
199 The New Construction Approach Adapted by Ashghal for ...

Ralstonia metallidurans, a bacterium speci¢cally adapted totoxic metals: towards a catalogue of metal-responsive genes
Max Mergeay a;b;�, Se¤bastien Monchy a;c, Tatiana Vallaeys a;d, Vanessa Auquier e,Abderra¢ Benotmane a, Philippe Bertin a, Sa¢yh Taghavi b;f , John Dunn f,
Daniel van der Lelie b;f , Ruddy Wattiez g
a Laboratories for Microbiology and Radiobiology, Belgium Nuclear Research Center, SCK/CEN, B-2400 Mol, Belgiumb Environmental Technology Expertise Center, Flemish Institute for Technological Research, Vito, B-2400 Mol, Belgiumc Unite¤ d’e¤tude de la Conformation des Macromole¤cules Biologiques, Universite¤ Libre de Bruxelles, Brussels, Belgiumd De¤partement de Microbiologie, Institut National de la Recherche Agronomique (INRA), Jouy-en-Josas, France
e Centre de Biologie structurale et de Bioinformatique: Structure et Fonction des Membranes Biologiques, Universite¤ Libre de Bruxelles, Brussels, Belgiumf Biology Department, Bldg. 463, Brookhaven National Laboratories (BNL), Upton, NY 11973-5000, USA
g Service de Chimie Biologique, Universite¤ de Mons-Hainaut, Mons, Belgium
Received 18 December 2002; received in revised form 9 February 2003; accepted 21 February 2003
First published online 9 May 2003
Abstract
Ralstonia metallidurans, formerly known as Alcaligenes eutrophus and thereafter as Ralstonia eutropha, is a L-Proteobacteriumcolonizing industrial sediments, soils or wastes with a high content of heavy metals. The type strain CH34 carries two large plasmids(pMOL28 and pMOL30) bearing a variety of genes for metal resistance. A chronological overview describes the progress made in theknowledge of the plasmid-borne metal resistance mechanisms, the genetics of R. metallidurans CH34 and its taxonomy, and theapplications of this strain in the fields of environmental remediation and microbial ecology. Recently, the sequence draft of the genome ofR. metallidurans has become available. This allowed a comparison of these preliminary data with the published genome data of the plantpathogen Ralstonia solanacearum, which harbors a megaplasmid (of 2.1 Mb) carrying some metal resistance genes that are similar to thosefound in R. metallidurans CH34. In addition, a first inventory of metal resistance genes and operons across these two organisms could bemade. This inventory, which partly relied on the use of proteomic approaches, revealed the presence of numerous loci not only on thelarge plasmids pMOL28 and pMOL30 but also on the chromosome. It suggests that metal-resistant Ralstonia, through evolution, areparticularly well adapted to the harsh environments typically created by extreme anthropogenic situations or biotopes.; 2003 Federation of European Microbiological Societies. Published by Elsevier Science B.V. All rights reserved.
Keywords: Ralstonia metallidurans ; Heavy metal ; Resistance gene; P1-ATPase; Genome; Proteomics ; HME-RND e¥ux system; Plasmid; Mobile geneticelement ; Regulation
Contents
1. Introduction: Ralstonia metallidurans : general description and historical overview . . . . . . . 3861.1. Strain CH34 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3861.2. Plasmids . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3861.3. Overview of metal resistance research . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3871.4. Microbial ecology and environmental biotechnology . . . . . . . . . . . . . . . . . . . . . . . . . 3871.5. Genetic tools and features . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3871.6. From the genome of R. metallidurans towards an inventory of metal-relevant
functions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 387
0168-6445 / 03 / $22.00 ; 2003 Federation of European Microbiological Societies. Published by Elsevier Science B.V. All rights reserved.doi :10.1016/S0168-6445(03)00045-7
* Corresponding author. Tel. : +32 (14) 332727; Fax: +32 (14) 314793. E-mail address: [email protected] (M. Mergeay).
FEMSRE 778 2-6-03 Cyaan Magenta Geel Zwart
FEMS Microbiology Reviews 27 (2003) 385^410
www.fems-microbiology.org

2. Taxonomy of the genus Ralstonia with emphasis on the relationships between metal-resistantspecies and phytopathogenic species . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 388
3. Genomics of the genus Ralstonia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3893.1. Main features of the genome of R. solanacearum and R. metallidurans . . . . . . . . . . . . 3893.2. Inventory of metal resistance genes or gene equivalents in R. solanacearum and
R. metallidurans . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3894. HME-RND gene clusters in R. metallidurans . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3905. The P1-ATPases . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 392
5.1. Prediction of structural characteristics of R. metallidurans P1-ATPases . . . . . . . . . . . . 3935.2. Phylogeny . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3945.3. Metal ¢xation motifs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 395
6. CDF, CHR and ars-like genes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3957. Large plasmids and mobile genetic elements of R. metallidurans : content in heavy metal
resistance genes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3968. Proteomic approaches in R. metallidurans . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3979. Plasmid-borne copper resistance genes in R. metallidurans . . . . . . . . . . . . . . . . . . . . . . . . 39810. Plasmid-borne genes for resistance to lead in R. metallidurans . . . . . . . . . . . . . . . . . . . . . 40011. Regulatory genes in the heavy metal resistance gene clusters . . . . . . . . . . . . . . . . . . . . . . . 401
11.1. The R and S two-component family . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 40111.2. The merR family . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 40211.3. The arsR family . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 40211.4. The cnrYXH family . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 40411.5. Other regulatory genes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 405
12. Concluding remarks . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 405
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 406
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 406
1. Introduction: Ralstonia metallidurans :general description and historicaloverview
1.1. Strain CH34
Metal-resistant bacteria [1], formerly known as Alcali-genes eutrophus [2], then Ralstonia sp., Ralstonia eutrophaor R. eutropha-like and ¢nally Ralstonia metallidurans [3],carry plasmid-borne czc (resistance to cadmium, zinc andcobalt) [2,4], ncc (nickel, cobalt and cadmium) [5] or cnr(cobalt and nickel) [6] cation e¥ux metal resistance oper-ons. Members of this genus are mostly found in industrialand polluted biotopes, prevailing in metallurgic wastes.The ¢rst strain of this group (and still the reference strain),R. metallidurans CH34, was found in the late 1970s insediments of a decantation basin of a zinc factory in Bel-gium, near Lie'ge [1]. The strain was found to be highlyresistant to Zn(II), Cd(II), and Co(II) and later to Ni(II),Cu(II), CrO234 , Hg(II) and Pb(II). Mutants lacking theresistance to Zn(II), Cd(II) and Co(II) were easily ob-tained among survivors at 37‡C, the maximum tempera-ture where growth could be observed [1,2]. This suggesteda possible curing of plasmid-borne genetic determinantsfor the resistance to these metals. Transfer of metal resis-tance markers to other strains supported this view [7^9].The strain proved also to be accessible to genetic analysisand to act as a good recipient of foreign genes in plasmid-
mediated mobilization of chromosomal genes between dif-ferent bacteria [10]. As a facultative chemolithotrophicbacterium able to grow aerobically in the presence of H2and CO2, strain CH34 was shown to possess two hydro-genases, one soluble and one particle-bound, which at thattime was considered typical for A. eutrophus [2]. The taxo-nomic proximity between metal-resistant strains such asCH34 and the A. eutrophus hydrogenotrophs was furthersupported by a variety of genetic and phenotypic tests.Yet, as discussed below, the taxonomic status of A. eutro-phus and related strains [11,12] was recently revised, andthe bacteria were eventually reclassi¢ed in the genus Ral-stonia [13].
1.2. Plasmids
The plasmid-borne character of the resistance to Cd(II),Zn(II), Co(II) and Ni(II) in strain CH34 was demon-strated via appropriate curing and transfer experiments:resistance to Zn(II), Cd(II) and Co(II) was linked to theplasmid pMOL30 (V250 kb) while resistance to Co(II)and Ni(II) as well as resistance to chromate was linkedto plasmid pMOL28 (180 kb) [14]. In addition, both plas-mids were shown to encode resistance to Tl(I) and Hg(II),the latter being encoded by the two mercury transposonsTn4378 (pMOL28) and Tn4380 (pMOL30) [15]. The Czc3
mutants that were obtained among the survivors at 37‡Cwere a ¢rst indication of the mutator phenotype that was
FEMSRE 778 2-6-03 Cyaan Magenta Geel Zwart
M. Mergeay et al. / FEMS Microbiology Reviews 27 (2003) 385^410386

further described in the strain CH34 and other R. metal-lidurans strains [14,16,17].
1.3. Overview of metal resistance research
As far as the resistance to heavy metals is concerned, the¢rst studies focused on the czc locus (resistance to Cd(II),Zn(II) and Co(II)) located on pMOL30 [4,18^21]), to themercury transposons Tn4378 (pMOL28) and Tn4380(pMOL30) [15], to the resistance to Ni(II), and then tothe corresponding cnr locus on pMOL28 [6,22^24]. Resis-tance to chromate was also studied early on [25,26]. In therelated strain R. metallidurans 31A (formerly known asAlcaligenes xylosoxidans 31A or sometimes as Achromo-bacter xylosoxidans 31A) [3,12], ncc-mediated resistanceto Ni(II) was found to be very high with a minimum in-hibitory concentration (MIC) value of 40 mM (the MICvalue for Ni(II) conferred by the cnr genetic determinantin strain CH34 is not higher than 2.5 mM) [5]. Of all themetals for which plasmid-borne resistance of heterotrophicbacteria has been reported, such a high level resistance wasonly observed for Ni(II) [27]. A similar high level of resis-tance to Ni(II) has been described in the strain Arthro-bacter ilicis that was isolated in nickel-rich sediments ofa metallurgical factory in Tyrol, Austria [28].Other plasmid-borne determinants of R. metallidurans
were described much later, including the pbr locus (resis-tance to lead) [29] and the cop locus (resistance to copper)[30], both of which are located on pMOL30. In the sameperiod of the late 1990s, fundamental studies on theczc operon [31^33], the cnrYXH regulatory region ofpMOL28 [34,35], the chr chromate resistance operons(located chromosomally and on pMOL28) [36], and thenickel resistance determinant nreB of R. metallidurans31A (the third route to resistance to Ni(II) besides cnrand ncc) [5,37,38] were undertaken. Although not directlylinked to metal resistance as such, various additional re-ports on siderophores and the oxidative stress responsepoint to other aspects of metal metabolism in R. metalli-durans [39^45]
1.4. Microbial ecology and environmental biotechnology
The knowledge about the mechanisms of resistance toheavy metals and especially their regulation stimulatedfurther work in microbial ecology [46^51] and in environ-mental biotechnology [52]. Industrial applications of met-al-resistant bacteria and their genes include the use ofbiosensors for monitoring the concentration of heavy met-als in a variety of substrates and soils [53^58], the develop-ment of bioreactors to remove heavy metals from pollutede¥uents or soils [59,60], and bioaugmentation [61] andphytoremediation [62] (Y. Wang, W. Hai, J. Vander Ley-den and M. Mergeay, unpublished). A recombinant strainof R. metallidurans CH34, expressing a metallothioneingene, was successfully used to immobilize metals in soil,
allowing revegetation [63,64]. The ncc-nre nickel resistancesystem of R. metallidurans 31A was e⁄ciently expressed inthe endophytic bacteria Burkholderia cepacia L.S.2.4 andHerbaspirillum seropedicae LMG2284. The heterologousexpression of ncc-nre-encoded nickel resistance was ac-companied by nickel removal from the culture medium.This capacity to remove nickel through sequestration orbioprecipitation processes and consequently lowering thefree nickel concentration o¡ers interesting bene¢ts forthese endophytic bacteria and their host plants. Lupinusluteus L, when grown on a nickel-enriched substrate andinoculated with B. cepacia L.S.2.4 : :ncc-nre, showed a sig-ni¢cant increase (30%) of the nickel concentration in theroots, whereas the nickel concentration in the shoots re-mained comparable with that of the control plants [62].
1.5. Genetic tools and features
Conjugation experiments, the isolation and character-ization of R-prime plasmids [10,65], the construction ofa cosmid library [39], and the development of electropora-tion [66] allowed the circularity of the chromosome to beestablished and various loci to be mapped [10,16,65,67,68].Genetic and physical maps of the pMOL28 plasmid andits conjugation-pro¢cient derivative pMOL50 were pro-duced [14]. The formation of R-prime plasmids by retro-transfer and the capture of small plasmids was also ob-served and studied in these conjugation studies [69,70].Finally, several mobile genetic elements present in theCH34 genome were trapped on appropriate positive selec-tion vectors and characterized [34,71,72] (M. Hassan, per-sonal communication).
1.6. From the genome of R. metallidurans towards aninventory of metal-relevant functions
Recently, the sequencing of the genome of R. metalli-durans CH34 was carried out by the Joint Genome Insti-tute in California (http://www.jgi.doe.gov/JGI_microbial/html/ralstonia/ralston_homepage.html) and a ¢rst draftof the sequence was released in 2000. The available se-quence still contains some gaps, some of them being closedthanks to the joint e¡orts of our laboratories (ftp://ftp.genome.bnl.gov/pub/Sequences/Rmetallidurans/). Yet it al-ready allows making a ¢rst inventory of functions relatedto the resistance to heavy metals as well as to the adapta-tion to harsh biotopes.Previous reviews have addressed the plasmid-borne re-
sistance to heavy metals as well as the biology and theenvironmental applications of R. metallidurans [17,52,73^83]. In the present review, we want to take the opportunityto combine the availability of genome sequence data andthe ¢rst results of proteomic approaches to generate aninventory of the genes and protein functions involved inbacterial adaptation and resistance to heavy metals.Although we already had a fairly good view of the plas-
FEMSRE 778 2-6-03 Cyaan Magenta Geel Zwart
M. Mergeay et al. / FEMS Microbiology Reviews 27 (2003) 385^410 387

mid-borne functions, chromosomal genes look far morediversi¢ed and abundant than ¢rst expected.This tentative inventory is preceded by an update on the
taxonomic and genomic situation of R. metallidurans withspecial reference to other well-known species of the genusRalstonia : the hydrogenotroph R. eutropha and the plantpathogen R. solanacearum. Thereafter the followingthemes will be addressed in the inventory:b the major families of metal resistance mechanisms: theRND (resistance^nodulation^cell division) [84^86] andCDF (cation di¡usion facilitation) detoxi¢cation sys-tems, the e¥ux P1-ATPases, and the systems focusingmore speci¢cally on mercury, copper, chromate, or ar-senic/antimony oxyanions
b the pMOL28- and pMOL30-borne genes involved inmetal resistance with special emphasis on the resistanceto copper and to lead
b ¢nally, a special section will be devoted to the regula-tory genes involved in metal resistance.
2. Taxonomy of the genus Ralstonia with emphasis on therelationships between metal-resistant species andphytopathogenic species
The need for a taxonomic reassessment came when it
was clear that A. eutrophus could no longer be clusteredwith Alcaligenes faecalis [13] and had to be transferred tothe genus Ralstonia. The metal-resistant hydrogenotrophssuch as strain CH34 proved to be closely related to clinicalisolates known as ‘CDC-IVc2’ that were later renamedRalstonia paucula [87]. A ¢rst molecular study of metal-resistant strains con¢rmed a narrow clustering of metal-resistant strains isolated from polluted or industrial envi-ronments [12]. Finally, a polyphasic study based on 16srRNA, whole cell protein pro¢ling, fatty acid analysis, andother data made it possible to divide the bacteria carryingczc, ncc or cnr genes over three Ralstonia species: R. me-tallidurans, R. basilensis and R. campinensis [3].Fig. 1 shows a dendrogram of the Ralstonia genus and
the taxonomic relationship between the two species forwhich extensive genomic data are available: R. solanacea-rum [88] and R. metallidurans (a draft of the genome hasbeen released by the Joint Genome Institute). R. solana-cearum is a plant pathogen: most isolates of R. solanacea-rum are specialized in the colonization and the infection ofplants and may be considered to colonize plant-boundniches. The genus Ralstonia is remarkably diverse in termsof ecological niches and specializations or capabilities :plant pathogenesis (R. solanacearum), nodulation and ni-trogen ¢xation in tropical legumes (R. taiwanensis) [89],nosocomial infections (R. pickettii, gilardii, paucula) [87],
Fig. 1. Polyphasic taxonomy of metal-resistant Ralstonia (czc and ncc). A dendrogram of the genus Ralstonia is shown [3,13,87,89] (Goris, personalcommunication). Various species are shown with indication of remarkable or speci¢c functions. Species characteristic of or found in metal-rich industrialbiotopes (R. metallidurans, campinensis and basilensis) are shown in bold and display metal resistance genotypes (czc : resistance to Cd(II), Zn(II) andCo(II) ; ncc : resistance to Ni(II), Co(II) and Cd(II)). czc and ncc genotypes are mostly de¢ned via positive hybridization with czc or ncc probes [11,12].The plant pathogen R. solanacearum is shown in gray bold.
FEMSRE 778 2-6-03 Cyaan Magenta Geel Zwart
M. Mergeay et al. / FEMS Microbiology Reviews 27 (2003) 385^410388

hydrogenotrophy (R. eutropha), colonization of metal-richbiotopes (R. metallidurans, campinensis, basilensis), degra-dation of recalcitrant aromatic compounds and man-madechemicals (R. oxalatica, eutropha, basilensis), and the syn-thesis of polyhydroxyalkanoates (R. eutropha, oxalatica,metallidurans). This diversity of functions, specializations,and range of biotopes in the genus Ralstonia illustrateswhy it was originally assigned to the large bacterial groupof Proteobacteria (in reference to the Greek god Proteus,famous for his diversity of shapes). In the same perspec-tive, natural genetic engineering is clearly at stake to pro-mote the functional diversity in this genus.
3. Genomics of the genus Ralstonia
3.1. Main features of the genome of R. solanacearum andR. metallidurans
The genome of R. solanacearum is made up of two largereplicons: a 3.5-Mb chromosome and a 2.1-Mb megaplas-mid [88]. Pathogenesis is linked with large clusters of genesthat are located on both replicons.It is interesting to compare this situation with the ge-
nome organization of the soil-dwelling hydrogenotrophR. eutropha H16. In this bacterium, pulse ¢eld electropho-resis studies have indeed shown the presence of two majorreplicons of 4.1 Mb and 2.9 Mb, and of a 0.44-Mb mega-plasmid that contains the genetic determinants for hydro-genotrophy and enables anaerobic growth at the expenseof nitrate [90]. These genetic determinants are chro-mosome-encoded in R. metallidurans CH34 as they arepresent in the plasmid-free derivative of R. metalliduransCH34 [2,16]. Reversely, the 0.44-Mb megaplasmid (pHG1)of R. eutropha H16 does not appear to contain any gene-tic determinants for heavy metal resistance genes (E.Schwartz, personal communication). Conjugational map-ping using two di¡erent conjugative plasmids (pULB113,an RP4: :miniMu derivative, and pMOL50, a derivative ofpMOL28) able to mobilize chromosomal determinantsshowed that these determinants are linked to other house-keeping gene determinants on the chromosome [16]. It isnot yet known if the genome of R. metallidurans containsa structure like the megaplasmid of R. solanacearum, ormore generally two major replicons as is the case for bothR. solanacearum G1000 and R. eutropha H16 [88,90].Strain CH34 contains two large plasmids of 180 andV250 kb [14,75] that are especially rich in metal resistancegenes and in open reading frames (ORFs) similar to well-known metal resistance genes. Within the scope of thisreview, we will refer to the chromosome of R. metallidu-rans CH34 for all genes and functions that are not locatedon the plasmids pMOL28 and pMOL30, and this withoutprejudice of the possible presence of two major repliconsas in R. solanacearum or R. eutropha.The genomes of R. solanacearum and R. metallidurans
appear to share a large number of orthologous sequences(Table 1). In addition, the major replicons of R. solana-cearum contain a number of genes that display substantialsequence similarity with genes of other bacteria (Table 1):the 3.5-Mb chromosome of R. solanacearum contains upto 65% of genes displaying good identity with those ofR. metallidurans while this percentage drops to 30% inthe megaplasmid. These ¢gures are much higher than the¢gures obtained in the comparison of R. solanacearumwith the available genomes of other L- and Q-Proteobac-teria (Table 1).
3.2. Inventory of metal resistance genes or gene equivalentsin R. solanacearum and R. metallidurans
The genome of R. solanacearum has been extensivelytested for the presence of equivalents of known metal re-sistance genes and especially those represented in the ge-nome of R. metallidurans. This search was carried out bylooking at ars- (arsenite, arsenate), chr- (chromate), cop-(Cu(II)), czc- (Cd(II),Zn(II),Co(II)), cnr- (Co(II),Ni(II)),mer- (Hg(II) and organomercurials), pbr- (Pb(II)), andsil- (Ag(I)) like genes as well as at genes encoding P-ATP-ases, although it is not always easy to de¢ne those in-volved in metal resistance. The main observation is thatmost of the putative metal resistance genes are linked tothe megaplasmid of R. solanacearum as reported in Fig. 2.On the chromosome, there are two loci governed by amerR-like regulator: only one locus has recognizablestructural genes for metal transport (P-ATPase); thereare also ¢ve arsR/cadC paralogues but none of them canbe associated with a metal resistance function. The chro-mosome contains an arsR-arsC (ArsC stands for reductaseof arsenate to arsenite) cluster (as the megaplasmid) but atpresent there is no indication for a possible relationshipwith resistance to arsenicals.Fig. 2 reports the localization of the metal resistance
genes and of the IS elements on the R. solanacearum mega-plasmid. This localization may give some important clueson the genetic mobility of metal resistance genes and theirpossible involvement in horizontal gene transfer events.The map of the R. solanacearum megaplasmid shows the
presence of many clusters of putative metal resistance
Table 1Distribution of R. solanacearum proteins conserved in other organisms
Chromosome Megaplasmid
Ralstonia metallidurans 2148 661Neisseria meningitidis 856 124Xylella fastidiosa 792 137Pseudomonas aeruginosa 1558 588Escherichia coli 1133 322Unknown proteins 315 433
The number of orthologous genes in various organisms was determinedfor each R. solanacearum protein having 35% or more sequence identityover at least 80% of its total length and an expected value of BLASTPbelow 1036.
FEMSRE 778 2-6-03 Cyaan Magenta Geel Zwart
M. Mergeay et al. / FEMS Microbiology Reviews 27 (2003) 385^410 389

genes that are ordered as they are observed on chromo-somal contigs of R. metallidurans and much less as theyare observed on the large plasmids pMOL28 andpMOL30. Fig. 2 indicates the clusters of metal resistancegenes of R. solanacearum that have an equivalent with thesame putative gene order in R. metallidurans. As an exam-ple, the order of the cop genes is similar to that observedin R. metallidurans (see below).There are also four clusters with czc-like genes (HME-
RND e¥ux according to the nomenclature proposed inanother paper in this issue [84] : RND stands for resis-tance^nodulation^cell division [86] and HME for the classof RND that is specialized in e¥ux of heavy metal cations[91]) with four paralogues for czcA/silA. The number ofputative HME-RND genes in R. solanacearum is higherthan for B. cepacia, Pseudomonas aeruginosa or Escheri-
chia coli but much less than was observed in R. metallidu-rans as shown in the next section.
4. HME-RND gene clusters in R. metallidurans
At least 12 (putative) genes encoding a HME-RND [86]e¥ux system (CzcA orthologues and paralogues) arepresent on the genome of R. metallidurans : one inpMOL28 (cnrA), three on pMOL30, and eight on thechromosome (Fig. 3). This high number of (putative)CzcA paralogues and of corresponding gene clusters oroperons supports the specialization of R. metallidurans inthe adaptation to high concentrations of heavy metals,although it is not yet possible to assign functions or metalspeci¢cities and concentration ranges for most of them.
Fig. 2. Circular representation of the R. solanacearum megaplasmid. The outer circle scale is marked with bases and IS distribution [88] and ori (replica-tion origin). The inner circle shows the homology between R. solanacearum and R. metallidurans. The black square represents regions with high identity(proteins having 35% or more sequence identity over at least 80% of their total length and an expect value of BLASTP below 1036) with the R. metalli-durans genome. The distribution of putative homologues of heavy metal genes is depicted by arrows. Inner black clusters indicate putative orthologuesof R. metallidurans heavy metal resistance genes. Outer gray clusters indicate putative paralogues of R. metallidurans heavy metal resistance genes.
FEMSRE 778 2-6-03 Cyaan Magenta Geel Zwart
M. Mergeay et al. / FEMS Microbiology Reviews 27 (2003) 385^410390

Fig. 4 shows the general phylogeny of czcA paraloguestaking into account czcA orthologues in various bacteria.This high number of paralogues is a strong argument infavor of the long history of adaptation of R. metalliduransto metal-rich biotopes. In HME1, besides the putativeorthologue of R. solanacearum and other duly assignedorthologues from other bacteria, one recognizes the well-known plasmid-borne czc operon and a putative chromo-somal counterpart (fused contigs 280^465, data depositedin ftp://ftp.genome.bnl.gov/pub/Sequences/Rmetallidur-ans/) with which no phenotype or any other characteristicis associated.The HME2 group (nickel detoxi¢cation systems) in-
cludes the pMOL28 cnr operon and the pTOM9 ncc oper-
on of R. metallidurans 31A [4], but also a pMOL30-bornecluster that is probably not functional, at least as far asnickel resistance is concerned, because no nickel resistanceis associated with pMOL30.The HME3 type a and b groups are not yet assigned to
any function: according to Nies [84], HME3a czcA pa-ralogues are closely related and might process monovalentcations. Two of them belong to the same contig 647.Group HME3b also contains three paralogues: one standsalone (contig 695), the second (contig 711) is likely inac-tive as the corresponding gene contains a transposableelement inserted around 300 nucleotides from the startcodon. The third (contig 696) belongs to an operon thatappears to be associated with a large genetic element of
Fig. 3. R. metallidurans gene clusters containing czcA paralogues or orthologues. Bold: R. metallidurans loci and contigs. Gray bold: R. solanacearum.Other bacteria are shown in regular black. From left to right, column 1: R. metallidurans contig number or microbial locus; column 2: location (chro-mosomal, plasmid, MPL (megaplasmid of R. solanacearum [88]); column 3: czc-like gene clusters. Black: basic czcABC [20] structural genes or pa-ralogues; gray arrows: two-component regulatory genes [32] ; white: unknown functions; other ¢llings: other classes of genes sharing high identity orencoding the same function; double arrow (?) indicates the extremity of a contig; column 4: HME-RND class [84] ; column 5: phenotypes and observa-tions. Metal resistance genotypes: czc : Cd, Zn, Co [20] ; czr : Cd, Zn [120] ; czt : Cd, Zn [149] ; cnr : Co, Ni [6] ; ncc : Ni, Co, Cd [5] ; nre : Ni [5] ; sil : Ag[96] ; cus : Cu, Ag [141]. Mnemonics for other genotypes: hel : genes involved in virulence; cybB : cytochrome b561 [150], ORF-7266 : the correspondingprotein is similar to nodulin 21, a methionine-rich polypeptide observed in plant nodules [151] ; phnB : metabolism of phosphonates [152]. R. metallidu-rans genes that could be associated with a phenotype, a proteomic or a transcriptomic feature are marked in bold and with enlarged letters. Contigs280 and 465 are now linked (see text).
FEMSRE 778 2-6-03 Cyaan Magenta Geel Zwart
M. Mergeay et al. / FEMS Microbiology Reviews 27 (2003) 385^410 391

the Tn4371 family (biphenyl transposon) [92,93]. An ele-ment of this family is also present in the chromosome ofR. solanacearum [88,94].The HME4 group includes two operons that show gen-
otypic and phenotypic similarity with known RND sys-tems involved in copper and silver detoxi¢cation: cus inE. coli [95] and sil in a Salmonella typhimurium plasmid[96]: a cus-like operon was found on contig 619 and a sil-like cluster was present on plasmid pMOL30. Both sys-tems were at least partially revealed by proteomics inR. metallidurans. Challenging with silver helped to revealCusB and CusC. Challenging with copper revealed a poly-peptide corresponding to the pMOL30 silB gene (V. Au-quier, G. Vandenbussche and J.-M. Ruysschaert, personalcommunication), CusB and CusC (Fig. 3, Table 2). Pro-teomics appears to be a promising tool in the detection ofB and C components of RND systems. An interesting
challenge will reside in the choice of appropriate inducersto reveal HME3 type a and b proteins.
5. The P1-ATPases
P1-type ATPases are ancient proteins found in all or-ganisms [97,98]. An average of two to three genes encod-ing P-type ATPases are currently found in fully sequencedgenomes. For instance, two are found in E. coli, three inListeria species, Synechocystis, Bacillus subtilis or Helico-bacter pylori, ¢ve in Mycobacterium tuberculosis. Threegenes are found in R. solanacearum, one on the megaplas-mid (Fig. 2) and two on the chromosome. In contrast, theR. metallidurans genome contains up to 11 putative genesencoding P-type ATPases among which at least eight met-al-transporting P1-ATPases (also called CPx-ATPases)
Fig. 4. Phylogenetic representation of R. metallidurans czcA (putative) orthologues/paralogues and corresponding orthologues in R. solanacearum andother bacteria. Phylogenetic analyses were performed using the ‘TreeTop ^ Phylogenetic Tree Prediction’ program available at: http://www.genebee.msu.su/services/phtree_reduced.html. The topological algorithm in phylogram representation was used to display the phylogenetic tree. Building of prob-able phylogenetic trees is based on the matrix of pair distances between sequences. These distances (scale at the bottom of the ¢gure) are calculatedfrom counting residual change weights in aligned sequences. Branch lengths correspond to the phylogenetic distance between sequences. Bootstrap val-ues are expressed in percentages (after generation of 100 trees) and placed at nodes. NCBI accession numbers are given in brackets. On the right partof the ¢gure, the HME-RND classes to which the putative czc orthologues and paralogues would belong [84]. R. solanacearum megaplasmid proteinsRSp0493, RSp0530, RSp0928 and RSp1040 correspond to the Fig. 2 clusters czcSRCBA, czcCBA-like, silCBA-like and cnrTczcSRBAC respectively.Contig numbers correspond to R. metallidurans czc paralogues with unassigned function (see Fig. 3).
FEMSRE 778 2-6-03 Cyaan Magenta Geel Zwart
M. Mergeay et al. / FEMS Microbiology Reviews 27 (2003) 385^410392
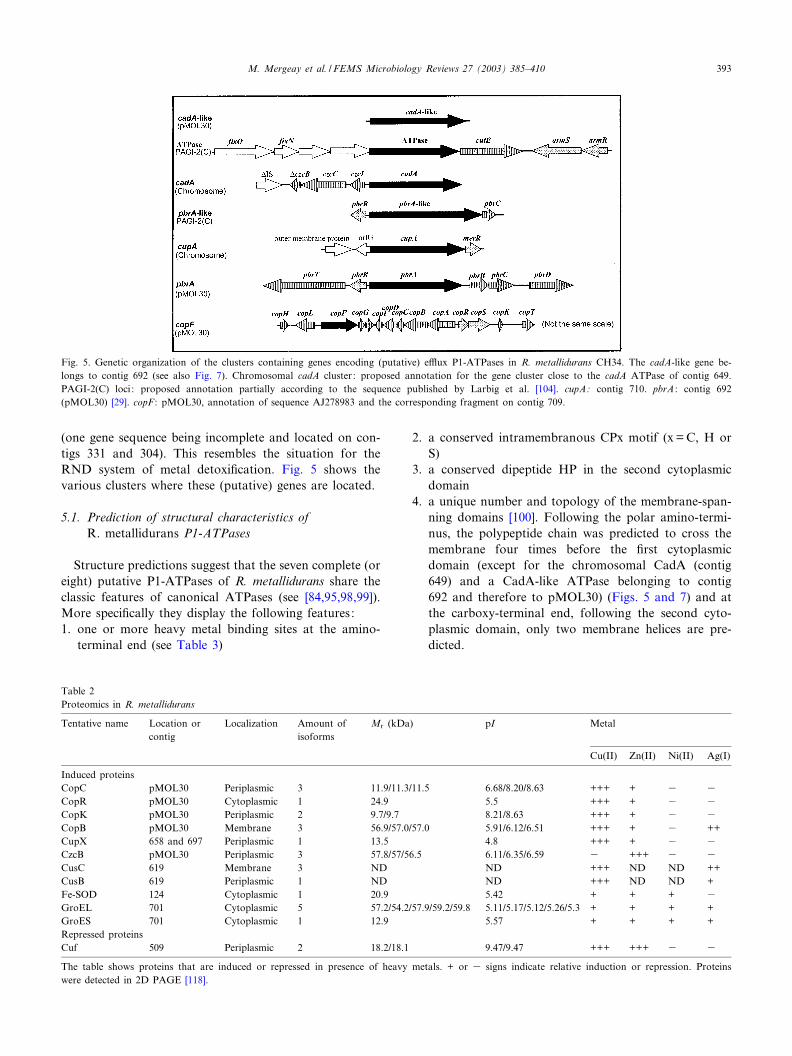
(one gene sequence being incomplete and located on con-tigs 331 and 304). This resembles the situation for theRND system of metal detoxi¢cation. Fig. 5 shows thevarious clusters where these (putative) genes are located.
5.1. Prediction of structural characteristics ofR. metallidurans P1-ATPases
Structure predictions suggest that the seven complete (oreight) putative P1-ATPases of R. metallidurans share theclassic features of canonical ATPases (see [84,95,98,99]).More speci¢cally they display the following features:1. one or more heavy metal binding sites at the amino-terminal end (see Table 3)
2. a conserved intramembranous CPx motif (x =C, H orS)
3. a conserved dipeptide HP in the second cytoplasmicdomain
4. a unique number and topology of the membrane-span-ning domains [100]. Following the polar amino-termi-nus, the polypeptide chain was predicted to cross themembrane four times before the ¢rst cytoplasmicdomain (except for the chromosomal CadA (contig649) and a CadA-like ATPase belonging to contig692 and therefore to pMOL30) (Figs. 5 and 7) and atthe carboxy-terminal end, following the second cyto-plasmic domain, only two membrane helices are pre-dicted.
Fig. 5. Genetic organization of the clusters containing genes encoding (putative) e¥ux P1-ATPases in R. metallidurans CH34. The cadA-like gene be-longs to contig 692 (see also Fig. 7). Chromosomal cadA cluster : proposed annotation for the gene cluster close to the cadA ATPase of contig 649.PAGI-2(C) loci : proposed annotation partially according to the sequence published by Larbig et al. [104]. cupA: contig 710. pbrA : contig 692(pMOL30) [29]. copF : pMOL30, annotation of sequence AJ278983 and the corresponding fragment on contig 709.
Table 2Proteomics in R. metallidurans
Tentative name Location orcontig
Localization Amount ofisoforms
Mr (kDa) pI Metal
Cu(II) Zn(II) Ni(II) Ag(I)
Induced proteinsCopC pMOL30 Periplasmic 3 11.9/11.3/11.5 6.68/8.20/8.63 +++ + 3 3
CopR pMOL30 Cytoplasmic 1 24.9 5.5 +++ + 3 3
CopK pMOL30 Periplasmic 2 9.7/9.7 8.21/8.63 +++ + 3 3
CopB pMOL30 Membrane 3 56.9/57.0/57.0 5.91/6.12/6.51 +++ + 3 ++CupX 658 and 697 Periplasmic 1 13.5 4.8 +++ + 3 3
CzcB pMOL30 Periplasmic 3 57.8/57/56.5 6.11/6.35/6.59 3 +++ 3 3
CusC 619 Membrane 3 ND ND +++ ND ND ++CusB 619 Periplasmic 1 ND ND +++ ND ND +Fe-SOD 124 Cytoplasmic 1 20.9 5.42 + + + 3
GroEL 701 Cytoplasmic 5 57.2/54.2/57.9/59.2/59.8 5.11/5.17/5.12/5.26/5.3 + + + +GroES 701 Cytoplasmic 1 12.9 5.57 + + + +Repressed proteinsCuf 509 Periplasmic 2 18.2/18.1 9.47/9.47 +++ +++ 3 3
The table shows proteins that are induced or repressed in presence of heavy metals. + or 3 signs indicate relative induction or repression. Proteinswere detected in 2D PAGE [118].
FEMSRE 778 2-6-03 Cyaan Magenta Geel Zwart
M. Mergeay et al. / FEMS Microbiology Reviews 27 (2003) 385^410 393

5.2. Phylogeny
Phylogenetic approaches demonstrated that the eightgenes grouped within three functionally, structurally, andphylogenetically distinct clusters [98,101^103]. One ofthose, harbored by the PAGI-2(C) genomic island [104],belongs to a separate cluster of ATPases presenting somesimilarities with FixI proteins, but for which metal speci-¢city has not yet been determined. This ATPase is in factpart of a locus which also includes an armRS two-compo-nent regulatory operon, four genes whose products displaysimilarities to cytochrome oxidases, and a cutE homo-logue. Three out of the seven P-type ATPase sequencesfound in the R. metallidurans genome encode transportproteins belonging to the copper/silver type of transporters(Fig. 6): one lies on the chromosome (contig 710) and isprovisionally called cupA, the other is the pMOL30-bornecopF (copper resistance), and the third is still incomplete.The proteins corresponding to the ¢rst two genes belongedto distinct branches of the Cu/Ag type of transporters:copF had no counterpart in the genome of the relatedplant pathogen R. solanacearum, but is rather phylogeneti-cally linked with silP, a plasmid-borne S. typhimuriumATPase involved in silver resistance [96], and to a lesserextent with the Enterococcus hirae copB ATPase involvedin copper export [105,106]. The three corresponding pro-teins share an extended CXXC motif in which the numberof amino acids between the two cysteines has been in-creased to three (conserved as YFCSXSC in the threesequences, X being A or N). Fig. 6 shows that R. metal-lidurans cupA clusters with a chromosomal R. solanacea-rum orthologue and is not necessarily the counterpart ofthe pMOL30 copF: their heavy metal motifs are also quitedi¡erent (Table 3). The real function of the chromosomal
cupA ATPase is not known but could resemble the detox-i¢cation function of the chromosomal E. coli CopA ATP-ase (see [107,108]). The CupA denomination is used inreference to the chromosomal E. coli CopA ATPase andto avoid confusion with the CopA periplasmic proteins(encoded in the copSRABCD gene clusters that are foundon both plasmid pMOL30 and chromosome of R. metal-lidurans). The possible eighth (incomplete) ATPase(contigs 304 and 331) might also belong to the Cu/Agfamily.The four remaining ATPase genes of R. metallidurans
appeared to be related to the Cd Zn Hg Pb transport P1-ATPases. Among these, a cadA-like gene located down-stream of the czc locus (resistance to Co(II), Zn(II) andCd(II)) of pMOL30 grouped with the Zn/Cd ATPasesencountered in R. capsulatus and Caulobacter crescentus,and, to a lesser extent, with the Cd ATPases from diverseFirmicutes. This cadA-like gene does not seem to belongto an operon: £anking genes do not give any clue. Aseparate cluster grouped around a P-type ATPase ofP. aeruginosa. This branch comprised the lead ATPasepbrA of R. metallidurans CH34 [29] and the proteins cor-responding to a pbrA-like gene located on the PAGI ge-nomic island [104], and to a chromosomal cadA (contig649). cadA insertional mutants made in the plasmid-freederivative of R. metallidurans CH34 resulted in increasedsensitivity to thallium, Cd(II) and lead (T. Vallaeys, M.Mergeay and A. Bossus, unpublished). In the presence ofpMOL30, the same kind of mutants displayed only aslight sensitivity to Cd(II). This suggests that anotherpMOL30 locus complements the increased sensitivity toheavy ions and that the pMOL30 czc system alone doesnot confer the maximal resistance to Cd(II) (T. Vallaeys,M. Mergeay and A. Bossus, unpublished).
Table 3Heavy metal binding motifs of six (putative) P1-ATPases of R. metallidurans CH34
pbrA (pMOL30 lead resistance operon pbrTRABCD)1. residues 31^94, ARYRIENMDCPTEEALIRDKLAKLPGVGGLEFNLMQRTLAVSHQMASPGPIEEALSAIGMRAVR2. residues 104^167, TVLAIHQMDCPTEEALIRSKLAGIPGVASLEFNLVQRTLHVWHAADTLSHVLTALKSLGFDAEVR
pbrA-like PAGI-2(C)1. residues 62^125, TVFHVSNMDCRNEEALVRRTLEGMPGVERLLFDLPQRLLTISHREVSADALEQALNSVGMKAQA2. residues 133^196, TTYRIENMDCPSEEKLIRSHLGAVEGIQDLDFDLAERTLSVKHLAQARAQMEQVLASIGMRAKE3. residues 233^296, VTYRIENMDCPTEEALIRDRLGKLPGVTALDFNLMQRVLGVQHTLATSAPIEKALASIGMQAAR4. residues 304^367, TVLRIAKMDCPTEESLIRGKLQGMPGVQGMDFNLMQRTLTVRHTPDAIKPAVEAIESLGMEAEV
Chromosomal CadA (contig 649)1. residues 21^55(*), HGRGHDHDHPHSRDHSHDHSHEHGHGHKHDHDHAH2. residues 92^154, AAYRIEAMDCPTEETLIRNKIGGMAGVAALDFNLMQRVLTVHHTLDSLDPVVKAIDSLGMKAEPL
CupA chromosomal putative Cu-ATPase (contig 710)1. residues 13^77, WRLPVEGMTCASCVRRVETALAKVPGVHDVAVNLATEQATLQADSPEVLNAAAKAVADAGYDVPR2. residues 80^145, IELNIADMTCASCVGRVERALRAVPGVVDAQVNLATERATVTVLRGAADVATLAAAVERAGYGATP
CopF (pMOL30 copper resistance cluster copKTSRABCDIGFLH)residues 10^34(*), HEHSAHGGASPGDHDAHHDHSHAGH
CadA-like (unknown phenotype or function) (contig 692)residues 156^181(*), HADHEHREGVKDHAHGEGEGHEAHAH
Relevant C, H and EE residues are underlined.The metal binding sites can be composed of a conserved motif (about 60 residues long) or a histidine-rich amino-terminus (marked with an asterisk)[148] or both.
FEMSRE 778 2-6-03 Cyaan Magenta Geel Zwart
M. Mergeay et al. / FEMS Microbiology Reviews 27 (2003) 385^410394

5.3. Metal ¢xation motifs
Table 3 reports putative metal binding motifs for sixATPases of R. metallidurans. The CXEE motifs ratherthan CXXC motifs were found in all R. metalliduransP-type ATPases belonging to the PbrA type of transport-ers (PbrA, chromosomal CadA and PbrA-like of thePAGI-2(C) genomic island). This motif was present aspart of a fully conserved MDCXXEEXLXR motif whichcould be repeated up to three times in the protein sequenceas is the case in the pbrA-like gene of the PAGI-2(C)island. The chromosomal CadA harbored both a CXXEEand a histidine-rich metal ¢xation motif. Conversely thechromosomal copper ATPase harbored the canonicalHMA motif CXXC, in two close copies in the N-terminalpart of the protein sequence. Similarly, a copy of the so-
called extended CXXXC site was found in CopF, in addi-tion to a histidine-rich type of metal binding motif.
6. CDF, CHR and ars-like genes
CDF stands for cation di¡usion facilitator: CzcD en-coded by the pMOL30 czc operon is the ¢rst prokaryoticrepresentative of this family to have been described[19,85,109,110].Two paralogues of CzcD are represented in contigs 705
and 708 of the R. metallidurans draft sequence but do notseem to be integrated in metal responses. Inside the czccluster, czcD is integrated in an operon czcDRSE that isinvolved in the regulation of the resistance to Zn(II),Cd(II) and Co(II), with czcR and S as classic representa-
Fig. 6. Phylogenetic representation of P1-ATPases. The proposed phylogeny shows three major branches: Cd(II), Zn(II) and Pb(II) ATPases, Cu(II)ATPases and ATPases with unknown functions with the position of seven (putative) R. metallidurans and three (putative) R. solanacearum ATPases.Phylogenetic trees were built using the Phylip package. Proteins similar to individual ATPases selected were identi¢ed by searching the NCBI proteindatabase using BLASTP. Multiple alignments in Phylip format were performed using the ClustalW software at http://www.infobiogen.fr/services/analy-seq/cgi-bin/clustalw_in.pl. The alignments were analyzed with ‘protdist’ (Phylip package), which uses protein sequences to compute a distance matrix.The distance for each pair of species estimates the total branch length between the two species, and was used in the distance matrix program NEIGH-BOR. The protdist output was then analyzed with the ‘NEIGHBOR’ program (Phylip package), which implements the neighbor-joining method.NEIGHBOR constructs a tree by successive clustering of lineages, setting branch lengths as the lineages join. Bootstrap analysis was performed usingboth ‘protpars’ (to generate 100 random trees) and ‘consense’ (to compute a consensus tree). The resulting tree was displayed using ‘treeview’ in an un-rooted format. NCBI accession numbers are given in brackets.
FEMSRE 778 2-6-03 Cyaan Magenta Geel Zwart
M. Mergeay et al. / FEMS Microbiology Reviews 27 (2003) 385^410 395

tives of the two-component sensor/activator system (seebelow). CzcD is involved in direct detoxi¢cation and inthe regulation of another metal e¥ux system [84].ChrA proteins characterize the CHR family of trans-
porters [111]. There are two ChrA orthologues: one ischromosome-bound (cluster chrBAF) [112], the other ispMOL28 plasmid-borne (cluster chrIBACEF) [25,112]. Incontig 665, a third putative chrA is tightly linked to tworegulation genes of the two-component sensor/activatorfamily. It is a typical con¢guration for metal-respondinggenes that warrant further investigation. A fourth chrA(putative paralogue) was seen in contig 393 and couldnot yet be assigned to a metal-processing function.Both chr clusters govern the resistance to chromate at
di¡erent toxicity levels [36]. Outside these resistance mech-anisms, one putative gene encoding a protein that is sim-ilar to the Pseudomonas chromate reductase is located incontig 628.
R. metallidurans does not appear to display an otherwiseanticipated resistance to arsenic oxyanions [84]. However,contig 596 contains a peculiar assembly of ars-like genes inthe order arsR-varsR-ORF-arsC-arsB/C-arsC-arsH. Thelatter gene, arsH, was ¢rst found in a defective transposonof a Yersinia enterocolitica plasmid conferring resistance toarsenite and arsenate [113], later in Acidithiobacillus fer-rooxidans [114].The striking feature of the R. metallidurans ars assembly
is the presence of three adjacent genes encoding a putativearsenate reductase activity. The corresponding phylogeny(data not shown) indicates that the three putative reduc-tases are quite distinct. Maybe, if they are functional theywould correspond to di¡erent oxyanionic substrates. Itwould be interesting to see if non-arsenic oxyanions wouldreveal one of these putative reductases via proteomics.
7. Large plasmids and mobile genetic elements ofR. metallidurans : content in heavy metalresistance genes
R. metallidurans strains often harbor conjugative plas-mids with metal resistance genes: the frequency of self-transfer is sometimes very high [52] but their host rangeseems to be limited to some species of the Ralstonia genus:R. metallidurans, R. eutropha, R. oxalatica, R. basilensis,and R. campinensis ([52], M. Mergeay, A. Sadouk and J.Gerits, unpublished). The markers that are known to betransferable are essentially those that are linked to thedeterminants of Zn(II) and Cd(II) (czc), Ni(II) (ncc), andcopper (cop) resistance. As far as pMOL28 and pMOL30from R. metallidurans CH34 are concerned, they are notself-transferable (or only at a very low frequency, e.g. as isthe case for pMOL28) but they can be e⁄ciently mobilizedby such ubiquitous plasmids as RP4 or other IncP/P1plasmids [15,115]. Mobilization processes, based on theformation of cointegrates after transposition, and the sub-
sequent homologous recombination to resolve the cointe-grates in replicons, made it possible to demonstrate thetransposition activity of the mercury transposons Tn4378(pMOL28) and Tn4380 (pMOL30) [15].The sequence of pMOL28 was obtained from the JGI
draft sequence of R. metallidurans. In pMOL28, metal re-sistance genes are con¢ned in a rather compact region ofthe plasmid including the mercury transposon Tn4378(merRTPADE), at least six ORFs involved in the resis-tance to chromate (chrI chrBACEF) [36], and the cnr-YXHCBA cnrT cluster involved in the resistance toNi(II) and Co(II) (Fig. 7) (for cnrT, see [84]). This ensem-ble received the name of ‘serpentine regulon’ referring tothe presence of nickel, chromate and mercury in serpen-tine-rich sediments that are thought to be a potential res-ervoir of metal-resistant strains [77,116].Genetic and sequencing data on resistance to Cd(II),
Zn(II) and Co(II) (czc) [20,32], resistance to Pb(II) [29],and resistance to Cu(II) [30] helped to assign contigs 663,692, and part of 709 to pMOL30. These contigs also con-tain some other putative genes for metal resistance withunknown functions. Contigs 663 (with czc) and 692 (withpbr) are linked (see ftp://ftp.genome.bnl.gov/pub/Sequen-ces/Rmetallidurans/). Fig. 7 shows the various clusters ofmetal resistance genes that are associated with pMOL30,including the mercury transposon Tn4380 [15].In pMOL30, according to the sequence data currently
available and the present annotation and functional anal-ysis (e.g. via mutational analysis, transcriptomics by real-time reverse transcription polymerase chain reaction [30]or proteomics; see Table 2) [117,118]), four functionalclusters have been identi¢ed: the czc cluster (czcNIC-BADRSE), allowing plasmid-borne resistance to Zn(II),Cd(II) and Co(II), the pbr cluster (pbrTRABCD, confer-ring resistance to lead), the cop cluster (copTKSRABC-DIGFLH, conferring resistance to copper), and the mer-cury transposon Tn4380 (merRTPADE). This transposonwas mapped on pMOL30 by classic genetic methods (de-letion mapping, restriction mapping, etc.) but did not ap-pear as such in the JGI draft of the R. metallidurans CH34genomic sequence. Both mercury transposons Tn4378(pMOL28) and Tn4380 (pMOL30) carry merE ORFs[119]. These merE ORFs, just like the correspondingMerE polypeptides of Tn501 and Tn21 transposons, con-tain the LTCPCHL motif. Another likely non-functionalmer cluster is located near czc as well as a paralogue ofmgtC (magnesium transport, Q. Dong, personal commu-nication). Other clusters (located on contig 709; Fig. 7)that must be mentioned are a putative ncc/cnr clusterwith no adjacent regulation genes (and which probablydoes not confer nickel resistance) and a putative sil cluster,again with no adjacent regulation genes but that doesseem to respond to the presence of copper in a proteomicsassay (V. Auquier, G. Vandenbussche and J.-M. Ruys-schaert, personal communication)Perhaps the most surprising feature of R. metallidurans
FEMSRE 778 2-6-03 Cyaan Magenta Geel Zwart
M. Mergeay et al. / FEMS Microbiology Reviews 27 (2003) 385^410396
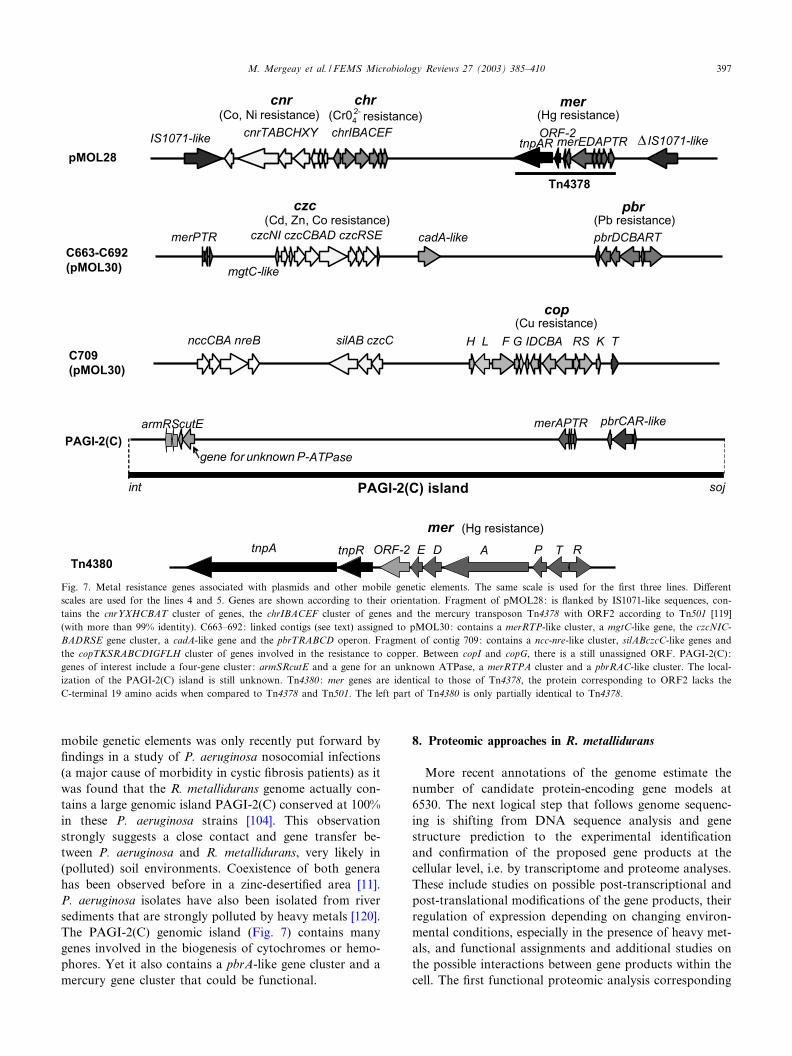
mobile genetic elements was only recently put forward by¢ndings in a study of P. aeruginosa nosocomial infections(a major cause of morbidity in cystic ¢brosis patients) as itwas found that the R. metallidurans genome actually con-tains a large genomic island PAGI-2(C) conserved at 100%in these P. aeruginosa strains [104]. This observationstrongly suggests a close contact and gene transfer be-tween P. aeruginosa and R. metallidurans, very likely in(polluted) soil environments. Coexistence of both generahas been observed before in a zinc-deserti¢ed area [11].P. aeruginosa isolates have also been isolated from riversediments that are strongly polluted by heavy metals [120].The PAGI-2(C) genomic island (Fig. 7) contains manygenes involved in the biogenesis of cytochromes or hemo-phores. Yet it also contains a pbrA-like gene cluster and amercury gene cluster that could be functional.
8. Proteomic approaches in R. metallidurans
More recent annotations of the genome estimate thenumber of candidate protein-encoding gene models at6530. The next logical step that follows genome sequenc-ing is shifting from DNA sequence analysis and genestructure prediction to the experimental identi¢cationand con¢rmation of the proposed gene products at thecellular level, i.e. by transcriptome and proteome analyses.These include studies on possible post-transcriptional andpost-translational modi¢cations of the gene products, theirregulation of expression depending on changing environ-mental conditions, especially in the presence of heavy met-als, and functional assignments and additional studies onthe possible interactions between gene products within thecell. The ¢rst functional proteomic analysis corresponding
Fig. 7. Metal resistance genes associated with plasmids and other mobile genetic elements. The same scale is used for the ¢rst three lines. Di¡erentscales are used for the lines 4 and 5. Genes are shown according to their orientation. Fragment of pMOL28: is £anked by IS1071-like sequences, con-tains the cnrYXHCBAT cluster of genes, the chrIBACEF cluster of genes and the mercury transposon Tn4378 with ORF2 according to Tn501 [119](with more than 99% identity). C663^692: linked contigs (see text) assigned to pMOL30: contains a merRTP-like cluster, a mgtC-like gene, the czcNIC-BADRSE gene cluster, a cadA-like gene and the pbrTRABCD operon. Fragment of contig 709: contains a ncc-nre-like cluster, silABczcC-like genes andthe copTKSRABCDIGFLH cluster of genes involved in the resistance to copper. Between copI and copG, there is a still unassigned ORF. PAGI-2(C):genes of interest include a four-gene cluster: armSRcutE and a gene for an unknown ATPase, a merRTPA cluster and a pbrRAC-like cluster. The local-ization of the PAGI-2(C) island is still unknown. Tn4380 : mer genes are identical to those of Tn4378, the protein corresponding to ORF2 lacks theC-terminal 19 amino acids when compared to Tn4378 and Tn501. The left part of Tn4380 is only partially identical to Tn4378.
FEMSRE 778 2-6-03 Cyaan Magenta Geel Zwart
M. Mergeay et al. / FEMS Microbiology Reviews 27 (2003) 385^410 397

to a global analysis of the R. metallidurans proteome hasbeen initiated in our laboratories as a prelude for thelarge-scale study of heavy metal response. Current proteo-mic analysis is based on two-dimensional gel electropho-resis (2-DE) and on mass spectrometry analysis [121,122].The main challenge of heavy metal proteomics is the de-tection of membrane proteins, which are overrepresentedin all the mechanisms conferring metal resistance. In thiscontext, a soluble and membrane protein 2-DE database isbeing constructed. This work shows the importance of theproteomic approach for genome analyses through (i) theidenti¢cation of previously undetected ORFs, (ii) the iden-ti¢cation of proteins not represented by the draft genomesequence, and (iii) the characterization of protein interac-tions, for instance between gene products originating fromtwo di¡erent contigs [118].Moreover, the ¢rst di¡erential proteomic analysis ap-
plied to R. metallidurans revealed new plasmid-borne pro-teins as well as a series of chromosomal gene clusters in-volved in the response to high concentrations of heavymetals. Table 2 shows a selection of proteins that wereinduced or repressed in the presence of metals, especiallyCu(II). The speci¢city of the protein response has beenstudied by comparison with other metals such as Ag(I),Zn(II), and Ni(II). The copper concentration used (0.8mM) is a typical restrictive concentration at which onlythe pMOL30-bearing strains can grow: it is not surprisingthat most of the induced proteins are plasmid-borne as the
regulator CopR (see Sections 9 and 11.1), the membrane-bound proteins CopB, and the periplasmic CopC andCopK proteins (as described below). The proteomic ap-proach also reveals the presence of di¡erent isoforms ofCopC, CopK, and CopB. The function of these isoformsremains unknown and is currently being studied in ourlaboratories. The expression of chromosomally derivedperiplasmic proteins with unknown functions was alsomodulated in the presence of Cu(II), i.e. CupX (induced)and Cuf (repressed). Table 2 also shows the presence ofheat shock and oxidative stress proteins. These proteinsare used as reference points on the 2-DE gel landscape.
9. Plasmid-borne copper resistance genes inR. metallidurans
Resistance to copper is found on the large plasmidpMOL30 and is transmissible by mobilization ofpMOL30 to a plasmid-free derivative of R. metalliduransCH34. The presence of pMOL30 increases the MIC tocopper by a factor of two to three in the transconjugants.Plasmid-borne resistance to copper was cloned and se-quenced (accession number: AJ278983). Genomic analysisshows that both the chromosome of R. metallidurans andpMOL30 share the six genes copSR-copABCD likely tocorrespond to a basic resistance mechanism [107] that isalso observed in a plasmid (provided with genes
Fig. 8. Copper resistance operons in R. metallidurans. The structure of the plasmid-borne cop cluster is presented in the top cartouche. Similar copS-RABCD genes located on chromosomal material of R. metallidurans (contig 711) and on the megaplasmid of R. solanacearum [88] are shown aligned inthe central rectangular cartouche. The alignment of similar genes in E. coli [123] and P. syringae [128] is shown in the lower part of the ¢gure. OnlycopSRABCD are conserved between all species. The E. coli plasmid pRJ1004-borne pcoE gene has no equivalent in other bacterial cop operons [124].
FEMSRE 778 2-6-03 Cyaan Magenta Geel Zwart
M. Mergeay et al. / FEMS Microbiology Reviews 27 (2003) 385^410398

pcoABCDpcoRSE) of an E. coli strain found in the fecesof copper-fed pigs [107,123,124], and in plasmids of Pseu-domonas syringae strains isolated after copper-based anti-fungal treatments of crop vegetables in California [125^129]. Genes of P. syringae variety tomato have the ordercopABCD-copRS [126,130], which is similar to the pcooperon structure where the transcription of both the cop/pcoABCD structural genes and the two-component copRSregulatory genes [131,132] is in the same orientation. Incontrast, the transcription of regulatory and structuralgenes is divergent in the Ralstonia genus (Fig. 8). As isshown in Fig. 9 for copC taken as a representative exam-ple of the six genes, the copSRABCD genes of R. metal-lidurans (chromosomally and on plasmid pMOL30) andtheir R. solanacearum counterparts are phylogeneticallytightly clustered. Intriguingly, the copSR-copABCD genesof pMOL30 are included in a larger cluster of genes thatrespond to copper: copTKSRABCDIFGLH (Figs. 7 and8). Transcriptomic approaches have also shown that these13 genes are transcribed in the presence of copper [30]. Adi¡erential proteomic approach also shows plasmid-borneproteins corresponding to periplasmic CopK and CopC,outer membrane-bound CopB, and cytoplasmic CopR(Table 2). In this respect, CopT is probably a paralogueof PbrT [29], seemingly an importer of lead as described inSection 10, and CopK is revealed as a new periplasmiccopper binding protein [117] (Table 2) while CopF is aP1-ATPase and CopG is likely to be involved in the ex-pression of CopF.
As shown in Fig. 9, the phyletic position of CopIbranches far away with respect to the periplasmic CopC.Its function is unknown although both proteins appear tobe related to Cu-containing oxidoreductases.CopH shares identity with the putative gene product of
czcE [32] but the function of both proteins is still un-known. This is also the case for the putative membraneprotein CopL.It is di⁄cult to understand the function of such complex
protein assemblies induced by copper. Yet, two alreadyknown mechanisms seem to be involved: the e¥ux ofcytoplasmic copper (thiol pool) that is mediated by aP1-ATPase (CopF) and the detoxi¢cation of periplasmiccopper via the copSR-copABCD-mediated e¥ux (by anal-ogy with what was observed or deduced in the E. coli pcosystem [107] and the P. syringae cop system). It is to berecalled that the chromosome also contains functionalcopSR-copABCD genes as well as a genetic determinantfor a putative Cu P1-ATPase, provisionally called cupA(see Section 5.2).The combinatory and regulatory integration (as seen for
the large gene cluster in pMOL30) of di¡erent resistancemechanisms that otherwise are not genetically linked (as isthe case on the chromosome) may explain the higher re-sistance to copper conferred by the pMOL30 plasmid. Onthe other hand, the complete deciphering of the copperresistance mechanism should also entail further studieson the isoforms for CopC, CopB and CopK, and theelucidation of their possible functions (Table 2).
Fig. 9. Phylogenetic representation of CopC proteins. Includes CopC of R. metallidurans (chromosomal and plasmid-borne), R. solanacearum, Pseudo-monas £uorescens, P. syringae, Xanthomonas campestris, E. coli strain K12, E. coli strain O157:H7, S. typhimurium, Salmonella enterica, Y. pestis, Deino-coccus radiodurans, Corynebacterium glutamicum, and CopI of R. metallidurans pMOL30. The same method was used for the phylogenetic analysis ofthe ATPases and copC. Using the R. metallidurans chromosome and pMOL30 copC (shown in bold) as reference, several copC homologues were foundin di¡erent organisms. NCBI accession numbers are given in brackets. copC genes were clustered in three di¡erent groups: R. solanacearum (shown inbold gray), P. £uorescens, P. syringae, X. campestris for the ¢rst group phylogenetically close to R. metallidurans ; E. coli strain K12, E. coli strainO157:H7, S. typhimurium, S. enterica, Y. pestis form the second group, and D. radiodurans, C. glutamicum which seems to be phylogenetically apart.R. metallidurans pMOL30 copI is also included in this phylogeny but does not group with the other genes.
FEMSRE 778 2-6-03 Cyaan Magenta Geel Zwart
M. Mergeay et al. / FEMS Microbiology Reviews 27 (2003) 385^410 399

10. Plasmid-borne genes for resistance to lead inR. metallidurans
Lead resistance was early recognized to be plasmid-borne in R. metallidurans [115], although it was only re-cently described at the molecular level [29]. However, dataabout lead resistance have also been reported for Arthro-bacter sp. and other Firmicutes that were isolated from thewaste site of an Australian factory of lead batteries [133].Pseudomonas marginalis showing extracellular lead exclu-sion and Bacillus megaterium demonstrating intracellularcytoplasmic lead accumulation were isolated from otherlead-contaminated soils [134]. Pb(II)-resistant strains ofStaphylococcus aureus and Citrobacter freundii have alsobeen isolated that accumulated the metal as an intracellu-
lar lead phosphate [135], though the molecular mechanismof detoxi¢cation remains to be elucidated. E¥ux of Pb(II)has also been reported for the CadA ATPase of S. aureusand the ZntA ATPase of E. coli [101,102].The lead resistance operon, pbr, of R. metallidurans
CH34 is unique, in the way that it combines functionsinvolved in uptake, e¥ux, and accumulation of Pb(II)[29] (Fig. 10). A MerR-like regulator, PbrR, controls tran-scription of the pbrABCD structural genes from the PpbrApromoter (Table 4), with the ¢rst gene downstream of thepbr operator/promoter encoding a P-type ATPase, like thezntA operon [136]. The PbrA Pb(II) ATPase is phyloge-netically grouped with the CadA-type Cd ATPases and theZntA Zn/Cd ATPase, and they form a group distinct fromthe Cu/Ag type ATPases. However, unlike CadA andZntA, we have previously seen that PbrA possesses twoHMA motifs with the amino acid sequence Cys-Pro-Thr-Glu-Glu instead of the consensus sequence Cys-X-X-Cys[29].In contrast to the cad and znt operons, which both
appear to comprise a regulatory gene plus an e¥ux ATP-ase only, additional proteins are required for maximalPb(II) resistance in R. metallidurans CH34. These arePbrT, PbrB, PbrC, and PbrD. The ¢rst step in pbr-en-coded Pb(II) resistance involves a Pb(II) uptake systemencoded by PbrT. Expression of PbrT in the absence ofPbrABCD results in Pb(II) hypersensitivity, probably dueto increased Pb(II) uptake into the cytoplasm. The resultof this Pb(II) uptake would be to reduce the interaction offree Pb(II) with side chains of membrane and periplasmicproteins, which would cause extensive cellular damage.Once Pb(II) has entered the cytoplasm, it can be exportedby the PbrA Pb(II) e¥ux ATPase or be bound by thePbrD protein, which may function as a chaperone forPb(II). PbrD is not absolutely required for Pb(II) resis-tance, but cells lacking PbrD show a decreased accumu-lation of Pb(II) compared to wild-type cells, and this ac-cumulation may protect against free exported Pb(II) andthe futile cycle of Pb(II) uptake and export [29].The PbrA Pb(II) e¥ux ATPase has been shown to be
Table 4Comparison of the promoter regions of di¡erent R. metallidurans CH34 genes that are under control of MerR- and PbrR-like regulators
335 310
PpbrA pMOL30 TTGACTCTATAGTAACTAGAGGGTG TTAAATCGGCAACGCGAGATGAAPpbrA-like PAGI-2(C) TTGACTCTATATCTACTAGAGGTTT TCTAATGATGGCATCCGGGGAAAPzntA TTGACTCTGGAGTCGACTCCAGAGTG TATCCTTCGGTTAATGAGAAAAAPcupA chromosome TTGACCTTCCAATGGTGGTAAGGTT CACAGTGCAGTTGTCGCGCTCCGTPforR C518 TTGACATTAAAGTTGACTTTAAGCT TAGGGTTACGCAGTCGTCAACGCAPorfA C671 TTGACGTTAACGTTAGCGTCAAGTC TAGCGCCTTATAACGACCAGAGACPmerT pMOL30(near czc) TTGACTCCGTACTCAGGTACGGGATT TAGAGTCACACTCGGGCAAACAGPmerT PAGI-2(C) TTGACTCCGTAGTTGACTACGGAAG TAACGTTACAGCATCAAGCGAAGPmerT Tn501 TTGACTCCGTACATGAGTACGGAAG TAAGGTTACGCTATCCAATTTCAPmerT Tn4380 TTGACTCCGTACATGAGTACGGAAG TAAGGTTACGCTATCCAATTTCAPmerT Tn4378 TTGACTCCGTACATGAGTACGGAAG TAAGGTTACGCTATCCAATTTCA
As comparison the PzntA (Zn resistance in E. coli) [136] and PmerT (Hg resistance of Tn501) [119] promoters are included. Bold: positions of 335 and310 regions; inverted repeats are underlined. Cnnn indicates the contig numbers of the newly identi¢ed merR-pbrR regulatory genes.
Fig. 10. Proposed mechanism for pMOL30 pbr-mediated resistance tolead. Model for the pbrTRABCD operon-encoded Pb resistance of R. me-tallidurans CH34. The model involves the following proteins: PbrT,which transports Pb into the cytoplasm; PbrA, the Pb e¥ux ATPase;PbrB and PbrC, the Pb transport-facilitating lipoprotein and its prolipo-protein signal peptidase, respectively ; and PbrD, a protein involved incytoplasmic Pb sequestration. The PbrR protein, which mediates Pb-in-ducible transcription from its divergent promoter, regulates the expres-sion of the pbr operon (from [29] with permission of the authors).
FEMSRE 778 2-6-03 Cyaan Magenta Geel Zwart
M. Mergeay et al. / FEMS Microbiology Reviews 27 (2003) 385^410400

functional and able to compensate for the Pb(II) uptakedriven by PbrT. However, for full Pb(II) resistance PbrBand PbrC are required. PbrB and related proteins may bepart of a new family of transporters assisting resistance(TAR) proteins. Comparison of PbrB with the EMBLdatabase resulted in 16 hits. However, with the exceptionof the BcrC protein of Bacillus licheniformis, all otherswere hypothetical membrane (lipo)proteins. The BcrC pro-tein of B. licheniformis encodes a hydrophobic membraneprotein that, together with the BcrB membrane protein,functions as membrane components of the bacitracin re-sistance ABC transporter [137]. Inactivation of BcrC re-sults in bacitracin sensitivity, and inactivation of PbrBresults in Pb(II) sensitivity. The PbrB lipoprotein maypromote transfer of Pb(II) from the periplasm to the outermembrane. This would result in a decreased Pb(II) uptakeby PbrT.At increased concentrations of Pb(II), metal removal
from the solution was observed for strain CH34 duringthe late exponential phase and the stationary phase, to-gether with a progressive increase of the pH (up to 9). Atthis increased pH the formation of lead complexes withhydroxide and carbonates will be strongly favored andshould play an important role in avoiding re-entry ofPb(II). A similar phenomenon has been described for theczc Cd(II)-Zn(II)-Co(II) resistance system of R. metallidu-rans CH34 [52]
11. Regulatory genes in the heavy metal resistance geneclusters
Regulatory genes for resistance to heavy metals in Ral-stonia are mostly strongly linked to structural genes, oftenupstream of the structural genes with a divergent tran-scription orientation or downstream of the structuralgenes but having the same orientation. By far the mostfrequent combination of regulatory genes belongs to thelarge two-component sensor and regulator family usingphosphorylation signals. But a few other families are dis-cussed too, including the well-known merR family. ThearsR/cadC/smtB family of regulators is also dealt withalthough the correlation of the arsRC gene clusters ob-served in R. solanacearum genomes or of the ars clusterof R. metallidurans (contig 596) with an arsenic resistancephenotype has still to be demonstrated. Some detailsabout new families of regulators such as cnr/nccYXHand the possibly new family czcI are included in the dis-cussion. Yet there are still quite a lot of genes found in themetal resistance clusters, generally membrane-bound, witha possible regulatory function for which no details orplausible mechanisms are currently available.
11.1. The R and S two-component family
Two-component RS systems are one of the major mech-
anisms responsible for the coordinated adaptation of liv-ing organisms in response to environmental challenges.Each system consists of a sensor kinase, generally an in-tegral membrane protein provided with a signal recogni-tion device, and a response regulator transcription factor,which activates or represses gene expression. In the pres-ence of an external stimulus, e.g. a threshold in extracel-lular metal concentration, the sensor kinase autophos-phorylates at an internal histidine and then transfers thehigh-energy phosphate group to an aspartyl residue on theresponse regulator. Two-component systems exist in or-ganisms belonging to all three domains of life, includingnon-animal Eucarya and Archaea. However, outside thedomain of Bacteria, two-component regulatory systemsare less abundant and seem to have been acquired byhorizontal gene transfer. In bacteria, such systems are usu-ally organized as a pair of adjacent genes included in alarge operon. They mediate adaptive responses to a widespectrum of environmental stimuli and play a crucial rolein metabolic processes such as aerobic respiration, osmo-regulation, sporulation, phosphate or nitrogen regulation,chemotaxis and virulence (for a review, see [138]).In many organisms, such as E. coli [139], R. solanacea-
rum [88] and P. aeruginosa [140], genes and/or operonsinvolved in e¥ux mechanisms are under the control ofvarious regulators, including two-component systems.R. metallidurans contains a high number of such proteins,with 42 sensor kinases and 67 response regulators so faridenti¢ed. This is more than the number found in the ge-nome of E. coli, i.e. 29 and 37, respectively, and that ofR. solanacearum, i.e. 37 and 55, respectively, but less thanin the genome of P. aeruginosa, i.e. 55 and 89, respectively.RS-mediated regulation of metal resistance genes is a com-mon feature of operons with RND genes (tricomponentcation/proton antiporter e¥ux), especially those displayinga Cd(II) (czc) [32], silver (sil) [96], or copper (cus) [141]resistance phenotype, but are also associated with cop/pco-mediated resistance to copper [131]. An associationwith a putative chrA should also be mentioned (see Section6).In R. metallidurans, at least seven two-component sys-
tems are associated with genes of the RND family, includ-ing ¢ve czc-like operons. A large diversity can be observedin terms of operon structure (Fig. 3). For example, some Rand S genes are divergently expressed from their structuralresistance genes, sometimes the R and S genes constitutean operon where both genes overlap, while they are eitherlocated downstream [32] or upstream of the structural re-sistance genes of czc-like clusters.Sensor kinases associated with czc-like and cop operon
components are highly similar to proteins of homologoussystems in various microorganisms, e.g. SilRS in S. ty-phimurium and CusRS in E. coli, and belong to type IA,which also includes the osmoregulation EnvZ/OmpR sys-tem and the KdpD/KdpE potassium transport in E. coli[142] (Fig. 11). The histidine site of phosphorylation is
FEMSRE 778 2-6-03 Cyaan Magenta Geel Zwart
M. Mergeay et al. / FEMS Microbiology Reviews 27 (2003) 385^410 401

located in the H-box domain and the kinase domain con-sists of several conserved consensus motifs. Yet, a fewresidues are conserved for all the S proteins involved inthe regulation of (putative) metal resistance genes (a Phein position 3 and an Arg in position 50 in Fig. 11a) andare not found in the other tested systems (Fig. 11a).Similarly, an alignment of their response regulator
counterparts demonstrates the presence of highly con-served residues, including not only amino acids knownto be involved in the catalytic transfer of the phosphorylgroup but also some residues located at their vicinity thatmay play an important role in the anchoring of both com-ponents during this process [143] (Fig. 11b). The observa-tion that these sensors and regulators form distinct clustersin a phylogenetic tree containing all two-component sys-tems identi¢ed in R. metallidurans further supports theirclose evolutionary relationship. However, such a compar-ative phylogenetic analysis of RS systems involved in met-al resistance with the corresponding regulated genes sug-gests that some of them may have evolved independentlyfrom each other (data not shown). With this respect, RSsystems and their target genes may constitute distinct evo-lutionary modules.Neither a metal binding site nor a histidine-rich domain,
known to be frequently associated with metal^protein in-teractions, was found to be conserved in the response reg-ulators or sensor kinases. This suggests that the mecha-nism by which bacterial cells sense the presence of heavymetals in their environment remains unclear at the proteinlevel. Nevertheless, as far as metal speci¢city is concerned,it seems that the systems (sensor and kinase) involved incopper resistance (cop and cus) remain clustered together,which is not the case for the RS genes that are implicatedin the regulation of the other HME-RND genes.At the direct protein level, at least one regulator (CopR
in P. syringae) has been described and further studied[132]. In R. metallidurans, proteomics helped to revealthe plasmid-borne CopR regulator (Table 2). We are con-¢dent that additional proteins with similar characteristicswill be found in the near future.
11.2. The merR family
In R. solanacearum, seven merR paralogues (of which¢ve are on the chromosome) have been identi¢ed. Onlytwo of these, one on the chromosome and one on themegaplasmid, are adjacent to a putative gene for metalresistance, in both cases a P-ATPase. There is no putativecluster conferring mercury resistance in this phytopatho-gen.Within the genome of R. metallidurans CH34, nine
MerR-like regulators have been identi¢ed. These regula-tors may be divided into four major groups based on se-quence similarity and functional linkage to structural re-sistance proteins.The ¢rst group consists of the MerR regulators, of
which the corresponding genes are found on pMOL28(Tn4378), on PAGI-2(C) and close to the czc operon),and on pMOL30 (Tn4380). They show close similaritywith the merR gene product of Tn501 and are alwayslinked to other, structural mercury resistance genes, whilethe merR-like gene close to the czc operon is linked to amerTP cassette. Owing to the absence of a merA gene, thecorresponding merRTP operon does not encode a func-tional Hg resistance system. In contrast, the PAGI-2(C)merR is linked to a merTPA Hg resistance cassette andcould encode a functional Hg resistance system; thepMOL28 merR is the regulator for the Tn4378-encodedHg resistance (merRTPAD operon) Interestingly, thefour MerR proteins of Tn4378 (pMOL28), Tn4380(pMOL30) [15] (Fig. 7) and Tn501 [144] are identicaland recognize identical PmerT promoters. This opens thepossibility for ¢ne-tuning the gene expression between thetwo major Hg resistance determinants located on the mer-cury resistance transposons of strain CH34.The second group of regulators consists of PbrR, the
regulator of the CH34 lead resistance operon that is lo-cated on pMOL30, and two regulators found on PAGI-2(C) and the chromosomal contig 710. As for thepbrTRABCD operon, the pbrR-like gene of PAGI-2(C) islinked to a heavy metal e¥ux ATPase, most similar to aCd e¥ux ATPase, and a pbcC-like gene (Fig. 5).The pbrR-like gene on contig 710 was not linked to any
structural heavy metal resistance gene. However, we can-not rule out that this regulator is involved in heavy metalresistance, since zntR [136] and cueR [145] are physicallydistant on the chromosome from their cognate promoters.A second merR-like gene of contig 710 is linked to a Cu
ATPase-type gene. This MerR-like regulator is more sim-ilar to ZntR (Fig. 12), and forms the third group. Thecorresponding gene is located downstream of cupA (seeFigs. 5 and 6, and Table 3). However, the ATPase andthe MerR-like regulator form one unit that is transcribedfrom a promoter, PcupA (contig 710), that has the proper-ties of a typical MerR-recognized promoter (Table 4).The Mer-like regulators of contigs 518 and 671 form the
last group. These two regulators are not physically linkedto any structural heavy metal resistance gene, and theirinvolvement in heavy metal resistance might be question-able.
11.3. The arsR family
arsR/cadC paralogues are represented in both R. metal-lidurans and R. solanacearum. In this latter bacterium,three of them are associated with the chromosome, whilethree are located on the megaplasmid. On each R. solana-cearum replicon, an association of (putative) arsR-arsC isobserved (see Fig. 2 for the arsRC of the megaplasmid). InR. metallidurans, only one complete arsR sensu stricto isobserved in the intriguing putative ars cluster describedabove. Eight other ‘arsR’ paralogues were identi¢ed that
FEMSRE 778 2-6-03 Cyaan Magenta Geel Zwart
M. Mergeay et al. / FEMS Microbiology Reviews 27 (2003) 385^410402

Fig. 11. a: Multiple sequence alignments of putative S genes from two-component sensor/kinase regulator systems. Amino acid alignment of the C-ter-minal part of sensor kinases belonging to the corresponding czc and czc-like operons. This includes the sequence of two other members of the type Iafamily [142], i.e. EnvZ and KdpD. Only the sequence downstream of the L237 residue in EnvZ and the corresponding sequence of other sensor kinasesare shown. The histidine box is black underlined as well as the downstream X region and the positive residues located at the extremity of the X region.Various conserved domains, i.e. N, G1, F, G2, G3, are also gray underlined. b: Multiple sequence alignments of putative R genes from two-componentsensor/kinase regulator systems. Amino acid alignment of the N-terminal part of the regulator belonging to the corresponding czc and czc-like operons.This includes the sequence of two other members of the OmpR family, i.e. OmpR and KdpE. Only the sequence up to the E111 residue in OmpR andthe corresponding sequence of other response regulators are shown. Conserved (gray arrows) and variable (gray dotted line) residues of loops 1^5 andhelix 1 supposed to be involved in the interaction surface of response regulators of the OmpR family with their sensor counterpart [143] are boxed.Black arrows point to catalytic residues.
FEMSRE 778 2-6-03 Cyaan Magenta Geel Zwart
M. Mergeay et al. / FEMS Microbiology Reviews 27 (2003) 385^410 403

belong to the cadC branch [146] : two are located on thePAGI-2(C) genomic island (Fig. 7), one of them beingassociated with the pbrA-like gene (see also Fig. 5). An-other cadC-like gene is adjacent to the (mutated) HME3b-RND operon located on contig 711 (see Fig. 3). Therefore,this would be an example of a new category of metal re-sistance genes regulated by the family arsR.
11.4. The cnrYXH family
RND-mediated resistance to Ni(II) has been reported inR. metallidurans plasmids pMOL28 (strain CH34) [6,24]and pTOM9 (strain 31A) [5] and is controlled by thegene clusters cnrYXHCBAT and nccYXHCBAN, respec-tively. In both gene clusters, nccY, nccX, and nccH areconsidered regulatory genes, of which the latter gene,cnrH, controls a c factor of RNA polymerase belongingto the ECF (environmental control factor) network. Mu-tations in cnrY were found to derepress the expression ofresistance to Ni(II) and Co(II). Two phenotypes were as-sociated with this derepression: the MIC of Ni(II) andCo(II) increases from 2.5 mM to 8 mM, and from 5 to10 mM respectively. The CnrY mutants were also resistantto Zn(II) and can be positively selected through this phe-notype [6,34,72].Mutants in cnrX are also constitutive but they did not
display a higher MIC to Ni(II) or Co(II) [6].Detailed molecular studies showed the presence of at
least two promoters that are recognized by CnrH: a pro-moter located upstream of cnrY [34,35], and another onelocated at the end of cnrX, a few bases upstream of the
start codon of cnrH (cnrHp) [34]. The former promoter,cnrYp, controls the transcription of cnrYXH mainly in theabsence of Ni(II) : yet no transcription of cnrCBA is ob-served in such conditions, suggesting that the transcriptionfrom cnrYp results in the synthesis of a complex of closelyinteracting proteins (CnrY, CnrX and CnrH) attached tothe inner membrane, allowing no access of the cECP factorCnrH to the transcription apparatus. In the presence ofNi(II), there is evidence for transcription from the cnrHppromoter that extends to the structural cnrCBA genes andallows the expression of full resistance to Ni(II) andCo(II). This active transcription from cnrHp seems to gotogether with decreased transcription from cnrYp : mem-brane proteins CnrY and CnrX are no longer producedand repression is no longer possible. Phenotypically, CnrYacts as a repressor although it is a membrane-bound pro-tein with no direct genotropic e¡ect. There is also an in-dication that transcription starts from a site between cnrHand cnrC [35] but the most active transcription in thepresence of otherwise toxic concentrations of Ni(II) clearlyoccurs from cnrHp [34]. The protein^protein interactionbetween CnrY and CnrH was con¢rmed using the Bacte-rioMatch Two-Hybrid system from Stratagene (S. Taghavi,unpublished results). In order to show these interactions itwas necessary to add a linker, consisting of [(Gly-Gly-Gly-Ser)2U] between the 3P end of the bacteriophage VcI re-pressor and the cnrY coding region. In addition, cnrY (thepredicted anti-c factor) was modi¢ed by replacing two Pheresidues of the predicted transmembrane region by Gluresidues in order to destroy this transmembrane domain,allowing access of the two-hybrid complex to the tran-
Fig. 12. Phylogenetic representation of the MerR-like proteins found in R. metallidurans. As references, the MerR protein of Tn501 and the ZntR pro-tein were taken. Numbers after the Mer proteins refer to the corresponding contigs.
FEMSRE 778 2-6-03 Cyaan Magenta Geel Zwart
M. Mergeay et al. / FEMS Microbiology Reviews 27 (2003) 385^410404

scription apparatus. The mutation resulted in a functionalVcI^CnrY hybrid protein. In addition, a RNA polymer-ase^cnrH translational fusion was constructed. Introduc-tion of both constructs in the appropriate E. coli reporterstrain resulted in ampicillin resistance plus L-galactosidaseactivity. This phenotype was not observed for the controlexperiments with strains carrying vector sequences only orwith constructs that lacked the CnrY or CnrH fusion pro-teins. This clearly demonstrates the interaction betweenCnrY and CnrH and supports the model of an anti-cECPfactor, CnrY, controlling the activity of CnrH, the cECP
factor.This mode of regulation is unique among metal resis-
tance operons and so far has only been observed in therelated plasmid-borne ncc resistance operon (strain R. me-tallidurans 31A).
11.5. Other regulatory genes
There are, in particular for the intricate plasmid-borneclusters, still a few genes with unknown function or whichhave been given a putative regulatory function.The genes czcN, nccN and copL encode membrane pro-
teins that are likely to be involved in regulation althoughtheir precise role remains enigmatic. nccN mutants de-creased the full resistance to nickel in R. metallidurans31A [5].
czcI is involved in the regulation of plasmid-borne Czc-mediated resistance to Zn(II), Cd(II) and Co(II). A pa-ralogue of czcI might also act as a regulator in a smallcluster containing a czcC-like gene and the proposed cadAgene encoding a P1-ATPase (contig 649; Fig. 5).
12. Concluding remarks
The present inventory of genes and operons that areinvolved in the resistance to heavy metals in R. metallidu-rans suggests that the genome of this bacterium containsmore genes related to this kind of function than any othergenome sequenced before. This is a striking notion whenwe look at the underlying genetic determinants for HME-RND e¥ux systems, the P1-ATPases, and the plasmid-mediated heavy metal resistance. It also supports the re-£ection from the molecular taxonomy data that R. metal-lidurans is specialized in the adaptation to heavy metals orto biotopes that are either occasionally or constitutivelyrich in heavy metals. It may therefore be postulated thatthe greater part of this specialization took place during aperiod largely before the development of anthropogenicactivities. At the present time, R. metallidurans CH34and its related strains are found almost exclusively inanthropogenic, industrial biotopes mostly bound to thewastes of metallurgical activities. The colonization by R.metallidurans and related bacteria of such biotopes may bethree centuries old if we refer to the European Industrial
Revolution that was determinant or even a few millenniaold if we refer to the very ¢rst developments of metallur-gical industry (copper, tin, mercury, etc.). In this respect,one wonders what the ‘natural’ biotopes of R. metallidu-rans could be [62]. A possible clue towards volcanic bio-topes could derive from the fact that R. metalliduransstrains are often hydrogenotrophs, and that serpentinerocks and dusts are rich in heavy metals and derivefrom the hydratation of eruptive olivine rocks. This hy-dratation releases measurable amounts of hydrogen thatcould help to support the growth of hydrogenotrophs atthe expense of atmospheric CO2 and air [147].This kind of speculation should mainly attract the at-
tention to the necessity to look at the microbial ecologyand taxonomy of harsh environments or any sites wherethe selection pressure would enhance the diversity and theexpression of metal resistance genes.The present catalogue of genes will surely be improved
with the completion of the genome. The closure of thegenome will be essential to evaluate the genetic linkagebetween metal resistance determinants and mobile geneticelements (plasmids, transposons, IS elements, genomic is-lands) and the role of mobile genetic elements in the emer-gence or in the acquisition of genetic determinants formetal resistance and processing. It will also help to checkthe presence, in R. metallidurans, of a second large repli-con as observed in R. solanacearum and in R. eutropha. Inthis respect, it is remarkable that, in R. solanacearum,most of the (putative) metal resistance genetic determi-nants are located on the megaplasmid and not on thechromosome. Importantly, with four HME-RND systems,R. solanacearum appears to be only second to R. metalli-durans in diversity of metal resistance genes.The catalogue points to the overlooked importance of
chromosomal genes (with respect to the situation observedin the plant pathogen R. solanacearum ^ and very likely inR. eutropha [90]), and within the context of this review, theterm ‘chromosomal’ actually refers to the genetic supportof genes that are carried in R. solanacearum by the chro-mosome and the 2.1-Mb megaplasmid [88]). Yet the im-portance of ‘ordinary’ plasmids in R. metallidurans shouldnot be underestimated, for at least three reasons:b pMOL28 and mainly pMOL30 have a uniquely highamount of metal resistance genes in comparison to allcurrently known plasmids from other bacteria
b they are remarkable for the composition of metal resis-tance gene clusters: those contain equivalents of chro-mosomal operons £anked by additional genes
b ¢nally, R. metallidurans plasmids carry various genes forwhich no real equivalents are known in other genomes,such as is copK (Table 2), cnrY and some other genesfrom the pbr, cop, cnr and czc plasmid-borne loci.The present catalogue of genes points mainly to cadmi-
um, cobalt, zinc, copper, and chromate at the plasmid andchromosomal levels, to mercury, nickel, and lead at theplasmid level, and to silver ^ at least at the chromosomal
FEMSRE 778 2-6-03 Cyaan Magenta Geel Zwart
M. Mergeay et al. / FEMS Microbiology Reviews 27 (2003) 385^410 405

level. As far as other metals are concerned, the presence ofsome plasmids mediates an increased tolerance to thalliumbut this metal does not seem to induce plasmid-bornegenes. Yet, chromosomal lux-based gene fusions that arespeci¢cally induced by thallium have been reported inR. metallidurans CH34 and AB2 (P. Corbisier and A. Bos-sus, personal communication).As this inventory mainly relies on the comparison of
already known genes or mechanisms either in Ralstoniagenomes or in other genomes, genuine novel genes orfunctions may be easily overlooked. Proteomics is of in-valuable help in this respect: it helps to assign functions toputative genes and it helps to detect new proteins, func-tions and metal signatures or domains. Consequently, thefuture of such catalogues lies in the combination of pro-teomics and functional genomics. It would give a non-em-pirical basis for the further development of biotechnolog-ical tools (including protein engineering by site-directedmutagenesis, etc.) to detect, concentrate, process or re-move heavy metals, including the heavy radionuclides,from waste storage or disposal sites.
Acknowledgements
Thanks are due to the Joint Genome Institute (US De-partment of Energy and University of California) for thesequencing of the genome of R. metallidurans and formaking it available to the scienti¢c community prior topublication. We are also deeply grateful to Geo¡reyHind for support and encouragement, and to BurkhardTu«mmler, Jacques Cove's, Dietrich Nies, Nigel Brown, Ed-ward Schwartz and Christopher Rensing for communicat-ing recent results and manuscripts. We are especiallygrateful to Christian Boucher for welcoming S.M. andM.M. in his laboratory in Toulouse and extensive ex-change of information about the R. solanacearum genome,to Je¤ro“me Gouzy for help in the comparisons of Ralstoniagenomes and the construction of Figure 2, to ArianeToussaint for the long-term collaboration and crucial dis-cussions about the genome of R. metallidurans, to JohanGoris and Peter Vandamme for the updated version ofFigure 1, to Patrick De Boever for decisive advice in theelaboration of the manuscript and Paul Janssen for carefulreading and insightful criticisms. The TOURNESOL ex-change program between French and Belgian scienti¢c in-stitutions supported the visit of S.M. in Toulouse. S.M. isalso grateful to Shoshana Wodak and Raphael Leplae forsupport and fruitful discussions. R.W. is a Research Asso-ciate at the National Funds for Scienti¢c Research of Bel-gium (FNRS). V.A. is supported by FRIA (Fonds de laRecherche Scienti¢que pour l’Industrie et l’Agriculture).D.v.d.L. and S.T. are currently supported by LaboratoryDirected Research and Development funds at the Brook-haven National Laboratory under contract with the USDepartment of Energy.
References
[1] Mergeay, M., Houba, C. and Gerits, J. (1978) Extrachromosomalinheritance controlling resistance to cadmium, cobalt, copper andzinc ions: evidence from curing in a Pseudomonas [proceedings].Arch. Int. Physiol. Biochem. 86, 440^442.
[2] Mergeay, M., Nies, D., Schlegel, H.G., Gerits, J., Charles, P. andVan Gijsegem, F. (1985) Alcaligenes eutrophus CH34 is a facultativechemolithotroph with plasmid-bound resistance to heavy metals.J. Bacteriol. 162, 328^334.
[3] Goris, J., De Vos, P., Coenye, T., Hoste, B., Janssens, D., Brim, H.,Diels, L., Mergeay, M., Kersters, K. and Vandamme, P. (2001) Clas-si¢cation of metal-resistant bacteria from industrial biotopes as Ral-stonia campinensis sp. nov., Ralstonia metallidurans sp. nov. and Ral-stonia basilensis Steinle et al., 1998 emend. Int. J. Syst. Evol.Microbiol. 51, 1773^1782.
[4] Nies, D., Mergeay, M., Friedrich, B. and Schlegel, H.G. (1987) Clon-ing of plasmid genes encoding resistance to cadmium, zinc, and co-balt in Alcaligenes eutrophus CH34. J. Bacteriol. 169, 4865^4868.
[5] Schmidt, T. and Schlegel, H.G. (1994) Combined nickel-cobalt-cad-mium resistance encoded by the ncc locus of Alcaligenes xylosoxidans31A. J. Bacteriol. 176, 7045^7054.
[6] Liesegang, H., Lemke, K., Siddiqui, R.A. and Schlegel, H.G. (1993)Characterization of the inducible nickel and cobalt resistance deter-minant cnr from pMOL28 of Alcaligenes eutrophus CH34. J. Bacte-riol. 175, 767^778.
[7] Mergeay, M., Gerits, J. and Houba, C. (1978) [Transmissible resis-tance factor to cobalt in Pseumononas hydrogenomonas]. C.R. Se¤ancesSoc. Biol. Fil. 172, 575^579.
[8] Friedrich, B., Hogrefe, C. and Schlegel, H.G. (1981) Naturally occur-ring genetic transfer of hydrogen-oxidizing ability between strains ofAlcaligenes eutrophus. J. Bacteriol. 147, 198^205.
[9] Gerstenberg, C., Friedrich, B. and Schlegel, H.G. (1982) Physicalevidence for plasmids in autotrophic, especially hydrogen-oxidizingbacteria. Arch. Microbiol. 133, 90^96.
[10] Lejeune, P., Mergeay, M., Van Gijsegem, F., Faelen, M., Gerits, J.and Toussaint, A. (1983) Chromosome transfer and R-prime plasmidformation mediated by plasmid pULB113 (RP4: :mini-Mu) in Alca-ligenes eutrophus CH34 and Pseudomonas £uorescens 6.2. J. Bacteriol.155, 1015^1026.
[11] Diels, L. and Mergeay, M. (1990) DNA probe-mediated detection ofresistant bacteria from soils highly polluted by heavy metals. Appl.Environ. Microbiol. 56, 1485^1491.
[12] Brim, H., Heyndrickx, M., de Vos, P., Wilmotte, A., Springael, D.,Schlegel, H.G. and Mergeay, M. (1999) Ampli¢ed rDNA restrictionanalysis and further genotypic characterisation of metal-resistant soilbacteria and related facultative hydrogenotrophs. Syst. Appl. Micro-biol. 22, 258^268.
[13] Yabuuchi, E., Kosako, Y., Yano, I., Hotta, H. and Nishiuchi, Y.(1995) Transfer of two Burkholderia and an Alcaligenes species toRalstonia gen. nov. Proposal of Ralstonia pickettii (Ralston, Palleroniand Doudoro¡ 1973) comb. nov., Ralstonia solanacearum (Smith1896) comb. nov. and Ralstonia eutropha (Davis 1969) comb. nov.Microbiol. Immunol. 39, 897^904.
[14] Taghavi, S., Mergeay, M. and van der Lelie, D. (1997) Genetic andphysical maps of the Alcaligenes eutrophus CH34 megaplasmidpMOL28 and its derivative pMOL50 obtained after temperature-in-duced mutagenesis and mortality. Plasmid 37, 22^34.
[15] Diels, L., Faelen, M., Mergeay, M. and Nies, D. (1985) Mercurytransposons from plasmids governing multiple resistance to heavymetals in Alcaligenes eutrophus CH34. Arch. Int. Physiol. Biochem.93, 27^28.
[16] Sadouk, A. and Mergeay, M. (1993) Chromosome mapping in Alca-ligenes eutrophus CH34. Mol. Gen. Genet. 240, 181^187.
[17] Mergeay, M. (2000) Bacteria adapted to industrial biotopes: the met-al resistant Ralstonia. In: Bacterial Stress Responses (Hengge-Aronis,R. and Storz, G., Ed.), pp. 403^414. ASM Press, Washington, DC.
FEMSRE 778 2-6-03 Cyaan Magenta Geel Zwart
M. Mergeay et al. / FEMS Microbiology Reviews 27 (2003) 385^410406

[18] Nies, D.H. and Silver, S. (1989) Metal ion uptake by a plasmid-freemetal-sensitive Alcaligenes eutrophus strain. J. Bacteriol. 171, 4073^4075.
[19] Nies, D.H. (1992) CzcR and CzcD, gene products a¡ecting regulationof resistance to cobalt, zinc, and cadmium (czc system) in Alcaligeneseutrophus. J. Bacteriol. 174, 8102^8110.
[20] Nies, D.H., Nies, A., Chu, L. and Silver, S. (1989) Expression andnucleotide sequence of a plasmid-determined divalent cation e¥uxsystem from Alcaligenes eutrophus. Proc. Natl. Acad. Sci. USA 86,7351^7355.
[21] Nies, D.H. (1995) The cobalt, zinc, and cadmium e¥ux systemCzcABC from Alcaligenes eutrophus functions as a cation-protonantiporter in Escherichia coli. J. Bacteriol. 177, 2707^2712.
[22] Sensfuss, C. and Schlegel, H.G. (1988) Plasmid pMOL28-encodedresistance to nickel is due to speci¢c e¥ux. FEMS Microbiol. Lett.55, 295^298.
[23] Siddiqui, R.A., Schlegel, H.G. and Meyer, M. (1988) Inducible andconstitutive expression of pMOL28-encoded nickel resistance in Al-caligenes eutrophus N9A. J. Bacteriol. 170, 4188^4793.
[24] Siddiqui, R.A., Benthin, K. and Schlegel, H.G. (1989) Cloning ofpMOL28-encoded nickel resistance genes and expression of the genesin Alcaligenes eutrophus and Pseudomonas spp. J. Bacteriol. 171,5071^5078.
[25] Nies, A., Nies, D.H. and Silver, S. (1989) Cloning and expression ofplasmid genes encoding resistances to chromate and cobalt in Alca-ligenes eutrophus. J. Bacteriol. 171, 5065^5070.
[26] Nies, A., Nies, D.H. and Silver, S. (1990) Nucleotide sequence andexpression of a plasmid-encoded chromate resistance determinantfrom Alcaligenes eutrophus. J. Biol. Chem. 265, 5648^5653.
[27] Stoppel, R.D. and Schlegel, H.G. (1995) Nickel-resistant bacteriafrom anthropogenically nickel-polluted and naturally nickel-perco-lated ecosystems. Appl. Environ. Microbiol. 61, 2276^2285.
[28] Margesin, R. and Schinner, F. (1997) Heavy metal resistant Arthro-bacter sp. ^ a tool for studying conjugational plasmid transfer be-tween gram-negative and gram-positive bacteria. J. Basic Microbiol.37, 217^227.
[29] Borremans, B., Hobman, J.L., Provoost, A., Brown, N.L. and vanDer Lelie, D. (2001) Cloning and functional analysis of the pbr leadresistance determinant of Ralstonia metallidurans CH34. J. Bacteriol.183, 551^568.
[30] Vallaeys, T., Benotmane, A., Noe«l-Georis, I., Borremans, B.,Monchy, S., Wattiez, R., Mergeay, M., Taghavi, S. and van derLelie, D. The organisation of plasmid-borne genes for resistance tocopper and of their chromosomal counterparts in Ralstonia metalli-durans. (in preparation).
[31] Rensing, C., Pribyl, T. and Nies, D.H. (1997) New functions for thethree subunits of the CzcCBA cation-proton antiporter. J. Bacteriol.179, 6871^6879.
[32] van der Lelie, D., Schwuchow, T., Schwidetzky, U., Wuertz, S., Bae-yens, W., Mergeay, M. and Nies, D.H. (1997) Two-component reg-ulatory system involved in transcriptional control of heavy-metalhomoeostasis in Alcaligenes eutrophus. Mol. Microbiol. 23, 493^503.
[33] Grosse, C., Grass, G., Anton, A., Franke, S., Santos, A.N., Lawley,B., Brown, N.L. and Nies, D.H. (1999) Transcriptional organizationof the czc heavy-metal homeostasis determinant from Alcaligeneseutrophus. J. Bacteriol. 181, 2385^2393.
[34] Tibazarwa, C., Wuertz, S., Mergeay, M., Wyns, L. and van DerLelie, D. (2000) Regulation of the cnr cobalt and nickel resistancedeterminant of Ralstonia eutropha (Alcaligenes eutrophus) CH34.J. Bacteriol. 182, 1399^1409.
[35] Grass, G., Grosse, C. and Nies, D.H. (2000) Regulation of the cnrcobalt and nickel resistance determinant from Ralstonia sp. strainCH34. J. Bacteriol. 182, 1390^1398.
[36] Juhnke, S., Peitzsch, N., Hu«bener, N., Grosse, C. and Nies, D. (2002)New genes involved in chromate resistance in Ralstonia metalliduransstrain CH34. Arch. Microbiol. 179, 15^25.
[37] Grass, G., Fan, B., Rosen, B.P., Lemke, K., Schlegel, H.G. and
Rensing, C. (2001) NreB from Achromobacter xylosoxidans 31A isa nickel-induced transporter conferring nickel resistance. J. Bacteriol.183, 2803^2807.
[38] Stoppel, R.D., Meyer, M. and Schlegel, H.G. (1995) The nickel re-sistance determinant cloned from the enterobacterium Klebsiella oxy-toca : conjugational transfer, expression, regulation and DNA homol-ogies to various nickel-resistant bacteria. Biometals 8, 70^79.
[39] Gilis, A., Khan, M.A., Cornelis, P., Meyer, J.M., Mergeay, M. andvan der Lelie, D. (1996) Siderophore-mediated iron uptake in Alca-ligenes eutrophus CH34 and identi¢cation of aleB encoding the ferriciron-alcaligin E receptor. J. Bacteriol. 178, 5499^5507.
[40] Gilis, A., Corbisier, P., Baeyens, W., Taghavi, S., Mergeay, M. andvan der Lelie, D. (1998) E¡ect of the siderophore alcaligin E on thebioavailability of Cd to Alcaligenes eutrophus CH34. J. Ind. Micro-biol. Biotechnol. 20, 61^68.
[41] Roux, M., Sarret, G., Pignot-Paintrand, I., Fontecave, M. andCoves, J. (2001) Mobilization of selenite by Ralstonia metalliduransCH34. Appl. Environ. Microbiol. 67, 769^773.
[42] Roux, M. and Coves, J. (2002) The iron-containing superoxide dis-mutase of Ralstonia metallidurans CH34. FEMS Microbiol. Lett. 210,129^133.
[43] Melchior, F., Ledrich, M.L., Foucaud, L. and Falla, J.A. (2001)Rapid identi¢cation of Ralstonia eutropha strain CH34 using mono-clonal antibodies. Hybrid Hybridomics 20, 325^332.
[44] Garg, R.P., Yindeeyoungyeon, W., Gilis, A., Denny, T.P., Van DerLelie, D. and Schell, M.A. (2000) Evidence that Ralstonia eutropha(Alcaligenes eutrophus) contains a functional homologue of the Ral-stonia solanacearum Phc cell density sensing system. Mol. Microbiol.38, 359^367.
[45] De Mot, R. and Parret, A.H. (2002) A novel class of self-su⁄cientcytochrome P450 monooxygenases in prokaryotes. Trends Microbiol.10, 502^508.
[46] Top, E., Mergeay, M., Springael, D. and Verstraete, W. (1990) Geneescape model: transfer of heavy metal resistance genes from Escheri-chia coli to Alcaligenes eutrophus on agar plates and in soil samples.Appl. Environ. Microbiol. 56, 2471^2479.
[47] Top, E.M., De Rore, H., Collard, J.-M., Gellens, V., Slobodkina, G.,Verstraete, W. and Mergeay, M. (1995) Retromobilization of heavymetal resistance genes in unpolluted and heavy metal polluted soil.FEMS Microbiol. Ecol. 18, 191^203.
[48] Weekers, F., Jacques, P.H., Springael, D., Mergeay, M., Diels, L.and Thonart, P.H. (1998) E¡ect of drying on bioremediation bacteriaproperties. Appl. Biochem. Biotechnol. 70^72, 311^322.
[49] Weekers, F., Rodriguez, C., Jacques, P., Mergeay, M. and Thonart,P. (2001) Dissemination of catabolic plasmids among desiccation-tol-erant bacteria in soil microcosms. Appl. Biochem. Biotechnol. 91^93,219^232.
[50] Diels, L., Springael, D., van der Lelie, N., Top, E. and Mergeay, M.(1993) Use of DNA probes and plasmid capture in a search for newinteresting environmental genes. Sci. Total Environ. 139^140, 471^478.
[51] Bossier, P., Top, E.M., Huys, G., Kersters, K., Boonaert, C.J., Rou-xhet, P.G. and Verstraete, W. (2000) Modi¢cation of the aggregationbehaviour of the environmental Ralstonia eutropha-like strain AE815is re£ected by both surface hydrophobicity and ampli¢ed fragmentlength polymorphism (AFLP) patterns. Environ. Microbiol. 2, 51^58.
[52] Diels, L., Dong, Q., van der Lelie, D., Baeyens, W. and Mergeay, M.(1995) The czc operon of Alcaligenes eutrophus CH34: from resis-tance mechanism to the removal of heavy metals. J. Ind. Microbiol.14, 142^153.
[53] Corbisier, P. (1997) Bacterial metal-lux biosensors for a rapid deter-mination of the heavy metal bioavailability and toxicity in solidsamples. Res. Microbiol. 148, 534^536.
[54] Corbisier, P., van der Lelie, D., Borremans, B., Provoost, A., deLorenzo, V., Brown, N.L., Lloyd, J.R., Hobman, J.L., Csoregi, E.,Johansson, G. and Mattiasson, B. (1999) Whole cell- and protein-
FEMSRE 778 2-6-03 Cyaan Magenta Geel Zwart
M. Mergeay et al. / FEMS Microbiology Reviews 27 (2003) 385^410 407

based biosensors for the detection of bioavailable heavy metals inenvironmental samples. Anal. Chim. Acta 387, 235^244.
[55] Peitzsch, N., Eberz, G. and Nies, D.H. (1998) Alcaligenes eutrophusas a bacterial chromate sensor. Appl. Environ. Microbiol. 64, 453^458.
[56] Tibazarwa, C., Corbisier, P., Mench, M., Bossus, A., Solda, P., Mer-geay, M., Wyns, L. and van der Lelie, D. (2001) A microbial bio-sensor to predict bioavailable nickel in soil and its transfer to plants.Environ. Pollut. 113, 19^26.
[57] van der Lelie, D., Corbisier, P., Baeyens, W., Wuertz, S., Diels, L.and Mergeay, M. (1994) The use of biosensors for environmentalmonitoring. Res. Microbiol. 145, 67^74.
[58] Heijerick, D.G., Janssen, C.R., Karlen, C., Wallinder, I.O. and Ley-graf, C. (2002) Bioavailability of zinc in runo¡ water from roo¢ngmaterials. Chemosphere 47, 1073^1080.
[59] Diels, L., Van Roy, S., Somers, K., Willems, I., Doyen, W., Mergeay,M., Springael, D. and Leysen, R. (1995) The use of bacteria immo-bilized in tubular membrane reactors for heavy metal recovery anddegradation of chlorinated aromatics. J. Membr. Sci. 100, 249^258.
[60] Diels, L., De Smet, M., Hooyberghs, L. and Corbisier, P. (1999)Heavy metals bioremediation of soil. Mol. Biotechnol. 12, 149^158.
[61] Dong, Q., Springael, D., Schoeters, J., Nuyts, G., Mergeay, M. andDiels, L. (1998) Horizontal transfer of bacterial heavy metal resis-tance genes and its applications in activated sludge systems. WaterSci. Technol. 37, 465^468.
[62] Lodewyckx, C., Taghavi, S., Mergeay, M., Vangronsveld, J., Clijs-ters, H. and van der Lelie, D. (2001) The e¡ect of recombinant heavymetal resistant endophytic bacteria in heavy metal uptake by theirhost plant. Int. J. Phytoremediat. 3, 173^187.
[63] Valls, M., Atrian, S., de Lorenzo, V. and Fernandez, L.A. (2000)Engineering a mouse metallothionein on the cell surface of Ralstoniaeutropha CH34 for immobilization of heavy metals in soil. Nat. Bio-technol. 18, 661^665.
[64] Valls, M. and de Lorenzo, V. (2002) Exploiting the genetic and bio-chemical capacities of bacteria for the remediation of heavy metalpollution. FEMS Microbiol. Rev. 26, 327^338.
[65] Springael, D., van Thor, J., Goorissen, H., Ryngaert, A., De Baere,R., Van Hauwe, P., Commandeur, L.C., Parsons, J.R., De Wachter,R. and Mergeay, M. (1996) RP4: :Mu3A-mediated in vivo cloningand transfer of a chlorobiphenyl catabolic pathway. Microbiology142, 3283^3293.
[66] Taghavi, S., van der Lelie, D. and Mergeay, M. (1994) Electropora-tion of Alcaligenes eutrophus with (mega) plasmids and genomicDNA fragments. Appl. Environ. Microbiol. 60, 3585^3591.
[67] Springael, D., Diels, L., Hooyberghs, L., Kreps, S. and Mergeay, M.(1993) Construction and characterization of heavy metal-resistanthaloaromatic-degrading Alcaligenes eutrophus strains. Appl. Environ.Microbiol. 59, 334^339.
[68] Springael, D., Diels, L. and Mergeay, M. (1994) Transfer and ex-pression of PCB-degradative genes into heavy metal resistant Alcali-genes eutrophus strains. Biodegradation 5, 343^357.
[69] Mergeay, M., Lejeune, P., Sadouk, A., Gerits, J. and Fabry, L.(1987) Shuttle transfer (or retrotransfer) of chromosomal markersmediated by plasmid pULB113. Mol. Gen. Genet. 209, 61^70.
[70] Szpirer, C., Top, E., Couturier, M. and Mergeay, M. (1999) Retro-transfer or gene capture: a feature of conjugative plasmids, withecological and evolutionary signi¢cance. Microbiology 145, 3321^3329.
[71] Dong, Q., Sadouk, A., van der Lelie, D., Taghavi, S., Ferhat, A.,Nuyten, J.M., Borremans, B., Mergeay, M. and Toussaint, A. (1992)Cloning and sequencing of IS1086, an Alcaligenes eutrophus insertionelement related to IS30 and IS4351. J. Bacteriol. 174, 8133^8138.
[72] Collard, J.M., Provoost, A., Taghavi, S. and Mergeay, M. (1993) Anew type of Alcaligenes eutrophus CH34 zinc resistance generated bymutations a¡ecting regulation of the cnr cobalt-nickel resistance sys-tem. J. Bacteriol. 175, 779^784.
[73] Mergeay, M. (1991) Towards an understanding of the genetics ofbacterial metal resistance. Trends Biotechnol. 9, 17^24.
[74] Collard, J.M., Corbisier, P., Diels, L., Dong, Q., Jeanthon, C., Mer-geay, M., Taghavi, S., van der Lelie, D., Wilmotte, A. and Wuertz, S.(1994) Plasmids for heavy metal resistance in Alcaligenes eutrophusCH34: mechanisms and applications. FEMS Microbiol. Rev. 14,405^414.
[75] Taghavi, S., Mergeay, M., Nies, D. and van der Lelie, D. (1997)Alcaligenes eutrophus as a model system for bacterial interactionswith heavy metals in the environment. Res. Microbiol. 148, 536^551.
[76] Nies, D.H. (1999) Microbial heavy-metal resistance. Appl. Microbiol.Biotechnol. 51, 730^750.
[77] Nies, D.H. (2000) Heavy metal-resistant bacteria as extremophiles:molecular physiology and biotechnological use of Ralstonia sp.CH34. Extremophiles 4, 77^82.
[78] Nies, D.H. (1992) Resistance to cadmium, cobalt, zinc, and nickel inmicrobes. Plasmid 27, 17^28.
[79] Nies, D.H. and Silver, S. (1995) Ion e¥ux systems involved in bac-terial metal resistances. J. Ind. Microbiol. 14, 186^199.
[80] Silver, S. (1996) Bacterial resistances to toxic metal ions ^ a review.Gene 179, 9^19.
[81] Silver, S. and Phung, L.T. (1996) Bacterial heavy metal resistance:new surprises. Annu. Rev. Microbiol. 50, 753^789.
[82] Silver, S. (1998) Genes for all metals ^ a bacterial view of the periodictable. The 1996 Thom Award Lecture. J. Ind. Microbiol. Biotechnol.20, 1^12.
[83] Mergeay, M. (1995) Heavy metal resistances in microbial ecosystems.In: Molecular Microbial Ecology Manual, pp. 1^17. Kluwer Aca-demic, Dordrecht.
[84] Nies, D. (2003) E¥ux-mediated heavy metal resistance in prokary-otes. FEMS Microbiol. Rev. 27, 313^339.
[85] Paulsen, I.T. and Saier, M.H.Jr. (1997) A novel family of ubiquitousheavy metal ion transport proteins. J. Membr. Biol. 156, 99^103.
[86] Tseng, T.T., Gratwick, K.S., Kollman, J., Park, D., Nies, D.H.,Go¡eau, A. and Saier Jr., M.H. (1999) The RND permease super-family: an ancient, ubiquitous and diverse family that includes hu-man disease and development proteins. J. Mol. Microbiol. Biotech-nol. 1, 107^125.
[87] Vandamme, P., Goris, J., Coenye, T., Hoste, B., Janssens, D.,Kersters, K., De Vos, P. and Falsen, E. (1999) Assignment of Centersfor Disease Control group IVc-2 to the genus Ralstonia as Ralstoniapaucula sp. nov. Int. J. Syst. Bacteriol. 49, 663^669.
[88] Salanoubat, M., Genin, S., Artiguenave, F., Gouzy, J., Mangenot, S.,Arlat, M., Billault, A., Brottier, P., Camus, J.C., Chandler, M.,Choisne, N., Claudel-Renard, S., Cumac, S., Demange, N., Gaspin,C., Lavie, M., Noisan, A., Robert, C., Saurin, W., Schiex, T., Siguier,P., Thebault, P., Whalen, M., Wincher, P., Levy, M., Weisenbach, J.and Boucher, C.A. (2002) Genome sequence of the plant pathogenRalstonia solanacearum. Nature 415, 497^502.
[89] Chen, W.M., Laevens, S., Lee, T.M., Coenye, T., De Vos, P., Mer-geay, M. and Vandamme, P. (2001) Ralstonia taiwanensis sp. nov.,isolated from root nodules of Mimosa species and sputum of a cystic¢brosis patient. Int. J. Syst. Evol. Microbiol. 51, 1729^1735.
[90] Schwartz, E. and Friedrich, B. (2001) A physical map of the mega-plasmid pHG1, one of three genomic replicons in Ralstonia eutrophaH16. FEMS Microbiol. Lett. 201, 213^219.
[91] Dong, Q. and Mergeay, M. (1994) Czc/Cnr e¥ux: a three-componentchemiosmotic antiport pathway with a 12-transmembrane-helix pro-tein. Mol. Microbiol. 14, 185^187.
[92] Springael, D., Kreps, S. and Mergeay, M. (1993) Identi¢cation of acatabolic transposon, Tn4371, carrying biphenyl and 4-chlorobiphe-nyl degradation genes in Alcaligenes eutrophus A5. J. Bacteriol. 175,1674^1681.
[93] Merlin, C., Springael, D., Mergeay, M. and Toussaint, A. (1997)Organisation of the bph gene cluster of transposon Tn4371, encodingenzymes for the degradation of biphenyl and 4-chlorobiphenyl com-pounds. Mol. Gen. Genet. 253, 499^506.
FEMSRE 778 2-6-03 Cyaan Magenta Geel Zwart
M. Mergeay et al. / FEMS Microbiology Reviews 27 (2003) 385^410408

[94] Toussaint, A., Merlin, C., Monchy, S., Benotmane, M., Leplae, R.,Coulson, A., Mergeay, M. and Springael, D. (2003) The biphenylcatabolic transposon Tn4371, a member of a new family of genomicislands related to IncP and Ti plasmids. Appl. Environ. Microbiol.(accepted).
[95] Rensing, C. and Grass, G. (2003) Escherichia coli mechanisms ofcopper homoeostasis in a changing environment. FEMS Microbiol.Rev. 27, 197^213.
[96] Gupta, A., Matsui, K., Lo, J.F. and Silver, S. (1999) Molecularbasis for resistance to silver cations in Salmonella. Nat. Med. 5,183^188.
[97] Scarborough, G.A. (1999) Structure and function of the P-typeATPases. Curr. Opin. Cell Biol. 11, 517^522.
[98] Rensing, C., Ghosh, M. and Rosen, B.P. (1999) Families of soft-metal-ion-transporting ATPases. J. Bacteriol. 181, 5891^5897.
[99] Palmgren, M.G. and Axelsen, K.B. (1998) Evolution of P-type ATP-ases. Biochim. Biophys. Acta 1365, 37^45.
[100] Solioz, M.A.V.C. (1996) CPx-type ATPases: a class of P-type ATP-ases that pump heavy metals. Trends Biochem. Sci. 21, 237^241.
[101] Rensing, C., Mitra, B. and Rosen, B.P. (1998) A Zn(II)-translocat-ing P-type ATPase from Proteus mirabilis. Biochem. Cell Biol. 76,787^790.
[102] Rensing, C., Sun, Y., Mitra, B. and Rosen, B.P. (1998) Pb(II)-trans-locating P-type ATPases. J. Biol. Chem. 273, 32614^32617.
[103] Rensing, C., Fan, B., Sharma, R., Mitra, B. and Rosen, B.P. (2000)CopA: An Escherichia coli Cu(I)-translocating P-type ATPase.Proc. Natl. Acad. Sci. USA 97, 652^656.
[104] Larbig, K.D., Christmann, A., Johann, A., Klockgether, J.,Hartsch, T., Merkl, R., Wiehlmann, L., Fritz, H-J. and Tuemmler,B. (2002) Gene islands integrated into tRNAGly genes confer ge-nome diversity on a Pseudomonas aeruginosa clone. J. Bacteriol. 184,6665^6680.
[105] Odermatt, A., Suter, H., Krapf, R. and Solioz, M. (1993) Primarystructure of two P-type ATPases involved in copper homeostasis inEnterococcus hirae. J. Biol. Chem. 268, 12775^12779.
[106] Solioz, M. and Odermatt, A. (1995) Copper and silver transport byCopB-ATPase in membrane vesicles of Enterococcus hirae. J. Biol.Chem. 270, 9217^9221.
[107] Lee, S.M., Grass, G., Rensing, C., Barrett, S.R., Yates, C.J., Stoya-nov, J.V. and Brown, N.L. (2002) The Pco proteins are involved inperiplasmic copper handling in Escherichia coli. Biochem. Biophys.Res. Commun. 295, 616^620.
[108] Franke, S., Grass, G. and Nies, D.H. (2001) The product of theybdE gene of the Escherichia coli chromosome is involved in detox-i¢cation of silver ions. Microbiology 147, 965^972.
[109] Anton, A., Grosse, C., Reissmann, J., Pribyl, T. and Nies, D.H.(1999) CzcD is a heavy metal ion transporter involved in regulationof heavy metal resistance in Ralstonia sp. strain CH34. J. Bacteriol.181, 6876^6881.
[110] Kunito, T., Kusano, T., Oyaizu, H., Senoo, K., Kanazawa, S. andMatsumoto, S. (1996) Cloning and sequence analysis of czc genes inAlcaligenes sp. strain CT14. Biosci. Biotechnol. Biochem. 60, 699^704.
[111] Nies, D.H., Koch, S., Wachi, S., Peitzsch, N. and Saier Jr., M.H.(1998) CHR, a novel family of prokaryotic proton motive force-driven transporters probably containing chromate/sulfate antiport-ers. J. Bacteriol. 180, 5799^5802.
[112] Juhnke, S., Peitzsch, N., Hu«bener, N., Grosse, C. and Nies, D.(2002) New genes involved in chromate resistance in Ralstonia me-tallidurans strain CH34. Arch. Microbiol. 179, 15^25.
[113] Neyt, C., Iriarte, M., Thi, V.H. and Cornelis, G.R. (1997) Virulenceand arsenic resistance in Yersiniae. J. Bacteriol. 179, 612^619.
[114] Butcher, B.G., Deane, S.M. and Rawlings, D.E. (2000) The chro-mosomal arsenic resistance genes of Thiobacillus ferrooxidans havean unusual arrangement and confer increased arsenic and antimonyresistance to Escherichia coli. Appl. Environ. Microbiol. 66, 1826^1833.
[115] Diels, L., Sadouk, A. and Mergeay, M. (1989) Large plasmids gov-erning multiple resistance to heavy metals : a genetic approach. Tox-icol. Environ. Chem. 23, 79^89.
[116] Baker, A.J.M. (1987) Metal tolerance in plants. New Phytol. 106,93^111.
[117] Wattiez, R., Auquier, V., Noe«l-Georis, I., Vallaeys, T., Vandenbus-sche, G., Ruysschaert, J.-M., Falmagne, P. and Mergeay, M. (2003)Proteomic analysis of copper response in Ralstonia metalliduransCH34 unravels new resistance mechanisms. (in preparation).
[118] Noe«l-Georis, I., Vallaeys, T., Chauvaux, R., Monchy, S., Falmagne,P., Mergeay, M. and Wattiez, R. (2003) Global analysis of theRalstonia metallidurans proteome: prelude for the large scale studyof heavy metal response. (submitted).
[119] Liebert, C.A. and Summers, A.O. (1999) Transposon Tn21, £agshipof the £oating genome. Microbiol. Mol. Biol. Rev. 63, 507^522.
[120] Hassan, M.T., van der Lelie, D., Springael, D., Romling, U.,Ahmed, N. and Mergeay, M. (1999) Identi¢cation of a gene cluster,czr, involved in cadmium and zinc resistance in Pseudomonas aeru-ginosa. Gene 238, 417^425.
[121] O’Connor, C.D., Adams, P., Alefounder, P., Farris, M., Kinsella,N., Li, Y., Payot, S. and Skipp, P. (2000) The analysis of microbialproteomes: strategies and data exploitation. Electrophoresis 21,1178^1186.
[122] Washburn, M. and Yates III, J.R. (2000) Analysis of the microbialproteome. Curr. Opin. Microbiol. 3, 292^297.
[123] Brown, N.L., Rouch, D.A. and Lee, B.T. (1992) Copper resistancedeterminants in bacteria. Plasmid 27, 41^51.
[124] Brown, N.L., Barrett, S.R., Camakaris, J., Lee, B.T. and Rouch,D.A. (1995) Molecular genetics and transport analysis of the cop-per-resistance determinant (pco) from Escherichia coli plasmidpRJ1004. Mol. Microbiol. 17, 1153^1166.
[125] Bender, C.L. and Cooksey, D.A. (1986) Indigenous plasmids inPseudomonas syringae pv. tomato conjugative transfer and role incopper resistance. J. Bacteriol. 165, 534^541.
[126] Bender, C.L. and Cooksey, D.A. (1987) Molecular cloning of cop-per resistance genes from Pseudomonas syringae pv. tomato. J. Bac-teriol. 169, 470^474.
[127] Cooksey, D.A. (1993) Copper uptake and resistance in bacteria.Mol. Microbiol. 7, 1^5.
[128] Cooksey, D.A. (1994) Molecular mechanisms of copper resistanceand accumulation in bacteria. FEMS Microbiol. Rev. 14, 381^386.
[129] Cha, J.S. and Cooksey, D.A. (1991) Copper resistance in Pseudo-monas syringae mediated by periplasmic and outer membrane pro-teins. Proc. Natl. Acad. Sci. USA 88, 8915^8919.
[130] Mellano, M.A. and Cooksey, D.A. (1988) Nucleotide sequence andorganization of copper resistance genes from Pseudomonas syringaepv. tomato. J. Bacteriol. 170, 2879^2883.
[131] Mills, S.D., Jasalavich, C.A. and Cooksey, D.A. (1993) A two-com-ponent regulatory system required for copper-inducible expressionof the copper resistance operon of Pseudomonas syringae. J. Bacte-riol. 175, 1656^1664.
[132] Mills, S.D., Lim, C.K. and Cooksey, D.A. (1994) Puri¢cation andcharacterization of CopR, a transcriptional activator protein thatbinds to a conserved domain (cop box) in copper-inducible pro-moters of Pseudomonas syringae. Mol. Gen. Genet. 244, 341^351.
[133] Trajanovska, S., Britz, M.L. and Bhave, M. (1997) Detection ofheavy metal ion resistance genes in gram-positive and gram-negativebacteria isolated from a lead-contaminated site. Biodegradation 8,113^124.
[134] Roane, T.M. (1999) Lead resistance in two bacterial isolates fromheavy metal-contaminated soils. Microb. Ecol. 37, 218^224.
[135] Levinson, H.S. and Mahler, I. (1998) Phosphatase activity and leadresistance in Citrobacter freundii and Staphylococcus aureus. FEMSMicrobiol. Lett. 161, 135^138.
[136] Brocklehurst, K.R., Hobman, J.L., Lawley, B., Blank, L., Marshall,S.J., Brown, N.L. and Morby, A.P. (1999) ZntR is a Zn(II)-respon-
FEMSRE 778 2-6-03 Cyaan Magenta Geel Zwart
M. Mergeay et al. / FEMS Microbiology Reviews 27 (2003) 385^410 409

sive MerR-like transcriptional regulator of zntA in Escherichia coli.Mol. Microbiol. 31, 893^902.
[137] Podlesek, Z., Comino, A., Herzog-Velikonja, B., Zgur-Bertok, D.,Komel, R. and Grabnar, M. (1995) Bacillus licheniformis bacitracin-resistance ABC transporter : relationship to mammalian multidrugresistance. Mol. Microbiol. 16, 969^976.
[138] Koretke, K.K., Koretke, L.A.N., Warren, P.V., Rosenberg, M. andBrown, J.R. (2000) Evolution of two-component signal transduc-tion. Mol. Biol. Evol. 17, 1956^1970.
[139] Blattner, F.R., Plunkett III, G., Bloch, C.A., Perna, N.T., Burland,V., Riley, M., Collado-Vides, J., Glasner, J.D., Rode, C.K., May-hew, G.F., Gregor, J., Davis, N.W., Kirkpatrick, H.A., Goeden,M.A., Rose, D.J., Mau, B. and Shao, Y. (1997) The complete ge-nome sequence of Escherichia coli K-12. Science 277, 1453^1474.
[140] Stover, C.K., Pham, X.Q., Erwin, A.L., Mizoguchi, S.D., Warrener,P., Hickey, M.J., Brinkman, F.S., Hufnagle, W.O., Kowalik, D.J.,Lagrou, M., Garber, R.L., Goltry, L., Tolentino, E., Westbrock-Wadman, S., Yuan, Y., Brody, L.L., Coulter, S.N., Folger, K.R.,Kas, A., Larbig, K., Lim, R., Smith, K., Spencer, D., Wong, G.K.,Wu, Z., Paulsen, I.T., Reizer, J., Saier, M.H., Hancock, R.E., Lory,S. and Olson, M.V. (2000) Complete genome sequence of Pseudo-monas aeruginosa PA01, an opportunistic pathogen. Nature 406,959^964.
[141] Grass, G. and Rensing, C. (2001) Genes involved in copper homeo-stasis in Escherichia coli. J. Bacteriol. 183, 2145^2147.
[142] Kim, D. and Forst, S. (2001) Genomic analysis of the histidinekinase family in bacteria and archea. Microbiology 147, 1197^1212.
[143] Hoch, J.A. and Varughese, K.I. (2001) Keeping signals straight inphosphorelay signal transduction. J. Bacteriol. 183, 4941^4949.
[144] Lund, P.A., Ford, S.J. and Brown, N.L. (1986) Transcriptional reg-
ulation of the mercury-resistance genes of transposon Tn501. J. Gen.Microbiol. 132, 465^480.
[145] Adaikkalam, V. and Swarup, S. (2002) Molecular characterizationof an operon, cueAR, encoding a putative P1-type ATPase and aMerR-type regulatory protein involved in copper homeostasis inPseudomonas putida. Microbiology (UK) 148, 2857^2867.
[146] Busenlehner, L.S., Penella, M.A. and Giedroc, D.P. (2003) TheSmtB/ArsR family of metalloregulatory transcriptional repressors:structural insights into prokaryotic metal resistance. FEMS Micro-biol. Rev. 27, 131^143.
[147] Zierenberg, R.A., Adams, M.W. and Arp, A.J. (2000) Life in ex-treme environments: hydrothermal vents. Proc. Natl. Acad. Sci.USA 97, 12961^12962.
[148] Tong, L., Nakashima, S., Shibasaka, M., Katsuhara, M. and Kasa-mo, K. (2002) A novel histidine-rich CPx-ATPase from the ¢lamen-tous cyanobacterium Oscillatoria brevis related to multiple-heavy-metal cotolerance. J. Bacteriol. 184, 5027^5035.
[149] Rossbach, S., Kukuk, M.L., Wilson, T.L., Feng, S.F., Pearson,M.M. and Fisher, M.A. (2000) Cadmium-regulated gene fusionsin Pseudomonas £uorescens. Environ. Microbiol. 2, 373^382.
[150] Nakamura, H., Murakami, H., Yamato, I. and Anraku, Y. (1988)Nucleotide sequence of the cybB gene encoding cytochrome b561 inEscherichia coli K12. Mol. Gen. Genet. 212, 1^5.
[151] Delauney, A.J., Cheon, C.I., Snyder, P.J. and Verma, D.P. (1990) Anodule-speci¢c sequence encoding a methionine-rich polypeptide,nodulin-21. Plant Mol. Biol. 14, 449^451.
[152] Kulakova, A.N., Kulakov, L.A., Akulenko, N.V., Ksenzenko, V.N.,Hamilton, J.T. and Quinn, J.P. (2001) Structural and functionalanalysis of the phosphonoacetate hydrolase (phnA) gene region inPseudomonas £uorescens 23F. J. Bacteriol. 183, 3268^3275.
FEMSRE 778 2-6-03 Cyaan Magenta Geel Zwart
M. Mergeay et al. / FEMS Microbiology Reviews 27 (2003) 385^410410
Copyright © 2022 FDOKUMEN




















