
Quaternary Phosphorites from the Continental Margin Off Chennai, Southeast India: Analogs of Ancient...
13
JOURNAL OF SEDIMENTARY RESEARCH,VOL. 70, NO. 5, SEPTEMBER, 2000, P. 1197–1209 Copyright q 2000, SEPM (Society for Sedimentary Geology) 1073-130X/00/070-1205/$03.00 QUATERNARY PHOSPHORITES FROM THE CONTINENTAL MARGIN OFF CHENNAI, SOUTHEAST INDIA: ANALOGS OF ANCIENT PHOSPHATE STROMATOLITES V. PURNACHANDRA RAO, 1 K. MOHAN RAO, 2 AND D.S.N. RAJU 3 1 National Institute of Oceanography (NIO), Dona Paula 403 004, Goa, India; e-mail: [email protected] 2 NIO Regional Centre, Lawsons Bay Colony, Visakhapatnam 530 017, India 3 Oil and Natural Gas Commission, Dehra Dun, India ABSTRACT: Pleistocene phosphorites occur on the continental margin off Chennai abundantly in the depth range of 186–293 m. They are associated with outer-shelf glauconites and carbonate skeletals includ- ing large shells of molluscs and rhodoliths. These are primary phos- phate deposits and are similar to phosphate stratiform stromatolites. This is the first report of phosphate stratiform stromatolites during the Quaternary and modern times, and they provide a Quaternary analog for ancient phosphorites. Thin sections of phosphorites exhibit laminated phosphatized micro- bial mats consisting of mechanically deposited clastic particles or of phosphatic molds. Clay particles are scattered throughout the matrix. SEM studies reveal that the laminated phosphate matrix is made of thin sheets of tightly packed apatite globules, densely packed inter- twined microfilaments, or filament molds. Phosphate with obscured laminae is composed of phosphate tubules. Cell-like structures resem- bling coccoid cyanobacteria or their botryoidal aggregates are com- mon. Carbonate fluorapatite and low-magnesium and high-magnesium calcites are the major mineral phases. The structural CO 2 content of apatite ranges from 4 to 5.5%. Quartz, feldspar, and goethite are ac- cessory minerals. The microbial mats were formed on the outer shelf, most probably during conditions of low sea level in the Quaternary, and microbial processes played a major role in direct phosphatization of these mats. The primary phosphorus source seems to have been the continental supply and degradation of organic matter associated with benthic microbial communities. The phosphate stratiform stromatolites thus formed were subsequently reworked into a shelf-margin depres- sion and resulted in the occurrence of condensed phosphorites. INTRODUCTION Ancient phosphorite deposits differ from the Quaternary and modern phosphorites in size, grade, and facies association. Ancient phosphorite deposits are larger in size, are mostly granular (pellets and ooids), and contain high concentrations of phosphate. Numerous stromatolitic (colum- nar) phosphorites formed during Precambrian–Cambrian times have been reported to be of replacement type (see Cook and Shergold 1986). Exten- sive reworking and/or benthic microbial communities might have played a major role in their formation. In contrast, Cambrian pure apatite stromat- olites are present on the African craton, Sahara (Bertrand-Sarfati et al. 1997). Mesozoic phosphate stromatolites interpreted as primary deposits have been found on the Alpine–Mediterranean paleo-margins (Fo ¨ llmi 1990; Krajewski et al. 1994; Martin-Algarra and Sanchez-Navas 1995). Soudry and Panczer (1994) reported Eocene stromatolitic phosphorites from Ne- gev, Israel. These are also columnar phosphate stromatolites. On the other hand, Quaternary and modern phosphorite deposits are smaller in size and occur as scattered nodules and pellets at the sediment–water interface (with- in the top few centimeters of sediments) and at or near to the boundary of the oxygen-minimum zone (Burnett 1977; Baturin 1982; Glenn et al. 1994a). Most of the modern phosphorites occur on western margins asso- ciated with upwelling (Kazakov 1937; Burnett and Riggs 1990). The sourc- es of phosphorus for modern phosphorites have been presumed to be sed- imented plankton organic matter, fish debris, and the iron redox cycle phos- phate pump. Phosphate stromatolites, however, are unknown from Quater- nary and modern phosphorites. Bentor (1980) suggested that modern phosphorites are poor analogs of their ancient counterparts. Recent studies, however, emphasize some relationships between the two. For example, Fi- lippelli and Delaney (1992) demonstrated that the phosphorus fluxes in ancient and modern phosphogenic environments are similar. Glenn et al. (1994b) showed that the abundance of modern phosphorites off Peru is much greater than previously estimated. Here, we describe laminated, microbially influenced phosphorite deposits off Chennai, by documenting microstructures and ultrastructures and min- eralogy, that share similarities to previously reported phosphate stromato- lites. These Quaternary phosphorites, however, do not share the macro- scopic, columnar habit of most ancient phosphate stromatolites. Both co- lumnar and stratiform stromatolites form from sediment trapping and bind- ing and precipitation because of metabolic activity (Walter 1976). In general, stratiform stromatolites with more or less planar lamination are synonymous with cryptalgal laminites (see Monty 1976). GEOLOGICAL SETTING The study region is located off Chennai on the southeastern continental margin of India (Fig. 1). The onshore region off Chennai is a tectonic depression bound by the Eastern Ghats on the west. The depression consists of phosphorites and phosphatic sediments of Upper Cretaceous to Lower Eocene successions (Laskar and Narasimhan 1968; Sastry et al. 1977). The sediments on the inner continental shelf off Chennai are clayey, whereas those on the outer shelf are relict sandy sediments (Subbarao 1964). Sandy clays consisting of a mixture of biogenic and terrigenous sandy clays occur on the continental slope. Phosphorites are associated with the relict sedi- ments of the outer shelf and upper slope. Faunal elements such as larger bivalve and gastropod shells belonging to the Upper Cretaceous to Eocene sequence have also been described from the continental shelf (Vaz 1995). Wind-driven upwelling is limited to only monsoon months (July–August). A southward current (0.15 to 0.3 m/s) induced by cyclonic eddies prevails on the continental shelf and slope. The western boundary current has pres- ently been traced about 186 km away from the coast (M.T. Babu, personal communication 2000). We describe bathymetry and seismic data, and de- tailed petrography and mineralogy of the phosphorites, in the study region and explain the origin and occurrence of phosphorites at these depths. RESULTS Bathymetric and Seismic data The bathymetric data of the three profiles V13, V14, and V15 normal to the coast off Chennai (Fig. 1) indicate that the shelf is wider (46 km) in the central profile (V14) and narrower (V13, 41 km; V15, 35 km) on either side. The average gradients of the shelf along these profiles were 1 in 205, 135, and 130, respectively. The outer-shelf region is rugged, and the shelf break lies at about 200, 340, and 270 m for V13, V14 and V15, respectively. An isolated topographic high of ; 15 m relief, with an ero- sional landward valley, is present on the upper slope (around 350 m) along the profile V15. The high-resolution shallow seismic record for profile V14 revealed three
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Quaternary Phosphorites from the Continental Margin Off Chennai, Southeast India: Analogs of Ancient...

JOURNAL OF SEDIMENTARY RESEARCH, VOL. 70, NO. 5, SEPTEMBER, 2000, P. 1197–1209Copyright q 2000, SEPM (Society for Sedimentary Geology) 1073-130X/00/070-1205/$03.00
QUATERNARY PHOSPHORITES FROM THE CONTINENTAL MARGIN OFF CHENNAI,SOUTHEAST INDIA: ANALOGS OF ANCIENT PHOSPHATE STROMATOLITES
V. PURNACHANDRA RAO,1 K. MOHAN RAO,2 AND D.S.N. RAJU3
1 National Institute of Oceanography (NIO), Dona Paula 403 004, Goa, India;e-mail: [email protected]
2 NIO Regional Centre, Lawsons Bay Colony, Visakhapatnam 530 017, India3 Oil and Natural Gas Commission, Dehra Dun, India
ABSTRACT: Pleistocene phosphorites occur on the continental marginoff Chennai abundantly in the depth range of 186–293 m. They areassociated with outer-shelf glauconites and carbonate skeletals includ-ing large shells of molluscs and rhodoliths. These are primary phos-phate deposits and are similar to phosphate stratiform stromatolites.This is the first report of phosphate stratiform stromatolites during theQuaternary and modern times, and they provide a Quaternary analogfor ancient phosphorites.
Thin sections of phosphorites exhibit laminated phosphatized micro-bial mats consisting of mechanically deposited clastic particles or ofphosphatic molds. Clay particles are scattered throughout the matrix.SEM studies reveal that the laminated phosphate matrix is made ofthin sheets of tightly packed apatite globules, densely packed inter-twined microfilaments, or filament molds. Phosphate with obscuredlaminae is composed of phosphate tubules. Cell-like structures resem-bling coccoid cyanobacteria or their botryoidal aggregates are com-mon. Carbonate fluorapatite and low-magnesium and high-magnesiumcalcites are the major mineral phases. The structural CO2 content ofapatite ranges from 4 to 5.5%. Quartz, feldspar, and goethite are ac-cessory minerals. The microbial mats were formed on the outer shelf,most probably during conditions of low sea level in the Quaternary,and microbial processes played a major role in direct phosphatizationof these mats. The primary phosphorus source seems to have been thecontinental supply and degradation of organic matter associated withbenthic microbial communities. The phosphate stratiform stromatolitesthus formed were subsequently reworked into a shelf-margin depres-sion and resulted in the occurrence of condensed phosphorites.
INTRODUCTION
Ancient phosphorite deposits differ from the Quaternary and modernphosphorites in size, grade, and facies association. Ancient phosphoritedeposits are larger in size, are mostly granular (pellets and ooids), andcontain high concentrations of phosphate. Numerous stromatolitic (colum-nar) phosphorites formed during Precambrian–Cambrian times have beenreported to be of replacement type (see Cook and Shergold 1986). Exten-sive reworking and/or benthic microbial communities might have played amajor role in their formation. In contrast, Cambrian pure apatite stromat-olites are present on the African craton, Sahara (Bertrand-Sarfati et al.1997). Mesozoic phosphate stromatolites interpreted as primary depositshave been found on the Alpine–Mediterranean paleo-margins (Follmi 1990;Krajewski et al. 1994; Martin-Algarra and Sanchez-Navas 1995). Soudryand Panczer (1994) reported Eocene stromatolitic phosphorites from Ne-gev, Israel. These are also columnar phosphate stromatolites. On the otherhand, Quaternary and modern phosphorite deposits are smaller in size andoccur as scattered nodules and pellets at the sediment–water interface (with-in the top few centimeters of sediments) and at or near to the boundary ofthe oxygen-minimum zone (Burnett 1977; Baturin 1982; Glenn et al.1994a). Most of the modern phosphorites occur on western margins asso-ciated with upwelling (Kazakov 1937; Burnett and Riggs 1990). The sourc-es of phosphorus for modern phosphorites have been presumed to be sed-imented plankton organic matter, fish debris, and the iron redox cycle phos-
phate pump. Phosphate stromatolites, however, are unknown from Quater-nary and modern phosphorites. Bentor (1980) suggested that modernphosphorites are poor analogs of their ancient counterparts. Recent studies,however, emphasize some relationships between the two. For example, Fi-lippelli and Delaney (1992) demonstrated that the phosphorus fluxes inancient and modern phosphogenic environments are similar. Glenn et al.(1994b) showed that the abundance of modern phosphorites off Peru ismuch greater than previously estimated.
Here, we describe laminated, microbially influenced phosphorite depositsoff Chennai, by documenting microstructures and ultrastructures and min-eralogy, that share similarities to previously reported phosphate stromato-lites. These Quaternary phosphorites, however, do not share the macro-scopic, columnar habit of most ancient phosphate stromatolites. Both co-lumnar and stratiform stromatolites form from sediment trapping and bind-ing and precipitation because of metabolic activity (Walter 1976). Ingeneral, stratiform stromatolites with more or less planar lamination aresynonymous with cryptalgal laminites (see Monty 1976).
GEOLOGICAL SETTING
The study region is located off Chennai on the southeastern continentalmargin of India (Fig. 1). The onshore region off Chennai is a tectonicdepression bound by the Eastern Ghats on the west. The depression consistsof phosphorites and phosphatic sediments of Upper Cretaceous to LowerEocene successions (Laskar and Narasimhan 1968; Sastry et al. 1977). Thesediments on the inner continental shelf off Chennai are clayey, whereasthose on the outer shelf are relict sandy sediments (Subbarao 1964). Sandyclays consisting of a mixture of biogenic and terrigenous sandy clays occuron the continental slope. Phosphorites are associated with the relict sedi-ments of the outer shelf and upper slope. Faunal elements such as largerbivalve and gastropod shells belonging to the Upper Cretaceous to Eocenesequence have also been described from the continental shelf (Vaz 1995).Wind-driven upwelling is limited to only monsoon months (July–August).A southward current (0.15 to 0.3 m/s) induced by cyclonic eddies prevailson the continental shelf and slope. The western boundary current has pres-ently been traced about 186 km away from the coast (M.T. Babu, personalcommunication 2000). We describe bathymetry and seismic data, and de-tailed petrography and mineralogy of the phosphorites, in the study regionand explain the origin and occurrence of phosphorites at these depths.
RESULTS
Bathymetric and Seismic data
The bathymetric data of the three profiles V13, V14, and V15 normalto the coast off Chennai (Fig. 1) indicate that the shelf is wider (46 km)in the central profile (V14) and narrower (V13, 41 km; V15, 35 km) oneither side. The average gradients of the shelf along these profiles were 1in 205, 135, and 130, respectively. The outer-shelf region is rugged, andthe shelf break lies at about 200, 340, and 270 m for V13, V14 and V15,respectively. An isolated topographic high of ; 15 m relief, with an ero-sional landward valley, is present on the upper slope (around 350 m) alongthe profile V15.
The high-resolution shallow seismic record for profile V14 revealed three
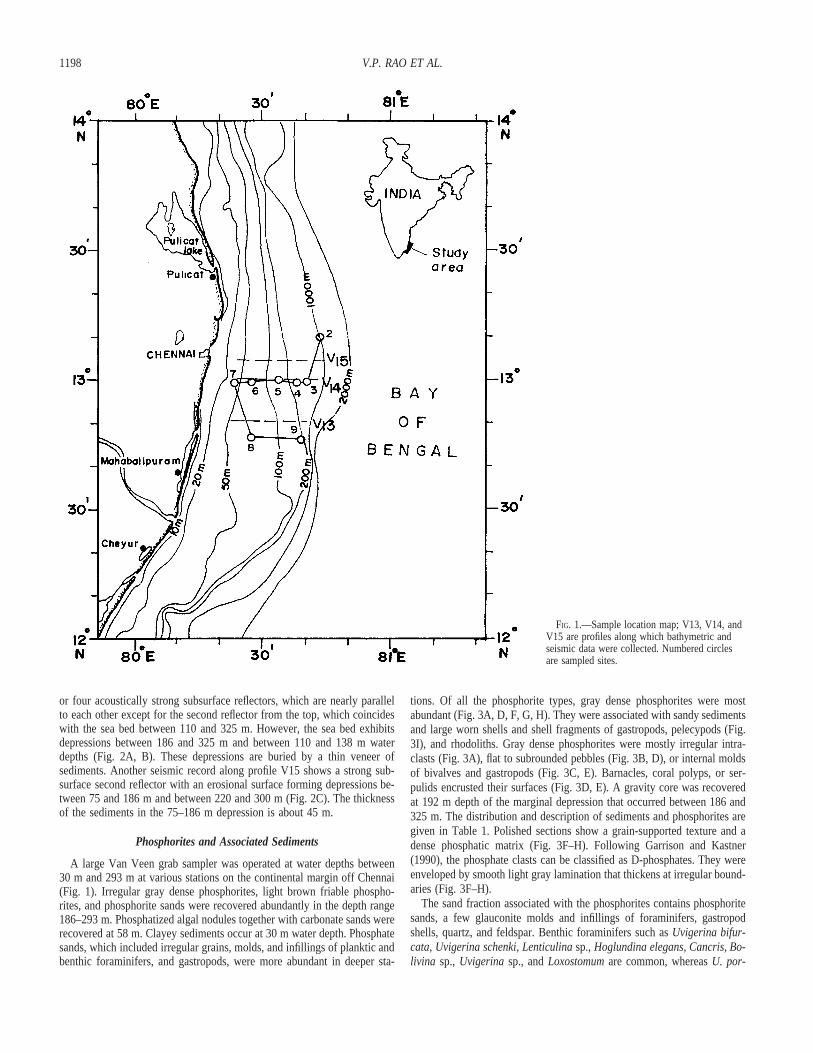
1198 V.P. RAO ET AL.
FIG. 1.—Sample location map; V13, V14, andV15 are profiles along which bathymetric andseismic data were collected. Numbered circlesare sampled sites.
or four acoustically strong subsurface reflectors, which are nearly parallelto each other except for the second reflector from the top, which coincideswith the sea bed between 110 and 325 m. However, the sea bed exhibitsdepressions between 186 and 325 m and between 110 and 138 m waterdepths (Fig. 2A, B). These depressions are buried by a thin veneer ofsediments. Another seismic record along profile V15 shows a strong sub-surface second reflector with an erosional surface forming depressions be-tween 75 and 186 m and between 220 and 300 m (Fig. 2C). The thicknessof the sediments in the 75–186 m depression is about 45 m.
Phosphorites and Associated Sediments
A large Van Veen grab sampler was operated at water depths between30 m and 293 m at various stations on the continental margin off Chennai(Fig. 1). Irregular gray dense phosphorites, light brown friable phospho-rites, and phosphorite sands were recovered abundantly in the depth range186–293 m. Phosphatized algal nodules together with carbonate sands wererecovered at 58 m. Clayey sediments occur at 30 m water depth. Phosphatesands, which included irregular grains, molds, and infillings of planktic andbenthic foraminifers, and gastropods, were more abundant in deeper sta-
tions. Of all the phosphorite types, gray dense phosphorites were mostabundant (Fig. 3A, D, F, G, H). They were associated with sandy sedimentsand large worn shells and shell fragments of gastropods, pelecypods (Fig.3I), and rhodoliths. Gray dense phosphorites were mostly irregular intra-clasts (Fig. 3A), flat to subrounded pebbles (Fig. 3B, D), or internal moldsof bivalves and gastropods (Fig. 3C, E). Barnacles, coral polyps, or ser-pulids encrusted their surfaces (Fig. 3D, E). A gravity core was recoveredat 192 m depth of the marginal depression that occurred between 186 and325 m. The distribution and description of sediments and phosphorites aregiven in Table 1. Polished sections show a grain-supported texture and adense phosphatic matrix (Fig. 3F–H). Following Garrison and Kastner(1990), the phosphate clasts can be classified as D-phosphates. They wereenveloped by smooth light gray lamination that thickens at irregular bound-aries (Fig. 3F–H).
The sand fraction associated with the phosphorites contains phosphoritesands, a few glauconite molds and infillings of foraminifers, gastropodshells, quartz, and feldspar. Benthic foraminifers such as Uvigerina bifur-cata, Uvigerina schenki, Lenticulina sp., Hoglundina elegans, Cancris, Bo-livina sp., Uvigerina sp., and Loxostomum are common, whereas U. por-

1199QUATERNARY PHOSPHORITES OFF CHENNAI, SOUTHEAST INDIA
FIG. 2.—Seismic profiles showing shelf-margin depressions (thin arrows) at different depths. Thick arrow shows the location of the gravity core in the marginal depression.
recta, Cellanthus, and Pseudorotalia schroeteriana are rare. Planktic for-aminifers such as Sphaeroidinella dehiscens, Orbulina universa, Pullen-iatina finalis, and Pulleniatina obliqueloculata are also found.
Petrology of the Phosphorites
Microstructure.—Thin sections of phosphorites exhibit light browncryptocrystalline phosphate matrices and detrital particles. Micro-laminatedphosphatized algal mats consist of mechanically deposited clastic particlesor of phosphatic molds appearing as smooth light to dark brown bands(Fig. 4A, B). Clastic particles are either bound to algal laminae or to dark
phosphatic laminae (Fig. 4B–D). The longer axes of clastic particles areoriented parallel to the laminae. Clay- and silt-size terrigenous and biogenicparticles are disseminated throughout the anisotropic phosphatic matrix.Algal fragments and filaments are abundant in the matrix (Fig. 4E). Micro-unconformities were found adjacent to the patches of smooth phosphaticmatrix (Fig. 4F, G). At places the laminae are indistinct, but the clasticparticles are still aligned (Fig. 4H). Sediment particles bound by microfil-aments appear as coated grains. Unphosphatized benthic foraminifers, ben-thic and planktic foraminifers infilled with green and opaque material, algalfragments, quartz, feldspar, and fragmented fine shell material are also pre-sent in the matrix (Fig. 4B–H). Some glauconite grains seem to be abraded

1200 V.P. RAO ET AL.
FIG. 3.—Morphology of the phosphorites: A) Irregular, B) subrounded, and D) flat pebbles of gray dense phosphorites; C, E) Phosphate internal molds of molluscs;encrusted barnacles and coral polyps can be seen in Parts D and E; F–H) Polished sections showing grain-supported texture and irregular algal coating at the peripheralparts; pelecypod embedded in phosphorite can also be seen in Part G; I) worn shell fragments associated with phosphorites.
or fractured, and others show peripheral alteration to reddish brown col-oration. The surfaces of carbonate skeletals are also ferruginized. Exceptin a few foraminifers, whose chambers are filled with phosphatized mate-rial, the boundaries between clastic particles and the adjacent phosphaticmatrix are conspicuous. Microfossils embedded in phosphorites are thesame as that in associated sediments. Sphaeroidinella dehiscens and Pul-leniatina obliqueloculata and several species of Uvigerina, especially Uvi-gerina schenki, are common. The thin laminae at the peripheral parts ofthe phosphate intraclasts are also of algal origin.
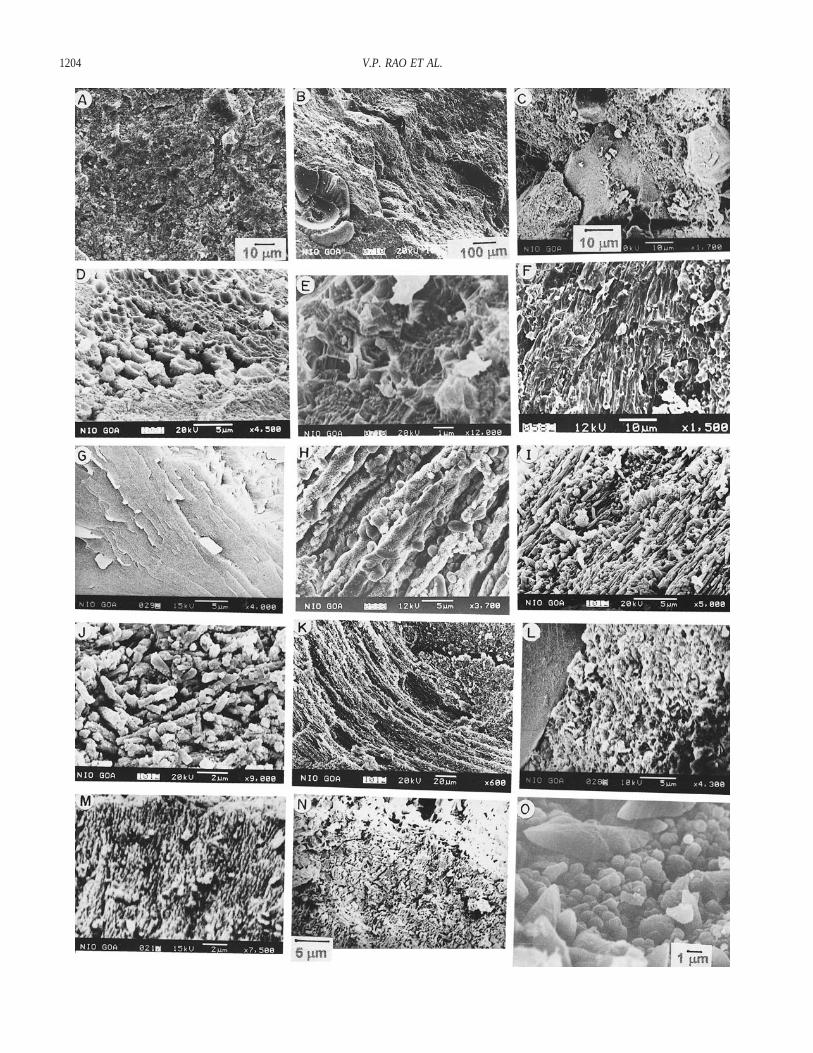
Ultrastructure.—SEM studies on freshly broken surfaces of the densegray phosphorites revealed compact internal structures of phosphates con-sisting of several fine- and coarse-grained clastic particles (Fig. 5A). Lam-inated phosphate matrix is characteristic of phosphorites, which can occurin three distinct types. In the first type, phosphatic laminae are about 10–400 mm thick, with detrital particles interspersed between the laminae (Fig.5B, C). On further magnification, these laminae are seen to be composedof tightly packed to diffuse hollow apatite globules ; 1 mm in diameterthat are oriented parallel to the lineation (Fig. 5D). The globules resemblemicrobial cells. This arrangement of cells is similar to a biofilm, whereinmicrobial cells form a thin sheet around a grain (Krumbein 1986). Someof these cells exhibit double-walled structure (Fig. 5E). In the second type,
the laminated matrix consist of densely packed intertwined, phosphatizedultrafine filaments with parallel alignment (Fig. 5F–I). Some of these appearto be hollow. Similar smaller isodiametric, nonbranching filaments result-ing in dense mats have been reported in stromatolites (Jones et al. 1997).Phosphate globules adhere to sheaths of some of these filaments (Fig. 5H).Molds of phosphatic filaments ; 10 mm in diameter are rare. Adjacentareas show partly decomposed filaments with fine detritus. In the third type,submicron-size phosphate tubules follow obscure lamination (Fig. 5J) thatcorresponds to less dense matrix. Some tubules also show granular surfacetexture. Detrital carbonate particles are wrapped by hollow phosphatic fil-aments ; 10 mm in diameter (Fig. 5K). Microlaminated phosphate wrapsthe grains in the Negev phosphorites as phosphatized remains of organicmat (Soudry and Champetier 1983), which could also be seen in Figure5L. The matrix consists of erect phosphate filaments and filaments withobscured internal structure (Fig. 5M, N), similar to but smaller than thatin Recent Phormidium (Walter 1976). Clustered apatite globules form athin lamina between the detrital grains (Fig. 5O).
In porous areas, the phosphate is mostly characterized by a cluster ofcell-like units of apatite globules 1 to 4 mm in size (Fig. 6A). The apatiteglobules aggregate into larger globose bodies (Fig. 6B, C) that are inter-nally hollow. Clustering of cell-like units (Fig. 6B) resembles the colonies
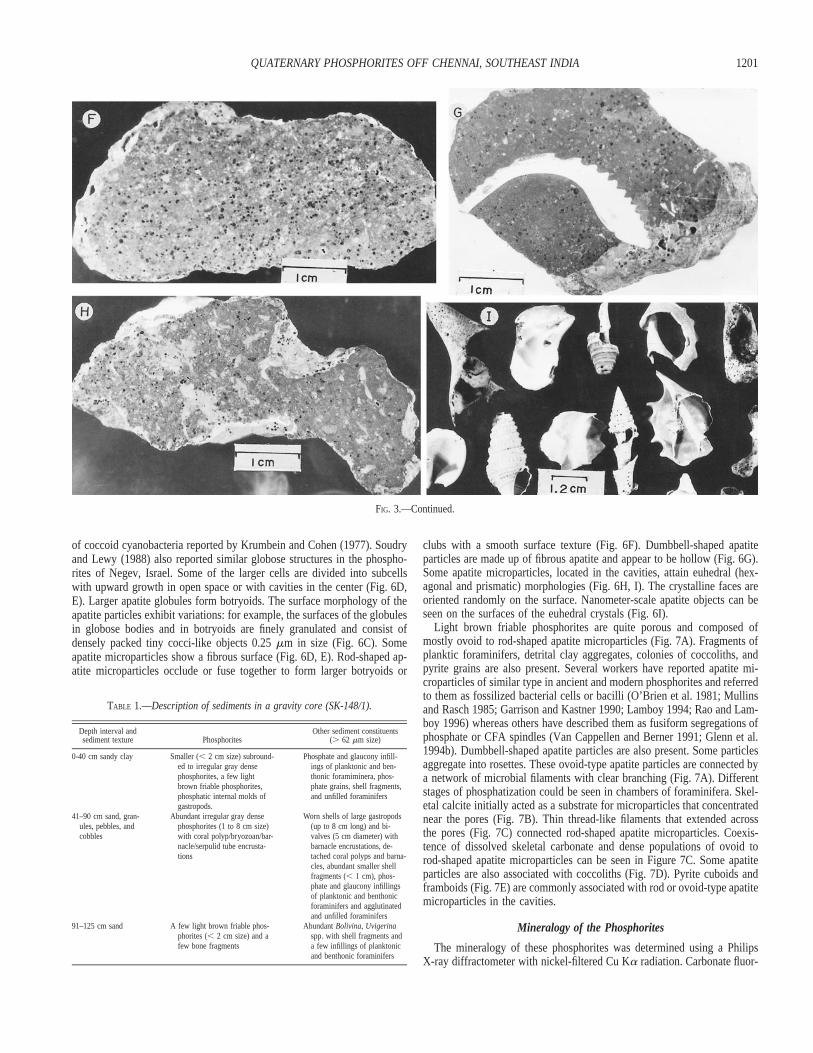
1201QUATERNARY PHOSPHORITES OFF CHENNAI, SOUTHEAST INDIA
FIG. 3.—Continued.
TABLE 1.—Description of sediments in a gravity core (SK-148/1).
Depth interval andsediment texture Phosphorites
Other sediment constituents(. 62 mm size)
0-40 cm sandy clay Smaller (, 2 cm size) subround-ed to irregular gray densephosphorites, a few lightbrown friable phosphorites,phosphatic internal molds ofgastropods.
Phosphate and glaucony infill-ings of planktonic and ben-thonic foramiminera, phos-phate grains, shell fragments,and unfilled foraminifers
41–90 cm sand, gran-ules, pebbles, andcobbles
Abundant irregular gray densephosphorites (1 to 8 cm size)with coral polyp/bryozoan/bar-nacle/serpulid tube encrusta-tions
Worn shells of large gastropods(up to 8 cm long) and bi-valves (5 cm diameter) withbarnacle encrustations, de-tached coral polyps and barna-cles, abundant smaller shellfragments (, 1 cm), phos-phate and glaucony infillingsof planktonic and benthonicforaminifers and agglutinatedand unfilled foraminifers
91–125 cm sand A few light brown friable phos-phorites (, 2 cm size) and afew bone fragments
Abundant Bolivina, Uvigerinaspp. with shell fragments anda few infillings of planktonicand benthonic foraminifers
of coccoid cyanobacteria reported by Krumbein and Cohen (1977). Soudryand Lewy (1988) also reported similar globose structures in the phospho-rites of Negev, Israel. Some of the larger cells are divided into subcellswith upward growth in open space or with cavities in the center (Fig. 6D,E). Larger apatite globules form botryoids. The surface morphology of theapatite particles exhibit variations: for example, the surfaces of the globulesin globose bodies and in botryoids are finely granulated and consist ofdensely packed tiny cocci-like objects 0.25 mm in size (Fig. 6C). Someapatite microparticles show a fibrous surface (Fig. 6D, E). Rod-shaped ap-atite microparticles occlude or fuse together to form larger botryoids or
clubs with a smooth surface texture (Fig. 6F). Dumbbell-shaped apatiteparticles are made up of fibrous apatite and appear to be hollow (Fig. 6G).Some apatite microparticles, located in the cavities, attain euhedral (hex-agonal and prismatic) morphologies (Fig. 6H, I). The crystalline faces areoriented randomly on the surface. Nanometer-scale apatite objects can beseen on the surfaces of the euhedral crystals (Fig. 6I).
Light brown friable phosphorites are quite porous and composed ofmostly ovoid to rod-shaped apatite microparticles (Fig. 7A). Fragments ofplanktic foraminifers, detrital clay aggregates, colonies of coccoliths, andpyrite grains are also present. Several workers have reported apatite mi-croparticles of similar type in ancient and modern phosphorites and referredto them as fossilized bacterial cells or bacilli (O’Brien et al. 1981; Mullinsand Rasch 1985; Garrison and Kastner 1990; Lamboy 1994; Rao and Lam-boy 1996) whereas others have described them as fusiform segregations ofphosphate or CFA spindles (Van Cappellen and Berner 1991; Glenn et al.1994b). Dumbbell-shaped apatite particles are also present. Some particlesaggregate into rosettes. These ovoid-type apatite particles are connected bya network of microbial filaments with clear branching (Fig. 7A). Differentstages of phosphatization could be seen in chambers of foraminifera. Skel-etal calcite initially acted as a substrate for microparticles that concentratednear the pores (Fig. 7B). Thin thread-like filaments that extended acrossthe pores (Fig. 7C) connected rod-shaped apatite microparticles. Coexis-tence of dissolved skeletal carbonate and dense populations of ovoid torod-shaped apatite microparticles can be seen in Figure 7C. Some apatiteparticles are also associated with coccoliths (Fig. 7D). Pyrite cuboids andframboids (Fig. 7E) are commonly associated with rod or ovoid-type apatitemicroparticles in the cavities.
Mineralogy of the Phosphorites
The mineralogy of these phosphorites was determined using a PhilipsX-ray diffractometer with nickel-filtered Cu Ka radiation. Carbonate fluor-

1202 V.P. RAO ET AL.
FIG. 4.—Thin-section micrographs: A) phosphatized algal laminae; B) phosphate mold of a mat; C) dark brown laminae with adhered clastic particles; D) clastic particlestrapped by the laminae; E) algal fragments and microfilaments in the matrix; F, G) micro-unconformities (arrows); H) aligned clastic particles at intervals with indistinctlaminae.

1203QUATERNARY PHOSPHORITES OFF CHENNAI, SOUTHEAST INDIA
apatite and calcite were the major mineral phases in light brown friablephosphorites, gray dense phosphorites, and phosphorite grains (Fig. 8).High-magnesium calcite was also present in gray dense phosphorites andwas the predominant phase in phosphatic molds of macrofossils. Quartz,feldspar, and aragonite were the dominant accessory minerals in bulk sam-ples. The peripheral algal coating was found to be rich in high-magnesiumcalcite. The CO2 content of apatite, determined by the peak pair methodof Gulbrandsen (1970) and Schuffert et al. (1990), varied from 4.5 to 5.5%and the a cell dimension of the apatite was 9.32 A. Relatively high CO2
values indicate carbonate substitution for phosphate in the apatite structure.The acid-insoluble residue was variable in phosphorites, with the highestvalue (21.5%) in gray dense phosphorite followed by that in light brownfriable phosphorites (20%), phosphorite grains (16%), and phosphaticmolds of macrofossils (8.5%). Goethite, illite, quartz, and feldspar werepresent in the acid-insoluble residue. The P2O5 content of the phosphoritevaried from 5 to 30%, and was higher (27 to 30%) in light brown friablephosphorites and gray dense phosphorites than in phosphorite grains (11%)and phosphatic macro-molds (5%) (Vaz 1995; Rao et al. 1998).
DISCUSSION
Age and Type of Phosphorites
The age of Sphaeroidinella dehiscens and Pulleniatina obliqueloculataranges from Pliocene to Recent. Absence of Pliocene markers may be takento suggest a Pleistocene to Recent age.
The presence of laminated microbial mats is a characteristic feature ofthese gray dense phosphorites. The laminated fabrics of these mats devel-oped from trapping and binding of clastic particles (Figs. 4A–D, 5B, C).Mineral precipitation can also produce lamination. Both fungi and cyano-bacteria and other types of bacteria build microbial mats. The laminaedescribed here comprise thin sheets of microbial cells (biofilm; Fig. 5D,E) or straight microbial filaments or tubules (Fig. 5F, K). Because branch-ing of filaments is common in fungi, these relatively straight filaments arelikely related to cyanobacteria. Diameters of phosphatic filaments and phos-phatic tubules (2 to 10 mm) are within the size range of cyanobacteria.Moreover, the erect phosphate filaments (Fig. 5M, N) and colonies of bac-terial cells (Figs. 5O; 6A–C) indicate their possible affinity to cyanobac-teria. Cyanobacteria have been reported to be laminae builders (Soudry andPanczer 1994). The uniform size of phosphate filaments and their stacking(Fig. 5G–I, 6K) suggest precipitation of phosphate within the mat. Glau-conite grains, carbonate skeletals (Fig. 4), quartz, and feldspar on the sur-faces of different laminae suggest alternate periods of mat growth and grainsupply. Following the definition of Walter (1976) for stromatolites, thefeatures described above correspond to phosphate stratiform stromatolitesor phosphate cryptalgal laminites. The phosphate stromatolites reported forPaleozoic (Bertrand-Sarfati et al. 1997) and Mesozoic times (Krajewski etal. 1994; Martin Algarra and Sanchez-Navas 1995; Follmi 1996) exhibitmostly columnar macrostromatolitic or microstromatolitic structures wherecyanophytes are the primary group responsible for their formation. Awra-mik et al. (1976) reported changes in fossil morphotypes of microbiotathrough Precambrian to modern times and modern microbial mats com-posed of communities of cyanophytes, eucaryotic algae, fungal structures,and phytosynthetic bacteria. That these stratiform stromatolites do not sharethe columnar structures of older phosphate stromatolites may be becauseof the different microbial forms inhabited. The phosphate stratiform stro-matolites reported here are the first to be reported during the Quaternaryand modern times.
Depositional Environment
Phosphate stromatolites have been reported in both shallow coastal set-tings and deeper dysaerobic environments with low net sedimentation rates,current action, and ample supply of organic matter (Bertrand-Sarfati et al.
1997; Martin-Algarra and Sanchez-Navas 1995). Abundant fine and coarsedetrital material in the mats (Fig. 4) indicates that mat formation may notbe characteristic of low sedimentation rates and deeper waters. The micro-unconformities (Fig. 4F) probably suggest that these were periods of sed-iment starvation, stromatolite growth, and episodic flux of sediments. Thelive Uvigerina and Hoeglundina occur on the outer shelf or upper slope,and their characteristic depth range is around 100–120 m (Murray 1991).Their presence in phosphorites implies that the microbial mats may haveoriginally accumulated and been phosphatized at around these depths, orsubsequently when outer shelf experienced conditions of low sea level andhigh energy during one of the glacial stages of the Pleistocene. We favorthe latter possibility, for the following reasons. (1) The constituents of thephosphorites include (already) glauconitized molds and infillings of fora-minifers and their broken fragments (Fig. 3), fresh benthic skeletals (Cel-lanthus and Pseudorotalia), and algal fragments (Fig. 4E) of shallow-waterorigin, implying that these constituents were transported to the site underagitated conditions. (2) The phosphatized mats, once formed, were dis-rupted into irregular phosphate clasts (Fig. 3A). (3) Encrusted barnacles(Fig. 3D), coral polyps (Fig. 3E), and high-magnesium-calcite-rich algaearound phosphate intraclasts (Fig. 3F–H) indicate current-swept environ-ments even after phosphatization. (4) Because abundant phosphorites occurat subsurface depths (Table 1) and are associated with large shells, wornshells, and shell fragments (Fig. 3I) of shallow-water origin, it is also nec-essary to accommodate reworking of the phosphorites into the shelf-margindepressions (185–325 m). This implies that the phosphorites were initiallyformed at depths between 60 and , 185 m and in association with high-energy conditions (see points 1, 2, and 3 above). We therefore suggest thatthe microbial mats formed at the outer shelf, possibly when it was subjectedto conditions of low sea level. Middle rare earth element (MREE)–enrichedpatterns (Rao et al., unpublished data) provide evidence that these phos-phate stratiform stromatolites have originated in a coastal setting dominatedby currents. In shallow marine settings, calcification of stromatolites isusually far more widespread than phosphatization. In order to understandthe formation of phosphate stromatolites, we will evaluate the sources ofphosphorus, processes of formation, and environmental conditions duringformation.
Sources of Phosphorus
Microbial mats (Figs. 4, 5) were abundant in the phosphorites. Hence,decomposition of organic mats might have been a local pore-water sourceof phosphorus. The presence of iron-oxide coatings on minerals and ske-letals, altered glauconites, and goethite in the phosphorites indicates thatthese may have provided some phosphate adsorbed onto FeOOH, which isreleased to the pore waters during Fe-redox cycling (see Berner 1990;O’Brien et al. 1990). The presence of illite and other clay particles inphosphorites could also be a source of phosphorus, because the phosphateadsorption capacity of illite far exceeds that of montmorillonite and kao-linite (Edzwald et al. 1976). Bremner (1980) showed that the phosphateadsorbed on mica and clay particles is a source for Namibian phosphorites.We assume that phosphatization of microbial mats occurred on the outershelf during conditions of lowered sea level in the Pleistocene (see above).Because Cretaceous phosphorites occur on the coastal region of Chennaiand outcrops of Cretaceous faunal elements have been identified on theshelf (Vaz 1995), it is likely that the intense subaerial weathering of theseformations may have supplied some phosphorus to the site of phosphoriteformation. Abundant algal fragments (Fig. 4E) present in phosphorites mayhave supplied some phosphorus. Pevear (1966) indicated that high produc-tivity of benthic algae in shallow-water conditions provide high phosphorussupply to the sediments. Follmi et al. (1993) documented nonlinearities inphosphogenesis for different periods originating in sedimentary settingsranging from nearshore to slope environments. He suggested that the in-creased continental weathering rates mobilize continental phosphorus and
1204 V.P. RAO ET AL.

1205QUATERNARY PHOSPHORITES OFF CHENNAI, SOUTHEAST INDIA
FIG. 6.—A) Globular apatite microparticles similar to microbial cells in the porous area; B) cell-like units grouped into globose bodies resembling cyanobacterial colonies;C) high-magnification of Part B showing central cavity and granular surface texture; D, E) apatite particles showing fibrous (D) to granular (E) surface texture and centralcavity; F, G) rod-shaped to dumbbell-shaped apatite particles with smooth to fibrous texture; H, I) apatite microparticles within cavity showing randomly oriented apatitecrystals; nanometer-size apatite objects can also be seen on apatite particles.
←
FIG. 5.—Ultrastructures of gray dense phosphorites: A) dense phosphate matrix with fine clastic particles; B, C) thick phosphate laminae with intermittent clay andcarbonate particles; D) mat showing a thin sheet formed by the arrangement of closely packed globular apatite particles; E) microbial cells showing double wall structure;F–I) straight to intertwined phosphatized microbial filaments forming laminae; J) phosphate tubules; K) bundle of phosphate filaments wrapping the grain; L) thickphosphate coating around the grain; M, N) erect filaments similar to Phormidium; O) cluster of apatite microparticles forming a microlamina between clastic particles.
accelerate flux rates of phosphorus into the oceans. This transfer of P maysometimes be coupled with episodic events in which large benthic microbialcommunities are buried by bypassing sediments and consequently are rapidlyphosphatized. A similar mechanism may have supplied abundant phosphorus.The remote sources of phosphorus include upwelling. Although weak up-welling occurs on this margin at present, there might have been intense up-welling or warm-water currents (inducing productivity) close to the coastduring the Pleistocene, when ocean circulation was somewhat vigorous. Theconcentrations of certain trace elements (Ba, Ni, Cu, Zn, and V) in ourphosphorites, however, are lower than in the average shale, average phos-phorite, and phosphorites and phosphate stromatolites formed under the in-
fluence of upwelling (Rao et al., unpublished work). This indicates that plank-ton organic matter may not be the prime source of metals. Sr and Nd isotopesof these phosphorites also indicate excess influx of source metals and eutro-phic conditions in a confined environment (Rao et al., unpublished work).We therefore assume that abundant particulate and dissolved P and otherelements (derived from continental sources and reworking of the shelf) wereavailable at the time of phosphorite formation.
Processes of Phosphatization
Two processes are involved in phosphate precipitation. The first is re-lated to extracellular precipitation of apatite. Hollow microbial filaments

1206 V.P. RAO ET AL.
FIG. 7.—Ultrastructures of light brown friable phosphorites. A) Ovoid to rod-shaped apatite microparticles; a microfilament showing branched pattern, resembling fungalstructure can also be seen; B, C) foraminiferal chamber showing apatite microparticles located close to the skeletal pores (B), carbonate substrate is dissolved and particlesextend across the pores; thick population of apatite particles replacing the carbonate substrate in Part C; D) molding of coccoliths by apatite microparticles; E) framboidpyrite in association with rod-shaped apatite microparticles.
and cells (Fig. 5D, E, F, I, K), and globose bodies with a central cavity(Fig. 6B–E) may represent phosphatic external molds of microbial bodies.The distinct and well preserved structures, however, suggest that phosphateprecipitation might have taken place very soon after mat organisms diedand prior to complete decay of the organic compounds. The exact timingof precipitation is controversial. Krumbein (1986) proposed that decay oforganic matter associated with microbial mats is necessary; this raises pH,and reducing microenvironmental conditions developed within the mat mayfavor precipitation. On the other hand, Martill (1994) proposed that instan-taneous autophosphatization can occur within a few hours or days after thedeath of the organisms. Experimental evidence suggests that precipitationcan occur essentially instantaneously via an amorphous phosphatic phasethat later recrystallizes to apatite (Gulbrandsen et al. 1984; Krajewski et al.1994). Amorphous calcium phosphate is stable in the presence of organicsubstances of the bacterial sheaths, especially polysaccharides. Phosphatecan nucleate in the extracellular mucilage of bacteria. Krumbein (1986)suggested that the chemo-organotrophic bacteria living within and on thecyanobacterial glycocalyx actually initiate mineralization. Martin Algarraand Sanchez-Navas (1995) showed colloform morphologies of the phos-phate stromatolites and suggested that the mucilaginous sheath has provid-ed a substrate facilitating rapid precipitation of amorphous phosphate phas-
es. They further stated that the stromatolite-forming bacteria are capableof adsorbing and concentrating dissolved phosphate from degraded organicmatter and seawater, and can lead to PO4
3- supersaturation required toprecipitate an amorphous precursor. Thus phosphate nucleation and precip-itation can probably begin prior to substantial decay of the microbial mats,as suggested by the microtextural details preserved in the phosphorites. Thedifferent morphology of the filaments and surface textures of the microbialfilaments and cells (Fig. 6C–F) may be due to the nature and chemicalconstituents of the organic structures associated with the sheath. Pentecostand Riding (1986) suggested that sheath development is environmentallyinduced, and that sheath thickness (2–10 mm) and properties vary widelyand could be species-specific. Submicron-size cocci-like granules on thesurfaces of larger particles (Fig. 6C) and nanometer-scale objects found onthe crystal faces of apatite (Fig. 6I) may be inorganic, although earlierworkers reported these objects as nannobacteria (Soudry and Lewy 1988).Experimental studies by Kirkland et al. (1999) demonstrated that, both insterile conditions and in the presence of dissolved organic compounds,calcite formed euhedral crystals 50–800 nm in diameter and smaller , 50nm anhedral or rounded particles or protocrystals. The , 50 nm particlesare, however, more common in minerals formed in organic-rich environ-ments. Kirkland et al. (1999) interpreted these smaller rounded nanno-fea-

1207QUATERNARY PHOSPHORITES OFF CHENNAI, SOUTHEAST INDIA
FIG. 8.—X-ray diffractograms of phosphorites. A) Light brown friable phosphorite; B) Gray dense phosphorite; C) phosphate macro-mold; D) Phosphorite grains. CFA,carbonate fluorapatite; Lmc, low-magnesium calcite; Hmc, high-magnesium calcite; Ara, aragonite; Qtz, quartz.

1208 V.P. RAO ET AL.
tures as inorganic precipitates. It is not understood why only some micro-bial bodies exhibited crystalline faces (Fig. 6H, I); it could be due to theconfinement of the environment rather than to aging, as suggested by Gul-brandsen et al. (1984). Phosphatization of microbial mats was a contem-poraneous process, and thus phosphate deposition was a primary depositrather than a carbonate replacement. Because of the episodic flux of sedi-ments onto microbial mats, microorganisms associated with the mat playeda major role in fixing dissolved P, regulating microenvironment at the sed-iment–water interface, and precipitating phosphate. Several workers (Soud-ry and Champetier 1983; Southgate 1986; Soudry and Lewy 1988) havereported microbial influences in phosphate mineralization. Apatite has beensynthesized from organic matter in nonsterile conditions, suggesting thatbacterial mediation is necessary in apatite mineralization (Lucas and Prevot1984; Prevot et al. 1989; Hirschuler et al. 1990).
The second process of phosphate precipitation is evidenced by the elon-gated forms of bacilli types (rod-shaped) and ovoid types of apatite micro-particles, which occur in all types of phosphorites but are most abundantin light brown friable phosphorites (Fig. 7). These particles do not showcentral cavities, and phosphate may have precipitated intracellularly.O’Brien et al. (1981) considered the cell-like structures to be a post-mortemtransformation of slowly growing P-rich bacterial cells. Several others re-ported similar type of particles in both ancient and modern phosphoritesand regarded them as fossilized microbes (Mullins and Rasch 1985; Gar-rison and Kastner 1990; Lamboy 1994; Rao and Lamboy 1996), which arecapable of concentrating phosphate intracellularly. Some workers, however,have considered these particles to be inorganic pellets resulting from rapidnucleation of phosphate in the pore water (Glenn et al. 1994b). Van Cap-pellen and Berner (1991) demonstrated experimentally that such crystalscan be grown in sea-water-like solutions under sterile conditions. The factthat these particles were connected by a framework of branched filamentsand cells containing flagellates at their tips (Fig. 7A, C) argues for a mi-crobial origin. Pyrite cuboids on top of cell-like structures and pyrite fram-boids (Fig. 7E) indicate reducing conditions. It is known that under oxicconditions, microbes tend to accumulate and store dissolved phosphate asan end product of phosphorylation and release substantial amounts of phos-phate to the interstitial waters upon transfer to the anoxic environment(Gachter and Mayer 1993). Similarly, microbiota modify the chemistry ofthe pore-water environment and affect the localization of apatite precipi-tation in sediments. Different stages showing sparsely populated (Fig. 7B)to somewhat densely populated phosphatized bacteria (Fig. 7C) within theforaminiferal chamber and their intimate connection with the substrate sug-gest that these microbes not only control the microenvironment but alsohelp to modify the substrate by leading to its dissolution and then fillingthe cavities.
The ultrastructures reported here are similar to those of the Cambrianphosphate stromatolites of west Africa and phoscrete profiles in Australia(Bertrand-Sarfati et al. 1997; Southgate 1986), Late Cretaceous and Eoceneshallow-water phosphorites in the Negev, Israel (Soudry and Lewy 1988;Soudry and Panczer 1994), and phosphate stromatolites reported in pelagicsettings (Martin-Algarra and Sanchez-Navas 1995). Similar microbial bod-ies have also been found in many Recent phosphorites from continentalmargins (O’Brien et al. 1981; Lamboy 1990; Rao and Lamboy 1995, 1996).These findings suggest that phosphate might have been precipitated directlyon microbial bodies.
The structural CO2 content of the apatite is considered to be an indicatorof water temperature at the time of formation, with higher CO2 values (4–6%) at higher temperatures (Gulbrandsen 1970). The CO2 contents in thephosphorites were 4 to 5.5%, and thus these might have formed under warmshallow shelfal conditions. Alternately, higher CO2 values indicate replace-ment-type phosphorites whereas lower values indicate authigenic phospho-rites (Gulbrandsen 1970). Contrastingly, higher CO2 values here are asso-ciated with authigenic phosphorites, suggesting that the CO2 content ofapatite is not a reliable indicator for determining the authigenic and re-
placement-type phosphorites. Birch (1980) also argued against a close re-lationship between the CO2 content and type of phosphorite. Glenn et al.(1988) correlated the CO2 content of apatite with carbonate alkalinity inthe pore waters and suggested that higher carbonate alkalinity and slowprecipitation rates favor incorporation of carbonate (see also Jarvis et al.1994). Carbonate alkalinity is expected to be higher at the site of phos-phorite formation because of shallow shelfal conditions and carbonate dis-solution (Fig. 7B, C). Petrography indicates, however, that the phosphatewas precipitated rapidly on the organic remains. The utility of CO2 contentof the apatite as an indicator therefore needs reevaluation.
SUMMARY
The Paleozoic (Bertrand-Sarfati et al. 1997), Mesozoic (Martin-Algarraand Sanchez-Navas 1995), and Paleogene (Soudry and Panczer 1994) phos-phate stromatolites and Quaternary phosphate stromatolites reported hereare all primary phosphate deposits. Despite their occurrence at differentdepths on the paleo-margins, the processes that control phosphate stromat-olite formation appear to be similar. The stratiform phosphate stromatolitesreported here, however, do not share the macroscopic or microscopic co-lumnar habit of the ancient phosphate stromatolites. The planar cryptalgallaminites are considered to be precursors of columnar stromatolites. Thiswork therefore reveals that the replicates (analogs) of ancient phosphoritesdo exist in the Quaternary. These phosphorites were formed at conditionsof low sea level on the outer shelf, and upwelling may not be the primarymechanism that supplied P to the site of phosphorite formation. Continentalsupply seems to be the major source of phosphorus. The primary phos-phorites thus formed were disrupted into phosphate clasts and subsequentlytransported to the depression at 186–300 m. As a result, the sediments inthe depression consist of an admixture of phosphate clasts. larger shells,rhodoliths, phosphate, and glaucony-dominated sandy sediments. Thesephosphorites are thus similar to condensed Mesozoic phosphorite depositsfrom the Mediterranean paleo-margins (Follmi 1990, 1996; Martin-Algarraand Sanchez-Navas 1995).
ACKNOWLEDGMENTS
We thank Dr. E. Desa, Director, National Institute of Oceanography, Goa for theencouragement. Dr. M. Dileep Kumar has gone through the manuscript and offeredsuggestions. This work was financed by the Department of Science and Technology,New Dehli, project no. ESS/CA/A1-25/97. We thank JSR reviewers Dr. David Kid-der, Dr. Dawn Sumner, and Dr. Gabriel Filippelli for their valuable comments. Thisis NIO contribution no. 3536.
REFERENCES
AWRAMIK, S.M., MARGULIS, L., AND BARGHOORN, E.S., 1976, Evolutionary processes in the for-mation of stromatolites, in Walter, M.R., ed., Stromatolites: Amsterdam, Elsevier, Devel-opments in Sedimentology 20, p. 149–162.
BATURIN, G.N., 1982, Phosphorites on the Sea Floor: Origin, Composition and Distribution:Amsterdam, Elsevier, Developments in Sedimentology 33, 345 p.
BENTOR, Y.K., 1980, Phosphorites—unsolved problems, in, Bentor, Y.K., ed., Marine Phos-phorites: Society of Economic Paleontologists and Mineralogists, Special Publication 29, p.3–18.
BERNER, R.A., 1990, Diagenesis of phosphorus in sediments from non-upwelling areas, inBurnett, W.C., and Riggs, S.R., eds., Phosphate Deposits of the World, vol. 3: Neogene toModern Phosphorites: Cambridge, U.K., Cambridge University Press, p. 27–32.
BERTRAND-SARFATI, J., FLICOTEAUX, R., MOUSSINE-POUCHKINE, A., AND AHMED, A.A.K., 1997, Low-er Cambrian apatitic stromatolites and phospharenites related to the glacio-eustatic cratonicrebound (Sahara, Algeria): Journal of Sedimentary Research, v. 67, p. 957–974.
BIRCH, G.F., 1980, A model of penecontemporaneous phosphatization by diagenetic and au-thigenic mechanisms from the western margin of southern Africa, in Bentor, Y. K., ed.,Marine Phosphorites: Society of Economic Paleontologists and Mineralogists, Special Pub-lication 29, p. 79–100.
BREMNER, J.M., 1980, Concretionary phosphorite from SW Africa: Geological Society of Lon-don, Journal, v. 137, p. 773–786.
BURNETT, W.C., 1977, Geochemistry and origin of phosphorite deposits from off Peru andChile: Geological Society of America, Bulletin, v. 88, p. 813–823.
BURNETT, W.C., AND RIGGS, S.R., 1990, Phosphate Deposits of the World, vol. 3: Neogene toModern Phosphorites: Cambridge, U.K., Cambridge University Press, 464 p.

1209QUATERNARY PHOSPHORITES OFF CHENNAI, SOUTHEAST INDIA
COOK, P.J., AND SHERGOLD, J.H., 1986, Phosphate Deposits of the World: Proterozoic and Cam-brian Phosphorites: Cambridge, U.K., Cambridge University Press, 385 p.
EDZWALD, J.K., TOENSING, D.C., AND LEUNG, M.C., 1976, Phosphate adsorption reactions withclay minerals: Environment Science Technology, v. 10, p. 485–490.
FILIPPELLI, G.M., AND DELANEY, M.L., 1992, Similar phosphorus fluxes in ancient phosphoritedeposits and a modern phosphogenic environment: Geology, v. 20, p. 709–712.
FOLLMI, K.B., 1990, Condensation and phosphogenesis: example of the helvetic mid-Cretaceous(Northern Tethyan margin), in Notholt, A.J.G., and Jarvis, I., eds., Phosphorite Researchand Development: Geological Society of London, Special Publication 52, p. 237–252.
FOLLMI, K.B., 1996, The phosphorous cycle, phosphogenesis and marine phosphate-rich de-posits: Earth-Science Reviews, v. 40, p. 55–124.
FOLLMI, K.B., WEISSERT, H., AND LINI, A., 1993, Nonlinearities in phosphogenesis and phos-phorus–carbon coupling and their implications for global change, in Wollast, R., Mackenzie,F.T., and Chou, L., eds., Interactions of C, N, P, and S Biogeochemical Cycles and GlobalChange: NATO ASI Series, 14, Berlin, Springer-Verlag, p. 447–474.
GACHTER, R., AND MEYER, J.S., 1993, The role of microorganisms in mobilisation and fixationof phosphorous in sediments: Hydrobiologia, v. 253, p. 103–121.
GARRISON, R.E., AND KASTNER, M., 1990, Phosphatic sediments and rocks recovered from thePeru margin during ODP Leg 112, in Suess, E., Von Huene, R., et al., eds., Ocean DrillingProgram, Proceedings, Scientific Results Leg 112: College Station, Texas, v. 112, p. 111–134..
GLENN, C.R., ARTHUR, M.A., YEH, H.W., AND BURNETT, W.C., 1988, Carbon isotope compositionand lattice bound carbonate of Peru–Chile margin phosphorites: Marine Geology, v. 80, p.287–307.
GLENN, C.R., FOLLMI, K.B., RIGGS, S.R., BATURIN, G.N., GRIMM, K.A., TRAPPE, J., ABED, A.M.,GALLI-OLIVIER, C., GARRISON, R.E., ILYIN, A., JEHL, C., ROHRLICH, V., SADAQAH, R., SCHID-LOWSKI, M., SHELDON, R.E., AND SIEGMUND, H., 1994a, Phosphorus and phosphorites: Sedi-mentology and environment of formation: Eclogae Geologicae Helvetiae, v. 87, p. 747–788.
GLENN, C.R., ARTHUR, M.A., RESIG, M.J., BURNETT, W.C., DEAN, W.E., AND JAHNKE, R.A., 1994b,Are modern and ancient phosphorites really so different?, in Iijima, A., Abed, A.M., andGarrison, R.E., eds., Phosphatic and Glauconitic Sediments of the Tertiary and Mesozoic:29th International Geological Congress, Proceedings, Part C; Utrecht, VSP, p. 159–188.
GULBRANDSEN, R.A., 1970, Relation of carbon dioxide content of apatite of the PhosphoriaFormation to regional facies: U.S. Geological Survey, Professional Paper 700-B, p. 9–13.
GULBRANDSEN, R.A., ROBERTSON, C.E., AND NEIL, S.T., 1984, Time and crystallisation of apatitein seawater: Geochimica et Cosmochimica Acta, v. 48, p. 213–218.
HIRSCHULER, A., LUCAS, J., AND HUBERT, J.C., 1990, Apatite genesis: a biologically induced orbiologically controlled mineral formation process: Geomicrobiology, v. 7, p. 47–57.
JARVIS, I., BURNETT, W.C., NATHAN, Y., ALMBAYDIN, F., ATTIA, K.M., CASTRO, L.N., FLICOTEAUX,R., HILMY, M.E., HUSAIN, V., QUTAWNA, A.A., SERJANI, A., AND ZANIN, Y.N., 1994, Phospho-rite geochemistry: State-of-the-art and environmental concerns: Eclogae Geologicae Helve-tiae, v. 7, p. 643–700.
JONES, B., RENAULT, R.W., AND ROSEN, M.R., 1997, Biogenicity of silica precipitation aroundgeysers and hot-spring vents, North Island, New Zealand: Journal of Sedimentary Research,v. 67, p. 88–104.
KAZAKOV, A.V., 1937, The phosphorite facies and the genesis of phosphorites: GeologicalInvestigations of Agricultural Ores: Scientific Institute of Fertilizers and Insecto-FungicidesTransactions, v. 142, 95–113.
KIRKLAND, B.L., LYNCH, F.L., RAHNIS, M.A., FOLK, R.L., MOLINEUX, I.J., AND MCLEAN, R.J.C.,1999, Alternative origins for nannobacteria-like objects in calcite: Geology, v. 27, p. 347–350.
KRAJEWSKI, K.P., VAN CAPPELLEN, P., TRICHET, J., KUHN, O., LUCAS, J., MARTIN-ALGARRA, A.,PREVOT, L., TEWARI, V.V., GASPER, I., KNIGHT, R.I., AND LAMBOY, M., 1994, Biological pro-cesses and apatite formation in sedimentary environments: Eclogae Geologicae Helvetiae,v. 87, p. 701–745.
KRUMBEIN, W.E., 1986, Biotransfer of minerals by microbes and microbial mats, in Leadbeater,B.S.C., and Riding, R., eds., Biomineralisation in Lower Plants and Animals: Oxford, U.K.,Oxford University Press, p. 55–72.
KRUMBEIN, W.E., AND COHEN, Y., 1977, Primary production, mat formation and lithification:Contribution of oxygenic and facultative anoxygenic cyanobacteria, in Flugel, E., ed., FossilAlgae: Berlin, Springer-Verlag, p. 37–56.
LAMBOY, M., 1990, Microstructure of a phosphatic crust from a Peruvian continental margin:phosphatized bacteria and associated phenomena: Oceanologica Acta, v. 13, p. 439–451.
LAMBOY, M., 1994, Nannostructure and genesis of phosphorites from ODP Leg 112, the Perumargin: Marine Geology, v. 118, p. 5–22.
LASKAR, B., AND NARASIMHAN, A.S., 1968, Rock phosphate deposits in the Cretaceous rocks ofSouth India: Geological Society of India, Memoir 2, p 105–119.
LUCAS, J., AND PREVOT, L., 1984, Apatite synthesis by bacterial activity from phosphatic organicmatter and several calcium carbonates in natural freshwater and seawater: Chemical Geol-ogy, v. 42, p. 101–118.
MARTILL, D.M., 1994, La fossilisation instantanee: La Recherche, v. 25, p. 996–1002.MARTIN-ALGARRA, A., AND SANCHEZ NAVAS, A. 1995, Phosphate stromatolites from condensed
cephalopod limestones, Upper Jurassic, Southern Spain: Sedimentology, v. 42, p. 893–919.MONTY, C.L.V., 1976, The origin and development of cryptalgal structures, in Walter, M.R.,
Stromatolites: Amsterdam, Elsevier, Developments in Sedimentology 20, p. 193–250.MULLINS, H.T., AND RASCH, R.F., 1985, Sea floor phosphorites along the central California
continental margin: Economic Geology, v. 80, p. 696–715.MURRAY, J.W., 1991, Ecology and Palaeoecology of Benthic Foraminifera: New York, Long-
man, 398 p.O’BRIEN, G.W., HARRIS, J.R., MILNES, A.R., AND VEEH, H.H., 1981, Bacterial origin of East
Australian continental margin phosphorites: Nature, v. 294, p. 442–444.O’BRIEN, G.W., VEEH, H.H., MILNES, A.R., HEGGIE, D.T., RIGGS, S.R., CULLEN, D.J., MARSHALL,
J.F., AND COOK, P.J., 1990, Sedimentation dynamics and redox iron-cycling: Controllingfactors for the apatite–glauconite association on the East Australian continental margin, inNotholt, A.J.G., and Jarvis, I., eds., Phosphorite Research and Development: GeologicalSociety of London, Special Publication 52, p. 61–86.
PENTECOST, A., AND RIDING, R., 1986, Calcification in cyanobacteria, in Leadbeater, B.S.C., andRiding, R., eds., Biomineralisation in Lower Plants and Animals, Oxford, U.K., OxfordUniversity Press, p. 55–72.
PEVEAR, D.R., 1966. The estuarine formation of United States Atlantic coastal plain phospho-rites: Economic Geology, v. 61, p. 251–256.
PREVOT, L., EL FALEH, E.M., AND LUCAS, J., 1989, Details on synthetic apatites formed throughbacterial mediation, mineralogy and chemistry of the products, In Lucas, J., Cook, P.J., andPrevot, L., eds., Apatite et Phosphorites: Sciences Geologie, Bulletin, v. 42, p. 237–254.
RAO, V.P., AND LAMBOY, M., 1995, Phosphorites from the Oman Margin, ODP Leg 117: Ocean-ologica Acta, v. 18, p. 289–307.
RAO, V.P., AND LAMBOY, M., 1996, Genesis of apatite in the phosphatized limestones of thewestern continental shelf of India: Marine Geology, v. 136, p. 41–53.
RAO, V.P., RAO, K.M., VORA, K.H., ALMEIDA, F., SUBRAMANIAM, M.M., AND SOUZA, C.G.A.,1998, A potential phosphorite deposit on the continental margin off Chennai: Current Sci-ence, v. 74, p. 574–577.
SASTRY, V.V., RAJU, A.T.R., SINHA, R.N., VENKATACHALA, B.S., AND BANERJEE, R.K., 1977, Bio-stratigraphy and evolution of the Cauvery Basin, India: Geological Society of India, Journal,v. 18, p. 355–377.
SCHUFFERT, J., KASTNER, M., EMANUELE, G., AND JAHNKE, R.A., 1990, Carbonate-ion substitutionin francolite: A new equation: Geochimica et Cosmochimica Acta, v. 54, p. 2323–2328.
SOUDRY, D., AND CHAMPETIER, Y., 1983, Microbial processes in the Negev phosphorites (south-ern Israel): Sedimentology, v. 30, p. 411–423.
SOUDRY, D., AND LEWY, Z., 1988, Microbially influenced formation of phosphate nodules andmega-fossil moulds (Negev, southern Israel): Palaeogeography, Palaeoclimatology, Palaeoe-cology, v. 64, p. 15–34.
SOUDRY, D., AND PANCZER, G., 1994, Stromatolitic phosphorites in the Eocene of the Negev(Southern Israel), in Bertrand-Sarfati, J., and Monty, C.L.V., eds., Phanerozoic StromatolitesII: Dordrecht, The Netherlands, Kluwer, 255–276.
SOUTHGATE, P.N., 1986, Cambrian phoscrete profiles, coated grains and microbial processes inphosphogenesis, Georgina Basin, Australia: Journal of Sedimentary Petrology, v. 56, p. 429–441.
SUBBARAO, M., 1964, Some aspects of continental shelf sediments off the east coast of India:Marine Geology, v. 1, p. 59–87.
VAN CAPPELLEN, P., AND BERNER, R.A., 1991, Fluorapatite crystal growth from modified seawater solution: Geochimica et Cosmochimica Acta, v. 55, p. 1219–1234.
VAZ, G.G., 1995, Phosphatic nodules in the outer continental shelf off Madras, Bay of Bengal:Indian Journal of Marine Sciences, v. 24, p. 8–12.
WALTER, M.R., 1976, Stromatolites: Amsterdam, Elsevier, Developments in Sedimentology 20,790 p.
Received 2 April 1999; accepted 24 January 2000.




















