
Dodecanol, metabolite of entomopathogenic fungus ... - Nature
Upload
independentCategory
view
3download
0
Production and Characterization of β-glucosidase Obtainedby the Solid-State Cultivation of the Thermophilic FungusThermomucor indicae-seudaticae N31
Josiani de Cassia Pereira & Rodrigo Simões Ribeiro Leite &
Heloiza Ferreira Alves do Prado & Daniela Alonso Bocchini Martins &Eleni Gomes & Roberto da Silva
Received: 7 May 2014 /Accepted: 15 October 2014# Springer Science+Business Media New York 2014
Abstract In this paper, several agro-industrial wastes (soybean meal and wheat straw, rice andpeanut husks, corn cob and corn stover, and sugarcane bagasse) were tested for the productionof β-glucosidase by the cultivation of thermophilic fungus Thermomucor indicae-seudaticaeN31 in solid-state fermentation (SSF). Among the tested substrates, the highest yields wereobtained in soybean meal. Other fermentation parameters were also evaluated, such as initialpH, merge substrates, and fermentation time, as well as the physicochemical characterizationof the enzyme. The best results were obtained after 192 h of fermentation with the initial pHadjusted to 6.0. The substrate mixture did not improve the enzyme production by microor-ganism. The β-glucosidase showed best catalytic activity at pH 4.5 and at 75 °C and remainedstable in the pH range from 4.5 to 9.5 and the temperature range 40–75 °C. The enzymeshowed 80 % of its activity at a concentration of 15 mM glucose and remained stable up to20 % ethanol.
Keywords Thermomucor indicae-seudaticaeN31 .β-glucosidase . Solid-state fermentation
Introduction
Cellulose is the most abundant organic compound in the biosphere, accounting for more thanhalf of all organic carbon on the planet [1]. Regarding this, the development of technologiesthat use this polysaccharide as a source of energy to replace fossil fuels has been offundamental importance for human maintenance on the planet [2].
Appl Biochem BiotechnolDOI 10.1007/s12010-014-1332-1
J. de Cassia Pereira (*) : H. F. A. do Prado :D. A. Bocchini Martins : E. Gomes : R. da SilvaLaboratory of Biochemistry and Applied Microbiology, Universidade Estadual Paulista (UNESP),Av. Cristóvão Colombo, 2265, 15054-000 São José do Rio PretoSão Paulo, Brazile-mail: [email protected]
R. S. R. LeiteBiological na Environment Sciences Faculty, FCBA, Universidade Federal da Grande Dourados (UFGD),Km 12, Rodovia Dourados—Itahum, Dourados, MS 79804-970, Brazil

The enzymatic hydrolysis of the cellulose material involves the synergistic action of at leastthree different enzymes: endoglucanase hydrolyzes the cellulose chains internally, resulting in arapid reduction in degree of polymerization; exoglucanase removes cellobiose units from theends of the molecule; and β-glucosidase hydrolyzes cellobiose to glucose and othercellodextrins. All these three enzymes suffer from complex catabolite repression by the finalproducts of hydrolysis. The cellobiose inhibits the action of endo- and exoglucanases, and thus,the β-glucosidase contributes to the continuity of this process, preventing the accumulation ofthis disaccharide on the medium, thereby playing a crucial role in the enzymatic degradation ofcellulose [3]. The presence of β-glucosidase in the winemaking process favors the release offlavor compounds from their glycosidic precursors. Such compounds are collectively calledterpenes; among them are nerol, α-terpineol, geraniol, linalool, citronellol, and also others that,when glycosylated, have low volatility and contribute little to the aroma of wine [4]. Endog-enous β-glucosidase of grape does not work efficiently during the winemaking process due tothe low stability of these enzymes [5]. An alternative to increase the aroma of wines is the use ofmicrobial enzymes that can stand the requirements of the process. Thus, different studies havebeen carried out to find enzymes that can be used in winemaking [5–8]. The microbial β-glucosidases can also be used by unglycosylated isoflavones present in soybean, favoring theuptake of these compounds by the human intestinal tract. In this way, the β-glycosidasescontribute to increase digestibility of foods derived from soybeans [9]. Previous studies havereported that a diet rich in isoflavones contributes to the control and prevention of many chronicdiseases, such as cancer (breast, prostate, and colon), osteoporosis, menopausal symptoms,cardiovascular, and other diseases [10].
The different possibilities for the industrial application of β-glycosidases justify theincessant search for new sources of microbial enzymes that can be produced with highstructural stability in fermentation media of low economic value [11–14]. Recently, a signif-icant production of β-glucosidase was observed from thermophilic fungus Thermomucorindicae-seudaticae N31 grown in solid-state fermentation. The absence of reports in thescientific literature of this fact encouraged us to study the production of this enzyme by thefungus in different agro-industrial residues. The physicochemical characteristics and theindustrial applicability of β-glucosidase were also evaluated. The use of these residues forthe production of β-glucosidase and other cellulases and hemicellulases has attracted muchattention because of the reduced cost of enzyme production and also the reduction ofenvironmental pollution caused by the accumulation of lignocellulose [3].
Materials and Methods
Microorganism
The thermophilic fungus T. indicae-seudaticae N31 was isolated from piles of sugarcanebagasse in Olimpia, SP, Brazil. The culture was maintained on Sabouraud Dextrose agar(Oxoid) at 4 °C.
Evaluation of Parameters Solid-State Fermentation
Different agro-industrial wastes were used in order to select the best substrate for theproduction of β-glucosidase by the microorganism, including wheat bran, soybeanmeal, rice husk, sugarcane bagasse, corn cob, and corn stover. The best substrate forthe production of the enzyme was mixed in different proportions with inert substrate (rice
Appl Biochem Biotechnol

husk) in different proportions (5:0 g, 4:1 g, 3:2 g, 2.5:2.5 g, 2:3 g, 1:4 g, 0:5 g). Afterdetermination of the parameters previously described, samples were taken every 48 h to acessthe β-glucosidase production by the fungus on the basis of fermentation time; the last samplewas taken after 336 h of cultivation.
Inoculum
The fungus was grown in 250-ml Erlenmeyer flasks containing 40 ml of agar SabouraudDextrose maintained at a temperature of 45 °C for 48 h. The suspension of the microorganismwas obtained by gently scraping the surface of the culture medium using 50 ml of nutrientsolution (0.1 % ammonium sulfate, 0.1 % magnesium sulfate heptahydrate, 0.1 % ammoniumnitrate). Inoculation of the fungus on the substrates was given by transferring 5 ml of thissuspension in Erlenmeyer flasks containing 250 ml of 5 g substrate with final moisture to 80%.The fermentationmedia (substrates) and all materials used for inoculation of the microorganismwere properly autoclaved at 121 °C for 30 min to ensure uniformity of the culture.
Enzyme Extraction
For the enzyme extraction, 50 ml of distilled water in fermentation media was added and keptunder agitation for 1 h. Subsequently, the material was centrifuged (10,000×g/20 min) at 4 °C,and the supernatant (enzyme extract) was used in the tests described below.
β-Glucosidase Assays
The β-glucosidase activity was determined by adding 50 μl of enzyme extract, 250 μL ofbuffer 0.1 M sodium acetate, pH 5.0, and 250 μl of p-nitrophenyl β-D-glucopyranoside(4 mmol l−1 pNPβG, Sigma) reacting for 10 min at 60 °C. The reaction was stopped byadding 2 ml of 2 M sodium carbonate. The released p-nitrophenol was quantified byspectrophotometry at 410 nm. One unit of enzyme activity was defined as the amount ofenzyme required to release 1 μmol of p-nitrophenol per minute of reaction.
Enzyme Characterization
pH and Temperature Effects
The optimum pH was determined by measuring the enzyme activity at 60 °C at different pHvalues (3.0 to 8.0); McIlvaine buffer 0.1 M was used for the assays. The optimum temperaturewas determined by measuring the enzyme activity at different temperatures (30–80 °C), withthe respective optimum pH of the enzyme. The pH stability of the enzyme was assessed byincubating it for 24 h at 25 °C at different pH values. The buffers used were 0.1 M McIlvaine(3.0 to 8.0), Tris–HCl 0.1 M (8.0 to 8.5), and 0.1 M Glycine-NaOH (8.5 to 10.5). The thermalstability was studied by incubating the enzyme for 1 h at temperature values ranging from 30to 70 °C. The residual activities were determined under optimal conditions of pH andtemperature of the enzyme.
Determination of Kinetic Parameters
The kinetic parameters were calculated for the enzymatic activity of β-glucosidase at pH 5.0 insodium acetate and temperature conditions at 60 °C. The activities were performed in the
Appl Biochem Biotechnol

absence and presence of glucose; different glucose concentrations, according to the finalvolume, were used in testing the activity. The Km and Vmax values were obtained graphicallyusing Lineweaver-Burk plots.
Glucose and Ethanol Effects on the Enzyme Activity
The effect of glucose and ethanol on the enzyme activity was determinated by using pNPβG(2 mmol l−1) as the substrate. The enzymatic activity was quantified with the addition of thesecompounds at different concentrations in the reaction mixture (0–100 mmol l−1 glucose and 0–40 % ethanol).
Analyze Statistic
Statistical analyses of the data included a one-way ANOVA followed by Fisher’s F post hoctest (when the assumptions of normality and homogeneity of variances were satisfied) with a5 % significance level. Tukey’s test parametric analysis of variance were applied to non-normal data. All analyses were run in the STATISTICA 7.0 package.
Results and Discussion
Evaluation of Parameters in the Solid-State Fermentation
Among the organic residues tested in this study, soybean meal and wheat bran were better forthe production of β-glucosidase by the fungus T. indicae-seudaticae N31, with about 2.35 and1.83 U/ml respectively, after 96-h incubation (Table 1). Regarding the analysis of variance, theresults were significant with ANOVA F=318; p<0.05.
Leite et al. [14] highlighted the β-glucosidase production by the filamentous fungusThermoascus aurantiacus. However, the highest yield was obtained from cultivation in wheatbran. According to Ferrarezi et al. [15], the fungus T. indicae-seudaticae N31 has significantproduction of lipase, which probably favored the growth of microorganisms in soybean mealand therefore contributed to the higher enzyme production in this residue. Soybean meal is richin cellulose and hemicellulose substrate complex, containing about 40 % cellulose and 30 %xylan. The high concentration of plant cell wall polysaccharides in the composition of theculture medium can favor the production of cellulolytic enzymes complex [16]. To reducecompaction of the fermentation medium and promote mass transportation, rice husk (substraterich in lignin and quite inert to fungus growth) was mixed with soybean meal in different
Table 1 β-glicosidase productionon the different agro-industrialwastes as substrates after 96 h ofcultivation
Substrates U/ml
Soybean meal 2.35±0.09
Wheat straw 1.83±0.05
Peanut husk 0.81±0.05
Corn cob 0.52±0.04
Corn stover 0.24±0.03
Rice husk 0.15±0.04
Sugarcane bagasse 0.13±0
Appl Biochem Biotechnol

proportions. The results demonstrate the increased production of the enzyme, about 2.8 U/mlin the cultivation carried out with 4 g of soybean meal and 1 g of rice husk (Table 2).Furthermore, analysis of variance showed that the results were significant, with ANOVA F=32; p<0.05.
The mixture of organic residues can be used in solid-state fermentation (SSF) for differentpurposes, such as facilitating the mass transfer during the fermentation process, increasing orreducing the absorption of water by the solid matrix, and favoring the balance of carbon andnitrogen composition of culture medium, among others. In this context, different studies havereported significant production of enzymes by filamentous fungi growing on mixtures of agro-industrial waste by solid-state fermentation [17, 16]. Later, the tests described, the cultivationtime for enzyme production was evaluated for a total of 336 h of fermentation. The increasedproduction of β-glucosidase, about 4.18 U/ml (41.8 U/g), was obtained after 192 h of cultureusing the 4:1 ratio (g) of soybean meal to rice husk (Fig. 1). The analysis of variance of theresults were significant, with ANOVA F=80; p<0.05.
The production of β-glucosidase presented by the fungus T. indicae-seudaticae N31 isquite significant when compared with some other fungi reported in the literature. Leite et al.[18] obtained approximately 1.3 U/ml of β-glucosidase after 120 h of cultivating the fungusAureobasidium pullulans in solid-state fermentation using wheat bran as substrate. Brijwaniet al. [16] reported production of 1.78 U/ml of β-glucosidase after 96 h of fermentation by co-cultivation of fungi Trichoderma reesei and Aspergillus oryzae in solid-state fermentationusing a mixture of soybean hulls and wheat bran in the ratio of 4:1, respectively.
pH and Temperature Effects on the Enzymatic Activities
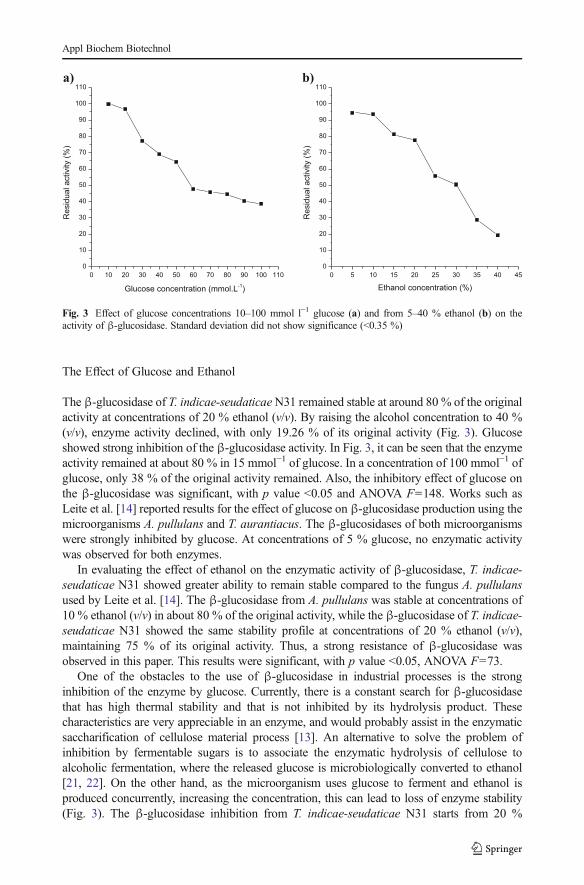
The β-glucosidase produced by the fungus T. indicae-seudaticaeN31 showed highest catalyticactivity at an optimum pH of 4.5 and an optimum temperature of 75 °C. The enzyme showedsignificant stability to pH and temperature at pH 4.5 to 9.5 (after 24 h of incubation) andretained its original activity when incubated for 1 h at 75 °C (Fig. 2). Previous studiesconfirmed the major activity of β-glucosidase produced by fungi at pHs close to that describedin this work [14, 19]. According to the analysis of variance, all the results were significantswith p value <0.05 and ANOVA F=414, F=15.9 (optimum pH and pH stability, respectively),F=282 and F=155 (optimum temperature and temperature stability, respectively).
Enzymes produced by thermophilic strains generally exhibit high structural stability,favoring their application in different industrial processes: a fact that justifies the constantinvestigation of microorganisms that have the ability to grow at elevated temperatures [20].Previous works describe high values of great expressive and stability of β-glucosidaseproduced by thermophilic fungi, which corroborates the data obtained in this study [14].
Table 2 β-glucosidase productionin medium composed of differentproportions of soybean meal andrice husk after 96 h of fermentation
Substrate proportion (g)Soybean meal : rice husk
U/ml
5:0 2.23±0.23
4:1 2.8±0.3
3:2 1.45±0.11
2.5:2.5 1.79±0.13
2:3 1.13±0.14
1:4 0.36±0.105
0:5 0.06±0.03
Appl Biochem Biotechnol
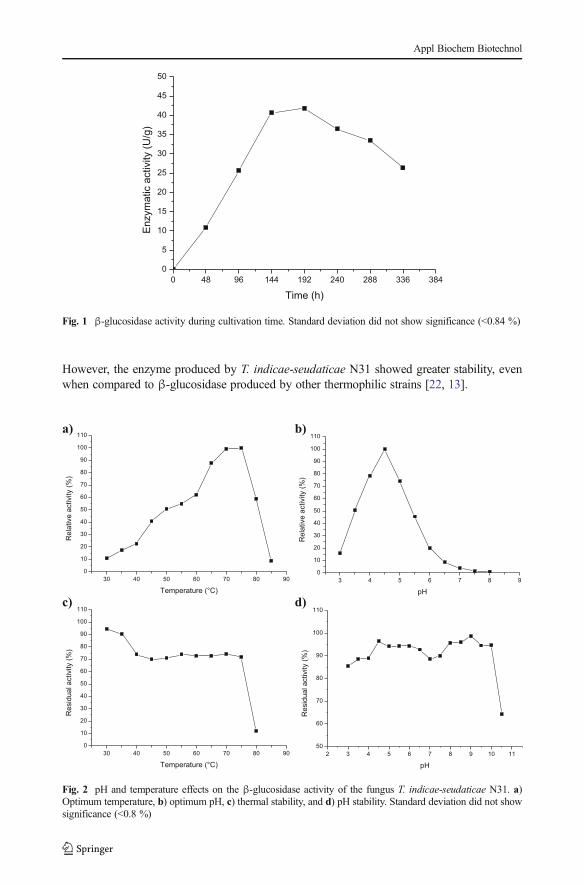
However, the enzyme produced by T. indicae-seudaticae N31 showed greater stability, evenwhen compared to β-glucosidase produced by other thermophilic strains [22, 13].
0 48 96 144 192 240 288 336 3840
5
10
15
20
25
30
35
40
45
50
Enzy
mat
ic a
ctiv
ity (U
/g)
Time (h)
Fig. 1 β-glucosidase activity during cultivation time. Standard deviation did not show significance (<0.84 %)
30 40 50 60 70 80 900
10
20
30
40
50
60
70
80
90
100
110
2 3 4 5 6 7 8 9 10 1150
60
70
80
90
100
110
30 40 50 60 70 80 900
10
20
30
40
50
60
70
80
90
100
110
3 4 5 6 7 8 90
10
20
30
40
50
60
70
80
90
100
110
Rel
ativ
e ac
tivity
(%)
Temperature (°C)
Res
idua
l act
ivity
(%)
pH
Res
idua
l act
ivity
(%)
Temperature (°C)
Rel
ativ
e ac
tivity
(%)
pHc)
a) b)
d)
Fig. 2 pH and temperature effects on the β-glucosidase activity of the fungus T. indicae-seudaticae N31. a)Optimum temperature, b) optimum pH, c) thermal stability, and d) pH stability. Standard deviation did not showsignificance (<0.8 %)
Appl Biochem Biotechnol
The Effect of Glucose and Ethanol
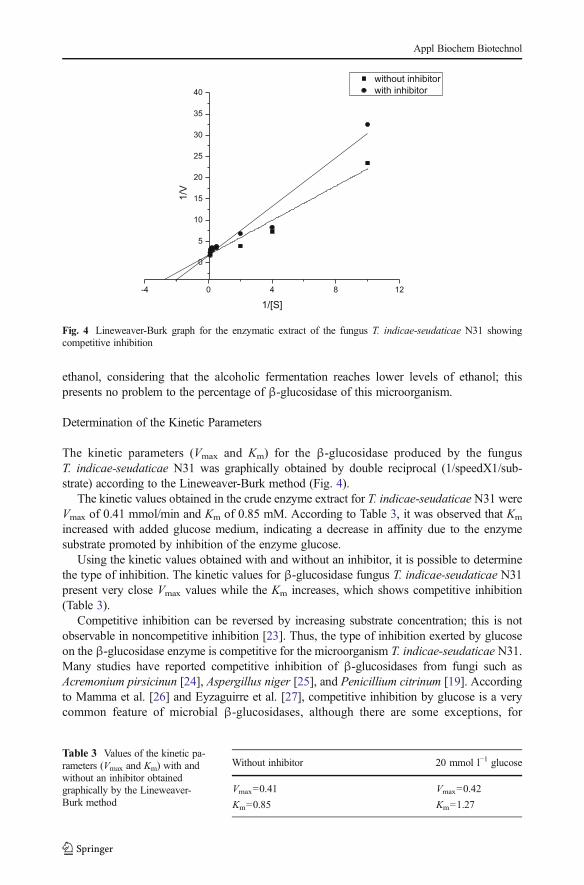
The β-glucosidase of T. indicae-seudaticaeN31 remained stable at around 80 % of the originalactivity at concentrations of 20 % ethanol (v/v). By raising the alcohol concentration to 40 %(v/v), enzyme activity declined, with only 19.26 % of its original activity (Fig. 3). Glucoseshowed strong inhibition of the β-glucosidase activity. In Fig. 3, it can be seen that the enzymeactivity remained at about 80 % in 15 mmol−1 of glucose. In a concentration of 100 mmol−1 ofglucose, only 38 % of the original activity remained. Also, the inhibitory effect of glucose onthe β-glucosidase was significant, with p value <0.05 and ANOVA F=148. Works such asLeite et al. [14] reported results for the effect of glucose on β-glucosidase production using themicroorganisms A. pullulans and T. aurantiacus. The β-glucosidases of both microorganismswere strongly inhibited by glucose. At concentrations of 5 % glucose, no enzymatic activitywas observed for both enzymes.
In evaluating the effect of ethanol on the enzymatic activity of β-glucosidase, T. indicae-seudaticae N31 showed greater ability to remain stable compared to the fungus A. pullulansused by Leite et al. [14]. The β-glucosidase from A. pullulans was stable at concentrations of10 % ethanol (v/v) in about 80 % of the original activity, while the β-glucosidase of T. indicae-seudaticae N31 showed the same stability profile at concentrations of 20 % ethanol (v/v),maintaining 75 % of its original activity. Thus, a strong resistance of β-glucosidase wasobserved in this paper. This results were significant, with p value <0.05, ANOVA F=73.
One of the obstacles to the use of β-glucosidase in industrial processes is the stronginhibition of the enzyme by glucose. Currently, there is a constant search for β-glucosidasethat has high thermal stability and that is not inhibited by its hydrolysis product. Thesecharacteristics are very appreciable in an enzyme, and would probably assist in the enzymaticsaccharification of cellulose material process [13]. An alternative to solve the problem ofinhibition by fermentable sugars is to associate the enzymatic hydrolysis of cellulose toalcoholic fermentation, where the released glucose is microbiologically converted to ethanol[21, 22]. On the other hand, as the microorganism uses glucose to ferment and ethanol isproduced concurrently, increasing the concentration, this can lead to loss of enzyme stability(Fig. 3). The β-glucosidase inhibition from T. indicae-seudaticae N31 starts from 20 %
0 10 20 30 40 50 60 70 80 90 100 1100
10
20
30
40
50
60
70
80
90
100
110
0 5 10 15 20 25 30 35 40 450
10
20
30
40
50
60
70
80
90
100
110
Res
idua
l act
ivity
(%)
Glucose concentration (mmol.L-1)
Res
idua
l act
ivity
(%)
Ethanol concentration (%)
a) b)
Fig. 3 Effect of glucose concentrations 10–100 mmol l−1 glucose (a) and from 5–40 % ethanol (b) on theactivity of β-glucosidase. Standard deviation did not show significance (<0.35 %)
Appl Biochem Biotechnol
ethanol, considering that the alcoholic fermentation reaches lower levels of ethanol; thispresents no problem to the percentage of β-glucosidase of this microorganism.
Determination of the Kinetic Parameters
The kinetic parameters (Vmax and Km) for the β-glucosidase produced by the fungusT. indicae-seudaticae N31 was graphically obtained by double reciprocal (1/speedX1/sub-strate) according to the Lineweaver-Burk method (Fig. 4).
The kinetic values obtained in the crude enzyme extract for T. indicae-seudaticae N31 wereVmax of 0.41 mmol/min and Km of 0.85 mM. According to Table 3, it was observed that Km
increased with added glucose medium, indicating a decrease in affinity due to the enzymesubstrate promoted by inhibition of the enzyme glucose.
Using the kinetic values obtained with and without an inhibitor, it is possible to determinethe type of inhibition. The kinetic values for β-glucosidase fungus T. indicae-seudaticae N31present very close Vmax values while the Km increases, which shows competitive inhibition(Table 3).
Competitive inhibition can be reversed by increasing substrate concentration; this is notobservable in noncompetitive inhibition [23]. Thus, the type of inhibition exerted by glucoseon the β-glucosidase enzyme is competitive for the microorganism T. indicae-seudaticae N31.Many studies have reported competitive inhibition of β-glucosidases from fungi such asAcremonium pirsicinun [24], Aspergillus niger [25], and Penicillium citrinum [19]. Accordingto Mamma et al. [26] and Eyzaguirre et al. [27], competitive inhibition by glucose is a verycommon feature of microbial β-glucosidases, although there are some exceptions, for
-4 0 4 8 12
0
5
10
15
20
25
30
35
40without inhibitorwith inhibitor
1/V
1/[S]
Fig. 4 Lineweaver-Burk graph for the enzymatic extract of the fungus T. indicae-seudaticae N31 showingcompetitive inhibition
Table 3 Values of the kinetic pa-rameters (Vmax and Km) with andwithout an inhibitor obtainedgraphically by the Lineweaver-Burk method
Without inhibitor 20 mmol l−1 glucose
Vmax=0.41 Vmax=0.42
Km=0.85 Km=1.27
Appl Biochem Biotechnol

example, β-glucosidase from the fungus Lenzites trabea [28] and Scytalidium lignicola [29]which showed no competitive inhibition.
Conclusions
The microorganism T. indicae-seudaticae N31 showed significant potential for growth andproduction of β-glucosidase when grown in SSF using soybean meal as substrate. This factconfirms the possibility of using this waste to reduce the cost of production of microbialenzymes; besides, it contributes to the decrease in the accumulation of these compounds in theenvironment. The β-glucosidase produced by the microorganism T. indicae-seudaticae N31presents optimal conditions for activity at pHs near neutrality and under elevated temperatures,in addition to showing a broad spectrum of stability to both pH and temperature. Moreover, theβ-glucosidase has high stability against high concentrations of glucose and ethanol. Suchcharacteristics are highly desirable in industrial applications. Finally, the results obtained so farstimulate the continuing work, particularly with regard to the studies of lignocellulosicsaccharification to obtain fermentable sugars that can be converted into ethanol secondgeneration materials.
Acknowledgments The authors wish to thank Fundação de Amparo à Pesquisa do Estado de São Paulo(FAPESP) and Conselho Nacional de Pesquisa (CNPq) for financial support.
References
1. Bhattacharya, D., Germinario, L. T., & Winter, W. T. (2008). Carbohydrate Polymers, 73, 371–377.2. Schuchardt, U., Ribeiro, M., & Gonçalves, A. R. (2001). Quimica Nova, 24, 247–251.3. Daroit, D. J., Silveira, S. T., Hertz, P. F., & Brandelli, A. (2007). Process Biochemistry, 42, 904–908.4. Bhatia, Y., Mishra, S., & Bisaria, V. S. (2002). CRC Critical Reviews in Biotechnology, 22, 375–407.5. Barbagallo, R. N., Spagna, G., Palmeria, R., Restuccia, C., & Giudicib, P. (2004). Enzyme and Microbial
Technology, 35, 58–66.6. Hernandez, L. F., Espinosa, J. C., Fernandez-Gonzalez, M., & Briones, A. (2003). International Journal of
Food Microbiology, 80, 171–176.7. Spagna, G., et al. (2002). Journal of the Science of Food and Agriculture, 82, 1867–1874.8. Villena, M. A., Iranzo, J. F. U., Gundllapalli, S. B., Otero, R. R. C., & Pérez, A. I. B. (2006). Enzyme
Microbial Technology, 39, 229–234.9. Otieno, D. O., & Shah, N. P. (2007). Journal of Applied Microbiology, 103, 910–912.10. Chung, H., Hogan, S., Zhang, L., Rainey, K., & Zhou, K. (2008). Journal of Agricultural and Food
Chemistry, 56, 23.11. Parry, N. J., Beever, D. E., Owen, E., Vandenbergue, I., & Van Beeumen, J. (2001). Biochemistry Journal,
353, 117–127.12. Iembo, T., et al. (2002). Applied Biochemistry and Microbiology, 38, 549–552.13. Zanoelo, F. F., Polizeli, M. L., & Jorge, J. A. (2004). Journal of Industrial Microbiology and Biotechnology,
31, 170–176.14. Leite, R. S. R., Alves-Prado, H. F., Cabral, H., Pagnocca, F. C., Gomes, E., & Da-Silva, R. (2008). Enzyme
and Microbial Technology, 43, 391–395.15. Ferrarezi, A. L., Chiachio-Egea, T., Zortarelli-Filho, I. J., Bonilla-Rodrigues, G. O., Da-Silva, R., & Gomes,
E. (2009). New Biotechnology, 25, 89.16. Brijwani, K., Oberoi, H. S., & Vadlani, P. V. (2009). Process Biochemistry, 45, 120–128.17. Jecu, L. (2000). Industrial Crop Production, 11, 1–5.18. Leite, R. S. R., Bocchini, D. A., Martins, E. S., Silva, D., Gomes, E., & Silva, R. (2007). Applied
Biochemistry and Biotechnology, 137, 281–288.19. I-Son, N., Chen-Wei, L., Shuang-Pi, C., Jiun-Ly, C., Po, T. C., Chii-Gong, T., Su-May, Y., & Tuan-Hua, D.
H. (2010). Bioresource Technology, 10, 1310–1317.
Appl Biochem Biotechnol

20. Gomes, E., Guez, M. A. U., Martin, N., & Da-Silva, R. (2007). Quimica Nova, 30, 136–145.21. Cardona, C. A., & Sanchez, O. J. (2007). Bioresource Technology, 98, 2415–2457.22. Saha, B. C., Freer, S. N., & Bothast, R. J. (1994). Applied and Environmental Microbiology, 60, 3774–3780.23. Marques, P. R. B. O., & Yamanaka, H. (2008). Quimica Nova, 7, 1791–1799.24. Pitson, S. M., Seviour, R. J., & McDougall, B. M. (1997). Enzyme and Microbial Technology, 21, 182–190.25. Le Traon-Masson, M.-P., & Pellerin, P. (1998). Enzyme Microbial Technology, 22, 374–382.26. Mamma, D., Hatzinikolaou, D. G., & Christakopoulos, P. (2004). Journal of Molecular Catalysis B:
Enzymatic, 27, 183–190.27. Eyzaguirre, J.; Hidalgo, M.; Leschot, A. (2005). In: Taylor & Francis Group, LLC. Handbook of
Carbohydrate Engineering, Charpter 23, 645–675.28. Herr, D., Baumer, F., & Dellweg, H. (1978). European Journal of Applied Microbiology and Biotechnology,
5, 29–36.29. Desai, J. D., Ray, R. M., & Patel, N. P. (1983). Biotechnology and Bioengineering, 25, 307–313.
Appl Biochem Biotechnol
Copyright © 2022 FDOKUMEN




















