
Protective Effect of Supplemental Low Intensity White Light on Ultraviolet-B Exposure-Induced...
9
Regular paper Protective effect of supplemental low intensity white light on ultraviolet-B exposure-induced impairment in cyanobacterium Spirulina platensis: formation of air vacuoles as a possible protective measure Subramanyam Rajagopal 1,2, *, Cosmin Sicora 1 , Zsuzsanna Va´rkonyi 1 , La´szlo´ Musta´rdy 1 & Prasanna Mohanty 3 1 Institute of Plant Biology, Biological Research Center, 521, 6701 Szeged, Hungary; 2 School of Life Sciences, and Center for the Study of Early Events in Photosynthesis, Arizona State University, Tempe, AZ 85287- 4501, USA; 3 School of Life Sciences, Jawaharlal Nehru University, New Delhi, 110 067, India; *Author for correspondence (e-mail: [email protected]; phone: +1-480-965-8725; fax: +1-480-965-6899) Received 14 October 2004; accepted in revised form 14 February 2005 Key words: Chl a antennas, photosynthetic active radiation, phycobilisomes, ultrastructure, ultraviolet-B, vacuoles Abstract Intact trichomes of Spirulina platensis were exposed to 1–5 h of low (0.2 mW cm )2 ) or high (0.6 mW cm )2 ) intensity UV-B (280–320 nm) radiation, alone or with photosynthetically active radiation (PAR) of sup- plemental 50 lEm )2 s )1 white light (WL). The mitigating effect of supplemental WL on UV-B induced alterations in Spirulina were investigated by monitoring time-dependent change in photosystem (PS) II mediated O 2 evolution, absorption, circular dichroism (CD) spectra, and ultrastructure. At low intensity, UV-B induced loss in PS II-catalyzed O 2 evolution, but caused no change in the absorption spectrum. At high intensity, UV-B caused a decrease in absorption by phycobilisomes (PBsomes), which was only partly prevented by the presence of low-intensity supplemental WL. The CD spectral analysis revealed that UV-B exposure caused time-dependent enhancement of the negative psi-type bands at 452 and 689 nm, reflecting alterations in the macroaggregation of chlorophyll–protein complexes. This enhancement of negative PS II- type bands was substantially arrested by the presence of supplemental WL exposure, even when UV-B exposure was continued for 5 h. These changes in UV-B-induced CD spectrum suggest alterations in the antenna structure of Spirulina involving both PBsomes and Chlorophyll a. Thus, supplemental low intensity WL arrests, to large extent, the macroaggregation of pigment–protein complexes. Furthermore, the electron micrographs of Spirulina revealed that UV-B exposure caused disorganization of the cellular ultrastructure, while the inclusion of supplemental WL enhanced the formation of air vacuoles in Spirulina. We suggest that the formation of vacuoles by supplemental WL is a protective feature against UV-B. Introduction Cyanobacteria, the oxygenic photosynthetic prokaryotes, are exposed to various types of environmental stresses. In recent years, due to enhanced levels of ultraviolet (UV)-B in the solar spectrum, studies on the effects of UV-B radiation on cyanobacteria have gained prominence because of the prospects of understanding UV-B damage and repair mechanisms in these organisms (Sinha et al. 1995; Lao and Glazer 1996; Araoz and Hader 1997; Campbell et al. 1998; Mate et al. Photosynthesis Research (2005) 85: 181–189 Ó Springer 2005
Transcript of Protective Effect of Supplemental Low Intensity White Light on Ultraviolet-B Exposure-Induced...

Regular paper
Protective effect of supplemental low intensity white light on ultraviolet-Bexposure-induced impairment in cyanobacterium Spirulina platensis:formation of air vacuoles as a possible protective measure
Subramanyam Rajagopal1,2,*, Cosmin Sicora1, Zsuzsanna Varkonyi1, Laszlo Mustardy1
& Prasanna Mohanty31Institute of Plant Biology, Biological Research Center, 521, 6701 Szeged, Hungary; 2School of Life Sciences,and Center for the Study of Early Events in Photosynthesis, Arizona State University, Tempe, AZ 85287-4501, USA; 3School of Life Sciences, Jawaharlal Nehru University, New Delhi, 110 067, India; *Author forcorrespondence (e-mail: [email protected]; phone: +1-480-965-8725; fax: +1-480-965-6899)
Received 14 October 2004; accepted in revised form 14 February 2005
Key words: Chl a antennas, photosynthetic active radiation, phycobilisomes, ultrastructure, ultraviolet-B,vacuoles
Abstract
Intact trichomes of Spirulina platensis were exposed to 1–5 h of low (0.2 mW cm)2) or high (0.6 mW cm)2)intensity UV-B (280–320 nm) radiation, alone or with photosynthetically active radiation (PAR) of sup-plemental 50 lE m)2 s)1 white light (WL). The mitigating effect of supplemental WL on UV-B inducedalterations in Spirulina were investigated by monitoring time-dependent change in photosystem (PS) IImediated O2 evolution, absorption, circular dichroism (CD) spectra, and ultrastructure. At low intensity,UV-B induced loss in PS II-catalyzed O2 evolution, but caused no change in the absorption spectrum. Athigh intensity, UV-B caused a decrease in absorption by phycobilisomes (PBsomes), which was only partlyprevented by the presence of low-intensity supplemental WL. The CD spectral analysis revealed that UV-Bexposure caused time-dependent enhancement of the negative psi-type bands at 452 and 689 nm, reflectingalterations in the macroaggregation of chlorophyll–protein complexes. This enhancement of negative PS II-type bands was substantially arrested by the presence of supplemental WL exposure, even when UV-Bexposure was continued for 5 h. These changes in UV-B-induced CD spectrum suggest alterations in theantenna structure of Spirulina involving both PBsomes and Chlorophyll a. Thus, supplemental lowintensity WL arrests, to large extent, the macroaggregation of pigment–protein complexes. Furthermore,the electron micrographs of Spirulina revealed that UV-B exposure caused disorganization of the cellularultrastructure, while the inclusion of supplemental WL enhanced the formation of air vacuoles in Spirulina.We suggest that the formation of vacuoles by supplemental WL is a protective feature against UV-B.
Introduction
Cyanobacteria, the oxygenic photosyntheticprokaryotes, are exposed to various types ofenvironmental stresses. In recent years, due toenhanced levels of ultraviolet (UV)-B in the solarspectrum, studies on the effects of UV-B radiation
on cyanobacteria have gained prominence becauseof the prospects of understanding UV-B damageand repair mechanisms in these organisms (Sinhaet al. 1995; Lao and Glazer 1996; Araoz andHader 1997; Campbell et al. 1998; Mate et al.
Photosynthesis Research (2005) 85: 181–189 � Springer 2005

1998; Sah et al. 1998; Rajagopal et al. 1998, 1999,2000; Vass et al. 2000; Tichy et al. 2003). Of thetotal solar energy ranging from 280 to 700 nm,absorbed by cyanobacterial cells, almost half isabsorbed by the phycobilisomes (PBsomes), whichare the major light-harvesting supramolecularpigment–protein complexes in cyanobacteria, andthe remaining by the chlorophyll (Chl)–proteincomplexes of the cells (Lao and Glazer 1996).
UV-B (280–320 nm) radiation is reaching theearth’s surface due to depletion of the stratosphericozone layer by anthrogenic factors (Frederic 1993;Murthy and Rajagopal 1995; Vass 1997). It pre-dominantly influences the primary photosyntheticprocess by causing damage to the photosystem(PS)II reaction centers (RCs) (Murthy and Rajag-opal 1995; Vass 1997). It has been shown that UV-B irradiation impairs growth (Strid et al. 1994),suppresses Chl biosynthesis (Murthy and Rajag-opal 1995), and inhibits electron transport (Ra-jagopal and Murthy 1996a; Kolli et al. 1998) andnet photosynthesis (Van et al. 1976; Brandle et al.1977). Of the two photosystems, PS II is known tobe more sensitive than PSI to UV-B irradiation(Melis et al. 1992). Degradation of the PS II RCproteins upon UV-B illumination has been char-acterized by many workers (Renger et al. 1989;Melis et al. 1992; Friso et al. 1994a, b; Mate et al.1998; Vass et al. 2000). UV-B-induced loss of wateroxidation capacity of PS II occurs largely due toloss of the D1 and D2 proteins of the PS II RC(Friso et al. 1994a, b). The vulnerability of the D1andD2 proteins to UV-B has been demonstrated inthe cyanobacterium Synechococcus (Mate et al.1998). The quinones of the photosynthetic electrontransport chain, which absorb UV-B, have beenreported to be the targets of UV-B radiation (Vass1997; Kolli et al. 1998).
Synechococcus and Spirulina platensis exposedto UV-B showed inhibited O2 evolution andaltered spectral characteristics of PBsomes due topartial uncoupling of energy transfer fromPBsomes to Chl a (Rajagopal and Murthy 1996a,b; Kolli et al.1998). Exposure of SynechococcusPBsomes to high intensity of UV-B has beenshown to bring about gradual but unspecificdamages to the linker and anchor polypeptides(Pandey et al. 1997). Quantum yield of photo-damage to PBsomes caused by UV-B is muchhigher than that caused by visible light in Anaba-ena sp. (Lao and Glazer 1996). UV-B also induces
structural and functional photodamage to thePBsomes (Sinha et al. 1995; Lao and Glazer 1996;Araoz and Hader 1997; Lorenz et al. 1997; Pandeyet al. 1997; Sah et al. 1998; Rajagopal et al. 1998,1999; Jha et al. 2000). Earlier studies indicated thatexposure of Synechococcus to a high intensity ofUV-B for short duration mainly causes the disso-ciation of PBsomes (Rajagopal and Murthy 1996b;Sah et al. 1998). Earlier, we reported that exposureof Spirulina platensis trichomes to moderate-intensity UV-B (1.9 mW m)2) for 9 h caused theloss of the anchor polypeptide (85.5 kDa)(Rajagopal et al. 1998) and Chl antenna complexes(Rajagopal et al. 2000). These effects wereobserved in response to both in vitro and in vivoexposure to UV-B (Rajagopal et al. 1998, 2000).
Since plants receive white light (WL) along withenhanced levels of UV-B radiation under naturalconditions, the WL retards UV-B injury. There areonly a few reports on the protection of PS IIagainst UV-B-induced damage when given incombination of photosynthetical active radiation(PAR) light in cyanobacteria (Vass et al. 1999;Ivanov et al. 2000; Sicora et al. 2003). Earlier, wereported that the exposure of isolated Spirulinaplatensis PBsomes to UV-B along with PAR pro-vided protection against UV-B-induced alterationsin energy transfer within the PBsomes (Rajagopalet al. 1999). Exposure of intact cells of Synecho-coccus to high-intensity UV-B along with WL wasshown to cause unspecific damage to cell ultra-structure (Chauhan et al. 1998). Although theeffects of UV-B on the cyanobacterial photosyn-thetic apparatus have been studied by many labs,there are only limited reports on the effects of UV-B administered in combination with PAR. Spiru-lina platensis are normally grown in open ponds,and these trichomes receive UV-B along with PAR;no studies on the effect of supplemental WL onUV-B damage have been carried out in Spirulina.
In the present study, we have used low and highintensities of UV-B (0.2 and 0.6 mW cm)2), incombination with supplemental low-intensity WL(50 lE m)2), to monitor the ultrastructure andspectral characteristics in Spirulina platensis, acommercially important filamentous cyanobacte-rium. Our results demonstrate that (a) intensity-dependent alterations in the photosynthetic appa-ratus occur upon exposure of cells to UV-B, (b)supplemental WL provides protection againstUV-B-induced damage in PBsomes and Chl a
182

antenna and, c) WL causes the formation of gasvacuoles, which may lower the UV-B dose receivedby the cells.
Materials and methods
Spirulina platensis cells were grown in Zarrouk’smedium at 26 ± 2 �C in continuous WL of50 lE m)2 s)1 to early-exponential phase (A730 nm
of 0.6–0.8; 5 lg Chl/ml), as earlier reported byRajagopal et al. (1998). The culture medium wasgently bubbled with air containing 5% CO2. Thecells were exposed to two different UV-B intensi-ties (0.2 or 0.6 mW cm)2) and UV-B along withsupplemental WL (PAR, 50 lE m)2 s)1) for dif-ferent intervals of time (1–5 h) at 26 ± 2 �C, andcontrol cells were kept in WL as in growth con-ditions with continuous stirring. UV-B irradiationwas performed in open, flat glass containers whichheld 300 ml of cell suspension (5 lg of Chl/ml) at adepth of 3 cm. The UV-B radiation was adminis-tered to cells by means of Fotodyne Inc. (USA),UV-B tubes (catalogue no. 3–4408) which havemaximum emission at 312 nm with 40 nm half-band-width. The UV-B tubes were used in com-bination with 4 layers of 0.1 mm cellulose acetatefilters (Clarfoil, Courtaulds Chemicals) to screenout any UV-C contamination.
Absorption spectra were recorded by a Schi-madzu-2000 double beam spectrophotometer onequal Chl basis 5 lg/ml).
Circular dichroism (CD) spectra were recordedusing a Jobin Yvon CD6 dichrograph. The Chlcontent of the samples was adjusted to 5 lg/ml.The optical path length of the cell in a sampleholder was 1 cm, and the distance of the samplefrom the photomultiplier was 5 cm. The spectrawere recorded in 1 nm steps with an integrationtime of 0.3 s and a band-pass of 2 nm (Simidjievet al. 1997). Spectral signals were averaged andthe baseline from the buffer was subtracted. CDwas measured in absorbance units, but data wereplotted in relative units for easier comparison.Each experiment was repeated on at least threeindependent batches, with identical tendencies andsimilar results.
PSII activity was measured polarographicallyby monitoring the amount of O2 evolved, withH2O as an electron donor and p-benzoquinone(pBQ, 1 mM) as an electron acceptor (Kolli et al.
1998). The measurements were made in a Clark-type O2 electrode from Hansatech, UK (modelDW2). The Chl content was adjusted to 10 lg/ml.Chl was estimated according to MacKinney(1941).
For the transmission electron microscopy(TEM) studies, the Spirulina cells were exposed toboth intensities of UV-B alone and UV-B alongwith supplemental WL for different time periods(1–5 h), and after exposure, the Spirulina cells werecollected by centrifugation at 6000 · g for 10 min.The pre-fixation was carried out in 2% glutaral-dehyde solution made in 100 mM phosphate buf-fer, pH 7.0 at 4 �C. Following pre-fixation, thecells were washed 2–3 times in phosphate buffer,and post-fixed in 2% osmium tetraoxide, for 1 h inice bath in a fume hood. The cells were washed 5–6times with phosphate buffer and dehydrated ingraded series of ethanol solutions (30, 50, 70, 90%,and 3 changes of 100% for 10 min each). Theywere then transferred to propylenoxide 3 timeseach for 10 min. The cells were then embedded inaraldite and thin sections (0.1 lm) were cut in aReichert OMU 2 ultramicrotome and mounted oncopper grids. The sections were stained withuranyl acetate (saturated solution in 60% ethanol)and lead citrate, and examined under a TEM(Tesla BS540) (Simidjiev et al. 1997). Theoriginal magnification of the negatives was12,000·.
Light intensity was measured with a Li-Corradiometer (model LI-189, LiCor, USA). UV-Birradiance was measured with the help of a VLX-312 UV meter (Vilber-Loumat, France). The UVmeter was protected with a UV-B interference fil-ter with 100% transmittance at 312 nm and a halfbandwidth of 5 nm at 50% transmittance, asdescribed elsewhere (Sah et al. 1998). Thus, con-taminating radiation in the UV-A and UV-Cspectral ranges was negligible.
Results and discussion
Trial experiments were carried out to determine asuitable level of UV-B radiation in the presence ofPAR, such that the cyanobacterial culture couldrecover and grow upon transfer to normal growthconditions. From our previous experiments, wenoted that a moderate intensity of UV-B(1.9 mW m)2) plus supplemental WL-exposed
183
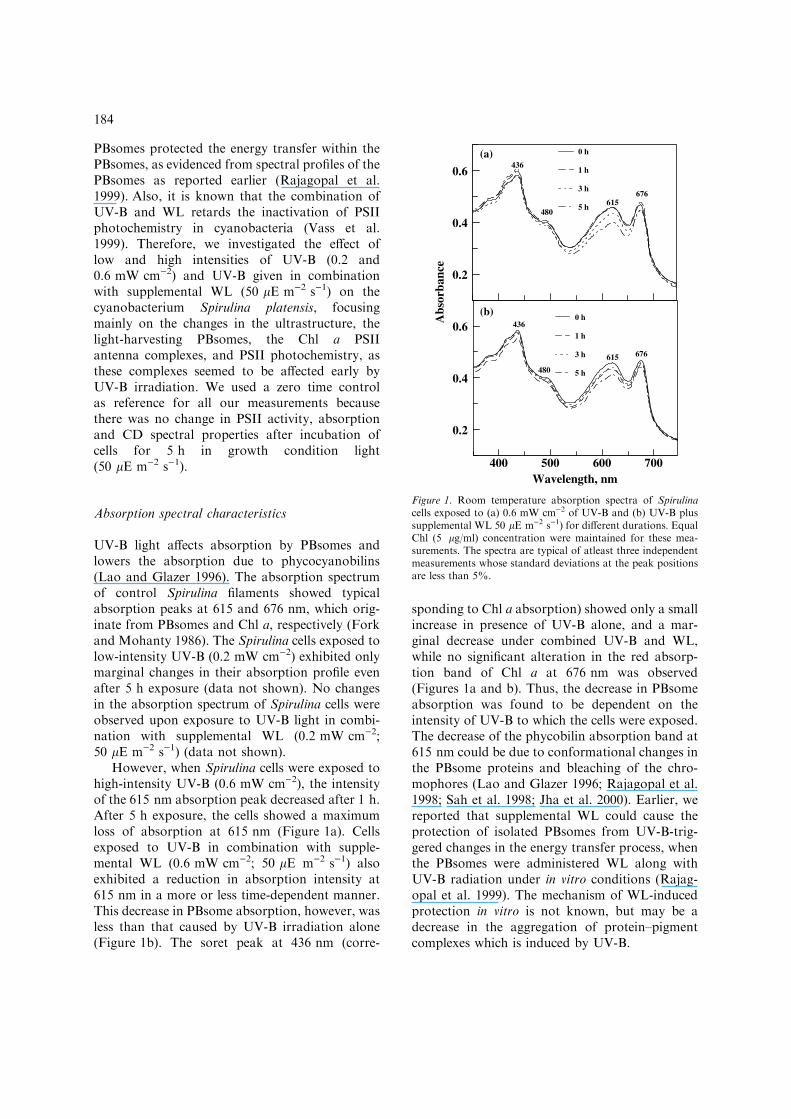
PBsomes protected the energy transfer within thePBsomes, as evidenced from spectral profiles of thePBsomes as reported earlier (Rajagopal et al.1999). Also, it is known that the combination ofUV-B and WL retards the inactivation of PSIIphotochemistry in cyanobacteria (Vass et al.1999). Therefore, we investigated the effect oflow and high intensities of UV-B (0.2 and0.6 mW cm)2) and UV-B given in combinationwith supplemental WL (50 lE m)2 s)1) on thecyanobacterium Spirulina platensis, focusingmainly on the changes in the ultrastructure, thelight-harvesting PBsomes, the Chl a PSIIantenna complexes, and PSII photochemistry, asthese complexes seemed to be affected early byUV-B irradiation. We used a zero time controlas reference for all our measurements becausethere was no change in PSII activity, absorptionand CD spectral properties after incubation ofcells for 5 h in growth condition light(50 lE m)2 s)1).
Absorption spectral characteristics
UV-B light affects absorption by PBsomes andlowers the absorption due to phycocyanobilins(Lao and Glazer 1996). The absorption spectrumof control Spirulina filaments showed typicalabsorption peaks at 615 and 676 nm, which orig-inate from PBsomes and Chl a, respectively (Forkand Mohanty 1986). The Spirulina cells exposed tolow-intensity UV-B (0.2 mW cm)2) exhibited onlymarginal changes in their absorption profile evenafter 5 h exposure (data not shown). No changesin the absorption spectrum of Spirulina cells wereobserved upon exposure to UV-B light in combi-nation with supplemental WL (0.2 mW cm)2;50 lE m)2 s)1) (data not shown).
However, when Spirulina cells were exposed tohigh-intensity UV-B (0.6 mW cm)2), the intensityof the 615 nm absorption peak decreased after 1 h.After 5 h exposure, the cells showed a maximumloss of absorption at 615 nm (Figure 1a). Cellsexposed to UV-B in combination with supple-mental WL (0.6 mW cm)2; 50 lE m)2 s)1) alsoexhibited a reduction in absorption intensity at615 nm in a more or less time-dependent manner.This decrease in PBsome absorption, however, wasless than that caused by UV-B irradiation alone(Figure 1b). The soret peak at 436 nm (corre-
sponding to Chl a absorption) showed only a smallincrease in presence of UV-B alone, and a mar-ginal decrease under combined UV-B and WL,while no significant alteration in the red absorp-tion band of Chl a at 676 nm was observed(Figures 1a and b). Thus, the decrease in PBsomeabsorption was found to be dependent on theintensity of UV-B to which the cells were exposed.The decrease of the phycobilin absorption band at615 nm could be due to conformational changes inthe PBsome proteins and bleaching of the chro-mophores (Lao and Glazer 1996; Rajagopal et al.1998; Sah et al. 1998; Jha et al. 2000). Earlier, wereported that supplemental WL could cause theprotection of isolated PBsomes from UV-B-trig-gered changes in the energy transfer process, whenthe PBsomes were administered WL along withUV-B radiation under in vitro conditions (Rajag-opal et al. 1999). The mechanism of WL-inducedprotection in vitro is not known, but may be adecrease in the aggregation of protein–pigmentcomplexes which is induced by UV-B.
0.6
0.4
0.2
0 h
1 h
3 h
5 h
700600500400
0.6
0.4
0.2
Wavelength, nm
0 h
1 h
3 h
5 h
615 676
676615
436
480
480
436Abs
orba
nce
(a)
(b)
Figure 1. Room temperature absorption spectra of Spirulinacells exposed to (a) 0.6 mW cm)2 of UV-B and (b) UV-B plussupplemental WL 50 lE m)2 s)1) for different durations. EqualChl (5 lg/ml) concentration were maintained for these mea-surements. The spectra are typical of atleast three independentmeasurements whose standard deviations at the peak positionsare less than 5%.
184
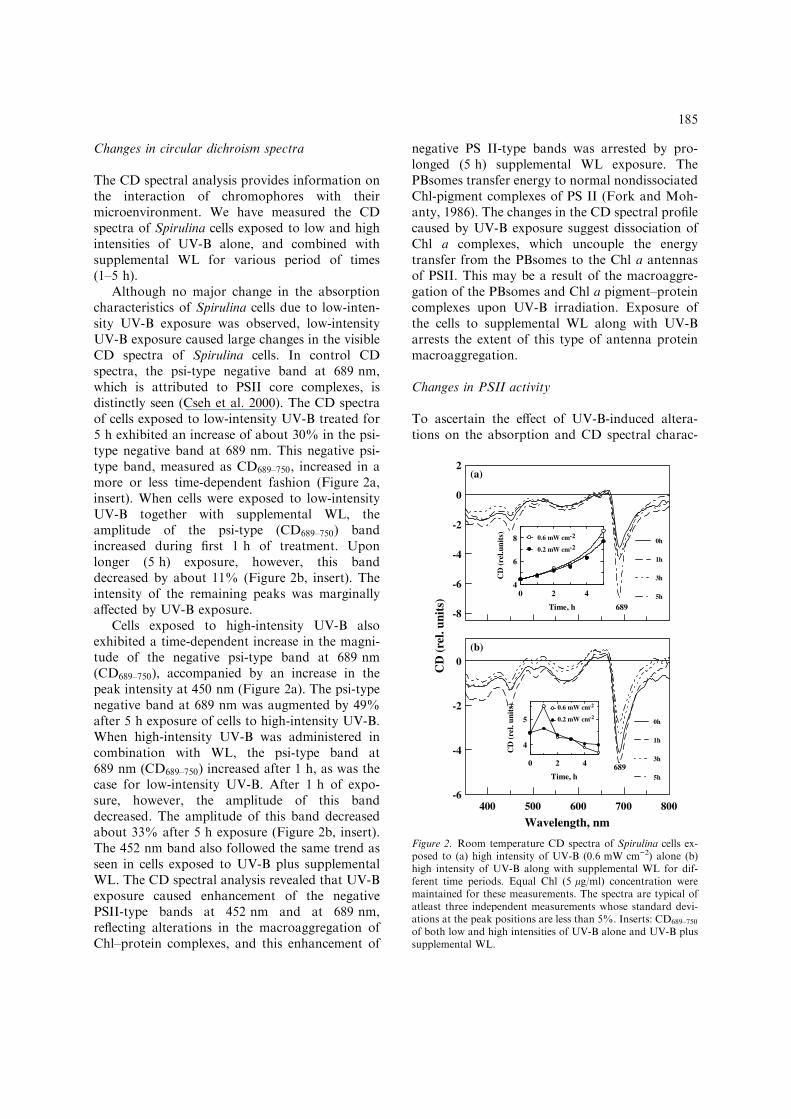
Changes in circular dichroism spectra
The CD spectral analysis provides information onthe interaction of chromophores with theirmicroenvironment. We have measured the CDspectra of Spirulina cells exposed to low and highintensities of UV-B alone, and combined withsupplemental WL for various period of times(1–5 h).
Although no major change in the absorptioncharacteristics of Spirulina cells due to low-inten-sity UV-B exposure was observed, low-intensityUV-B exposure caused large changes in the visibleCD spectra of Spirulina cells. In control CDspectra, the psi-type negative band at 689 nm,which is attributed to PSII core complexes, isdistinctly seen (Cseh et al. 2000). The CD spectraof cells exposed to low-intensity UV-B treated for5 h exhibited an increase of about 30% in the psi-type negative band at 689 nm. This negative psi-type band, measured as CD689–750, increased in amore or less time-dependent fashion (Figure 2a,insert). When cells were exposed to low-intensityUV-B together with supplemental WL, theamplitude of the psi-type (CD689–750) bandincreased during first 1 h of treatment. Uponlonger (5 h) exposure, however, this banddecreased by about 11% (Figure 2b, insert). Theintensity of the remaining peaks was marginallyaffected by UV-B exposure.
Cells exposed to high-intensity UV-B alsoexhibited a time-dependent increase in the magni-tude of the negative psi-type band at 689 nm(CD689–750), accompanied by an increase in thepeak intensity at 450 nm (Figure 2a). The psi-typenegative band at 689 nm was augmented by 49%after 5 h exposure of cells to high-intensity UV-B.When high-intensity UV-B was administered incombination with WL, the psi-type band at689 nm (CD689–750) increased after 1 h, as was thecase for low-intensity UV-B. After 1 h of expo-sure, however, the amplitude of this banddecreased. The amplitude of this band decreasedabout 33% after 5 h exposure (Figure 2b, insert).The 452 nm band also followed the same trend asseen in cells exposed to UV-B plus supplementalWL. The CD spectral analysis revealed that UV-Bexposure caused enhancement of the negativePSII-type bands at 452 nm and at 689 nm,reflecting alterations in the macroaggregation ofChl–protein complexes, and this enhancement of
negative PS II-type bands was arrested by pro-longed (5 h) supplemental WL exposure. ThePBsomes transfer energy to normal nondissociatedChl-pigment complexes of PS II (Fork and Moh-anty, 1986). The changes in the CD spectral profilecaused by UV-B exposure suggest dissociation ofChl a complexes, which uncouple the energytransfer from the PBsomes to the Chl a antennasof PSII. This may be a result of the macroaggre-gation of the PBsomes and Chl a pigment–proteincomplexes upon UV-B irradiation. Exposure ofthe cells to supplemental WL along with UV-Barrests the extent of this type of antenna proteinmacroaggregation.
Changes in PSII activity
To ascertain the effect of UV-B-induced altera-tions on the absorption and CD spectral charac-
2
0
-2
-4
-6
-8
0h
1h
3h
5h
800700600500400
0
-2
-4
-6
Wavelength, nm
0h
1h
3h
5h
689
420
8
6
4
Time, h
CD
(rel
.uni
ts)
0.6 mW cm
0.2 mW cm
420
5
4
Time, h
CD
(re
l. un
its) 0.6 mW cm
0.2 mW cm
689
CD
(rel
. uni
ts)
-2
-2
-2
-2
(a)
(b)
Figure 2. Room temperature CD spectra of Spirulina cells ex-posed to (a) high intensity of UV-B (0.6 mW cm)2) alone (b)high intensity of UV-B along with supplemental WL for dif-ferent time periods. Equal Chl (5 lg/ml) concentration weremaintained for these measurements. The spectra are typical ofatleast three independent measurements whose standard devi-ations at the peak positions are less than 5%. Inserts: CD689–750
of both low and high intensities of UV-B alone and UV-B plussupplemental WL.
185
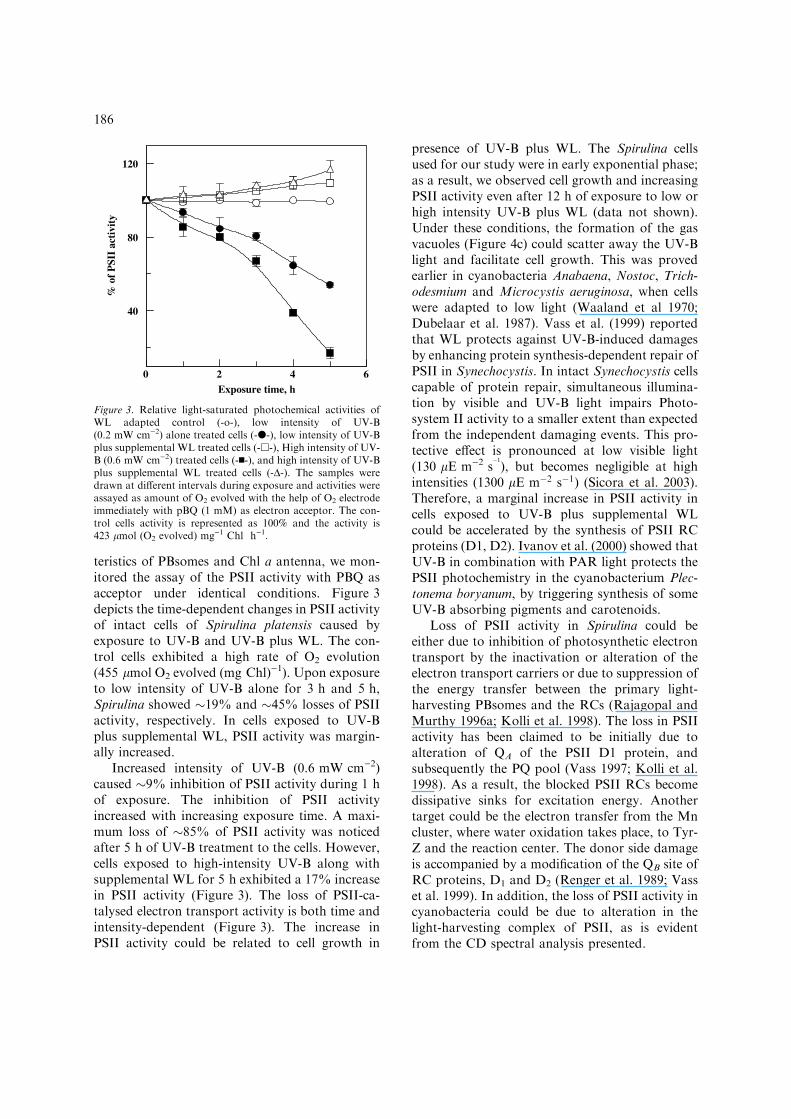
teristics of PBsomes and Chl a antenna, we mon-itored the assay of the PSII activity with PBQ asacceptor under identical conditions. Figure 3depicts the time-dependent changes in PSII activityof intact cells of Spirulina platensis caused byexposure to UV-B and UV-B plus WL. The con-trol cells exhibited a high rate of O2 evolution(455 lmol O2 evolved (mg Chl))1). Upon exposureto low intensity of UV-B alone for 3 h and 5 h,Spirulina showed �19% and �45% losses of PSIIactivity, respectively. In cells exposed to UV-Bplus supplemental WL, PSII activity was margin-ally increased.
Increased intensity of UV-B (0.6 mW cm)2)caused �9% inhibition of PSII activity during 1 hof exposure. The inhibition of PSII activityincreased with increasing exposure time. A maxi-mum loss of �85% of PSII activity was noticedafter 5 h of UV-B treatment to the cells. However,cells exposed to high-intensity UV-B along withsupplemental WL for 5 h exhibited a 17% increasein PSII activity (Figure 3). The loss of PSII-ca-talysed electron transport activity is both time andintensity-dependent (Figure 3). The increase inPSII activity could be related to cell growth in
presence of UV-B plus WL. The Spirulina cellsused for our study were in early exponential phase;as a result, we observed cell growth and increasingPSII activity even after 12 h of exposure to low orhigh intensity UV-B plus WL (data not shown).Under these conditions, the formation of the gasvacuoles (Figure 4c) could scatter away the UV-Blight and facilitate cell growth. This was provedearlier in cyanobacteria Anabaena, Nostoc, Trich-odesmium and Microcystis aeruginosa, when cellswere adapted to low light (Waaland et al 1970;Dubelaar et al. 1987). Vass et al. (1999) reportedthat WL protects against UV-B-induced damagesby enhancing protein synthesis-dependent repair ofPSII in Synechocystis. In intact Synechocystis cellscapable of protein repair, simultaneous illumina-tion by visible and UV-B light impairs Photo-system II activity to a smaller extent than expectedfrom the independent damaging events. This pro-tective effect is pronounced at low visible light(130 lE m)2 s
)1
), but becomes negligible at highintensities (1300 lE m�2 s�1) (Sicora et al. 2003).Therefore, a marginal increase in PSII activity incells exposed to UV-B plus supplemental WLcould be accelerated by the synthesis of PSII RCproteins (D1, D2). Ivanov et al. (2000) showed thatUV-B in combination with PAR light protects thePSII photochemistry in the cyanobacterium Plec-tonema boryanum, by triggering synthesis of someUV-B absorbing pigments and carotenoids.
Loss of PSII activity in Spirulina could beeither due to inhibition of photosynthetic electrontransport by the inactivation or alteration of theelectron transport carriers or due to suppression ofthe energy transfer between the primary light-harvesting PBsomes and the RCs (Rajagopal andMurthy 1996a; Kolli et al. 1998). The loss in PSIIactivity has been claimed to be initially due toalteration of QA of the PSII D1 protein, andsubsequently the PQ pool (Vass 1997; Kolli et al.1998). As a result, the blocked PSII RCs becomedissipative sinks for excitation energy. Anothertarget could be the electron transfer from the Mncluster, where water oxidation takes place, to Tyr-Z and the reaction center. The donor side damageis accompanied by a modification of the QB site ofRC proteins, D1 and D2 (Renger et al. 1989; Vasset al. 1999). In addition, the loss of PSII activity incyanobacteria could be due to alteration in thelight-harvesting complex of PSII, as is evidentfrom the CD spectral analysis presented.
6420
120
80
40
Exposure time, h
% o
f P
SII
acti
vity
Figure 3. Relative light-saturated photochemical activities ofWL adapted control (-o-), low intensity of UV-B(0.2 mW cm)2) alone treated cells (-d-), low intensity of UV-Bplus supplemental WL treated cells (-h-), High intensity of UV-B (0.6 mW cm)2) treated cells (-n-), and high intensity of UV-Bplus supplemental WL treated cells (-D-). The samples weredrawn at different intervals during exposure and activities wereassayed as amount of O2 evolved with the help of O2 electrodeimmediately with pBQ (1 mM) as electron acceptor. The con-trol cells activity is represented as 100% and the activity is423 lmol (O2 evolved) mg)1 Chl h)1.
186
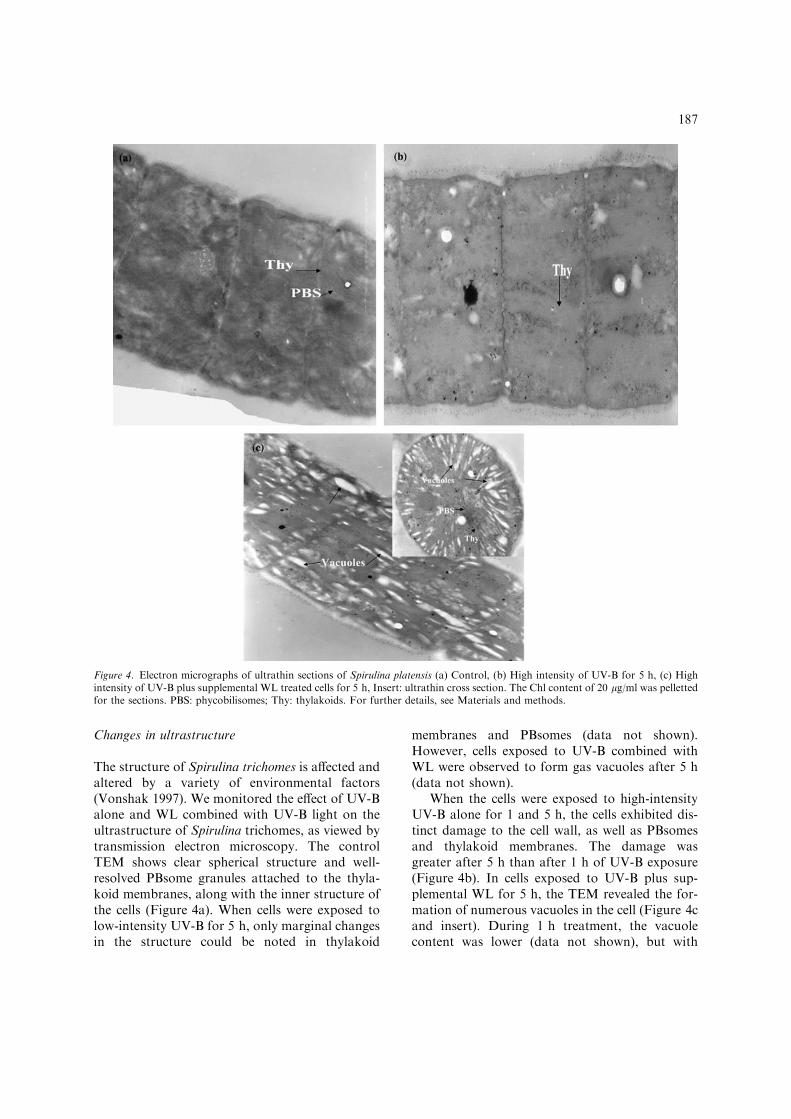
Changes in ultrastructure
The structure of Spirulina trichomes is affected andaltered by a variety of environmental factors(Vonshak 1997). We monitored the effect of UV-Balone and WL combined with UV-B light on theultrastructure of Spirulina trichomes, as viewed bytransmission electron microscopy. The controlTEM shows clear spherical structure and well-resolved PBsome granules attached to the thyla-koid membranes, along with the inner structure ofthe cells (Figure 4a). When cells were exposed tolow-intensity UV-B for 5 h, only marginal changesin the structure could be noted in thylakoid
membranes and PBsomes (data not shown).However, cells exposed to UV-B combined withWL were observed to form gas vacuoles after 5 h(data not shown).
When the cells were exposed to high-intensityUV-B alone for 1 and 5 h, the cells exhibited dis-tinct damage to the cell wall, as well as PBsomesand thylakoid membranes. The damage wasgreater after 5 h than after 1 h of UV-B exposure(Figure 4b). In cells exposed to UV-B plus sup-plemental WL for 5 h, the TEM revealed the for-mation of numerous vacuoles in the cell (Figure 4cand insert). During 1 h treatment, the vacuolecontent was lower (data not shown), but with
Figure 4. Electron micrographs of ultrathin sections of Spirulina platensis (a) Control, (b) High intensity of UV-B for 5 h, (c) Highintensity of UV-B plus supplemental WL treated cells for 5 h, Insert: ultrathin cross section. The Chl content of 20 lg/ml was pellettedfor the sections. PBS: phycobilisomes; Thy: thylakoids. For further details, see Materials and methods.
187

increasing exposure time, the vacuole content wasenhanced. The gas vacuoles look like cylindricaltubes, and could be filled with various gases.Earlier reports showed that the gas vacuoles werepermeable to different gases – H2, N2, O2, CO2,CH4 and Ar – in Anabaena (Walsby 1994). Wepresume that in our case also the gas vacuolescould be filled by several different gases.
Van Eykelenburg (1979) has reported that anincrease in light intensity or temperature lead to anincrease in vacuole concentration in Spirulina plat-ensis. Generally, cyanobacteria species, in particu-lar, Spirulina produce gas vacuoles in response tochanges in culture medium conditions, such asdilution, increase in salt or increase in light intensity(Allen 1984). The cynobacteria Nostoc, Anabaenaand Trichodesmium form gas vacuoles under lowlight (Waaland et al. 1970). It was also proven thatthe vacuoules are in fact filled with gas; they have arefractive index much lower than that of the cyto-plasm which surrounds them, and therefore theyscatter light. Consequently, they reduce the amountof light absorbed by the photosynthetic pigments.In another report on the cyanobacterium Micro-cystis aeruginosa, the vacuoles are likely to scatterthe WL and reduce the dose of WL received (Du-belaar et al. 1987). The results cited above supportour findings that, inSpirulina cells exposed toUV-Bin combination with WL, the gas vacuoles seem tobe scattering away part of the UV-B light and sub-sequently protecting photochemical activity, themacroaggregation of PBsomes and Chl–pigmentcomplexes. Thus, the formation of gas vacuolesunder UV-B plus supplemental WL seems to pro-vide protection to UV-B-induced cell damage,which appears to be an additional protectivemechanism against UV-B injury.
Conclusions
We show that the presence of a low flux density ofPAR protects the Spirulina cells from UV-B lighteven at moderately high levels in more than oneway, such as arresting the increase of macroag-gregation in PBsomes and core complexes of Chlproteins, which is associated with the negative psi-type band in the CD spectrum, and by scatteringaway the UV-B by gas vacuoles, as shown in theelectron micrographs.
Acknowledgements
The authors thank Dr Zolton Gombos for the giftof Spirulina platensis culture and also thank DrImre Vass for providing us with the UV-B lightsource. We are indebted to Dr Gyozo Garab forhelpful discussions. We also thank Craig Jolleyand Dr Nimmagadda Venkata Sridevi for criticalreading of this manuscript. This work was sup-ported by grants from OTKA (HungarianResearch Fund, T34188 and T42696).
References
Allen MM (1984) Cyanobacterial cell inclusions. Ann RevMicrobiol 38: 1–25
Araoz R and Hader DP (1997) Ultraviolet radiation inducesboth degradation and synthesis of phycobilisomes in Nostocsp.: a spectroscopic and biochemical approach. FEMSMicrobiol Ecol 23: 301–313
Brandle JR, Campbell WF, Sisson WB and Caldwell MM(1977) Net photosynthesis, electron transport capacity, andultrastructure of Pisum sativum L. exposed to ultraviolet-Bradiation. Plant Physiol 60: 165–169
Campbell D, Eriksson MJ, Oquist G, Gustafsson P and ClarkeAK (1998) The cyanobacterium Synechocystis resists UV-Bby exchanging Photosystem II reaction-center D1 proteins.Proc Natl Acad Sci USA 95: 364–369
Chauhan S, Pandey R and Singhal GS (1998) Ultraviolet-Binduced change in ultrastructure and D1/D2 proteins incyanobacteria Synechococcus sp. PCC 7942. Photosynthetica35: 161–167
Cseh Z, Rajagopal S, Tsonev T, Busheva M, Papp E and GarabG (2000) Thermal effect in chloroplast thylakoid membranes.Thermal and light stability of pigment arrays with differentlevels of structural complexity. Biochemistry 39: 15250–15257
Dubelaar GB, Visser JW and Donze M (1987) Anomalousbehaviour of forward and perpendicular light scattering of acyanobacterium owing to intracellular gas vacuoles. Cytom-etry 8: 405–412
Fork DC and Mohanty P (1986) Fluorescence and othercharacteristics of blue-green algae (cyanobacteria), red algae,and cryptomonads. In: Govindjee, Amez J and Fork DC(eds) Light Emission by Plants and Bacteria, pp 451–496.Academic Press, Orlando, Florida
FrederickJE(1993)Ultraviolet sunlight reaching theearth’s surface:a review of recent research. Photochem Photobiol 57: 175–178
Friso G, Spetea C, Giacometti GM, Vass I and Barbato R(1994a) Degradation of Photosystem II reaction centre D1protein induced by UV-B radiation in isolated thylakoids –identification and characterization of C- and N-terminalbreakdown products. Biochim Biophys Acta 1184: 78–84
Friso G, Barbato R, Giacometti GM and Barber J (1994b)Degradation of D2 protein due to UV-B radiation of thereaction centre of Photosystem II. FEBS Lett 399: 217–221
Ivanov A, Miskiewicz E, Clarke AK, Greenberg BM andHuner NP (2000) Protection of Photosystem II against UV-Band UV-B radiation in the cyanobacterium Plectonema
188

boryanum: the role of growth temperature and growthirradiance. Photochem Photobiol 72: 772–779
Jha IB, Kolli BK, Sah JF, Garab G and Mohanty P (2000) Lowdose UV-B induced modification of chromophore conforma-tion and it’s interaction with microenvironment in cyano-bacterial phycobilisomes. Ind J Biochem Biophys 37: 486–490
Kolli BK, Tiwari S and Mohanty P (1998) Ultraviolet-Binduced damages to Photosystem II in intact filaments ofSpirulina platensis. Z Naturforsch 53c: 369–377
Lao K and Glazer AN (1996) Ultraviolet-B photodestruction ofa light harvesting complex. Proc Natl Acad Sci USA 93:5258–5263
Lorenz M, Schubert H and Forster RM (1997) In vitro and invivo effects of ultraviolet-B radiation on the energy transferphycobilisomes. Photosynthetica 33: 517–527
MacKinney G (1941) Absorption of light by chlorophyllsolutions. J Biol Chem 140: 315–322
Mate Z, Sass L, Szekeres M, Vass I and Nagy F (1998) UV-B-induced differential transcription of psbA genes encoding theD1 protein of Photosystem II in the cyanobacteriumsynechocystis 6803. J Biol Chem 273: 17439–17444
Melis A, Nemson JA and Harrison MA (1992) Damage tofunctional components and partial degradation of PSIIreaction centre protein upon chloroplast exposure to ultra-violet-B radiation. Biochim Biophys Acta 1109: 312–320
Murthy SDS and Rajagopal S (1995) UV-B radiation inducedalterations the bioenergetic processes of photosynthesis.Photosynthetica 31: 481–487
Pandey R, Chauhan S and Singhal GS (1997) UV-B inducedphotodamage to phycobilisomes of Synechococcus sp. PCC7942. J Photochem Photobiol (B. Biol) 40: 228–232
Rajagopal S and Murthy SDS (1996a) Ultraviolet-B radiationinduced alterations in photosynthetic electron transportactivities of the cyanobacterium Synechococcus 6301. Photo-synthetica 32: 281–284
Rajagopal S and Murthy SDS (1996b) Short-term effect ofultraviolet-B radiation on Photosystem 2 photochemistry inthe cyanobacterium Synechococcus 6301. Biol Plant 38:129–132
Rajagopal S, Jha IB, Murthy SDS and Mohanty P (1998)Ultraviolet-B effects on Spirulina platensis cell: modificationof chromophore–protein interaction and energy transfercharacteristics of phycobilisomes. Biochem Biophys ResCommun 249: 172–177
Rajagopal S, Mohanty P and Murthy SDS (1999) Protection ofenergy transfer process in isolated phycobilisome by whitelight from ultraviolet-B induced damage in the cyanobacte-rium, Spirulina platensis. Photosynthetica 36: 617–620
Rajagopal S, Murthy SDS and Mohanty P (2000) Effect ofultraviolet-B radiation on intact cells of the cyanobacteriumSpirulina platensis: characterization of the alterations in thyla-koid membranes. J Photochem Photobiol (B. Biol) 54: 61–66
Renger G, Volker HJ, Eckert R, Fromme R, Hohm-Veit S andHohm-Veit and Graber P (1989) On the mechanism of
Photosystem II deterioration by UV-B irradiation. Photo-chem Photobiol 49: 97–105
Sah JF, Kolli BK, Srivastava M and Mohanty P (1998) Effectsof ultraviolet-B radiation on phycobilisomes of Synechococ-cus PCC 7942: alterations in conformation and energytransfer characteristics. Biochem Mol Biol Int 44: 245–257
Sicora C, Mate Z, Vass I (2003) The interaction of visible UV-Blight during photodamage and repair of Photosystem II.Photosynth Res 75: 127–137
Simidjiev I, Barzda V, Mustardy L and Garab G (1997)Isolation of lamellar aggregates of the light-harvestingchlorophyll a/b protein complex of Photosystem II withlong-range chiral order and structural flexibility. AnalBiochem 250: 169–175
Sinha RP, Lebert M, Kumar A, Kumar HD and Hader DP(1995) Disintegration of phycobilisomes in a rice fieldcyanobacterium Nostoc sp. following UV irradiation. Bio-chem and Mol Biol Int 37: 697–706
Strid A, Chow WS and Anderson JM (1994) Ultraviolet-Bdamage and protections at the molecular level in plants.Photosynth Res 39: 475–489
Tichy M, Lupınkova L, Sicorac C, Vass I, Kuvikova S, Prasil Oand Komenda J (2003) Synechocystis 6803 mutants express-ing distinct forms of the Photosystem II D1 protein fromSynechococcus 7942: relationship between the psbA codingregion and sensitivity to visible and UV-B radiation. BiochimBiophys Acta 1605: 55–66
Van TK, Garrard LA and West SH (1976) Effects of UV-Bradiation on net photosynthesis of some crop plants. CropSci 16: 715–718
Van Eykelenburg C (1979) The ultrasturcture of Spirulinaplatensis in relation to temperature and light intensity.Antonie Van Leeuwenhoek 45: 369–90
Vass I (1997) Adverse effects of UV-B light on the structureand functions of the photosynthetic apparatus. In: Pessara-kali M (ed) Handbook of Photosynthesis, pp 931–946.Marcel Dekker, New York
Vass I, Kirilovsky D and Etienne AL (1999) UV-B radiation-induced donor- and acceptor-side modifications of Photo-system II in the cyanobacterium Synechocystis sp. PCC 6803.Biochemistry 38: 12786–12794
Vass I, Kirilovsky D, Perwoska I, Mate Z, Nagy F and EtienneAL (2000) UV-B radiation induced exchange of the D1reaction centre subunits produced from the psbA2 and psbA3genes in the cyanobacterium Synechocystis sp. PCC 6803.Eur J Biochem 267: 2640–2648
Vonshak A (1997) Use of Spirulina platensis (Arthrospira):Physiology, Cell biology and Biotechnology. In: Vonshak A(ed) 205 pp. Taylor and Francis.
Waaland JR, Waaland SD and Branton D (1970) Gas vacuoles:light shielding in blue-green algae. J Cell Biol 48: 212–215
Walsby AE (1994) Gas vesicles. Microbiol Rev 58: 94–144
189




















