
Protein farnesylation inhibitors cause donut-shaped cell nuclei attributable to a centrosome...
6
Protein farnesylation inhibitors cause donut-shaped cell nuclei attributable to a centrosome separation defect Valerie L. R. M. Verstraeten a,b,c , Lana A. Peckham a , Michelle Olive d , Brian C. Capell e , Francis S. Collins e , Elizabeth G. Nabel d,e , Stephen G. Young f , Loren G. Fong f , and Jan Lammerding a,1 a Cardiovascular Division, Department of Medicine, Brigham and Women’s Hospital and Harvard Medical School, Cambridge, MA 02139; b Department of Dermatology, Maastricht University Medical Center, 6202 AZ, Maastricht, The Netherlands; c GROW-School for Oncology and Developmental Biology, Maastricht University Medical Center, 6200 MD, Maastricht, The Netherlands; d National Heart, Lung, and Blood Institute and e National Human Genome Research Institute, National Institutes of Health, Bethesda, MD 20892; and f Department of Medicine, University of California, Los Angeles, CA 90095 Edited by Mark T. Groudine, Fred Hutchinson Cancer Research Center, Seattle, WA, and approved February 4, 2011 (received for review December 27, 2010) Despite the success of protein farnesyltransferase inhibitors (FTIs) in the treatment of certain malignancies, their mode of action is incompletely understood. Dissecting the molecular pathways affected by FTIs is important, particularly because this group of drugs is now being tested for the treatment of Hutchinson–Gilford progeria syndrome. In the current study, we show that FTI treat- ment causes a centrosome separation defect, leading to the forma- tion of donut-shaped nuclei in nontransformed cell lines, tumor cell lines, and tissues of FTI-treated mice. Donut-shaped nuclei arise dur- ing chromatin decondensation in late mitosis; subsequently, cells with donut-shaped nuclei exhibit defects in karyokinesis, develop aneuploidy, and are often binucleated. Binucleated cells proliferate slowly. We identified lamin B1 and proteasome-mediated degrada- tion of pericentrin as critical components in FTI-induced “donut formation” and binucleation. Reducing pericentrin expression or ectopic expression of nonfarnesylated lamin B1 was sufficient to elicit donut formation and binucleated cells, whereas blocking pro- teasomal degradation eliminated FTI-induced donut formation. Our studies have uncovered an important role of FTIs on centrosome separation and define pericentrin as a (indirect) target of FTIs af- fecting centrosome position and bipolar spindle formation, likely explaining some of the anticancer effects of these drugs. cell division | nuclear envelope | doughnut-shaped nuclei | antitumor P rotein farnesylation is a posttranslational modification that facilitates the binding of proteins to membrane surfaces. Protein farnesyltransferase catalyzes the addition of a 15-carbon farnesyl lipid to proteins containing a carboxyl-terminal CaaX motif consisting of a cysteine (C) followed by two aliphatic amino acids (aa) and a terminal amino acid residue (X), which is often alanine, serine, methionine, or glutamine (1). Among the most familiar examples of farnesylated proteins are the Ras family of proteins, which require the farnesyl lipid for anchoring the pro- tein to the plasma membrane and for proper protein function. Because mutations in RAS oncogenes are involved in ∼30% of all human cancers and are associated with poor prognosis and treatment outcome (2, 3), protein farnesyltransferase was seen as an attractive target for anticancer drug therapy, prompting the development of many protein farnesyltransferase inhibitors (FTIs). Interestingly, FTIs exhibit significant efficacy in tumor cells in animal models whether or not they have RAS mutations, suggesting that the efficacy of FTIs cannot be attributed solely to their effects on Ras processing and raising the possibility that other mechanisms underlie their anticancer properties. Recently, FTIs have found another potential application in Hutchinson– Gilford progeria syndrome (HGPS). HGPS is a pediatric proge- roid disorder caused by a mutant form of prelamin A that (unlike mature lamin A) retains its farnesyl lipid anchor. Based on salu- tary effects of FTIs on disease phenotypes in mouse models of progeria (4–6), FTIs are now being tested in children with HGPS. Over the past few years, investigators interested in using FTIs for progeria have recorded anecdotal accounts of misshapen cell nuclei when cells are treated with FTIs (7–9). In the current studies, we investigated the effects of FTIs on nuclear shape and structure in more detail. Results Inhibiting Protein Farnesylation Causes Donut-Shaped Nuclei in Vivo and in Vitro. Treatment of primary human skin fibroblasts with an FTI resulted in a high frequency of cells with donut-shaped nuclei (i.e., nuclei with a central hole devoid of chromatin and lined by a nuclear lamina) (Fig. 1). 3D reconstruction based on con- focal images confirmed that the central void penetrates the entire nucleus and is continuous with the cytoplasm; mitochondria and cytoskeletal structures freely pass through the donut hole (Fig. 1 A and B and Movie S1). FTI treatment induced donut- shaped nuclei in fibroblasts from healthy controls and in human cervical cancer (HeLa) cells, human epithelial larynx cancer (HEp2) cells, mouse embryonic fibroblasts (MEFs), and mouse vascular smooth muscle cells (Fig. 1C), indicating that the effect does not depend on the expression of a mutant form of lamin A. To determine if the formation of donut-shaped nuclei is a com- pound-related effect linked to a specific FTI or the direct result of inhibiting protein farnesylation, we evaluated nuclear shape after treatment with (i ) different FTIs; (ii ) drugs, such as statins and aminobisphosphonates, that inhibit protein prenylation by af- fecting other enzymatic steps (8, 10, 11); (iii ) triple-drug combi- nations of the aforementioned drugs, which are currently being tested in children with HGPS; and (iv) shRNA-mediated knock- down of the catalytic β-subunit of protein farnesyltransferase (FNTB). The migration of HDJ-2 on SDS/polyacrylamide gels was used to assess the level of enzyme inhibition (4, 5). Impor- tantly, even low levels of inhibition of protein farnesylation resulted in donut-shaped nuclei, regardless of the treatment ap- proach (Fig. 1D and Figs. S1–S3). These data indicate that the reduction in protein farnesylation, and not any compound-related effect, accounts for the nuclear shape abnormality. To determine if an FTI could also elicit donut-shaped cell nuclei in vivo, we treated mice with an FTI for 6–12 mo (6). FTI treatment resulted in donut-shaped cell nuclei in the intestines and the skin, con- Author contributions: V.L.R.M.V. and J.L. designed research; V.L.R.M.V., L.A.P., M.O., B.C.C., L.G.F., and J.L. performed research; F.S.C., E.G.N., S.G.Y., and L.G.F. contributed new reagents/analytic tools; V.L.R.M.V., L.A.P., and J.L. analyzed data; and V.L.R.M.V., S.G.Y., and J.L. wrote the paper. The authors declare no conflict of interest. This article is a PNAS Direct Submission. 1 To whom correspondence should be addressed. E-mail: [email protected]. edu. This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10. 1073/pnas.1019532108/-/DCSupplemental. www.pnas.org/cgi/doi/10.1073/pnas.1019532108 PNAS Early Edition | 1 of 6 MEDICAL SCIENCES
Transcript of Protein farnesylation inhibitors cause donut-shaped cell nuclei attributable to a centrosome...

Protein farnesylation inhibitors cause donut-shapedcell nuclei attributable to a centrosomeseparation defectValerie L. R. M. Verstraetena,b,c, Lana A. Peckhama, Michelle Olived, Brian C. Capelle, Francis S. Collinse,Elizabeth G. Nabeld,e, Stephen G. Youngf, Loren G. Fongf, and Jan Lammerdinga,1
aCardiovascular Division, Department of Medicine, Brigham and Women’s Hospital and Harvard Medical School, Cambridge, MA 02139; bDepartment ofDermatology, Maastricht University Medical Center, 6202 AZ, Maastricht, The Netherlands; cGROW-School for Oncology and Developmental Biology,Maastricht University Medical Center, 6200 MD, Maastricht, The Netherlands; dNational Heart, Lung, and Blood Institute and eNational Human GenomeResearch Institute, National Institutes of Health, Bethesda, MD 20892; and fDepartment of Medicine, University of California, Los Angeles, CA 90095
Edited by Mark T. Groudine, Fred Hutchinson Cancer Research Center, Seattle, WA, and approved February 4, 2011 (received for review December 27, 2010)
Despite the success of protein farnesyltransferase inhibitors (FTIs)in the treatment of certain malignancies, their mode of actionis incompletely understood. Dissecting the molecular pathwaysaffected by FTIs is important, particularly because this group ofdrugs is now being tested for the treatment of Hutchinson–Gilfordprogeria syndrome. In the current study, we show that FTI treat-ment causes a centrosome separation defect, leading to the forma-tion of donut-shaped nuclei in nontransformed cell lines, tumor celllines, and tissues of FTI-treated mice. Donut-shaped nuclei arise dur-ing chromatin decondensation in late mitosis; subsequently, cellswith donut-shaped nuclei exhibit defects in karyokinesis, developaneuploidy, and are often binucleated. Binucleated cells proliferateslowly. We identified lamin B1 and proteasome-mediated degrada-tion of pericentrin as critical components in FTI-induced “donutformation” and binucleation. Reducing pericentrin expression orectopic expression of nonfarnesylated lamin B1 was sufficient toelicit donut formation and binucleated cells, whereas blocking pro-teasomal degradation eliminated FTI-induced donut formation. Ourstudies have uncovered an important role of FTIs on centrosomeseparation and define pericentrin as a (indirect) target of FTIs af-fecting centrosome position and bipolar spindle formation, likelyexplaining some of the anticancer effects of these drugs.
cell division | nuclear envelope | doughnut-shaped nuclei | antitumor
Protein farnesylation is a posttranslational modification thatfacilitates the binding of proteins to membrane surfaces.
Protein farnesyltransferase catalyzes the addition of a 15-carbonfarnesyl lipid to proteins containing a carboxyl-terminal CaaXmotif consisting of a cysteine (C) followed by two aliphatic aminoacids (aa) and a terminal amino acid residue (X), which is oftenalanine, serine, methionine, or glutamine (1). Among the mostfamiliar examples of farnesylated proteins are the Ras family ofproteins, which require the farnesyl lipid for anchoring the pro-tein to the plasma membrane and for proper protein function.Because mutations in RAS oncogenes are involved in ∼30% of allhuman cancers and are associated with poor prognosis andtreatment outcome (2, 3), protein farnesyltransferase was seenas an attractive target for anticancer drug therapy, promptingthe development of many protein farnesyltransferase inhibitors(FTIs). Interestingly, FTIs exhibit significant efficacy in tumorcells in animal models whether or not they have RAS mutations,suggesting that the efficacy of FTIs cannot be attributed solely totheir effects on Ras processing and raising the possibility thatother mechanisms underlie their anticancer properties. Recently,FTIs have found another potential application in Hutchinson–Gilford progeria syndrome (HGPS). HGPS is a pediatric proge-roid disorder caused by a mutant form of prelamin A that (unlikemature lamin A) retains its farnesyl lipid anchor. Based on salu-tary effects of FTIs on disease phenotypes in mouse models ofprogeria (4–6), FTIs are now being tested in children with HGPS.
Over the past few years, investigators interested in using FTIsfor progeria have recorded anecdotal accounts of misshapen cellnuclei when cells are treated with FTIs (7–9). In the currentstudies, we investigated the effects of FTIs on nuclear shape andstructure in more detail.
ResultsInhibiting Protein Farnesylation Causes Donut-Shaped Nuclei in Vivoand in Vitro. Treatment of primary human skin fibroblasts with anFTI resulted in a high frequency of cells with donut-shaped nuclei(i.e., nuclei with a central hole devoid of chromatin and linedby a nuclear lamina) (Fig. 1). 3D reconstruction based on con-focal images confirmed that the central void penetrates the entirenucleus and is continuous with the cytoplasm; mitochondriaand cytoskeletal structures freely pass through the donut hole(Fig. 1 A and B and Movie S1). FTI treatment induced donut-shaped nuclei in fibroblasts from healthy controls and in humancervical cancer (HeLa) cells, human epithelial larynx cancer(HEp2) cells, mouse embryonic fibroblasts (MEFs), and mousevascular smooth muscle cells (Fig. 1C), indicating that the effectdoes not depend on the expression of a mutant form of lamin A.To determine if the formation of donut-shaped nuclei is a com-pound-related effect linked to a specific FTI or the direct result ofinhibiting protein farnesylation, we evaluated nuclear shape aftertreatment with (i) different FTIs; (ii) drugs, such as statins andaminobisphosphonates, that inhibit protein prenylation by af-fecting other enzymatic steps (8, 10, 11); (iii) triple-drug combi-nations of the aforementioned drugs, which are currently beingtested in children with HGPS; and (iv) shRNA-mediated knock-down of the catalytic β-subunit of protein farnesyltransferase(FNTB). The migration of HDJ-2 on SDS/polyacrylamide gelswas used to assess the level of enzyme inhibition (4, 5). Impor-tantly, even low levels of inhibition of protein farnesylationresulted in donut-shaped nuclei, regardless of the treatment ap-proach (Fig. 1D and Figs. S1–S3). These data indicate that thereduction in protein farnesylation, and not any compound-relatedeffect, accounts for the nuclear shape abnormality. To determineif an FTI could also elicit donut-shaped cell nuclei in vivo, wetreated mice with an FTI for 6–12 mo (6). FTI treatment resultedin donut-shaped cell nuclei in the intestines and the skin, con-
Author contributions: V.L.R.M.V. and J.L. designed research; V.L.R.M.V., L.A.P., M.O.,B.C.C., L.G.F., and J.L. performed research; F.S.C., E.G.N., S.G.Y., and L.G.F. contributednew reagents/analytic tools; V.L.R.M.V., L.A.P., and J.L. analyzed data; and V.L.R.M.V.,S.G.Y., and J.L. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.1To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1019532108/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1019532108 PNAS Early Edition | 1 of 6
MED
ICALSC
IENCE
S
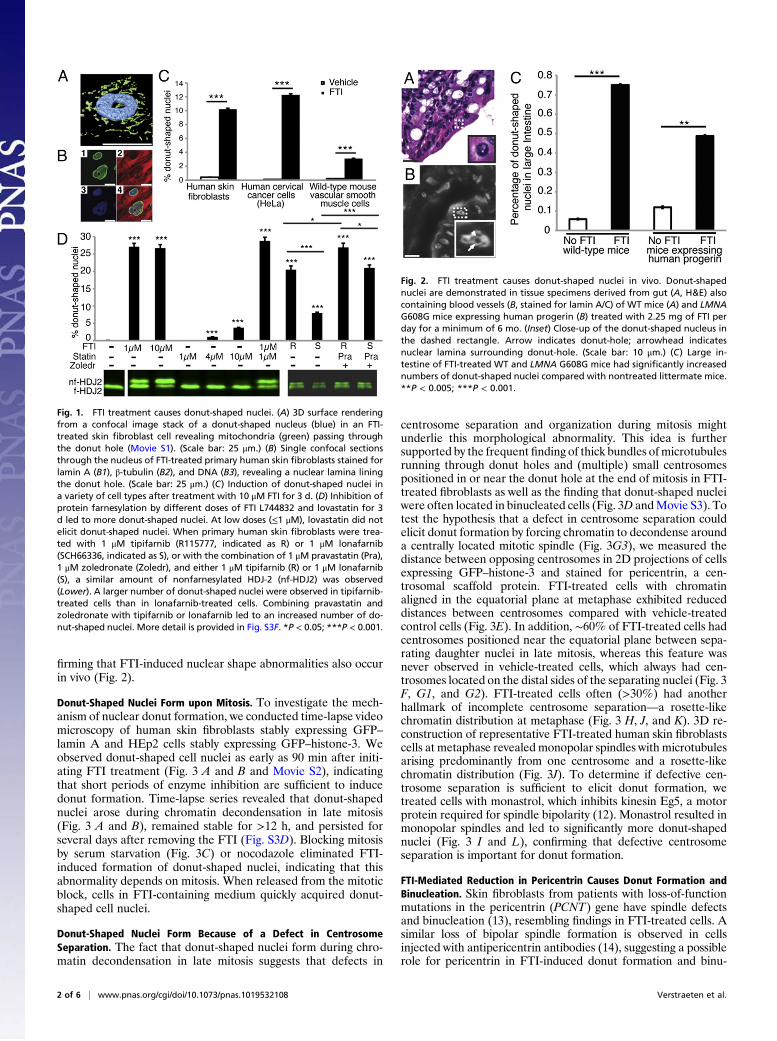
firming that FTI-induced nuclear shape abnormalities also occurin vivo (Fig. 2).
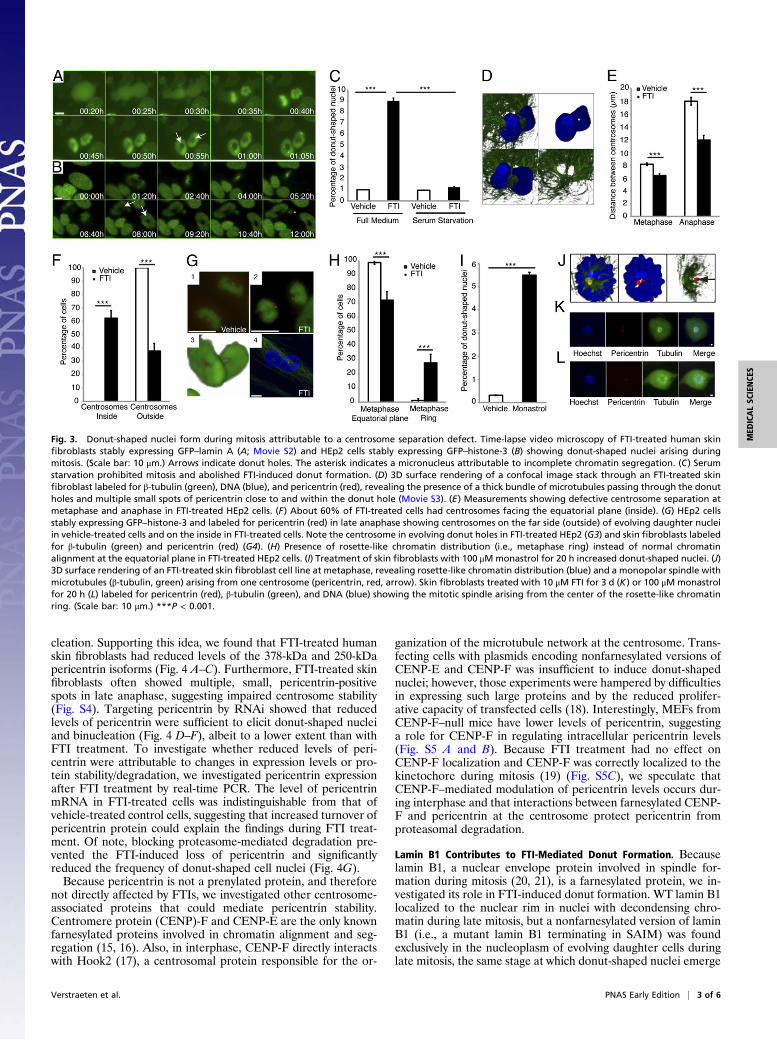
Donut-Shaped Nuclei Form upon Mitosis. To investigate the mech-anism of nuclear donut formation, we conducted time-lapse videomicroscopy of human skin fibroblasts stably expressing GFP–lamin A and HEp2 cells stably expressing GFP–histone-3. Weobserved donut-shaped cell nuclei as early as 90 min after initi-ating FTI treatment (Fig. 3 A and B and Movie S2), indicatingthat short periods of enzyme inhibition are sufficient to inducedonut formation. Time-lapse series revealed that donut-shapednuclei arose during chromatin decondensation in late mitosis(Fig. 3 A and B), remained stable for >12 h, and persisted forseveral days after removing the FTI (Fig. S3D). Blocking mitosisby serum starvation (Fig. 3C) or nocodazole eliminated FTI-induced formation of donut-shaped nuclei, indicating that thisabnormality depends on mitosis. When released from the mitoticblock, cells in FTI-containing medium quickly acquired donut-shaped cell nuclei.
Donut-Shaped Nuclei Form Because of a Defect in CentrosomeSeparation. The fact that donut-shaped nuclei form during chro-matin decondensation in late mitosis suggests that defects in
centrosome separation and organization during mitosis mightunderlie this morphological abnormality. This idea is furthersupported by the frequent finding of thick bundles ofmicrotubulesrunning through donut holes and (multiple) small centrosomespositioned in or near the donut hole at the end of mitosis in FTI-treated fibroblasts as well as the finding that donut-shaped nucleiwere often located in binucleated cells (Fig. 3D andMovie S3). Totest the hypothesis that a defect in centrosome separation couldelicit donut formation by forcing chromatin to decondense arounda centrally located mitotic spindle (Fig. 3G3), we measured thedistance between opposing centrosomes in 2D projections of cellsexpressing GFP–histone-3 and stained for pericentrin, a cen-trosomal scaffold protein. FTI-treated cells with chromatinaligned in the equatorial plane at metaphase exhibited reduceddistances between centrosomes compared with vehicle-treatedcontrol cells (Fig. 3E). In addition, ∼60% of FTI-treated cells hadcentrosomes positioned near the equatorial plane between sepa-rating daughter nuclei in late mitosis, whereas this feature wasnever observed in vehicle-treated cells, which always had cen-trosomes located on the distal sides of the separating nuclei (Fig. 3F, G1, and G2). FTI-treated cells often (>30%) had anotherhallmark of incomplete centrosome separation—a rosette-likechromatin distribution at metaphase (Fig. 3 H, J, and K). 3D re-construction of representative FTI-treated human skin fibroblastscells at metaphase revealed monopolar spindles with microtubulesarising predominantly from one centrosome and a rosette-likechromatin distribution (Fig. 3J). To determine if defective cen-trosome separation is sufficient to elicit donut formation, wetreated cells with monastrol, which inhibits kinesin Eg5, a motorprotein required for spindle bipolarity (12). Monastrol resulted inmonopolar spindles and led to significantly more donut-shapednuclei (Fig. 3 I and L), confirming that defective centrosomeseparation is important for donut formation.
FTI-Mediated Reduction in Pericentrin Causes Donut Formation andBinucleation. Skin fibroblasts from patients with loss-of-functionmutations in the pericentrin (PCNT) gene have spindle defectsand binucleation (13), resembling findings in FTI-treated cells. Asimilar loss of bipolar spindle formation is observed in cellsinjected with antipericentrin antibodies (14), suggesting a possiblerole for pericentrin in FTI-induced donut formation and binu-
Fig. 1. FTI treatment causes donut-shaped nuclei. (A) 3D surface renderingfrom a confocal image stack of a donut-shaped nucleus (blue) in an FTI-treated skin fibroblast cell revealing mitochondria (green) passing throughthe donut hole (Movie S1). (Scale bar: 25 μm.) (B) Single confocal sectionsthrough the nucleus of FTI-treated primary human skin fibroblasts stained forlamin A (B1), β-tubulin (B2), and DNA (B3), revealing a nuclear lamina liningthe donut hole. (Scale bar: 25 μm.) (C) Induction of donut-shaped nuclei ina variety of cell types after treatment with 10 μM FTI for 3 d. (D) Inhibition ofprotein farnesylation by different doses of FTI L744832 and lovastatin for 3d led to more donut-shaped nuclei. At low doses (≤1 μM), lovastatin did notelicit donut-shaped nuclei. When primary human skin fibroblasts were trea-ted with 1 μM tipifarnib (R115777, indicated as R) or 1 μM lonafarnib(SCH66336, indicated as S), or with the combination of 1 μM pravastatin (Pra),1 μM zoledronate (Zoledr), and either 1 μM tipifarnib (R) or 1 μM lonafarnib(S), a similar amount of nonfarnesylated HDJ-2 (nf-HDJ2) was observed(Lower). A larger number of donut-shaped nuclei were observed in tipifarnib-treated cells than in lonafarnib-treated cells. Combining pravastatin andzoledronate with tipifarnib or lonafarnib led to an increased number of do-nut-shaped nuclei. More detail is provided in Fig. S3F. *P < 0.05; ***P < 0.001.
Fig. 2. FTI treatment causes donut-shaped nuclei in vivo. Donut-shapednuclei are demonstrated in tissue specimens derived from gut (A, H&E) alsocontaining blood vessels (B, stained for lamin A/C) of WT mice (A) and LMNAG608G mice expressing human progerin (B) treated with 2.25 mg of FTI perday for a minimum of 6 mo. (Inset) Close-up of the donut-shaped nucleus inthe dashed rectangle. Arrow indicates donut-hole; arrowhead indicatesnuclear lamina surrounding donut-hole. (Scale bar: 10 μm.) (C) Large in-testine of FTI-treated WT and LMNA G608G mice had significantly increasednumbers of donut-shaped nuclei compared with nontreated littermate mice.**P < 0.005; ***P < 0.001.
2 of 6 | www.pnas.org/cgi/doi/10.1073/pnas.1019532108 Verstraeten et al.
cleation. Supporting this idea, we found that FTI-treated humanskin fibroblasts had reduced levels of the 378-kDa and 250-kDapericentrin isoforms (Fig. 4 A–C). Furthermore, FTI-treated skinfibroblasts often showed multiple, small, pericentrin-positivespots in late anaphase, suggesting impaired centrosome stability(Fig. S4). Targeting pericentrin by RNAi showed that reducedlevels of pericentrin were sufficient to elicit donut-shaped nucleiand binucleation (Fig. 4 D–F), albeit to a lower extent than withFTI treatment. To investigate whether reduced levels of peri-centrin were attributable to changes in expression levels or pro-tein stability/degradation, we investigated pericentrin expressionafter FTI treatment by real-time PCR. The level of pericentrinmRNA in FTI-treated cells was indistinguishable from that ofvehicle-treated control cells, suggesting that increased turnover ofpericentrin protein could explain the findings during FTI treat-ment. Of note, blocking proteasome-mediated degradation pre-vented the FTI-induced loss of pericentrin and significantlyreduced the frequency of donut-shaped cell nuclei (Fig. 4G).Because pericentrin is not a prenylated protein, and therefore
not directly affected by FTIs, we investigated other centrosome-associated proteins that could mediate pericentrin stability.Centromere protein (CENP)-F and CENP-E are the only knownfarnesylated proteins involved in chromatin alignment and seg-regation (15, 16). Also, in interphase, CENP-F directly interactswith Hook2 (17), a centrosomal protein responsible for the or-
ganization of the microtubule network at the centrosome. Trans-fecting cells with plasmids encoding nonfarnesylated versions ofCENP-E and CENP-F was insufficient to induce donut-shapednuclei; however, those experiments were hampered by difficultiesin expressing such large proteins and by the reduced prolifer-ative capacity of transfected cells (18). Interestingly, MEFs fromCENP-F–null mice have lower levels of pericentrin, suggestinga role for CENP-F in regulating intracellular pericentrin levels(Fig. S5 A and B). Because FTI treatment had no effect onCENP-F localization and CENP-F was correctly localized to thekinetochore during mitosis (19) (Fig. S5C), we speculate thatCENP-F–mediated modulation of pericentrin levels occurs dur-ing interphase and that interactions between farnesylated CENP-F and pericentrin at the centrosome protect pericentrin fromproteasomal degradation.
Lamin B1 Contributes to FTI-Mediated Donut Formation. Becauselamin B1, a nuclear envelope protein involved in spindle for-mation during mitosis (20, 21), is a farnesylated protein, we in-vestigated its role in FTI-induced donut formation. WT lamin B1localized to the nuclear rim in nuclei with decondensing chro-matin during late mitosis, but a nonfarnesylated version of laminB1 (i.e., a mutant lamin B1 terminating in SAIM) was foundexclusively in the nucleoplasm of evolving daughter cells duringlate mitosis, the same stage at which donut-shaped nuclei emerge
Fig. 3. Donut-shaped nuclei form during mitosis attributable to a centrosome separation defect. Time-lapse video microscopy of FTI-treated human skinfibroblasts stably expressing GFP–lamin A (A; Movie S2) and HEp2 cells stably expressing GFP–histone-3 (B) showing donut-shaped nuclei arising duringmitosis. (Scale bar: 10 μm.) Arrows indicate donut holes. The asterisk indicates a micronucleus attributable to incomplete chromatin segregation. (C) Serumstarvation prohibited mitosis and abolished FTI-induced donut formation. (D) 3D surface rendering of a confocal image stack through an FTI-treated skinfibroblast labeled for β-tubulin (green), DNA (blue), and pericentrin (red), revealing the presence of a thick bundle of microtubules passing through the donutholes and multiple small spots of pericentrin close to and within the donut hole (Movie S3). (E) Measurements showing defective centrosome separation atmetaphase and anaphase in FTI-treated HEp2 cells. (F) About 60% of FTI-treated cells had centrosomes facing the equatorial plane (inside). (G) HEp2 cellsstably expressing GFP–histone-3 and labeled for pericentrin (red) in late anaphase showing centrosomes on the far side (outside) of evolving daughter nucleiin vehicle-treated cells and on the inside in FTI-treated cells. Note the centrosome in evolving donut holes in FTI-treated HEp2 (G3) and skin fibroblasts labeledfor β-tubulin (green) and pericentrin (red) (G4). (H) Presence of rosette-like chromatin distribution (i.e., metaphase ring) instead of normal chromatinalignment at the equatorial plane in FTI-treated HEp2 cells. (I) Treatment of skin fibroblasts with 100 μMmonastrol for 20 h increased donut-shaped nuclei. (J)3D surface rendering of an FTI-treated skin fibroblast cell line at metaphase, revealing rosette-like chromatin distribution (blue) and a monopolar spindle withmicrotubules (β-tubulin, green) arising from one centrosome (pericentrin, red, arrow). Skin fibroblasts treated with 10 μM FTI for 3 d (K) or 100 μM monastrolfor 20 h (L) labeled for pericentrin (red), β-tubulin (green), and DNA (blue) showing the mitotic spindle arising from the center of the rosette-like chromatinring. (Scale bar: 10 μm.) ***P < 0.001.
Verstraeten et al. PNAS Early Edition | 3 of 6
MED
ICALSC
IENCE
S

(Fig. 4K and Fig. S6). Importantly, transient expression of laminB1-SAIM was sufficient to induce formation of donut-shapednuclei in the absence of an FTI (Fig. 4J). Also, lamin B1-deficient MEFs (Lmnb1Δ/Δ) and cells in which lamin B1 expres-sion was knocked down by RNAi did not develop donut-shapednuclei in the presence of an FTI (Fig. 4H and I), providing furthersupport for a role of lamin B1 in the formation of donut-shapednuclei. In contrast, MEFs lacking lamin B2 (Lmnb2−/−), MEFslacking exclusively lamin A [“lamin C-only cells” (22)], or MEFsexpressing a nonfarnesylated version of prelamin A (23) formeddonut-shaped nuclei when incubated with an FTI (Fig. 4H andFig. S7). Thus, formation of donut-shaped cell nuclei requireslamin B1 but not the other lamins.
Donut-Shaped Nuclei Are Mostly Found in Binucleated Cells. FTItreatment increased the number of binucleated cells, with most ofthem also having donut-shaped nuclei (Fig. 5 A and B). In fact,>70% of donut-shaped nuclei were found in binucleated cells(Fig. 5C). Bayesian analysis revealed that the probability of ob-serving donut-shaped nuclei in binucleated cells was ∼3.5 timeshigher than would be predicted for independent events, in-dicating that the formation of binucleated cells and donut-shapednuclei is correlated (Fig. 5 D and E and SI Materials and Meth-ods). To assess the functional consequences of donut-shapednuclei and increased binuclear cells, we followed the fate of thesecells for up to 4 d (Fig. 5 F and G). Binucleated cells had a lowerpropensity to divide, whereas mononucleated cells with donut-shaped nuclei divided at the same rate as cells with normallyshaped nuclei (Fig. 5G).
Donut-Shaped Nuclei Divide Abnormally and Exhibit Aneuploidy.Time-lapse video microscopy revealed that ∼50% of cells withdonut-shaped nuclei divided abnormally (Fig. 5H), with most ofthe abnormal divisions yielding cells with donut-shaped nuclei ormultiple nuclei (Fig. 5I and Fig. S8A). Aside from defects in cy-tokinesis, FTI treatment also interfered with karyokinesis (Fig. 5J,Fig. S8E, and Movie S4). Consequently, donut-shaped nucleiexhibitedmore aneuploidy than cells with normal cell nuclei (Fig. 5K and L). Interestingly, despite the dramatic effects of FTI treat-ment on nuclear shape, we did not observe increased apoptosis inFTI-treated primary skin fibroblasts. Conversely, the nuclei of FTI-treated HEK293 cells exhibited DNA fragmentation.
DiscussionDespite well-documented antiproliferative and proapoptoticeffects of FTIs on cancer cells (24), their mode of antitumor ac-tivity has remained elusive, in part because of the large number(>100) of protein substrates for protein farnesyltransferase (25,26). Induction of apoptosis and loss of bipolar spindle formationwith arrest of cells in G2-M phase with a rosette-like chromatindistribution have been reported previously in the setting of FTItreatment (24, 27, 28), without any mechanism. Our studies con-firm that FTI treatment causes loss of bipolar spindle formationbut show that different cell types, including HeLa and HEp2cancer cells, go on to develop donut-shaped nuclei and bi-nucleated cells. Donut-shaped nuclei could also be detected invivo in tissues of FTI-treated mice. Interestingly, the PKCβII-selective inhibitor βIIV5-3, a drug used for treating prostate can-cer, acts partly by reducing pericentrin levels (29). Hence, reducingpericentrin expression could constitute one mechanism for the
Fig. 4. FTI-induced loss of pericentrin causes formation of donut-shaped nuclei and binucleation. (A) Reduced expression of the large (378 kDa) and small(250 kDa) pericentrin (PCNT) isoforms in FTI-treated primary human skin fibroblasts. MW, molecular weight. Quantification of 378-kDa (B) and 250-kDa (C)PCNT expression levels normalized to β-tubulin and compared with primary skin fibroblasts treated with vehicle alone. (D) RNA interference directed againstPCNT resulted in reduced levels of the 378-kDa PCNT product in HeLa cells. Knockdown of PCNT significantly increased binucleation (E) and donut formation(F). (G) Blocking proteasome-mediated degradation reduced donut formation in FTI-treated primary skin fibroblasts. PI, protease inhibitor. (H) Donut for-mation in MEFs lacking lamin B1 (Lmnb1Δ/Δ) or lamin B2 (Lmnb2−/−) and WT controls. (I) Formation of donut-shaped nuclei in HeLa cells after RNAi directedagainst lamin B1, indicating that lamin B1 is required for FTI-induced donut formation. (J) Percentage of cells with donut-shaped nuclei after expression ofa nonfarnesylated version of lamin B1 (LaB1-SAIM) or WT lamin B1. Expression of LaB1-SAIM and, to a lesser extent, lamin B1 increased donut-shaped nucleieven without FTI treatment. (K) Localization of GFP-labeled WT lamin B1 (Upper) or LaB1-SAIM (Lower) in late mitosis showing that nonfarnesylated lamin B1is mislocalized to the nucleoplasm (more detail is provided in Fig. S6). (Scale bar: 10 μm.) *P < 0.05; **P < 0.005; ***P < 0.001.
4 of 6 | www.pnas.org/cgi/doi/10.1073/pnas.1019532108 Verstraeten et al.

antiproliferative effects of FTIs. Alternatively, spindle defectscould arise from FTI-mediated loss of farnesyltransferase bindingto tubulin (30). Importantly, cells with donut-shaped nuclei did notexhibit apoptosis or senescence and continued to divide, but witha large fraction dividing abnormally and becoming aneuploid.We identified lower pericentrin levels and lamin B1 as critical
for FTI-induced donut formation and binucleation. The precisemechanism by which FTIs reduce pericentrin levels remains tobe identified, opening an attractive avenue for future investi-gation. An earlier study (31) ascribed proteasomal inhibitoryactivity to FTIs in a breast cancer cell line; however, here, wereport data suggesting that FTIs increase proteasomal degrada-tion of pericentrin, in keeping with earlier reports of increasedprotein degradation in FTI-treated cells (8, 32). Blocking pro-teasome-mediated degradation prevented the FTI-induced lossof pericentrin and prevented the formation of donut-shaped cellnuclei (Fig. 4G).Following an ongoing clinical trial in Europe that uses a statin
and an aminobisphosphonate, which target different steps of pre-nylation (8, 11, 33), a triple-drug regimen of anFTI, a statin, and anaminobisphosphonate is currently being tested in theUnited Statesas a treatment for progeria. For that reason, we examined theeffects of lonafarnib, pravastatin, and zoledronate on donut for-mation in human skin fibroblasts. We found that the triple-drugregimen induced more donut-shaped nuclei than the FTI alone.Interestingly, low levels of lovastatin [10 nM, similar to plasmalevels achieved in patients being treated for hypercholesterolemia(34)] had no perceptible effect on protein farnesylation and did notyield donut-shaped nuclei. In any case, we believe that physicianstreating patients who have progeria with an FTI, or the triple-drugcombination regimen, should be cognizant of the effects of FTIs on
nuclear morphology and pericentrin levels, particularly becauseloss of pericentrin has been reported to cause dwarfism (13).
Materials and MethodsMice and FTI Treatment. WT mice and mice expressing human progerin froma BAC transgene received a 5-g ball of dough each morning containing 2.25mg of tipifarnib (R115777; Zarnestra) for up to 12 mo (6). All animal usecomplied with the Animal Care and Use Committee guidelines under Pro-tocol G-03-05 (National Institutes of Health).
Cell Culture, FTI, Lovastatin, and Monastrol Treatment. For FTI treatment, cellswere treated daily with 10 μM FTI L744832 (Biomol) or an equal volume ofvehicle (DMSO; 0.025%final concentration) for at least 48 h in the case ofMEFsand for 72 h for other cell lines (35). For lovastatin treatment, cells were treateddaily with 10 nM and 1, 4, 10, or 20 μM lovastatin (Calbiochem) or equal vol-umes of vehicle (DMSO) for 72 h. Cells were pretreated with 10 μg/mL cyclo-heximide (Sigma) for 3 h and thereafter treated daily with 10 μM FTI and 10μg/mL cycloheximide for 3 d to determine whether only newly synthesizedproteins are affected by FTI treatment. Skin fibroblasts were treated for 20 hwith 100 μMmonastrol (M8515; Sigma) or vehicle (DMSO),fixed, and evaluatedfor donut-shaped nuclei. Primary skin fibroblasts were cotreated daily with10 μM lactacystin (Cayman Chemical), which is a proteasome inhibitor, and10 μM FTI or vehicle (DMSO) alone for a period of 3 d to evaluate if FTI-mediatedprotein degradationunderlies the formation ofdonut-shapednuclei.
Time-Lapse Video Microscopy. HEp2 cells stably expressing GFP–histone-3(HEp2-GFP–H3) were transfected with a mCherry–β-tubulin plasmid, andSV40-transformed human skin fibroblasts stably expressing GFP–lamin Awere imaged on a temperature-controlled dish holder on a motorized stageof an Olympus IX-70 microscope or confocal Zeiss LSM-710 microscope.
Immunofluorescence Microscopy. Cells were fixed, processed, and incubatedwith primary antibodies against lamin A/C, CENP-E, CENP-F, vimentin, peri-centrin, prelamin A, β-tubulin, macroH2A (generously provided by P. Adams,
Fig. 5. FTI treatment results in donut-shaped nuclei and binucleated cells that divide abnormally. FTI-treated primary human skin fibroblasts showing donut-shaped nuclei in binucleated cells in phase-contrast images (A) and in single confocal sections after staining for lamin A (B1), vimentin (B2), and DNA (B3,Hoechst). (B4) Merged image. (Scale bar: 10 μm.) (C) Distribution of donut-shaped nuclei between mononucleated and binucleated cells. (D) FTI treatmentcaused increased numbers of binucleated cells in two primary human skin fibroblasts (Fibro) lines and in HEp2 cells. (E) Large fraction of binucleated cellscontained donut-shaped nuclei. (F) Photoconversion of Dendra2–histone-4 from green to red to track individual nuclei over time. (G) Follow-up study of cellfate for 4 d in FTI-treated skin fibroblasts after photoconversion of mononucleated and binucleated cells with and without donut-shaped nuclei showinga lower probability of cell division in binucleated cells compared with mononucleated cells, regardless of the presence of donut-shaped nuclei. (H and I) Time-lapse video microscopy studies of HEp2 cells showing that ∼50% of cells with donut-shaped nuclei divide abnormally, often yielding new donut-shaped nuclei(I, Upper; 55%) or an abnormal number of daughter nuclei (I, Lower; 21%). Arrows indicate daughter nuclei. (Scale bar: 10 μm.) (J) Single confocal sectionsshowing defective karyokinesis in an FTI-treated skin fibroblast after labeling for lamin A (J1), vimentin (J2), and DNA (J3, Hoechst). (Scale bar: 25 μm.)(K) Inactive X-chromosomes in female FTI-treated human skin fibroblasts labeled for the histone macroH2A that preferentially binds to the inactive X-chromosome (arrows). (Scale bar: 10 μm.) (L) Quantification of primary female human skin fibroblasts containing more than one inactive X-chromosomein response to FTI treatment, revealing higher levels of aneuploidy in cells with donut-shaped nuclei. Costaining of macroH2A with Ki67 was used to identifynoncycling cells. *P < 0.05; ***P < 0.001.
Verstraeten et al. PNAS Early Edition | 5 of 6
MED
ICALSC
IENCE
S

University of Edinburgh, Glasgow, United Kingdom), or Mitotracker (Invi-trogen). They were then counterstained with Hoechst 33342.
Immunohistochemistry. Tissue sections of gut and skin derived from FTI-treated and nontreated WT mice and LMNA G608G mice were processed forH&E staining or immunostaining with a lamin A/C antibody.
Western Blot Analyses. Western blot analyses are discussed in SI Materialsand Methods.
RNAi Against FNTB, Pericentrin, and Lamin B1. HeLa cells and SV40-transformed human skin fibroblasts were transfected with shRNA plasmids toreduce expression of FNTB, pericentrin, and lamin B1. The knockdown ofeach gene was verified by Western blot analysis and real-time PCR assay.
Constructs specifying nonfarnesylated versions of lamin B1, CENP-E, andCENP-F are discussed in SI Materials and Methods.
Photoconversion of Histone 4–Dendra2. Primary skin fibroblasts were elec-troporatedwith aplasmid forDendra2-taggedhistone-4, plated, treateddailywith 10 μM FTI for 3 d, and then photoconverted from green to red as de-scribed in SI Materials andMethods. Data were acquired in four independentexperiments, and at least 60 cells were evaluated for each category.
Statistical Analysis. Experiments were performed at least three independenttimes. Images were blinded to the person performing the analysis andrandomized before the evaluation. Statistical analyses were performed withPRISM 3.0 and INSTAT software (GraphPad) by an unpaired Student’s t testwith Welch’s correction to allow for different variances. To compare donutformation (a binomial event) between different conditions, results wereanalyzed by a χ2 distribution and Fisher’s exact test. For these results, datawere expressed as the sample proportion ± SE. All other data were ex-
pressed as mean ± SEM. For all experiments, a two-tailed P value ≤0.05 wasconsidered significant.
Image Acquisition and Manipulation. Phase-contrast and fluorescence imageswere acquired with a CoolSNAP HQ digital CCD-camera (Photometrics, RoperScientific) mounted on an Olympus IX70 inverted microscope and controlledby IPLab 4.0 (BD Biosciences) image acquisition software. Experiments tocount the fraction of donuts, as well as time-lapse video microscopyexperiments, were imaged with an Olympus LCPlanF 20× phase-contrastobjective (0.40 N.A.). Immunofluorescence samples were imaged with anOlympus UApo/340 40× water immersion objective (1.15 N.A.). For analysisof Western blots, X-ray films were digitized on an Epson Perfection 2450scanner with linear intensity settings. Digital images were processed withAdobe Photoshop (version 6.0; Adobe Systems, Inc.) by adjusting the linearimage intensity display range, and fluorescence gray-scale images werecolorized in ImageJ (National Institutes of Health), Adobe Photoshop, orIPLab (BD Biosciences) 4.0 by selecting a color plane appropriate for the-chromophore.
A more detailed description of experimental protocols can be found in SIMaterials and Methods.
ACKNOWLEDGMENTS. We thank the following investigators for provid-ing reagents: Dr. A. Ishov, Dr. D. Bader, Dr. C. Stewart, Dr. F. Gertler,Dr. T. Glover, Dr. P. Adams, Dr. R. Goldman, Dr. T. Yen, Dr. D. Cleveland,Dr. K. Roux, and Dr. I. Raska. We thank the following investigators forhelpful discussions: Dr. K. Roux and Dr. R. Prince. This work was supportedby National Institutes of Health Grants HL082792, NS059348, AG035626,HL086683, and HL089781; the Ellison Medical Foundation Senior ScholarProgram; the Progeria Research Foundation; and fellowships from theNetherlands Genomics Initiative 2007/01129/MW (to V.L.R.M.V.) and theAmerican Heart Association 09POST2080264 (to V.L.R.M.V.).
1. Sousa SF, Fernandes PA, Ramos MJ (2008) Farnesyltransferase inhibitors: A detailedchemical view on an elusive biological problem. Curr Med Chem 15:1478–1492.
2. Cox AD, Der CJ (2002) Ras family signaling: Therapeutic targeting. Cancer Biol Ther 1:599–606.
3. Downward J (2003) Targeting RAS signalling pathways in cancer therapy. Nat RevCancer 3:11–22.
4. Fong LG, et al. (2006) A protein farnesyltransferase inhibitor ameliorates disease ina mouse model of progeria. Science 311:1621–1623.
5. Yang SH, et al. (2006) A farnesyltransferase inhibitor improves disease phenotypes inmice with a Hutchinson-Gilford progeria syndrome mutation. J Clin Invest 116:2115–2121.
6. Capell BC, et al. (2008) A farnesyltransferase inhibitor prevents both the onset andlate progression of cardiovascular disease in a progeria mouse model. Proc Natl AcadSci USA 105:15902–15907.
7. GlynnMW, Glover TW (2005) Incomplete processing of mutant lamin A in Hutchinson-Gilford progeria leads to nuclear abnormalities, which are reversed byfarnesyltransferase inhibition. Hum Mol Genet 14:2959–2969.
8. Toth JI, et al. (2005) Blocking protein farnesyltransferase improves nuclear shape infibroblasts from humans with progeroid syndromes. Proc Natl Acad Sci USA 102:12873–12878.
9. Verstraeten VL, et al. (2006) Compound heterozygosity for mutations in LMNAcauses a progeria syndrome without prelamin A accumulation. Hum Mol Genet 15:2509–2522.
10. Sinensky M, Beck LA, Leonard S, Evans R (1990) Differential inhibitory effects oflovastatin on protein isoprenylation and sterol synthesis. J Biol Chem 265:19937–19941.
11. Varela I, et al. (2008) Combined treatment with statins and aminobisphosphonatesextends longevity in a mouse model of human premature aging. Nat Med 14:767–772.
12. Mayer TU, et al. (1999) Small molecule inhibitor of mitotic spindle bipolarityidentified in a phenotype-based screen. Science 286:971–974.
13. Rauch A, et al. (2008) Mutations in the pericentrin (PCNT) gene cause primordialdwarfism. Science 319:816–819.
14. Doxsey SJ, Stein P, Evans L, Calarco PD, Kirschner M (1994) Pericentrin, a highlyconserved centrosome protein involved in microtubule organization. Cell 76:639–650.
15. Putkey FR, et al. (2002) Unstable kinetochore-microtubule capture and chromosomalinstability following deletion of CENP-E. Dev Cell 3:351–365.
16. Schafer-Hales K, et al. (2007) Farnesyl transferase inhibitors impair chromosomalmaintenance in cell lines and human tumors by compromising CENP-E and CENP-Ffunction. Mol Cancer Ther 6:1317–1328.
17. Moynihan KL, Pooley R, Miller PM, Kaverina I, Bader DM (2009) Murine CENP-Fregulates centrosomal microtubule nucleation and interacts with Hook2 at thecentrosome. Mol Biol Cell 20:4790–4803.
18. Hussein D, Taylor SS (2002) Farnesylation of Cenp-F is required for G2/M progressionand degradation after mitosis. J Cell Sci 115:3403–3414.
19. Ashar HR, et al. (2000) Farnesyl transferase inhibitors block the farnesylation of CENP-E and CENP-F and alter the association of CENP-E with the microtubules. J Biol Chem275:30451–30457.
20. Goodman B, et al. (2010) Lamin B counteracts the kinesin Eg5 to restrain spindle poleseparation during spindle assembly. J Biol Chem 285:35238–35244.
21. Tsai MY, et al. (2006) A mitotic lamin B matrix induced by RanGTP required for spindleassembly. Science 311:1887–1893.
22. Fong LG, et al. (2006) Prelamin A and lamin A appear to be dispensable in the nuclearlamina. J Clin Invest 116:743–752.
23. Davies BS, et al. (2010) An accumulation of non-farnesylated prelamin A causescardiomyopathy but not progeria. Hum Mol Genet 19:2682–2694.
24. Sebti SM, Der CJ (2003) Opinion: Searching for the elusive targets offarnesyltransferase inhibitors. Nat Rev Cancer 3:945–951.
25. Hougland JL, et al. (2010) Identification of novel peptide substrates for proteinfarnesyltransferase reveals two substrate classes with distinct sequence selectivities. JMol Biol 395:176–190.
26. Maurer-Stroh S, et al. (2007) Towards complete sets of farnesylated andgeranylgeranylated proteins. PLOS Comput Biol 3:e66.
27. Crespo NC, et al. (2002) The farnesyltransferase inhibitor, FTI-2153, inhibits bipolarspindle formation during mitosis independently of transformation and Ras and p53mutation status. Cell Death Differ 9:702–709.
28. Crespo NC, Ohkanda J, Yen TJ, Hamilton AD, Sebti SM (2001) The farnesyltransferaseinhibitor, FTI-2153, blocks bipolar spindle formation and chromosome alignment andcauses prometaphase accumulation during mitosis of human lung cancer cells. J BiolChem 276:16161–16167.
29. Kim J, et al. (2008) Centrosomal PKCbetaII and pericentrin are critical for humanprostate cancer growth and angiogenesis. Cancer Res 68:6831–6839.
30. Zhou J, et al. (2009) The protein farnesyltransferase regulates HDAC6 activity ina microtubule-dependent manner. J Biol Chem 284:9648–9655.
31. Efuet ET, Keyomarsi K (2006) Farnesyl and geranylgeranyl transferase inhibitorsinduce G1 arrest by targeting the proteasome. Cancer Res 66:1040–1051.
32. Yang SH, Andres DA, Spielmann HP, Young SG, Fong LG (2008) Progerin elicits diseasephenotypes of progeria in mice whether or not it is farnesylated. J Clin Invest 118:3291–3300.
33. Meta M, Yang SH, Bergo MO, Fong LG, Young SG (2006) Protein farnesyltransferaseinhibitors and progeria. Trends Mol Med 12:480–487.
34. Lamson M, et al. (2002) Pharmacokinetics of lovastatin extended-release dosage form(Lovastatin XL) in healthy volunteers. Biopharm Drug Dispos 23:143–149.
35. Verstraeten VL, Ji JY, Cummings KS, Lee RT, Lammerding J (2008) Increasedmechanosensitivity and nuclear stiffness in Hutchinson-Gilford progeria cells: Effectsof farnesyltransferase inhibitors. Aging Cell 7:383–393.
6 of 6 | www.pnas.org/cgi/doi/10.1073/pnas.1019532108 Verstraeten et al.




















