
Redefining Darius: A New Perspective on the Battle of Gaugamela
Upload
independentCategory
view
0download
0
RPL
ACstistarmtnstmibmcaptbnbfsticgc3
KBe
o
dOE0
Ed
Review Article
6
edefining the Persistent Infection in Root Canals:ossible Role of Biofilm Communities
uis Chávez de Paz, DDS, MS, PhD
Amtptofegbmdmikvtnr(cwce
bmams(abcvpreistp
ta(
bstracturrent concepts suggest that persisting infections sub-equent to endodontic therapy are caused by one orwo bacterial species that are “too robust” to be elim-nated by conventional treatment measures. As a con-equence, numerous studies are exploring the charac-eristics of these “most” resistant organisms to definen effective treatment strategy to eradicate them fromoot canals. By taking an ecological perspective, theain objective of this review is to present evidence that
he nature of persisting endodontic infections dependsot on the robustness of the organisms in the infectedite, but on their capability of adapting their physiologyo the new environmental conditions set by the treat-ent. Changes in the environment, such as an increase
n pH by calcium hydroxide or the effect of antimicro-ials, are capable of triggering genetic cascades thatodify the physiological characteristics of bacterial
ells. Surface adherence by bacteria to form biofilms isgood example of bacterial adaptation and one that isertinent to endodontic infections. Increasing informa-ion is now available on the existence of polymicrobialiofilm communities on root canal walls, coupled withew data showing that the adaptive mechanisms ofacteria in these biofilms are significantly augmentedor increased survival. This ecological view on the per-isting infection problem in endodontics suggests thathe action of individual species in persisting endodonticnfections is secondary when compared to the adaptivehanges of a polymicrobial biofilm community under-oing physiological and genetic changes in response tohanges in the root canal environment. (J Endod 2007;3:652–662)
ey Wordsacterial adaptation, Enterococcus faecalis, microbialcology, pathogens, physiological changes
From the Department of Oral Biology, Faculty of Odontol-gy, Malmö University, Malmö, Sweden.
Address requests for reprints to Dr. Luis Eduardo Cháveze Paz Villanueva, Department of Oral Biology, Faculty ofdontology, Malmö University, SE-205 06 Malmö, Sweden.-mail address: [email protected]/$0 - see front matter
Copyright © 2007 by the American Association ofndodontists.oi:10.1016/j.joen.2006.11.004
a
52 Chávez de Paz
traditional concept that explains infectious processes occurring in humans sug-gests that diseases are produced as the result of the aggressive invasion of harmful
icroorganisms, which battle with the human host’s defenses, triggering mechanismshat release antibodies and immune cells. The impact of such an approach generates aredisposition to search for those “most dangerous” microorganisms that can cause/
rigger the most severe damage to the host. In line with this view, infectious processesf the oral cavity were proposed to be caused by a relatively small number of organisms
rom the diverse collection of species found in the human mouth (1). In caries, forxample, the frequent isolation of Streptococcus mutans from carious lesions (2– 4)enerated a considerable number of studies to explore the ex vivo features of thisacterium. Research findings showing the significant acid-tolerant capabilities of S.utans defined this organism as “the” agent responsible for initial enamel and dentine
emineralization. Similarly, in periodontal disease, the frequent recovery of proteolyticicroorganisms from deep periodontal pockets, such as Porphyromonas gingivalis,
ncreased the attention of periodontists to these bacteria because they were consideredey etiological agents of the disease (5, 6). The main disadvantage with this traditionaliew of the infectious process, especially in oral infections, is that the determination ofrue cause-and-effect relationships is not always possible. Consequently, the predomi-ance of certain microorganisms at a given site may be the result of the disease itselfather than that of the initiating agent (7). Recently, the “ecological plaque hypothesis”8 –13) has improved on these classic infectious concepts to explain the etiology ofaries and periodontal disease. This hypothesis suggests that the organisms associatedith the disease may also be present at sound sites, but at levels too low to represent alinical threat. In other words, disease is produced as the result of changes in the localnvironmental conditions that will shift the balance of the resident flora.
Root canal infections have a different nature than that of caries or periodontitisecause they become established in originally sterile compartments of the oral cavity. Inany cases, this led to the concept that the etiology of root canal infections involves onlysingle pathogen. For example, the predominance of certain proteolytic black-pig-ented anaerobic organisms in cultures from infected root canals associated with acute
ymptoms suggested that these organisms are foremost etiological agents in such cases14, 15). Recently, the frequent recovery of Enterococcus faecalis in root canalsssociated with persistent infections brought about an intense research interest in thisacterium. E. faecalis has become the ideal organism to test different irrigants, medi-aments, and antiseptic solutions used in endodontics ex vivo, with findings that re-ealed its innate resistance capacity (16 –18). This extensive interest in E. faecalis,erhaps driven by its ability to grow under almost any laboratory condition (19),esulted in the concept that the organism is the sole etiological agent for chronicndodontic infections. Consequently, the focus on E. faecalis resulted in much lessnformation on the existence of other organisms in such infections that may possessimilar tolerating characteristics to E. faecalis and that would shed light on the exis-ence of a polymicrobial persisting community. Thus, it is not surprising that ecologicalarameters in root canal infections are not often discussed.
From an ecological perspective, the root canal can be considered a highly con-rolled environment with a limited number of niches. Although niches are composed byvariety of environmental factors that limit the growth of one species relative to others20), the main limiting factors in root canal niches that influence bacterial colonization
re, for instance, oxygen and nutrient availability (21). After root canal treatment, otherJOE — Volume 33, Number 6, June 2007

letiec
ristlngnapp“ut
uscpt(dm
sbedmphgs
riiipcudvmmswiipcrbosp
O
vckicopodrcctsscgawoi
ccmbt(bvtdo
F
Review Article
J
imiting factors become involved, such as pH and the short-/long-termffects of the antibacterial medicaments applied. The limiting factors forhree different niches in root canals are depicted in Fig. 1, from whicht can be reasonably assumed that bacterial survival in such controllednvironments, especially after root canal treatment, is based on theapacity of organisms to adapt to the existing conditions.
Although traditional views suggest that the organisms survivingoot canal treatment are a selected group of the “most robust” organ-sms, the application of ecological parameters indicates that bacterialurvival after root canal treatment will depend not on the robustness ofhe organisms, but on how good an adaptor the organism is to the newimiting factors in their corresponding niches. Furthermore, as in everyatural microenvironment, the adaptive capabilities of individual or-anisms are exponentially augmented when growing in biofilm commu-ities. As proposed in this review, data now exist (Fig. 2) that provide anrgument for the inclusion of the biofilm concept in the etiology ofersisting endodontic infections. The foundation for this ecological ap-roach to endodontic infections suggests that the most dangerouspathogen” is not an individual species, but a polymicrobial entity thatndergoes physiological and genetic changes triggered by changes inhe root canal environment.
Pathogens and VirulenceThe accepted definitions of microbial pathogen and microbial vir-
lence were formulated largely from the study of infections caused by aingle etiological agent, although both represent combinations of highlyomplex parameters. As a result, classifications of microbial pathogenslace the primary responsibility for causing a determined disease at allimes on the microorganism [for a review see Casadevall and Pirofski22)]. Likewise, the virulence of a pathogen is generally defined as theegree of pathogenicity or ability of the organism to cause diseaseeasured by an experimental procedure (23).
The theory of the single pathogen as the etiological agent of apecific disease is derived from Robert Koch’s Postulates, which areased on the work accomplished by Koch and his coworkers on dis-ases, such as, tuberculosis, diphtheria, anthrax, and cholera, that wereetermined to be caused by specific microbial entities at a time when theost prominent medical and scientific communities denied their im-
ortance [see Kaufmann and Schaible (24)]. By current standards,owever, there are very few microorganisms to which the term patho-en can be applied invariably (25). A common example is group A
igure 1. Different niches and limiting factors in the root canal environment.
treptococci, which are etiological agents of acute rheumatic fever, l
OE — Volume 33, Number 6, June 2007
heumatic heart disease, post-streptococcal glomerulonephritis, andnvasive infections causing at least 517,000 deaths each year (26). Notncluded in such a traditional “pathogenic” categorization are the var-ous microorganisms that we encounter on a daily basis coexistingeacefully with humans and from which a small number of them may beapable of a pathogenic phase. For example, Escherichia coli is ubiq-itous, asymptomatically colonizing the human intestines and is widelyistributed in the environment, yet, after experiencing specific geneticariations, this organism can cause epidemic dysentery and neonataleningitis, among other diseases (27). Another example is Listeriaonocytogenes, which is well adapted as a saprophyte for peaceful
urvival in soil and decaying vegetation, but it has also another phasehere it acts as an intracellular invader capable of causing serious
nfections in humans (28 –30). Current research suggests that the abil-ty of these opportunistic organisms to switch from the harmless to theathogenic state appears to occur in response to environmentalhanges that are mediated through complex regulatory pathways, whicheversibly modulate the expression of virulence factors. The advent ofacterial genomic studies has significantly increased our understandingf the pathogenic state of many microbes with the on/off virulencewitch, in fact, constituting a valuable marker of individual microbialathogens (31–33).
ral PathogensHistorically, much of the earlier research into dental caries and the
arious periodontal diseases was focused on correlating a single spe-ific organism with the disease to satisfy Koch’s Postulates. We nownow that not all diseases are the result of the action of a single organ-
sm and this is particularly true of the oral cavity where all of the mi-robial diseases associated with tissue destruction involve more thanne type of organism and are, therefore, “mixed” infections (34). Thisolymicrobial nature of oral disease has its basis in the characterizationf the organisms present in dental plaque and their potential roles inental caries, gingivitis, and periodontitis (35–37). Early data (38)ecognized the association of mutans streptococci, including Strepto-occus mutans and S. sobrinus, with the initial phase of human dentalaries because their acidogenic and aciduric properties permitted themo create a low-pH environment in dental plaque after the ingestion ofugars. In addition, lactobacilli and certain acid tolerant non-mutanstreptococci can now be considered virulent with respect to dentalaries (39, 40). In periodontal disease, the use of animal models sug-ested that Actinomyces naeslundii was involved in the destructivelveolar bone loss characteristic of advanced periodontitis (41),hereas evidence was also presented in the 1970s that black-pigmentedrganisms, such as Porphyromonas gingivalis, were directly involved
n periodontitis (36).Marsh (7) proposed the “ecological plaque hypothesis” to explain
hanges in the ecology of dental plaque that lead to the development ofaries or periodontal disease. This hypothesis constitutes a dynamicodel in which plaque-mediated diseases are the consequence of im-
alances in the resident microflora resulting from an enrichment withinhe microbial community of the above mentioned “oral pathogens”9 –11, 13). In caries, for example, potentially cariogenic bacteria maye found naturally in dental plaque, but at neutral pH and with a con-entional low-sugar diet, the levels of such potentially cariogenic bac-eria are clinically insignificant. If the intake of fermentable carbohy-rates increases, the low pH provoked in plaque favors the proliferationf acidogenic and aciduric bacteria, such as mutans streptococci and
actobacilli, which promote enamel demineralization.
Persistent Infection in Root Canals 653

FtarO
Review Article
6
igure 2. Preparation of a tooth apex associated with chronic infection as observed with scanning electron microscope. Overview in (a) at �40 magnification showshe main root canal obtured with gutta percha (arrow). Magnifications of �150 in (b) and �300 in (c) show a gap between the gutta-percha and the root canal wall,
section is marked in (c) for higher magnification. Images (d), (e), and (f) show the marked section at magnifications of �2,500, �7,000, and �10,000,espectively. Accumulations of bacterial cells adhered in the gap between the gutta percha and the root canal wall are observed. The specimen was provided by Dr.
lle Henningsson, Department of Endodontics, Malmö University, and the images were obtained with a Hitachi electron microscope.54 Chávez de Paz JOE — Volume 33, Number 6, June 2007

E
mvtotismaifirctiFftcrtt1tfinof4
fa
pmiwtwpaiafivtrwmlloaii
ot(ntgFabcbtfe(lgatcschilb(oer
Fbwcm1cpd
Review Article
J
ndodontic PathogensCurrently, there is no substantial evidence indicating that certain
icroorganisms of the microbial flora in root canal infections are moreirulent than others. With this in mind, Sundqvist and Figdor (42) statedhat a proper definition for endodontic pathogens should include everyrganism capable of inducing the tissue destruction in apical periodon-itis. In reality, however, the majority of endodontic-microbiology stud-es refer to the endodontic pathogen as the bacterium isolated from aymptom-associated root canal that grows in the laboratory in a specificedia. By this approach, the most frequently recovered species will
ssume the role of major endodontic pathogen. In persistent root canalnfections, for example, the frequent occurrence of monocultures of E.aecalis has raised suspicion that this bacterium may be the sole organ-sm persisting in the root canals. Considering that mono-infectionsarely if ever occur in nature, it is possible that the apparent pureultures of E. faecalis could be the result of sampling and culturingechniques that favor it over other organisms at the site that were eithern low numbers or were physiologically inactive or dormant (see later).or instance, in a commonly cited study (43), from the total 100 root-illed teeth with apical periodontitis sampled E. faecalis was reported ashe most frequently recovered organism (32%), although in 32% of theases with persistent lesion no microbe could be isolated. In yet nineoot-filled teeth without periapical lesion that showed bacterial growth,he organism was found in one case. In a similar study, 25 root-filledeeth requiring retreatment were sampled and E. faecalis was found in4 of those 20 teeth with bacterial growth (44). However, it would seem
hat this study was focused primarily in proving the occurrence of E.aecalis in root-filled teeth rather than in exploring the microbial floran persisting infections. Similarly, in a recent study using a sophisticatedested PCR technique, the target bacterium E. faecalis was found in 41f 50 (82%) untreated root canals and in 38 of 50 (76%) treatment-ailure–associated root canals (45). As in other related works (46 –
E. f
aeca
lisL.
par
acas
eiS.
ang
inos
usPlanktonic cells Biofilms
pH 7 pH 10.5 pH 10.5
Viable cells: 98-100%
Viable cells: 98-100%
Viable cells: 98-100%
Viable cells: 81-93%
Viable cells: 90-98%
Viable cells: 0-3%
Viable cells: 90-98%
Viable cells: 70-98%
Viable cells: 84-95%
igure 3. Fluorescence micrographs using Live/Dead fluorescence staining foracterial viability. Cells stained fluorescent green represent viable cells,hereas cells stained fluorescent red are nonviable or damaged. In the firstolumn, images show planktonic cells of three root canal strains at neutraledia (pH 7). The middle column shows planktonic cells after exposure to pH
0.5 for 4 hours, and the right column shows biofilm cells exposed to alkalinehallenge (pH 10.5) for 4 hours. Bars, 2 �m. Images are published withermission of Blackwell Publishing. International Endodontic Journal, Cháveze Paz et al. (65).
9), PCR methodology seems to be exclusively directed to find only E. o
OE — Volume 33, Number 6, June 2007
aecalis, ignoring the rest of the flora present that may be as importants E. faecalis in provoking the treatment failures.
On the other hand, recent investigations have confirmed theolymicrobial nature of root canal infections (50, 51). In a study withonkeys (50), different combinations of bacteria were experimentally
noculated in root canals and periapical lesions were induced. The teethere treated endodontically and followed-up radiographically and his-
ologically for 2 to 2.5 years. In the root canals with bacteria presenthen the root filling was removed, 30 of the 31 canals had persistingeriapical lesions. Importantly, more of these nonhealed lesions weressociated with various combinations of bacterial strains, that is, mixednfections, than single strains. Previously, the same research group (52)lso found that when an “eight-strain collection” of species, derivedrom one infected root canal, was re-inoculated in equal proportionsnto other monkey teeth, species such as Bacteroides oralis (now Pre-otella oralis) dominated in mixed infections and showed a more po-ent capacity for tissue destruction. Furthermore, B. oralis could not bee-isolated from inoculated root canals after the experimental periodhen inoculated as a pure culture. In another study using the tissue cageodel implanted subcutaneously in the backs of rabbits, the same col-
ection of eight bacterial strains from monkey root canals were inocu-ated in different combinations and individual species. The combinationf B. oralis, Fusobacterium necrophorum, Peptostreptococcusnaerobius, and Streptococcus milleri was the most predominant and
nduced higher titers of circulating antibodies than that obtained withndividual inoculations, such as E. faecalis (53).
Even if we accept the polymicrobial nature of root canal infections,ne of the major problems in understanding endodontic infections is
hat we still extrapolate between individual organisms growing in liquidplanktonic) cultures and the in vivo situation. A significant literatureow exists demonstrating that the physiology of a bacterium in plank-
onic culture is profoundly different from that of the same organismrowing on a surface in a biofilm [see review by Costerton et al. (54)].or instance, planktonic bacteria are more sensitive to antimicrobialgents because of their ease of diffusion within the bulk fluid, whereasiofilm bacteria are notably resistant to these agents (55–59). In thisontext, the study of biofilms in root canal infections has includediofilms formed by mixed cultures of anaerobic bacteria in extracted
eeth (60, 61) or by pure cultures of E. faecalis (62, 63). Biofilms ofive root canal isolates have also been used to test the antimicrobialfficacy of endodontic irrigants, such as sodium hypochlorite (NaOCl)2.25%), 0.2% chlorhexidine, 10% povidone iodine, and 5 ppm col-oidal silver, with NaOCl shown to be the most effective agent of thisroup (64). In addition, our research group tested the alkaline toler-nce of species isolated from chronically infected root canals and foundhat E. faecalis and other Gram-positive organisms, such as Lactoba-illus paracasei, Olsenella uli, or Streptococcus gordonii, sharedimilarly high alkaline-tolerant capabilities when growing in planktoniconditions. S. anginosus, S. oralis, and F. nucleatum, on the otherand, were greatly affected by the alkaline stress (see Fig. 3) (65). Of
mportance, however, was the observation that this difference in alka-ine tolerance was not apparent when the strains were tested in biofilmsecause all seven strains showed a similar high tolerance to alkaline pHFig. 3). These findings not only show the capacity of root canal bacteriather than E. faecalis to adapt to alkaline stress, but also provide furthervidence that bacteria in surface-adhered biofilm consortia are moreesistant to environmental stress than when grown in liquid culture.
Physiological Status of BacteriaThe previous discussion relative to the adaptation and survival of
ral bacteria under environmental stress indicates the importance
Persistent Infection in Root Canals 655

oaotss(agspw1
ccvahtrcsimtmrctrctltb
pitfaoienfbtt(L(dop(saaap
awgocTr(to
wcif
pisnetuOawrcosfa8aamhte
B
f
F
Review Article
6
f the physiological state of bacteria with respect to the potential level ofctivity in disease processes. However, the exact description of the statusf a microorganism can be complex given the numerous terms usedo describe different physiological states, such as, dead, moribund,tarved, dormant, resting, quiescent, viable-but-not-culturable, injured,ublethally damaged, inhibited, resuscitable, living, active, and vital66). Many of these terms are used conceptually and do not reflect thectual knowledge of the state of the organism in question (67). That thisap in information exists is apparent from recent bacterial genomicequencing data that indicate how little we still know about bacterialhysiology. This can be seen in studies with Haemophilus influenzae,hich has 736 genes of unknown function in its genome out of a total of,743 genes (68).
Viability of bacteria is conventionally defined as the capacity ofells to perform all cell functions necessary for survival under givenonditions (66). The simplest method we have used to assess bacterialiability is the plate count method, where the number of viable cellspproximates the number of colony-forming units. Bacteria, therefore,ave been classified into two physiological groups: those that can andhose that cannot readily be grown to detectable levels in vitro (67). Inoot canal infections, we have cultivated microbial strains from rootanal samples in the laboratory, using growth media that contained apecific substrate, and then identified the cultivated bacteria at the phys-ological, biochemical, and, more recently, at the molecular level. The
etabolic properties of these bacterial isolates were then used to inferhe potential roles of these and related microorganisms in the environ-
ent. Under some circumstances, however, such methods may under-epresent the number of viable bacteria for a variety of reasons, such asases where slightly damaged organisms are present (69), the labora-ory growth media used are deficient for one or more essential nutrientsequired for the growth of some bacteria in the sample (70), or viableells are present that have lost their ability to form colonies (71). Fur-hermore, if the bacteria exist in a biofilm they may assume a status ofow-metabolic activity similar to stationary-phase planktonic growth forhe majority of time (72, 73). The bacteria in such low active states maye undetectable by regular culture techniques.
Currently, a variety of methods have been developed to assess thehysiological status of bacterial cells, including metabolic activity, thentegrity of the cell, or the presence of nucleic acids (Fig. 4). Perhapshe most useful are fluorescent probes that target different cellularunctions, such as membrane potential, enzyme activity, nucleic acids,nd membrane integrity [see review by Joux and Lebaron (74)]. Basedn the detection of the amount of rRNA in bacterial cells, fluorescencen situ hybridization (FISH) has been applied to identify and, to anxtent, determine the physiological state of different species in theiratural environments (75). One weakness of FISH is the generation of
alse negative results with bacteria possessing low physiological activityecause they exhibit low amounts of rRNA, resulting in low signal in-ensity (76). In endodontics, FISH has been used to visualize and iden-ify bacteria from periapical lesions of asymptomatic root-filled teeth77). Alternative fluorescent probes to test bacterial viability are theive/Dead BacLight kit and the tetrazolium salts 2-(4-iodophenyl)-3-4-nitrophenyl)-5-phenyl tetrazolium chloride (INT) and 5-cyano-2,3-itolyl tetrazolium chloride (CTC). The Live/Dead kit tests the integrityf the cell membrane by applying two nucleic acid stains, SYTO-9 andropidium iodide (PI), which can simultaneously detect dead/injuredfluorescent red by stain with PI) and intact cells (fluorescent green bytaining with SYTO-9) (78, 79). This fluorescent probe has been used tossess the viability of root canal strains ex vivo (65) and to determine theutoaggregation and coaggregation of bacteria isolated from teeth withcute endodontic infections (80). Furthermore, the outcome of such a
rocedure can be seen in Fig. 5 with a histological preparation of a root u56 Chávez de Paz
pex specimen obtained after surgical removal of a tooth associatedith persistent radiographic signs and symptoms. The filamentous or-anisms observed in the histological images (Fig. 5a and b), are alsobserved under the fluorescent microscope, where their viabilityan be assessed (cells stained fluorescent green in Fig. 5c and d).he tetrazolium salts INT and CTC are often used as markers of bacte-ial respiratory activity, as well as viability [for a review see Creach et al.81)]. With these relatively simple methods, a good correlation be-ween the number of INT/CTC-positive cells and the CFU count can bebtained.
In conclusion, the physiological state of bacteria in root canals,hether they are viable, dormant, injured, or in another state of their lifeycle, is a crucial marker that measures their involvement in periapicalnflammation rather than only the identification of species from DNAragments.
Mechanisms of AdaptationThere are a number of different mechanisms used by bacteria that
ermit them to adapt to the environment. Biofilm formation (55), phys-ological modification (82), stress response (83), and the creation ofubpopulations of cells (84) are among some of the adaptive mecha-isms used by bacteria along with various mechanisms involving thexchange of genetic material between bacteria (85). The exploration ofhese mechanisms can aid us in understanding the bacterial survivalnder the limiting environments, such as that found in the root canal.ne of the most relevant features of adaptation for oral bacteria is thedhesion to surfaces that leads to the formation of plaque biofilms,hich serves not only to aid in their retention in the oral cavity, but also
esults in increased survival (86). Interestingly, this ability to formomplex biofilm communities is not lost when oral organisms colonizether sites in the human body. For example, species from viridians oraltreptococci, such as Streptococcus oralis and S. gordonii, have beenound to form plaque biofilms on the endocardium and valve leafletsnd are thus considered major etiological agents in endocarditis (87,8). Similarly, oral microorganisms are able to colonize root canals bydhering to the dentine walls as shown in microphotographs of a toothpex associated with a chronic infection taken with a scanning electronicroscope (Fig. 6). Aggregations of microorganisms can be seen ad-
ering to the inner walls of an accessory canal under high magnifica-ion, thus demonstrating the retention of these biofilm communitiesven after root canal treatment.
iofilmsGiven that surface-associated microbial communities are the main
orm of colonization and retention by oral bacteria in the mouth, it is not
igure 4. Different approaches used in the assessment of bacterial viability.
nreasonable to assume that biofilms also form in root canals having the
JOE — Volume 33, Number 6, June 2007

scbaFs(sfsttasihbrtdds
e
wcacw
mpiat(aTcf(actcib
Fbbcpa
Review Article
J
ame properties as the parent communities colonizing the enamel andementum surfaces (see Figs. 2 and 6). Biofilms form when planktonicacteria in a natural liquid phase are deposited on a surface containingn organic conditioning polymeric matrix or “conditioning film” (seeig. 7). In this dynamic process many other organisms co-adhere to theurface and grow with some cells detaching from the biofilm over time89 –92). Biofilm formation in root canals, as hypothesized by Sven-äter and Bergenholtz (93), is probably initiated at some time after theirst invasion of the pulp chamber by planktonic oral organisms afterome tissue breakdown. At this point, the inflammatory lesion frontagehat moves successively toward the apex will provide the fluid vehicle forhe invading planktonic organisms so these can multiply and continuettaching to the root canal walls. Interestingly, bacteria have been ob-erved to detach from inner root canal surfaces and occasionally massn the inflammatory lesion per se (94). This observation could explainow the inflammatory lesion front serves as a fluid source for bacterialiofilm detachment and colonization of other inaccessible sites in theoot canal. Thus, when biofilms are formed on surfaces located beyondhe reach of mechanical removal and the effects of antimicrobials, host-erived proteins from remaining necrotic tissues and bacterially pro-uced adhesive substances will provide the proper prerequisites for theurvival of microbes.
Biofilms in root canals have been confirmed by examinations of
a
dc
igure 5. Histological preparation of a section of a tooth associated with chronacteria, stained with a Taylor-modified Brown and Brenn staining. The high my neutrophils. Images (c), (d), and (e) show sections stained with Live/Dead Baells represent viable cells, whereas fluorescent red cells are injured or dead ceroduced are material of the manuscript in preparation (Chávez de Paz LE, Ripexes).
xtracted teeth with periapical lesions. For example, when sections p
OE — Volume 33, Number 6, June 2007
ere viewed by transmission electron microscopy, dense aggregates ofocci and rods embedded in an extracellular matrix were observedlong the walls (95), whereas studies using scanning electron micros-opy showed microcolonies of cocci, rods, and filaments on root canalalls (93, 96, 97). Such root canal biofilms can be seen in Figs. 2 and 6.
Introducing this biofilm concept to endodontic microbiology is aajor step forward in our understanding of root canal infections, es-
ecially those of the persistent kind, because microorganisms growingn biofilms are better protected from adverse environmental changesnd other antimicrobial agents (56, 98). Apart from the physical pro-ection provided by the extracellular matrix [see review by Branda et al.99)], additional protection is afforded by physiological changes initi-ted by the bacteria after their adhesion to the surface (55, 100, 101).hese phenotypic changes by the biofilm bacteria usually result in in-reased resistance to antimicrobial agents, in some cases up to 1,000-old greater than that of the same microorganisms living planktonically56, 57). Evidence already exists showing that biofilms of oral bacteriare more resistant to chlorhexidine, amine fluoride, amoxycillin, doxy-ycline, and metronidazole than planktonic cells (58, 59). As men-ioned previously, our group found that the viability of susceptible rootanal strains to alkaline stress in planktonic cultures was considerablyncreased when these strains were exposed to the same alkaline stress iniofilms (65). The increased resistance of the strains in biofilms com-
b
x 1000
e
ction. Overview in (a) shows the site of the root canal with a condensation ofation (�1,000) in (b) shows a number of filamentous organisms surroundedt, where the viability of filamentous bacteria could be noticed. Fluorescent greene histological specimen was provided by Dr. Domenico Ricucci and the imagesD, Svensäter G. Viability of bacteria in histological specimens of infected root
ic infeagnificcLighlls. Thcucci
ared to planktonic cultures of the same organism thus raises questions
Persistent Infection in Root Canals 657
Faatw
F
Review Article
6
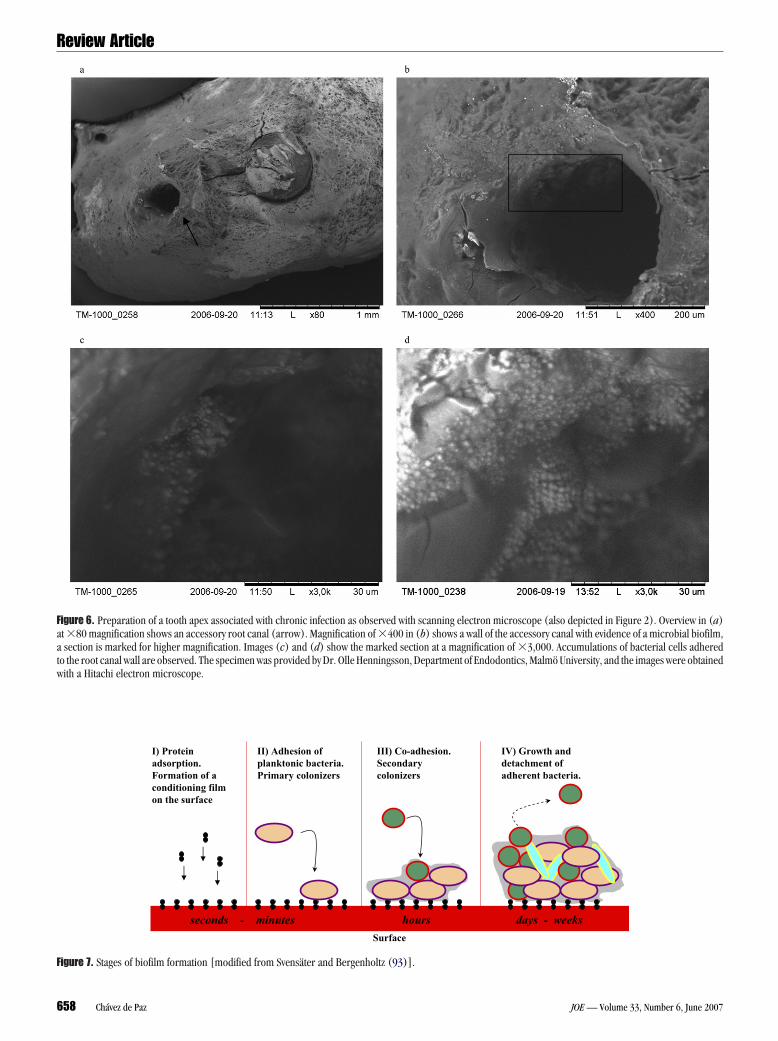
igure 6. Preparation of a tooth apex associated with chronic infection as observed with scanning electron microscope (also depicted in Figure 2). Overview in (a)t �80 magnification shows an accessory root canal (arrow). Magnification of �400 in (b) shows a wall of the accessory canal with evidence of a microbial biofilm,section is marked for higher magnification. Images (c) and (d) show the marked section at a magnification of �3,000. Accumulations of bacterial cells adhered
o the root canal wall are observed. The specimen was provided by Dr. Olle Henningsson, Department of Endodontics, Malmö University, and the images were obtained
ith a Hitachi electron microscope.I) Protein adsorption.Formation of a conditioning film on the surface
II) Adhesion of planktonic bacteria. Primary colonizers
III) Co-adhesion.Secondarycolonizers
IV) Growth and detachment of adherent bacteria.
hours days - weeksseconds - minutes
Surface
igure 7. Stages of biofilm formation [modified from Svensäter and Bergenholtz (93)].
58 Chávez de Paz JOE — Volume 33, Number 6, June 2007

al
tpdhtit“apc
M
gmllppdevosescwwnganW
ntmfspCrst
P
rsoacftrppteapctcm
F
opo
Fgci� z de P
Review Article
J
s to the validation of studies using exclusively liquid-grown cultures inaboratory tests.
During the various stages of biofilm development, as well ashroughout the various sections of the biofilm, cells are in differenthysiological states. Cells at the base of the film, for example, may beead or lysing, whereas those near the surface may be actively growing;owever, even with such physiological diversity, it can be argued thathe majority of time cells in biofilms are in a status equivalent to cellsn the stationary phase of growth (72, 73, 102). From the perspective ofhe persisting root canal flora, however, one might imagine that suchstationary-phase” cells might maintain a low but sufficient metabolicctivity to provoke periapical inflammation. Thus, from the metabolicerspective, they will not be “dead” but would, theoretically, be able toontribute to the persistence of inflammation.
orphological ChangesAdaptation of cells to environmental shift in some cases may trig-
er phenotypic changes in cell morphology. Figure 8 illustrates theorphologic changes undertaken by a root canal strain of Lactobacil-
us paracasei grown in biofilms exposed to a commercial rinsing so-ution of sodium fluoride at a concentration of 0.2%. Column A showshase contrast and Live/Dead images of the 1-day-old biofilms cells of L.aracasei growing in optimal conditions in peptone yeast glucose me-ia (PYG) at 37°C. In Column B, images of biofilms of L. paracasei,xposed to a 0.2% sodium fluoride solution for 1 day and then reacti-ated in PYG at 37°C for 1 day, show changes in the cellular morphologyf L. paracasei. Similar changes were observed with the root canaltrain of Streptococcus anginosus. The explanation for this phenom-non is scant; however, such changes are consistent with stress re-ponses triggering changes while the cells are dividing. For example,olonies of an antibiotic-resistant strain of Pseudomonas aeruginosaere found to have different morphologies when exposed to kanamycinith the agent resulting in the formation of smaller and rougher colo-ies than wild-type cells. However, when this P. aeruginosa strain wasrown on antibiotic-free media for 5 days, colonies appeared smoothnd large just as their wild-type. The authors suggested that these phe-otypic variations observed in the resistant variants were transient (82).
igure 8. One-day-old biofilms of Lactobacillus paracasei isolated from an infrowing in optimal conditions, 37°C in the rich media peptone yeast glucoseells of L. paracasei after exposure for 1 day with a commercial sodium flun PYG. As observed, the cellular morphology of the biofilms cells of L. para
m2. The images are material from the manuscript in preparation (Cháve
ith the advent of tools such as confocal laser scanning microscopy, l
OE — Volume 33, Number 6, June 2007
umerous structural proteins have been shown to be involved in bac-erial morphological alterations. Some of these proteins have a good
olecular similarity to tubulin and are thus capable of forming scaf-olding-like filamentous assemblies in bacteria (103, 104). For in-tance, the structural proteins, crescentin and MreB, were recently im-licated as scaffolds responsible for maintaining shape withinaulobacter crescentus (105, 106). In any case, it is clear that envi-onmental stress results in phenotypic changes to bacterial cells anduch modifications constitute important markers of bacterial adapta-ion.
roteomicsA variety of environmental changes can trigger bacterial adaptation
esulting in the coordinated induction of stress proteins or “heat-hock” proteins. Such proteins have been well characterized for diverseral species, such as S. mutans and S. oralis among others, as gener-ted when exposed to both heat and acid stress (83, 107–109). Intra-ellular expression of different proteins was also reported for E.acealis upon alkaline stress (110, 111). These specific bacterial adap-ive pathways also include the expression of cytosolic proteins that areeleased outside the cell as shown by the release of glyceraldehyde-3-hosphate dehydrogenase (GAPDH) from S. gordonii after a change inH from 6.5 to 7.5 (112). Furthermore, in a recent study, we observed
hat selected cytosolic proteins were as well induced and released to thexternal media in cultures of root canal isolates exposed to sublethallkaline stress (65). The proteins Dnak, Hpr, and fructose-1,6-bisphos-hate aldolase (FBA) were released extracellularly at high levels by rootanal strains of S. anginosus, S. oralis, and S. gordonii. Interestingly,hese copious protein excretions were more intense with planktonicultures than with biofilms, and thus such sublethal effects of a stressay be altered when an organism assumes growth in a biofilm.
ormation of SubpopulationsBacterial adaptation also includes the creation of subpopulations
f cells at the population level. This generation of diversity within aopulation is achieved either by genetic pathways involving constitutiver transient mutators and contingency loci, or by modifications of cel-
oot canal associated with persistent symptoms. (a) Shows the biofilm cells). Biofilm cells were stained with Live/Dead BacLight. (b) Shows the biofilmsolution (0.2%) and after 1 day of reactivation in optimal conditions, 37°Cafter stress was reduced, from an original area of 3 �m2 to an area of 1.5
az LE, Hamilton IR, Svensäter G. Dormancy of oral bacteria in biofilms).
ected r(PYG
oridecasei
ular phenotypes (113, 114). Groups of cells have been found to persist
Persistent Infection in Root Canals 659

atrbisdtdawdtotcfcotctscsu
twuwiTsscoitos
awtctfaForf
eB
tMw
Review Article
6
fter exposure to lethal doses of antibiotics and new growing popula-ions appear in the culture (115, 116). These persister cells (1) mayepresent cells in some protected part of their cell cycle, (2) are capa-le of rapid adaptation, (3) are in a dormant state, or (4) are unable tonitiate programmed cell death in response to the stimulus (84). Thus,uch persisters cells represent a recalcitrant subpopulation that will notie and are capable of initiating a new population with normal suscep-ibility once the antibacterial effect has been dissipated (117–119). Toate, these cells have been reported to occur only after the exposure ofbacterial population to high doses of a single antimicrobial agent,hich triggered the appearance of persister cells exhibiting multiplerug resistance (120, 121). The frequency of persister occurrence andhe mechanism(s) involved in their appearance is unclear, althoughne hypothesis with E. coli suggests that persister cells are regulated byhe expression of chromosomal toxin–antitoxin genes (122). In thisase, the operon HipA seems to be responsible for tolerance to cipro-loxacin and mitomycin C in stationary-phase planktonic cells and E.oli biofilms (122). It was also previously proposed that the expressionf toxins drives bacteria reversibly into the slow growing, multiple drug-olerant phenotypes by “shutting down” antibiotic targets (84). In theontext of root canal bacteria, the formation of such persisting popula-ions that are capable of surviving imposed endodontic treatment mea-ures, as the increase of alkaline levels resulting from application ofalcium hydroxide (65), would explain how organisms are able tourvive and remain in the environment until the effects of noxious stim-li have dissipated.
Concluding Remarks and Directions forFuture Research
The survival of bacteria in root canals after treatment is based onhe capability of the individual organisms to adapt to the environmentithin the consortium. Therefore, the study of the adaptive mechanismssed by organisms to survive in such a highly controlled environment,ith limiting nutrients plus the effects of the antibacterial medicaments,
s important to our understanding of persisting root canal infections.he ability of the organisms in such infections to form biofilms can beeen as the most important adaptive mechanism used by bacteria tourvive the environmental changes resulting from the treatment proto-ol. Thus, from the realization that all oral microorganisms are capablef forming a biofilm and that such surface-associated communities existn root canals, it is possible to apply the “biofilm concept” to clinicalreatment; that is, efforts should not be directed to specific individualrganisms, but to a group of well-adapted organisms undoubtedly pos-essing increased resistance to a variety of antimicrobial agents.
We should obtain much better understanding of the characteristicsnd properties of bacterial biofilms in root canals and the degree tohich such microbial communities enhance survival from environmen-
al changes. A glance at how a biofilm consortium of root canal strainsope with alkaline stress is shown in Chávez de Paz (123). Prospec-ively, the application of molecular biology will be invaluable to identi-ying, for instance, biofilm-expressed genes by root canal colonizers,nd permit the identification of genes involved in adaptation signaling.rom this knowledge we should come to identify key adaptive eventsccurring in the root canal microflora while organisms are exposed tooot canal treatment measures. This path of research should aid ininding mechanisms that can block such processes.
AcknowledgmentsThe work was supported by a research grant from the Knowl-
dge Foundation (KK-stiftelsen, Biofilms—Research Center for
iointerfaces). The author expresses gratitude to Dr. Ian R. Hamil-60 Chávez de Paz
on, Professor Emeritus, Department of Oral Biology, University ofanitoba, Canada, who contributed exceptional guidance and helpith editing throughout this review.
References1. Loesche WJ. Chemotherapy of dental plaque infections. Oral Sci Rev 1976;
9:65–107.2. Bowden GH. Microbiology of root surface caries in humans. J Dent Res
1990;69:1205–10.3. Loesche WJ. Role of Streptococcus mutans in human dental decay. Microbiol Rev
1986;50:353– 80.4. Marsh PD. Microbial community aspects of dental plaque. In: Newman HN, Wilson
M, eds. Dental plaque revisited. Cardiff, UK: BioLine; 1999:237–53.5. Moore WE, Moore LV. The bacteria of periodontal diseases. Periodontology
1994;5:66 –77.6. Socransky SS, Haffajee AD, Cugini MA, Smith C, Kent RL Jr. Microbial complexes in
subgingival plaque. J Clin Periodontol 1998;25:134 – 44.7. Marsh PD. Are dental diseases examples of ecological catastrophes? Microbiology
2003;149:279 –94.8. Bowden GH, Hardie JM, McKee AS, Marsh PD, Fillery ED, Slack GL. The micro-
flora associated with developing carious lesions of the distal surfaces of theupper first premolars in 13–14 year old children. In: Stiles HM, Loesche WJ,O’Brien TC, eds. Microbial Aspects of Dental Caries. Washington, DC: InformationRetrieval; 1976:233– 41.
9. Marsh PD. Sugar, fluoride, pH and microbial homeostasis in dental plaque. ProcFinn Dent Soc 1991;87:515–25.
10. Marsh PD. Microbial ecology of dental plaque and its significance in health anddisease. Adv Dent Res 1994;8:263–71.
11. Marsh PD, Bradshaw DJ. Physiological approaches to the control of oral biofilms.Adv Dent Res 1997;11:176 – 85.
12. Marsh PD, Featherstone A, McKee AS, et al. A microbiological study of early cariesof approximal surfaces in schoolchildren. J Dent Res 1989;68:1151– 4.
13. Newman HN. Plaque and chronic inflammatory periodontal disease. A question ofecology. J Clin Periodontol 1990;17:533– 41.
14. Chávez de Paz Villanueva LE. Fusobacterium nucleatum in endodontic flare-ups.Oral Surg Oral Med Oral Pathol Oral Radiol Endod 2002;93:179 – 83.
15. Haapasalo M, Ranta H, Ranta K, Shah H. Black-pigmented Bacteroides spp. inhuman apical periodontitis. Infect Immun 1986;53:149 –53.
16. Dahlén G, Samuelsson W, Molander A, Reit C. Identification and antimicrobialsusceptibility of enterococci isolated from the root canal. Oral Microbiol Immunol2000;15:309 –12.
17. Eddy RS, Joyce AP, Roberts S, Buxton TB, Liewehr F. An in vitro evaluation of theantibacterial efficacy of chlorine dioxide on E. faecalis in bovine incisors. J Endod2005;31:672–5.
18. Portenier I, Waltimo T, Ørstavik D, Haapasalo M. The susceptibility of starved,stationary phase, and growing cells of Enterococcus faecalis to endodontic medi-caments. J Endod 2005;31:380 – 6.
19. Siren EK, Haapasalo MP, Waltimo TM, Orstavik D. In vitro antibacterial effect ofcalcium hydroxide combined with chlorhexidine or iodine potassium iodide onEnterococcus faecalis. Eur J Oral Sci 2004;112:326 –31.
20. Kassen R, Rainey PB. The ecology and genetics of microbial diversity. Annu RevMicrobiol 2004;58:207–31.
21. Sundqvist G. Ecology of the root canal flora. J Endod 1992;18:427–30.22. Casadevall A, Pirofski LA. Host-pathogen interactions: redefining the basic concepts
of virulence and pathogenicity. Infect Immun 1999;67:3703–13.23. Curtis MA, Slaney JM, Aduse-Opoku J. Critical pathways in microbial virulence.
J Clin Periodontol 2005;32 Suppl 6:28 –38.24. Kaufmann SH, Schaible UE. 100th Anniversary of Robert Koch’s Nobel Prize for the
discovery of the tubercle bacillus. Trends Microbiol 2005;13:469 –75.25. Isenberg HD. Pathogenicity and virulence: another view. Clin Microbiol Rev
1988;1:40 –53.26. Carapetis JR, Steer AC, Mulholland EK, Weber M. The global burden of group A
streptococcal diseases. Lancet Infect Dis 2005;5:685–94.27. Wirth T, Falush D, Lan R, et al. Sex and virulence in Escherichia coli: an evolutionary
perspective. Mol Microbiol 2006;60:1136 –51.28. Dussurget O, Pizarro-Cerda J, Cossart P. Molecular determinants of Listeria mono-
cytogenes virulence. Annu Rev Microbiol 2004;58:587– 610.29. Roberts AJ, Wiedmann M. Pathogen, host and environmental factors contributing to
the pathogenesis of listeriosis. Cell Mol Life Sci 2003;60:904 –18.30. Vazquez-Boland JA, Kuhn M, Berche P, et al. Listeria pathogenesis and molecular
virulence determinants. Clin Microbiol Rev 2001;14:584 – 640.31. Gray MJ, Freitag NE, Boor KJ. How the bacterial pathogen Listeria monocytogenes
mediates the switch from environmental Dr. Jekyll to pathogenic Mr. Hyde. Infect
Immun 2006;74:2505–12.JOE — Volume 33, Number 6, June 2007

Review Article
J
32. Hallet B. Playing Dr. Jekyll and Mr. Hyde: combined mechanisms of phase variationin bacteria. Curr Opin Microbiol 2001;4:570 – 81.
33. Raskin DM, Seshadri R, Pukatzki SU, Mekalanos JJ. Bacterial genomics and patho-gen evolution. Cell 2006;124:703–14.
34. Hamilton IR, Bowden GH. Oral microbiology. In: Alexander M, Bloom BR, Hop-wood DA, et al., eds. Encyclopedia of microbiology, 2nd ed. San Diego, CA: Aca-demic Press, 2000:466 – 81.
35. Kleinberg I. A mixed-bacteria ecological approach to understanding the role of theoral bacteria in dental caries causation: an alternative to Streptococcus mutansand the specific-plaque hypothesis. Crit Rev Oral Biol Med 2002;13:108 –25.
36. Socransky SS. Microbiology of periodontal disease—present status and future con-siderations. J Periodontol 1977;48:497–504.
37. van Winkelhoff AJ, van Steenbergen TJ, de Graaff J. The role of black-pigmentedBacteroides in human oral infections. J Clin Periodontol 1988;15:145–55.
38. Bowden GH. Which bacteria are cariogenic in humans? In: Johnson NW, ed. Dentalcaries. Cambridge, UK: Cambridge University Press, 1991:266 – 86.
39. Sansone C, Van Houte J, Joshipura K, Kent R, Margolis HC. The association of mutansstreptococci and non-mutans streptococci capable of acidogenesis at a low pH withdental caries on enamel and root surfaces. J Dent Res 1993;72:508 –16.
40. van Houte J, Lopman J, Kent R. The final pH of bacteria comprising the predominantflora on sound and carious human root and enamel surfaces. J Dent Res1996;75:1008 –14.
41. Socransky SS, Hubersak C, Propas D. Induction of periodontal destruction in gno-tobiotic rats by a human oral strain of Actinomyces naeslundii. Arch Oral Biol1970;15:993–5.
42. Sundqvist G, Figdor D. Life as an endodontic pathogen. Ecological differencesbetween the untreated and root-filled root canals. Endodontic Topics 2003;6:3–28.
43. Molander A, Reit C, Dahlén G, Kvist T. Microbiological status of root-filled teeth withapical periodontitis. Int Endod J 1998;31:1–7.
44. Peciuliene V, Balciuniene I, Eriksen HM, Haapasalo M. Isolation of Enterococcusfaecalis in previously root-filled canals in a Lithuanian population. J Endod2000;26:593–5.
45. Gomes BP, Pinheiro ET, Sousa EL, et al. Enterococcus faecalis in dental root canalsdetected by culture and by polymerase chain reaction analysis. Oral Surg Oral MedOral Pathol Oral Radiol Endod 2006;102:247–53.
46. Kaufman B, Spångberg L, Barry J, Fouad AF. Enterococcus spp. in endodonticallytreated teeth with and without periradicular lesions. J Endod 2005;31:851– 6.
47. Sedgley C, Nagel A, Dahlén G, Reit C, Molander A. Real-time quantitative polymerasechain reaction and culture analyses of Enterococcus faecalis in root canals. JEndod 2006;32:173–7.
48. Williams JM, Trope M, Caplan DJ, Shugars DC. Detection and quantitation of E.faecalis by real-time PCR (qPCR), reverse transcription-PCR (RT-PCR), and culti-vation during endodontic treatment. J Endod 2006;32:715–21.
49. Zoletti GO, Siqueira JF Jr, Santos KR. Identification of Enterococcus faecalis inroot-filled teeth with or without periradicular lesions by culture-dependent and-independent approaches. J Endod 2006;32:722– 6.
50. Fabricius L, Dahlén G, Sundqvist G, Happonen RP, Möller ÅJ. Influence of residualbacteria on periapical tissue healing after chemomechanical treatment and rootfilling of experimentally infected monkey teeth. Eur J Oral Sci 2006;114:278 – 85.
51. Rôças IN, Siqueira JF Jr, Aboim MC, Rosado AS. Denaturing gradient gel electro-phoresis analysis of bacterial communities associated with failed endodontic treat-ment. Oral Surg Oral Med Oral Pathol Oral Radiol Endod 2004;98:741–9.
52. Fabricius L, Dahlén G, Hölm SE, Möller ÅJ. Influence of combinations of oralbacteria on periapical tissues of monkeys. Scand J Dent Res 1982;90:200 – 6.
53. Dahlén G, Fabricius L, Hölm SE, Möller ÅJ. Interactions within a collection of eightbacterial strains isolated from a monkey dental root canal. Oral Microbiol Immunol1987;2:164 –70.
54. Costerton JW, Lewandowski Z, DeBeer D, Caldwell D, Korber D, James G. Biofilms,the customized microniche. J Bacteriol 1994;176:2137– 42.
55. Costerton JW, Cheng KJ, Geesey GG, et al. Bacterial biofilms in nature and disease.Annu Rev Microbiol 1987;41:435– 64.
56. Gilbert P, Das J, Foley I. Biofilm susceptibility to antimicrobials. Adv Dent Res1997;11:160 –7.
57. Johnson SA, Goddard PA, Iliffe C, et al. Comparative susceptibility of resident andtransient hand bacteria to para-chloro-meta-xylenol and triclosan. J Appl Microbiol2002;93:336 – 44.
58. Larsen T. Susceptibility of Porphyromonas gingivalis in biofilms to amoxicillin,doxycycline and metronidazole. Oral Microbiol Immunol 2002;17:267–71.
59. Shani S, Friedman M, Steinberg D. The anticariogenic effect of amine fluorides onStreptococcus sobrinus and glucosyltransferase in biofilms. Caries Res 2000;34:260 –7.
60. Barrieshi KM, Walton RE, Johnson WT, Drake DR. Coronal leakage of mixed an-aerobic bacteria after obturation and post space preparation. Oral Surg Oral Med
Oral Pathol Oral Radiol Endod 1997;84:310 – 4.OE — Volume 33, Number 6, June 2007
61. Clegg MS, Vertucci FJ, Walker C, Belanger M, Britto LR. The effect of exposure toirrigant solutions on apical dentin biofilms in vitro. J Endod 2006;32:434 –7.
62. Dunavant TR, Regan JD, Glickman GN, Solomon ES, Honeyman AL. Comparativeevaluation of endodontic irrigants against Enterococcus faecalis biofilms. J Endod2006;32:527–31.
63. George S, Kishen A, Song KP. The role of environmental changes on monospe-cies biofilm formation on root canal wall by Enterococcus faecalis. J Endod2005;31:867–72.
64. Spratt DA, Pratten J, Wilson M, Gulabivala K. An in vitro evaluation of the antimi-crobial efficacy of irrigants on biofilms of root canal isolates. Int Endod J2001;34:300 –7.
65. Chávez de Paz LE, Bergenholtz G, Dahlén G, Svensäter G. Response to alkaline stressby root canal bacteria in biofilms. Int Endod J 2006. Accepted for publication.
66. Breeuwer P, Abee T. Assessment of viability of microorganisms employing fluores-cence techniques. Int J Food Microbiol 2000;55:193–200.
67. Kell DB, Kaprelyants AS, Weichart DH, Harwood CR, Barer MR. Viability and activityin readily culturable bacteria: a review and discussion of the practical issues. An-tonie Van Leeuwenhoek 1998;73:169 – 87.
68. Mushegian AR, Koonin EV. A minimal gene set for cellular life derived by compar-ison of complete bacterial genomes. Proc Natl Acad Sci USA 1996;93:10268 –73.
69. Blackburn CW, McCarthy JD. Modifications to methods for the enumeration anddetection of injured Escherichia coli O157:H7 in foods. Int J Food Microbiol2000;55:285–90.
70. Ward DM, Weller R, Bateson MM. 16S rRNA sequences reveal uncultured inhabit-ants of a well-studied thermal community. FEMS Microbiol Rev 1990;6:105–15.
71. Xu HS, Roberts N, Singleton FL, Attwell RW, Grimes DJ, Colwell RR. Survival andviability of non-culturable Escherichia coli and Vibrio cholerae in the estuarineand marine environment. Microbial Ecol 1982;8:313–23.
72. Nyström T. Not quite dead enough: on bacterial life, culturability, senescence, anddeath. Arch Microbiol 2001;176:159 – 64.
73. Nyström T. Conditional senescence in bacteria: death of the immortals. Mol Micro-biol 2003;48:17–23.
74. Joux F, Lebaron P. Use of fluorescent probes to assess physiological functions ofbacteria at single-cell level. Microbes Infect 2000;2:1523–35.
75. Moter A, Gobel UB. Fluorescence in situ hybridization (FISH) for direct visualizationof microorganisms. J Microbiol Methods 2000;41:85–112.
76. Poulsen LK, Ballard G, Stahl DA. Use of rRNA fluorescence in situ hybridization formeasuring the activity of single cells in young and established biofilms. Appl EnvironMicrobiol 1993;59:1354 – 60.
77. Sunde PT, Olsen I, Gobel UB, et al. Fluorescence in situ hybridization (FISH) fordirect visualization of bacteria in periapical lesions of asymptomatic root-filledteeth. Microbiology 2003;149:1095–102.
78. Boulos L, Prevost M, Barbeau B, Coallier J, Desjardins R. LIVE/DEAD BacLight:application of a new rapid staining method for direct enumeration of viable and totalbacteria in drinking water. J Microbiol Methods 1999;37:77– 86.
79. Leuko S, Legat A, Fendrihan S, Stan-Lotter H. Evaluation of the LIVE/DEAD BacLightkit for detection of extremophilic archaea and visualization of microorganisms inenvironmental hypersaline samples. Appl Environ Microbiol 2004;70:6884 – 6.
80. Khemaleelakul S, Baumgartner JC, Pruksakom S. Autoaggregation and coaggrega-tion of bacteria associated with acute endodontic infections. J Endod 2006;32:312– 8.
81. Creach V, Baudoux AC, Bertru G, Rouzic BL. Direct estimate of active bacteria: CTCuse and limitations. J Microbiol Methods 2003;52:19 –28.
82. Drenkard E, Ausubel FM. Pseudomonas biofilm formation and antibiotic resistanceare linked to phenotypic variation. Nature 2002;416:740 –3.
83. Svensäter G, Sjögren B, Hamilton IR. Multiple stress responses in Streptococcusmutans and the induction of general and stress-specific proteins. Microbiology2000;146:107–17.
84. Lewis K. Persister cells and the riddle of biofilm survival. Biochemistry (Mosc)2005;70:267–74.
85. Sedgley CM, Clewell DB. Bacterial plasmids in the oral and endodontic microflora.Endod Topics 2004;9:37–51.
86. Bowden GH, Hamilton IR. Survival of oral bacteria. Crit Rev Oral Biol Med1998;9:54 – 85.
87. Presterl E, Grisold AJ, Reichmann S, Hirschl AM, Georgopoulos A, Graninger W.Viridans streptococci in endocarditis and neutropenic sepsis: biofilm formationand effects of antibiotics. J Antimicrob Chemother 2005;55:45–50.
88. Scheld MW, Sande MA. Endocarditis and intravascular infections. In: Mandell GL,Douglas RG, Dolin R, eds. Principles and practice of infectious diseases, 4th ed. NewYork: Churchill Livingstone, 1995:740 – 83.
89. Bowden GH, Ellwood DC, Hamilton IR. Microbial ecology of the oral cavity. In:Alexander M, ed. Advances in microbial ecology. New York: Plenum. 1979:
135–217.Persistent Infection in Root Canals 661

1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
Review Article
6
90. Brecx M, Theilade J, Attstrom R. An ultrastructural quantitative study of the signif-icance of microbial multiplication during early dental plaque growth. J PeriodontalRes 1983;18:177– 86.
91. Gilbert P, Evans DJ, Brown MR. Formation and dispersal of bacterial biofilms in vivoand in situ. J Appl Bacteriol 1993;74 Suppl:67S–78S.
92. Lee SF, Li YH, Bowden GH. Detachment of Streptococcus mutans biofilm cells byan endogenous enzymatic activity. Infect Immun 1996;64:1035– 8.
93. Svensäter G, Bergenholtz G. Biofilms in endodontic infections. Endod Topics2004;9:27–36.
94. Nair PN, Henry S, Cano V, Vera J. Microbial status of apical root canal system ofhuman mandibular first molars with primary apical periodontitis after “one-visit”endodontic treatment. Oral Surg Oral Med Oral Pathol Oral Radiol Endod2005;99:231–52.
95. Nair P. Light and electron microscopic studies on root canal flora and periapicallesions. J Endod 1987;13:29 –39.
96. Mölven O, Olsen I, Kerekes K. Scanning electron microscopy of bacteria in theapical part of root canals in permanent teeth with periapical lesions. Endod DentTraumatol 1991;7:226 –9.
97. Sen BH, Piskin B, Demirci T. Observation of bacteria and fungi in infected rootcanals and dentinal tubules by SEM. Endod Dent Traumatol 1995;11:6 –9.
98. Brown MR, Gilbert P. Sensitivity of biofilms to antimicrobial agents. J Appl Bacteriol1993;74 Suppl:87S–97S.
99. Branda SS, Vik S, Friedman L, Kolter R. Biofilms: the matrix revisited. TrendsMicrobiol 2005;13:20 – 6.
00. Fletcher M. The physiological activity of bacteria attached to solid surfaces. AdvMicrobiol Physiol 1991;32:53– 85.
01. Goodman AE, Marshall KC. Genetic responses of bacteria at surfaces. In: CostertonJW, Lappin-Scott HM, eds. Microbial biofilms. Cambridge, UK: CambridgeUniversityPress, 1995:80 –95.
02. Wimpenny JW, Leistner L, Thomas LV, Mitchell AJ, Katsaras K, Peetz P. Submergedbacterial colonies within food and model systems: their growth, distribution andinteractions. Int J Food Microbiol 1995;28:299 –315.
03. Beveridge TJ. Understanding the shapes of bacteria just got more complicated. MolMicrobiol 2006;62:1– 4.
04. Weiss DS. Bacterial cell division and the septal ring. Mol Microbiol 2004;54:588 –97.
05. Ausmees N, Kuhn JR, Jacobs-Wagner C. The bacterial cytoskeleton: an intermediatefilament-like function in cell shape. Cell 2003;115:705–13.
06. Dye NA, Pincus Z, Theriot JA, Shapiro L, Gitai Z. Two independent spiral structures
control cell shape in Caulobacter. Proc Natl Acad Sci USA 2005;102:18608 –13.62 Chávez de Paz
07. Hamilton IR, Svensäter G. Acid-regulated proteins induced by Streptococcus mu-tans and other oral bacteria during acid shock. Oral Microbiol Immunol1998;13:292–300.
08. Welin J, Wilkins JC, Beighton D, et al. Effect of acid shock on protein expression bybiofilm cells of Streptococcus mutans. FEMS Microbiol Lett 2003;227:287–93.
09. Wilkins JC, Beighton D, Homer KA. Effect of acidic pH on expression of surface-associated proteins of Streptococcus oralis. Appl Environ Microbiol 2003;69:5290 – 6.
10. Flahaut S, Hartke A, Giard JC, Auffray Y. Alkaline stress response in Enterococcusfaecalis: adaptation, cross-protection, and changes in protein synthesis. Appl En-viron Microbiol 1997;63:812– 4.
11. Giard JC, Laplace JM, Rince A, et al. The stress proteome of Enterococcus faecalis.Electrophoresis 2001;22:2947–54.
12. Nelson D, Goldstein JM, Boatright K, et al. pH-regulated secretion of a glyceralde-hyde-3-phosphate dehydrogenase from Streptococcus gordonii FSS2: purification,characterization, and cloning of the gene encoding this enzyme. J Dent Res2001;80:371–7.
13. Foster PL. Stress responses and genetic variation in bacteria. Mutat Res2005;569:3–11.
14. Kussell E, Leibler S. Phenotypic diversity, population growth, and information influctuating environments. Science 2005;309:2075– 8.
15. Lewis K. Riddle of biofilm resistance. Antimicrob Agents Chemother 2001;45:999 –1007.
16. Moyed HS, Bertrand KP. hipA, a newly recognized gene of Escherichia coli K-12 thataffects frequency of persistence after inhibition of murein synthesis. J Bacteriol1983;155:768 –75.
17. Balaban NQ, Merrin J, Chait R, Kowalik L, Leibler S. Bacterial persistence as aphenotypic switch. Science 2004;305:1622–5.
18. Harrison JJ, Turner RJ, Ceri H. Persister cells, the biofilm matrix and tolerance tometal cations in biofilm and planktonic Pseudomonas aeruginosa. Environ Micro-biol 2005;7:981–94.
19. Keren I, Kaldalu N, Spoering A, Wang Y, Lewis K. Persister cells and tolerance toantimicrobials. FEMS Microbiol Lett 2004;230:13– 8.
20. Roberts ME, Stewart PS. Modelling protection from antimicrobial agents in biofilmsthrough the formation of persister cells. Microbiology 2005;151:75– 80.
21. Stewart PS. Mechanisms of antibiotic resistance in bacterial biofilms. Int J MedMicrobiol 2002;292:107–13.
22. Keren I, Shah D, Spoering A, Kaldalu N, Lewis K. Specialized persister cells and themechanism of multidrug tolerance in Escherichia coli. J Bacteriol 2004;186:8172– 80.
23. Chávez de Paz LE. On bacteria persisting root canal treatment [PhD thesis]. Goth-
enburg, Sweden: Gothenburg University, 2005.JOE — Volume 33, Number 6, June 2007
Copyright © 2022 FDOKUMEN




















