
Pulmonary Protein dependent fate of hepatic cells under nicotine induced stress and curcumin...
14
Pulmonary, Gastrointestinal and Urogenital Pharmacology Protein dependent fate of hepatic cells under nicotine induced stress and curcumin ameliorated condition Satyam Banerjee a , Krishna Chattopadhyay b , Jasmeet Kaur Chhabra c , Brajadulal Chattopadhyay a, ⁎ a Department of Physics, Jadavpur University, Raja S.C. Mullick Road, Kolkata-700032, India b Department of Chemical Technology, Calcutta University, 92 A.P.C. Road, Kolkata-700009, India c Immuno Biology Division, Indian Institute of Toxicology Research, M. G. Marg, Lucknow-226001, India abstract article info Article history: Received 1 July 2011 Received in revised form 1 February 2012 Accepted 9 February 2012 Available online 23 February 2012 Keywords: Apoptosis Cell proliferation Curcumin Nicotine Protein-restriction Signaling molecule Nicotine is mainly metabolized in liver. Its abuse elicits acute phase response by activating macrophages to produce pro-inflammatory cytokines, which play critical role in apoptosis or cell proliferation. The protective pharmacological mechanism of curcumin against nicotine-induced toxicity on protein malnourished liver is still remaining unclear. This study investigated the ameliorative mechanism of curcumin against nicotine- induced toxicity and also fate of liver particularly under protein restricted condition. Female Albino-rats maintained under normal/protein-restricted diets, were subcutaneously injected with nicotine tartrate (2.5 mg/kg body weight/day) and orally supplemented with curcumin (80 mg/kg body weight/day) for 21 days. The animals were then sacrificed to dissect out liver and proceed with further experiments. Interac- tions of nicotine with DNA both in vivo and in vitro were observed by thermal denaturation and DNA ladder- ing assays. Effects of nicotine on hepatic cells were monitored by differential staining, comet assay, cytokine profiling, mRNA and protein expression. Nicotine caused more intense DNA damage, promoted hepatic cell death through up-regulating pro-apoptotic proteins and signaling molecules in protein malnourished indi- viduals. Through up-regulation of anti-apoptotic proteins and proliferation promoting molecules, nicotine dysregulated homeostasis in normal protein condition. Curcumin significantly ameliorated the nicotine- induced toxicity in both conditions and regulated the imbalance between cell survival and death induced by nicotine. The protein content present in the nicotine induced hepatic cell decides either cell-survival path- way or cytotoxic pathway. © 2012 Elsevier B.V. All rights reserved. 1. Introduction Large populations in developing countries are deprived of proper nutrition and addicted to tobacco consumption (Ortner et al., 2002). Malnutrition is a recurring problem particularly protein-energy mal- nutrition is widely observed in India and other developing countries (Müller and Krawinkel, 2005). Nicotine, the major addictive alkaloid of tobacco is mainly metabolized in liver (Hukkanen et al., 2005; Nakajima et al., 1998), though its toxic effects on hepatic cells remain unclear. Low dietary protein possesses a constraint on the metabolic activity thereby resulting in impaired detoxification machinery of liver (Nakajima et al., 1998). Based on human autopsy samples from smokers, the highest affinity for nicotine is in the liver, kidney, spleen and lung (Benowitz et al., 2009). Nicotine abuse elicits acute phase response in liver by activating monocytes and macrophages to produce pro-inflammatory cytokines like IL-6, TNF-α, etc.(Song et al., 1999) whose over exposure eventually contribute to acute liver injuries (Hoek and Pastorino, 2002). IL-6 phos- phorylates Signal Transducers and Activators of Transcription (STAT) by activating Janus Kinases (Jaks) (Streetz et al., 2000). Phosphorylated STATs form dimers which function as transcription factors for them- selves and induce expression of one of its member's proteins e.g. STAT 3. The induction of Suppressor of Cytokine Signaling (SOCS) genes is facilitated by the STATs, with several putative STAT-binding sites in dif- ferent SOCS promoter regions. Constitutive activation of STAT 3 has been reported in various carcinomas (Macha et al., 2011; Shukla et al., 2010) whereas over expression of SOCS3 has been proved to be detri- mental for cell survival (Provost et al., 2005). SOCS proteins have been shown to be negative regulators of cytokine signaling (Alexander and Hilton, 2004). TNF-α is an enigmatic cytokine controlling signaling pathways to- wards cell proliferation and cell death. Complex-I and complex-II are the two complexes formed by TNF-α by binding with TNF-R (Wullaert et al., 2007). Complex I activates IKK, which in turn phos- phorylates IκBα resulting in ubiquitination and further proteosomal degradation of IκBα. This leads to activation of NFκB, which com- prises of homodimers of p65 or heterodimers of p50 and p65. These dimers then translocate to the nucleus and bind to the promoter European Journal of Pharmacology 684 (2012) 132–145 ⁎ Corresponding author. Tel.: + 91 9433343917. E-mail address: [email protected] (B. Chattopadhyay). 0014-2999/$ – see front matter © 2012 Elsevier B.V. All rights reserved. doi:10.1016/j.ejphar.2012.02.009 Contents lists available at SciVerse ScienceDirect European Journal of Pharmacology journal homepage: www.elsevier.com/locate/ejphar
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Pulmonary Protein dependent fate of hepatic cells under nicotine induced stress and curcumin...

European Journal of Pharmacology 684 (2012) 132–145
Contents lists available at SciVerse ScienceDirect
European Journal of Pharmacology
j ourna l homepage: www.e lsev ie r .com/ locate /e jphar
Pulmonary, Gastrointestinal and Urogenital Pharmacology
Protein dependent fate of hepatic cells under nicotine induced stress and curcuminameliorated condition
Satyam Banerjee a, Krishna Chattopadhyay b, Jasmeet Kaur Chhabra c, Brajadulal Chattopadhyay a,⁎a Department of Physics, Jadavpur University, Raja S.C. Mullick Road, Kolkata-700032, Indiab Department of Chemical Technology, Calcutta University, 92 A.P.C. Road, Kolkata-700009, Indiac Immuno Biology Division, Indian Institute of Toxicology Research, M. G. Marg, Lucknow-226001, India
⁎ Corresponding author. Tel.: +91 9433343917.E-mail address: [email protected] (B. Chatto
0014-2999/$ – see front matter © 2012 Elsevier B.V. Alldoi:10.1016/j.ejphar.2012.02.009
a b s t r a c t
a r t i c l e i n f oArticle history:Received 1 July 2011Received in revised form 1 February 2012Accepted 9 February 2012Available online 23 February 2012
Keywords:ApoptosisCell proliferationCurcuminNicotineProtein-restrictionSignaling molecule
Nicotine is mainly metabolized in liver. Its abuse elicits acute phase response by activating macrophages toproduce pro-inflammatory cytokines, which play critical role in apoptosis or cell proliferation. The protectivepharmacological mechanism of curcumin against nicotine-induced toxicity on protein malnourished liver isstill remaining unclear. This study investigated the ameliorative mechanism of curcumin against nicotine-induced toxicity and also fate of liver particularly under protein restricted condition. Female Albino-ratsmaintained under normal/protein-restricted diets, were subcutaneously injected with nicotine tartrate(2.5 mg/kg body weight/day) and orally supplemented with curcumin (80 mg/kg body weight/day) for21 days. The animals were then sacrificed to dissect out liver and proceed with further experiments. Interac-tions of nicotine with DNA both in vivo and in vitro were observed by thermal denaturation and DNA ladder-ing assays. Effects of nicotine on hepatic cells were monitored by differential staining, comet assay, cytokineprofiling, mRNA and protein expression. Nicotine caused more intense DNA damage, promoted hepatic celldeath through up-regulating pro-apoptotic proteins and signaling molecules in protein malnourished indi-viduals. Through up-regulation of anti-apoptotic proteins and proliferation promoting molecules, nicotinedysregulated homeostasis in normal protein condition. Curcumin significantly ameliorated the nicotine-induced toxicity in both conditions and regulated the imbalance between cell survival and death inducedby nicotine. The protein content present in the nicotine induced hepatic cell decides either cell-survival path-way or cytotoxic pathway.
© 2012 Elsevier B.V. All rights reserved.
1. Introduction
Large populations in developing countries are deprived of propernutrition and addicted to tobacco consumption (Ortner et al., 2002).Malnutrition is a recurring problem particularly protein-energy mal-nutrition is widely observed in India and other developing countries(Müller and Krawinkel, 2005). Nicotine, the major addictive alkaloidof tobacco is mainly metabolized in liver (Hukkanen et al., 2005;Nakajima et al., 1998), though its toxic effects on hepatic cells remainunclear. Low dietary protein possesses a constraint on the metabolicactivity thereby resulting in impaired detoxification machinery ofliver (Nakajima et al., 1998). Based on human autopsy samples fromsmokers, the highest affinity for nicotine is in the liver, kidney, spleenand lung (Benowitz et al., 2009).
Nicotine abuse elicits acute phase response in liver by activatingmonocytes and macrophages to produce pro-inflammatory cytokineslike IL-6, TNF-α, etc. (Song et al., 1999) whose over exposure eventually
padhyay).
rights reserved.
contribute to acute liver injuries (Hoek and Pastorino, 2002). IL-6 phos-phorylates Signal Transducers andActivators of Transcription (STAT) byactivating Janus Kinases (Jaks) (Streetz et al., 2000). PhosphorylatedSTATs form dimers which function as transcription factors for them-selves and induce expression of one of its member's proteins e.g. STAT3. The induction of Suppressor of Cytokine Signaling (SOCS) genes isfacilitated by the STATs, with several putative STAT-binding sites in dif-ferent SOCS promoter regions. Constitutive activation of STAT 3 hasbeen reported in various carcinomas (Macha et al., 2011; Shukla et al.,2010) whereas over expression of SOCS3 has been proved to be detri-mental for cell survival (Provost et al., 2005). SOCS proteins have beenshown to be negative regulators of cytokine signaling (Alexander andHilton, 2004).
TNF-α is an enigmatic cytokine controlling signaling pathways to-wards cell proliferation and cell death. Complex-I and complex-II arethe two complexes formed by TNF-α by binding with TNF-R(Wullaert et al., 2007). Complex I activates IKK, which in turn phos-phorylates IκBα resulting in ubiquitination and further proteosomaldegradation of IκBα. This leads to activation of NFκB, which com-prises of homodimers of p65 or heterodimers of p50 and p65. Thesedimers then translocate to the nucleus and bind to the promoter

133S. Banerjee et al. / European Journal of Pharmacology 684 (2012) 132–145
region of genes having NF-κB consensus site (Tripathi and Aggarwal,2006). Complex II induces pro-apoptotic protein BAX that leads torelease of cytochrome C and reactive oxygen species (ROS) from themitochondria. Cytochrome C along with a cascade of caspases causesapoptosis whereas ROS induces necrosis of the cell (Wullaert et al.,2007).
Curcumin, an active ingredient in the rhizome of Curcuma longaexhibits a variety of pharmacological effects including anti-tumor,anti-inflammatory and anti-genotoxic activities (Aggarwal et al.,2003; Menon and Sudheer, 2007). Our previous study showed thatnicotine could bind to DNA causing significantly higher genotoxiceffect in protein-restricted condition (Bandyopadhyaya et al., 2008;Banerjee et al., 2010).
The number of smokers in female population is increasing day byday who are also deprived of healthy diet. Nicotine seems to be agreater risk factor in women than man (Prescott et al., 1998; Zangand Wynder, 1996). Smoking affects the reproductive health ofwomen and causes infertility, birth defects, ectopic pregnancy andspontaneous abortion (De Mallo et al., 2001). The present investiga-tions elucidate the nicotine induced signaling network and fate ofliver cells under normal and protein restricted conditions and theantagonistic effects of curcumin over nicotine induced changes hence-forth in female population.
2. Materials and methods
2.1. Chemicals
Nicotine hydrogen tartrate and curcumin were purchased (2008)from Sigma Chemicals Company, St. Louis, USA. All other analyticalgrade chemicals were supplied (2008 and 2009) by SpectrochemPvt. Ldt. India and Genei, Bangalore, India.
2.2. Diet and treatments
Thirty adult (40–45 days old) female albino rats (Rattus norvegi-cus) having body weight 120–130 g of Wistar strain were procuredfrom the animal housing facility and acclimatized under standardconditions of temperature and humidity with 12 h light/dark cycles.They were maintained in accordance to the guidelines of InstructionalAnimal Ethics Committee of Jadavpur University, Kolkata, India as de-scribed earlier (Bandyopadhyaya et al., 2008). The animals were fedstandard pellet diet (Hindustan Lever ltd., India) and water ad libituma week prior to start with normal/protein restricted diet. Thirty ratswere divided equally (n=5) into the following groups. Group 1: an-imals fed with normal protein diet (18% casein, 70% carbohydrate, 7%fat, 4% salt mixture and 1% vitamin mixture) and received no nicotinetreatment. Group 2: animals fed with normal protein diet and treatedwith nicotine, Group 3: animals fed with normal protein diet andtreated with nicotine+curcumin, Group 4: animals fed with proteinrestricted diet (5% casein, 83% carbohydrate, 7% fat, 4% salt mixtureand 1% vitamin mixture) and received no nicotine treatment. Group5: animals fed with protein-restricted diet and treated with nicotine.Group 6: animals fed with protein restricted diet and treated withnicotine+curcumin. Nicotine tartrate was used for nicotine treat-ment and administered subcutaneously daily at the dose of 2.5 mg/kg of body weight in 0.5 ml physiological saline and curcumin wasadministered orally at the dose of 80 mg/kg body weight/day. Com-positions of diets were prepared as described by Hawk et al. (1954)and selection of nicotine dose, route of administration and the periodof dosing were adopted from earlier study of Mandal et al. (2004).The dose of curcumin was selected from the study of Kalpana andMenon (2004). Nicotine and curcumin administration continued for21 days. The animals in groups 1 and 4 received subcutaneous injec-tion of 0.5 ml physiological saline without nicotine. One week priorto treatment till completion of the treatments, animals were kept in
their respective dietary regime. After the completion of treatments,animals were kept fasting overnight and sacrificed next day by decapi-tation. Blood was collected from heart immediately after decapitationand serum was separated by centrifugation and stored at −20 °Cprior to further analysis. Livers were dissected out and wiped clearwith tissue paper to remove adhering blood and tissue fluid and storedin vacuum desiccators at −20 °C as such without pulverization or ho-mogenization in buffer in order to prevent exposure to auto-oxidationenvironment.
2.3. DNA extraction
50 mg of liver tissue was homogenized in homogenizing buffercontaining 1% SDS, 50 mM EDTA, Proteinase K (100 μg/μl) and incu-bated at 45 °C for 45 min. To the homogenate, equal volumes of satu-rated phenol and sevag (24:1 mix of chloroform and isoamyl alcohol)was added and centrifuged at 8000 g for 10 min at 4 °C in SuperspinPlastocraft Centrifuge. The supernatant was collected and 1.5 volumesof sevag was added to it and mixed properly. The mixture was thencentrifuged at 8000 g for 10 min at 4 °C and the supernatant wascollected. Two times volumes of chilled absolute ethanol and 0.2 vol-umes of ammonium acetate (10 M) was added to the supernatantand the mixture was incubated at −20 °C for 1 h. The nuclear precip-itate was obtained by spinning the mixture at 14,000 g for 25 min at4 °C, which dissolved in 600 μl TE buffer (pH 8). RNase (50 μg/μl)was added to it and incubated at 37 °C for 45 min. Equal volumes ofphenol and sevag was added to it, mixed properly and the mixturewas centrifuged at 8000 g for 10 min at 4 °C. The supernatant was col-lected carefully and 1.5 volumes of sevag was added to it, mixed, andcentrifuged again at 8000 g for 10 min at 4 °C. Finally the supernatantwas collected carefully and 2 volumes of chilled absolute ethanol and0.2 volumes of ammonium acetate (10 M)was added to it and incubat-ed at−20 °C for 1 h. The DNA precipitate was collected by centrifugingthe mixture at 14,000 g for 30 min and dissolved in TE buffer (pH 8).The purity of isolated DNA and its concentration was checked and esti-mated by spectrophotometrically.
2.4. Thermal denaturation assay
Binding interactions of nicotine and curcumin with DNA (both invivo and in vitro) were studied by thermal denaturation experimentusing liver DNA isolated from animals of each group. The absorbanceratios of DNA at 260/280 nm were found to be 1.79–1.84 indicatingthat the isolated DNA were satisfactorily free from proteins (Müllerand Rajewsky, 1980). Working solutions for DNA were prepared inMillipore-purified water buffered at pH 8 using Tris–EDTA (10 mMTris; 1 mM EDTA). For in vitro study, 1 ml working solution contain-ing 15 μg DNA (isolated from rats of normal diet control group) wastaken in quartz cuvette and nicotine was added to it such that thefinal concentration of nicotine in the mixture became 100 μM. Opticaldensities (OD) of the mixture were monitored at 260 nm at differenttemperatures (20–90 °C) using Jasco V-650 spectrophotometer withTeflon stoppered quartz absorption cell thermostated with a circulat-ing water heating/cooling accessory. Similar experiments were doneby adding different final concentrations of nicotine (200, 300, 400and 500 μM) and also 500 μM final concentration of nicotine alongwith 100 μM final concentration of curcumin respectively to 1 ml ofworking solution containing 15 μg DNA (isolated from control ratsof normal diet group). All nicotine–DNA and nicotine–DNA–curcuminsolutions were incubated at 37 °C for 1 h prior to examination. Heat-ing was applied at 5 °C min−1 from 20 °C until thermal denaturationof the DNA was completed, as judged from the increase in absorption.Thermal denaturation temperature (Tm) (defined as the mid-point ofthe hyperchromic transition) was determined from the optical absor-bance versus temperature curves. The change in Tm (ΔTm) followinginteraction of rat liver DNA with added nicotine and curcumin was

134 S. Banerjee et al. / European Journal of Pharmacology 684 (2012) 132–145
evaluated from: ΔTm=[Tm (DNA)−Tm (DNA–drug)]. In vivo thermaldenaturation assay of the DNA samples isolated from the animals ofeach group were studied by observing the OD at different tempera-tures similarly.
2.5. DNA laddering assay
DNA was isolated from liver tissues of animals in control groups(Group 1) as described early. The experiment was conducted by add-ing nicotine (final concentration 1, 1.5 and 2 mM respectively) to15 μg DNA each and 2 mM final concentration of nicotine alongwith 1 mM final concentration of curcumin to 15 μg DNA and incubat-ed at room temperature for 1 h. The interaction products were re-solved in 0.8% agarose gel electrophoresis. DNA isolated from theliver tissues of the animals in each groups were directly analyzed in0.8% agarose gel electrophoresis to monitor the effects of nicotineand curcumin on DNA in vivo. All the gels were visualized with ethi-dium bromide staining in BIO View UV Light transilluminator.
2.6. Differential staining
Paraffin sections of dissected livers from different groups of ani-mals were prepared to perform differential staining. Tissue sectionswere deparaffinized in xylene and rehydrated in various percentagesof ethyl alcohol. The sections were then washed with phosphate buff-er saline (PBS, pH 7.4), permeabilized with 0.25% Triton X-100 andagain washed with PBS. RNase (100 μg/ml) treatment was given at37 °C for 20 min. The sections were then incubated in PBS containingpropidium iodide (25 μg/ml) for 30 min at 39 °C. After washing inPBS, the sections were counter stained with bisbenzimide (5 μg/ml)and photograph was taken under fluorescent microscope (×200).An independent examiner performed blinded differential counting.Five tissue sections from each group were stained to obtain consistentresults. On the basis of morphology and stain the percentage of apo-ptosis and necrosis in control and treated groups were evaluated[percentage of necrotic or apoptotic cells=(total number of necroticor apoptotic cells / total number of cells)×100]. The mean percent-ages of cells±S.E.M. of five observations were plotted.
2.7. Comet assay
The procedure for Comet assay was followed with some modifica-tion of the method by Kido et al. (2006) and as described byBandyopadhyaya et al. (2008). Liver tissues (50 mg) were minced,suspended at 1 ml/g in chilled homogenizing buffer (0.075 M NaCland 0.024 MEDTA) and gently homogenized at 1000 g using LaboratoryStirrer (REMI-RQ127A) for 10 min at 0 °C to obtain nuclear precipitate.The precipitate was then resuspended in 1 ml chilled homogenizingbuffer. 100 μl of 2% regular melting point agarose (Genei, India) wasquickly layered on a pre-cooled fully frosted slide and covered with acoverslip and allowed to solidify. The nuclear preparation was mixed1:1 (v/v) with 2% low melting point agarose (Genei, India). The coverslip was removed carefully and a second layer of 100 μl of the mixturewas pipette out on the slide, covered with the cover slip again andallowed to gel at 4 °C for 15 min. The slide (without cover slip) was im-mersed in a freshly prepared and chilled lysing solution (2.5 M NaCl,100 mM EDTA, 10 mM Tris–HCl, 1% Sarkosyl, 10% DMSO and 1% TritonX-100, at pH 10; DMSO added just before use) and kept at 4 °C for 2 h.Slides were then placed on a horizontal gel electrophoresis platformin alkaline electrophoresis solution (300 mM NaOH and 1 mM EDTA,pH 13) and left in dark for 15 min to allow unwinding of the DNA tooccur. The DNA was then electrophoresed at 4 °C in the dark for15 min at 1 V/cm and approximately 250 mA. The slides were gentlyrinsed in neutralization buffer (0.4 M Tris–HCl, pH 7.5) and stainedwith 50 μl of 20 μg/ml ethidium bromides and covered with a cover
slip. The photomicrograph of each slide was taken by Leica FluorescentMicroscope (Model 300 FX) at 400× magnification.
2.8. Measurement of comet parameters
Measurement of the comet head diameter, tail length, and percent-age of DNA damage were performed as described by Helma and Uhl(2000). A total of 50 cells were screened per animal and examinedin a fluorescencemicroscope (Leica 3000-FXwith 400×magnification).Quantification of DNA damage for each cell was determined by Image Jsoftware as:
Total DNA in comet ¼ Total comet areað Þ � mean DNA intensityð ÞTotal DNA in comet head ¼ Total head areað Þ � mean DNA intensityð Þ%DNA damage ¼ Total DNA in cometð Þ− Total DNA in comet headð Þ
Total DNA in cometð Þ � 100
2.9. ELISA
Quantification of nicotine and curcumin induced IL-6 and TNF-αin serum of normal and protein restricted rats were performedusing Quantikine Rat IL-6 and TNF-α Immunoassay Kit (Minneapolis,U.S.A.). This assay employs the quantitative sandwich enzyme immu-noassay technique. A monoclonal antibody specific for rat IL-6/TNF-αhad been pre-coated onto a microplate. Standard controls and sam-ples were pipetted into wells. If rat IL-6/TNF-α were present, itwould bind by the immobilized antibody. After washing away any un-bound substances, an enzyme-linked polyclonal antibody specific forrat IL-6/TNF-α was added to the wells. Following a wash to removeany unbound antibody, a substrate solution was added to the wells.The enzyme reaction yielded a blue product that turned into yellowwhen the stop solution was added. The intensity of the color mea-sured was in proportion to the amount of rat IL-6/TNF-α bound inthe initial step. The sample values were then read off from the stan-dard curve.
2.10. RNA extraction, quantitation and RT-PCR analysis
Total RNA was isolated from liver of control and treated rats main-tained under normal and protein restricted diets using Qiagen RNAeasymini kit (Qiagen, Hilden, Germany) as per the manufacturer' instruc-tions. The expression profile for mRNA was evaluated by qRT2-PCRtechnique using QuantiTect SYBR green RT-PCR kit (Qiagen, Germany)and by conventional Reverse Transcriptase PCR using Qiagen RT-PCRkit (Qiagen, Germany). The reaction conditions for qRT2-PCR were50 °C for 30 min for reverse transcription; initial PCR activation at95 °C for 15 min followed by 40 cycles of denaturation at 94 °C for15 s, annealing at a 5 °C below the Tm of primers for 30 s and extensionof 30 s at 72 °C. The reaction conditions for RT-PCR were 50 °C for30 min for reverse transcription; initial PCR activation at 95 °C for15 min followed by 30 cycles of denaturation at 94 °C for 15 s, anneal-ing at a 5 °C below the Tm of primers for 30 s, extension of 1 min at72 °C final extension of 10 min at 72 °C. One micro liter of templateRNA containing 50 ng of total RNA was used in a 50 μl RT-PCR cocktail.All reactions were performed in triplicates. Relative quantificationswere performed in Light Cycler Real-Time PCR Machine using LightCycler 480 relative quantification software release 1.2.0625. GAPDHwas used as reference control. The amounts of the detected gene prod-uctswere normalizedwith that of GAPDHbefore statistical analysis. Theresults are expressed as the target/reference ratio of each sample nor-malized by the target/reference ratio of the calibrator. The RT-PCRprod-ucts were resolved on 1.2% agarose gel, stained with ethidium bromideand visualized under UV-transilluminator. Primers (Operon Biotechno-logis, Gmbh, Germany) used for RT-PCR were:

135S. Banerjee et al. / European Journal of Pharmacology 684 (2012) 132–145
IL-6: Forward 5′GCCCTTCAGGAACAGCTATG3′Reverse 5′ACCACAGTGAGGAATGTCCA3′;
STAT3: Forward 5′ TCTTAGGGCCTGGTGTGAAC3′Reverse 5′CACTCCGAGGTCAGATCCAT3′;
SOCS3: Forward 5′ CCTCAAGACCTTCAGCTCCA3′Reverse 5′GGCTGGATTTTTGTGCTTGT3′;
p65: Forward 5′CTCCTTTTCTCAAGCCGATG3′Reverse 5′GACAGATGCCAGGTCTGTGA3′;
IκBα: Forward 5′ TTGGTCAGGTGAAGGGAGAC3′Reverse 5′ACAAGTCCACGTTCCTTTGG3′;
Bcl-2: Forward 5′ATGATAACCGGGAGATCGTG3′Reverse 5′ CTCACTTGTGGCCCAGGTAT3′;
Bax: Forward 5′GAAGACAGGGGCCTTTTTGT3′Reverse 5′CAGCCCATCTTCTTCCAGAT3′
Fig. 1. A. In vitro thermal denaturation of liver DNA treated with different concentra-tions of nicotine.Binding interactions of nicotine with DNAwere studied by thermal de-naturation experiments using liver DNA isolated from control group animals of normaldiet. For in vitro study, 1 ml working solution (10 mM Tris; 1 mM EDTA, pH 8) contain-ing 15 μg DNAwas taken in quartz cuvette and 100 μMnicotine was added to it. Opticaldensities (OD) of the mixture were monitored at 260 nm at different temperatures(20–90 °C) using Jasco V-650 spectrophotometer with Teflon stoppered quartz absorp-tion cell thermostated with a circulating water heating/cooling accessory. All nicotine–DNA solutions were incubated at 37 °C for 1 h prior to examination. Heating wasapplied at 5 °C min−1 from 20 °C until thermal denaturation of the DNA was complet-ed, as judged from the increase in absorption. Thermal denaturation temperature (Tm)defined as the mid-point of the hyperchromic transition was determined from the op-tical absorbance versus temperature curves. The change in Tm (ΔTm) following interac-tion of rat liver DNA with added nicotine and curcumin was evaluated from: ΔTm=[Tm(DNA)−Tm (DNA–drug)]. Similar experiments were done by adding 200, 300, 400 and500 μM nicotine to DNA.B. In vitro thermal denaturation of liver DNA treated with nic-otine and curcumin.The interactions of nicotine and curcumin with DNA were studiedsimilarly as described in Fig. 1A. In this experiment 500 μM nicotine along with 100 μMcurcumin respectively was added to 1 ml of working solution containing 15 μg DNAand performed the experiment similarly.
2.11. Western blotting
Effects of nicotine and curcumin on the protein expression level ofBax, Bcl-2, IκBα and p65 in normal and protein-restricted liver wereassessed by Western blot analysis. Cytosolic and nuclear fractions ofliver tissues were prepared by using Lysis Buffer A (20 mM HEPES,100 mMNaCl, 1 mMDTT, 1 mM EDTA, 0.05% Triton X-100, 1× ProteaseInhibitor cocktail, pH 7.9) and Lysis Buffer B (20 mM HEPES, 1.5 mMMgCl2, 0.4 M NaCl, 1 mM DTT, 1 mM EDTA, 0.05% Triton X-100, 25%Glycerol, 1× protease inhibitor cocktail, pH 7.9). The tissue (30 mg)was minced in PBS (pH 7.4) containing 1× protease inhibitor cocktailand homogenized on low power using 30 s bursts until fully homoge-nized. The homogenate was transferred to pre-cooled centrifuge tubeand centrifuged at 2900 g for 10 min at 4 °C. Supernatant (a) was col-lected and the pellet was dissolved in lysis buffer A which again centri-fuged at 3500 g for 10 min at 4 °C. Supernatant (b) was collected andthe pellet was dissolved in lysis buffer B, vortexed for 10 s and incubat-ed on ice for 20 min. After incubation, it was centrifuged at 40,000 g for15 min at 4 °C and the supernatant (c) was collected. Combination ofsupernatants (a) and (b) was the cytoplasmic extract and supernatant(c) was the nuclear extract. Proteins were precipitated by additionof 8 volumes ice-cold acetone to the supernatants, incubated overnightat −20 °C and pelleted by centrifugation at 12,000 g for 20 min at 4 °C.The pellets were dissolved in buffer, containing 8 M urea, 5% β-mercaptoethanol, 10% SDS, 2% CHAPS and 50mM DTT. Protein concen-trationwas estimated by Bradfordmethod andequal amount of proteins(20 μg)were separated on12% SDS-PAGE,which then electrophoretical-ly transferred on nitrocellulose membranes. After blocking with 2% bo-vine serum albumin (BSA) in TBST (137 mM NaCl, 25 mM Tris, 1 mMNa2EDTA and 0.1% Tween 20), the membranes were incubated withrabbit anti-mouse (Bax, Bcl-2, IκBα and p65) antibodies (Santa CruzBiotech, California, USA) at 4 °C overnight at a dilution of 1:200 to1:1000. After washing three times with TBST, the membranes were in-cubated with HRP conjugated goat anti-rabbit IgG antibody (1:2000 di-lution) at room temperature for 3 h, and then washed three times withTBST before incubation with a chemiluminescent peroxides substrate(Sigma, St. Louis, USA). Reactions were visualized using an enhancedchemiluminescence system. This experiment was repeated two times.
2.12. Statistical analysis
All experiments were repeated twice and data (n=10 animals)were averaged and tabulated as mean±standard deviation (S.D.).The statistical analysis of the data estimated from each group of allconditions was done by One-way analysis of variance (ANOVA) andall pair wise Multiple Comparison Procedures (Holm–Sidak method)by using Sigma Stat (version 3.2). Significance levels were examinedat Pb0.01 for significant and Pb0.001 for more significant.
3. Results
3.1. Nicotine–DNA adduction caused subsequent shift in DNA meltingcurve, which was reverted by curcumin
In vitro interaction study showed that nicotine alone and togetherwith curcumin affected the DNA melting temperature as seen inFig. 1A and B. It was observed that the hyperchromicity of normal
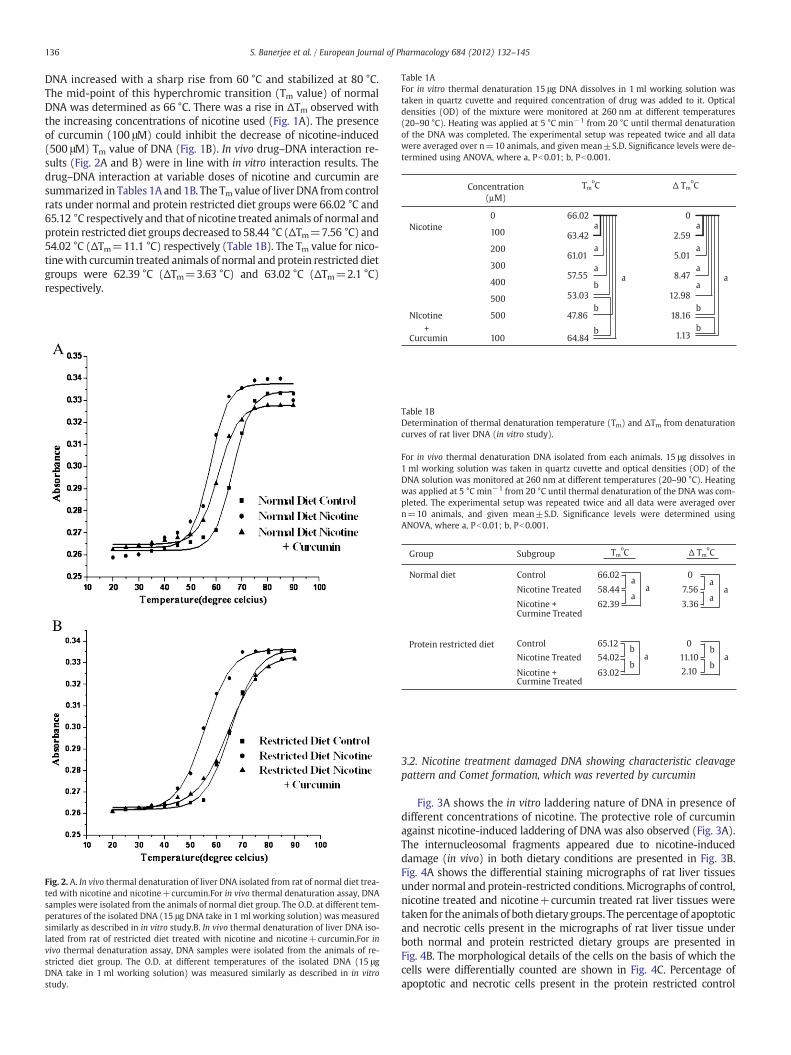
Table 1AFor in vitro thermal denaturation 15 μg DNA dissolves in 1 ml working solution wastaken in quartz cuvette and required concentration of drug was added to it. Opticaldensities (OD) of the mixture were monitored at 260 nm at different temperatures(20–90 °C). Heating was applied at 5 °C min−1 from 20 °C until thermal denaturationof the DNA was completed. The experimental setup was repeated twice and all datawere averaged over n=10 animals, and given mean±S.D. Significance levels were de-termined using ANOVA, where a, Pb0.01; b, Pb0.001.
Concentration
(µM)
TmoC Δ Tm
oC
Nicotine
0 0
100
200
300
400
66.02
63.42
61.01
57.55
53.03
2.59
5.01
8.47
12.98
a
a
a
aa
a
a
a
ba
136 S. Banerjee et al. / European Journal of Pharmacology 684 (2012) 132–145
DNA increased with a sharp rise from 60 °C and stabilized at 80 °C.The mid-point of this hyperchromic transition (Tm value) of normalDNA was determined as 66 °C. There was a rise in ΔTm observed withthe increasing concentrations of nicotine used (Fig. 1A). The presenceof curcumin (100 μM) could inhibit the decrease of nicotine-induced(500 μM) Tm value of DNA (Fig. 1B). In vivo drug–DNA interaction re-sults (Fig. 2A and B) were in line with in vitro interaction results. Thedrug–DNA interaction at variable doses of nicotine and curcumin aresummarized in Tables 1A and 1B. The Tm value of liver DNA from controlrats under normal and protein restricted diet groups were 66.02 °C and65.12 °C respectively and that of nicotine treated animals of normal andprotein restricted diet groups decreased to 58.44 °C (ΔTm=7.56 °C) and54.02 °C (ΔTm=11.1 °C) respectively (Table 1B). The Tm value for nico-tinewith curcumin treated animals of normal and protein restricted dietgroups were 62.39 °C (ΔTm=3.63 °C) and 63.02 °C (ΔTm=2.1 °C)respectively.
Fig. 2. A. In vivo thermal denaturation of liver DNA isolated from rat of normal diet trea-ted with nicotine and nicotine+curcumin.For in vivo thermal denaturation assay, DNAsamples were isolated from the animals of normal diet group. The O.D. at different tem-peratures of the isolated DNA (15 μg DNA take in 1 ml working solution) was measuredsimilarly as described in in vitro study.B. In vivo thermal denaturation of liver DNA iso-lated from rat of restricted diet treated with nicotine and nicotine+curcumin.For invivo thermal denaturation assay, DNA samples were isolated from the animals of re-stricted diet group. The O.D. at different temperatures of the isolated DNA (15 μgDNA take in 1 ml working solution) was measured similarly as described in in vitrostudy.
NIcotine
Curcumin
500
500
100
47.86
64.84
18.16
1.13
b
b
b
b+
Table 1BDetermination of thermal denaturation temperature (Tm) and ΔTm from denaturationcurves of rat liver DNA (in vitro study).
For in vivo thermal denaturation DNA isolated from each animals. 15 μg dissolves in1 ml working solution was taken in quartz cuvette and optical densities (OD) of theDNA solution was monitored at 260 nm at different temperatures (20–90 °C). Heatingwas applied at 5 °C min−1 from 20 °C until thermal denaturation of the DNA was com-pleted. The experimental setup was repeated twice and all data were averaged overn=10 animals, and given mean±S.D. Significance levels were determined usingANOVA, where a, Pb0.01; b, Pb0.001.
Subgroup TmoC Δ Tm
oCGroup
Normal diet Control
Nicotine Treated
Nicotine +Curmine Treated
Nicotine Treated
Nicotine +Curmine Treated
66.02
58.44
62.39
0
7.56
3.36
Protein restricted diet Control 65.12
54.02
63.02
0
11.10
2.10
a
aa
a
aa
b
ba
b
ba
3.2. Nicotine treatment damaged DNA showing characteristic cleavagepattern and Comet formation, which was reverted by curcumin
Fig. 3A shows the in vitro laddering nature of DNA in presence ofdifferent concentrations of nicotine. The protective role of curcuminagainst nicotine-induced laddering of DNA was also observed (Fig. 3A).The internucleosomal fragments appeared due to nicotine-induceddamage (in vivo) in both dietary conditions are presented in Fig. 3B.Fig. 4A shows the differential staining micrographs of rat liver tissuesunder normal and protein-restricted conditions. Micrographs of control,nicotine treated and nicotine+curcumin treated rat liver tissues weretaken for the animals of both dietary groups. The percentage of apoptoticand necrotic cells present in the micrographs of rat liver tissue underboth normal and protein restricted dietary groups are presented inFig. 4B. The morphological details of the cells on the basis of which thecells were differentially counted are shown in Fig. 4C. Percentage ofapoptotic and necrotic cells present in the protein restricted control

Fig. 3. A. Photomicrographs of gel electrophoresis showing DNA laddering (in vitroassay)DNA isolated from the liver tissues of the animals in control groups and isolatedDNA (15 μg) was treated with different concentrations of nicotine and curcumin. Thetreated DNA was analyzed in 0.8% agarose gel electrophoresis to monitor the effectsof nicotine and curcumin on DNA in vitro. All the gels were visualized with ethidiumbromide staining in BIO View UV Light transilluminator and photograph was taken.B.Photomicrographs of gel electrophoresis showing DNA laddering (in vivo assay)DNAsample (15 μg) isolated from the liver tissues of individual animals in each groupswere directly analyzed in 0.8% agarose gel electrophoresis to monitor the effects ofnicotine and curcumin on DNA in vivo. All the gels were visualized with ethidium bro-mide staining in BIO View UV Light transilluminator and photograph was taken.
137S. Banerjee et al. / European Journal of Pharmacology 684 (2012) 132–145
group (12.4±1.2% and 16.6±1.8% respectively) was relatively morethan those of normal diet control group (8±1.2% and 11.6±2.7%respectively). Exposure to nicotine increased the percentage of apoptosisand necrosis in both dietary conditions, where the percentage of celldeath was significantly higher in protein restricted nicotine treatedgroup (30±3.4% for apoptosis and 39±3% for necrosis) than normaldiet nicotine treated group (17±1.7% for apoptosis and 25.2±2% fornecrosis). Exposure to combination of nicotine and curcumin resultedin decrease of apoptosis (10.4±1.8% for normal diet and 23±2.2% forprotein restricted diet) and necrosis (18.8±2.3% for normal diet and20.4±1.2% for protein restricted diet) in the rat liver tissues.
The results of Comet assay are shown in Fig. 5A. In this figure, thecomet photographs of the rat liver DNAmaintained under both normalprotein diet and protein restricted diets are included. Comet assayshowed retrenched head diameter, elongated tail length (Fig. 5B), re-duced head diameter vs. tail length ratio (Fig. 5C) and heightened per-centage of DNA damages (Fig. 5D), which were significantly higherin protein-restricted condition. The percentages of DNA damages as ob-served in comet assay were 5.96% in normal diet control group and13.69% in protein restricted control group respectively. Nicotine signif-icantly elevated the percentage of DNA damages to 52.84% in protein-restricted condition and 36.76% in normal condition (Fig. 5D). Curcuminadministrationmore significantly prevented the nicotine-induced dam-age in the protein-restricted group (Fig. 5D).
3.3. Nicotine heightened expression of the Th1 cytokines from their basalserum levels and curcumin prevented this elevation
ELISA was performed to determine the concentration of IL-6 andTNF-α in serum of rats treated with nicotine and curcumin in bothdietary conditions. ELISA results indicated that the protein expressionprofile of IL-6 (Fig. 6A) and TNF-α (Fig. 6B) in basal level of proteinrestricted serum was more as compared to that of serum of normaldiet. Nicotine increased the concentration of IL-6 by 2 folds in normaldiet condition and 2.5 folds in protein restricted condition (Fig. 6A)approximately. The concentration of TNF-α was found to be 2.5 foldsmore in normal diet serum and 1.5 folds more in protein restrictedserum (Fig. 6B) due to nicotine treatment. Curcumin treatment sup-pressed the protein levels of both the cytokines and thereby broughtdown their levels approximately close to that in the control of theirrespective groups as seen from Fig. 6. The effects of curcumin in downregulation of the cytokineswere found to bemore prominent in proteinrestricted condition.
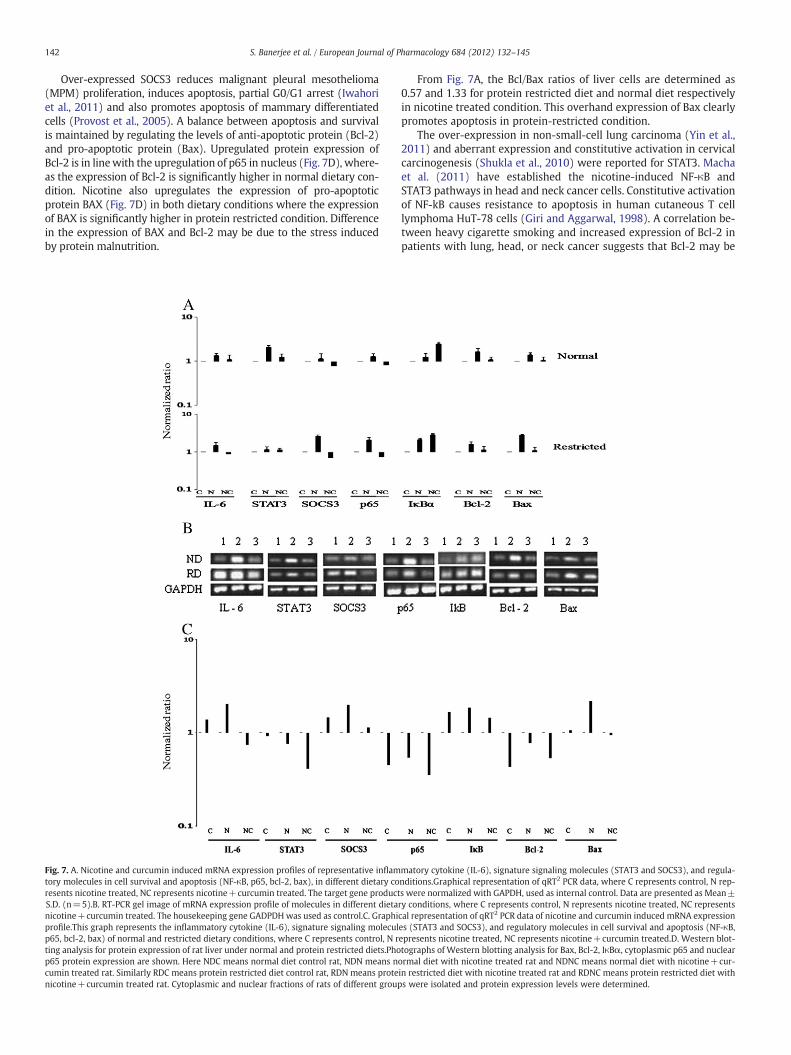
3.4. Nicotine and curcumin regulated a signaling cascade involving in-flammatory cytokines, signature signaling molecules, transcription fac-tors and apoptosis-associated molecules that modulate cell survivaland death, variably in both the dietary conditions
To monitor the effects of nicotine and curcumin on the signalingmolecules we measured their relative mRNA expressions by Real TimePCR. Consistent over expression of IL-6, STAT3 and SOCS3 were ob-served in liver of nicotine-treated rats under both dietary conditions,in comparison to their respective controls (Fig. 7A and B). But the ex-pression levels of IL-6 and SOCS3 in restricted diet condition were com-paratively higher than that of normal diet condition. At the same timethe expression level of STAT3 was found to be less in protein restrictedcondition than that of normal diet condition. Exposure to curcuminresulted in down regulation of these molecules in comparison to theirnicotine-induced expression in both dietary conditions (Fig. 7A andB). The variation in the expressions of IL-6, STAT3 and SOCS3 indicatedthat the basal level of IL-6 and SOCS3 was higher and that of STAT3waslower in protein restricted condition as compared to normal diet condi-tion (Fig. 7C). Nicotine induced STAT3/SOCS3 ratio in protein-restrictedliver was found to be less as compared to normal liver (Fig. 7A). Down-regulation of IL-6 and STAT3 by curcuminwasmore effective in protein-restricted condition whereas SOCS3 was down regulated more in nor-mal diet condition.
The mRNA expression levels of p65, IκBα, Bcl-2 and Bax were ele-vated significantly after nicotine treatment in both dietary conditionsas observed in Fig. 7A and B. Curcumin induced down-regulation ofBcl-2, Bax and p65 whereas over-expressed IκBα in both dietary con-ditions. Comparative study between their expressions in differentdietary conditions revealed that the basal levels of p65 and Bcl-2were lower whereas IκBα and Bax were higher in protein restrictedcondition compared to normal protein condition (Fig. 7C). Nicotineinduced Bcl-2/Bax ratio in protein restricted liver was lower in com-parison to the ratio that of normal liver. Curcumin induced down-regulation of Bcl-2 and Bax were more effective in protein-restrictedcondition than normal protein condition. Also curcumin induced ex-pression level of p65 was found to be lower but that of IκBα washigher in liver under restricted protein condition compared to normalprotein condition.
3.5. Nicotine and curcumin induced expression of apoptosis associatedmolecules and regulation of NFκB activation in both dietary conditions
The effects of nicotine and curcumin on the expression levels ofpro-apoptotic protein Bax, anti-apoptotic protein Bcl-2 and transcrip-tion factor NFκB under both dietary conditions were determined byWestern blot analysis (Fig. 7D). The expression of Bcl-2 protein was
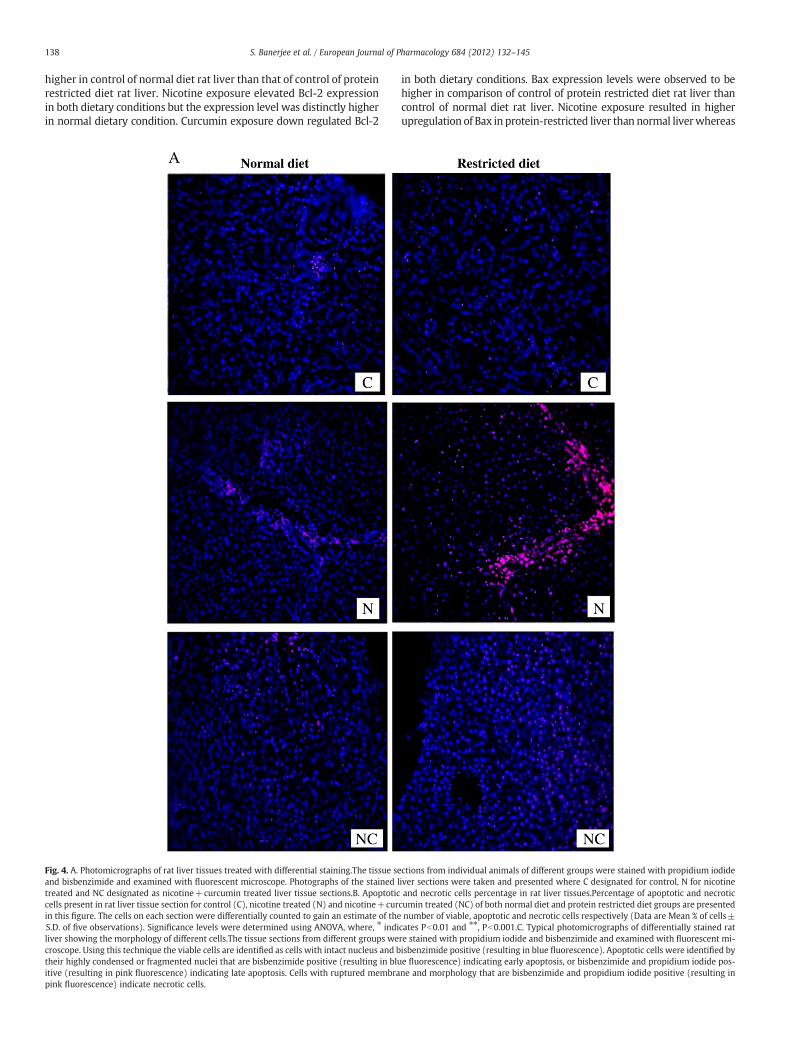
138 S. Banerjee et al. / European Journal of Pharmacology 684 (2012) 132–145
higher in control of normal diet rat liver than that of control of proteinrestricted diet rat liver. Nicotine exposure elevated Bcl-2 expressionin both dietary conditions but the expression level was distinctly higherin normal dietary condition. Curcumin exposure down regulated Bcl-2
Fig. 4. A. Photomicrographs of rat liver tissues treated with differential staining.The tissue seand bisbenzimide and examined with fluorescent microscope. Photographs of the stained ltreated and NC designated as nicotine+curcumin treated liver tissue sections.B. Apoptoticcells present in rat liver tissue section for control (C), nicotine treated (N) and nicotine+curin this figure. The cells on each section were differentially counted to gain an estimate of theS.D. of five observations). Significance levels were determined using ANOVA, where, ⁎ indiliver showing the morphology of different cells.The tissue sections from different groups wecroscope. Using this technique the viable cells are identified as cells with intact nucleus and btheir highly condensed or fragmented nuclei that are bisbenzimide positive (resulting in bluitive (resulting in pink fluorescence) indicating late apoptosis. Cells with ruptured membrapink fluorescence) indicate necrotic cells.
in both dietary conditions. Bax expression levels were observed to behigher in comparison of control of protein restricted diet rat liver thancontrol of normal diet rat liver. Nicotine exposure resulted in higherupregulation of Bax in protein-restricted liver than normal liverwhereas
ctions from individual animals of different groups were stained with propidium iodideiver sections were taken and presented where C designated for control, N for nicotineand necrotic cells percentage in rat liver tissues.Percentage of apoptotic and necrotic
cumin treated (NC) of both normal diet and protein restricted diet groups are presentednumber of viable, apoptotic and necrotic cells respectively (Data are Mean % of cells±
cates Pb0.01 and ⁎⁎, Pb0.001.C. Typical photomicrographs of differentially stained ratre stained with propidium iodide and bisbenzimide and examined with fluorescent mi-isbenzimide positive (resulting in blue fluorescence). Apoptotic cells were identified bye fluorescence) indicating early apoptosis, or bisbenzimide and propidium iodide pos-ne and morphology that are bisbenzimide and propidium iodide positive (resulting in
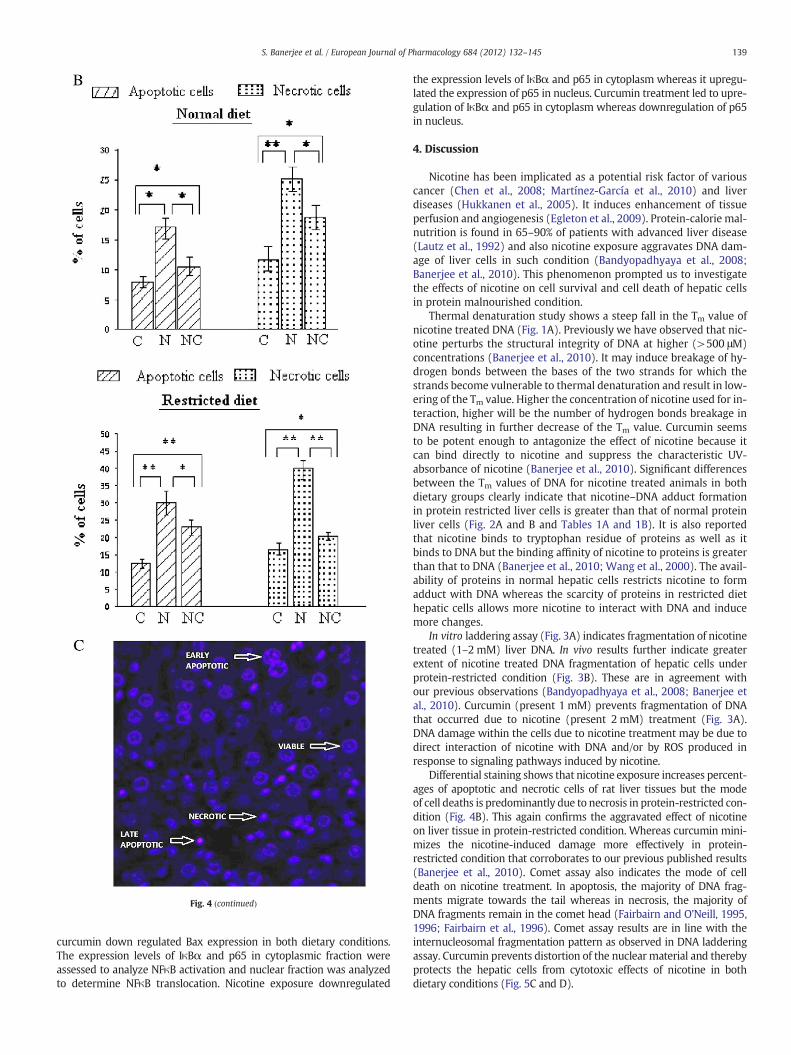
Fig. 4 (continued)
139S. Banerjee et al. / European Journal of Pharmacology 684 (2012) 132–145
curcumin down regulated Bax expression in both dietary conditions.The expression levels of IκBα and p65 in cytoplasmic fraction wereassessed to analyze NFκB activation and nuclear fraction was analyzedto determine NFκB translocation. Nicotine exposure downregulated
the expression levels of IκBα and p65 in cytoplasm whereas it upregu-lated the expression of p65 in nucleus. Curcumin treatment led to upre-gulation of IκBα and p65 in cytoplasm whereas downregulation of p65in nucleus.
4. Discussion
Nicotine has been implicated as a potential risk factor of variouscancer (Chen et al., 2008; Martínez-García et al., 2010) and liverdiseases (Hukkanen et al., 2005). It induces enhancement of tissueperfusion and angiogenesis (Egleton et al., 2009). Protein-calorie mal-nutrition is found in 65–90% of patients with advanced liver disease(Lautz et al., 1992) and also nicotine exposure aggravates DNA dam-age of liver cells in such condition (Bandyopadhyaya et al., 2008;Banerjee et al., 2010). This phenomenon prompted us to investigatethe effects of nicotine on cell survival and cell death of hepatic cellsin protein malnourished condition.
Thermal denaturation study shows a steep fall in the Tm value ofnicotine treated DNA (Fig. 1A). Previously we have observed that nic-otine perturbs the structural integrity of DNA at higher (>500 μM)concentrations (Banerjee et al., 2010). It may induce breakage of hy-drogen bonds between the bases of the two strands for which thestrands become vulnerable to thermal denaturation and result in low-ering of the Tm value. Higher the concentration of nicotine used for in-teraction, higher will be the number of hydrogen bonds breakage inDNA resulting in further decrease of the Tm value. Curcumin seemsto be potent enough to antagonize the effect of nicotine because itcan bind directly to nicotine and suppress the characteristic UV-absorbance of nicotine (Banerjee et al., 2010). Significant differencesbetween the Tm values of DNA for nicotine treated animals in bothdietary groups clearly indicate that nicotine–DNA adduct formationin protein restricted liver cells is greater than that of normal proteinliver cells (Fig. 2A and B and Tables 1A and 1B). It is also reportedthat nicotine binds to tryptophan residue of proteins as well as itbinds to DNA but the binding affinity of nicotine to proteins is greaterthan that to DNA (Banerjee et al., 2010; Wang et al., 2000). The avail-ability of proteins in normal hepatic cells restricts nicotine to formadduct with DNA whereas the scarcity of proteins in restricted diethepatic cells allows more nicotine to interact with DNA and inducemore changes.
In vitro laddering assay (Fig. 3A) indicates fragmentation of nicotinetreated (1–2 mM) liver DNA. In vivo results further indicate greaterextent of nicotine treated DNA fragmentation of hepatic cells underprotein-restricted condition (Fig. 3B). These are in agreement withour previous observations (Bandyopadhyaya et al., 2008; Banerjee etal., 2010). Curcumin (present 1 mM) prevents fragmentation of DNAthat occurred due to nicotine (present 2 mM) treatment (Fig. 3A).DNA damage within the cells due to nicotine treatment may be due todirect interaction of nicotine with DNA and/or by ROS produced inresponse to signaling pathways induced by nicotine.
Differential staining shows that nicotine exposure increases percent-ages of apoptotic and necrotic cells of rat liver tissues but the modeof cell deaths is predominantly due to necrosis in protein-restricted con-dition (Fig. 4B). This again confirms the aggravated effect of nicotineon liver tissue in protein-restricted condition. Whereas curcumin mini-mizes the nicotine-induced damage more effectively in protein-restricted condition that corroborates to our previous published results(Banerjee et al., 2010). Comet assay also indicates the mode of celldeath on nicotine treatment. In apoptosis, the majority of DNA frag-ments migrate towards the tail whereas in necrosis, the majority ofDNA fragments remain in the comet head (Fairbairn and O'Neill, 1995,1996; Fairbairn et al., 1996). Comet assay results are in line with theinternucleosomal fragmentation pattern as observed in DNA ladderingassay. Curcumin prevents distortion of the nuclear material and therebyprotects the hepatic cells from cytotoxic effects of nicotine in bothdietary conditions (Fig. 5C and D).

140 S. Banerjee et al. / European Journal of Pharmacology 684 (2012) 132–145
Protein malnutrition also leads to various physical complexitiescharacterized by persistent or relapsing fatigue. High expressionlevel of IL-6 is generally related to fatigue scores and greater magni-tude of TNF-α can also be observed in these conditions (Gaab et al.,2005; Glaser et al., 1999). Protein deficiency induces stress in liverleading to over-expression of Th1 expressed cytokines. ELISA resultsof our experiment confirm the previous reports in protein-restrictedcondition (Fig. 6A and B). Mahapatra et al. (2011) have reportedthat nicotine augments the secretion of pro-inflammatory cytokines
Fig. 5. A. Photomicrographs of Comet assay of liver DNA of rats.Comet assay were done by umal diet and protein restricted diet groups. Comet photomicrographs of each liver DNA, wshown here.B. Determination of head diameter and tail length of Comet picture of liver Dwere determined using ANOVA, where, ⁎ indicates Pb0.01 and ⁎⁎, Pb0.001.C. Determinatioand protein restricted diets.Comet head diameter vs. tail length ratios of two different dietsshown here. Significance levels were determined using ANOVA, where, ⁎ indicates Pb0.01 annormal and protein restricted diets.A total of 50 cells were screened per animal and examintion of DNA damage, head diameter and tail length for each cell was determined by Image JANOVA, where, ⁎ indicates Pb0.01 and ⁎⁎, Pb0.001.
and generates excess amount of ROS in macrophages. According toSong et al. (1999), nicotine increases IL-6 mRNA expression in liverand spleen. Our findings show that nicotine induced expression ofthese cytokines in serum is significantly higher in protein restrictedcondition as compared to normal protein condition. Stress and nicotinetherefore synergistically heightens the immunological and acute phaseresponse in protein restricted condition.
Significantly higher level of IL-6 in protein restricted (control/treated) condition increases STAT3 expression (Fig. 7A and B),
sing liver DNA from control, nicotine, and nicotine+curcumin treated rats of both nor-here C for control, N for nicotine treated and NC for nicotine+curcumin treated areNA from rats maintained with normal and protein restricted diets.Significance levelsn of head diameter vs. tail length ratio of liver DNA from rats maintained with normal, where C for control, N for nicotine treated and NC for nicotine+curcumin treated ared ⁎⁎, Pb0.001.D. Determination of % of damage of liver DNA from rats maintained withed in a fluorescence microscope (Leica 3000-FX with 400× magnification). Quantifica-software. Data are presented as Mean±S.D. Significance levels were determined using
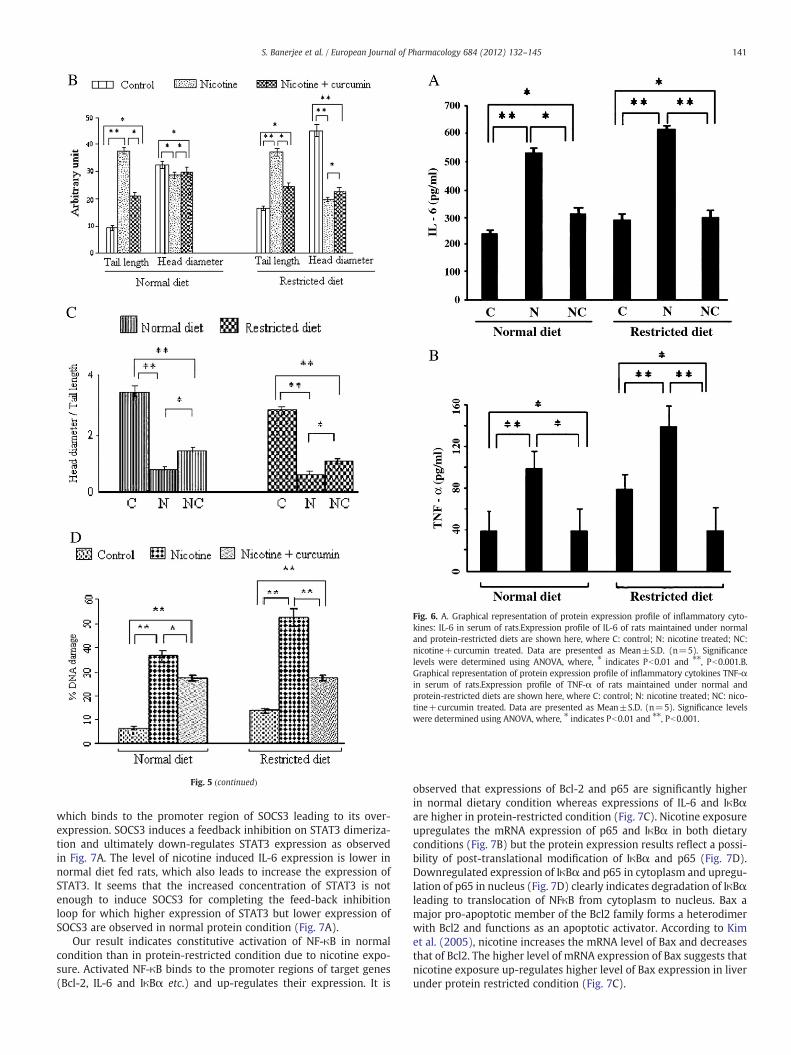
Fig. 6. A. Graphical representation of protein expression profile of inflammatory cyto-kines: IL-6 in serum of rats.Expression profile of IL-6 of rats maintained under normaland protein-restricted diets are shown here, where C: control; N: nicotine treated; NC:nicotine+curcumin treated. Data are presented as Mean±S.D. (n=5). Significancelevels were determined using ANOVA, where, ⁎ indicates Pb0.01 and ⁎⁎, Pb0.001.B.Graphical representation of protein expression profile of inflammatory cytokines TNF-αin serum of rats.Expression profile of TNF-α of rats maintained under normal andprotein-restricted diets are shown here, where C: control; N: nicotine treated; NC: nico-tine+curcumin treated. Data are presented as Mean±S.D. (n=5). Significance levelswere determined using ANOVA, where, ⁎ indicates Pb0.01 and ⁎⁎, Pb0.001.
Fig. 5 (continued)
141S. Banerjee et al. / European Journal of Pharmacology 684 (2012) 132–145
which binds to the promoter region of SOCS3 leading to its over-expression. SOCS3 induces a feedback inhibition on STAT3 dimeriza-tion and ultimately down-regulates STAT3 expression as observedin Fig. 7A. The level of nicotine induced IL-6 expression is lower innormal diet fed rats, which also leads to increase the expression ofSTAT3. It seems that the increased concentration of STAT3 is notenough to induce SOCS3 for completing the feed-back inhibitionloop for which higher expression of STAT3 but lower expression ofSOCS3 are observed in normal protein condition (Fig. 7A).
Our result indicates constitutive activation of NF-κB in normalcondition than in protein-restricted condition due to nicotine expo-sure. Activated NF-κB binds to the promoter regions of target genes(Bcl-2, IL-6 and IκBα etc.) and up-regulates their expression. It is
observed that expressions of Bcl-2 and p65 are significantly higherin normal dietary condition whereas expressions of IL-6 and IκBαare higher in protein-restricted condition (Fig. 7C). Nicotine exposureupregulates the mRNA expression of p65 and IκBα in both dietaryconditions (Fig. 7B) but the protein expression results reflect a possi-bility of post-translational modification of IκBα and p65 (Fig. 7D).Downregulated expression of IκBα and p65 in cytoplasm and upregu-lation of p65 in nucleus (Fig. 7D) clearly indicates degradation of IκBαleading to translocation of NFκB from cytoplasm to nucleus. Bax amajor pro-apoptotic member of the Bcl2 family forms a heterodimerwith Bcl2 and functions as an apoptotic activator. According to Kimet al. (2005), nicotine increases the mRNA level of Bax and decreasesthat of Bcl2. The higher level of mRNA expression of Bax suggests thatnicotine exposure up-regulates higher level of Bax expression in liverunder protein restricted condition (Fig. 7C).
142 S. Banerjee et al. / European Journal of Pharmacology 684 (2012) 132–145
Over-expressed SOCS3 reduces malignant pleural mesothelioma(MPM) proliferation, induces apoptosis, partial G0/G1 arrest (Iwahoriet al., 2011) and also promotes apoptosis of mammary differentiatedcells (Provost et al., 2005). A balance between apoptosis and survivalis maintained by regulating the levels of anti-apoptotic protein (Bcl-2)and pro-apoptotic protein (Bax). Upregulated protein expression ofBcl-2 is in linewith the upregulation of p65 in nucleus (Fig. 7D), where-as the expression of Bcl-2 is significantly higher in normal dietary con-dition. Nicotine also upregulates the expression of pro-apoptoticprotein BAX (Fig. 7D) in both dietary conditions where the expressionof BAX is significantly higher in protein restricted condition. Differencein the expression of BAX and Bcl-2 may be due to the stress inducedby protein malnutrition.
Fig. 7. A. Nicotine and curcumin induced mRNA expression profiles of representative inflamtory molecules in cell survival and apoptosis (NF-κB, p65, bcl-2, bax), in different dietary coresents nicotine treated, NC represents nicotine+curcumin treated. The target gene productS.D. (n=5).B. RT-PCR gel image of mRNA expression profile of molecules in different dietanicotine+curcumin treated. The housekeeping gene GADPDH was used as control.C. Graphiprofile.This graph represents the inflammatory cytokine (IL-6), signature signaling moleculp65, bcl-2, bax) of normal and restricted dietary conditions, where C represents control, N rting analysis for protein expression of rat liver under normal and protein restricted diets.Phop65 protein expression are shown. Here NDC means normal diet control rat, NDN means ncumin treated rat. Similarly RDC means protein restricted diet control rat, RDN means protenicotine+curcumin treated rat. Cytoplasmic and nuclear fractions of rats of different group
From Fig. 7A, the Bcl/Bax ratios of liver cells are determined as0.57 and 1.33 for protein restricted diet and normal diet respectivelyin nicotine treated condition. This overhand expression of Bax clearlypromotes apoptosis in protein-restricted condition.
The over-expression in non-small-cell lung carcinoma (Yin et al.,2011) and aberrant expression and constitutive activation in cervicalcarcinogenesis (Shukla et al., 2010) were reported for STAT3. Machaet al. (2011) have established the nicotine-induced NF-κB andSTAT3 pathways in head and neck cancer cells. Constitutive activationof NF-kB causes resistance to apoptosis in human cutaneous T celllymphoma HuT-78 cells (Giri and Aggarwal, 1998). A correlation be-tween heavy cigarette smoking and increased expression of Bcl-2 inpatients with lung, head, or neck cancer suggests that Bcl-2 may be
matory cytokine (IL-6), signature signaling molecules (STAT3 and SOCS3), and regula-nditions.Graphical representation of qRT2 PCR data, where C represents control, N rep-s were normalized with GAPDH, used as internal control. Data are presented as Mean±ry conditions, where C represents control, N represents nicotine treated, NC representscal representation of qRT2 PCR data of nicotine and curcumin induced mRNA expressiones (STAT3 and SOCS3), and regulatory molecules in cell survival and apoptosis (NF-κB,epresents nicotine treated, NC represents nicotine+curcumin treated.D. Western blot-tographs of Western blotting analysis for Bax, Bcl-2, IκBα, cytoplasmic p65 and nuclearormal diet with nicotine treated rat and NDNC means normal diet with nicotine+cur-in restricted diet with nicotine treated rat and RDNC means protein restricted diet withs were isolated and protein expression levels were determined.

Fig. 7 (continued)
143S. Banerjee et al. / European Journal of Pharmacology 684 (2012) 132–145
a target of carcinogens found in tobacco smoke (Gallo et al., 1995).Significantly lower fragmentation of DNA, constitutive activation ofNF-κB and increased expressions of p65, STAT3 and Bcl-2 observedin our experiments clearly support the cellular survival in nicotinetreated normal liver.
The anticancer potential of curcumin stems from its ability to sup-press proliferation of a wide variety of tumor cells and down-regulates several transcription factors. Curcumin attenuates the ex-pression of IL-6 and IL-8 in the TNF-α-treated HaCaT cells (Park etal., 2010) and inhibits Th1 cytokine profile in CD4+ T cells (Kang etal., 1999). These results corroborate with our findings showing thatcurcumin down regulates the protein expression level of IL-6 andTNF-α in serum of rats in both dietary conditions (Fig. 6A and B).It is already established that nicotine increases lipid peroxidation. Itsignificantly increases Malondialdehyde (MDA) levels in plasma andliver tissues (Chattopadhyay and Chattopadhyay, 2008; Chattopadhyayet al., 2010). We have also observed that curcumin down regulatesROS levels in serum and protects hepatic cell membranes of nicotine in-duced rats from lipid peroxidation (communicated data). Suppression ofIL-6 level in serum leads to suppression of signaling molecules (STAT3andSOCS3) that are up-regulated through JAK-STAT pathway. Curcumininduces down regulation of STAT3 leading to reduction in dysregulatedsurvival of cells that is supported by constitutive expression of STAT3. Italso protects cells from dysregulated apoptosis supported by over-expressed SOCS3. Curcumin supplemented down regulation of TNF-αleads to over expression of IκBα and suppression in the expression of
NF-κB (Fig. 6 and 7). IκBα (an NF-κB target gene) also terminates NF-κB activation at the transcriptional level. The increased synthesis ofIκBα shuts down NF-κB-induced gene expression by IκBα-mediatednuclear export of the DNA-binding subunits, thereby acting within anegative feedback loop (Arenzana-Seisdedos et al., 1997). Curcuminsupplementation leads to simultaneous co-expression of IκBα and p65in cytoplasm (Fig. D) along with downregulation of p65 in the nucleus(Fig. 7D). This clearly supports the phenomenon of curcumin mediatedNF-κB inactivation and downregulation of its target genes IL-6 and Bcl-2. Also down-regulation of TNF-α inhibits the expression of pro-apoptotic molecule (Bax) in presence of curcumin (Figs. 6 and 7).
5. Conclusion
Nicotine promotes cell survival in hepatic cells maintained undernormal protein condition. On the other hand, it acts as a cytotoxicagent promoting hepatic cell death, which may lead to liver cirrhosis.Thus, the protein content of the nicotine induced hepatic cell decideseither the survival pathwayor cytotoxic pathway. Curcumin significant-ly ameliorates nicotine-induced changes in both dietary conditions andregulates the imbalance between cell survival and death induced bynicotine, which is shown in Fig. 8. Curcumin supplementation alongwith chemotherapy and reduced protein intake may prove therapeuti-cally beneficial in treating cancers that were resistant to chemotherapyand radiation therapy due to over expression of anti-apoptotic proteins.

Fig. 8. Mechanism of the pathway of nicotine and curcumin interaction with liver in different dietary conditions.Nicotine induced IL-6 binds to IL-6 receptor gp130 inducing phos-phorylation of STAT3 by Jaks and STAT3 dimerization. STAT3 dimers translocates to the nucleus and binds to STAT binding sequence TTNNNNNAA. STAT3 functions as transcriptionfactor for STAT3 itself and binding of STAT3 on their promoter region upregulates STAT3. SOCS3 promoter region has STAT3 binding site and over-expression of STAT3 promotesSOCS3 expression. SOCS3 induces a feedback inhibition on phosphorylation of STAT3 by Jaks, inhibiting STAT3 dimerization and nuclear translocation. Over expression of STAT3 isobserved in hepatic cells maintained under normal dietary conditions whereas over expression of SOCS3 is observed in hepatic cells maintained under protein restricted condition.Nicotine induced TNF-α binds to its receptor inducing phosphorylation, ubiquitination of IκB by IKK, activating p65-p50 (NF-κB) dimers to translocate to the nucleus. NF-κB func-tions as transcription factor for IL-6, Bcl-2 (anti-apoptotic) and IκBα thus upregulating their expression. TNF-α induced IL-6 binds to gp130 further inducing its response throughJAK-STAT signaling pathway. Binding of TNF-α to TNF-R1 leads to complex II formation and caspase 8 is activated. Caspase 8 initiates the mitochondrial pathway by cleaving Bid totBid which leads to insertion of Bak and Bax (pro-apoptotic) in the mitochondria and finally mitochondrial permeabilization. These events result in the mitochondrial release ofcytochrome C and ROS. Cytochrome C contributes to apoptosis whereas ROS contributes to necrosis. Over expression of Bcl-2 is observed in hepatic cells under normal dietarycondition whereas over expression of Bax is observed in hepatic cells under protein restricted condition. Curcumin supplementation down regulated IL-6 and TNF-α thus down-regulating the signaling molecules down the line. Curcumin upregulated the expression of IκBαwhich translocates to the nucleus, binds to p65 and importing it back to its inactivestate in the cytoplasm.
144 S. Banerjee et al. / European Journal of Pharmacology 684 (2012) 132–145
Conflict of interest
There is no conflict of interest in this research work.
Acknowledgment
The support for this work from Biophysics Laboratory (PhysicsDepartment, Jadavpur University, Kolkata, India) is gratefully acknowl-edged. We are grateful to Professor Soumen Basak, Head, Chemical Sci-ences Department, Saha Institute of Nuclear Physics, Kolkata, India forhis kind help and cooperation in conducting some experiments in hisown laboratory. We are also very much thankful to Dr. B.N Paul, Head,Immunobiology Division, Indian Institute of Toxicology Research, Luck-now, India for allowing us to conduct a part of this research work. Weextend our thanks to Ms. Vani Mishra, Immunology Division, IndianInstitute of Toxicology Research, Lucknow, India for her support. Wealso acknowledge the support of Dr. S. Biswas, IICB, Kolkata, India.The authors are also thankful to Jadavpur University for giving legalapproval of the work.
References
Aggarwal, B.B., Kumar, A., Bharti, A.C., 2003. Anticancer potential of curcumin: preclin-ical and clinical studies. Anticancer. Res. 23, 363–398.
Alexander, W.S., Hilton, D.J., 2004. The role of suppressors of cytokine signaling (SOCS)proteins in regulation of the immune response. Annu. Rev. Immunol. 22, 503–529.
Arenzana-Seisdedos, F., Turpin, P., Rodriguez, M., Thomas, D., Hay, R.T., Virelizier, J.L.,Dargemont, C., 1997. Nuclear localization of I kappa B alpha promotes active trans-port of NF-kappa B from the nucleus to the cytoplasm. J. Cell Sci. 110, 369–378.
Bandyopadhyaya, G., Sinha, S., Chattopadhyay, B.D., Chakraborty, A., 2008. Protectiverole of curcumin against nicotine induced genotoxicity on rat liver under restricteddietary protein. Eur. J. Pharmacol. 588, 151–157.
Banerjee, S., Bandhyopadhyaya, G., Chattopadhyay, K., Chattopadhyay, B.D., 2010. Ame-lioration of nicotine induced damage of blood cells in protein malnourished femalerats by curcumin. Int. J. Pharmacol. 6, 409–420.
Benowitz, N.L., Hukkanen, J., Jacob, P.I.I.I., 2009. Nicotine chemistry, metabolism, kineticsand biomarkers. Handb. Exp. Pharmacol. 192, 29–60.
Chattopadhyay, K., Chattopadhyay, B.D., 2008. Effect of nicotine on lipid profile, peroxida-tion and antioxidant enzymes in female rats with restricted dietary protein. Indian J.Med. Res. 127, 571–576.
Chattopadhyay, K., Mandal, S., Chattopadhyay, B.D., Ghosh, S., 2010. Ameliorative effectof sesame lignans on nicotine toxicity in rats. Food Chem. Toxicol. 48, 3215–3220.
Chen, R.J., Ho, Y.S., Guo, H.R., Wang, Y.J., 2008. Rapid activation of Stat3 and ERK1/2 bynicotine modulates cell proliferation in human bladder cancer cells. Toxicol. Sci.104, 283–293.
De Mallo, B.P.R., Pinto, G.R., Botelho, C., 2001. The influence of smoking on fertility,pregnancy and lactation. J. Pediatr. 77, 257–264.
Egleton, R.D., Brown, K.C., Dasgupta, P., 2009. Angiogenic activity of nicotinic acetylcho-line receptors. Implications in tobacco related vascular diseases. Pharmacol. Ther.121, 205–223.
Fairbairn, D.W., O'Neill, K.L., 1995. Necrotic DNA degradation mimics apoptotic nucle-osomal fragmentation comet tail length. In vitro Cell Dev. Biol. 31, 171–173.
Fairbairn, D.W., O'Neill, K.L., 1996. The neutral comet assay is sufficient to identify anapoptotic ‘window’ by visual inspection. Apoptosis 1, 91–94.
Fairbairn, D.W., Walburger, D.K., Fairbairn, J.J., O'Neill, K.L., 1996. Key morphologicchanges and DNA strand breaks in human lymphoid cells discriminating apoptosisfrom necrosis. J. Scan Microsc. 18, 407–416.
Gaab, J., Rohleder, N., Heitz, V., Engert, V., Schad, T., Schurmeyer, T.H., Ehlert, U., 2005.Stress-induced changes in LPS-induced pro-inflammatory cytokine production inchronic fatigue syndrome. Psychoneuroendocrinology 30, 188–198.
Gallo, O., Bianchi, S., Porfirio, B., 1995. Bcl-2 overexpression and smoking history inhead and neck cancer. J. Natl. Cancer Inst. 87, 1024–1025.
Giri, D.K., Aggarwal, B.B., 1998. Constitutive activation of NF-kappaB causes resistance to ap-optosis in human cutaneous T cell lymphoma HuT-78 cells. Autocrine role of tumor ne-crosis factor and reactive oxygen intermediates. J. Biol. Chem. 273, 14008–14014.
Glaser, R., Kiecolt-Glaser, J.K., Marucha, P.T., MacCallum, R.C., Laskowski, B.F., Malarkey,W.B., 1999. Stress-related changes in pro-inflammatory cytokine production inwounds. Arch. Gen. Psychiatry 56, 450–456.
Hawk, P.B., Oser, B.L., Summerson, W.H., Oser, B.L.Ltd., 1954. Practical PhysiologicalChemistry, 13th Ed. Mc Graw Hill Book Comp, New York, London.
Helma, C., Uhl, M., 2000. A public domain image-analysis program for the single-cellgel-electrophoresis (Comet) assay. Nutr. Res. 466, 9–15.

145S. Banerjee et al. / European Journal of Pharmacology 684 (2012) 132–145
Hoek, J.B., Pastorino, J.G., 2002. Ethanol, oxidative stress, and cytokine-induced livercell injury. Alcohol 27, 63–68.
Hukkanen, J., Jacob, P., Benowitz, N.L., 2005. Metabolism and disposition kinetics ofnicotine. Pharmacol. Rev. 57, 79–115.
Iwahori, K., Serada, S., Fujimoto, M., Nomura, S., Osaki, T., Lee, C.M., Mizuguchi, H.,Takahashi, T., Ripley, B., Okumura, M., Kawase, I., Kishimoto, T., Naka, T., 2011.Overexpression of SOCS3 exhibits preclinical antitumor activity against malignantpleural mesothelioma. Int. J. Cancer 129, 993–1005.
Kalpana, C., Menon, V.P., 2004. Modulatory effects of curcumin on lipid peroxidation andantioxidant status during nicotine-induced toxicity. Pol. J. Pharmacol. 56, 581–586.
Kang, B.Y., Song, Y.J., Kim, K.M., Choe, Y.K., Hwang, S.Y., Kim, T.S., 1999. Curcumin in-hibits Th1 cytokine profile in CD4+ T cells by suppressing interleukin-12 produc-tion in macrophages. Br. J. Pharmacol. 128, 380–384.
Kido, R., Sato, I., Tsuda, S., 2006. Detection of in vivo DNA damage induced by ethanol inmultiple organs of pregnant mice using the alkaline single cell gel electrophoresis(Comet) assay. J. Vet. Med. Sci. 68 (1), 41–47.
Kim, K.H., Joo, K.J., Park, H.J., Kwon, C.H., Jang, M.H., Kim, C.J., 2005. Nicotine inducesapoptosis in TM3 mouse Leydig cells. Fertil. Steril. 83, 1093–1099.
Lautz, H.U., Selberg, O., Korber, J., 1992. Protein–calorie malnutrition in liver cirrhosis.Clin. Investig. 70, 478–486.
Macha, M.A., Matta, A., Chauhan, S.S., Michael, K.W.S., Ralhan, R., 2011. Guggulsterone(GS) inhibits smokeless tobacco and nicotine-induced NF-κB and STAT3 pathwaysin head and neck cancer cells. Carcinogenesis 32, 368–380.
Mahapatra, S.K., Bhattacharjee, S., Chakraborty, S.P., Majumdar, S., Roy, S., 2011. Alter-ation of immune functions and Th1/Th2 cytokine balance in nicotine-induced mu-rine macrophages: Immunomodulatory role of eugenol and N-acetylcysteine. Int.Immunopharmacol. 11, 485–495.
Mandal, K., Dasgupta, S., Chattopadhyay, B.D., 2004. Nicotine induced alterations on re-productive organs of female rats under protein deficient condition. Indian J. Exp.Biol. 42, 330–332.
Martínez-García, E., Irigoyen, M., González-Moreno, O., Corrales, L., Teijeira, A., Salvo, E.,Rouzaut, A., 2010. Repetitive nicotine exposure leads to a more malignant andmetastasis-prone phenotype of SCLC: a molecular insight into the importance ofquitting smoking during treatment. Toxicol. Sci. 116, 467–476.
Menon, V.P., Sudheer, A.R., 2007. Antioxidant and anti-inflammatory properties of cur-cumin. Adv. Exp. Med. Biol. 595, 105–125.
Müller, O., Krawinkel, M., 2005. Malnutrition and health in developing countries. CMAJ173, 279–286.
Müller, R., Rajewsky, M.S., 1980. Immunological quantification by high affinity anti-bodies of 06-ethyldeoxyguanosine in DNA exposed to N-ethyl-N-nitrosourea. Can-cer Res. 40, 887–896.
Nakajima, M., Iwata, K., Yamamoto, T., Funae, Y., Yoshida, T., Kuroiwa, Y., 1998. Nicotinemetabolism in liver microsomes from rats with acute hepatitis or cirrhosis. DMD26, 36–41.
Ortner, R., Schindler, S.D., Kraigher, D., Mendelsohn, A., Fischer, G., 2002. Womenaddicted to nicotine. Arch Womens Ment Health 4, 103–109.
Park, K., Lee, J.H., Cho, H.C., Cho, S.Y., Cho, J.W., 2010. Down-regulation of IL-6, IL-8,TNF-α and IL-1β by glucosamine in HaCaT cells, but not in the presence of TNF-α. Oncologia 1, 289–292.
Prescott, E., Hippe, M., Schnorr, P., Hein, H.O., Vestbo, L., 1998. Smoking and risk ofmyocardial infarction in women and men: longitudinal population study. Br.Med. J. 316, 1043–1047.
Provost, F.L., Miyoshi, K., Vilotte, J.L., Bierie, B., Robinson, G.W., Hennighausen, L., 2005.SOCS3 promotes apoptosis of mammary differentiated cells. Biochem. Biophys.Res. Commun. 338, 1696–1701.
Shukla, S., Shishodia, G., Mahata, S., Hedau, S., Pandey, A., Bhambhani, S., Batra, S., Basir,S.F., Das, B.C., Bharti, A.C., 2010. Aberrant expression and constitutive activation ofSTAT3 in cervical carcinogenesis: implications in high-risk human papillomavirusinfection. Mol. Cancer 9, 282–299.
Song, D.K., Im, Y.B., Jung, J.S., Suh, H.W., Huh, S.O., Song, J.H., Kim, Y.H., 1999. Central in-jection of nicotine increases hepatic and splenic interleukin 6 (IL-6) mRNA expres-sion and plasma levels in mice: involvement of the peripheral sympatheticnervous system. FASEB J. 13, 1259–1267.
Streetz, K.L., Luedde, T., Manns, M.P., Trautwein, C., 2000. Interleukin 6 and liver regen-eration. Gut 47, 309–312.
Tripathi, P., Aggarwal, A., 2006. NF-kB transcription factor: a key player in the genera-tion of immune response. Curr. Sci. 90, 519–531.
Wang, Y., Cheng, Y., Sun, H.F., 2000. Interaction of nicotine and bovine serum albumin.Chin. Chem. Lett. 11, 247–250.
Wullaert, A., Loo, G.V., Heyninck, K., Beyaert, R., 2007. Hepatic tumor necrosis factorsignaling and nuclear factor-κB: effects on liver homeostasis and beyond. Endocr.Rev. 28, 365–386.
Yin, Z.J., Jin, F.G., Liu, T.G., Fu, E.Q., Xie, Y.H., Sun, R.L., 2011. Overexpression of STAT3potentiates growth, survival, and radioresistance of non-small-cell lung cancer(NSCLC) cells. J. Surg. Res. 171, 675–683.
Zang, E.A., Wynder, E.L., 1996. Differences in lung cancer risk between men andwomen: examination of the evidence. J. Natl. Cancer Inst. 88, 183–192.




















