efforts in the lab. The probability that low-density maize macrofossilspreserved on a site will be found and identified is dependent on high levels of effortbeing expended on each of these factors. The classic open-air eastern NorthAmerican example is the 19 maize fragments found at the Holding site in Illinois,with a pooled mean of two AMS dates on fragments of 2037 ± 41 BP (cal 2r range= 166 BC–AD 52, median probability = 45 BC). These were recovered throughflotation of 5,340 liters of soil and subsequent intensive identification efforts in thelab (Riley et al. 1994). The component from which the maize was recovered isinterpreted to have included seven houses surrounding a central courtyard and 143pit features (Riley et al. 1994)—an intensive occupation relative to contempora-neous sites in the Northeast. A good example from the Northeast is the Grand Bankssite in southern Ontario. This intensively excavated, deeply stratified site, at whichflotation samples were taken from every excavation unit, level, and feature,produced the earliest direct AMS-dated maize in southern Ontario (Crawford et al.1997a; Smith and Crawford 1997). Such complete sampling and intensiveidentification efforts long have been the norm in many archaeological programsin the Northeast (e.g., Parker 1996) but not in others (e.g., Petersen and Cowie2002). Although clearly an important component of any subregional investigation ofthe history of maize, given all the variables affecting macrofossil incorporation intothe archaeological record and all the variables affecting their preservation, recovery,and identification, macrofossils cannot stand alone as evidence for maize use anddomestication.
Microfossil maize evidence
Microfossils, defined here as phytoliths and starch grains, are an additional line ofevidence for examining the histories of maize. Phytoliths are silica structuresformed in and between plant cells (Piperno 2006). Although not every plantproduces phytoliths in the structures consumed by humans, grasses, includingmaize, do. Starch grains are particles produced by plants to store energy (Messner2011a). Both of these microscopic structures preserve in the archaeological recordand can be identified to the species level through comparison with those producedby modern plants (e.g., Hart and Matson 2009; Messner 2011a). Though used only
188 J Archaeol Res (2013) 21:175–216
123

recently in the Northeast (Hart et al. 2003; Messner 2011a), the analysis ofphytoliths and starch in the building of regional histories of maize is routine in otherportions of the Western Hemisphere (e.g., Chavez and Thompson 2006; Pipernoet al. 2009; Ranere et al. 2009). Most relevant to the histories of maize use in theNortheast are maize cobs for phytoliths and starch produced in kernels.
More than a half-century of microfossil research notwithstanding (Piperno 2006;Torrence and Barton 2006), the approach continues to sustain its critics andskeptics, particularly in the arena of phytolith studies (e.g., Rovner 1999).Skepticism most often hinges on issues of overlapping morphotypes across relatedspecies that potentially result in misidentifications or false positives for domesti-cates and the asserted inability of morphotypes to reliably distinguish between wildand domestic varieties. Studies of archaeologically derived starches suffer morefrom taphonomic and preservation issues than identification issues (Cortella andPochettino 1994; Crowther 2012), although similar issues of wild versus domesticvarieties arise here as well (see Messner 2008). We take the position in this contextthat resolution of issues surrounding regional variations in morphologies and thecontinued validation of the ability of morphotypes to isolate domesticates from wildvarieties, coupled with ongoing taphonomic research (Crowther 2012; Messner2008, 2011a; Raviele 2010, 2011), support the systematic use of microfossil data asan independent line of evidence for inception and adoption trajectories. This isparticularly true when we find that such independent lines of evidence arecorroborative or point to new or altered research hypotheses.
Hart and Matson (2009) demonstrate that rondel phytoliths produced in maizecobs from traditional Northeast varieties can be statistically discriminated fromrondel phytoliths produced in the inflorescences of grasses native to the Northeast(also see Hart et al. 2003, 2007a). Piperno (2006) demonstrates that crossbodyphytoliths produced primarily in leaves can be used to discriminate between maizeand other grasses. The great potential of these methods is further demonstrated byHart et al. (2011b), who show that inflorescence rondel phytoliths mirror Zeataxonomy for species and subspecies of teosinte. This work also demonstrates thatrondel phytoliths can be used to discriminate between Mexican maize and teosinte;maize evolved from a subspecies of teosinte Zea mays ssp. parviglumis.
Messner (2008, 2011a, b) compiled a comparative collection of starch grainsfrom 100 species of angiosperms from 63 genera representing 25 families known tohave been exploited by Native Americans in the Northeast. Messner (2008, p. 5)demonstrated that starch from maize kernels can be distinguished from starchproduced by related grasses native to eastern North America. This effort isunprecedented in North America and has enabled other researchers to use starchgrains extracted from secure archaeological contexts to contribute to newexplorations of maize history (e.g., Raviele 2010).
The extraction of maize microfossils from charred cooking residues—formedduring water-based cooking and encrusted on the interior of pottery sherds—has thepotential to overcome some of the uncertainty of macrofossil preservation andrecovery in reconstructing subregional histories of maize (Crowther 2012; Hart et al.2007a; Messner 2011a; Messner et al. 2008; Raviele 2010). These same residuescan be directly dated via AMS (Lovis 1990) to provide precise and accurate dates
J Archaeol Res (2013) 21:175–216 189
123

for maize use (Hart et al. 2003, 2007a; see Hart and Lovis 2007 for the onlysystematic evaluation of AMS dating of residues relative to other datable material).The deposition of microfossils in cooking residues has only recently been thesubject of experimental investigation. Raviele (2011) determined that cob phytolithsare most likely to be deposited in cooking residues when cut green maize cobs arecooked. Starch is most likely to be deposited when mature dried kernels areremoved from the cob before cooking, whether whole, cracked, or ground. Bothphytoliths and starches are likely to occur when green kernels are cut from the cobbefore cooking. Earlier work by Thompson and Mulholland (1994) suggests that theremoval of kernels from cobs displaces small, phytolith-containing pieces of chaffand that these phytoliths are subsequently deposited in cooking residues.
Up to now, experiments have not demonstrated that greater or lesser amounts ofphytoliths and/or starch grains in cooking residues reflect the amounts of maizecooked (Raviele 2011). Yet the large numbers of microstructures produced by anygiven maize ear and their ability to be deposited in cooking residues and survivecharring, and chemical extraction centuries to millennia after their deposition in theresidues, indicate their importance as a source of information on maize use in anygiven vessel. Important is Crowther’s (2012) observation that charred cookingresidues can be ideal contexts for the preservation of starch grains.
The recovery of maize microfossils from such residues depends on water-basedmaize cooking in a container, a context that results in the cooking residue beingcharred and incorporating and preserving microfossils, preservation of the containeror parts thereof and the adhering residue in the archaeological record, andappropriate extraction and identification methods and techniques. Obviously, othermethods of cooking maize may not result in such preservation. It is also possible torecover microfossils from contexts such as soils from hearths, other features, andmiddens (Bozarth 1993; Mulholland 1993), as well as the microcracks andmicrotopography of stone tools (Messner et al. 2008; Piperno and Holst 1998).Other than the pioneering work with starch grain analysis in the Delaware Rivervalley by Messner and colleagues (Messner 2008, 2011a; Messner et al. 2008), theselatter methods have not been used extensively in the Northeast.
Microfossils extracted from charred cooking residues now account for the earliestdirect-dated evidence for maize in the Northeast (Table 1). For Michigan, Raviele(2010) reports on the recovery of maize starch from cooking residues AMS dated asearly as 2120 ± 40 BP (cal 2r = 352–41 BC, median probability = 146 BC) fromthe Schultz site in the Saginaw River valley. Hart et al. (2007a) report maizephytoliths from cooking residue directly AMS dated to 2270 ± 35 BP (cal 2r =399–208 BC, median probability = 296 BC) from the Vinette site in the FingerLakes region. Significantly, phytoliths were recovered from other residues atVinette, and residues from other sites in the region demonstrate the more or lesscontinuous presence of maize through the 15th century AD when maizemacrofossils are common in the archaeological record (Table 1; also see Hartet al. 2003; Thompson et al. 2004). Messner (2008, 2011a) and Messner et al. (2008)report the recovery of maize starch from post-AD 950 cooking residues and stonetools from the Delaware River Water Gap region.
190 J Archaeol Res (2013) 21:175–216
123

Given the results of microfossil analyses of residues in the Northeast and nearbyregions to date (Boyd and Surette 2010; Hart et al. 2003, 2007a; Messner 2011a, b;Messner et al. 2008; Raviele 2010; Thompson et al. 1994, 2004), the construction ofthe history of maize use in any portion of the Northeast must be consideredincomplete until microfossil evidence is available.
Stable carbon isotopes: Human bone
First used in the 1970s for archaeological applications in the Northeast (e.g., Vogeland van der Merwe 1977), stable carbon isotope analysis (SCIA) of human bonecontinues to be used as an aid in reconstructing histories of maize as a measure ofmaize consumption by individuals (Allegretto 2007; Greenlee 2006; Harrison andKatzenberg 2003; Hart et al. 2011a; Rose 2008; van der Merwe et al. 2003a). Themethods and theories of SCIA have been reviewed at length elsewhere (e.g., Tykot2006). Collagen was the traditional material on which SCIA was performed; sincethe late 1990s bone and tooth-enamel apatite (carbonates) also have been assayed.Experiments indicate that collagen reflects the protein component of the diet,whereas carbonates reflect the whole diet (Ambrose 1993; Ambrose and Norr 1993;Tieszen and Fagre 1993; but see Greenlee 2006). As a result, depleted d13C valueson collagen from an individual who consumed maize, but not in quantitiescontributing substantially to protein intake (maize is only approximately 12 %protein), will result in a false negative for maize consumption. The spacing betweencollagen and apatite values can be used to determine whether consumption consistedof C3 protein and C4 carbohydrate or C3 carbohydrate and protein from C4 terrestrialor aquatic/marine-plant-eating animals (Katzenberg 2006). Without taking apatiteand collagen values into account, it is questionable how valuable SCIA is forreconstructing complete histories of maize consumption in a given region.
In addition to the material assayed, sampling is an important factor in theinterpretation of SCIA. Small samples most probably reflect the norms of diets atany given time or place. If the norm is little or no maize consumption, small samplesare likely to miss individuals who consumed detectable amounts of maize.Similarly, if the norm was the consumption of large amounts of maize, individualswho consumed little maize are likely to be missed. Regional interpretations of maizeconsumption based on small samples will be incomplete at best.
The pioneering applications of SCIA in the Northeast that were designed toassess regional histories of maize consumption were based on small samples(Schwarcz et al. 1985; Stothers and Bechtel 1987), were not always chronologicallyand serially complete (Vogel and van der Merwe 1977), and were based solely oncollagen. Although they advanced knowledge of regional histories of maize use,they provided only hints of gross trends of consumption in different regions. Theconfounding effects of marine C4 food sources prevented a confident assessment ofmaize’s contribution to diets on the Atlantic Coast of New England (Bourque andKrueger 1994; Bridges 1994; Little and Schoeninger 1995).
Enhancing earlier results, Harrison and Katzenberg (2003) and Katzenberg(2006) report on the analysis of apatite in many of the same bone samples used in
J Archaeol Res (2013) 21:175–216 191
123

previous southern Ontario studies that analyzed collagen (e.g., Katzenberg et al.1995). The new evidence indicates that maize was consumed by at least somepeople in relatively small amounts by c. AD 500, or perhaps a few centuries earlier,and that consumption gradually increased until c. AD 1000, after which it increasedmore rapidly (Katzenberg 2006, p. 270). Given the small sample sizes, this evidenceof early consumption probably indicates a common dietary pattern.
Studies of relatively large samples have been limited to single post-AD 1000sites; they do not address issues of potential early maize consumption (e.g., Schurrand Redmond 1991) but do contribute to subregional understandings of maizeconsumption levels, population mobility, and potentially migration (e.g., Cook andSchurr 2009). Dewar et al. (2010) and Watts et al. (2011) demonstrate that somepost-AD 1100 populations in the western Lake Erie basin consumed high levels ofmaize while maintaining seasonal mobility. Single-site analysis also has providedinformation on the intensification of maize consumption. Stable carbon isotopeanalysis of dentin collagen and tooth enamel at the 14th century AD MoatfieldOssuary near Toronto indicates a statistically significant enrichment of d13C valuesduring a 25–50-year period that reflects increased consumption of maize over one ortwo human generations (van der Merwe et al. 2003a, b).
Greenlee’s (2002, 2006) large-scale chronological study of the Middle OhioRiver basin suggests an abrupt regional change to the consumption of large amountsof maize at AD 900, with subregional variation in consumption amounts. Greenlee’ssample is very large (n = 380) and chronologically continuous but limited tocollagen analysis. In addition, the pre-AD 900 sample (n = 67) is much smallerthan the post-AD 900 sample (n = 313). Of the pre-AD 900 samples, 52 fallbetween 300 BC (the earliest direct-dated evidence for maize in the Northeast) andAD 900, and these are primarily from burial mound contexts. As a result, theseemingly abrupt increase in maize consumption evident in Greenlee’s analysis maybe an artifact of sampling, reflecting only the protein portion of the diet and perhapsprimarily elite members of pre-AD 900 local populations. Hart et al. (2005)demonstrate that 89 post-AD 950 d13C collagen values with good chronologicalcontrol from the upper Ohio River basin (Farrow 1986; Greenlee 1990, 2002; Scuilli1995; Valko 2001) show no trend for enriched values through time.
The southern Michigan SCIA data reveal two different results of broader analyticimport: the degree of individual variation that might be present in maizeconsumption after its incorporation into local economies and the importance oflongitudinal regional studies. Human bone collagen from the Frazer-Tyra site(n = 33), AD 1200–1400, resulted in d13C values of individuals ranging from –20.1to –14.3. These values indicate that even after its adoption, maize consumption wasvariable; it was an important but not necessarily dominant part of some diets in thislocal population (Muhammad 2010). This finding has important implications forhow populations are sampled for SCIA analysis, as well as the sample sizesnecessary for such analyses; small numbers of individuals may not accurately reflectvariation in maize consumption. Brandt’s (1996) longitudinal analysis of d13C andd15N values of bone collagen from 41 individuals from 11 sites dated c. 3000 BC toAD 1300, while suffering from small sample sizes, nonetheless reveals significanttemporal data on consumption norms. Mean d13C values from four sites dating from
192 J Archaeol Res (2013) 21:175–216
123

3000 BC to c. AD 500 (ranging from –20.5 % to –19.8 %) suggest almost noconsumption of C4 plants in the southern lower peninsula, although a fifth sitedisplays a low C4 diet input. From c. AD 500 to AD 900/1100, enriched mean d13Cvalues reveal increasing C4 plant consumption, initially in the southeastern part ofthe lower peninsula, by c. AD 900 in the Saginaw Valley, and ultimately in northernMichigan (Brandt 1996, pp. 66–76, table 3.1). Such subregional temporal trends,detectable only with a longitudinal sample, reveal differential rates of consumptioneven over small areas and are compatible with regional bulk d13C analyses (e.g.,Hart et al. 2012).
Stable carbon isotopes: Cooking residues
Used to correct 14C dates for isotopic fractionation (Walker 2005), bulk d13C valuesare routinely reported by radiocarbon labs. Maize is the only C4 plant of dietaryconsequence in the Northeast. Therefore, it would seem logical that d13C values oncharred cooking residue from a pot in which maize had been cooked would be moreenriched than those on residue from a pot in which only C3 resources had beencooked (Hastorf and DeNiro 1985). Morton and Schwarcz (2004) suggested thatbulk d13C values reflect the proportion of the resources cooked in a pot that maizeconstituted, assuming a linear relationship, and that a specific bulk d13C value cutpoint can be used to identify maize use in a pot. A series of experimentsdemonstrated that these assumptions are not accurate (Hart et al. 2003, 2007b, 2009;Lovis et al. 2011; Schulenberg 2002); the relationship between maize proportionsand bulk d13C values is nonlinear dependent on the mix of C3 resources and formsof maize cooked in the pot.
Several variables that affect bulk d13C values can mask the presence of maizeand make the interpretation of individual depleted bulk d13C values dependent onprior knowledge about what resources were cooked in a pot and in what proportions(Hart et al. 2007b). These factors include the amount of carbon provided by anygiven resource, the dissolution rates of resources cooked in the pot, and the amountof cooking time (Hart et al. 2007b, 2009). The use of hardwood ash innixtamalization for hominy production also can deplete bulk d13C values, resultingin false-negative interpretations for maize (Lovis et al. 2011). As a result, depletedbulk d13C values cannot be used as evidence that maize was not cooked in a givenpot or by a particular component of a site.
Large regional sets of bulk d13C values on residues can be used in conjunctionwith other lines of evidence as an aid to reconstructing regional histories of maizeuse if the samples are well distributed through time, even if the values remaindepleted through all or most of the sequence. Complementing the microfossilrecord, Hart et al. (2012) demonstrate a significant linear trend for enriched valuesover the course of some 1,300 years in central New York that reflects increasedmaize use and/or changes in the forms of maize cooked (e.g., from whole kernels tomeal), and/or changes in the resources cooked with maize (e.g., wild rice to deer).This trend is almost perfectly negatively correlated with a trend for pottery vesselwall thinning adjusted for vessel girth after AD 200, strongly suggesting that the
J Archaeol Res (2013) 21:175–216 193
123

trend in vessel wall thinning represents increased concern about thermal conduc-tivity with increased processing of maize (Hart 2012). An analysis of bulk d13Cvalues on residues in Michigan failed to produce a similar trend, largely because ofsampling issues involved in compiling a dataset from unrelated studies, which werechronologically incomplete and weighted toward the early end of the sequence (Hartet al. 2012). Of particular interest, however, is the presence of at least one highlyenriched value of –17.5 % at c. 289 cal BC (Beld 1993; Hart et al. 2012),corroborating regional microbotanical data for maize use.
Maize and villages
There has been a traditional correlation between the advent of maize-basedagriculture and the transition to settled village life in the Northeast. For much of the20th century the adoption of maize agriculture was seen as the catalyst for settledvillage life and cultural elaborations. Some saw the appearance of maizemacrofossils in the local archaeological record and the concurrent appearance ofnucleated villages to represent the migration of agriculturists into areas withhunting-gathering populations (e.g., Snow 1995) or the quick transition of mobilehunter-gatherers into settled agriculturists (e.g., Ritchie 1969). These interpretationsreflect misreadings of the probabilities of maize macrofossils entering andpreserving in the archaeological record.
There is ample ethnographic and ethnohistorical evidence for maize agricultureamong populations that maintain mobility. Raymond and DeBoer (2006), forexample, describe populations in Amazonia that spent as little as 3–4 weeks oncultivating and harvesting maize while maintaining their coeval hunting andgathering rounds. Will and Hyde (1917) describe how populations along the upperMissouri River left maize fields after the first or second hoeing to hunt bison andreturn only at harvest time. Kinietz (1940) relates that Chippewa populations livingat Sault Ste. Marie and the region between lakes Superior and Huron left their maizefields after planting, dispersed along the shores of Lake Huron, and returned atharvest time. As described by Day (1978), the Western Abanaki of interior NewEngland and southern Quebec left their maize fields for extended fishing trips,returning occasionally to weed.
These few examples demonstrate how maize could be adopted into domesticatedlandscapes and perpetuated as a valued, if not focal, resource in the absence ofpermanent nucleated villages. Rather, maize was incorporated into the resourcemixes without becoming a dominant resource for long periods of time, a pattern thatcan be demonstrated archaeologically across varying time scales and regions. Forexample, Vierra and Ford (2006) suggest that populations in the northern RioGrande region of New Mexico integrated maize into mobile foraging economies by1000 BC, planting it in a variety of settings near other resources such as pinon. Itwas not until AD 600–900 that maize became a focal resource.
In the Northeast, Chilton (1999, 2006, 2008) suggests that after AD 1000, maize-growing populations in the central Connecticut River valley maintained seasonallymobile subsistence patterns. These ‘‘mobile farmers,’’ like some Algonquian
194 J Archaeol Res (2013) 21:175–216
123

populations recorded in the ethnohistoric record for New England, dispersed afterplanting their crops and returned at harvest time or remained at a site during thegrowing season and dispersed after the harvest; nucleated villages did not develop inthis region until near the time of European incursions. More recently, Dewar et al.(2010) and Watts et al. (2011) have documented high levels of maize consumptionamong some mobile local populations in the western Lake Erie basin severalcenturies before the advent of nucleated village life. In southeastern Ontario, maizemacrofossils are present in the archaeological record at least half a millenniumbefore the advent of nucleated villages (Birch 2010; Warrick 1996; Williamson1990). In central New York and the Saginaw River basin of Michigan, maize wasadopted more than a millennium before the advent of nucleated villages (Hart et al.2007a; Raviele 2010). In the West Branch of the Susquehanna River basin, maizemacrofossils became ubiquitous in the archaeological record several centuries priorto the advent of nucleated villages, which suggests intensive use at hamlets (Hartand Asch Sidell 1996; Nass 2001; Stewart 1993). In the Delaware River basin,hamlets or household clusters apparently persisted throughout the period after c. AD900 when maize macrofossils are found (Custer 1986; Kraft 1986, 2001; Stewart1993). Dispersed villages were present in southern coastal New England by the 14thcentury AD (Leveillee et al. 2006; Waller 2000, 2009), several centuries after maizemacrofossils first became evident in the region (Largy and Morenon 2008).
Wild rice
Wild rice is one of several widely and intensively exploited annual plants that whilelacking morphological changes and therefore not having been ‘‘domesticated’’ in thetraditional sense, it also may have spread across the Northeast through purposefulhuman intervention (Arzigian 2000; Crawford and Smith 2003). To the northgenerally, and particularly as far west as Wisconsin and Minnesota (regions notformally considered here), the introduction of maize had far less impact than themore intensive extraction of wild rice (Arzigian 2000; Moffat and Arzigian 2000).The incorporation of maize into domesticated riverine/wetland domesticatedlandscapes, however, may have been facilitated by wild rice, especially so in themore northern, glaciated areas with poorly evolved drainage systems and abundantwetlands. Maize founder populations were introduced into varying domesticatedlandscapes, some where wild rice may have been an important grain resource.
It is well established that wild rice (primarily Zizania aquatica) was a significantcomponent of indigenous economies in the extreme western Great Lakes region ofWisconsin and Minnesota and northward into Canada (e.g., Arzigian 2000; Johnson1969). Thompson et al. (1994), for example, reported the recovery of wild ricephytoliths from charred cooking residues dated to c. 1800 BP in Minnesota. Wildrice microfossils were recovered along with maize microfossils from charredcooking residues from several sites dating as early as AD 500 in southwesternOntario and southern Manitoba (Boyd and Surette 2010). Hensley (1994) andCrawford and Smith (2003) discuss low incidences of early wild rice in the GreenRiver of Kentucky.
J Archaeol Res (2013) 21:175–216 195
123

In southern Michigan, as in other areas of the Northeast where microfossilanalysis of carbonized food residues has taken place (Hart et al. 2007a; Raviele2010), maize is more often associated with wild rice prior to AD 1000. In theseveral centuries prior to that date, wild rice macrofossils increased in abundance atcertain key locales such as the Schultz site (Lovis et al. 2001). Macrofossil evidencefor wild rice use across eastern Lake Erie and southern Ontario is scant, although ithas been recovered from the Grand Banks site and several post-AD 1100 sites(Crawford and Smith 2003, p. 203). Site locations near wetlands indicate theprobability of wild rice exploitation during the first several centuries after theearliest evidence for maize (Crawford and Smith 2003). Farther to the east, there isidiosyncratic early evidence for wild rice use. A direct date of 2386 ± 48 BP (cal2r = 750–385 BC, median probability = 484 BC) was obtained on a wild rice seedrecovered from the Parslow Field site in the Mohawk River basin of New York(Hart et al. 2003, pp. 634–635). There also are isolated seeds of wild rice from thepost-AD 800 Memorial Park and Catawissa Bridge sites in the Susquehanna Basinof Pennsylvania (Hart and Asch Sidell 1996; King 1992). In New England, 81 wildrice seeds were recovered from the post-AD 1100 Headquarters site in Vermont(Asch Sidell 2008). Wild rice also may have been a component of diets in coastalMassachusetts as well (Leveillee et al. 2006; Waller 2000). There does not appear tobe any direct evidence of wild rice in the Ohio River basin.
A multiphase burial at the Dunn Farm site in Michigan, which was initially cross-dated typologically to between c. 2500 and 3000 BP uncal (Ford and Brose 1975)but subsequently direct dated to 1500 ± 60 BP (cal 2r range = AD 431–637,median probability = AD 558; Brose and Hambacher 1999, p. 176), is among theearliest evidence for wild rice macrofossils in the western Great Lakes. Scatteredevidence for wild rice macrofossils occurs across the Northeast until c. AD 1000,with some abundant remains 500–700 years earlier (Crawford 2011; Lovis et al.2001; Smith and Crawford 1997). Wild rice macrofossils became more commononly after c. AD 1000 (Crawford and Smith 2002, 2003), predating the introductionof the common bean (Phaseolus vulgaris) as a nutritionally complementary foodcrop throughout much of the region (Hart et al. 2002).
Overall, however, wild rice macrofossils are neither common nor ubiquitous,even on post-AD 1000 sites in the Northeast (Crawford and Smith 2003). Someweight, therefore, has been placed on the spatial association of known modern wildrice stands with precontact archaeological sites, suggesting the potential for siteplacement proximal to wild rice patches (Crawford 2011; Lee et al. 2004). Thisphenomenon is more common in southeastern Ontario than elsewhere.
There has been abundant discussion of the specific habitat conditions required forwild rice stands to develop (Arzigian 2000; Jenks 1901; Johnson 1969), withhydrologic (flow rate, depth, clarity, stability), solarization, temperature, and bottomconditions combining to make unique habitats conducive to wild rice growth. It ispossible that interior lakes may be more stable environments for wild rice thaneither lower riverine or Great Lakes coastal zones that are subject to fluctuations ofbase level or annual flood regimes (cf. Lovis et al. 2001). Also of importance is thetiming of wild rice maturation, which allows for earlier harvesting than maturemaize but potentially coincides with its green phase. It was common among the
196 J Archaeol Res (2013) 21:175–216
123

Ojibwa in the western Great Lakes to harvest wild rice in August (Vennum 1988).Vennum further suggested maturation from late August through October. Regard-less, there would be reduced competition for labor during their respective peakproduction and harvest periods.
Pollen has been used extensively to assess the presence of wild rice in variouscontexts (McAndrews 1969, 1984, 1994). More recently, pollen has contributed toan enhanced circumstantial record of wild rice use at Cootes Paradise, Ontario, awetland near a series of logistic procurement sites (Lee et al. 2004). Analysispresented in Lee et al. (2004) indicates dense stands of wild rice after 3000 BP,which may certainly have been exploited by local human populations. As of thiswriting, there are no published microfossil analyses for this region. Among the moresignificant aspects of new microfossil analyses in the Northeast has been theobservation that wild rice and maize often are associated in carbonized cookingresidues (Hart et al. 2003; Raviele 2010). At the Schultz site in Michigan, forexample, residues with associated rice and maize microfossils have been dated to asearly as 170 BC–AD 30 (Raviele 2010).
The nutritional component of wild rice is a potentially significant considerationin assessments of its relationship to maize as a primary food resource. Uncookedwild rice is high in essential amino acids, but some are reduced when rice isprocessed and cooked (US Department of Agriculture 2011). Although proteinscomplementing maize might be derived from other mixed-economy food resourcessuch as meat, fish, or nuts, mixes of maize and rice could provide rather completecomplementary nutrients in the absence of other foodstuffs. By comparison, wildrice loses essential amino acids when cooked, whereas the common bean retains oreven increases in certain nutrients when cooked (US Department of Agriculture2011), which suggests some nutritional advantage to cooking maize with commonbeans rather than wild rice.
Given the late arrival of the common bean in the Northeast (Hart et al. 2002), itslow abundance in the archaeological record, the significant early association ofmaize and wild rice microfossils in several regions, and the substantial essentialamino acid value of wild rice, its incorporation into diets after c. 1 AD may havebeen one initial catalyst for increased maize use in mixed economies. The commonbean may well have supplanted wild rice or other protein-rich enablers because itnot only retains its essential amino acids when cooked but unlike wild rice, it couldbe cropped with maize in the same fields.
Native crops
Throughout much of the Northeast, maize was adopted into domesticatedlandscapes that included cultivation of native seed and grain-bearing annualsconstituting the Eastern Agricultural Complex. At least some of the EAC starchyseed annuals such as goosefoot (Chenopodium berlandieri) and erect knotweed(Polygonum erectum), and oily seed annuals such as sumpweed/marshelder (Ivaannua), cucurbits (gourd and squash: Cucurbita pepo), and sunflower (Helianthusannuus) had undergone selection as a result of cultivation by local populations in the
J Archaeol Res (2013) 21:175–216 197
123

Midwest and Midsouth by c. 4000–3000 BP (e.g., Smith 2006; Smith and Yarnell2009). Some of these plants had been introduced beyond their natural biogeographicranges by at least this time (e.g., Asch Sidell 2008; Cowan 1978).
The more direct issues with which we are confronted include the degree to whichsuch indigenous grains and seeds were preadapted by local regional populations formore intensive incorporation of maize into their economies, provided a platform forthe establishment of processing and storage technologies that could be applied tomaize, potentially provided appropriate nutritional supplements for maize inaddition to essential amino acids derived from other sources, and, finally,understanding the role EAC seeds and grains may or may not have played inpost-maize intensive economies.
The nutritional contributions of various EAC crops, both seeds and grains, havebeen addressed in a variety of contexts (e.g., Asch and Asch 1985). As expected,oily seeds from cucurbits (including Lagenaria gourds), sumpweed/marshelder, andsunflower are nutritionally rich in protein (Asch and Asch 1978; King 1985, 1999)and would have been important early complements to maize. Starchy grains such aslittle barley also are high in nutritional value (Gordon 2007). Little barley may havebeen significant due to seasonal maturation and storage capability in the spring,which made it available as a maize complement even before the early ripening ofgreen corn. Maygrass also has high nutritional value (Asch and Asch 1985; Critesand Terry 1984).
To date, the majority of research on the role of indigenous EAC cultigens hasconcentrated on their earliest inception and evidence for intensive selection; thisfocus has resulted in discernible morphological changes rather than systematicassessment of their role in maize-based economies. Some have argued for completereplacement of EAC plants with the advent of intensive maize agriculture (Cowan1998), although current data do not support such a categorical interpretation (e.g.,Simon 2000). Little barley, for example, occurs in post-maize contexts and inassociation with other indigenous seeds in the Susquehanna River basin (e.g., Hartand Asch Sidell 1996). The early (c. 4200 BP) and continued presence of cucurbitsas far north as the Saginaw drainage of Michigan (Lovis and Monaghan 2008;Monaghan et al. 2006; Simon 2011) and their early occurrence in Maine (Petersenand Asch Sidell 1996) demonstrate their well-known and longstanding importancein indigenous horticulture to the northern limits of potential production. This also istrue of sunflower, with its distribution into southwestern Michigan by c. 2500 BPand its presence in post-AD 1000 Saginaw drainage contexts (Parker 1990, 1996)and ultimately as far north and east as Quebec (Yarnell 1978). Other EAC seeds andgrains are minimally represented across the region of interest although they are stillpresent in post-AD 1000 contexts (e.g., Asch Sidell 2008).
Of particular note in this context is that the EAC seeds and grains, importantacross the riverine Midwest and Midsouth, including the southernmost componentsof the Middle Ohio and upper Mississippi River drainages, were of far lessimportance at the northern margins of maize production. Several significant EACplant species, such as sumpweed/marshelder, did not occur in these areas; if they didoccur, they were present both in lower abundance and with lower individualproductivity.
198 J Archaeol Res (2013) 21:175–216
123

Eastern Agricultural Complex seeds and grains are variably present as precursorsto maize in the regional archaeological records of the Northeast. EAC species insouthern Michigan occurred as early as 4000–3000 BP, including rind and bothcarbonized and uncarbonized seeds of Cucurbita sp. recovered from several sites inthe Saginaw drainage basin (Lovis and Monaghan 2008; Monaghan et al. 2006).They continued to occur sporadically throughout subsequent time periods,ultimately in the form of pumpkin seeds in at least one 18th century ritual context(the Fletcher site; Mainfort 1979, p. 340). Intentional chenopodium harvest can bedemonstrated by c. AD 500 in the Saginaw drainage (Lovis et al. 1996); by c. AD1000 some seeds display metric and morphological evidence of selection (Parker1996). Sunflower was present by c. 500 BC in southwestern Michigan, by c. 1000AD in the Saginaw drainage (Parker 1996), and in 15th century contexts at theSchwerdt site in southwestern Michigan (Walz 1992). The recovery of a singlesumpweed seed from the Saginaw drainage is important because this species is notindigenous to the region and likely was brought in by human transport.
The presence of EAC crops in Ohio cannot be easily separated from adjacentparts of Kentucky and West Virginia, where chenopodium, sumpweed, squash, andsunflower were all broadly present in local economies by c. 1500 BC (e.g.,Gremillion 1993, 1994, 1996b; see also Simon 2000, 2009 for a summary). Yetthere were differential biogeographic distributions of certain EAC species fromnorth to south. By c. AD 600–700 chenopod and maygrass dominated subregionalassemblages, with limited presence of little barley and knotweed but regularoccurrences of cultivated sunflower and squash. Several scholars (e.g., Simon 2000;Wymer 1993) interpret the regional data to reveal an inverse temporal relationshipbetween indigenous crop plants, both starchy and oily, and maize, suggesting theirreplacement by a maize/bean crop complex. Likewise, there is a long history ofexploitation of EAC species in the upper Ohio River region, including chenopods,knotweed, maygrass, and sumpweed, dating to at least the 15th century BC.Morphological indications of cultivation of chenopod seeds are evident by c. AD200. Use of these seed-bearing annuals is apparent on some sites in the region untilat least the 15th century AD (McConaughy 2008).
The earliest evidence for EAC crops in the West Branch of the SusquehannaValley are direct dates on C. pepo gourd and squash from Memorial Park of5404 ± 552 BP and 2820 ± 75 BP, respectively (Hart and Asch Sidell 1997). Arelatively large number (n = 336) of little barley seeds were recovered from thesite, with the highest count coming from features assigned to the AD 874component (n = 119; two cultivated chenopod varieties also were recovered fromthese contexts). Consistent with Memorial Park data, King (1992) reports therecovery of large numbers of little barley seeds from two strata at Catawissa Bridge(n = 52 and 167, respectively) and a few wild rice seeds (n = 2 in both strata). Inthe upper Susquehanna Basin, a suite of seeds from annual plants belonging to EACspecies, but showing no morphological traits associated with extended cultivation,were recovered from 2900–2150 BP contexts at the Broome Tech site (Asch Sidell2008). There are additional sites dating to after AD 800–900 in the Middle andUpper Susquehanna Basin from which up to several hundred seeds of EAC plantswere recovered, some with morphological characteristics indicating cultivated
J Archaeol Res (2013) 21:175–216 199
123

varieties (Asch Sidell 2008). Squash phytoliths were recovered from the Fortin 2site in residue AMS dated to 1525 ± 35 BP (cal 2r = AD 432–605; medianprobability = AD 542; Thompson et al. 2004). Grass (cf. little barley) phytolithswere recovered from charred residue adhering to steatite sherds from the HuntersHome site in New York’s Finger Lakes region that were directly dated as early as3530 ± 30 BP (cal 2r = 1942–1759 BC, median probability = 1845 BC; Hart et al.2008).
New England displays a long record of pre- and post-maize chenopodiumexploitation. None of the material analyzed by Asch Sidell (2008; also see Georgeand Dewar 1999) dating from c. 6400–200 BP, however, has morphologicalcharacteristics suggesting cultivation. George and Dewar (1999) report potentiallycultivated chenopodium from the Burnham–Shepard site in the central ConnecticutValley dating to the 14th century.
Summary and discussion
The adoption of maize by local human populations in the Northeast involvedprocesses that resulted in a mosaic of regional histories. Interactions between humanbehavior, maize genetics, human exploitation of other food resources, andenvironmental variables determined whether founding populations of maizesurvived, as did the spread and long-term success of maize populations. Interactionsbetween human behavior, characteristics of maize kernels and cobs, and subsequenttaphonomic processes determined whether or not maize was preserved in thearchaeological record. These factors, coupled with the methods and techniquesemployed by archaeologists in both recovery and analysis, determine whether maizemacro- and microfossils that are preserved on an archaeological site are recoveredand identified in the lab. Chemical interactions between consumed maize and thehuman body determine whether maize consumption can be identified through SCIAof collagen and apatite, and sample sizes determine how accurately SCIA resultsreflect variation in prehistoric maize consumption. Complex interactions of maizewith C3 resources during water-based cooking determine bulk d13C values ofcharred cooking residues, making an interpretation of any given depleted valueimpossible. Establishing the history of maize in a particular region, then, must relyon more than the simple correlation of single lines of evidence of human use. Theinterplay of multiple lines of evidence permits archaeologists to create robusthistories and understand the dynamic interactions between settlement systems andmaize adoption and intensification.
On a gross regional scale, the history of maize in the Northeast is nowsubstantially different from what was known a few decades ago. The crop waspresent by at least 300 BC—ample time for the local evolution of Northern Flint.Maize founder populations were introduced into varying domesticated landscapes,some with existing agroecologies consisting of one to several EAC plants, otherswhere seed-bearing plants were exploited but apparently not cultivated, and otherswhere wild rice may have been an important grain resource. These latter tended tobe in formerly glaciated subregions with relatively poorly evolved drainage systems.
200 J Archaeol Res (2013) 21:175–216
123

Throughout the Northeast, maize did not become a focal resource until manycenturies later. During the intervening centuries, local human populations remainedat least seasonally mobile. It was not until after maize became a focal resource thatnucleated villages became common. As recent SCIA results in the western LakeErie basin demonstrate, even after maize became a significant component of localhuman diets, some populations remained at least seasonally mobile. As demon-strated by the SCIA record in the Middle Ohio River subregion, even after maizebecame a focal resource in particular subregions, it did not become one for all localhuman populations. For some groups, it was not a significant dietary component forall individuals within the population. In at least some areas of New England, maizemay not have been a focal resource until European incursions. Across the entireNortheast, the earliest records of maize occur in contexts of population mobility.
Available evidence for the history of maize across subregions of the Northeast isvery uneven. Given the complexities of the processes of maize adoption,perpetuation, and intensification and the interactions of these processes with theformation of the archaeobotanical and isotope records, multiple lines of evidenceare needed to build subregional histories for the crop. For example, while there isnow a fairly complete record of maize use in the Finger Lakes–Mohawk subregionbased on microfossils extracted from cooking residues, the absence of complemen-tary macrofossil and human bone SCIA records prevents the construction of a morecomplete history of the crop. Similarly, there is a relatively poor record of thehistories of EAC crops in this region. Although we know that little barley (oranother grass) and squash were in economic use early based on phytoliths, there is apoor to nonexistent macrobotanical record. By contrast, the directly datedmacrofossil record in the Grand River valley of southern Ontario is exemplaryfor both maize and other plants, and it is complemented by a small but fairlycomplete chronological SCIA record of human bone collagen and apatite from thebroader subregion. The absence of a microfossil record, however, precludes theestablishment of a complete history for maize in the region—it is not possible todetermine how long the crop was present before the macrofossil record began.
New England, on the other hand, has only a late macrofossil record, lackscomplementary datasets from microfossils, and has only a limited record of SCIAon human bone. It remains unclear, therefore, what portion of the history of maize isrepresented by the extant database. The same issues are evident in a number of othersubregions, that is, there is good evidence for when maize became an importantresource, but little or no data exist for the centuries before then. In the Middle OhioRiver basin, for example, there is an early directly dated macrofossil but none afterthat until maize became ubiquitous in the archaeological record. The human boneSCIA record is large and chronologically continuous but limited to collagen. Inaddition, the pre-AD 900 sample is much smaller than the post-AD 900 sample andit is primarily from burial mound contexts. As a result, the seemingly abruptincrease in maize consumption evident in the record may be an artifact of sampling,reflecting only the protein portion of the diet and perhaps only elite members oflocal populations. As Ambrose et al. (2003) demonstrated for Mound 72 at Cahokia,elite members of populations could have different maize consumption levels thannonelite members. In the lower Upper Ohio River basin, other than the probable
J Archaeol Res (2013) 21:175–216 201
123

early evidence for maize at Meadowcroft Rockshelter, there is a lack of anysubstantiated evidence in the form of microfossils, directly dated macrofossils, orSCIA of human bone upon which to build subsequent histories of maize untilaround AD 900.
The existing record does indicate, however, that the extensive and often intensiveagricultural systems that characterize many late precontact subsistence patterns inthe Northeast were the result of long evolutionary processes. Available evidencedoes suggest that these processes are in line with the expectations of SBT. Maizewas adopted by dispersed, mobile human populations. Some of these populationswere already exploiting native grasses such as wild rice and little barley. Theresulting maize metapopulations and their respective demes were scattered acrossthe physical and fitness landscapes, exposed to varying selection regimes. As inphase II of SBT, individual demes/metapopulations would have ascended highfitness peaks that resulted in increased productivity and, therefore, preservation ofmacrofossils in the archaeological record at open-air sites such as Grand Banks insouthern Ontario. Productive landraces of maize spread through seed flow as inphase III of SBT, eventually resulting in the mosaic of Northern Flint landracesevident in the historical record. Throughout this process the fitness of maize andother species in the domesticated landscapes across the Northeast changed,increasing and decreasing their exploitation by local human populations. Whethernative grasses continued to be exploited depended on local conditions and themarginal costs of each species in the domesticated landscape, all of which created acontinually shifting mosaic of maize use.
In sum, then, although we have a much clearer understanding of the gross historyof maize in the Northeast, the records for subregional histories are incomplete. Thisincompleteness prevents comprehensive understanding of the range of variation inthe histories of maize in the Northeast. Based on current knowledge of the processesof maize adoption, perpetuation, and intensification, the construction of suchhistories is critical to a complete understanding of why and how maize eventuallybecame the dominant grain crop across much of the Northeast.
Future research directions
Although future research directions are not that different from those we haveemployed and summarized, it is evident to us that there should be a shift inemphasis. For example, rather than focusing on the adoption of maize as a primaryor dominant agricultural crop in precontact economies across the Northeast, anoperating assumption of much ongoing research, we believe the question shouldrelate more directly to the degree and rate at which maize was incorporated intomixed economies. Such a shift in focus would include a better understanding of theamount of variation and the varying temporal role of indigenous seeds and grains,both terrestrial and aquatic, in these economies. How do the subregional histories ofpre- and early maize adoption as they relate to behavior and technology act to fostermore rapid or relatively more intensive incorporation of maize into local
202 J Archaeol Res (2013) 21:175–216
123

economies? Did maize factor into more variable and flexible nutritional decisionsthan we might currently believe?
Clearly linked to these altered directions is the central issue of the developmentof the Northern Flint landraces that dominates the post-AD 1000 florescence ofmaize agriculture. Micro- and macrobotanical data from multiple subregions nowestablish that maize was introduced more than 2000 years ago—maybe even 2500years ago—well before the earliest evidence for Northern Flint. What, then, are theselective trajectories that led to the evolution of Northern Flint and its landracesover a millennium of low-level production? Certainly, a millennium of local-levelselection is sufficient to allow for a hypothesis of its in situ indigenous developmentin the Northeast. Recent developments in ancient DNA extraction from charredgrains (Bunning et al. 2012) may provide opportunities to enhance our understand-ings of maize evolution in the Northeast.
There is also an overarching methodological component to our synoptic endgame. The cumulative available data on the regional ages for inception, the contextsfor introduction, trajectories of adoption, and impact of maize certainly allow forinterpretations that suggest the presence of regional variation across all threedimensions. The lingering question is, of course, the degree of inferential confidencewith which such outcomes are presented. Certainly we can point to regional recordsthat vary in the ages for first presence of maize. There are C and N isotope data onhuman bone that reveal regional differences in the degree to which maize wasconsumed in the diet, both collectively and individually. And there are regionaldifferences in temporal trajectories for bulk d13C measurements on carbonizedresidues adhering to cooking containers. All of these observations, we should note,are consistent with the expectations of SBT.
That said, the methodological advances of the last several decades—approacheseither independent of and/or complementary to the primary use of carbonizedpreserved macrobotanical remains—have been variably applied (and even variablywell applied) in different regions. There is variation in both time duration and thedegree to which practitioners in some regions have used flotation recovery, resultingin better macrofossil records. Some regions have no direct AMS-dated maizemacrofossils, continuing a dubious reliance on contextually associated datablematerials. Most have no analyses incorporating microfossils recovered from directlydated cooking residues or other contexts. In certain contexts and for various reasons,human bone is no longer readily accessible for isotopic assay, preventingassessment of correspondence between resource records and diet. Finally, there istaphonomy, as related to individual plants, including maize, and, more generally,the level of regional preservation potential. All these disparities should be evident inour attenuated regional summary.
As the Michigan and central New York data clearly indicate, the application ofsplit-sample analyses in a systematic and intensive fashion at the subregional levelcan markedly alter regional histories of maize (and other plants). This reflects a flawthat lies at the crux of current macroregional interpretation and that, to our mind,should form a focus for concerted future research: the identification of how localarchaeological records of maize and related plants can benefit from strategicallydirected research programs that reflect judicious inquiry into isotopic analysis of
J Archaeol Res (2013) 21:175–216 203
123

human bone and carbonized food residues, partially destructive direct dating ofcurated maize macrofossils, and the systematic identification of microfossils,including additional insights into the specifics of their taphonomy under differentconditions. As an aside, it is our experience, borne out by the contemporaryliterature, that microfossil analysis has not only come of age but is an indispensablecomplement to other lines of inquiry; any lingering suspicions of its accuracy and aconsequent reticence to apply it should finally be summarily dispensed with.
While we recognize that there will continue to be advances in analytic methodsand that the differential application of new analytic approaches on sample sets withvarying access constraints will always yield data unevenness, it is currently essentialthat the evident disparities at least be reduced in magnitude and made moreregularly compatible for comparative purposes. Only when such concordance isbetter achieved will the inferential strength of interpretations of the history of maizehave scientific credibility.
Acknowledgments We thank Gary Feinman and the seven peer reviewers for their constructivecomments on the original version of this article.
References cited
Abrams, M. D., and Nowacki, G. J. (2008). Native Americans as active and passive promoters of mastand fruit trees in the eastern USA. The Holocene 18: 1123–1137.
Adovasio, J. M., and Johnson, W. C. (1981). The appearance of cultigens in the upper Ohio Valley: Aview from Meadowcroft Rockshelter. Pennsylvania Archaeologist 51(1–2): 63–80.
Allegretto, K. O. (2007). Adopting Maize in the Eastern Woodlands of North America, Ph.D. dissertation,Department of Anthropology, Brandeis University, Waltham, MA.
Ambrose, S. H. (1993). Isotopic analysis of paleodiets: Methodological and interpretive considerations. InSandford, M. K. (ed.), Investigations of Ancient Human Tissue: Chemical Analyses in Anthropology,Gordon and Breach, Langhorne, PA, pp. 59–130.
Ambrose, S. H., and Norr, L. (1993). Experimental evidence of the carbon isotope ratios of whole diet anddietary protein of bone collagen and carbonate. In Lambert, J. B., and Grupe, G. (eds.), PrehistoricHuman Bone: Archaeology at the Molecular Level, Springer, Berlin, pp. 1–37.
Ambrose, S. H., Buikstra, J., and Krueger, H. W. (2003). Status and gender differences in diet at Mound72, Cahokia, revealed by isotopic analysis of bone. Journal of Anthropological Archaeology 22:217–226.
Arzigian, C. (2000). Middle Woodland and Oneota contexts for wild rice exploitation in southwesternWisconsin. Midcontinental Journal of Archaeology 25: 245–268.
Asch, D. L., and Asch, N. B. (1978). The economic potential of Iva annua and its prehistoric importancein the lower Illinois Valley. In Ford, R. I. (ed.), The Nature and Status of Ethnobotany,Anthropological Papers No. 67, Museum of Anthropology, University of Michigan, Ann Arbor,pp. 301–341.
Asch, D. L., and Asch, N. B. (1985). Prehistoric plant cultivation in west-central Illinois. In Ford, R. I.(ed.), Prehistoric Food Production in North America, Anthropological Papers No. 75, Museum ofAnthropology, University of Michigan, Ann Arbor, pp. 149–203.
Asch Sidell, N. (2002). Paleoethnobotanical indicators of subsistence and settlement change in theNortheast. In Hart, J. P., and Rieth, C. B. (eds.), Northeast Subsistence-Settlement Change: AD700–1300, New York State Museum Bulletin 496, University of the State of New York, Albany,pp. 241–264.
Asch Sidell, N. (2008). The impact of maize-based agriculture on prehistoric plant communities in thenortheast. In Hart, J. P. (ed.), Current Northeast Paleoethnobotany II, New York State MuseumBulletin 512, University of the State of New York, Albany, pp. 29–51.
204 J Archaeol Res (2013) 21:175–216
123

Azar, C., Mather, D. E., and Hamilton, R. I. (1997). Maize landraces of the St. Lawrence-Great Lakesregion of North America. Euphytica 98: 141–148.
Badstrue, L., Bellon, M., Berthaud, J., Ramırez, A., Flores, D., and Juarez, X. (2007). The dynamics offarmers’ maize seed supply practices in the Central Valleys of Oaxaca, Mexico. World Development35: 1579–1593.
Beld, S. G. (1993). Site 20IA37 (Arthurburg Hill Earthworks), Lyons Township, Ionia County, Michigan.In Beld, S. G. (ed.), Lyons Township Archaeological Survey, S-92-313, Archaeological SurveyCompletion Report submitted to the Michigan Office of the State Archaeologist, Alma College,Alma, MI, pp. 3–82.
Bendremer, J. C. M., Kellogg, E. A., and Largy, T. B (1991). A grass-lined maize storage pit and earlymaize horticulture in central Connecticut. North American Archaeologist 12: 325–349.
Birch, J. (2010). Coalescent Communities in Iroquoian Ontario, Ph.D. dissertation, Department ofAnthropology, McMaster University, Hamilton, Ontario.
Black, B. A., Ruffner, C. M., and Abrams, M. D. (2006). Native American influences on the forestcomposition of the Allegheny Plateau, northwest Pennsylvania. Canadian Journal of ForestResources 36: 1266–1275.
Bourque, B. J., and Krueger, H. W. (1994). Dietary reconstruction from human bone isotopes for fivecoastal New England populations. In Sobolik, K. D. (ed.), Paleonutrition: The Diet and Health ofPrehistoric Americans, Occasional Paper No. 22, Center for Archaeological Investigations,Southern Illinois University, Carbondale, pp. 195–209.
Boyd, M., and Surette, C. (2010). Northernmost precontact maize in North America. American Antiquity75: 117–133.
Bozarth, S. R. (1993). Maize (Zea mays) cob phytoliths from a central Kansas Great Bend aspectarchaeological site. Plains Anthropologist 38: 279–286.
Brandt, K. L. (1996). The Effects of Early Maize Agriculture on Native North American Populations:Evidence from the Teeth and Skeleton, Ph.D. dissertation, Department of Anthropology, Universityof Michigan, Ann Arbor.
Bridges, P. S. (1994). Prehistoric diet and health in a coastal New York skeletal sample. NortheastAnthropology 8: 13–23.
Brose, D. S., and Hambacher, M. J. (1999). The Middle Woodland in northern Michigan. In Halsey, J. R.(ed.), Retrieving Michigan’s Buried Past: The Prehistoric and Historic Archaeology of the GreatLakes State, Cranbrook Institute of Science, Bloomfield Hills, MI, pp. 173–192.
Brown, W. L. (1947). The Northern Flint corns. Annals of the Missouri Botanical Garden 34: 1–20.Bunning, S. L., Jones, G., and Brown, T. A. (2012). Next generation sequencing of DNA in 3300-year-old
charred cereal grains. Journal of Archaeological Science 39: 2780–2784.Camus-Kulandaivelu, L., Veyrieras, J.-B., Madur, D., Combes, V., Fourmann, M., Barraud, S., Dubreuil,
P., Gouesnard, B., Manicacci, D., and Charcosset, A. (2006). Maize adaptation to temperate climate:Relationship between population structure and polymorphism in the Dwarf8 gene. Genetics 172:2449–2463.
Cassedy, D., and Webb, P. (1999). New data on the chronology of maize horticulture in eastern New Yorkand southern New England. In Hart, J. P. (ed.), Current Northeast Paleoethnobotany, New YorkState Museum Bulletin 494, University of the State of New York, Albany, pp. 85–99.
Ceci, L. (1990). Radiocarbon dating ‘‘village’’ sites in coastal New York: Settlement pattern change in theMiddle to Late Woodland. Man in the Northeast 39: 1–28.
Chavez, S. J., and Thompson, R. G. (2006). Early maize on the Copacabana Peninsula: Implications forthe archaeology of the Lake Titicaca Basin. In Staller, J. E., Tykot, R. H., and Benz, B. F. (eds.),Histories of Maize: Multidisciplinary Approaches to the Prehistory, Linguistics, Biogeography,Domestication, and Evolution of Maize, Academic Press, Burlington, MA, pp. 415–428.
Chilton. E. A. (1999). Mobile farmers of pre-contact southern New England: The archaeological andethnohistoric evidence. In Hart, J. P. (ed.), Current Northeast Paleoethnobotany, New York StateMuseum Bulletin 494, University of the State of New York, Albany, pp. 157–176.
Chilton, E. A. (2006). The origin and spread of maize (Zea mays) in New England. In Staller, J. E., Tykot,R. H., and Benz, B. F. (eds.), Histories of Maize: Multidisciplinary Approaches to the Prehistory,Linguistics, Biogeography, Domestication, and Evolution of Maize, Academic Press, Burlington,MA, pp. 539–548.
Chilton, E. A. (2008). So little maize, so much time: Understanding maize adoption in New England. InHart, J. P. (ed.), Current Northeast Paleoethnobotany II, New York State Museum Bulletin 512,University of the State of New York, Albany, pp. 53–59.
J Archaeol Res (2013) 21:175–216 205
123

Conard, N., Asch, D. L., Asch, N. B., Elmore, D., Grove, H., Rubin, M., Brown, J. A., Wiant, M. D.,Farnsworth, K. B., and Cook, T. G. (1984). Accelerator radiocarbon dating of evidence forprehistoric horticulture in Illinois. Nature 308: 443–447.
Cook, R. A., and Schurr, M. R. (2009). Eating between the lines: Mississippian migration and stablecarbon isotope variation in Fort Ancient populations. American Anthropologist 111: 344–359.
Cortella, A. R., and Pochettino, M. L. (1994). Starch grain analysis as a microscopic diagnostic feature inthe identification of plant material. Economic Botany 48: 171–181.
Cowan, C. W. (1978). The prehistoric use and distribution of maygrass in eastern North America:Cultural and phytogeographical implications. In Ford, R. I. (ed.), The Nature and Status ofEthnobotany, Anthropological Papers No. 67, Museum of Anthropology, University of Michigan,Ann Arbor, pp. 263–288.
Cowan, C. W. (1998). Eastern Agricultural Complex. In Gibbon, G. E. (ed.), Archaeology of PrehistoricNative America: An Encyclopedia, Routledge, New York, pp. 237–239.
Crawford, G. W. (2011). People and plant interactions in the northeast. In Smith, B. D. (ed.), TheSubsistence Economies of Indigenous North American Societies: A Handbook, SmithsonianInstitution Scholarly Press, Washington, DC, pp. 431–448.
Crawford, G. W., and Smith, D. G. (2002). Early Late Woodland in southern Ontario: An update(1996–2000). In Hart, J. P. (ed.), Northeast Subsistence-Settlement Change: AD 700–1300. NewYork State Museum Bulletin 496, University of the State of New York, Albany, pp. 117–134.
Crawford, G. W., and Smith, D. G. (2003). Paleoethnobotany in the Northeast. In Minnis, P. E. (ed.),People and Plants in Ancient North America, Smithsonian Books, Washington, DC, pp. 172–257.
Crawford, G. W., Smith, D. G., and Bowyer, V. E. (1997a). Dating the entry of corn (Zea mays) into thelower Great Lakes. American Antiquity 62: 112–119.
Crawford, G. W., Smith, D. G., Desloges, J. R., and Davis, A. M. (1997b). Floodplains and agriculturalorigins: A case study in south-central Ontario, Canada. Journal of Field Archaeology 25: 123–137.
Crawford, G. W., Saunders, D., and Smith, D. G. (2006). Pre-contact maize from Ontario, Canada:Context, chronology, and plant association. In Staller, J. E., Tykot, R. H., and Benz, B. F. (eds.),Histories of Maize: Multidisciplinary Approaches to the Prehistory, Linguistics, Biogeography,Domestication, and Evolution of Maize, Academic Press, Burlington, MA, pp. 549–563.
Crites, G. D., and Terry, R. D. (1984). Nutritive value of maygrass. Economic Botany 38: 114–120.Crowther, A. (2012). The differential survival of native starch during cooking and implications for
archaeological analyses: A review. Archaeological and Anthropological Sciences 4: 221–235.Custer, J. F. (1986). Late Woodland cultural diversity in the Middle Atlantic: An evolutionary
perspective. In Custer, J. F. (ed.), Late Woodland Cultures of the Middle Atlantic Region, Universityof Delaware Press, Newark, pp. 143–168.
Day, G. M. (1978). Western Abinaki. In Trigger, B. G. (ed.), Handbook of North American Indians,Volume 15: Northeast, Smithsonian Institution Press, Washington, DC, pp. 148–159.
Dewar, G., Ginter, J. K., Shook, B. A., Ferris, N., and Henderson, H. (2010). A bioarchaeological study ofa Western Basin tradition cemetery on the Detroit River. Journal of Archaeological Science 37:2245–2254.
Doebley, J. F., Goodman, M. M., and Stubb, C. W. (1986). Exceptional genetic divergence of NorthernFlint corn. American Journal of Botany 73: 64–69.
Dyer, G. A., and Taylor, J. E. (2008). A crop population perspective on maize seed systems in Mexico.Proceedings of the National Academy of Sciences 105: 470–475.
Environmental Protection Agency. (2008). The Great Lakes Atlas, electronic document, http://www.epa.gov/greatlakes/atlas/glat-ch2.html, accessed January 11, 2012.
Eswaran, V. (2002). A diffusion wave out of Africa: The mechanism of the modern human revolution?Current Anthropology 43: 749–774.
Farrow, D. C. (1986). A study of Monongahela subsistence patterns based on mass spectrometric analysis.Midcontinental Journal of Archaeology 11: 153–180.
Ford, R. I. (1985). Patterns of prehistoric food production in North America. In Ford, R. I. (ed.),Prehistoric Food Production in North America, Anthropological Papers No. 75, Museum ofAnthropology, University of Michigan, Ann Arbor, pp. 341–366.
Ford, R. I. (1987). Dating early maize in the eastern United States. Paper presented at the 10thEthnobotany Meetings, Gainesville, FL.
Ford, R. I., and Brose, D. S. (1975). Prehistoric wild rice from the Dunn Farm site, Leelanau County,Michigan. The Wisconsin Archeologist 56: 9–15.
206 J Archaeol Res (2013) 21:175–216
123

Fritz, G. J. (1990). Multiple pathways to farming in precontact eastern North America. Journal of WorldPrehistory 4: 387–435.
Fritz, G. J. (1993). Early and Middle Woodland period ethnobotany. In Scarry C. M. (ed.), Foraging andFarming in the Eastern Woodlands, University Press of Florida, Gainesville, pp. 39–56.
George, D. R., and Dewar, R. E. (1999). Chenopodium in Connecticut prehistory: Wild, weedy,cultivated, or domesticated? In Hart, J. P. (ed.), Current Northeast Paleoethnobotany, New YorkState Museum Bulletin 494, University of the State of New York, Albany, pp. 121–132.
Gillespie, J. H. (1998). Population Genetics: A Concise Guide, Johns Hopkins University Press,Baltimore, MD.
Goette, S., Williams, M., Johannessen, S., and Hastorf, C. A. (1994). Toward reconstructing ancientmaize: Experiments in processing and charring. Journal of Ethnobiology 14: 1–21.
Goldberg, P., and Arpin, T. L. (1999). Micromorphological analysis of sediments from MeadowcroftRockshelter, Pennsylvania: Implications for radiocarbon dating. Journal of Field Archaeology 26:325–342.
Gordon, A. A. (2007). Hordeum pusillum Nutt., little barley, graminae. In Fritz, G. J. (ed.), LaboratoryGuide to Archaeological Plant Remains from Eastern North America, electronic document,http://artsci.wustl.edu/*gjfritz/index.html, accessed January 2, 2012.
Greenlee, D. M. (1990). Environmental and Temporal Variability in d13C Values in Late PrehistoricSubsistence Systems in the Upper Ohio Valley, Master’s thesis, Department of Anthropology,University of Washington, Seattle.
Greenlee, D. M. (2002). Accounting for Subsistence Variation Among Maize Farmers in Ohio ValleyPrehistory, Ph.D. dissertation, Department of Anthropology, University of Washington, Seattle.
Greenlee, D. M. (2006). Dietary variation and prehistoric maize farming in the middle Ohio Valley. InStaller, J. E., Tykot, R. H., and Benz, B. F. (eds.), Histories of Maize: Multidisciplinary Approachesto the Prehistory, Linguistics, Biogeography, Domestication, and Evolution of Maize, AcademicPress, Burlington, MA, pp. 215–237.
Gremillion, K. J. (1993). Plant husbandry at the Archaic/Woodland transition: Evidence from the ColdOak shelter, Kentucky. Midcontinental Journal of Archaeology 18: 161–189.
Gremillion, K. J. (1994). Evidence of plant domestication from Kentucky caves and rockshelters. InGreen, W. (ed.), Agricultural Origins and Development in the Midcontinent, Report No. 19, Officeof the State Archaeologist, University of Iowa, Iowa City, pp. 87–103.
Gremillion, K. J. (1996a). Diffusion and adoption of crops in evolutionary perspective. Journal ofAnthropological Archaeology 15: 183–204.
Gremillion, K. J. (1996b). Early agricultural diet in eastern North America: Evidence from two Kentuckyrockshelters. American Antiquity 61: 520–536.
Grobman, A., Bonavia, D., Dillehay, T. D., Piperno, D. R., Iriarte, J., and Holst, I. (2012). Preceramicmaize from Paredones and Huaca Prieta, Peru. Proceedings of the National Academy of Sciences109: 1755–1759.
Hall, R. L. (1980). An interpretation of the two-climax model of Illinois prehistory. In Browman, D. L.(ed.), Early Native America, Mouton, The Hague, pp. 401–462.
Hallauer, A. R., and Miranda, J. B. (1988). Quantitative Genetics in Maize Breeding, 2nd ed., Iowa StateUniversity Press, Ames.
Harrison, R. G., and Katzenberg, M. A. (2003). Paleodiet studies using stable carbon isotopes from boneapatite and collagen: Examples from southern Ontario and San Nicolas Island, California. Journal ofArchaeological Science 22: 227–244.
Hart, J. P. (1999a). Maize agriculture evolution in the eastern woodlands of North America: A Darwinianperspective. Journal of Archaeological Method and Theory 6: 137–180.
Hart, J. P. (ed.). (1999b). Current Northeast Paleoethnobotany, New York State Museum Bulletin 494,University of the State of New York, Albany.
Hart, J. P. (2001). Maize, matrilocality, migration and northern Iroquoian evolution. Journal ofArchaeological Method and Theory 8: 151–182.
Hart, J. P. (2008a). Evolving the three sisters: The changing histories of maize, bean, and squash in NewYork and the greater Northeast. In Hart, J. P. (ed.), Current Northeast Paleoethnobotany II, NewYork State Museum Bulletin 512, University of the State of New York, Albany, pp. 87–99.
Hart, J. P. (ed.) (2008b). Current Northeast Paleoethnobotany II, New York State Museum Bulletin 512,University of the State of New York, Albany.
Hart, J. P. (2012). Pottery wall thinning as a consequence of increased maize processing: A case studyfrom central New York. Journal of Archaeological Science 39: 3470–3474.
J Archaeol Res (2013) 21:175–216 207
123

Hart, J. P., and Asch Sidell, N. (1996). Prehistoric agricultural systems in the West Branch of theSusquehanna River basin, AD 800 to AD 1350. Northeast Anthropology 52: 1–30.
Hart, J. P., and Asch Sidell, N. (1997). Additional evidence for early cucurbit use in the northern EasternWoodlands east of the Allegheny Front. American Antiquity 62: 523–537.
Hart, J. P., and Lovis, W. A. (2007). A multi-regional analysis of AMS and radiometric dates fromcarbonized food residues. Midcontinental Journal of Archaeology 32: 201–261.
Hart, J. P., and Matson, R. G. (2009). The use of multiple discriminant analysis in classifying prehistoricphytolith assemblages recovered from cooking residues. Journal of Archaeological Science 36:74–83.
Hart, J. P., and Means, B. K. (2002). Maize and villages: A summary and critical assessment of currentNortheast early late prehistoric evidence. In Hart, J. P., and Rieth, C. B. (eds.), NortheastSubsistence-Settlement Change: AD 700–1300, New York State Museum Bulletin 496, University ofthe State of New York, Albany, pp. 342–358.
Hart, J. P., and Rieth, C. B. (eds.) (2002). Northeast Subsistence-Settlement Change: AD 700–1300, NewYork State Museum Bulletin 496, University of the State of New York, Albany.
Hart, J. P., Asch, D. L., Scarry, C. M., and Crawford, G. W. (2002). The age of the common bean(Phaseolus vulgaris L.) in the northern Eastern Woodlands of North America. Antiquity 76:377–385.
Hart, J. P., Thompson, R. G., and Brumbach, H. J. (2003). Phytolith evidence for early maize (Zea mays)in the northern Finger Lakes region of New York. American Antiquity 68: 619–640.
Hart, J. P., Nass, J. P., and Means, B. K. (2005). Monongahela subsistence-settlement change?Midcontinental Journal of Archaeology 30: 327–365.
Hart, J. P., Brumbach, H. J., and Lusteck. R. (2007a). Extending the phytolith evidence for early maize(Zea mays ssp. mays) and squash (Cucurbita sp.) in central New York. American Antiquity 72:563–583.
Hart, J. P., Lovis, W. A., Schulenberg, J. K., and Urquhart, G. R. (2007b). Paleodietary implications fromstable carbon isotope analysis of experimental cooking residues. Journal of Archaeological Science34: 804–813.
Hart, J. P., Reber, E. A., Thompson, R. G., and Lusteck, R. (2008). Taking variation seriously: Evidencefor steatite vessel use from the Hunter’s Home site, New York. American Antiquity 76: 729–741.
Hart, J. P., Urquhart, G. R., Feranec, R. S., and Lovis, W. A. (2009). Nonlinear relationship between bulkd13C and percent maize in carbonized cooking residues and the potential of false negatives indetecting maize. Journal of Archaeological Science 36: 2206–2212.
Hart, J. P., Anderson, L. M., and Feranec, R. S. (2011a). Additional evidence for cal seventh-century ADmaize consumption at the Kipp Island site, New York. In Rieth, C. B., and Hart, J. P. (eds.), CurrentResearch in New York State Archaeology: AD 700–1300, New York State Museum Record 2,University of the State of New York, Albany, pp. 27–40.
Hart, J. P., Matson, R. G., Thompson, R. G., and Blake, M. (2011b). Teosinte inflorescence phytolithassemblages mirror Zea taxonomy. PLoS ONE 6(3): e18349.
Hart, J. P., Lovis, W. A. Jeske, R. J., and Roberts, J. D. (2012). Stable carbon isotope values on encrustedcooking residues and chronology: Assessing regional maize use histories. American Antiquity 77:315–325.
Hastorf, C., and DeNiro, M. J. (1985) Reconstruction of prehistoric plant production and cookingpractices by a new isotopic method. Nature 315: 489–491.
Hatch, J. W. (ed.). (1980). The Fisher Farm Site: A Late Woodland Hamlet in Context, Occasional PapersNo. 12, Department of Anthropology, Pennsylvania State University, University Park.
Hensley, C. K. (1994). The Archaic Settlement System of the Middle Green River Valley, Kentucky, Ph.D.dissertation, Department of Anthropology, Washington University, St. Louis, MO.
Hyde, G. E. (1962). Indians of the Woodlands from Prehistoric Times to 1725, University of OklahomaPress, Norman.
Jenks, A. E. (1901). The wild rice gatherers of the upper lakes. Annual Report of the Bureau of Ethnology19: 1013–1137.
Johannessen, C. L., Wilson, M. R., and Davenport, W. A. (1970). Domestication of maize: Process orevent? The Geographical Review 60: 398–413.
Johnson, E. (1969). Archeological evidence for utilizaton of wild rice. Science 163: 276–277.Jugenheimer, R. W. (1976). Corn Improvement, Seed Production, and Uses, John Wiley & Sons, New
York.
208 J Archaeol Res (2013) 21:175–216
123

Katzenberg, M. A. (2006). Prehistoric maize in southern Ontario: Contributions from stable carbonisotope studies. In Staller, J. E., Tykot, R. H., and Benz, B. F. (eds.), Histories of Maize:Multidisciplinary Approaches to the Prehistory, Linguistics, Biogeography, Domestication, andEvolution of Maize, Academic Press, Burlington, MA, pp. 263–273.
Katzenberg, M. A., Schwarcz, H. P., Knyf, M., and Melbye, F. J. (1995). Stable isotope evidence formaize horticulture and paleodiet in southern Ontario, Canada. American Antiquity 60: 335–350.
Kays, R. W., Jansen, P. A., Knecht, E. M., Vohwinkel, R., and Wikelski, M. (2011). The effect of feedingtime on dispersal of virola seeds by toucans determined from GPS tracking and accelerometers. ActaOecologica 37: 625–631.
King, F. B. (1985). Early cultivated cucurbits in eastern North America. In Ford, R. I. (ed.), PrehistoricFood Production in North America, Anthropological Papers No. 75, Museum of Anthropology,University of Michigan, Ann Arbor, pp. 73–98.
King, F. B. (1987). Prehistoric Maize in Eastern North America: An Evolutionary Evaluation, Ph.D.dissertation, Department of Anthropology, University of Illinois, Urbana-Champaign.
King, F. B. (1992). Floral remains. In East, T. C., Adovasio, J. A., Johnson, W. C., and Pedler, D. R.(eds.), The Prehistory of the Catawissa Bridge Replacement Site (36CO9), Columbia County,Pennsylvania, Cultural Resource Management Program, Department of Anthropology, University ofPittsburgh, Pittsburgh, PA, pp. 11.1–11.8.
King, F. B. (1999). Changing evidence for prehistoric plant use in Pennsylvania. In Hart, J. P. (ed.),Current Northeast Paleoethnobiology, New York State Museum Bulletin 494, University of theState of New York, Albany, pp. 11–26.
Kinietz, W. V. (1940). The Indians of the Western Great Lakes, University of Michigan Press, Ann Arbor.Knapp, T. D. (2008). An unbounded future? Ceramic types, ‘‘cultures,’’ and scale in late prehistoric
research. In Miroff, L. E., and Knapp, T. D. (eds.), Iroquoian Archaeology and Analytical Scale,University of Tennessee Press, Knoxville, pp. 101–130.
Kraft, H. C. (1986). Late Woodland settlement patterns in the Upper Delaware Valley. In Custer, J. F.(ed.), Late Woodland Cultures of the Middle Atlantic Region, Associated University Presses,Cranbury, NJ, pp. 102–115.
Kraft, H. C. (2001). The Lenape-Delaware Indian Heritage 10,000 BC to AD 2000, Lenape Books,Union, NJ.
Largy, T., and Morenon, E. P. (2008). Maize agriculture in coastal Rhode Island: Imaginative, illusive, orinvasive? In Hart, J. P. (ed.), Current Northeast Paleoethnobotany II, New York State MuseumBulletin 512, University of the State of New York, Albany, pp. 73–86.
Lee, G.-A., Davis, A. M., Smith, D. G., and McAndrews, J. H. (2004). Identifying fossil wild rice(Zizania) pollen from Cootes Paradise, Ontario: A new approach using scanning electronmicroscopy. Journal of Archaeological Science 31: 411–421.
Leveillee, A., Waller Jr., J., and Ingham, D. (2006). Dispersed villages in Late Woodland period south-coastal Rhode Island. Archaeology of Eastern North America 34: 71–89.
Little, E. A. (2002). Kautantouwit’s legacy: Calibrated dates on prehistoric maize in New England.American Antiquity 67: 109–119.
Little, E. A., and Schoeninger, M. J. (1995). The Late Woodland diet on Nantucket Island and theproblem of maize in coastal New England. American Antiquity 60: 351–368.
Lopinot, N. H. (1992). Spatial and temporal variability in Mississippian subsistence: The archaeologicalrecord. In Woods, W. I. (ed.), Late Prehistoric Agriculture: Observations from the Midwest, Studiesin Illinois Archaeology No. 8, Illinois Historic Preservation Agency, Springfield, pp. 44–94.
Louette, D., Charrier, A., and Berthaud, J. (1997). In situ conservation of maize in Mexico: Geneticdiversity and maize seed management in a traditional community. Economic Botany 51: 20–38.
Lovis, W. A. (1990). Curatorial considerations for systematic research collections: AMS dating a curatedceramic assemblage. American Antiquity 55: 382–387.
Lovis, W. A., and Monaghan, G. W. (2008). Chronology and evolution of the Green Point flood plain andassociated Cucurbita pepo. In Hart, J. P. (ed.), Current Northeast Paleoethnobotany II, New YorkState Museum Bulletin 512, University of the State of New York, Albany, pp. 141–150.
Lovis, W. A., Egan, K. C., Monaghan, G. W., Smith, B. A., and Prahl, E. J. (1996). Environment andsubsistence at the Marquette Viaduct locale of the Fletcher site. In Holman, M., Brashler, J., andParker, K. (eds.), Investigating the Archaeological Record of the Great Lakes State: Essays in Honorof Elizabeth Baldwin Garland, New Issues Press, Kalamazoo, MI, pp. 251–305.
J Archaeol Res (2013) 21:175–216 209
123

Lovis, W. A., Egan-Bruhy, K. C., Smith, B. A., and Monaghan, G. W. (2001). Wetlands and emergenthorticultural economies in the upper Great Lakes: A new perspective from the Schultz site.American Antiquity 66: 615–632.
Lovis, W. A., Urquhart, G. R., Raviele, M. E., and Hart, J. P. (2011). Hardwood ash nixtamalization maylead to false negatives for the presence of maize by depleting bulk d13C in carbonized residues.Journal of Archaeological Science 38: 2726–2730.
Mainfort, R. C. (1979). Indian Social Dynamics in the Period of European Contact, AnthropologicalSeries 1:4, Publications of the Museum, Michigan State University, East Lansing.
Matsuoka, Y., Vigoroux, Y., Goodman, M. M., Sanchez, J., Buckler, E., and Doebley, J. (2002). A singledomestication for maize shown by multilocus microsatellite genotyping. Proceedings of theNational Academy of Sciences 99: 6080–6084.
McAndrews, J. H. (1969). Paleobotany of a wild rice lake in Minnesota. Canadian Journal of Botany 47:1671–1679.
McAndrews, J. H. (1984). Late Quaternary vegetation history of Rice Lake, Ontario, and the McIntyresite. In Johnston, R. B. (ed.), The McIntyre Site: Archaeology, Subsistence and Environment,Archaeological Survey of Canada, Mercury Series, Paper 126, National Museum of Man, Ottawa,pp. 179–196.
McAndrews, J. H. (1994). Pollen diagrams from southern Ontario applied to archaeology. In MacDonald,R. (ed.), Great Lakes Archaeology and Paleoecology: Exploring Interdisciplinary Initiatives for theNineties, University of Waterloo, Waterloo, ON, pp. 179–195.
McCann, J. C. (2004). Maize and Grace: Africa’s Encounter with a New World Crop: 1500–2000,Harvard University Press, Cambridge, MA.
McConaughy, M. A. (2008). Current issues in paleoethnobotanical research from Pennsylvania andvicinity. In Hart, J. P. (ed.), Current Northeast Paleoethnobotany II, New York State MuseumBulletin 512, University of the State of New York, Albany, pp. 9–28.
Mercer, K. L., and Perales, H. R. (2010). Evolutionary response of landraces to climate change in centersof crop diversity. Evolutionary Applications 3: 480–493.
Messner, T. C. (2008). Woodland Period People and Plant Interactions: New Insights from Starch GrainAnalysis, Ph.D. dissertation, Department of Anthropology, Temple University, Philadelphia, PA.
Messner, T. C. (2011a). Acorns and Bitter Roots: Starch Grain Research in the Prehistoric EasternWoodlands, University of Alabama Press, Tuscaloosa.
Messner, T. C. (2011b). Acorns and bitter roots: Starch grain research in the Eastern Woodlands,supplemental material. Electronic document, http://uap.firebrandtech.com/uap/MiscFiles/Messner_SupMaterials.pdf, accessed May 4, 2012.
Messner, T. C., Dickau, R., and Harbison, J. (2008). Starch grain analysis: Methodology and applicationsin the Northeast. In Hart, J. P. (ed.), Current Northeast Paleoethnobotany II, New York StateMuseum Bulletin 512, University of the State of New York, Albany, pp. 111–127.
Miksicek, C. H. (1987). Formation processes of the archaeobotanical record. In Schiffer, M. D. (ed.),Advances in Archaeological Method and Theory, Vol. 10, Academic Press, San Diego, CA,pp. 211–247.
Moffat, C. R., and Arzigian, C. M. (2000). New data on the Late Woodland use of wild rice in northernWisconsin. Midcontinental Journal of Archaeology 25: 49–81.
Monaghan, G. W., Lovis, W. A., and Egan-Bruhy, K. C. (2006). The earliest Cucurbita from the GreatLakes region, northern USA. Quaternary Research 65: 216–222.
Moore, C. R., and Dekle, V. G. (2010). Hickory nuts, bulk processing and the advent of early horticulturaleconomies in eastern North America. World Archaeology 42: 595–608.
Morton, J. D., and Schwarcz, H. (2004). Paleodietary implications from stable isotopic analysis ofresidues on prehistoric Ontario ceramics. Journal of Archaeological Science 31: 503–517.
Mt Pleasant, J. (2006). The science behind the three sisters mound system: An agronomic assessment ofan indigenous agricultural system in the Northeast. In Staller, J. E., Tykot, R. H., and Benz, B. F.(eds.), Histories of Maize: Multidisciplinary Approaches to the Prehistory, Linguistics, Biogeog-raphy, Domestication, and Evolution of Maize, Academic Press, Burlington, MA, pp. 529–538.
Muhammad, A. J. (2010). A Bioarchaeological Study of a Late Woodland Population from Michigan:Frazer-Tyra Site (20SA9), Ph.D. dissertation, Department of Anthropology, Wayne State University,Detroit, MI.
Mulholland, S. C. (1993). A test of phytolith analysis at Big Hidatsa, North Dakota. In Pearsall, D. M.,and Piperno, D. R. (eds.), Current Approaches in Phytolith Analysis: Applications in Archaeology
210 J Archaeol Res (2013) 21:175–216
123

and Paleoecology, MASCA Research Papers in Science and Archaeology, Vol. 10, UniversityMuseum of Archaeology and Anthropology, University of Pennsylvania, Philadelphia, pp. 131–145.
Myers, T. P. (2006). Hominy technology and the emergence of Mississippian societies. In Staller, J. E.,Tykot, R. H., and Benz, B. F. (eds.), Histories of Maize: Multidisciplinary Approaches to thePrehistory, Linguistics, Biogeography, Domestication, and Evolution of Maize, Academic Press,Burlington, MA, pp. 497–510.
Nass, J. P. (2001). Subsistence-settlement change and continuity in western Pennsylvania. In Sullivan, L.P., and Prezzano, S. C. (eds.), Archaeology of the Appalachian Highlands, University of TennesseePress, Knoxville, pp. 177–197.
Niklas, K. J. (1997). The Evolutionary Biology of Plants, University of Chicago Press, Chicago.Parker, K. E. (1990). Botanical remains. In Garland, E. B. (ed.), Late Archaic and Early Woodland
Adaptation in the Lower St. Joseph River Valley, Berrien County, Michigan: The US 31 BerrienCounty Freeway Project, Michigan Cultural Resource Investigation Series, Vol. 2, MichiganDepartment of Transportation, Lansing.
Parker, K. E. (1996). Three corn kernels and a hill of beans: the evidence for prehistoric horticulture inMichigan. In Holman, M. B., Brashler, J. G., and Parker, K. E. (eds.), Investigating theArcheological Record of the Great Lakes State: Essays in Honor of Elizabeth Baldwin Garland,New Issues Press, Kalamazoo, MI, pp. 307–339.
Petersen, J. B., and Asch Sidell, N. (1996). Mid-Holocene evidence of Cucurbita sp. from central Maine.American Antiquity 61: 685–698.
Petersen, J. B., and Cowie, E. R. (2002). From hunter-gatherer camp to horticultural village: Lateprehistoric indigenous subsistence and settlement in New England. In Hart, J. P., and Rieth, C. B.(eds.), Northeast Subsistence-Settlement Change: AD 700–1300, New York State Museum Bulletin496, University of the State of New York, Albany, pp. 265–288.
Piperno, D. R. (2006). Phytoliths: A Comprehensive Guide for Archaeologists and Paleoecologists,AltaMira Press, Lanham, MD.
Piperno, D. R., and Holst. I. (1998). The presence of starch grains on prehistoric stone tools from thehumid neotropics: Indications of early tuber use and agriculture in Panama. Journal ofArchaeological Science 25: 765–776.
Piperno, D. R., Ranere, A. G., Holst, I., and Dickau, R. (2009). Starch grain and phytolith evidence forearly ninth millennium BP maize from the central Balsas River valley, Mexico. Proceedings of theNational Academy of Sciences 106: 5019–5024.
Pressoir, G., and Berthaud, J. (2004). Patterns of population structure in maize landraces from the CentralValleys of Oaxaca in Mexico. Heredity 92: 88–94.
Provine, W. B. (1986). Sewall Wright and Evolutionary Biology, University of Chicago Press, Chicago.Ranere, A. G., Piperno, D. R., Holst, I., Dickau, R, and Iriarte, J. (2009). The cultural and chronological
context of early Holocene maize and squash domestication in the central Balsas River valley,Mexico. Proceedings of the National Academy of Sciences 106: 5014–5018.
Raviele, M. E. (2010). Assessing Carbonized Archaeological Cooking Residues: Evaluation of MaizePhytolith Taphonomy and Density through Experimental Residue Analysis, Ph.D. dissertation,Department of Anthropology, Michigan State University, East Lansing.
Raviele, M. E. (2011). Experimental assessment of maize phytolith and starch taphonomy in carbonizedcooking residues. Journal of Archaeological Science 38: 2708–2713.
Raymond, J. S., and DeBoer, W. R. (2006). Maize on the move. In Staller, J. E., Tykot, R. H., and Benz,B. F. (eds.), Histories of Maize: Multidisciplinary Approaches to the Prehistory, Linguistics,Biogeography, Domestication, and Evolution of Maize, Academic Press, Burlington, MA,pp. 337–342.
Rebourg, C., Chastanet, M., Gouesnard, B., Welcker, C., Dubreuil, P., and Charcosset, A. (2004). Maizeintroduction into Europe: The history reviewed in the light of molecular data. Theoretical andApplied Genetics 106: 895–903.
Riley, T. J., Walz, G. R., Bareis, C. J., Fortier, A. C., and Parker, K. (1994). Accelerator massspectrometry (AMS) dates confirm early Zea mays in the Mississippi River valley. AmericanAntiquity 59: 490–497.
Rindos, D. A. (1984). The Origins of Agriculture, Academic Press, New York.Ritchie, W. A. (1969). The Archaeology of New York State, revised edition, Natural History Press, Garden
City, NY.Ritchie, W. A., and Funk, R. E. (1973). Aboriginal Settlement Patterns in the Northeast, New York State
Museum Memoir 29, University of the State of New York, Albany.
J Archaeol Res (2013) 21:175–216 211
123

Rose, F. (2008). Intra-community variation in diet during the adoption of a new staple crop in the EasternWoodlands. American Antiquity 73: 413–439.
Rousset, R. (2004). Genetic Structure and Selection in Subdivided Populations, Princeton UniversityPress, Princeton, NJ.
Rovner, I. (1999). Phytolith analysis. Science 283: 487.Scarry, C. M. (1993). Variability in Mississippian crop production strategies. In Scarry, C. M. (ed.),
Foraging and Farming in the Eastern Woodlands, University Press of Florida, Gainesville,pp. 78–90.
Schulenberg, J. K. (2002). The Point Peninsula to Owasco Transition in Central New York, Ph.D.dissertation, Department of Anthropology, Pennsylvania State University, University Park.
Schurr, M. R., and Redmond, B. G. (1991). Stable isotope analysis of incipient maize horticulturalistsfrom Gard Island 2 site. Midcontinental Journal of Archaeology 16: 69–84.
Schwarcz, H. P., Melbye, J., Katzenberg, M. A., and Knyf, M. (1985). Stable isotopes in human skeletonsof southern Ontario: Reconstructing paleodiet. Journal of Archaeological Science 12: 187–206.
Scuilli, P. W. (1995). Biological indicators of diet in Monongahela populations. PennsylvaniaArchaeologist 65(2): 1–18.
Simon, M. L. (2000). Regional variations in plant use strategies in the Midwest during the LateWoodland. In Emerson, T. E., McElrath, D. L., and Fortier, A. C. (eds.), Late Woodland Societies:Tradition and Transformation across the Midcontinent, University of Nebraska Press, Lincoln,pp. 37–75.
Simon, M. L. (2009). A regional and chronological synthesis of Archaic period plant use in themidcontinent. In Emerson, T. E., McElrath, D. L., and Fortier, A. C. (eds.), Archaic Societies:Diversity and Complexity across the Midcontinent, State University of New York Press, Albany,pp. 81–114.
Simon, M. L. (2011). Evidence for variability among squash seeds from the Hoxie site (11CK4), Illinois.Journal of Archaeological Science 38: 2079–2093.
Simon, M. L., and Parker, K. E. (2006). Prehistoric plant use in the American Bottom: New thoughts andinterpretations. Southeastern Archaeology 25: 212–257.
Smalley, J., and Blake, M. (2003). Sweet beginnings: Stalk sugar and the domestication of maize. CurrentAnthropology 44: 675–689.
Smith, B. D. (1992). Rivers of Change: Essays on Early Agriculture in Eastern North America,Smithsonian Institution Press, Washington, DC.
Smith, B. D. (2001). Low-level food production. Journal of Archaeological Research 9: 1–43.Smith, B. D. (2006). Eastern North America as an independent center of plant domestication. Proceedings
of the National Academy of Sciences 103: 1223–1228.Smith, B. D., and Yarnell, R. A. (2009). Initial formation of an indigenous crop complex in eastern North
America at 3800 BP. Proceedings of the National Academy of Sciences 106: 6561–6566.Smith, D. G., and Crawford, G. W. (1997). Recent developments in the archaeology of the Princess Point
complex in southern Ontario. Canadian Journal of Archaeology 21: 9–32.Snow, D. R. (1995). Migration in prehistory: The northern Iroquoian case. American Antiquity 60: 59–79.Staller, J. E., Tykot, R. H., and Benz, B. F. (eds.) (2006). Histories of Maize: Multidisciplinary
Approaches to the Prehistory, Linguistics, Biogeography, Domestication, and Evolution of Maize,Academic Press, Burlington, MA.
Stewart, R. M. (1993). Comparison of Late Woodland cultures: Delaware, Potomac, and SusquehannaRiver valleys, middle Atlantic region. Archaeology of Eastern North America 21: 163–178.
Stothers, D. M., and Bechtel, S. K. (1987). Stable carbon isotope analysis: An inter-regional perspective.Archaeology of Eastern North America 15: 137–154.
Stuiver, M., and Reimer, P. J. (1993). Extended 14C database and revised CALIB radiocarbon calibrationprogram. Radiocarbon 35: 215–230.
Styles, B. W. (2011). Animal use by aboriginal societies in the Northeast. In Smith, B. D. (ed.), TheSubsistence Economies of Indigenous North American Societies: A Handbook, SmithsonianInstitution Scholarly Press, Washington, DC, pp. 449–482.
Terrell, J. E. (2008). Domesticated landscapes in the prehistoric Northeast. In Hart, J. P. (ed.), CurrentNortheast Paleoethnobotany II, New York State Museum Bulletin 512, University of the State ofNew York, Albany, pp. 199–204.
Terrell, J. E., and Hart, J. P. (2008). Domesticated landscapes. In David, B., and Thomas, J. (eds.),Handbook of Landscape Archaeology, Left Coast Press, Walnut Creek, CA, pp. 328–332.
212 J Archaeol Res (2013) 21:175–216
123

Terrell, J. E., Hart, J. P., Barut, S., Cellinese, N., Curet, A., Denham, T., Kusimba, C. M., Latinis, K.,Oka, R., Palka, J. Pohl, M. E. D., Pope, K. O., Williams, P. R., Haines, H., and Staller, J. E. (2003).Domesticated landscapes: The subsistence ecology of plant and animal domestication. Journal ofArchaeological Method and Theory 10: 323–368.
Thompson, R. G., and Mulholland, S. C. (1994). Identification of corn in food residues on utilizedceramics at the Shea site (32CS101). The Phytolitharien Newsletter 8(2): 7–11.
Thompson, R. G., Kluth, R. A., and Kluth, D. W. (1994). Tracing the use of Brainerd ware through opalphytolith analysis of food residues. The Minnesota Archaeologist 53: 86–98.
Thompson, R. G., Hart, J. P., Brumbach, H. J., and Lusteck, R. (2004). Phytolith evidence for twentieth-century BP maize in northern Iroquoia. Northeast Anthropology 68: 25–40.
Tieszen, L. L., and Fagre, T. (1993). Effect of diet quality and composition on the isotopic composition ofrespiratory CO2, bone collagen, bioapatite, and soft tissue. In Lambert, J. B., and Grupe, G. (eds.),Prehistoric Human Bone: Archaeology at the Molecular Level, Springer, Berlin, pp.121–155.
Torrence, R., and Barton, H. (2006). Ancient Starch Research, Left Coast Press, Walnut Creek, CA.Trigger, B. G. (ed.) (1976). Handbook of North American Indians, Volume 15: Northeast, Smithsonian
Institution Press, Washington, DC.Tykot, R. H. (2006). Stable isotope analysis and human health. In Staller, J. E., Tykot, R. H., and Benz, B.
F. (eds.), Histories of Maize: Multidisciplinary Approaches to the Prehistory, Linguistics,Biogeography, Domestication, and Evolution of Maize, Academic Press, Burlington, MA,pp. 131–142.
United Nations. (2001). FRA 2000: Global Ecological Zoning for the Global Forest ResourcesAssessment, Final Report 2000, Working Paper 56, Forest Resources Assessment Program, ForestryDepartment, Food and Agriculture Organization of the United Nations, Rome, ftp://ftp.fao.org/docrep/fao/006/ad652e/ad652e00.pdf, accessed January 11, 2012.
United States Department of Agriculture. (2011). USDA National Nutrient Data Base for StandardReference. Nutrient Data Laboratory, Agriculture Research Service, Food and Nutrition InformationCenter and Information Systems Division, National Agricultural Library, Washington, DC,electronic document, http://www.nal.usda.gov/fnic/cgi-bin/nut_s.pl, accessed January 20, 2012.
Valko, A. L. (2001). The Prehistoric Diet and Nutritional Status of the Wylie Site Inhabitants, M.A.Thesis, Department of Anthropology, Kent State University, Kent, OH.
van der Merwe, N. J., Pfeiffer, S., Williamson, R. F., Oakberg Allegretto, K., and Katzenberg, M. A.(2003a). The Moatfield Ossuary: Isotopic dietary analysis of an Iroquoian community, using dentaltissue. Journal of Anthropological Archaeology 22: 245–261.
van der Merwe, N. J., Pfeiffer, S., Williamson, R. F., and Thomas, S. C. (2003b). The diet of theMoatfield people. In Williamson, R. F., and Pfeiffer, S. (eds.), Bones of the Ancestors: TheArchaeology and Osteobiography of the Moatfield Ossuary, Mercury Series, Archaeology Paper163, Canadian Museum of Civilization, Gatineau, QC, pp. 205–222.
van Heerwaarden, J., van Eeuwijk, F. A., and Ross-Ibarra, J. (2010). Genetic diversity in a cropmetapopulation. Heredity 104: 28–39.
Vennum, T. (1988). Wild Rice and the Ojibwa People, Minnesota Historical Society Press, St. Paul.Vierra, B. J., and Ford, R. I. (2006). Early maize in the northern Rio Grande valley, New Mexico. In
Staller, J. E., Tykot, R. H., and Benz, B. F. (eds.), Histories of Maize: Multidisciplinary Approachesto the Prehistory, Linguistics, Biogeography, Domestication, and Evolution of Maize, AcademicPress, Burlington, MA, pp. 497–510.
Vigouroux,Y., Glaubitz, J. C., Matsuoka, Y., Goodman, M. M., Sanchez G. J., and Doebley, J. (2008).Population structure and genetic diversity of New World maize races assessed by DNAmicrosatellites. American Journal of Botany 95: 1240–1253.
Vogel, J. C., and van der Merwe, N. J. (1977). Isotopic evidence for early maize cultivation in New YorkState. American Antiquity 42: 238–242.
Wagner, G. E. (1987). The Uses of Plants by Fort Ancient Indians, Ph.D. Dissertation, Department ofAnthropology, Washington University, St. Louis, MO.
Walker, M. (2005). Quaternary Dating Methods, Wiley, Chichester, UK.Waller, Jr., J. N. (2000). Late Woodland settlement and subsistence in southern New England revisited:
The evidence from coastal Rhode Island. North American Archaeologist 21: 139–153.Waller, Jr., J. N. (2009). ‘‘Growne Up in that Very Place’’: Twelfth to fourteenth century AD social
organization in the territory of the Narragansett. Paper presented at the 74th Annual Meeting of theSociety for American Archaeology, Atlanta, GA.
J Archaeol Res (2013) 21:175–216 213
123

Walz, G. R. (1992). The paleoethnobotany of the Schwerdt site (20AE127), an early fifteenth centuryencampment in the lower Kalamazoo Valley, Allegany County, Michigan. The MichiganArchaeologist 38(1–2): 121–138.
Warrick, G. A. (1996). Evolution of the Iroquoian longhouse. In Coupland, G., and Banning, E. B. (eds.),People Who Lived in Big Houses: Archaeological Perspectives on Large Domestic Structures,Prehistory Press, Madison, WI, pp. 11–26.
Watts, C. M., White, C. D., and Longstaffe, F. J. (2011). Childhood diet and Western Basin traditionfoodways at the Krieger site, southwestern Ontario, Canada. American Antiquity 76: 446–472.
Will, G. F., and Hyde, G. F. (1917). Corn Among the Indians of the Upper Missouri, W. H. Miner, St.Louis, MO.
Williamson, R. F. (1990). The Early Iroquoian period in southern Ontario. In Ellis, C. J., and Ferris, N.(eds.), The Archaeology of Southern Ontario to AD 1650, Occasional Publication 5, OntarioArchaeological Society, London, pp. 291–320.
Winterhalder, B., and Goland, C. L. (1997). An evolutionary ecology perspective on diet, risk, and plantdomestication. In Gremillion, K. J. (ed.), People, Plants, and Landscapes: Studies in Paleobotany,University of Alabama Press, Tuscaloosa, pp. 123–160.
Wright, J. V. (1966). The Ontario Iroquois Tradition, Bulletin 210, National Museum of Canada, Ottawa,ON.
Wright, P. (2003). Preservation or destruction of plant remains by carbonization? Journal ofArchaeological Science 30: 577–583.
Wright, S. (1932). The roles of mutation, inbreeding, crossbreeding and selection in evolution.Proceedings of the Sixth International Congress of Genetics 1: 356–366.
Wright, S. (1978). Evolution and the Genetics of Populations, Volume 4: Variability Within and AmongPopulations, University of Chicago Press, Chicago.
Wymer, D. A. (1992). Trends and disparities: The Woodland paleoethnobotanical record of the Mid-OhioValley. In Seeman, M. (ed.), Cultural Variability in Context: Woodland Settlements of the Mid-OhioValley, Special Paper 7, Midcontinental Journal of Archaeology, Kent State University Press, Kent,OH, pp. 65–76.
Wymer, D. A. (1993). Cultural change and subsistence: The Middle and Late Woodland transition in themid-Ohio Valley. In Scarry, C. M. (ed.), Foraging and Farming in the Eastern Woodlands,University Press of Florida, Gainesville, pp. 138–156.
Yarnell, R. A. (1978). Domestication of sunflower and sumpweed in eastern North America. In Ford, R. I.(ed.), The Nature and Status of Ethnobotany, Anthropological Paper No. 67, Museum ofAnthropology, University of Michigan, Ann Arbor, pp. 289–299.
Bibliography of recent literature
Arzigian, C. (2009). The origins of agriculture in the Midwest: The Eastern Agricultural Complex. InMcManamon, F. P. (ed.), Archaeology in America, An Encyclopedia, Vol. 2, Greenwood Press,Westport, CT, pp. 36–39.
Brashler, J. G., Garland, E. B., Holman, M. B., Lovis, W. A, .and Martin, S. R. (2000). Adaptivestrategies and socioeconomic systems in northern Great Lakes riverine environments: The LateWoodland of Michigan. In Emerson, T., McElrath, D., and Fortier, A. (eds.), Late WoodlandSocieties: Tradition and Transformation across the Midcontinent, University of Nebraska Press,Lincoln, pp. 543–582.
Buckmaster, M. M. (2004). The northern limits of ridge field agriculture: An example from MenomineeCounty. In Lovis, W. A. (ed.), An Upper Great Lakes Archaeological Odyssey: Essays in Honor ofCharles E. Cleland, Cranbrook Institute of Science, Bloomfield Hills, MI pp. 30–42.
Bush, L. L. (2004). Boundary Conditions: Macrobotanical Remains and the Oliver Phase of CentralIndiana, AD 1200–1450, University of Alabama Press, Tuscaloosa.
Chilton, E. S. (2009). Pre-contact maize horticulture in New England: A summary of the archaeologicalevidence. In Chilton, E. S., and Rainey, M. L. (eds.), Nantucket and Other Native Places, StateUniversity of New York Press, Albany, pp. 159–179.
Chilton, E. S. (2010). Mobile farmers and sedentary models: Horticulture and cultural transitions in LateWoodland and contact period New England. In Alt, S. (ed.), Ancient Complexities: New
214 J Archaeol Res (2013) 21:175–216
123

Perspectives in Pre-Columbian North America, University of Utah Press, Salt Lake City,pp. 96–103.
Chilton, E. A., Largy, T. B., and Curran, K. (2000). Evidence for prehistoric maize horticulture at the PineHills site, Deerfield, Massachusetts. Northeast Anthropology 59: 23–46.
Cook, R. A. (2008). Sunwatch: Fort Ancient Developments in the Mississippian World, University ofAlabama Press, Tuscaloosa.
Foreman, J. L. (2011). Seasonal subsistence in Late Woodland southwestern Ontario: An examination ofthe relationships between resource availability, maize agriculture, and faunal procurement andprocessing strategies. Electronic Thesis and Dissertation Repository, Paper 280, http://ir.lib.uwo.ca/etd/280, accessed November 22, 2011.
Fritz, G. J. (2011). The role of ‘‘tropical’’ crops in early North American agriculture. In Smith, B. D. (ed.),The Subsistence Economies of Indigenous North American Societies: A Handbook, SmithsonianInstitution Scholarly Press, Washington, DC, pp. 503–516.
Holman, M. B., and Lovis, W. A. (2008). Social and environmental constraints on mobility in the lateprehistoric upper Great Lakes, USA. In Barnard, H., and Wendrich, W. (eds.), The Archaeology ofMobility: Old World and New World Nomadism, Cotsen Institute of Archaeology, University ofCalifornia, Los Angeles, pp. 280–306.
Howey, M. C. (2007). Using multi-criteria cost surface analysis to explore past regional landscapes: Acase study of ritual activity and social interaction in Michigan, AD 1200–1600. Journal ofArchaeological Science 34: 1830–1846.
Howey, M. C., and O’Shea, J. M. (2006). Bear’s journey and the study of ritual in archaeology. AmericanAntiquity 71: 261–282.
Lovis, W. A. (2012). Revisiting the Johnson site (20CN46): A winter site in a cedar swamp on MullettLake, Michigan. In Brashler, J. G., and Lovis, W. A. (eds.), Papers in Memory of Margaret B.‘‘Sister Peg’’ Holman, special issue of The Michigan Archaeologist, in press.
Lovis, W. A., and Bogdan-Lovis, E. A. (2004). Ridged fields, catastrophic wet season planting, andgermination rates: Notes from Libby’s garden. The Wisconsin Archeologist 85(1): 78–83.
Martin, S. W. (2004). Lower Great Lakes maize and enchainment in the first millennium AD. OntarioArchaeology 77/78: 135–159.
Martin, S. W. (2006). Earmarked: Maize, Materiality and Agricultural Frontiers in the Lower GreatLakes Region of North America, Ph.D. dissertation, Department of Archaeology, CambridgeUniversity, Cambridge.
Means, B. K. (2007). Circular Villages of the Monongahela Tradition, University of Alabama Press,Tuscaloosa.
Means, B. K. (2012). Villagers and farmers of the middle and upper Ohio River valley, 11th to 17thcenturies AD: The Fort Ancient and Monongahela traditions. In Pauketat, T. (ed.), The OxfordHandbook of North American Archaeology, Oxford University Press, Oxford, pp. 297–309.
Miroff, L. E. (2008). A local-level analysis of social reproduction and transformation in the ChemungValley: The Thomas/Luckey site. In Miroff, L. E., and Knapp, T. D. (eds.), Iroquoian Archaeologyand Analytical Scale, University of Tennessee Press, Knoxville, pp. 69–100.
Mt Pleasant, J. (2010). Estimating productivity of traditional Iroquoian cropping systems from fieldexperiments and historical literature. Journal of Ethnobiology 30: 52–79.
Mullins, H. T., Patterson, W. P., Teece, M. A., and Burnett, A. W. (2011). Holocene climate andenvironmental change in central New York (USA). Journal of Paleolimnology 45: 243–256.
Nass, J. P., and Hart, J. P. (2000). Subsistence-settlement change during the late prehistoric period in theupper Ohio River valley: New models and old constructs. In Genheimer, R. A. (ed.), Cultures BeforeContact: The Late Prehistory of Ohio and Surrounding Regions, Ohio Archaeological Council,Columbus, pp. 124–157.
Nolan, K. C., and Cook, R. A. (2010). An evolutionary model of social change in the middle Ohio Valley:Was social complexity impossible during the Late Woodland but mandatory during the lateprehistoric? Journal of Anthropological Archaeology 29: 62–79.
O’Shea, J. M., and Milner, C. M. (2002). Material indicators of territory, identity, and interaction in aprehistoric tribal system. In Parkinson, W. A. (ed.), The Archaeology of Tribal Societies,Archaeological Series 15, International Monographs in Prehistory, Ann Arbor, MI, pp. 200–226.
Ounjian, G. L. (1998). Glen Meyer and Prehistoric Neutral Paleoethnobotany, Ph.D. dissertation,Department of Anthropology, University of Toronto, Toronto, ON.
J Archaeol Res (2013) 21:175–216 215
123

Scarry, C. M. (2003). Patterns of wild plant utilization in the prehistoric eastern woodlands. In Minnis, P.E. (ed.), People and Plants in Ancient Eastern North America, Smithsonian Books, Washington,DC, pp. 50–104.
Scarry, C. M., and Yarnell, R. A. (2011). Native American domestication and husbandry of plants in easternNorth America. In Smith, B. D. (ed.), The Subsistence Economies of Indigenous North AmericanSocieties: A Handbook, Smithsonian Institution Scholarly Press, Washington, DC, pp. 483–502.
Smith, B. A., and Egan, K. C. (1990). Middle and Late Archaic faunal and floral exploitation at the WeberI site (20SA581), Michigan. Ontario Archaeology 50: 39–54.
Smith, B. D. (2006). Eastern North America as an independent center of plant domestication. Proceedingsof the National Academy of Sciences 103: 1223–1228.
Smith, B. D., and Cowan, C. W. (2003). Domesticated crop plants and the evolution of food productioneconomies in eastern North America. In Minnis, P. E. (ed.), People and Plants in Ancient EasternNorth America, Smithsonian Books, Washington, DC, pp. 105–125.
Stein, N. R. (2007). Native Peoples and Subsistence in Late Woodland and Early Contact PeriodSouthern New England, Ph.D. dissertation, Department of Anthropology, Brown University,Providence, RI.
Vickery, K. D., Sunderhaus, T. S., and Genheimer, R. A. (2000). Preliminary report on excavations at theFort Ancient State Line site, 33 Ha 58, in the Central Ohio valley. In Genheimer, R. A. (ed.), CulturesBefore Contact: The Late Prehistory of Ohio and Surrounding Regions, Ohio Archaeological Council,Columbus, pp. 272–329.
Wagner, G. E. (2003). Eastern woodlands anthropogenic ecology. In Minnis, P. E. (ed.), People andPlants in Ancient Eastern North America, Smithsonian Books, Washington, DC, pp. 126–171.
Zalucha, L. A. (1997). Paleoethnobotanical analysis of the Statz site. In Meinholz, N. M., and Kold, J. L.(eds.), The Statz Site (47 Da-642): A Late Woodland Community and Archaic Lithic Workshop inDane County, Wisconsin, Archaeology Research Series No. 5, State Historical Society of Wisconsin,Madison, pp. 171–191.
216 J Archaeol Res (2013) 21:175–216
123

Copyright of Journal of Archaeological Research is the property of Springer Science & Business Media B.V.and its content may not be copied or emailed to multiple sites or posted to a listserv without the copyrightholder's express written permission. However, users may print, download, or email articles for individual use.




















