
Identification and characterization of secreted and pathogenesis-related proteins in Ustilago maydis
Upload
independentCategory
view
6download
0
Proteomic Analysis of Early Melanosomes: Identification of Novel
Melanosomal Proteins
Venkatesha Basrur,†,⊥ Feng Yang,‡,⊥ Tsuneto Kushimoto,† Youichiro Higashimoto,†
Ken-ichi Yasumoto,† Julio Valencia,§ Jacqueline Muller,| Wilfred D. Vieira,† Hidenori Watabe,†
Jeffrey Shabanowitz,‡ Vincent J. Hearing,† Donald F. Hunt,X and Ettore Appella*,†
Laboratory of Cell Biology, National Cancer Institute, National Institutes of Health, Bethesda, Maryland 20892,Department of Chemistry, University of Virginia, Charlottesville, Virginia 22901, Center for Biologics
Evaluation and Research, Food and Drug Administration, Rockville, Maryland 20852 and Pathology Section,National Heart, Lung and Blood Institute, National Institutes of Health, Bethesda, Maryland 20892
Received August 20, 2002
Melanin is a heterogeneous biopolymer produced only by specific cells termed melanocytes, whichsynthesize and deposit the pigment in specialized membrane-bound organelles known as melanosomes.Although melanosomes have been suspected of being closely related to lysosomes and platelets, thetotal number of melanosomal proteins is still unknown. Thus far, six melanosome-specific proteinshave been identified, and the challenge is to characterize the complete proteome of the melanosometo further understand its mechanism of biogenesis. In this report, we used mass spectrometry andsubcellular fractionation to identify protein components of early melanosomes. Using this approach,we have identified all 6 of the known melanosome-specific proteins, 56 proteins that are shared withother organelles, and confirmed the presence of 6 novel melanosomal proteins using western blottingand by immunohistochemistry.
Keywords: melanoma • proteomics • melanosome
IntroductionMelanin is a complex pigmented biopolymer that is synthe-
sized in specialized membrane-bound organelles called mel-anosomes that are produced exclusively by melanocytes andby retinal pigment epithelial cells (reviewed in refs 1 and 2).The biogenesis of melanosomes and the regulation of melaninsynthesis have been studied extensively by means of electronmicroscopy, biochemical, and immunohistochemical methods.On the basis of their morphology and degree of melanization,melanosomes have been classified into four developmentalstages (I-IV). Various melanosome-specific proteins, such astyrosinase, Trp1/Tyrp1, Trp2/DCT, OA1 and MART1, aredelivered to stage I melanosomes following their processingthrough the endoplasmic reticulum (ER) and Golgi network,via several adapter protein complexes.3-5 Another melanosome-specific protein, gp100/Pmel17, which is a structural matrixprotein, is delivered directly to the stage I melanosomes withoutbeing processed through the Golgi network.6 Even though
active melanogenic enzymes are found in the sorting vesiclesand can be detected in stage I melanosomes, melanin synthesisdoes not begin until after gp100 is processed from its mem-brane bound into its soluble form and is integrated into thefibrillar matrix characteristic of stage II melanosomes.4,7 StageIII and IV melanosomes are mature forms that are laden withincreasing amounts of dark melanin pigment. As the pigmentis synthesized and deposited on the melanosome fibrils, themelanosomes move from the perinuclear region toward theperiphery of the melanocytes, i.e., the dendrite tips.8,9 Thisintracellular movement of melanosomes is mediated in partby microtubular motor myosin Va that is recruited by rab27avia its interaction with melanophilin, a process that is only nowbeing clarified at the molecular level.10-12 Once they arrive atthe dendritic tips, melanosomes are captured by actin filamentsand are transferred to adjoining keratinocytes.9,13
Despite the cloning and characterization of several key struc-tural and enzymatic proteins that make up these organelles,the mechanisms involved in the biogenesis of melanosomes,the regulation of melanin biosynthesis and their transfer to ker-atinocytes are still only partially understood. The presence ofmelanosomes uniquely in melanocytes does not reveal whetherthey are generated by a novel or an existing cellular mechanismfor organelle biogenesis. The observation that many humandiseases affect pigmentation and lysosomal function suggeststhat both types of organelles are closely related.14-16 However,Raposo et al. have recently reported that melanosomes repre-sent a distinct lineage of organelles, separable from conven-tional endosomes and lysosomes, within pigmented cells.17
* To whom correspondence should be addressed. Ettore Appella, M. D.,Laboratory of Cell Biology, Building 37, Room 1B03, National CancerInstitute, National Institutes of Health, Bethesda, MD 20892 USA. Phone:(301) 402-4177. Fax: (301) 496-7220. Email: [email protected].
† Laboratory of Cell Biology, National Cancer Institute, National Institutesof Health.
‡ Department of Chemistry, University of Virginia.§ Center for Biologics Evaluation and Research, Food and Drug Admin-
istration.| Pathology Section, National Heart, Lung and Blood Institute, National
Institutes of Health.⊥ V.B. and F.Y. contributed equally to this work.X Department of Chemistry and Pathology, University of Virginia.
10.1021/pr025562r CCC: $25.00 2003 American Chemical Society Journal of Proteome Research 2003, 2, 69-79 69Published on Web 11/08/2002

To date, the identification and cloning of melanosomalproteins has involved the genetic approach of characterizinggenes involved with melanosome/lysosome-related organelledysfunctions, such as Hermansky-Pudlak syndrome (HPS),Griscelli syndrome (GS), and Chediak-Higashi syndrome (CHS)or the use of different mouse coat color mutants. Over the past40 years, only 6 proteins have been identified as melanosome-specific proteins, whereas more than 100 distinct gene productsare thought to play direct or indirect roles in regulating theformation and function of these organelles in melanocytes.Hence, a more direct approach to comprehensively identify allof the proteins that constitute these organelles is likely toprovide useful insights into the complex mechanisms governingmammalian pigmentation and melanosome biogenesis.
With the increasing availability of larger genomic and pro-teomic databases, the mass spectrometric approach to proteinidentification is fast becoming the preferred technique. Becausethe tandem mass spectrometer is able to sequence complexmixtures of proteins with great sensitivity, it can be used toanalyze the proteomes of complex organelles, such as melano-somes. Hitherto, melanosomes have resisted such a direct anal-ysis, as it was difficult to prepare sufficient quantities of highlypurified, relatively melanin-free melanosomes. Stage III and IVmelanosomes, which can be readily purified because they sedi-ment to the bottom of sucrose density gradients, are not suit-able for such analysis because the endogenous melanin rendersmany of the melanosomal proteins insoluble by covalentmodification.18,19 Previously, we reported the capability of freeflow electrophoresis to purify early (stage I and II) melanosomesfrom mixtures partially purified by sucrose density gradientcentrifugation.4 Those purified early melanosomes were char-acterized using electron microscopy, immunohistochemical,and biochemical methods which clearly demonstrated that theprocessing of membrane bound gp100 into a soluble form is acritical step in the transition from stage I to stage II melano-somes, and that this then stabilizes melanogenic enzymefunction allowing the start of melanin synthesis.4
In this report, we have used mass spectrometry in conjunc-tion with free-flow electrophoresis (FFE) of sucrose densitygradient fractions to identify protein components of earlymelanosomes as they hold the key to understanding thebiogenesis and sorting mechanisms involved. Using this ap-proach, we have identified all 6 of the known melanosomalproteins and several novel proteins whose presence in mel-anosomes has been confirmed by western blotting and byimmunohistochemistry.
Materials and MethodsCell Culture and Purification of Early Melanosomes from
Human MNT1 Cells. The pigmented MNT1 cells were culturedat 37 °C in an atmosphere of 95% air, 5% CO2 in 150 cm2 culturedishes in minimal essential medium (all culture reagents werefrom Life Technologies, Grand Island, NY), as previouslydetailed.4
Confluent monolayers of MNT1 cells were harvested with0.05% trypsin, 0.53 mM EDTA and were washed once in 0.25M sucrose by centrifugation at 1000g for 5 min at 4 °C. Theywere then homogenized on ice using 20 strokes of a glass:glasstissue grinder and centrifuged at 1000g for 10 min at 4 °C.Tissue homogenates were then fractionated using variousprotocols, as detailed below and outlined in the Figures.
Melanosomes at various stages of maturation were separatedas described previously4 using sucrose density gradient cen-
trifugation (Gradient 1). Briefly, the homogenate was layeredon a discontinuous gradient of 1.0, 1.2, 1.4, 1.5, 1.6, 1.8, and2.0 M sucrose in 10 mM HEPES buffer, pH 7.0. Centrifugationwas carried out at 100 000 ×g in a Beckman SW28 swinging-bucket rotor for 1 h at 4 °C and melanosomes banding at 1.0-1.2 M sucrose were carefully removed and were further purifiedby FFE. Early melanosomes recovered from the 1.0-1.2 Msucrose interface were injected into the right inlet of anOctopus-PZE FFE apparatus (Weber GmbH, Kirchheim, Ger-many) at 2.0 mL/hr. FFE was performed at 1000-1100 V andapproximately 110-125 mA using 0.25 M sucrose in TEA, pH7.4, as the chamber buffer and an elution flow rate of 3-4 mL/min. The temperature of the sample was kept at 4 °C, and thechamber was maintained at 10 °C. Fractions were collected andanalyzed as detailed below.
Subsequently, a more effective separation method was devel-oped. Briefly, the MNT1 homogenate was resuspended in 2 Msucrose and was layered at the bottom of the 1.0-2.0 M sucrosestep gradient noted above (this reverse gradient is labeledGradient 2). The gradient was centrifuged at 100 000 ×g for 1h as noted above and the 1.0 M fraction was carefully recovered.That 1.0 M sample was then layered carefully in the middle ofa 3 part gradient (Gradient 3) consisting of equal volumes of0.8, 1.0, and 1.2 M sucrose. That gradient was again centrifugedat 100 000 ×g for 1 h and all 3 sucrose fractions were recoveredand analyzed by electron microscopy and by Western blottingas detailed below.
Protein Digestion and Mass Spectrometry. Enriched stageI and II melanosomes were processed for the purpose of se-quencing in two ways (Figure 1). First, we used SDS-PAGE andin gel digestion followed by 1D LC-MS/MS. Briefly, melano-some preparations were solubilized directly in sample bufferand 15 µg were separated on 10% NuPAGE gels according tothe manufacturer’s specifications. Gels were stained with colloi-dal Coomassie stain (Invitrogen, Carlsbad, CA) for 6 h and weredestained in water. The lane containing the sample was cutinto 4-mm slices, irrespective of the extent of staining, fromtop to bottom of the gel and destained for further 3 h in 30%methanol. In gel proteolysis with modified, sequencing gradetrypsin (Promega, Madison, WI) was carried out essentially asdescribed previously. Briefly, the gel slices were diced into 1mm cubes and washed with 150 µL 50% acetonitrile in 0.1 Mammonium bicarbonate buffer, pH 8.0. The diced cubes weredried and re-swollen in a minimal volume of 0.1 M ammoniumbicarbonate buffer containing 0.5 µg trypsin. Digestion wascarried out for 16 h at 37 °C with an additional aliquot of 0.25µg trypsin added after 12 h. Peptides were extracted with 150µL 60% acetonitrile containing 0.1% TFA for 30 min, followedby a further extraction with acetonitrile. All extracts were pooledand concentrated to about a 10-µL volume in a vaccufuge.About 2 µL of the digest was separated on an Aquasil C18reverse phase column (15 µm tip × 75 µm id × 5 cm Picofritcolumn, New Objectives, Inc) using an acetonitrile/1% aceticacid gradient system (5-75% acetonitrile over 25 min followedby 95% acetonitrile for 3 min) at a flow rate of 300 nl/min.Collision induced dissociation spectra for the most abundantpeaks were collected using an ion-trap mass spectrometer (LCQClassic, ThermoFinnigan). The mass spectrometer was set foranalyzing the positive ions and was operated on double playmode in which the instrument was set up to automaticallyacquire a full scan and a tandem MS/MS spectrum (relativecollision energy ∼30%) on the most intense ion from the fullMS scan. Dynamic exclusion was set to collect two MS/MS
research articles Basrur et al.
70 Journal of Proteome Research • Vol. 2, No. 1, 2003

spectra on the most abundant peak and then exclude it for 2min. An initial database search against a subset of indexedhuman database was carried out using TurboSEQUEST. Theresults of TurboSEQUEST were manually verified and any unin-terpreted collision induced dissociation (CID) spectra weremanually searched using the MS-Tag provision of ProteinProspector (http://prospector.ucsf.edu)21 against a subset ofhuman/mouse proteins in the nonredundant or SwissProtdatabase.
In solution digestion of proteins and removal of melaninfollowed by 2D LC-MS/MS was performed in the followingmanner: Melanosome proteins from 4 × 108 cells weresolubilized in 100 µL of ammonium bicarbonate buffer (100mM, pH 8.5), vortexed for 2 min, sonicated for 30 min, andthen pelleted by centrifugation at 14000g for 10 min. Thesupernatant was digested with modified trypsin (5 µg) (Prome-ga) at 37 °C for 4 h and then at room temperature for anadditional ∼18 h. The digest was then acidified and frozen at-35 °C until analyzed. The membrane-protein pellet wassolubilized in 10% SDS (50 µL) and 10 M urea (50 µL), anddelipidated with a methanol/chloroform mixture as describedpreviously.22 The above solution was treated with methanol (260µL), mixed briefly, treated with chloroform (87 µL) and stirredto yield a single phase. The addition of water (174 µL), withvigorous mixing, afforded two phases that were separated intotwo layers, with a precipitate at the interface, by centrifugation
(10000g for 2 min). The bulk of the upper aqueous methanolphase was removed and the pellet was washed quickly withmethanol (40 µL) to remove pigmented material and was thentreated with 150 µL of methanol and 100 µL of water. Theresulting suspension was mixed vigorously and then centrifuged(10000g for 1 min). The supernatant was removed and theprotein pellet was dried under vacuum on a Speed Vac andredissolved in a mixture of 10 M urea (35 µL) and 10% SDS (25µL), reduced with 40 mM dithiothreitol (10 µL) at 37 °C for 1h, and carboxyamidomethylated with 400 mM iodoacetamide(12 µL) in the dark at room temperature for 1 h. For proteolysis,the sample was diluted with 100 mM ammonium bicarbonate(400 µL) and digested with endo-lys-C (2.6 µg)(Roche) at 37 °Cfor 15 h at pH 8.5. Following dilution of the sample with anadditional aliquot of 100 mM ammonium bicarbonate (2 mL),the mixture was then incubated with trypsin (5 µg) at 37 °C for12 h, acidified to pH 3 with glacial acetic acid and desaltedusing a Poros 20 R2 column (360 × 200 µm) (PerseptiveBiosystems, Cambridge, MA). Peptides were eluted from thecolumn with 50 µL acetonitrile (80%) in 0.1% acetic acid. Theeluate was then evaporated to dryness on a Speed Vac and re-suspended in 0.5% acetic acid (pH 3) before loading it onto aPoros 20 HS cation exchange column (360 × 200 µm). Afterwashing the HS column with 100 µL acetonitrile (40%) in 1%acetic acid (pH 3) and then with 100 µL 0.1% acetic acid,peptides were eluted stepwise with 100 µL of 0 mM, 75 mMand 500 mM KCl in 5 mM potassium phosphate buffer (pH 3)containing 5% acetonitrile.
For 2D LC-tandem MS, aliquots of the above digests wereloaded onto nano-HPLC precolumns (360 × 100 µm fused silicapacked with 5-cm C18 beads (YMC ODS-AQ; Waters) andwashed with 0.1% acetic acid. Precolumns were then connectedto analytical columns (360 × 50 µm fused silica packed with7-cm C18 beads (YMC ODS-AQ; Waters),23 and the sampleswere analyzed on a ThermoFinnigan, LCQ Deca mass spec-trometer equipped with a nano-HPLC microelectrospray ion-ization source as described previously.24
Antibodies. Polyclonal antibodies to VAT-1 (Cys-MQEKKN-VGKVLLVPGPEKQN), oculospanin (Ac-MEEGERSPLLSQETA-GQKPLSVHR-Cys) and FLJ20420 (Ac-MGGTTSTRRVTFEAD-ENE-Cys) were raised in rabbits against the respective KLH-conjugated specific peptide sequences shown in parentheses.Sera from immunized rabbits were affinity purified using eachappropriate peptide coupled with SulfoLink (Pierce ChemicalCo, Rockford, IL). Specificities of antibodies were confirmedby ELISA and by immunoblot assays.
Other antibodies used in this study included: RPEP7h fortyrosinase;25 HMB45 for gp100;26 RPEP13h for gp100;6 PDP forsyntenin (a kind gift from Dr. Rouleau, Center for Research inNeuroscience, McGill University);27 and R-flotillin-1 (BD Bio-sciences, San Diego, CA).
Western Blotting. Proteins (5 µg protein/lane) were sepa-rated on 8% or 14% SDS gels and were transferred electro-phoretically to poly(vinylidene difluoride) membranes (Immo-bilon-P, Millipore Corp., Bedford, MA). The blots were incubatedwith primary antibodies (at 1/500 or 1/1000 dilution) for 1 hat 23 °C, washed several times, and subsequent visualizationof antibody binding was carried out with Enhanced ChemiLu-minescence (Amersham Corp., Arlington Heights, IL), accordingto the manufacturer’s instructions.
Electron Microscopy. Specimens were fixed overnight at 4°C in 2% glutaraldehyde and 2% paraformaldehyde in 0.1 Msodium cacodylate buffer, pH 7.3. The samples were then
Figure 1. Outline of melanosome purification. This flowchartshows the method used to isolate melanosomes at variousstages of maturation from MNT1 melanoma cells; the protocolinvolves homogenization of the cells, purification over a sucrosedensity gradient (Gradient 1) by ultracentrifugation, and (forStage I and II melanosomes) further purification by FFE.4 Thepurified samples were then analyzed by SDS-PAGE followed bytandem mass spectrometry, or by direct solubilization and 2D-LC tandem mass spectrometry as indicated.
Proteomic Analysis of Early Melanosomes research articles
Journal of Proteome Research • Vol. 2, No. 1, 2003 71
stored in PBS at 4 °C until they were embedded in epoxy resin.Thin sections were cut, stained with uranyl acetate and leadcitrate, and examined using a Zeiss EM 912 electron micro-scope.
For immuno-electron microscopy, fractions were reacted for1 h at 23 °C with the primary antibody (at 1:100 dilution),
washed twice in PBS, then incubated with the appropriate 10nm gold-labeled goat anti-rabbit or anti-mouse IgG (at 1:25dilution) for 1 h at 4 °C. Controls included samples with noprimary antibody. The fractions were washed again twice inPBS, then fixed and processed for electron microscopy asdescribed above.
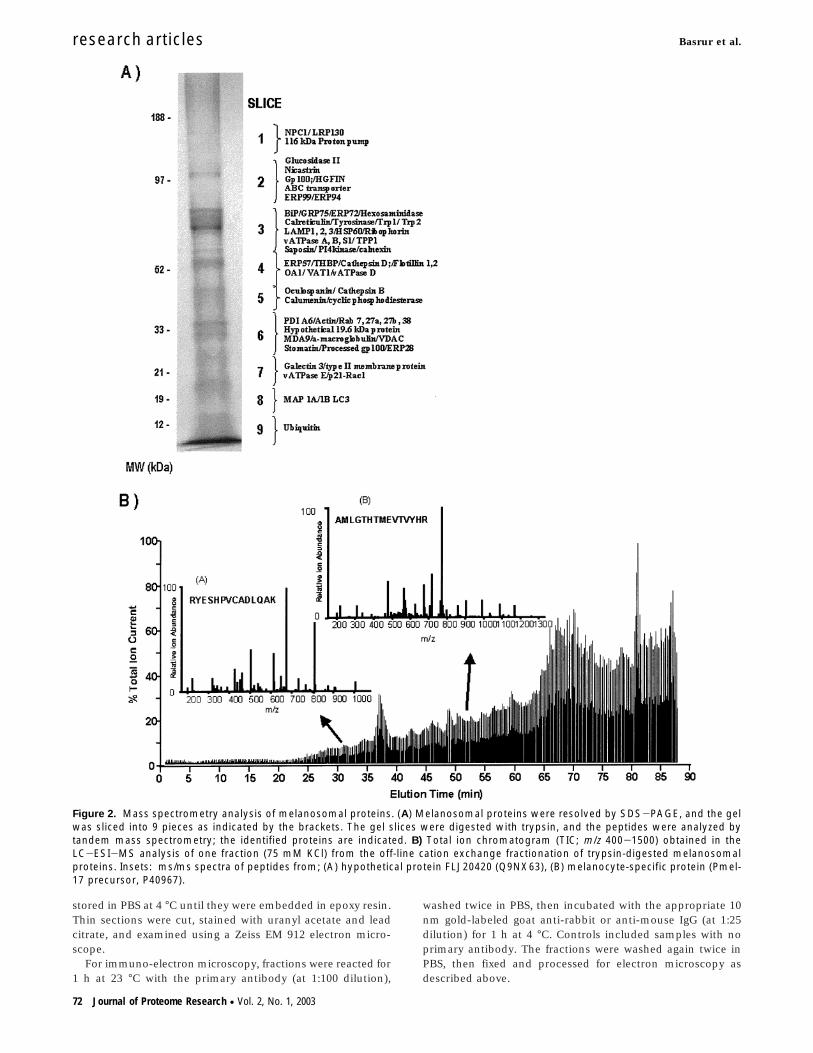
Figure 2. Mass spectrometry analysis of melanosomal proteins. (A) Melanosomal proteins were resolved by SDS-PAGE, and the gelwas sliced into 9 pieces as indicated by the brackets. The gel slices were digested with trypsin, and the peptides were analyzed bytandem mass spectrometry; the identified proteins are indicated. B) Total ion chromatogram (TIC; m/z 400-1500) obtained in theLC-ESI-MS analysis of one fraction (75 mM KCl) from the off-line cation exchange fractionation of trypsin-digested melanosomalproteins. Insets: ms/ms spectra of peptides from; (A) hypothetical protein FLJ20420 (Q9NX63), (B) melanocyte-specific protein (Pmel-17 precursor, P40967).
research articles Basrur et al.
72 Journal of Proteome Research • Vol. 2, No. 1, 2003
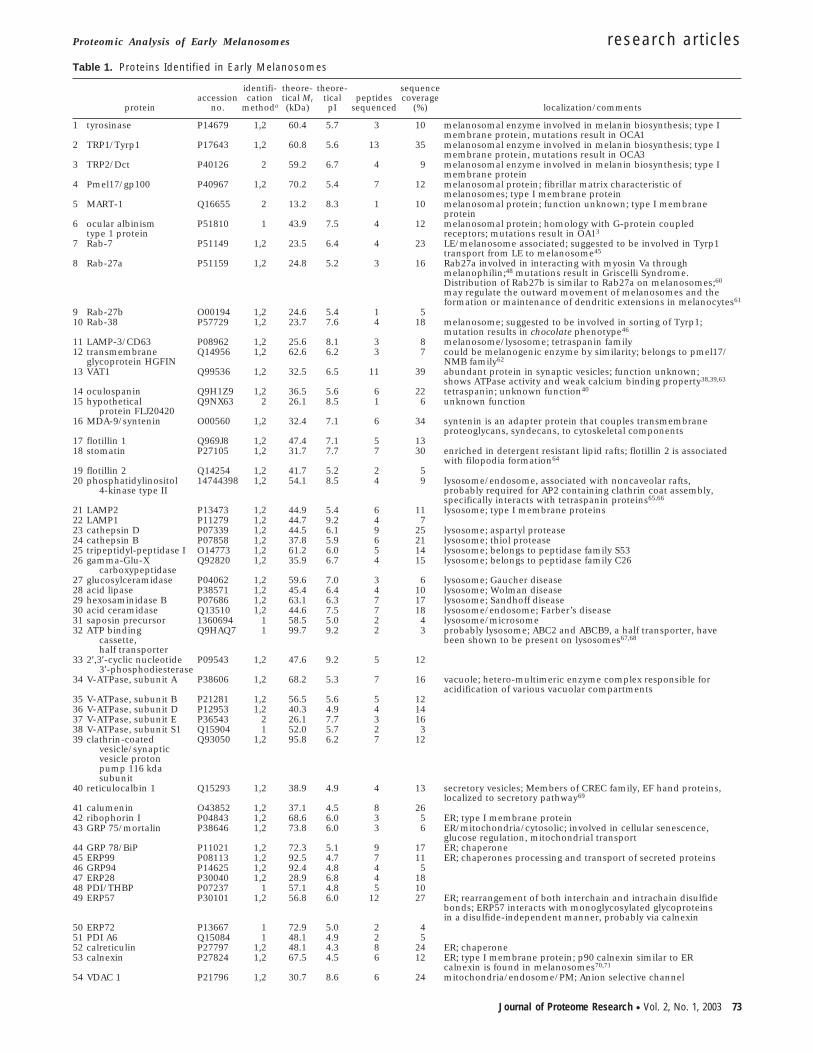
Table 1. Proteins Identified in Early Melanosomes
proteinaccession
no.
identifi-cation
methoda
theore-tical Mr(kDa)
theore-tical
pIpeptides
sequenced
sequencecoverage
(%) localization/comments
1 tyrosinase P14679 1,2 60.4 5.7 3 10 melanosomal enzyme involved in melanin biosynthesis; type Imembrane protein, mutations result in OCA1
2 TRP1/Tyrp1 P17643 1,2 60.8 5.6 13 35 melanosomal enzyme involved in melanin biosynthesis; type Imembrane protein, mutations result in OCA3
3 TRP2/Dct P40126 2 59.2 6.7 4 9 melanosomal enzyme involved in melanin biosynthesis; type Imembrane protein
4 Pmel17/gp100 P40967 1,2 70.2 5.4 7 12 melanosomal protein; fibrillar matrix characteristic ofmelanosomes; type I membrane protein
5 MART-1 Q16655 2 13.2 8.3 1 10 melanosomal protein; function unknown; type I membraneprotein
6 ocular albinismtype 1 protein
P51810 1 43.9 7.5 4 12 melanosomal protein; homology with G-protein coupledreceptors; mutations result in OA13
7 Rab-7 P51149 1,2 23.5 6.4 4 23 LE/melanosome associated; suggested to be involved in Tyrp1transport from LE to melanosome45
8 Rab-27a P51159 1,2 24.8 5.2 3 16 Rab27a involved in interacting with myosin Va throughmelanophilin;48 mutations result in Griscelli Syndrome.Distribution of Rab27b is similar to Rab27a on melanosomes;60
may regulate the outward movement of melanosomes and theformation or maintenance of dendritic extensions in melanocytes61
9 Rab-27b O00194 1,2 24.6 5.4 1 510 Rab-38 P57729 1,2 23.7 7.6 4 18 melanosome; suggested to be involved in sorting of Tyrp1;
mutation results in chocolate phenotype46
11 LAMP-3/CD63 P08962 1,2 25.6 8.1 3 8 melanosome/lysosome; tetraspanin family12 transmembrane
glycoprotein HGFINQ14956 1,2 62.6 6.2 3 7 could be melanogenic enzyme by similarity; belongs to pmel17/
NMB family62
13 VAT1 Q99536 1,2 32.5 6.5 11 39 abundant protein in synaptic vesicles; function unknown;shows ATPase activity and weak calcium binding property38,39,63
14 oculospanin Q9H1Z9 1,2 36.5 5.6 6 22 tetraspanin; unknown function40
15 hypotheticalprotein FLJ20420
Q9NX63 2 26.1 8.5 1 6 unknown function
16 MDA-9/syntenin O00560 1,2 32.4 7.1 6 34 syntenin is an adapter protein that couples transmembraneproteoglycans, syndecans, to cytoskeletal components
17 flotillin 1 Q969J8 1,2 47.4 7.1 5 1318 stomatin P27105 1,2 31.7 7.7 7 30 enriched in detergent resistant lipid rafts; flotillin 2 is associated
with filopodia formation64
19 flotillin 2 Q14254 1,2 41.7 5.2 2 520 phosphatidylinositol
4-kinase type II14744398 1,2 54.1 8.5 4 9 lysosome/endosome, associated with noncaveolar rafts,
probably required for AP2 containing clathrin coat assembly,specifically interacts with tetraspanin proteins65,66
21 LAMP2 P13473 1,2 44.9 5.4 6 11 lysosome; type I membrane proteins22 LAMP1 P11279 1,2 44.7 9.2 4 723 cathepsin D P07339 1,2 44.5 6.1 9 25 lysosome; aspartyl protease24 cathepsin B P07858 1,2 37.8 5.9 6 21 lysosome; thiol protease25 tripeptidyl-peptidase I O14773 1,2 61.2 6.0 5 14 lysosome; belongs to peptidase family S5326 gamma-Glu-X
carboxypeptidaseQ92820 1,2 35.9 6.7 4 15 lysosome; belongs to peptidase family C26
27 glucosylceramidase P04062 1,2 59.6 7.0 3 6 lysosome; Gaucher disease28 acid lipase P38571 1,2 45.4 6.4 4 10 lysosome; Wolman disease29 hexosaminidase B P07686 1,2 63.1 6.3 7 17 lysosome; Sandhoff disease30 acid ceramidase Q13510 1,2 44.6 7.5 7 18 lysosome/endosome; Farber’s disease31 saposin precursor 1360694 1 58.5 5.0 2 4 lysosome/microsome32 ATP binding
cassette,half transporter
Q9HAQ7 1 99.7 9.2 2 3 probably lysosome; ABC2 and ABCB9, a half transporter, havebeen shown to be present on lysosomes67,68
33 2′,3′-cyclic nucleotide3′-phosphodiesterase
P09543 1,2 47.6 9.2 5 12
34 V-ATPase, subunit A P38606 1,2 68.2 5.3 7 16 vacuole; hetero-multimeric enzyme complex responsible foracidification of various vacuolar compartments
35 V-ATPase, subunit B P21281 1,2 56.5 5.6 5 1236 V-ATPase, subunit D P12953 1,2 40.3 4.9 4 1437 V-ATPase, subunit E P36543 2 26.1 7.7 3 1638 V-ATPase, subunit S1 Q15904 1 52.0 5.7 2 339 clathrin-coated
vesicle/synapticvesicle protonpump 116 kdasubunit
Q93050 1,2 95.8 6.2 7 12
40 reticulocalbin 1 Q15293 1,2 38.9 4.9 4 13 secretory vesicles; Members of CREC family, EF hand proteins,localized to secretory pathway69
41 calumenin O43852 1,2 37.1 4.5 8 2642 ribophorin I P04843 1,2 68.6 6.0 3 5 ER; type I membrane protein43 GRP 75/mortalin P38646 1,2 73.8 6.0 3 6 ER/mitochondria/cytosolic; involved in cellular senescence,
glucose regulation, mitochondrial transport44 GRP 78/BiP P11021 1,2 72.3 5.1 9 17 ER; chaperone45 ERP99 P08113 1,2 92.5 4.7 7 11 ER; chaperones processing and transport of secreted proteins46 GRP94 P14625 1,2 92.4 4.8 4 547 ERP28 P30040 1,2 28.9 6.8 4 1848 PDI/THBP P07237 1 57.1 4.8 5 1049 ERP57 P30101 1,2 56.8 6.0 12 27 ER; rearrangement of both interchain and intrachain disulfide
bonds; ERP57 interacts with monoglycosylated glycoproteinsin a disulfide-independent manner, probably via calnexin
50 ERP72 P13667 1 72.9 5.0 2 451 PDI A6 Q15084 1 48.1 4.9 2 552 calreticulin P27797 1,2 48.1 4.3 8 24 ER; chaperone53 calnexin P27824 1,2 67.5 4.5 6 12 ER; type I membrane protein; p90 calnexin similar to ER
calnexin is found in melanosomes70,71
54 VDAC 1 P21796 1,2 30.7 8.6 6 24 mitochondria/endosome/PM; Anion selective channel
Proteomic Analysis of Early Melanosomes research articles
Journal of Proteome Research • Vol. 2, No. 1, 2003 73

Confocal Microscopy and Immunofluorescence. Dual label-ing using immunofluorescence methods and laser scanningconfocal microscopy was employed to evaluate the subcellularlocalization of proteins identified in this study, at the followingdilutions: VAT-1 (1:20), oculospanin (1:25), FLJ20420 (1:25),HMB-45 (1:500), RPEP7 (1:500), RPEP13 (1:500), and flotillin-1(1:10). Chamber slides containing subconfluent MNT-1 mela-noma cells, were stained by the double indirect immunofluo-rescence method, as previously described in detail.4 Briefly,MNT-1 cells were washed twice with 1X PBS, and were thenfixed with 4% paraformaldehyde for 15 min at 4 °C. All proce-dures were performed at room temperature (23-25 °C), exceptwhere indicated. After fixation, cells were washed twice with1X PBS and were immediately permeabilized with 0.01% Triton×100 for 3 min. Nonspecific binding was blocked with 5%normal serum for 1 h. Chamber slides were incubated with amixture containing a polyclonal and a monoclonal antibodyovernight at 4 °C. The polyclonal antibodies were reacted withsecondary antibodies (goat anti-rabbit IgG; Vector; dilution1:100) labeled with Texas red. The monoclonal antibodies werereacted with secondary antibodies (goat anti-mouse IgG; Vec-tor; dilution 1:100) labeled with fluorescein, followed by nuclearcounterstaining with DAPI (Vector Labs, Burlingame, CA).Reactivity was classified into three categories, according to whe-ther they showed green, red, or yellow fluorescence. The latterwas indicative of colocalization of the red and green fluorescentsignals. All preparations were examined with a confocalmicroscope (model TCS-SP/DMRE; Leica, Heidelberg, Ger-many), equipped with HeNe, Argon and krypton laser sources.
ResultsPurification of Early Melanosomes and Protein Identifica-
tion by Mass Spectrometry. Figure 1 shows a schematicsummarizing the approach used to purify and characterizemelanosomal proteins. Basically, the procedure takes advantageof the density of small amounts of melanin in early melano-
somes to purify those organelles over stepwise density gradients(from 1.0 to 2.0 M sucrose). Earlier, we reported the use of FFEto further separate the 1.0 M sucrose fraction (which containsStage I and Stage II early melanosomes) into tyrosinase-richand protein-rich fractions.4 The specific activity of tyrosinasewas enriched about 80-fold in the early melanosomes and thefraction was negative for mitochondria, Golgi, and early en-dosome markers, as determined by Western blot analysis.Ultrastructural analysis further confirmed that the fraction washighly enriched for stage II melanosomes. This FFE-purifiedearly melanosome fraction was used for protein identificationby mass spectrometry.
Our early repeated attempts to establish a reproducibleprofile of melanosomal proteins using 2D PAGE were unsuc-cessful due to the presence of small amounts of the charged,insoluble polymer, melanin. The pigment, by virtue of its abilityto covalently modify proteins,18,19,28,29 renders them insolubleand because many of the melanosomal proteins are membranebound, under the mild solubilization conditions required forthe IEF step of 2D electrophoresis, they are not efficientlysolubilized. Hence, two different approaches were undertakento comprehensively identify the proteome of the melanosome.In the first approach, FFE-enriched melanosomes were sepa-rated by SDS-PAGE, followed by in gel digestion and tandemmass spectrometry. As shown in Figure 2A, the melanosomalextract was resolved by SDS-PAGE into a relatively largenumber of protein bands, although many of those were poorlyresolved from each other. Thus, we cut the gel into 9 slicesand analyzed those slices by the in gel digestion techniquewhich gave positive identification of 61 proteins (this experi-ment was repeated twice with comparable results). The proteincharacteristics and the peptides sequenced for each proteinusing this approach are listed in Table 1 (method 1). However,due to limited resolution of proteins due to the minor amountsof melanin and the presence in the digests of many otherprotein bands in low abundance, we resorted to a second
Table 1 (Continued)
proteinaccession
no.
identifi-cation
methoda
theore-tical Mr(kDa)
theore-tical
pIpeptides
sequenced
sequencecoverage
(%) localization/comments
55 glucosidase II R subunit Q9P0 X0 1,2 109.4 5.8 6 7 ER56 Hsp60 P10809 1,2 61.1 5.7 5 10 mitochondria; chaperone57 CD59/protectin P13987 2 14.1 6.0 2 19 PM/vesicles58 ACTA2 protein/actin,
cardiacQ13707 1,2 36.8 5.2 7 24
59 Ras-related C3botulinum toxinsubstrate 1/p21-Rac1
P15154 2 21.4 8.8 4 30 PM; its homologue cdc42 is suggested to be involved infilopodia formation to facilitate the transfer of melanosomesto keratinocytes13
60 type II membraneprotein
Q9Y2B0 1,2 20.7 4.8 2 16 probably ER/lysosome; contains a signal peptide (1-21),saposin B motif (64-171) and C-terminal HDEL sequence
61 ubiquitin P02248 1,2 8.5 6.6 3 44 apart from signaling proteins for proteosome mediateddegradation, ubiquitin has also been shown to be involved inprotein trafficking within the cell72
62 microtubule-associatedproteins 1A/1Blight chain 3
Q9GZQ8 1 14.5 9.0 1 6 predominantly expressed in neurons and suggested to regulatethe microtubule binding activity of MAP1A/1B
63 galectin 3 Q96J47 1,2 26.1 8.6 5 22 cytosolic/nuclear64 Niemann-Pick C1
proteinO15118 1,2 142.1 5.2 4 3
65 homologue tohypothetical19.5 kDa protein
12847703 1 31.2 8.8 2 10 human fragments: XP•087115: doesn’t have either of thepeptides sequenced. But completely identical to the mouseprotein from the middle to the end of the protein T34524,CAB59180: Have the second peptide sequenced
66 nicastrin Q92542 1,2 78.4 5.7 4 767 LRP 130 P42704 1,2 145.2 5.5 2 2 believed to interact with proteins involved in cytoskeletal
rearrangement and vesicle trafficking68 R-2-macroglobulin P01023 1,2 163.2 6.0 3 2 found at ∼30 kDa range
a 1 ) SDS-PAGE followed by in gel digestion and 1D LC-MS/MS; 2 ) In solution digestion followed by 2D LC-MS/MS.
research articles Basrur et al.
74 Journal of Proteome Research • Vol. 2, No. 1, 2003
approach, which is based on solubilization of proteins aftermelanin removal.
Melanosomes were successfully solubilized as detailed in theMethods section and 2D-LC-tandem MS was performed. Anexample chromatogram depicting the identification of gp100and FLJ20410 using this approach is shown in Figure 2B. Table1 shows the complete repertoire of melanosomal proteinsidentified using these approaches. Sequencing was performedon at least 3 different melanosome preparations and only thoseproteins which were consistently observed either by 1D-LCand/or by 2D-LC approaches are reported in Table 1. Usingboth techniques, we have now identified a total of 68 proteinsin melanosomes which include all 6 of the known melanosome-specific proteins (tyrosinase, Tyrp1, Dct, OA1, gp100, andMART1) whose presence (except for OA1) had already beenconfirmed by Western blot analysis.4
It is evident from these results that in addition to the knownmelanosomal proteins, there are a number of other proteinspresent in melanosomes, most of which are membrane-boundor membrane-associated proteins. These include several typeI membrane proteins (LAMP-1 and LAMP-2), tetraspanins(CD63 and oculospanin), lipid raft associated proteins (stoma-tin, flotillin-1 and flotillin-2), vacuolar proton ATPases (VATPaseA, D, B, E, and S1 and the clathrin coated vesicle/synapticvesicle proton pump), and cytoskeletal and associated proteinsinvolved in vesicular trafficking (actin, MAP1B LC3 and LRP130,
small GTPase family members or related proteins such as rab7,rab27b, rab38, and p21-rac1).
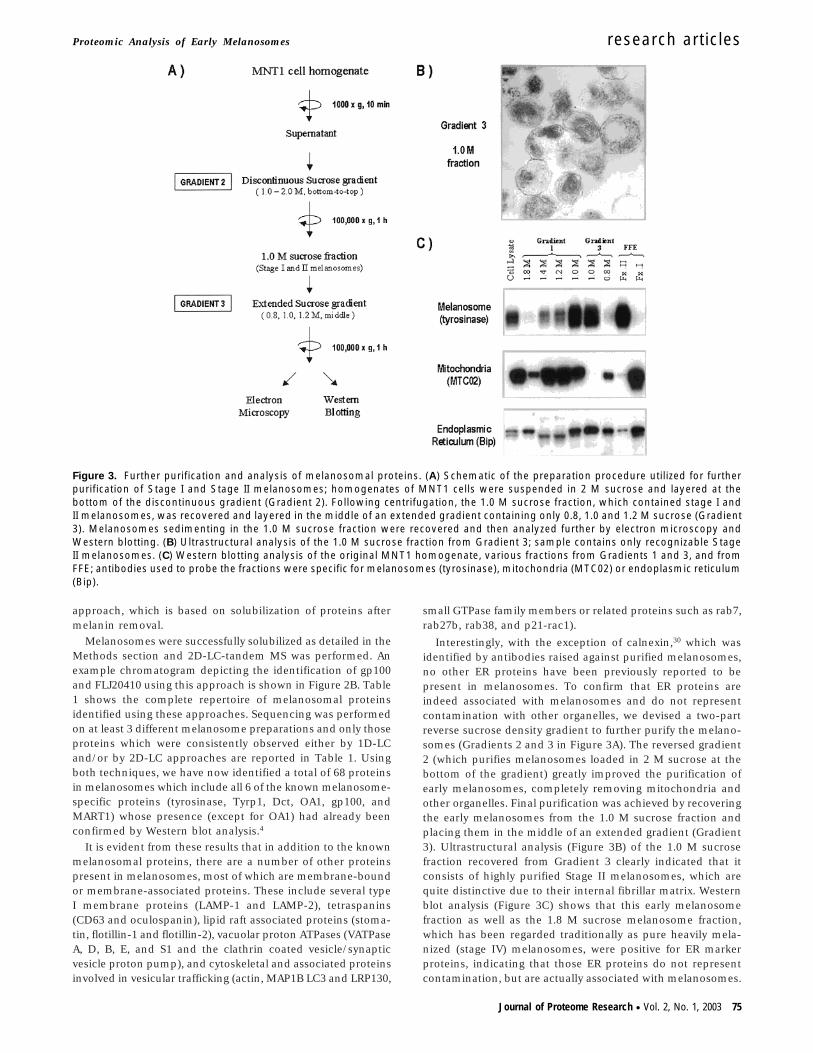
Interestingly, with the exception of calnexin,30 which wasidentified by antibodies raised against purified melanosomes,no other ER proteins have been previously reported to bepresent in melanosomes. To confirm that ER proteins areindeed associated with melanosomes and do not representcontamination with other organelles, we devised a two-partreverse sucrose density gradient to further purify the melano-somes (Gradients 2 and 3 in Figure 3A). The reversed gradient2 (which purifies melanosomes loaded in 2 M sucrose at thebottom of the gradient) greatly improved the purification ofearly melanosomes, completely removing mitochondria andother organelles. Final purification was achieved by recoveringthe early melanosomes from the 1.0 M sucrose fraction andplacing them in the middle of an extended gradient (Gradient3). Ultrastructural analysis (Figure 3B) of the 1.0 M sucrosefraction recovered from Gradient 3 clearly indicated that itconsists of highly purified Stage II melanosomes, which arequite distinctive due to their internal fibrillar matrix. Westernblot analysis (Figure 3C) shows that this early melanosomefraction as well as the 1.8 M sucrose melanosome fraction,which has been regarded traditionally as pure heavily mela-nized (stage IV) melanosomes, were positive for ER markerproteins, indicating that those ER proteins do not representcontamination, but are actually associated with melanosomes.
Figure 3. Further purification and analysis of melanosomal proteins. (A) Schematic of the preparation procedure utilized for furtherpurification of Stage I and Stage II melanosomes; homogenates of MNT1 cells were suspended in 2 M sucrose and layered at thebottom of the discontinuous gradient (Gradient 2). Following centrifugation, the 1.0 M sucrose fraction, which contained stage I andII melanosomes, was recovered and layered in the middle of an extended gradient containing only 0.8, 1.0 and 1.2 M sucrose (Gradient3). Melanosomes sedimenting in the 1.0 M sucrose fraction were recovered and then analyzed further by electron microscopy andWestern blotting. (B) Ultrastructural analysis of the 1.0 M sucrose fraction from Gradient 3; sample contains only recognizable StageII melanosomes. (C) Western blotting analysis of the original MNT1 homogenate, various fractions from Gradients 1 and 3, and fromFFE; antibodies used to probe the fractions were specific for melanosomes (tyrosinase), mitochondria (MTC02) or endoplasmic reticulum(Bip).
Proteomic Analysis of Early Melanosomes research articles
Journal of Proteome Research • Vol. 2, No. 1, 2003 75

Although mitochondria tend to co-purify with early melano-somes in Gradient 1, they were completely removed from earlymelanosomes purified through Gradients 2 and 3, as assessedby Western blot analysis and by electron microscopy. Inaddition, the 1.0 M fraction from Gradient 3 was highlyenriched for the melanosome and ER markers (tyrosinase andBiP, respectively), but was completely negative for the mito-chondrial marker (MTC02).
Subcellular Localization of Novel Melanosomal Proteins.Because it is not practically feasible to validate all proteinsidentified in our preparation (Table 1) as melanosome specificor associated, we focused on a few of them. To determine ifthey are bona fide melanosomal proteins and to determine ifthey co-purify with tyrosinase, the most stringent marker ofmelanosomes, we used Western blotting with specific antibod-ies to those proteins (Figure 4). Fractions examined includedwhole cell lysates, the 1.8, 1.4, 1.2, and 1.0 M sucrose fractionsfrom Gradient 1, the 1.0 and 0.8 M sucrose fractions fromGradient 3, and the purified FFE fractions (the tyrosinase-richFraction II and the protein-rich Fraction I). Western blotanalysis clearly showed co-purification of VAT-1, oculospanin,syntenin and flotillin with tyrosinase-rich melanosomes. Theapparent decreases in the intensities of reactive bands in the1.8 M fraction (containing Stage IV melanized melanosomes),especially with respect to tyrosinase, oculospanin, and syntenin,is probably due to epitope masking by melanin deposition, aswe have previously discussed.4 FLJ20420 and flotillin were notappreciably enriched in the tyrosinase containing fractions,which suggests that while they are associated with melano-somes, they may not be melanosome-specific proteins, butmight just be generally present in many membrane-boundorganelles.
To confirm the subcellular localization of these proteins inmelanosomes, we used RPEP13 and HMB45, antibodies thatrecognize GP100 in Stage I or in Stage II melanosomes,respectively,4 with confocal immunohistochemistry. Antibodiesto tyrosinase, Tyrp1 and Dct, which are traditionally used asspecific markers of melanosomes, and which recognize theseproteins when they are in transit in the ER and other vesicularsorting structures, were also included in our analysis (Figure5). Dual labeling showed that tyrosinase, TYRP1, VAT-1, ocu-lospanin, flotillin, and FLJ20420 co-localized (cf yellow color)with Stage I melanosomes (left column) and/or Stage IImelanosomes (middle and right columns). The punctate dis-tribution and extensive co-localization of VAT-1, oculospanin,flotillin, and FLJ20420 with the melanosomal markers indicatestheir association with early melanosomes. Note that the ERmarkers Bip and KDEL are not only found in the perinucleararea and co-localized with gp100 in early melanosomes, butare also readily detectable in mature melanosomes in theextremities of the melanocytes.
Discussion
This study has demonstrated the utility of combining FFEwith sensitive mass spectrometry techniques to analyze com-plex subcellular components, such as the melanosome, whichshed light on its biogenesis. The combination of the twoapproaches for protein identification, such as in gel or insolution digestion followed by 1D-LC or 2D-LC MS, respec-tively, has resulted in the reproducible identification of proteinsvarying over a wide dynamic range in abundance (from themost abundant Tyrp1 to the low abundant MART1). Table 1details the number of peptides which have been identified foreach protein and whether they were identified by both tech-niques. In addition, the coupling of subcellular localizationusing Western blot analysis and confocal immunohistochem-istry allows the further characterization of novel proteins notonly in melanosomes, but in any organelle of interest. Ideally,one would like to have a suitable control to use for comparisonbut since the melanosome is such a unique organelle, therereally is not such a control. To get around this problem wehave used Western blotting and immunohistochemistry toshow that the six novel proteins identified in this study do infact colocalize with melanosomes.
Proteomic mapping of early melanosomes has identified allsix of the known melanosomal proteins as well as a number ofother common constituents from lysosomes, endosomes andER. This agrees with a number of earlier studies suggesting thatearly Stage I melanosomes are derived directly from the ER-Golgi/endosome network and/or from lysosomes. A link be-tween lysosomes and melanosomes has been widely acceptedin the field and hence finding lysosomal and ER proteins wasnot surprising. Interestingly, a number of novel protein com-ponents were also identified, as summarized in Figure 6. Hence,our approach opens the way for the first time to address theirfunctions in the melanosome and allows new proteins identi-fied (such as VAT-1, oculospanin, syntenin, FLJ20420, andflotillin) to be analyzed for their roles in melanosome structureand/or function.
Our results have already provided some important insightsinto the cellular machinery used to generate this specificorganelle, which shows an admirable conservation of cellularprocesses and energy. At this stage of the analysis, it hasbecome clear that the majority of components of melanosomes
Figure 4. Novel protein analysis in melanosome fractions.Fractions as detailed for Figure 3 were analyzed by Westernblotting using antibodies specific for tyrosinase, VAT-1, ocu-lospanin, syntenin, FLJ20420, and flotillin, as noted in the Figure.
research articles Basrur et al.
76 Journal of Proteome Research • Vol. 2, No. 1, 2003

derive from both lysosomal and ER vesicles, are incorporatedinto early melanosomes, and that only a limited number ofadditional specific proteins are to be found that provide themelanosome with its unique architecture and function. Ourfindings, therefore, are consistent with the ultrastructuralmorphology of melanosomes which begin life as small amor-phous spherical organelles often seen blebbing from the ER.31-34
These Stage I melanosomes have been shown to be positivefor GP100 and ER markers.4 As other melanosome-specificproteins (such as tyrosinase, TYRP1, and DCT) are deliveredvia sorting vesicles to melanosomes, these vesicles coalescewith the maturing melanosomes and the size of that organellegradually increases. Once the organelle matures to Stage II andbecomes competent to produce melanin, the size of themelanosome remains relatively constant and further growthby vesicle fusion is minimal.35-37
The functions of the known melanosome-specific proteinsare relatively well-characterized as structural or enzymaticcomponents that play roles in the synthesis and deposition ofmelanin. In that context, it is interesting to speculate about
the putative functions of other proteins identified in thisanalysis as to what might be their function in melanosomes.For example, VAT-1 was identified more than a decade ago asan abundant membrane protein from Torpedo cholinergicsynaptic vesicles; since then, it has been reported that VAT-1exhibits ATPase38 and low affinity calcium binding activity.39
Whether VAT-1 plays a role in melanocytes with respect to theexocytosis of melanosomes is an exciting possibility to examinein future studies.
As another example, oculospanin belongs to the tetraspaninsuperfamily, which includes 25 other members.40 Interestingly,oculospanin has a sorting signal sequence at its amino terminus(GERSPLL) that closely resembles the sorting signal sequencefound in the carboxyl terminus of GP100 (GENSPLL) respon-sible for its targeting to the melanosome. Because one of thestriking features of the tetraspanins is their ability to formmulti-molecular complexes with each other and with othersurface proteins which has led to the concept of a “tetraspaninweb”, 40 oculospanin along with CD63 may be involved in the
Figure 5. Subcellular localization of novel proteins. The subcellular localization of novel proteins was assessed by confocalimmunohistochemistry. The marker used for Stage I melanosomes was RPEP13 (red, left column), whereas HMB45 was used as amarker for Stage II melanosomes (green, middle and right columns). Antibodies for melanosomes (tyrosinase and Tyrp1) and for ER(KDEL and Bip) are shown, as is the localization of novel proteins (VAT-1, oculospanin, syntenin, FLJ20402, and flotillin). Yellow indicatescolocalization of both antibodies. Shown are the merged images of the red (Texas Red) and green (FITC) antibody stains; yellowindicated colocalization of both antibodies noted.
Proteomic Analysis of Early Melanosomes research articles
Journal of Proteome Research • Vol. 2, No. 1, 2003 77

recruitment of organelle specific proteins and/or membraneremodeling which is crucial for melanosome movement andtransfer.
Further, a number of small GTPases were identified in ourstudy. Many Rab GTPases have only very recently been shownto play important roles in melanosome protein sorting andmovement within melanocytes.41-44 For example, rab7 andrab38 have been implicated in the intracellular sorting ofTyrp1,45,46 which is distinct from the sorting of tyrosinase andDct to that organelle. The localization pattern of rab27b, amember of the rab27 subfamily, which consists of rab27a andrab27b, closely resembles that of rab27a in melanocytes42,47,48
which suggests that the two rab27 proteins are functionalhomologues. Recently, it was demonstrated that rab27a, whichlocalizes on the melanosome membrane, acts as a receptor formyosin Va through its interaction with melanophilin.10,11,47-49
We do not know whether the a and b forms of rab27 havecomplementary or distinct functions in the melanosome andthe obvious possibility that they bind distinct motor proteinshas not escaped us.
Other proteins identified by our analysis that are interestingto discuss include VDAC, and several subunits of vacuolarproton ATPases. Regulation of pH within melanosomes is animportant regulatory factor for melanin production.52-55 Theobservation of melanosomes with aberrant pH in pinkeyed-dilution melanocytes led Puri et al. to suggest that the p protein,a protein with 12 predicted membrane-spanning domains, isan ionic transporter responsible for maintaining low pH withinmelanocyte, which is critical for melanogenesis. In our early
melanosome preparations, we were unable to detect the pprotein by mass spectrometry, although obviously it might bedelivered there at a later stage in melanosome maturation. Toour knowledge, our study provides the first direct evidence ofthe presence of vATPases and VDAC, two pH regulators, whichmight modulate melanosomal pH and, thus, control rates ofmelanin biosynthesis.
Finally, all six known melanosomal proteins serve as mela-noma-specific immune targets56-58 and an interesting challengeis to determine if some of these novel melanosomal proteinscan also serve that function. However, all studies using theknown melanosomal proteins to elicit immune responses tointerfere with the growth of tumors in melanoma patients havebeen encouraging but have not yet been significantly success-ful. The recent demonstration that flotillin,59 one of the novelmelanosomal proteins identified in our study, is up-regulatedin melanoma cells raises the possibility that these proteinsmight prove useful in eliciting immune responses in melanomapatients.
Continuation of this work will be important to further resolvethe patterns of expression of these novel melanosomal proteinsin melanocytes and to elucidate their roles in melanosomestructure and function. Complete characterization of the fullrepertoire of melanosomal proteins should resolve the uniquestructural and functional features of the melanosome, whichis the ultimate goal of proteomics.
Acknowledgment. This work was supported in part byfunding from USPHS GM 37537 (to D.F.H.).
Figure 6. Schematic of melanosomal proteins identified in this study. Hypothetical figure depicting melanosomal proteins identifiedin this study (as listed in Table 1), grouped according to their anticipated function (e.g., pH regulation) or their presence in otherorganelles. The functions of some proteins shown are well-defined, others are conjectural based on predicted properties.
research articles Basrur et al.
78 Journal of Proteome Research • Vol. 2, No. 1, 2003

References
(1) Prota, G. Melanins and Melanogenesis; Academic Press: NewYork, 1992.
(2) Nordlund, J. J.; Boissy, R. E.; Hearing, V. J.; King, R. A.; Ortonne,J. P. The Pigmentary System: Physiology and Pathophysiology;Oxford Univ Press: New York, 1998.
(3) Shen, B.; Samaraweera, P.; Rosenberg, B.; Orlow, S. J. PigmentCell Res. 2001, 14, 243-248.
(4) Kushimoto, T.; Basrur, V.; Matsunaga, J.; Vieira, W. D.; Muller, J.;Appella, E.; Hearing, V. J. Proc. Natl. Acad. Sci. U.S.A. 2001, 98,10 698-10 703.
(5) Marks, M. S.; Seabra, M. C. Nature Rev: Mol. Cell Biol. 2001, 2,1-11.
(6) Kobayashi, T.; Urabe, K.; Orlow, S. J.; Higashi, K.; Imokawa, G.;Kwon, B. S.; Potterf, S. B.; Hearing, V. J. J. Biol. Chem. 1994, 269,29 198-29 205.
(7) Jimbow, K.; Hua, C.; Gomez, P. F.; Hirosaki, K.; Shinoda, K.;Salopek, T.; Matsusaka, H.; Jin, H. Y.; Yamashita, T. Pigment CellRes. 2000, 13, 110-117.
(8) Wu, X.; Hammer, J. A., III Pigment Cell Res. 2000, 13, 241-247.(9) Virador, V.; Muller, J.; Wu, X.; Abdel-Malek, Z. A.; Yu, Z.-X.;
Ferrans, V. J.; Kobayashi, N.; Wakamatsu, K.; Ito, S.; Hammer, J.A.; Hearing, V. J. FASEB J. 2001, 16, 105-107.
(10) Hume, A. N.; Collinson, L. M.; Hopkins, C. R.; Strom, M.; Barral,D. C.; Bossi, G.; Griffiths, G. M.; Seabra, M. C. Traffic 2002, 3,193-202.
(11) Nagashima, K.; Torii, S.; Yi, Z.; Igarashi, M.; Okamoto, K.;Takeuchi, T.; Izumi, T. FEBS Lett. 2002, 517, 233-238.
(12) Barral, D. C.; Ramalho, J. S.; Anders, R.; Hume, A. N.; Knapton,H. J.; Tolmachova, T.; Collinson, L. M.; Goulding, D.; Authi, K.S.; Seabra, M. C. J. Clin. Invest. 2002, 110, 247-257.
(13) Scott, G.; Leopardi, S.; Printup, S.; Madden, B. C. J. Cell Sci. 2002,115, 1441-1451.
(14) Swank, R. T.; Novak, E. K.; McGarry, M. P.; Zhang, Y.; Li, W.;Zhang, Q.; Feng, L. Pigment Cell Res. 2000, 13, 59-67.
(15) Spritz, R. A. Pigment Cell Res. 2000, 13, 15-20.(16) Huizing, M.; Anikster, Y.; Gahl, W. A. Traffic 2000, 1, 823-835.(17) Raposo, G.; Tenza, D.; Murphy, D. M.; Berson, J. F.; Marks, M. S.
J. Cell Biol. 2001, 152, 809-823.(18) Salinas, C.; Garcıa-Borron, J. C.; Solano, F.; Lozano, J. A. Biochim.
Biophys. Acta 1994, 1204, 53-60.(19) Ito, S.; Kato, T.; Fujita, K. Biochem. Pharmacol. 1988, 37, 1707-
1710.(20) Cuomo, M.; Nicotra, M. R.; Apollonj, C.; Fraioli, R.; Giacomini,
P.; Natali, P. G. J. Invest. Dermatol. 1991, 96, 446-451.(21) Clauser, K. R.; Baker, P. R.; Burlingame, A. L. Anal. Chem. 1999,
71, 2871-2882.(22) le Coutre, J.; Whitelegge, J. P.; Gross, A.; Turk, E.; Wright, E. M.;
Kaback, H. R.; Faul, K. F. Biochemistry 2000, 39, 4237-4242.(23) Martin, S. E.; Shabanowitz, J.; Hunt, D. F.; Marto, J. A. Anal. Chem.
2000, 72, 4266-4274.(24) Ficarro, S. B.; McCleland, M. L.; Stukenberg, P. T.; Burke, D. J.;
Ross, M. M.; Shabanowitz, J.; Hunt, D. F.; White, F. M. NatureBiotech. 2002, 20, 301-305.
(25) Jimenez, M.; Tsukamoto, K.; Hearing, V. J. J. Biol. Chem. 1991,266, 1147-1156.
(26) Adema, G. J.; de Boer, A. J.; Hullenaar, R. V.; Benijn, M.; Ruiter,D. J.; Vogel, A. M.; Figdor, C. G. Am. J. Path. 1993, 143, 1579-1585.
(27) Jannatipour, M.; Dion, P.; Khan, S.; Jindal, H.; Fan, X.; Laganiere,J.; Chishti, A. H.; Rouleau, G. A. J. Biol. Chem. 2001, 276, 33 093-33 100.
(28) Donatien, P. D.; Orlow, S. J. Eur. J. Biochem. 1995, 232, 159-164.(29) Mani, I.; Sharma, V.; Tamboli, I.; Raman, G. Pigment Cell Res.
2001, 14, 170-179.(30) Dakour, J.; Vinayagamoorthy, T.; Jimbow, K.; Chen, H.; Luo, D.;
Dixon, W. T.; Munoz, V. Exp. Cell Res. 1993, 209, 288-300.(31) Seiji, M.; Fitzpatrick, T. B.; Birbeck, M. S. C. J. Invest. Dermatol.
1961, 36, 243-252.(32) Seiji, M.; Iwashita, S. J. Invest. Dermatol. 1965, 45, 305-314.(33) Maul, G. G. J. Ultra. Res. 1969, 26, 163-176.(34) Orlow, S. J.; Boissy, R. E.; Moran, D. J.; Pifko-Hirst, S. J. Invest.
Dermatol. 1993, 100, 55-64.
(35) Diment, S.; Eidelman, M.; Rodriguez, G. M.; Orlow, S. J. J. Biol.Chem. 1995, 270, 4213-4215.
(36) Orlow, S. J. J. Invest. Dermatol. 1995, 105, 3-7.(37) Orlow, S. J.; Brilliant, M. H. Exp. Eye Res. 1999, 68, 147-154.(38) Linial, M.; Levius, O. Neurosci. Lett. 1993, 152, 155-157.(39) Levius, O.; Linial, M. Cell. Mol. Neurobiol. 1993, 13, 493-492.(40) Boucheix, C.; Rubinstein, E. Cell. Mol. Life Sci. 2001, 58, 1189-
1205.(41) Araki, K.; Horikawa, T.; Chakraborty, A. K.; Nakagawa, K.; Itoh,
H.; Oka, M.; Funasaka, Y.; Pawelek, J. M.; Ichihashi, M. PigmentCell Res. 2000, 13, 332-336.
(42) Bahadoran, P.; Aberdam, E.; Mantoux, F.; Bille, K.; Busca, R.;Yalman, N.; de Saint Basile, G.; Casaroli, R.; Ortonne, J. P.; Ballotti,R. J. Cell Biol. 2001, 152, 843-849.
(43) Deacon, S. W.; Gelfand, V. I. J. Cell Biol. 2001, 152, F21-F23.(44) Hume, A. N.; Collinson, L. M.; Rapak, A.; Gomes, A. Q.; Hopkins,
C. R.; Seabra, M. C. J. Cell Biol. 2001, 152, 795-808.(45) Gomez, P. F.; Luo, D.; Hirosaki, K.; Shinoda, K.; Yamashita, T.;
Suzuki, J. I.; Otsu, K.; Ishikawa, K.; Jimbow, K. J. Invest. Dermatol.2001, 117, 81-90.
(46) Loftus, S. K.; Larson, D. M.; Baxter, L. L.; Antonellis, A.; Chen, Y.;Wu, X.; Jiang, Y.; Bittner, M.; Hammer, J. A.; Pavan, W. J. Proc.Natl. Acad. Sci. U.S.A. 2002, 99, 4471-4476.
(47) Fukuda, M.; Kuroda, T. S.; Mikoshiba, K. J. Biol. Chem. 2002, 277,12 432-12 436.
(48) Wu, X. S.; Rao, K.; Zhang, H.; Wang, F.; Sellers, J. R.; Matesic, L.E.; Copeland, N. G.; Jenkins, N. A.; Hammer, J. A. Nature CellBiol. 2002, 4, 271-278.
(49) Provance, D. W.; James, T. L.; Mercer, J. A. Traffic 2002, 3, 124-132.
(50) Scott, G.; Zhao, Q. J. Invest. Dermatol. 2001, 116, 296-304.(51) Scott, G. Pigment Cell Res. 2002, 15, 322-330.(52) Puri, N.; Gardner, J. M.; Brilliant, M. H. J. Invest. Dermatol. 2000,
115, 607-613.(53) Fuller, B. B.; Spaulding, D. T.; Smith, D. R. Exp. Cell Res. 2001,
262, 197-208.(54) Ancans, J.; Thody, A. J. FEBS Lett. 2000, 478, 57-60.(55) Manga, P.; Orlow, S. J. Pigment Cell Res. 2001, 14, 362-367.(56) de Vries, T. J.; Smeets, M.; de Graaf, R.; Hou-Jensen, K.; Brocker,
E. B.; Renard, N.; Eggermont, A. M.; van Muijen, G. N. P.; Ruiter,D. J. J. Pathol. 2001, 193, 13-20.
(57) de Vries, T. J.; Trancikova, D.; Ruiter, D. J.; van Muijen, G. N. P.Brit. J. Cancer 1998, 78, 1156-1161.
(58) Orlow, S. J.; Silvers, W. K.; Zhou, B. K.; Mintz, B. Cancer Res. 1998,58, 1521-1523.
(59) Hazarika, P.; George, S.; Prieto, V. G.; McCarty, M.; Bar-Eli, M.Pigment Cell Res. 2002, 15 (Suppl 9), 66-67.
(60) Ramalho, J. S.; Tolmachova, T.; Hume, A. N.; McGuigan, A.;Gregory-Evans, C. Y.; Huxley, C.; Seabra, M. C. BMC Genet. 2001,2, 2.
(61) Chen, Y.; Samaraweera, P.; Sun, T. T.; Kreibich, G.; Orlow, S. J. J.Invest. Dermatol. 2002, 118, 933-940.
(62) Weterman, M. A. J.; Ajubi, N.; van Dinter, I. M.; Degen, W. G.;van Muijen, G. N.; Ruiter, D. J.; Bloemers, H. P. J. Int. J. Cancer1995, 60, 73-81.
(63) Linial, M.; Miller, K.; Scheller, R. H. Neuron 1989, 2, 1265-1273.(64) Hazarika, P.; Dham, N.; Patel, P.; Cho, M.; Weidner, D.; Goldsmith,
L.; Duvic, M. J. Cell. Biochem. 1999, 75, 147-159.(65) Arneson, L. S.; Kunz, J.; Anderson, R. A.; Traub, L. M. J. Biol. Chem.
1999, 274, 17 794-17 805.(66) Yauch, R. L.; Hemler, M. E. Biochem. J. 2000, 351, 629-637.(67) Zhou, C.; Zhao, L.; Inagaki, N.; Guan, J.; Nakajo, S.; Hirabayashi,
T.; Kikuyama, S.; Shioda, S. J. Neurosci. 2001, 21, 849-857.(68) Zhang, F.; Zhang, W.; Liu, L.; Fisher, C. L.; Hui, D.; Childs, S.;
Dorovini-Zis, K.; Ling, V. J. Biol. Chem. 2000, 275, 23 287-23 294.(69) Honore, B.; Vorum, H. FEBS Lett. 2000, 466, 11-18.(70) Dakour, J.; Jimbow, K.; Vinayagamoorthy, T.; Luo, D.; Chen, H.
Melanoma Res. 1993, 3, 331-336.(71) Salopek, T.; Jimbow, K. J. Invest. Dermatol. 1996, Symp Proc, 195-
202.(72) Hicke, L. Cell 2001, 106, 527-530.
PR025562R
Proteomic Analysis of Early Melanosomes research articles
Journal of Proteome Research • Vol. 2, No. 1, 2003 79
Copyright © 2022 FDOKUMEN




















